Настоящее изобретение относится к полипептидам, в особенности к эритропоэтину, конъюгированному с гидроксиалкилкрахмалом (HAS), и в особенности с гидроксиэтилкрахмалом.
Применение полипептидов, особенно ферментов или цитокинов, для введения их в систему кровообращения с целью достижения конкретного физиологического эффекта представляет собой хорошо известный инструмент в современной медицине.
Эритропоэтин (EPO) представляет собой гликопротеиновый гормон, необходимый для созревания эритроидных клеток-предшественников в эритроциты. У взрослых людей он образуется в почках. EPO необходим для регулирования уровня эритроцитов в кровотоке. Условия, определяемые низким уровнем кислорода в тканях, провоцируют усиление биосинтеза EPO, что, в свою очередь, стимулирует эритропоэз. Снижение функциональной активности почек, которая отмечается при хронической почечной недостаточности, в типичном случае может, например, приводить к снижению биосинтеза EPO и к сопутствующему снижению уровня эритроцитов.
Эритропоэтин представляет собой кислый гликопротеидный гормон массой примерно 34000 Да. Человеческий эритропоэтин представляет собой полипептид из 166 аминокислот, который в природе обычно представлен в форме мономера (Lin et al, PNAS 82, 7580-7584, EP 148605 B2, EP 411678 B). Идентификация, клонирование и экспрессия генов, кодирующих эритропоэтин, описана, например, в патенте США 4703008. Выделение и очистка рекомбинантного эритропоэтина из культуральной среды, используемой для выращивания клеток млекопитающих, содержащих плазмиды для получения рекомбинантного эритропоэтина, описаны, например, в патенте США 4667016.
В основном в данной области техники известно, что биологическая активность EPO in vivo в основном зависит от уровня содержащихся сиаловых кислот, которые присоединяются к EPO (см., например, EP 428267 B1). Теоретически 14 молекул сиаловой кислоты могут присоединиться к одной молекуле EPO на терминальных участках боковых цепей углеводов, присоединенных к N- и О-сайтам гликозилирования. Для получения высокосиалилированных препаратов EPO требуются чрезвычайно сложные стадии очистки.
В литературе имеется детальная информация по различным аспектам эритропоэтина (см., например, Krantz, Erythropoietin, 1991, Blood, 77(3):419-34 (обзор) and Cerami, Beyond erythropoiesis: novel application for recombinant human erythropoietin, 2001, Semin Hematol., (3 Suppl 7):33-9 (обзор).
Ряд имеющихся проблемы, связанных с применением полипептидов и ферментов, заключается в том, что указанные белки зачастую проявляют неудовлетворительную стабильность. В особенности известно, что эритропоэтин обладает относительно коротким периодом полувыведения из плазмы (Spivak and Hogans, 1989, Blood 73, 90; McMahon et al., 1990, Blood 76, 1718). Данный аспект указывает на то, что терапевтические уровни в плазме быстро снижаются и должны проводиться повторные внутривенные введения. Кроме того, в некоторых случаях наблюдается иммунный ответ против вводимых пептидов.
В основном считается, что стабильность полипептидов может быть улучшена и иммунный ответ против указанных полипептидов снижен, если полипептид связать с полимерными молекулами. В WO 94/28024 показано, что физиологически активный полипептид, модифицированный полиэтиленгликолем (PEG), демонстрирует сниженную иммуногенность и антигенность и циркулирует в кровотоке значительно дольше, чем неконъюгированные белки, то есть имеет более длительный клиренс.
Однако конъюгатам PEG-лекарственное средство свойственны некоторые недостатки, определяемые тем, что они не имеют природной структуры, которая может распознаваться элементами путей деградации in vivo. В этой связи кроме PEG-конъюгатов были разработаны другие конъюгаты и белковые полимерные образования. В литературе описано множество методов сшивания различных белков и макромолекул, таких как метод с использованием полимеразы (см., например, Wong, Chemistry of protein conjugation and cross-linking, 1993, CRCS, Inc.).
Гидроксиэтилкрахмал (HES) представляет собой производное природного амилопектина, которое подвергается деградации в организме под действием α-амилазы. Получение HES-пептидных конъюгатов описано в данной области техники (см., например, HES-hemoglobin-conjugates в DE 2616086 или DE 2646854).
В DE 2646854 раскрываются способы конъюгирования гемоглобина с HES. В рамках данных методик HES подвергают реакции с перйодатом натрия, приводящей к образованию диальдегидов, которые связывают с гемоглобином. В отличие от указанной методики, в документе DE 2616086 раскрывается конъюгирование гемоглобина с HES по процедуре, в которой вначале сшивающий агент (например, бромциан) связывают с HES и затем гемоглобин связывают с промежуточным продуктом.
HES представляет собой замещенное производное углеводного полимера амилопептида, который присутствует в кукурузном крахмале в концентрации до 95 мас.%. HES демонстрирует полезные биологические свойства и используется в качестве кровезаменителя и при лечении по методу гемодилюции в клинических условиях (Sommermeyer et al., 1987, Krankenhauspharmazie, 8(8), 271-278, и Weidler et al., 1991, Arzneim.-Forschung/Drug Res., 41, 494-498).
Амилопептид состоит из двух фрагментов глюкозы, причем в главной цепи имеются α-1,4-гликозидные связи, а в сайтах ветвления найдены α-1,6-гликозидные связи. Физико-химические свойства данной молекулы в основном определяются типом гликозидных связей. Из-за точечных разрывов α-1,4-гликозидных связей образуются спиральные структуры с шестью глюкозными мономерами на виток структуры.
Физико-химические, а также биохимические свойства полимера могут быть модифицированы путем замещения. Введение гидроксиэтильной группы может быть осуществлено путем щелочного гидроксиэтилирования. При соответствующей адаптации реакционных условий можно использовать различную реакционную способность в отношении гидроксиэтилирования соответствующей гидроксильной группы в незамещенном глюкозном мономере. В этой связи любой специалист со средним уровнем знаний в данной области может в некотором ограниченном диапазоне оказывать воздействие на характер замещения.
Соответственно HES в основном характеризуется определенным распределением по молекулярной массе и степени замещения. Имеются две возможности характеристики степени замещения:
1. Степень замещения может быть описана относительно части замещенных глюкозных мономеров в сравнении с соответствующим показателем для всех глюкозных фрагментов (DS).
2. Степень замещения может быть охарактеризована как "молярное замещение" (MS), когда описывают количество гидроксиэтильных групп, приходящихся на фрагмент глюкозы.
Растворы HES присутствуют в виде полидисперсных композиций, где каждая молекула отличается от другой степенью полимеризации, числом и характером сайтов ветвления, а также характером замещения. В этой связи HES представлен смесью соединений с разной молекулярной массой. Следовательно, конкретный раствор HES может быть определен по средней молекулярной массе с помощью статистических методов. В данном контексте Mn вычисляют как среднее арифметическое значение, зависящее от числа молекул. Альтернативно MW,как среднее значение молекулярной массы, отражает единицу, которая зависит от массы HES.
Известные в настоящее время конъюгаты HES-лекарственное средство имеют недостаток, связанный с тем, что HES не конъюгирует сайт-специфичным образом относительно лекарственного средства. В этой связи конъюгирование приводит к получению очень гетерогенного продукта, включающего много компонентов, которые могут быть неактивными из-за деструкции трехмерной структуры на стадии конъюгирования.
Резюмируя, следует отметить, что все еще имеется потребность в более усовершенствованных полипептидах с улучшенной стабильностью и/или биологической активностью. Указанное особенно относится в эритропоэтину, в случае которого изоформы с высокой степенью присоединенных сиаловых кислот и в этой связи с более высокой активностью должны быть отделены при очистке от изоформ с низким уровнем содержания сиаловых кислот (см. EP 428267 B1). В этой связи было бы очень выгодно, если бы были доступны способы производства, которые обеспечивали бы получение высокоактивных полипептидов без потребности в тщательной очистке. К сожалению, получение пептидов в бактериях или в клетках насекомых часто затруднено, поскольку полипептиды зачастую не продуцируются в таких случаях в соответствующей складчатой, нативной конфигурации и не имеют соответствующего гликозилирования.
Следовательно, объектом настоящего изобретения является получение полипептидных производных, в особенности производных эритропоэтина, обладающих высокой биологической активностью in vivo, которые могут быть легко получены и со сниженной стоимостью. Кроме того, другим объектом настоящего изобретения является разработка способа получения полипептидных производных, который было бы легко осуществлять и который приводил бы к получению продуктов с высокой биологической активностью. Еще одним объектом настоящего изобретения является получение фармацевтической композиции, включающей полипептидные производные с высокой биологической активностью.
В соответствии с одним аспектом настоящего изобретения указанная проблема решается с помощью конъюгата гидроксиалкилкрахмал (HAS) - эриторопоэтин (EPO) (HAS-EPO), содержащего одну или несколько молекул HAS, где каждая HAS конъюгирована с EPO через
а) углеводный фрагмент или
b) тиоэфир.
Конъюгат HAS-EPO по настоящему изобретению обладает преимуществом, связанным с тем, что он демонстрирует повышенную биологическую активность в сравнении с эритропоэтином до конъюгирования. Кроме того, он демонстрирует более высокую биологическую активность, чем стандартный препарат EPO BRP. Указанное свойство связано главным образом с тем, что HAS-EPO в меньшей степени или вовсе не распознается системами удаления в печени и почках, и в этой связи указанный конъюгат циркулирует в системе кровообращения в течение длительного периода времени. Кроме того, поскольку HAS подвергается сайт-специфичному воздействию, минимизируется риск нарушения биологической активности EPO in vivo путем конъюгирования HAS с EPO.
Конъюгат HAS-EPO по настоящему изобретению включает в основном два компонента, а именно: эритропоэтин (EPO)-полипептид и гидроксиалкилкрахмал (HAS), присоединенный к нему.
EPO может быть человеческого происхождения (см., например, Inoue, Wada, Takeuchi, 1994, An improved method for the purification of human erythropoietin with high in vivo activity from the urine of anemic patients, Biol Pherm Bull. 17(2), 180-4; Miyake, Kung, Goldwasser, 1977, Purification of human erythropoietin., J Biol Chem., 252(15), 5558-64) или может происходить из другого млекопитающего в качестве источника и может быть получен путем выделения и очистки из природных источников, таких как почка человека, эмбриональная печень человека, предпочтительно почки обезьяны. Кроме того, выражение "эритропоэтин" или "EPO" относится к варианту EPO, где одна или несколько аминокислот (например, от 1 до 25, предпочтительно от 1 до 10, более предпочтительно от 1 до 5, наиболее предпочтительно от 1 до 2) заменены на другую аминокислоту и который проявляет эритропоэтическую активность (см., например, EP 640619 B1). Определение эритропоэтической активности описано в литературе (например, измерение активности in vitro приведено в Fibi et al., 1991, Blood, 77, 1203 ff; Kitamura et al., 1989, J.Cell Phys., 140, 323-334; описание метода измерения активности EPO in vivo см. в Ph.Eur. 2001, 911-917; Ph. Eur. 2000, 1316 Erythropoietini solutio concentrata, 780-785; European Pharmacopoeia (1996/2000); European Pharmacopoeia 1996, Erythropoietin concentrated solution, Pharmaeuropa., 8, 371-377; Fibi, Hermentin, Pauly, Lauffer, Zettimeissl., 1995, N- and O-glycosylation muteins of recombinant human erythropoietin secreted from BHK-21 cells, Blood, 85(5), 1229-36) (EPO и модифицированные формы EPO инъецируют самкам мышей NMRI (что эквивалентно по белку 50 нг/мышь) в дни 1, 2 и 3, на 4 день отбирают образцы крови и определяют число ретикулоцитов). Другие публикации, в которых описаны тесты измерения активности, включают: Barbone, Aparicio, Anderson, Natarajan, Ritchie, 1994, Reticulocytes measurement as a bioassay for erythropoietin, J. Pharm. Biomed, Anal., 12(4), 515-22; Bowen, Culligan, Beguin, Kendal, Villis, 1994, Estimation of effective and total erythropoiesis in myelodysplasia using serum transferrin receptors and erythropoietin concentrations, with automated reticulocyte parameters, Leukemi, 8(1), 151-5; Delorme, Lorenzini, Giffin, Martin, Jacobsen, Boone, Elliot, 1992, Role of glycosylation on the secretion and biological activity of erythropoietin, Biochemisrty, 31(41), 9871-6; Higuchi, Oh-eda, Kuboniwa, Tomonoh, Shimonaka, Ochi, 1992; Role of sugar chains in the expression of the biological activity of human erythropoietin, J. Biol. Chem., 267(11), 7703-9; Yamaguchi, Akai, Kawanishi, Ueda, Masuda, Sasaki, 1991, Effects of site-directed removal of N-glycosylation sites in human erythropoietin on its production and biological properties, J. Biol. Chem., 266(30), 20434-9; Takeuchi, Inoue, Strickland, Kubota, Wada, Shimizu, Hoshi, Kozutsumi, Takasaki, Kobata, 1989, Relationship between sugar chains structure and biological activity of recombinant human erythropoietin produced in Chinese hamster ovary cells, Prod, Natl. Acad. Sci. USA, 85(20), 7819-22; Kurtz, Eckardt, 1989, Assay methods for erythropoietin, Nephron., 51(1), 11-4 (German); Zucali, Sulkowski, 1989, Purification of human urinary erythropoietin on controlled-pore glass and silicic acid, Exp. Hematol., 13(3), 833-7; Krystal, 1983, Physical and biological characterization of erythroblast enhancing factor (EEF), a late acting erythropoietin stimulator in serum distinct from erythropoietin, Exp. Hematol., 11(1), 18-31.
Предпочтительно EPO получают рекомбинантными способами. Указанный вариант включает продукцию в эукариотических или прокариотических клетках, предпочтительно в клетках млекопитающих, насекомых, дрожжей, бактериальных клетках или в клетках любого другого типа, которые удобны для получения EPO рекомбинантными методами. Кроме того, EPO может экспрессироваться в трансгенных животных, например в жидкостях организма (таких как молоко, кровь и т.п.), в яйцах трансгенных птиц, в особенности у сельскохозяйственных птиц, предпочтительно у кур, или в трансгенных растениях.
Получение полипептидов по рекомбинантной технологии известно специалистам в данной области техники. В целом такая продукция включает трансфекцию клетки-хозяина соответствующим экспрессирующим вектором, культивирование клеток-хозяев в условиях, позволяющих осуществлять продукцию полипептида и выделение и очистку полипептида из клеток-хозяев. Подробная информация содержится в ряде публикаций (см., например, Kristal, Pankratz, Farber, Smart, 1986, Purification of human erythropoietin to homogeneity by a rapid five-step procedure, Blood, 67(1), 71-9; Quelle, Caslake, Burkert, Wojchowski, 1989, High-level expression and purification of a recombinant human erythropoietin produced using a baculovirus vector, Blood, 74(2), 652-7; EO 640619 B1 and EP 668351 B1).
В предпочтительном варианте осуществления настоящего изобретения EPO имеет аминокислотную последовательность человеческого EPO (см., EP 148605 B2).
EPO может включать одну или несколько углеводных боковых цепей (предпочтительно 1-4, предпочтительно 4), присоединяемых к EPO через N- и/или O-связанное гликозилирование, т.е. EPO гликозилируется. Обычно в случае продукции EPO в эукариотических клетках полипептид подвергается посттрансляционному гликозилированию. Следовательно, углеводные боковые цепи могут присоединяться к EPO в процессе синтеза в клетках млекопитающего, особенно человека, клетках насекомых или дрожжей. Структура и свойства гликозилированного EPO широко исследовались в данной области (см. EP 428267 B1; EP 640619 B1, Rush, Derby, Smith, Merry, Rogers, Rohde, Katta, 1995, Microgeterogeneity of erythropoietin carbohydrate structure, Anal Chem., 67(8), 1442-52; Takeuchi, Kobata, 1991, Structure and functional roles of the sugar chains of human erythropoietin, Glycobiology, 1(4), 337-46 (обзор)).
HAS может быть непосредственно конъюгирован с EPO или, альтернативно, он может быть конъюгирован через молекулу линкера. Природа линкерной молекулы зависит от способа, посредством которого HAS присоединяется к EPO. Возможные функциональные группы линкеров показаны в таблице 1 и в приведенном ниже описании. Некоторые линкеры коммерчески доступны (например, от Pierce, доступной от компании Perbio Science Deutschland GmbH, Bonn, Germany). Некоторые подходящие линкеры описаны в таблице 2. Природа линкера и его назначение подробно описаны ниже в разделе, относящемся к способу получения HES-EPO.
В соответствии с предпочтительным вариантом осуществления настоящего изобретения в аспекте конъюгата HAS-EPO указанный HAS подвергают конъюгированию с EPO через углеводный фрагмент.
В контексте настоящего описания термин "углеводный фрагмент" относится к гидроксиальдегидам или гидроксикетонам, а также к их химическим модификациям (см. Rompp Chemielexicon, Thieme Stuttgart, Germany, 9th edition 1990, Volume 9, pages 2281-2285, и цитированная в работе литература). Кроме того, указанный термин также относится к производным природных углеводных фрагментов типа глюкозы, галактозы, маннозы, сиаловой кислоты и т.п. Указанный термин также включает химически окисленные природные углеводные фрагменты, которые имеют открытую циклическую структуру.
Углеводный фрагмент может быть присоединен непосредственно к полипептидному скелету EPO. Предпочтительно углеводный фрагмент представляет собой часть углеводной боковой цепи. В данном случае другие углеводные фрагменты могут присутствовать между углеводным фрагментом, к которому присоединяется HAS, и полипептидным скелетом EPO. Более предпочтительно углеводный фрагмент представляет собой терминальный фрагмент углеводной боковой цепи.
В предпочтительном варианте изобретения HAS подвергают конъюгированию с галактозным остатком углеводных боковых цепей, предпочтительно с терминальным галактозным остатком углеводной боковой цепи. Указанный галактозный остаток может быть открыт для конъюгирования за счет удаления терминальных сиаловых кислот с последующим окислением (см. приведенное ниже описание).
В другом более предпочтительном варианте HAS подвергают конъюгированию с остатком сиаловой кислоты в углеводных боковых цепях, предпочтительно с терминальным остатком сиаловой кислоты в углеводной боковой цепи.
Кроме того, HAS может быть подвергнут конъюгированию с EPO через тиоэфир. Как будет описано подробно ниже, атом S может происходить из SH-группы, присоединенной к EPO как в природном, так и в неприродном варианте.
В предпочтительном варианте осуществления изобретения атом S может содержаться в SH-группе, которую вводят в окисленный углеводный фрагмент HES, предпочтительно окисленный углеводный фрагмент, который представляет собой часть углеводной боковой цепи EPO (см. приведенное ниже описание).
Предпочтительно атом S в тиоэфире получают из природного цистеина или из добавленного цистеина. Более предпочтительно EPO имеет аминокислотную последовательность человеческого EPO, а природные цистеины представляют собой цистеин 29 и/или 33. В более предпочтительном варианте HAS подвергают конъюгированию с цистеином 29, а цистеин 33 замещают другой аминокислотой. Альтернативно HAS может быть конъюгирован с цистеином 33, а цистеин 29 заменяют другой аминокислотой.
В контексте настоящего описания термин "добавленные цистеины" следует понимать таким образом, что полипептиды, предпочтительно EPO, включают цистеиновый остаток, который не присутствует в полипептиде дикого типа.
В контексте данного аспекта изобретения цистеин может представлять собой дополнительную аминокислоту, добавляемую на N- или С-конце EPO.
Кроме того, добавленный цистеин может быть внесен путем замены природной аминокислоты цистеином. Подходящие способы известны в технике (см. выше). Предпочтительно в контексте данного аспекта изобретения EPO представляет собой человеческий EPO и замененный аминокислотный остаток представляет собой серин 126.
Второй компонент HAS-EPO представляет собой гидроксиалкилкрахмал (HAS).
В контексте настоящего описания термин "гидроксиалкилкрахмал" используют для указания производных крахмала, которые содержат замещение гидроксиалкильными группами. В данном контексте алкильная группа может быть замещена. Предпочтительно гидроксиалкил содержит 2-10 атомов углерода, более предпочтительно 2-4 атома углерода. Термин "гидроксиалкилкрахмал" в этой связи предпочтительно включает гидроксиэтилкрахмал, гидроксипропилкрахмал и гидроксибутилкрахмал, где предпочтительными являются гидроксиэтилкрахмал и гидроксипропилкрахмал.
Гидроксиалкильная группа(ы) HAS содержит по меньшей мере одну OH-группу.
Выражение "гидроксиалкилкрахмал" также включает производные, в которых алкильная группа представляет собой моно- или полизамещенную группу. В данном контексте предпочтительно, чтобы алкильная группа была замещена галогеном, в особенности фтором, или арильной группой, при условии, что HAS остается водорастворимым. Кроме того, терминальная гидроксигруппа гидроксиалкила может быть этерифицирована с образованием простого или сложного эфира. Дополнительно следует отметить, что алкильная группа в гидроксиалкилкрахмале может быть линейной или разветвленной.
Кроме того, вместо алкила могут также использоваться линейные или разветвленные замещенные или незамещенные алкеновые группы.
Гидроксиэтилкрахмал (HES) наиболее предпочтителен для всех вариантов осуществления настоящего изобретения.
В контексте настоящего описания гидроксиэтилкрахмал может иметь значение средней молекулярной массы (средняя масса) от 1 до 300 кДа, где средняя молекулярная масса, равная 5-100 кДа, является более предпочтительной. Гидроксиэтилкрахмал может также демонстрировать степень молярного замещения от 0,1 до 0,8 и соотношение между С2:С6-замещением в диапазоне 2-20 относительно гидроксиэтильных групп.
HAS-EPO может включать 1-12, предпочтительно 1-9, 1-6 или 1-3, наиболее предпочтительно 1-4 молекул HAS на одну молекулу EPO. Количество HAS-молекул на одну молекулу EPO может быть определено путем количественного анализа углеводного состава с использованием GC-MS после гидролиза продукта и дериватизации полученных моносахаридов (см. Chaplin and Kennedy (eds.), 1986, Carbohydrate Analysis: a practical approach, IRL Press Practical approach series (ISBN 0-947946-44-3), в особенности Chapter 1, Monosaccharides, page 1-36; Capter 2, Oligosaccharides, page 37-53, Chapter 3, Neutral Polysaccharides, page 55-96).
HAS-EPO конъюгат по настоящему изобретению может демонстрировать по существу ту же биологическую активность in vitro, что и рекомбинантный нативный EPO, поскольку его биологическая активность in vitro основана на измерении лишь аффинности по связыванию с рецептором EPO. Способы определения биологической активности in vitro известны в данной области техники (см. приведенное выше описание).
Кроме того, HAS-EPO демонстрирует бульшую активность in vivo, чем EPO, используемый в качестве исходного материала для конъюгирования (неконъюгированный EPO). Способы определения биологической активности in vivo известны в технике (см. приведенное выше описание). Кроме того, в примерах 9 и 10 даны тесты для определения активности EPO in vivo и in vitro.
Конъюгат HAS-EPO может демонстрировать активность in vivo, составляющую от 110 до 500%, предпочтительно от 300 до 400%, или от 110% до 300%, предпочтительно от 110% до 200%, более предпочтительно от 110% до 180% или от 110% до 150%, наиболее предпочтительно от 110% до 140%, если активность in vivo неконъюгированного EPO принять за 100%.
В сравнении с высокосиалилированной формой EPO в виде препарата Амген (Amgen) (см. EP 428267 B1), HAS-EPO демонстрирует активность in vivo, равную предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 85% или по меньшей мере 95%, по меньшей мере 150%, по меньшей мере 200% или по меньшей мере 300% от активности высокосиалилированного EPO, если активность in vivo высокосиалилированного EPO принять за 100%. Наиболее предпочтительно указанная форма демонстрирует активность по меньшей мере на уровне 95% от активности in vivo высокосиалилированного EPO.
Высокая биологическая активность in vivo конъюгата HAS-EPO по настоящему изобретению в основном связана с тем фактом, что конъюгат HAS-EPO остается дольше в кровотоке, чем неконъюгированный EPO, поскольку он хуже распознается системами удаления печени и поскольку снижен почечный клиренс из-за более высокой молекулярной массы. Способы определения in vivo периода полувыведения EPO из кровотока известны в данной области техники (Sytkowski, Lunn, Davis, Feldman, Siekman, 1998, Human erythropoietin dimers with markedly enhanced in vivo activity, Proc. Natl. Acad. Sci. USA, 95(3), 1184-8).
В этой связи большим преимуществом настоящего изобретения является тот факт, что HAS-EPO представляет собой форму, которая может вводиться реже, чем препараты EPO, коммерчески доступные в настоящее время. В то время как стандартные препараты EPO должны вводиться по меньшей мере 3 дня, конъюгат HAS-EPO по настоящему изобретению предпочтительно вводится два раза в неделю или предпочтительно один раз в неделю.
Все раскрытые ниже варианты осуществления изобретения, относящиеся к способу производства HAS-EPO по настоящему изобретению, которые относятся к свойствам EPO или HAS, применимы также и к конъюгату HAS-EPO по настоящему изобретению.
Гидроксиалкилкрахмал представляет собой эфирное производное крахмала. В контексте настоящего изобретения, кроме указанных производных в виде простого эфира, могут также использоваться другие производные крахмала. Например, могут использоваться производные, которые включают этерифицированные гидроксигруппы. Такие производные могут представлять собой производные незамещенных моно- или дикарбоновых кислот, включающих 2-12 атомов углерода, или их замещенные производные. Особенно подходят для использования производные незамещенных монокарбоновых кислот, включающие 2-6 атомов углерода, в особенности уксусной кислоты. В контексте настоящего изобретения предпочтительны ацетилкрахмал, бутилкрахлмал или пропилкрахмал.
Кроме того, предпочтительны производные незамещенных дикарбоновых кислот, включающих 2-6 атомов углерода.
В случае производных дикарбоновых кислот полезно, если вторая карбоксигруппа в дикарбоновой кислоте также этерифицирована. Кроме того, в контексте настоящего изобретения также пригодны производные сложных моноалкиловых эфиров дикарбоновых кислот.
В случае замещенных моно- или дикарбоновых кислот могут быть предпочтительны те же замещающие группы, что и указанные выше для случая замещенных алкильных остатков.
Методики этерификации известны в данной области техники (см., например, Klemm D. et al., Comprehensive Cellulose Chemistry Vol. 2, 1998, Whiley-VCH, Wienheim, New York, especially chapter 4.4, Esterification of Celulose (ISBN 3-527-29489-9).
