Область применения.
Настоящее изобретение относится к биотехнологии и может быть использовано для клонирования и высокоэффективной экспрессии целевых белков в клетках Escherichia coli, в частности для экспрессии потенциально токсичных продуктов.
Предлагаются системы для регулируемого синтеза гетерологичного белка, характеризующиеся высоким максимальным уровнем экспрессии целевого гена в условиях индукции и большим диапазоном регуляции, которые представляют собой штамм E. coli с геном белка-антирепрессора бактериофага N15, находящимся в хромосоме под контролем индуцируемого промотора, стабильно трансформированный вектором экспрессии, содержащим тот же, что и в штамме-хозяине, или другой индуцируемый промотор, репликационный ген repA и ген белка-репрессора фага N15.
Актуальность
Escherichia coli является одним из наиболее привлекательных объектов, используемых для продукции гетерологичных белков вследствие того, что она способна к быстрому росту до высоких концентраций на простых средах, ее генетика достаточно хорошо охарактеризована, доступен большой набор клонирующих и экспрессионных векторов и мутантных штаммов.
Идеальный экспрессионный вектор должен обеспечивать высокий уровень синтеза продукта в случае индукции при минимально низком базовом уровне экспрессии, что необходимо, в первую очередь, при экспрессии продуктов, которые могут быть токсичны для клетки.
Несмотря на это, большинство современных систем для синтеза гетерологичных белков в E. coli обладают относительно узким диапазоном регулирования. Так, например, диапазон регуляции синтеза продукта в коммерческой системе pQE фирмы Qiagen составляет всего 100-200 раз. Вследствие этого системы, обладающие низким базовым уровнем экспрессии, не позволяют достичь высокого уровня синтеза продукта при полной индукции, а системы с высоким максимальным уровнем имеют обычно и достаточно высокий базовый уровень экспрессии, что делает их мало пригодными для получения чужеродных полипептидных продуктов, которые могут быть токсичными для бактериальной клетки.
Таким образом, задача создания экспрессионных систем, сочетающих высокий максимальный уровень экспрессии продукта с большим диапазоном регуляции (т.е. низким базовым уровнем), продолжает оставаться актуальной.
Уровень техники
Регуляция уровня синтеза целевого белка в существующих экспрессионных системах достигается за счет использования промоторов, уровень активности которых (т.е. количество актов инициации синтеза мРНК за единицу времени) может быть искусственно изменен, и/или путем изменения числа копий вектора, определяемого участком, который ответственен за репликацию.
А) Регулируемые промоторы
Большинство используемых промоторов для экспрессии генов в E. coli получены из грамм положительных бактерий и бактериофагов. Классическим примером регуляции экспрессии генов у прокариот в природе является лактозный оперон, поэтому именно его компоненты используются во многих регулируемых промоторах. Сам lac промотор, как и сходный с ним lacUV5, является достаточно слабым и редко используется для высокоэффективного синтеза продукта (Yanisch-Perron et al., 1985). Синтетические промоторы tac и trc, включающие (-35) район trp промотора и (-10) район lac промотора, являются значительно более сильными и обеспечивают уровни экспрессии до 15-30% общего клеточного белка (de Boer et al., 1983; Brosius et al., 1985). Такие промоторы контролируются репрессором лактозного оперона: синтез продукта достигается внесением в среду аналога лактозы IPTG. Сходную структуру и регуляцию имеет гибридный промотор Т5 в векторах серии pQE фирмы Qiagen.
Одной из альтернатив регуляции экспрессии является использование позднего промотора фага Т7 (Tabor and Richardson, 1985; векторы серии рЕТ фирмы Novagen), специфически транскрибируемого РНК полимеразой фага. При этом ген Т7 РНК-полимеразы клонируется in trans под контролем промотора lacUV5, индуцируемого IPTG. Хотя эта система обеспечивает уровни экспрессии до 30-50% общего белка (Baneyx, 1999), она не лишена недостатков. Например, высокие уровни мРНК могут вызывать деградацию рибосом, а базовый уровень экспрессии Т7 РНК полимеразы может приводить к нестабильности плазмиды (Miroux and Walker, 1996). Более того, даже «пустые» рЕТ векторы могут быть токсичны для клетки в присутствии IPTG (Miroux and Walker, 1996).
Общим же недостатком всех производных lac промотора следует считать достаточно высокий базовый уровень экспрессии, препятствующий высокоэффективной экспрессии белков, которые могут быть токсичны для клетки. Так, диапазон регуляции Plac промотора составляет всего 80-100 раз, что обуславливает достаточно высокий уровень его активности в отсутствие индуктора (Guzman et al., 1995, Yanisch-Perron et al., 1985). Эта проблема сохраняется и для векторов серии рЕТ, поскольку ген Т7 РНК полимеразы в этих системах клонируется под контролем имеющего достаточно высокий базовый уровень экспрессии промотора lacUV5, индуцируемого IPTG.
Другим широко используемым регулируемым промотором является araPBAD, промотор арабинозного оперона Е. coli, коммерциализованный фирмой Intirogen. Этот промотор обладает широким диапазоном регуляции, однако он является менее эффективным, чем, например, tac промотор (Guzman et al., 1995). Кроме того, недостатком этой системы является гетерогенность клеточной популяции в условиях неполной индукции промотора, когда в некоторых клетках наблюдается полная индукция, в то время как в других - ее отсутствие (Siegele and Hu, 1997).
Наряду с бактериальными промоторами для регулируемой экспрессии используются также узнаваемые РНК-полимеразой клетки-хозяина промоторы фагов, в частности PL промотор фага лямбда. Репрессия этого промотора достигается за счет экспрессии температурочувствительного мутанта лямбда-репрессора CI857, индукция происходит при повышении температуры с 30°С до 42°С (Elvin et al., 1990). Основным недостатком этой системы экспрессии также является высокий базовый уровень экспрессии, обусловленный как неполной репрессией мутантным CI белком, так и тем, что PL промотор является холодоиндуцибельным (Giladi et al., 1995).
Б) Плазмидный репликон
Для достижения высокой «дозы гена» большинство коммерческих экспрессионных систем (см. таблицу 1) используют плазмиды с ослабленным контролем репликации, число копий которых составляет 15-60 (производные pMB1/ColE1) или несколько сот копий (например, плазмиды серии pUC, производные рМВ1) на хромосому (Yanisch-Perron et al., 1985, см. обзор Baneyx, 1999).
