Область изобретения
Данное изобретение относится к составам, содержащим ингибиторы матриксных металлопротеиназ, которые пригодны для заживления ран и устранения результатов воздействия старения на кожу. Эти ингибиторы представляют собой пептиды, имеющие последовательности, соответствующие вырезаемому участку проферментных форм матриксных металлопротеиназ.
Предшествующий уровень техники
На состояние кожи всегда воздействуют такие факторы, как влажность, ультрафиолетовое излучение, косметические средства, старение, заболевания, стресс и привычки в еде. В результате, могут возникнуть различные кожные проблемы. С возрастом кожа также становится менее упругой, что проявляется в образовании морщин. Старение обычно сопровождается утончением и общей деградацией кожи. По мере естественного старения кожи происходит уменьшение количества клеток и кровеносных сосудов, питающих кожу. Также имеет место уплощение кожно-эпидермального соединения, что приводит к ослаблению его механических свойств. В результате, пожилые люди становятся более восприимчивыми к образованию волдырей при механических травмах или болезненных процессах (см. Oikarinen (1990) "The Aging of Skin: Chronoaging Versus Photoaging", Photodermatal. Photoimmunol. Photomed., Vol.7, pp 3-4).
Кожа содержит сложную сеть волокон эластина, которые ответственны за поддержание ее эластичных свойств. При избыточном воздействии солнечного излучения система эластичных волокон становится гиперпластической, дезорганизованной и, в конечном счете, разрушается. Этот процесс известен как актинический эластоз, который является главной причиной образования морщин, обесцвечивания и рыхлости кожи на открытых участках тела. При формировании новых фибробластов, эндотелиальных клеток и кератиноцитов кожа может восстанавливаться сама. Однако по мере старения кожа постепенно утрачивает эту способность. Поэтому для преждевременно состарившейся кожи требуются агенты, способные ускорять процессы роста и восстановления.
Заживление ран также ускоряется при повышении клеточной пролиферации и миграции некоторых типов клеток. Механизмы, задействованные в процессах заживления ран, часто подразделяются на четыре фазы: гемостаз, воспаление, пролиферация и созревание. Во время воспаления происходит скопление лейкоцитов для борьбы с бактериями, и повышается проницаемость кровеносных сосудов, что приводит к опуханию. Если инфекция не развивается, количество лейкоцитов уменьшается. Лейкоциты заменяются моноцитами. Макрофаги и лимфоциты высвобождают факторы роста (цитокины), а также ряд химических веществ, таких как гистамин, серотонин и простагландины. Эти вещества помогают регулировать процесс заживления ран. В фазе пролиферации возникают новые фибробласты, эндотелиальные клетки и кератиноциты, образуется соединительная ткань, появляются новые кровеносные сосуды и восстанавливается поврежденная ткань. Фибробласты начинают доминировать примерно через неделю, при этом воспаление спадает и прочность ткани вокруг поврежденного участка быстро возрастает. Во время фазы созревания откладывается коллаген, и образуется рубцовая ткань. Эта фаза созревания может продолжаться длительное время, в течение которого восстанавливаются ткани различных типов. Для оптимального восстановления кожи и ассоциированных тканей необходимо достаточное снабжение их различными витаминами и микроэлементами, а также питательными веществами и кислородом.
Хронические раны или безболезненные, незаживающие раны могут возникать по различным причинам, включая инфекции, наличие инородных тел или токсичных раздражающих веществ, ожоги, продолжительное механическое воздействие на кожу и недостаточное кровоснабжение в связи с нарушенной циркуляцией крови. В хронических ранах окружение раны влияет на тканевой гомеостаз таким образом, что либо заживления не происходит, либо оно начинается, но в дальнейшем приостанавливается. Факторами, препятствующими процессам заживления в хронических ранах, являются некроз ткани, дегидратация, отек хронической раны, фиброзное уплотнение и болезни мелких кровеносных сосудов.
Одной из главных причин незаживления хронических ран является действие особого класса протеиназ, называемых матриксными металлопротеиназами (ММП), которые разрушают новое сформированное ложе раны (Vaalamo et al., 1997; Weckroth et al., 1996; DiColandrea et al., 1998; Moses et al., 1996). Обычно защиту ложа раны от разрушения этими матриксными металлопротеиназами выполняют четыре тканевых ингибитора металлопротеиназ (ТИМП 1-4), которые образуют очень специфические ингибиторные комплексы с матриксными металлопротеиназами (Olson et al., 1997; Taylor et al., 1996; Howard et al., 1991). Это значит, что каждый ТИМП ингибирует только определенный подкласс матриксных металлопротеиназ. В хронических ранах наблюдается высокое соотношение матриксной металлопротеиназы к ТИМП, в результате чего большинство матриксных металлопротеиназ не ингибируются (Vaalamo et al., 1996; Saarialho-Kere, 1998). Фактически, при повышенном уровне протеиназ молекулы ТИМП сами могут подвергаться гидролизу. Среди природных тканевых ингибиторов металлопротеиназ нет таких, которые в одиночку были бы способны ингибировать все типы матриксных металлопротеиназ.
Для контроля активности матриксных металлопротеиназ было предложено несколько подходов, включая использование небольших молекул (Levy et al., 1998; Wojtowicz-Praga et al., 1997; Duivenvoorden, et al., 1997), пептидных ингибиторов (Odake et al., 1994) и антител к матриксным металлопротеиназам (Su et al., 1995). Однако идеальное средство для заживления ран и замедления процессов старения должно не только обеспечивать оптимальное ингибирование металлопротеиназ, но и способствовать росту и восстановлению поврежденных тканей.
Краткое изложение изобретения
Настоящее изобретение предоставляет композиции, содержащие пептиды, которые могут использоваться как противовозрастные и ранозаживляющие агенты. Пептиды согласно настоящему изобретению могут не только ингибировать металлопротеиназы, но также стимулировать клеточную пролиферацию и миграцию в некоторых типах клеток, включая фибробласты, эндотелиальные клетки и кератиноциты. В рамках данного изобретения рассматриваются также различные лосьоны топического действия, повязки и композиции, а также способы использования пептидов для замедления процессов старения и для заживления ран.
Представленное изобретение относится, таким образом, к пептидным ингибиторам матриксных металлопротеиназ. Эти пептидные ингибиторы имеют аминокислотные последовательности, идентичные или родственные участку связывания, соединяющему два глобулярных домена матриксных металлопротеиназ. Известно несколько типов матриксных металлопротеиназ и их последовательностей, в частности матриксная металлопротеиназа-1, матриксная металлопротеиназа-2, матриксная металлопротеиназа-3, матриксная металлопротеиназа-4, матриксная металлопротеиназа-5, матриксная металлопротеиназа-6, матриксная металлопротеиназа-7, матриксная металлопротеиназа-8, матриксная металлопротеиназа-9, матриксная металлопротеиназа-10, матриксная металлопротеиназа-11, матриксная металлопротеиназа-12 и матриксная металлопротеиназа-13. Данное изобретение включает в себя ингибиторы, обладающие аминокислотной последовательностью участка связывания какой-либо из матриксных металлопротеиназ. Например, пептидные ингибиторы, заявленные в данном изобретении, могут иметь аминокислотные последовательности, соответствующие любой области примерно с 70 по 120 аминокислоту последовательности матриксной металлопротеиназы-2 (SEQ ID NO: 14) и аналогичным областям всех других матриксных металлопротеиназ.
Настоящее изобретение представляет пептиды, характеризующиеся одной из следующих формул (I), (II), (III):

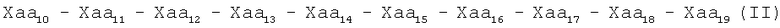
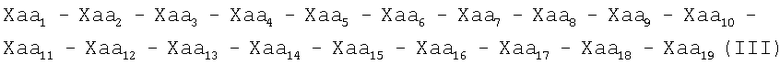
где
Хаа1, Хаа4 и Хаа6 независимо друг от друга обозначают неполярные аминокислоты;
Хаа2 является основной аминокислотой;
Хаа3 является цистеиноподобной аминокислотой;
Хаа5 является полярной или алифатической аминокислотой;
Хаа7 является кислой аминокислотой,
Хаа8 является алифатической или полярной аминокислотой;
Хаа9 является алифатической, неполярной или основной аминокислотой; и
Хаа10 является полярной, кислой, основной или неполярной аминокислотой;
Хаа11 является полярной или ароматической аминокислотой;
Хаа12 является полярной, основной, алифатической или неполярной аминокислотой;
Хаа13 является ароматической, алифатической, полярной или кислой аминокислотой;
Xaa14 является ароматической, неполярной или полярной аминокислотой;
Хаа15 является неполярной или кислой аминокислотой;
Хаа16 является основной, полярной или неполярной аминокислотой;
Xaa17 является основной, полярной, алифатической, неполярной или кислой аминокислотой;
Хаа18 является неполярной или алифатической аминокислотой;
Хаа19 является основной или алифатической аминокислотой; и
указанный пептид способен ингибировать активность матриксной металлопротеиназы-1, матриксной металлопротеиназы-2, матриксной металлопротеиназы-3, матриксной металлопротеиназы-4, матриксной металлопротеиназы-5, матриксной металлопротеиназы-6, матриксной металлопротеиназы-7, матриксной металлопротеиназы-8 или матриксной металлопротеиназы-9, матриксной металлопротеиназы-10, матриксной металлопротеиназы-11, матриксной металлопротеиназы-12 и матриксной металлопротеиназы-13. В некоторых вариантах осуществления данного изобретения этот пептид может ингибировать активность матриксной металлопротеиназы-2, матриксной металлопротеиназы-3, матриксной металлопротеиназы-7, матриксной металлопротеиназы-8 или матриксной металлопротеиназы-9.
В качестве неполярной аминокислоты может выступать, например, метионин, глицин или пролин. В качестве основной аминокислоты может выступать, в частности, гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин или 2,4-диаминомасляная кислота. Цистеиноподобные аминокислоты включают, в частности, цистеин, гомоцистеин, пеницилламин или β-метилцистеин.
К алифатическим аминокислотам относятся, в частности, аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота. К кислым аминокислотам относятся, в частности, аспарагиновая кислота или глутаминовая кислота. К полярным аминокислотам относятся, в частности, аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин, а к неполярным аминокислотам, такие как метионин, глицин или пролин. В рамках данного изобретения к ароматическим аминокислотам относятся, в частности, фенилаланин, тирозин, триптофан, фенилглицин, нафтилаланин, β-2-тиенилаланин, 1,2,3,4-тетрагидроизохинолин-3-карбоновая кислота, 4-хлорофенилаланин, 2-фторфенилаланин, 3-фторфенилаланин, 4-фторфенилаланин, пиридилаланин или 3-бензотиенилаланин.
Настоящее изобретение относится также к пептидам с формулами (IV) (SEQ ID NO: 18):
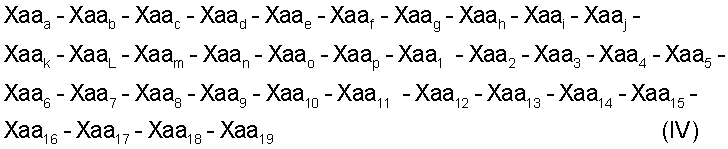
где:
Xaaa обозначает пролин;
Хааb обозначает глутамин или глутаминовую кислоту;
Хаас обозначает треонин;
Xaad обозначает глицин;
Хааe обозначает аспарагиновую кислоту или глутаминовую кислоту;
Хааf обозначает лейцин;
Xaag обозначает аспарагиновую кислоту;
Хааh обозначает глутамин или серин;
Хааi обозначает аспарагин или аланин;
Xaaj обозначает треонин;
Хааk обозначает изолейцин или лейцин;
ХааL обозначает глутаминовую кислоту или лизин;
Хааm обозначает треонин или аланин;
Хааn обозначает метионин;
Хаао обозначает аргинин;
Хаар обозначает лизин или треонин;
Xaa1 обозначает пролин;
Хаа2 обозначает аргинин;
Хаа3 обозначает цистеин;
Хаа4 обозначает глицин;
Хаа5 обозначает валин или аспарагин
Хаа6 обозначает пролин;
Хаа7 обозначает аспарагиновую кислоту;
Xaa8 обозначает валин или лейцин;
Хаа9 обозначает аланин или глицин;
Хаа10 обозначает аспарагин или аргинин;
Xaa11 обозначает тирозин или фенилаланин;
Xaa12 обозначает аспарагин или глутамин;
Xaa13 обозначает фенилаланин или треонин;
Xaa14 обозначает фенилаланин;
Xaa15 обозначает пролин или глутаминовую кислоту;
Xaa16 обозначает аргинин или глицин;
Xaa17 обозначает лизин или аспарагиновую кислоту;
Xaa18 обозначает пролин или лейцин;
Xaa19 обозначает лизин; и
указанный пептид способен ингибировать активность металлопротеиназы. Примером матриксных металлопротеиназ могут служить матриксная металлопротеиназа-1, матриксная металлопротеиназа-2, матриксная металлопротеиназа-3, матриксная металлопротеиназа-4, матриксная металлопротеиназа-5, матриксная металлопротеиназа-6, матриксная металлопротеиназа-7, матриксная металлопротеиназа-8, матриксная металлопротеиназа-9, матриксная металлопротеиназа-10, матриксная металлопротеиназа-11, матриксная металлопротеиназа-12 и матриксная металлопротеиназа-13. Предпочтительные пептиды ингибируют матриксную металлопротеиназу-2 или матриксную металлопротеиназу-9.
Участки связывания, из которых пептидные ингибиторы согласно настоящему изобретению могут быть получены, имеют, например, аминокислотные последовательности в пределах от около 70 до около 120 положения SEQ ID NO: 14 и аналогичные области других матриксных металлопротеиназ. В некоторых вариантах осуществления данного изобретения пептидные ингибиторы имеют аминокислотные последовательности в пределах от около 77 до около 110 положения SEQ ID NO: 14, и аналогичных областей других матриксных металлопротеиназ. Примеры некоторых пептидных ингибиторов включают ингибиторы, содержащие аминокислотные последовательности SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12 или SEQ ID NO: 13.
Пептиды настоящего изобретения могут обладать различным сродством к различным матриксным металлопротеиназам. Например, в одном из вариантов осуществления данного изобретения эти пептидные ингибиторы могут ингибировать матриксную металлопротеиназу-2 со значением ki в пределах примерно от 1,0 мкМ до 500,0 мкМ. В другом варианте осуществления данного изобретения эти пептидные ингибиторы могут ингибировать матриксную металлопротеиназу-2 со значением ki в пределах примерно от 1,0 мкМ до 400,0 мкМ. В еще одном варианте осуществления данного изобретения эти пептидные ингибиторы могут ингибировать матриксную металлопротеиназу-2 со значением ki в пределах примерно от 1,0 мкМ до 50,0 мкМ.
Настоящее изобретение также предоставляет композиции, содержащие терапевтически эффективное количество пептида согласно настоящему изобретению и фармацевтически приемлемый носитель. Составы для обработки ран и кожные лосьоны также рассматриваются в настоящем изобретении.
Объектом данного изобретения является также способ лечения ран или устранения результатов воздействия старения, предусматривающий применение терапевтически эффективного количества пептида, соответствующего одной из следующих формул I, II, III или IV:
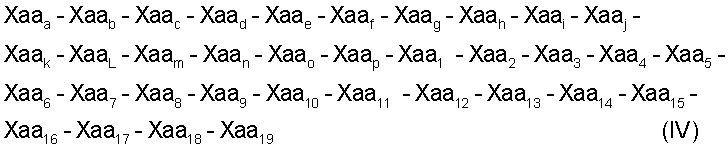
(SEQ ID NO: 21)
где:
Xaa1, Xaa4 и Хаа6 независимо друг от друга обозначают неполярные аминокислоты;
Хаа2 является основной аминокислотой;
Хаа3 является цистеиноподобной аминокислотой;
Хаа5 является полярной или алифатической аминокислотой;
Хаа7 является кислой аминокислотой,
Xaa8 является алифатической или полярной аминокислотой;
Хаа9 является алифатической, неполярной или основной аминокислотой; и
Хаа10 является полярной, кислой, основной или неполярной аминокислотой;
Xaa11 является полярной или ароматической аминокислотой;
Xaa12 является полярной, основной, алифатической или неполярной аминокислотой;
Хаа13 является ароматической, алифатической, полярной или кислой аминокислотой;
Xaa14 является ароматической, неполярной или полярной аминокислотой;
Xaa15 является неполярной или кислой аминокислотой;
Xaa16 является основной, полярной или неполярной аминокислотой;
Xaa17 является основной, полярной, алифатической, неполярной или кислой аминокислотой;
Xaa18 является неполярной или алифатической аминокислотой;
Xaa19 является основной или алифатической аминокислотой;
Хааa обозначает пролин;
Хааb обозначает глутамин или глутаминовую кислоту;
Хаас обозначает треонин;
Хааd обозначает глицин;
Хаае обозначает аспарагиновую кислоту или глутаминовую кислоту;
Хааf обозначает лейцин;
Xaag обозначает аспарагиновую кислоту;
Хааh обозначает глутамин или серин;
Хааi обозначает аспарагин или аланин;
Xaaj обозначает треонин;
Xaak обозначает изолейцин или лейцин;
XaaL обозначает глутаминовую кислоту или лизин;
Xaam обозначает треонин или аланин;
Xaan обозначает метионин;
Xaaо обозначает аргинин; и
Хааp обозначает лизин или треонин;
причем указанный пептид способен ингибировать активность матриксной металлопротеиназы.
В качестве неполярной аминокислоты в заявленных в данном изобретении пептидах может выступать, например, метионин, глицин или пролин. В качестве основной аминокислоты может выступать, например, гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин и 2,4-диаминомасляная кислота. В качестве цистеиноподобной аминокислоты может выступать, например, цистеин, гомоцистеин, пеницилламин или β-метил цистеин. В качестве алифатической аминокислоты может выступать, например, аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота. В качестве кислой аминокислоты может выступать, например, аспарагиновая кислота или глутаминовая кислота. В качестве полярной аминокислоты может выступать аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетиллизин, метионин сульфоксид или гомосерин, а в качестве неполярной аминокислоты, такие как метионин, глицин или пролин. В качестве ароматической аминокислоты может выступать фенилаланин, тирозин, триптофан, фенилглицин, нафтилаланин, β-2-тиенилаланин, 1,2,3,4-тетрагидроизохинолин-3-карбоновая кислота, 4-хлорофенилаланин, 2-фторфенилаланин, 3-фторфенилаланин, 4-фторфенилаланин, пиридилаланин или 3-бензотиенилаланин.
В другом варианте осуществления данного изобретения предлагается способ лечения ран или устранения результатов воздействия старения, предусматривающий применение терапевтически эффективного количества пептида, имеющего последовательность SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12 или SEQ ID NO: 13.
Описание чертежей
На Фиг.1 приведены согласно CLUSTAL Х (версия 1.8) множественное выравнивание последовательностей вырезаемых связывающих участков выбранных проферментных форм белка ММП. На Фиг.1А показано выравнивание, на котором ярко выделены консервативные остатки, где символ '*' обозначает полную идентичность между последовательностями, символ ':' обозначает положения, для которых степень консервативности составляет 7/9, а символ '.' обозначает положения, для которых степень идентичности составляет более 80% с преимущественно консервативными заменами. На Фиг.1В гетерогенные положения выделены жирным шрифтом.
На Фиг.2 показана структура проферментной формы ММП-1 (Банк данных для белков файл 1FBL.ENT). Область расположения последовательностей SEQ ID NO: 2-10, приведенных в таблице 1, охватывает короткий участок между двумя крупными доменами. При активации фермента этот участок вырезается.
На Фиг.3 показана трехмерная модель матриксной металлопротеиназы-9. Вырезаемый участок, образующий N-конец активной протеиназы, обозначен штриховкой. Два иона цинка изображены в виде сфер. Вырезаемый доменный пептид может связываться с матриксной металлопротеиназой вблизи от места его нормального расположения в проферменте. Это связывание (также рядом с каталитическим цинком) стерически блокирует часть активного участка. Эта блокировка препятствует связыванию с субстратом.
На Фиг.4 приведен график ингибирования активности ММП-9 вырезаемым доменным пептидом 19-mer (SEQ ID NO: 11). ММП-9 смешивался с пептидом 19-mer (SEQ ID NO: 11) перед проведением ФРПЭ анализа. Использовались следующие концентрации пептида 19-mer (SEQ ID NO: 11): 0 мМ (черные кружки), 0.01 мМ (белые кружки), 0.03 мМ (черные квадраты), 0.06 мМ (белые квадраты), 0.125 мМ (черные треугольники), 0.25 мМ (белые треугольники), 0.5 мМ (крестики), 1 мМ (перевернутые черные треугольники), 2 мМ (перевернутые белые треугольники).
На Фиг.5 приведен график ингибирования активности ММП-9 пептидом вырезаемого домена 10-mer (SEQ ID NO: 13). ММП-9 смешивался с пептидом 10-mer (SEQ ID NO: 13) перед проведением ФРПЭ анализа. Использовались следующие концентрации пептида 10-mer (SEQ ID NO: 13): 0 мМ (черные треугольники), 0.25 мМ (белые треугольники), 0.5 мМ (перевернутые белые треугольники), 1.0 мМ (перевернутые черные треугольники), 2.0 мМ (крестики).
На Фиг.6 показано ингибирование активности ММП-9 пептидом вырезаемого домена 9-mer (SEQ ID NO: 12). ММП-9 смешивался с пептидом 9-mer (SEQ ID NO: 12) перед проведением ФРПЭ анализа. Использовались следующие концентрации пептида 9-mer (SEQ ID NO: 12): 0 мМ (черные треугольники), 0.25 мМ (белые треугольники), 0.5 мМ (перевернутые белые треугольники), 1.0 мМ (перевернутые черные треугольники), 2.0 мМ (крестики).
На Фиг.7 показано ингибирование активности ММП-9 пептидом вырезаемого домена 19-mer (SEQ ID NO: 11). ММП-9 смешивался с пептидом 19-mer (SEQ ID NO: 11) перед проведением флуоресцентного анализа коллагена. Использовались следующие концентрации пептида 19-mer (SEQ ID NO: 11): 0 мМ (черные кружки), 0.06 мМ (белые ромбики), 0.1 мМ (белые квадраты), 0.25 мМ (белые кружки), 0.5 мМ (крестики).
На Фиг.8 показано продолжительное ингибирование активности ММП-9 пептидом вырезаемого домена 19-mer (SEQ ID NO: 11). ММП-9 смешивался с пептидом 19-mer (SEQ ID NO: 11) перед проведением флуоресцентного анализа коллагена. Использовались следующие концентрации пептида 19-mer (SEQ ID NO: 11): 0 мМ (черные кружки), 0.06 мМ (белые ромбики), 0.1 мМ (белые квадраты), 0.25 мМ (белые кружки), 0.5 мМ (крестики).
На Фиг.9 показано продолжительное ингибирование активности ММП-9 пептидом вырезаемого домена 10-mer (SEQ ID NO: 13). ММП-9 смешивался с пептидом 10-mer (SEQ ID NO: 13) перед проведением флуоресцентного анализа коллагена. Использовались следующие концентрации пептида 10-mer (SEQ ID NO: 13): 0 мМ (белые кружки), 0.1 мМ (белые ромбики), 0.2 мМ (белые квадраты), 0.4 мМ (крестики).
На Фиг.10 показано продолжительное ингибирование активности ММП-9 пептидом вырезаемого домена 9-mer (SEQ ID NO: 12). ММП-9 смешивался с пептидом 9-mer (SEQ ID NO: 12) перед проведением флуоресцентного анализа коллагена. Использовались следующие концентрации пептида 9-mer (SEQ ID NO: 12): 0 мМ (черные кружки), 0.06 мМ (белые ромбики), 0.1 мМ (белые квадраты), 0.25 мМ (белые кружки), 0.5 мМ (крестики).