В другом аспекте настоящее изобретение относится к способу получения конъюгата гидроксиалкилкрахмала (HAS)-эритропоэтина (EPO) (HAS-EPO), включающему стадии:
а) получения EPO, способного реагировать с модифицированным HAS,
b) получение модифицированного HAS, способного взаимодействовать с EPO со стадии а), и
c) проведения реакции EPO со стадии а) с HAS со стадии b), посредством чего образуется HAS-EPO, включающий одну или несколько молекул HAS, где каждый из HAS конъюгирует с EPO через
i) углеводный фрагмент или
ii) тиоэфир.
Способ по настоящему изобретению имеет то преимущество, что образуется конъюгат HAS-EPO, который демонстрирует высокую биологическую активность. Кроме того, способ по настоящему изобретению имеет то достоинство, что может быть получено эффективное производное EPO со сниженной стоимостью, поскольку данный способ не включает дорогостоящие и длительные по времени стадии очистки, приводящие к низкому конечному выходу, то есть в данном способе нет необходимости очищать продукт от недостаточно сиалилированных EPO, которые, как известно, характеризуются низкой биологической активностью in vivo или вовсе ее отсутствием. В особенности пример 20 показывает, что получаемый в результате нескольких стадий модификации HES-EPO обладает трехкратной активностью в сравнении со стандартным препаратом BRP-EPO.
Соответственно на первой стадии способа по настоящему изобретению получают EPO, который способен взаимодействовать с модифицированным HAS.
В контексте настоящего описания термин "получение" следует интерпретировать таким образом, что после соответствующей стадии становится доступной молекула (стадии а) EPO на стадии b) HAS) с желательными свойствами.
В случае стадии а) данный термин включает выделение и очистку EPO из природных источников, а также получение в клетках-хозяевах или организмах рекомбинантными методами и, при необходимости, модификацию получаемого при этом EPO.
В том, что касается EPO, выполняющего функцию исходного материала по настоящему изобретению, к нему относятся те же требования, что и в отношении эритропоэтина, представляющего собой часть конъюгата HAS-EPO по настоящему изобретению. В данном контексте предпочтительные варианты, раскрываемые выше, применимы также к способу по настоящему изобретению.
Следовательно, в предпочтительном варианте указанный EPO имеет аминокислотную последовательность человеческого EPO.
Предпочтительно EPO получают рекомбинантным способом. Указанный способ включает получение в эукариотических или прокариотических клетках, предпочтительно в клетках млекопитающего, насекомых, дрожжей, бактерий или в клетках любого другого типа, которые удобны для получения EPO рекомбинантными методами. Кроме того, EPO может быть экспрессирован в трансгенных животных (например, в жидкостях организма типа молока, крови и т.п.), в яйцах трансгенных птиц, особенно сельскохозяйственных птиц, предпочтительно кур, или в трансгенных растениях.
Получение полипептида рекомбинантными меиодами известно в данной области техники. В целом указанный способ получения включает трансфекцию клеток-хозяев соответствующим экспрессирующим вектором, культивирование клеток-хозяев, позволяющие осуществлять продукцию полипептида, а также выделение и очистку полипептида из клеток-хозяев (Kristal, Pankratz, Farber, Smart, 1986, Purification of human erythropoietin to homogeneity by a rapid five-step procedure, Blood, 67(1), 71-9; Quelle, Caslake, Burkert, Wojchowski, 1989, High-level expression and purification of a recombinant human erythropoietin produced using a baculovirus vector, Blood, 74(2), 652-7; EP 640619 B1 и EP 668351 B1).
EPO может включать одну или несколько углеводных боковых цепей, присоединенных к EPO путем N- или O-связанного гликозилирования, т.е. EPO в данном случае подвергается гликозилированию. Обычно, когда EPO продуцируется в эукариотических клетках, полипептид подвергается посттрансляционному гликозилированию. Следовательно, углеводные боковые цепи могут быть присоединены к EPO в процессе образования в клетках млекопитающего, особенно человека, в клетках насекомых или дрожжей, которые могут представлять собой клетки трансгенного животного (см. выше), либо могут быть экстрагированы из животного или все еще находиться в организме животного.
Указанные углеводные боковые цепи могут быть химически или энзиматически модифицированы после экспрессии в соответствующих клетках, например, путем удаления или добавления одного или нескольких углеводных фрагментов (см., например, Dittmar, Conradt, Hauser, Hofer, Lindenmaier, 1989, Advanced in Protein design; Bloecker, Collins, Schmidt, and, Schomburg eds., GBF-Monographs, 12, 231-246, VCH Publishers, Wienheim, New York, Cambridge).
Другой аспект способа по настоящему изобретению относится к получению HAS-EPO, включающему одну или несколько молекул HAS, где указанный HAS конъюгируют с EPO через углеводный фрагмент (i) или через тиоэфир (ii). Следовательно, EPO, образуемый на стадии а), должен иметь свойства, позволяющие осуществлять конъюгирование через углеводный фрагмент и/или тиоэфир. В этой связи EPO после стадии а) может предпочтительно содержать
(1) по меньшей мере одну реакционноспособную группу, присоединенную непосредственно или через линкерную молекулу к сульфидным группам или углеводным фрагментам, которые способны взаимодействовать с HES или с модифицированным HES,
(2) по меньшей мере один углеводный фрагмент, с которым может быть конъюгирован модифицированный HAS, и/или
(3) одну свободную SH-группу.
В том, что касается указанного выше пункта (1), EPO на стадии а) предпочтительно получают путем конъюгирования соответствующей линкерной молекулы с SH-группой(ами) или углеводными фрагментами EPO. Пример такого модифицированного EPO приведен в примере 4, 2.1. Важно отметить, что добавление линкерной молекулы не повреждает EPO. Однако это известно специалистам в данной области техники.
В том, что касается указанного выше пункта 2, в предпочтительном варианте осуществления изобретения модифицированный HAS подвергают конъюгированию с EPO через углеводный фрагмент.
Углеводный фрагмент может быть соединен непосредственно с полипептидным скелетом EPO. Предпочтительно углеводный фрагмент представляет собой часть углеводной боковой цепи. В данном случае другие углеводный фрагменты могут присутствовать между углеводным фрагментом, к которому присоединяется HAS, и полипептидным скелетом EPO. Более предпочтительно углеводный фрагмент представляет собой терминальный фрагмент углеводной боковой цепи.
Следовательно, в предпочтительном варианте осуществления настоящего изобретения модифицированный HAS присоединяют (через линкер или нет, см. ниже) к углеводородным цепям, присоединенным к N- и/или О-сайтам гликозилирования EPO.
Однако в область настоящего изобретения включается также тот аспект, что EPO содержит другой(ие) углеводный(ые) фрагмент(ы), с которым(ими) подвергают конъюгированию модифицированный HAS. Методики присоединения углеводных фрагментов к полипептидам либо ферментного характера, либо путем генно-инженерной процедуры с последующей экспрессией в соответствующих клетках, известны специалистам в данной области (Berger, Greber, Mosbach, 1986, Galactosyltransferase-dependent sialylation of complex and endo-N-acetylglucosaminidase H-treated core N-glycans in vitro, FEBS Lett., 203(1), 64-8; Dittmar, Conradt, Hauser, Hofer, Lindenmaier, 1989, Advanced in Protein design; Bloecker, Collins, Schmidt, and, Schomburg eds., GBF-Monographs, 12, 231-246, VCH Publishers, Wienheim, New York, Cambridge).
В предпочтительном варианте осуществления способа по настоящему изобретению окисляют углеводный фрагмент с тем, чтобы он обладал способностью взаимодействовать с модифицированным HAS. Указанное окисление может быть проведено химическим или ферментативным способом.
Способы химического окисления углеводных фрагментов полипептидов известны в данной области техники и включают обработку перйодатом (Chamow et al., 1992, J. Biol. Chem., 267, 15916-15922).
При проведении химического окисления становится принципиально возможным окислить любой углеводный фрагмент, независимо от того, локализован он в концевом положении или нет. Однако при выборе мягких условий обработки (1 мМ перйодата, 0°С, тогда как жесткие условия включают: 10 мМ перйодата, 1 час при комнатной температуре) достигается возможность окислить предпочтительно терминальный углеводный фрагмент, например сиаловую кислоту или галактозу в боковой цепи углевода.
Альтернативно углеводный фрагмент может быть окислен ферментативным способом. Ферменты, применяемые для окисления отдельных углеводных фрагментов, известны в технике и включают, например, в случае галактозы, фермент галактозооксидазу.
В случае проведения окисления терминальных фрагментов галактозы возникает необходимость постепенно удалять терминальные сиаловые кислоты (частично или полностью), если EPO продуцируется в клетках, способных осуществлять реакцию присоединения сиаловых кислот к углеводным цепям, например, в клетках млекопитающих или в клетках, которые были генетически модифицированы, с целью придать им способность присоединять сиаловые кислоты к углеводным цепям. Химические или ферментативные способы удаления сиаловых кислот известны в данной области техники (Chaplin and Kennedy (eds.), 1986, Carbohydrate Analysis: a practical approach, в особенности Chapter 5 Montreuill, Glycoproteins, pages 175-177; IRL Press Practical approach series (ISBN 0-947946-44-3)).
Однако в область настоящего изобретения включается также вариант, при котором углеводный фрагмент, к которому должен быть присоединен модифицированный HAS, присоединяется к EPO в ходе стадии (а). В таком случае желательно присоединять галактозу, что может быть достигнуто с использованием галактозилтрансферазы. Такие способы известны в данной области техники (Berger, Greber, Mosbach, 1986, Galactosyltransferase-dependent sialylation of complex and endo-N-acetylglucosaminidase H-treated core N-glycans in vitro, FEBS Lett., 203(1), 64-8).
В наиболее предпочтительном варианте осуществления настоящего изобретения EPO на стадии (а) модифицируют путем окисления по меньшей мере одной терминальной сахаридной единицы, предпочтительно галактозы, в одной или большем числе боковых углеводных цепей EPO, предпочтительно после частичного или полного (ферментативного и/или химического) удаления терминальной сиаловой кислоты, при необходимости (см. приведенное выше описание).
Соответственно и предпочтительно модифицированный HAS подвергают конъюгированию с окисленной терминальной сахаридной единицей углеводной цепи, предпочтительно с галактозой.
Кроме того, модифицированный HAS может быть предпочтительно конъюгирован с терминальной сиаловой кислотой, которую предпочтительно окисляют на стадии (а) по способу настоящего изобретения.
В другим предпочтительном варианте осуществления настоящего изобретения (см. пункт (3) выше) EPO включает по меньшей мере одну SH-группу.
В соответствии с предпочтительным вариантом указанная SH-группа может быть присоединена к предпочтительно окисленному углеводному фрагменту, например, путем использования производного гидроксиламина, например, гидрохлорида 2-(аминоокси)этилмеркаптана (Bauer L. et al., 1965, J. Org. Chem., 30, 949) или с использованием гидразидного производного, например гидразида тиогликолевой кислоты (Whitesides et al., 1977, J. Org. Chem., 42, 332). Способы осуществления реакции конъюгирования указанных молекул с окисленным углеводным фрагментом EPO могут быть аналогичны способам, описанным в процедурах примеров 8 и 9.
В соответствии с еще одним предпочтительным вариантом осуществления настоящего изобретения свободная SH-группа представляет часть природного цистеина или добавленного цистеина.
EPO млекопитающих содержит несколько цистеинов, которые в норме образуют дисульфидные связи. Однако при замене по меньшей мере одного цистеина другой аминокислотой (например, рекомбинантными методами) становится возможным получать EPO, в котором по меньшей мере один из природных цистеинов включает свободную SH-группу. Способы замены аминокислот известны в данной области техники (Elliot, Lorenzini, Chang, Barzilay, Delorme, 1997, Mapping of active site of recombinant human erithropoietin, Blood, 89(2), 493-502; Boissel, Lee, Presnell, Cohen, Bunn, 1993, Erithropoietin structure-function relationship. Mutant proteins that test a model of tertiary structure, J. Biol. Chem., 268(21), 15983-93)).
Предпочтительно EPO имеет аминокислотную последовательность человеческого EPO и природные цистеины представляют собой цистеин 29 и/или 33.
Соответственно в предпочтительном варианте осуществления настоящего изобретения цистеин 33 заменяют другой аминокислотой и на стадии с) модифицированный HAS подвергают конъюгированию с цистеином 29.
В другом предпочтительном варианте осуществления настоящего изобретения цистеин 29 заменяют другой аминокислотой и на стадии с) модифицированный HAS подвергают конъюгированию с цистеином 33.
В контексте настоящего описания термин "добавленные цистеины" означает, что полипептид, предпочтительно EPO, включает остаток цистеина, который не присутствует в полипептиде дикого типа. Указанного состояния можно достигнуть при добавлении (например, рекомбинантными методами) остатка цистеина на N- или С-конце путем замены (например, рекомбинантными способами) природной аминокислоты цистеином. Соответствующие способы известны специалистам в данной области техники (см. приведенное выше описание).
Предпочтительно добавленный цистеин вводят путем замены природной аминокислоты цистеином.
В предпочтительном варианте осуществления настоящего изобретения EPO представляет собой человеческий EPO и замененный аминокислотный остаток представляет собой серин 126.
Предпочтительно модифицированный HAS подвергают конъюгированию на стадии с) с добавленным цистеином.
На стадии b) в способе настоящего изобретения получают модифицированный HAS, который способен взаимодействовать с EPO со стадии а).
В контексте настоящего описания HAS может быть предпочтительно модифицирован на своем восстановленном конце. Указанный вариант имеет то преимущество, что химическая реакция может быть легко контролироваться и специалист со средним уровнем знаний в данной области может иметь гарантии того, что группа HAS подверглась модификации в ходе реакции. Поскольку в HAS вводится лишь одна группа, то может быть предотвращено сшивание между различными молекулами EPO за счет многофункциональных молекул HAS и в ходе других побочных реакций.
Соответственно модифицированный HAS может обладать способностью взаимодействовать:
(1) по меньшей мере с одной группой, присоединенной непосредственно или через линкерную молекулу к сульфидным группам или углеводным фрагментам EPO,
(2) по меньшей мере с одним углеводным фрагментом, который предпочтительно окисляют и/или
(3) по меньшей мере с одной свободной SH-группой.
В том, что касается указанного выше пункта (1), то модификация HAS будет зависеть от группы, присоединяемой к EPO. Механизм указанной реакции известен в данной области техники. Такого рода реакция проиллюстрирована в примере 4, 2.1.
В том, что касается приведенных выше пунктов (2) и (3), известно в данной области техники несколько способов модификации HAS. Принципиальная основа данного метода заключается в том, что либо модифицируют реакционноспособную группу HAS с тем, чтобы придать ей способность взаимодействовать с углеводным фрагментом или SH-группой, либо проводят конъюгирование линкерной молекулы с HAS, который содержит реактивную группу, способную взаимодействовать с углеводным фрагментом или с SH-группой.
В случае пункта (2) модифицированный HAS может обладать способностью взаимодействовать с окисленными углеводными фрагментами, предпочтительно с терминальным сахаридным остатком, более предпочтительно с галактозой или с терминальной сиаловой кислотой.
Известно несколько способов модификации HAS с тем, чтобы придать ему способность взаимодействовать с окисленным, предпочтительно терминальным, сахаридным остатком. Как отмечалось выше, указанная модификация может быть введена региоселективно на восстановленном конце HES-цепи. В данном случае на первой стадии окисляют альдегидную группу до лактона. Такие модификации включают, без ограничения, добавление гидрозидной, амино (а также гидроксиламино), семикарбазидной или тиоловой функций к HAS, либо непосредственно, либо через линкер. Указанные методики поясняются более детально в примерах 2-4. Существенно также, что механизм таких реакций известен в технике (см., например, DE 19628705 A1; Hpoe et al., 1981, Carbohydrate Res., 91, 39; Fissekis et al., 1960, Journal of Medicinal and Pharmaceutical Chemistry, 2, 47; Frie et al., 1998, дипломная работа, Fachhochschule Hamburg, DE).
В контексте данного описания добавление гидразидной или гидроксиламинофункции является предпочтительным. В таком случае при проведении реакции на стадии с) по способу настоящего изобретения предпочтительно в условиях pH 5,5, гарантируется, что модифицированный HAS будет селективно взаимодействовать с окисленным углеводным фрагментом EPO без меж- или внутримолекулярного сшивания EPO путем образования имина на основе боковых цепей лизина и окисленного сахаридного остатка.
В том, что касается пункта (3), известно также несколько способов модификации HAS, в результате чего он приобретает способность взаимодействовать со свободной SH-группой. Предпочтительно указанную модификацию вводят региоселективно на восстановленном конце HES-цепи. Указанные способы включают, без ограничения, добавление малеимидной, дисульфидной или галогенацетамидной функции к HAS. Указанные методики поясняются более подробно в примерах 2-4.
Другие детали указанных методик описаны в литературе (Chamov et al., 1992, J. Biol Chem., 267, 15916; Thorpe et al., 1984, Eur. J. Biochem., 140, 62; Greenfield et al., 1990, Cancer Research, 50, 6600), а также из литературы, цитированной в примерах 2, 1.3.
Другие возможные функции перечислены в таблице 1, в которой дается систематический обзор возможных линкерных молекул. Более того, следует отметить, что механизм указанных реакций известен в данной области техники.
Несколько линкерных молекул, которые могут быть полезны в контексте настоящего описания, известны в данной области техники или коммерчески доступны (например, от компании Perbio Science Deutschland GmbH, Bonn, Germany). Примеры приведены в таблице 2.
На стадии с) способа настоящего изобретения EPO со стадии а) подвергают реакции с HAS со стадии b), в результате чего получают HAS-EPO, включающий одну или несколько молекул HAS, где HAS подвергают конъюгированию с EPO через углеводный фрагмент или через тиоэфир.
В принципе, детали способов, используемых для проведения реакции EPO с модифицированным HAS, зависят от индивидуальной модификации EPO и/или HAS и они известны в данной области техники (см., например, Rose, 1994, J. Am. Chem. Soc., 116, 30, O'Shannessay and Wichek, 1990, Analytical Biochemistry, 191, 1; Thorpe et al., 1984, Eur. J. Biochem., 140, 63; Chamov et al., 1992, J. Biol Chem., 267, 15916).
Репрезентативные способы настоящего изобретения подробно описаны в примерах 2-4, особенно в примере 4.
Стадия с) может быть осуществлена в реакционной среде, включающей по меньшей мере 10 мас.% воды.
Реакционная среда в данном предпочтительном варианте осуществления способа настоящего изобретения включает по меньшей мере 10 мас.% воды, предпочтительно, по меньшей мере 50%, более предпочтительно, по меньшей мере 80%, например, 90% или до 100%. Уровень содержания органических растворителей вычисляется соответствующим образом. Следовательно, реакция происходит в водной фазе. Предпочтительной реакционной средой является вода.
Одним преимуществом данного варианта осуществления способа настоящего изобретения является то, что необязательно использовать токсикологически критические растворители, так что при этом не возникает необходимости удалять такие растворители по окончании процесса производства с тем, чтобы избежать загрязнения данным растворителем. Кроме того, нет необходимости в проведении дополнительных контрольных тестов качества в том, что касается содержания остаточных токсикологически критических растворителей. Предпочтительно использовать в качестве органических растворителей токсикологически некритические растворители, такие как этанол или пропиленгликоль.
Другое преимущество осуществления способа настоящего изобретения заключается в том, что удается избежать необратимых или обратимых структурных изменений, которые индуцируются органическими растворителями. Следовательно, получаемые по способу настоящего изобретения полипептиды отличаются от полипептидов, получаемых в органических растворителей, таких как ДМСО.
Кроме того, было неожиданно обнаружено, что конъюгирование HAS с лекарственными средствами в водном растворе сводит к минимуму или позволяет полностью избежать побочных реакций. Следовательно, данный вариант осуществления способа настоящего изобретения ведет к получению улучшенных продуктов с точки зрения их большей чистоты.
В контексте настоящего изобретения термин "гидроксиалкилкрахмал" используется для обозначения производных крахмала, которые включают замещение гидроксиалкильными группами. В данном контексте алькильная группа может быть замещена. Предпочтительно гидроксиалкил содержит 2-10 атомов углерода, более предпочтительно 2-4 атома углерода. В этой связи "гидроксиалкилкрахмал" предпочтительно включает гидроксиэтилкрахмал, гидроксипропилкрахмал и гидроксибутилкрахмал, где гидроксиэтилкрахмал и гидроксипропилкрахмал являются предпочтительными.
Гидроксиалкильная группа(ы) HAS содержит по меньшей мере одну OH-группу.
Гидроксиэтилкрахмал (HES) является наиболее предпочтительным для всех вариантов осуществления способа настоящего изобретения.
Выражение "гидроксиалкилкрахмал" также включает производные, в которых алкильная группа является моно- или полизамещенной. В данном контексте предпочтительно, чтобы алкильная группа была замещена атомом галогена, в особенности фтором, или арильной группой, при условии что HAS остается водорастворимым. Кроме того, терминальная гидроксигруппа в гидроксиалкиле может быть этерифицирована с получением сложного или простого эфира. Дополнительно следует указать, что алкильная группа гидроксиалкилкрахмала может быть линейной или разветвленной.
Кроме того, вместо алкила могут использоваться также линейные или разветвленные замещенные или незамещенные алкиленовые группы.
В контексте настоящего описания гидроксиэтилкрахмал может иметь среднюю молекулярную массу (средняя масса) 1-300 кДа, где средняя молекулярная масса 5-100 кДа является более предпочтительной. Гидроксиэтилкрахмал может также демонстрировать степень молярного замещения от 0,1 до 0,8 и коэффициент соотношения для С2:С6-замещения в диапазоне от 2 до 20 для гидроксиэтильных групп.
HAS-EPO, получаемый по способу настоящего изобретения, может быть очищен и охарактеризован следующим образом:
Выделение HAS-EPO может быть проведено с использованием известных процедур очистки природных и рекомбинантных EPO (например, путем вытеснительной хроматографии, ионообменной хроматографии, RP-ВЭЖХ, хроматографии на гидроксиапатите, хроматографии, основанной на гидрофобном взаимодействии, с использованием процедуры, описанной в примере 20.8, или их сочетаний).
Ковалентное присоединение HAS к EPO-полипептиду может быть подтверждено путем анализа углеводного состава после гидролиза модифицированного белка (соотношение гидроксиэтилглюкозы и маннозы, присутствующих в трех сайтах N-гликозилирования в EPO).
Выявление модификации HAS по N-присоединенным олигосахаридам EPO может быть осуществлено при удалении HAS-модифицированных N-гликанов с последующей оценкой наличия предполагаемого сдвига к большей подвижности при проведении электрофореза в SDS-PAGE +/- по результатам вестерн-блоттинга.
HAS-модификация EPO по цистеиновым остаткам может быть продемонстрирована по неспособности выявить после протеолиза соответствующий Cys-пептид при проведении анализа методом RP-ВЭЖХ и MALDI/TOF-MS фрагментов, полученных после протеолиза HAS-модифицированного продукта (Zhou et al., 1998, Application of capillary electrophoresis, liquid chromatography, electrospray-mass spectrometry and matrix-assisted laserdesorption/ionozation-time of flight-mass spectrometry to the characterization of recombinant human erythropoietin. Electrophoresis, 19(13), 2348-55). Выделение HAS-содержащей фракции после протеолитического расщепления Cys-модифицированного EPO позволяет провести анализ, подтверждающий наличие искомого продукта в данной фракции соответствующего пептида, в рамках традиционного метода определения аминокислотного состава.
Все описанные выше варианты осуществления настоящего изобретения в аспекте HAS-EPO, которые относятся к свойствам EPO или HAS, применимы также к способу получения конъюгата HAS-EPO по настоящему изобретению.
Настоящее изобретение также относится к HAS-EPO, получаемому по способу настоящего изобретения. Предпочтительно указанный HAS-EPO имеет свойства, определенные для описанного выше HAS-EPO по настоящему изобретению.
Настоящее изобретение также относится к HAS-EPO по настоящему изобретению для применения в способе лечения человека или животного.
Кроме того, настоящее изобретение относится к фармацевтической композиции, включающей HAS-EPO по настоящему изобретению. В предпочтительном варианте фармацевтическая композиция включает также по меньшей мере один фармацевтически приемлемый разбавитель, адъювант и/или носитель, используемые в терапии эритропоэтином.
Фармацевтическая композиция предпочтительно применяется для лечения анемических нарушений или нарушений кроветворной функции или заболеваний, связанных с ними.
Термин "терапевтически эффективное количество" в контексте настоящего описания относится к количеству, которое обеспечивает терапевтический эффект в данных условиях и в данном режиме введения. Введение изоформ эритропоэтина предпочтительно проводится парентеральными способами. Конкретный выбираемый способ зависит от состояния пациента, которого предстоит лечить. Введение изоформ эритропоэтина предпочтительно проводят в виде композиции, содержащей соответствующий носитель, такой как человеческий сывороточный альбумин, соответствующий разбавитель, такой как забуференный солевой раствор, и/или соответствующий адъювант. Требуемая дозировка будет представлять собой количества, достаточные для повышения гематокрита у пациента, и будет варьировать в зависимости от тяжести состояния, подлежащего лечению, способа введения и т.п.
Объектом лечения фармацевтической композицией по настоящему изобретению предпочтительно является повышение уровня гемоглобина более чем 6,8 ммоль/л в крови. Для этой цели фармацевтическая композиция может вводиться таким способом, что содержание гемоглобина повышается от 0,6 ммоль/л до 1,6 ммоль/л в неделю. Если содержание гемоглобина превышает уровень 8,7 ммоль/л, то предпочтительно лечение следует прервать до снижения уровня гемоглобина ниже уровня 8,1 ммоль/л.
Композиция по настоящему изобретению предпочтительно используется в составе препарата, пригодного для подкожной или внутривенной парентеральной инъекции. Пригодные для этой цели наполнители и носители включают, например, дигидрофосфат натрия, гидрофосфат динатрия, хлорат натрия, полисорбат 80, HSA и воду для инъекций. Указанная композиция может вводиться три раза в неделю, предпочтительно два раза в неделю, более предпочтительно один раз в неделю и наиболее предпочтительно раз в две недели.
Предпочтительно фармацевтическую композицию вводят в количестве 0,01-10 мкг/кг массы тела пациента, более предпочтительно 0,1-5 мкг/кг, 0,1-1 мкг/кг или 0,2-0,9 мкг/кг, наиболее предпочтительно 0,3-0,7 мкг/кг и, еще наиболее предпочтительно 0,4-0,6 мкг/кг массы тела.