Описаны векторы, основанные на совместимом с рМВ1 репликоне р15А, имеющие 10-12 копий на хромосому, но в силу своей низкой копийности они дают невысокий выход продукта и обычно используются для экспрессии вспомогательных белков (Baneyx, 1999). В лабораторных условиях такие плазмиды случайно распределяются между дочерними клетками при делении и теряются с частотой 10-5-10-6 на генерацию, в основном вследствие мультимеризации (Summers, 1998). Однако если плазмида содержит ген, продукт которого вреден для клетки, то видимая скорость потери плазмиды может возрастать в тысячи раз, поскольку бесплазмидные клетки будут обладать селективным преимуществом в росте. Помимо этого, по той же причине может происходить отбор и накопление спонтанных мутаций в клонируемом гене.
Простейшее решение проблемы потери плазмиды состоит во введении в нее гена устойчивости к антибиотику и соответствующего антибиотика в среду. Недостатком этого подхода является, во-первых, возможность инактивации антибиотика в среде, сопровождаемая снижением селективного давления, а во-вторых, его присутствие в продукте, что может быть недопустимо с точки зрения биобезопасности.
Известен ряд альтернативных стратегий, основанных на использовании постсегрегационного киллинга бесплазмидных клеток (например, hok/sok система плазмиды R1 (Gerdes et al., 1990), ccdA/B плазмиды F (Jaffe A et al., 1985), phd/doc фага P1 (Lehnherr et al., 1993), которые также не лишены недостатков, в частности их использование предполагает необходимость экспрессии дополнительных плазмидных генов, что снижает скорость роста культур.
Одним из наиболее перспективных направлений в плане возможности регулирования числа копий плазмидного вектора представляется использование системы «репрессор-антирепрессор». Она предполагает включение в состав векторной конструкции репликонов, содержащих ген репрессора, экспрессия которого определяет низкий уровень репликации плазмиды и соответственно число копий. Этот уровень может быть повышен путем введения в систему экспрессии белка-антирепрессора. Примером такой конструкции служит описанная ранее миниплазмида pNC10, включающая ген репликационного белка repA и ген репрессора сВ фага N15 (Ravin et al., JMB 2003). Данная плазмида является низкокопийной (3-5 копий на хромосому) за счет того, что СВ репрессор, ген которого включен в ее состав, связывается с операторными последовательностями, перекрывающими промотор repA. Геном фага N15 содержит ген антирепрессора antA (ген 31 в соответствии с нумерацией, приведенной в работе Ravin et al., 2000), продукт которого может инактивировать СВ репрессор (Ravin et al., 1999). Репрессия промотора repA в плазмиде pNC10 при желании может сниматься антирепрессором фага N15, что приводит к увеличению числа копий плазмиды в 50-100 раз (Mardanov et al., в печати). Однако возможности использования данной конструкции, а также и других плазмид, основанных на репликоне фага N15, в экспрессионных системах не исследованы.
Раскрытие изобретения
В настоящем изобретении ставилась задача создания систем для экспрессии гетерологичных белков в Е. coli, сочетающих высокий уровень экспрессии продукта с большим диапазоном регуляции (низким базовым уровнем).
В основу решения поставленной задачи было положено использование регулируемого промотора в сочетании с плазмидным репликоном, число копий которого может регулироваться в широком диапазоне с помощью системы «репрессор-антирепрессор». Такой подход позволяет существенно расширить диапазон регуляции синтеза продукта за счет перемножения эффектов активации промотора и эффекта дозы гена. Стабильное наследование вектора в неселективных условиях предполагалось обеспечить за счет использования механизма правильного распределения реплицированных плазмидных клеток между дочерними клетками перед делением.
Фактически задача была решена путем а) конструирования пригодного для включения целевого гена вектора, основанного на репликоне фага N15 и содержащего индуцируемый промотор; б) создания штамма Е. coli, клетки которого содержат в составе хромосомы ген антирепрессора фага N15 (antA) под контролем регулируемого (тем же, что и промотор вектора, или другим индуктором) промотора и sop оперона фага N15 и в) введения сконструированного вектора, в который предварительно встроен подлежащий экспрессии целевой ген, в штамм-реципиент с получением стабильно трансформированного рекомбинантного штамма E. coli.
Выбор в качестве основы для создания новой экспрессионной системы генетических элементов умеренного бактериофага N15 был обусловлен целом рядом его специфических особенностей. Во-первых, в лизогенном состоянии фаг не интегрируется в хромосому, а представляет собой линейную плазмиду, имеющую ковалентно замкнутые концы (теломеры). Во-вторых, N15 обладает набором собственных репликационных генов, в частности в результате анализа определенной полной нуклеотидной последовательности фага N15 был идентифицирован ответственный за процесс репликации фага ген repA. В-третьих, геном фага N15 содержит ген репрессора сВ, продукт которого связывается с промотором гена repA, контролирует уровень его экспрессии и, тем самым, обеспечивает низкое число копий профага, а также ген антирепрессора antA (ген 31 в соответствии с нумерацией, приведенной в работе Ravin et al., 2000), продукт которого может инактивировать СВ репрессор, в результате чего число копий плазмиды возрастает в 50-100 раз. В-четвертых, фаг N15 обладает собственной системой сегрегационной стабильности, обеспечивающей его стабильное наследование в неселективных условиях. Стабильное наследование плазмиды N15 обеспечивается за счет активного распределения реплицированных молекул между дочерними клетками при делении (partition), обусловленного генами sopA и sopB профага. Sop оперон N15 гомологичен соответствующим оперонам плазмид F и Р1, однако имеется ряд уникальных особенностей, связанных с линейностью плазмиды N15. Так N15 имеет не один, а четыре cis-действующих центромерных сайта, расположенных в различных районах генома, в частности, один из сайтов расположен в гене repA. Миниплазмида, включающая гены repA и сВ, стабильно наследуется в неселективных условиях (частота потери <0.01% на генерацию), если sop гены N15 экспрессируются in trans.
Таким образом, геном N15 содержит все необходимое для создания искусственной стабильной системы экспрессии с широкими возможностями регуляции процесса: участок размером около 6 т.п.н., способный обеспечивать репликацию низкокопийной миниплазмиды, ген антирепрессора (ant А), в результате экспрессии которого число копий плазмиды может быть увеличено в 50-100 раз, и оперон (sop), за счет экспрессии которого in trans достигается ее стабильное наследование.
