Данное изобретение относится к новым (β-)глюканазам (и, таким образом, также к целлюлазам) из гриба Talaromyces, в частности Talaromyces emersonii, и к их применению для расщепления глюкана в целлюлозе.
Предпосылка изобретения
Состав клеточной стенки растений сложен и изменчив. Полисахариды главным образом выявляют в форме длинных цепей целлюлозы (основного структурного компонента клеточной стенки растений), гемицеллюлозы (содержащей различные цепи β-ксилана, такие как ксилоглюканы) и пектина.
Бета-глюкан (или более точно, (1→3)(1→4)β-D-глюкан), М.м. от 10 до 14 кДа, является компонентом гемицеллюлозы растений (М.м. примерно 700 кДа) и состоит из основной цепи остатков глюкопиранозы, соединенных β-1,4 и 1,3-связями. Другим компонентом является ксилан, который состоит из β-1,4-связанных остатков ксилозы, необязательно замещенных боковыми цепями, такими как остатки арабинозы и/или глюкуроновой кислоты.
Основные различия существуют между однодольными (например, злаковые и травы) и двудольными (например, клевер, рапс и соя) и между семенами и вегетативными частями растения. Однодольные характеризуются наличием арабиноксиланового комплекса в качестве основной цепи гемицеллюлозы, а основная структура гемицеллюлозы у двудольных представляет собой ксилоглюкановый комплекс. У двудольных обнаружены более высокие концентрации пектина, чем у однодольных. В семенах, как правило, высокое содержание пектиновых веществ, но относительно низкое содержание целлюлозного материала.
Ферменты, разрушающие целлюлозу, используют для обработки растительного материала в корме, а также в пищевых добавках, или в качестве корма или пищевой добавки благодаря их способности действовать на основные компоненты клеточной стенки растений.
Большинство разрушающих целлюлозу ферментов, имеющихся в промышленности, по-видимому, являются глюканазами с относительно низкой молекулярной массой и средней стабильностью при высоких температурах. Однако для некоторых применений желательно использовать глюканазу с относительно высокой термостабильностью. В том случае, если глюканазу необходимо использовать в качестве пищевой добавки для животных, то высокая термостабильность предпочтительна из-за высокотемпературных условий, применяемых во время гранулирования корма для животных.
Сущность изобретения
В данном изобретении представлены новые β-глюканазы (или целлюлазы), которые способны расщеплять β-D-глюкан (или целлюлозу), такой, который присутствует в растительном материале.
В самом широком смысле изобретение относится к β-глюканазам (или целлюлазам) из Talaromyces, такого как Talaromyces emersonii (гриб). Указанные β-глюканазы могут расщеплять β-D-глюкан или целлюлозу, например, они являются β-1,4-эндоглюканазами, или обладают активностью EC 3.2.1.4.
Глюканазы могут иметь:
а. оптимум рН ниже 7,0 или 5,4, такой как ниже 5,0, например от 4,4 до 5,2 или от 3,0 до 6,0 или 7,0; или
b. оптимум температуры, по меньшей мере, 72, 75 или даже 81°С, такой как от 78 до 85°С или от 83 до 87°С.
Более конкретно данное изобретение относится к (выделенному) полипептиду β-глюканазы, содержащему:
(i) аминокислотную последовательность SEQ ID NO: 2, 4 или 6; или
(ii) вариант (i), который способен расщеплять β-D-глюкан; или
(iii) фрагмент (i) или (ii), который способен расщеплять β-D-глюкан.
Согласно другому аспекту изобретение относится к полинуклеотиду, который содержит:
(а) последовательность нуклеиновой кислоты SEQ ID NO: 1, 3 или 5, или последовательность, кодирующую полипептид согласно изобретению;
(b) последовательность, которая комплементарна, или которая гибридизуется с любой последовательностью, охарактеризованной в (а);
(с) фрагмент любой последовательности по п.(а) или (b);
(d) последовательность, обладающая, по меньшей мере, 60% идентичностью с любой последовательностью, которая определена в (а), (b) или (c); или
(e) последовательность, которая является вырожденной в результате свойства генетического кода по отношению к любой из последовательностей, которые определены в пп. с (а) по (d).
Изобретение также относится к следующему:
- вектор (например, экспрессирующий), который содержит полинуклеотид согласно изобретению и который может обладать способностью экспрессировать полипептид согласно изобретению;
- линия клеток или штамм, содержащий вектор согласно изобретению;
- способ получения полипептида согласно изобретению, причем указанный способ включает в себя поддержание линии клеток или штамма согласно изобретению в условиях, подходящих для достижения экспрессии полипептида, и, при необходимости, выделение полипептида;
- способ расщепления β-D-глюкана, причем указанный способ включает в себя осуществление контакта материала, содержащего β-D-глюкан, с полипептидом согласно изобретению;
- способ идентификации соединения, которое модулирует активность β-глюканазы, причем указанный способ включает в себя осуществление контакта полипептида согласно изобретению с тестируемым соединением в присутствии β-D-глюкана и наблюдение или регистрацию любой модуляции активности;
- способ лечения субъекта с гиперлипидемией, причем указанный способ включает в себя введение субъекту эффективного количества полипептида согласно изобретению; и
- применение полипептида в производстве лекарственного средства для лечения или профилактики гиперлипидемии.
Краткое описание последовательностей
SEQ ID NO: 1 представляет собой последовательность ДНК первой β-глюканазы (СЕА) согласно изобретению из Talaromyces emersonii;
SEQ ID NO: 2 является аминокислотной последовательностью первой β-глюканазы (СЕА);
SEQ ID NO: 3 является последовательностью ДНК второй β-глюканазы (СЕВ) из того же организма;
SEQ ID NO: 4 является аминокислотной последовательностью второй β-глюканазы (СЕВ);
SEQ ID NO: 5 является последовательностью ДНК третьей β-глюканазы (СЕС) также из того же самого организма;
SEQ ID NO: 6 является аминокислотной последовательностью третьей β-глюканазы (СЕС); и
SEQ ID NO: 7 и 8 представляют собой праймеры ПЦР, которые гибридизуются с SEQ ID NO: 1 (и были использованы для того, чтобы повторно клонировать вставки кДНК в ходе генетического анализа позитивных трансформантов).
Подробное описание изобретения
А. Полинуклеотиды.
Данное изобретение относится к полинуклеотиду (например, выделенному и/или очищенному), кодирующему полипептиды согласно изобретению. Таким образом, данное изобретение относится к полинуклеотиду, кодирующему β-глюканазу, аминокислотная последовательность которой указана в SEQ ID NO: 2, 4 или 6. Кроме того, данное изобретение относится к полинуклеотиду, кодирующему полипептид, обладающий значительной гомологией аминокислотной последовательности с аминокислотной последовательностью, указанной в SEQ ID NO: 2, 4 или 6. Также в изобретение включен полинуклеотид, выбранный из следующей группы:
(а) полинуклеотид, содержащий нуклеотидную последовательность, указанную в SEQ ID NO: 1, 3 или 5, или комплементарную ей последовательность;
(b) полинуклеотид, содержащий нуклеотидную последовательность, способную гибридизоваться (например, избирательно) с нуклеотидной последовательностью, указанной в SEQ ID NO: 1, 3 или 5, или ее фрагмент;
(с) полинуклеотид, содержащий нуклеотидную последовательность, способную гибридизоваться (например, избирательно) с последовательностью, комплементарной нуклеотидной последовательности, указанной в SEQ ID NO: 1, 3 или 5, или ее фрагмент; и/или
(d) полинуклеотид, содержащий полинуклеотидную последовательность, которая является вырожденной в результате свойства генетического кода по отношению к полинуклеотиду, охарактеризованному в (а), (b) или (с).
Полинуклеотид согласно изобретению также включает в себя полинуклеотид, который:
(а) кодирует полипептид, обладающий активностью β-глюканазы, и этот полинуклеотид представляет собой
(1) кодирующую последовательность SEQ ID NO: 1, 3 или 5;
(2) последовательность, которая избирательно гибридизуется с комплементом последовательности, определенной в (1); или
(3) последовательность, которая является вырожденной в результате свойства генетического кода по отношению к последовательности, определенной в (1) или (2); или
(b) является последовательностью, комплементарной полинуклеотиду, определенному в (а).
Гибридизующиеся последовательности
Термин "способен к гибридизации" означает, что полинуклеотид-мишень согласно изобретению может гибридизоваться с нуклеиновой кислотой, используемой в качестве зонда (например, нуклеотидной последовательностью, указанной в SEQ ID NO: 1, 3 или 5, или ее фрагментом или ее комплементом) на уровне, значимо превышающем фон. Изобретение также включает в себя нуклеотидные последовательности, которые кодируют β-глюканазу или ее варианты, а также нуклеотидные последовательности, которые им комплементарны. Нуклеотидная последовательность может представлять собой РНК или ДНК, и таким образом, включает геномную ДНК, синтетическую ДНК или кДНК. Предпочтительно нуклеотидная последовательность является последовательностью ДНК и наиболее предпочтительно последовательностью кДНК. Обычно полинуклеотид согласно изобретению содержит непрерывную последовательность нуклеотидов, которая при селективных условиях способна соответствующим образом гибридизоваться с кодирующей последовательностью или последовательностью, комплементарной кодирующей последовательности SEQ ID NO: 1, 3 или 5. Такие нуклеотиды можно синтезировать способами, известными в данной области1.
Полинуклеотид согласно изобретению может гибридизоваться с кодирующей последовательностью или последовательностью, комплементарной кодирующей последовательности SEQ ID NO: 1, 3 или 5 (соответствующим образом) на уровне, значимо превышающем фон. Фоновая гибридизация может иметь место, например, вследствие наличия в библиотеке кДНК других кДНК. Уровень сигнала (например, создаваемого при взаимодействии между полинуклеотидом согласно изобретению и кодирующей последовательностью или комплементом кодирующей последовательности SEQ ID NO: 1, 3 или 5) обычно, по меньшей мере, в 10 раз, предпочтительно, по меньшей мере, в 100 раз интенсивнее, чем взаимодействия между другими полинуклеотидами и кодирующей последовательностью SEQ ID NO: 1, 3 или 5. Интенсивность взаимодействия можно, например, измерить, с помощью радиоактивного мечения зонда, например, 32Р. Избирательной гибридизации обычно можно достичь, используя условия низкой жесткости (0,3 М хлорид натрия и 0,03 М цитрат натрия примерно при 40°С), средней жесткости (например, 0,3 М хлорид натрия и 0,03 М цитрат натрия примерно при 50°С) или высокой жесткости (например, 0,3 М хлорид натрия и 0,03 М цитрат натрия примерно при 60°С). Гибридизацию можно проводить при любых подходящих условиях, известных в данной области1, и в качестве инструкции условиями низкой жесткости могут быть 2 × SSC при 55°С, условиями средней жесткости могут быть от 0,5 до 1,0 × SSC при 60°С и условиями высокой жесткости могут быть 0,1 или 0,2 × SSC при 60°С или выше (например, при 68°С), во всех случаях в 0,5% SDS.
Модификации
Полинуклеотиды согласно изобретению могут содержать ДНК или РНК. Они могут быть одноцепочечными или двухцепочечными. Они также могут быть полинуклеотидами, которые содержат внутри себя один или несколько синтетических или модифицированных нуклеотидов. В данной области известен ряд разных типов модификаций полинуклеотидов. Модификации включают метилфосфатную или фосфоротиоатную основную цепь и/или присоединение акридина или цепей полилизина на 3'- или 5'- концах молекулы. Следует понимать, что для целей данного изобретения описанные здесь полинуклеотиды могут быть модифицированы любым способом, имеющимся в данной области.
Следует понимать, что специалисты в данной области, используя обычные способы, могут делать нуклеотидные замены, которые не влияют на последовательности полипептида, кодируемого полинуклеотидами согласно данному изобретению, чтобы отразить использование кодонов любым конкретным организмом-хозяином, например организмом, в котором необходимо экспрессировать полипептиды согласно изобретению.
Кодирующую последовательность SEQ ID NO: 1, 3 или 5 можно модифицировать нуклеотидными заменами, например, от или в пределах 1, 2 или 3 до 10, 25, 59 или 100 замен. Полинуклеотид альтернативно или дополнительно можно модифицировать одной или несколькими инсерциями, и/или делециями, и/или удлинением любого или обоих концов. Модифицированный полинуклеотид, как правило, кодирует полипептид, который обладает β-глюканазной активностью. Можно произвести вырожденные замены и/или можно сделать замены, результатом которых при трансляции модифицированной последовательности могла бы быть консервативная замена аминокислоты, например, как обсуждается при обращении к полипептидам далее.
Гомологи
Нуклеотидная последовательность, которая способна избирательно гибридизоваться (например, комплементарная последовательность) с кодирующей последовательностью ДНК SEQ ID NO: 1, 3 или 5, может обладать 50% или 60%, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 98% или, по меньшей мере, 99% идентичностью последовательности (или гомологией) с кодирующей последовательностью SEQ ID NO: 1, 3 или 5. Это может иметь место на протяжении района, по меньшей мере, из 20, предпочтительно, по меньшей мере, из 30, например, по меньшей мере, из 40, по меньшей мере, из 60, более предпочтительно, по меньшей мере, из 100 следующих друг за другом нуклеотидов, или оптимально на протяжении полной длины SEQ ID NO: 1, 3 или 5. Для SEQ ID NO: 1 идентичность последовательностей предпочтительно составляет, по меньшей мере, 75% или 80%, для SEQ ID NO: 3 предпочтительно, по меньшей мере, 85% и для SEQ ID NO: 5 предпочтительно, по меньшей мере, 85%.
Любую комбинацию указанных выше степеней гомологии и минимальных размеров можно использовать для того, чтобы определить полинуклеотиды согласно изобретению, при этом предпочтительны более жесткие комбинации (т.е. более высокая гомология при больших длинах участков). Таким образом, например, полинуклеотид, который, по меньшей мере, на 80% или 90% гомологичен на протяжении 25, предпочтительно на протяжении 30 нуклеотидов, составляет один аспект изобретения, как и полинуклеотид, который, по меньшей мере, гомологичен на 90% на протяжении 40 нуклеотидов.
Гомологи полинуклеотидных (или белковых) последовательностей обычно обладают, по меньшей мере, 70% гомологией, предпочтительно, по меньшей мере, 80%, 90%, 95%, 97% или 99% гомологией, например, на протяжении района, по меньшей мере, из 15, 20, 30, 100 и более следующих друг за другом нуклеотидов (или аминокислот). Гомологию можно рассчитать на основе идентичности аминокислот (иногда называемой "строгой гомологией").
Пакет программ UWGCG, например, предоставляет программу BESTFIT, которую можно использовать для расчета гомологии (например, используя ее при настройках по умолчанию5). Алгоритмы PILEUP и BLAST можно использовать для расчета гомологии или выравнивания последовательностей (а именно для идентификации эквивалента или соответствующих последовательностей, например при их настройках по умолчанию6,7).
Компьютерная программа для выполнения BLAST-анализа находится в открытом доступе через National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/). Указанный алгоритм включает в себя сначала идентификацию пары последовательностей с высоким количеством очков (HSP) благодаря идентификации коротких слов длиной W в запрашиваемой последовательности, которые либо соответствуют, либо удовлетворяют некоторому пороговому количеству положительных очков Т при выравнивании со словом такой же длины в последовательности базы данных. Т называют порогом количества очков для слов в определенной окрестности6,7. Указанные исходные совпадения соседних слов в окрестности действуют как начало для инициации поисков, для того чтобы найти содержащие их HSP. Поиск совпадения слов продолжают в обоих направлениях вдоль каждой последовательности до тех пор, пока суммарное количество очков выравнивания можно увеличивать. Продолжение поиска совпадения слов в каждом направлении прекращают, когда: суммарное количество очков выравнивая уменьшается на количество Х по сравнению с его достигнутым максимальным значением; суммарное количество очков опускается до нуля или ниже, вследствие суммирования одного или нескольких отрицательно оцениваемых выравниваний остатков; или достигается конец какой-либо последовательности. Параметры W, T и Х алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLAST по умолчанию используется длина слов (W), равная 11, выравнивания в матрице подсчета очков BLOSUM 628 (B) - 50, ожидание (Е) - 10, M=5, N=4 и сравнение обоих нитей.
Алгоритм BLAST выполняет статистический анализ сходства между двумя последовательностями9. Одной из мер сходства, даваемых алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), которая является показанием вероятности, с которой случайно может иметь место совпадение между двумя нуклеотидными или аминокислотными последовательностями. Например, последовательность считается сходной с другой последовательностью, если наименьшая суммарная вероятность при сравнении первой последовательности со второй последовательностью примерно составляет меньше 1, предпочтительно примерно меньше 0,1, более предпочтительно примерно меньше 0,01 и наиболее предпочтительно примерно меньше 0,001.
Праймеры и зонды
Полинуклеотиды согласно изобретению включают и могут быть использованы в качестве праймера, например ПЦР-праймера, праймера для альтернативной реакции амплификации, зонда, или полинуклеотиды можно клонировать в векторах. Такие праймеры, зонды или другие фрагменты будут иметь длину, по меньшей мере, равную или до 20, например, по меньшей мере, 25, 30 или 40 нуклеотидов. Как правило, они будут составлять в длину до 40, 50, 60, 70, 100, 150, 200 или 300 нуклеотидов, или даже до длины на нескольких нуклеотидов (как, например, 5 или 10 нуклеотидов) короче кодирующей последовательности SEQ ID NO: 1, 3 или 5.
В общем, праймеры будут получены синтетическими способами, включающими в себя поэтапную сборку требуемой последовательности нуклеиновой кислоты по одному нуклеотиду за один раз. Способы выполнения этого с использованием автоматизированных методов легкодоступны в данной области. Примеры праймеров согласно изобретению указаны в SEQ ID NO: 7 и 8.
Более длинные полинуклеотиды, как правило, будут получены с использованием рекомбинантных способов, например с использованием способов клонирования на основе ПЦР (полимеразной цепной реакции). Этот способ будет включать в себя получение пары праймеров (например, длиной примерно 15-30 нуклеотидов) к району β-глюканазы, который необходимо клонировать, осуществление контакта праймеров с мРНК или кДНК, полученной из клетки-мишени (например, дрожжевой, бактериальной, растительной, прокариотической клетки или клетки гриба), предпочтительно штамма Talaromyces, выполнение полимеразной цепной реакции при условиях, которые вызывают амплификацию требуемого района, выделение амплифицированного фрагмента (например, очисткой реакционной смеси в агарозном геле) и извлечение амплифицированной ДНК. Праймеры можно сконструировать так, чтобы они содержали подходящие сайты узнавания ферментами рестрикции, с тем, чтобы амплифицированную ДНК можно было клонировать в подходящем клонирующем векторе.