На Фиг.11 приведены профили элюирования ВЭЖХ для типичной реакции сплайсинга. Стрелки указывают, что площадь первого пика уменьшается в процессе реакции (пик проферментной формы ММП-9), в то время как площади двух следующих пиков (зрелый ММП-9 и N-концевой продукт расщепления, соответственно) увеличиваются.
На Фиг.12 показано превращение проферментной формы ММП-9 в N-концевой и С-концевой домены под действием стромилизина. Про-ММП-9 реагировала со стромилизином в присутствие пептида 19-mer (SEQ ID NO: 11) в следующих концентрациях: 0 мкМ (черные кружки), 0.5 мкМ (белые квадраты) или 1.0 мкМ (черные квадраты). В указанные моменты времени отбиралась аликвота и подвергалась ВЭЖХ. Площадь пика проферментной формы матриксной металлопротеиназы суммировалась и принималась за 100 процентов для пробы, взятой в нулевой момент времени. Белые кружки обозначают проферментную форму матриксной металлопротеиназы, инкубируемую в буферном растворе, не содержащем стромилизина или пептида 19-mer (SEQ ID NO: 11).
На Фиг.13А приведены данные изотермического титрометрического калориметрического анализа взаимодействия ингибиторного пептида 19-mer (SEQ ID NO: 11) с ферментом ММП-9. Каждый пик обозначает количество тепла, выделившегося после введения и последующей реакции связывания. На Фиг.13В изображена изотерма связывания, полученная путем интегрирования по времени величин пиков для каждой инъекции из Фиг.13А.
На Фиг.14 приведены данные изотермического титрометрический калориметрического анализа взаимодействия ингибиторного пептида 19-mer (SEQ ID NO: 11) с ферментом ММП-2. На Фиг.14А показаны исходные данные изотермического калориметрического анализа для титрования пептида 19-mer (SEQ ID NO: 11) (1 мМ) в растворе, содержащем ММП-2 (20 мкМ), 20 мМ какодилата (рН 6.8) и 10 мМ NaCl при температуре 25°С. Каждый пик обозначает количество тепла, выделившегося после введения и последующей реакции связывания. На Фиг.14В изображена изотерма связывания, полученная путем интегрирования по времени величин пиков для каждой инъекции из Фиг.14А.
На Фиг.15 изображена изотерма поверхностного плазменного резонансного связывания при пропускании раствора, содержащего пептид 19-mer (SEQ ID NO: 11), по поверхности иммобилизованного ММП-9.
На Фиг.16 изображена гистограмма, отображающая процентное соотношение живых клеток по сравнению с положительным контролем на кожной модели после обработки пептидом в двух различных концентрациях. Первый образец, обработанный солевым фосфатным буферным раствором (PBS), служил в качестве положительного контроля для определения степени жизнеспособность клеток, принимаемой за 100%. Второй образец служил в качестве отрицательного контроля, в котором клетки подвергались воздействию 1% Triton-X100, демонстрируя, что данный анализ позволяет регистрировать гибель клеток. Следующие три образца обрабатывались пептидами 9-mer (SEQ ID NO: 12), 10-mer (SEQ ID NO: 13) и 19-mer (SEQ ID NO: 11) в концентрации 500 мкМ. Последние три образца обрабатывались пептидами 9-mer (SEQ ID NO: 12), 10-mer (SEQ ID NO: 13) и 19-mer (SEQ ID NO: 11) в концентрации 2 мМ. Приведенные данные даны для усредненных значений по трем экспериментам.
На Фиг.17 графически отображена динамика заживления ран у диабетических мышей db/db. На графике показано изменение относительной средней площади раны у мышей в зависимости от количества дней, прошедших после ее нанесения при обработке либо обычным солевым раствором (белые кружки) или 20 мкг/мл пептида 19-mer (SEQ ID NO: 11) (черные кружки). Приведенные данные даны для усредненного относительного диаметра раны, полученного по результатам измерения для десяти экспериментальных животных.
На Фиг.18 изображена гистограмма пролиферации нормальных человеческих кожных фибробластов (Clonetics, CC-2509) в присутствие пептида 19-mer (SEQ ID NO: 11) и без него. Клеточный рост определялся по оптической плотности (OD) на длине волны 490 нм для трех различных концентраций пептида. Столбик "19mer6" характеризует рост клеток в присутствие пептида 19-mer (SEQ ID NO: 11) при концентрации 1×10-6 М. Столбик "19mer5" характеризует рост клеток в присутствие пептида 19-mer (SEQ ID NO: 11) при концентрации 1×105 М. Столбик "19mer4" характеризует рост клеток в присутствие пептида 19-mer (SEQ ID NO: 11) при концентрации 1×10-4 М. Контрольные клетки выращивались без добавления пептида.
На Фиг.19 также изображена гистограмма пролиферации нормальных человеческих кератиноцитов в присутствие пептида 19-mer (SEQ ID NO: 11) и без него. Как видно из Фиг.19, добавление пептида 19-mer (SEQ ID NO: 11) приводит к усилению роста кератиноцитов в зависимости от применяемой дозы. Контрольные клетки без добавления пептида 19-mer имели наименьшую клеточную плотность. Клетки, получавшие лишь 1×10-5 М пептида 19-mer (SEQ ID NO: 11, обозначенные "19mer5" на Фиг.19), имели значительно большую клеточную плотность (Р>0.01), чем клетки, не получавшие пептида 19-mer. Клетки, получавшие 1×10-4 М пептида 19-mer (обозначенные "19mer4" на Фиг.19), демонстрировали еще более интенсивный клеточный рост (Р>0.001). Однако клетки, получавшие 1×10-6 М пептида 19-mer (обозначенные "19mer6" на Фиг.19), демонстрировали достаточно слабую клеточную пролиферацию (Р>0.05), что было признано статистически незначимым.
На Фиг.20 изображена 48-луночная хемотаксическая камера (Neuroprobe, Inc.), используемая для измерения миграции нормальных человеческих кожных фибробластов (НЧКФ).
На Фиг.21А и В показана миграция нормальных человеческих кожных фибробластов (НЧКФ). На Фиг.21А изображена мембрана для измерения миграции с 8 мкм порами (выглядящими как кружки) без клеток НЧКФ (300-кратное увеличение). На Фиг.21В изображена та же мембрана после миграции НЧКФ при добавлении препарата положительного контроля (плазматический фибронектин 1.25 мкг/мл). Эта фотография сделана со 300-кратным увеличением. Ядра клеток НЧКФ окрашены в фиолетовый цвет. Некоторые клетки НЧКФ мигрировали через мембрану, в то время как другие застряли в 8 мкм порах.
На Фиг.22 изображена гистограмма, отображающая процентное соотношение мигрирующих нормальных человеческих кожных фибробластов (НЧКФ) по отношению к мигрирующим НЧКФ в экспериментах с положительным контролем (плазматический фибронектин) при различных концентрациях пептида 19-mer (SEQ ID NO: 11). Поскольку некоторые хемотаксические вещества обладают очень узким диапазоном активных концентраций, в начальных экспериментах использовались десятикратные разведения пептида 19-mer. На гистограмме приведены средние значения по трем отдельным экспериментам (данные для концентрации 100 мкг/мл отображают среднее значение по шести экспериментам). Концентрации 1000 и 100 мкг/мл пептида 19-mer (SEQ ID NO: 11) являлись хемотаксическими для фибробластов (55±3% и 46±3% в положительном контроле соответственно). Количество мигрирующих фибробластов при использовании концентраций пептида 19-mer (SEQ ID NO: 11) меньше 100 мкг/мл было лишь примерно в два раза выше, чем в эксперименте с отрицательным контролем и что было признано статистически незначимым.
На Фиг.23 изображена гистограмма, отображающая процентное соотношение мигрирующих нормальных человеческих кожных фибробластов (НЧКФ) по отношению к мигрирующим НЧКФ в экспериментах с положительным контролем (фибронектин) при различных концентрациях пептида 19-mer (SEQ ID NO: 11) и его производных. При концентрации 100 мкг/мл ацетилированный пептид 19-mer (SEQ ID NO: 11) (Ac-19-mer) инициировал миграцию НЧКФ до примерно той же степени, что и неацетилированный пептид 19-mer, но при более высоких концентрациях Ас-19-mer действовал менее эффективно. Пептиды 9-mer (SEQ ID NO: 12) и 10-mer (SEQ ID NO: 13) активизировали миграцию НЧКФ только при концентрации 1000 мкг/мл. Интересно, что пептиды 14-mer TMRKPRCGNPDVAN (SEQ ID NO: 19) и 17-mer TLKAMRKPRCGNPDVAN (SEQ ID NO: 20) не обладают хемотаксической активностью для НЧКФ ни при каких концентрациях (данные не показаны). Начало последовательностей пептидов 14-mer TMRKPRCGNPDVAN (SEQ ID NO: 19) и 17-mer TLKAMRKPRCGNPDVAN (SEQ ID NO: 20), полученных из ферментов матриксных металлопротеиназ, лишь немного смещено к N-концу. Следовательно, аминокислотная последовательность пептида существенна для индукции миграции НЧКФ.
На Фиг.24 изображена гистограмма, свидетельствующая, что добавление пептида 19-mer (SEQ ID NO: 11) приводит к усилению синтеза коллагена в клетках человеческих кожных фибробластов. Контрольные человеческие кожные фибробласты, не получавшие пептида 19-mer (SEQ ID NO: 11), производили лишь небольшое количество коллагена. Напротив, клетки, получавшие лишь 1×10-6 М пептида 19-mer (SEQ ID NO: 11, обозначенные "19mer6" на Фиг.24) или 1×10-5 М пептида 19-mer (SEQ ID NO: 11, обозначенные "19mer5" на Фиг.24), имели значительно большую клеточную плотность (Р>0.001), чем клетки, не получавшие пептида 19-mer.
Подробное описание изобретения
Объектом настоящего изобретения являются ингибиторы матриксных металлопротеиназ, пригодные для противодействия результатам воздействия старения на кожу и для стимулирования процессов заживления ран. В целом, представленные ингибиторы и композиции способствуют заживлению ран, предотвращают рубцевание, улучшают цвет кожи, снижают образование морщин и стимулируют развитие гладкой здоровой кожи.
Матриксные металлопротеиназы синтезируются in vivo в виде неактивных проферментов. Протеолитическое расщепление профермента приводит к активации и образованию зрелой матриксной металлопротеиназы. Отщепляемая пептидная последовательность является лидерной последовательностью профермента длиной около от 100 до 110 аминокислот, которая располагается на аминоконце белка. Согласно данному изобретению, эти лидерные пептиды профермента могут блокировать активный участок матриксных металлопротеиназ и ингибировать активность матриксной металлопротеиназы. Применение лидерных пептидов профермента матриксной металлопротеиназы снижает скорость разрушения внеклеточного матрикса и ускоряет процесс заживления ран.
Большинство стратегий ингибирования заключается в подавлении ферментативной активности матриксных металлопротеиназ с помощью небольших органических молекул. Эти составы зачастую оказываются токсичными для организма и содержат искусственные молекулы, не встречающиеся в природе. Использование природных пептидов для активированных матриксных металлопротеиназ обеспечивает высокую степень контроля за уровнем протеиназной активности, не оказывая токсического побочного действия. В отличие от стратегий ингибирования с использованием небольших молекул пептиды, заявленные в данном изобретении, могут использоваться для ингибирования активации отдельных или всех классов матриксных металлопротеиназ одновременно. Эти пептиды могут непосредственно наноситься на кожу, вводиться в рану, или их можно привязывать или наносить посредством кожных покрытий или перевязочного материала для ран.
Данное изобретение обеспечивает высокую степень контроля за уровнем протеиназной активности для заживления хронических ран и улучшения последствий старения. Например, поскольку при заживлении хронических ран требуется некоторый уровень протеиназной активности (Agren et al., 1999), специалисту в данной области может потребоваться лишь частичное ингибирование протеиназной активности. Регулируя тип и количество применяемого ингибиторного пептида, можно контролировать степень ингибирования матриксной металлопротеиназы.
Пептиды
Согласно настоящему изобретению пептиды, имеющие последовательности, родственные лидерной последовательности проферментной формы матриксной металлопротеиназы в области вырезаемого участка, пригодны для заживления ран и для стимулирования развития здоровой кожи. Эти пептиды ингибируют активность различных типов матриксных металлопротеиназ и способствуют клеточному росту и миграции фибробластов и кератиноцитов.
Положение, по которому отщепляется лидерная последовательность проферментной формы матриксной металлопротеиназы, располагается около 110 аминокислотного положения в аминокислотной последовательности профермента. Пептидные ингибиторы данного изобретения имеют последовательности, родственные любой области внутри профермента в пределах примерно с 70 по 120 аминокислоту. Подобные пептиды ингибируют активность многих типов матриксных металлопротеиназ. Данные пептиды могут также предотвращать активацию проферментной формы матриксных металлопротеиназ, а также ингибировать ферментативную активность зрелых матриксных металлопротеиназ. Пептиды, содержащие последовательности, обладающие более высокой консервативностью среди различных матриксных металлопротеиназ, например последовательности, расположенные ближе к N-концевой области вырезаемого участка, могут использоваться для создания ингибиторов, в целом обладающих эффективностью в отношении широкого спектра матриксных металлопротеиназ. В то же время пептиды, содержащие менее консервативные последовательности, например последовательности, расположенные ближе к С-концевой области вырезаемого участка, могут использоваться для создания ингибиторов, обладающих специфичностью в отношении определенных матриксных металлопротеиназ.
Следовательно, пептиды с последовательностями из любой лидерной области проферментной формы матриксной металлопротеиназы рассматриваются согласно настоящему изобретению как ингибиторы матриксных металлопротеиназ, так же, как и варианты этих пептидов, у которых одна или несколько аминокислот заменены аминокислотами, присутствующими в природной матриксной металлопротеиназе. Смеси пептидов, имеющих различные последовательности, также рассматриваются в данном изобретении.
В целом, пептидные последовательности, варианты пептидов и смеси пептидов составляются и применяются таким образом, чтобы оптимизировать заживление ран, восстановление кожи, предотвращение рубцевания или устранение морщин и предотвращение их образования. Следовательно, состав данных пептидов можно варьировать, снижая или повышая степень ингибирования, с тем, чтобы стимулировать процессы заживления и противодействия старению.
Размер пептидного ингибитора может варьироваться. Как правило, пептид, содержащий лишь порядка пяти аминокислот, может оказаться слишком коротким для обеспечения оптимального ингибирования. Однако пептиды, состоящие из более чем восьми-девяти аминокислот, являются достаточно длинными для обеспечения ингибирования. Поэтому, хотя общая длина не является критическим параметром, в рамках данного изобретения часто используются пептиды длиной более восьми аминокислот. Также в рамках данного изобретения используются пептиды длиной более девяти аминокислот. Кроме того, в рамках данного изобретения используются пептиды длиной более десяти аминокислот. Более того, в рамках данного изобретения также используются пептиды длиной более чем примерно пятнадцать аминокислот. На максимальный размер пептида не накладывается никаких определенных ограничений. Однако, обычно оказывается дешевле синтезировать более короткие, чем более длинные пептиды. Поэтому длина пептидных ингибиторов согласно настоящему изобретению обычно не превышает ста аминокислот. Длина большинства пептидных ингибиторов, используемых в рамках данного изобретения, не превышает примерно пятидесяти аминокислот. Также в рамках данного изобретения используются пептидные ингибиторы, содержащие не более чем порядка тридцати аминокислот. Также могут использоваться пептиды с длиной менее чем примерно двадцать пять аминокислот. Аналогично, в рамках данного изобретения могут использоваться пептиды, длина которых составляет не более двадцати трех аминокислот. Примером пептида, используемого в рамках данного изобретения, может служить SEQ ID NO: 11 с девятнадцатью аминокислотами.
Последовательности проферментных форм нескольких типичных матриксных металлопротеиназ приблизительно с 70 по 120 аминокислоту приведены в таблице 1.
Каждый из пептидов, перечисленных в таблице 1, а также пептиды с SEQ ID NO: 1, 11, 12 и 13, рассматриваются в качестве пептидных ингибиторов настоящего изобретения. Более того, варианты и производные пептидов, имеющих любую из последовательностей SEQ ID NO: 1-13, также могут использоваться в качестве пептидных ингибиторов. Подобные варианты и производные пептидов могут иметь одну или несколько аминокислотных замен, делеций, вставок или иных модификаций при условии, что эти варианты или производные пептидов смогут ингибировать матриксную металлопротеиназу.
В качестве аминокислотных остатков отдельных пептидов могут выступать кодируемые генетически L-аминокислоты, природные не кодируемые генетически L-аминокислоты, синтетические L-аминокислоты или D-энантиомеры любой из вышеперечисленных групп. В рамках данного изобретения используются традиционные условные обозначения для двадцати кодируемых генетически L-аминокислот и обычных некодируемых аминокислот, которые приведены в таблице 2.
У пептидов, включенных в объем данного изобретения, одна или несколько аминокислот могут быть замещены другими аминокислотами, обладающими сходными химическими и/или физическими свойствами, при условии, что эти варианты или производные пептидов будут сохранять способность ингибировать активность матриксной металлопротеиназы, стимулировать рост клеток фибробластов или кератиноцитов или стимулировать миграцию клеток фибробластов.
Аминокислоты, которые могут замещать друг друга, обычно принадлежат одному и тому же классу или подклассу. Как известно специалистам в данной области, аминокислоты можно разбить на три основных класса: гидрофильные аминокислоты, гидрофобные аминокислоты и цистеиноподобные аминокислоты, в зависимости, главным образом, от характеристик боковой цепи аминокислоты. Эти основные классы могут быть далее разбиты на подклассы. К гидрофильным аминокислотам относятся аминокислоты, обладающие кислой, основной или полярной боковыми цепями, а к гидрофобным аминокислотам относятся аминокислоты, обладающие ароматической или неполярной боковыми цепями. Неполярные аминокислоты в свою очередь можно подразделить на подклассы, включающие в себя, в частности, алифатические аминокислоты. Ниже приводятся определения классов аминокислот, которые используются в рамках данного изобретения:
"Гидрофобной аминокислотой" называется аминокислота, имеющая незаряженную при физиологических значениях рН и отталкиваемую водным раствором боковую цепь. Примерами генетически кодируемых гидрофобных аминокислот являются изолейцин, лейцин и валин. Примером не кодируемой генетически гидрофобной аминокислоты является t-бутилаланин.
"Ароматической аминокислотой" называется гидрофобная аминокислота, имеющая боковую цепь, содержащую по меньшей мере одно кольцо, имеющее сопряженную π-электронную систему (ароматическую группу). Ароматическая группа может быть также замещена замещающими группами, такими как алкил, алкенил, алкинил, гидроксил, сульфонил, нитро- и аминогруппы и другие. Примеры генетически кодируемых ароматических аминокислот включают фенилаланин, тирозин и триптофан. Примеры наиболее распространенных не кодируемых генетически ароматических аминокислот включают фенилглицин, 2-нафтилаланин, β-2-тиенилаланин, 1,2,3,4-тетрагидроизохинолин-3-карбоновая кислота, 4-хлорофенилаланин, 2-фторфенилаланин, 3-фторфенилаланин и 4-фторфенилаланин.
"Неполярной аминокислотой" называется гидрофобная аминокислота, имеющая боковую цепь, обычно незаряженную при физиологических значениях рН и не являющуюся полярной. Примерами генетически кодируемых неполярных аминокислот являются глицин, пролин и метионин. Примером не кодируемой генетически неполярной аминокислоты может служить циклогексилаланин.
"Алифатической аминокислотой" называется неполярная аминокислота, имеющая насыщенную или ненасыщенную неразветвленную цепь, разветвленную или циклическую углеводородную боковую цепь. Примеры генетически кодируемых алифатических аминокислот включают аланин, лейцин, валин и изолейцин. Примером не кодируемой генетически алифатической аминокислоты является циклогексилаланин.
"Гидрофильной аминокислотой" называется аминокислота, имеющая боковую цепь, притягиваемую водным раствором. Примеры генетически кодируемых гидрофильных аминокислот включают серин и лизин. Примером не кодируемых генетически гидрофильных аминокислот являются цитруллин и гомоцистеин.
"Кислой аминокислотой" называется гидрофильная аминокислота, имеющая боковую цепь со значением рК меньше 7. Кислые аминокислоты при физиологических значениях рН обычно имеют отрицательно заряженные в результате потери иона водорода боковые цепи. Примерами генетически кодируемых кислых аминокислот являются аспарагиновая кислота (аспартат) и глутаминовая кислота (глутамат).
"Основной аминокислотой" называется гидрофильная аминокислота, имеющая боковую цепь со значением рК более 7. Основные аминокислоты при физиологических значениях рН обычно имеют положительно заряженные в результате присоединения иона гидроксония боковые цепи. Примеры генетически кодируемых основных аминокислот включают аргинин, лизин и гистидин. Примером не кодируемых генетически основных аминокислот являются нециклические аминокислоты орнитин, 2,3-диаминопропионовая кислота, 2,4-диаминомасляная кислота и гомоаргинин.
"Полярной аминокислотой" называется гидрофильная аминокислота, имеющая незаряженную при физиологических значениях рН боковую цепь, но содержащую межатомную связь, в которой пара обобществленных электронов смещена к одному из атомов. Примеры генетически кодируемых полярных аминокислот включают аспарагин и глутамин. Примеры не кодируемых генетически полярных аминокислот включают цитруллин, N-ацетил лизин и метионин сульфоксид.
"Цистеиноподобной аминокислотой" называется аминокислота, имеющая боковую цепь, способную к образованию ковалентной связи с боковой цепью другого аминокислотного остатка, например дисульфидного мостика. Обычно боковые цепи цистеиноподобных аминокислот содержат по меньшей мере одну тиольную (SH) группу. Примером генетически кодируемой цистеиноподобной аминокислоты является цистеин. Примерами не кодируемых генетически цистеиноподобных аминокислот являются гомоцистеин и пеницилламин.
Как могут отметить специалисты в данной области, приведенная выше классификация не является абсолютной. Некоторые аминокислоты обладают несколькими характерными свойствами, и поэтому могут быть включены в более чем одну категорию. Так, к примеру, тирозин имеет как ароматическое кольцо, так и полярную гидроксильную группу. Таким образом, тирозин обладает двумя характерными свойствами и может быть причислен к категории как ароматических, так и полярных аминокислот. Аналогичным образом цистеин, помимо способности образовывать дисульфидные мостики, также обладает характеристиками неполярных аминокислот. Таким образом, не являясь строго классифицированным как гидрофобная или неполярная аминокислота, во многих случаях цистеин может использоваться для придания пептиду гидрофобных свойств.
Некоторые часто встречающиеся аминокислоты, которые не кодируются генетически, но могут присутствовать или замещать другие аминокислоты в пептидах и их аналогах, описанных в данном изобретении, включают (но не ограничиваются этим перечнем) β-аланин (b-Ala) и другие омега-аминокислоты, такие как 3-аминопропионовая кислота (Dap), 2,3-диаминопропионовая кислота (Dpr), 4-аминомасляная кислота и так далее; α-аминоизомасляная кислота (Aib); ε-аминогексановая кислота (Alia); δ-аминовалериановая кислота (Ava); метилглицин (MeGly); орнитин (Orn); цитруллин (Cit); t-бутилаланин (t-BuA); t-бутилглицин (t-BuG); N-метилизолейцин (MeIle); фенилглицин (Phg); циклогексилаланин (Cha); норлейцин (Nle); 2-нафтилаланин (2-Nal); 4-хлорофенилаланин (Phe (4-Cl)); 2-фторфенилаланин (Phe (2-F)); 3-фторфенилаланин (Phe (3-F)); 4-фторфенилаланин (Phe (4-F)); пеницилламин (Pen); 1,2,3,4-тетрагидроизохинолин-3-карбоновая кислота (Tic); β-2-тиенилаланин (Thi); метионин сульфоксид (MSO); гомоаргинин (hArg); N-ацетил лизин (AcLys); 2,3-диаминомасляная кислота (Dab); 2,3-диаминомасляная кислота (Dbu); ρ-аминофенилаланин (Phe (pNH2)); N-метил валин (MeVal); гомоцистеин (hCys) и гомосерин (hSer). Эти аминокислоты также попадают в определенные выше категории.