Первый аспект настоящего изобретения связан с конструированием экспрессионного вектора, основанного на репликоне бактериофага N15 и содержащего оперативно связанные репликационный ген repA; ген, кодирующий белок-репрессор СВ, функционально активные в клетке Е. coli регулируемый промотор и терминатор транскрипции, в котором также предусмотрена возможность встраивания гетерологичного гена, предпочтительно путем включения в его состав полилинкера. В одном из конкретных воплощений (вектор pN15E4, фиг.1) в качестве индуцируемого промотора был использован промотор арабинозного оперона Е. coli araPBAD, а в качестве терминатора - терминатор ТО фага лямбда; в другом (вектор pN15E5t, фиг.6) - гибридный промотор РТ5-lac, индуцируемый IPTG, и терминатор ТО фага лямбда. Однако в состав предлагаемого вектора экспрессии могут быть включены и любые другие терминирующие области и регулируемые промоторы, и выбор среди известных последовательностей этого типа по существу ограничивается только их способностью проявлять функциональную активность в клетках Е. coli.
Вторым аспектом изобретения является получение рекомбинантного вектора экспрессии, предусматривающее встраивание в вектор-носитель, в частности в полилинкерную область плазмидных конструкций pN15E4 и pN15E5t, фрагмента ДНК, кодирующего целевой белок, так, чтобы указанный фрагмент был оперативно связан с содержащимся в векторе индуцируемым промотором. Конкретными примерами таких рекомбинантных конструкций являются сконструированные плазмиды pN15E4-dsRed, экспрессирующую красный флуоресцентный белок, и pN15E5t-lacZ, обеспечивающую синтез в E. coli бета-галактозидазы. Очевидно, что с помощью предлагаемого рекомбинантного вектора может быть осуществлен синтез любого белкового продукта, однако особое значение приобретает возможность продукции с его помощью соединений, токсичных для клетки. С целью подтверждения указанной возможности в рамках данного изобретения в векторе pN15E4 был клонирован ген эндонуклеазы рестрикции Ecl18Ki и получена рекомбинантная плазмида pN15E4-Ecl. В отличие стандартного вектора рQE60, в котором клонирование этого гена невозможно вследствие высокого базового уровня экспрессии, сконструированный рекомбинантный вектор pN15E4 позволил клонировать ген Ecl18Ki и получить в клетках E. coli функционально активную рестриктазу (см. пример 5).
При трансформации названными конструкциями традиционно используемых штаммов Е. coli, например штамма DH10B, уровень экспрессии продукта мог быть увеличен только за счет активации промотора; при полной индукции выход белка повышался примерно в 200 раз. С тем, чтобы при использовании сконструированных векторов могли быть реализованы оба возможные варианта регуляции синтеза продукта (как путем индукции промотора в составе вектора, так и за счет увеличения числа копий гена в результате снятия действия белка-репрессора), в хромосомы клеток-хозяев был введен ген антирепрессора (ant A).
Соответственно третий аспект изобретения относится к созданию штаммов Е. coli, в хромосомы клеток которых интегрирован ген антирепрессора фага N15 (antA) под контролем регулируемого промотора, функционально активного в бактериальной клетке. Примерами таких штаммов являются сконструированные при осуществлении настоящего изобретения штаммы Е. coli DH31sop и Е. coli DH31soplacI. Первый из них отличается от штамма DH10B, из которого он получен (см. пример 2), содержанием в хромосоме гена antA под контролем промотора araPBAD и sop оперона фага N15 (под контролем его собственного промотора), предназначенного для обеспечения стабильного наследования введенного в клетки штамма вектора в неселективных условиях. В хромосомы клеток второго штамма, кроме этого, дополнительно интегрирован ген репрессора лактозного оперона (lacI).
Четвертый аспект изобретения связан с получением систем экспрессии на основе предлагаемых векторных конструкций и штаммов.
В результате трансформации штамма Е. coli DH31sop рекомбинантными векторными конструкциями, производными от pN15E4, получены системы экспрессии гетерологичных белков, в которых в условиях индукции арабинозой реализуются оба возможных варианта повышения уровня синтеза продукта: за счет активации araPBAD в составе вектора, приводящей к повышению уровня экспрессии включенного в вектор целевого гена, и за счет активации araPBAD в составе хромосомы, имеющей следствием накопление в клетке белка-антирепрессора и увеличение числа копий вектора («дозы гена»). При этом было установлено (см. пример 4), что первый вариант регуляции процесса приводит к увеличению синтеза продукта примерно в 200 раз (это согласуется с ранее опубликованными данными о характеристиках araPBAD промотора [Guzman et al., 1995]), а активация репликации антирепрессором (второй вариант) позволяет увеличить число копий вектора в 10 раз. Соответственно перемножение этих двух эффектов должно было привести к возможности получения 2000-кратного диапазона регуляции, что и было подтверждено экспериментально.
Усовершенствованным вариантом описанной выше системы является система с разделением двух возможных видов регуляции (через активацию экспрессии гена и увеличение числа его копий). Для этого ген антирепрессора в хромосоме клетки - хозяина и целевой ген в экспрессионном векторе помещают под контроль разных промоторов. Так, в сконструированной экспрессионной системе, включающей штамм DH31soplacI и рекомбинантный вектор экспрессии на основе pN15E5t синтез антирепрессора регулируется araPBAD, который активируется арабинозой, а экспрессия целевого гена регулируется гибридным рТ5-lac промотором, репрессируемым Lad репрессором, ген которого интегрирован в хромосому штамма-хозяина, и индуцируемым IPTG (пример 8). Использование системы этого типа позволяет осуществлять плавную регуляцию синтеза продукта в зависимости от концентрации индуктора.
Краткое описание чертежей.
Фиг.1 - Структура вектора pN15E4.
repA - репликационный ген бактериофага N15
36, 35, 34 - гены бактериофага N15, нумерация соответствует приведенной в работе (Ravin et al., 2000)
сВ - ген репрессора бактериофага N15
araC - ген, кодирующий регуляторный белок арабинозного оперона Е. coli
Km - ген устойчивости к канамицину
araPBAD - промотор арабинозного оперона Е. coli, araPBAD
Т0 - терминатор Т0 фага лямбда.
На карте вектора отмечены уникальные сайты рестрикции.