Такие способы можно использовать для того, чтобы получить всю или часть описанной здесь последовательности β-глюканазы. Геномные клоны, соответствующие кДНК SEQ ID NO: 1, 3 или 5, или гену β-глюканазы, содержащему, например, интроны и промоторные районы, также находятся в рамках изобретения и также могут быть получены аналогичными способами (например, рекомбинантными способами, ПЦР, способами клонирования), исходя из геномной ДНК клетки грибов, дрожжевой, бактериальной, растительной или прокариотической клетки.
Полинуклеотиды или праймеры могут нести обнаруживаемую метку, например, радиоактивную или нерадиоактивную метку. Подходящие метки включают радиоизотопы, такие как 32Р или 35S, ферментные метки или другие белковые метки, такие как биотин. Такие метки можно присоединять к полинуклеотидам или праймерам согласно изобретению и можно выявлять, используя по существу известные способы.
Полинуклеотиды, меченые или немеченые, можно использовать в основанных на нуклеиновых кислотах тестах для выявления или секвенирования β-глюканазы или ее варианта в образце (например, образце гриба). Такие тесты для выявления, как правило, включают в себя осуществление контакта образца (например, образца гриба), (предположительно) содержащего ДНК, с зондом или праймером согласно изобретению в условиях гибридизации и выявление любого дуплекса, образованного между зондом и нуклеиновой кислотой в образце. Такое выявление можно осуществить, используя такие способы, как ПЦР, или посредством иммобилизации зонда на твердой подложке, удаления нуклеиновой кислоты в образце, которая не гибридизуется с зондом, и затем регистрацией нуклеиновой кислоты, которая гибридизовалась с зондом. Альтернативно нуклеиновую кислоту образца можно иммобилизовать на твердой подложке, и можно регистрировать количество зонда, связанного с такой подложкой.
Зонды согласно изобретению можно удобно упаковать в форме тест-набора в подходящей таре. В таких наборах зонд может быть связан с твердой подложкой, в тех случаях, когда форма анализа, для которого предназначен набор, требует такого связывания. Набор также может содержать подходящие реагенты для обработки образца, который необходимо исследовать, гибридизации зонда с нуклеиновой кислотой в образце, реагенты для контроля, инструкции и тому подобное.
Предпочтительно полинуклеотид согласно изобретению получают из того же самого организма, что и полипептид, такого как гриб, в частности гриб рода Talaromyces.
Полинуклеотид согласно изобретению также включает варианты последовательности SEQ ID NO: 1, 3 или 5, которые обладают активностью β-глюканазы. Варианты могут быть образованы присоединениями, заменами и/или делециями, и могут обладать способностью расщеплять полимер β-D-глюкана.
Получение полинуклеотидов
Полинуклеотиды, которые не обладают 100% идентичностью с SEQ ID NO: 1, 3 или 5, но попадают в рамки изобретения, можно получить рядом способов. Такие варианты описанной здесь β-глюканазной последовательности можно получить, например, посредством зондирования библиотек геномных ДНК, полученных для ряда организмов, например, организмов, которые обсуждались как источники полипептидов согласно изобретению. Кроме того, можно получить другие гомологи β-глюканазы из грибов, растений или прокариот, и такие гомологи и их фрагменты, как правило, будут обладать способностью гибридизоваться с SEQ ID NO: 1, 3 или 5. Такие последовательности можно получить зондированием библиотек кДНК или библиотек геномной ДНК других видов, и зондированием таких библиотек зондами, содержащими все или части SEQ ID NO: 1, 3 или 5 в условиях от средней до высокой жесткости (как описано ранее). Зонды на основе нуклеиновых кислот, содержащие все или части SEQ ID NO: 1, 3 или 5, можно использовать для исследования библиотек кДНК других видов, таких как виды, описанные в качестве источников полипептидов согласно изобретению.
Видовые гомологи также можно получить, используя вырожденную ПЦР, в которой будут использоваться праймеры, сконструированные к последовательностям-мишеням в пределах вариантов и гомологов, кодирующих консервативные аминокислотные последовательности. Праймеры могут содержать одно или несколько вырожденных положений, и их будут использовать в условиях более низкой жесткости, чем условия, используемые для клонирования последовательностей с праймерами с единственной последовательностью против известных последовательностей.
Альтернативно, такие полинуклеотиды можно получить сайт-направленным мутагенезом β-глюканазных последовательностей или их вариантов. Это может быть полезным, например, в том случае, когда требуются молчащие замены кодонов в последовательностях, чтобы оптимизировать предпочтительность кодонов для конкретной клетки-хозяина, в которой полинуклеотидные последовательности экспрессируются. Другие изменения последовательностей могут требоваться для того, чтобы ввести сайты узнавания ферментами рестрикции, или для того, чтобы изменить свойство или функцию полипептидов, кодируемых полинуклеотидами.
Изобретение включает в себя двухцепочечные полинуклеотиды, содержащие полинуклеотид согласно изобретению и комплементарную ему последовательность.
Данное изобретение также относится к полинуклеотидам, кодирующим полипептиды согласно изобретению, описанные ниже. Поскольку такие полинуклеотиды будут использоваться в качестве последовательностей для рекомбинантного получения полипептидов согласно изобретению, нет необходимости в том, чтобы они обладали способностью гибридизоваться с последовательностью SEQ ID NO: 1, 3 или 5, хотя, как правило, это будет желательным. В остальном такие полинуклеотиды при необходимости можно метить, использовать и получать, как описано выше.
В. Полипептиды
Данное изобретение относится к β-глюканазам (или целлюлазам) и их вариантам (например, очищенным (в значительной степени) и/или выделенным). Полипептиды согласно изобретению могут по существу состоять из аминокислотной последовательности SEQ ID NO: 2, 4 или 6, или варианта указанной последовательности. Полипептиды также могут кодироваться полинуклеотидом согласно изобретению, который описан выше.
Полинуклеотид согласно изобретению может быть в изолированной или в значительной степени очищенной форме. Следует понимать, что полипептид может быть смешан с носителями или разбавителями, которые не препятствуют достижению планируемой цели и/или функции полипептида, и, тем не менее, считаться по существу изолированным. Как правило, он будет включать в себя полипептид в препарате, в котором более 20%, например, более 30%, 40%, 50%, 80%, 90%, 95% или 99% по массе полипептида в препарате составляет полипептид согласно изобретению. Для очистки и/или синтеза белков согласно изобретению можно применять обычные способы1. Для некоторых композиций (например, для нефармацевтических применений) количество присутствующего полипептида может быть небольшим, например от 0,01 до 10%, как, например, от 0,1 до 5%, или 2%, или даже от 0,2 до 1%.
Предпочтительно полипептид согласно изобретению получают из микроорганизма, который имеет ген, кодирующий фермент с активностью β-глюканазы (или целлюлазы). Более предпочтительно микроорганизм является грибом, или оптимально нитчатым грибом. Предпочтительными организмами являются организмы рода Talaromyces, такие как организмы вида Talaromyces emersonii.
Активность
Полипептид согласно изобретению может обладать одним или несколькими из следующих характеристик, а именно он:
(1) обладает β-глюканазной (или целлюлазной) активностью;
(2) имеет пределы оптимума рН от 3 до 6,5 или 7,0, такие как от 4 до 5,5 или 6,0, оптимально от 4,5 до 5,0;
(3) имеет оптимальную активность при температуре от 30 до 100°С, как, например, от 60 до 95°С, оптимально от 75 или 80 до 90°С. Предпочтительно оптимальная температура составляет, по меньшей мере, 75°С, как, например, по меньшей мере, 85°С.
(4) имеет молекулярную массу (дегликозилированная форма) от 20 до 60 кДа, предпочтительно от 20 до 25, от 35 до 45 или от 23 до 50 кДа, оптимально от 36 до 40 кДа или (гликозилированная форма) от 40 до 45 кДа.
(5) имеет изоэлектрическую точку от 3,0 до 3,6 или 5,0; от 3,8 до 4,5; от 4,5 до 5,0 или от 6,0 до 7,0; и/или
(6) обладает гидролитической активностью по отношению к β-глюкану злаковых или проявляет гидролитическую активность ниже рН 7,0.
Полипептид может обладать активностью ЕС.3.2.1.4 (или активностью эндоглюканазы). В общем, это может быть эндогидролиз 1,4-β-D-глюкозидной связи в целлюлозе. "Активность β-глюканазы" представляет собой способность расщеплять целлюлозу или полимер β-глюкана (например, которые обнаружены в растениях, например овсе и ячмене). Таким образом, активность обеспечивает расщепление между близлежащими концом глюкопиранозы и/или неконцевыми единицами. Предпочтительно расщепление происходит по месту связи [глюкоза(1-4), (1-3) или (1-6) глюкоза]. Полипептид предпочтительно может расщеплять в положении между двумя близлежащими единицами (например, незамещенной глюкозы). Таким образом, он может обладать эндоактивностью (т.е. являться эндоглюканазой). Полимерный субстрат может быть замещенным или незамещенным. Предпочтительно полипептид не будет обладать активностью ксиланазы.
Полипептид также может обладать активностью целлюлазы, то есть он активен по отношению к целлюлозе, или может расщеплять целлюлозу. Так как β-глюкан является компонентом целлюлозы, все глюканазы подпадают под более широкий термин целлюлазы. Поэтому полипептиды согласно данному изобретению являются целлюлазами, так как они относятся к подклассу глюканаз. Другими типами классов активности в пределах целлюлаз, отличными от эндоглюканазы (ЕС 3.2.1.4, как указано выше) являются экзоглюканаза/целлобиогидролаза (ЕС 3.2.1.91), β-глюкозидаза (ЕС 3.2.1.21), эндо-1,6-глюканаза (ЕС 3.2.1.75), экзо-1,3-глюканаза (ЕС 3.2.1.58), маннаназа (ЕС 3.2.1.78) и эндогликоцерамидаза (ЕС 3.2.1.123). Считают, что некоторые полипептиды обладают одной, двумя или большим количеством указанных дополнительных активностей, на основании их структуры и последовательности при сравнении с известными ферментами и семейством гидролаз, к которому они относятся. Таким образом, в описании в случае соответствующего контекста активность глюканазы может означать активность целлюлазы. Список (целлюлазных) активностей полипептидов приведен далее в примере 11.
Предпочтительно полипептид относится к одному из семейств 5, 7 или 45 в соответствии с классификацией гидролаз гликозидов (CAZy). Полипептид может быть нуклеофилом/донором протонов glu/glu или asp/asp.
Варианты и гомологи
Полипептид согласно изобретению может содержать аминокислотную последовательность, указанную в SEQ ID NO: 2, 4 или 6, или в значительной степени гомологичную последовательность, или фрагмент любой последовательности, и может обладать активностью β-глюканазы. В общем, предпочтительна аминокислотная последовательность природного происхождения, показанная в SEQ ID NO: 2, 4 или 6.
В частности, полипептид согласно изобретению может включать в себя:
а. полипептидную последовательность SEQ ID NO: 2, 4 или 6;
b. ее вариант природного происхождения или видовой гомолог; или
с. белок с последовательностью, идентичной, по меньшей мере, на 70, по меньшей мере, на 80, по меньшей мере, на 90, по меньшей мере, на 95, по меньшей мере, на 98 или, по меньшей мере, на 99% с последовательностью по (а) или (b).
Вариант может быть вариантом природного происхождения, например, в клетках грибов, бактерий, дрожжей или растений, и который может функционировать в значительной степени сходным образом с белком SEQ ID NO: 2, 4 или 6, например, он обладает активностью β-глюканазы. Подобным образом, видовой гомолог белка будет эквивалентом белка, который имеет природное происхождение у другого вида и который может функционировать как фермент β-глюканаза. Варианты включают аллельные варианты либо из того же самого штамма, что и полипептид согласно изобретению, либо из другого штамма, но того же рода, или того же самого вида.
Вариант и видовые гомологи можно получить посредством следующих описанных здесь процедур получения полипептида SEQ ID NO: 2, 4 или 6, и проводя такие процедуры на подходящем источнике клеток, например, на клетке бактерий, дрожжей, грибов или растений. Также можно использовать зонд, который охарактеризован выше для того, что исследовать библиотеки, полученные из клеток дрожжей, бактерий, грибов или растений для того, чтобы получить клоны, содержащие варианты или видовые гомологи. Клоны можно обрабатывать обычными способами, чтобы создать полипептид согласно изобретению, который в таком случае можно получить по существу известными рекомбинантными или синтетическими способами.
Полипептид согласно изобретению предпочтительно обладает, по меньшей мере, 70% идентичностью последовательности с белком SEQ ID NO: 2, 4 или 6, более предпочтительно, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 97% или, по меньшей мере, 99% идентичностью последовательности при этом, например, на протяжении района, по меньшей мере, из 60, по меньшей мере, из 100, 150, 200 или 300 следующих друг за другом аминокислот, или на протяжении всей длины SEQ ID NO: 2, 4 или 6. В случае SEQ ID NO: 2 идентичность последовательностей предпочтительно составляет, по меньшей мере, 70%, в случае SEQ ID NO: 4, по меньшей мере, 60% и в случае SEQ ID NO: 6, по меньшей мере, 65%.
Таким образом, последовательность полипептида SEQ ID NO: 2, 4 или 6 и вариантов и видовых гомологов можно модифицировать, чтобы получить полипептиды согласно изобретению. Можно осуществить замены аминокислот, например, от или на уровне 1, 2, или 3 до 10, 20, 30, 50 или 100 замен. Также можно сделать такое же количество делеций и инсерций. Указанные изменения можно произвести вне районов, необходимых для функционирования полипептида, и поэтому в результате можно получить еще активный фермент. Модифицированный полипептид, как правило, сохраняет активность как β-глюканаза.
Полипептиды согласно изобретению включают фрагменты вышеупомянутых полипептидов полной длины и их вариантов, включая фрагменты последовательности, указанной в SEQ ID NO: 2, 4 или 6. Такие фрагменты обычно сохраняют активность β-глюканазы. Фрагменты могут составлять в длину, по меньшей мере, 50, 100, 150, 200 ил 250 аминокислот, или могут быть на это количество аминокислот короче последовательности полной длины (которая указана в SEQ ID NO: 2, 4 или 6).
Полипептиды согласно изобретению при необходимости можно получить синтетическими способами, хотя, обычно их будут получать рекомбинантным способом, как описано ниже. Полипептиды можно модифицировать, например, присоединением остатков гистидина или Т7-метки, чтобы способствовать их идентификации и очистке, или присоединением сигнальной последовательности для активации их секреции из клетки.
Термин "варианты" относится к полипептидам, которые имеют такой же существенный признак или основные функциональные возможности, как и β-глюканаза (или целлюлаза), и включает в себя аллельные варианты. Существенным признаком β-глюканазы является то, что она представляет собой фермент, который проявляет активность ЕС 3.2.1.4, или то, что она расщепляет 1→3, 1→4 и/или 1→6 связи в β-D-глюкане, таком как (β)-глюкан зерновых или овсяной полбы. Предпочтительно вариант полипептида обладает такой же активностью, как β-глюканаза. Полипептид, имеющий такой же существенный признак, как и β-глюканза, можно идентифицировать, используя анализ расщепления целлюлозы или β-глюкана, например как описано далее.
Варианты SEQ ID NO: 2, 4 или 6 также включают последовательности, которые отличаются от SEQ ID NO: 2, 4 или 6, но которые не обязательно получены из белка β-глюканазы природного происхождения. Указанные варианты можно описать, как последовательности, имеющие некоторый % гомологии с SEQ ID NO: 2, 4 или 6, или имеющие ряд замен в пределах этой последовательности. Альтернативно вариант может кодироваться полинуклеотидом, который гибридизуется с SEQ ID NO: 1, 3 или 5.
Варианты можно определить подобно определению вариантов SEQ ID NO: 1, 3 или 5. Таким образом, варианты могут включать в себя вариантные последовательности, полученные из других штаммов Talaromyces. Другие варианты из других штаммов Talaromyces можно идентифицировать при исследовании β-D-глюканазной активности и клонировании и секвенировании, как указано ранее. Варианты могут включать в себя делецию, модификацию или присоединение отдельных аминокислот или групп аминокислот в последовательности белка при условии, что пептид сохраняет основные выполняемые функции β-глюканазы.
Консервативные замены можно сделать, например, в соответствии со следующей таблицей. Аминокислоты в одном и том же блоке во второй колонке и предпочтительно на одной и той же линии в третьей колонке можно взаимно заменять. Предпочтительно замены не влияют на укладку и активность полипептида.
Более короткие полипептидные последовательности входят в рамки изобретения. Например, считается, что пептид длиной, по меньшей мере, 50 аминокислот или до 60, 70, 80, 100, 150 или 200 аминокислот входит в рамки изобретения при условии, что он проявляет основные биологические функции β-глюканазы. В частности, но не только, данный аспект изобретения охватывает ситуацию, когда белок является фрагментом полной последовательности белка и может содержать или представлять собой район, связывающий β-D-глюкан (или маннозу), или район, расщепляющий β-D-глюкан (или целлюлозу).
Модификации
Полипептиды согласно изобретению могут быть химически модифицированными, например модифицированными пост-трансляционно. Например, полипептиды могут быть гликозилированы (один или несколько раз, одним и тем же или разными сахарами) или содержать модифицированные аминокислотные остатки. Их также можно модифицировать присоединением остатков гистидина (чтобы обеспечить их очистку) или присоединением сигнальной последовательности (чтобы обеспечить встраивание в мембрану клеток). Полипептид может иметь одно или несколько удлинений (N)-амино- или (С)-карбоксильных концов, таких как аминоконцевой остаток метионина, небольшой линкерный пептид длиной примерно до 20-25 остатков или (небольшое) удлинение, которое облегчает очистку, такое как полигистидин или Т7-метка, антигенный эпитоп или связывающий домен (например, мальтозу)14 (например, на С-конце). Указанные удлинения можно присоединять посредством линкера или без него.
Полипептид изобретения можно метить регистрируемой меткой. Регистрируемой меткой может быть любая подходящая метка, которая позволяет выявить полипептид. Подходящие метки включают радиоизотопы, например 125I, 35S, ферменты, антибиотики, полинуклеотиды и линкеры, такие как биотин.
