ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области слитых конструктов (fusion constructs) и их применению для лечения рака и в регенеративной медицине. Конкретно, изобретение относится к конструктам, включающим тканевые ингибиторы металлопротеиназ (ТИМП), закрепленные (GPI-anchored) гликозилфосфатидилинозитом (ГФИ). Дополнительно, слитые конструкты по настоящему изобретению являются эффективными регенеративными средствами, полезными в области лечения ран.
Уровень техники изобретения
ТИМП в исследовании рака
Лечение рака остается важной задачей и использует различные терапевтические подходы и стратегии, предлагая различные степени успеха. Один известный подход - увеличить чувствительность раковых клеток к иммуноопосредованному лизису. Чувствительность опухолей к иммуноопосредованному лизису связывали с биологией матриксных металлопротеиназ (ММП), и определенно, с экспрессией ММП клеточной поверхностью клетки - мишени опухоли. Матриксные металлопротеиназы (ММП) ухудшают компоненты внеклеточного матрикса (ВКМ) и вовлечены в модернизацию ткани, инвазию опухоли и метастаз (Egeblad & Werb, 2002; Itoh & Nagase, 2002). Активность ММП также связана с эффективностью как перфорин/гранзим-(perforin/granzyme-), так и FAS-опосредованного апоптоза (рассмотрены в Egeblad & Werb, 2002). Показано, что активность ММП регулируется на многих уровнях, включая четыре эндогенных ингибитора, тканевые ингибиторы матриксных металлопротеиназ (ТИМП-1, -2, -3 и -4) (Bode & Maskos, 2003). Баланс в естественных условиях (in vivo) между ММП и ТИМП определяет, имеет ли место матриксная ресорбция или депонирование (Nagase & Woessner, 1999).
Эндогенные ингибиторы тканевой металлопротеиназы (ТИМП) показывают разнообразные физиологическо-биологические функции, включая замедление роста опухоли, метастаз и апоптоза. Эти разнообразные биологические активности ТИМП связывали частично со стехиометрией белковых взаимодействий ТИМП/ММП/ клеточная поверхность. Пополнение цитотоксических лимфоцитов представляет один возможный путь защиты от опухолей. Хотя цитотоксические Т лимфоциты (ЦТЛ) и естественные клетки-киллеры (ЕКК), которые проникают и узнают опухолевые клетки, идентифицированы, эффективный антиопухолевый иммунитет часто не в состоянии эффективно развиться. Эта неэффективность является одной из причин, которая предотвращает полное удаление остаточных опухолевых клеток после неполной хирургической резекции, или из-за продвинутой стадии болезни или местной неоперабельности. Этиология этого функционального дефицита цитотоксических лимфоцитов в настоящее время неясна.
В общем, антиопухолевые эффекты ЦТЛ и клеток ЕКК опосредованы либо через перфорин/гранзим-, либо Fas-опосредованные (CD95/CSD95L) апоптотические пути (Kagi и др., 1994). Перфориновый путь опосредован цитотоксинами, секретированными в течение времени узнавания целевых клеток ЦТЛ и ЕКК (Kagi и др., 1994). CD95 или Fas-рецептор смерти принадлежит регулятору семейства клеток смерти белков и имеет центральное значение в иммуноопосредованном апоптозе опухолевых клеток (Nagata, 1999). Человеческий FAS/CD95/Apo-1 является единственным трансмембранным гликопротеиновым рецептором (325 аминокислот, 45-48 kDa). Лиганд FAS (лиганд FAS, FASL, CD95L) является составным мембранным белком и является трансмембранным гликопротеином типа II. FASL является членом семейства фактора некроза опухолей (ФНО), которое включает ФНОα, α- и β-цепи лимфотоксина (LT), лиганда CD40 и лиганда CD30. Действие FAS опосредуется через FADD (домен смерти, ассоциированный с FAS)/MORTI, белок адаптера, который имеет домен смерти на С-конце и связан с цитоплазматическим доменом смерти FAS. Как найдено, много опухолей устойчивы к апоптозу, опосредованному FAS (Frost и др., 2003; рассмотрено в Igney & Krammer, 2002).
В качестве модельной системы, чтобы испытать конструкты ТИМП-ГФИ по настоящему изобретению, клеточные линии почечно-клеточного рака использовали в качестве примера. Почечно-клеточный рак (ПКР) является седьмой ведущей причиной рака. У приблизительно одной трети пациентов с ПКР есть метастатическая болезнь при представлении и до 50% рецидива после нефрэктомии (Vogelzang & Stadler, 1998). ПКР трудно лечить, и иммунологические методы лечения, такие как интерферон-альфа и интерлейкин-2 обычно более эффективны, чем химиотерапия или облучение (Vogelzang & Stadler, 1998).
Цитотоксические лимфоциты представляют потенциальный компонент в защите против опухолей, включая ПКР.
Один член семейства ТИМП, ТИМП-1, является ингибитором ММП широкого действия (Bode and Maskos, 2003). Это - растворимый белок, который может быть обнаружен на поверхности клеток только через ассоциацию с белками, связанными с поверхностью (Brew и др., 2000; Klier и др., 2001). Полная роль ТИМП-1 в биологии рака остается субъектом противоречивых сообщений (Brand, 2002). К настоящему времени признано, что ТИМП-1 играет роль в ангиогенезе, миграции клеток и пролиферации (Brand, 2002). Недавно, было показано, что белок ТИМП-1, закрепленный ГФИ, показал ярко выраженное подавление миграции эндотелиальных клеток в ответ на основной фактор роста фибробластов, основной ФРФ (bFGF), (Djafarzadeh R и др., 2004).
Обычные стратегии и подходы в терапии рака все еще страдают от проблемы, такой, что опухолевые клетки трудно удалять, как только опухоль развилась. Первичные опухоли обычно удаляются из пациента хирургией. Однако, в некоторых случаях не все области доступны для хирурга, и, таким образом, опухолевые клетки остаются в теле, где они могут развиться во вторичные опухоли. Это - результат неполной хирургической резекции первичной опухоли.
Настоящее изобретение, следовательно, обеспечивает эффективное противораковое средство и стратегию, чтобы уменьшить или смягчить пролиферацию опухолевых клеток в человеке, в особенности у пациента, который был подвергнут неполной хирургической резекции первичной опухоли. Противораковое средство по настоящему изобретению полезно для лизиса опухолевых клеток как клеточных линий в пробирке, так и в тканях в естественных условиях.
Роль ТИМП в регенеративной медицине
Настоящее изобретение, кроме того, полезно в области регенеративной медицины. Одна значительная область регенеративной медицины касается процесса заживления ран. Заживление ран относится к естественной восстановительной реакции на повреждение ткани и включает сложный каскад клеточных событий, который, в конечном счете, производит замену покрытия, воссоздание и восстановление прочности травмированной ткани. Этот процесс, обычно, включает задействование и пролиферацию различных типов клеток, выработку клеточного матрикса и увеличение иммунного надзора.
Заживление ран протекает последовательно и может быть разделено на четыре общих фазы: воспаление, грануляция, (пере-)эпителизация и реконструкция ткани. Каждая фаза процесса заживления раны регулируется специальными путями трансдукции сигнала. Во время заживления раны происходит увеличение экспрессии факторов роста и цитокинов; в частности, описано увеличение уровней ФНО, интерлейкина-1 (IL-1) и IL-6. Во время начальной фазы воспаления, которая включает эффекторные белки IL-1, ФНО-α и ЦСЖ (цереброспинальная жидкость), как макрофаги, так и нейтрофилы действуют, чтобы сформировать сгусток фибрина. Во время фазы грануляции фибробласты разрастаются, мигрируют к ране и секретируют внеклеточный матрикс (ВКМ). Эффекторные белки, вовлеченные в эту последнюю фазу, включают ММП и факторы роста: тромбоцитпроизводный, фибробластов, эпидермальный и эндотелия сосудов (соответственно PDGF, FGF, EGF и VEGF). Третья фаза в заживлении раны, переэпителизация, характеризуется разрастанием кератиноцитов, которые мигрируют в рану, а также увеличением миофибробластов, которые ответственны за сокращение раны. Результатом этой фазы, которая вовлекает эффекторные белки ММП, фактор роста кератиноцитов, трансформирующий фактор роста, фактор, стимулирующий колонии гранулоцитов/макрофагов, эпидермальный фактор роста, урокиназу и активатор плазминогена тканевого типа (соответственно ММПs, KGF, TGF, GM-CSF, EGF и uРА и tPA), является переэпителизация поверхности раны, рассечение струпа и формирование барьера. Наконец, во время реконструкции ткани фибробласты производят коллагеновый матрикс, приводящий к формированию рубцовой ткани, апоптозу фибробластов и переключению от активации к дифференцированию кератиноцитов. Известные эффекторные белки, вовлеченные в эту последнюю фазу заживления раны, включают TGF-β1, ММП и ТИМП.
Таким образом, эффекторные клетки, ответственные за большинство аспектов заживления ран, являются фибробластами и кератиноцитами и ММП, которые играют важную роль в миграции фибробластов (ММП-1, -2, -3 и -13) и кератиноцитов (ММП-1, -2, -3 и -10) (Singer & Clark, 1999) в дополнении к образованию рубцов. Каждый из ММП имеет разную специфичность по субстрату в пределах ВКМ и играет важную роль в деградации ВКМ и обновлении. Семейство ММП включает, среди прочего, коллагеназы (ММП-1, ММП-8, ММП-13, ММП-18), стромелизины (ММП-3, ММП-10, ММП-11), желатиназы (ММП-2, ММП-9), матрилизин (ММП-7), металлоэластазу (ММП-12) и ряд мембраносвязанных матричных металлопротеиназ (МТ-ММП). Поскольку функция ММП протеолитически расщепить окружающий ВКМ, баланс между этой протеазной активностью и депонированием ВКМ во время заживления раны, то есть - восстановление поврежденной ткани, должен быть оптимально выдержан. Контроль активности ММП модулируется белками ТИМП, которые производятся большинством клеток, и действуют так, чтобы ингибировать ММП в отношении 1:1. Где этот тонкий баланс между протеолитическим распадом и депонированием ВКМ нарушен, там нарушения, такие как патологическое заживление раны, могут приводить, например, к хроническим ранам, чрезмерному рубцеванию или келоидному рубцу.
Следовательно, существует потребность управлять или влиять на физиологический баланс между протеазной активностью и депонированием ВКМ во время процесса заживления раны.
В другом варианте слитого конструкта по настоящему изобретению обеспечивают эффективное регенеративное средство для лечения состояний, определяемых нарушенным балансом между протеазной активностью ММП и депонированием ВКМ, как например, в келоидных рубцах или хронических ранах, которые обычно связывают с повышенными уровнями ММП. Дополнительно, слитые конструкты по настоящему изобретению обеспечивают эффективное регенеративное средство, которое может уменьшить, минимизировать или ингибировать образование рубцов во время процесса заживления раны.
Сущность изобретения
Настоящее изобретение обеспечивает новые противоопухолевые средства и методы лечения рака.
Настоящее изобретение основано на удивительной находке, что ТИМП, закрепленный якорем ГФИ, эффективно уменьшает или ослабляет пролиферацию раковых клеток, и вызывает лизис раковых клеток в клеточных линиях в пробирке и в естественных условиях. Структурные и функциональные определяющие факторы ТИМП соединяли с якорем гликозилфосфатидилинозита (ГФИ) и, необязательно, с муцином, чтобы получить очень эффективное химиотерапевтическое средство. Этот подход использует белки ТИМП, закрепленные гликозилфосфатидилинозитом (ГФИ), для включения в поверхностные мембраны после очистки и добавления к раковым клеткам. Слияние ТИМП-ГФИ с доменом муцина далее увеличивает представление белков ТИМП на поверхностной клеточной мембране и делает слитый конструкт более эффективным в придании раковым клеткам чувствительности к иммуноопосредованной деструкции.
В следующих примерах настоящее изобретение демонстрирует, что у ТИМП есть потенциал для ингибирования роста опухолевых клеток и сокращения развития опухоли в клеточных линиях в пробирке и в тканях в естественных условиях. Соединение ТИМП с якорем ГФИ и экзогенное введение ГФИ-фиксированного ТИМП, приводит к эффективной вставке белка ТИМП в клеточные мембраны раковых клеток. Поверхностная экспрессия ГФИ-фиксированного ТИМП-1 вызывала множество биологических эффектов в линиях раковых клеток с потенциальной терапевтической значимостью, такой как стимулирование FAS-опосредованного апоптотического пути в раковых клетках. Как показано в следующих примерах, подавление пролиферации раковых клеток зависело от дозы.
Белок ГФИ-фиксированный ТИМП-1 также блокировал секрецию проММП-2 и проММП-9 и резко изменял ассоциацию разнообразных ММП поверхностью клеток. Наиболее конкретно, обычно линии опухолевых клеток, устойчивые к FAS-апоптозу, становились чувствительными к FAS/CD95-опосредованному лизису. Обработка ГФИ-ТИМП приводит к низкоуровневой регуляции антиапоптотического белка BCL2 и соответствующему увеличению проапоптотического белка ВАХ. Этот сдвиг к более высокой концентрации проапоптотических белков может быть одной из причин повышенной чувствительности FAS-опосредованного апоптоза раковых клеток, поверхность которых включает ТИМП.
Согласно вышеуказанному подходу, ГФИ-фиксированные белки ТИМП или полипептиды по настоящему изобретению, как было доказано, являются особенно полезными в терапевтических применениях при лечении остаточного рака после неполной хирургической резекции первичной опухоли, такой как рак молочной железы, остеогенная саркома, почечно-клеточный рак или злокачественные опухоли головного мозга, например глиобластома.
Кроме того, так как опухолевые клетки, включая почечно-клеточный рак (ПКР), в сущности, устойчивы к FAS-опосредованному лизису, настоящее изобретение обеспечивает эффективное средство делать опухолевые клетки, восприимчивыми к FAS-опосредованному апоптозу.
Следовательно, в первом аспекте настоящее изобретение относится к слитому конструкту (ТИМП-ГФИ или ТИМП-муцин-ГФИ), включающему аминокислотную последовательность тканевого ингибитора металлопротеиназ (ТИМП) или его биологически активного фрагмента, в котором указанный ТИМП или его биологически активный фрагмент связаны с аминокислотной последовательностью домена муцина, за которой следует аминокислотная последовательностью якоря гликозилфосфатидилинозита (ГФИ).
В предпочтительном варианте 3'-конец ТИМП связан непосредственно с последовательностью якоря ГФИ и не содержат домен муцина.
Термин "муцин" относится к семейству больших, сильно гликозилированных белков. Один класс муцинов представляет собой муцины, связанные с мембраной, из-за наличия домена, охватывающего мембрану, который способствует удержанию в плазменной мембране, в то время как другой класс муцинов секретируется на поверхностях слизистой оболочки. Гены муцина кодируют мономеры муцина, которые обычно синтезируются как палочкообразные стебли апомуцина, которые посттрансляционно модифицируются избыточным гликозилированием. Две отчетливо различных области найдены в зрелых муцинах. Одна область включает область с концевыми амино- и карбоксигруппами, которая является слабо гликозилированной, но богата остатками цистеина, которые вероятно вовлечены в установление дисульфидных связей в пределах и среди мономеров муцина. Вторая центральная область образована многократными тандемными повторами последовательностей 10-80 остатков, в которых больше половины аминокислотных остатков являются серином или треонином.
Обычно муцины секретируются как массивные скопления белков, имеющих молекулярные массы примерно 1-10 миллионов дальтон. В пределах этих скоплений мономеры связаны друг с другом, главным образом, нековалентными взаимодействиями, хотя межмолекулярные дисульфидные мостики могут также играть роль в этом процессе. По меньшей мере, различают 19 человеческих генов муцина, включая MUC1, 2, 3А, 3В, 4, 5АС, 5В, 6-9,11-13 и 15-19.
Муцин, как понимают в настоящем изобретении, предпочтительно является доменом муцина, связанным с мембраной, и включает предпочтительно аминокислотную последовательность, выбранную из группы, состоящей из MUC1, MUC3A, MUC3B, MUC4, MUC11, MUC12, MUC16, и MUC17, или его варианта или участка (вышеупомянутые муцины рассмотрены в Moniaux N. и др., 2004). В другом предпочтительном варианте применяют стебель муцина, который выделяют из связанных с поверхностью хемокина CXCL16 или фракталкина (CX3CL1).
Фракталкин является членом большого и сложного генного суперсемейства хемокинов, которое состоит преимущественно из секретированных, провоспалительных молекул. Типичная стебельная структура хемокинов частично удерживается дисульфидными мостиками между позиционно сохраняемыми остатками цистеина. Для большинства пептидов хемокина структурной особенностью семейства является распределение четырех цистеинов в молекуле, то есть характерный цистеиновый мотив: СХС, СС и С, где С представляет собой цистеин и X является любым аминокислотным остатком. Четыре различных семейства хемокинов были идентифицированы на основе наблюдений, что пептиды хемокина могут отличаться организацией остатков цистеина, расположенных около N-конца молекулы. Сам фракталкин определяет одно из семейств хемокинов и отличается структурно от других семейств хемокинов, так как цистеины N-конца фракталкина разделены тремя остатками (то есть мотив СХ3С), а также привязывается к клеточной мембране удаленным С-концом трансмембранного якоря, который включает муциноподобный домен или муциноподобный стебель. Таким образом, домены муцина и фракталкина, содержащиеся в пределах слитых конструктов по настоящему изобретению, являются подходящими для достижения улучшенной фиксации белка ТИМП в клеточной мембране.
ТИМП, как используется в настоящем изобретении, предпочтительно получают из млекопитающего; более предпочтительно из человека (четыре ТИМП рассмотрены в Mannello F. и др., 2001). Примеры белков ТИМП, которые могут использоваться в соответствии с настоящим изобретением, включают ТИМП-1, ТИМП-2, ТИМП-3 или ТИМП-4 и их соответствующие варианты в других организмах, таких как мыши, кролики, собаки, кошки, овцы и коровы.
ГФИ-якорь, как используется в настоящем изобретении, предпочтительно получают из антигена, связанного с функцией лимфоцитов (LFA-3), или его участка и включает ГФИ-сигнальную последовательность, которая опосредует ассоциацию мембран.
Настоящее изобретение также относится к молекуле нуклеиновой кислоты, такой как РНК или ДНК, включая нуклеиновокислотную последовательность, которая кодирует конструкт ГФИ-фиксированный ТИМП по изобретению.
В дальнейшем аспекте настоящего изобретения молекула нуклеиновой кислоты по изобретению содержится в экспрессионной плазмиде, векторе или клетке-хозяине для экспрессии молекулы нуклеиновой кислоты по изобретению.
Настоящее изобретение также относится к применению слитых конструктов ТИМП-ГФИ или ТИМП-муцин-ГФИ по изобретению для лечения рака, особенно остаточного рака после хирургического удаления первичной опухоли.
В дальнейшем аспекте настоящего изобретения слитые конструкты ТИМП-ГФИ или ТИМП-муцин-ГФИ по изобретению содержатся в фармацевтическом препарате или медикаменте. В дальнейшем варианте слитые конструкты ТИМП-ГФИ или ТИМП-муцин-ГФИ по изобретению обычно применяют как противораковое средство или лекарство. В предпочтительном варианте противораковые препараты по изобретению вводятся или наносятся локально на место резекции опухолевой массы у больных с "высокорисковой" опухолью с высоким риском остаточных раковых клеток и повышенным уровнем местного рецидива, и у больных, имеющих очевидную остаточную опухоль из-за продвинутой стадии болезни или местной неоперабельности. Предпочтительно, слитый конструкт вводят распылением в рану и/или инъекцией в области, которые недоступны для хирургии.
Настоящее изобретение также относится к in vivo методу для торможения пролиферации раковых клеток, включающему стадии воздействия на линию раковой клетки эффективного количества слитых конструктов ТИМП-муцин-ГФИ и ТИМП-ГФИ.
В дальнейшем варианте настоящее изобретение обеспечивает новые средства и методы для лечения состояний, определяемых нарушенным балансом между нормальной физиологической активностью ММП протеазы и депонированием ВКМ, которые приводят к патологическому заживлению раны. В одном варианте настоящее изобретение обеспечивает средства и методы, пригодные для лечения келоидных или гипертрофических рубцующихся и хронических ран, обычно связанных с повышенным уровнем ММП. Кроме того, настоящее изобретение также обеспечивает эффективные средства и методы для снижения, минимизации или ингибирования формирования рубцов во время процесса заживления раны.
Определения
Термин "ТИМП", как используется здесь, означает эндогенный тканевый ингибитор металлопротеиназ, который, как известно, вовлекается в физиологическо-биологические функции, включая ингибирование активной матриксной металлопротеиназы, регулирование активации проММП, роста клеток и модуляции ангиогенеза. Человеческое "семейство ТИМП" содержит четыре члена, ТИМП-1, ТИМП-2, ТИМП-3 и ТИМП-4. Один предпочтительный член, используемый в настоящем изобретении, ТИМП-1, является секретированным белком, который может быть обнаружен на поверхности клетки через ее взаимодействие с поверхностными белками (Bode & Maskos, 2003).
Термин " слитый конструкт " или " слитый конструкт ТИМП", как используется здесь, относится как к молекуле нуклеиновой кислоты, так и к молекуле аминокислоты, закодированной таким образом.
Изобретение конкретно относится к нуклеиновым кислотам, содержащим нуклеотидную последовательность, включающую последовательность, определенную в соответствии с номером SEQ ID NOS:l-5, или ее гомологом или ее уникальными фрагментом.
В настоящем изобретении последовательность молекулы нуклеиновой кислоты, которая кодирует образующийся белок, считается гомологичной второй молекуле нуклеиновой кислоты, если нуклеотидная последовательность первой молекулы нуклеиновой кислоты является, по меньшей мере, на приблизительно 70%, предпочтительно, по меньшей мере, на приблизительно 80%, и более предпочтительно, по меньшей мере, на приблизительно 90%, гомологичной последовательности второй молекулы нуклеиновой кислоты. Гомология между двумя нуклеиновокислотными последовательностями может быть легко определена, используя известный алгоритм BLASTN (Altschul и другие, 1990) с параметрами настройки по умолчанию. Как дальнейший пример, другой известный тест на установление гомологии двух нуклеиновокислотных последовательностей - гибридизируются ли они при нормальных условиях гибридизации, предпочтительно при строгих условиях гибридизации.
Учитывая нуклеиновокислотную последовательность, раскрытую здесь, квалифицированный человек (специалист в данной области) может легко проектировать структуры нуклеиновых кислот, имеющих специфические функции в различных применениях. Например, специалист может получить олигонуклеотиды или полинуклеотиды для использования в качестве инициаторов в методиках амплификации нуклеиновой кислоты, таких как полимеразно-цепная реакция (PCR), лигазная цепная реакция (LCR), реакция репарации цепи (RCR), PCR-проба на лигирование олигонуклеотидов (PCR-OLA) и т.п. Олигонуклеотиды, полезные в качестве зондов в исследованиях гибридизации, такой, как in situ гибридизация, могут быть получены. Известны многочисленные методы для маркировки таких зондов радиоизотопами, флуоресцентными метками, ферментами и связывающими фрагментами (например, биотином), таким образом, зонды по изобретению могут быть легко приспособлены для легкого обнаружения.
Олигонуклеотиды также могут быть разработаны и получены для других целей. Например, изобретение дает возможность разработки антисенсорных олигонуклеотидов и триплексобразующих олигонуклеотидов для применения в исследовании взаимоотношений «структура-функция». Гомологичная рекомбинация может быть осуществлена адаптацией описанной нуклеиновой кислоты для применения в качестве нацеливающего средства.