Фиг.2 - Кассеты экспрессии, интегрированные в хромосому штамма Е. coli DH31sop
sopA, sopB - гены sop оперона бактериофага N15
antA - ген антирепрессора бактериофага N15
araC - ген, кодирующий регуляторный белок арабинозного оперона Е. coli
araPBAD - промотор арабинозного оперона Е. coli
Psop - промотор sop оперона бактериофага N15
Фиг.3 - Определение уровня стабильности наследования плазмиды pN15E4 в неселективных условиях.
Показана зависимость доли клеток, содержащих вектор pN15E4, от числа клеточных генераций, прошедших при выращивании штаммов DH10B/pN15E4 и DH31sop/pN15E4 в среде LB, не содержащей канамицина.
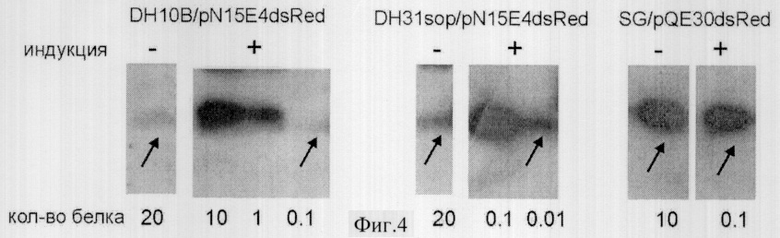
Фиг.4 - Вестерн блот анализ уровня продукции репортерного белка dsRed в экспрессионных системах DH10B/pN15E4-dsRed, DH31sop/pN15E4-dsRed и SG/pQE60-dsRed.
Белковые препараты были выделены из указанных штаммов до (-) и через 3 часа после (+) индукции промотора. Количества общего бактериального белка, нанесенного на дорожки, указаны под фотографиями (1 ед соответствует 3.2 мкг белка).
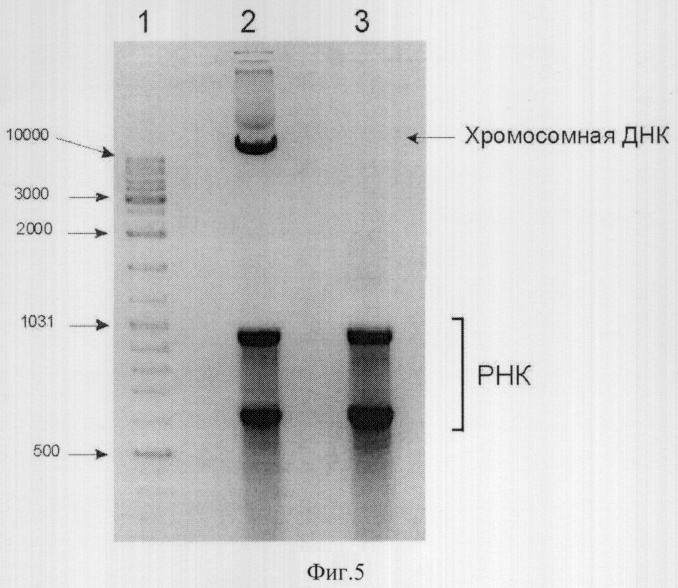
Фиг.5 - Электрофоретический анализ ДНК, выделенной из штамма DH31sop/pN15E4-Ecl до и после индукции синтеза рестриктазы Ecll8KiR.
1 - маркер молекулярного веса, длины фрагментов указаны в нуклеотидах.
2 - препарат ДНК выделенной из штамма DH31sop/pN15E4-Ecl до индукции синтеза рестриктазы.
3 - препарат ДНК выделенной из штамма DH31sop/pN15E4-Ecl через 2 часа после индукции синтеза рестриктазы внесением арабинозы в среду.
Отмечено положение геномной ДНК и одновременно выделяемой РНК.
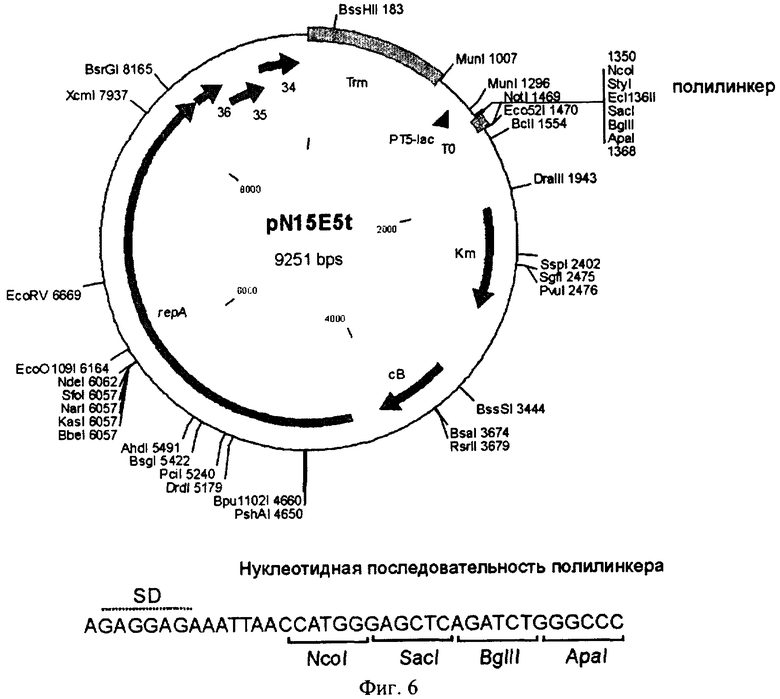
Фиг.6 - Структура вектора pN15E5t.
repA - репликационный ген бактериофага N15
36, 35, 34 - гены бактериофага N15, нумерация соответствует приведенной в работе (Ravin et al., 2000)
cB - ген репрессора бактериофага N15
Km - ген устойчивости к канамицину
РТ5-lac - промотор фага Т5, перекрывающийся lac-операторами
Trrn - терминатор рибосомального оперона E. coli
Т0 - терминатор Т0 фага лямбда.
На карте вектора отмечены уникальные сайты рестрикции.