Полипептиды могут быть модифицированы таким образом, чтобы они содержали аминокислоты неприродного происхождения или чтобы повысить стабильность полипептида. В том случае, когда белки или пептиды получают синтетическими способами, такие аминокислоты можно вводить во время получения. Белки или пептиды также можно модифицировать после получения синтетическим или рекомбинантным способом.
Полипептиды согласно изобретению также можно получить с использованием D-аминокислот, или они могут содержать D-аминокислоты (одну или несколько).
В данной области известен ряд модификаций боковых цепей, и такие модификации можно осуществить по отношению к боковым цепям белков или пептидов согласно данному изобретению. Такие модификации включают, например, модификации аминокислот восстановительным алкилированием в результате реакции с альдегидом с последующим восстановлением NaBH4, амидированием метилацетимидатом или ацилирование уксусным ангидридом.
Последовательности, представленные в данном изобретении, также можно использовать в качестве исходных материалов для конструирования ферментов "второго поколения". Глюканазы "второго поколения" являются глюканазами, измененными способами мутагенеза (например, сайт-направленного мутагенеза), которые обладают свойствами, отличными от свойств глюканаз дикого типа или рекомбинантных глюканаз, таких как глюканазы, получаемые согласно данному изобретению. Например, оптимум температуры или рН, удельная активность, сродство к субстрату или термостабильность можно изменять для того, чтобы они лучше подходили для применения в определенных способах.
Аминокислоты, необходимые для проявления активности глюканаз согласно изобретению и поэтому предпочтительно подлежащие замене, можно идентифицировать согласно способам, известным в данной области, такими как сайт-направленный мутагенез или мутагенез на основе сканирования аланина10. В последнем способе мутации вводят в каждый остаток молекулы и полученные в результате мутантные молекулы проверяют в отношении биологической активности (например, глюканазной активности), чтобы идентифицировать остатки аминокислот, которые являются критическими для активности молекулы. Сайты взаимодействия фермент-субстрат также можно определить при анализе кристаллической структуры, которую определяют такими способами, как ядерный магнитный резонанс, кристаллография или фото-аффинное мечение11,12,13 или молекулярное моделирование.
Предполагается, что клетки-хозяева дрожжей и грибов обеспечивают такие посттрансляционные модификации (например, протеолитический процессинг, миристилирование, гликозилирование, укорочение и фосфорилирование тирозина, серина или треонина), которые могут быть необходимы для придания оптимальной биологической активности рекомбинантным продуктам экспрессии согласно изобретению.
Полипептиды согласно изобретению могут быть получены в такой форме, в которой они находятся вне их естественной клеточной среды. Таким образом, их можно в значительной степени изолировать или очистить, как обсуждалось выше, или они могут быть в клетке, в которой они не встречаются в природе, например клетке гриба другого вида, животного, дрожжей или бактерий.
С. Аспекты рекомбинации
Изобретение также относится к векторам, содержащим полинуклеотид согласно изобретению, включая клонирующие и экспрессирующие векторы, и способам выращивания, трансформации или трансфекции такими векторами подходящей клетки-хозяина, например, в условиях, при которых происходит экспрессия полипептида согласно изобретению. Также изобретение относится к клеткам-хозяевам, содержащим полинуклеотид или вектор согласно изобретению, в которых полинуклеотид является гетерологичным по отношению к геному клетки-хозяина. Термин "гетерологичный" обычно по отношению к клетке-хозяину означает, что полинуклеотид не является полинуклеотидом природного происхождения в геноме клетки-хозяина, или что полипептид в естественных условиях не продуцируется данной клеткой. Предпочтительно клеткой-хозяином является дрожжевая клетка, например клетка дрожжей рода Kluyveromyces или Saccharomyces, или клетка грибов, например рода Aspergillus.
Полинуклеотиды согласно изобретению могут быть включены в рекомбинантный реплицирующийся вектор, например клонирующий или экспрессирующий вектор. Вектор можно использовать для того, чтобы реплицировать нуклеиновую кислоту в совместимой клетке-хозяине. Таким образом, в следующем варианте изобретение относится к способу получения полинуклеотидов согласно изобретению путем введения полинуклеотида согласно изобретению в реплицируемый вектор, введения вектора в совместимую клетку-хозяина и выращивания клетки-хозяина в условиях, которые вызывают репликацию вектора. Вектор можно извлекать из клетки-хозяина. Подходящие клетки-хозяева описаны далее в связи с экспрессирующими векторами.
Векторы
Полинуклеотид согласно изобретению можно встроить в экспрессирующую кассету. Вектором, в который встраивают экспрессирующую кассету или полинуклеотид согласно изобретению, может быть любой вектор, который легко можно подвергнуть процедурам рекомбинации ДНК, и выбор вектора часто будет зависеть от клетки-хозяина, в которую его необходимо ввести. Таким образом, вектором может быть автономно реплицирующийся вектор, т.е. вектор, который существует в экстрахромосомном виде, репликация которого независима от репликации хромосом, например плазмида. Альтернативно, вектором может быть вектор, который при введении в клетку-хозяина интегрируется в геном клетки-хозяина и реплицируется вместе с хромосомой(ми), в которую он был интегрирован.
Предпочтительно полинуклеотид согласно изобретению в векторе оперативно связан с регуляторной последовательностью, которая способна обеспечивать экспрессию кодирующей последовательности клеткой-хозяином, т.е. вектор является экспрессирующим вектором. Термин "оперативно связан" относится к размещению по соседству, при котором описанные компоненты находятся во взаимосвязи, позволяющей им функционировать соответствующим образом. Регуляторная последовательность, такая как промотор, энхансер или другой сигнал регуляции экспрессии, "оперативно связанный" с кодирующей последовательностью, расположен таким образом, что экспрессия кодирующей последовательности достигается в условиях, сочетающихся с регуляторными последовательностями.
Вектором может быть плазмида, космида, вирус или фаговый вектор, обычно снабженный началом репликации, необязательно промотором для экспрессии полинуклеотида и необязательно энхансером и/или регулятором промотора. Может присутствовать последовательность терминатора, как и последовательность полиаденилирования. Вектор может содержать один или несколько селектируемых маркерных генов, например ген устойчивости к ампициллину (в случае бактериальной плазмиды) или ген устойчивости к неомицину (в случае вектора млекопитающих). Векторы можно использовать in vitro, например, для получения РНК или использовать для того, чтобы трансфицировать или трансформировать клетку-хозяина. Векторы могут содержать два и более полинуклеотидов согласно изобретению, например для сверхэкспрессии.
Последовательность ДНК, кодирующую полипептид, предпочтительно вводят в подходящего хозяина в виде части экспрессирующей кассеты (или конструкции), в которой последовательность ДНК оперативно связана с сигналами экспрессии, которые способны направлять экспрессию последовательности ДНК в клетке-хозяине. Для трансформации подходящего хозяина экспрессирующей конструкцией имеются способы трансформации, которые хорошо известны специалистам3,4. Экспрессирующую конструкцию можно использовать для трансформации хозяина в виде части вектора, несущего селектируемый маркер, или экспрессирующей конструкцией в виде отдельной молекулы можно совместно трансформировать вместе с вектором, несущим селектируемый маркер. Вектор может содержать один или несколько генов селектируемых маркеров.
Предпочтительные селектируемые маркеры15,16 включают, но не ограничены маркерами, которые комплементируют дефект в клетке-хозяине, или придают устойчивость к лекарственному средству. Указанные маркеры включают, например, универсальные маркерные гены, которые можно использовать для трансформации большинства нитчатых грибов и дрожжей, такие как гены или кДНК ацетамидазы (гены или кДНК amdS, niaD, facA из A. nidulans, A. oryzae или A. niger), или гены, придающие устойчивость к антибиотикам, подобную устойчивости к G418, гигромицину, блеомицину, канамицину, флеомицину или беномилу (benA). Альтернативно можно использовать специфичные селектируемые маркеры, такие как ауксотрофные маркеры, которые требуют соответствующих мутантных линий хозяина: например, URA3 (из S. cerevisiae или аналогичные гены из других дрожжей), pyrG или pyrA (из A. nidulans или A. niger), argB (из A. nidulans или A. niger) или trpC. В предпочтительном варианте селектируемый маркер удаляют из трансформированной клетки-хозяина после введения экспрессирующей конструкции для того, чтобы получить трансформированные клетки-хозяева, способные продуцировать полипептид, который не содержит продуктов селектируемых маркерных генов21,22.
К другим маркерам относится АТФ-синтетаза, субъединица 9 (oliC), оротидин-5'-фосфатдекарбоксилаза (pvrA), бактериальный ген устойчивости к G418 (который также можно использовать у дрожжей, но не у грибов), ген устойчивости к ампициллину (E. coli), ген устойчивости к неомицину (Bacillus) и ген uidA E. coli, кодирующий β-глюкуронидазу (GUS). Векторы можно использовать in vitro, например, для получения РНК, или использовать для того, чтобы трансфицировать или трансформировать клетку-хозяина.
В случае большинства нитчатых грибов и дрожжей вектор или экспрессирующую конструкцию предпочтительно интегрируют в геном клетки-хозяина для того, чтобы получить стабильные трансформанты. Однако в случае некоторых дрожжей также имеются подходящие эписомные векторы, в которые может быть включена экспрессирующая конструкция для стабильной экспрессии на высоком уровне, примеры таких векторов включают векторы, полученные из плазмид 2 мкм и pKD1 Saccharomyces и Kluyveromyces, соответственно, или векторы, содержащие последовательность АМА (например, АМА1 из Aspergillus3,20). В том случае, когда экспрессирующие конструкции интегрируют в геном клеток-хозяев, конструкции интегрируют либо в случайный локус генома, либо в предварительно определенный локус-мишень с использованием гомологичной рекомбинации, и в этом случае локус-мишень предпочтительно содержит высокоэкспрессируемые гены. Высокоэкспрессируемым геном является ген, мРНК которого может составлять, по меньшей мере, 0,01% (мас./мас.) суммарной клеточной мРНК, например, в условиях индукции, или альтернативно ген, генный продукт которого может составлять, по меньшей мере, 0,2% (мас./мас.) суммарного клеточного белка, или в случае секретируемого генного продукта он может секретироваться на уровне, по меньшей мере, 0,05 г/л. Ряд примеров подходящих высокоэкспрессируемых генов приведен далее.
Вектор или экспрессирующая конструкция для данной клетки-хозяина может содержать следующие элементы, оперативно связанные друг с другом в последовательном порядке от 5'- до 3'-конца относительно кодирующей нити последовательности, кодирующей полипептид согласно изобретению:
(1) последовательность промотора, способного управлять транскрипцией последовательности ДНК, кодирующей полипептид, в данной клетке-хозяине;
(2) необязательно сигнальная последовательность, способная управлять секрецией полипептида из данной клетки-хозяина в культуральную среду;
(3) последовательность ДНК, кодирующая зрелую и предпочтительно активную форму полипептида; и предпочтительно также
(4) район терминации транскрипции (терминатор), способный терминировать транскрипцию ниже последовательности ДНК, кодирующей полипептид.
Ниже последовательности ДНК, кодирующей полипептид, может располагаться 3'-нетранслируемый район, содержащий один или несколько сайтов терминации транскрипции (например, терминатор). Происхождение терминатора менее значимо. Терминатор, например, может быть собственным для последовательности ДНК, кодирующей полипептид. Однако предпочтительно используют терминатор дрожжей в дрожжевых клетках-хозяевах и терминатор нитчатого гриба используют в клетках-хозяевах нитчатого гриба. Более предпочтительно терминатор является эндогенным по отношению к клетке-хозяину (в которой последовательность ДНК, кодирующую полипептид, необходимо экспрессировать).
Повышенной экспрессии полинуклеотида, кодирующего полипептид согласно изобретению, можно также добиться путем селекции гетерологичных регуляторных районов, например районов промотора, лидера секреции и/или терминатора, которые могут служить для усиления экспрессии, и при необходимости повышения уровней секреции представляющего интерес белка из экспрессирующего хозяина и/или для того, чтобы предусмотреть индуцируемый контроль экспрессии полипептида согласно изобретению.
Помимо промотора, естественного для гена, кодирующего полипептид согласно изобретению, можно использовать другие промоторы для того, чтобы управлять экспрессией полипептида согласно изобретению. Промотор можно выбирать по его эффективности в управлении экспрессией полипептида согласно изобретению в требуемом экспрессирующем хозяине.
Промоторы/энхансеры и другие сигналы регуляции экспрессии можно выбрать так, чтобы они были совместимы с клеткой-хозяином, для которой предназначен экспрессирующий вектор. Например, можно использовать прокариотические промоторы, в частности промоторы, подходящие для применения в штаммах E. coli. В том случае, когда экспрессию осуществляют в клетках млекопитающих, можно использовать промоторы млекопитающих. Также можно использовать тканеспецифичные промоторы, например промоторы, специфичные для гепатоцитов. Также можно использовать вирусные промоторы, например длинный концевой повтор вируса лейкоза мышей Молони (MMLV LTR), промотор LTR вируса саркомы Рауса (RSV), промотор SV40 (например, большого Т-антигена), промотор IE цитомегаловируса человека (CMV), промоторы вируса простого герпеса или промоторы аденовируса, промоторы HSV, такие как промоторы IE HSV или промоторы HPV, в частности, регуляторный район выше (URR) HPV. Дрожжевые промоторы включают промоторы GAL4 и ADH S. cerevisiae, промотор nmt1 и adh S. pombe. Промоторы млекопитающих включают промотор металлотионеина, промотор, который можно индуцировать в ответ на тяжелые металлы, такие как кадмий, и промоторы β-актина. Особенно предпочтительны тканеспецифичные промоторы, в частности промоторы, специфичные для эндотелиальных и нейронных клеток (например, промоторы DDAHI и DDAHII).
Можно использовать множество промоторов15,16, которые способны управлять транскрипцией в клетках-хозяевах согласно изобретению. Предпочтительно последовательность промотора получают из высокоэкспрессируемого гена, который охарактеризован ранее. Примеры предпочтительных высокоэкспрессируемых генов, из которых предпочтительно получают промоторы, и/или которые находятся в предпочтительном предварительно определенном локусе-мишени для интеграции экспрессирующих конструкций, включают, но не ограничены генами, кодирующими гликолитические ферменты, такие как триозофосфатизомеразы (TPI), глицеральдегидфосфатдегидрогеназы (GAPDH), фосфоглицераткиназы (PGK), пируваткиназы (PYK), алкогольдегидрогеназы (ADH), а также генами, кодирующими амилазы, глюкоамилазы, протеазы, ксиланазы, целлобиогидролазы, β-галактозидазы, алкоголь(метанол)оксидазы, факторы элонгации и рибосомные белки. Конкретные примеры подходящих высокоэкспрессируемых генов включают, например, ген LAC4 из Kluyveromyces sp., гены метанолоксидазы (AOX и MOX) из Hansenula и Pichia, соответственно, гены глюкоамилазы (glaA) из A. niger и A. awamori, ген ТАКА-амилазы A. oryzae, ген gpdA A. nidulans и гены целлобиогидролазы T. reesei.
Примерами сильных конститутивных и/или индуцибельных промоторов, которые предпочтительны для применения в экспрессирующих хозяевах - грибах15,16, являются промоторы, которые получают из генов грибов ксиланазы (xlnA), фитазы, АТФ-синтетазы, субъединицы 9 (oliC), триозофосфатизомеразы (tpi), алкогольдегидрогеназы (AdhA), α-амилазы (amy), амилоглюкозидазы (AG - из гена glaA), ацетамидазы (amdS) и промоторы глицеральдегид-3-фосфатдегидрогеназы (gpd).
Примерами сильных дрожжевых промоторов являются промоторы, получаемые из генов алкогольдегидрогеназы, лактазы, 3-фосфоглицераткиназы и триозофосфатизомеразы.
Примерами сильных бактериальных промоторов являются промоторы α-амилазы и SPo2, а также промоторы генов внеклеточных протеаз.
Промоторы, подходящие для растительных клеток, включают промоторы нопалинсинтазы (nos), октопинсинтазы (ocs), маннопинсинтазы (mas), малой субъединицы риболозы (rubisco ssu), гистона, актина риса, фазеолина, вируса мозаики цветной капусты (CMV) 35S и 19S и цирковируса. Все указанные промоторы легко доступны в данной области.
Вектор, кроме того, может включать в себя последовательности, фланкирующие полинуклеотид, дающий начало РНК, которые содержат последовательности, гомологичные эукариотическим геномным последовательностям, предпочтительно геномным последовательностям млекопитающих или вирусным геномным последовательностям. Это позволит вводить полинуклеотиды согласно изобретению в геном эукариотических клеток или вирусов посредством гомологичной рекомбинации. В частности, плазмидный вектор, содержащий экспрессирующую кассету, фланкированную вирусными последовательностями, можно использовать для того, чтобы получить вирусный вектор, подходящий для доставки полинуклеотидов согласно изобретению в клетку млекопитающего. Другие примеры подходящих вирусных векторов включают векторы вируса простого герпеса18,19 и ретровирусы, включая лентивирусы, аденовирусы, аденоассоциированные вирусы и вирусы HPV (такие как HPV-16 или HPV-18). Способы переноса генов с использованием указанных вирусов известны специалистам в данной области. Ретровирусные векторы, например, можно использовать для того, чтобы стабильно интегрировать полинуклеотид, дающий начало антисмысловой РНК, в геном хозяина. Дефектные по репликации аденовирусные векторы напротив сохраняются в виде эписом и поэтому обеспечивают временную экспрессию.
Вектор может содержать полинуклеотид согласно изобретению, ориентированный в антисмысловом направлении, чтобы обеспечить продукцию антисмысловой РНК. Это можно использовать для снижения при необходимости уровней экспрессии полипептида.
Клетки-хозяева и экспрессия
В следующем аспекте изобретение относится к способу получения полипептида согласно изобретению, который включает в себя культивирование клетки-хозяина (например, трансформированной или трансфицированной экспрессирующим вектором, как описано выше) в условиях, обеспечивающих экспрессию (вектором) кодирующей последовательности, кодирующей полипептид, и необязательно извлечение экспрессированного полипептида. Полинуклеотиды согласно изобретению можно включить в рекомбинантный реплицируемый вектор, например экспрессирующий вектор. Вектор можно использовать для того, чтобы реплицировать нуклеиновую кислоту в совместимой клетке-хозяине. Таким образом, в следующем варианте изобретение относится к способу подготовки полинуклеотида согласно изобретению путем введения полинуклеотида согласно изобретению в реплицируемый вектор, введения вектора в совместимую клетку-хозяина и выращивания клетки-хозяина в условиях, в которых осуществляется репликация вектора. Вектор можно извлечь из клетки-хозяина. Подходящие клетки-хозяева включают бактерии, такие как E. coli, дрожжи, линии клеток млекопитающих и другие линии эукариотических клеток, например клетки насекомых, такие как клетки Sf9, и клетки грибов (например, нитчатых).