Белок, кодированный нуклеиновой кислотой по настоящему изобретению, далее включает функциональные гомологи. Белок считают функциональным гомологом другого белка для определенной функции, как описано ниже, если у гомолога есть та же самая функция, что и у другого белка. Гомолог может быть, например, фрагментом белка или заменой, дополнением или делеционным мутантом белка.
Определение того, являются ли две аминокислотные последовательности существенно гомологичными для цели настоящего изобретения, основывается на исследовании FASTA согласно Pearson & Lipman (1988). Например, аминокислотную последовательность первого белка считают гомологичной последовательности второго белка, если аминокислотная последовательность первого белка имеет, по меньшей мере, приблизительно 70% идентичность аминокислотной последовательности, предпочтительно, по меньшей мере, приблизительно 80% идентичность и более предпочтительно, по меньшей мере, приблизительно 95% идентичность последовательности второго белка.
Возможность замены одной аминокислоты в последовательности эквивалентной аминокислотой известна. Группы аминокислот, известных как эквивалентные, включают
(a) Ala (A), Ser (S), Thr (Т), Pro (Р), Gly (G);
(b) Asn (N), Asp (D), Glu (E), GIln (Q);
c) His (H), Arg(R), Lys(K);
d) Met (M), Leu (L), Ile (I), Val (V); и
(e) Phe (F), Tyr (Y), Trp (W).
Замены, дополнения и/или делеции в аминокислотных последовательностях могут быть сделаны до тех пор, пока белок, кодированный нуклеиновой кислотой по изобретению, продолжает удовлетворять функциональным критериям, описанным здесь. Аминокислотная последовательность, которая является существенно той же самой, как другая последовательность, но отличается от другой последовательности одной или более замен, добавлений и/или делеций, как полагают, является эквивалентной последовательностью.
Предпочтительно, меньше 20%, более предпочтительно меньше 10% и еще более предпочтительно меньше 5% остатков аминокислот в последовательности заменяются, добавляются или удаляются из белка, кодированного нуклеиновой кислотой по изобретению.
Термин "ММП", как используется здесь, обозначает матриксную металлопротеиназу, которая принадлежит суперсемейству ММП, как представлено, по меньшей мере, 26 внеклеточными матрикс-деградирующими металлоэндопептидазами, которые действуют во время развития ткани и дифференцирования, клеточной инфильтрации, заживления ран, и как регуляторы иммунного ответа.
Термин "ГФИ", как используется здесь, обозначает гликоинозитфосфолипиды, в частности гликозилфосфатидилинозит, как описано в Medof и другие, 1996. Эти фосфолипидоподобные якоря имеют общую структуру для закрепления на мембране независимо от функции белка. ГПИ-фиксирующие единицы составляют из линейного гликана, содержащего фосфоэтаноламин, трех остатков маннозы и неацетилированого глюкозамина, связанного с инозитфосфолипидом. Последовательность ГФИ содержит сигналы, которые направляют ГФИ фиксацию.
Термин "муцин" или "домен муцина", как используется здесь, обозначает мембраносвязанный или немембранный компонент гликопротеина. Обычно, мембраносвязанные муцины имеют гидрофобные последовательности или трансмембранные домены, ответственные за их фиксацию в двойном слое липида и, необязательно, содержат один или несколько доменов, подобных фактору Виллебрандта, которые функционируют в олигомеризации мономеров муцина и заполнении секреторных пузырьков. Термин "муцин" или "домен муцина", как используется здесь, также охватывает стебли муцина или муциноподобные домены, такие как стебли муцина, обычно находимые в хемокинах CXCL16 или фракталкине (CX3CL1).
Слитый конструкт "ТИМП-ГФИ", как используется здесь, относится к ТИМП, который непосредственно связан с последовательностью ГФИ-линкера. Слитой конструкт ТИМП-ГФИ получен заменой эндогенной концевой последовательности 3'-мРНК или кДНК ТИМП на концевую последовательность 3'-мРНК или кДНК белков естественно закрепленных якорем ГФИ (то есть последовательности, которые содержат сигналы, которые направляют фиксацию ГФИ).
"ТИМП-муцин-ГФИ" или "ТИМП-муц-ГФИ", как используется здесь, относится к ТИМП, который непосредственно слит с доменом муцина, за которым следует последовательность ГФИ-линкера. Слитый конструкт ТИМП-муцин-ГФИ разработан, как описано для ТИМП-ГФИ, но включает аминокислотную последовательность домена муцина между аминокислотными последовательностями ТИМП и ГФИ. По аналогии, "ТИМП-фракталкин-ГФИ" или "ТИМП-фрак-ГФИ" относится к ТИМП, который непосредственно слит с доменом фракталкина, за которым следует последовательность ГФИ-линкера.
Термин "ПКР" обозначает почечно-клеточный рак, который, как полагают, является прогрессирующей опухолью с ограниченными терапевтическими вариантами из-за устойчивости опухоли к современным химиотерапевтическим средствам и радиации. ПКР служит модельной системой в настоящем изобретении, чтобы показать противораковую активность ГФИ-фиксированного ТИМП. Линиями клеток модели, применяемыми в настоящем изобретении, являются клеточные линии ПКР-26 и ПКР-53, которые были созданы от пациентов со стадией I и стадией IV рака клеток.
Термин "FAS" обозначает член семейства рецепторов фактора некроза опухолей/фактора роста нервов, которое вызывает апоптоз, независимый от ФНО-α. Другие сокращения, известные в технике для FAS, являются Apol (=белок1, вызывающий апоптоз) и CD95.
Термин "регенерация" вообще относится к восстановлению целостности травмированных или иначе поврежденных тканей. Этот термин может включать процессы заживления ран, репарации ткани и другие типы восстанавливающих активностей, встречающихся в местоположении, где физиологическое поражение и последующее повреждение ткани имели место.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Семейство ТИМП и белковая инженерия клеточных поверхностей
Тканевые ингибиторы металлопротеиназ (ТИМП) известны, как главные клеточные ингибиторы подсемейства матричной металлопротеиназы (ММП), показывающие различные степени эффективности против различных членов ММП, а также различные структуры экспрессии ткани и способы регулирования. ТИМП обычно модулируют активность растворимых ММП, связанных с матриксом и ассоциированных с клетками. Все четыре ММП млекопитающих имеют много широких общих черт, но показывают отличительные структурные особенности, биохимические свойства и структуры экспрессии, которые предполагают, что каждый ТИМП имеет особую функцию в естественных условиях.
Белок ТИМП-1 является наиболее широко экспрессируемым и изучаемым членом семейства ТИМП. Другие члены семейства ТИМП включают ТИМП-2, ТИМП-3 и ТИМП-4. Белки ТИМП не только разделяют общие структурные особенности, включая ряд сохраняющих остатков цистеина, которые образуют дисульфидные мостики, существенные для естественной белковой конформации (Brew и др., 2000), но у них также есть широко перекрывающиеся биологические активности. Сохраненная N-концевая область белков ТИМП необходима для функциональных ингибирующих активностей, в то время как дивергентные С-концевые области, как думают, модулируют эффективность ингибирования и связывания средств с ММП (Maskos & Bode, 2003). Однако, кроме их способности действовать как ингибиторы ММП, различные члены семейства ТИМП могут также проявлять дополнительные биологические активности, включая регулирование пролиферации и апоптоза в дополнение к модуляции ангиогенных и воспалительных реакций.
Как было найдено, ТИМП-1 ингибирует большинство ММП (кроме ММП-2 и -14) и избирательно ингибирует ММП-8. ТИМП-1 производится и секретируется в растворимой форме множеством типов клеток и широко распределяется по всему телу. Это сильно гликозилированный белок с молекулярной массой 28,5 kDa. ТИМП-1 ингибирует активные формы ММП и комплексы с проформой ММП9. Подобно ММП9, экспрессия ТИМП-1 чувствительна ко многим факторам. Повышенный синтез ТИМП-1 вызван широким разнообразием реагентов, которые включают: бета TGF, EGF, PDGF, FGFb, РМА, все-транс-ретиновая кислота (РА), IL1 и IL11.
ТИМП-2 является 21 kDa гликопротеином, который экспрессируется многими типами клеток. Он образует нековалентный, стехиометрический комплекс как со скрытым, так и с активным ММП. ТИМП-2 показывает предпочтение ингибированию ММП-2.
ТИМП-3 обычно связывается с ВКМ и ингибирует активность ММП-1, -2, -3, -9 и -13. ТИМП-3 показывает 30% аминокислотную гомологию ТИМП-1 и 38% гомологию ТИМП-2. Как показано, ТИМП-3, вызывает отделение трансформированных клеток от ВКМ и ускоряет морфологические изменения, связанные с трансформацией клеток.
Из-за высокого сродства связывания с ВКМ ТИМП-3 уникален среди ТИМП. Как было показано, ТИМП-3 вызывает отделение трансформированных клеток от ВКМ и ускоряет морфологические изменения, связанные с трансформацией клеток.
ТИМП-3 содержит домен, связывающий глюкозаминогликан (ГАГ), включающий шесть аминокислот (Lys30, Lys26, Lys22, Lys42, Arg20, Lys45), которые, как думают, ответственны за ассоциацию с поверхностью клетки. ТИМП-3 - это единственный ТИМП, который обычно ингибирует ТАСЕ (ФНОα-превращающий фермент), другую металлопротеазу, которая выделяет растворимый ФНО, и ответственен за переработку рецептора IL-6, таким образом, играя центральную роль в процессе заживления раны.
ТИМП-4 ингибирует все известные ММП, а предпочтительно ингибирует ММП-2 и -7. ТИМП-4 показывает 37% аминокислотную идентичность ТИМП-1 и 51% гомологию ТИМП-2 и ТИМП-3. ТИМП-4 секретируется внеклеточно, в основном, в сердечной и мозговой ткани и, кажется, функционирует тканеспецифичным образом относительно внеклеточного матриксного (ВКМ) гомеостаза.
Белковая инженерия клеточных поверхностей - это потенциально сильная технология, через которую поверхностным белковым составом клеток можно управлять без генной передачи. Заменой мРНК-производной последовательности кДНК, полученной из ГФИ-фиксированного белка, который содержит сигнальный домен ГФИ для участка карбоксильного конца белка интереса можно получить слитый конструкт, который кодирует ГФИ-фиксированный белок.
Этот подход предлагает многократные преимущества перед более традиционными подходами генной передачи. Например, метод применим к клеткам, которые трудно или невозможно трансфецировать (например, первичный капиллярный эндотелий, первичные клетки-мишени и т.д.). Количество белка, добавленного на поверхность клетки, можно регулировать и количественно определять (клеточный сортер с возбуждением флуоресценции (FACS) или иммунофлуоресценция). Кроме того, белки, многократно закрепленные ГФИ, могут быть последовательно или одновременно вставлены в те же самые клетки. Через молекулярную инженерию можно экспрессировать дополнительную эпитопную метку, который помогает очищать белок, а также контролировать реагент во время экспериментов. Средство может быть введено непосредственно в опухоль или околоопухолевую область и определен эффект селективного рекрутмента лейкоцитов на рост опухоли или FAS-опосредованный апоптоз.
Слитый конструкт ТИМП для лечения рака
Прогноз злокачественных опухолей, главным образом, зависит от их клинической и патологической стадии. В то время как большинство раков (например, первичные и вторичные опухоли) могут быть полностью хирургически удалены в большинстве случаев, повторная резекция продвинутой стадии рака часто невозможна, что связано с ранним рецидивом болезни и повышенной летальностью, обусловленной болезнью.
В частности, в раке молочной железы продвинутая стадия болезни с расширенным размером опухоли (>2 см) связана с возникновением отдаленного метастаза и ограниченным выживанием. Большой объем опухоли, как также полагают, является критическим параметром для наличия остаточного рака, который представляет также высокий риск местного рецидива и распространения отдаленного метастаза. Аналогично, опухоли головного мозга, такие как глиобластома (астроцитома IV степени), являются другой мыслимой целью для наблюдения опухоли, потому что полное хирургическое удаление почти всегда невозможно и местный рецидив встречается в 95% всех случаев в течение одного года после первичной хирургии.
Чтобы решить проблему, которая связана с остаточным раком, необходимо идентифицировать новый выбор терапии рака, который особенно полезен для лечения остаточного рака после неполной хирургической резекции.
В качестве решения настоящее изобретение обеспечивает ГФИ-фиксированный ТИМП, который может быть применен локально в края резекции, чтобы привлечь иммунные клетки и сосредоточить наблюдение остаточной опухоли на остаточных раковых клетках.
С этой целью белки ТИМП фиксируются ГФИ и, когда очищены и добавлены к раковым клеткам, включаются в их поверхностные мембраны и полностью функциональны. Заменяя эндогенную концевую последовательность 3'-мРНК на концевые последовательности 3'-мРНК белков, естественно закрепленных якорем ГФИ (то есть последовательность, которая направляет ГФИ-фиксацию), фактически любой белок ТИМП может быть экспрессирован как ГФИ-фиксированное производное.
В настоящем изобретении включение очищенного белка ГФИ-ТИМП в поверхностные мембраны линий опухолевой клетки демонстрируется инкубацией клеточных линий с очищенным ТИМП-1-ГФИ, ТИМП-1-муцин-ГФИ или рекомбинантный человеческим (rh)ТИМП-1 контрольным белком. Как детализировано ниже, поверхностная экспрессия с ГФИ-фиксированным ТИМП-1 приводит к сильному поверхностному сигналу для ТИМП-1.
Как используется здесь, термин "выделенный и/или очищенный" относится к in vivo выделению ДНК или молекулы полипептида из ее естественной клеточной среды, и из ассоциации с другими компонентами клетки, такими как нуклеиновая кислота или полипептиды так, чтобы она могла быть секвенирована, воспроизведена и/или экспрессирована. Например, "выделенная ГФИ-фиксирующая последовательность" является РНК или ДНК, содержащей больше 9, предпочтительно 36 и более предпочтительно 45 или больше последовательных нуклеотидных оснований, которые кодируют, по меньшей мере, часть связывающей последовательности, или ее вариант, или РНК или ДНК, комплементарные к ней, которые являются комплементарными или гибридизованными, соответственно, с РНК или ДНК, кодирующей связывающую последовательность, и остается устойчиво связанной при строгих условиях, как определено методами, известными в технике. Таким образом, РНК или ДНК "выделяют" так, что она является свободной, по меньшей мере, от одной загрязняющей нуклеиновой кислоты, с которой она обычно связывается в естественном источнике РНК или ДНК, и предпочтительно существенно свободной от любой другой РНК или ДНК млекопитающих.
Как используется здесь, термин "рекомбинантная нуклеиновая кислота", например "рекомбинантная последовательность ДНК", относится к нуклеиновой кислоте, например, к ДНК, которая была получена или выделена из любого адекватного источника ткани, который может быть впоследствии химически изменен in vitro так, что ее последовательность естественно не встречается, или соответствует естественно встречающимся последовательностям, которые не позиционированы так как они были бы позиционированы в геноме, который не был трансформирован экзогенной ДНК. Примером такой ДНК, "полученной" из источника, была бы последовательность ДНК, которую идентифицируют как полезный фрагмент в пределах данного организма, и которая затем химически синтезируется в чрезвычайно чистой форме. Примером такой ДНК, "выделенной" из источника, была бы полезная последовательность ДНК, которая вырезается или удаляется из указанного источника химическим способом, например применением рестрикционной эндонуклеазы, так, чтобы ею можно было далее управлять, например, усиливать для использования в изобретении в соответствии с известной методологией генной инженерии.
В отличие от обычных полипептидных якорей, которые имеют различные трансмембранные последовательности и соединяются с определенными цитоплазматическими удлинителями, эти фосфолипидоподобные якоря используют общую структуру как общий механизм мембранного прикрепления независимо от функции белка.
ГФИ-фиксированные единицы составляют из линейного гликана, содержащего фосфоэтаноламин, трех остатков маннозы и неацетилированного глюкозамина, связанного с инозитфосфолипидом. Их заранее готовят в эндоплазматической сети (ЭС) и добавляют к первичным трансляционным продуктам во время их транслокации через мембрану ЭС. Продукты, модифицированные ГФИ, гликозилируются в ЭС и комплексе Гольджи, и затем транспортируются к клеточной поверхности.
Предпочтительные ГФИ-связывающие последовательности, которые могут использоваться в настоящем изобретении, получают из ГФИ-якорей, которые выделяют из, например, ферментов, таких как щелочная фосфатаза, ацетилхолинэстераза, 5-нуклеотидаза (СО73); молекулы адгезии, такие как антиген, ассоциированный с функцией лимфоцитов (LFA-3; CD58), молекула адгезии невральных клеток (NCAM); комплементарные регуляторные белки, такие как фактор, ускоряющий распад (DAF или CD-55) или другие, такие как Fey рецептор типа III В (Fc-y-RIII или CD16b), Thy-1 (CD90), Qa-2, Ly-6A, мембранный ингибитор реактивного лизиса (MIRL или CD59). Для цели настоящего изобретения антиген, ассоциированный с функцией лимфоцитов (LFA-3), предпочтителен. Специалисты признают, что также любые другие из известных ГФИ-якорей могут использоваться в практике настоящего изобретения.
Для строительства ТИМП-ГФИ в слитом конструкте может использоваться либо последовательность ТИМП полной длины, либо функционально активный ее участок, который сохраняет активность ТИМП. Аналогично может использоваться также участок последовательности ГФИ, пока участок учитывает включение белка ТИМП в поверхностную клеточную мембрану раковых клеток.
Далее, описано много вариантов, касающихся слитых конструктов ТИМП, в результате чего конструкты были произведены и представлены для лечения рака и как средства в области регенеративной медицины. В первом варианте молекулу ТИМП выбирают из группы, состоящей из ТИМП-1, ТИМП-2, ТИМП-3 и ТИМП-4, и предпочтительно связывают с последовательностью ГФИ.
В другом предпочтительном варианте последовательность ГФИ составляет 36 аминокислот в длину.
В еще одном варианте молекулу ТИМП выбирают из группы, состоящей из ТИМП-1, ТИМП-2, ТИМП-3 и ТИМП-4, и связывают с доменом муцина или доменом фракталкина, за которым следует последовательность ГФИ.
В дальнейшем варианте белок ТИМП, который выбирают из группы, состоящей из ТИМП-1, ТИМП-2, ТИМП-3 и ТИМП-4, и связывают с последовательностью ГФИ или связывают с доменом муцина или доменом фракталкина, за которым следует последовательность ГФИ, укорачивают на С-конце. В предпочтительном варианте указанную молекулу ТИМП укорачивают до первых 50, 50-100 или 50-152 N-концевых аминокислотных остатков. Более предпочтительно, молекула ТИМП укорачивается до первых 152 N-концевых аминокислотных остатков и является молекулой ТИМП-1. Термин "укороченный" относится к нуклеиновокислотной или аминокислотной последовательности ТИМП, которая содержит меньше чем полное число нуклеиновокислотных оснований или аминокислотных остатков, найденных в естественной нуклеиновокислотной последовательности или белке ТИМП, или к нуклеиновокислотной или аминокислотной последовательности, которая была удалена из нежелательных последовательностей.
В еще одном дальнейшем варианте слитый конструкт ТИМП определяется последовательностью, выбранной из группы, состоящей из SEQ ID NOS: 1, 2, 3, 4 и 5. Полученный конструкт затем может быть экспрессирован в любой подходящей линии клеток или клетках хозяина, чтобы получить функциональный полипептид или белок ТИМП. С этой целью может использоваться любой из подходящих известных векторов или плазмид, чтобы экспрессировать ГФИ-фиксированные белки ТИМП по настоящему изобретению. Как описано более подробно ниже, клетки-мишени рака, обработанные белком ТИМП-ГФИ (и для контроля белком rhТИМП-1) узнавались белковыми конструктами и, как следствие, погибали из-за FAS-опосредованного апоптоза.
В предпочтительном варианте, и в качестве примера, один вектор, используемый для экспрессии слитых конструктов по настоящему изобретению, содержит промотор для человеческого фактора элонгации 1 альфа, за которым следует сайт многократного клонирования и внутренний сайт связывания рибосомы, который позволяет бицистронную экспрессию конструкта и дигидрофолатредуктазы (DHFR), используемой как маркер селекции (Mack, et al., P.N.A.S. USA 92:7021, 1995). 3'-Конец (карбоксильный конец) белка ТИМП связывается либо непосредственно с последовательностью ГФИ-связи (например, полученной из антигена 3, связанного с функцией лимфоцитов (LFA-3)), либо с муциноподобным доменом, выделенным из CXCL16 или фракталкина (CX3CL1), за которым следует ГФИ-сигнал. Как указано выше, эти области муцина в значительной степени состоят из остатков серин/треонин/глицин/пролина, которые, как показано, облегчают межклеточные взаимодействия. Образующиеся слитые конструкты трансфецировали в клетки яичника китайского хомячка с дефицитом дигидрофолатредуктазы (DHFR) и селекцию выполняли, как описано (Mack, et al., P.N.A.S. USA 92:7021-7025, 1995). В предпочтительном варианте трансфектанты могут быть подвергнуты воздействию метотрексата, чтобы увеличить скорость экспрессии амплификацией гена.
В дальнейшем варианте конструкта ТИМП-ГФИ может далее быть слит с доменом муцина, чтобы увеличить эффективность включения белков ТИМП-ГФИ в мембраны. Муцины - это мембраносвязанные или немембранные гликопротеиновые компоненты, которые были сначала идентифицированы в секретированной слизи, выстилающей поверхности эпителия желез. Мембраносвязанные муцины показывают гидрофобные последовательности или трансмембранные домены, ответственные за их фиксацию в двойном слое липида.
В настоящее время в общей сложности 21 ген получил название MUC: MUC1-2, MUC3A, MUC3B, MUC4, MUC5AC, MUC5B, MUC6-13, MUC15-20 (Moniaux N, и другие, 2004). Пять общих признаков муцина: (1) секреция в слой слизи, (2) О-гликопротеин высокого молекулярного веса, (3) наличие тандемных повторяющихся совокупностей, кодированное единственным и центрально размещенным большим экзоном, (4) наличие предсказанного пептидного домена, содержащего высокий процент остатков серина и треонина, и (5) сложный характер экспрессии мРНК. При одном исключении (MUC7) секреторные муцины (MUC2, MUC5AC, MUC5B и MUC6) обладают одним или несколькими доменами, подобными фактору фон Виллебрандта, пептидами, богатыми цистеином, которые функционируют в олигомеризации мономеров муцина и в заполнении секреторных пузырьков. Как правило, секретированные муцины экспрессируются исключительно специализированными эпителиальными клетками, секретируются в слизи, и показывают ограниченный характер экспрессии в пределах человеческого тела. Четыре секреторных муцина, также называемые гель-образующие муцины, имеют общую архитектуру с высоким уровнем подобия профактору Виллебрандта. Они, как также известно, укрывают пять доменов D из-за их гомологии доменам D фактора фон Виллебрандта.
Мембраносвязанные муцины состоят из MUC 1, MUC3A, MUC3B, MUC4, MUC11, MUC12, MUC16 и MUC17. Закрепленные на мембране муцины содержат SEA (белок спермы морского ежа, энтерокиназа и агрин) модуль, за исключением MUC4. MUC3A-B, MUC4, MUC11-12 и MUC17 содержат два - три домена, подобных эпидермальному фактору роста (EGF). Примерами мембраносвязанных муцинов, которые могут использоваться в настоящем изобретении, являются MUC1, MUC3, MUC4 и MUC12. В предпочтительном варианте изобретения применяют муцин-стебель, связанного с поверхностью хемокина CXCL16 или фракталкина (CX3CL1). CXCL16 является членом подсемейства СХС хемокинов. В отличие от других членов этой подгруппы CXCL16 структурно отличен и имеет четыре разных домена: домен хемокина, привязанный к клеточной поверхности через муциноподобный стебель, который в свою очередь присоединен к трансмембранным и эндоплазматическим доменам. Фракталкин (CX3CL1) имеет структуру, подобную структуре CXCL16, и как CXCL16, так и фракталкин действуют как молекулы адгезии, когда экспрессируются на клеточной поверхности и при отщеплении от клеточной поверхности клетки, растворимые хемокины действуют как хемоаттрактанты.