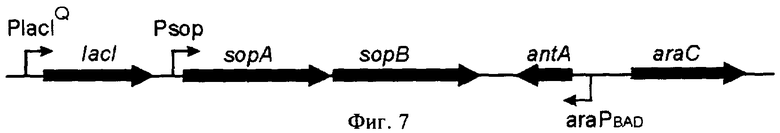
Фиг.7 - Кассеты экспрессии, интегрированные в хромосому штамма Е. coli DH31soplacI.
lacI - ген репрессора лактозного оперона Е. coli.
sopA, sopB - гены sop оперона бактериофага N15
antA - ген антирепрессора бактериофага N15
araC - ген, кодирующий регуляторный белок арабинозного оперона Е. coli.
araPBAD - промотор арабинозного оперона Е. coli
Psop - промотор sop оперона бактериофага N 15
PlacIQ - промотор lacI гена Е. coli (мутантный вариант, характеризующийся повышенной активностью).
Осуществление изобретения.
Пример 1. Конструирование экспрессионного вектора pN15E4.
В качестве основы для конструирования экспрессионного вектора была использована низкокопийная кольцевая миниплазмида pCN10 (Ravin et al., 2003).
В качестве регулируемого промотора при создании экспрессионного вектора использовался промотор арабинозного оперона Е. coli araPBAD, который имеет широкий диапазон регуляции (200 раз) и обеспечивает высокий уровень синтеза белка при индукции арабинозой [Guzman et al., 1995].
Фрагмент, содержащий область промотора, получали из плазмиды pBAD24 [Guzman, 1995]. Данную плазмиду предварительно модифицировали путем встраивания в нее по сайтам NheI и HindIII фрагмента ДНК, содержащего набор клонирующих сайтов и терминатор транскрипции ТО фага лямбда. Указанный фрагмент был получен в результате проведения ПЦР с использованием праймеров Т0-1 (5'-CGG СТА GCA GGA GGA ACT AGT CCA TGG GAG CTC AGA TCT GGG CCC GAC TCC TGT TGA TAG АТС CAG ТА) и Т0-2 (5' - CGA AGC CTG CGG CCG CAT TCT TAC CAA TAA AAA ACG CCC GG) и ДНК pQE60 (Qiagen) в качестве матрицы. ПЦР проводили при следующих условиях: (1) 94°С - 30 с, (2) 50°С - 60 с, (3) 70°С - 90 с, шаги 1-3 повторяли 40 раз.
После электрофоретического анализа выделяли ПЦР-фрагмент размером 158 п.н., который лигировали с pBAD24, предварительно обработанной рестриктазами NheI и HindIII. Полученную плазмиду, которая была отобрана по результатам рестрикционного анализа, обозначали pBAD24-T0 и использовали для получения фрагмента, содержащего araPBAD промотор. Для этого ее обрабатывали рестриктазами ClaI и NotI, выделяли фрагмент размером 1437 п.н., который содержал araC - araPBAD - полилинкер - Т0, и клонировали его в плазмиде pNC10 по сайтам Bsp119I и NotI (фиг.1). Полученный экспрессионный вектор был обозначен как pN15E4 и использован в дальнейшей работе.
Пример 2. Получение штамма Е. coli DH31sop
Конструирование штамма Е. coli, хромосомы клеток которого содержат ген антирепрессора N15 (antA) под контролем промотора araPBAD, а также sop оперон фага N15 (под контролем его собственного промотора), производили по схеме, предложенной в работе (Platt et al., 2000). На первом этапе в плазмиду pCD11PSK (Plan et al., 2000), при помощи которой впоследствии осуществлялось интегрирование чужеродного генетического материала в хромосому, клонировали два фрагмента ДНК: (1) PciI-HindIII фрагмент из плазмиды pBAD24-31 [Ravin et al., 1999], содержащий ген araC и ген antA под контролем промотора araPBAD и (2) HpaI - HindIII фрагмент из плазмиды pUC1-6 inv [Ravin and Lane, 1999], содержащий sop оперон фага N15. Для этого оба указанных фрагмента лигировали с препаратом ДНК pCD11PSK, обработанным рестриктазами PciI и HindIII, продуктами лигирования трансформировали штамм DH5αpir116 и отбирали нужные рекомбинантные клоны с помощью рестрикционного анализа. В результате была получена плазмида pCD31sop. Эта плазмида, как и исходный вектор pCD11PSK, содержит сайт интеграции в хромосому фага λ (attP сайт), ген устойчивости к хлорамфениколу, а также сайт инициации репликации плазмиды R6K. Для интеграции этой плазмиды в хромосому использовалась вспомогательная плазмида pPICK (Platt et al., 2000), содержащая ген устойчивости к канамицину, ген int интегразы фага лямбда, ген pir, кодирующий необходимый для репликации pCD11PSK белок π (Platt et al., 2000), а также репликационный ген, кодирующий белок с температурно-чувствительной мутацией. Этой плазмидой трансформировали штамм Е. coli DH10В (Grant et al., 1990). Плазмиду pCD31sop вводили в полученный штамм DH10B/pPICK, трансформанты отбирали при 30°С на чашках с канамицином и хлормфениколом. При этой температуре обе плазмиды реплицируются, экспрессируется интеграза фага лямбда и происходит интеграция плазмиды pCD31 sop в хромосому E. coli.
Отбор клеток, в которых произошла интеграция, осуществляли при 42°С на селективной среде с хлорамфениколом. При этой температуре плазмида pPICK не реплицируется и теряется, следовательно, отсутствует белок π, вследствие чего плазмида pCD31 sop также не реплицируется. Соответственно, на среде с хлорамфениколом при 42°С растут только клетки, в которых произошла интеграция плазмиды pCD31sop в хромосому. Из отобранных таким образом клеток был получен штамм Е. coli DH31sop, в хромосоме которого интегрирована кассета экспрессии araPBAD - antA, и sop оперон фага N15 (фиг.2). Отличительным признаком этого штамма является присутствие в его хромосоме описанных выше элементов, по остальным характеристикам (морфологическим, культуральным и физиолого-биохимическим) он соответствует исходному штамму DH10B (Grant et al., 1990).
Пример 3. Определение стабильности наследование вектора pN15E4 в штамме Е. coli DH31sop при выращивании в среде без антибиотика.