Классификации вышеописанных кодируемых и не кодируемых генетически аминокислот приведены в таблице 3 ниже. Следует отметить, что таблица 3 служит только для пояснения и не означает исчерпывающий список аминокислотных остатков, которые могут быть включены в состав пептидов или их аналогов, описанных в данном изобретении. Другие аминокислотные остатки, которые пригодны для получения пептидов и их аналогов, описанных в данном изобретении, можно найти, например, в Fasman, 1989, CRC Practical Handbook of Biochemistry and Molecular Biology, CRC Press, Inc., и в приводимых там ссылках. Не указанные здесь аминокислоты можно легко классифицировать по вышеописанным категориям на основании их известных характеристик и/или характерных химических и/или физических свойств и сравнения их с конкретно определенными аминокислотами.
В пептидах согласно настоящему изобретению любая аминокислота может быть заменена другой аминокислотой из того же класса с образованием варианта или производного соответствующего пептида, при условии, что такие варианты или производные пептидов сохраняют способность ингибировать активность матриксной металлопротеиназы.
В одном из вариантов осуществления данного изобретения пептидные ингибиторы, заявленные в данном изобретении, включают в себя любой из пептидов формул I, II или III.
где
Xaa1, Xaa4 и Хаа6 независимо друг от друга обозначают неполярные аминокислоты, например метионин, глицин или пролин;
Хаа2 является основной аминокислотой, такой, например, как гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин, и 2,4-диаминомасляная кислота;
Хаа3 является цистеиноподобной аминокислотой, такой, например, как цистеин, гомоцистеин, пеницилламин или β-метил цистеин;
Хаа5 является полярной или алифатической аминокислотой, например полярной аминокислотой, такой как аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетиллизин, метионин сульфоксид или гомосерин или алифатической аминокислотой, такой как аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота;
Хаа7 является кислой аминокислотой, такой, например, как аспарагиновая кислота или глутаминовая кислота;
Xaa8 является алифатической или полярной аминокислотой, например алифатической аминокислотой, такой как аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота, или полярной аминокислотой, такой как аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин;
Хаа9 является алифатической, неполярной или основной аминокислотой, например, алифатической аминокислотой, такой как аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота, неполярной аминокислотой, такой как метионин, глицин или пролин, или основной аминокислотой, такой как гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин и 2,4-диаминомасляная кислота;
Хаа10 является полярной, кислой, основной или неполярной аминокислотой, например полярной аминокислотой, такой как аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин, кислой аминокислотой, такой как аспарагиновая кислота или глутаминовая кислота, основной аминокислотой, такой как гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин и 2,4-диаминомасляная кислота, или неполярной аминокислотой, такой как метионин, глицин или пролин;
Xaa11 является полярной или ароматической аминокислотой, например полярной аминокислотой, такой как аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин, или ароматической аминокислотой, такой как фенилаланин, тирозин, триптофан, фенилглицин, нафтилаланин, β-2-тиенилаланин, 1,2,3,4-тетрагидроизохинолин-3-карбоновая кислота, 4-хлорофенилаланин, 2-фторфенилаланин, 3-фторфенилаланин, 4-фторфенилаланин, пиридилаланин или 3-бензотиенилаланин;
Xaa12 является неполярной, основной, алифатической или неполярной аминокислотой, например неполярной аминокислотой, такой аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин, или основной аминокислотой, такой как гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин, и 2,4-диаминомасляная кислота, или алифатической аминокислотой, такой как аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота, или неполярной аминокислотой, такой как метионин, глицин или пролин;
Хаа13 является ароматической, алифатической, полярной или кислой аминокислотой, например ароматической аминокислотой, такой как фенилаланин, тирозин, триптофан, фенилглицин, нафтилаланин, β-2-тиенилаланин, 1,2,3,4-тетрагидроизохинолин-3-карбоновая кислота, 4-хлорофенилаланин, 2-фторфенилаланин, 3-фторфенилаланин, 4-фторфенилаланин, пиридилаланин или 3-бензотиенил аланин, или алифатической аминокислотой, такой как аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота, или полярной аминокислотой, такой как аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин, или кислой аминокислотой, такой как аспарагиновая кислота или глутаминовая кислота;
Xaa14 является ароматической, неполярной или полярной аминокислотой, например ароматической аминокислотой, такой как фенилаланин, тирозин, триптофан, фенилглицин, нафтилаланин, β-2-тиенилаланин, 1,2,3,4-тетрагидроизохинолин-3-карбоновая кислота, 4-хлорофенилаланин, 2-фторфенилаланин, 3-фторфенилаланин, 4-фторфенилаланин, пиридилаланин или 3-бензотиенил аланин, или неполярной аминокислотой, такой как метионин, глицин или пролин, или полярной аминокислотой, такой как аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин;
Xaa15 является неполярной или кислой аминокислотой, например неполярной аминокислотой, такой как метионин, глицин или пролин, или кислой аминокислотой, такой как аспарагиновая кислота или глутаминовая кислота;
Xaa16 является основной, полярной или неполярной аминокислотой, например основной аминокислотой, такой как гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин и 2,4-диаминомасляная кислота; или полярной аминокислотой, такой как аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин, или неполярной аминокислотой, такой как метионин, глицин или пролин;
Xaa17 является основной, полярной, алифатической, неполярной или кислой аминокислотой, например основной аминокислотой, такой как гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин, и 2,4-диаминомасляная кислота, или полярной аминокислотой, такой как аспарагин, глутамин, серин, треонин, тирозин, цитруллин, N-ацетил лизин, метионин сульфоксид или гомосерин, или алифатической аминокислотой, такой как аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота или неполярной аминокислотой, такой как метионин, глицин или пролин, кислой аминокислотой, такой как аспарагиновая кислота или глутаминовая кислота;
Xaa18 является неполярной или алифатической аминокислотой, например неполярной аминокислотой, такой как метионин, глицин или пролин, или алифатической аминокислотой, такой как аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота; и
Xaa19 является основной или алифатической аминокислотой, например основной аминокислотой, такой как гистидин, лизин, аргинин, 2,3-диаминопропионовая кислота, орнитин, гомоаргинин, ρ-аминофенилаланин и 2,4-диаминомасляная кислота, или алифатической аминокислотой, такой как аланин, валин, лейцин, изолейцин, t-бутилаланин, t-бутилаланин, N-метилизолейцин, норлейцин, N-метилвалин, циклогексилаланин, β-аланин, N-метилглицин или α-аминоизомасляная кислота.
В некоторых вариантах осуществления:
Xaa1 обозначает пролин,
Хаа2 обозначает аргинин,
Хаа3 обозначает цистеин,
Хаа4 обозначает глицин,
Xaa5 обозначает валин или аспарагин,
Хаа6 обозначает пролин,
Хаа7 обозначает аспарагиновую кислоту,
Xaa8 обозначает валин, лейцин или серин,
Хаа9 обозначает аланин, глицин или гистидин,
Xaa10 обозначает аспарагин, аспарагиновую кислоту, гистидин, аргинин, глутамин или глицин,
Xaa11 обозначает тирозин или фенилаланин,
Xaa12 обозначает аспарагин, серин, аргинин, глутамин, валин или метионин,
Xaa13 обозначает фенилаланин, валин, лейцин, треонин, серин или глутаминовую кислоту,
Xaa14 обозначает фенилаланин, метионин или треонин,
Xaa15 обозначает пролин или глутаминовую кислоту,
Xaa16 обозначает аргинин, аспарагин или глицин,
Xaa17 обозначает лизин, треонин, серин, изолейцин, метионин, глицин, аспарагиновую кислоту или аспарагин,
Xaa18 обозначает пролин или лейцин, и
Xaa19 обозначает лизин, валин или аргинин.
В рамках данного изобретения желательными также являются пептиды, включающие в себя последовательности SEQ ID NO: 1-13. Одним из примеров желательного пептида может служить пептид, состоящий из девятнадцати аминокислот, обладающий SEQ ID NO: 11 (PRCGNPDVANYNFFPRKPK). Этот пептид (SEQ ID NO: 11) охватывает вырезаемый участок ММП-2. Два меньших пептида (PRCGNPDVA (SEQ ID NO: 12) и NYNFFPRKPK (SEQ ID NO: 13)), представляющие собой примерно половину пептида SEQ ID NO: 11, также являются желательными пептидами. Все три пептида в различной степени ингибируют активность ММП-9 и других матриксных металлопротеиназ.
Одиночный пептид с последовательностью, идентичной последовательности вырезаемого участка матриксной металлопротеиназы, может использоваться для ингибирования активности только одной или лишь небольшого числа матриксных металлопротеиназ. Составы на основе таких одиночных пептидов способны ингибировать одну или несколько, но обычно не все, матриксные металлопротеиназы. Подобное частичное ингибирование активности матриксной металлопротеиназы может способствовать заживлению ран. Альтернативно, два или более пептида можно сочетать для воздействия на две или более матриксные металлопротеиназы, что может обеспечить более полное ингибирование активности матриксной металлопротеиназы.
Специалист в данной области может сконструировать подходящий пептидный ингибитор или комбинацию пептидных ингибиторов для достижения требуемого качества и степени ингибирования, пользуясь известными способами в сочетании со способами, изложенными в рамках данного изобретения. "Качество" ингибирования означает тип ингибируемой матриксной металлопротеиназы. Различные матриксные металлопротеиназы могут иметь несколько различающихся субстратов и участков, на которые они воздействуют. "Степень" ингибирования означает общую степень ингибирования для всех матриксных металлопротеиназ. Варьируя тип и количество используемого пептидного ингибитора, можно варьировать качество и степень ингибирования. Специалисту в данной области не составит труда выполнить модификации пептидов, описанных в рамках данного изобретения, и добиться требуемого типа и степени ингибирования данной матриксной металлопротеиназы.
Например, специалист в данной области может сравнить и произвести выравнивание пептидных последовательностей, приведенных на Фиг.1, и сконструировать соответствующий пептидный ингибитор требуемого качества и степени ингибирования. В одном из вариантов осуществления, представленном в примере, сопоставляемые аминокислотные последовательности для трех матриксных металлопротеиназ, характерных для участка поврежденной ткани, mmp2, mmp9 и mmp1, сравниваются с целью выявления областей гомологии и областей расхождения в последовательности.

На этом выравнивании последовательностей жирным шрифтом обозначены аминокислоты, присутствующие в данном месте в белке ММП-1 и отсутствующие в белках ММП-2 или ММП-9, а подчеркнутым шрифтом обозначены аминокислоты, присутствующие в данном месте в белке ММП-1 и только в одном из белков ММП-2 или ММП-9.
В одном из вариантов осуществления представляется желательным ингибировать ММР-2 и ММР-9, но сохранить активность белка ММП-1 примерно на прежнем уровне для ингибирования развития опухолей или обеспечения заживления хронических ран. Исходя из приведенного выше выравнивания последовательностей, специалист в данной области может сконструировать пептид, содержащий аминокислоты, присутствующие в последовательностях проферментных форм ММР-2 и ММР-9, но отсутствующие в последовательности проферментной формы ММР-1, для получения пептида, способного ингибировать ММП-2 и ММП-9, но не ингибировать ММП-1. Такой пептид имеет формулу IV.
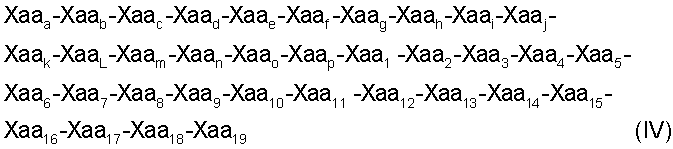
(SEQ ID NO: 18)
где:
Хааа обозначает пролин;
Хааb обозначает глутамин или глутаминовую кислоту;
Хаас обозначает треонин;
Xaad обозначает глицин;
Хааe обозначает аспарагиновую кислоту или глутаминовую кислоту;
Хааf обозначает лейцин;
Xaag обозначает аспарагиновую кислоту;
Хааh обозначает глутамин или серин;
Xaai обозначает аспарагин или аланин;
Xaaj обозначает треонин;
Xaak обозначает изолейцин или лейцин, предпочтительно изолейцин;
XaaL обозначает глутаминовую кислоту или лизин, предпочтительно глутаминовую кислоту;
Xaam обозначает треонин или аланин;
Хааn обозначает метионин;
Хаао обозначает аргинин;
Хаар обозначает лизин или треонин;
Хаа1 обозначает пролин;
Xaa2 обозначает аргинин;
Хаа3 обозначает цистеин;
Хаа4 обозначает глицин;
Хаа5 обозначает валин или аспарагин, предпочтительно аспарагин;
Хаа6 обозначает пролин;
Хаа7 обозначает аспарагиновую кислоту;
Xaa8 обозначает валин или лейцин, предпочтительно лейцин;
Хаа9 обозначает аланин или глицин, предпочтительно глицин;
Хаа10 обозначает аспарагин или аргинин;
Xaa11 обозначает тирозин или фенилаланин, предпочтительно тирозин;
Xaa12 обозначает аспарагин или глутамин;
Xaa13 обозначает фенилаланин или треонин;
Xaa14 обозначает фенилаланин;
Xaa15 обозначает пролин или глутаминовую кислоту, предпочтительно пролин;
Xaa16 обозначает аргинин или глицин, предпочтительно аргинин,
Xaa17 обозначает лизин или аспарагиновую кислоту;
Xaa18 обозначает пролин или лейцин, предпочтительно лейцин; и
Xaa19 обозначает лизин.
Модификации пептидов
Данное изобретение также предусматривает модифицирование пептидных ингибиторов для их стабилизации, облегчения их поглощения и всасывания, а также улучшения иных характеристик или свойств пептидов, известных специалистам в данной области. В частности, пептидные ингибиторы могут быть циклизованы, заряды на пептидных ингибиторах могут быть нейтрализованы, а сами пептиды могут быть соединены с другими химическими компонентами.
Пептиды могут быть циклизованы любым способом, известным специалистам в данной области. Например, N- и С-концы могут быть сконденсированы с образованием пептидной связи согласно известным процедурам. Функциональные группы, присутствующие в боковых цепях аминокислот в пептидах, также могут участвовать в циклизации пептидов, заявленных в данном изобретении. Примерами функциональных групп, способных образовывать ковалентные связи, являются -СООН и -ОН; -СООН и -NH2; и -СООН и -SH. Пары аминокислот, которые могут использоваться для циклизации пептида, включают Asp и Lys; Glu и Lys; Asp и Arg; Glu и Arg; Asp и Ser; Glu и Ser; Asp и Thr; Glu и Thr; Asp и Cys; и Glu и Cys. Другим примером аминокислотных остатков, способных к образованию ковалентных связей друг с другом, являются цистеиноподобные аминокислоты, такие как Cys, hCys, β-метил-Cys и Pen, которые могут образовывать дисульфидные мостики друг с другом. Примерами цистеиноподобных аминокислотных остатков могут служить Cys и Pen. Другие пары аминокислот, которые могут использоваться для циклизации пептида, будут очевидны специалистам в данной области.
Группы, используемые для циклизации пептида, не обязательно должны быть аминокислотами. К функциональным группам, способным к образованию ковалентной связи с аминоконцом пептида, относятся карбоновые кислоты и сложные эфиры. К функциональным группам, способным к образованию ковалентной связи с карбоксильным концом пептида, относятся -ОН, -SH, -NH2 и -NHR, где R представляет собой (С1-С6) алкильную, (C1-С6) алкенильную или (C1-С6) алкинильную группу.
Всевозможные реакции между двумя боковыми цепями с функциональными группами, подходящими для образования таких связей, а также условия реакции, подходящие для образования таких связей, хорошо известны специалистам в данной области. Условия реакции, используемые для циклизации пептидов, обычно выбираются достаточно мягкими во избежание разрушения или иного повреждения пептида. Подходящие группы, используемые при необходимости для защиты различных функциональных групп, хорошо известны специалистам (смотрите, например, Greene & Wuts, 1991, 2nd, John Wiley & Sons, NY), равно как и различные схемы реакций для получения таких защищенных молекул.
В одном из вариантов осуществления заряды на N- и С-концах эффективно удаляются. Это можно осуществить любым способом, известным специалистам в данной области, например, путем ацетилирования N-конца или амидирования С-конца.
Способы получения циклических или иных модификаций пептидов хорошо известны специалистам (смотрите, например, Spatola, 1983, Vega Data 1(3) для общего обзора); Spatola, 1983, "Peptide Backbone Modifications" (Модификации пептидного скелета) в книге: Chemistry and Biochemistry of Amino Acids, Peptides and Proteins (Химия и биохимия аминокислот, пептидов и белков) (Weinstein, ed.), Marcel Dekker, New York, стр.267 (общий обзор); Morley, 1980, Trends Pharm. Sci. 1:463-468; Hudson et al., 1979, Int. J. Prot. Res. 14:177-185 (-CH2NH-, -CH2CH2-); Spatola et al., 1986, Life Sci. 38:1243-1249 (-CH2-S); Harm, 1982, J. Chem. Soc. Perkin Trans. I. 1:307-314 (-CH=CH-, цис и транс); Almquist et al., 1980, J. Med. Chem. 23:1392-1398 (-COCH2-); Jennings-White et al., Tetrahedron. Lett. 23:2533 (-COCH2-); Европейская патентная заявка ЕР 45665 (1982) СА:97:39405 (-СН(ОН)CH2-); Holladay et al., 1983, Tetrahedron Lett. 24:4401-4404 (-С(ОН)CH2-); и Hruby, 1982, Life Sci. 31:189-199 (-CH2-S-).
Заживление ран и противодействие старению
Пептиды, заявленные в данном изобретении, могут использоваться для заживления ран, устранения результатов воздействия старения на кожу и в особенности для заживления хронических ран. Отдельные пептиды, варианты пептидов, производные пептидов и их смеси (т.е. смеси пептидов, имеющих различные последовательности) можно комбинировать при приготовлении состава для ускорения заживления ран и профилактики или лечения кожных проблем, связанных со старением. Для оптимального заживления ран и восстановления кожи может потребоваться некоторая активность матриксной металлопротеиназы. Таким образом, от композиций и составов настоящего изобретения не обязательно требуется максимальное ингибирование матриксных металлопротеиназ. Напротив, активность пептидного ингибитора варьируют для оптимизации процессов заживления и стимулирования развития здоровой кожи. Варьируя тип, содержание и количество ингибиторных пептидов можно обеспечить большую или меньшую степень ингибирования, стимулируя тем самым процессы заживления и развития здоровой кожи.
Для стимулирования развития здоровой кожи и/или лечения ран пептиды, заявленные в данном изобретении, могут наноситься на кожу или вводиться в рану любым способом, выбранным специалистом в данной области. В частности, данные пептиды могут быть приготовлены в виде терапевтической композиции, содержащей терапевтически эффективное количество одного или нескольких пептидов и фармацевтический носитель. Подобный состав может наноситься на кожу или вводиться в рану в виде крема, аэрозолей, пены, геля или в любой другой форме. В другом варианте осуществления пептиды согласно настоящему изобретению могут быть включены в состав кожных покрытий или перевязочного материала, содержащих терапевтически эффективное количество одного или нескольких пептидов, нанесенных путем пропитки, ковалентного связывания или иным образом на материал покрытия или перевязочный материал. В одном из вариантов осуществления кожное покрытие или перевязочный материал позволяют осуществлять высвобождение пептидного ингибитора. Высвобождение пептидного ингибитора может осуществляться неконтролируемым или контролируемым образом. Таким образом, кожные пластыри или перевязки для ран, описанные в данном изобретении, позволяют осуществлять медленное или спланированное по времени высвобождение пептидного ингибитора в рану. В качестве кожных покрытий и перевязочных материалов может использоваться любой материал, применяемый в данной области, в том числе бинты, марлю, стерильные обертки (sterile wrapping), гидрогели, гидроколлоиды и аналогичные материалы.
Терапевтически эффективное количество пептида, заявленного в данном изобретении, определяется как количество пептида, которое ингибирует матриксную металлопротеиназу до той степени, которая необходима для стимулирования развития здоровой кожи и/или заживления ран. В частности, при составлении терапевтической или фармацевтической композиции количество пептидов согласно настоящему изобретению может варьировать в пределах примерно от 0.001% до 35% массы композиции. Пептиды могут составлять примерно от 0.5% до 20% массы препарата. Альтернативно, пептиды могут составлять примерно от 1.0% до 10% массы препарата. Терапевтически эффективное количество пептидного ингибитора зависит от способа введения препарата. Например, для внутривенного введения терапевтически эффективное количество пептида может составлять от 30 до 112,000 мкг на килограмм веса тела. Однако, количество пептидного ингибитора, требуемого для развития здоровой кожи или лечения раны, может зависеть не только от способа введения, но также от диагноза, возраста и состояния пациента и, в конечном счете, определяется лечащим врачом.
Дозировку и способ применения можно варьировать в зависимости от расположения участка кожи или ткани, который предполагается обработать, и/или от степени серьезности раны. Используемые дозировки пептидов и пептидных конъюгатов могут быть определены путем сопоставления их активностей in vitro и in vivo на животных моделях, описываемых ниже. Это соединение удобно вводить в виде единичной дозы препарата; содержащей, к примеру, примерно от 0.001 мкг до 10 мг, предпочтительно примерно от 0.01 мкг до 5 мг, более предпочтительно примерно от 0.10 мкг до 1 мг, и еще более предпочтительно примерно от 1.0 мкг до 500 мкг пептида на единичную дозу препарата. Нужная доза может быть введена в один прием, в несколько приемов или путем непрерывной инфузии. Нужная доза может также назначаться к применению через определенные интервалы времени, например два, три, четыре или более раз в день. Специалисту в данной области не составит труда приготовить и назначить эффективный состав на основе представленной здесь информации.
Пептидные ингибиторы, заявленные в данном изобретении, могут быть включены в состав фармацевтических композиций и вводиться в организм млекопитающих, в частности человека, в различных лекарственных формах, приспособленных для выбранного способа введения, т.е. перорально или парентерально, внутривенно, внутримышечно, топически или подкожно.
Таким образом, пептидные ингибиторы могут вводиться системно, например, внутривенно или внутрибрюшинно посредством инфузии или инъекции. Растворы пептидного ингибитора могут готовиться в воде, в которую при необходимости добавляют нетоксичное поверхностно-активное вещество. Дисперсии также могут готовиться в глицерине, жидком полиэтиленгликоле, триацетине, их смесях, а также в маслах. При обычных условиях хранения и использования эти препараты содержат консервант для предотвращения роста микроорганизмов.