В целом, в составе дозы вводят предпочтительно от 10 мкг до 200 мкг, предпочтительно от 15 мкг до 100 мкг.
Настоящее изобретение также относится к применению HAS-EPO по настоящему изобретению при получении лекарственного средства, применяемого для лечения анемических нарушений или нарушений кроветворной функции или связанных с ними заболеваний.
В соответствии с другим аспектом настоящего изобретения имеющаяся проблема решается путем использования конъюгата гидроксиалкилкрахмал (HAS)-полипептид (HAS-полипептид), включающего одну или несколько молекул HAS, где каждую молекулу HAS подвергают конъюгированию с полипептидом через
а) углеводный фрагмент или
b) тиоэфир.
HAS-полипептид по настоящему изобретению обладает преимуществом, заключающимся в том, что он демонстрирует улучшенную биологическую стабильность в сравнении с полипептидом до конъюгирования. Указанное свойство связано прежде всего с тем, что HAS-полипептид в меньшей степени или вообще не распознается системами удаления в печени или почках и, в этой связи дольше остается в системе кровообращения. Кроме того, поскольку HAS присоединяется в сайт-специфическом режиме, минимизируется риск разрушения биологической активности полипептида in vivo в ходе конъюгирования HAS с полипептидом.
HAS-полипептид по настоящему изобретению содержит в основном два компонента, а именно: полипептид и присоединенный к нему гидроксиалкилкрахмал (HAS).
Полипептид может быть любого происхождения: из организма человека или животного. В предпочтительном варианте полипептид имеет в качестве источника человеческий организм.
Полипептид может представлять собой цитокин, в особенности эритропоэтин, антитромбин (АТ), такой как АТ III, интерлейкин, в особенности интерлейкин-2, IFN-бета, IFN-альфа, G-CSF, CSF, интерлейкин-6 и терапевтические антитела.
В соответствии с предпочтительным вариантом осуществления настоящего изобретения полипептид представляет собой антитромбин (АТ), предпочтительно АТ III (Levy JH, Weisinger A, Ziomek CA, Echelard Y, Recombinant Antithrombin: Production and Role in Cardiovascular Disorder, Seminars in Thrombosis and Hemostasis 27, 4 (2001) 405-416; Edmunds T, Van Patten SM, Pollock J, Hanson E, Bernasconi R, Higgins E, Manavalan P, Ziomek C, Meade H, McPherson J, Cole ES, Transgenically Produced Human Antithrombin: Structural and Functional Comparison to Human Plasma-Derived Antithrombin, Blood 91, 13 (1998) 4661-4671; Minnema MC, Chang ACK, Jansen PM, Lubbers YTP, Pratt BM, Whittaker BG, Taylor FB, Hack CE, Friedman B, Recombinant human antithrombin III improves survival and attenuates inflammatory responses in baboons lethally challenged with Escherichia coli; Blood, 95, 4 (2000) 1117-1123; Van Patten SM, Hanson EH, Bernasconi R, Zhang K, Manavaln P, Cole ES, McPherson JM, Edmunds T, Oxidation of Methionine Residues in Antithrombin, J Biol. Chemistry 274, 15 (1999) 10268-10276).
В соответствии с другим предпочтительным вариантом полипептид представляет собой человеческий IFN-бета, в частности IFN-бета 1а (например, Avonex®, REBIF®) и IFN-бета 1b (например, BETASERON®).
Другой предпочтительный полипептид представляет человеческий G-CSF (гранулоцитарный колониестимулирующий фактор). (См., например, Nagata et al., The chromosomal gene structure and two mRNAs for human granulocyte colony-stimulating factor, EMBO J. 5: 575-581, 1986; Souza et al., Recombinant human granulocyte colony-stimulating factor: effects in normal and leukemic myeloid cells, Science 232 (1986) 61-65; and Herman et al., Characterization, formulation, and stability of Neupogen® (Filgrastim), a recombinant human granulocyte colony-stimulating factor, in: Formulation, characterization, and stability of protein drugs, Rodney Pearlman and Y. John Wang, eds., Plenum Press, New York, 1996, 303-328).
В том, что касается эритропоэтина, все указанные выше варианты также применимы и к нему.
Предпочтительно полипептид получают рекомбинантным способом. Указанный способ включает продукцию в эукариотических или прокариотических клетках, предпочтительно в клетках млекопитающего, насекомых, дрожжей, бактерий или в клетках любого другого типа, который удобен для получения полипептида рекомбинантными методами. Кроме того, полипептид может экспрессироваться в трансгенных животных (например, в жидкостях организма, таких как молоко, кровь и т.п.), в яйцах трансгенных птиц, особенно сельскохозяйственных птиц, предпочтительно кур, или в трансгенных растениях.
Получение полипептидов рекомбинантным способом известно в данной области техники. В основном такой способ включает трансфекцию клеток-хозяев соответствующим экспрессирующим вектором, культивирование клеток-хозяев в условиях, которые позволяют получать полипептид и затем выделять и очищать полипептид из клеток-хозяев. Подробная информация приведена в ряде работ (см., например, Kristal, Pankratz, Farber, Smart, 1986, Purification of human erythropoietin to homogeneity by a rapid five-step procedure, Blood, 67(1), 71-9; Quelle, Caslake, Burkert, Wojchowski, 1989, High-level expression and purification of a recombinant human erythropoietin produced using a baculovirus vector, Blood, 74(2), 652-7; EO 640619 B1 и EP 668351 B1).
Полипептид может включать одну или несколько углеводных боковых цепей, присоединенных к полипептиду путем N- или O-связанного гликозилирования, т.е. полипептид является гликозилированным. Обычно в том случае, когда полипептид получают в эукариотических клетках, указанный полипептид подвергается посттрансляционному гликозилированию. Впоследствии углеводные боковые цепи могут быть присоединены к полипептиду в процессе биосинтеза в клетках млекопитающих, в особенности человека, в клетках насекомых или дрожжей.
HAS может быть непосредственно подвергнут конъюгированию с полипептидом или, альтернативно, через линкерную молекулу. Природа линкерной молекулы зависит от способа присоединения HAS к полипептиду. Коммерчески доступно несколько линкеров (например, от Pierce, см. выше). Природа линкера и его функция описаны более подробно ниже, в разделе, относящемся к обсуждению и способу получения полипептида HES.
В соответствии с предпочтительным вариантом, относящимся к конъюгату HAS-полипептид по настоящему изобретению, HAS подвергают конъюгированию с полипептидом через углеводный фрагмент. Предпочтительно указанное относится к тому случаю, когда полипептид представляет собой антитромбин, предпочтительно АТ III.
В контексте настоящего описания термин "углеводный фрагмент" относится к гидроксиальдегидам или гидроксикетонам, а также к их химически модифицированным формам (см. Rompp Chemielexicon, Thieme Verlag Stuttgart, Germany, 9th edition 1990, Volume 9, pages 2281-2285 и цитированная в работе литература). Кроме того, указанный термин также относится к производным природных углеводных фрагментов типа глюкозы, галактозы, маннозы, сиаловой кислоты и т.п. Указанный термин также включает химически окисленные природные углеводные фрагменты, которые имеют открытую циклическую структуру.
Углеводный фрагмент может быть присоединен непосредственно к полипептидному скелету. Предпочтительно углеводный фрагмент представляет собой часть углеводной боковой цепи. В данном случае могут присутствовать другие углеводные фрагменты между углеводным фрагментом, к которому присоединяется HAS, и полипептидным скелетом. Более предпочтительно углеводный фрагмент представляет собой терминальный фрагмент боковой цепи углевода.
В более предпочтительном варианте изобретения HAS подвергают конъюгированию с галактозным остатком боковых цепей углевода, предпочтительно с терминальным галактозным остатком углеводной боковой цепи. Указанный галактозный остаток может быть открыт для конъюгирования за счет удаления терминальных сиаловых кислот с проведением впоследствии окисления (см. приведенное ниже описание).
В другом более предпочтительном варианте HAS подвергают конъюгированию с остатком сиаловой кислоты в углеводных боковых цепях, предпочтительно с терминальным остатком сиаловой кислоты в углеводной боковой цепи.
Кроме того, HAS может быть подвергнут конъюгированию с полипептидом через тиоэфир. Как будет подробно описано ниже, атом S может быть получен из SH-группы, присоединенной к полипептиду как в природном, так и неприродном варианте.
В предпочтительном варианте осуществления изобретения атом S может быть получен из SH-группы, которую вводят в окисленный углеводный фрагмент HES, предпочтительно окисленный углеводный фрагмент, который представляет собой часть углеводной боковой цепи полипептида (см. приведенное ниже описание).
Предпочтительно атом S в тиоэфире получают из природного цистеина или из добавленного цистеина.
В контексте настоящего описания термин "добавленные цистеины" означает, что полипептиды включают цистеиновый остаток, который не присутствует в полипептиде дикого типа.
В контексте данного аспекта изобретения цистеин может представлять собой дополнительную аминокислоту, добавляемую на N- или С-терминальном конце полипептида.
Кроме того, добавленный цистеин может быть внесен путем замены природной аминокислоты цистеином.
Второй компонент HAS-полипептида представляет собой HAS.
В контексте настоящего описания термин "гидроксиалкилкрахмал" используют для обозначения производных крахмала, которые содержат замещение гидроксиалкильными группами. В данном контексте алкильная группа может быть замещена. Предпочтительно гидроксиалкил содержит 2-10 атомов углерода, более предпочтительно 2-4 атома углерода. Термин "гидроксиалкилкрахмал" в этой связи предпочтительно включает гидроксиэтилкрахмал, гидроксипропилкрахмал и гидроксибутилкрахмал, где гидроксиэтилкрахмал и гидроксипропилкрахмал являются предпочтительными.
Гидроксиалкильная группа(ы) HAS содержит по меньшей мере одну OH-группу.
Термин "гидроксиалкилкрахмал" также включает производные, в которых алкильная группа представляет собой моно- или полизамещенную группу. В данном контексте предпочтительно, чтобы алкильная группа была замещена атомом галогена, в особенности фтором, или арильной группой, при условии что HAS остается водорастворимым. Кроме того, терминальная гидроксигруппа гидроксиалкила может быть этерифицирована с образованием простого или сложного эфира. Дополнительно следует отметить, что алкильная группа в гидроксиалкилкрахмале может быть линейной или разветвленной.
Кроме того, вместо алкила могут также использоваться линейные или разветвленные, замещенные или незамещенные алкеновые группы.
Гидроксиэтилкрахмал (HES) наиболее предпочтителен для всех вариантов осуществления настоящего изобретения.
В контексте настоящего описания гидроксиэтилкрахмал может иметь среднее значение молекулярной массы (средняя масса) от 1 до 300 кДа, где средняя молекулярная масса, равная 5-100 кДа, является более предпочтительной. Гидроксиэтилкрахмал может также демонстрировать степень молярного замещения от 0,1 до 0,8 и коэффициент С2:С6-замещения в диапазоне 2-20 относительно гидроксиэтильных групп.
HAS-полипептид может включать 1-12, предпочтительно 1-9, 1-6 или 1-3, наиболее предпочтительно 1-4 молекул HAS в расчете на одну полипептидную молекулу. Количество молекул HAS на одну полипептидную молекулу может быть определено путем количественного анализа углеводного состава с использованием GC-MS после гидролиза продукта и дериватизации полученных моносахаридов (см. Chaplin and Kennedy, 1986, Carbohydrate Analysis (eds.): a practical approach ed., Chapter 1, Monosaccharides page 1-36; Chapter 2, Oligosaccharides, page 37-53, Chapter 3, Neutral Polysaccharides, page 55-96, IRL Press Practical approach series (ISBN 0-947946-44-3).
Все раскрываемые ниже варианты осуществления способа по настоящему изобретению в аспекте получения HAS-полипептида, касающиеся свойств полипептида или HAS, применимы также к HAS-полипептиду по настоящему изобретению. Кроме того, все раскрытые выше варианты, описывающие HAS-EPO или препараты, которые относятся к пептидам в целом или к HAS, относятся также к HAS-полипептиду по настоящему изобретению.
Гидроксиэтилкрахмал представляет собой эфирное производное крахмала. Кроме указанных эфирных производных, в рамках настоящего изобретения могут использоваться также другие производные крахмала. Например, могут использоваться производные, которые включают этерифицированные гидроксигруппы. Указанные производные могут представлять собой, например, производные незамещенных моно- или дикарбоновых кислот, содержащих 2-12 атомов углерода, или их замещенные производные. Особенно полезными являются производные незамещенных монокарбоновых кислот, содержащих 2-6 атомов углерода, в особенности уксусной кислоты. В данном контексте предпочтительны ацетилкрахмал, бутилкрахмал или пропилкрахмал.
Кроме того, предпочтительны производные незамещенных дикарбоновых кислот, содержащих 2-6 атомов углерода.
В случае производных дикарбоновых кислот полезно, чтобы вторая карбоксильная группа дикарбоновой кислоты была также этерифицирована. Кроме того, пригодны также производные сложных моноалкиловых эфиров дикарбоновых кислот для применения в рамках настоящего изобретения.
В случае замещенных моно- или дикарбоновых кислот замещающие группы могут быть предпочтительны те же самые, что и указывалось выше для случая замещенных алкильных остатков.
Методики этерификации крахмала известны в данной области техники (см., например, Klemm D. et al.,.Comprehensive Cellulose Chemistry Vol. 2, 1998, Whiley-VCH, Weinheim, New York, в особенности chapter 4.4, Esterification of Cellulose (ISBN 3-527-29489-9).
В другом аспекте настоящее изобретение относится к способу получения конъюгата гидроксиалкилкрахмал (HAS)-полипептид (HAS-полипептид), включающему стадии:
а) получения полипептида, способного реагировать с модифицированным HAS,
b) получения модифицированного HAS, способного взаимодействовать с полипептидом со стадии а), и
c) проведения реакции полипептида со стадии а) с HAS со стадии b), в результате чего образуется HAS-полипептид, включающий одну или несколько молекул HAS, где HAS подвергают конъюгированию с полипептидом через
i) углеводный фрагмент или
ii) тиоэфир.
Способ по настоящему изобретению имеет преимущество, заключающееся в том, что образуется конъюгат HAS-полипептид, который демонстрирует высокую биологическую активность. Кроме того, способ по настоящему изобретению имеет то достоинство, что может быть получено эффективное производное полипептида со сниженной стоимостью, поскольку данный способ не включает дорогостоящие и длительные по времени стадии очистки, приводящие к низкому конечному выходу.
Соответственно на первой стадии способа по настоящему изобретению получают полипептид, который способен взаимодействовать с модифицированным HAS.
В контексте настоящего описания термин "получение" следует интерпретировать таким образом, что после соответствующей стадии становится доступной молекула (на стадии а) полипептида, на стадии b) HAS) с желательными свойствами.
В случае стадии а) данный термин включает выделение и очистку полипептида из природных источников, а также получение рекомбинантными методами в клетках-хозяевах или организме и, при необходимости, модификацию получаемого при этом полипептида.
В том, что касается полипептида, выполняющего функцию исходного материала по настоящему изобретению, к нему применимы те же требования, что и в отношении эритропоэтина, представляющего собой часть конъюгата HAS-полипептид по настоящему изобретению. В данном контексте предпочтительные варианты, раскрываемые выше, применимы также к способу по настоящему изобретению.
Предпочтительно полипептид получают рекомбинантным способом. Указанный способ включает продукцию в эукариотических или прокариотических клетках, предпочтительно в клетках млекопитающего, насекомых, дрожжей, бактерий или в клетках любого другого типа, которые удобны для рекомбинантного получения полипептида. Кроме того, полипептид может быть экспрессирован в трансгенных животных (например, в жидкостях организма, таких как молоко, кровь и т.п.), в яйцах трансгенных птиц, особенно сельскохозяйственных птиц, предпочтительно кур, или в трансгенных растениях.
Получение полипептида рекомбинантным способом известно в данной области техники. В основном указанный способ получения включает трансфекцию клеток-хозяев соответствующим экспрессирующим вектором, культивирование клеток-хозяев в условиях, позволяющих осуществлять продукцию полипептида, выделение и очистку полипептида из клеток-хозяев (Kristal, Pankratz, Farber, Smart, 1986, Purification of human erythropoietin to homogeneity by a rapid five-step procedure, Blood, 67(1), 71-9; Quelle, Caslake, Burkert, Wojchowski, 1989, High-level expression and purification of a recombinant human erythropoietin produced using a baculovirus vector, Blood, 74(2), 652-7; EO 640619 B1 и EP 668351 B1).
Полипептид может включать одну или несколько боковых цепей углевода, присоединяемых к полипептиду посредством N- или O-связанного гликозилирования, т.е. полипептид подвергается гликозилированию. Обычно, когда полипептид продуцируется в эукариотических клетках, указанный полипептид подвергается посттрансляционному гликозилированию. Впоследствии боковые цепи углевода могут быть присоединены к полипептиду в процессе образования в клетках млекопитающих, особенно человека, в клетках насекомых или дрожжей, которые могут представлять собой клетки трансгенного животного или растения (см. выше).
Указанные углеводные боковые цепи могут быть химически или энзиматически модифицированы после экспрессии в соответствующих клетках, например, путем удаления или добавления одного или большего числа углеводных фрагментов (см., например, Dittmar, Conradt, Hauser, Hofer, Lindenmaier, 1989, Advanced in Protein design; Bloecker, Collins, Schmidt, and Schomburg eds., GBF-Monographs, 12, 231-246, VCH Publishers, Wienheim, New York, Cambridge).
Другой аспект способа по настоящему изобретению относится к получению HAS-полипептида, включающего одну или несколько молекул HAS, где указанный HAS подвергают конъюгированию с полипептидом через углеводный фрагмент (i) или через тиоэфер (ii). Следовательно, полипептид, получаемый на стадии а), должен иметь свойства, позволяющие осуществлять конъюгирование через углеводный фрагмент и/или тиоэфир. В этой связи полипептид после стадии а) может предпочтительно содержать:
(1) по меньшей мере одну реакционноспособную группу, присоединенную либо непосредственно, либо через линкерную молекулу к сульфидным группам или углеводным фрагментам, которые способны взаимодействовать с HES или с модифицированным HES,
(2) по меньшей мере один углеводный фрагмент, с которым может быть конъюгирован модифицированный HAS и/или
(3) одну свободную SH-группу.
В том, что касается приведенного выше пункта (1), полипептид со стадии а) предпочтительно получают путем конъюгирования соответствующей линкерной молекулы с SH-группой(ами) или углеводными фрагментами полипептида. Такого рода модифицированный полипептид проиллюстрирован в примере 4, 2.1. Очень важно иметь гарантии того, что добавление линкерной молекулы не разрушает полипептид. Однако это известно специалистам в данной области техники.
В том, что касается указанного выше пункта 2, то в предпочтительном варианте осуществления настоящего изобретения модифицированный HAS подвергают конъюгированию с полипептидом через углеводный фрагмент.
Углеводный фрагмент может быть присоединен непосредственно к полипептидному скелету. Предпочтительно углеводный фрагмент представляет собой часть углеводной боковой цепи. В данном случае могут присутствовать другие углеводные фрагменты между углеводным фрагментом, к которому присоединяется HAS, и полипептидным скелетом. Более предпочтительно углеводный фрагмент представляет собой терминальный фрагмент углеводной боковой цепи.
Следовательно, в предпочтительном варианте осуществления настоящего изобретения модифицированный HAS присоединяют (через линкер или иным способом, см. ниже) к углеводным цепям, присоединенным к N- и/или О-сайтам гликозилирования полипептида.
Однако в область настоящего изобретения включается также тот факт, что полипептид содержит другой(ие) углеводный(ые) фрагмент(ы), с которым(ими) подвергают конъюгированию модифицированный HAS. Методики присоединения углеводных фрагментов к полипептидам либо ферментативным способом, либо с помощью генноинженерной процедуры, с последующей экспрессией в соответствующих клетках, известны в данной области техники (Berger, Greber, Mosbach, 1986, Galactosyltransferase-dependent sialylation of complex and endo-N-acetylglucosaminidase H-treated core N-glycans in vitro, FEBS Lett., 203(1), 64-8; Dittmar, Conradt, Hauser, Hofer, Lindenmaier, 1989, Advanced in Protein design; Bloecker, Collins, Schmidt, and, Schomburg eds., GBF-Monographs, 12, 231-246, VCH Publishers, Weinheim, New York, Cambridge).
В предпочтительном варианте осуществления способа по настоящему изобретению окисляют углеводный фрагмент с тем, чтобы он стал способным к взаимодействию с модифицированным HAS. Указанное окисление может быть проведено химическим или энзиматическим способом.
Способы химического окисления углеводных фрагментов полипептидов известны в данной области техники и включают обработку перйодатом (Chamow et al., 1992, J. Biol. Chem., 267, 15916-15922).
Химическое окисление дает принципиальную возможность окислить любой углеводный фрагмент независимо от того, где он локализован: в терминальном положении или нет. Однако при выборе мягких условий реакции (1 мМ перйодата, 0°С, тогда как жесткие условия включают: 10 мМ перйодата, 1 час при комнатной температуре) достигается возможность окислить предпочтительно терминальный углеводный фрагмент, например, сиаловую кислоту или галактозу в боковой цепи углевода.
Альтернативно углеводный фрагмент может быть окислен ферментативным способом. Ферменты, применяемые для окисления отдельных углеводных фрагментов, известны в технике, например, в случае галактозы таким ферментом является галактозооксидаза.
В случае окисления терминальных фрагментов галактоз возникает необходимость постепенно удалять терминальные сиаловые кислоты (частично или полностью), если полипептид продуцируется в клетках, способных осуществлять реакцию присоединения сиаловых кислот к углеводным цепям, например, в клетках млекопитающих или в клетках, которые были генетически модифицированы, с тем чтобы придать им способность присоединять сиаловые кислоты к углеводным цепям. Химические или ферментативные способы удаления сиаловых кислот известны в данной области техники (Chaplin and Kennedy (eds.), 1996, Carbohydrate Analysis: a practical approach, в сообенности Chapter 5 Montreuill, Glycoproteins, pages 175-177; IRL Press Practical approach series (ISBN 0-947946-44-3)).
Однако в область настоящего изобретения включается также вариант, в соответствии с которым углеводный фрагмент, к которому должен быть присоединен модифицированный HAS, присоединяется к полипептиду на стадии (а). В данном случае желательно присоединять галактозу, что может быть достигнуто с использованием галактозилтрансферазы. Такие способы известны в данной области техники (Berger, Greber, Mosbach, 1986, Galactosyltransferase-dependent sialylation of complex and endo-N-acetylglucosaminidase H-treated core N-glycans in vitro, FEBS Lett., 203(1), 64-8).
В наиболее предпочтительном варианте осуществления настоящего изобретения полипептид на стадии а) модифицируют путем окисления по меньшей мере одной сахаридной единицы, предпочтительно галактозы, в одной или большем числе углеводных боковых цепей полипептида, предпочтительно после частичного или полного (ферментативного и/или химического) удаления терминальной сиаловой кислоты, при необходимости (см. приведенное выше описание).
Впоследствии предпочтительно модифицированный HAS подвергают конъюгированию с окисленной терминальной сахаридной единицей углеводной цепи, предпочтительно с галактозой.
В другим предпочтительном варианте осуществления настоящего изобретения (см. пункт (3) выше) полипептид включает по меньшей мере одну свободную SH-группу.
В соответствии с предпочтительным вариантом свободная SH-группа представляет собой часть природного цистеина или добавленного цистеина.
Способы замены аминокислот известны в данной области техники (Elliot, Lorerzini, Chang, Barzilay, Delorme, 1997, Mapping of the active site of recombinant human erithropoietin, Blood, 89(2), 493-502; Boissel, Lee, Presnell, Cohen, Bunn, 1993, Erithropoietin structure-function relationship. Mutant proteins that test a model of tertiary structure, J. Biol. Chem., 268(21), 15983-93)).
В контексте настоящего описания термин "добавленные цистеины" означает, что полипептид включает остаток цистеина, который не присутствует в полипептиде дикого типа. Указанную форму можно достигнуть при добавлении (например, рекомбинантными методами) остатка цистеина на N- или С-конце полипептида или путем замены (например, рекомбинантными методами) природной аминокислоты цистеином. Соответствующие способы известны специалистам в данной области техники (см. приведенное выше описание).
Предпочтительно добавленный цистеин вводят путем замены природной аминокислоты цистеином.
Предпочтительно модифицированный HAS подвергают конъюгированию на стадии с) с добавленным цистеином.
На стадии b) в способе настоящего изобретения получают модифицированный HAS, который способен взаимодействовать с полипептидом со стадии а).
В данном контексте настоящего описания HAS может быть предпочтительно модифицирован на своем восстановленном конце. Указанный вариант имеет то преимущество, что химическая реакция может легко контролироваться, и специалист со средним уровнем знаний в данной области может быть уверен, что группа HAS модифицируется в ходе реакции. Поскольку в HAS вводится лишь одна группа, то может быть предотвращено сшивание между различными полипептидными молекулами под действием многофункциональных молекул HAS и в ходе других побочных реакций.
Соответственно модифицированный HAS может обладать способностью взаимодействовать:
(1) по меньшей мере с одной группой, присоединенной непосредственно или через линкерную молекулу к сульфидным группам или углеводным фрагментам полипептида,
(2) по меньшей мере с одним углеводным фрагментом, который предпочтительно окислен, и/или
(3) по меньшей мере с одной свободной SH-группой.
В том, что касается указанного выше пункта (1), то модификация HAS будет зависеть от группы, присоединяемой к полипептиду. Механизм указанной реакции известен в данной области техники. Соответствующая иллюстрация дана в примере 4, 2.1.
В том, что касается указанных выше пунктов (2) и (3), в данной области техники известно несколько способов модификации HAS. Принципиальная основа данного метода заключается в том, что подвергают модификации любую реактивную группу HAS с тем, чтобы придать ей способность взаимодействовать с углеводным фрагментом или с SH-группой, либо проводят конъюгирование линкерной молекулы с HAS, который содержит реактивную группу, способную взаимодействовать с углеводным фрагментом или с SH-группой.
В том, что касается пункта (2), следует отметить, что модифицированный HAS может обладать способностью взаимодействовать с окисленными углеводными фрагментами, предпочтительно с терминальным сахаридным остатком, более предпочтительно с галактозой или терминальной сиаловой кислотой.