Предпочтительно полипептид получают в виде секретируемого белка, и в этом случае последовательность ДНК, кодирующая зрелую форму полипептида, в экспрессирующей конструкции оперативно связана с последовательностью ДНК, кодирующей сигнальную последовательность. Предпочтительно сигнальная последовательность является собственной (гомологичной) по отношению к последовательности ДНК, кодирующей полипептид. Альтернативно сигнальная последовательность является чужеродной (гетерологичной) по отношению к последовательности ДНК, кодирующей полипептид, и в этом случае сигнальная последовательность предпочтительно является эндогенной по отношению к клетке хозяина, в которой экспрессируется последовательность ДНК. Примерами подходящих сигнальных последовательностей для дрожжевых клеток-хозяев являются сигнальные последовательности, полученные из генов α-фактора дрожжей. Подобным образом подходящей сигнальной последовательностью для клеток-хозяев нитчатых грибов является, например, сигнальная последовательность, полученная из гена амилоглюкозидазы (AG) нитчатого гриба, например гена glaA A. niger. Указанную последовательность можно использовать в комбинации с собственным промотором амилоглюкозидазы (также называемой (глюко)амилазой), а также в комбинации с другими промоторами. Также можно использовать гибридные сигнальные последовательности в ситуациях согласно данному изобретению.
Предпочтительными гетерологичными лидерными последовательностями для секреции являются последовательности, происходящие из гена амилоглюкозидазы (AG) грибов (glaA - как версия из 18, так и версия из 24 аминокислот, например, из Aspergillus), гена α-фактора (дрожжи, например Saccharomyces и Kluyveromyces) или гена α-амилазы (Bacillus).
Векторами можно трансформировать или трансфицировать подходящую клетку-хозяина, как описано выше, чтобы обеспечить экспрессию полипептида согласно изобретению. Указанный способ может включать в себя культивирование клетки-хозяина, трансформированной экспрессирующим вектором, который описан выше, в условиях, обеспечивающих экспрессию вектором кодирующей последовательности, кодирующей полипептид.
Таким образом, следующий аспект изобретения относится к клеткам-хозяевам, трансформированным или трансфицированным полинуклеотидом или вектором согласно изобретению, или содержащим полинуклеотид или вектор согласно изобретению. Предпочтительно полинуклеотид включен в вектор для репликации и экспрессии полинуклеотида. Клетки будут выбраны так, чтобы они были совместимыми с указанным вектором, и они могут быть, например, прокариотическими (например, бактериальными) клетками, клетками грибов, дрожжевыми или растительными клетками.
Также можно выбрать гетерологичного хозяина, в котором полипептид согласно изобретению продуцируется в форме, которая по существу не содержит других разрушающих целлюлозу ферментов. Этого можно достичь в результате подбора хозяина, который обычно не продуцирует такие ферменты, такого как Kluyveromyces lactis.
Изобретение охватывает способы получения полипептида согласно изобретению посредством рекомбинантной экспрессии последовательности ДНК, кодирующей полипептид. Для этой цели последовательность ДНК согласно изобретению можно использовать для амплификации генов и/или обмена сигналов экспрессии, таких как промоторы, последовательности сигналов секреции, для того, чтобы обеспечить экономичную продукцию полипептида в подходящей гомологичной или гетерологичной клетке-хозяине. Гомологичная клетка-хозяин имеет здесь значение клетки-хозяина, которая относится к тому же самому виду или которая является вариантом в пределах того же самого вида, что и вид, из которого получают последовательность ДНК.
Подходящими клетками-хозяевами предпочтительно являются прокариотические микроорганизмы, такие как бактерии, или более предпочтительно эукариотические организмы, например, грибы, такие как дрожжи или нитчатые грибы, или растительные клетки. Как правило, дрожжевые клетки предпочтительнее, чем клетки грибов, поскольку ими легче манипулировать. Однако некоторые белки либо недостаточно секретируются из дрожжей, либо в некоторых случаях не процессируются должным образом (например, гипергликозилирование у дрожжей). В указанных случаях следует выбирать гриб в качестве организма-хозяина.
Клетка-хозяин может сверхэкспрессировать полипептид, и способы конструирования для получения сверхэкспрессии хорошо известны3. Таким образом, хозяин может иметь две или больше копий кодирующего полинуклеотида (и таким образом, вектор может соответственно иметь две или больше копий).
Бактерии из рода Bacillus очень подходят в качестве гетерологичных хозяев благодаря их способности секретировать белки в культуральную среду. Другими бактериями, подходящими в качестве хозяев являются бактерии из рода Streptomyces и Pseudomonas. Предпочтительной дрожжевой клеткой-хозяином для экспрессии последовательности ДНК, кодирующей полипептид, является клетка рода Saccharomyces, Kluyveromyces, Hansenula, Pichia, Yarrowia и Schizosaccharomyces. Более предпочтительно дрожжевая клетка-хозяин выбрана из группы, состоящей из видов Saccharomyces cerevisiae, Kluyveromyces lactis (также известного как Kluyveromyces marxianus вариант lactis), Hansenula polymorpha, Pichia pastoris, Yarrowia lipolytica и Schizosaccharomyces pombe.
Однако наиболее предпочтительными хозяевами являются клетки грибов (например, нитчатых). Предпочтительные клетки-хозяева нитчатых грибов выбраны из группы, состоящей из родов Aspergillus, Trichoderma, Fusarium, Disporotrichum, Penicillium, Acremonium, Neurospora, Thermoascus, Myceliophtora, Sporotrichum, Thielavia и Talaromyces. Наиболее предпочтительной клеткой-хозяином нитчатых грибов является клетка вида Aspergillus oryzae, Aspergillus sojae, Aspergillus nidulans или видов из группы Aspergillus niger (которые охарактеризованы Raper and Fennell, The Genus Aspergillus, The Williams and Wilkins Company, Baltimore, pp 293-344, 1965). Указанные виды включают, но не ограничены этим Aspergillus niger, Aspergillus awamori, Aspergillus tubigensis, Aspergillus aculeatus, Aspergillus foetidus, Aspergillus nidulans, Aspergillus japonicus, Aspergillus oryzae и Aspergillus ficuum, и, кроме того, группа состоит из видов Trichoderma reesei, Fusarium graminearum, Penicillium chrysogenum, Acremonium alabamense, Neurospora crassa, Myceliophtora thermophilum, Sporotrichum cellulophilum, Disporotrichum dimorphosporum и Thielavia terrestris.
Примерами предпочтительных экспрессирующих хозяев в рамках данного изобретения являются грибы, такие как грибы вида Aspergillus (описанные в EP-A-184438 и EP-A-284603) и вида Trichoderma; бактерии, такие как бактерии вида Bacillus (описанные в EP-A-134048 и EP-A-253455), например, Bacillus subtilis, Bacillus licheniformis, Bacillus amyloliquefaciens, вида Pseudomonas; и дрожжи, такие как дрожжи вида Kluyveromyces (описанные в EP-A-096430, например, Kluyveromyces lactis в EP-A-301670) и вида Saccharomyces, например Saccharomyces cerevisiae.
Клетки-хозяева согласно изобретению включают клетки растений, и поэтому изобретение распространяется на трансгенные организмы, такие как растения и их части, которые содержат одну или несколько клеток согласно изобретению. Клетки могут гетерологично экспрессировать полипептид согласно изобретению или могут гетерологично содержать один или несколько полинуклеотидов согласно изобретению. Следовательно, трансгенное (или генетически модифицированное) растение может содержать встроенную в его геном (например, стабильно) последовательность, кодирующую один или несколько полипептидов согласно изобретению. Трансформацию растительных клеток можно осуществлять с использованием известных способов, например, используя плазмиду Ti или Ri из Agrobacterium tumefaciens. Таким образом, плазмида (или вектор) может содержать последовательности, необходимые для того, чтобы инфицировать растение, и можно использовать производные плазмид Ti и/или Ri.
Альтернативно можно осуществить непосредственное инфицирование части растения, такой как лист, корень или стебель. В данном способе растению, которое необходимо инфицировать, может быть нанесена рана, например, при разрезании растения бритвой или пунктировании растения иглой или натирании растения абразивным материалом. Затем в рану инокулируют Agrobacterium. Затем растение или часть растения выращивают на подходящей культуральной среде и дают возможность развиваться во взрослое растение. Регенерации трансформированных клеток в генетически модифицированные растения можно достичь, используя известные способы, например отбором трансформированных ростков с использованием антибиотика и субкультивированием ростков на среде, содержащей соответствующие питательные вещества, гормоны растений и тому подобное.17
Культура клеток-хозяев и рекомбинантная продукция
Изобретение также включает в себя клетки, которые были модифицированы для того, чтобы экспрессировать β-глюканазу или ее вариант. Такие клетки включают временные или предпочтительно стабильные линии клеток высших эукариот, такие как клетки млекопитающих или клетки насекомых, клетки низших эукариот, такие как клетки дрожжей и грибов (например, нитчатых) или прокариотические клетки, такие как бактериальные клетки.
В случае белков согласно изобретению также возможна временная экспрессия в линии клеток или на мембране, как, например, в бакуловирусной системе экспрессии. Такие системы, которые адаптированы для экспрессии белков согласно изобретению, также включены в рамки данного изобретения.
Согласно данному изобретению получение полипептида согласно изобретению можно осуществить посредством культивирования экспрессирующих хозяев-микроорганизмов, которые были трансформированы одним или несколькими полинуклеотидами согласно изобретению, в обычной питательной среде для ферментации.
Рекомбинантные клетки-хозяева согласно изобретению можно культивировать, используя способы, известные в данной области. Для каждой комбинации промотора и клетки-хозяина существуют условия культивирования, которые способствуют экспрессии последовательности ДНК, кодирующей полипептид. После достижения необходимой плотности клеток или титра полипептида культивирование прекращают и извлекают полипептид известными способами.
Среда для ферментации может содержать известную культуральную среду, содержащую источник углерода (например, глюкозу, мальтозу, мелассу, целлюлозу, β-глюкан и т.д.), источник азота (например, сульфат аммония, нитрат аммония, хлорид аммония, и т.д.), источники органического азота (например, дрожжевой экстракт, солодовый экстракт, пептон и т.д.) и источники неорганических питательных веществ (например, фосфат, магний, калий, цинк, железо и т.д.). Необязательно можно включать индуктор (например, целлюлозу, β-глюкан, мальтозу или мальтодекстрин).
Подбор соответствующей среды может быть основан на выборе экспрессирующего хозяина и/или основан на требованиях по отношению к регуляции экспрессирующей конструкции. Такие среды известны специалистам в данной области. Среда при необходимости может содержать дополнительные компоненты, дающие преимущества трансформированным экспрессирующим хозяевам над другими потенциально загрязняющими микроорганизмами.
Ферментацию можно выполнять в течение периода, равного 0,5-30 дням. Это может быть периодический, непрерывный процесс или процесс с периодической подпиткой, соответственно при температуре в пределах от 0 до 45°С и, например, при рН от 2 до 10. Предпочтительными условиями ферментации является температура в пределах от 20 до 37°С и/или рН от 3 до 9. Подходящие условия обычно выбирают на основе выбора экспрессирующего хозяина и белка, который необходимо экспрессировать.
После ферментации при необходимости клетки можно извлекать из бульона для ферментации посредством центрифугирования или фильтрования. После остановки ферментации или после извлечения клеток затем можно извлечь полипептид согласно изобретению и при необходимости очистить и выделить обычными способами.
D. Применения полипептидов и способы обработки растения или материалов, содержащих целлюлозу
Полипептиды согласно изобретению, которые обладают β-глюканазной (или целлюлазной) активностью, можно использовать для обработки материала грибов и растений, включая пульпу растений и экстракты растений. Например, полипептиды можно использовать для обработки зерновых культур, овощей, фруктов или их экстрактов. Их также можно использовать в композициях для промывки (жидких или твердых, например, порошках) и/или в композициях детергентов. Полипептид согласно изобретению удобно комбинировать с подходящими носителями (твердыми или жидкими) или разбавителями, включая буферы, чтобы получить композицию/ферментный препарат. Полипептид можно соединять или смешивать с носителем, например иммобилизовать на твердом носителе. Таким образом, в следующем аспекте данное изобретение относится к композиции, содержащей полипептид согласно изобретению. Композиция может быть в форме, подходящей для упаковки, транспортировки и/или хранения, предпочтительно в форме, в которой сохраняется глюканазная активность. Композиции включают пасту, жидкость, эмульсию, порошок, хлопья, гранулы, таблетки или другую прессованную форму.
Композиция, кроме того, может содержать дополнительные ингредиенты, такие как один или несколько ферментов, например пектиназы, включая (например, эндо)-арабинаназу и рамногалатуроназу, другие целлюлазы, ксиланазы, галактаназы, маннаназы и/или ксилоглюканазы. Полипептид обычно готовят в виде стабильной жидкой или сухой формы. Обычно продукт готовят в виде композиции, которая необязательно содержит, например, стабилизирующий буфер и/или консервант. Композиции также могут содержать другие ферменты, способные расщеплять растительный материал или целлюлозу, например, другие целлюлазы, например (β-D-)глюканазы. В случае некоторых применений предпочтительной может быть иммобилизация фермента на твердом матриксе или включение на частицы или в частицы твердого носителя. Композиция также может содержать множество других ферментов, расщепляющих растительный материал, например (другие) целлюлазы и другие пектиназы.
Следовательно, полипептиды и композиции согласно изобретению можно использовать в способе обработки растительного материала, для того чтобы разрушить или модифицировать целлюлозные компоненты (например, β-D-глюкан) клеточной стенки в материале растений или грибов. Следовательно, в следующем аспекте изобретение относится к способу разрушения или модифицирования растительной клетки или целлюлозы, при этом указанный способ включает в себя осуществление контакта клетки растения, гриба или целлюлозы с полипептидом или композицией согласно изобретению.
Изобретение также относится к способу обработки растительного материала, и указанный способ включает в себя осуществление контакта растительного материала с полипептидом или композицией согласно изобретению, чтобы разрушить или модифицировать целлюлозу в растительном материале. Предпочтительно растительным материалом является пульпа растений или экстракт растений.
В частности, разрушение предпочтительно включает в себя расщепление субъединиц β-глюкана целлюлозного компонента клеточной стенки растений. Растительным материалом предпочтительно является злаковая культура, овощ, фрукт или пульпа или экстракт овощей или фруктов. Данное изобретение, кроме того, относится к обработанному растительному материалу, получаемому посредством осуществления контакта растительного материала с полипептидом или композицией согласно изобретению.
Данное изобретение также относится к способу уменьшения вязкости растительного экстракта, и этот способ включает в себя осуществление контакта растительного экстракта с полипептидом или композицией согласно изобретению в количестве, эффективном при разрушении целлюлозы (или β-D-глюкана), присутствующей в растительном экстракте.
Растительные и содержащие целлюлозу материалы включают пульпу растений, части растений и растительные экстракты. В контексте данного изобретения экстракт из растительного материала представляет собой любое вещество, которое можно получить из растительного материала посредством экстракции (механической и/или химической), обработки или другими способами разделения. Экстракт может представлять собой сок, нектар, материал основы или полученные из них концентраты. Полипептид можно использовать при разжижении (например, полном) и/или вымачивании (улучшенном), например, при получении (фруктовых) соков. Растительный материал может содержать или его можно получать из овощей, например моркови, сельдерея, лука, бобов или бобовых растений (соя, соевые бобы, горох), или фруктов, например плода типа яблока или плода с семенами (яблоки, груши, айва и т.д.), винограда, томатов, цитрусовых (апельсин, лимон, лайм, мандарин), дынь, слив, вишни, черной смородины, красной смородины, малины, клубники, клюквы, ананаса и других тропических фруктов, деревьев или их частей (например, пыльцы сосновых деревьев) или злаковых (овса, ячменя, пшеницы, кукурузы, риса).
Таким образом, полипептиды согласно изобретению можно использовать для обработки растительного материала, включая мягкие ткани растений и растительные экстракты. Полипептиды также можно использовать для обработки жидких или твердых пищевых продуктов или съедобных ингредиентов пищевых продуктов.
Обычно полипептиды согласно изобретению используют в виде композиции/ферментного препарата, который описан выше. Композиции, как правило, будут добавляться к мягким тканям растений, получаемым, например, механической обработкой, такой как измельчение или размол растительного материала. Инкубацию композиции с растением обычно будут проводить в течение периода времени от 10 минут до 5 часов, как, например, от 30 минут до 2 часов, предпочтительно около 1 часа. Температура обработки предпочтительно составляет 10-55°С, например от 15 до 25°С, оптимально около 20°С, и можно использовать 10-300 г, предпочтительно 30-70 г, оптимально около 50 г фермента на тонну материала, который необходимо обработать. Все используемые ферменты или их композиции можно добавлять к пульпе растения последовательно или одновременно. В зависимости от композиции препарата фермента растительный материал можно сначала подвергнуть вымачиванию (например, до пюре) или разжижению. Используя полипептиды согласно изобретению, можно улучшить параметры обработки, такие как выход при экстракции, вязкость экстракта и/или качество экстракта.
Альтернативно или дополнительно к сказанному выше полипептид согласно изобретению можно добавлять в сырой сок, полученный из отжатой или разжиженной пульпе растения. Обработка сырого сока будет проводиться способом, подобным обработке пульпы растения в отношении дозы, температуры и времени выдерживания. Опять таки, можно включать другие ферменты, такие как ферменты, обсуждаемые ранее. Типичные условия инкубации описаны в предыдущем параграфе. После того как сырой сок проинкубировали с полипептидами согласно изобретению, сок центрифугируют или (ультра)фильтруют, чтобы получить конечный продукт.
Композицию, содержащую полипептид согласно изобретению, также можно использовать в ходе приготовления фруктовых или овощных пюре.