Фармацевтические лекарственные формы, пригодные для инъекции, инфузии или топического применения могут включать в себя стерильные водные растворы или дисперсии или стерильные порошки, содержащие активный ингредиент и пригодные для приготовления для немедленного приема стерильных инъекционных или инфузионных растворов или дисперсий, при необходимости заключенных в липосомы. Во всех случаях, дозировочная форма в конечном виде должна оставаться стерильной, текучей и стабильной в условиях производства и хранения. В качестве жидкого носителя или средства доставки может служить растворитель или жидкая дисперсионная среда, содержащая, к примеру, воду, этанол, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и тому подобное), растительные масла, нетоксичные сложные эфиры глицерина и их соответствующие смеси. Требуемая текучесть может быть обеспечена, например, благодаря образованию липосом, путем поддержания требуемого размера частиц в случае дисперсий или благодаря использованию поверхностно-активных веществ. Защита от воздействия микроорганизмов может быть обеспечена посредством использования различных антибактериальных и противогрибковых агентов, таких, например, как парабены, хлорбутанол, фенол, сорбиновая кислота, тимеросал, и тому подобные. В некоторых случаях, специалист в данной области может посчитать нужным включить в состав средства изотонические агенты, такие, например, как сахара, буферы или хлорид натрия. Пролонгированная абсорбция инъекционных препаратов может быть обеспечена за счет включения в состав средства агентов, замедляющих абсорбцию, например моностеарата алюминия и желатина.
Стерильные инъекционные растворы готовятся путем введения пептида или пептидного конъюгата в требуемом количестве в соответствующий растворитель с добавлением при необходимости других вышеперечисленных ингредиентов и последующей стерилизацией путем фильтрования. В случае стерильных порошков для приготовления стерильных инъекционных растворов методы приготовления должны включать в себя вакуумную сушку и сублимационную технику, что позволяет получить порошок, содержащий активный ингредиент вместе с необходимыми дополнительными ингредиентами, присутствующими в предварительно стерилизованных и профильтрованных растворах.
В некоторых случаях пептидные ингибиторы могут также вводиться перорально, в сочетании с фармацевтически допустимым средством доставки, таким как инертный разжижитель или усвояемый съедобный носитель. Они могут быть заключены в твердые или мягкие желатиновые капсулы, спрессованы в таблетки или непосредственно добавляться в пищу пациента. Для перорального терапевтического применения пептидный ингибитор может сочетаться с одним или несколькими наполнителями и использоваться в форме принимаемых внутрь таблеток, сосательных таблеток, пастилок, капсул, эликсиров, суспензий, сиропов, вафель и тому подобного. Подобные составы и препараты должны содержать по меньшей мере 0.1% активного вещества. Процент содержания активного вещества в составах и препаратах может, конечно, варьировать и предпочтительно составляет примерно от 2 до 60% массы данной единичной дозы препарата. Количество активного вещества в таких терапевтически пригодных композициях выбирается таким образом, чтобы получить уровень эффективной дозировки.
Таблетки, пастилки, пилюли, капсулы и тому подобное могут также содержать связующие вещества, такие как трагакантовая камедь, гуммиарабик, кукурузный крахмал или желатин; наполнители, такие как фосфат кальция; разрыхлители, такие как кукурузный крахмал, картофельный крахмал, альгиновая кислота и тому подобное; смазывающие вещества, такие как стеарат магния; и подсластители, такие как сахароза, фруктоза, лактоза или аспартам, а также ароматизаторы, такие как мятное масло, винтергриновое масло или вишневые вкусовые добавки. Если единичной дозой препарата является капсула, она может содержать, помимо материалов вышеописанных типов, жидкие носители, такие как растительное масло или полиэтиленгликоль. Различные другие материалы могут присутствовать в качестве покрытий или оболочки или для иной модификации физической формы твердой единичной дозы препарата. Например, таблетки, пилюли или капсулы могут быть покрыты желатином, воском, шеллаком, сахаром и иными подобными веществами. Сироп или эликсир могут содержать активное вещество, сахарозу или фруктозу в качестве подсластителей, метил и пропилпарабен в качестве консервантов, красители и ароматизаторы, такие как вишневые или апельсиновые ароматизаторы. Разумеется, любой материал, используемый при приготовлении любой единичной дозы препарата, должен быть фармацевтически приемлемым и по существу нетоксичным в используемых количествах. Кроме того, пептидный ингибитор может быть включен в препараты и устройства пролонгированного высвобождения.
Используемые твердые носители включают в себя тонко диспергированные твердые вещества, такие как тальк, глина, микрокристаллическая целлюлоза, кремнезем, глинозем и тому подобное. Используемые жидкие носители включают в себя воду, спирты или гликоли или смеси воды, спирта и гликоля, в которых присутствующие компоненты могут быть растворены или диспергированы в эффективной концентрации, с добавлением, при необходимости, нетоксичных поверхностно-активных веществ. Для улучшения свойств состава при конкретном применении в него могут добавляться вспомогательные вещества, такие как ароматизаторы и дополнительные противомикробные агенты.
Загустители, такие как синтетические полимеры, жирные кислоты, соли и сложные эфиры жирных кислот, жирные спирты, модифицированная целлюлоза или модифицированные минеральные вещества, также могут использоваться вместе с жидкими носителями для получения паст, гелей, мазей, мыл и иных продуктов для непосредственного нанесения на кожу пациента.
Как правило, пептиды согласно настоящему изобретению применяются местно для заживления ран и для стимулирования развития здоровой кожи. Активные пептиды при местном применении могут наноситься на выбранную ткань любым способом, прямым или непрямым, в виде аэрозолей, пены, порошков, кремов, желе, паст, суппозиториев или растворов. Термин паста, используемый в данном документе, включает в себя кремы и другие вязкие составы для намазывания, которые могут наноситься непосредственно на кожу или использоваться для смазки бинтов или перевязочного материала. Пептиды, заявленные в данном изобретении, могут быть ковалентно связаны, прочно адсорбированы или иным образом нанесены на кожное покрытие или перевязочный материал для раны. Для облегчения заживления после хирургической операции активные пептиды, заявленные в данном изобретении, могут наноситься непосредственно на поврежденные ткани или вводиться в имплантируемые протетические приспособления. Эти композиции могут наноситься непосредственно на кожу или рану посредством аэрозоля, такого как пена или туман, вместе с другими агентами.
Составы на основе данных пептидов могут включать в себя эмульсии пептидов в воске, масле, эмульгаторе, воде и/или существенно не растворимом в воде материале, образующем в присутствие воды гель. Такой состав придает эмульсии ряд полезных качеств, при сохранении своей вязкой консистенции она не распадается, если ее подвергнуть обычным процедурам стерилизации, например стерилизации паром, поскольку гель стабилизирует эмульсию. Она также лучше удерживает воду, чем обычный гель, поскольку вода удерживается и эмульсией, и гелем.
Состав может также содержать увлажнитель для снижения парциального давления водяного пара в креме или лосьоне с целью снижения скорости высыхания крема или лосьона. Подходящие для этого увлажнители смешиваются с водой в широком диапазоне концентраций и, как правило, являются пригодными для применения на коже. Для этих целей особенно хорошо подходят полиолы, и примером подходящих полиолов могут служить монопропиленгликоль или глицерин. Полиол может составлять примерно 20-50% (по массе) всего состава; диапазон концентраций также может составлять порядка 30-40%. Такое сравнительно высокое содержание полиола позволяет пасте при любой степени высыхания оставаться мягкой и податливой, поскольку глицерин может выступать в качестве пластификатора для полимера. Таким образом, например, при нанесении пасты на повязку, после того, как паста потеряет воду, ее можно будет легко удалить с кожи без разрезания повязки. Преимущество полиола также заключается в способности подавлять пролиферацию бактерий в пасте при ее контакте с кожей или раной, в особенности инфицированными ранами.
В состав смеси могут входить и другие ингредиенты. К ингредиентам, которые могут использоваться при приготовлении смеси, относятся: оксид цинка, ихтаммол, каламин, сульфадиазин серебра, хлоргексидин ацетат, каменноугольный деготь, хлоргексидин глюконат, салициловая кислота, метронидазол или прочие антибактериальные агенты или их комбинации. Прочие ингредиенты также могут быть включены в состав крема.
Эти ингредиенты могут быть включены в соответствующих концентрациях, например содержание оксида цинка может достигать 15 мас.%; обычно используемая концентрация оксида цинка составляет 6-10%, возможно в сочетании с другими ингредиентами, такими как ихтаммол (0-3 мас.%) и/или каламин (0-15 мас.%). Ихтаммол или каламин также могут применяться отдельно. Хлоргексидин ацетат может использоваться в концентрациях до 1 мас.%; типичной является концентрация 0,5 мас.%.
Одним из примеров восков, используемых для получения эмульсий, является глицерил моностеарат или комбинация глицерил моностеарата и PEG 100 стеарата, коммерчески выпускаемого фирмой CITHROL GMS/AS/NA корпорации Croda Universal Ltd. Эта комбинация содержит как воск, так и эмульгатор (PEG 100 стеарат), особенно хорошо совместимый с воском, для образования водных эмульсий. Для повышения стабильности эмульсии в состав средства может быть включен второй эмульгатор, например, PEG20 стеарат, например CITHROL 1OMS, производимый корпорацией Croda Universal Ltd. Суммарная концентрация эмульгатора в креме обычно составляет порядка 3-15%. При использовании двух эмульгаторов, один из них может присутствовать в более высокой концентрации, чем другой.
Не растворимый в воде материал должен образовывать гель при взаимодействии с водой, входящей в состав. Этот материал должен поэтому быть гидрофильным, но растворяться в воде лишь в незначительной степени. Такой материал может, например, представлять собой полимерный материал, способный абсорбировать воду, но не растворимый в воде. Также могут использоваться неполимерные материалы, способные образовывать гель при взаимодействии с водой и сохраняющие стабильность при повышенных температурах, например глины, такие как каолин или бентонит. Некоторые полимеры, используемые в настоящем изобретении, являются суперабсорбентами, например полимеры, описанные в публикации WO 92/16245, которые содержат гидрофильные производные целлюлозы, частично сшитые с образованием трехмерной структуры. Подходящие сшитые производные целлюлозы включают такие гидроксиалкилцеллюлозы, в которых алкильная группа содержит от 1 до 6 атомов углерода, например гидроксиэтилцеллюлозу или гидроксипропилцеллюлозу, а также карбоксицеллюлозы, например карбоксиметилгидроксиэтилцеллюлозу или карбоксиметилцеллюлозу. Примером частично сшитого полимера является карбоксиметилцеллюлоза натрия, выпускаемая под названием AKUCELL X181 фирмой Akzo Chemicals B.V. Этот полимер является суперабсорбентом, так как он может поглощать воду в количестве, по крайней мере, в десять раз превышающем собственную массу. Сшитая структура этого полимера препятствует его растворению в воде, но вода легко абсорбируется и удерживается внутри трехмерной структуры полимера с образованием геля. Вода медленнее уходит из такого геля, чем из раствора, и это позволяет замедлить или предотвратить высыхание крема. Содержание полимера в составе средства обычно меньше чем 10%, в частности оно находится в диапазоне от около 0,5 до около 5,0 мас.% или от около 1,0% до около 2%.
Полученный состав может быть стерилизован, и его компоненты следует выбирать, варьируя содержание полимера, для того, чтобы обеспечить требуемые реологические свойства конечного продукта. То есть, если продукт требуется стерилизовать, то его состав должен быть выбран таким образом, чтобы придать продукту относительно высокую вязкость/эластичность перед стерилизацией. Если определенные компоненты состава не должны подвергаться стерилизации, состав может быть стерилизован перед добавлением этих компонентов, либо каждый компонент может быть стерилизован отдельно. Этот состав затем может быть приготовлен путем смешивания стерилизованных ингредиентов в стерильных условиях. При стерилизации компонентов по отдельности с последующим их смешиванием содержание полимера следует выбирать таким образом, чтобы обеспечить требуемые реологические свойства конечного продукта. Содержание эмульсии определяет обрабатывающие свойства состава и его восприятие на ощупь, более высокое содержание эмульсии приводит к увеличенной способности распределяться и кремообразности.
Состав может быть упакован в тюбики, баночки или иные подходящие формы контейнеров для хранения, либо распределен по субстрату, который впоследствии упаковывается. Подходящим субстратом являются перевязочные материалы, включая пленочные повязки, и бинты.
Приводимые ниже примеры служат для иллюстрации, но не ограничивают данное изобретение.
ПРИМЕР 1: Пептидные ингибиторы
Общие материалы
Все пептиды были синтезированы фирмой Sigma-Genosys, Inc. Полученные пептиды очищались в компании до степени гомогенности более 95% методом ВЭЖХ. Материал объединенных элюированных пиков обессаливался и лиофилизировался. Методом масс спектроскопии подтверждались молекулярная масса и чистота пептида. Если не указано иное, все химические реактивы приобретались у фирм Sigma Chemical Corp. или Fluka Chemical Co. Активный фермент ММП-9 приобретался у фирмы Calbiochem.
Молекулярное моделирование
При молекулярном моделировании использовались две программы визуализации, Swiss PDB Viewer (Guex и Peitsch, 1997) и Rasmol (Sayle и Milner-White, 1995). Моделирование осуществлялось на персональном компьютере Compaq с операционной системой Windows 95, а также рабочей станцией Silicon Graphics, Inc. Octane UNIX. Кроме того, на станции Octane использовался специальный пакет для молекулярного моделирования Cerius 2, предоставленный Molecular Simulations, Inc. Файлы трехмерной структуры были взяты из банка данных для белков в следующем формате (имя файла, ссылка): ММР-1 (1FBL, Li et al., 1995), MMP-2 (1GEN, Libson et al., 1995), ММП-8 (1JAO, 1JAN, Grams, et al., 1995; Reinemer et al., 1994), MMP-9 (1MMQ, Browner et al., 1995), ТИМП-2/МТ-1 MMP complex (1BUV, Femandez-Catalan et al., 1998), ТИМП-2 (1BR9, Tuuttila et al., 1998) и ТИМП-1/ММР complex (1UEA, Gomis-Ruth et al., 1997; Huang et al., 1996; Becker et al., 1995). Эти файлы использовались для анализа трехмерной структуры белков, а также служили источником данных о первичной последовательности.
Исследование ингибирования
Было проведено два ферментативных анализа. При первом анализе измерялся ферментативный гидролиз флуоресцеинированного коллагена под действием ММП-9 как функция времени. Флуоресцеинированный коллаген (Molecular Probes, Inc.) с концентрацией 5 мкМ добавлялся в реакционный буферный раствор (50 мМ Tris-HCl (рН 7.6), 150 мМ NaCl, 5 мМ CaCl2, 0.1 мМ NaN3) и помещался в кварцевую флюорометрическую кювету Spectrosil™. Матриксная металлопротеиназа с концентрацией 0.1 мкМ смешивалась с различными количествами пептидов в различных концентрациях и инкубировалась при температуре 25°С в течение 10 минут для осуществления связывания. Смесь белков добавлялась к коллагеновому субстрату и быстро перемешивалась. Интенсивность эмиссионной флуоресценции на длине волны 520 нм измерялась как функция времени (длина волны возбуждения 495 нм) на флуорометре Shimadzu RF5301 (Lakowicz, 1983). Анализ высвобождения флуоресцеина использовался для определения константы ингибирования (Ki) пептидного ингибитора ([I]) согласно Сегелю (1993) с использованием метода Диксона (1/v против [I]), по следующей формуле:
где Km - константа Михаэлиса, Vmax - максимальная скорость реакции, и [S] - концентрация субстрата.
При втором анализе использовалась методика флуоресцентного резонансного переноса энергии (ФРПЭ). Субстратный пептид (Calbiochem) из семи аминокислот связывался с карбоксиконцевым динитрофенильным акцептором, и аминоконцевым 2-аминобензо-антранилоильным (Abz) донором. Расщепление этого субстрата под действием ММП-9 приводило к высвобождению флуоресцентного продукта (365 нм возбуждение, 450 нм эмиссия). Пептид с концентрацией 5 мкМ добавлялся в реакционный буферный раствор (50 мМ Tris-HCl (рН 7.6), 150 мМ NaCl, 5 мМ CaCl2, 0.1 мМ NaN3) и помещался в ячейку черной 96-луночной плашки для микротитрования, предварительно блокированную 1% раствором BSA. Матриксная металлопротеиназа с концентрацией 0.1 мкМ смешивалась с пептидом 9-mer (SEQ ID NO: 12), 10-mer (SEQ ID NO: 13) или 19-mer (SEQ ID NO: 11) в различных концентрациях и инкубировалась при температуре 25°С в течение 10 минут для осуществления связывания. Смесь белков добавлялась к флуоресцентному пептидному субстрату и быстро перемешивалась. Интенсивность флуоресценции как функция времени измерялась с помощью устройства Dynex MFX для флуоресцентного считывания плашек для микротитрования. Выявляли зависимость интенсивности флуоресценции от молей расщепленного пептида путем построения стандартной кривой по Abz содержащему не ФРПЭ-пептиду. Константы ингибирования определялись по кривым, как указано выше. Другие матриксные металлопротеиназы исследовались аналогичным образом с использованием специфических субстратных ФРПЭ пептидов (все получены от Calbiochem).
Исследование подавления активации
В этом анализе измеряется степень превращения профермента в зрелую матриксную металлопротеиназу. Проферментная форма ММП-9 (100 мкг) смешивалась с 0.5 мкг стромилизина в растворе PBS. Реакционный раствор инкубировался при температуре 35°С. Аликвоты отбирались из реакционного раствора через каждые 80 минут.
Каждую аликвоту разводили в EDTA до конечной концентрации 1 мМ, вводили в колонку ВЭЖХ BioSelect 125 и хроматографировали в PBS. В нулевой момент времени (инъекция) возникает одиночный пик, который элюируется из колонки приблизительно через 750 секунд. Величина этого пика уменьшается как функция времени, и при этом появляется два новых пика. Первый пик элюируется приблизительно через 800 секунд и соответствует зрелой форме ММП-9. Второй пик элюируется приблизительно через 1100 секунд и соответствует N-концевому фрагменту проферментной формы. Площади пиков определялись путем интегрирования по профилю элюирования, и на графике отмечались значения изменения площади в процентах.
Изотермическая титрационная калориметрия
Изотермическая титрационная калориметрия (ИТК) осуществлялась с помощью ИТК-инструмента производства MicroCal, Inc. Титрование проводилось путем введения 5 мкл раствора ингибиторного пептида (в диапазоне концентраций от 0.5 мМ до 2.0 мМ) в перемешиваемую реакционную ячейку объемом 1.4 мл. Концентрация ММП-9 варьировала в пределах от 50 до 80 мкМ на ячейку. Как ингибитор, так и фермент находились в растворе 20 мМ какодилата натрия (рН 5.5-7.0), 40 мМ NaCl или 20 мМ Tris-HCl (pH 7.0-7.5), 40 мМ NaCl. Титрования проводилось при температуре от 20°С до 40°С. Типичные условия проведения титрования предусматривали 10-секундный период инъекции с последующей 240-секундной задержкой перед следующей инъекцией при общем количестве инъекций, равном 40. Для учета теплоты, выделяющейся при разведении и смешивании, производилось пустое титрование ингибиторного пептида в буферный раствор.
Независимый набор множества участков связывания является наиболее распространенной моделью для экспериментального определения связывания. Формула для аналитического определения общего количества теплоты дается следующим уравнением (Freire et al., 1990):
где Q - общее количество теплоты, V - объем ячейки, ΔН - энтальпия, М - концентрация макромолекул (связываемый партнер в ячейке), n - стехиометрия связывания, L - концентрация лиганда (связываемый партнер в шприце) и К - константа ассоциации. Данные были подобраны для этой модели с помощью пятой версии Origin (MicroCal, Inc.).
Поверхностный плазмонный резонанс
Устройство BiaCore-X для поверхностного плазмонного резонанса (ППР) (BiaCore, Inc.) использовалось для измерения взаимодействия между пептидом (Р) 19-mer (SEQ ID NO: 11) и ММП-9. В этих экспериментах карбоксиметилдекстрановый сенсорный чип (СМ-5, Lofas et al., 1993) активировался 50 мМ N-гидроксисукцинимидом, 0,2 М N-этил-N'-(диметиламинопропил)-карбодиимидом со скоростью потока 10 мкл в минуту в течение десяти минут. ММП-9 при концентрации 75 нг/мкл связывался с активированной поверхностью со скоростью потока 10 мкл в минуту в течение десяти минут. В завершение поверхность инактивировалась пропусканием 1 М этаноламин-HCl со скоростью 10 мкл в минуту в течение пяти минут по поверхности датчика. Пептид 19-mer (SEQ ID NO: 11) в концентрации от 10 до 50 нМ пропускался по поверхности датчика со скоростью 20 мкл в минуту. Фазы ассоциации и диссоциации изотерм связывания сглаживались автоматическим выполнением быстрого преобразования Фурье перед моделированием констант скорости. Изотермы связывания определялись с учетом одновременно прямой (ka) и обратной (ka) константы скорости:
(Karlsson и Fait, 1997) где [Р], [ММР-9] и [Р˜ММР-9] концентрации свободного пептида, свободного ММП-9 и комплекса соответственно. Константа равновесного связывания (КA) при этом определяется следующим образом:
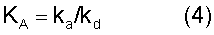
Уравнение 3 в терминах ППР сигнала (Morion et al., 1995) выражается следующим образом:
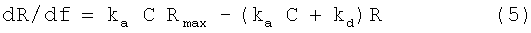
где R представляет собой ППР сигнал (в единицах отклика, ЕО) в момент времени t, Rmax - максимальная способность связывания ММП-9 в ЕО, и С - концентрация хелатного пептида. Кинетический анализ (O'Shannessy et al., 1993) осуществлялся с помощью программы Origin (MicroCal, Inc.).
Определение жизнеспособности
Относительная токсичность пептидов 9-mer (SEQ ID NO: 12), 10-mer (SEQ ID NO: 13) и 19-mer (SEQ ID NO: 11) определялась на модели кожи Epiderm™ (MatTek Corp.). Перед добавлением пептидов отдельные контейнеры с образцами кожи предварительно инкубировались в течение двух часов в культуральной среде при температуре 37°С и 5% содержании СО2. Затем контейнеры с образцами переносились на 6-луночные плашки, содержащие свежую среду. Все пептиды растворялись в PBS до конечной концентрации 10 мМ, и 100 мкл каждого пептидного раствора пипетировали на поверхность контейнера с образцом Epiderm. Инкубация проводилась в течение 12 часов при температуре 37°С и 5% содержании СО2. После завершения периода инкубации контейнеры с образцами три раза промывались раствором PBS и переносились на 24-луночную плашку, содержащую по 300 мкл аналитической среды МТТ на одну ячейку (концентрация МТТ составляла 1 мг/мл). Колориметрический анализ проводился в течение трех часов (инкубация при температуре 37°С и 5% содержании СО2). Контейнеры с образцами затем переносились на 24-луночную культуральную плашку, содержащую 2 мл изопропанола на одну ячейку. Экстракция окрашенного осадка проводилась в течение четырех часов при комнатной температуре. Оптическая плотность измерялась на длине волны 570 нм и 650 нм для каждого образца. Процент жизнеспособности для каждого образца по отношению к PBS контролю рассчитывался следующим образом:
Для каждого пептида проводилось три эксперимента.
Результаты
Последовательность матриксной металлопротеиназы-2 (SEQ ID NO: 14) приведена ниже для облегчения определения различных доменов и областей в матриксных металлопротеиназах.
Строгое попарное выравнивание аминокислотных последовательностей девяти матриксных металлопротеиназ было выполнено с помощью программы CLUSTAL™ (Higgins et al., 1992). Это выравнивание позволило определить положения как консервативных, так и неконсервативных аминокислот, фланкирующих участок активационного расщепления протеиназы. Для сопоставления бралось произвольное количество аминокислот как с амино-, так и с карбоксиконца от участка активационного расщепления. Выравнивание последовательностей матриксных металлопротеиназ (Таблица 1), показанное на Фиг.1, указывает на то, что все участки активации матриксной металлопротеиназы могут быть выравнены статистически значимым образом. Эти области, выбранные для сопоставления примерно соответствуют последовательности аминокислот с 70 по 120, при том, что в среднем сигнальная последовательность матриксной металлопротеиназы включает аминокислоты 1-20, пропептидный домен соответствует аминокислотам 21-100, а зрелый активный фермент включает область со 101 аминокислоты до конца. Последовательность 19-mer (SEQ ID NO: 11), выбранная для изучения, содержится внутри области выравнивания. В частности, в белке ММП-2 пептид 19-mer (SEQ ID NO: 11) соответствует аминокислотам 100-118.