Предпочтительно, домен муцина вставляют между 3'-концом последовательности ТИМП и 5'-концом последовательности ГФИ-якоря любым из известных обычных методов генной инженерии. Полученный слитой конструкт ТИМП-муцин-ГФИ по изобретению затем может быть трансфецирован и экспрессирован в любой подходящей известной линии клеток или клетке хозяина. Специалист признает, что любые другие муцины или домены муцина являются подходящими для цели настоящего изобретения.
В предпочтительном варианте изобретения фракталкин, слитый с молекулой ТИМП, включает аминокислоты 100-342 из CX3CL1, за которыми следует последовательность ГФИ. В даже более предпочтительном варианте изобретения молекулой ТИМП является молекула ТИМП-1, укороченная до первых 152 N-концевых аминокислот (SEQ ID NO: 5).
Хотя применение ТИМП-1 (Bode & Maskos, 2003) для получения ГФИ-фиксированного белка ТИМП является предпочтительным в настоящем изобретении, специалист признает, что также другие белки ТИМП могут использоваться в практике настоящего изобретения. Дальнейшими примерами человеческих ТИМП, которые полезны, являются ТИМП-2, ТИМП-3 и ТИМП-4 (Mannello F. и др., 2001).
Используемые ТИМП получают из человеческих источников и применяют, чтобы лечить человеческие раковые клетки. Специалист также признает, что также гомологи ТИМП, в частности ТИМП-1, в других организмах, отличных от человека, будут иметь подобный эффект в лизисе опухолевых клеток. Например, в некоторых вариантах последовательность ТИМП-1, полученную из животного, такого как собака, кошка, мышь, кролик, корова или овца, или птицы, может использоваться для строительства слитого конструкта ТИМП-ГФИ по настоящему изобретению. Химера ТИМП-ГФИ будет затем применена к участку опухоли точно так же, как описано для человеческого индивидуума.
Опухоли и раковые клетки, которые могут быть обработаны ГФИ-фиксированным ТИМП, включают следующие раковые заболевания, (но не ограничиваются ими): рак молочной железы, почечный рак, рак простаты, лейкозы, семиномы, меланомы, тератомы, лимфомы, нейробластомы, глиомы, ректальный рак, внутриматочный рак, почечный рак, рак надпочечников, рак щитовидной железы, рак крови, рак кожи, рак мозга, рак шейки матки, кишечный рак, рак печени, рак толстой кишки, рак желудка, рак кишки, желудочно-кишечный рак, рак лимфатических узлов, рак пищевода, колоректальный рак, рак поджелудочной железы, рак носоглотки/«ухо, нос и горло» (ENT), рак матки, овариальный рак и рак легкого, и их метастазы.
Для лечения остаточного рака ТИМП-ГФИ может быть назначен и нанесен на место резекции массы опухоли у "высокорисковых" раковых больных с высоким риском остаточных раковых клеток и повышенным уровнем местного рецидива и у больных с очевидной остаточной опухолью из-за продвинутой стадии болезни или местной неоперабельности. Предпочтительно, слитый конструкт назначают при концентрации белка 0,5-5 мкг/мл, более предпочтительно 0,5-1 мкг/мл или 1-2 мкг/мл. Концентрация приблизительно 1 мкг/мл ТИМП-ГФИ или ТИМП-муцин-ГФИ наиболее предпочтительна. Белок может назначаться человеку любыми применимыми путями. Предпочтительно, лечение выполняют во время хирургии таким образом, что слитый конструкт распыляют в рану или вводят в области, которые не доступны для хирургии. С этой целью слитый конструкт ГФИ-фиксированный ТИМП по настоящему изобретению может быть компонентом фармацевтического состава или медикамента, который далее включает один или больше традиционно известных носителей, разбавителей и наполнителей.
Как можно заключить из примеров ниже, ГФИ-фиксированный ТИМП, кажется, вызывает противоопухолевую активность, то есть лизис опухолевых клеток, не апоптозом, вызванным клетками цитотоксических Т лимфоцитов и естественными клетками-киллерами (ЦТЛ и ЕКК), но скорее вторым путем, который включает FAS/ СD95-опосредованный апоптоз (для дальнейших подробностей, см. примеры 5 и 6; фигуры 5 и 6). Кроме того, в то время как много опухолевых клеток устойчивы к FAS-опосредованному апоптозу, обработка ТИМП-1-ГФИ, но не контрольным rhТИМП-1, делает линии клеток чувствительными к FAS-опосредованному апоптозу.
Было также найдено, что обработка белком ТИМП-1-ГФИ снижает BCL2 и повышает экспрессию белка ВАХ. Белки BCL2 представляют семейство белков, вовлеченных в контроль апоптоза. Некоторые члены этого семейства (такие как BCL2 и BCL-XL) являются антиапоптотическими, в то время как другие (такие как Bad или ВАХ) являются проапоптотическими. Чувствительность клеток к апоптотическим стимулам зависит от баланса между про- и антиапоптотическими членами семейства BCL2. Кроме того, эффект обработки ТИМП-1-ГФИ на экспрессию BCL2 и ВАХ был определен, и показано, что обработка раковых клеток ТИМП-1-ГФИ увеличивало экспрессию проапоптотического ВАХ и уменьшало экспрессию антиапоптотического BCL2.
Подобные результаты были получены для слитых конструктов ТИМП, кодированных SEQ ID NO: 1, 2, 3, 4 и 5 соответственно.
Суммируя вышесказанное, пригодность методологии производить от микрограммовых до миллиграммовых количеств рекомбинантных ГФИ-фиксированных белков ТИМП в соединении с включением этих молекул в поверхности раковых клеток обеспечивает эффективный метод для лечения рака.
Слитые конструкты ТИМП для применения в регенеративной медицине
Кроме того, слитый конструкт по настоящему изобретению подходит для применения в регенеративной медицине, особенно в области заживления ран. Как описано выше, белки ТИМП, которые соединены с ГФИ или с муцин-ГФИ или фракталкин-ГФИ, эффективно включаются в клеточную поверхностную мембрану, где они сосредотачивают функциональные домены на клеточных поверхностях независимо от белково-белковых взаимодействий. Слитые конструкты ТИМП по настоящему изобретению обычно весьма устойчивы и показывают усиленные и новые биологические активности.
Как описано выше, как ММП, так и ФНО-альфа-превращающий энзим (ТАСЕ) играют критическую роль в процессе заживления ран. Повышенные уровни ММП связаны с различными нарушениями излечивания ран, среди прочего с наличием хронической раны. Поскольку ТИМП является естественным ингибитором ММП, слитые конструкты по настоящему изобретению могут также использоваться как эффективные терапевтические средства для того, чтобы управлять процессом заживления раны, например, в регенеративной медицине и подходят для того, чтобы лечить нарушения, характеризующиеся повышенным уровнем ММП.
Таким образом, настоящее изобретение предоставляет средства и методы, пригодные для использования в регенеративной медицине и/или, чтобы лечить нарушения, характеризующиеся повышенным уровнем ММП. В предпочтительном варианте изобретения слитые конструкты по настоящему изобретению используются, чтобы лечить или предотвращать чрезмерное рубцевание и патологическое заживление ран, включая келоидные или гипертрофические рубцующиеся и/или хронические раны. В дальнейшем предпочтительном варианте изобретения слитый конструкт по настоящему изобретению используется, чтобы ингибировать или предотвращать формирование рубцовой ткани.
Типичная реакция на заживление раны характеризуется движением специализированных клеток в участок раны. Тромбоциты и воспалительные клетки обычно являются первыми клетками, которые прибывают на место раны, и эти молекулы обеспечивают важные функции и химические сигналы, включая цитокины, которые необходимы для притока соединительных тканевых клеток и других заживляющих факторов. Термин "рана" означает разрушение нормальной физиологической структуры и функции. Таким образом, процесс заживления раны относится к сложной и динамической последовательности событий, в конечном счете, приводящей к восстановлению физиологической непрерывности и функции.
Когда рана заживает, рубец обычно развивается на его месте. В течение нормального заживления раны восстанавливаются простые ткани, такие как жир, соединительная ткань и эпителий.
Однако, поскольку кожа - более сложный орган, который образуется из двух зародышевых слоев, она заживает через образование, главным образом, фиброзной ткани, то есть рубца.
При нормальном заживлении раны протеолитическая активность ММП регулируется различными механизмами, включая транскрипцию генов, продукцию фермента и местную секрецию эндогенных ингибиторов ТИМП. Во время заживления раны существует физиологический баланс между активностями ММП и ТИМП. Однако матриксные металлопротеазы, как известно, имеют повышенные уровни в хронических ранах, и такие высокие концентрации ММП, как известно, ухудшают процесс заживления раны. Различные типы клеток, включая макрофаги, фибробласты, нейтрофилы, эпителиальные клетки и эндотелиальные клетки, синтезируют ММП в присутствии определенных биохимических сигналов, таких как воспалительные цитокины. ММП способны переваривать почти все компоненты внеклеточного матрикса, выбирая необходимый баланс между белком, разрушающим активности ММП и другой клеточной активностью, которая синтезирует и депонирует компоненты белка ткани.
Фигура 8 обеспечивает краткий обзор процесса модернизации ткани, фиброза и других факторов, вовлеченных в коррекцию этого процесса. После острой и хронической воспалительной реакции на специфическое повреждение, паразитную инфекцию или аутоиммунную реакцию, экспрессируются и секретируются фиброгенные факторы, таким образом, приводя к активации фибробластов и кератиноцитов. Эти фиброгенные факторы включают, среди прочего, TGF-β, IL-1β, IL-1α, MOB, TGF-α, IL-4, IL-13, bFGF, ФНО-α и PDGF-BB. Возможно, двумя, наиболее важными из этих цитокинов являются: ФНО, который является митогенным для фибробластов и вызывает ангиогенез и секретируется макрофагами, тучными клетками и Т лимфоцитами; и трансформирующий фактор роста (TGF-α), который является митогенным для кератиноцитов и фибробластов, стимулирует миграцию кератиноцитов, и секретируется макрофагами, Т лимфоцитами и кератиноцитами. Стадия, включающая ФНО и секрецию TGF, отмечает переход от воспалительной фазы процесса заживления раны к процессу реконструкции ткани, то есть к фазе пролиферации.
При активации фибробласты секретируют интерлейкин-6 (IL-6), который в комбинации с TGF-β приводит к пролиферации фибробластов. TGF-β также вызывает дифференцирование фибробластов. Результатом этих процессов пролиферации и дифференцирования является общее увеличение коллагена, фибронектина, ТИМП, ММП, так же как других белков ВКМ и приводит к увеличению продукции ВКМ и уменьшению обновления ВКМ.
Настоящее изобретение основано на неожиданной находке, что описанные слитые конструкты, то есть белки ТИМП или их мутанты, слитые с якорем ГФИ, якорем муцин-ГФИ или фракталкин-ГФИ, могут использоваться как сильное средство, чтобы влиять на уровень экспрессии и/или активность цитокинов и других важных ферментов, вовлеченных в процесс заживления раны. Таким образом, настоящее изобретение предлагает эффективные регенеративные средства и методы управления процессом заживления раны (например, чтобы влиять на, ингибировать или предотвращать формирование рубцовой ткани) и для того, чтобы лечить другие известные дисфункции, связанные с процессом заживления раны.
Патологическое заживление раны
Келоидное и гипертрофическое рубцевание характеризуется накоплением лишнего коллагена, и отличаются друг от друга их физическим видом. Как келоидные, так и гипертрофические рубцы - это раны, которые заживают сверхбыстро на внешней поверхности кожи. Келоидный рубец типично продолжает увеличиваться вне размера и формы раны, в то время как гипертрофический рубец увеличивается в пределах физических границ первоначальной раны. Гипертрофическое рубцевание обычно заметно вскоре после повреждения ткани, тогда как келоидные рубцы могут сформироваться в пределах года после ранения. Однако почти все случаи патологического рубцевания связаны с физиологическими повреждениями, включая татуировки, ожоги, инъекции, укусы, прививки, травмы, хирургия или инфекция.
Гипертрофические рубцы и келоиды могут быть описаны как изменения типичного процесса заживления раны. В типичной ране анаболические и катаболические процессы достигают равновесия спустя приблизительно 6-8 недель после первоначального ранения. Во время созревания рубца прочность кожи на растяжение улучшается, поскольку волокна коллагена постепенно поперечно связываются. В этой точке рубец обычно гиперемирован и может быть утолщенным. Однако начальная рубцовая ткань имеет тенденцию спадать постепенно в течение месяцев, давая более зрелый рубец, который является типично плоским, белым, гибким и возможно вытянутым внешне. Когда есть дисбаланс между анаболическими и катаболическими фазами процесса заживления раны, больше коллагена производится, чем разрушается, и рубец может, поэтому, расти во всех направлениях.
Единый оптимальный метод для лечения гипертрофической и келоидной рубцовой ткани еще не разработан, таким образом, норма рецидива этих патологических рубцов значительна.
В итоге, цитокины и определенные ферменты, включая ММП и ТИМП, играют критическую роль в процессе заживления раны и в формировании рубцовой ткани. Кроме того, патологическая экспрессия ММП и цитокинов часто связывается с патологическим заживлением раны.
Таким образом, настоящее изобретение предлагает эффективные регенеративные средства и методы управления процессом заживления раны и/или лечения дисфункций, обычно связанных с заживлением раны. Определенно, слитые конструкты по настоящему изобретению могут быть использованы, чтобы эффективно управлять, изменять, ингибировать или даже предотвращать эти нежелательные процессы. Слитые конструкты по настоящему изобретению могут быть составлены как фармацевтические препараты и применены на месте ранения. В одном варианте место ранения создается хирургией, ожогом, инъекцией, укусом, прививкой, травмой, хирургией или инфекцией. В другом варианте слитые конструкты, применяемые для приготовления медикамента, который будет нанесен на место ранения, выбирают из группы, состоящей из ТИМП-1-ГФИ, ТИМП-2-ГФИ, ТИМП-3-ГФИ, ТИМП-4-ГФИ, ТИМП-1-муц-ГФИ, ТИМП-2-муц-ГФИ, ТИМП-3-муц-ГФИ, ТИМП-4-муц-ГФИ или их мутантов.
Составление конструктов ТИМП и способы введения
Фармацевтические препараты на основе конструктов ТИМП по настоящему изобретению могут быть составлены обычным способом, используя один или больше физиологически приемлемых носителей или наполнителей. Методы и составы обычно могут быть найдены в Remrnington's Pharmaceutical Sciences, Meade Publishing Co., Eastone, Pa. В целях инъекции соединения по изобретению могут быть рецептированы в жидком растворе, предпочтительно в физиологически совместимом буфере, таком как раствор Хенка или раствор Рингера. Кроме того, соединения могут быть составлены в твердой форме и повторно растворены или суспендированы немедленно перед использованием. Лиофилизированные формы также пригодны.
В дополнение к этим составам соединения могут также быть составлены как депо-препараты. Эти долгодействующие депо-препараты могут быть введены имплантацией (например, подкожно или внутримышечно) или инъекцией. Таким образом, составы могут быть составлены с подходящими полимерными или гидрофобными материалами (например, как эмульсия в приемлемом масле) или как ионообменная смола, или как слаборастворимое производное, такое как слаборастворимая соль. Другие подходящие системы доставки включают микросферы, которые предлагают возможность местной и неразрушающей доставки препаратов в широком временном интервале. Эта специфическая технология использует микросферы, имеющие прекапиллярный размер, которые может быть введен через коронарный зонд в любую выбранную часть ткани, например сердце или другие органы, не вызывая результирующее воспаление или ишемию. Вводимое терапевтическое средство медленно выделяется из этих микросфер и легко принимается клетками, присутствующими в окружающей ткани (например, поврежденными или раковыми клетками).
Для местного введения олигомеры по изобретению могут быть составлены в мази, бальзамы, гели или кремы, общеизвестные в технике. Моющий раствор, содержащий олигомер, может использоваться локально, чтобы лечить рану или воспаление или вообще ускорить процесс заживления.
Конструкты ТИМП по настоящему изобретению могут быть объединены при получении медикамента так, что образующийся медикамент включает больше чем один, предпочтительно два и еще более предпочтительно три различных конструкта ТИМП. С этим подходом усиленные и новые биоактивности различных членов семейства ТИМП могут быть предпочтительно объединены и нацелены на поверхность клетки, что может привести к синергетическому эффекту. Например, слитый конструкт ТИМП-1 ингибирует большинство ММП, кроме ММП-2 и ММП-14. Поэтому любой из конструктов ТИМП-1 может быть объединен с любым из конструктов ТИМП-2 или ТИМП-4, которые оба предпочтительно ингибируют ММП-2. Поэтому посредством этого объединения более полное ингибирование семейства ММП может быть достигнуто.
В одном варианте изобретения составы по настоящему изобретению, следовательно, включают конструкт ТИМП-1, конструкт ТИМП-2 и/или конструкт ТИМП-4. В предпочтительном варианте изобретения состав включает конструкт ТИМП-1, выбранный из группы, состоящей из укороченного ТИМП-1-ГФИ, кодированного SEQ ID NO: 1, укороченного ТИМП-1-фрак-ГФИ, кодированного SEQ ID NO: 5, и укороченного ТИМП-1-муц-ГФИ, кодированного SEQ ID NO: 2, и конструкты ТИМП-2 и/или ТИМП-4. Предпочтительно, конструкт ТИМП-2 кодируется SEQ ID NO: 3.
В дальнейшем варианте состав включает конструкт ТИМП-3 по настоящему изобретению, предпочтительно кодированный SEQ ID NO: 4, который ингибирует ТАСЕ (ФНОα-превращающий фермент) вместе с, по меньшей мере, одним из конструктов ТИМП, выбранным из группы, состоящей из укороченного ТИМП-1-ГФИ, кодированного SEQ ID NO: 1, укороченного ТИМП-1-фрак-ГФИ, укороченного ТИМП-1-муц-ГФИ, конструкта ТИМП-2 и ТИМП-4. Предпочтительно, конструкты ТИМП-1 и ТИМП-2 кодируются SEQ ID NO: 1,2, 3 и 5 соответственно.
Краткое описание чертежей
Фигура 1. Включение ТИМП-1-ГФИ в клеточные мембраны ПКР
А. Чтобы продемонстрировать повторное включение белка ГФИ-ТИМП-1 в клеточные мембраны, очищенный ТИМП-1-ГФИ или контрольный rhТИМП-1 добавляли к нативным клеткам ПКР-26, ПКР-53 и А498. ТИМП-1 обнаруживали на клеточной поверхности клеточным сортером с возбуждением флуоресценции (FACS анализ). Серые гистограммы - это изотип-зависимое окрашивание, линейные гистограммы представляют окрашивание антителами к ТИМП-1.
B. Чтобы продемонстрировать ГФИ-связь после инкубации с 200 нг/мл или 700 нг/мл ТИМП-1-ГФИ или rhТИМП-1, клетки обрабатывали фосфолипазой С (PLC) (60 нг/мл) и затем анализировали FACS. Серые гистограммы представляют изотипный контроль.
C. Иммуносорбентный анализ (ELISA) человеческого ТИМП-1 применяли, чтобы определить количество белка ТИМП-1, выделенного клетками ПКР, обработанными ТИМП-1-ГФИ (как показано в В), после обработки фосфолипазой С (PLC).
Фигура 2. ТИМП-1-ГФИ ингибирует выделение проММП-2 и проММП-9 из клеток ПКР-53
Зимографию применяли, чтобы изучить секрецию белков ММП-2 и ММП-9 из ПКР-53. Клетки обрабатывали повышенным количеством ТИМП-1-ГФИ или контрольного rhТИМП-1, и после 48 часов свободную от сыворотки культуральную надосадочную жидкость (супернатант) удаляли и анализировали желатиназной зимографией.
Фигура 3. Поверхностная экспрессия ММП после обработки ТИМП-1-ГФИ
После инкубации клеток ПКР-53 с 700 нг/мл белка ТИМП-1-ГФИ в течение 24 часов анализ FACS выполняли, используя определенные антитела, направленные против: (А) ТИМП-1, ММП-1, ММП-2, ММП-3, ММП-7, ММП-8 и ММП-9, или (В) ММП-12, ММП-13, ММП-14, ММП-16, HLA-A2 (НВ82), pan HLA Class 1 (W6/32) и ICAM-1. Как дополнительный контроль ТИМП-1-ГФИ отщепляли от поверхности после одного часа обработки фосфолипазой С (PLC). Серые гистограммы - это изотип-зависимое окрашивание, линейные гистограммы представляют образцы, обработанные ТИМП-1-ГФИ.
C. Секрецию ряда ММП из ПКР-53 проверяли, используя Вестерн-блоттинг и моноклональные антитела, направленные против ММП-1, ММП-3, ММП-7, ММП-8, ММП-12 и ММП-13. Питательные среды (свободные от сыворотки) были взяты через 24 часа после обработки ПКР-53 либо rhТИМП-1, либо ТИМП-1-ГФИ (700 нг/мл) и сравнивались с необработанными контрольными клетками.
D. Эффект секвестирования ММП клеточной поверхностью на проникновение ПКР-53 через модельную мембрану Matragel был оценен. Оптимальная миграция клеток ПКР-53 к фактору роста эндотелия сосудов (VEGF) (4 нг/мл) была установлена как начальная или "ноль", а величину 100% ингибирования устанавливают для величины миграции необработанных клеток ПКР-53 без VEGF сигнала. Клетки ПКР-53 предварительно обрабатывались rhТИМП-1 или ТИМП-1-ГФИ (350 нг/мл или 700 нг/мл). После одного часа клетки промывали и вносили в камеру миграции. Влияние на миграцию затем оценивали. Представленные данные представляют среднее число из четырех лунок и двух экспериментов.
Фигура 4. Влияние белка rhТИМП-1 и ТИМП-1-ГФИ на пролиферацию линий ПКР
Влияние увеличивающихся уровней белков ТИМП-1-ГФИ или контрольного rhТИМП-1 на пролиферации ПКР-53 (А), А498 (В) и ПКР-26 (С) измеряли, используя пробу МТТ. МТТ (3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолийбромид) добавляли после 24 часов, 48 часов или 72 часов, как указано.
Фигура 5. ТИМП-1-ГФИ не влияет на восприимчивость ПКР к перфорин-опосредованному апоптозу
Клетки ПКР оставили необработанными (■), обрабатывали ТИМП-1-ГФИ (700 нг/мл) (0) или белком rhТИМП-1 (о) в течение 24 часов и инкубировали либо с цитотоксическими Т лимфоцитами CTL JB4 (А), либо с естественными клетками-киллерами, NK-линиями (В) (NKL для ПКР-53 и А498 или NK-92 для ПКР-26). Показаны представительные примеры трех независимых экспериментов с похожими результатами.
Фигура 6. ТИМП-1-ГФИ не увеличивает экспрессию FAS, но делает клетки, чувствительными к FAS-опосредованому апоптозу
А. Клетки ПКР-53, ПКР-26 или А498 обрабатывали или не обрабатывали ТИМП-1-ГФИ (700 нг/мл) или rhТИМП-1 в течение 24 часов, и окрашивали человеческим анти-FAS (L-958) и анализировали на поверхностную экспрессию FAS поточной цитометрией. Изотип-зависимая окраска моноклональными антителами показана как серые гистограммы.