Известно несколько способов модификации HAS с целью придать ему способность взаимодействовать с окисленным, предпочтительно терминальным сахаридным остатком. Как указывалось выше, указанная модификация может быть введена региоселективно на восстановленном конце HES-цепи. В данном случае на первой стадии окисляют альдегидную группу до лактона. Такие модификации включают, без ограничения, добавление гидразидной, амино (а также гидроксиламино), семикарбазидной или тиоловой функций к HAS, либо непосредственно, либо через линкер. Указанные методики поясняются более детально в примерах 2-4. Более того, в данной области техники известен механизм таких реакций сам по себе (см., например, DE 19628705 A1; Hpoe et al., 1981, Carbohydrate Res., 91, 39; Fissekis et al., 1960, Journal of Medicinal and Pharmaceutical Chemistry, 2, 47; Frie et al., 1998, дипломная работа, Fachhochschule Hamburg, DE).
В контексте настоящего описания добавление гидразидной или гидроксиламинофункции является предпочтительным. В данном случае при предпочтительном варианте проведения реакции на стадии с) способа настоящего изобретения при pH 5,5 гарантируется, что модифицированный HAS будет взаимодействовать селективно с окисленным углеводным фрагментом полипептида без наличия меж- или внутримолекулярного сшивания полипептида за счет образования имина на основе боковых цепей лизина и окисленного сахаридного остатка.
В том, что касается пункта (3), известно также несколько способов модификации HAS, который приобретает способность взаимодействовать со свободной SH-группой. Предпочтительно указанную модификацию вводят региоселективно на восстановленном конце HES-цепи. Указанные способы включают, без ограничения, добавление малеимидной, дисульфидной или галогенацетамидной функции к HAS. Указанные методики поясняются более подробно в примерах 2-4.
Другие детали указанных методик описаны в литературе (Chamov et al., 1992, J. Biol Chem., 267, 15916; Thorpe et al., 1984, Eur. J. Biochem., 140, 63; Greenfield et al., 1990, Cancer Research, 50, 6600), а также из литературы, цитированной в примере 2, 1.3.
В таблице 1 перечислены другие возможные функции в рамках систематического обзора возможных линкерных молекул. Кроме того, в данной области техники известны механизмы данной реакции сами по себе.
Несколько линкерных молекул, которые могут быть полезны в контексте настоящего изобретения, известны в данной области или являются коммерчески доступными (например, от Pierce, доступной от компании Perbio Science Deutschland GmbH, Bonn, Germany).
На стадии с) способа по настоящему изобретению полипептид со стадии а) подвергают реакции с HAS со стадии b), в результате чего получают HAS-полипептид, включающий одну или несколько молекул HAS, где HAS подвергают конъюгированию с полипептидом через углеводный фрагмент или через тиоэфир.
В принципе, детали способов проведения реакции полипептида с модифицированным HAS зависят от конкретной модификации полипептида и/или HAS и известны в данной области (см., например, Rose, 1994, J. Am. Chem. Soc., 116, 30, O'Shannessay and Wichek, 1990, Analytical Biochemistry, 191, 1; Thorpe et al., 1984, Eur. J. Biochem., 140, 63; Chamov et al., 1992, J. Biol Chem., 267, 15916).
Детали осуществления способов по настоящему изобретению описаны в примерах 2-4, особенно в примере 4.
Стадия с) может быть проведена в реакционной среде, включающей по меньшей мере 10 мас.% воды.
Реакционная среда в данном предпочтительном варианте осуществления способа настоящего изобретения включает по меньшей мере 10 мас.% воды, предпочтительно, по меньшей мере 50%, более предпочтительно, по меньшей мере 80%, например, 90% или до 100%. Содержание органических растворителей вычисляется соответствующим образом. Следовательно, реакция происходит в водной фазе. Предпочтительной реакционной средой является вода.
Одним преимуществом данного варианта осуществления способа настоящего изобретения является то, что нет необходимости использовать токсикологически критические растворители, так что в этой связи не возникает потребности удалять такие растворители после завершения процесса производства с тем, чтобы избежать загрязнения данным растворителем. Кроме того, нет необходимости проводить дополнительные контрольные тесты качества относительно содержания остаточных токсикологически критических растворителей. Предпочтительно использовать в качестве органических растворителей токсикологически некритические растворители, такие как этанол или пропиленгликоль.
Другое преимущество осуществления способа настоящего изобретения заключается в том, что удается избежать необратимых или обратимых структурных изменений, которые индуцируются органическими растворителями. Следовательно, получаемые по способу данного изобретения полипептиды отличаются от полипептидов, получаемых в органических растворителях, таких как ДМСО.
Кроме того, было неожиданно обнаружено, что проведение реакции конъюгирования HAS с лекарственными средствами в водном растворе сводит к минимуму или позволяет избежать побочных реакций. Следовательно, данный вариант осуществления способа настоящего изобретения ведет к получению улучшенных продуктов с точки зрения их большей чистоты.
В контексте настоящего изобретения термин "гидроксиалкилкрахмал" используется для обозначения производных крахмала, которые содержат замещение гидроксиалкильными группами. В данном контексте алькильная группа может быть замещена. Предпочтительно гидроксиалкил содержит 2-10 атомов углерода, более предпочтительно 2-4 атома углерода. В этой связи "гидроксиалкилкрахмал" предпочтительно включает гидроксиэтилкрахмал, гидроксипропилкрахмал и гидроксибутилкрахмал, где гидроксиэтилкрахмал и гидроксипропилкрахмал являются предпочтительными.
Гидроксиалкильная группа(ы) HAS содержит по меньшей мере одну OH-группу.
Гидроксиэтилкрахмал (HES) является наиболее предпочтительным для всех вариантов осуществления способа по настоящему изобретению.
Выражение "гидроксиалкилкрахмал" также включает производные, в которых алкильная группа является моно- или полизамещенной. В данном контексте предпочтительно, чтобы алкильная группа была замещена атомом галогена, в особенности фтором, или арильной группой, при условии что HAS остается водорастворимым. Кроме того, терминальная гидроксигруппа в гидроксиалкиле может быть этерифицирована с получением сложного или простого эфира. Дополнительно следует отметить, что алкильная группа гидроксиалкилкрахмала может быть линейной или разветвленной.
Кроме того, вместо алкила могут использоваться также линейные или разветвленные, замещенные или незамещенные алкиленовые группы.
В контексте настоящего описания гидроксиэтилкрахмал может иметь среднее значение молекулярной массы (средняя масса) 1-300 кДа, где средняя молекулярная масса 5-100 кДа является более предпочтительной. Гидроксиэтилкрахмал может также демонстрировать степень молярного замещения от 0,1 до 0,8 и коэффициент соотношения для С2:С6-замещения в диапазоне от 2 до 20 относительно гидроксиэтильных групп.
HAS-полипептид, получаемый по способу настоящего изобретения, может быть выделен, очищен и охарактеризован следующим образом.
Выделение HAS-полипептида может быть проведено с использованием известных процедур очистки природных и рекомбинантных полипептидов (например, путем вытеснительной хроматографии, ионообменной хроматографии, RP-ВЭЖХ, хроматографии на гидроксиапатите, хроматографии, основанной на гидрофобном взаимодействии, а также с использованием процедуры, описанной в примере 20.8, или их сочетаний).
Ковалентное присоединение HAS к полипептиду может быть подтверждено при проведении анализа углеводного состава после гидролиза модифицированного белка.
Подтверждение наличия модификации HAS по N-присоединенным олигосахаридам полипептида может быть осуществлено при удалении HAS-модифицированных N-гликанов с последующей оценкой наличия предполагаемого сдвига к большей подвижности при электрофорезе в SDS-PAGE +/- по результатам вестерн-блоттинга.
HAS-модификация полипептида по цистеиновым остаткам может быть продемонстрирована по неспособности выявить соответствующий Cys-пептид после протеолиза при проведении анализа методом RP-ВЭЖХ и MALDI/TOF-MS протеолитических фрагментов, полученных после протеолиза HAS-модифицированного продукта (Zhou et al., 1998, Application of capillary electrophoresis, liquid chromatography, electrospray-mass spectrometry and matrix-assisted laserdesorption/ionozation-time of flight-mass spectrometry to the characterization of recombinant human erythropoietin. Electrophoresis, 19(13), 2348-55). Выделение HAS-содержащей фракции после протеолитического расщепления Cys-модифицированного полипептида позволяет выявить наличие искомого продукта в данной фракции соответствующего пептида путем традиционного анализа аминокислотного состава.
Все описанные выше варианты осуществления настоящего изобретения в аспекте HAS-полипептида по настоящему изобретению, касающиеся свойств полипептида или HAS, применимы также к способу получения конъюгата HAS-полипептид по настоящему изобретению. Кроме того, все описанные выше варианты, касающиеся HAS-EPO или способов получения, относящихся к полипептид в целом или к HAS, применимы также к способу получения конъюгата HAS-полипептид по настоящему изобретению.
Настоящее изобретение также относится к HAS-полипептиду, получаемому по способу настоящего изобретения. Предпочтительно данный HAS-полипептид обладает свойствами, определенными для указанного выше HAS-полипептида по настоящему изобретению.
В соответствии с предпочтительным вариантом осуществления настоящего изобретения используемый HAS имеет следующую формулу (I):
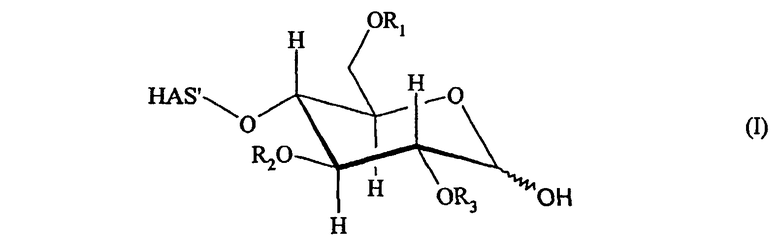
где R1, R2 и R3 обозначают независимо атом водорода или линейную или разветвленную гидроксиалкильную группу. Термин "гидроксиалкилкрахмал" в контексте настоящего описания не ограничивается соединениями, в которых терминальный углеводный фрагмент включает гидроксиалкильные группы R1, R2 и/или R3, как показано, например, для ясности в формуле (I), но также относится к соединениям, в которых по меньшей мере одна гидроксильная группа присутствует в любом месте или на терминальном углеводном фрагменте, и/или в оставшейся части молекулы. HAS' замещен гидроксиалкильной группой R1, R2 и R3. В данном контексте алкильная группа может быть линейной или разветвленной алкильной группой, которая может быть соответствующим образом замещена. Предпочтительно гидроксиалкильная группа содержит 1-10 атомов углерода, более предпочтительно 1-6 атомов углерода, более предпочтительно 1-4 атома углерода и, еще более предпочтительно 2-4 тома углерода. В этой связи термин "гидроксиалкил" предпочтительно включает гидроксиэтилкрахмал, гидроксипропилкрахмал и гидроксибутилкрахмал, где гидроксиэтилкрахмал и гидоксипропилкрахмал являются более предпочтительными, и особенно предпочтительным является гидроксиэтилкрахмал.
HAS и предпочтительно HES могут взаимодействовать со сшивающим соединением, которое реагирует с HAS, предпочтительно с HES, и с полипептидами, такими как указанные выше полипептиды.
Реакция между HAS и сшивающим соединением может происходить на восстановленном конце HAS или на восстановленном конце HAS при его окислении. В этой связи HAS может взаимодействовать, имея структуру формулы (I)
и/или в случае окисления восстановленного конца в соответствии с формулой (IIa)
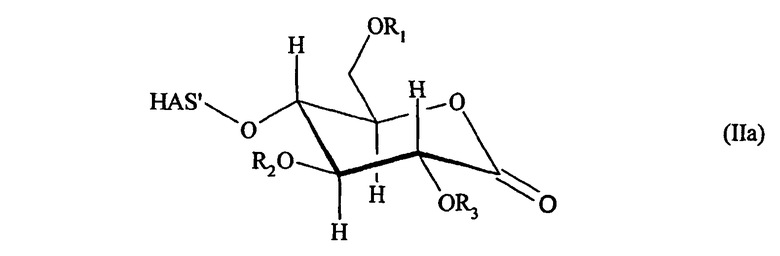
и/или в соответствии с формулой (IIb)
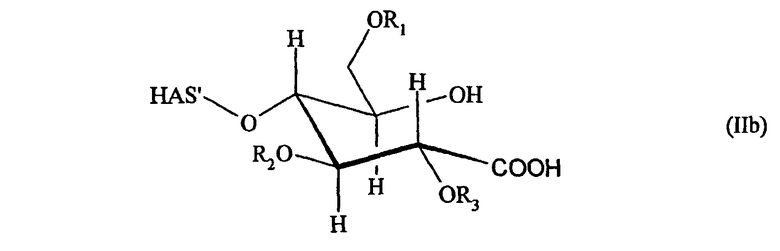
Если HAS формулы (I) взаимодействует со сшивающим соединением, то реакция предпочтительно проходит в водной среде. Если HAS формулы (IIa) и (IIb) взаимодействует со сшивающим соединением, то реакция предпочтительно проходит в неводной среде, такой как полярный апротонный растворитель, или в смеси растворителей, такой как ДМСО и/или в ДМФА.
Если конъюгат HAS-полипептид по настоящему изобретению получают путем реакции производного HAS, включающего HAS и сшивающее соединение, с окисленным углеводным фрагментом полипептида, то сшивающее соединение предпочтительно представляет собой соединение
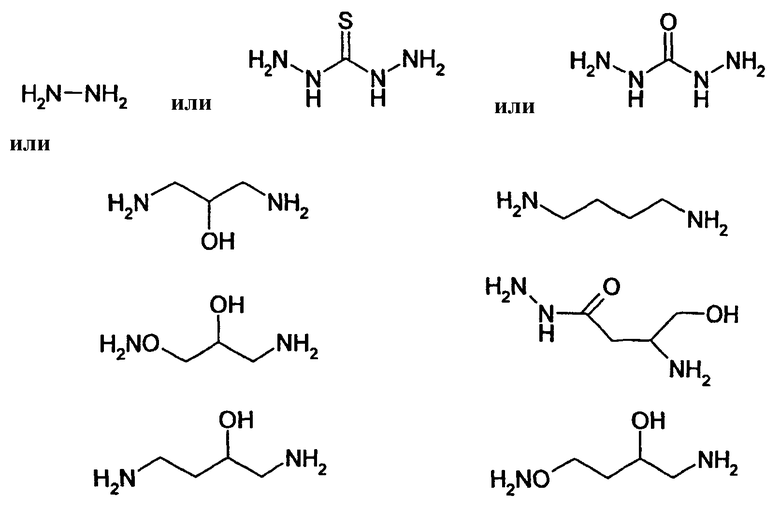
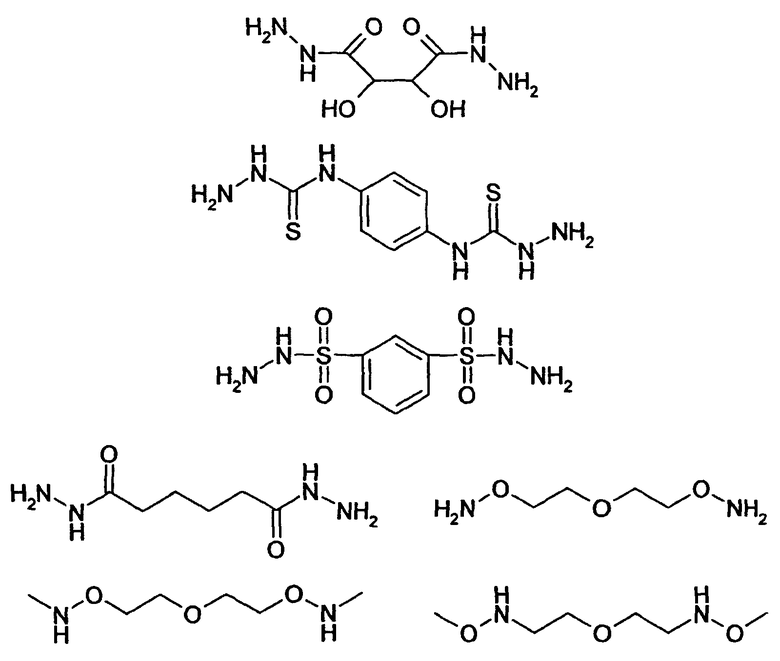
Если HAS-полипептидный конъюгат по настоящему изобретению получают путем реакции производного HAS, включающего HAS и по меньшей мере одно сшивающее соединение, с тиогруппой полипептида, предпочтительно проводить реакцию по его восстановленному концу при необязательном окислении с первым сшивающим соединением, которое предпочтительно представляет собой соединение
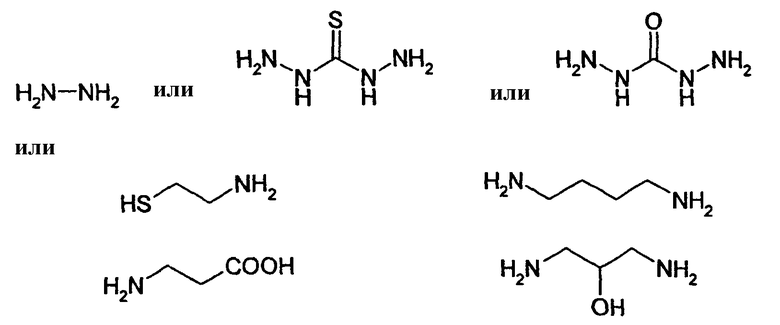
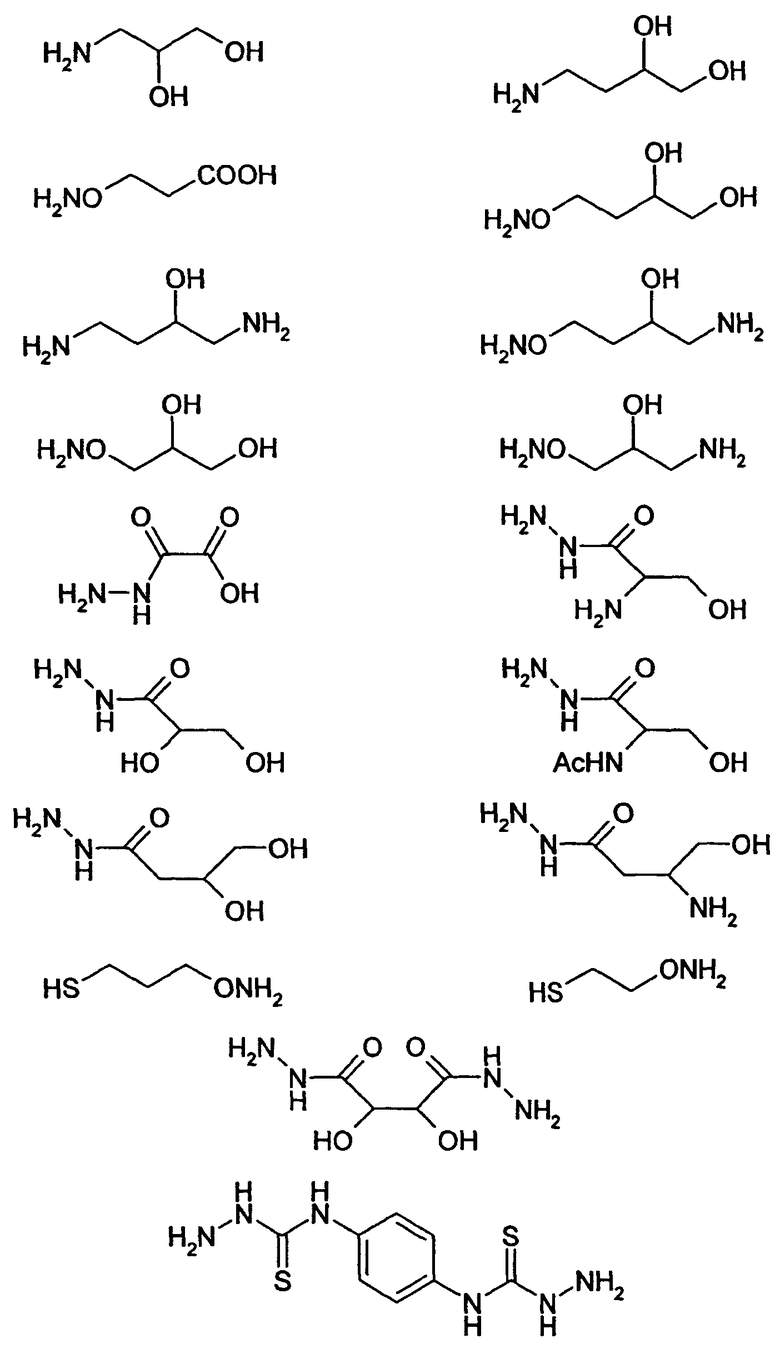
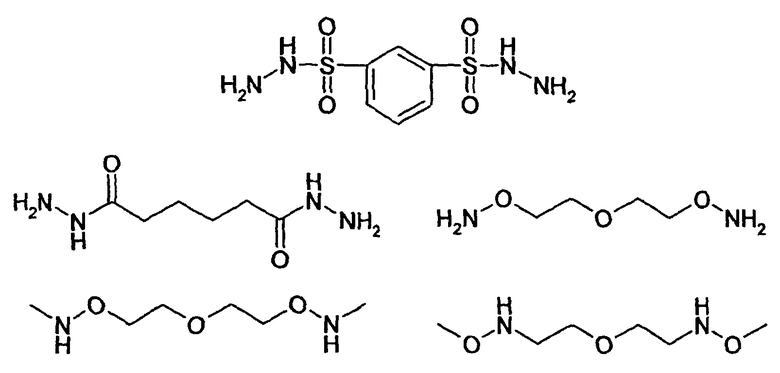
и далее проводят реакцию полученного HAS-производного со вторым сшивающим соединением, которое способно взаимодействовать с HAS-производным и тиогруппой полипептида. Если, например, используют HAS-производное, которое включает в качестве функциональной группы, взаимодействующей со вторым сшивающим соединением, структуру -NH-, как подробно описано выше, то предпочтительными являются следующие типы вторых сшивающих соединений с функциональными группами F1 и F2:
Особенно предпочтительные примеры первого сшивающего соединения, включают:
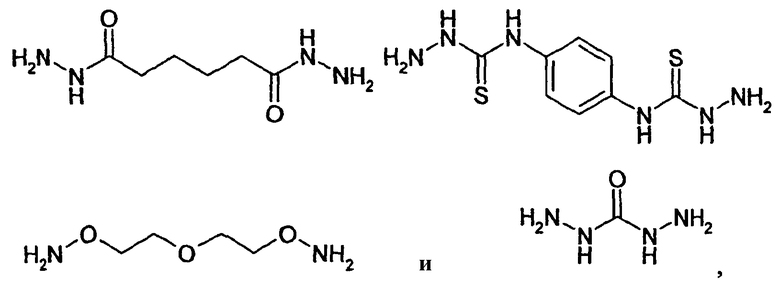
и соединения
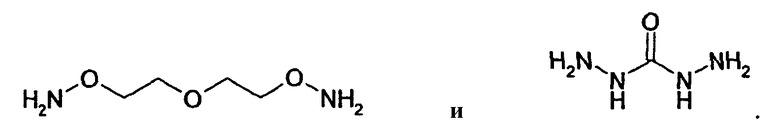
которые являются особенно предпочтительными, и приведенная ниже вторая группа включает сшивающие соединения
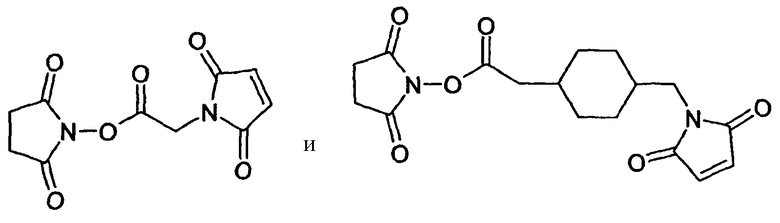
которые являются предпочтительными, при этом соединения
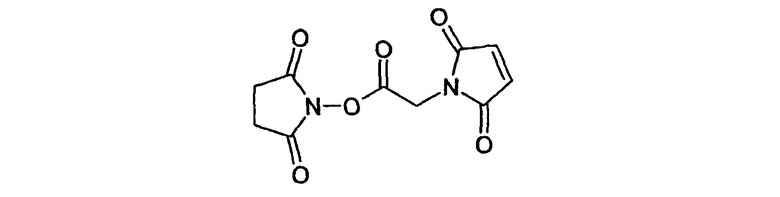
являются особенно предпочтительными.
В зависимости от соответствующих условий реакции, используемых растворителей или смеси растворителей и/или природы остатков R' и/или R" в соединении R'-NH-R", с которым HAS взаимодействует в водной среде, становится возможным, чтобы гидроксиалкилкрахмальное производное, получаемое по такому способу или способам, описанным выше, могло иметь указанную ниже структуру (IIIa):
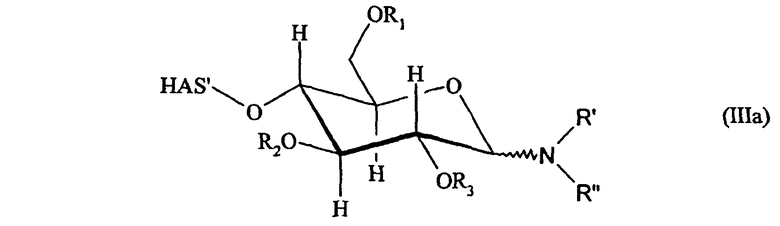
В этой связи настоящее изобретение также относится к описанным выше гидроксиалкилкрахмальным производным, которые имеют структуру формулы (IIIa).
Возможно также, чтобы, например, в том случае, когда R' обозначает водород, указанное гидроксиалкилкрахмальное производное, получаемое по способу или способам, описанным выше, имело структуру (IIIa) или (IIIb), где (IIIa) и (IIIb) могли бы оба присутствовать в реакционной смеси в состоянии определенного равновесия:
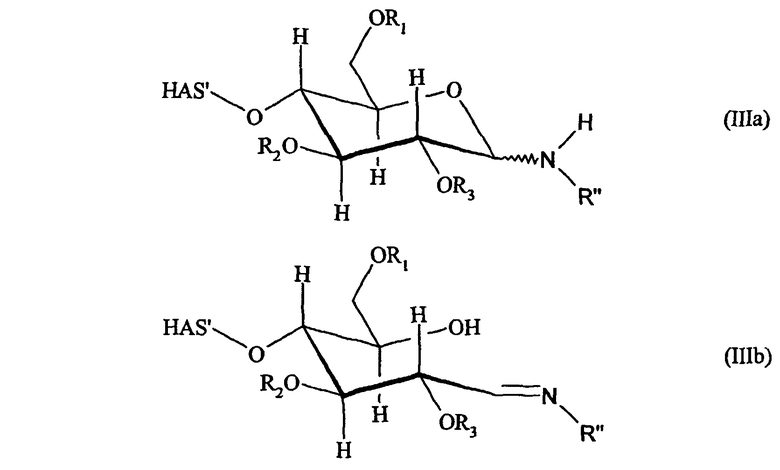
В этой связи настоящее изобретение также относится к описанному выше производному гидроксиалкилкрахмала формулы (IIIb).