Выравнивание последовательностей матриксной металлопротеиназы указывает на то, что центральная область активационного домена, PRCGVPDV (SEQ ID NO: 1), является высококонсервативной, в то время как последовательности, фланкирующие эту область, обладают более высокой вариабельностью. Гетерогенность последовательности может использоваться для конструирования последовательностей пептидов, способных ингибировать специфические ММП ферменты или комбинации матриксных металлопротеиназ, просто выбирая соответствующие аминокислоты (основываясь на этом выравнивании). Кроме того, длину конкретного пептида можно варьировать с целью модулирования его эффективности.
Трехмерная структура проферментной формы ММП-1 представлена на Фиг.2, на которой видно, что области активации, показанные в таблице 1 и на Фиг.1, образуют мостик, соединяющий два крупных глобулярных домена. Этот вырезаемый участок определяется как короткий неструктурированный домен, соединяющий пропептидный домен с доменом активного фермента. На одном из этапов активации эта последовательность разрезается на две. Это также область, чувствительная к активации, опосредованной хлоридом ртути in vitro.
Активация устраняет стерическое блокирование (осуществляемое пропептидным доменом) и открывает доступ к активному участку зрелого фермента. N-конец располагается вблизи от каталитического иона цинка, который совершенно необходим для обеспечения ферментативной активности. Структура активного фермента ММП-9 показана на Фиг.3, где ионы цинка изображены в виде сплошных шаров. Второй ион цинка является структурным, то есть, он влияет на стабильность белка, но не на катализ. С-концевая половинка пептида 19-mer (SEQ ID NO: 11) теперь располагается на аминоконце фермента, она изображена в левой части на Фиг.3 в виде поднимающейся части последней петли (заштрихована). Моделирование взаимодействия активационного доменного пептида с поверхностью активированной матриксной металлопротеиназы указывает на то, что этот пептид (особенно при включении более длинной N-концевой области) может взаимодействовать с областью активного участка, блокируя при этом доступ субстрата к активному участку. Таким образом, он может выступать в роли небольшого продомена или ферментативного "кэпа".
Известно, что ферменты могут быть расщеплены на фрагменты и эти фрагменты могут воссоединяться с образованием активного фермента. Различные пептидные домены воссоединяются и удерживаются вместе нековалентными межмолекулярными силами. Классическим примером такого пептид-белкового взаимодействия служит взаимодействие между S-пептидом и S-белком рибонуклеазы (Levit и Berger, 1976). S-пептид связывается с S-белком рибонуклеазы в соответствующем положении с образованием комплекса, восстанавливающего ферментативную активность РНКазы-S.
Согласно настоящему изобретению пептиды активационного домена могут повторно связываться с активированной матриксной металлопротеиназой в той же области, где они присутствуют в проферментной форме матриксной металлопротеиназы, образуя неактивный комплекс. Эффективность такого связывания может быть измерено (смотрите ниже). Более того, пептид 19-mer (SEQ ID NO: 11) может связывать цинк благодаря своему цистеиновому остатку, опять-таки препятствуя катализу.
Ингибирование ферментативной активности матриксной металлопротеиназы
Первые исследования ингибирования осуществлялись с пептидом из 19 аминокислот (SEQ ID NO: 11), полученным из области вырезаемого домена ММП-2. Этот пептид был выбран из области CLUSTAL выравнивания, демонстрирующей наибольшую степень консервативности. Последовательность пептида 19-mer (SEQ ID NO: 11) строго консервативна на N-концевом, но обладает высокой степенью вариабельности на С-концевом участке. Два меньших пептида, представляющих собой N-концевую и С-концевую половинки этого пептида, также были исследованы. Эти две половины, грубо говоря, разделяют пептид на консервативную N-концевую часть (9-mer (SEQ ID NO: 12)) и неконсервативную С-концевую часть (10-mer (SEQ ID NO: 13)). Это позволяет исследовать не только общую эффективность ингибирования, но и ее селективность.
Все три пептида во всех флуоресцентных анализах продемонстрировали способность ингибировать фермент ММП-9. Во всех исследованных случаях пептид 19-mer (SEQ ID NO: 11) оказывался более эффективным ингибитором фермента, чем две половинки пептида. Пептид 9-mer (SEQ ID NO: 12) оказывался более эффективным ингибитором, чем С-концевой пептид 10-mer (SEQ ID NO: 13). Эти результаты указывают на то, что цистеин, возможно, необходим в качестве лиганда цинка, или что N-концевые области требуются для осуществления стерического блокирования активного участка фермента. Эта гипотеза может быть проверена в соответствующих экспериментах на ингибиторных пептидах, содержащих более длинную N-концевую последовательность (то есть аминокислоты, расположенные перед остатком 100). Типичная кривая ингибирования при титровании ММП-9 пептидом 19-mer (SEQ ID NO: 11) показана на Фиг.4.
Аналогичные исследования ингибирования, проведенные с пептидами 10-mer (SEQ ID NO: 13) и 9-mer (SEQ ID NO: 12), показаны на Фиг.5 и 6 соответственно. При проведении ФРПЭ анализов все пептиды демонстрировали способность ингибировать фермент ММП-9 с константой ингибирования (Ki) в пределах от 45,2 до 327,7 мкМ (смотрите таблицу 4). Выбор субстрата (ФРПЭ пептид или флуоресцеинированный коллаген) мало влияет на относительную эффективность ингибирования для трех пептидов, во всех случаях наблюдалась следующая устойчивая тенденция: 19-mer (SEQ ID NO: 11)>9-mer (SEQ ID NO: 12)>10-mer (SEQ ID NO: 13). Типичные кривые реакций для титрования ММП-9 соответствующими пептидами показаны на Фиг.7-9.
В целом, полученные константы ингибирования были несколько меньше для коллагеннового субстрата, в пределах от 30,3 до 221,3 мкМ для коллагена и от 45,2 до 327,7 мкМ для ФРПЭ-пептида. Эти данные указывают на то, что пептидные ингибиторы несколько более эффективны при использовании в качестве субстрата коллагена, что предположительно связано с тем, что ингибиторный пептид блокирует активный участок и, поскольку коллаген значительно крупнее чем ФРПЭ-пептид, более эффективно препятствует его доступу к активному участку фермента. Менее крупный ФРПЭ-пептид легче может найти доступ к активному участку, даже в присутствие ингибиторного пептида.
Типичные ферментативные анализы (показаны на Фиг.4-7) обычно проводились в течение 30-40 минут. При увеличении времени эксперимента было показано, что пептид 19-mer (SEQ ID NO: 11) эффективно ингибирует катализируемый ММП-9 гидролиз коллагена более 1000 минут (Фиг.8). Пептид 10-mer (SEQ ID NO: 13) менее эффективно препятствует разрушению коллагена при больших временах (Фиг.9) чем пептид 9-mer (SEQ ID NO: 12) (Фиг.10). При этом пептид 19-mer (SEQ ID NO: 11) опять демонстрирует наибольшую степень ингибирования.
Аналогичные ферментативные анализы проводились с другими ММП ферментами для проверки эффективности пептида 19-mer (SEQ ID NO: 11). В этих анализах использовались ФРПЭ пептиды, содержащие в своей последовательности специфические вырезаемые участки матриксной металлопротеиназы. Пептид 19-mer (SEQ ID NO: 11) способен эффективно ингибировать различные матриксные металлопротеиназы. Эффективность пептида 19-mer (SEQ ID NO: 11) в отношении различных матриксных металлопротеиназ следующая: ММП-2>ММП-3>ММП-8>ММП-7>ММП-9>ММП-1, с константой ингибирования в пределах от 3,1 мкМ (ММП-2) до 41,1 мкМ (ММП-1). Эти данные приведены в таблице 4.
Антисплайсинговая активность пептида 19-mer (SEQ ID NO: 11)
Матриксные металлопротеиназы биологически синтезируются в неактивной проферментной форме. Протеолитическое расщепление профермента, часто посредством отдельного класса мембраносвязанных матриксных металлопротеиназ, приводит к активации матриксной металлопротеиназы. Длина лидерной последовательности профермента составляет приблизительно 100 аминокислот (она несколько варьирует у разных матриксных металлопротеиназ) и находится на аминоконце белка. Ингибирование активации профермента может оказаться эффективным методом снижения ферментативной активности матриксных металлопротеиназ в хронических ранах. Если эти ферменты будут неспособны функционировать, скорость разрушения матрикса будет снижена, что в свою очередь может привести к более быстрому заживлению хронических ран.
Очевидно, что пептиды активационного домена (пептид 19-mer (SEQ ID NO: 11), 9-mer (SEQ ID NO: 12) и 10-mer (SEQ ID NO: 13)) способны ингибировать ферментативную активность различных матриксных металлопротеиназ. Помимо этой активности, пептид 19-mer (SEQ ID NO: 11) препятствует активации неактивной проферментной формы ММП-9. Таким образом, пептид 19-mer (SEQ ID NO: 11) может снижать общий уровень активности матриксных металлопротеиназ в коже и внутри экссудата хронических ран путем ингибирования уже имеющихся активированных матриксных металлопротеиназ или препятствуя активации новосинтезированных проферментных форм матриксных металлопротеиназ.
На Фиг.11 приведены результаты типичного анализа сплайсинга. Первый пик, элюируемый приблизительно через 700 секунд, соответствует проферментной форме ММП-9. По мере протекания реакции сплайсинга интенсивность этого пика уменьшается (как указано стрелкой вниз) и появляется два новых пика. Первый новый пик, элюируемый приблизительно через 800 секунд, соответствует зрелой и активной ММП-9. Второй новый пик, элюируемый приблизительно через 1050 секунд, соответствует продомену. По мере протекания реакции сплайсинга интенсивность этих двух пиков увеличивается (как указано стрелками вверх). После завершения реакции проферментная форма ММП-9 перестает регистрироваться. Титрование стандартной реакции сплайсинга пептидом 19-mer (SEQ ID NO: 11) предотвращает превращение проферментной формы ММП-9 в продомен и активный фермент. На Фиг.12 показаны результаты этого титрования. Сплайсинг может ингибироваться пептидом 19-mer (SEQ ID NO: 11) в микромолярных концентрациях, степень ингибирования зависит от дозы.
Изотермическая титрационная калориметрия
Калориметрический анализ использовался для определения, образует ли или нет пептид 19-mer (SEQ ID NO: 11) стабильный нековалентный комплекс с активной ММП-9. Эти данные позволяют понять механизм ферментного ингибирования и антиактивационных свойств пептида 19-mer (SEQ ID NO: 11). На Фиг.13 показаны данные изотермического калориметрического эксперимента по определению взаимодействия между пептидом 19-mer (SEQ ID NO: 11) ингибитором матриксной металлопротеиназы и ферментом ММП-9. Пептид растворялся в 20 мМ какодилате (рН 6.8), 20 мМ NaCl до конечной концентрации 1 мМ. ММП-9 разводился в том же буферном растворе до конечной концентрации 20 мкМ. Последовательность стандартных инъекций выполнялась так, как описано выше. Результаты определения взаимодействия между ферментом ММП-9 и пептидом 19-mer (SEQ ID NO: 11) приведены ниже:
Стехиометрия: 0,975±0,02
ΔН (kcal/mol): -26,1±1,45
ΔS (cal mol-1K-1): -11,6±2,2
KA (M-1): 1,65×106±4,5×104
Эти результаты указывают на то, что взаимодействие между пептидом 19-mer (SEQ ID NO: 11) и ферментом ММП-9 определяется энтальпийным фактором, то есть ΔН отрицательно. Данная реакция энтропийно невыгодна, о чем свидетельствует отрицательное значение ΔS. Однако энтальпийный член превышает член TΔS, следовательно, общая свободная энергия (ΔG) отрицательна.
Реакция пептида 19-mer (SEQ ID NO: 11) с ферментом ММП-2 наблюдалась и оказалась определяемой энтальпийным фактором и энтропийно невыгодной. Результаты изотермического калориметрического анализа, представленные на Фиг.14, были получены путем титрования пептида 19-mer (SEQ ID NO: 11) ферментом ММП-2. В этих экспериментах были получены следующие значения.
Стехиометрия: 0,99±0,03
ΔН (kcal/mol): -15,4±2,05
ΔS (cal mol-1 K-1): -21,1±1,8
KA (M-1): 2,40×106±3,7×104
Таким образом, реакции связывания энтропийно невыгодны. Это предположительно связано с потерей конфигурационной энтропии в результате связывания. Следует учесть, что полностью гибкий пептид обладает большим числом степеней свободы. Во всех случаях связывания стехиометрия взаимодействия пептида с ММП составляет 1:1, что указывает на то, что одна молекула пептида 19-mer (SEQ ID NO: 11) взаимодействует с одной молекулой матриксной металлопротеиназы.
Поверхностный плазмонный резонанс
Связывание пептида 19-mer (SEQ ID NO: 11) с ферментом ММП-9 кинетически изучалось с использованием метода поверхностного плазмонного резонанса (ППР). Сенсорный чип изготовлялся путем связывания активного фермента ММП-9 с поверхностью чипа СМ-5 (BIACore, Inc.) с использованием стандартных химических реакций, рекомендованных изготовителем. Раствор пептида 19-mer (SEQ ID NO: 11) пропускали по поверхности ММП-9 в приборе BIACore-X™, и связывание и диссоциация регистрировались в реальном масштабе времени. Типичная изотерма связывания показана на Фиг.15. Фаза ассоциации (30-430 секунд) лучше всего соответствовала модели одиночного участка связывания и приводила к константе скорости ассоциации (ka) 2.2×104 М-1с-1. Фаза диссоциации (440-700 секунд) также соответствовала модели одиночного участка связывания и приводила к константе скорости диссоциации (kd) 4.1×10-3 s-1. Рассчитанная константа равновесного связывания (КA=ka/kd) 5.3×106 хорошо согласуется с термодинамическими данными. В эксперименте наблюдался массовый транспортный эффект порядка 100 единиц отклика в начале фазы диссоциации, что не было отражено в модели. Таким образом, связывание пептида 19-mer (SEQ ID NO: 11) с ММП-9 является кинетически и термодинамически выгодным.
Определение жизнеспособности
В отличие от многих низкомолекулярных ингибиторов матриксных металлопротеиназ, три выбранных для данного исследования пептида не являются токсичными для клеток в экспериментах на модели кожи EpiDerm™. На Фиг.16 видно, что пептид, примененный в двух концентрациях (500 мкМ и 2 мМ) приводит только к незначительному снижению жизнеспособности по сравнению с PBS контролем. Суммарная средняя жизнеспособность клеток после применения пептидов составляла 97,6% (для пептида 19-mer (SEQ ID NO: 11)), 89,6% (для пептида 10-mer (SEQ ID NO: 13)) и 95,8% (для пептида 9-mer (SEQ ID NO: 12)). Эти результаты указывают на то, что данные пептиды, используемые для заживления хронических ран, не являются токсичными для клеток млекопитающих. Данные, приведенные на Фиг.16, дают среднее значение для трех образцов. Стандартное отклонение для жизнеспособности составляло от 2,2 до 3,7 в данном исследовании и не обнаруживало корреляции с дозой или конкретным видом используемого пептида. Жизнеспособность несколько снижалась при повышении концентрации пептида.
Эти результаты указывают на то, что данные пептиды не являются токсичными в экспериментах на модели кожи EpiDerm™, что кинетические и энтропийные факторы благоприятствуют образованию связанных комплексов между ними и матриксными металлопротеиназами, и что они способны ингибировать ферментативную активность и предотвращать активацию матриксных металлопротеиназ.
ПРИМЕР 2: Использование пептидных ингибиторов для заживления ран
Методы
Раны наносились мышам C57BL6/KsJ db/db с помощью 4 мм биопсийного перфоратора. Мыши были получены из лаборатории Jackson Laboratories, возраст животных на момент начала эксперимента составлял 3-7 месяцев. Все мыши перед нанесением раны подвергались анестезии. Две ранки наносились в верхней части спины каждого животного путем снятия кожи с нижележащих структур и проталкивания перфоратора через изолированную кожу. Типичная ранка имела среднюю глубину 1,7 мм, и размер от 1,3 до 2,2 мм. При нанесении раны мышечная ткань не затрагивалась. Непосредственно после нанесения раны обрабатывались либо физиологическим раствором (животные контрольной группы), либо 5 мкл пептида 19-mer (SEQ ID NO: 11) в концентрации 20 мкг/мл.
Каждый день участок раны фотографировался цифровым аппаратом, и площадь раны определялась путем компьютерного интегрирования фотографий. Все процедуры обработки ран и последующего анализа данных осуществлялись слепым методом (см., например, Brown et al., 1994). Площадь раны в момент ее нанесения (нулевой день) для всех ран произвольно принималась равной 1 в относительных единицах; так что в дальнейшем площади раны преобразовывались в относительные площади ран путем деления абсолютного значения площади раны в день n на площадь раны в нулевой день.
Результаты:
Как можно видеть на Фиг.17, однократное применение пептида 19-mer (в момент нанесения раны, нулевой день) значительно ускоряет процесс полного затягивания раны на тестируемой модели диабетических мышей. В среднем раны, обработанные пептидом 19-mer, затягивались на девятый день после нанесения по сравнению с 14 днями у контрольных животных, подвергавшихся обработке солевым раствором. Кроме того, при обработке раны пептидом 19-mer (SEQ ID NO: 11) наблюдалось уменьшение воспаления поврежденной области в первый день после нанесения раны. Также следует отметить, что при обработке раны пептидом 19-mer (SEQ ID NO: 11) процесс затягивания начинался раньше, чем при обработке раны солевым раствором (5 дней против 8 дней).
ПРИМЕР 3: Стимуляция роста фибробластов с помощью пептидных ингибиторов
В этом примере приводятся данные, показывающие, что пептиды согласно настоящему изобретению стимулируют пролиферацию фибробластов.
Материалы и методы
На клеточной линии человеческих кожных фибробластов (Clonetics, Walkersville, MD, нормальные человеческие неонатальные кожные фибробласты, номер в каталоге СС-2509) исследовалась способность заявленного в данном изобретении пептида стимулировать клеточную пролиферацию. Интенсивность пролиферативного ответа человеческих кожных фибробластов на пептид 19-mer (SEQ ID NO: 11) измерялась в 96-луночной системе анализа с использованием не содержащей сыворотки среды в качестве контроля. Маточный раствор, содержащий 0,5 г/л пептида 19-mer (SEQ ID NO: 11), готовился в воде и затем разводился не содержащей сыворотки средой Игл, модифицированной Дульбекко (DMEM, Sigma Chemical Co., St. Louis, МО) до получения растворов, содержащих данный пептид в концентрации 1×10-4 М, 1×10-5 М и 1×10-6 М. Клетки высевались на 96-луночные плашки при концентрации 1×103 клеток в 100 мкл среды DMEM, содержащей 10% эмбриональной бычьей сыворотки (FBS, Sigma Chemical Co., St. Louis, МО). Плашки инкубировались в течение 24 часов при температуре 37°С в увлажненной атмосфере с 5% содержанием СО2. После инкубации среду отсасывали, и ячейки дважды промывали 100 мкл не содержащей сыворотки среды DMEM. Затем среду отсасывали и по 100 мкл раствора, содержащего 1×10-4 М, 1×10-5 М или 1×10-6 М пептида 19-mer (SEQ ID NO: 11), добавляли в ячейки (по 20 ячеек для каждой концентрации). Кроме того, 100 мкл носителя (не содержащей сыворотки среды DMEM) добавляли в 10 ячеек в качестве контроля. Все ячейки инкубировались в течение 28 часов при температуре 37°С в увлажненной атмосфере с 5% содержанием СО2. После инкубации во все ячейки добавляли по 20 мкл раствора Cell Titer 96 Aqueous One Solution. Плашки осторожно перемешивали и помещали обратно в инкубатор на 45 минут, и спектрофотометрическая абсорбция измерялась для каждой ячейки при 490 нм. Результаты анализировались статистически методом одностороннего дисперсионного анализа.
Результаты:
Как можно видеть на Фиг.18, добавление пептида 19-mer (SEQ ID NO: 11) приводит к усилению роста фибробластов в зависимости от применяемой дозы. В контрольном опыте, без добавления пептида 19-mer, клетки имели наименьшую плотность. Клетки, получавшие лишь 1×10-5 М пептида 19-mer (SEQ ID NO: 11, обозначенные "19mer5" на Фиг.18), имели значительно большую клеточную плотность (Р>0.01), чем клетки, не получавшие пептида 19-mer. Клетки, получавшие 1×10-4 М пептида 19-mer (обозначенные "19mer4" на Фиг.18), демонстрировали еще более интенсивный клеточный рост (Р>0.001). Однако клетки, получавшие 1×10-6 М пептида 19-mer (обозначенные "19mer6" на Фиг.18), демонстрировали достаточно слабую клеточную пролиферацию (Р>0.05), что было признано статистически незначимым.
Таким образом, между контрольными клетками и клетками фибробластов, обработанных пептидом 19-mer (SEQ ID NO: 11), наблюдается статистически значимое различие в интенсивности клеточного роста. На основании этих результатов пептид 19-mer (SEQ ID NO: 11) представляется хорошим пролиферационным агентом для фибробластов.
ПРИМЕР 4: Стимуляция роста кератиноцитов пептидными ингибиторами
В этом примере приводятся данные, показывающие, что пептиды настоящего изобретения стимулируют пролиферацию кератиноцитов.
Материалы и методы
На клеточную линию человеческих кожных кератиноцитов (Clonetics Walkersville, MD, нормальные человеческие неонатальные эпидермальные кератиноциты, номер в каталоге сс-2503) воздействовали пептидом 19-mer (SEQ ID NO: 11), чтобы выяснить, может ли этот пептид стимулировать пролиферацию кератиноцитов. Пролиферативный ответ этих человеческих кожных кератиноцитов на пептид 19-mer (SEQ ID NO: 11) измерялся на 96-луночной системе анализа с использованием основной питательной среды для кератиноцитов (КВМ, Clonetics, номер в каталоге СС-3103) в качестве контроля. Маточный раствор, содержащий 0,5 г/л пептида 19-mer, готовился в воде и затем разводился средой КВМ до получения растворов, содержащих данный пептид в концентрации 1×10-4 М, 1×10-5 М и 1×10-6 М. Клетки высевались на 96-луночную плашку при концентрации 2.5×103 клеток на 100 мкл среды КВМ. Плашки инкубировались в течение 24 часов при температуре 37°С в увлажненной атмосфере с 5% содержанием CO2. После инкубации по 100 мкл раствора, содержащего 1×10-4 М, 1×10-5 М или 1×10-6 М пептида 19-mer (SEQ ID NO: 11), добавляли в ячейки (по 10 ячеек для каждой концентрации). Кроме того, 100 мкл носителя КВМ добавляли в 10 ячеек в качестве контроля. Плашку инкубировали в течение 48 часов при температуре 37°С в увлажненной атмосфере с 5% содержанием CO2. После инкубации во все ячейки добавляли по 20 мкл раствора Cell Titer 96 Aqueous One Solution. Плашку осторожно перемешивали и помещали обратно в инкубатор на 3 часа. Спектрофотометрическая абсорбция измерялась для каждой ячейки при 490 нм. Результаты анализировались статистически методом одностороннего дисперсионного анализа.