Три линии клеток ПКР обрабатывали ТИМП-1-ГФИ или rhТИМП-1 с последующей инкубацией L-957. Связывание аннексин V-флуоресцеинизотиоцианата (FITC) применяли, чтобы обнаружить жизнеспособный и ранний апоптоз поточной цитометрией.
Низкий уровень апоптоза в клетках ПКР-53 в (В) подтверждали, используя более чувствительный цитоплазматический нуклеосомный иммуносорбентный анализ ELISA. Растущие уровни апоптоза были обнаружены после инкубации L-957 с увеличивающимися уровнями ТИМП-1 -ГФИ-, но не rhТИМП-1.
Фигура 7. Экспрессия BCL-2 и ВАХ в ПКР с последующей обработкой ТИМП-1-ГФИ или rhТИМП-1, анализированная внутренним окрашиванием FACS и Вестерн-блоттингом.
Клетки ПКР-53 (А), ПКР-26 (В) и А498 (С) преинкубировали с ТИМП-1-ГФИ или rhТИМП-1 в течение 24 часов, затем обрабатывали L-957, активируя FAS-антитело в течение дополнительных 16 часов. Клетки затем анализировали моноклональными антителами против BCL-2 и против ВАХ, используя поточную цитометрию. Параллельно белки извлекали и измеряли Вестерн-блоттингом. Сигналы, полученные из ВАХ и BCL-2, нормализовали до уровней β-актина с последующей денситометрией. Результаты FACS представлены как гистограммы с величинами в круглой скобке, соответствующими интенсивности флуоресценции (MFI) либо BCL-2 или ВАХ, либо соответствующих изотипных антител.
Фигура 8. Обзор модернизации ткани и фиброза
Схематическая диаграмма, изображающая сложное взаимодействие факторов, вовлеченных в тонкий баланс продукции ВКМ и обновления во время процесса заживления раны.
Фигура 9. Влияние слитых конструктов ТИМП на продукцию фибронектина фибробластами в присутствии rhТИМП-1
Сливные фибробласты культивировали в присутствии или отсутствии rhТИМП-1 и ТИМП-1-ГФИ; экспрессированный и секретированный фибронектин определяли количественно Вестерн-блоттинг анализом, используя антиантитела на фибронектин (β-актин служил контролем). rhТИМП-1 (при 700 нг/мл) не приводил к какому-либо значительному уменьшению экспрессии фибронектина, в то время как ТИМП-1-ГФИ (при 700 нг/мл) сильно снижал фибронектин, который секретировался фибробластами.
Фигура 10. Влияние слитого конструкта ТИМП на продукцию фибронектина фибробластами в присутствии ФНО-α
Фибробласты культивировали в присутствии (Фигура 10А) или отсутствии (Фигура 10В) 10 нг/мл ФНО-α, активирующего фибробласт, вместе с ТИМП-1-ГФИ (350 нг/мл) или ТИМП-1-ГФИ (700 нг/мл), денатурированными ТИМП-1-ГФИ и rhТИМП-1-ГФИ соответственно. При концентрации 350 нг/мл ТИМП-1-ГФИ транскрибированная РНК фибронектина значительно снижалась, независимо от того, присутствовал ли ФНО-α в питательной среде.
Фигура 11. Влияние слитых конструктов ТИМП на продукцию IL-6 фибробластами
РНК фибронектина, транскрибированная фибробластами, была оценена Нозерн-блоттинг анализом, используя зонд для РНК интерлейкина-6 (IL-6). Таким образом, фибробласты культивировали в присутствии (Фигура 11А) или отсутствии (Фигура 11В) 10 нг/мл ФНО-α, активирующего фибробласт, вместе с ТИМП-1-ГФИ (350 нг/мл) или ТИМП-1-ГФИ (700 нг/мл), денатурированными ТИМП-1-ГФИ и rhТИМП-1-ГФИ соответственно. При концентрации 350 нг/мл ТИМП-1-ГФИ транскрибированная РНК IL-6 значительно снижалась, независимо от того, присутствовал ли ФНО-α в питательной среде.
Фигура 12. Влияние слитых конструктов ТИМП на продукцию коллагена фибробластами
РНК фибронектина, транскрибированная фибробластами, была оценена Нозерн-блоттинг анализом, используя зонды для Коллагена 1А1 (Фигура 12А и В), Коллагена 4А2 (Фигура 12С и D), Коллагена 16А1 (Фигура 12Е и F) соответственно. Таким образом, фибробласты культивировали в присутствии (Фигура 12А, С, Е) или отсутствии (Фигура 12В, D, F) 10 нг/мл ФНО-α, активирующего фибробласт, вместе с ТИМП-1-ГФИ (350 нг/мл) или ТИМП-1-ГФИ (700 нг/мл), денатурированными ТИМП-1-ГФИ и rhТИМП-1-ГФИ соответственно. При концентрации 350 нг/мл ТИМП-1-ГФИ все три транскрибированных РНК коллагена значительно снижались, независимо от того, присутствовал ли ФНО-α в питательной среде.
Фигура 13. Влияние слитых конструктов ТИМП на продукцию TGF-β фибробластами
РНК фибронектина, транскрибированная фибробластами, была оценена Нозерн-блоттинг анализом, используя зонд для TGF-β. Таким образом, фибробласты культивировали в присутствии (Фигура 13А) или отсутствии (Фигура 13 В) 10 нг/мл ФНО-α, активирующего фибробласт, вместе с ТИМП-1-ГФИ (350 нг/мл) или ТИМП-1-ГФИ (700 нг/мл), денатурированными ТИМП-1-ГФИ и rhТИМП-1-ГФИ соответственно. При концентрации 350 нг/мл ТИМП-1-ГФИ почти никакая PHK TGF-β не может быть обнаружена, независимо от того, присутствовал ли ФНО-α в питательной среде и независимо от содержания фетальной телячьей сыворотки (FCS) вереде.
ПРИМЕРЫ
В следующих примерах противоопухолевые эффекты ГФИ-фиксированного ТИМП на раковые клетки описаны более подробно. Хотя описанные эксперименты выполнены с человеческим ТИМП-1, изобретение не должно быть ограничено этим типом ТИМП.
В следующих примерах ГФИ-якорь соединяли с ТИМП-1, чтобы сосредоточить определенные концентрации этого ингибиторного белка на поверхности трех клеточных линий почечно-клеточных раков (ПКР-26, ПКР-53 и А498) независимо от белково-белковых взаимодействий клеточной поверхности. Как показано в дальнейшем, экзогенно добавленный ТИМП-1-ГФИ эффективно внедрялся в мембрану клетки ПКР и резко изменял ассоциацию ММП с поверхностью клетки. Обработка ТИМП-1-ГФИ ингибировала пролиферацию ПКР и делала обычно FAS-устойчивые клетки ПКР чувствительными к FAS-индуцированному апоптозу, но не изменяла перфорин-опосредованный лизис цитотоксическими эффекторными клетками. Повышенная чувствительность к FAS-опосредованному апоптозу коррелировала со сдвигом в балансе про- и антиапоптотических белков семейства BCL-2.
Клеточные линии ПКР-26 (Schendel и другие, 1993) и ПКР-53 основаны от местных больных со стадией 1 и стадией IV светлоклеточных раков соответственно. Таким образом, они представляют две клинических крайности ПКР. Противоопухолевые цитотоксические Т лимфоциты выделяли из опухоли обоих больных. Хотя эти естественные эффекторные клетки неспособны регулировать рост опухоли in vivo, окрашивание поверхностными маркерами ПКР-26 и ПКР-53 выявило хорошую поверхностную экспрессию главного комплекса гистосовместимости (ГКГ) класса 1 и обе линии, как было показано, индуцируют алло- и антиопухолевые специфичные ЦТЛ in vitro ((Schendel и другие, 2000) и DJS, неопубликованное наблюдение). А498 были первоначально изолированы из опухоли 52-летнего мужчины и являются хорошо изученным примером ПКР (Giard и др., 1973).
Пример 1
Включение экзогенно добавленного ТИМП-1-ГФИ в поверхность ПКР-53
Белок ГФИ-фиксированный ТИМП-1 производили и выделяли, как ранее описано (Djafarzadeh и другие, 2004). Включение очищенного белка ГФИ-ТИМП-1 в поверхностные мембраны клеточных линий ПКР-53, ПКР-26 или А498 ПКР было продемонстрировано инкубацией клеточных линий с 700 нг/мл очищенного ТИМП-ГФИ или рекомбинантного человеческого контрольного белка (rh)ТИМП-1 в течение одного часа. Поверхностно-связанный белок ТИМП-1 затем детектировали, используя FACS анализ.
(Фигура 1А). Добавление контрольного rhТИМП-1 не приводило к изменению в сдвиге FACS, однако, ГФИ-фиксированный ТИМП-1 приводил к сильному поверхностному сигналу для ТИМП-1.
Чтобы продемонстрировать, что экзогенно добавленный белок был закреплен якорем ГФИ, клетки ПКР-53 сначала инкубировали с белком ТИМП-1-ГФИ (200 или 700 нг/мл) и затем обрабатывали 60 нг/мл фосфолипазы С (PLC). Анализ FACS продемонстрировал полную потерю сигнала ТИМП-1 клеточной поверхности после обработки PLC (Фигура 1В). Чтобы измерить эффективность интеграции ТИМП-1-ГФИ, ТИМП-1, освобожденный от мембраны, собирали в промывных буферах и определяли количественно, используя ТИМП-1 специфический иммуносорбентный анализ ELISA (Фигура 1С). Результаты показывают, что 66% исходного антигена ТИМП-1 возвращали из образца 200 нг/мл, в то время как 31% мог быть выделен из инкубации 700 нг/мл.
Пример 2
Белок ТИМП-1-ГФИ блокирует выделение проММП-2 и проММП-9 из ПКР-53
Увеличенная экспрессия ММП-2 и ММП-9 коррелирует с плохим прогнозом ПКР (Hemmerlein и другие, 2004). Стадия IV клеточной линии светлоклеточного рака ПКР-53 нерегулируемо секретирует проММП-2 и проММП-9 (Фигура 2). Эффект увеличивающихся поверхностных уровней ТИМП-1 на нерегулируемое выделение белков ММП-2 и ММП-9 проверяли желатиназной зимографией (Djafarzadeh и другие, 2004; Klier и другие, 2001). Белок rhТИМП-1 при 600 или 1200 нг/мл не имел никакого эффекта на секрецию проММП-2 или проММП-9. Напротив, начиная с 10 нг/мл, обработка ТИМП-1-ГФИ показала концентрационно-зависимое снижение выделения проММП-2 и проММП-9 в питательные ростовые среды.
Пример 3
Обработка ТИМП-1-ГФИ приводит к увеличению поверхностной экспрессии матриксных металлопротеиназ
На основе результатов экспериментов желатиназной зимографии, возможно, что ТИМП-1-ГФИ может действовать, чтобы связывать ММП на поверхности клетки. ТИМП-1 связывает большинство активных форм ММП, исключая ММП-14 и ММП-16 (Brew и другие, 2000; Lang и другие, 2004). После инкубации ПКР-53 с 700 нг/мл белка ТИМП-1-ГФИ в течение 24 часов анализы FACS, используя определенные антитела к ММП-1, ММП-2, ММП-3, ММП-7, ММП-8, ММП-9, ММП-12, ММП-13, ММП-14, ММП-15 и ММП-16, показали, за исключением ММП-14, увеличение среднеканальной интенсивности флюоресценции (MFI) для каждого из ММП. Контрольный белок rhТИМП-1 не имел никакого очевидного эффекта на сигнал FACS (данные не показаны). На поверхностную экспрессию других белков, включая ГКГ класс 1 (pan class 1 и HLA-A2) и ICAM-1, не влияла обработка ТИМП-1-ГФИ. Расщепление ПКР-53, обработанных ТИМП-1-ГФИ, фосфолипазой С (PLC) в течение одного часа (как выполнено в Фигуре 1) не показала увеличения ММП (Фигуры 3А и 3В). 3636 Накопление ММП на клеточной поверхности отражало сокращение секреции проММП-2 и проММП-9, показанное на Фигуре 2. Чтобы далее проверить эту очевидную блокаду выделения ММП, эксперименты Вестерн-блоттинга были выполнены, используя моноклональные антитела, направленные против ММП-1, ММП-3, ММП-7, ММП-8, ММП-12 и ММП-13 на питательных средах (свободных от сыворотки) полученный из контрольных клеток ПКР-53 или клеток, обработанных 24 часа 700 нг/мл rhТИМП-1 или ТИМП-1-ГФИ (Фигура 3С). Присутствие каждой из ММП детектировали в питательных средах из контрольных клеток ПКР-53. Инкубация с rhТИМП-1 не устраняла секрецию ММП. Напротив, ТИМП-1-ГФИ, как оказалось, полностью блокирует выделение каждой изученной ММП.
Чтобы оценить эффект этого поверхностного накопления ММП на способность клеток заселять ВКМ, способность клеток к миграции/инвазии проверяли, используя анализ в модифицированной камере Бойдена с мембранами, покрытыми ВКМ. Полная способность клеток ПКР-53 вторгаться во ВКМ не была очень явной (данные не показаны). Чтобы увеличить инвазию, применяли повышенные уровни фактора роста эндотелия сосудов (VEGF) в более низких ячейках и эксперименты продолжали 48 часов. Оптимальный отклик инвазии замечен при 4 нг/мл VEGF (данные не показаны) и этот отклик устанавливали как начальную инвазию или нулевой % ингибирования (Фигура 3D). Клетки ПКР-53, предварительно обработанные в течение 30 минут 350 или 700 нг/мл rhТИМП-1 или ТИМП-1-ГФИ, затем промывали и вносили в верхнюю ячейку камеры Бойдена. Относительное увеличение или уменьшение миграции/инвазии затем определяли (см. Материалы и Методы). В то время как обработка rhТИМП-1 частично блокировала инвазию клеток, ТИМП-ГФИ при 700 нг/мл полностью блокировал инвазию клеток ПКР-53 (Фигура 3D).
Пример 4
ГФИ-фиксированный ТИМП-1 влияет на пролиферацию ПКР
Чтобы оценить влияние поверхностной инженерии с ТИМП-1 на пролиферацию ПКР, выполняли анализ с МТТ (3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолийбромид). Экзогенно добавленный белок ТИМП-1-ГФИ, как найдено, выявляет зависимое от дозы снижение пролиферации ПКР-53 и А498 за 24, 48 и 72 часа (Фигура 4В). Клетки ПКР-26 распространяются чрезвычайно медленно (48+ часов, степень удвоения) и общая тенденция предполагала подавление пролиферации (Фигура 4С). Дополнительные регуляторы, использующие фосфатидилинозит при равной молярной концентрации с реагентом ТИМП-1-ГФИ (Сигма, Германия, Номер Р6636), не приводили к значительным изменениям в пролиферации клеток (данные не показаны).
Пример 5
Клеточно-опосредованная цитотоксичность
Активность ММП связывалась с чувствительностью целевых клеток к апоптозу, вызванному цитотоксической активностью Т-лимфоцитов (как перфорин/гранзимный, так и FAS-опосредованный апоптоз) (Egeblad & Werb, 2002). Влияние ТИМП-1-ГФИ на клеточно-зависимый лизис ПКР проверяли, используя апоптоз, вызванный аллогенными ЦТЛ и ЕКК клетками. В анализе, опосредованном аллогенными ЦТЛ, целевые клетки ПКР-53, А498 и ПКР-26 обрабатывали 700 нг/мл rhТИМП-1 или ТИМП-1-ГФИ, затем метили Cr51 и инкубировали с клеточными линиями аллогенных CD8+CTL JB4 (Фигура 5А) или ЕКК (Фигура 5В). Клеточные линии опухоли ПКР узнавались и эффективно подвергались лизису как клетками ЦТЛ, так и ЕКК. Обработка ТИМП-1 или ТИМП-1-ГФИ не изменяла восприимчивость трех линий ПКР к апоптозу, опосредованному как клетками ЦТЛ, так и ЕКК.
Пример 6
Обработка ТИМП-ГФИ делает ПКР чувствительными к FAS-опосредованному лизису
Эксперименты ЦТЛ/ЕКК показали, что обработка ТИМП-1-ГФИ не влияла на перфорин/гранзим-опосредованный литический путь, измеряемый анализом выделения хрома. Этот путь действует быстро, используя способность секреции сохраненных цитотоксинов инициировать апоптоз, и представляет один из двух главных иммуно-инициированных механизмов смерти клетки (Trapani и другие, 2000). Второй путь включает FAS/CD95-лигирование. Затем определяли влияние обработки ТИМП-1-ГФИ на FAS-опосредованный апоптоз.
FAS-экспрессию на линиях ПКР сначала оценивали поточной цитометрией, используя анти-FAS моноклональные антитела (L-958) (Н. Engelmann, неопубликованные результаты). Необработанные клетки и клетки, обработанные 700 нг/мл ТИМП-1-ГФИ или контрольного белка rhТИМП-1 в течение 24 часов, окрашивали L-958 и анализировали. FAS-белок сильно экспрессировался всеми тремя линиями ПКР. Обработка ТИМП-1-ГФИ или rhТИМП-1 не влияла на экспрессию FAS поверхностью клеток (Фигура 6А).
Анти-FAS моноклональные антитела (L-958) могут вызывать апоптоз в клетках, экспрессирующих FAS (Н. Englemann, неопубликованные результаты). Закрепление аннексин V-флюоресцеинизоцианата (FITC) и включение пропидийодида (ПИ) в клетки ПКР после обработки L-957 применяли, чтобы обнаружить апоптоз поточной цитометрией. Как показано в Фигуре 6В, ПКР-26 и ПКР-53 в значительной степени устойчивы к FAS-опосредованному апоптозу. Интенсивность флюоресценции (MFI) необработанных клеток и клеток, обработанных L-957, похожа. Небольшое увеличение интенсивности замечено в ПКР-53 в ответ на обработку L-957. Эти наблюдения соответствуют предыдущим сообщениям, что ПКР обычно устойчив к FAS-опосредованному апоптозу (Frost и другие, 2003). Обработка ТИМП-1-ГФИ (L-957+ТИМП-1-ГФИ), но не контрольным rhТИМП-1 (L-957 + rhТИМП-1), делало клеточные линии чувствительными к FAS-опосредованному апоптозу (Фигура 6В). Как найдено, А498 были более чувствительными к активации анти-FAS моноклональных антител (обнаружено повышенной интенсивностью флюоресценции аннексин-MFI в образцах, обработанных L-957), обработка ТИМП-1-ГФИ, но не rhТИМП-1, значительно увеличивала апоптоз клеток А498.
В то время как клетки ПКР-26 и А498 показали резкое увеличение FAS-индуцированного апоптоза, за которым следует обработка ТИМП-1-ГФИ, клеточная линия ПКР-53 показала менее явное увеличение чувствительности (изменение интенсивностью флюоресценции (МFI) от 62 до 93). Чтобы подтвердить ТИМП-1-ГФИ/FAS-индуцированный апоптоз ПКР-53, применяли второй иммуносорбентный анализ ELISA, основанный на обнаружении цитоплазматического хроматина. Преимущества этого анализа включают недостаточную субъективность в интерпретации результатов и его повышенную чувствительность относительно окрашивания аннексином V. ELISA анализ хроматина в состоянии обнаружить только 300 апоптотических клеток и измеряет события апоптоза значительно позже раннего наличия аннексина V на поверхности клетки. Как найдено FACS анализом аннексина V (Фигура 6В), обработка ПРК-53 одним L-957 вызывала небольшое увеличение апоптоза (Фигура 6С). Однако ТИМП-1-ГФИ обработка клеток ПКР-53 резко увеличивала чувствительность клеток к анти-FAS-индуцированному апоптозу зависимым от дозы образом.
Результаты показывают, что обработка раковых клеток ГФИ-фиксированным ТИМП вызывает лизис раковых клеток FAS-индуцированным апоптозом. Поэтому ГФИ-фиксированный ТИМП полезен в качестве противоопухолевого средства.
Пример 7
ТИМП-1-ГФИ обработка ПРК снижает BCL-2 и увеличивает экспрессию белка ВАХ
Белки BCL-2 представляют семейство белков, вовлеченных в контроль апоптоза (рассмотрено у Igney & Krammer, 2002). Некоторые члены этого семейства (такие как BCL-2 и BCL-XL) являются антиапоптотическими, в то время как другие (такие как Bad или ВАХ) являются проапоптотическими. Чувствительность клеток к апоптотическим стимулам может зависеть от баланса между про- и антиапоптотическими членами семейства BCL-2 (Igney & Krammer, 2002). Эффект ТИМП-1-ГФИ обработки на экспрессии BCL-2 и ВАХ затем определяли. После 24 часов прединкубации с 700 нг/мл ТИМП-1-ГФИ или контрольного rhТИМП-1, клетки ПКР стимулировались 1 мкг/мл L-957 (или контрольного моноклонального антитела) в течение дополнительных 16 часов. Уровень белка BCL-2 и ВАХ затем определяли, используя внутриклеточный FACS и Вестерн-блоттинг. Во всех трех клеточных линиях обработка ТИМП-1-ГФИ увеличивала экспрессию проапоптотического ВАХ, и уменьшала экспрессию антиапоптотического BCL-2. Подобная картина отмечена в Вестерн-блоттинг анализе (Фигура 7А, В и С).
Пример 8
Включение экзогенно добавленного ТИМП-1-муцин-ГФИ в поверхность ПКР-53
Белок ТИМП-1-муцин-ГФИ производили и выделяли, как описано в примере 1. Включение очищенного белка ГФИ-ТИМП-1 в поверхностные мембраны ПКР-53, ПКР-26 или А498 клеточных линий ПКР показано инкубацией клеточных линий с 700 нг/мл очищенного ТИМП-1-муцин-ГФИ или рекомбинантного человеческого (rh)ТИМП-1 контрольного белка в течение одного часа. Связанный с поверхностью белок ТИМП-1-муцин-ГФИ затем детектировали, используя FACS анализ. Конструкт ТИМП-1-муцин-ГФИ эффективно включался в поверхностную мембрану и эффективно вызывает противоопухолевую активность.
Пример 9
ТИМП-ГФИ для лечения остаточного рака в человеке
Реагент ТИМП-1-ГФИ или ТИМП-муцин-ГФИ применяют в количестве 1 мкг/мл местно в область резекции после хирургического вырезания опухоли. Неоперабельную опухоль, глиобластому (астроцитома grad IV WHO), хирургически удаляют и реактивы вводят перед закрытием раны.
Пример 10
ТИМП-ГФИ для лечения остаточного рака в человеке
Количество 1 мкг/мл реагента ТИМП-1-ГФИ или ТИМП-муцин-ГФИ применяют местно в область резекции после хирургического вырезания опухоли. Опухоль, рак молочной железы в продвинутой стадии, хирургически удаляют и реактивы вводят перед закрытием раны, особенно, если есть клинический риск местного рецидива.
Пример 11
Оценка ТИМП-ГФИ в моделях метастаза опухоли
Оценивали влияние ТИМП-1-ГФИ на метастаз опухоли. Используя мышиную модель, Т клетки лимфомы, которая эффективно метастазировала в печени, предварительно обрабатывали ТИМП-1-ГФИ или rhТИМП-1 контрольным белком. Получающуюся опухоль затем вводили через вену хвоста и распределение опухоли в печени определяли три и семь дней спустя. Результаты демонстрируют, что клетки, обработанные ТИМП-1-ГФИ, показывают значительно сниженный уровень микрометастаза относительно контрольных клеток, обработанных ТИМП.