Более того, настоящее изобретение относится также к описанному выше производному гидроксиалкилкрахмала, который присутствует в смеси структур формул (IIIa) и (IIIb).
В зависимости от условий реакции и/или химической природы соединения R'-NH-R", используемого в реакции, могут присутствовать соединения формулы (IIIa), в которых атом N находится в экваториальном или аксиальном положении, а также может присутствовать смесь обеих форм в состоянии их определенного равновесного распределения.
В зависимости от условий реакции и/или химической природы соединения R'-NH-R", используемого в реакции, могут присутствовать соединения формулы (IIIb), в которых имеется двойная C-N связь в E- или Z-конфигурации, а также может присутствовать смесь обеих форм в состоянии их определенного равновесного распределения.
В некоторых случаях может быть желательно стабилизировать соединение формулы (IIIa). Это особенно справедливо в том случае, когда соединение формулы (IIIa) получают и/или используют в водном растворе. В качестве способа стабилизации особенно предпочтительно проведение ацилирования соединения формулы (IIIa), особенно в том случае, когда R' обозначает водород. В качестве ацилирующего реагента могут использоваться все приемлемые реагенты с получением желательного производного гидроксиалкилкрахмала формулы (IVa)
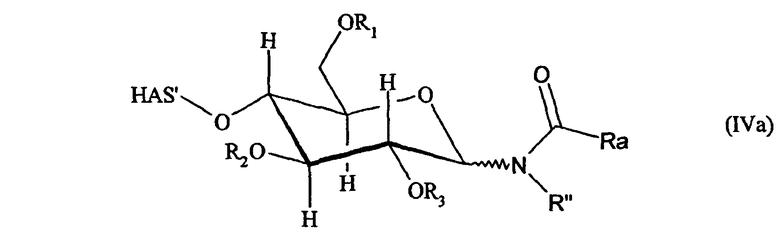
В соответствии с особенно предпочтительными вариантами осуществления настоящего изобретения остаток Ra, представляющий собой часть ацилирующего реагента, является метилом. В качестве ацилирующих реагентов предпочтительно используют ангидриды карбоновых кислот, галогенангидриды карбоновых кислот и активированные сложные эфиры карбоновых кислот.
В этой связи настоящее изобретение также относится к производному гидроксиалкилкрахмала, получаемому по описанному выше способу, где указанное производное имеет структуру формулы (IVa).
Реакцию ацилирования проводят в температурном диапазоне от 0 до 30°С, предпочтительно в диапазоне от 2 до 20°С и особенно предпочтительно в диапазоне от 4 до 10°С.
В других случаях может быть желательно осуществить стабилизацию соединения формулы (IIIb). Такой подход особенно справедлив для того случая, когда соединение формулы (IIIb) получают и/или используют в водном растворе. В качестве способа стабилизации особенно предпочтительно проведение реакции восстановления соединения формулы (IIIb), особенно в том случае, когда R' обозначает водород. В качестве восстановителя могут использоваться все подходящие реагенты, что приводит к получению желательного производного гидроксиалкилкрахмала формулы (IVb)
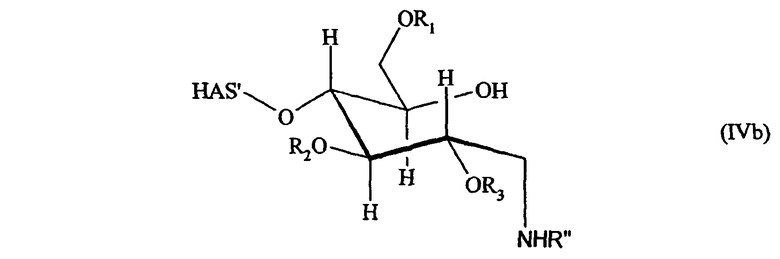
В соответствии с особенно предпочтительными вариантами осуществления настоящего изобретения в качестве восстановителей используют боргидриды, такие как NaCNBH3 или NaBH4.
В этой связи настоящее изобретение также относится к производному гидроксиалкилкрахмала, получаемому по описанному выше способу, где указанное производное имеет структуру формулы (IVb).
Реакцию восстановления проводят в температурном диапазоне от 4 до 100°С, предпочтительно в диапазоне от 10 до 90°С и особенно предпочтительно в диапазоне от 25 до 80°С.
Настоящее изобретение также относится к смесям соединений (IIIa) и (IIIb), (IVa) и (IVb), (IIIa) и (IVa), (IIIa) и (IVb), (IIIb) и (IVa), (IIIb) и (IVb), (IIIa) и (IIIb) и (IVa), (IIIa) и (IIIb) и (IVb), (IVa) и (IVb) и (IIIa), и (IVa) и (IVb) и (IIIb), где (IIIa) и/или (IVa) могут независимо присутствовать в конфигурации, в которой атом N может находиться в экваториальном или аксиальном положении, и/или где может присутствовать (IIIb), в котором имеется двойная C-N связь в E- или Z-конфигурации.
Настоящее изобретение также относится к HAS-полипептиду по настоящему изобретению, используемому в способе лечения человека или животного.
Кроме того, настоящее изобретение относится к фармацевтической композиции, включающей HAS-полипептид по настоящему изобретению. В предпочтительном варианте фармацевтическая композиция включает также по меньшей мере один фармацевтически приемлемый разбавитель, адъювант и/или носитель, используемые в терапии эритропоэтином.
Настоящее изобретение также относится к использованию HAS-полипептида по настоящему изобретению при получении лекарственного средства, используемого для лечения анемических нарушений или гематопоэтических дисфункций или связанных с ними заболеваний.
Настоящее изобретение ниже поясняется с помощью прилагаемых чертежей, таблиц и примеров, которые не следует интерпретировать в плане ограничения области настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1
На фиг.1 показаны результаты анализа в SDS-PAGE двух конъюгатов HES-EPO
Фиг.2
На фиг.2 приведены данные, показывающие, что HES конъюгируют с углеводным фрагментом боковой цепи углевода, при этом показано расщепление HAS-модифицированных форм EPO N-гликозидазой полипептида
CA, USA)
Фиг.3
На фиг.3 приведены результаты анализа в SDS-PAGE конъюгата HES-EPO, получаемого по процедуре примера 17.1.
значения молекулярной массы (кДа) белкового маркера сверху вниз:
245, 123, 77, 42, 30, 25,4 и 17.
Фиг.4
На фиг.4 приведены результаты анализа в SDS-PAGE конъюгата HES-EPO, получаемого по процедуре примера 17.3.
значения молекулярной массы (кДа) белкового маркера сверху вниз:
245, 123, 77, 42, 30, 25,4 и 17.
Фиг.5
На фиг.5 приведены результаты анализа в SDS-PAGE конъюгата HES-EPO, получаемого по процедуре примеров 17.4 и 17.5.
значения молекулярной массы (кДа) белкового маркера сверху вниз:
245, 123, 77, 42, 39, 25,4 и 17.
Фиг.6
На фиг.6 приведены результаты анализа в SDS-PAGE HES-EPO конъюгатов, получаемых по процедуре примеров 19.1 и 19.4.
молекулярные массы (в кДа) белкового маркера составляют сверху вниз:
245, 123, 77, 42, 30, 25,4 и 17;
Фиг.7
На фиг.7 приведены результаты анализа в SDS-PAGE HES-EPO конъюгатов, получаемых по процедуре примеров 19.2, 19.3, 19.5 и 19.6.
молекулярные массы (в кДа) белкового маркера составляют сверху вниз:
245, 123, 77, 42, 30, 25,4 и 17;
основанной на методике примера 13.3b;
основанной на методике примера 13.1b;
основанной на методике примера 13.3а;
основанной на методике примера 13.1а;
Фиг.8
На фиг.8 приведены результаты анализа в SDS-PAGE HES-EPO конъюгатов, получаемых по процедуре примеров 19.7, 19.8, 19.9, 19.10, 19.11 и 19.12.
молекулярные массы (в кДа) белкового маркера составляют сверху вниз:
245, 123, 77, 42, 30, 25,4 и 17;
Фиг.9
Результаты анализа в SDS-PAGE EPO-GT-1, подвергнутого мягкой кислотной обработке: в течение 5 минут = линия 2; 10 минут = линия 3; 60 минут = линия 4 и необработанный EPO = линия 1; показано смещение мобильности EPO после удаления N-гликанов (+PNGASE).
Фиг.10
Результаты анализа в HPAEC-PAD олигосахаридов, выделенных из необработанного EPO и из EPO, инкубированных в течение 5 минут, 10 минут и 60 минут в условиях мягкого кислотного гидролиза. Римские цифры I-V указывают положение при элюции: I = десиалилированной двуантенной структуры, II = трисиалилированных трехантенных структур (два изомера); III = антенной (tennary) структуры; II = трисиалилированных трехантенных структур (два изомера); III = тетрасиалилированной тетраантенной структуры + 2 N-ацетиллактозаминовых повтора; IV = тетрасиалилированной тетраантенной структуры + 1 N-ацетиллактозаминовый повтор; V = тетрасиалилированной тетраантенной структуры без N-ацетиллактозаминового повтора. В скобках показана область элюции олигосахаридных структур без сиаловой кислоты и при наличии 1-4 сиаловых кислот.
Фиг.11
Результаты анализа в HPAEC-PAD N-связанных олигосахаридов после десиалилирования; показано положение в ходе элюции N-ацетилнейраминовой кислоты; номера 1-9 указывают положение стандартных олигосахаридов: 1 = двухантенный; 2 = трехантенный (2-4 изомера), 3 = трехантенный (2-6 изомеров), 4 = тетраантенный; 5 = трехантенный плюс 1 повтор; 6 = тетраантенный плюс 1 повтор; 7 = трехантенный плюс 2 повтора; 8 = тетраантенный плюс 2 повтора и 9 = тетраантенный плюс 3 повтора.
Фиг.12
Результаты анализа в SDS-PAGE EPO после обработки в мягких условиях и необработанного EPO, которые подвергают окислению перйодатом по остаткам сиаловой кислоты. 1 = окисление перйодатом, но без кислотной обработки; 2 = окисление перйодатом плюс 5 минут кислотной обработки; 3 = окисление перйодатом плюс 10 минут кислотной обработки; 4 = окисление перйодатом, но без кислотной обработки; 5 = Стандарт EPO BRP без окисления перйодатом и без кислотной обработки.
Фиг.13
Результаты анализа в HPAEC-PAD нативных олигосахаридов, выделенных из необработанного EPO и из EPO, инкубированного в течение 5 минут и 10 минут в условиях мягкого кислотного гидролиза, с последующей обработкой перйодатом. В скобках 1-5 показана область элюции олигосахаридных структур без сиаловой кислоты и при наличии 1-4 сиаловых кислот.
Фиг.14
Результаты анализа в SDS-PAGE HES-модификации с течением времени EPO-GT-1-А: аликвоты по 20 мкл EPO-GT-1-А подвергают реакции с производным Х, полученным с использованием модифицированного гидроксиламином HES, в течение 30 минут, 2, 4 и 17 часов. Линия 1 = время реакции 30 минут; линия 2 = время реакции 2 часа; линия 3 = время реакции 4 часа; линия 4 = время реакции 17 часов; линия 5 = EPO-GT-1-А без модификации HES. На левом чертеже показано смещение подвижности EPO-GT-1-А при увеличении времени инкубации в присутствии модифицированного гидроксиламином HES-производного (скорость течения: 1 мл/мин); Х: линия 1 = время реакции 30 минут; линия 2 = время реакции 2 часа; линия 3 = время реакции 4 часа; линия 4 = время реакции 17 часов; линия 5 = EPO-GT-1-А при наличии HES-модификации. Чертеж с правой стороны показывает результат анализа тех же образцов после их обработки N-гликозидазой.
Фиг.15
Результаты анализа в SDS-PAGE фракций конъюгатов HES-EPO, полученных при элюции с сефарозы (Q-Sepharose). Концентрируют в концентраторе Speed Vac 1% фракции, полученной при прохождении потока через колонку, и 1% фракции, элюируемой при высокой концентрации соли, и затем наносят на гели в буфере для образца. EPO белок окрашивают красителем кумасси голубым. А = образец I; B = образец II; C = образец III; К = контроль EPO-GT-1; А1, В1, С1 и К1 относятся к фракциям, полученным при свободном прохождении потока; А2, В2, С2 и К2 указывают на фракции, элюируемые при высокой концентрации соли.
Фиг.16а
Результаты анализа в SDS-PAGE HES-модифицированного образца EPO А2 (см. фиг.15), контрольного образца EPO К2 и EPO-GT-1 препарата EPO, расщепленных в присутствии N-гликозидазы, с тем чтобы удалить N-связанные олигосахариды. Все образцы EPO демонстрируют смещение мобильности в сторону форм с низкой молекулярной массой, которые не содержат или содержат О-гликан. Наблюдается низкий коэффициент соотношения полос для О-гликозированных и негликозилированных белков в случае HES-модифицированного образца EPO А2 после де-N-гликозилирования, а также отмечается размытая белковая полоса в области 30 кДа, что преимущественно отражает наличие HES-модификации по сиаловой кислоте О-гликанового остатка (см. стрелку, помеченную звездочкой).
Фиг.16b
Результаты анализа в SDS-PAGE HES-модифицированного образца EPO А2после гидролиза в мягких условиях (см. фиг.15), контрольного образца EPO К2 и EPO-GT-1А, необработанных или расщепленных в присутствии N-гликозидазы для удаления N-связанных олигосахаридов (см. фиг.16а). Как высокомолекулярная форма А2 до, так и форма А после N-гликозидазной обработки (см. участки со скобками, помеченными и не помеченными стрелками), исчезают при кислотной обработке образцов. Стандарт EPO BRP, который анализируют для сравнения, не подвергают кислотной обработке в мягких условиях.
Фиг.17
Результаты анализа в HPAEC-PAD N-связанного олигосахаридного материала, высвобождаемого из HES-модифицированного образца А, из EPO-GT-1-А и из контрольного образца EPO, инкубированного с немодифицированным HES (К). Римские цифры I-V указывают положение при элюции: I = дисиалилированной двухантенной структуры, II = трисиалилированных трехантенных структур (два изомера); III = тетрасиалилированной тетраантенной структуры + 2 N-ацетиллактозаминовых повтора; IV = тетрасиалилированной тетраантенной структуры + 1 N-ацетиллактозаминовый повтор; V = тетрасиалилированной тетраантенной структуры без N-ацетиллактозаминового повтора; в скобках показана область элюции ди-, три- и тетрасиалилированных N-гликанов, как описано в пояснении к фиг.10 и 13.
Фиг.18
Результаты анализа в HPAEC-PAD N-связанного олигосахаридного материала, высвобождаемого из HES-модифицированного образца А, из EPO-GT-1А и из контрольного образца EPO (К), инкубированного с немодифицированным HES. Показано время удерживания смеси стандартных олигосахаридов: номера 1-9 указывают положение при элюции стандартных олигосахаридов: 1 = двухантенного; 2 = трехантенного (2-4 изомера); 3 = трехантенного (2-6 изомеров); 4 = тетраантенного; 5 = трехантенного плюс 1 повтор; 6 = тетраантенного плюс 1 повтор; 7 = трехантенного плюс 2 повтора; 8 = тетраантенного плюс 2 повтора и 9 = тетраантенного плюс 3 повтора.
Фиг.19-25
На фиг.19-25 показаны масс-спектры MALDI/TOF энзиматически высвобожденных и химически десиалилированных N-гликанов, выделенных из препаратов HES-модифицированного EPO и контрольного EPO. Основные сигналы при значениях м/z, равных 1809,7, 2174,8, 2539,9, 2905,0 и 3270,1 ([M+Na]+), соответствуют ди- и тетраантенным N-гликановым структурам комплексного типа, в отсутствие или при наличии одного или двух N-ацетиллактозаминовых повторов, сопровождаемых слабыми сигналами из-за потери фукозы или галактозы, вызванной условиями кислотной обработки, используемыми для десиалилирования образцов, подвергаемых МС анализу.
Фиг.19
MALDI/TOF спектр: десиалилированные олигосахариды HES-модифицированного ЕРО А2.
Фиг.20
MALDI/TOF спектр: десиалилированные олигосахариды EPO-GT-1A.
Фиг.21
MALDI/TOF спектр: десиалилированные олигосахариды EPO К2.
Фиг.22
MALDI/TOF спектр: десиалилированные олигосахариды EPO-GT-1.
Фиг.23
MALDI/TOF спектр: десиалилированные олигосахариды EPO-GT-1, подвергнутые кислотному гидролизу в течение 5 минут.
Фиг.24
MALDI/TOF спектр: десиалилированные олигосахариды EPO-GT-1, подвергнутые кислотному гидролизу в течение 10 минут.
Фиг.25
MALDI/TOF спектр: десиалилированные олигосахариды EPO-GT-1, подвергнутые кислотному гидролизу в течение 60 минут.
ПРИМЕРЫ
Пример 1
Получение рекомбинантного EPO
А) Получение в клетках млекопитающего
Рекомбинантный EPO получают в CHO клетках следующим образом.
Плазмиду, несущую кДНК для EPO, клонируют в эукариотическом векторе экспрессии (pCR3 и впоследствии названный pCREPO). Сайт-специфический мутагенез был выполнен с использованием стандартных методов, как описано в литературе (Grabenhorst, Nimtz, Costa et al., 1998. In vivo specificity of human alpha 1,3/4-fucosyltransferases III-VII in the biosynthesis of Lewis(x) and sialyl Lewis(x) motifs on complex-type N-glycans -Coexpression studies from BHK-21 cells together with human beta-trace protein, J. Biol. Chem., 273(47), 30985-30994).
CHO клетки, стабильно экспрессирующие человеческий EPO или его аминокислотные варианты (например, Cys-29→Ser/Ala или Cys-33→Ser/Ala, Ser-126→Ala и т.п.), получают по методике осаждения фосфатом кальция и отбирают с помощью G418-сульфата, как описано в литературе (Grabenhorst et al.). Через три дня после трансфекции клетки подвергают субкультивированию в пропорции 1:5 и отбирают на среде ДМЕМ, содержащей 10% PBS и 1,5 г/л G418-сульфата.
При использовании указанной процедуры обычно имеют уровень выживания 100-500 клонов, которые далее размножают в селективной среде еще в течение 2-3 недель. Супернатанты клеточных культур из выросших до слияния монослоев анализируют далее на уровень экспрессии EPO по методике вестерн-блоттинга и в рамках IEF-вестерн-блоттинг-анализа.
ЕРО получают из стабильных субклонов в роллерных колбах или в 21 перфузионном реакторе. Различные гликоформы EPO с варьирующими количествами NeuAc (например, 2-8, 4-10, 8-12 остатками NeuAc) выделяют по известным методикам с использованием сочетаний различных хроматографических процедур, как описано ниже.
Литература:
Grabenhorst, Conradt, 1999, The cytoplasmic, transmembrane, and stem regions of glycosyltransferases specify their in vivo functional sublocalization and stability in the Golgi., J Biol Chem., 274(51), 36107-16; Grabenhorst, Schlenke, Pohl, Nimtz, Conradt, 1999, Genetic engineering of recombinant glycoproteins and the glycosylation pathway in mammalian host cells, Glycoconj J., 16(2), 81-97; Mueller, Schlenke, Nimtz, Conradt, Hauser, 1999, Recombinant glycoprotein protein product quality in proliferation-controlled BHK-21 cells, Biotechnology and bioengineering, 65(5), 529-536; Schlenke, Grabenhorst, Nimtz, Conradt, 1999, Construction and characterization of stably transfected BHK-21 cells with human-type sialylation characteristic, Cytotechnology, 30(1-3), 17-25.
B) Получение в клетках насекомого
Рекомбинантный EPO получают в клетках клеточной линии насекомых SF9 и SF21 после инфицирования клеток рекомбинантным бакуловирусным вектором, содержащим кДНК для EPO, под контролем полигедринового промотора, как описано в литературе.
Клетки, выращенные в бессывороточной культуральной среде, инфицируют с плотностью клеток 2×106 или ×107 клеток на мл и определяют титр EPO каждый день в супернатантах клеточных культур. EPO очищают хроматографией на сефарозе (Blue Sepharose), ионообменной хроматографией на сефарозе (Q-Sepharose) и в конце RP-ВЭЖХ в С4-фазе.
Чистоту продукта проверяют в SDS-PAGE и проводят анализ путем N-терминального секвенирования - подробного структурного анализа углеводов (N- и О-гликозилирования) в соответствии с опубликованными процедурами.
Литература:
Grabenhorst, Hofer, Nimtz, Jager, Conradt, 1993, Biosynthesis and secretion of human interleukin 2 glycoprotein variants from baculovirus-infected Sf21 cells. Characterization of polypeptides and posttranslational modifications, Eur J Biochem., 215(1), 189-97; Quelle, Caslake, Burkert, Wojchowski, 1989, High-level expression and purification of a recombinant human erythroproietin produced using a baculovirus vector, Blood, 74(2), 652-7.
Пример 2
Получение реактивных HES-производных
SH-реактивный HES
1.1 Реакция ЕМСН с оксо-HES12KD с образованием SH-реактивного HES12KD В

0,144 г (0,012 ммоль) оксо-HES12KD (Fresenius German патент DE 19628705 A1) растворяют в 0,3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота смесь 34 мг (0,15 ммоль) EMCH (Perbio Science, Deutschland GmbH, Bonn, Germany) в 1,5 мл ДМСО. После перемешивания в течение 19 часов при 60°С реакционную смесь добавляют к 16 мл смеси этанола и ацетона, 1:1. Осадок собирают центрифугированием, перерастворяют в 3 мл ДМСО и снова осаждают, как описано. SH-реактивный HES12KD B получают после центрифугирования и сушки в вакууме. Реакция конъюгирования с тио-ЕРО описана в примере 3, 2.2.
Альтернативы:
В данной реакции могут использоваться все сшивающие линкеры, которые демонстрируют наличие гидразидной или малеимидной функций, разделенных спейсером. Другие примеры молекул с данной группой, доступные от Perbio Science, Deutschland GmbH, Bonn, Germany, показаны в таблице 2 со знаком "А". Кроме того, может использоваться другая группа сшивающих линкеров, демонстрирующих наличие активированной дисульфидной функции вместо малеимидной функции.
1.2 Галогенацетамидные призводные HES-гликозаминов
а) Образование гликозамина (Manger, Wong, Rademacher, Dwek, 1992, Biochemistry, 31, 10733-10740; Manger, Rademacher, Dwek, 1992, Biochemistry, 31, 10724-10732)
1 мл образца HES12KD растворяют в 3 мл насыщенного бикарбоната аммония. Добавляют дополнительное количество бикарбоната аммония для насыщения раствора в ходе инкубации в течение 120 часов при 30°С. Амино-HES12KD C обессоливают путем непосредственной лиофилизации реакционной смеси.
b) Ацилирование гликозамина С ангидридом хлоруксусной кислоты
1 мг амино-HES12KD C растворяют в 1 мл 1 М бикарбоната натрия и охлаждают на льду. К указанной смеси добавляют кристаллы твердого ангидрида хлоруксусной кислоты (˜5 мг) и реакционной смеси дают нагреться до комнатной температуры. Отслеживают рН реакционной смеси и, если значение рН опускается ниже 7,0, добавляют дополнительное количество основания. Через два часа при комнатной температуре добавляют вторую аликвоту основания и ангидрида. Через шесть часов продукт хлорацетамид-HES D1 (X=Cl) обессоливают пропусканием через смешанный слой ионообменных смол амберлит (Amberlite MB-3(H)(OH)).
c) Ацилирование гликозамина бромуксусным ангидридом (Black, Kiss, Tull, Withers, 1993, Carbohydr., Res., 250, 195)
Получают бромуксусный ангидрид по методике, описанной в литературе (Thomas, 1977, Methodes Enzymol., 46. 362). 1 мг образца амино-HES12KD C растворяют в 0,1 мл сухого ДМФА, охлаждают на льду и добавляют 5 мг бромуксусного ангидрида. Реакционную смесь медленно доводят до комнатной температуры и раствор перемешивают в течение 3 часов. Далее указанную реакционную смесь добавляют к 1 мл смеси этанола и ацетона, 1:1, при -20°С. Осадок собирают центрифугированием, перерастворяют в 0,1 мл ДМФА и снова осаждают, как описано. Бромацетамид-HES D2 (X=Br) получают путем центрифугирования и сушки в вакууме. Реакция конъюгирования с тио-EPO описана в примере 3, 1.2.
d) Соответствующее иодистое производное D3 (X=I) синтезируют по методике получения D2. Вместо бромуксусного ангидрида используют N-сукцинимидиловый иодацетат и все стадии синтеза проводят в темноте.
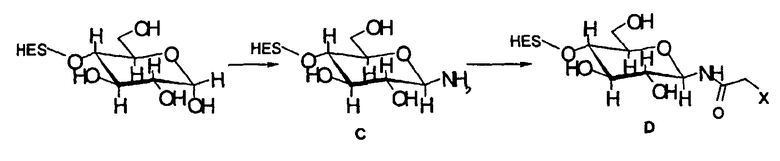
Альтернативы:
С целью ацилирования аминогруппы могут использоваться другие галогенсодержащие кислоты, например:
- бромангидриды или хлорангидриды;
- сложные эфиры, например, сложный N-гидроксисукцинимидный эфир, сложные эфиры на основе замещенных фенолов (п-нитрофенол, пентафторфенол, трихлорфенол и т.п.).
Кроме того, могут использоваться сшивающие линкеры, имеющие реактивную аминогруппу и галогенацетильную функцию, разделенные спейсером. Примером таких линкеров является SBAP. Указанная молекулярная форма доступна от Perbio Science, Deutschland GmbH, Bonn, Germany. Данные молекулы помечены в таблице 2 знаком "D". Относительно использования сшивающих линкеров для лигирования амино-HES с тио-EPO без выделения производных галогенацетамид-HES см. примечения, приведенные в примере 3, 1.2.