Результаты
Как можно видеть на Фиг.19, добавление пептида 19-mer (SEQ ID NO: 11) приводит к усилению роста кератиноцитов в зависимости от применяемой дозы. Контрольные клетки без добавления пептида 19-mer имели наименьшую клеточную плотность. Клетки, получавшие лишь 1×10-5 М пептида 19-mer (SEQ ID NO: 11, обозначенные "19mer5" на Фиг.19), имели значительно большую клеточную плотность (Р>0.01), чем клетки, не получавшие пептида 19-mer. Клетки, получавшие 1×10-4 М пептида 19-mer (обозначенные "19mer4" на Фиг.19), демонстрировали еще более интенсивный клеточный рост (Р>0.001). Однако, клетки, получавшие 1×10-6 М пептида 19-mer (обозначенные "19mer6" на Фиг.19), демонстрировали достаточно слабую клеточную пролиферацию (Р>0,05), что было признано статистически незначимым.
Таким образом, между контрольными клетками и клетками кератиноцитов, обработанных пептидом 19-mer (SEQ ID NO: 11), наблюдается статистически значимое различие в интенсивности клеточного роста. Следовательно, пептид 19-mer (SEQ ID NO: 11) является, по-видимому, хорошим пролиферационным агентом для кератиноцитов.
ПРИМЕР 5: Стимуляция миграции фибробластов с помощью пептидных ингибиторов
В этом примере приводятся данные, свидетельствующие, что пептиды, заявленные в настоящем изобретении, могут стимулировать миграцию фибробластов.
Материалы и методы
Нормальные человеческие кожные фибробласты (НЧКФ, Biowhittaker, Walkersville, MD) высевались во флаконы Т75 со средой FBM (500 мл, Biowhittaker) содержащей инсулин, hFGF-b, GA-1000 и эмбриональную бычью сыворотку (10 мл) и пересевались до 12 раз. Для анализа миграции клетки НЧКФ промывали один раз 10 мл солевого буферного раствора Hank (HBSS). Затем для снятия клеток НЧКФ с флакона во флакон Т75 добавляли не более чем на пять минут три миллилитра трипсина (0,25%) в EDTA. Клетки НЧКФ в растворе трипсина добавляли к 7 мл среды FBM, не содержащей дополнительных добавок. Затем клетки НЧКФ центрифугировали в течение 5 минут и удаляли надосадочную жидкость. Клетки НЧКФ ресуспендировали в 10 мл среды FBM для подсчета. Затем клетки НЧКФ центрифугировали еще раз в течение 5 минут и удаляли надосадочную жидкость Клетки НЧКФ затем ресуспендировали в концентрации 1×106 клеток на мл в среде FBM, не содержащей дополнительных добавок. При измерении миграции важно использовать именно среду FBM, поскольку полная среда содержит факторы роста фибробластов и сыворотку, которые способны активизировать миграцию фибробластов.
Синтез и очистка пептидов производились на микрохимическом оборудовании в университете Эмори (Emory) или компанией SigmaGenesis. После хроматографирования методом ВЭЖХ все пептиды подвергались масс-спектрометрическому анализу для определения их чистоты. Приблизительно 0,5-2,0 миллиграммов пептида использовалось для приготовления свежих маточных растворов (5 мг/мл в PBS) для каждого опыта по измерению миграционной способности. Маточные растворы пептида в PBS использовались для приготовления более разбавленных растворов (1 мг/мл, 100 мкг/мл, 10 мкг/мл, 1 мкг/мл, 100 нг/мл, 10 нг/мл) в среде FBM (без дополнительных добавок) для измерения миграции. Мембраны из не содержащего поливинилпирролидона поликарбоната с размером пор 8 мкм (Neuroprobe, Inc., Gaithersburg, MD) промывались 90% этанолом в течение 15 минут и затем четыре раза деионизированной водой. Мембраны затем помещали в стеклянную тарелку, содержащую водный раствор желатина с концентрацией 5 мкг/мл. Стеклянную тарелку помещали в водяную баню при температуре около 90°С на один час. Мембраны извлекали из раствора желатина и высушивали в течение одного часа в инкубаторе при температуре 37°С.
Для анализа миграции использовалась 48-луночная хемотаксическая камера (Neuroprobe, Inc., Gaithersburg, MD) (см. Фиг.20). По двадцать восемь микролитров каждого исследуемого раствора добавляли в нижние ячейки камеры (содержащей четыре ячейки). Обработанная желатином мембрана аккуратно помещалась сверху на нижние ячейки. Прокладку и верхнюю часть камеры затем аккуратно помещали на верхнюю часть мембраны. Аппарат закреплялся с помощью шести барашковых винтов. Раствор НЧКФ, содержащий заданную плотность клеток (2×105 клеток на мл), готовился путем добавления 320 мкл раствора с концентрацией 1×106 клеток на мл к 1,28 мл среды FBM (без дополнительных добавок). Пятьдесят микролитров раствора НЧКФ с концентрацией 2×105 клеток на мл добавляли в верхнюю часть каждой ячейки камеры. Камеру затем помещали в инкубатор при температуре 37°С и 5% содержании CO2 на три часа. После этого камеру извлекали, откручивали барашковые винты и ставили камеру в перевернутом положении на бумажное полотенце. Нижнюю часть камеры снимали, и мембрана, через которую мигрировали клетки НЧКФ, оказывалась сверху. Мембрану аккуратно снимали. Клетки, не мигрировавшие через мембрану, соскребали с мембраны с помощью клеточного скребка, смочив одну сторону мембраны раствором PBS (Neuroprobe, Inc.). Мембрану фиксировали в метаноле и окрашивали красящими растворами Diff-Quik I и II. Клеточные ядра НЧКФ окрашивались в фиолетовый цвет. Мембрану помещали на предметное стекло, погрузив в кедровое масло, и закрывали покровным стеклом. Клетки в трех отдельных полях, соответствующих различным камерам, подсчитывались с помощью оптического микроскопа Цейса (25x объектив × 10х окуляр × 1.25х). Среднее количество мигрировавших клеток НЧКФ в отрицательном контроле вычитали из значения, полученного для экспериментального раствора (пептид), и данных положительного контроля. Эти результаты затем выражались в проценте мигрировавших клеток НЧКФ по отношению к данным положительного контроля (плазматический фибронектин 1,25 мкг/мл), согласно следующей формуле.

Результаты
Мембрана, используемая для определения степени миграции фибробластов (НЧКФ), имела 8 мкм поры, показана на Фиг.21А (вверху). Раствор хемоаттрактанта инициирует миграцию фибробластов через мембрану. При миграции фибробластов через мембрану они прикрепляются к мембране со стороны хемоаттрактанта. Как показано на Фиг.21В (внизу), ядра фибробластов окрашены в фиолетовый цвет для визуального подсчета. Клетки, застрявшие внутри поры (пора выглядит темно-фиолетовой), включались в общее количество посчитанных клеток. При измерении миграции НЧКФ в положительном контроле количество клеток в поле при подсчете составляло от 27 до 60. В качестве положительного контроля при анализе миграции НЧКФ использовался раствор плазматического фибронектина (1,25 мкг/мл в среде FBM). Фибронектин представляет собой молекулу, способствующую образованию внеклеточного матрикса во время заживления раны. Фибронектин, связывающийся с рецептором фибробластов α4β1 интегрином, является хемотаксическим в очень узком диапазоне концентраций (0,8-1,6 мкг/мл). Postlethwaite, A.E.; Kang, A.H. "Fibroblast Chemoattractants" in Methods of Enzymology, vol.163, Academic Press: New York, 1988, 694-707. В отрицательном контроле (среда FBM без добавок) количество клеток в поле при подсчете составляло от 1 до 7 НЧКФ (менее 10% от результатов положительного контроля) (см. Фиг.22).
В анализе миграции НЧКФ использовалось несколько концентраций пептида 19-mer. В связи с узостью диапазона концентраций, в котором плазматический фибронектин является хемотаксическим для НЧКФ, использовались десятикратные разведения пептида для исследования широкого диапазона концентраций. Как показано на Фиг.23, при концентрациях пептида 19-mer свыше 0,1 мг/мл значительное количество клеток НЧКФ мигрировали через мембрану (55%±3% и 46%±3% для 1000 мкг/мл и 100 мкг/мл соответственно). При концентрации пептида 19-mer ниже 100 мкг/мл он обладает лишь незначительной хемотаксичностью. Поскольку такие концентрации пептида 19-mer индуцировали миграцию только 20% НЧКФ, что лишь примерно в два раза больше, чем для отрицательного контроля, эти данные были признаны незначимыми. Никакого различия в данных по миграции под действием пептидов 19-mer, полученных из двух различных коммерческих источника, обнаружено не было (данные не показаны).
Для определения влияют ли изменения аминокислотной последовательности на миграцию фибробластов были синтезированы и очищены различные вариации последовательности 19-mer. Были исследованы следующие пептиды: 9-mer PRCGNPDVA (SEQ ID NO: 12), 10-mer NYNFFPKKPK (SEQ ID NO: 13), 14-mer TMRKPRCGNPDVAN (SEQ ID NO: 19) и 17-mer TLKAMRKPRCGNPDVAN (SEQ ID NO: 20). Последовательности пептидов 14-mer и 17-mer представляют собой последовательности, взятые соответственно у ММП-2 и ММП-9 из области, смещенной ближе к N-концу. Пептиды 9-mer (SEQ ID NO: 12) и 10-mer (SEQ ID NO: 13) ингибируют активность матриксных металлопротеиназ. Пептид 19-mer (SEQ ID NO: 11) также синтезировался с защитными ацетильными и амидными группами (Ас-19-mer) на N- и С-концах соответственно, чтобы определить, может ли добавление этих групп влиять на интенсивность миграции фибробластов. Ас-19-mer являлся хемотаксическим для фибробластов как при концентрации 100 (44±3%), так и 1000 (40±6%) мкг/мл, как показано на Фиг.23. Пептид 10-mer (SEQ ID NO: 13) являлся хемотаксическим при концентрации 1000 мкг/мл (30±2%), но не при концентрации 100 мкг/мл (11±0%, того же порядка, что и отрицательный контроль, как показано на Фиг.22). Пептид 9-mer (SEQ ID NO: 12) являлся хемотаксическим при концентрации 1000 мкг/мл (32±2%), но не при концентрации 100 мкг/мл (16±0%). Интересно, что добавление аминокислот на N-конце пептида 9-mer приводило к отсутствию миграции НЧКФ. Пептиды 14-mer TMRKPRCGNPDVAN (SEQ ID NO: 19) и 17-mer TLKAMRKPRCGNPDVAN (SEQ ID NO: 20) не вызывают миграцию НЧКФ при концентрациях в пределах от 1000 мкг/мл до 10 нг/мл (данные не показаны). Из-за широкой вариации в степени миграции НЧКФ для различных пептидов аминокислотная последовательность может быть существенна для стимулирования клеток НЧКФ к миграции.
Миграция фибробластов к пораженному участку является важным процессом для правильного заживления раны. В эксперименте по измерению миграции НЧКФ были продемонстрированы хемотаксические свойства пептида 19-mer (>100 мкг/мл) и его производных Ac-19-mer, 9-mer и 10-mer. Поскольку пептиды 9-mer и 10-mer индуцируют миграцию НЧКФ примерно в одинаковой степени, трудно установить, насколько отдельные аминокислоты важны для активации. Интересно, что пептиды 14-mer TMRKPRCGNPDVAN (SEQ ID NO: 19) и 17-mer TLKAMRKPRCGNPDVAN (SEQ ID NO: 20) не оказывают влияния на подвижность фибробластов. Несмотря на то, что точный механизм хемоаттракции пептида 19-mer неизвестен, установлено, что аминокислотная последовательность пептида существенна для активации фибробластов.
ПРИМЕР 6: Пептидные ингибиторы не стимулируют миграцию нейтрофилов
В этом примере приводятся данные, свидетельствующие, что пептид, заявленный в данном изобретении, не стимулирует миграцию нейтрофилов.
Материалы и методы
Синтез и очистка пептидов производились на микрохимическом оборудовании в университете Эмори или компанией SigmaGenesis. После хроматографирования методом ВЭЖХ все пептиды подвергались масс-спектрометрическому анализу для определения их чистоты. Приблизительно 0,5-2,0 миллиграммов полученного пептида использовалось для приготовления свежих маточных растворов (5 мг/мл в HBSS/HSA) для каждого анализа миграции. Маточные растворы пептида использовались для приготовления более разбавленных растворов (1 мг/мл, 100 мкг/мл, 10 мкг/мл, 1 мкг/мл, 100 нг/мл и 10 нг/мл) в HBSS/HSA для анализов.
Нейтрофилы выделялись из человеческой крови (собранной в пробирки Vacutainer™, содержащей EDTA) путем нанесения смеси кровь/EDTA (примерно 20 мл) поверх двух слоев Histopaque™ 1119 (нижний слой, 18 мл) и 1077 (верхний слой, 7 мл) в 50 мл пробирках Falcon. Пробирки центрифугировали на скорости 1800 об/мин в течение 30 минут (без остановки). После центрифугирования в пробирках наблюдалось четыре слоя (сверху вниз): плазма, 1077, 1119 и красные кровяные клетки. Промежуточный слой между слоями плазмы и 1077 содержал лимфоциты, моноциты и некоторое количество тромбоцитов. Слой плазмы, промежуточный слой и большая часть слоя 1077 отбирались и отбрасывались. Слой 1119, содержащий нейтрофилы, отбирался и помещался в чистую 50 мл пробирку. Добавлением солевого фосфатного буферного раствора Дульбекко (DPBS) объем доводили до 50 мл. Клетки центрифугировали в течение 15 минут на скорости 2100 об/мин. Затем клетки ресуспендировали в 10 мл раствора DPBS, переносили в 15 мл пробирки и центрифугировали снова в течение 10 минут на скорости 2000 об/мин. Этот процесс повторялся для вторичного промывания клеток. После удаления надосадочной жидкости красные кровяные клетки лизировали добавлением 6 мл холодной стерильной воды к каждой 15 мл порции надосадочной жидкости. Клетки смешивались с водой только в течение 30 секунд. Затем добавляли 6 мл холодного 2% стерильного солевого раствора. Клетки центрифугировали снова в течение 10 минут на скорости 2000 об/мин. Красная надосадочная жидкость (содержащая гемоглобин из красных кровяных клеток) удалялась. Этот процесс лизирования красных кровяных клеток повторялся еще два раза для удаления всех красных кровяных клеток. После удаления большей части красных кровяных клеток нейтрофилы ресуспендировали в 10 мл раствора HBSS/HSA для подсчета с трипановым синим. Клетки центрифугировали и ресуспендировали в HBSS/HSA при концентрации 1,25×106 клеток/мл.
Анализ миграции нейтрофилов осуществляли в двух вариантах на 24-луночной плашке с разделенными ячейками (размер пор в разделяющей мембране 3 мкм). Разделяющие мембраны предварительно смачивались 20-40 мкл раствора HBSS/HSA (0.04-0.4%). В нижнюю часть каждой ячейки добавляли хемотаксические растворы (500 мкл). Нейтрофилы (200 мкл раствора 1,25×106 клеток/мл) добавляли поверх мембраны в каждую ячейку. Ячейки затем опускали в хемотаксические растворы. Плашку на один час помещали в инкубатор при температуре 37°С и 5% содержании CO2). Количество клеток, мигрировавших через мембрану, подсчитывалось путем окраски трипановым синим.
Результаты
В экспериментах по миграции нейтрофилов при выделении количество жизнеспособных нейтрофилов составляло приблизительно 95%. Для данного эксперимента нейтрофилы ресуспендировали в растворе HBSS/0,4% HSA. В качестве положительного контроля для исследования миграции нейтрофилов использовался IL-8 (10 нМ в буферном растворе HBSS/0.4% HSA). В эксперименте использовались следующие концентрации пептида 19-mer 1 мг/мл, 100 мкг/мл, 10 мкг/мл и 1 мкг/мл.
Во второй колонке в таблице 5 приведены результаты первого эксперимента. В качестве отрицательного и положительного контроля использовались соответственно HBSS/HSA и IL-8.
Пептид 19-mer (SEQ ID NO: 11) индуцировал миграцию нейтрофилов примерно в той же степени, что и отрицательный контроль. Добавление IL-8 вызывало 81% увеличение миграции нейтрофилов через мембрану. В то время как результаты положительного контроля были вполне удовлетворительными, результаты отрицательного контроля (29%) представлялись слишком высокими. Поэтому во втором эксперименте количество HSA было снижено десятикратно до 0,04%. Как видно из третьей колонки таблицы 5, во втором эксперименте в отрицательном контроле количество мигрировавших нейтрофилов резко упало до 0%. Количество мигрирующих нейтрофилов в положительном контроле IL-8 также уменьшилось, но все еще оставалось достаточно большим (51%). Как и в первом эксперименте, пептид 19-mer не оказывал влияния на миграцию нейтрофилов.
Миграция фибробластов к пораженному участку является важным процессом для правильного заживления раны. В эксперименте по измерению степени миграции НЧКФ были продемонстрированы хемотаксические свойства пептида 19-mer (SEQ ID NO: 11). Однако пептид 19-mer не оказывал влияния на миграцию нейтрофилов. По-видимому, у фибробластов имеется рецептор, отсутствующий у нейтрофилов, который обусловливает узнавание пептида 19-mer, его связывание и хемотаксическую сигнализацию.
ПРИМЕР 7: Стимуляция продукции коллагена
Стимуляция синтеза коллагена в ответ на добавление пептида 19-mer (SEQ ID NO: 11) исследовалась на клеточной линии человеческих кожных фибробластов (Clonetics, Walkersville, MD, нормальные человеческие неонатальные кожные фибробласты, номер в каталоге СС-2509) с использованием аналитического набора Takara Biomedicals EIA (ТАК МК101), приобретенного у Panvera (Madison, WI). Клетки сначала выращивались на 96-луночных плашках в среде Игл, модифицированной Дульбекко (DMEM), с добавлением 10% эмбриональной бычьей сыворотки (FBS), при этом среда и сыворотка приобретались у компании Sigma Chemical Co, St. Louis, МО. В качестве контроля использовалась не содержащая сыворотки среда DMEM. Маточный раствор, содержащий 0,5 г/л пептида 19-mer (SEQ ID NO: 11), готовился в воде и затем разводился не содержащей сыворотки средой DMEM до получения растворов, содержащих данный пептид в концентрации 1×10-5 М и 1×10-6 М. Клетки высевались на 96-луночные плашки при концентрации 5×103 клеток в 100 мкл среды DMEM, содержащей 10% эмбриональной бычьей сыворотки (FBS, Sigma Chemical Co., St. Louis, МО). Плашки инкубировались в течение 24 часов при температуре 37°С в увлажненной атмосфере с 5% содержанием СО2. После инкубации среду отсасывали и ячейки дважды промывали 100 мкл не содержащий сыворотки среды DMEM. Затем среду отсасывали и по 100 мкл раствора, содержащего 1×10-5 М или 1×10-6 М пептида 19-mer (SEQ ID NO: 11), добавляли в ячейки (n=2 для каждой концентрации). Кроме того, 100 мкл носителя (не содержащей сыворотки среды DMEM) добавляли в 4 ячейки в качестве контроля. Все ячейки инкубировались в течение 48 часов при температуре 37°С в увлажненной атмосфере с 5% содержанием CO2.
Для анализа отбирали по 20 мкл надосадочной жидкости из каждой ячейки 96-луночной плашки. Стандартный буферный раствор и останавливающие растворы готовились непосредственно перед проведением анализа. 100 мкл раствора конъюгата антитело-POD (включенного в аналитический набор) добавляли в ячейки 96-луночной плашки, предварительно покрытые антителами (включенными в аналитический набор). 20 мкл стандартного и исследуемого раствора (из другой 96-луночной плашки, содержащей фибробласты) добавляли в соответствующие ячейки. Плашку слегка перемешивали, плотно закрывали и инкубировали в течение трех часов при температуре 37°С.
После инкубации каждую ячейку четыре раза аккуратно промывали буферным раствором PBS (400 мкл). После завершения промывки из всех ячеек полностью удаляли всю жидкость. В каждую ячейку добавляли по 100 мкл субстратного раствора (перекись водорода и тетраметилбензидин в буферном растворе, поставляются с аналитическим набором) и плашку инкубировали в течение 15 минут. В этот момент в каждую ячейку в том же порядке, что и субстрат, добавляли по 100 мкл останавливающего раствора (свежеприготовленного 1 н H2SO4). Плашку осторожно перемешивали и измеряли абсорбцию при 450 нм. Результаты анализировались статистически методом одностороннего дисперсионного анализа.
Результаты:
Как видно из Фиг.24 и Таблицы 6, добавление пептида 19-mer (SEQ ID NO: 11) приводит к усилению синтеза коллагена при обеих используемых концентрациях. Контрольные клетки без добавления пептида 19-mer (SEQ ID NO: 11) производили наименьшее количество коллагена. При концентрациях пептида 19-mer (SEQ ID NO: 11) 10-5 и 10-6 M клетки производили статистически значимое (Р<0.001) количество коллагена. Эти результаты указывают на то, что пептид 19-mer (SEQ ID NO: 11) стимулирует продукцию коллагена.
ССЫЛКИ:
Следующие ссылки, а также любые другие ссылки, упомянтые здесь, включены в настоящее изобретение посредством ссылки.
Agren, M.S. (1999). Matrix metalloproteinases (MMPs) are required for reepithelialization of cutaneous wounds. Arch. Dermatol. Res. 291, 583-590.
Becker, J.W., Marcy, A.I., Rokosz, L.L., Axel, M.G., Burbaum, J.J., Fitzgerald, P.M., Cameron, P.M., Esser, C.K., Hagmann, W.K., Hermes, J.D., and Springer, J.P. (1995). Stromelysin-1: Three dimensional structure of the inhibited catalytic domain and of the C-truncated proenzyme. Protein Sci. 4, 1966-76.
Brown, RL., Breeden, MP., and Greenhalgh, MD., (1994). PDGF and TGF-a act synergistically to improve wound healing in the genetically diabetic mouse. J. Surg. Res. 56, 562-570.
Browner, M.F., Smith, W.W., Castelhano, A.L. (1995). Matrilysin-inhibitor complexes: Common themes among 18 metalloproteinases. Biochemistry 34, 6602-10.
Calabrese, E.J. "Cell Migration/Chemotaxis: Biphasic Dose Responses," Critical Reviews tm Toxicology 2001, 31 (4&5), 615-624.
Calvin, М. "Cutaneous Wound Repair," Wounds 1998, 10(1), 12-32.
Clark, R.A.F. "Wound Repair. Overview and General Considerations," in The Molecular and Cellular Biology of Wound Repair, 2nd edition, Plenum Press: New York, 1996, pp.3-50.
Di Colandrea, T., Wang, L., Wille, J., D'Armiento, J., and Chada, K.K. (1998). Epidermal expression of collagenase delays wound healing in transgenic mice. J. Invest. Dermatol. 111, 1029-1033.
Duivenvoorden, W.C.M., Hirte, H.W., and Singh, G. (1997). Use of tetracycline as an inhibitor of matrix metalloproteinase activity secreted by human bone metastasizing cancer cells. Invasion and Metas. 17, 312-322.
Fernandez-Catalan, С., Bode, W., Huber, R, Turk, D., Calvete, J.J., Lichte, A., Tschesche, H., and Maskos, K. (1998). Crystal structure of the complex formed by membrane type-1 matrix metalloproteinase with the tissue inhibitor of metalloproteinases-2, the soluble progelatinase A receptor. EMBO J. 17, 5238-48.
Freire, E., van Osdol, WW., Mayorga, OL, and Sanchez-Ruiz, JM. (1990). Calorimetrically determined dynamics of complex unfolding transitions in proteins. Annu Rev Biophys Biophys Chem. 19, 159-88.