Пример 12
Анализ инвазии Матригеля
Влияние ТИМП-1-ГФИ на клеточные линии опухоли Примера 11 анализировался в ряду экспериментов Матригель. Результаты подтвердили, что ТИМП-1-ГФИ имел сильное воздействие на способность клеток линии опухоли Т клеток подвергаться инвазии Матригеля относительно rhТИМП-1.
Пример 13
Влияние слитых конструктов ТИМП на продукцию фибронектина фибробластами
Сливные фибробласты культивировали в присутствии или отсутствии rhТИМП-1 и ТИМП-1-ГФИ. Экспрессированный и секретированный фибронектин количественно определяли Вестерн-блоттинг анализом, используя антитела к фибронектину (β-актин служил контролем). Фигура 9 изображает результаты этого эксперимента и ясно демонстрирует, что rhТИМП-1 (при 700 нг/мл) не приводил к какому-либо значительному уменьшению в экспрессии фибронектина, в то время как ТИМП-1-ГФИ (при 700 нг/мл) сильно уменьшал фибронектин, который секретировался фибробластами.
Дополнительно, РНК фибронектина, транскрибированная фибробластами, оценивалась Нозерн-блоттинг анализом. Фибробласты культивировали в присутствии (Фигура 10А) или отсутствии (иллюстрация 10В) 10 нг/мл фибробласт-активирующего ФНО-α вместе с ТИМП-1-ГФИ (350 нг/мл) или ТИМП-1-ГФИ (700 нг/мл), денатурированным ТИМП-1-ГФИ и rhТИМП-1-ГФИ соответственно. Кроме того, в культуральной питательной среде присутствовали 0%, 1%, 5% или 10% фетальной телячьей сыворотки (FCS).
Эти результаты ясно демонстрируют, что при концентрации 350 нг/мл ТИМП-1-ГФИ, транскрибированная РНК фибронектина значительно уменьшалась, независимо от того, присутствовал ли ФНО-α в питательной среде или нет, и независимо от содержания FCS в питательной среде. При 700 нг/мл ТИМП-1-ГФИ РНК фибронектина едва поддавалась обнаружению.
Таким образом, слитый конструкт ТИМП-ГФИ эффективно ингибирует и синтез и секрецию фактора роста фибронектина в фибробластах.
Пример 14
Влияние слитого конструкта ТИМП на продукцию IL-6 фибробластами
Нозерн-блоттинг анализ Примера 13 повторяли, используя зонд для РНК IL-6. Таким образом, фибробласты культивировали в присутствии (Фигура 11А) или отсутствии (Фигура 11В) 10 нг/мл фибробласт-активирующего ФНО-α вместе с ТИМП-1-ГФИ (350 нг/мл) или ТИМП-1-ГФИ (700 нг/мл), денатурированного ТИМП-1-ГФИ и rhТИМП-1-ГФИ соответственно. Кроме того, 0%, 1%, 5% или 10% фетальной телячьей сыворотки (FCS) присутствовали в культуральной питательной среде (Фигура 11).
Результаты ясно демонстрируют, что уже при концентрации 350 нг/мл ТИМП-1-ГФИ транскрибированная РНК IL-6 поразительно снижалась, независимо от того, присутствовал ли ФНО-α в питательной среде, и независимо от содержания FCS в питательной среде.
Таким образом, слитый конструкт ТИМП-ГФИ эффективно ингибирует синтез и секрецию еще одного важного цитокина, вовлеченного в заживление раны, а именно IL-6, в фибробластах.
Пример 15
Влияние слитого конструкта ТИМП на продукцию коллагена фибробластами
Нозерн-блоттинг анализ, выполненный в Примерах 13 и 14, повторяли, используя зонды для Коллагена 1А1 (Фигура 12А и В), Коллагена 4А2 (Фигура 12С и D) и Коллагена 16А1 (Фигура 12Е и F) соответственно. Таким образом, фибробласты культивировали в присутствии (Фигура 12А, С и Е) или отсутствии (Фигура 12 В, D, F) 10 нг/мл фибробласт-активирующего ФНО-α вместе с ТИМП-1-ГФИ (350 нг/мл) или ТИМП-1-ГФИ (700 нг/мл), денатурированным ТИМП-1-ГФИ и rhТИМП-1-ГФИ соответственно. Кроме того, 0%, 1%, 5% или 10% FCS присутствовали в культуральной питательной среде.
Результаты ясно демонстрируют, что уже при концентрации 350 нг/мл ТИМП-1-ГФИ все три транскрибированных РНК коллагена были значительно уменьшены, независимо от того, присутствовал ли ФНО-α в питательной среде, и независимо от содержания FCS в питательной среде.
Таким образом, слитый конструкт ТИМП-ГФИ эффективно ингибирует синтез и секрецию коллагена, который является одним существенным белком в продукции ВКМ и модернизации ткани.
Пример 16
Влияние слитого конструкта ТИМП на продукцию TGF-β фибробластами
Нозерн-блоттинг анализ Примеров 13-15 повторяли, используя зонды для TGF-β. Таким образом, фибробласты культивировали в присутствии (Фигура 13А) или отсутствии (Фигура 13В) 10 нг/мл фибробласт-активирующего ФНО-α вместе с ТИМП-1-ГФИ (350 нг/мл) или ТИМП-1-ГФИ (700 нг/мл), денатурированного ТИМП-ГФИ и rhТИМП-1-ГФИ соответственно. Кроме того, 0%, 1%, 5% или 10% FCS присутствовали в культуральной питательной среде.
Результаты ясно демонстрируют, что уже при концентрации 350 нг/мл ТИМП-ГФИ, почти никакой PHK TGF-β не обнаружено, независимо оттого, присутствовал ли ФНО-α в питательной среде или нет, и независимо от содержания FCS в питательной среде.
Поэтому, как показано вышеупомянутым Примером, слитый конструкт ТИМП-ГФИ эффективно ингибирует синтез и секрецию еще одного важного цитокина, вовлеченного в заживление раны, а именно TGF-β, в фибробластах.
Пример 17
Генерация дальнейших слитых конструктов ТИМП
Дальнейшие слитые конструкты ТИМП производили и очищали согласно разделу "Материалы и Методы", предоставленному ниже. Конкретно, укороченный слитый конструкт ТИМП-1-ГФИ (SEQ ID NO:1), укороченный слитый конструкт ТИМП-1-муц-ГФИ (SEQ ID NO:2), конструкт ТИМП-2-ГФИ (SEQ ID NO:3), конструкт ТИМП-3-ГФИ и измененная форма ТИМП-3-ГФИ (SEQ ID NO:4) и укороченный слитый конструкт ТИМП-1-фракталкин-ГФИ (SEQ ID NO:5) были получены, экспрессированы и очищены.
Укороченный слитый конструкт ТИМП-1-ГФИ (SEQ ID NO:1) включает первые 152 аминокислоты человеческого белка ТИМП-1 (то есть удалены 126-207 аминокислоты С-конца), слитых с ГФИ-якорем из 36 аминокислот в длину. Укороченный слитый конструкт ТИМП-1-муц-ГФИ (SEQ ID NO:2) содержит первые 152 аминокислоты человеческого белка ТИМП-1, слитого с аминокислотами 256-380 человеческого CXCR16 (муцина), далее слитого с ГФИ-якорем из 36 аминокислот в длину. Получившийся слитый конструкт содержит 295 аминокислот и имеет молекулярный вес 32 111 kDa.
По аналогии с ТИМП-1-ГФИ полной длины, ТИМП-2-ГФИ (SEQ ID NO:3) и ТИМП-3-ГФИ состоят из человеческого ТИМП-2 и белка ТИМП-3 соответственно, слитых с ГФИ-якорем из 36 аминокислот в длину. Чтобы произвести измененную форму слитого конструкта ТИМП-3-ГФИ (SEQ ID NO:4), связывающий домен глюкозаминоглюканов (ГАГ) человеческого ТИМП-3, который, как думали, отвечает за ассоциацию белка с поверхностью клетки, изменяли следующими шестью заменами: R43A, К45А, К49А, К53А, К65А и К68А. Укороченный слитый конструкт ТИМП-1-фракталкин-ГФИ (SEQ ID NO:5) содержит N-концевой участок человеческого ТИМП-1 (аминокислоты 1-152), слитый с аминокислотами 100-342 человеческого CX3CL1, далее слитый с ГФИ-якорем из 36 аминокислот в длину.
Подробное изучение укороченных конструктов ТИМП-1-ГФИ (SEQ ID NO:1), ТИМП-1-муцин-ГФИ (SEQ ID NO:2) и ТИМП-1-фракталкин-ГФИ (SEQ ID NO:5)
а) Включение экзогенно добавленных укороченного ТИМП-1-ГФИ, укороченного ТИМП-1-муцин-ГФИ и укороченного ТИМП-1-фракталкин-ГФИ в клеточную поверхность
Укороченный ТИМП-1-ГФИ, укороченный ТИМП-1-муцин-ГФИ и укороченный ТИМП-1-фракталкин-ГФИ производили и выделяли согласно Djafarzadeh и др., 2004. Включение очищенных слитых конструктов в мембраны клеточной поверхности клеточных линий ПКР-53, ПКР-26 или А498 ПКР показано инкубацией клеточных линий в течение одного часа с 700 нг/мл очищенного укороченного ТИМП-1-ГФИ, укороченного ТИМП-1-муцин-ГФИ и укороченного ТИМП-1-фракталкин-ГФИ соответственно, и сравнивается с инкубацией соответствующего укороченного ТИМП-1 контрольного белка ТИМП-1, который имеет недостаток муцина, фракталкина и доменов ГФИ. Связанный с поверхностью белок затем детектировали, используя анализ FACS. Ожидалось, что добавление контрольного ТИМП-1 не будет приводить к какому-либо изменению в сдвиге FACS, однако, укороченный ТИМП-1-ГФИ, укороченный ТИМП-1-муцин-ГФИ и укороченный ТИМП-1-фракталкин-ГФИ соответственно фактически приводили к сильному поверхностному сигналу для ТИМП-1.
Чтобы продемонстрировать, что экзогенно добавленный белок был ГФИ-фиксированным, клетки ПКР-53 сначала инкубировали с укороченным ТИМП-1-ГФИ, укороченным ТИМП-1-муцин-ГФИ и укороченным ТИМП-1-фракталкин-ГФИ соответственно (200 или 700 нг/мл), и затем обрабатывали фосфолипазой С (PLC) (60 нг/мл). Анализ FACS, как ожидается, показывает полную потерю сигнала ТИМП-1 поверхностью клетки после расщепления PLC. Чтобы измерить эффективность интеграции фиксированных конструктов ТИМП, конструкты ТИМП-1, освобожденные от мембраны, собирали в промывных буферах и определяли количественно, используя ТИМП-1-специфический иммуносорбентный анализ ELISA. Ожидалось, что большая часть исходного антигена ТИМП-1 будет извлекаться из образца на 200 нг/мл.
b) Белки укороченный ТИМП-1-ГФИ, укороченный ТИМП-1-муцин-ГФИ и укороченный ТИМП-1-фракталкин-ГФИ блокируют выделение проММП-2 и проММП-9 из ПКР-53
Увеличенная экспрессия ММП-2 и ММП-9 обычно коррелирует с плохим прогнозом ПКР (Hemmerlein и др., 2004). Клеточная линия стадии IV светлоклеточного рака ПКР-53 значительно секретирует и проММП-2 и проММП-9 (Фигура 2). Влияние увеличивающихся поверхностных уровней ТИМП-1 на значительное выделение белков ММП-2 и ММП-9 проверяли, используя анализ желатиназной зимографией (Djafarzadeh и др., 2004; Klier и др., 2001). Белок rhТИМП-1 при 600 или 1200 нг/мл не имел никакого эффекта на секрецию проММП-2 или проММП-9. Напротив, начиная с 10 нг/мл, обработка укороченным ТИМП-1-ГФИ, укороченным ТИМП-1-муцин-ГФИ и укороченным ТИМП-1-фракталкин-ГФИ соответственно, как ожидали, показало зависимое от концентрации уменьшение выделения как проММП-2, так и проММП-9 в питательные ростовые среды, сопоставимое с обработкой ТИМП-1-ГФИ, описанной в Примере 2.
с) Белки укороченный ТИМП-1-ГФИ, укороченный ТИМП-1-муцин-ГФИ и укороченный ТИМП-1-фракталкин-ГФИ воздействуют на пролиферацию ПКР
Чтобы оценить влияние инженерии поверхности с помощью ТИМП-1 на пролиферацию ПКР, выполняли пробы МТТ. Экзогенно добавленные белки укороченный ТИМП-1-ГФИ, укороченный ТИМП-1-муцин-ГФИ и укороченный ТИМП-1-фракталкин-ГФИ соответственно, как ожидали, выявили зависимое от дозы уменьшение пролиферации ПКР-53 и А498 за 24, 48 и 72 часа, сопоставимое с Примером 4. Клетки ПКР-26, как ожидали, распространяются чрезвычайно медленно (48+ часов, норма удвоения); общая тенденция действительно предполагает подавление пролиферации.
Пример 18
Дальнейшая оценка других слитых конструктов Примера 17
Эксперименты Примеров 1-16 повторяли, используя слитые конструкты по Примеру 17; подобные результаты ожидались. В частности, слитые конструкты ТИМП-2 ингибировали большинство ММПs (кроме ММП-9) и предпочтительно ингибировали ММП-2. Слитый конструкт ТИМП-3 ингибировал ММП-1, -2, -3, -9 и -13, так же как ФНО-альфа-превращающий энзим (ТАСЕ). Измененная форма слитого конструкта ТИМП-3 (SEQ ID NO:4) исследовалась относительно его интеграционных свойств, которые дополнительно показали улучшенную способность включаться в клеточную мембрану (эти эксперименты были выполнены согласно методам, предоставленным Примером 8). Укороченные слитые конструкты ТИМП-1 (SEQ ID NO:1, 2, 5) показали биологические функции, сравнимые с функциями слитых конструктов ТИМП-1 полной длины, таким образом, поддерживая представление, что N-концевой участок ТИМП является существенным для их ингибиторной функции. Слитый конструкт фракталкина (SEQ ID NO:5) далее показал способность интеграции в мембраны, сопоставимую со способностью слитого конструкта муцина (SEQ ID NO:4).
Обсуждение результатов
Инженерия поверхности клетки ГФИ-фиксировнными белками по настоящему изобретению предлагает несколько преимуществ перед традиционными подходами переноса гена. 1) Метод применим к клеткам, которые являются трудными для трансфекции, например, к ПКР-резистентным клеткам FAS-опосредованного апоптоза, но также к первичным культурам, клеткам-предшественникам костного мозга и клеткам иммунной системы. 2) Метод может использоваться, когда только небольшое количество клеток доступно или когда клетки не могут легко размножаться. 3) Поверхность клетки может быть изменена независимо от типа клетки. 4) Количеством белка, в конечном счете, показанного на поверхности клетки, можно точно управлять. 5) Многочисленные ГФИ-фиксированные белки могут быть включены последовательно или одновременно в те же самые клетки.
Человеческий ПКР представляет собой прогрессирующую опухоль с ограниченными терапевтическими возможностями из-за устойчивости опухоли к современным химиотерапевтическим средствам и радиации. Иммунотерапии полезны для некоторых больных, предполагая, что ПКР может быть целью иммунного эффекторного механизма. Проникающие в опухоль лимфоциты, такие как CD8 + ЦТЛ или клетки ЕКК часто замечаются в тканях почечного рака и часто узнают аутологические опухолевые клетки при проверке in vitro (рассмотрено в Schendel и др., 1997). Несмотря на эти многообещающие наблюдения, опухоли обычно продолжают расти, указывая, что ПКР, возможно, приобретали устойчивость к цитотоксическим механизмам. Для того чтобы продуктивный «антиканцерный» эффект имел место, иммунная система должна не только узнавать опухоль, но раковые клетки должны также быть восприимчивыми к механизмам лизиса, используемым клетками ЦТЛ или клетками ЕКК. Раковые клетки развили различные механизмы, чтобы уклоняться от иммунной защиты, включая пониженную чувствительность к апоптозу (рассмотрено у Dunn и др., 2004).
Клетки ЦТЛ и ЕКК убивают клетки-мишени перфорин/гранзим- или FAS/FASL-зависимым апоптозом (Kagi и др., 1994). Относительная важность экзоцитоза гранул против FAS/FASL-опосредованных литических активностей контроля опухоли in vivo является спорной. В то время как много исследований указывают на доминирование механизма экзоцитоза гранул, другие исследования, использующие перфорин оглушенных мышей (Seki и др., 2002) предполагают, что FAS-зависимый апоптоз может быть более известным путем in vivo. Большинство опухолевых клеток, включая ПКР, в действительности, устойчиво к FAS-опосредованному лизису (Frost и др., 2003). Использование ГФИ-фиксированного ТИМП представляет перспективный терапевтический подход для того, чтобы делать опухолевые клетки, восприимчивыми к FAS-опосредованному апоптозу.
Мы показали в настоящем изобретении, что инженерия клеток экзогенным добавлением ГФИ-ТИМП-1 может выявить усиленные, также как и новые, биологические активности ТИМП-1. Внешне вводимый ТИМП-1-ГФИ эффективно внедрялся в клеточные мембраны ПКР и вызывал разные биологические эффекты в линиях ПКР с потенциальной терапевтической значимостью.
Кроме того, ГФИ-фиксированный белок ТИМП-1 резко изменял ассоциацию поверхности клетки с разными ММП, экспрессированными ПКР. Это было отражено пониженной секрецией ММП, включая проММП-2 и проММП-9, клетками ПКР. В то время как ТИМП-1 будет блокировать ферментативную активность ММП-2, она, как думают, не связана с проформой фермента. Все же данные, демонстрирующие блокаду секреции проММП-2 после обработки ТИМП-1-ГФИ, внушаются этим действием. Кажется, что присоединение якоря ГФИ к ТИМП-1 приводит к тому, что измененная поверхностная стехиометрия увеличивала способность белка ТИМП-1 связывать ММП.
Это очевидное повышенное связывание ТИМП-1-ГФИ также продемонстрировано ММП мембранного типа. В то время, как мало известно об ассоциации ТИМП-1 с ММП-15, связывание белка ТИМП-1-ГФИ с ММП-16, как было предсказано, не встречается ввиду довольно слабой авидности природного ТИМП-1 к этому белку (Lang и другие, 2004). Мутационный анализ петель ТИМП-1, важных для связывания ММП-16, показал, что малые, очевидно незначительные изменения в ТИМП-1 могут резко сдвигать его ингибиторно-связывающие характеристики. В этом случае измененная стехиометрия ТИМП-1 на клеточной поверхности кажется достаточной, чтобы сдвинуть его связывание с ММП-16. Связывание ММП на поверхности клетки также ассоциируется с пониженной способностью клеточной линии ПКР-53 подвергаться инвазии в ВКМ.
Как демонстрируется в настоящем изобретении, обработка ТИМП-1-ГФИ приводит к явному зависимому от дозы сокращению пролиферации линий ПКР. Возможно, наиболее важно, что линии ПКР, обычно устойчивые к FAS-апоптозу, делались чувствительными к FAS/CD95-опосредованному лизису после обработки белком ТИМП-ГФИ по изобретению. Однако средство не влияло на чувствительность к перфориновому пути. Это предполагает, что ГФИ-фиксированный ТИМП достигает противоопухолевого эффекта FAS-индуцированным апоптозом, а не перфорин/гранзим-опосредованным лизисом клетками ЦТЛ/ЕКК.
Путь FAS-апоптоза регулируется активацией каспазы, в то время как повреждение клеточной мембраны ЦТЛ/ЕКК, использующее экзоцитоз гранул, как измерено анализом выделения хрома, имеет место независимо от каспазы (Sayers и другие, 1998; Seki и другие, 2002; Trapani и другие, 2000). События, предшествующие активации каспазы, включают баланс между про- и антиапоптотическими белками семейства BCL-2. Как показано в настоящем изобретении, обработка ГФИ-ТИМП-1 приводило к низкоуровневому регулированию антиапоптотического белка BCL-2 и соответствующему увеличению проапоптотического белка ВАХ. Этот сдвиг к более высокой концентрации проапоптотических белков может быть одной из причин повышенной чувствительности клеток ПКР с ТИМП-1-модифицированой поверхностью к FAS-опосредованному апоптозу. Эти наблюдения представляют новую активность для ТИМП-1. Подобные активности также показали ТИМП-3 и другие ТИМП (ТИМП-2 и ТИМП-4) (данные не показаны).
Чрезмерная экспрессия ТИМП-1, -2 или -3 в клетках сосудов гладких мышц, используя аденовирусные векторы, как найдено, ингибирует их миграцию через модельные мембраны основания. Чрезмерная экспрессия ТИМП-1 не имела никакого влияния на пролиферацию клеток, в то время как ТИМП-2 вызвал зависимое от дозы ингибирование пролиферации клеток. Чрезмерная экспрессия ТИМП-3 также вызвала зависимое от дозы ингибирование пролиферации и, кроме того, приводила к апоптозу через деполяризацию митохондриальных мембран и утечку цитохрома С (Baker и другие, 1999; Baker и другие, 1998; Smith и другие, 1997). ТИМП-3 представляет собой единственный белок ТИМП, который выборочно связывается с поверхностью клеток, независимо от ассоциации с другими поверхностными белками (Majid и другие, 2002; Smith и другие, 1997). Сосредоточение ТИМП-1 на поверхности клетки через ГФИ-якорь приводит к новым биологическим активностям, которые, кажется, подражают эффектам, сообщенным для ТИМП-3.
ТИМП-3, как показано, делает чувствительными клетки меланомы к апоптозу, вызванному анти-FAS антителами, ФНО-α и TRAIL (родственный ФНО апоптозиндуцирующий лиганд). Механизм действия связывали с общей стабилизацией FAS, ФНО-рецептора I и TRAIL-рецептора I на поверхности клетки меланомы, обработанной ТИМП-3 (Ahonen и другие, 2003). Эта повышенная поверхностная экспрессия рецепторов связывалась с активацией каспазы-8 и каспазы-3 (Ahonen и другие, 2003). 4848
В экспериментах, детализированных в настоящем изобретении, клетки ПКР не показывали изменение поверхностной экспрессии FAS после обработки ТИМП-1-ГФИ (Фигура 6А). Кроме того, клетки ПКР оставались устойчивыми к ФНО-α-вызванному апоптозу (проверено от 100 до 10000 единиц на мл) независимо от обработки ТИМП-ГФИ (данные не показаны). Анализ FACS клеток ПКР впоследствии показал едва поддающиеся обнаружению уровни ФНО-рецептора I (р55) и ФНО-рецептора II (р75) на ПКР-53 и никакой экспрессии на линиях ПКР-26 или А498 (данные не показаны). Поверхностная экспрессия не изменялась при обработке ТИМП-1-ГФИ. Таким образом, повышенная чувствительность к FAS-опосредованному апоптозу после обработки ТИМП-1-ГФИ, кажется, не опосредуется через общую стабилизацию белковых рецепторов смерти на поверхности клетки. Ясно, что обработка ТИМП-1-ГФИ действительно изменяет баланс белков Bcl-2, чтобы установить "более проапоптотический" профиль экспрессии.
Результаты настоящего изобретения обеспечивают дополнительную связь между биологией опухоли, функцией ММП/ТИМП и путями апоптоза. Соединение белков ТИМП с ГФИ непосредственно или через домены муцина представляет сильное противоопухолевое средство, чтобы придать опухолевым клеткам, которые обычно устойчивы к FAS-опосредованному апоптозу, чувствительность к FAS-опосредованному апоптозу. Этим механизмом опухолевые клетки будут эффективно подвергаться лизису.