1.3 Галогенацетамидные производные амино-HES E(Rademacher, Dwek, 1992, Biochemistry, 31, 10724-10732)
а) Реакция 1,4-диаминобутана с оксо-HES12KD с образованием амино-HES12KD E(S.Frie, Diplomarbeit, Fachhoschule Hamburg, 1998)
1,44 г (0,12 ммоль) оксо-HES12KD растворяют в 3 мл сухого диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 1,51 мл (15 ммоль) 1,4-диаминобутана в 15 мл ДМСО. После перемешивания в течение 19 часов при 40°С реакционную смесь добавляют к 160 мл смеси этанола и ацетона, 1:1. Осадок амино-HES12KD E собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
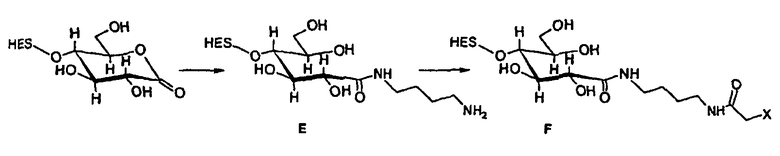
b) Хлорацетамид-HES12KD F1 получают по методике, описанной выше, в1.3, для хлорацетамид-HES12KD D1.
с) Бромацетамид-HES12KD F2 (X=Br) получают по методике, описанной выше, в1.3, для бромацетамид-HES12KD D2. Реакция конъюгирования с тио-EPO описана в примере 3, 1.2.
d) Соответствующее иодистое производное F3 (X=I) не выделяют перед его реакцией с тио-EPO. Указанный эксперимент описан в примере 3, 1.1.
Альтернативы:
См. пункт 1.2, приведенный выше.
2. CHO-реактивный HES
2.1 Гидразид-HES
а) Реакция гидразина с оксо-HES12KD
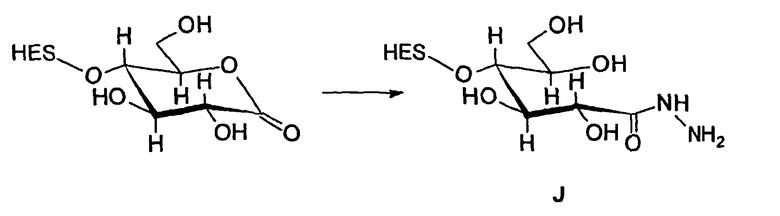
1,44 г (0,12 ммоль) оксо-HES12KD растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 0,47 мл (15 ммоль) гидразина в 15 мл ДМСО. После перемешивания в течение 19 часов при 40°С реакционную смесь добавляют к 160 мл смеси этанола и ацетона, 1:1. Осадок амино-HES12KD J собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 2 дней против 0,5% (объем/объем) водного раствора триэтиламина (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют. Реакция конъюгирования с окислением глико-EPO описана в примере 4, 2.2.
b) Реакция дигидразида адипиновой кислоты с оксо-HES12KD
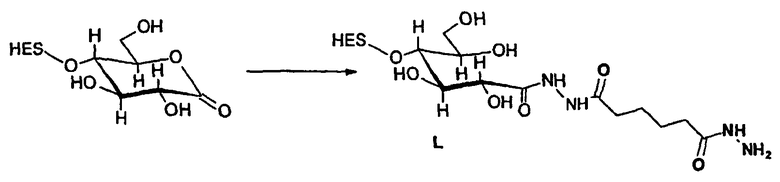
1,74 г (15 ммоль) дигидразида адипиновой кислоты растворяют в 20 мл абсолютного диметилсульфоксида (ДМСО) при 65°С и добавляют по каплям в атмосфере азота 1,44 г (0,12 ммоль) оксо-HES12KD, растворенного в 3 мл абсолютного ДМСО. После перемешивания в течение 68 часов при 60°С реакционную смесь добавляют к 200 мл воды. Раствор, содержащий L, диализуют в течение 2 дней против 0,5% (объем/объем) водного раствора триэтиламина и в течение 2 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют. Реакция конъюгирования с окислением глико-EPO описана в примере 4, 2.2.
Альтернативы:
Кроме того, могут использоваться производные, в которых 2 гидразидные группы разделены любым спейсером.
3. Другие амино-HES12KD-производные I и H (Rademacher, Dwek, 1992, Biochemistry, 31, 10724-10732)
Аммонолиз D или F проводят отдельно путем растворения образца 1 мг каждого из галогенацетамидов в 0,1 мл насыщенного карбоната аммония. Затем добавляют дополнительное количество твердого карбоната аммония для поддержания состояния насыщенности раствора в ходе инкубации в течение 120 часов при 30°С. Реакционную смесь добавляют к 1 мл смеси этанола и ацетона, 1:1, при -20°С. Осадок собирают центрифугированием, перерастворяют в 0,05 мл воды и снова осаждают, как описано. Продукт амино-HES H или I получают путем центрифугирования и последующей сушки в вакууме. Реакция конъюгирования с окисленным глико-EPO описана в примере 4, 4.1.
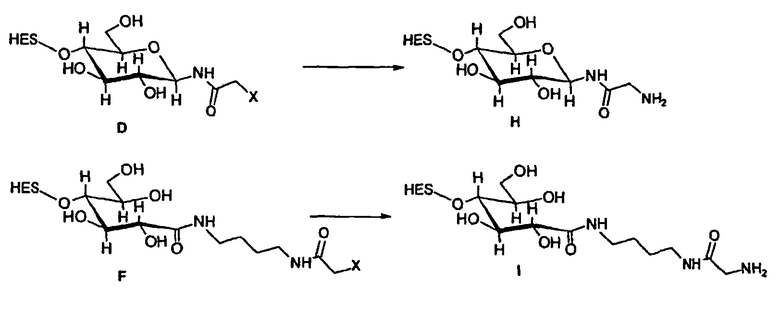
4. НES12KD K, модифицированный гидроксиламином
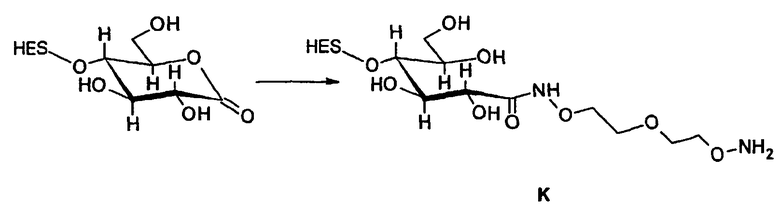
О-[2-(2-аминооксиэтокси)этил]гидроксиламин синтезируют по методике Boturyn et al. в 2 стадии из коммерчески доступного материала (Boturyn, Boudali, Constant, Defrancq, Lhomme, 1997, Tetrahedron, 53, 5485). 1,44 г (0,12 ммоль) оксо-HES12KD растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 2,04 г (15 ммоль) О-[2-(2-аминооксиэтокси)этил]гидроксиламина в 15 мл ДМСО. После перемешивания в течение 48 часов при 65°С реакционную смесь добавляют к 160 мл смеси этанола и ацетона, 1:1. Осажденный продукт К собирают центрифугированием, перерастворяют в 40 мл воды, диализуют против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют. Реакция конъюгирования с окисленным глико-EPO описана в примере 4, 3.1.
Альтернативы:
Кроме того, могут использоваться производные, в которых две гидроксиламиновые группы разделены любым спейсером.
5. Тио-НES12KD
5.1 Добавление к оксо-НES12KD
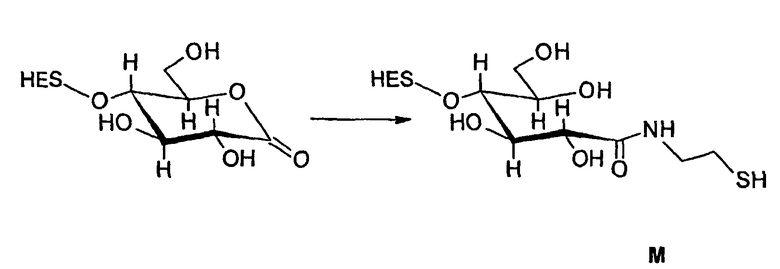
1,44 г (0,12 ммоль) оксо-HES12KD растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 1,16 г (15 ммоль) цистеамина в 15 мл ДМСО. После перемешивания в течение 24 часов при 40°С реакционную смесь добавляют к 160 мл смеси этанола и ацетона, 1:1. Осажденный продукт М собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 2 дней против 0,5% (объем/объем) водного раствора триэтиламина и в течение 2 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют. Реакция конъюгирования с окисленным глико-EPO описана в примере 4, 2.1.
Альтернативы:
Могут использоваться производные, в которых аминогруппа и тиофункция разделены любым спейсером. Кроме того, аминогруппа в производных может быть замещена гидразином, гидразидом или гидроксиламином. Тиофункция может быть защищена, например, за счет создания дисульфидного производного или формы тритильного производного. Однако в данном случае далее должна быть проведена стадия удаления защитной группы перед конъюгированием, которая приведет к высвобождению компонента, аналогичного М.
5.2 Модификация амино-HES12KD E, H или I
а) Модификация с использованием SATA/SATP
1,44 г (0,12 ммоль) амино-HES12KD E, H или I растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 139 мг (0,6 ммоль) SATA в 5 мл ДМСО. После перемешивания в течение 24 часов при комнатной температуре реакционную смесь добавляют к 160 мл смеси этанола и ацетона, 1:1. Осажденный продукт N собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 2 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Реакцию удаления защитной группы проводят в 50 мМ натрий-фосфатного буфера, содержащем 25 мМ ЭДТА и 0,5 М гидроксиламина, рН 7,5 в течение 2 часов при комнатной температуре и продукт O очищают путем диализа против 0,1 М натрий-ацетатного буфера, рН 5,5, содержащего 1 мМ ЭДТА. Реакцию удаления защитной группы проводят непосредственно перед реакцией конъюгирования, которая описана в примере 4, 2.1.
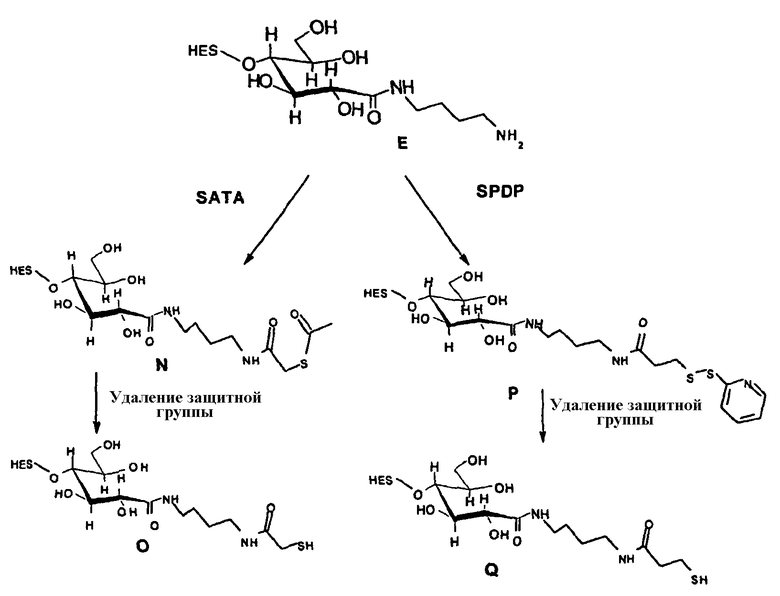
b) Модификация с использованием SPDP
1,44 г (0,12 ммоль) амино-HES12KD E, H или I растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 187 мг (0,6 ммоль) SPDP в 5 мл ДМСО. После перемешивания в течение 24 часов при комнатной температуре реакционную смесь добавляют к 160 мл смеси этанола и ацетона, 1:1. Осажденный продукт Р собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 2 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Реакцию удаления защитной группы проводят в растворе 12 мг дитиотрейтола (ДТТ) в 0,5 мл 100 мМ натрий-ацетатного буфера, содержащего 100 мМ хлорида натрия, при рН 4,5 в течение 30 минут при комнатной температуре и продукт Q очищают путем диализа против 0,1 М натрий-ацетатного буфера, рН 5,5, содержащего 1 мМ ЭДТА. Реакцию удаления защитной группы проводят непосредственно перед реакцией конъюгирования, которая описана в примере 4, 2.1.
Альтернативы
Для целей превращения амино- и тиоловых групп как в свободной форме, как и в защищенном виде может использоваться ряд доступных реагентов. После модификации продукты могут быть выделены. Альтернативно, как принято в случае использования сшивающих линкеров, они могут непосредственно использоваться для реакции конъюгирования, предпочтительно после стадии очищения. Для выделения и хранения тио-HES-производных может осуществляться синтез тио-HES-производных в защищенной форме. С этой целью могут использоваться все аналогичные SATA-производные, которые содержат активную сложноэфирную функцию и тиоэфирную функцию, разделенные спейсером. SATP, как другой представитель данной группы, приведен в таблице 2 с пометкой "H". Производные, которые аналогичны SPDP, могут иметь активную сложноэфирную функцию и тиоэфирную функцию, разделенные спейсером. Другие представители данной группы приведены в таблице 2 с пометкой "F". Дополнительные аналогичные производные могут иметь активную сложноэфирную функцию и тиоловую функцию, защищенную в виде тритильного производного, разделенные спейсером.
Пример 3
Реакции конъюгирования с тио-ЕРО
Реакция тио-ЕРО с модифицированным галогенацетамидом SH-реактивным HES
Пример проведения процедуры 1
Конъюгирование тио-EPO с амино-HES12KD (E, H или I) с использованием сшивающего линкера, содержащего NHS-активный сложный эфир, например SIA (Cumber, Forrester, Foxwell, Ross, Thorpe, 1985, Methods Enzymol., 112, 207)
Материалы
А. Боратный буфер. Композиция включает 50 мМ бората натрия, рН 8,3 и 5 мМ ЭДТА.
В. PBS, фосфатно-буферный раствор: 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4.
С. Амино-HES12KD E, H или I. Готовят в концентрации 1 мг/мл в боратном буфере.
D. Раствор сшивающего линкера: 14 мг SIA растворяют в 1 мл ДМСО.
E. Колонки для обессоливания с декстраном D-SaltTM, рабочий объем 2×5 мл (Perbio Science, Deutschland GmbH, Bonn, Germany)
F. Реактив для количественного определения белка кумасси (Coomassie®) (Perbio Science, Deutschland GmbH, Bonn, Germany)
G. Раствор тио-ЕРО: 5 мг/мл тио-EPO 1 в боратном буфере.
H. Микроконцентратор Microcon Ym-3 (амикон (amicon), Milipore GmbH, Eschborn, Germany).
Метод
100 мкг раствора SIA добавляют к 400 мкл раствора амино-HES12KD E и оставляют для реакции при перемешивании в течение 0,5 часа при комнатной температуре. Избыток сшивающего средства удаляют центрифугированием образца при 14000g в течение 60 минут с использованием микроконцентратора. После центрифугирования объем образца доводят до исходного значения боратным буфером и указанный процесс повторяют еще два раза. Оставшийся раствор добавляют к 1 мл раствора тио-ЕРО и реакционную смесь инкубируют в течение 16 часов при комнатной температуре. В конце инкубационного периода реакционную способность избытка йодацетамида гасят добавлением цистеина до конечной концентрации 10 мМ. Реакционную смесь наносят на колонку для обессоливания, уравновешенную буфером PBS, и во фракциях отслеживают содержание белка по методу анализа, включающему использование кумасси. Объединяют все фракции, содержащие белковый конъюгат, и получают конъюгат путем лиофилизации после диализа против воды в течение ночи.
Альтернативы
В данной реакции могут использоваться все сшивающие линкеры, которые имеют сукцинимидную или сульфосукцинимидную функцию и йодацетамидную функцию, разделенные спейсером. Другие примеры приведены в таблице 2. Они помечены знаком "C" и доступны от Perbio Science, Deutschland GmbH, Bonn, Germany.
1.2 Пример проведения процедуры 2
Конъюгирование тио-ЕРО 1 с бромацетамидом D2, F2 или йодацетамидом D3 SH-реактивного HES12KD (de Valasco, Merkus, Anderton, Verheul, Lizzio, Van der Zee, Van Eden, Hoffmann, Verhoef, Snippe, 1995, Infect. Immun., 63, 961)
Материалы
А. Фосфатный буфер. Композиция включает 100 мМ фосфата натрия, рН 6,1 и 5 мМ ЭДТА.
В. PBS, фосфатно-буферный раствор: 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4.
С. Бромацетамид SH-реактивного HES12KD D2. Готовят в концентрации 10 мг/мл в фосфатном буфере.
D. Колонки для обессоливания с декстраном D-SaltTM, рабочий объем 2×5 мл (Perbio Science, Deutschland GmbH, Bonn, Germany)
Е. Реактив для количественного определения белка кумасси (Coomassie®) (Perbio Science, Deutschland GmbH, Bonn, Germany)
F. Раствор тио-EPO: 5 мг/мл тио-EPO 1 в фосфатном буфере.
Метод
Объединяют 1 мл бромацетамида SH-реактивного HES12KD D2 и 1 мл раствора тио-ЕРО и реакционную смесь инкубируют в течение 48 часов при комнатной температуре. В конце инкубационного периода реакционную способность избытка бромацетамида гасят добавлением цистеина до конечной концентрации 10 мМ. Реакционную смесь наносят на колонку для обессоливания, уравновешенную буфером PBS. Отслеживают во фракциях содержание белка по методу анализа, включающему использование кумасси, и все фракции, содержащие белковый конъюгат, объединяют и получают конъюгат путем лиофилизации после диализа против воды в течение ночи.
Альтернативы:
Вместо выделения бромацетамида SH-реактивного HES12KD D2 может быть проведено связывание HES12KD (E, H, I) с линкером через сукцинимидную и бромацетамидную функцию (см. пункт 1.1, описанный выше). SBAP, как представитель указанной группы сшивающих линкеров, приведен в таблице 2 с пометкой знаком "D".
2. Реакция тио-ЕРО с модифицированным малеимидом SH-реактивным HES
2.1 Пример проведения процедуры 3
Конъюгирование тио-ЕРО с HES12KD с использованием сшивающего линкера, содержащего гидразидную и малеимидную функциональную группу, например, M2C2H.
Материалы
А. Раствор для хранения M2C2H: 10 мг/мл М2С2Н в ДМСО, используют в свежеприготовленном виде
B. HES12KD: в концентрации 10 мг/мл в 0,1 М натрий-ацетатного буфера, рН 5,5
С. Раствор тио-ЕРО: 5 мг/мл тио-ЕРО в фосфатном буфере, содержащем NaCl
D. Фосфат/NaCl: 0,1 М раствора фосфата натрия, 50 мМ NaCl, рН 7,0
E. Микроконцентратор Microcon Ym-3 (амикон (amicon), Milipore GmbH, Eschborn, Germany)
F. Колонка для гель-фильтрации с сефадексом (Sephadex® G-200) (1,5×45 см)
G. Реактив для количественного определения белка кумасси (Coomassie®) (Perbio Science, Deutschland GmbH, Bonn, Germany)
H. PBS, фосфатно-буферный раствор: 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4.
Метод
Раствор M2C2H добавляют к 400 мкл раствора HES12KD до конечной концентрации 1 мМ и оставляют для реакции при перемешивании в течение 2 часов при комнатной температуре. Избыток сшивающего средства удаляют центрифугированием образца при 14000g в течение 60 минут с использованием микроконцентратора. После центрифугирования объем образца доводят до исходного значения фосфатным буфером/NaCl и указанный процесс повторяют еще два раза. К M2C2H-модифицированному HES12KD добавляют 0,5 мл раствора тио-ЕРО и реакционную смесь инкубируют в течение 2 часов при комнатной температуре. В конце инкубационного периода реакционную способность избытка малеимида гасят добавлением цистеина до конечной концентрации 10 мМ. Реакционную смесь наносят на колонку с сефадексом (Sephadex® G-200) (1,5×45 см), уравновешенную буфером PBS, и собирают 1 мл фракций. Во фракциях отслеживают содержание белка по методу анализа, включающему использование реактива для количественного определения белка кумасси. Объединяют все фракции, содержащие белковый конъюгат, и получают конъюгат путем лиофилизации после диализа против воды в течение ночи.
Примечания по процедуре осуществления методики
Гидразоновый аддукт отличается несколько сниженной стабильностью при экстремальных значениях рН. В случае тех направлений применения, которые включают обработку при низком рН, авторы проводят восстановление гидразона с использованием 30 мМ цианборгидрата натрия в буфере PBS до гидразина. Однако для большинства направлений применения такая дополнительная стадия является необязательной.
2.2 Пример проведения процедуры 4
Конъюгирование тио-ЕРО с малеимидо-HES12KD B.
Материалы
А. Малеимидо-HES12KD В: в концентрации 10 мг/мл в 0,1 М натрий-ацетатного буфера, рН 5,5
В. Раствор тио-ЕРО: 5 мг/мл тио-ЕРО в фосфатном буфере, содержащем NaCl
С. Фосфат/NaCl: 0,1 М раствора фосфата натрия, 50 мМ NaCl, рН 7,0
D. Колонка для гель-фильтрации с сефадексом (Sephadex® G-200) (1,5×45 см)
Е. Реактив кумасси (Coomassie®) для количественного определения белка (Perbio Science, Deutschland GmbH, Bonn, Germany)
F. PBS, фосфатно-буферный раствор: 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4.
Метод
Объединяют 1 мл SH-реактивного HES12KD В и 1 мл раствора тио-ЕРО 1 и реакционную смесь инкубируют в течение 2 часов при комнатной температуре. В конце инкубационного периода реакционную способность избытка малеимида гасят добавлением цистеина до конечной концентрации 10 мМ. Реакционную смесь наносят на колонку с сефадексом (Sephadex® G-200) (1,5×45 см), уравновешенную буфером PBS, и собирают 1 мл фракций. Во фракциях отслеживают содержание белка по методу анализа, включающему использование реактива для количественного определения белка кумасси. Объединяют все фракции, содержащие белковый конъюгат, и получают конъюгат путем лиофилизации после диализа против воды в течение ночи.
2.3 Пример проведения процедуры 12
Конъюгирование тио-ЕРО с амино-HES12KD (E, H, I) с использованием сшивающего линкера, содержащего NHS-активный сложный эфир и малеимидную группу, например, SMCC.
Материалы
А. Микроконцентратор Microcon Ym-3 (амикон (amicon), Milipore GmbH, Eschborn, Germany)
В. PBS, фосфатно-буферный раствор: 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4
C. Амино-HES12KD E, H или I. Готовят в концентрации 10 мг/мл в буфере PBS.
D. Раствор SMCC: 1 мг SMCC растворяют в 50 мкл ДМСО.
Е. Колонки для обессоливания с декстраном D-SaltTM, рабочий объем 2×5 мл (Perbio Science, Deutschland GmbH, Bonn, Germany)
F. Реактив для количественного определения белка кумасси (Coomassie®) (Perbio Science, Deutschland GmbH, Bonn, Germany)
G. Раствор тио-ЕРО 1: 5 мг/мл тио-ЕРО 1 в буфере PBS.
Метод
К 50 мкл раствора SMCC добавляют 400 мкл раствора амино-HES12KD E и реакционную смесь оставляют для реакции в течение 80 минут при комнатной температуре и 10 минут при 46°C. Избыток сшивающего средства удаляют центрифугированием образца при 14000g в течение 60 минут с использованием микроконцентратора. Объем доводят до отметки 450 мкл буфером PBS и указанный процесс повторяют еще два раза. После последнего центрифугирования объем оставшегося раствора доводят до 450 мкл с помощью PBS и добавляют к 1 мл раствора тио-ЕРО, после чего реакционную смесь инкубируют в течение 16 часов при комнатной температуре. В конце инкубационного периода реакционную способность избытка малеимида гасят добавлением цистеина до конечной концентрации 10 мМ. Реакционную смесь наносят на колонку для обессоливания, уравновешенную буфером PBS. Во фракциях отслеживают содержание белка по методу анализа, включающему использование реактива для количественного определения белка кумасси, все фракции, содержащие белковый конъюгат, объединяют и получают конъюгат путем лиофилизации после диализа против воды в течение ночи.
Альтернативы:
В данной реакции могут использоваться все сшивающие линкеры, которые содержат сукцинимидную или сульфосукцинимидную функцию и малеимидную функцию, разделенные спейсером. Другие примеры молекул, которые относятся к данной группе, доступные от Perbio Science, Deutschland GmbH, Bonn, Germany, приведены в таблице 2 с пометкой "Е". Имеется еще одна группа сшивающих линкеров, которые содержат вместо малеимидной функции активированную дисульфидную функцию. Указанные сшивающие линкеры могут использоваться для конъюгирования. Однако дисульфидная связь в конъюгате расщепляется в восстановительных условиях. Представители данной группы отмечены в таблице 2 знаком "F". Третья группа сшивающих линкеров в качестве SH-реактивной группы использует вместо малеимидной функции винилсульфоновую функцию. Представитель такой группы "SVSB" помечен в таблице 2 знаком "G".
Пример 4
Реакции конъюгирования с окисленным EPO
Окисление глико-EPO
Окисление глико-EPO мета-перйодатом натрия: пример проведения процедуры 5
Материалы
А. Раствор глико-EPO: 10 мг/мл глико-EPO в ацетатном буфере
В. Раствор мета-перйодата натрия: 10 мМ или 100 мМ перйодата натрия в ацетатном буфере, используют в свежеприготовленном виде. Выдерживают в темноте. В случае использования данных растворов конечная концентрация перйодата натрия в окисленной смеси составляет 1 мМ или 10 мМ, соответственно.
С. Ацетатный буфер: 0,1 М натрий-ацетатного буфера, рН 5,5
D. Глицерин
E. Микроконцентратор Microcon Ym-3 (амикон (amicon), Milipore GmbH, Eschborn, Germany)
Метод
Все стадии выполняют в темноте.
К 1 мл охлажденного раствора глико-EPO добавляют 0,1 мл холодного раствора мета-перйодата натрия и проводят реакцию окисления в течение 1 часа в темноте. Если подвергаемый окислению глико-EPO содержит остатки сиаловой кислоты, то условия окисления включают следующее: 1 мМ перйодата натрия, 0°С. В ином случае используют 10 мМ перйодата натрия в условиях комнатной температуры. Для остановки реакции добавляют окисленный глицерин до конечной концентрации 15 мМ и инкубируют в течение 5 минут при температуре 0°С. Избыток реагентов и побочных продуктов реакции удаляют центрифугированием продукта при 14000g в течение 60 минут с использованием микроконцентратора. После центрифугирования объем образца доводят до исходного значения буфером, используемым на следующей стадии, т.е. ацетатным буфером. Указанный процесс повторяют еще два раза.