Для определения уровня стабильности наследования плазмиды pN15E4 в неселективных условиях ее вводили в два штамма: Е. coli DH10B (без sop оперона) и Е. coli DH31sop, содержащий sop оперон. Оба штамма, трансформированные pN14E4, выращивали до стационарного состояния в среде LB, содержащей 50 мг/мл канамицина. Затем культуру разводили в 106 раз и выращивали в бульоне-LB без канамицина до стационарного состояния, что соответствовало 20 генерациям. Затем клетки высевали на чашки без канамицина и делали реплики на агар с канамицином, где вырастали только колонии, содержащие pN15E4. В результате определяли долю клеток, содержащих pN15E4. Эксперимент повторяли на протяжении 40 генераций. Из результатов, представленных на фиг.3, следует, что уже через сорок генераций в штамме без sop оперона происходила потеря pN15E4 в 40% клеток. В штамме Е. coli DH31sop, содержащем sop оперон N15, потери вектора в те же сроки не наблюдалось.
Пример 4. Определение диапазона регуляции синтеза целевого белка для векторов pN15E4 и pQE60.
Для определения диапазона регуляции синтеза целевого белка в область полилинкера (по сайтам NcoI и BglII)_вектора pN15E4 встраивали ген красного флуоресцентного белка dsRed (Clontech). Полученный рекомбинантный вектор экспрессии (pN15E4-dsRed) вводили в два штамма: Е. coli DH10B, в котором возможна только индукция промотора, и в Е. coli DH31sop, в котором внесение арабинозы приводит к одновременной активации промотора и увеличению числа копий.
Культуры трансформированных клеток DH10B/pN15E4-dsRed и DH31sop/pN15E4-dsRed выращивали в LB-бульоне до середины логарифмической фазы роста (OD600=0,5), добавляли арабинозу до концентрации 0.2% (достаточной для полной индукции промотора) и выращивали клетки еще в течение 6 часов. Пробы для определения белка отбирали в начальный момент времени, а также через 1 и 6 часов после индукции.
Для сравнения аналогичный эксперимент проводили с коммерческим экспрессионным вектором pQE60 фирмы QIAGEN, в котором также клонировали dsRed. В качестве штамма-хозяина использовали рекомендованный фирмой штамм Е. coli SG, индукцию экспрессии dsRed осуществляли внесением в среду 1 mM IPTG.
Определение уровня синтеза dsRed осуществляли с помощью Вестерн-блот анализа. Электрофорез белков проводили в полиакриламидном геле (10%) в присутствии SDS. Перенос белков из геля на мембрану Hybond-P (Amersham) осуществляли методом электроблоттинга, следуя рекомендациям производителя. Для того чтобы исключить неспецифическое связывание антител с мембраной, последнюю обрабатывали 5% раствором сухого молока в буфере TBS-T. После этого мембрану гибридизовали с мышиными антителами к белку dsRed (разведение 1:5000), затем проводили гибридизацию со вторичными антителами, конъюгированными с пероксидазой хрена (Amersham). Детекцию специфических белковых комплексов проводили с использованием набора Western Blott ECL Plus (Amersham).
Как следует из полученных результатов (фиг.4), в экспрессионной системе DH10B/pN15E4-dsRed синтез белка после индукции увеличивается в 200 раз, тогда как в системе DH31sop/pN15E4-desRed - в 2000 раз. В то же время в экспрессионной системе SG/pQE-dsRed синтез продукта увеличивался только в 100 раз.
Максимальный уровень продукции белка в DH31sop/pN15E4-dsRed и в SG/pQE-dsRed был практически одинаков и составлял около 20% общего белка. При этом базовый уровень экспрессии в DH31sop/pN15E4-dsRed оказался в 20 раз ниже, чем в SG/PQE-dsRed (фиг.4).
Пример 5. Использование pN15E4 для клонирования и экспрессии гена эндонуклеазы рестрикции Ecl18Ki.
Ген эндонуклеазы рестрикции Ecl18Ki был амплифицирован с помощью ПЦР с использованием праймеров Ecli-NcoI-F (5'-ATCCATGGAACAAAGATTAAGTCCTGG) и Ecl-BamHI-R (5'-CAGGATCCTTAGTCAAAAAGAGCACTAA) и ДНК плазмиды pBCNR в качестве матрицы (любезно предоставлена А.С.Солониным, ИБФМ РАН) [Перцев и др., 1997]. ПЦР проводили при следующих условиях: (1) 94°С - 30 с, (2) 55°С - 60 с, (3) 72°С - 90 с, шаги 1-3 повторяли 40 раз.
Полученный ПЦР-фрагмент был обработан ферментами рестрикции NcoI и BamHI и лигирован с препаратами ДНК векторов pN15E4 и pQE60, гидролизованными NcoI и BglII. Лигирующую смесь использовали для трансформации штамма Е. coli DH31 sop (в случае pN15E4) и штамма SG (в случае pQE60). Колонии трансформантов, содержащих рекомбинантные плазмиды с геном Ecl18Ki (pN15E4-Ecl), были получены только при использовании вектора pN15E4. В случае pQE60 рост колоний трансформантов не наблюдался. Для проверки корректности клонирования Ecl18Ki и отсутствия мутаций, которые могли бы нарушать активность фермента, были выделены препараты плазмидной ДНК pN15E4-Ecl и проведено определение нуклеотидной последовательности вставки и окружающих участков вектора. В результате было установлено, что последовательность клонированного гена идентична исходной. Таким образом, возможность продукции клетками токсичного продукта полностью определяется низким базовым уровнем экспрессии в системе DH31sop/pN15E4-Ecl.
Кроме этого, был проведен дополнительный контрольный эксперимент, целью которого было подтвердить функциональную активность экспрессированной рестриктазы. Для этого культуру Е. coli DH31sop/pN15E4-Ecl выращивали при 37°С в LB бульоне до середины логарифмической фазы роста, затем добавляли арабинозу до 0.2%, что приводило к активации синтеза рестриктазы. Препараты суммарной клеточной ДНК выделяли до индукции, а также через 3 часа после ее начала и анализировали с помощью электрофореза. Как следует из результатов, представленных на фиг.5, индукция синтеза рестриктазы приводит к деградации высокомолекулярной хромосомной ДНК и исчезновению соответствующей полосы. Таким образом, синтезируемый в системе DH31sop/pN15E4-Ecl фермент является функционально активным.
Пример 6. Конструирование экспрессионного вектора pN15E5t.
В качестве регулируемого промотора при создании pN15E5t был использован гибридный промотор РТ5-lac, состоящий из промотора фага Т5 и двух lac-операторов. Этот промотор, входящий в состав экспрессионного вектора pQE60 (Qiagen), индуцируется IPTG и обеспечивает высокий уровень синтеза белка при индукции.