Gomis-Ruth, F.X., Maskos, К., Betz, M., Bergner, A., Huber, R., Suzuki, K., Yoshida, N., Nagase, BL, Brew, K., Bourenkov, G.P., Bartunik, H., and Bode, W. (1997). Mechanism of inhibition of the human matrix metalloproteinase stromelysin-1 by TIMP-1. Nature 389, 77-81.
Grams, F., Reinemer, P., Powers, J.C., Kleine, Т., Pieper, M., Tschesche, H., Huber, R., Bode, W. (1995). X-ray structures of human neutrophil collagenase complexed with peptide hydroxamate and peptide thiol inhibitors: Implications for substrate binding and rational drug design. Eur. J. Biochem. 228, 830-834.
Guex., N. and Peitsch, M.C. (1997). Swiss Model and the Swiss-Pdb Viewer: An environment for comparative protein modeling. Electrophoresis 18, 2714-2723.
Higgins, DG., Bleasby, AJ., and Fuchs, R. (1992). CLUSTAL V: improved software for multiple sequence alignment. Comput Appl Biosci., 8(2), 189-91.
Howard, E.W., Bullen, E.C., and Banda, M.J. (1991). Preferential inhibition of 72 and 92 kDa gelatinase by tissue inhibitor of metalloproteinase-2. J. Biol. Chem. 266, 13070-13075.
Huang, W., Suzuki, K., Nagase, H., Arumugam, S., Van Doren, S.R., and Brew, K. (1996). Folding and characterization of the amino terminal domain of human tissue inhibitor of metalloproteinases-1 (TIMP-1) expressed at high yield in E.coli. FEBS Lett. 384, 155-161.
Karlsson, R., and Falt, A. (1997). Experimental design for kinetic analysis of protein-protein interactions with surface plasmon resonance biosensors, J. Immunol. Meths. 200, 121-33.
Lakowicz, J.R. (1983). Principles of Fluorescence Spectroscopy, Chapter 10, Plenum Press, New York, London.
Levit, S., and Berger, A (1976). Ribonuclease S-peptide. A model for molecular recognition. J. Biol. Chem. 251, 1333-9.
Levy, D.E., Lapierre, F., Liang, W., Ye, W., Lange, C.W., Li, X., Grobelny, D., Casabonne, M., Tyrrell, D., Holme, К., Nadzan, A., and Galardy, R.E. (1998). Matrix metalloproteinase inhibitors: A structure activity study. J. Med. Chem. 41, 199-223.
Li, J., Brick, P., O'Hare, M.C., Skarzynski, Т., Lloyd, L.F., Curry, V.A, Clark, I.M., Bigg, H.F., Hazleman, B.L., Cawston, T.E. et al. (1995). Structure of full-length porcine synovial collagenase reveals a C-terminal domain containing a calciumlinked, four-bladed beta-propeller. Structure 3, pp.541-49.
Libson, A.M., Gittis, A.G., Collier, I.E., Marmer, B.L., Goldberg, G.I., and Lattman, E.E. (1995). Crystal structure of the haemopexin-like С terminal domain of gelatinase A. Nat. Struct Biol. 2, 938-42.
Lofas, S., Johnsson, В., Tegendahl, K., and Ronnberg, I. (1993). Dextran modified gold surfaces for surface plasmon resonance biosensors; immunoreactivity of immobilized antibodies and antibody-surface interaction studies. J. Colloid Interface Sci. 65, 423-431.
Morton, T.A., Myska, D.G., and Chaiken, I.M. (1995). Interpreting complex binding kinetics from optical biosensors: A comparison of analysis by linearization, the integrated rate equation, and numerical integration. Anal. Biochem. 227, 176-185.
Moses, M.A., Marikovsky, M., Harper, J.W., Vogt, P., Eriksson, E., Klagsbrun, M. and Langer, R. (1996). Temporal study of the activity of matrix metalloproteinases and their endogenous inhibitors during wound healing. J. Cell. Biochem. 60, 379-386.
Odake, S., Morita, Y., and Morikawa, T. (1994). Inhibition of matrix metalloproteinases by peptidyl hydroxamic acids. Biochem. Biophys. Res. Comm. 199, 1442-1446.
Ohtsuka, Y.; Lee, J.; Stamm, D.S.; Sanderson, I.R. "MIP-2 Secreted by Epithelial Cells Increases Neutrophil and Lymphocyte Recruitment in the Mouse Intestine. "Gut 2001, 49, 526-533.
Olson, M.W., Gervasi, D.C., Mobashery, S., and Fridman, R. (1997). Kinetic analysis of the binding of human matrix metalloproteinase 2 and 9 to tissue inhibitor of metalloproteinase (TIMP)-1 and TIMP-2. J. Biol. Chem. 272, 29975-29983.
O'Meara, SM.; Cullum, N.A.; Majid, J.; Sheldon, T.A. "Systemic Review of Antimicrobial Agents Used for Chrome Wounds," Brit J. Surg. 2001, 88, 4-21.
O'Shannessy, D.J., Brigham-Burke, M., Saneson, K.K, Hensley, P., and Brooks, I. (1993). Determination of rate and equilibrium binding constants for macromolecular interactions using surface plasmon resonance: use of non linear least squares analysis methods. Anal. Biochem. 212, 457-468;
Postlethwaite, A.E.; Kang, A.H. "Fibroblast Chemoattractants" in Methods of Enzymology, vol.163, Academic Press: New York, 1988, pp.694-707.
Reinemer, P., Grams, F., Huber, R., Kleine, Т., Schnierer, S., Pieper, M., Tschesche, H., Bode, W. (1994). Structural implications for the role of the N terminus in the superactivation of collagenases: A crystallographic study. FEBBS Lett. 338, 227-33.