1.2 Ферментативное окисление глико-EPO: пример проведения процедуры 6
Ферментативное окисление EPO описано в литературе (Chamow et al., 1992, J. Biol. Chen., 267, 15916-15922).
2. Конъюгирование с производными гидразина/гидразида
2.1 Пример проведения процедуры 7
Конъюгирование окисленного глико-EPO с тио-HES12KD M, O или Q с использованием сшивающего линкера, содержащего гидразидную и малеимидную функциональную группу, например, M2C2H (Perbio Science, Deutschland GmbH, Bonn, Germany).
Материалы
А. Раствор для хранения M2C2H: 10 мг/мл М2С2Н в ДМСО, используют в свежеприготовленном виде
В. Раствор окисленного глико-EPO из 6.1.1: 5 мг/мл глико-EPO в ацетатном буфере
С. Тио-HES12KD М, О или Q: 10 мг/мл в фосфатном буфере, содержащем NaCl
D. Ацетатный буфер: 0,1 М натрий-ацетатного буфера, рН 5,5
Е. Фосфат/NaCl: 0,1 М раствора фосфата натрия, 50 мМ NaCl, рН 7,0
F. Микроконцентратор Microcon Ym-3 (амикон (amicon), Milipore GmbH, Eschborn, Germany)
G. Колонка для гель-фильтрации с сефадексом (Sephadex® G-200) (1,5×45 см)
H. Реактив для количественного определения белка кумасси (Coomassie®) (Perbio Science, Deutschland GmbH, Bonn, Germany)
I. PBS, фосфатно-буферный раствор: 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4.
Метод
Раствор для хранения M2C2H добавляют к 1 мл окисленного глико-EPO до конечной концентрации 1 мМ и оставляют для реакции при перемешивании в течение 2 часов при комнатной температуре. Избыток сшивающего средства удаляют центрифугированием образца при 14000g в течение 60 минут с использованием микроконцентратора. После центрифугирования объем образца доводят до исходного значения фосфатным буфером/NaCl и указанный процесс повторяют еще два раза. К M2C2H-модифицированному глико-EPO добавляют 1 мл раствора тио-HES12KD М, О или Q и реакционную смесь инкубируют в течение 16 часов при комнатной температуре. В конце инкубационного периода реакционную способность избытка малеимида гасят добавлением цистеина. Реакционную смесь наносят на колонку с сефадексом (Sephadex® G-200) (1,5×45 см), уравновешенную буфером PBS, и собирают 1 мл фракций. Во фракциях отслеживают содержание белка по методу анализа, включающему использование реактива для количественного определения белка кумасси, все фракции, содержащие белковый конъюгат, объединяют и получают конъюгат путем лиофилизации после диализа против воды в течение ночи.
Примечания по процедуре осуществления методики
Гидразоновый аддукт отличается несколько сниженной стабильностью при экстремальных значениях рН. В случае тех направлений применения, которые включают обработку при низком рН, авторы проводят восстановление гидразона с использованием 30 мМ цианборгидрата натрия в буфере PBS до гидразина. Однако для большинства направлений применения такая дополнительная стадия является необязательной.
2.2 Пример проведения процедуры 8
Прямое конъюгирование окисленного глико-ЕРО с гидразидо-HES12KD L или J.
Материалы
А. Раствор окисленного глико-EPO из 6.1.1: 5 мг/мл глико-EPO в ацетатном буфере
В. Гидразидо-HES12KD L или J: 10 мг/мл в ацетатном буфере
С. Ацетатный буфер: 0,1 М натрий-ацетатного буфера, рН 5,5
D. Колонка для гель-фильтрации, наример, с сефадексом (Sephadex® G-200) (1,5×45 см)
Е. Реактив для количественного определения белка кумасси (Coomassie®) (Perbio Science, Deutschland GmbH, Bonn, Germany)
F. PBS, фосфатно-буферный раствор: 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4.
Метод
Объединяют 1 мл раствора гидразидо-HES12KD L или Q и добавляют 1 мл раствора окисленного глико-EPO и реакционную смесь оставляют для реакции при перемешивании в течение 16 часов при комнатной температуре. Реакционную смесь наносят на колонку с сефадексом (Sephadex® G-200) (1,5×45 см), уравновешенную буфером PBS, и собирают 1 мл фракций. Во фракциях отслеживают содержание белка по методу анализа, включающему использование реактива для количественного определения белка кумасси, все фракции, содержащие белковый конъюгат, объединяют и получают конъюгат путем лиофилизации после диализа против воды в течение ночи. Результат конъюгирования показан на фиг.24. Наблюдаемое смещение молекул указывает на то, что конъюгирование прошло удачно. Некоторое размазывание полос связано с гетерогенностью HES. Из фиг.25 видно, что HES конъюгирует с углеводным фрагментом боковой цепи углевода.
Примечания по процедуре осуществления методики
Гидразоновый аддукт отличается несколько сниженной стабильностью при экстремальных значениях рН. В случае тех направлений применения, которые включают обработку при низком рН, авторы проводят восстановление гидразона с использованием 30 мМ цианборгидрата натрия в буфере PBS до гидразина. Однако для большинства направлений применения такая дополнительная стадия является необязательной.
3. Конъюгирование с производными гидроксиламина (Rose, 1994, Am. Chem. Soc., 116, 30)
3.1 Пример проведения процедуры 9
Конъюгирование окисленного глико-EPO с гидроксиламино-HES12KD К
Материалы
А. Раствор окисленного глико-EPO из 6.1.1: 5 мг/мл глико-EPO в ацетатном буфере
В. Гидроксиламино-HES12KD К: 10 мг/мл в ацетатном буфере
С. Ацетатный буфер: 0,1 М натрий-ацетатного буфера, рН 5,5
D. Колонка для гель-фильтрации, наример, с сефадексом (Sephadex® G-200) (1,5×45 см)
Е. Реактив для количественного определения белка кумасси (Coomassie®) (Perbio Science, Deutschland GmbH, Bonn, Germany)
F. PBS, фосфатно-буферный раствор: 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4.
Метод
Объединяют 1 мл раствора гидроксиламино-HES12KD К и добавляют 1 мл раствора окисленного глико-EPO и реакционную смесь оставляют для реакции при перемешивании в течение 16 часов при комнатной температуре. Реакционную смесь наносят на колонку с сефадексом (Sephadex® G-200) (1,5×45 см), уравновешенную буфером PBS, и собирают 1 мл фракций. Во фракциях отслеживают содержание белка по методу анализа, включающему использование реактива для количественного определения белка кумасси, все фракции, содержащие белковый конъюгат, объединяют и получают конъюгат путем лиофилизации после диализа против воды в течение ночи. Результат конъюгирования показан на фиг.24. Наблюдаемое молекулярное смещение в полосе 2 указывает на то, что конъюгирование прошло удачно. Некоторое размазывание полос связано с гетерогенностью HES. Из фиг.25 видно, что HES конъюгирует с углеводным фрагментом боковой цепи углевода.
Пример 5
Характеристика N-гликанов EPO, обработанных галактозооксидазой
Рекомбинантный EPO или частично десиалилированные формы EPO (полученные путем ограниченного кислотного гидролиза) инкубируют с галактозооксидазой в присутствии каталазы при 37°С от 30 минут до 4 часов в 0,05 М Na-фосфатного буфера, рН 7,0. Ход реакции отслеживают путем отбора аликвот EPO по 50 мкг с последующей обработкой белка N-гликаназой полипептидов.
Высвобожденные N-связанные олигосахариды (тестирование проводят путем выявления в SDS-PAGE де-N-гликозилированного полипептида) подвергают картированию по методу HPAEC-PAD, как описано в литературе (Grabenhorst et al., 1999, Nimtz et al., 1993/1994; Schlenke et al., 1999) перед и после удаления сиаловых кислот. Количественное определение окисленных остатков галактозы в индивидуальных олигосахаридах EPO проводят на основании определения типичного смещения, отмечаемого в процессе HPAEC-PAD, и подтверждают результаты при исследовании олигосахаридных смесей по методу MALDI/TOF MS.
Пример 6
Характеристика HAS-модифицированного EPO
Отделение HAS-модифицированных форм EPO от непрореагировавшего EPO и молекул-предшественников HAS достигается путем гель-фильтрации с использованием, например, Ультрогеля АсА 44/54 (Ultrogel АсА 44/54) или аналогичных сред, применяемых для фильтрования. Альтернативно, непрореагировавший HAS удаляют путем выделения EPO иммуноаффинным методом на колонке 4 мл, содержащей моноклональные антитела, связанные с гелем Аффигель (Affigel; BioRad), с последующим отделением немодифицированного EPO гель-фильтрацией (например, с использованием матрицы, позволяющей осуществлять отделение глобулярных белков с молекулярной массой от 20 кДа до 200 кДа).
HAS-модифицированный EPO идентифицируют при анализе в SDS-PAGE (с использованием 12,5 или 10% гелей акриламида) путем определения их молекулярных масс в сравнении с молекулярной массой немодифицированного EPO в процессе окрашивания гелей красителем кумасси бриллиантовый синий. Более высокая молекулярная масса HAS-модифицированных полипептидов EPO может быть также идентифицирована посредством вестерн-блот-анализа образцов с использованием поликлональных антител, образованных против рекомбинантного человеческого EPO.
N-гликановая модификация форм EPO была продемонстрирована при успешном удалении EPO белка N-гликаназой полипептидов (рекомбинантная N-гликозидаза от Roche, Germany, используемая в количестве 25 единиц/мг EPO белка при 37°С в течение 16 часов); анализ в SDS-PAGE выявляет наличие типичного смещения белка EPO в положение миграции обработанного N-гликозидазой немодифицированного EPO с массой примерно 20 кДа.
Модификация одного десиалилированного и обработанного галактозооксидазой О-гликана EPO в положении Ser 126 была продемонстрирована в SDS-PAGE по миграции де-N-гликозилированного продукта путем выявления положения его миграции в сравнении с непрореагировавшим де-N-гликозилированным EPO. Если требуется, модифицированный EPO перед анализом в SDS-PAGE подвергают фракционированию методом RP-ВЭЖХ в C8-фазе. О-гликановую модификацию HAS в EPO также анализируют путем β-элиминации О-гликана и выявления де-О-гликозилированной формы EPO методом вестерн-блоттинга с использованием поликлональных антител, образованных против рекомбинантного человеческого EPO.
Пример 7
Количественное определение EPO и модифицированных форм EPO
Количественное определение EPO проводят на основе измерения УФ поглощения в соответствии с процедурой Европейской Фармакопеи (Phr. Eur, 2000, Erythropoietini solutio concentrata, 1316, 780-785) и сравнивают с международным стандартом BRP EPO. Альтернативно, концентрации EPO определяют методом RP-ВЭЖХ с использованием колонки RP-C4 и при измерении уровня адсорбции при длине волны 254 нм с использованием для калибровки 20, 40, 80 и 120 мкг стандартного препарата BRP EPO.
Пример 8
Биологическая активность in vitro HES-модифицированного рекомбинантного человеческого EPO
Активность очищенного HES-модифицированного EPO определяют в рамках биотеста на эритропоэтин, как описано в работе Krystal, 1984, Exp. Heamatol., 11, 649-660.
Анемию индуцируют у мышей NMRI обработкой гидрохлоридом фенилгидразина, собирают клетки селезенки и используют, как описано в работе Fibi et al., 1991, Blood, 77, 1203, ff. Варианты с разбавлениями EPO инкубируют с 3×105 клеток/ячейку в 96-ячеечных микротитрационных планшетах. После инкубирования в течение 24 часов при 37°С в увлажненной атмосфере (5% СО2) клетки в течение 4 часов подвергают мечению 1 мк Ci 3Н-тимидина в расчете на ячейку. Уровень включенной радиоактивности определяют путем счета в жидком сцинтилляторе. Для сравнения используют Международный стандартный препарат EPO (BRP-стандарт).
Альтернативно биологическую активность определяют в тесте in vitro с использованием EPO-чувствительной клеточной линии TF-1 (Kitamura et al., [J. cell Phys., 140. 323-334]). Экспоненциально растущие клетки отмывают от ростовых факторов и инкубируют в присутствии серийных разбавлений EPO еще в течение 48 часов. Пролиферацию клеток оценивают с использованием теста на восстановление, как описано в работе Mosmann, 1983, J.Immunol. Methods, 65, 55-63.
Пример 9
Определение in vivo активности EPO и HAS-модифицированных форм EPO
Определение in vivo активности проводят с использованием нормоцитных мышей путем измерения уровня повышения количества ретикулоцитов через 4 дня после введения животным предварительно определенной дозы EPO или модифицированных форм EPO. Тесты проводят с использованием стандарта BRP EPO, который калибруют против стандартного препарата WHO на EPO в тесте с использованием полицитемических мышей. Образцы EPO разбавляют фосфатно-буферным раствором, содержащим 1 мг/мл бычьего сывороточного альбумина (Sigma).
0,5 мл исследуемого раствора EPO в буферном растворе Дульбекко (который по эквиваленту белка EPO соответствует 100, 80, 40 или 20 МЕ/мл стандартного препарата BRP EPO) инъецируют подкожно мышам. Через 4 дня после инъекции отбирают образцы крови и окрашивают ретикулоциты акридином оранжевым, количественное определение ретикулоцитов проводят методом проточной цитометрии, определяя в целом 30000 клеток крови в течение 5 часов после отбора образца (см. Ph. Eur, 2000, Erythropoietini solutio concentrata, 1316, стр. 780-785 и European Pharmacoppoeia (1996/2000, приложение 2002)).
Пример 10
Определение периода полувыведения in vivo
Кроликам инъецируют внутривенно заданные количества HAS-модифицированной формы EPO. Через заданные интервалы времени отбирают образцы крови и получают сыворотку. Определяют уровень эритропоэтина в сыворотке путем биотеста in vitro или с использованием EPO-специфичного коммерческого набора для ELISA.
Пример 11
Фармакокинетика in vivo
При определении на мышах: Каждому животному вводят подкожно 300 МЕ EPO/кг. Через семь дней после обработки у каждого животного определяют гематокрит. У 9 из всех животных, которым был введен модифицированный EPO, отмечалось существенное повышение гематокрита, что представляет собой ожидаемый результат, определяемый относительно коротким периодом полувыведения необработанного EPO. Средний показатель изменения гематокрита в группе, принимавшей модифицированный EPO, отличался от соответствующего показателя в группе приема необработанного EPO и в контрольной группе.
При исследовании на кроликах: Кроликам вводят однократную дозу немодифицированного или HAS-модифицированного EPO в дозе, соответствующей 200 или до 800 нг/кг массы тела. Через 2, 6, 16, 24 и 48 часов образцы крови анализируют с использованием коммерческого EPO-специфичного набора ELISA для определения концентраций в плазме. Определяют среднее значение концентраций EPO в плазме и вычисляют средние значения исходных показателей периода полувыведения (α-фаза) и конечных показателей периода полувыведения (β-фаза) на основании результата анализа методом ELISA, как описано в работе Zettlmissl et al., 1989, J. Biol. Chem., 264, 21153-21159.
Литература:
Sytkowski, Lunn, Risinger, and Davis, 1999, An Erythropoietin Fusion Protein Comprised of Identical Repeating Domains Exhibitis Enhanced Biological Properties, J. Biol. Chem., 274, 24773-24778.
Пример 12
Оценка биологической активности in vitro HES-модифицированного рекомбинантного человеческого IL-2
Модифицированный IL-2 выделяют гель-фильтрацией в Ультрогеле (Ultrogel AcA 54). Аликвоты соответствующей фракции стерильно фильтруют и определяют биологическую активность IL-2 при использовании IL-2-зависимой клеточной линии клеток мышей (CTLL-2) [Gillis, Ferm, On, and Smith, 1978, J. Immunol., 120, 2027-2032]. Полученное значение активности сравнивают со стандартным препаратом IL-2, используемым в качестве международного эталона.
Пример 13: Образование производных гидроксиэтилкрахмала путем восстановительного аминирования неокисленного восстановленного конца
Пример 13.1. Реакция гидроксиэтилкрахмала с 1,3-диамино-2-гидроксипропаном

а) К раствору 200 мг гидроксиэтилкрахмала (HES18/0,4 (М.м. = 18000 Да, DS = 0,4)) в 5 мл воды добавляют 0,83 ммоль 1,3-диамино-2-гидроксипропана и 50 мг цианборгидрата натрия NaCNBH3. Полученную смесь инкубируют при 80°С в течение 17 часов. Реакционную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1 (объем/объем). Осадок собирают центрифугированием, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
b) Инкубирование смеси, полученной при добавлении 0,83 ммоль 1,3-диамино-2-гидроксипропана и 50 мг цианборгидрата натрия NaCNBH3 к раствору 200 мг гидроксиэтилкрахмала также возможно и проводится при 25°С в течение 3 дней.
Пример 13.2. Реакция гидроксиэтилкрахмала с 1,2-дигидрокси-3-аминопропаном
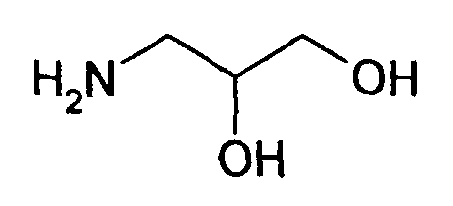
а) К раствору 200 мг гидроксиэтилкрахмала (HES18/0,4 (М.м. = 18000 Да, DS = 0,4)) в 5 мл воды добавляют 0,83 ммоль 1,2-дигидрокси-3-аминопропана и 50 мг цианборгидрата натрия NaCNBH3. Полученную смесь инкубируют при 80°С в течение 17 часов. Реакционную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1 (объем/объем). Осадок собирают центрифугированием, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Осуществление реакции 1,2-дигидрокси-3-аминопропана с HES подтверждается опосредованно при определении уровня формальдегида, получаемого при окислительном расщеплении 1,2-диола в реакционном продукте перйодатом, как описано G. Avigad, Anal. Biochem. 134 (1983) 449-504.
b) Инкубирование смеси, полученной при добавлении 0,83 ммоль 1,2-дигидрокси-3-аминопропана и 50 мг цианборгидрата натрия NaCNBH3 к раствору 200 мг гидроксиэтилкрахмала, также возможно и проводится при 25°С в течение 3 дней.
Пример 13.3. Реакция гидроксиэтилкрахмала с 1,4-диаминобутаном
а) К раствору 200 мг гидроксиэтилкрахмала (HES18/0,4 (М.м. = 18000 Да, DS = 0,4)) в 5 мл воды добавляют 0,83 ммоль 1,4-диаминобутана и 50 мг цианборгидрата натрия NaCNBH3. Полученную смесь инкубируют при 80°С в течение 17 часов. Полученную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1 (объем/объем). Осадок собирают центрифугированием, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
b) Инкубирование смеси, полученной при добавлении 0,83 ммоль 1,4-диаминобутана и 50 мг цианборгидрата натрия NaCNBH3 к раствору 200 мг гидроксиэтилкрахмала, также возможно и проводится при 25°С в течение 3 дней.
Пример 13.4. Реакция гидроксиэтилкрахмала с 1-меркапто-2-аминоэтаном
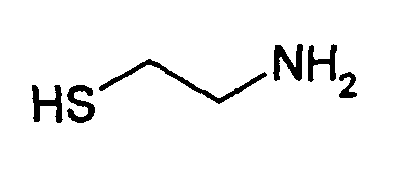
а) К раствору 200 мг гидроксиэтилкрахмала (HES18/0,4 (М.м. = 18000 Да, DS = 0,4)) в 5 мл воды добавляют 0,83 ммоль 1-меркапто-2-аминоэтана и 50 мг цианборгидрата натрия NaCNBH3. Полученную смесь инкубируют при 80°С в течение 17 часов. Реакционную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1 (объем/объем). Осадок собирают центрифугированием, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
b) Инкубирование смеси, полученной при добавлении 0,83 ммоль 1-меркапто-2-аминоэтана и 50 мг цианборгидрата натрия NaCNBH3 к раствору 200 мг гидроксиэтилкрахмала, также возможно и проводится при 25°С в течение 3 дней.
Пример 14: Образование производных гидроксиэтилкрахмала путем конъюгирования с неокисленным восстановленным концом
Пример 14.1 Реакция гидроксиэтилкрахмала с карбогидразидом
0,96 г HES18/0,4 (М.м. = 18000 Да, DS = 0,4) растворяют в 8 мл водного 0,1 М натрий-ацетатного буфера, pH 5,2 и добавляют 8 ммоль карбогидразида (Sigma, Aldrich, Taufkirchen, D). После перемешивания в течение 18 часов при 25°С реакционную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1 (объем/объем). Осажденный продукт собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 3 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 14.2 Реакция гидроксиэтилкрахмала с адипиновым дигидразидом
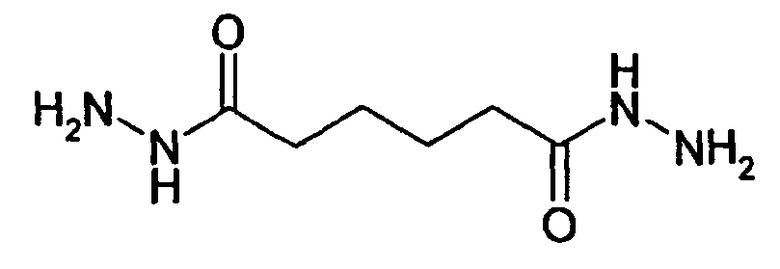
0,96 г HES18/0,4 (М.м. = 18000 Да, DS = 0,4) растворяют в 8 мл водного 0,1 М натрий-ацетатного буфера, pH 5,2 и добавляют 8 ммоль адипинового дигидразида (Lancaster Synthesis, Frankfurt/Main, D). После перемешивания в течение 18 часов при 25°С реакционную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1, (объем/объем). Осажденный продукт собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 3 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 14.3 Реакция гидроксиэтилкрахмала с 1,4-фенилен-бис-3-тиосемикарбазидом

0,96 г HES18/0,4 (М.м. = 18000 Да, DS = 0,4) растворяют в 8 мл водного 0,1 М натрий-ацетатного буфера, pH 5,2 и добавляют 8 ммоль 1,4-фенилен-бис-3-тиосемикарбазида (Lancaster Synthesis, Frankfurt/Main, D). После перемешивания в течение 18 часов при 25°С к реакционной смеси добавляют 8 мл воды и суспензию центрифугируют в течение 15 минут при 4500 об/мин. Прозрачный супернатант декантируют и затем добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1, (объем/объем). Осажденный продукт собирают центрифугированием, перерастворяют в 40 мл воды и центрифугируют в течение 15 минут при 4500 об/мин. Прозрачный супернатан диализуют в течение 3 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 14.4: Реакция гидроксиэтилкрахмала с О-[2-(2-аминоокси-этокси)этил]гидроксиламином
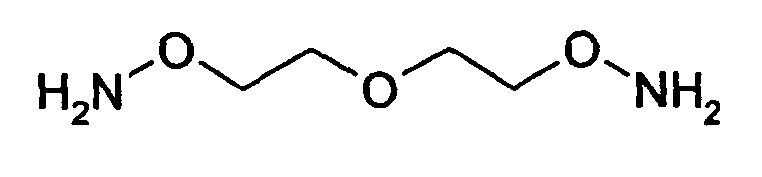
О-[2-(2-аминооксиэтокси)этил]гидроксиламин синтезируют по методу Boturyn et al., Tetrahedron 53 (1997) p.5485-5492, в 2 стадии из коммерчески доступного материала.
0,96 г HES18/0,4 (М.м. = 18000 Да, DS = 0,4) растворяют в 8 мл водного 0,1 М натрий-ацетатного буфера, pH 5,2 и добавляют 8 ммоль О-[2-(2-аминоокси-этокси)этил]гидроксиламина. После перемешивания в течение 18 часов при 25°С реакционную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1 (объем/объем). Осажденный продукт собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 3 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 15. Образование гидроксиэтилкрахмальных производных при реакции с восстановленным концом при его окислении
Пример 15.1. Реакция гидроксиэтилкрахмала с карбогидразидом

0,12 ммоль оксо-HES 10/0,4 (М.м. = 10000 Да, DS = 0,4, получен в соответствии с DE 19628705 1А) растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 15 ммоль карбогидразида (Sigma, Aldrich, Taufkirchen, D) в 15 мл ДМСО. После перемешивания в течение 88 часов при 65°С реакционную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1, (объем/объем). Осажденный продукт собирают центрифугированием, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 15.2. Реакция гидроксиэтилкрахмала с 1,4-фенилен-бис-3-тиосемикарбазидом
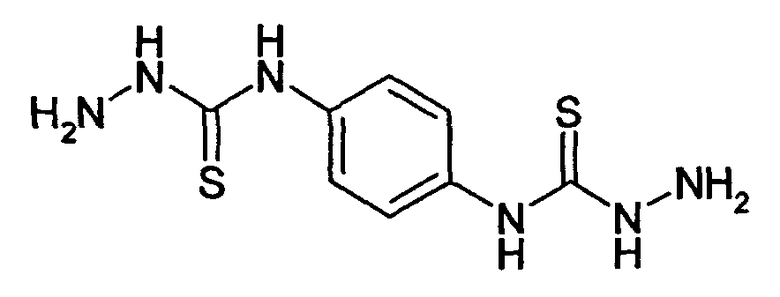
0,12 ммоль оксо-HES 10/0,4 (М.м. = 10000 Да, DS = 0,4, получают в соответствии с DE 19628705 1А) растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 15 мл 1,4-фенилен-бис-3-тиосемикарбазида (Lancaster Synthesis, Frankfurt/Main, D) в 15 мл ДМСО. После перемешивания в течение 88 часов при 65°С реакционную смесь добавляют к 160 мл охлажденной смеси ацетона и этанола, 1:1, (объем/объем). Осажденный продукт собирают центрифугированием, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 15.3. Реакция гидроксиэтилкрахмала с гидразином

1,44 г (0,12 ммоль) оксо-HES 10/0,4 (М.м. = 10000 Да, DS = 0,4, получают в соответствии с DE 19628705 1А) растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 0,47 мл (15 ммоль) гидразина в 15 мл ДМСО. После перемешивания в течение 19 часов при 40°С реакционную смесь добавляют к 160 мл смеси ацетона и этанола, 1:1, (объем/объем). Осажденный продукт собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 2 дней против 0,5% (объем/объем) триэтиламина в водном растворе в течение 2 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 15.4. Реакция гидроксиэтилкрахмала с гидроксиламином
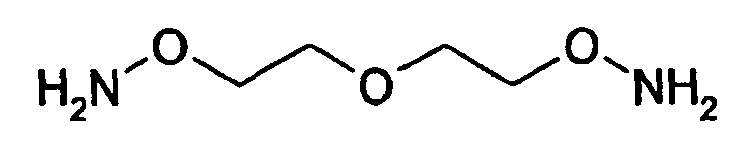
О-[2-(2-аминооксиэтокси)этил]гидроксиламин синтезируют по методике Boturyn et al. в 2 стадии из коммерчески доступного материала (Boturyn, Boudali, Constant, Defrancq, Lhomme, 1997, Tetrahedron 53, 5485).