Фрагмент, содержащий область промотора, получали из плазмиды pQE60 (Qiagen). Для этого ДНК pQE60 обрабатывали рестриктазой XhoI, образовавшиеся липкие концы застраивали с помощью фрагмента Кленова ДНК-полимеразы I, инактивировали ферменты нагреванием препарата до 70°С в течение 15 мин, а затем проводили обработку рестриктазой NcoI. Фрагмент длиной 112 п.н., содержащий промотор, выделяли из агарозного геля после электрофореза продуктов рестрикции.
ДНК вектора pN15E4 обрабатывали рестриктазой AflII, образовавшиеся липкие концы застраивали с помощью фрагмента Кленова ДНК-полимеразы I, инактивировали ферменты нагреванием препарата до 70°С в течение 15 мин, а затем проводили обработку рестриктазой NcoI. Фрагмент длиной 8200 п.н., содержащий репликон N15, выделяли из агарозного геля после электрофореза продуктов рестрикции. Фрагмент ДНК из pQE60, содержащий РТ5 промотор, лигировали с фрагментом вектора pN15E4, содержащим репликон N15, в результате чего была получена плазмида pN15E5.
Затем в полученной плазмиде pN15E5 клонировали фрагмент ДНК, содержащий терминатор транскрипции Trrn оперона рибосомных генов E. coli. Для этого ДНК плазмиды pRS551 (Simons et al., 1987) обрабатывали рестриктазами SmaI и StuI, фрагмент длиной 931 п.н., содержащий указанный терминатор транскрипции, выделяли из агарозного геля после электрофореза продуктов рестрикции. ДНК вектора pN15E5 подвергали неполному гидролизу рестриктазой MunI (в плазмиде имеется два сайта MunI), образовавшиеся липкие концы застраивали с помощью фрагмента Кленова ДНК-полимеразы I, а затем проводили обработку бактериальной щелочной фосфатазой. Линеаризованный вектор (фрагмент длиной 8316 п.н.) выделяли из агарозного геля после электрофореза продуктов рестрикции и лигировали с фрагментом ДНК, содержащим терминатор Trrn. Отбор рекомбинантов, содержащих вставку в прямой ориентации, проводили с помощью рестрикционного анализа и секвенирования вставки и прилегающих участков плазмиды. Полученный экспрессионный вектор был обозначен как pN15E5t (фиг.6) и использован в дальнейшей работе.
Пример 7. Получение штамма Е. coli DH31soplacI.
На первом этапе был получен фрагмент ДНК, содержащий ген lacIQ репрессора лактозного оперона E. coli. Для этого проводили ПЦР с использованием праймеров lac-F (5'-CTCGAGCCTTTCGTCTTCACCTCGAC) и lacI-R (5'-TGGTACCAATTGCGTTGCGCTCACTGC), и ДНК pZS4int-1 (Lutz and Bujard, 1997) качестве матрицы. ПЦР осуществляли при следующих условиях: (1) 94°С - 30 с, (2) 55°С - 60 с, (3) 72°С - 90 с, шаги 1-3 повторяли 40 раз.
После электрофоретического анализа выделяли ПЦР-фрагмент размером 1277 п.н, который обрабатывали рестриктазами KpnI и XhoI и лигировали с pCD31sop, предварительно обработанной теми же рестриктазами. Полученную плазмиду, которая была отобрана по результатам рестрикционного анализа, обозначали pCD31soplacI. Интеграцию этой плазмиды в хромосому штамма Е. coli DH10B проводили как описано в Примере 2. Таким образом, был получен штамм Е. coli DH31soplacI, в хромосому клеток которого интегрирована кассета экспрессии araPBAD - antA, sop оперон фага N15 и ген lacIQ репрессора лактозного оперона E. coli (фиг.7).
Пример 8. Определение диапазона регуляции синтеза целевого белка для вектора pN15E5t
Для определения диапазона регуляции синтеза целевого белка в область полилинкера (по сайтам NcoI и BglII) вектора pN15E5t встраивали ген lacZ из Е. coli, кодирующий β-галактозидазу. Полученный рекомбинантный вектор экспрессии (pN15E5t-lacZ) вводили в штамм DH31soplacI, который позволяет индуцировать промотор внесением в среду IPTG и увеличивать число копий вектора pN15E5t внесением в среду арабинозы.
Культуру трансформированных клеток DH31soplacI/pN15E5t-lacZ выращивали в LB-бульоне до середины логарифмической фазы роста (OD600=0,5), добавляли только IPTG до концентрации 0.1 мМ (достаточной для полной индукции РТ5-lac промотора) или одновременно IPTG до концентрации 0.1 мМ и арабинозу до концентрации 0.2% (достаточной для полной индукции araPBAD промотора) и выращивали клетки еще в течение 3 часов. Пробы для определения уровня активности β-галактозодазы отбирали в начальный момент времени, а также через 3 часа после начала индукции. Количественное определение уровня активности β-галактозодазы проводили по методу Миллера (Miller, 1972) и выражали в единицах Миллера (ME).
Для сравнения аналогичный эксперимент проводили с коммерческим экспрессионным вектором pQE60 фирмы QIAGEN, в котором также клонировали ген lacZ (pQE-lacZ). В качестве штамма-хозяина использовали рекомендованный фирмой штамм Е. coli SG, а также сконструированный штамм DH31soplacI.
Как следует из полученных результатов (таблица 2), в экспрессионной системе DH31soplacI/pN15E5t-lacZ уровень активности β-галактозодазы после индукции РТ5-lac промотора увеличивался в 3360 раз (от 3 М.Е. до 10090 М.Е.), а в случае одновременной индукции промотора и активации репликации он увеличивается в 15800 раз (с 3 М.Е. до 47430 М.Е.). В то же время в экспрессионной системе SG/pQE60-lacZ уровень активности β-галактозодазы увеличивался только в 30 раз (с 2625 до 71660 М.Е.), что было обусловлено значительно более высоким базовым уровнем экспрессии (2625 М.Е. против 3 М.Е. в случае DH31soplacI/pN15E5t). При использовании штамма DH31soplacI базовый уровень синтеза продукта, обеспечиваемый вектором pQE60 оказался ниже (70 ME), чем в SG (2625 М.Е.), но все равно существенно выше, чем в случае вектора pN15E5t (3 М.Е.). Максимальный уровень β-галактозодазы в экспрессионных системах DH31soplacI/pN15E5t-lacZ, DH31soplacI/pQE60-lacZ и SG/pQE60-lacZ был сопоставим (47430-71660 М.Е.).