Полипептид согласно изобретению также можно использовать в пивоварении, виноделии, производстве спирта или при выпечке. Поэтому полипептид можно использовать при приготовлении алкогольных напитков, таких как вино и пиво, например, чтобы улучшить фильтруемость или прозрачность вина. При выпечке полипептид может улучшить структуру теста, модифицировать его жесткость или эластичность, увеличить объем хлеба и/или структуру хлебного мякиша.
Полипептид согласно изобретению можно использовать в пивоварении. В пивоварении проблемы при фильтрации могут повышать стоимость сусла, в которое входит сверхмодифицированый солод, вследствие наличия β-глюканов, высвобождаемых при высоких температурах. Уменьшение скорости фильтрации сусла может быть одной из главных встречающихся проблем. Дополнительными проблемами являются стабильность коллоида и образование мутности в готовом пиве. Это может быть вызвано такими же углеводными комплексами, особенно при производстве пива с высокой плотностью. Полипептиды согласно изобретению могут обладать способностью улучшать фильтруемость сусла, например, после затирания сусла. Таким образом, меньше раствора сусла может оставаться в слое пивной дробины и будет повышен выход экстракта. Результатом более эффективного фильтрования может быть повышенная производительность пивоваренного завода. Таким образом, в общем полипептиды согласно изобретению можно использовать для повышения фильтруемости или скорости фильтрования пива (например, готового).
Полипептиды согласно изобретению также могут предотвращать или, по меньшей мере, уменьшать образование мутности при производстве пива. β-глюканы, обнаруженные в стенках эндосперма злаковых, могут оставаться в готовом пиве. Это может служить причиной мутности и отсутствия прозрачности. Полипептиды согласно изобретению могут предотвращать или, по меньшей мере, уменьшать накопление частиц в результате гидролиза β-глюкана. Следовательно, полипептиды согласно изобретению можно использовать для увеличения стабильности коллоида пива (например, готового).
Полипептиды благодаря их глюканазной активности находят применение в ряде областей промышленности. Применение может не только включать в себя получение спирта, но также биологическое образование метана, производство хлеба и пекарное производство, гигиену зубов (например, композиции для зубов и ротовой полости), обработку тканей, одежды или производство кожи, производство бумаги, фармацевтических препаратов, чая, производство или обработку текстильных изделий, использование в детергентных или моющих композициях, и переработку отходов. Поэтому одним из аспектов изобретения является продукты питания или пищевые продукты, содержащие полипептид, такие как алкогольные напитки, хлеб, тесто или чай. Для любого из указанных применений полипептид может быть приготовлен в составе подходящих композиций. Полипептид может присутствовать в водной композиции (например, горячей воде) предпочтительно с одним или несколькими фунгицидами для обработки растительного материала (например, луковиц), особенно для борьбы с паразитическими насекомыми, клещами и нематодами.
Так как полипептиды согласно изобретению могут разрушать глюкан, их можно добавлять к продуктам питания или пищевым продуктам (например, потребляемых человеком). Известно, что растворимый β-D-глюкан связан с понижением холестерина и триглицеридов в сыворотке, и поэтому полипептиды согласно изобретению можно использовать для снижения уровней холестерина и триглицеридов в сыворотке у человека или животных, например у субъектов при гиперлипидемии. Следовательно изобретение включает в себя фармацевтические и ветеринарные композиции, которые содержат полипептид согласно изобретению и фармацевтически или ветеринарно приемлемый носитель. Таким образом, полипептиды можно использовать в производстве лекарственного средства для предотвращения или лечения гиперлипидемии, или высоких уровней холестерина и триглицеридов, или возникающих вследствие этого заболеваний.
Полипептиды согласно изобретению также могут проявлять противогрибковую активность. Они могут обладать способностью разрушать клеточные стенки грибов, и, следовательно, их можно применять для лизиса клеточной стенки грибов для того, чтобы вскрыть клетки. При этом можно высвободить внутриклеточные белки. Таким образом, полипептиды можно использовать для получения экстрактов дрожжей и/или грибов.
Е. Корма для животных
Изобретение, кроме того, относится к пищевым продуктам или кормовой композиции, содержащей один или несколько полипептидов согласно изобретению. Полипептид может присутствовать в корме в концентрации, отличающейся от его естественной концентрации. Предпочтительные количества составляют от 0,1 до 100, как, например, от 0,5 до 50, предпочтительно от 1 до 10 мг на кг корма.
Изобретение также относится к способу получения кормовой композиции для животных, указанный способ включает в себя добавление полипептида согласно изобретению к одному или нескольким съедобным кормовым веществам или ингредиентам. Полипептиды можно добавлять к кормовым композициям для животных отдельно от пищевых веществ или ингредиентов, отдельно или в комбинации с другими кормовыми добавками. Полипептид может быть составной частью одного из кормовых веществ или ингредиентов.
Полипептиды согласно изобретению также можно добавлять к животным кормам, богатым целлюлозой, чтобы усилить разрушение клеточной стенки растений, что приводит к повышенной утилизации растительных питательных веществ животным. Полипептиды согласно изобретению можно добавлять в корм или силос в том случае, если предпочтительны предварительно замоченные или увлажненные корма. Преимущественно полипептиды согласно изобретению могут продолжать разрушать целлюлозу в корме in vivo. Полипептиды согласно изобретению, полученные из грибов, в частности, как правило, имеют более низкие оптимумы рН и способны высвобождать важные питательные вещества в таких кислых средах, как среда в желудке животных. Таким образом, целью изобретения также являются корма (например, для животных) или пищевые продукты, содержащие один или несколько полипептидов изобретения.
Полипептиды согласно изобретению также можно использовать при производстве заменителей молока из соевых бобов. Указанные заменители молока могут потребляться как человеком, так и животными. Обычно проблемой в ходе получения указанных заменителей молока является высокая вязкость кашицы соевых бобов, приводящая к необходимости в нежелательном разведении кашицы до концентрации сухих твердых веществ от 10 до 15%. Препарат фермента, содержащий полипептид согласно изобретению, можно добавлять в кашицу или во время обработки кашицы, делая возможным обработку при высокой концентрации сухих твердых веществ (обычно от 40 до 50%). Фермент также можно использовать для получения приятного на вкус продукта(тов), например, из соевых бобов.
Композиция, кроме того, может содержать (особенно в том случае, когда ее готовят для применения в корме для животных) один или несколько ионофоров, окисляющих агентов, поверхностно-активных веществ, защищенных от переваривания в желудке аминокислот, усилителей ферментов или ферментов, которые могут продуцироваться естественным образом в желудочно-кишечном тракте животных, которых предстоит кормить.
В случае добавления к кормам (включая силос) для жвачных животных или животных с однокамерным желудком (например, домашняя птица или свинья) корма могут содержать зерновые, такие как ячмень, пшеница, кукуруза, рожь или овес, или субпродукты из зерновых, такие как пшеничные отруби или кукурузные отруби, или другие растительные материалы, такие как соевые бобы и другие плоды бобовых. Фермент(ты) могут в значительной степени усиливать разрушение клеточных стенок растений, что приводит к лучшей утилизации животным растительных питательных веществ. В результате можно увеличить скорость роста и/или переработку корма. Полипептиды согласно изобретению можно добавлять к корму (непосредственно или в виде добавки или ингредиента), или вместо этого можно добавлять обработанную целлюлозу (например, глюкан).
Особенно предпочтительным способом (экзогенного) добавления β-глюканазы является добавление полипептида согласно изобретению в виде трансгенного растительного материала и/или семян (например, трансгенных). Таким образом, полипептид может быть синтезирован посредством гетерологичной экспрессии гена, например ген, кодирующий требуемый фермент, можно клонировать в экспрессирующем растительном векторе под контролем соответствующих сигналов экспрессии в растениях, например, тканеспецифичного промотора, такого как промотор, специфичный для семян. Затем экспрессирующим вектором, содержащим ген, кодирующий полипептид, можно трансформировать растительные клетки, и трансформированные клетки можно отобрать для регенерации целых растений. Полученные таким образом трансгенные растения можно выращивать и собирать, и части растений, содержащие гетерологичный (по отношению к растению) полипептид, можно включать в одну из композиций либо как таковые, либо после обработки. Общие способы (гетерологичной) экспрессии ферментов в (трансгенных) растениях, включая способы для экспрессии ферментов, специфичной для семян, известны23. Гетерологичный полипептид может находится в семенах трансгенных растений, или он может находиться в других частях растения, таких как корни, стебли, листья, древесина, цветки, кора и/или фрукт. Растение может быть однодольным или двудольным. Подходящие растения включают зерновые культуры, такие как овес, ячмень, пшеницу, кукурузу и рис. Предпочтительно полинуклеотид согласно изобретению стабильно включен в геном растения.
Добавление полипептида в форме материала трансгенного растения, например, в виде трансгенных семян, может требовать обработки растительного материала для того, чтобы сделать фермент доступным или, по меньшей мере, повысить его доступность. Такие способы обработки могут включать в себя различные механические способы (например, размалывание и/или измельчение) или термомеханические способы, такие как экструзия или экспансия.
Таким образом, данное изобретение также относится к способу стимуляции роста и/или переработки корма у животного с однокамерным желудком или нежвачного животного, указанный способ включает в себя кормление животного полипептидом согласно изобретению. Подходящие животные включают сельскохозяйственных животных с однокамерным желудком и/или нежвачных животных, таких как свиньи (или поросята), домашнюю птицу (такую как цыплята, индюшки), телят или телят, выращиваемых на убой, или водных (например, морских) животных (например, рыб).
Анализ ферментов, разрушающих целлюлозу
В рамки данного изобретения также входит применение полипептидов согласно изобретению в способах скрининга для идентификации соединений, которые могут действовать как агонисты или антагонисты, которые могут модулировать β-глюканазу. В общих чертах, такие способы скрининга могут включать в себя контактирование полипептида согласно изобретению с тестируемым соединением и затем измерение активности, или инкубацию полипептида согласно изобретению с тестируемым соединением и затем выявление какого-либо модулирования активности β-глюканазы. Агенты, которые связываются с полипептидами согласно данному изобретению, также можно идентифицировать посредством анализов связывания.
Активность модулятора можно определить, осуществляя контакт клеток, экспрессирующих полипептид согласно изобретению, с исследуемым веществом и наблюдая эффект, опосредованный полипептидами. Клетки, экспрессирующие полипептид, могут быть в условиях in vitro, и предпочтительно анализ проводят in vitro, используя клетки, экспрессирующие рекомбинантный полипептид.
Описанные здесь анализы и субстраты позволили идентифицировать и подтвердить β-глюканазную активность. Однако указанные анализы можно использовать для выявления других разрушающих целлюлозу ферментов, либо обладающих, либо не обладающих β-глюканазной активностью. Субстрат, который можно использовать для данного анализа может содержать β-глюкан.
Другой аспект изобретения относится к анализу для идентификации или выявления полипептида, который обладает способностью разрушать целлюлозу. Активность может быть глюканазной (например, β-глюканазной) или целлюлазной или ксилоглюканазной. Анализ может включать в себя:
(а) подготовку в качестве субстрата для испытываемого соединения (обычно полипептида) субстрата, описанного в предыдущем параграфе; и
(b) осуществление контакта субстрата с испытываемым соединением и выявление того, продуцируются ли какие-либо углеводы.
Количество указанных углеводов можно измерить. При необходимости это количество затем можно сравнить с количеством углеводов, продуцируемых в контрольном эксперименте в отсутствие испытываемого соединения.
Указанные выше анализы можно применять для идентификации модуляторов β-глюканазной активности.
Предпочтительные особенности и характеристики одного аспекта изобретения применимы к другому аспекту с необходимыми изменениями.
Теперь изобретение будет описано со ссылками на следующие примеры, которые предназначены только для иллюстрации, но не для ограничения.
ПРИМЕРЫ
Общие процедуры
Стандартные способы молекулярного клонирования, такие как выделение ДНК, гель-электрофорез, модификации нуклеиновых кислот ферментами рестрикции, Саузерн-анализы, трансформация E. coli, переносы колоний и гибридизация на фильтрах, и т.д. выполняли, используя стандартные технологии.1,2 Синтетические олигодезоксинуклеотиды получали из ISOGEN Bioscience (Maarssen, The Netherlands). Анализ последовательности ДНК выполняли на секвенаторе ДНК Applied Biosystems 373A согласно инструкциям поставщика.
Мечение ДНК и гибридизации проводили согласно системам прямого мечения нуклеиновых кислот и регистрации ECLTM (Amersham LIFE SCIENCE, Little Chalfont, England) или согласно стандартным способам радиоактивного мечения.1
Пример 1: Выделение РНК из T. emersonii и синтез кДНК
Штамм CBS 393.64 T. emersonii подвергали ферментации в условиях, индуцирующих целлюлазу. В нескольких временных точках собирали мицелий и надосадки культур посредством фильтрования с использованием материала для фильтрования Miracloth. Мицелий тщательно промывали деминерализованной водой и сжимали между бумажными полотенцами, чтобы удалить избыточную воду. Мицелий из выбранных временных точек (на основании измерений целлюлазы в надосадках культуры) сразу же замораживали и жидком азоте и измельчали до тонкого порошка, используя ступку и пестик. Полученный в результате порошок переносили в стерильную пробирку объемом 50 мл и взвешивали: на каждые 1-1,2 г размолотого мицелия добавляли 10 мл реагента TRIzol (Gibco/BRL) (максимум 25 мл на пробирку). Порошок мицелия сразу же солюбилизировали энергичным перемешиванием (встряхивание с перемешиванием, 1 мин), после чего следовала инкубация в течение 5 минут при комнатной температуре с периодическим перемешиванием. Добавляли хлороформ в объеме, равном 0,2 (от исходного объема TRIzol) (таким образом, 2 мл на каждые 10 мл исходно используемого реагента TRIzol), встряхивали и оставляли при комнатной температуре в течение 10 мин. Затем смесь центрифугировали при 4°С, 6000 g в течение 30 минут. Верхнюю водную фазу переносили в новую пробирку, и суммарную РНК осаждали добавлением изопропилового спирта объемом 0,5 (от исходного объема TRIzol) (таким образом, 5 мл изопропилового спирта на каждые 10 мл исходно используемого реагента TRIzol). После 10 минут осаждения при комнатной температуре РНК извлекали центрифугированием в течение 30 минут при 6000 g. После удаления надосадка осадок РНК промывали одним объемом 70% этанола. После удаления этанола осадок РНК сушили на воздухе. Высушенный осадок РНК растворяли в 3 мл буфера GTS (100 мМ трис-Cl, рН 7,5, 4 М тиоцианат гуанидина, 0,5% лаурилсаркозинат натрия). 10 мкл раствора РНК использовали для определения качества и концентрации нуклеиновых кислот.
Выполняли Нозерн-анализ3 и выделенную РНК далее очищали.1,3 Для выделения мРНК использовали модифицированный протокол из набора для очистки Pharmacia (катал. No. 27-9258-02)3 (используя самотек вместо центрифугирования). Для синтеза кДНК использовали набор для синтеза кДНК Stratagene в соответствии с инструкциями производителя, за исключением ряда оптимизаций для использования векторов pGBFIN, основные изменения при которых описаны ранее.3
Количество синтезированной кДНК определяли при осаждении ТХУ и затем анализировали электрофорезом в щелочных агарозных гелях.3
Пример 2: Получение библиотеки кДНК из мРНК T. emersonii
кДНК пула, полученного в примере 1, затупляли, лигировали с адаптерами и расщепляли ферментами рестрикции.3
Клонирование кДНК в экспрессирующем векторе pGBFIN-11 (смотри WO-99/32617 по вопросам конструирования вектора) требует наличия сайта EcoRI на 5'-конце и сайта XhoI на 3'-конце кДНК. Поэтому используемые последовательности олигонуклеотида, праймирующего первую нить, и адаптера (Pharmacia) выбирали так, чтобы они удовлетворяли необходимым условиям, установленным для экспрессирующего вектора.
Полученные кДНК разделяли посредством фракционирования по размеру, пропуская через матрикс SEPHAROSE CL-2B, после чего размер полученных отдельных пулов анализировали посредством гель-электрофореза в неденатурирующих условиях.3 Два пула кДНК, полученных при отсечении 0,5 т.п.н. и 1,0 т.п.н. соответственно, отбирали для конструирования библиотеки кДНК в pGBFIN-11. В случае pGBFIN-11 готовили пул полностью дважды расщепленного (EcoRI-XhoI) вектора pGBFIN-11 (фоновое лигирование <1%). Выбранные пулы кДНК лигировали с вектором pGBFIN-11 и трансформировали бактериальные клетки E. coli XL10-Gold, чтобы получить две исходные библиотеки кДНК. Частоты трансформирования обоими пулами составляли >1,0 × 106. Из фракции обеих библиотек кДНК E. coli случайным образом отбирали колонии и выделяли плазмидную ДНК. Анализ указанной плазмидной ДНК показал, что обе библиотеки кДНК имеют процентное содержание вставок от 90 до 95%.
Кроме того, колонии из фракции библиотеки переносили на фильтры, и затем полученные фильтры гибридизовали с геном gpdA T. emersonii, кодирующим ген глицеральдегид-3-фосфатдегидрогеназы. Затем выделяли плазмидную ДНК, и посредством рестрикционного анализа было показано, что все плазмиды содержали одиночные вставки в правильной ориентации. Секвенирование 5'-концов кДНК в пределах указанных плазмид, содержащих gpdA T. emersonii, показал, что >85% кДНК имело полную длину.
Пример 3: Трансформация A. niger экспрессирующей библиотекой
ДНК из библиотеки кДНК E. coli выделяли, как описано ранее. Суммарную плазмидную ДНК расщепляли в течение 4 часов при 37°C NotI, чтобы удалить плазмидные последовательности, полученные из E. coli. После очистки ДНК растворяли в стерильной деминерализованной воде.
Множественные трансформации A. niger DS2978 выполняли3, используя от 1,5 × 107 до 3,0 × 107 протопластов и 10 мкг плазмидной ДНК на трансформацию. Трансформанты отбирали по наличию маркера селекции amdS на основании роста на ацетамиде в качестве единственного источника N. Поскольку и селектируемый маркер amdS, и экспрессирующая кДНК кассета находятся во встроенном фрагменте, рост на ацетамиде является показателем наличия экспрессирующей кДНК кассеты.