Saariaho-Kere, U.K. (1998). Patterns of matrix metalloproteinase and TIMP expression in chronic ulcers. Arch. Dermatol. Res. 290 (suppl), 47-54.
Sayle, R.A. and Milner-White, E.J. (1995). RasMol: Biomolecular graphics for all. Trends in Biochemical Sciences 20, 374-376.
Segel, IH. (1993) Enzyme Kinetics: Behavior and analysis of rapid equilibrium and steady-state enzyme systems. Wiley Classics Library, John Wiley and Sons, Inc. New York.
Staiano-Coico, L.; Higgins, P.J.; Schwartz, S.B.; Zimm, A.J.; Gonclaves, J. "Wound Fluids: A Reflection of the State of Healing," Ostomy/Wound Management 2000, 46(S1A), 83S-93S.
Su, J-L., Becherer, D., Edwards, C., Bukhart, W., McMgeehan, G.M., and Champion, B.R. (1995). Monoclonal antibodies against human collagenase and stromelysin. Hybridoma. 14, 383-390.
Taylor, K.B., Windsor, J.L., Caterina, N.C.M., Bodden, M.K., and Engler, J.A. (1996). The mechanism of inhibition of collagenase by TIMP-1. J. Biol. Chem. 271, 23938-23945.
Tuuttila, A., Morgunov, E., Bergmann, U., Lindqvist, Y., Maskos, K., Fernandez-Catalan, C., Bode, W., Tryggvason, K., and Schneider, G. (1998). Three dimensional structure of human tissue inhibitor of metalloproteinases-2 at 2.1 Å resolution. J. Mol. Biol. 284, 1133-1140.
Vaalamo, M., Weckroth, M., Puolakkainen, P., Kere, J., Saarinen, P., Lauharanta, J., and Saarialho-Kere, U.K. (1996). Patterns of matrix metalloproteinase and TIMP-1 expression in chrome and normally healing human cutaneous wounds. Brit. J. Dermatol. 135, 52-59.
Vaalamo, M., Mattila, L., Johansson, N., Kariniemi, A-L., Karjalainen-Lindsberg, L., Kahari, V-M., and Saarialho-Kere, U.K. (1997). Distinct populations of stromal cells express collagenase-3 (MMP-13) and collagenase-1 (MMP-1) in chronic ulcers, but not in normally healing wounds. J. Investig. Dermatol. 109, 96-101.
Weckcroth, M., Vaheri, A., Lauharanta, J., Sorsa, Т., and Konttinen, Y.T. (1996). Matrix metalloproteinases, gelatinases, and collagenases in chronic leg ulcers. J. Investig. Dermatol. 108, 1119-1124.
Wojtowicz-Praga, S.M., Dickson, R.B., and Hawkins, M.J. (1997). Matrix metalloproteinase inhibitors. Investigational new Drugs. 15, 61-75.
Wysocki, A.; Staiano-Cioco, L.; Grinnell, F. "Wound fluid from chronic leg ulcers contains elevated levels of metalloproteinases MMP-2 and MMP-9," J. Invest. Dermatol. 1993, 101,64-68.
Wysocki, A.B. "Wound Fluids and the Pathogenesis of Chrome Wounds," J. Wound Ostomy Continence Nurs. 1996, 23(6), 283-290.





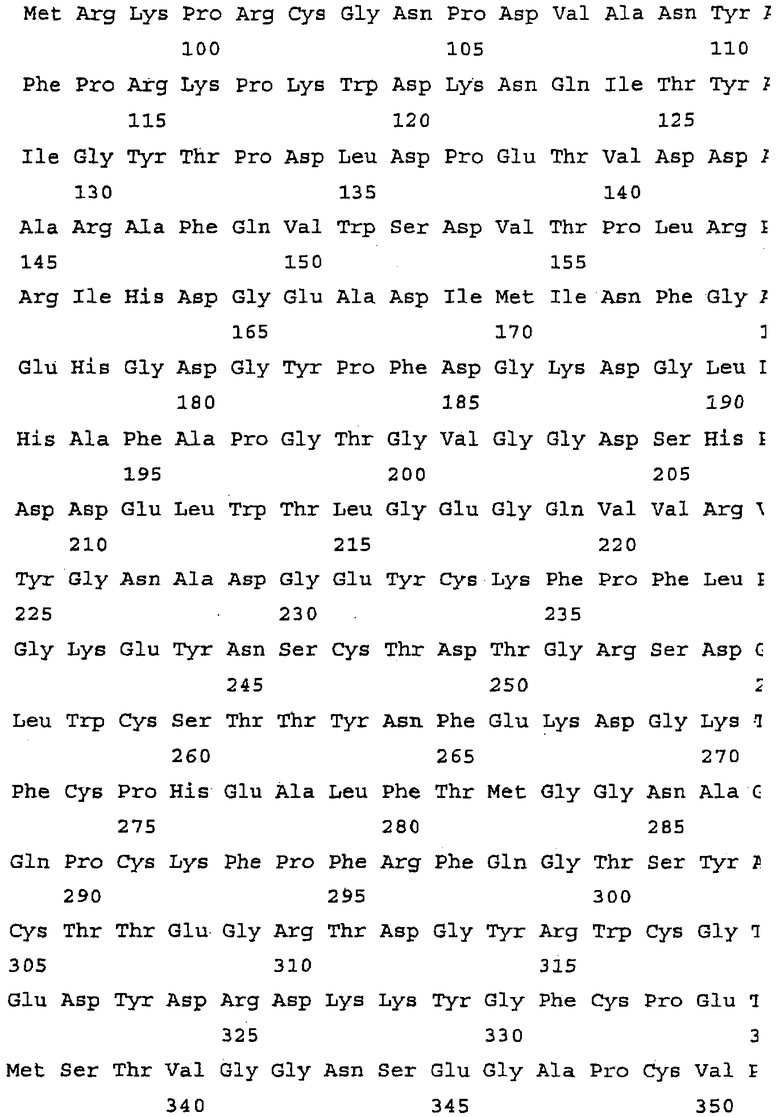










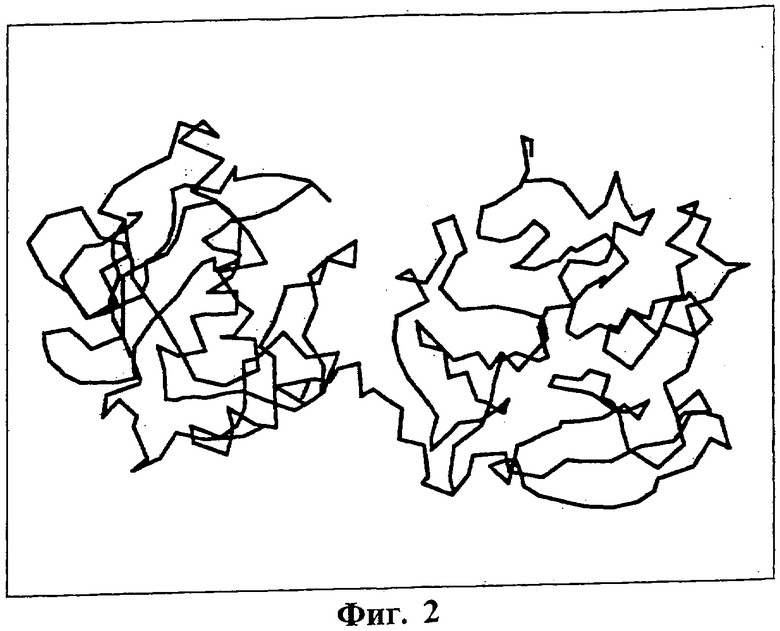
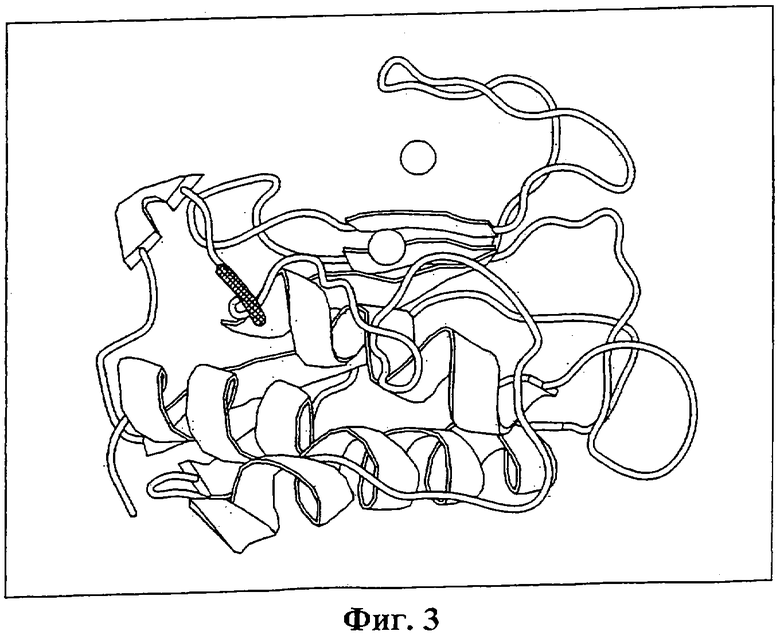
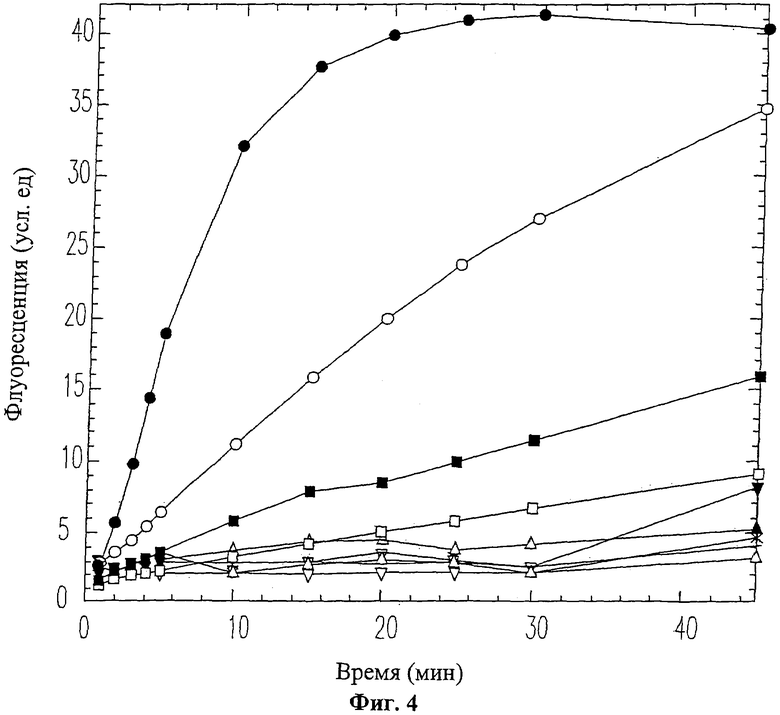
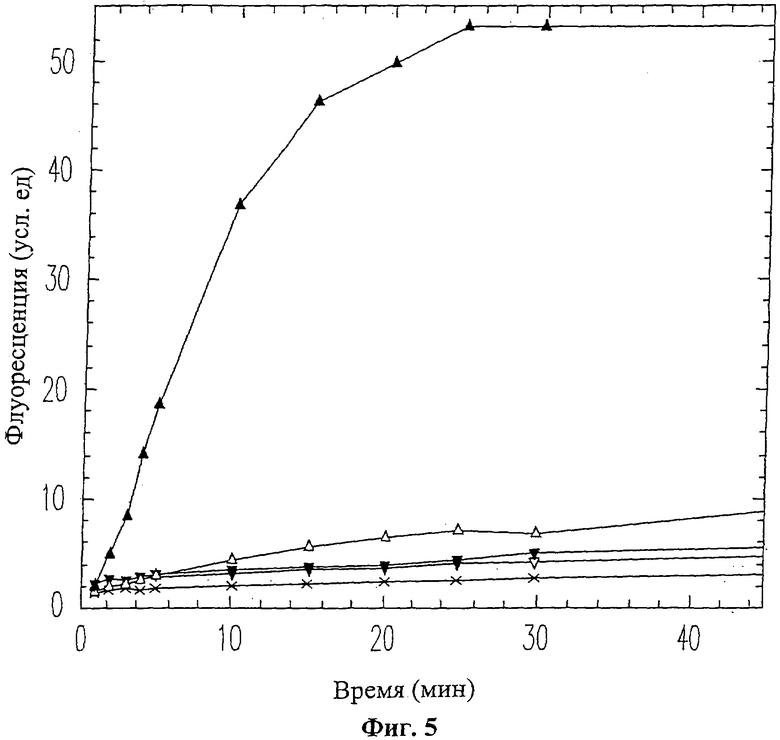
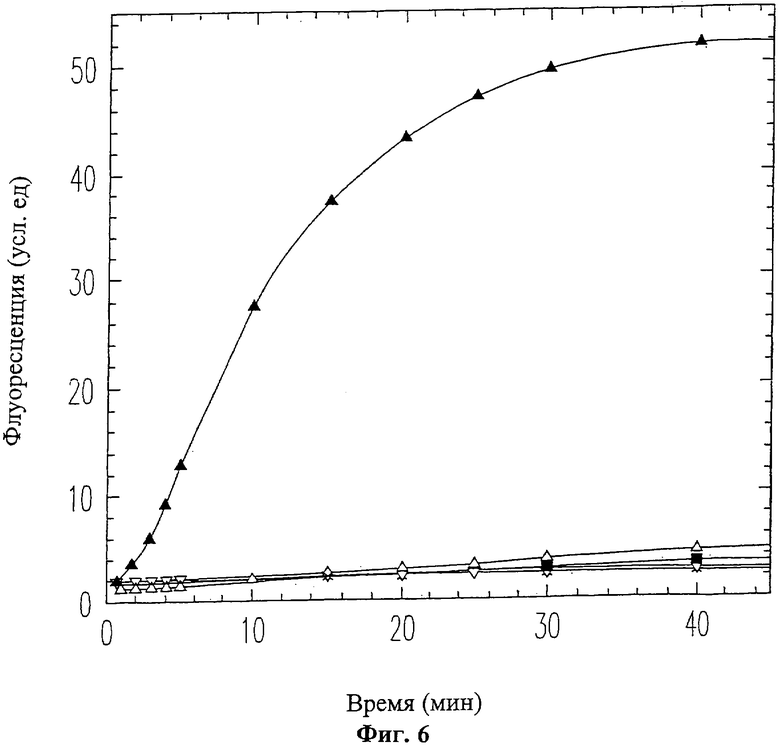
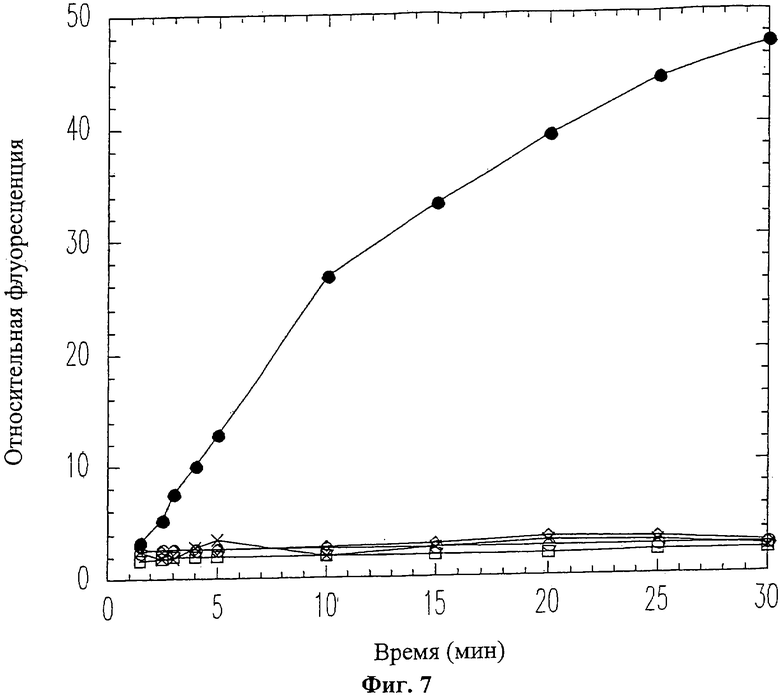
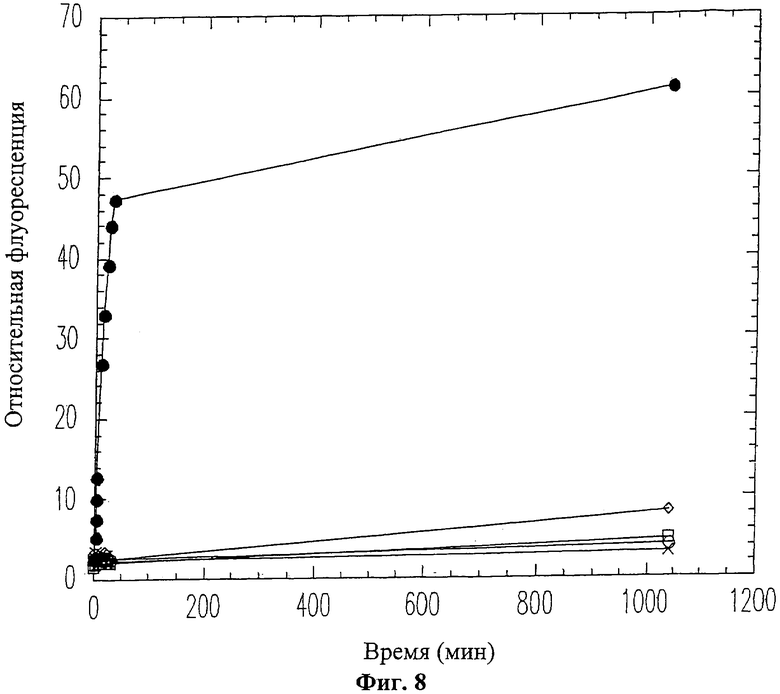
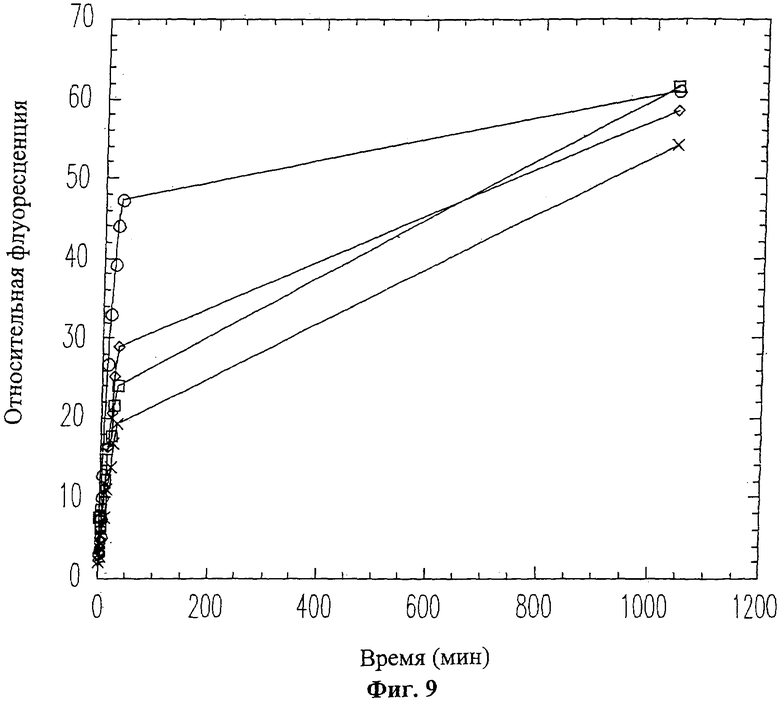
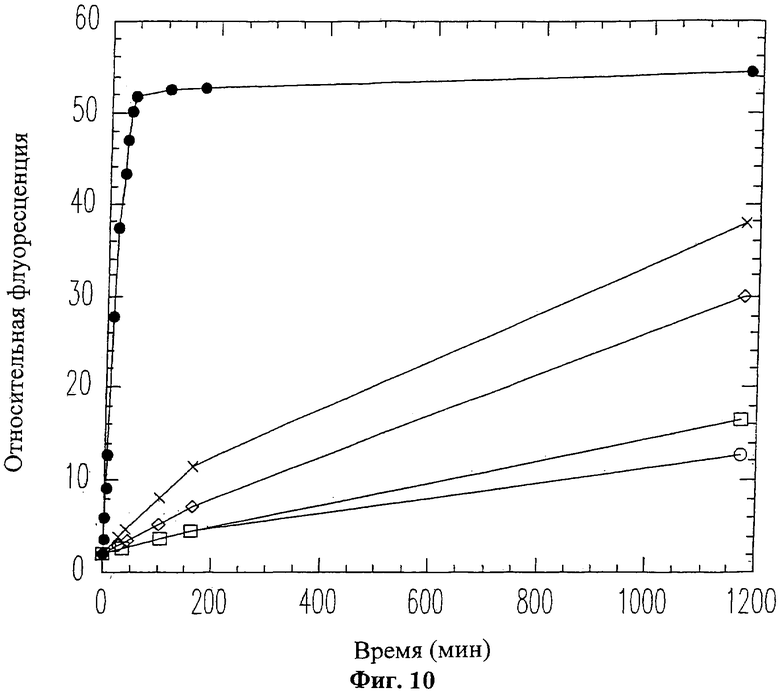
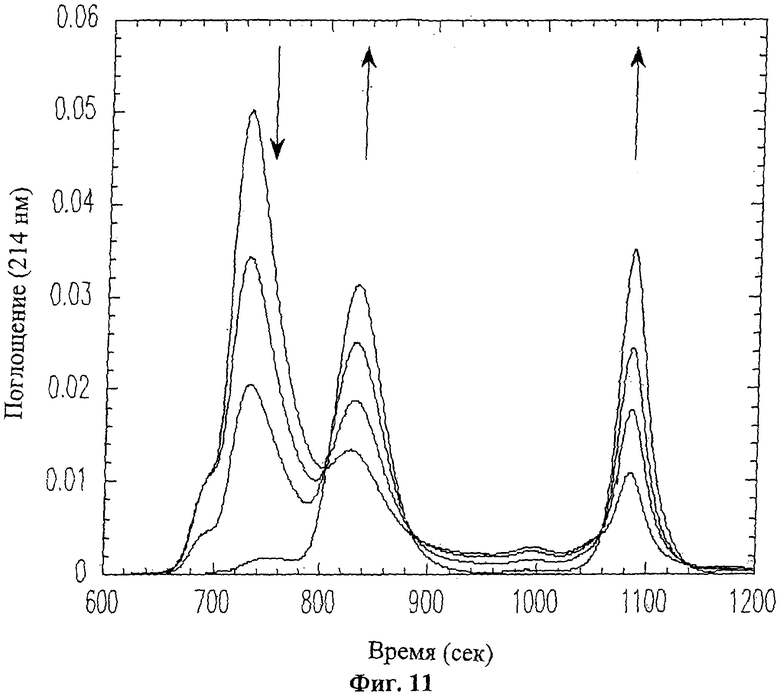

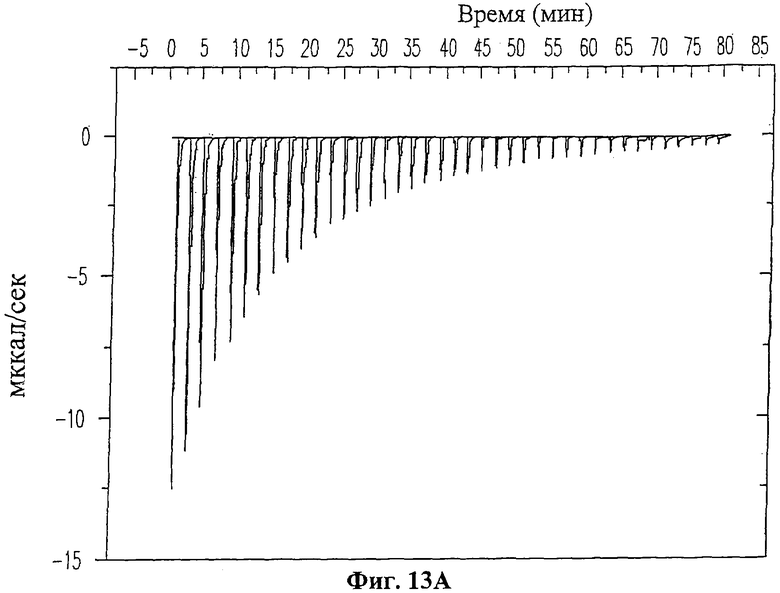
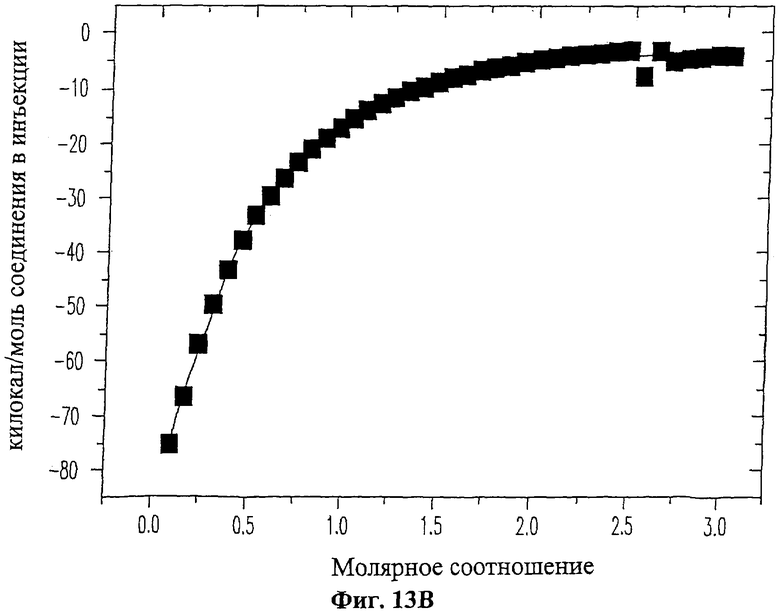
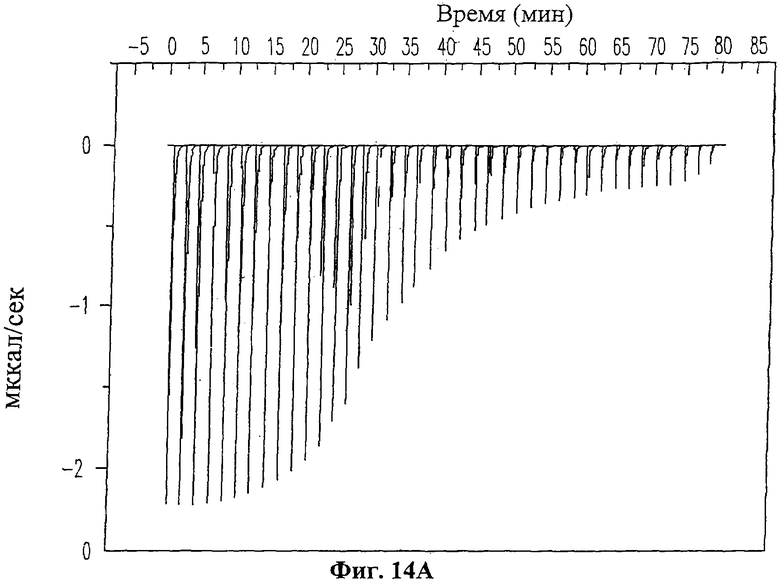
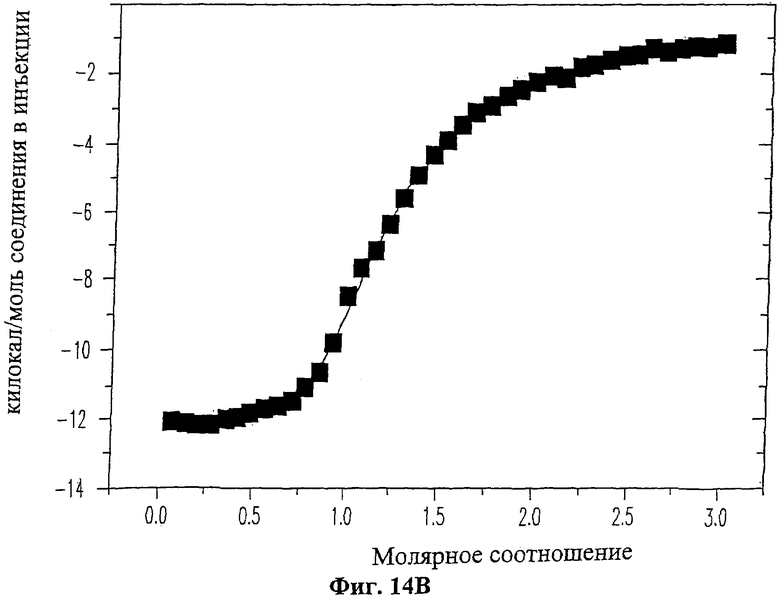
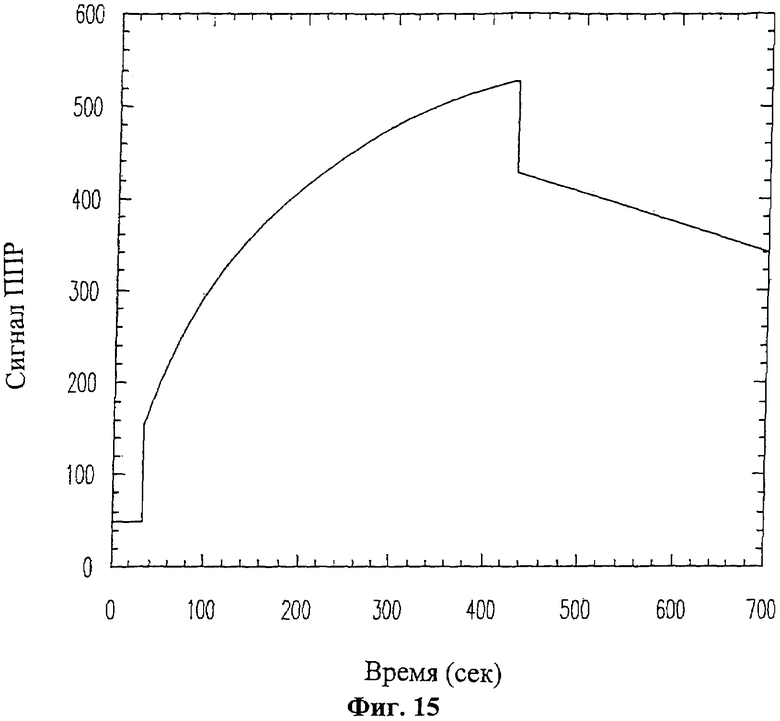
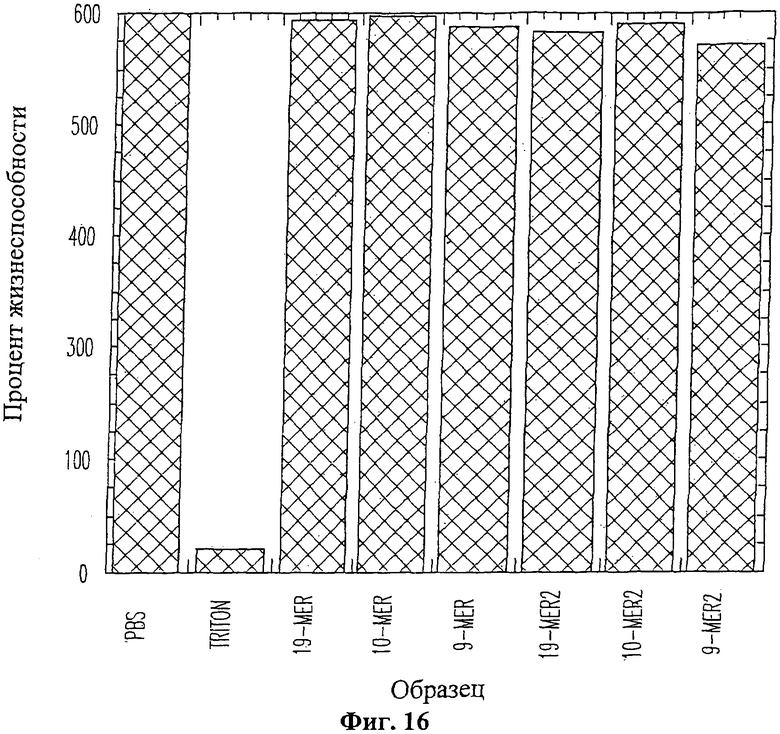
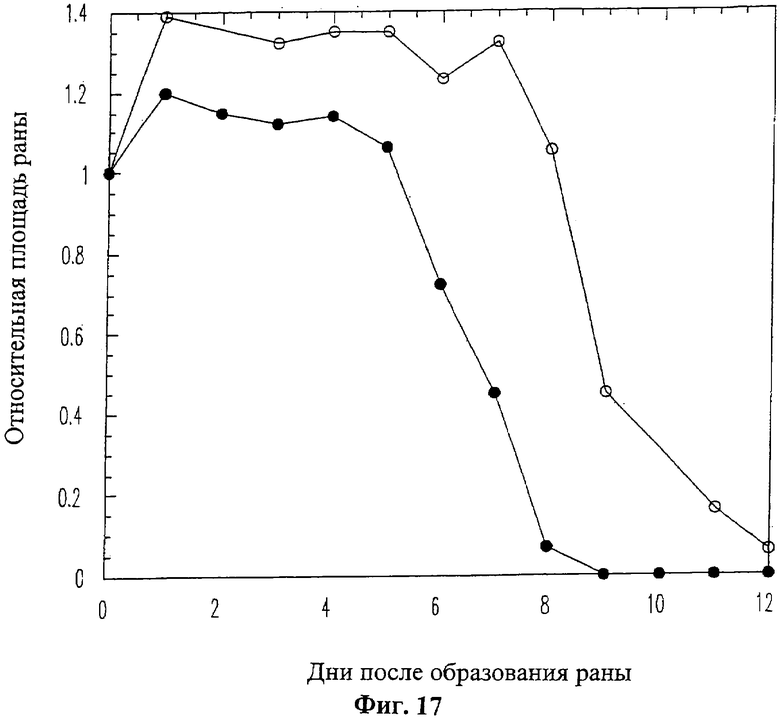
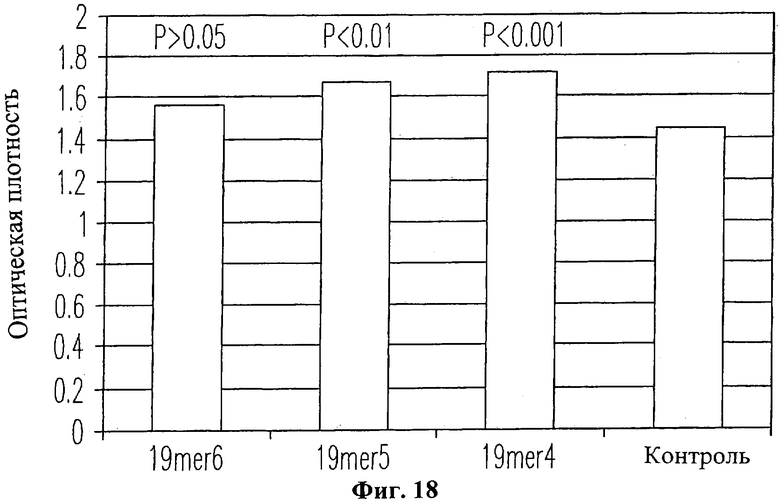
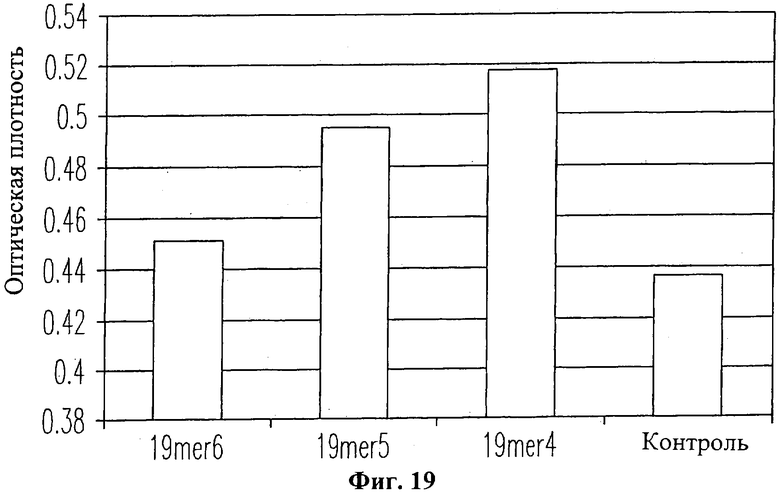

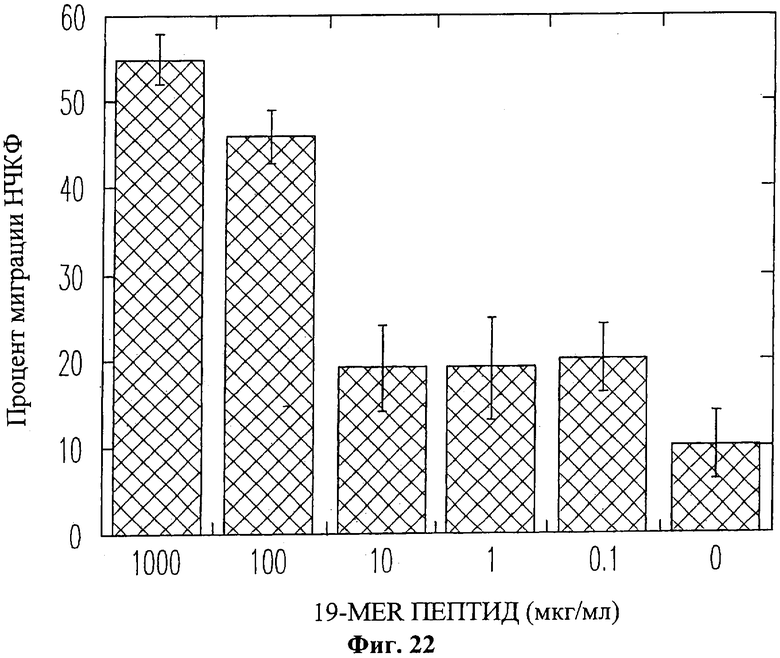

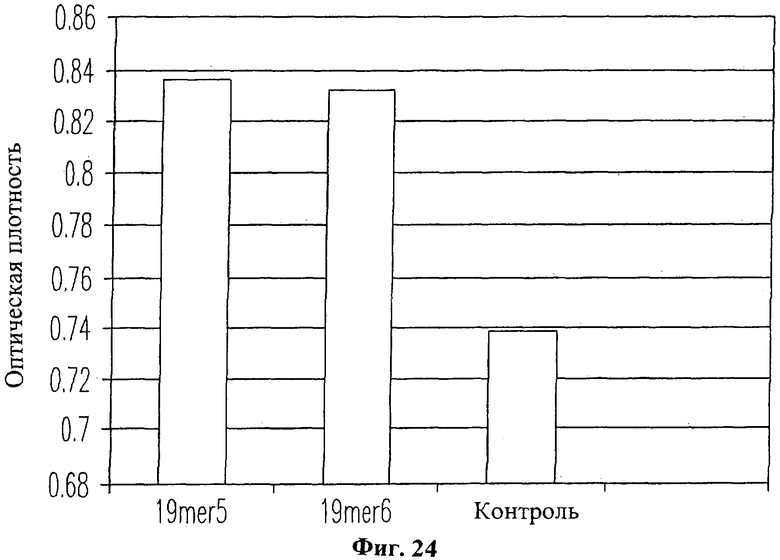
Изобретение относится к биотехнологии. Предложены пептиды, которые происходят из проферментных форм матриксных металлопротеиназ и являются ингибиторами матриксных металлопротеиназ. Аминокислотная последовательность представлена в описании. Описана композиция для стимулирования развития здоровой кожи, содержащая терапевтически эффективное количество пептидов. Описаны перевязочный материала для ран, способ стимулирования развития здоровой кожи и лосьон для обработки кожи для уменьшения результатов воздействия старения и для стимулирования развития здоровой кожи и лечения ран на основе композиции. Раскрыто применение композиции для изготовления лекарственного средства, способствующего заживлению ран, предотвращению рубцевания, снижению образования морщин, уменьшению результатов воздействия старения или улучшения цвета кожи. Использование изобретения позволяет получить новые противовозрастные и ранозаживляющие агенты. 6 н. и 9 з.п. ф-лы, 24 ил., 6 табл.
Приоритет по пунктам:
| WO 9010228 А, 07.09.1990 | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ МЕДИАТОРА МЕЖКЛЕТОЧНОЙ АДГЕЗИИ (ВАРИАНТЫ), СПОСОБ ИНГИБИРОВАНИЯ МЕЖКЛЕТОЧНОЙ АДГЕЗИИ, СПОСОБ ЛЕЧЕНИЯ ВОСПАЛИТЕЛЬНОГО ЗАБОЛЕВАНИЯ, СПОСОБ АНАЛИЗА ТЕСТИРУЕМОГО СОЕДИНЕНИЯ НА СПОСОБНОСТЬ ИНГИБИРОВАТЬ КЛЕТОЧНУЮ АДГЕЗИЮ, СПОСОБ ПОЛУЧЕНИЯ СОЕДИНЕНИЯ | 1991 |
|
RU2123338C1 |