ЛИТЕРАТУРА
1. Brosius J., M.Erfle and J.Storella. 1985. Spacing of the 210 and 235 regions in the tac promoter. Effect on its in vivo activity. J. Biol. Chem. 260: 3539-3541.
2. De Boer H.A., L.J.Comstock and M.Vasser. 1983. The tac promoter: a functional hybrid derived from the trp and lac promoters. Proc. Natl. Acad. Sci. USA 80: 21-25.
3. Elvin C.M., P.R.Thompson, M.E.Argall, P.Hendry, N.P.Stamford, P.E.Lilley and N.E.Dixon. 1990. Modified bacteriophage lambda promoter vectors for overproduction of proteins in Escherichia coli. Gene 87: 123-126.
4. Gerdes K., Thisted T., Martinussen J. 1990 Mechanism of post-segregational killing by the hok/sok system of plasmid R1: sok antisense RNA regulates formation of a hok mRNA species correlated with killing of plasmid-free cells. Mol Microbiol. 4(11): 1807-18.
5. Giladi H., Goldenberg G. and Oppenheim A.B. (1995) Enhanced activity of the bacteriophage lambda PL promoter at low temperatures. Proc. Natl. Acad. Sci. USA, 92: 2184-2188.
6. Grant S.G., Jessee J., Bloom F.R. and Hanahan, D. (1990) Differential plasmid rescue from transgenic mouse DNAs into Escherichia coli methylation-restriction mutants. Proc Natl Acad Sci USA 87: 4645-4649.
7. Guzman L.M., Belin D., Carson M.J., Beckwith J. Tight regulation, modulation and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995 Jul; 177 (14): 4121-30.
8. Jaffe A., T. Ogure, S.Hiraga. 1985. Effects of the ccd function of the F plasmid on bacterial growth. J. Bacteriol. 163: 841-849.
9. Lehnherr H., Maguin E., Jafri S., Yarmolinsky M.B. 1993. Plasmid addiction genes of bacteriophage P1: doc, which causes cell death on curing of prophage and phd, which prevents host death when prophage is retained. J Mol Biol. 233(3): 414-28.
10. Lutz R. and Bujard H. (1997) Independent and tight regulation of transcriptional units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2 regulatory elements. Nucleic Acids Res, 25, 1203-1210.
11. Mardanov A.V., Ravin N.V. (2006) Functional characterization of the repA replication gene of linear plasmid prophage N15. Res Microbiol. 157(2): 176-183.
12. Miller J.H. (1972). Experiments in molecular genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor N.Y.
13. Miroux B. and Walker J.E. (1996) Over-production of protein in Escherichia coli: mutant hosts that allow synthesis of some membrane proteins and globular proteins at high levels. J. Mol. Biol., 260: 289-298.
14. Pertzev A.V., Kravetz A.N., Mayorov S.G., Zakharova M.V., Solonin A.S. Isolation of a strain overproducing endonuclease Eco29kI: purification and characterization of the homogeneous enzyme. Biochemistry (Mosc). 1997 Jul; 62(7): 732-41.
15. Platt R., Drescher C., Park S.K., Phillips G.J. Genetic system for reversible integration of DNA constructs and lacZ gene fusions into the Escherichia coli chromosome. Plasmid. 2000 Jan; 43(1): 12-23.
16. Ravin N., Lane D. (1999) Partition of the linear plasmid, N15: functional interactions with the sop locus of the F plasmid. J. Bacteriol 181: 6898-6906.
17. Ravin N.V., Rech J., & Lane D. (2003). Mapping of functional domains in F plasmid partition proteins reveals a bipartite SopB-recognition domain in SopA. J. Mol. Biol., 329, 875-889.
18. Ravin N., Svarchevsky A., Deho G. (1999). The antiimunity system of phage-plasmid N15: identification of the antirepressor gene and its control by a small processed RNA. Mol. Microbiol 34: 980-994.
19. Ravin V., Ravin N., Casjens S., Ford M., Hatfull G., & Hendrix R. (2000). Genomic sequence and analysis of the atypical bacteriophage N15. J. Mol. Biol., 299, 53-73.
20. Siegele D.A., Hu J.C. Gene expression from plasmids containing the araBAD promoter at subsaturating inducer concentrations represents mixed populations. Proc Natl Acad Sci USA. 1997 Jul 22; 94(15): 8168-72.
21. Simons R.W., Houman F. and Kleckner N. (1987) Improved single and multicopy lac-based cloning vectors for protein and operon fusions. Gene. 53: 85-96.
22. Summers F. (1998) Timing, self-control and sense of direction are the secrets of multicopy plasmid stability. Mol. Microbiol. 29: 1137-1145.


Настоящее изобретение относится к области биотехнологии и решает задачу создания системы экспрессии целевых белков в клетках Escherichia coli, сочетающей высокий уровень биосинтеза продукта с большим диапазоном его регуляции. Сконструированы экспрессионные системы, представляющие собой штамм Е. coli с интегрированным в хромосому клеток геном антирепрессора фага N15, который находится под контролем функционально активного в Е. coli индуцируемого промотора, и sop-опероном фага N15, трансформированный рекомбинантным вектором, основанным на репликоне фага N15 и содержащим фрагмент ДНК, кодирующий целевой белок, под контролем того же, что и в составе хромосомы, или иного индуцируемого промотора. Предложенные векторы стабильно наследуются при выращивании бактерий в среде без антибиотика за счет механизма правильного распределения реплицированных плазмидных копий между дочерними клетками. Применение изобретения обеспечивает широкий диапазон регулирования выхода целевого продукта и соответственно возможность использования одной и той же системы экспрессии для получения широкого круга белковых продуктов вне зависимости от их функциональной активности. 3 н. и 9 з.п. ф-лы, 7 ил., 2 табл.
| RAVIN V | |||
| ет al., J | |||
| Mol | |||
| Biol., 299, 53-73, 2000 | |||
| RAVIN N.V | |||
| ет al., Nucleic Acids Res., 31, 6552-6560, 2003 | |||
| МАРДАНОВ А.В | |||
| и др., Доклады Академии наук, т.397, с.1-4, 2004. |