Примерно после 7-10 дней инкубации при 30°C очищали 10000 трансформантов: трансформанты Aspergillus niger переносили автоматизированным способом (отборочный автомат FlexysJcolony) из планшетов для трансформации в 96-луночные сводные планшеты (МР) MTP, содержащие 150 мкл на лунку отвержденной селективной среды (SM) (на 1000 мл: 0,52 г KCl, 1,52 г K2HPO4, 0,52 г MgSO4, 20 г глюкозы, 1 г ацетамида, 0,1 М буфер MES, 15 г агара, 1 мл раствора микроэлементов [раствор микроэлементов (содержащий в 1 литре): 2,2 г ZnSO4/7H2O, 1,1 г H3BO3, 0,5 г FeSO4/7H2O, 0,17 г CoCl2/6H2O, 0,16 г CuSO4/5H2O, 0,15 г NaMoO4/2H2O, 5,0 г EDTA, pH 6,5], рН 5,5). Трансформанты выращивали на SM в течение 5 дней при 34°C. Созданный таким образом набор MP использовали для того, чтобы 1) инокулировать MTP для роста и последующего расщепления ферментами, и 2) создания резервных планшетов (BP) библиотеки кДНК, которые хранили при -80°C.
Пример 4: Анализ экспрессирующей библиотеки T. emersonii
МР после выращивания в течение 5 дней использовали в качестве репликативной матрицы, и реплику помещали на планшеты со свежей селективной средой (SM), содержащей в литре: 0,52 г KCl, 1,52 г K2HPO4, 0,52 г MgSO4, 20 г глюкозы, 1 г ацетамида, 0,1 М буфер MES, 15 г агара, 1 мл раствора микроэлементов [раствор микроэлементов (на 1 литр): 2,2 г ZnSO4/7H2O, 1,1 г H3BO3, 0,5 г FeSO4/7H2O, 0,17 г CoCl2/6H2О, 0,16 г CuSO4/5H2O, 0,15 г NaMoO4/2H2O, 5,0 г EDTA, pH 6,5], pH 5,5.
После инокулирования планшеты инкубировали при 34°C в течение 48 час. Затем планшеты заполняли верхним агаром, содержащим карбоксиметилцеллюлозу (КМЦ) (5 г агарозы, 0,5 г КМЦ (Sigma, ссылка C4888), приготовленным в 1000 мл 50 мМ фосфатного буфера, pH 7). После затвердевания верхнего агара планшеты оставляли при 65°C в течение 4 часов. Для визуализации активности планшеты красили раствором Конго красного (10 г Конго красного в 1000 мл фосфатного буфера, pH 7) в течение 15 минут. Окрашивающий раствор удаляли и планшеты промывали 1 M NaCl (58,44 г в 1 литре дистиллированной воды). Эту последнюю стадию повторяли дважды. Позитивные клоны проявлялись в результате формирования бледного осветленного ореола на красном фоне.
Клоны, позитивные в отношении целлюлазы, после указанного первого скрининга (проявляющиеся по осветленному ореолу после скрининга с помощью Конго красного) снова инокулировали на свежую среду SM и выращивали в течение 5 дней при 34°C. Затем делали реплики с полученного таким образом матричного планшета на селективной среде и на селективной среде, содержащей 0,075% (мас./об.) AZCL-целлюлозы (Megazyme катал. ссылка I-AZCEL). SM-планшеты обрабатывали и оценивали, как описано ранее (рост при 34°C и последующий скрининг посредством нанесения покрывающего слоя, содержащего целлюлозу, и окрашивания Конго красным), тогда как планшеты с SM-AZCEL-целлюлозой инкубировали при 34°C в течение 48 час, и затем далее инкубировали при 65°C в течение 8 час. Планшеты с SM-AZCL-целлюлозой оценивали до и после инкубации при высокой температуре. Позитивные в отношении целлюлазы клоны в результате давали диффузный голубой ореол.
В конце концов, идентифицировали 20 позитивных в отношении целлюлазы клонов. Продуцирующие целлюлазу трансформанты Aspergillus, которые идентифицировали при анализе планшетов в отношении ксиланазы, выращивали во встряхиваемых флаконах для ферментации.3 Через 5 дней ферментации отбирали образцы среды и анализировали в отношении целлюлазной активности, как описано далее.
Пример 5: Генетический анализ позитивных трансформантов
Идентифицированные позитивные (повторно подтвержденные) трансформанты выращивали на жидкой среде, собирали мицелий и выделяли суммарную (хромосомную) ДНК, используя систему для выделения Puregene (Biozym B. V.), предназначенную для выделения ДНК из нитчатых грибов. Выделение и очистку ДНК выполняли согласно протоколу поставщика с небольшими модификациями: повторяли стадии 3 и 4 осаждения белка.
Хромосомную ДНК использовали в качестве матрицы в реакции ПЦР, используя праймеры 12207 (SEQ ID NO: 8) и 11937 (SEQ ID NO: 7), чтобы амплифицировать вставку(ки), присутствующую в экспрессирующей кассете, интегрированной в хромосомную ДНК.
Прямые ПЦР на трансформантах выполняли согласно адаптированной версии известного протокола4, по которому полученный мицелий затем обрабатывают GlucanexJ (Novo Nordisk) при концентрациях 5 мг/мл вместо NOVOzyme.
Реакционные смеси для ПЦР содержали буфер eLONGaseTM B (Life Technologies, Breda, The Netherlands), дНТФ (200 мкМ каждого), 1 мкл смеси ферментов eLONGase™, 1-5 мкл матрицы и 10-30 пмоль каждого из олигонуклеотидов в конечном объеме 50 мкл. Оптимальное количество олигонуклеотидов определяли экспериментально для каждой приобретенной партии. В среднем использовали от 10 до 30 пмоль. Реакции выполняли при следующих условиях цикла: 1 × (2 мин) 94°C, 35 × (1 мин 94°C, 1 мин 55°C, 6 мин 72°C), 1 × (7 мин 72°С). Образцы наносили на агарозные гели для анализа продуктов ПЦР.
Полученный таким образом продукт ПЦР субклонировали в клонирующем векторе E. coli pcr2.1 (Invitrogen, согласно инструкциям поставщика), получая в результате плазмиду pGBCEA-1.
Субклонированный продукт ПЦР секвенировали. Полученная в результате нуклеотидная последовательность кодирующего района изображена в SEQ ID NO 1, а рассчитанная аминокислотная последовательность белка - в SEQ ID NO 2. Данный белок был назван CEA.
Пример 6: Вискозиметрический скрининг экспрессирующей библиотеки кДНК с использованием аппарата для вискозиметрии Hamilton
Вискозиметрические измерения
Капиллярные вискозиметры являются наиболее широко используемым типом вискозиметров, применяемых для измерений вязких жидкостей. В общем, представляющую интерес жидкость заставляют течь сквозь капиллярную трубку при известной разнице давления. Затем измеряют скорость потока, обычно, отмечая время, требуемое для того, чтобы заданный объем жидкости перешел градуировочную отметку. В других типах капиллярных вискозиметров жидкость заставляют протекать через капилляр при предварительно определенной скорости потока, и измеряют получаемый при этом перепад давления на концах капилляра для того, чтобы определить вязкость жидкости. Контролируемое ферментативное расщепление полимерных жидких растворов можно использовать для определения ферментативной активности. В том случае, если известно численное значение взаимосвязи между вязкостью и концентрацией жидкости, можно оценить такие кинетические параметры, как константа Михаэлиса-Ментена.
I. Система регистрации.
(а) Настройка аппарата для вискозиметрии Hamilton
Аппарат для пипетирования Hamilton (Hamilton Workstation Microlab 2200, Hamilton Company, Reno, USA) контролировали с помощью компьютерной программы, названной Eclipse (Hamilton Company). Указанная компьютерная программа дает пользователю возможность направлять зонд-манипулятор в определенное положение в пределах рабочего пространства Hamilton. Разработали стандартную программу Eclipse для анализа вязкости. Указанная программа работает на 96-луночных MTP, при этом каждую лунку можно адресно отдельно анализировать. Использовали шприцы для того, чтобы отсосать или распределить жидкость образца в заданном Eclipse положении на протяжении канала иглы. По желанию устанавливали как скорость аспирации, так и скорость распределения (в сек/мл), а также объемы аспирации и распределения (в мкл). При введении функции определения положения в держателе можно оценить множество сосудов с реагентами эффективным времясберегающим способом. Перенос жидкости образца с одной стадии пипетирования на следующую устраняли посредством введения команды Eclipse промывать трубку жидкостью системы. Скорость аспирации, равная 10 сек/мл (иногда 15 сек/мл), подходила для того, чтобы различить большинство из вязкостей при просмотре высоты пиков. Обычно отсасывали и распределяли 200-250 мкл жидкости образца.
(b) Настройка Hamilton
Аспирация жидкости образца дает перепад давления на протяжении заполненной водой трубки. Величина перепада давления зависит как от скорости аспирации (или движения поршня), так и от вязкости жидкости образца. Наибольший перепад давления создается на конце иглы, так как радиус здесь наименьший (около 0,3 мм). Аспирация жидкости образца вызывает изменения давления, которые передаются от Т-коннектора к датчику давления. Датчик (Depex B. V., de Bilt, The Netherlands) превращает величину разрежения в электрический ток. Электрический ток регистрируется регистрирующим устройством Datataker один раз в секунду и потом храниться в памяти Datataker в цифровом формате. Компьютер, который связан с Datataker, делает возможным (с помощью компьютерной программы Delogger) загрузку памяти Datataker в файлы. Файлы в свою очередь могут считываться и анализироваться обычными программами крупноформатных таблиц (такими, как Microsoft Excel). Так как давление измеряли во времени, конечные выходные данные можно визуально представить посредством построения графика зависимости разрежения (в мВ) от времени (в секундах). При перемещении Т-коннектора в положение непосредственно за иглой сигнал давления меньше зависит от объема аспирации.
(с) Получение калибровочных кривых, позволяющих определять вязкость любой жидкости
Согласно закону Пуазеля измеряемый перепад давления должен быть прямо пропорционален вязкости, так как
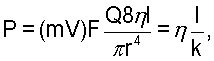 где
где 
С помощью указанной формулы можно связать выходные данные по разрежению в милливольтах с неизвестной вязкостью жидкости образца. Это можно сделать при аспирации определенного постоянного объема калибровочной жидкости (обычно глицерина, так как он охватывает широкие пределы вязкости и ведет себя подобно ньютоновской жидкости) при разных концентрациях. Созданное таким образом разрежение связано с выходом в мВ неизвестным фактором F. Так как известны абсолютные вязкости разных концентраций глицерина, график выходных данных разрежения в мВ для разных известных концентраций глицерина против известных абсолютных вязкостей тех же самых концентраций глицерина дает прямую линию, проходящую через начало координат (так как P=(mV)·F=η/k).
(d) Программа Eclipse, используемая для полномасштабного скрининга
Один цикл измерения состоит из аспирации образца объемом 250 мкл и субстратной смеси надосадка при скорости аспирации 10 сек/мл. Перед аспирацией жидкости образца насасывают 40 мкл воздуха, чтобы отделить систему от жидкости образца. После завершения аспирации жидкости образца следует распределение 290 мкл образца и воздуха. Указанный цикл измерения повторяли шесть раз, и затем следовала стадия промывки 5 мл, чтобы очистить трубку.
II. Разработка анализа
(e) Определение оптимальной субстратной смеси пектина и ксилана для скрининга
Использование более одного вязкого субстрата одновременно имело несколько преимуществ. В одно и то же время можно проводить скрининг разных типов ферментов. Это сохраняло большое количество усилий и времени, причем требовалось меньше субстрата. Кроме ксилана овсяной полбы было решено использовать пектин в качестве второго субстрата. Раствор 1:1 1% пектина и 7% ксилана овсяной полбы, по-видимому, был наиболее подходящим для скрининга. Так как в библиотеке было пропорциональное небольшое количество позитивных клонов, большинство пиков имело приблизительную высоту, равную 435 мВ. В случае образца, позитивного по ксиланазе, весь или большая часть ксилана расщеплялась, так что оставалось только 0,5% пектина. Поэтому клон, позитивный в отношении ксиланазы, давал пик высотой 280 мВ. В случае клона, расщепляющего пектин, высота пика составляла 220 мВ.
(f) Определение минимальной активности ксиланазы, необходимой для регистрации с помощью аппарата для вискозиметрии Hamilton
Скрининг проводили с очень небольшими объемами как субстрата, так и надосадка, поскольку отдельная лунка МТР максимально вмещает 360 мкл. Кроме того, чем больше добавляли надосадка, тем больше становилось разведение субстрата и тем больше снижалась исходная вязкость. В лунки МТР распределяли по 250 мкл 6% ксилана овсяной полбы. Готовили серийные разведения эталонной ксиланазы (ксиланаза A. tubigensis с активностью 685400 EXU/г, где 1 EXU=4,53 моль восстанавливающих сахаров/мин/г), и 30 мкл каждого разведения добавляли к субстрату. После инкубации в течение 24 часов при 50°C измеряли вязкость каждого образца. Самая маленькая регистрируемая концентрация фермента соответствовала 53,6 нг/мл или 0,0367 EXU/мл. Поэтому добавление 20 мкл надосадка к 300 мкл субстрата будет обеспечивать возможность регистрации очень низких активностей фермента.
(g) Подготовка скрининга для идентификации термостабильных ферментов
Для того чтобы из библиотеки отобрать только термостабильные ферменты и для того чтобы избежать помех, связанных с активностью ферментов хозяина - A. niger, планшеты с клонами, содержащими термостабильные испытываемые ферменты, подвергали термической обработке. Достоверность системы проверяли, используя пустые штаммы хозяина, штаммы хозяина, экспрессирующие термолабильные ферменты, и штаммы хозяина, экспрессирующие термостабильные ферменты. После роста штаммов и продукции фермента планшеты МТР нагревали при 72°С в течение 30 минут. Затем 20 мкл надосадка добавляли к 300 мкл 6% ксилана овсяной полбы. Вместе с негативными контролями (добавление 20 мкл воды) планшеты, содержащие образцы, герметично закрывали клейкой лентой и инкубировали при 60°C в сушильном шкафу. Высокая температура может увеличивать активность термостабильного фермента, но нарушает как фоновую мешающую активность фермента хозяина A. niger, так и активности нетермостабильных ферментов, экспрессируемых библиотекой. После инкубации в течение 20 часов не обнаружено заметного снижения высоты пиков для клонов с термолабильной ксиланазой, что свидетельствует о том, что активность ксиланазы хозяина полностью инактивирована. Дополнительно в качестве контроля были включены клоны Aspergillus niger с термолабильным ферментом ксиланазой xynB. В данном случае не регистрировали остаточной активности, хотя, с другой стороны, устойчивая к нагреванию ксиланаза, которую включали в качестве контроля, еще была активной. Тепловая инактивация в течение 30 минут при 72°C и время инкубации образца, равное 24 часам или меньше, давали авторам возможность специфично выявлять термостабильные ксиланазы T. emersonii.
III. Скрининг экспрессирующей библиотеки кДНК с использованием автоматического устройства для вискозиметрии Hamilton
(h) Получение реплик экспрессирующей библиотеки кДНК и экспрессия библиотеки
Один цикл получения реплик включает в себя двукратную инокуляцию как в селективную среду (SM) (содержащую в литре: 0,52 г KCl, 1,52 г K2HPO4, 0,52 г MgSO4, 20 г глюкозы, 1 г ацетамида, 0,1 М буфер MES, 15 г агара, 1 мл раствора микроэлементов [раствор микроэлементов (на литр): 2,2 г ZnSO4/7H2O, 1,1 г H3BO3, 0,5 г FeSO4/7H2O, 0,17 г CoCl2/6H2О, 0,16 г CuSO4/5H2O, 0,15 г NaMoO4/2H2O, 5,0 г EDTA, pH 6,5], pH 5,5), так и в жидкую ростовую среду (GM) (содержащую в литре 70 г глюкозы, 25 г гидролизата казеина, 12,5 г дрожжевого экстракта, 1 г KH2PO4, 0,5 г K2SO4, 2 г MgSO4, 0,03 г ZnCl2, 0,02 г CaCl2, 0,01 г MnSO4, 0,3 г FeSO4, pH 5,6). Библиотеку хранили в стандартных MTP, в которых спорулированные рекомбинантные колонии A. niger выросли в каждой лунке на SM в присутствии 10% глицерина. При хранении указанные планшеты (сводные планшеты, MP) держали в замороженном состоянии (-80°C). Перед получением реплик MP их размораживали в течение одного часа в стерилизованном вытяжном шкафу, чтобы избежать микробного загрязнения. Получение реплик MP осуществляли с помощью репликатора колоний PBA Flexys-Colony. Сводные планшеты после 5 дней роста использовали в качестве репликативных матриц и реплики высевали в 96-луночные MTP, наполненные жидкой ростовой средой (GM). Свежеинокулированные планшеты с агаризованной SM помещали в инкубатор при 32°C и затем хранили при -80°C после добавления в каждую лунку 150 мкл 10% глицерина. Продуцирующие GM-планшеты инкубировали в смоченном водой встряхивателе Tomtec Quadrastor 96000 (32°C). Растущий мицелий появлялся на планшетах через 2-3 дня. GM-планшеты инкубировали в течение 6 дней.
(i) Отбор образцов надосадков после завершения роста
После 6 дней роста оставшуюся жидкую ростовую среду GM (содержащую внеклеточные продукты экспрессии) извлекали и переносили в новые MTP. Надосадки переносили в новые МТР, используя 4-канальное автоматическое устройство для пипетирования Tecan. Средний объем извлеченного надосадка составлял от 120 до 140 мкл. Полученные планшеты с надосадками замораживали и позже анализировали в отношении активности ксиланазы.
(j) Подготовка для полномасштабного скрининга
Планшеты с надосадками из экспрессирующей библиотеки размораживали и помещали в закрытую водяную баню при 72°C на 35-40 минут. Используя 12-канальную пипетку и коммерческие предварительно приготовленные наконечники в одноразовых держателях MTP-формата (поставляемые Eppendorf), 20 мкл каждого надосадка переносили в МРТ, содержащий субстратную смесь, который перед добавлением надосадков нагревали до 60°C. Субстратная смесь состояла из смеси 1:1 1% пектина (смотри (k) далее) и 7% ксилана овсяной полбы (смотри (1) ниже), при конечном pH 4,12. Анализируемые MTP наполняли 310 мл субстрата/мл. Смешивания надосадков с субстратной смесью достигали при перемешивании наконечниками пипетки в лунках с образцом сразу после разнесения надосадков. Затем планшеты помещали в сушильный шкаф при 60°C на период инкубации, равный 18-24 часам. После инкубации MTP давали возможность охладиться, и проводили скрининг снижения вязкости, используя автоматическое устройство для вискозиметрии Hamilton (Hamilton Company, Reno, USA).