Относительно использования слитых конструктов, в частности конструкта ТИМП-1-ГФИ, конструкта ТИМП-1-муц-ГФИ и таких как определено SEQ ID NOs:1, 2, 3, 4 и 5, в области регенеративной медицины, например заживлении ран, конструкты ТИМП-ГФИ по настоящему изобретению, как здесь показано, эффективно ингибировали продукцию и секрецию важных ферментов и цитокинов (фибронектин, коллаген, IL-6, TGF-β), вовлеченных в процессы модернизации ткани и фиброза, которые приводят к повышенной продукции ВКМ. Таким образом, члены семейства ТИМП (ТИМП-1, ТИМП-2, ТИМП-3, ТИМП-4), если закрепляются в клеточной мембране посредством ГФИ-якоря или муцина или фракталкина и ГФИ, могут быть использованы, чтобы эффективно смодулировать процессы заживления раны, влияя на тонкий баланс между продукцией ВКМ и обновлением ВКМ.
Материалы и Методы
Клеточные линии и клеточная культура
Линии ПКР, ПКР-53 и ПКР-26 генерировали D.J.S. (Мюнхен, Германия) из образцов, взятых у больных. ПКР-53, ПКР-26 и А498 (American Type Culture Collection) (Giard и другие, 1973) культивировали в питательной среде RPMI1640 (GIBCO BRL, Life Technologies GmbH, Eggenstein, Germany), дополненной 2 мМ L-глутамина (Biochrom KG, Berlin), 1 мМ пирувата натрия (GIBCO BRL, Life Technologies GmbH, Eggenstein, Germany), 12% инактивированной температурой фетальной телячьей сывороткой FCS (Biochrom KG, Berlin, No. S01 15). Свежую питательную среду подавали каждый третий день, и культуры разлагали на компоненты, если клетки были объединенными.
Цитотоксические эффекторные клетки: JB4 представляет собой HLA-A2-аллореактивный цитотоксический Т эффекторный клон, произведенный в нашем собственном оборудовании (E.N.), и расширяется стимулированием каждые две недели, как описано (Milani и другие, 2005). Он используется в пробах цитотоксичности на 7 или 8 день после стимулирования. Человеческие лейкозные линии ЕКК (NKL) (Robertson и другие, 1996) и NK-92 (Gong и другие, 1994) были любезно предоставлены C.S. Falk (GSF-lnstitute of Molecular Immunology, Munich, Germany) и культивировались в питательной среде, содержащей 15% инактивированную температурой фетальную телячью сыворотку (FCS) и 100 ЕД/мл рекомбинантного интерлейкина-2 (IL-2). За день перед использованием в пробах цитотоксичности, культура была приведена к 0,3 х 105 клеток/мл в свежей питательной среде.
Анализ клеточным сортером с возбуждением флуоресценции (FACS анализ)
Клетки разъединяли 1,5 мМ этилендиаминтетрауксусной кислотой (ЭДТУК) (Biochrom А, Берлин, Германия Номер L2113) в 1 х фосфатно-буферном растворе (ФБР) и инкубировали в течение 60 минут на льду с антителами, специфическими для человека; ТИМП-1 (IM32L), ММП-1 (IM35L-100), ММП-3 (IM36L-100), ММП-8 (IM38L) (CALBIOCHEM, Merck Darmstadt, Germany); ММП-9 (IM 61-100), ММП-2 (IM 51L) (ONCOGENE, Bad Soden, Germany); ММП-7 (MAB907), ММП-12 (MAB917), ММП-14 (МАВ9181) (R&D Systems, Minneapolis, USA); ММП-13 (IM44L), ММП-15 (IM48L), ММП-16 (IM50L) (CALBIOCHEM, Merck Darmstadt, Germany) и lgGIK (SIGMA-ALDRICH, Taufkirchen, Germany No. M9269). ICAM-1 и HLA-антитела (W6/32 и HB82) были описаны ранее (Johnson и другие, 1988; Barnstable и другие, 1978; Parham & Brodsky, 1981). Анти-FAS (Н. Engelmann, неопубликованные данные), анти-ФНО-RI, анти-ФНО-RII и изотипные контрольные антитела использовались, как описано (Bigda и другие, 1994). Клетки промывали три раза 1×ФБР, инкубировали с ослиными анти-мышиными антителами, сопряженными с флуоресцеинизотиоцианатом (ФИЦ) (DAKO A/S, Glostrup, Denmark No. F0313) в течение 45 минут на льду, затем промывали три раза 1×ФБР и анализировали, используя поточный цитометр (FACSCalibur, Becton, Dickinson and Company, San Jose, USA) и программное обеспечение CellQuest. Антитела анти-ВСL-2 (ALX-804-225) и анти-ВАХ (ANC-357-040) получали от ALEXIS (Grunberg, Germany). 5050
Очистка белка ТИМП-1-ГФИ
Белок ТИМП-1-ГФИ производили и очищали как ранее описано (Djafarzadeh и др., 2004). Кратко, человеческий ТИМП-1 клонировали из комплементарной кДНК, используя человеческие hТИМП-1, специфические праймеры, слитые без кодона остановки трансляции с ГФИ-сигнальной последовательностью, клонированной из антигена, связанного с функцией лимфоцитов (LFA-3) (Kirby и другие, 1995; Medof и другие, 1996) и субклонировали в pEF-DHFR и стабильно вводили в клетки яичника китайского хомячка (СНО) с дефицитом DHFR и отбирали, как описано (Mack и другие, 1995). Слитый белок ТИМП-1-ГФИ очищали от клеток СНО экстракцией детергентом Тритон Х-100, с последующей колоночной очисткой, используя диэтиламиноэтилцеллюлозу, гепаринсефарозу и эксклюзионную хроматографию (Djafarzadeh и другие, 2004).
Иммуносорбентный анализ ТИМП-1
Иммуносорбентный анализ, специфический к человеческому ТИМП-1, по протоколу, примененному согласно указаниям изготовителя (МАВ970, R&D Systems), использовался, чтобы контролировать уровни ТИМП-1 в растворе.
Моноклональное антитело покрытия против человеческого ТИМП-1 (МАВ970), биотинилированное моноклональное антитело обнаружения против человеческого ТИМП-1 (BAF970) и белок rhТИМП-1 (970-ТМ) были приобретены от R&D Systems GmbH (Wiesbaden, Germany).
Включение ТИМП-1-ГФИ в клеточные мембраны
Клетки ПКР-53 (5-10×106 клеток/мл) инкубировали с 200-700 нг/мл очищенного rhТИМП-1-ГФИ при 37°С/5% СO2. Клетки затем промывали три раза холодным фосфатно-буферным раствором (ФБР) и анализировали FACS, используя моноклональные антитела против человеческого ТИМП-1 (см. выше).
Расщепление ГФИ-якоря фосфолипазой С
Клетки (5-10×106 клетк/мл) инкубировали с 200 или 700 нг белка ТИМП-1-ГФИ или rhТИМП-1 в бессывороточной питательной среде в течение 1 часа при 37°С/5% СO2. Клетки промывали три раза холодным ФБР и обрабатывали 60 нг/мл фосфолипазы С, специфической к фосфатидилинозиту (SIGMA-ALDRICH, Taufkirchen, Germany No. 661-9) в бессывороточной питательной среде в течение 30 минут при 37°С/5% СO2. Клетки промывали три раза, все супернатанты были собраны.
Пролиферация