(k) Приготовление 1% пектина, pH 4,1
Буфер на основе лимонной кислоты-фосфата (буфер Mc Ilvaine) готовили добавлением 0,2 М раствора Na2HPO4 к 250 мл 0,1 М раствора лимонной кислоты до получения значения pH 5,5. Его доводили до одного литра дистиллированной водой и при необходимости снова доводили рН. 0,5% раствор пектина готовили медленным добавлением 0,5 г высокометилированного пектина (тип Ruban Brun) в сосуд, содержащий 50 мл указанного выше буфера McIlvaine и 25 мл дистиллированной воды, при 60°C. Энергичное встряхивание обеспечивало хорошее растворение пектина. Раствор доводили до 100 мл и снова проверяли pH. Использованный в данном случае пектин понижал рН раствора, так что при этих условиях раствор имел 5,1. Если раствор пектина был мутным, полезным было центрифугирование раствора при 15 g в течение 15 минут, чтобы удалить любые нерастворившиеся частицы.
(l) Приготовление 7% обработанного щелочью ксилана овсяной полбы, pH 4,1
20 мл 2 М NaOH нагревали до 60°C в стакане объемом 150 мл. В отдельном стакане добавляли 7 г ксилана овсяной полбы (из компании Sigma) к 20 мл воды, так что происходило образование ярко коричневой рыхлой массы. При необходимости еще добавляли воду, но всего не более 30 мл. Используя стальную ложку, полученную массу, содержащую воду и ксилан овсяной полбы, медленно добавляли к горячему раствору NaOH. Было необходимо устройство для энергичного перемешивания, чтобы растворить ксилан в растворе гидроксида, при этом температуру сохраняли на постоянном уровне в 60°C. После того как растворялся весь ксилан, раствор становился темно-коричневым и был похож на очень вязкий, прозрачный сироп. Затем добавляли остаток воды, так чтобы было добавлено общее количество, равное 50 мл. Раствору давали возможность охладиться и добавляли 4 н. HCl до достижения требуемого значения pH (обычно pH 4,1-4,2). Раствор доводили до 100 мл и при необходимости снова доводили pH. Центрифугирование при 20000 g в течение 15 минут при 10°C давало прозрачный желтоватый надосадок (вязкость которого зависела от исходного количества добавленного ксилана овсяной полбы).
(m) Скрининг библиотеки
В результате скрининга библиотеки Talaromyces emersonii, клонированной в A. niger, получили 119 графиков, соответствующих 119 тестированным MTP. Каждый график показывал вязкость в 96 лунках, представленную в виде 96 пиков, которые являются мерой разрежения для каждой лунки в мВ. Графики анализировали в отношении наличия низких пиков, которые указывали на пониженную вязкость. Пики, которые были ниже, чем средняя высота пиков, отбирали для повторного тестирования. Некоторые планшеты давали очень немного изменений, так что отбор предполагаемых позитивных клонов для повторного тестирования был нетрудным. Другие планшеты показали большое количество изменений, поэтому для повторного тестирования часто отбирали более 5 или 6 предполагаемых клонов. Чтобы иметь позитивный контроль после инкубации, добавляли 10 мкл эндоксиланазы в случайно выбранный планшет в заданное положение. Используя указанные позитивные контроли, обнаружили, что в том случае, когда присутствует активность ксиланазы, следует ожидать высоты пика от 240 до 280 мВ.
Планшеты библиотеки Talaromyces, которые были тестированы, также содержали пять подтвержденных клонов с термостабильной ксиланазой, которые были выявлены до использования анализа регистрации с красителем, основанного на растворении красителей, которые были химически связаны с нерастворимым полимерным субстратом. Указанные пять клонов без исключения независимо были обнаружены при вискозиметрическом анализе уже в первом раунде скрининга. Все пики известных клонов были на уровне намного более низком, чем остальные пики (при 250 мВ). Всего для повторного тестирования отобрали 118 лунок с культурой, за исключением лунок с ранее обнаруженными клонами с ксиланазой.
(n) Повторное тестирование 118 предполагаемых клонов
Повторное тестирование было основано на вискозиметрическом анализе согласно принципу, используемому в случае полномасштабного скрининга. Однако при этом использовали больший объем для анализа, в таком случае можно было более четко различить активность термостабильного фермента и нерегулярные низкие пики. Два больших МТР (2 мл/лунку) заполняли 1,2 мл 9% ксилана овсяной полбы. В первые две лунки каждого ряда добавляли по 50 мкл воды (негативный контроль). В последнюю лунку каждого ряда добавляли 50 мкл стандартного раствора эндоксиланазы для того, чтобы также иметь позитивный контроль. С 3 по 11 лунку каждого ряда использовали для повторного тестирования предполагаемых клонов с ксиланазой (добавление 50 мкл надосадка). После перемешивания и инкубации в течение 24 часов при 60°C измеряли вязкость с помощью специально для этого предназначенной программы Eclipse (объем аспирации: 800 мкл, скорость аспирации: 10 сек/мл).
Идентифицировали несколько позитивных клонов, так как их пики были точно на таком же уровне, как и пики позитивных контролей, что свидетельствовало о полном расщеплении ксилана. Всего в повторном тесте при скрининге на основе вискозиметрии идентифицировано тринадцать предполагаемых штаммов. Вставки ДНК прямо клонировали в векторе pCR2.1 посредством ПЦР-амплификации, осуществляемой на хромосомной ДНК. Из 12 штаммов библиотеки с использованием полимеразы pwo и набора праймеров 2 (11937/12207, SEQ ID NO: 7 и 8) получено шестнадцать вставок кДНК. Ориентацию вставки кДНК в векторе pCR2.1 определяли расщеплением XhoI, которое имело место в векторе и на 3'-конце кДНК. Анализ последовательности ДНК осуществляли на 5'-конце вставок кДНК.
Повторное тестирование возможных разрушающих пектин ферментов выполняли подобно тестированию в случае ксиланазы, за исключением того, что вместо 9% ксилана овсяной полбы использовали 1% пектин. Повторно тестировали только десять возможных клонов, которые представляли собой клоны, дававшие очень низкие пики при первичном раунде скрининга, но не проявляли какой-либо активности во время повторного тестирования в отношении ксиланазы. Один клон воспроизводимо продуцировал фермент, разрушающий пектин. Измеренное значение давления в 200 мВ соответствовало вязкости чистой воды. Поскольку используемый пектин был не полностью метилирован, его могла расщеплять термостабильная полигалактуроназа, пектатлиаза или пектинлиаза.
(o) Преимущества скрининга на основе вискозиметрии
Разработке способа скрининга на основе вискозиметрии для идентификации термостабильных ферментов способствовало три ключевых фактора. Во-первых, возможность термической инактивации нативной ксиланазной активности хозяина позволяла намного быстрее установить соответствующие условия реакции. Во-вторых, тот факт, что в случае тестирования известных термолабильных ферментов термическая инактивация устраняла всю активность, так что отбирали только более термостабильные ферменты. В-третьих, повышенные температуры увеличивали ферментативные активности, так что скрининг становился даже более чувствительным. На основе ксилана овсяной полбы (Sigma) идентифицировали одиннадцать клонов, которые обладали способностью в значительной степени снижать вязкость, и на основе пектина был идентифицирован один четко установленный клон, разрушающий пектин. Кроме того, независимо снова были обнаружены пять клонов с термостабильной ксиланазой, которые были идентифицированы ранее.
Пример 7: Характеристика первой термостабильной β-глюканазы (СЕА) Talaromyces emersonii из трансформанта A. niger
Анализы и определения активности
Определение PAHBAH-единицы целлюлазы (CPU)
Одну единицу целлюлазной активности определяют как количество фермента, необходимое для высвобождения одного мкмоль восстанавливающих сахаров, продуцируемых в минуту при pH 5,0 и 60°C при концентрации субстрата - 5% карбоксиметилцеллюлозы (КМЦ) с использованием калибровочной кривой для глюкозы.
Согласно способу CPU ферментативную активность измеряют посредством регистрации восстанавливающих сахаров с использованием гидразида 4-гидроксибензойной кислоты (PAHBAH). Анализ основан на способе Lever, M., Powell, J. C., Killip, M., Small, C. W. (1973) J. Lab. Clin. Med. 82, 649-655 с небольшими модификациями. Модификация состоит в приготовлении реагента PAHBAH следующим образом: 0,05 М цитрат тринатрия, 0,1 М Na2SO3, 0,02 М CaCl2, 0,5 М NaOH и 0,1 М гидразида парагидроксибензойной кислоты (PAHBAH). Конечное значение pH было равно 12. Реагент, содержащий PAHBAH в щелочном растворе, который хранят при комнатной температуре, следует использовать в течение одного дня. В качестве эталонного восстанавливающего сахара использовали глюкозу (калибровочная кривая). Для калибровочной кривой в данном анализе конечные концентрации глюкозы составляли от 0 до 300 мМ. Активность CPU анализировали при смешивании 100 мкл раствора фермента с 400 мкл 5% КМЦ в 0,1 М натрий-ацетатном буфере (pH 5,0). Пробирки Эппендорф с субстратом (КМЦ) предварительно инкубировали в течение 5 минут при 60°C. Реакцию начинали добавлением раствора фермента. Через 15 минут реакцию останавливали добавлением 1,0 мл PAHBAH-реагента. Пробирки Эппендорф нагревали в течение 5 минут при 100°C и затем охлаждали на льду. Образцы центрифугировали с соответствующей скоростью, чтобы осадить любые твердые материалы (1 минута при полной скорости в микроцентрифуге Е Beckman). Оптическую плотность измеряли при 420 нм. Пустой контроль готовили добавлением 100 мкл 0,1 М натрий-ацетатного буфера вместо раствора фермента.
Определение единицы бета-глюканазы (BGU)
Единица бета-глюканазы представляет собой активность, необходимую для освобождения 0,258 мкмоль восстанавливающих сахаров (измеренных в виде эквивалента глюкозы) в минуту при pH 3,5 и 40°C при концентрации субстрата 0,5% β-глюкана ячменя.
Таким образом, кроме указанного выше определения активности целлюлазы (CPU), выполняли более специфичный анализ для выявления активности β-глюканазы. Принцип анализа состоит в определении скорости, с которой снижается вязкость раствора β-глюкана ячменя средней вязкости (Megazyme, Australia, 2/11 Ponderosa Parade, Warriewood NSW 2101) при добавлении определенного количества фермента. β-глюкан ячменя средней вязкости (Megazyme Australia) растворяли в 0,425 М натрий-цитратном буфере с pH 3,5 до концентрации 6,25 мг/мл. Субстрат инкубировали при 40°C в течение 10 минут. Затем добавляли небольшое количество фермента (в пределах 0,005-0,062 единиц/мл) и давали возможность реакции продолжаться. Во временной точке реакции, равной 60 минутам, определяли вязкость образца относительно стандартного образца, который инкубировали со стандартной эндоглюканазой с известной ферментативной активностью. Абсолютные активности стандарта определяли способом на основе восстанавливающих сахаров, используя Fe-III-гексацианид и 4,76 мг/мл β-глюкана ячменя в качестве исходной концентрации субстрата.
Определение единицы эндоксиланазы (EXU)
Единицу активности ксиланазы (EXU) определяют как количество фермента (эндо-1,4-β-ксиланазы Asp. niger, которая описана в EP-A-0463706 (Gist-brocades B. V.)), которое высвобождает 4,53 мкмоль восстанавливающих сахаров (измеряемых в виде эквивалентов ксилозы) в минуту в условиях анализа. Условия анализа включают в себя: 5 мг/мл арабиноксилана пшеничной муки (Megazyme, Australia, 2/11 Ponderosa Parade, Warriewood NSW 2101) в 100 мМ натрий-цитратном буфере (pH 3,5), температуру 40°C, время реакции 60 минут. Реакции останавливали добавлением 1 М NaOH. Регистрацию выполняли колориметрически при 420 нм после инкубирования образцов с Fe-III-гексацианидом в течение 15 минут в кипящей воде. Реагент на основе гексацианоферрата готовили растворением 1,17 г KFe(CN) и 19,5 г безводного карбоната натрия в 1 литре воды.
Кроме указанного выше определения абсолютного значения ксиланазной активности использовали относительный способ, при котором следили за снижением вязкости раствора арабиноксилана пшеницы (Megazyme, Australia, 2/11 Ponderosa Parade, Warriewood NSW 2101) при добавлении определенного количества фермента. Арабиноксилан пшеницы растворяли в 0,425 М натрий-цитратном буфере (pH 3,5) до концентрации 8,3 мг/мл. Субстрат инкубировали при 55°C в течение 10 минут. Затем добавляли небольшое количество фермента (в пределах 0,01-0,05 единиц/мл) и реакции давали возможность продолжаться. Во временной точке реакции, равной 60 минутам, определяли вязкость образца относительно стандарта, который инкубировали со стандартом эндоксиланазы Aspergillus niger известной EXU-активности (EP-A-0463706). Абсолютные значения активности стандарта в EXU определяли способом на основе восстанавливающих сахаров, используя Fe-III-гексацианид, как описано выше. Вязкость определяли вручную, используя вискозиметр с падающим шариком Haake.
Определение единицы целлюлазы (CXU)
Единицу активности бета-глюканазы (BGU) определяют как количество целлюлазы, которое гидролизует за один час количество гликозидных связей, эквивалентное продукции 0,5 мг глюкозы в условиях анализа. Условия анализа включают в себя: 9 мг/мл карбоксиметилцеллюлозы в 5 мМ натрий-ацетатном буфере (pH 4,6), температуру 37°С. Освободившуюся глюкозу определяли способом на основе восстанавливающих сахаров, используя реагент DNS.
Кроме указанного выше определения абсолютного значения целлюлазной активности использовали относительный способ, при котором следили за снижением вязкости раствора карбоксиметилцеллюлозы при добавлении определенного количества фермента. Карбоксиметилцеллюлозу растворяли в 5 мМ натрий-цитратном буфере (pH 4,6) до концентрации, которая зависит от вязкости партии. Субстрат инкубировали при 37°C в течение 10 минут. Затем добавляли небольшое количество фермента, и реакции давали возможность продолжаться. После протекания реакции в течение 60 минут определяли вязкость образца относительно стандарта, который инкубировали со стандартом с известной CXU-активностью.
Активность в зависимости от pH
β-глюканазу T. emersonii получали с использованием соответствующего трансформанта(тов) A. niger во встряхиваемых флаконах, как описано в примере 3, после удаления выросшего мицелия фильтрованием. Для данного эксперимента использовали фильтрат. Активность фильтрата измеряли при разных значениях pH при фиксированной температуре, равной 60°C. Активность измеряли CPU-способом. Вместо использования фиксированного значения pH, равного pH 5,0, КМЦ-субстрат разбавляли цитратным буфером соответствующего рН, чтобы получить рН измерения. Эксперимент повторяли дважды, и результаты показаны ниже в таблице 1. Обнаружено, что оптимальное значение pH для фермента составляет от pH 4,5 до 5,0, примерно при pH 4,8.
Профиль рН при 60°С
(измерения в двух повторах)
Активность в зависимости от температуры
Затем измеряли активность указанной β-глюканазы при разных температурах. Измерения активности проводили CPU-способом при рН 4,0. Вместо инкубации при фиксированной температуре, равной 60°С, инкубации проводили при разных температурах. Эксперимент проводили дважды, и результаты показаны ниже в таблице 2. Обнаружено, что оптимум температур составляет от 80 до 85°С. Топт., по-видимому, приблизительно равна 84°С (хотя она может составлять 82, 83 или 85°С, в зависимости от того, как проведены и интерполированы линии по точкам данных).
Температурный профиль при рН 4
(°С)
(измерения в двух повторах)
Активность по сравнению с другими ферментами
Кроме того, измеряли активность при 30-60°С, используя BGU-способ для β-глюканазы, и сравнивали с коммерчески доступным очищенным от клеток T. reesei бульоном, обладающим целлюлазной и глюканазной активностью, как стандартом. Бульон, не содержащий клеток T. reesei, состоял из смеси разных ферментов, среди которых имеется одна или несколько β-глюканаз, но в котором данные активности не охарактеризованы. Для сравнения активностей β-глюканазы T. emersonii согласно изобретению и целлюлаз/глюканаз T. reesei ферменты дозировали примерно при равной активности при 40°С. Активность β-глюканазы T. emersonii при 40°С принимали за единицу, и таким образом другие активности выражены относительно активности данного фермента. Результаты показаны ниже в таблице 3.
Относительная активность по сравнению с ферментами T. reesei
(°С)
При температурах выше 40°С активность целлюлазы/глюканазы T. reesei начинала выравниваться с активностью β-глюканазы T.emersonii. Выше 40°C активности расходились, при этом CEA была более активной. Также, несмотря на то, что относительная активность была выше при повышенных температурах, при средних температурах активность по-прежнему соответствовала смеси T. reesei. Это свидетельствует о широком диапазоне температур, при которых β-глюканаза является активной.
Очистка и удельная активность
Очистка β-глюканазы T. emersonii начиналась с фильтрата. Примерно 25 мл не содержащего клеток бульона высаливали на колонке PD 10 и наносили на анионообменную колонку resource Q объемом 6 мл, которую уравновешивали 10 мМ натрий-ацетатным буфером (рН 5,0). Элюирование выполняли линейным градиентом 10-300 мМ натрий-ацетатного буфера (pH 5,0). (Линейный градиент от A до B; скорость потока: 6 мл/мин; время прохождения: 54 мин; регистрация при длине волны: 280 нм, 254 нм, 214 нм). Активные фракции собирали (20 мл), концентрировали с помощью концентраторов Microsep (3,5 мл, 10 K) и дважды промывали 0,1 M натрий-фосфатным буфером (pH 5,0). Чистоту фракций анализировали посредством ВЭЖХ-SEC (эксклюзионной хроматографии по размеру) и SDS-ПААГ.
Измерения удельной активности
Рассчитанный коэффициент молярной экстинкции β-глюканазы при 280 нм составляет 81550 M-1 см-1. Концентрацию белка очищенного фермента получали на основе измерений E280, используя удельную оптическую плотность E2801см=2,33 для 1 мг/мл. Удельную активность очищенного образца определяли в BGU (субстрат: бета-глюкан ячменя), и она составляла 1027 BGU/мг. При использовании CPU-способа специфичная активность составляла 628 единиц/мг. Так как активность согласно вискозиметрическому способу давала более высокое значение по сравнению со способом на основе восстанавливающих сахаров, был сделан вывод о том, что β-глюканаза T. emersonii проявляет значительную эндоглюканазную активность. Типичная экзоглюканаза проявляла бы очень хорошую эффективность в анализе восстанавливающих сахаров, и очень плохую в вискозиметрическом анализе.
Изоэлектрическая точка
ИЭФ-ПААГ выполняли следующим образом. Оборудование представляло собой систему Phast (Pharmacia Biotech), IEF 3-9 PhastGel (Pharmacia Biotech). Гели разгоняли и красили (Кумасси) согласно стандартным способам для системы Phast. Изоэлектрическую точку определяли на PhastGel IEF3-9, и она оказалась равной 3,3.
Определение молекулярной массы
SDS-ПААГ выполняли следующим образом. Снова использовали оборудование Phast (Pharmacia Biotech); 12,5% гомогенные гели (Pharmacia Biotech); SDS-буферные полоски (Pharmacia Biotech). Обработка образца: один объем буфера (5 образцов) (500 мМ трис-HCl, pH 6,8, 10% SDS, 0,1% бромфенолового синего) смешивали с 4 объемами образца и кипятили в течение 3 минут. Гели разгоняли и красили (Кумасси) согласно стандартным способам для системы Phast. ВЭЖХ-эксклюзионную хроматографию по размеру выполняли, используя колонку: TSK G3000SW, катал. No. 05103 (TosoHaas). Способ: элюенты представляли собой: 0,1 М натрий-фосфатный буфер pH 7,0; скорость потока: 1 мл/мин; время прохождения: 30 мин, регистрация при длине волны: 280 нм. Дегликозилирование фермента: смесь 5 мкл очищенного фермента (7 мг/мл) с 20 мкл 0,5% SDS и 25 мкл 1% β-меркаптоэтанола. Полученную смесь кипятили в течение 4 минут. После охлаждения добавляли 20 мкл N-гликозидазы F (500 U/мл) и 20 мкл 3% тритона X-100 в 1 М натрий-фосфатном буфере, pH 7,0. Все это инкубировали в течение ночи, и дегликозилирование анализировали с помощью SDS-ПААГ.
Молекулярная масса, определенная в SDS-ПААГ-геле и HP-SEC, оказалась равной 43 кДа. После дегликозилирования в SDS-ПААГ была видна ровная полоса при 37 кДа.
Термостабильность
T50-термостабильность определяли, используя в качестве стандарта целлюлазы/глюканазы T. reesei. T50 является температурой, при которой после 20 минут инкубации теряется 50% активности. В таблице 4 показаны различия в T50-термостабильности между очищенной β-глюканазой T. emersonii и смесью целлюлаз/глюканаз T.reesei, используемой ранее в данном примере (исходная активность была принята за единицу: ферменты инкубировали в течение 20 минут, и после охлаждения измеряли активность, используя CPU-способ). Каждый эксперимент повторяли дважды, и поэтому в таблице 4 показаны двойные измерения. T50-термостабильность очищенной β-глюканазы находится около 93,4°C, а T50-термостабильность целлюлаз/глюканаз T. reesei - около 64,9°C.
Термостабильность по сравнению с ферментами T. reesei
(°С)
Пример 8: Измерения активности
Два штамма библиотеки, которые были идентифицированы при вискозиметрическом скрининге в примере 6 с использованием в качестве субстрата ксилана овсяной полбы, подвергали ферментации во встряхиваемых флаконах, используя среду GM (исходные штаммы маркировали AD009.21 и AD011.31). Кроме того, включали штамм, маркированный AD021.B6 (CEA), который идентифицировали в примерах 4 и 5 как штамм, экспрессирующий β-глюканазную активность. Провели несколько анализов, и результаты показаны ниже. Неожиданно штаммы AD009.21 и AD011.31 проявляли низкую ксиланазную активность. Вместо этого AD009.21 и AD011.31 по-видимому обладают целлюлазной активностью, даже несмотря на то, что клоны были идентифицированы при вискозиметрическом скрининге с использованием ксилана овсяной полбы. В частности βD009.21 очень активен на β-глюкане ячменя.
Ферментация целлюлазных штаммов во встряхиваемых флаконах;
Активность ксиланазы (EXU/мл), β-глюканазы (BGU/мл) и целлюлазы (CXU/мл)
Пределы регистрации были установлены на уровне ниже 10 для анализов EXU и BGU и 5 для анализа CXU.
Анализы ДНК штаммов AD 009.21 и AD 011.31 (которые описаны ранее в примере 6) выявили две новые β-глюканазы, названные CEB и CEC. Нуклеотидные последовательности кодирующих районов представляют собой SEQ ID NO: 3 и 5, а соответствующие аминокислотные последовательности представляют собой SEQ ID NO: 4 и 6 соответственно.
Штамм CEB проявляет высокую β-глюканазную активность. Штамм CEA проявляет относительно высокую целлюлазную активность, по сравнению со штаммом CEB. Соотношение между β-глюканазной активностью и целлюлазной активностью различно в случае CEA и CEB. Штамм CEC проявлял целлюлазную активность, а его β-глюканазная или ксиланазная активность была ниже пределов регистрации в данных анализах. Однако исследования гомологии строго подтвердили наличие β-глюканазной активности, и предполагается, что данная активность может быть намного выше при других значения рН.
Можно использовать более одного субстрата для скрининга. Неожиданно оказалось, что при вискозиметрическом скрининге вместо двух субстратов одновременно можно использовать три разных субстрата. Одним из субстратов была примесь полимера глюкозы, вероятно глюкана или целлюлозного материала. Показано, что использование ксилана овсяной полбы, пектина и материала, подобного целлюлозе, в результате приводило к идентификации трех типов ферментов, ксиланаз, пектиназ и целлюлаз в одном раунде скрининга. Вискозиметрический анализ обладает одним другим большим преимуществом по сравнению с обычным анализом, основанном на регистрации с помощью красителя. Анализ с регистрацией с помощью красителя основан на красителях, которые химически связываются с субстратом. Данный вид субстрата коммерчески доступен только в случае небольшого числа соединений. Однако вискозиметрический анализ может работать с любым типом вязкого субстрата, даже природными субстратами, которые не требуют какой-либо предварительной химической обработки. Указанный аспект важен как с коммерческой, так и промышленной точки зрения, можно проводить скрининг ферментативных активностей на субстратах, снижение вязкости которых (например, фруктовых соков) или увеличение вязкости которых (например, молочных продуктов) имеет коммерческое значение.
Пример 9: Эффективность эндоглюканазы Talaromyces emersonii (CEC) при выпекании хлеба
Получение формованного хлеба при стандартном способе выпечки выполняли смешиванием 3500 г пшеничной муки (смесь 80% пшеничной муки Kolibri и 20% пшеничной муки Ibis (Meneba, Holland) примерно при 21°C), 77 г прессованных дрожжей (Konings), 70 г соли, 25 т.п.н. аскорбиновой кислоты, 10 т.п.н. α-амилазы грибов FermizymeTMP200, (DSM N. V., Bakery Ingredients, Delft, The Netherlands) и разных количеств фермента эндоглюканазы CEC (из 4 разных клонов, идентифицированных своим номером AD) и 2030 мл воды (8-15°C) в спиральном миксере (Hobart) в течение 2 минут (при скорости 1) и примерно в течение 6 минут (при скорости 2), чтобы ввести 125 Вт-час (ватт-час) энергии. Температура теста составляла 28°C. Возможности механической обработки теста вручную анализировал квалифицированный пекарь.
Сразу после смешивания тесто делили на 6 кусков, каждый по 875 г, округляли и расстаивали в течение 35 минут в камере расстойки при 34°C и 85% ОВ (относительной влажности). В конце этого периода времени тесто формовали и укладывали в формы и давали окончательно расстояться в течение 75 минут в камере расстойки при 38°C и 87% ОВ. Затем полностью расстоявшееся тесто выпекали в электрической печи при 210°C в течение 30 минут. После охлаждения до комнатной температуры определяли объемы буханок хлеба способом определения с использованием рапсового семени. После хранения в течение 16-24 часов в герметичных полиэтиленовых пакетах при комнатной температуре опытный пекарь оценивал качество хлебного мякиша. Четыре штамма библиотеки, для которых было установлено, что они экспрессируют CEC, подвергали ферментации во встряхиваемых флаконах и проверяли в данном тесте выпекания хлеба. Результаты показаны в таблицах ниже.
буханки
(мл) (%)
(шкала 0-7)
56
189
4357
4289
106
104
Легче, не прилипает
Легче, некоторая липкость
6
6,5
буханки
(мл) (%)
(шкала 0-7)
56
189
4427
4419
108
108
Легче, не прилипает
Легче, некоторая липкость
6
6
буханки
(мл) (%)
(шкала 0-7)
56
189
4432
4622
108
113
Легче, не прилипает
Легче, некоторая липкость
5,5
6
буханки
(мл) (%)
(шкала 0-7)
56
189
4450
4553
108
111
Легче, не прилипает
Легче, некоторая липкость
5,5
6
Качество теста было очень хорошим. При более высоких дозировках эндоглюканазы СЕС липкость была небольшой при испытании во время ручной обработки теста. Однако эта небольшая липкость не влияла на возможность проводить механическую обработку теста. Все партии теста, содержащие эндоглюканазу СЕС, были очень эластичными и легко подвергались ручной обработке.
На основе полученных результатов при выпекании сделан вывод, что эндоглюканаза СЕС очень эффективно улучшает качество хлеба, и в отношении объема буханки и в отношении качества хлебного мякиша. Несмотря на большие объемы буханок структура хлебного мякиша еще оставалась очень равномерной и тонкой.
Пример 10. Сравнение эффективности фермента Talaromyces emersonii (CEC) при выпекании хлеба с эндоксиланазами Asp. niger
Эффективность CEC при выпекании в случае производства формованного голландского хлеба сравнивали с используемой в настоящее время эндоглюканазой грибов из Aspergillus niger. Указанную эндоглюканазу A. niger добавляли в чистой коммерчески доступной форме, т.е. FermizymeTM HS2000.
Точно повторяли процедуру примера 9, за исключением того, что использовали разные количества либо эндоглюканазы СЕС, либо FermizymeTM HS2000.
(мл) (%)
(мл) (%)
Из полученных результатов ясно, что эндоглюканаза СЕС увеличивает объем буханки в более высокой степени, чем объем, получаемый при введении FermizymeTM HS2000. Более того, необходимо меньше единиц эндоглюканазы/кг муки, чтобы достичь определенного уровня объема буханки, в том случае если вместо FermizymeTM HS2000 используют СЕС.
Пример 11
Сравнение некоторых молекулярных и биохимических свойств трех глюканаз представлено ниже в таблицах.
(37 кДа)
(эндоглюканаза)
(эндоглюканаза)
(эндоглюканаза)
+ предсказана на основе аминокислотной последовательности
EC 3.2.1.75
EC 3.2.1.58
EC 3.2.1.78
EC 3.2.1.123
эндо-1,6-глюканаза
экзо-1,3-глюканаза
маннаназа
эндогликоцерамидаза
считается маловероятной
EC 3.2.1.91
Целлобиогидролаза
СЕВ, СЕС все являются целлюлазами, которые проявляют активность ЕС 3.2.1.4. Многие целлюлазы также могут гидролизовать 1,4-связи в глюкане ячменя, что делает их особенно полезными для некоторых применений, таких, например, как удаление антипищевых факторов в корме. Так как активность измеряли с помощью анализа вязкости, вероятно, что целлюлаза проявляет эндоактивность. Однако нельзя исключать, что также гидролизуются 1,3-связи. Следовательно, по наблюдениям нельзя исключить активности EC 3.2.1.73, 3.2.1.39 и 3.2.1.6, даже, несмотря на то, что эти активности выявлены в семействе 16: характеристики семейств 7 и 16, по-видимому, довольно похожи.
Исключительно в качестве объяснения, целлюлазы (EC 3.2.1.4) обычно обладают способностью катализировать эндогидролиз 1,4-D-глюкозидной связи в целлюлозе. Систематическое название целлюлаз 1,4-(2,3;1,4)-D-глюкан-4-глюканогидролаза. Эндо-1,4-D-глюканаза и эндоглюканаза D являются синонимами 1,4-(1,3;1,4)-D-глюкан-4-глюканогидролазы. Целлюлаза также может гидролизовать 1,4-связи в D-глюканах, содержащих также 1,3-связи, что делает их особенно полезными для некоторых применений, таких, например, как удаление антипищевых факторов из корма. Такие ферменты, которые проявляют эндоглюканазную активность, в общем, называют глюканазами. Кроме того, гидролиза глюканов можно добиться с помощью лихеназы (1,3-1,4-D-глюкан-глюканогидролазы (EC 3.2.1.73)) и эндо-1,3(4)-глюканазы (1,3-(1,3;1,4)-D-глюкан-3(4)-глюканогидролазы (EC 3.2.1.6)).
Обзор целлюлазных активностей показан в таблице ниже.
Также будет гидролизовать 1,4-связи в β-D-глюканах, содержащих также 1,3-связи
1,4)-β-D-глюкан-4-глюканогидролаза
Эндоглюканаза D
β-1,3-глюканаза, Олиго-1,3-глюко-зидаза
β-1,6-глюкан-гидролаза
Бета-1,3-1,4-глюканаза,
Эндо-β-1,3(4)-глюканаза
Целлобио-гидролаза,
Экзоцелло-биогидролаза
ССЫЛКИ









| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИД И КОМПОЗИЦИЯ, ОБЛАДАЮЩИЕ АКТИВНОСТЬЮ КСИЛАНАЗЫ, ПРИМЕНЕНИЕ ПОЛИПЕПТИДА, ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ ПОЛИПЕПТИД, ВЕКТОР ЭКСПРЕССИИ, СОДЕРЖАЩИЙ ПОЛИНУКЛЕОТИД, СПОСОБ ПОЛУЧЕНИЯ ПОЛИНУКЛЕОТИДА, СПОСОБ ОБРАБОТКИ РАСТИТЕЛЬНОГО ИЛИ КСИЛАНСОДЕРЖАЩЕГО МАТЕРИАЛА | 2000 |
|
RU2291901C2 |
| ЦЕЛЛЮЛОЗНЫЕ БЕЛКИ СЛИЯНИЯ И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2458127C2 |
| НОВЫЕ РЕГУЛИРУЮЩИЕ ЭКСПРЕССИЮ ПОСЛЕДОВАТЕЛЬНОСТИ И ПРОДУКТ ЭКСПРЕССИИ В ОБЛАСТИ ГИФОМИЦЕТОВ | 2001 |
|
RU2272835C2 |
| ЛИПОЛИТИЧЕСКИЙ ФЕРМЕНТ ДЛЯ ПРИМЕНЕНИЯ В ХЛЕБОПЕЧЕНИИ | 2018 |
|
RU2763469C2 |
| ПРОТИВОМИКРОБНЫЕ ПОЛИПЕПТИДЫ ИЗ PSEUDOPLECTANIA NIGRELLA | 2002 |
|
RU2336278C2 |
| ГЕН ABFB-2 PENICILLIUM FUNICULOSUM | 2006 |
|
RU2388820C2 |
| МОДИФИЦИРОВАННЫЕ ФИТАЗЫ | 2003 |
|
RU2329301C2 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| ПОЛИПЕПТИДЫ С ПРОТИВОМИКРОБНОЙ АКТИВНОСТЬЮ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2005 |
|
RU2393224C2 |
| КЛОНИРОВАНИЕ, ЭКСПРЕССИЯ И ПРИМЕНЕНИЕ КИСЛЫХ ФОСФОЛИПАЗ | 2010 |
|
RU2567659C2 |
Изобретение относится к биотехнологии и представляет собой бета-глюканазу, получаемую из гриба Talaromyces emersonii, или композицию, обладающую активностью бета-глюканазы. Данный фермент или композиция используются при обработке растительного материала для разрушения или модификации целлюлозы, а также входят в состав пищевых продуктов и кормов для животных. Заявленное изобретение позволяет расширить ассортимент ферментов, обладающих бета-глюканазной активностью. 9 н. и 8 з.п. ф-лы, 15 табл.
i) аминокислотную последовательность SEQ ID NO: 2, 4 или 6; или
ii) вариант (i) обладающий, по меньшей мере, 70%, 75%, 80% или 85% идентичностью с аминокислотной последовательностью SEQ ID NO: 2, 4 или 6; или
iii) фрагмент (i) или (ii), который способен расщеплять β-D-глюкан.
a) оптимум рН ниже 5,4; или
b) оптимум температуры, по меньшей мере 80°С, как, например, 83-87°С.
(а) последовательность нуклеиновой кислоты SEQ ID NO: 1, 3 или 5 или последовательность, кодирующую полипептид по одному из пп.1-3;
(b) последовательность, которая комплементарна, или гибридизуется в жестких условиях с последовательностью по подпункту (а) на протяжении района, по меньшей мере, из 100 нуклеотидов, в частности на протяжении полной длины SEQ ID NO: 1, 3 или 5;
(c) модифицированную последовательность SEQ ID NO: 1, 3 или 5, имеющую до 100 нуклеотидных замещений;
(d) последовательность, обладающую, по меньшей мере, 70% идентичностью с последовательностью по подпунктам (а), (b) или (с); или
(e) последовательность, которая является вырожденной в результате свойства генетического кода по отношению к любой из последовательностей по подпунктам (a)-(d).
(a) последовательность, которая кодирует полипептид, обладающий активностью β-глюканазы, которая представляет собой
(1) кодирующую последовательность SEQ ID NO: 1, 3 или 5;
(2) последовательность, которая гибридизуется в жестких условиях с комплементом последовательности по подпункту (1); или
(3) последовательность, которая является вырожденной в результате свойства генетического кода по отношению к последовательности по подпунктам (1) или (2); или
(b) последовательность, комплементарную полинуклеотиду по подпункту (а).
| MOLONEY А.Р | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Приспособление для установки двигателя в топках с получающими возвратно-поступательное перемещение колосниками | 1917 |
|
SU1985A1 |
| US 3880742, 29.04.1975 | |||
| WO 9806858 A, 19.02.1998 | |||
| RU 2070921 C1, 27.12.1996. | |||