Клетки ПКР-53, А498 или ПКР-26 (30×103/100 мкл питательной среды) культивировали в 96-луночном титрационном микропланшете в течение 24 часов в стандартных условиях, давая крепко присоединенные и устойчиво растущие клетки. После отбрасывания супернатантов, 50 мкл питательной среды, содержащей ТИМП-1-ГФИ, буфер или rhТИМП-1 добавляли к клеткам и инкубировали от 24 до 72 часов. Затем добавляли 50 мкл 1 мг/мл раствора МТТ (3,5-диметилтиазол-2-ил-2,5-дифенилтетразолийбромида) (SIGMA-ALDRICH, Taufkirchen, Germany No. M2128). После 3 ч инкубации при 37°С кристаллы формазана растворяли добавлением 100 мкл изопропанола и 0,04 N НСl. Коэффициент поглощения затем измеряли при 550 нм, используя GENios plus TEC AN ELISA планшет-ридер. Для каждого эксперимента, по меньшей мере, 6 лунок анализировали на экспериментальное условие и пункт времени.
Зимография
Клетки ПКР-53 культивировали в 24-луночном планшете (5×104 клеток/лунку). Питательную среду обменивали в течение 24 часов с бессывороточной питательной средой, содержащей либо rhТИМП-1, либо увеличивающиеся количества ТИМП-1-ГФИ, и инкубировали в течение 24 часов, 48 часов и 72 часов. Клеточные супернатанты анализировали желатиновой зимографией, используя 10% гели натрийдодецилсульфат-полиакриламида (Invitrogen, Groningen, Netherlands, No. EC61755BOX), как описано (Djafarzadeh и другие, 2004). Рекомбинантный фермент ММП-9 (Amersham Biosciences, Uppsala, Sweden, No. RPN2634) использовали как надежный контроль.
Внеклеточная проба инвазии
Влияние обработки ТИМП-1-ГФИ против обработки rhТИМП-1 на способность клеток вторгаться в ВКМ оценивали, используя коммерческий анализ на инвазию клеток (Chemicon International Inc., Temecula, Калифорния, No. ECM 555). Клетки ПКР-53 сначала анализировали на их способность вторгаться в ВКМ. Клетки вторжения в основании вставки отделяли, подвергали лизису и обнаруживали краской CyQuant, как описано в сопроводительном протоколе. Увеличение уровней фактора роста сосудов эндотелия (ФРСЭ) (VEGF) от 2 нг/мл до 8 нг/мл использовали, чтобы увеличить инвазию. Оптимальная миграция была замечена при 4 нг/мл ФРСЭ (данные не показаны). Влияние обработки 350 нг/мл и 700 нг/мл контрольного rhТИМП-1 или ТИМП-1-ГФИ на миграцию затем определяли. Чтобы определить количественно потенциальное влияние средств ТИМП, начальная миграция ПКР-53 при 4 нг/мл ФРСЭ была установлена как 0. Величина 100% "ингибирования" ФРСЭ-индуцированной миграции была установлена как миграция/инвазия клеток ПКР-53 в отсутствии ФРСЭ. Конечные эффекты обработки rhТИМП или ТИМП-1-ГФИ на инвазию ПКР-53 были вычислены как изменение процента (отрицательное или положительное) относительно "максимальной" величины.
Обнаружение апоптоза с помощью аннексина-V
Обнаружение и определение количества апоптотических против некротических клеток на едином уровне клетки выполняли, используя Комплект окрашивания аннексин-V-FLUOS (Becton, Dickinson и Company, Heidelberg, Germany, No. 556547). Клетки ПКР-53 засевали при 1×106 клеток/лунку в 24-луночный планшет и позволяли закрепляться в течение ночи. Лунки затем ополаскивали 3 раза 1 × ФБР и добавляли 1 мл бессывороточной питательной среды RPMI 1640, за которой следовало 700 нг/мл ТИМП-1-ГФИ или rhТИМП-1. Клетки инкубировали в течение 24 часов при 37°С/5% СО2. После 24 часов добавляли 1 мкг/мл активирующего антитела L-957 против FAS (Н.Engelmann, неопубликованные данные) или изотипного контрольного моноклонального антитела и клетки затем далее инкубировали в течение 16 часов при 37°С/5% СO2. Клетки промывали ФБР, гранулировали и повторно суспендировали в окрашивающем растворе (аннексин-V-флуоресцеин-метящий реактив и пропидий йодид (ПИ) в буфере Hepes) в течение 15 минут при комнатной температуре. Клетки затем анализировали поточной цитометрией. Исследование динамики показало, что закрепление аннексина-V в клетках ПКР предшествует реакции ПИ.
Измерение апоптоза иммуносорбентным анализом ELISA, специфическим к хроматину
Апоптоз измеряли, используя ELISA Плюс комплект Обнаружения Смерти Клетки от Roche (Pensberg, Germany, No. 1774425). Клетки ПКР-53 высевали в 96-луночный планшет при концентрации 4×104 клеток/лунку и позволяли закрепляться в течение ночи. Лунки ополаскивали 3 раза 1 ×ФБР и 200 мкл бессывороточной питательной среды RPMI 1640 добавляли в каждую лунку, за которой следовало 700 нг/мл ТИМП-1-ГФИ или rhТИМП-1. Клетки инкубировали в течение 24 часов при 37°С/5% СО2. После 24 часов инкубации с ТИМП-1-ГФИ или rhТИМП-1 добавляли 1 мкг/мл активирующего моноклонального антитела L-957 против FAS или изотипного контрольного моноклонального антитела и клетки далее инкубировали в течение 16 часов при 37°С/5% СO2. Затем планшет центрифугировали, а супернатант тщательно удаляли. Шарик клеток помещали в 200 мл лизис-буфера, предоставленного изготовителем, на 30 минут и центрифугировали. Аликвоты супернатанта (20 мкл) использовали в иммуносорбентном анализе ELISA с антителами против ДНК и против гистонов, чтобы обнаружить наличие цитоплазматических нуклеосом.
Вестерн-блоттинг
Вестерн-блоттинг использовали для обнаружения ММП в бессывороточной ростовой среде. Используемые анти-ММП антитела описаны выше (анализ FACS). Рекомбинантные человеческие ММП стандарты вестерн-блоттинга приобретали от R&D Systems (Minneapolis, USA) и включали: ММП-1 (WBC024), ММП-2 (WBC025), ММП-3 (WBC015), ММП-8 (WBC017), ММП-12 (WBC019) и ММП-13 (WBC020). Вестерн-блоттинг также использовали для обнаружения BCL-2, ВАХ (см. выше для моноклональных антител mAbs) и β-актина (Acris Hiddenhausen, Germany, No. ab8227). Все белки детектировали, используя коммерческий комплект для Вестерн-блоттинг анализа от Chemiluminescent Immunodetection System (Invitrogen, Groningen, Netherlands).
Цитотоксичность, опосредуемая клетками
Клетки-мишени метили Cr51 в течение 1-2 часа, промывали и соинкубировали с эффекторными клетками при постоянном количестве клеток 2000 клеток/лунку в 96-луночном планшете с V-образными лунками. Двойные измерения четырех шагов титрования эффекторных клеток использовали во всех экспериментах. Самопроизвольные и максимальные выделения определяли инкубацией только клеток-мишеней и непосредственным подсчетом меченых клеток соответственно. После 4 часов инкубации при 37°С в увлажненной атмосфере 5% CO2 супернатанты собирали, переносили в микропланшет для сцинтилляции Lumaplate, высушивали в течение ночи и считали на сцинтилляционном счетчике микропланшетов TopCount (Packard, Meriden, Коннектикут). Для каждого отношения эффектор:мишень процент лизиса рассчитывали следующим образом: % специфического лизиса = (экспериментальное число импульсов в минуту - спонтанное число импульсов в минуту / максимальное число импульсов в минуту спонтанное число импульсов в минуту) × 100. Спонтанное выделение клеток-мишеней всегда <15% полного максимального выделения.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ПОДАВЛЕНИЯ АНГИОГЕНЕЗА | 2010 |
|
RU2786127C2 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ ИСХОДОВ ПЛОСКОКЛЕТОЧНЫХ ОПУХОЛЕЙ ГОЛОВЫ И ШЕИ | 2009 |
|
RU2426991C2 |
| УСТРОЙСТВА И СПОСОБЫ ДЛЯ ОТБОРА КЛЕТОК, УСТОЙЧИВЫХ К АПОПТОТИЧЕСКИМ СИГНАЛАМ, И ОБЛАСТИ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2630301C2 |
| АНТИТЕЛО, СЕЛЕКТИВНОЕ В ОТНОШЕНИИ РЕЦЕПТОРА ЛИГАНДА, ИНДУЦИРУЮЩЕГО АПОПТОЗ И СВЯЗАННОГО С ФАКТОРОМ НЕКРОЗА ОПУХОЛИ, И ЕГО ПРИМЕНЕНИЕ | 2002 |
|
RU2313537C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИНГИБИРОВАНИЯ КЛЕТОЧНОЙ АДГЕЗИИ ИЛИ НАПРАВЛЕННОСТИ ДИАГНОСТИЧЕСКИХ ИЛИ ТЕРАПЕВТИЧЕСКИХ АГЕНТОВ К RGD СВЯЗЫВАЮЩИМ САЙТАМ | 2010 |
|
RU2642625C2 |
| Способ прогнозирования течения светлоклеточного почечно-клеточного рака | 2022 |
|
RU2792250C1 |
| АНТАГОНИСТЫ ИНТЕГРИНОВЫХ РЕЦЕПТОРОВ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2721907C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ПРИМЕНИМЫЕ ДЛЯ МОДУЛЯЦИИ АНГИОГЕНЕЗА С ИСПОЛЬЗОВАНИЕМ ПРОТЕИНКИНАЗЫ RAF И RAS | 2000 |
|
RU2257911C2 |
| AXL-АНТИТЕЛА | 2008 |
|
RU2506276C2 |
| ПОЛИПЕПТИД СО СВОЙСТВАМИ АГОНИСТА РЕЦЕПТОРА ГОРМОНА РОСТА, КОДИРУЮЩАЯ ЕГО НУКЛЕИНОВАЯ КИСЛОТА, ВЕКТОР ДЛЯ ЕГО ЭКСПРЕССИИ И ПРОДУЦИРУЮЩАЯ ЕГО КЛЕТКА | 2005 |
|
RU2391353C2 |




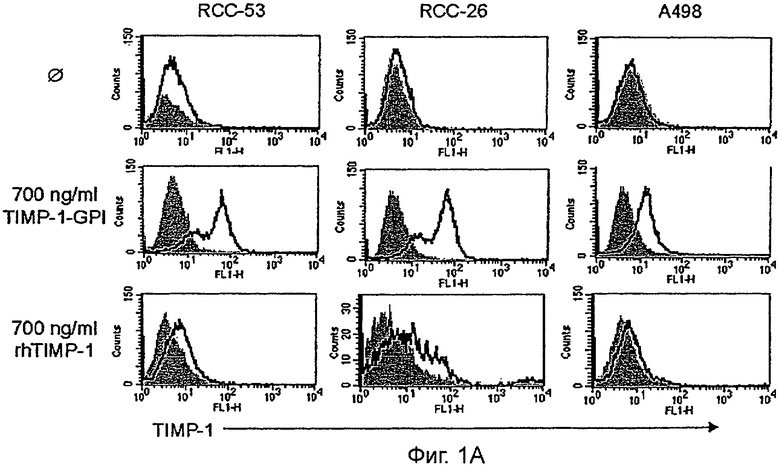
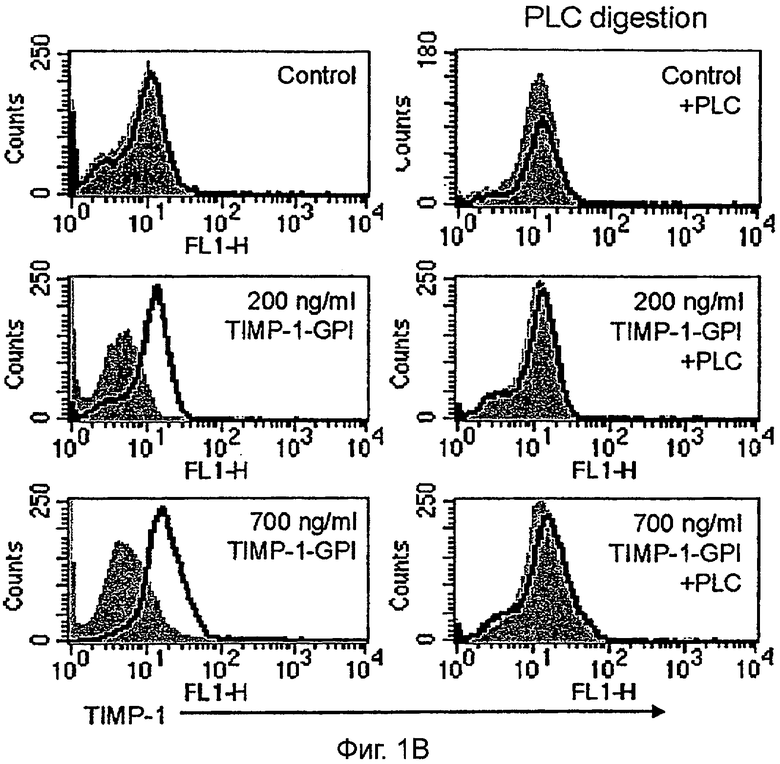
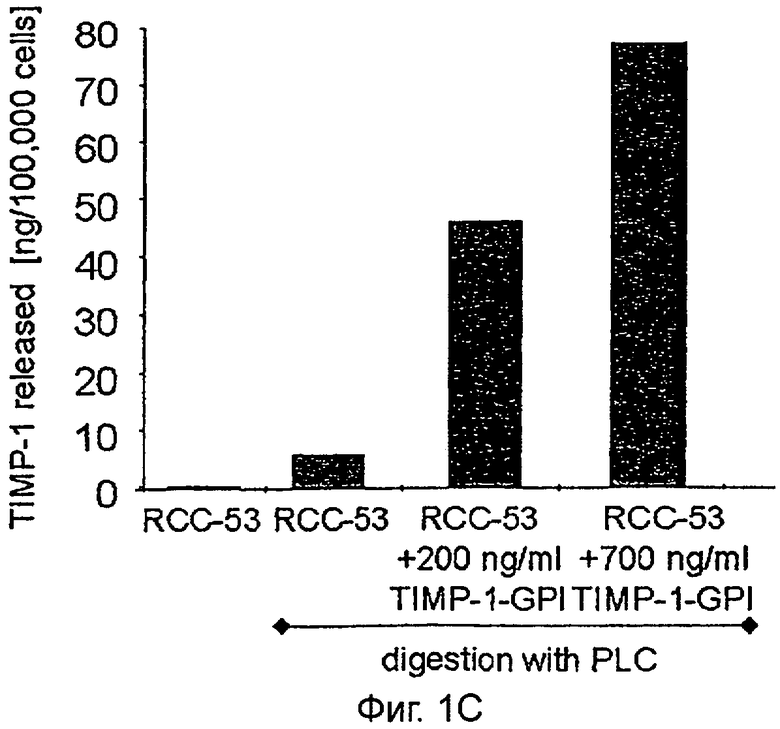
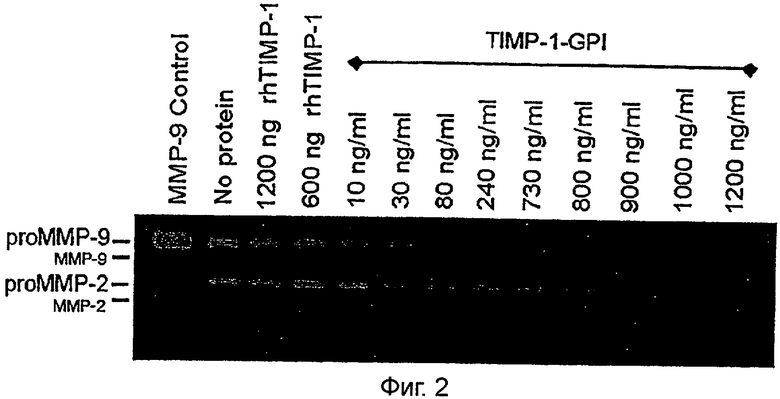
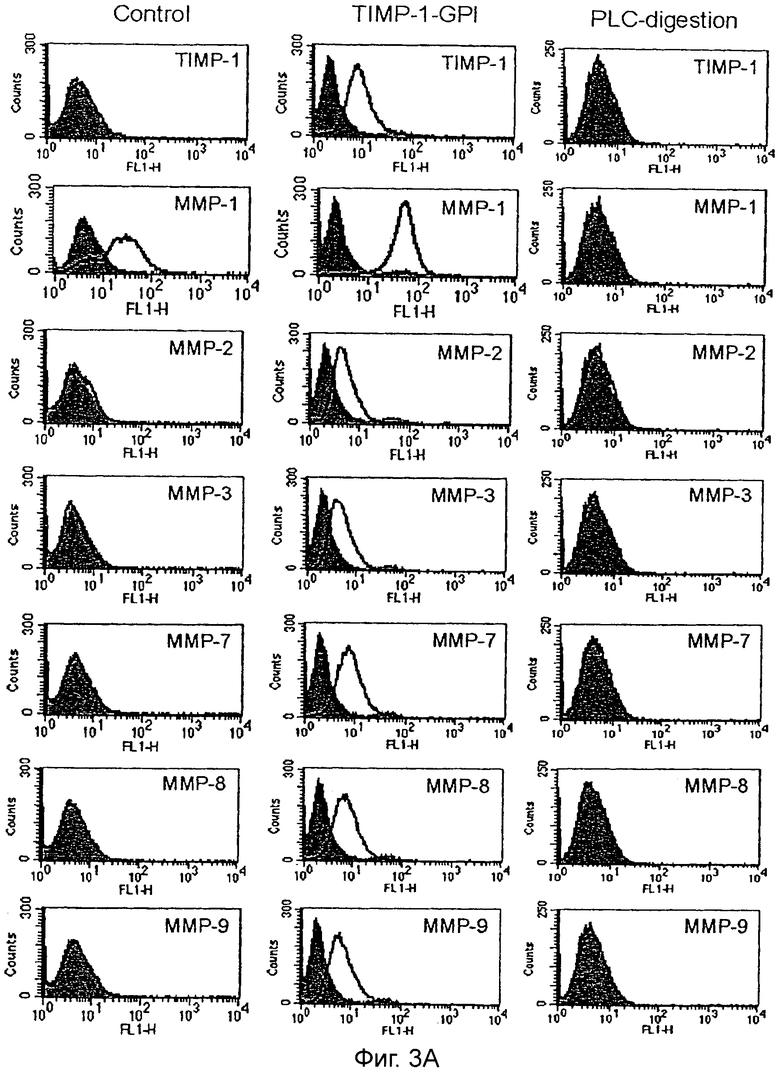
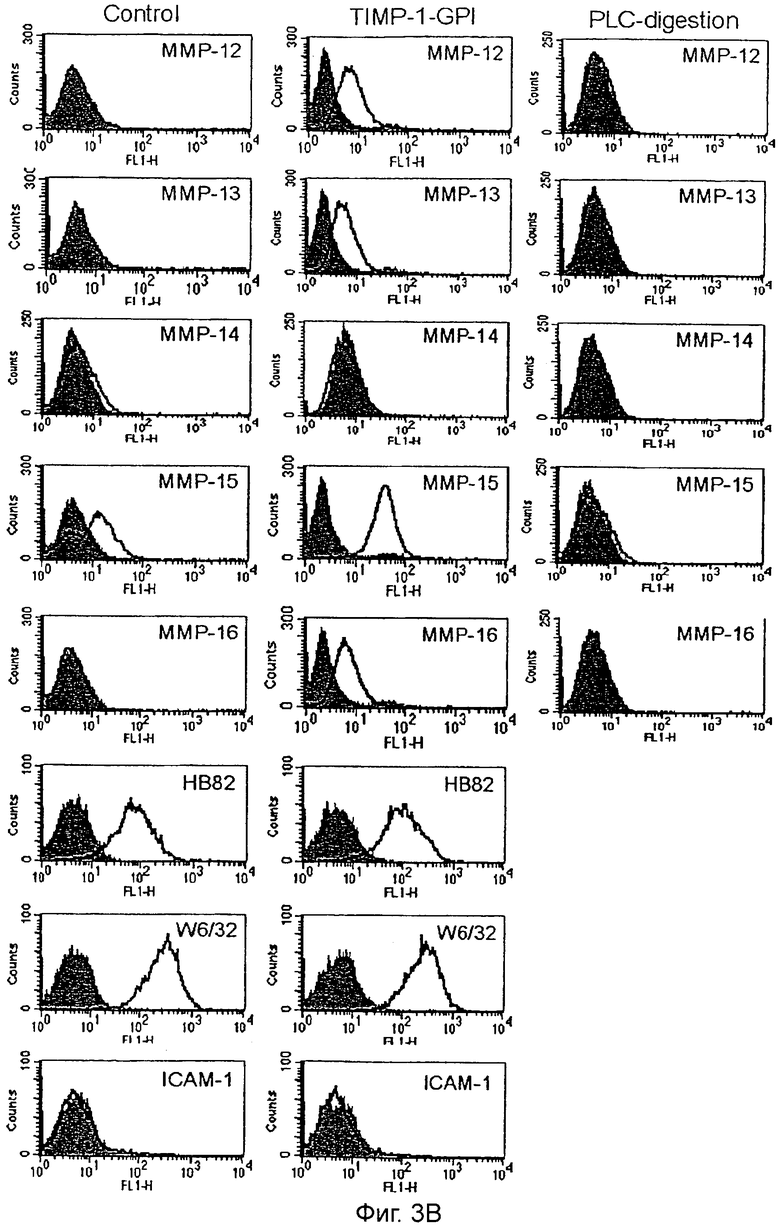
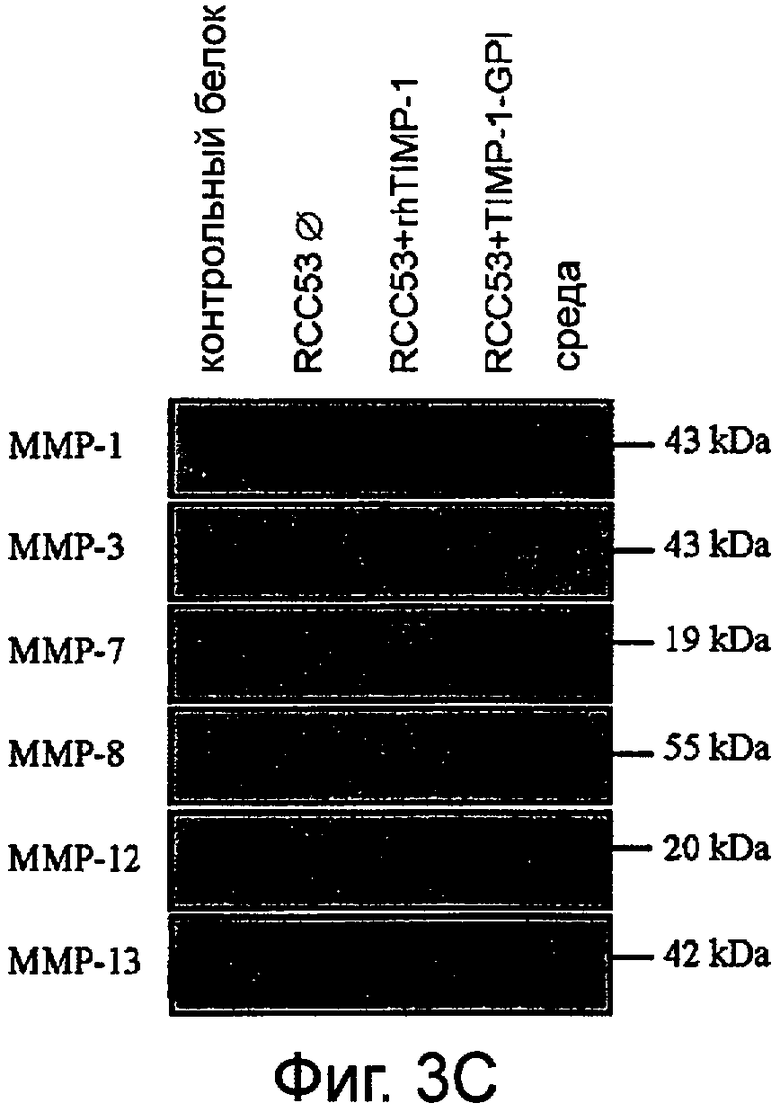
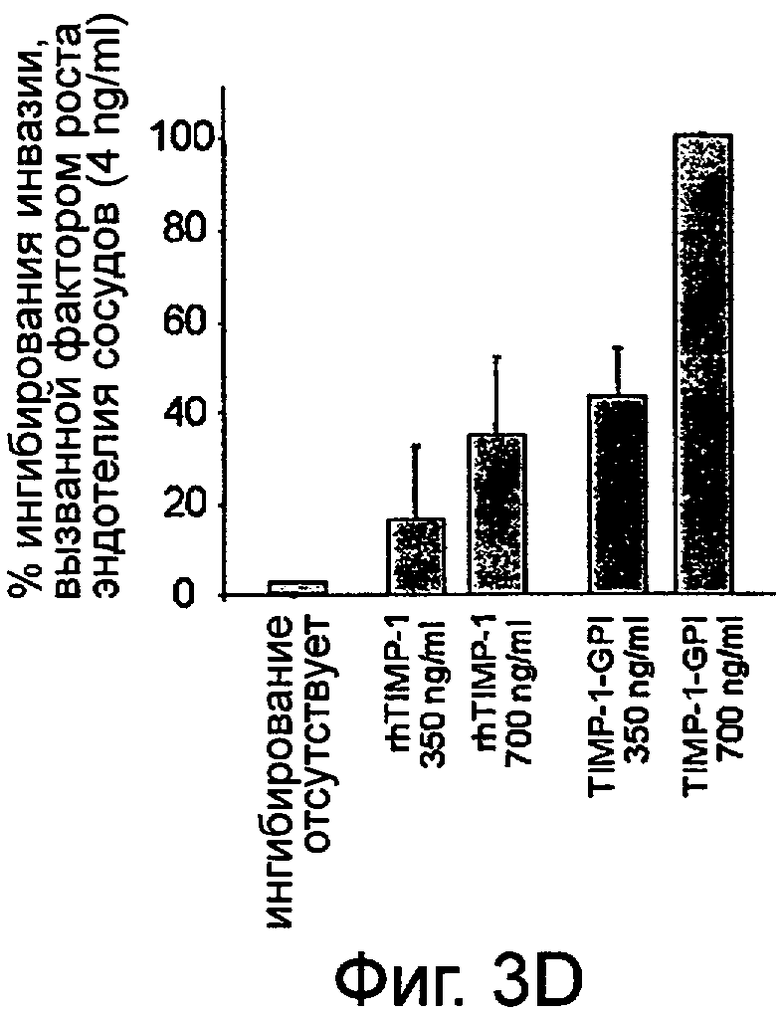
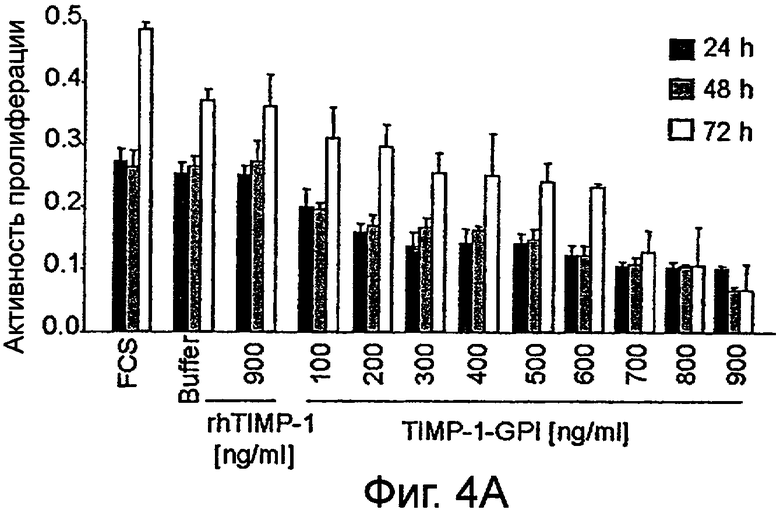
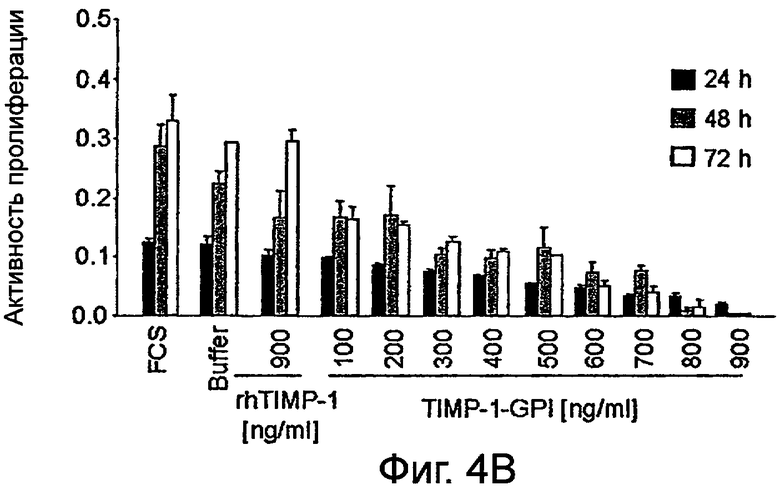
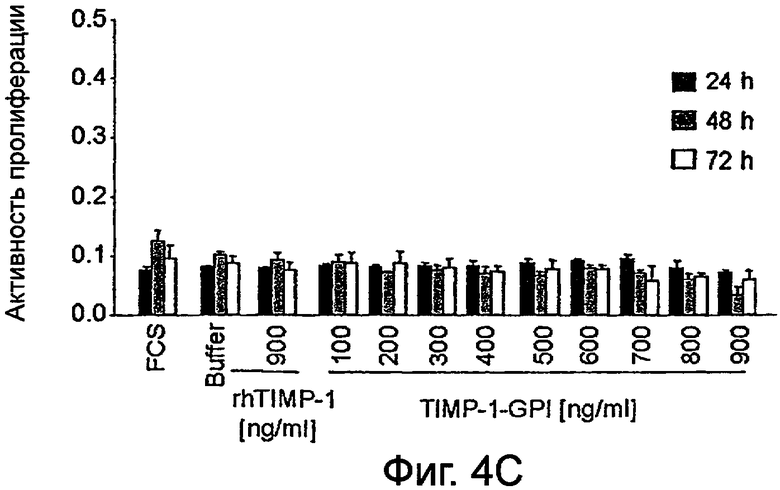
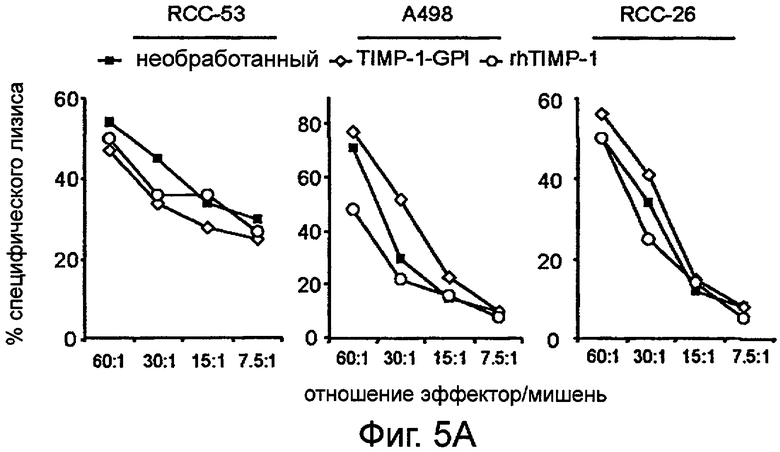
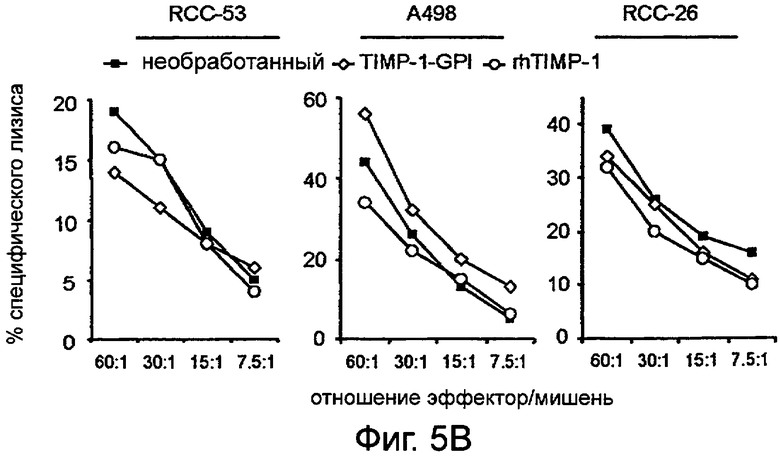
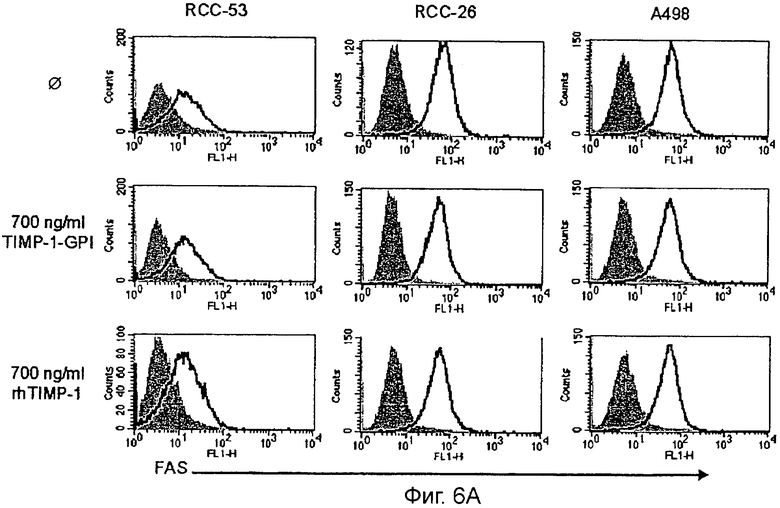
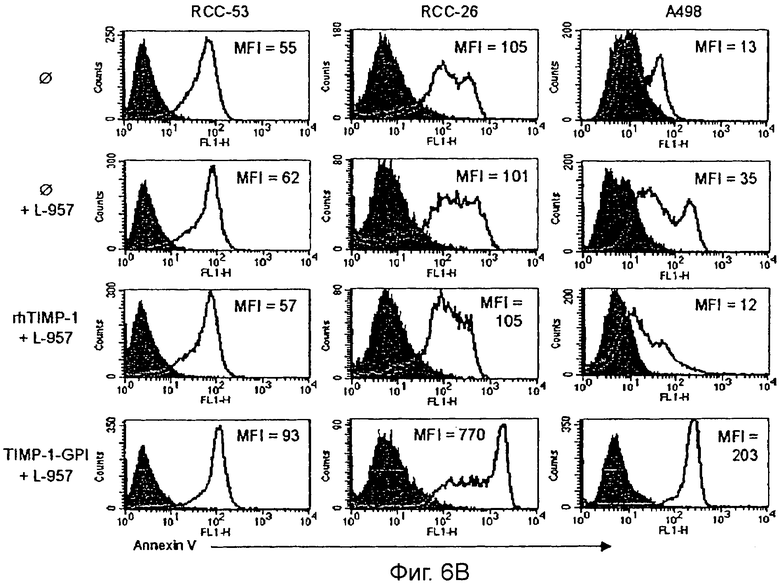
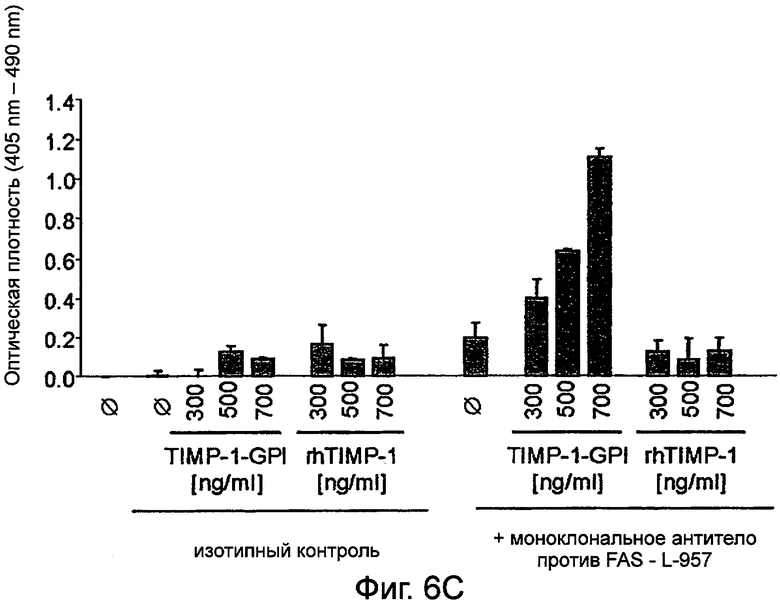
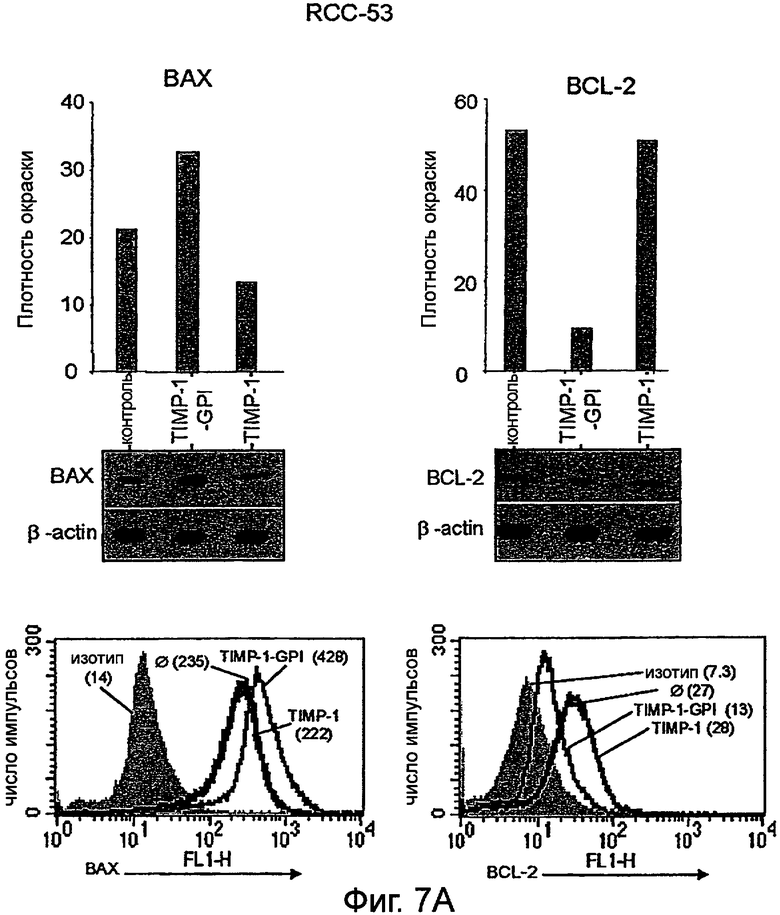
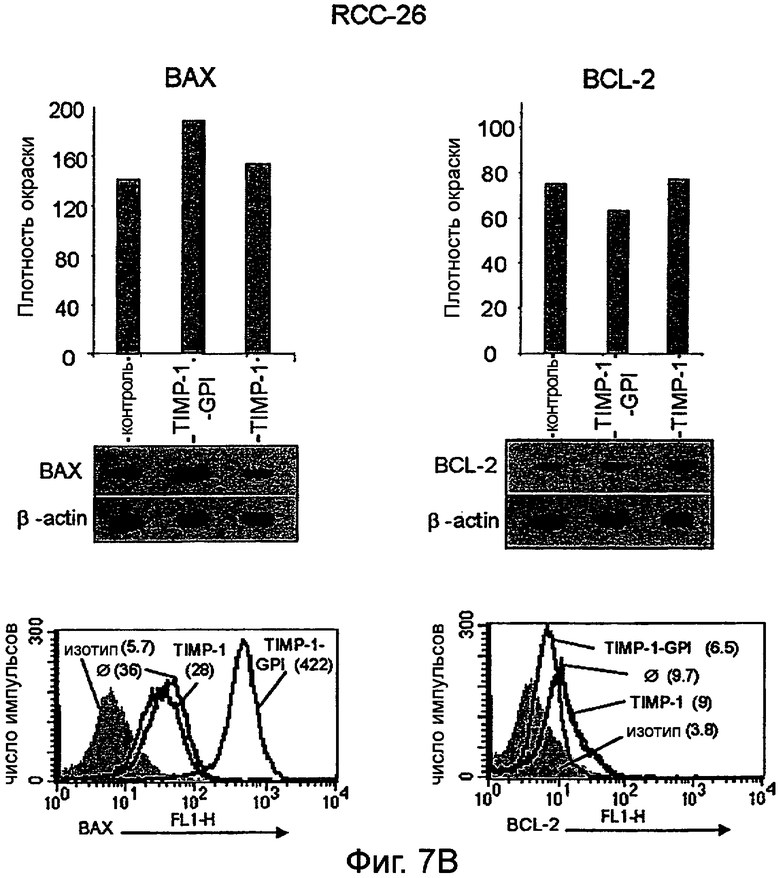
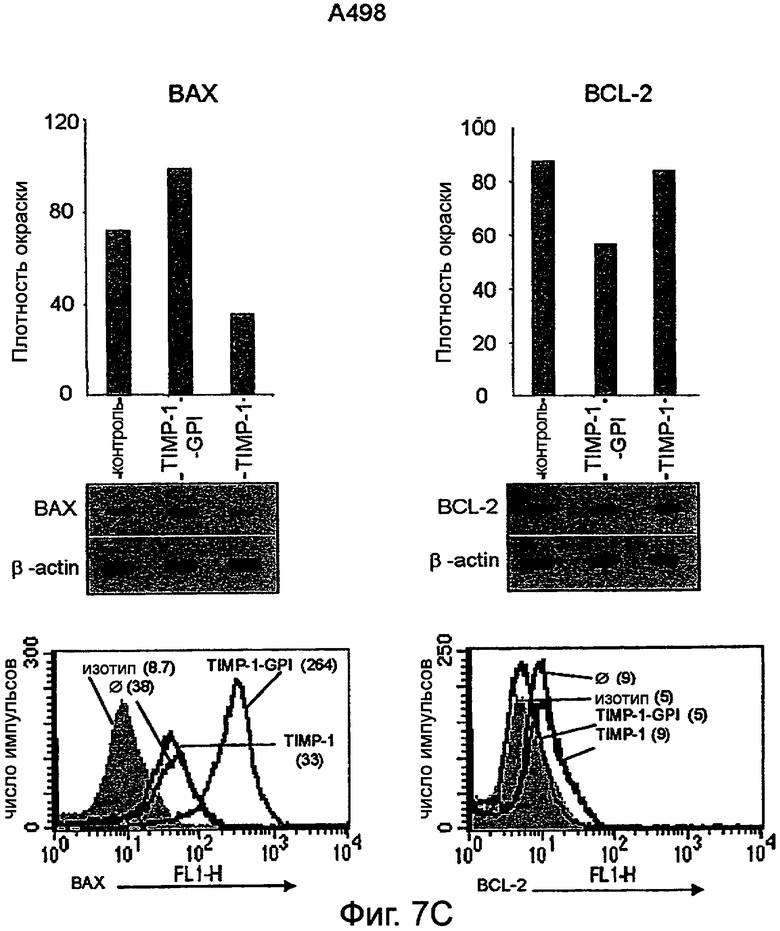
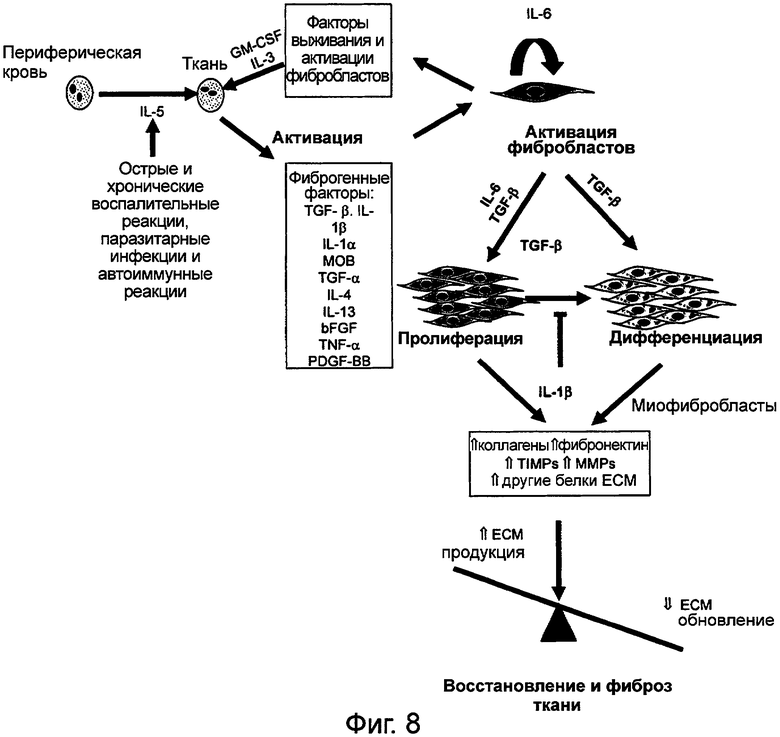
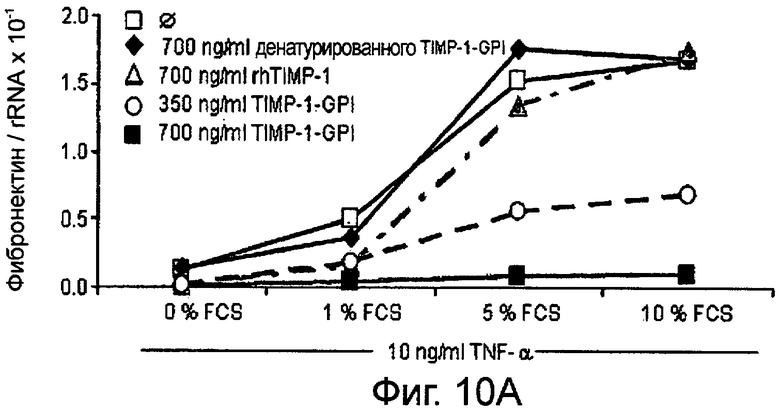
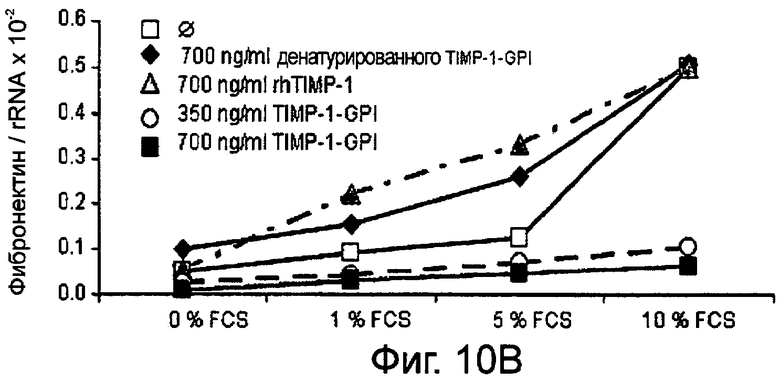
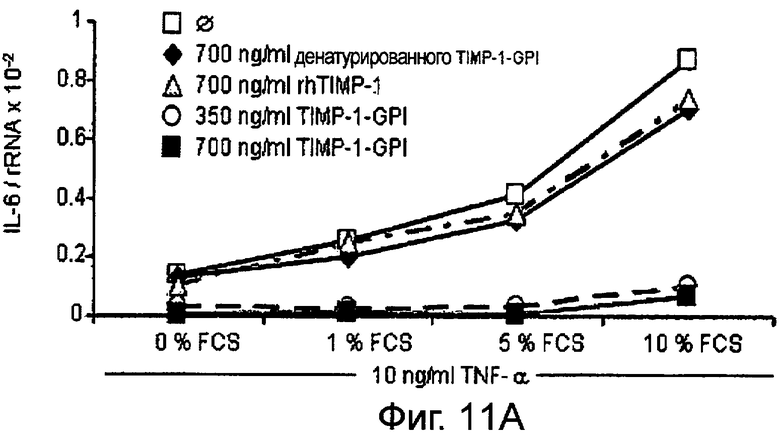
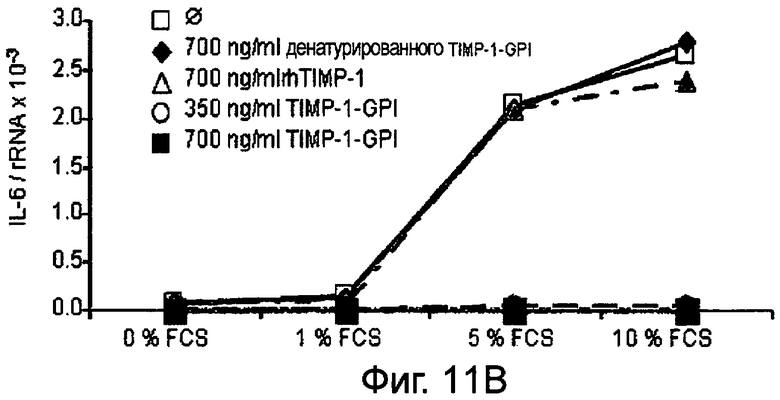
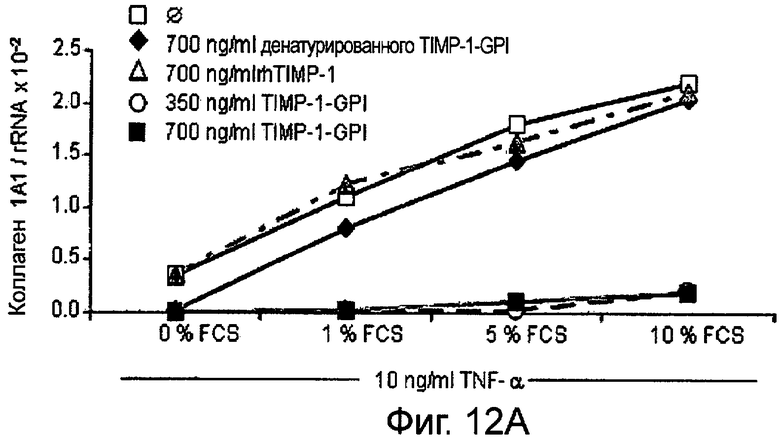
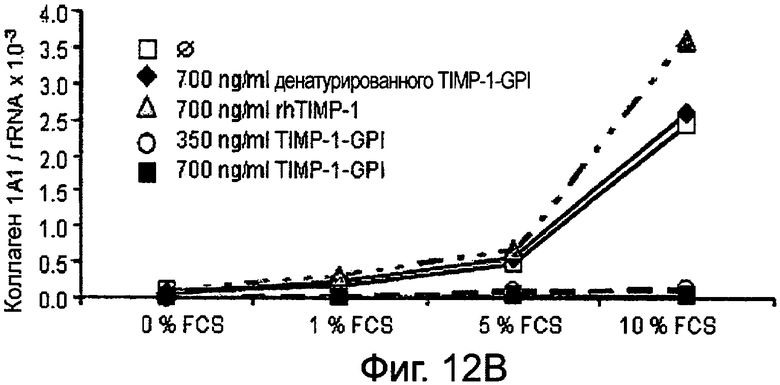
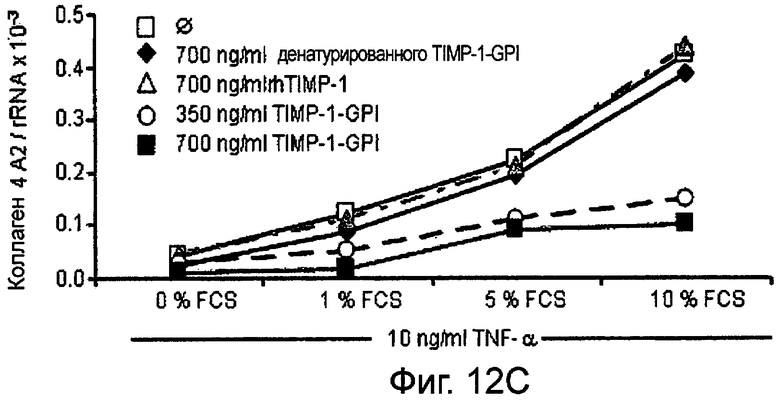
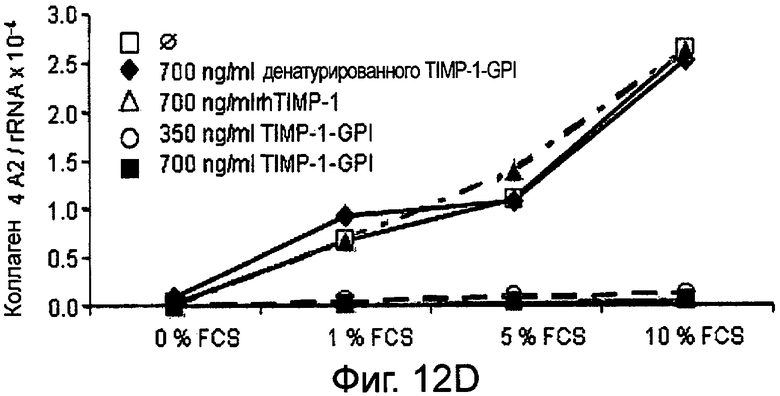
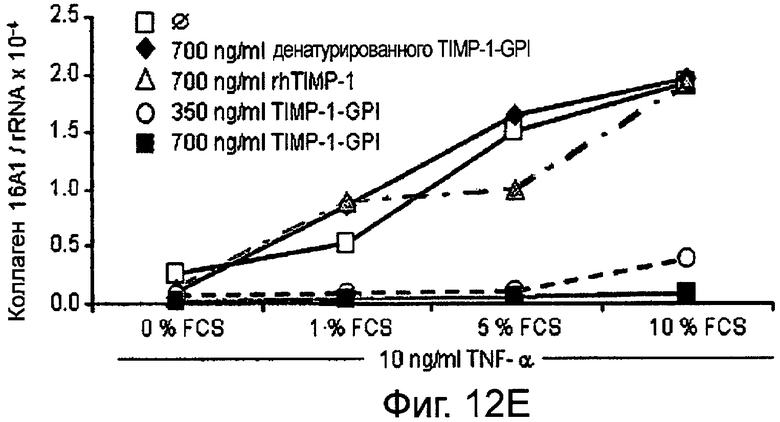
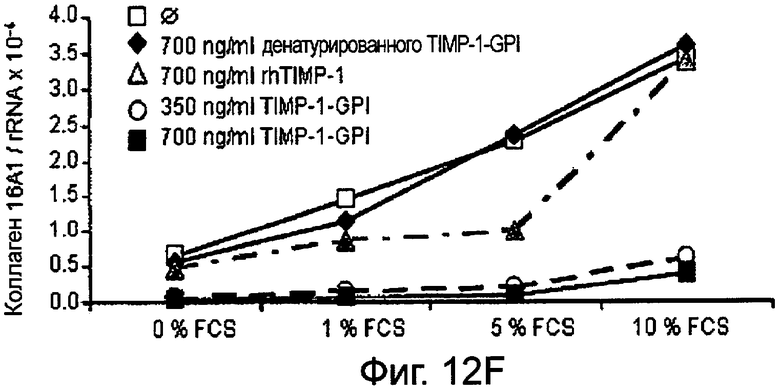
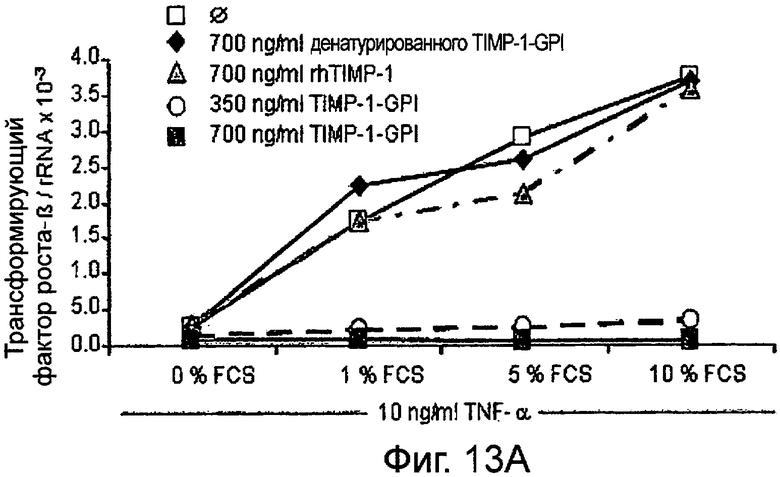
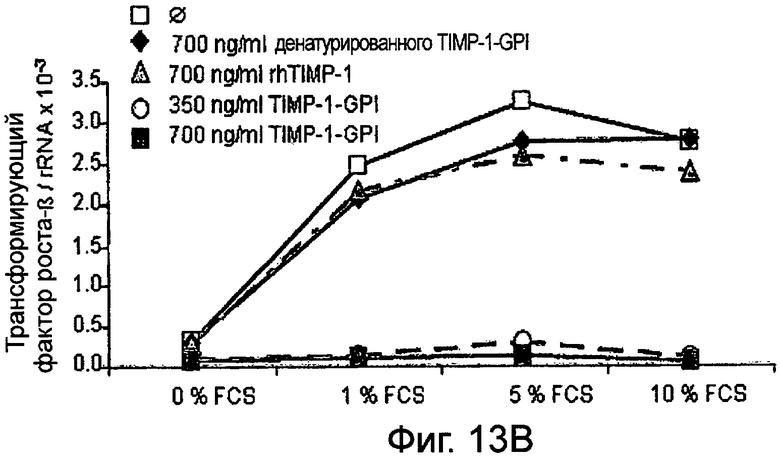
Изобретение относится к области биотехнологии, конкретно к получению слитых белков, и может быть использовано в медицине. Изобретение позволяет предотвратить образование рубца в процессе лечения повреждений кожи при использовании слитого конструкта, состоящего из аминокислотной последовательности тканевого ингибитора металлопротеиназ (ТИМП), связанного с якорем гликозилфосфатидилинозита (ГФИ). 2 н. и 8 з.п. ф-лы, 33 ил.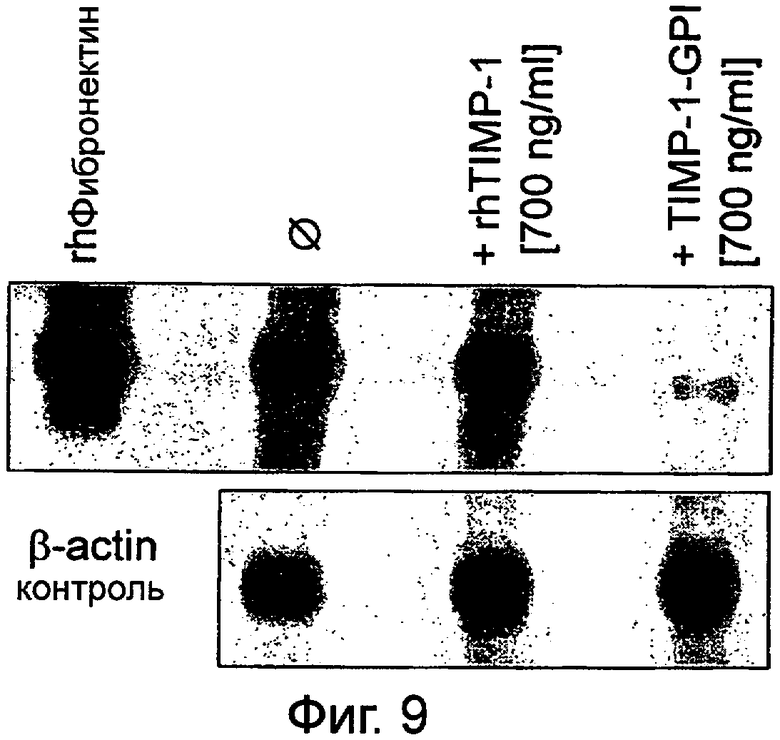
1. Применение слитого конструкта, включающего аминокислотную последовательность тканевого ингибитора металлопротеиназ (ТИМП) или его биологически активного фрагмента, где биологически активный фрагмент ингибирует активные матриксы металлопротеиназ и кодируется любой из последовательностей SEQ ID NO 1 или 4, в которой указанный ТИМП или его биологически активный фрагмент связан с якорем гликозилфосфатидилинозита (ГФИ) для приготовления лекарственного средства для лечения повреждений кожи для предотвращения или ингибирования образования рубца.
2. Применение по п.1, в котором указанный ТИМП выбран из группы, состоящей из ТИМП-1, ТИМП-2, ТИМП-3 или ТИМП-4.
3. Применение по п.1, в котором указанный ТИМП представляет собой человеческий ТИМП-1.
4. Применение по п.1, в котором указанный слитый конструкт содержит одну или больше сигнальных последовательностей ГФИ, чтобы направить фиксацию ГФИ.
5. Применение по п.1, в котором указанный ГФИ-якорь получают из антигена, связанного с функцией лимфоцитов (LFA-3), или его части.
6. Применение по п.1 для предотвращения или ингибирования образования рубца, в котором рубец представляет собой гипертрофический рубец или келоидное образование.
7. Применение по п.1, в котором лекарственное средство применяют вместе с детергентом, герметиком или носителем.
8. Способ лечения повреждения кожи для предотвращения или ингибирования образования рубца, включающий нанесение на участок поврежденной кожи или рубца фармацевтического состава, содержащего слитый конструкт, включающий аминокислотную последовательность тканевого ингибитора металлопротеиназ (ТИМП) или его биологически активного фрагмента, где биологически активный фрагмент ингибирует активные матриксы металлопротеиназ и кодируется любой из последовательностей SEQ ID NO 1 или 4, в которой указанный ТИМП или его биологически активный фрагмент связан с якорем гликозилфосфатидилинозита (ГФИ), и фармацевтически приемлемый носитель.
9. Способ по п.8, в котором рубец представляет собой гипертрофический рубец или келоидное образование.
10. Способ по п.8, в котором лекарственное средство применяют вместе с детергентом, герметиком или носителем.
| DJAFARZADEH R | |||
| et al., Exogenously added GPI-anchored tissue inhibitor of matrix metalloproteinase-1 (TIMP-1) displays enhanced and novel biological activities, Biol | |||
| Chem., 2004, v.385, n.7, p.655-663 | |||
| GILLARD J.A | |||
| et al., Matrix metalloproteinase activity and immunohistochemical profile of matrix metalloproteinase-2 and -9 and tissue inhibitor of |