1,44 г (0,12 ммоль) оксо-HES 10/0,4 (М.м. = 10000 Да, DS = 0,4, получают в соответствии с DE 19628705 1А) растворяют в 3 мл абсолютного диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 2,04 г (15 ммоль) О-[2-(2-аминооксиэтокси)этил]гидроксиламина в 15 мл ДМСО. После перемешивания в течение 48 часов при 65°С реакционную смесь добавляют к 160 мл смеси этанола и ацетона, 1:1, (объем/объем). Осажденный продукт собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 15.5. Реакция гидроксиэтилкрахмала с адипиновым дигидразидом
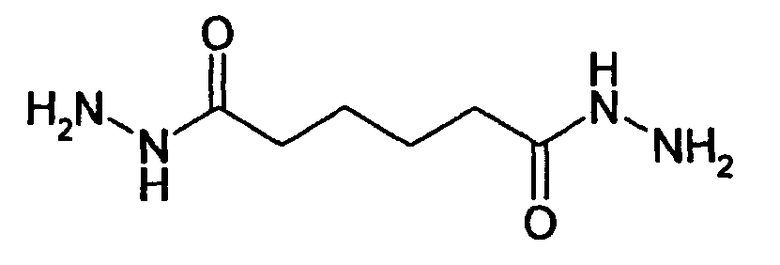
1,74 г (15 ммоль) адипинового дигидразида растворяют в 20 мл абсолютного диметилсульфоксида (ДМСО) при 65°С и добавляют по каплям в атмосфере азота 1,44 г (0,12 ммоль) оксо-HES 10/0,4 (М.м. = 10000 Да, DS = 0,4, получают в соответствии с DE 19628705 1А), растворенного в 3 мл абсолютного ДМСО. После перемешивания в течение 68 часов при 60°С реакционную смесь добавляют к 200 мл воды. Раствор, содержащий реакционный продукт, диализуют в течение 2 дней против 0,5% (объем/объем) триэтиламина в водном растворе и в течение 2 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 15.6. Реакция гидроксиэтилкрахмала с 1,4-диаминобутаном
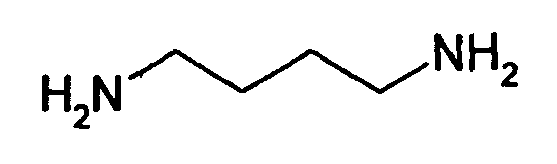
1,44 г (0,12 ммоль) оксо-HES 10/0,4 (М.м. = 10000 Да, DS = 0,4, получают в соответствии с DE 19628705 1А) растворяют в 3 мл сухого диметилсульфоксида (ДМСО) и добавляют по каплям в атмосфере азота к смеси 1,51 мл (15 ммоль) 1,4-диаминобутана в 15 мл ДМСО. После перемешивания в течение 19 часов при 40°С реакционную смесь добавляют к 160 мл смеси этанола и ацетона, 1:1, (объем/объем). Осадок амино-HES10KD/0,4 собирают центрифугированием, перерастворяют в 40 мл воды, диализуют в течение 4 дней против воды (трубка для диализа SnakeSkin, с границей отсечения молекулярной массы на уровне 3,5 кДа, Perbio Science, Deutschland GmbH, Bonn, Germany) и лиофилизируют.
Пример 16. Окисление эритропоэтина
Окисленный эритропоэтин получают по методике, описанной в примере 20. В качестве окисленного эритропоэтина используют EPO-GT-1-A, описанный в примере 20.11 (с) (EPO-GT-1, полученный без кислотного гидролиза путем обработки в условиях мягкого окисления перйодатом).
Пример 17: Конъюгирование гидроксиэтилкрахмальных производных с окисленным эритропоэтином из примера 4
Пример 17.1. Реакция окисленного эритропоэтина с реакционным продуктом из примера 14.1
Окисленный EPO (1,055 мкг/мкл) в 20 мМ буфера PBS доводят до значения pH 5,3 с использованием 5 М натрий-ацетатного буфера, pH 5,2. К 19 мкл раствора EPO добавляют 18 мкл раствора производного HES, полученного по процедуре примера 14.1 (M.м. 18 кДа; 18,7 мкг/мкл в 0,1 М натрий-ацетатного буфера, pH 5,2), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт очищают в SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях от компании Инвитроген (Invitrogen). Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента проиллюстрирован на фиг.3. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и количеством производных HES, присоединенных к белку.
Пример 17.2. Реакция окисленного эритропоэтина с реакционным продуктом из примера 14.3
Окисленный EPO (1,055 мкг/мкл) в 20 мМ буфера PBS доводят до значения pH 5,3 с использованием 5 М натрий-ацетатного буфера, pH 5,2. К 19 мкл раствора EPO добавляют 18 мкл раствора производного HES, полученного по процедуре примера 14.3 (М.м. 18 кДа; 18,7 мкг/мкл в 0,1 М натрий-ацетатного буфера, pH 5,2) и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген (Invitrogen, Carlcbad, CA, USA).
Пример 17.3. Реакция окисленного эритропоэтина с реакционным продуктом из примера 14.4
Окисленный EPO (1,055 мкг/мкл) в 20 мМ буфера PBS доводят до значения pH 5,3 с использованием 5 М натрий-ацетатного буфера, pH 5,2. К 19 мкл раствора EPO добавляют 18 мкл раствора производного HES, полученного по процедуре примера 14.4 (М.м. 18 кДа; 18,7 мкг/мкл в 0,1 М натрий-ацетатного буфера, pH 5,2) и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.4. Удачный результат конъюгирования показан в виде перемещения белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 17.4. Реакция окисленного эритропоэтина с продуктом реакции из примера 15.1
Окисленный EPO (1,055 мкг/мкл) в 20 мМ буфера PBS доводят до значения pH 5,3 с использованием 5 М натрий-ацетатного буфера, pH 5,2. К 19 мкл раствора EPO добавляют 18 мкл раствора производного HES, полученного по процедуре примера 15.1 (М.м. 10 кДа; 18,7 мкг/мкл в 0,1 М натрий-ацетатного буфера, pH 5,2), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.5. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 17.5. Реакция окисленного эритропоэтина с реакционным продуктом из примера 15.2
Окисленный EPO (1,055 мкг/мкл) в 20 мМ буфера PBS доводят до значения pH 5,3 с помощью 5 М натрий-ацетатного буфера, pH 5,2. К 19 мкл раствора EPO добавляют 18 мкл раствора производного HES, полученного по процедуре примера 15.1 (М.м. 10 кДа; 18,7 мкг/мкл в 0,1 М натрий-ацетатного буфера, pH 5,2) и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.5. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением молекулярного веса используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 18. Образование тио-EPO путем восстановления эритропоэтина
241,5 мкг эритропоэтина (EPO-GT-1, см. пример 20) в 500 мкл 0,1 М натрий-боратного буфера, 5 мМ ЭДТА, 10 мМ ДТТ (Lancaster, Morcambe, UK), pH 8,3, инкубируют в течение 1 часа при 37°С. ДТТ удаляют фильтрованием при центрифугировании при использовании концентратора на 0,5 мл VIVASPIN, 10 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с последующей промывкой 3 раза боратным буфером и дважды фосфатным буфером (0,1 M, 9,15 M NaCl, 50 мМ ЭДТА, pH 7,2).
Пример 19. Конъюгирование производных гидроксиэтилкрахмала с тиоэритропоэтиом с использованием сшивающего соединения
В каждом из приведенных ниже примеров в качестве сшивающего соединения используют сложный N-(альфа-малеимидоацетокси)сукцинимидный эфир (АМАС; AMAS).
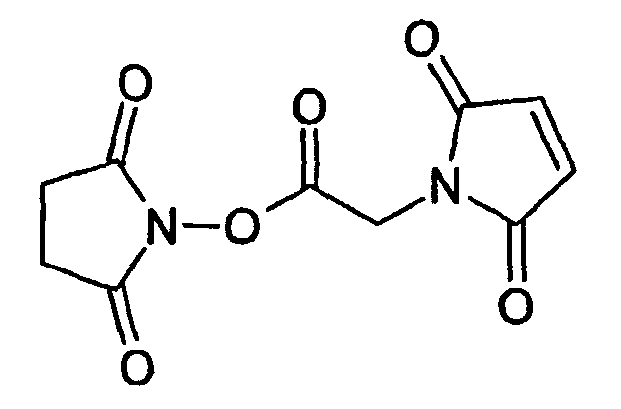
Пример 19.1. Реакция тиоэритропоэтина с реакционным продуктом из примера 14.1 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 14.1 и растворенного в 200 мкл 0,1 М натрий-фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2) добавляют 10 мкл 2,5 мкмоль раствора AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО. Прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. Оставшийся AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут фосфатным буфером.
К оставшемуся раствору добавляют 15 мкг тио-ЕРО, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25 С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.6. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.2. Реакция тиоэритропоэтина с реакционным продуктом из примера 14.2 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 14.2 и растворенного в 200 мкл 0,1 М натрий-фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО. Прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. Оставшийся AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-ЕРО, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере) и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют в SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.7. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярному веса используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.3. Реакция тиоэритропоэтина с реакционным продуктом из примера 14.3 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 14.3 и растворенного в 200 мкл 0,1 М натрий-фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО. Прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. Оставшийся AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-ЕРО, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере) и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.6. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.4. Реакция тиоэритропоэтина с реакционным продуктом из примера 14.4 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 14.4 и растворенного в 200 мкл 0,1 М натрий-фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО. Прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. Оставшийся AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-ЕРО, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере) и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.6. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.5. Реакция тиоэритропоэтина с реакционным продуктом из примера 13.1 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 13.1, при инкубации в условиях, включающих 80°С в течение 17 часов, а также 25°С в течение 3 дней, и растворенного в 200 мкл 0,1 М натрий-фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2) добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО. Прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. Оставшийся AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-EPO, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.7. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярномассы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.6. Реакция тиоэритропоэтина с реакционным продуктом из примера 13.3 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 13.3, при инкубации в условиях, включающих 80°С в течение 17 часов, а также 25°С в течение 3 дней, и растворенного в 200 мкл 0,1 М натрий-фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО. Прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. Оставшийся AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-EPO, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.7. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.7. Реакция тиоэритропоэтина с реакционным продуктом из примера 15.1 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 15.1 и растворенного в 200 мкл фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО и прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-EPO, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.8. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.8. Реакция тиоэритропоэтина с реакционным продуктом из примера 15.2 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 15.2, растворенного в 200 мкл фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО и прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-EPO, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.8. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.9. Реакция тиоэритропоэтина с реакционным продуктом из примера 15.3 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 15.3 и растворенного в 200 мкл фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО и прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-EPO, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.8. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.10. Реакция тиоэритропоэтина с реакционным продуктом из примера 15.4 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 15.4 и растворенного в 200 мкл фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО и прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-EPO, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPs (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.8. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.11. Реакция тиоэритропоэтина с реакционным продуктом из примера 15.5 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 15.5 и растворенного в 200 мкл фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО и прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-EPO, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.8. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 19.12. Реакция тиоэритропоэтина с реакционным продуктом из примера 15.6 и сшивающим соединением
К 50 нмоль HES-производного, полученного по методике примера 15.6 и растворенного в 200 мкл фосфатного буфера (0,1 М, 9,15 М NaCl, 50 мМ ЭДТА, pH 7,2), добавляют 10 мкл раствора 2,5 мкмоль AMAS (Sigma, Aldrich, Taufkirchen, D) в ДМСО и прозрачный раствор инкубируют в течение 80 минут при температуре 25°С и 20 минут при 40°С. AMAS удаляют фильтрованием при центрифугировании с использованием концентратора на 0,5 мл VIVASPIN, 5 кДа MWCO (VIVASCIENCE, Hannover, D) при 13000 об/мин с промывкой 4 раза в течение 30 минут в фосфатном буфере.
К оставшемуся раствору добавляют 15 мкг тио-EPO, полученного по методике примера 18 (1 мкг/мкл в фосфатном буфере), и смесь инкубируют в течение 16 часов при 25°С. После лиофилизации неочищенный продукт анализируют методом SDS-PAGE с использованием буфера NuPAGE 10% Bis-Tris гели/MOPS (Invitrogen, Carlsbad, CA, USA), как описано в инструкциях компании Инвитроген. Гель окрашивают красящим реактивом кумасси (Roti-Blue) (Roth, Karlsruhe, D) в течение ночи.
Результат эксперимента приведен на фиг.8. Удачный результат конъюгирования показан в виде миграции белковой полосы в сторону более высокой молекулярной массы. Расширение полосы связано с распределением по молекулярной массе используемых производных HES и с числом производных HES, присоединенных к белку.
Пример 20. Препаративное получение конъюгатов HES-EPO
Краткое описание
Конъюгаты HES-EPO синтезируют путем связывания HES-производных (среднее значение М.м. составляет 18000 Дальтон; степень замещения гидроксиэтилом составляет 0,4) с частично (в условиях мягкой обработки перйодатом) окисленными остатками сиаловой кислоты на олигосахаридных цепях рекомбинантного человеческого EPO. На основании структурного анализа углеводов было показано, что введенные модификации не воздействуют на структурную целостность ядерных олигосахаридных цепей, поскольку анализ методом MALDI/TOF-MS HES-модифицированных гликанов в условиях мягкой обработки кислотой выявил наличие интактных нейтральных цепей N-ацетиллактозаминового типа, которые не отличались от соответствующих цепей, наблюдаемых в немодифицированном EPO-продукте. Полученный результат указывает на то, что по меньшей мере 3 модифицированных HES-остатка присоединяются к молекуле EPO в препарате EPO, который подвергается модификации без предшествующего частичного удаления сиаловой кислоты. Вариант EPO, потерявший примерно 50% остатков сиаловой кислоты относительно предшествующего белка, демонстрирует аналогичную мобильность в SDS-PAGE, явно указывающую на высокую молекулярную массу (60-110 кДа против 40 кДа, характерного для стандарта BRP EPO). HES-модифицированный EPO стабилен в стандартных условиях ионообменой хроматографии при комнатнй температуре и pH 3-10.
Биотест на EPO в системе нормоцитных мышей указывает на то, что HES-модифицированный EPO обладает в 2,5-3,0 раза более высокой удельной активностью (МЕ/мг) в данном тесте в сравнении с международным эталонным стандартом BRP EPO, что следует из результатов определения белка по величине поглощения в УФ области в соответствии с методом Европейской Фармакопеи, и результатов определения белка EPO методом RP-ВЭЖХ, калиброванным относительно стандартного препарата BRP EPO.
Пример 20.1. Материалы и методы
(а) Высвобождение N-связанных олигосахаридов путем расщепления N-гликозидазой
Образцы инкубируют с 25 единицами (в соответствии со спецификацией от производителя, Roche Diagnostics, Germany) рекомбинантной PNG-азы F в течение ночи при 37°С. Полноту расщепления отслеживают по специфическому изменению мобильности белка в SDS-PAGE. Высвобожденные N-гликаны отделяют от полипептида путем добавления 3 объемов холодного 100% этанола с последующей инкубацией при -20°С в течение по меньшей мере 2 часов (Schroeter S et. al. 1999). Осажденный белок удаляют центрифугированием в течение 10 минут при 4°С со скоростью 13000 об/мин. Полученный осадок затем промывают два раза с использованием 500 мкл охлажденного льдом 75% этанола. Олигосахариды в объединенных супернатантах сушат в вакуумной центрифуге (концентратор Speed Vac, Savant Instruments Inc., USA). Образцы гликана перед использованием обессоливают с помощью картриджей Hypercarb (25 мг или 100 мг HyperCarb) следующим образом: колонки промывают 3×500 мкл 80% ацетонитрила (объем/объем) в 0,1% TFA с последующими промывками 3×500 мкл воды. Образцы разбавляют водой до конечного объема 300-600 мкл перед внесением в картридж, который затем тщательно промывают водой. Олигосахариды элюируют с использованием 1,2 мл (картриджи на 25 мг; берут 1,8 мл в случае картриджей на 100 мг) 25% ацетонитрила в воде, содержащей 0,1% трифторуксусной кислоты (объем/объем). Элюированные олигосахариды нейтрализуют с использованием 2 M NH4OH и сушат в концентраторе Speed Vac. В некоторых случаях обессоливание олигосахаридов, высвобожденных N-гликозидазой, проводят путем абсорбции расщепленной смеси из образцов, содержащих <100 мкг общего (глико)протеина, на картриджах Hypercarb на 100 мг.
(b) Анализ олигосахаридов с помощью лазерной десорбции на основе матричной/ионизационной масс-спектрометрии по времени пролета (MALDI/TOF/TOF-MS)
Используют прибор Bruker ULTRAFLEX для анализа по времени пролета (TOF-TOF): нативные десиалилированные олигосахариды анализируют с использованием 2,5-дигидроксибензойной кислоты в качестве УФ-поглощающего материала как в варианте положительного, так и отрицательного иона с использованием в обоих случаях рефлектрона. В случае анализов по методу MS-MS выбранные парные ионы подвергают индуцированной лазером диссоциации (LID) и полученные фрагменты ионов разделяют с помощью второго блока TOF прибора (LIFT). Образец растворов по 1 мкл в соответствующей концентрации 1-10 пмоль/мкл смешивают с равным количеством соответствующей матрицы. Указанную смесь наносят капельно на мишень из нержавеющей стали и сушат при комнатной температуре перед проведением анализа.
Пример 20.2. Получение и характеристика рекомбинантного человеческого EPO (EPO-GT-1)
EPO получают из рекомбинантных CHO-клеток при его экспрессии в них, как описано в литературе (Mueller PP et al., 1999, Dorner AJ et al., 1984), и исследуют характеристики полученных препаратов в соответствии с методами Европейской Фармакопеи (Ph. Eur. 4, Monography 01/2002:1316: Erythropoietin concentrated solution). Конечный продукт характеризуется содержанием сиаловой кислоты на уровне 12 нмоль (±1,5 нмоль) на нмоль белка. Определяют структуры N-связанных олигосахаридов методом HPAEC-PAD и MALDI/TOF-MS, как описано в литературе (Nimtz et al., 1999, Grabenhorst, 1999). Полученные EPO препараты содержат ди-, три- и тетрасиалилированные олигосахариды (2-12%, 15-28% и 60-80% соответственно, сульфатированные и пентасиалилированные цепи присутствуют в небольших количествах). Было показано, что в целом характеристики гликозилирования препаратов EPO аналогичны таковым для международного стандартного препарата BRP EPO.
Результаты изоэлектрофокусирования рекомбинантного EPO были сравнимы с результатами соответствующего анализа международного стандартного препарата BRP EPO, которые демонстрировали соответствующие изоформы. У 25% белка EPO отсутствовало О-гликозилирование в положении Ser126 в полипептидой цепи.
Пример 8.3. Получение частично десиалилированных форм EPO
Белок EPO-GT-1 (2,84 мг/мл) нагревают до 80°С в 20 мМ Na-фосфатном буфере, pH 7,0 и затем добавляют 100 мкл 1 н. H2SO4 в расчете на 1 мл раствора, инкубацию продолжают в течение 5 минут, 10 минут и 60 минут соответственно, что приводит к получению препаратов EPO с разной степенью сиалилирования. Количественное определение олигосахаридов с наличием 0-4 сиаловых кислот проводят после высвобождения олигосахаридов N-гликозидазой полипептидов и выделение N-связанных цепей проводят при обессоливании с использованием картриджей Hypercarb (25 мг HyperSep Hypercarb; Thermo-Hypersil-Keystone, UK). Препараты EPO нейтрализуют путем добавления 1 н. NaOH и затем замораживают в жидком N2 и хранят при температуре -20°С до дальнейшего использования.
Пример 20.4. Окисление перйодатом сиалилированных форм EPO
К 10 мг необработанного или обработанного кислотой в мягких условиях EPO, который растворен в 3,5 мл 20 мМ Na-фосфатного буфера, pH 7,0, добавляют 1,5 мл 0,1 M Na-ацетатного буфера, pH 5,5 и смесь охлаждают до 0°С на ледяной бане; добавляют 500 мкл 10 мМ перйодата Na и реакционную смесь выдерживают в темноте в течение 60 минут при 0°C. Затем добавляют 10 мкл глицерина и инкубацию продолжают еще в течение следующих 10 минут в темноте. Частично окисленные формы EPO отделяют от реагентов путем обессоливания с использованием концентраторов VIVASPIN (10000 MWCO, PES Vivascience AG, Hannover, Germany) в соответствии с рекомендациями производителя со скоростью 3000 об/мин в лабораторной центрифуге, снабженной фиксированный угловым ротором. После замораживания в жидком азоте препараты EPO хранят в конечном объеме 4 мл при температуре -20°С.
Аликвоты частично окисленного препарата EPO по 100 мкг подвергают обработке N-гликозидазой и выделяют олигосахариды с использованием картриджей Hypercarb, как описано. Олигосахариды подвергают десиалилированию путем мягкой обработки кислотой и анализируют методом HPAEC-PAD, сравнивая полученное значение времени удерживания с соответствующими показателями аутентичных препаратов стандартных олигосахаридов, как описано в литературе (Nimtz et al., 1990 и 1993).
Пример 20.5. Восстановление дисульфидов EPO с дитиоэритрейтолом
5 мг EPO-GT-1 инкубируют с 5 мл 0,1 М Трис/HCl буфера, pH 8,1, в присутствии 30 мМ дитиоэритрейтола (ДТТ) при 37°С в течение 60 минут, удаление ДТТ достигается путем использования концентратора Vivaspin при 4°С с проведением 4 циклов замены буфера. Конечный восстановленный препарат EPO замораживают в жидком азоте и хранят при -20°С в 50 мМ Na-ацетатном буфере, pH 5,5.
Пример 20.6. Анализ препарата EPO
Количественное определение белка в препарате EPO проводят путем измерения УФ поглощения при длине волны 280 нм в соответствии с методикой Европейской Фармакопеи (European Pharmacopeia 4, Monography 01/2002; 1316: Erythropoietin concentrated solution) в кювете с длиной пробега 1 см. Кроме того, количественное EPO проводят методом OA-ВЭЖХ с использованием колонки RP-C4 (Vydac Protein C4, Cat#214TP5410, Grace Vydac, Ca, US); методика ВЭЖХ была откалибрована с использованием эталонного препарата эритропоэтина BRP 1 (European Pharmacopeia, Conseil, de l'Europe B.P. 907-F67029, Strasbourg Cedex 1).
Пример 20.7. Окисление десиалилированного EPO галактозооксидазой
4,485 мг полностью десиалилированого EPO инкубируют в 20 мМ Na-фосфатном буфере, pH 6,8, в присутствии 16 мкл каталазы (6214 единиц/200 мл) и 80 мкл галактозооксидазы (2250 единиц/мл из Dactylium dendroides (Sigma-Aldrich, Steinheim, Germany)); инкубацию проводят при 37°С в течение ночи; добавляют 2 раза по 20 мкл галактозооксидазы через 4 часа и через 8 часов после начала инкубации.
Пример 20.8. Подготовка образцов EPO для биотестирования
Выделение и очистка образцов EPO из инкубационных сред белковых препаратов EPO, обработанных перйодатом или окисленных галактооксидазой, с активированным HES
Очистку препаратов EPO (удаление непрореагировавших производных HES) проводят при комнатной температуре. Смеси для инкубации EPO (примерно 5 мг по белку EPO) разбавляют 1:10 буфером А (20 мМ N-морфолинпропансульфоновой кислоты [MOPS/NaOH] в бидистиллированной воде, pH 8,0) и наносят на колонку, содержащую 3 мл сефарозы (Q-Sepharose HP (Pharmacia Code no. 17-1014-03m Lot no. 220211)), уравновешенную буфером А в количестве, равном 10 объемам колонки (CV), со скоростью течения 0,5 мл/мин. Колонку промывают 6-8 CV буфера А (скорость течения = 0,8 мл/мин) и проводят элюцию с использованием буфера B (20 мМ морфолинэтансульфоновой кислоты [MES/NaOH], 0,5 M NaCl в бидистиллированной воде, pH 6,5) со скоростью течения 0,5 мл/мин. EPO определяют по УФ поглощению при длине волны 280 нм и элюируют примерно в 6 мл. Колонку регенерируют с использованием 3 CV буфера С (20 мМ MES, 1,5 M NaCl в воде, доведенной до pH 6,5) и подвергают повторному уравновешиванию с использованием 10 CV буфера А (скорость течения 0,7 мл/мин).
Буферную замену в элюатах EPO, полученных на стадии элюции с Q-Sepharose, проводят с помощью концентратора Vivaspin и фосфатно-буферного раствора (PBS) в течение 3 циклов центрифугирования на образец; объем образцов доводят до 2 мл с помощью PBS, после чего образцы хранят при -20°С.
Получают лишь <25% частично десиалилированных и впоследствии окисленных в условиях мягкой обработки перйодатом форм EPO, которые подвергались HES-модификации из элюата с колонок с сефарозой (Q-Sepharose), поскольку в используемых условиях основные формы EPO не связываются с сефарозой (Q-Sepharose) и обнаруживаются в протекающем потоке вместе с непрореагировавшими HES-производными.
Пример 20.9. Анионообменная хроматография при высоком pH с пульсовым амперометрическим обнаружением (HPAEC-PAD)
Очищенные нативные и десиалилированные олигосахариды анализируют с помощью анионообменной хроматографии с высоким pH (HPAE) в системе Dionex BioLC (Dionex, USA), снабженной колонкой CarboPac PA1 (0,4×25 см) в сочетании с системой пульсового амперометрического выявления (PAD) (Schroter et al., 1999; Nimtz et al., 1999). Значения потенциалов детектора (Е) и длительность пульсовой обработки (Т) составляют: Е1: +50 мВ, Т1: 480 мсек; Е2: +500 мВ, Т2: 120 мсек; Е3: -500 мВ, Т3: 60 мсек, а диапазон выхода составляет 500-1500 нА. Затем олигосахариды инъецируют в колонку CarboPac PА1, уравновешенную 100% растворителем А. В случае элюции десиалилированных олигосахаридов (скорость течения: 1 мл/мин) указанную элюцию проводят при использовании линейного градиента (0-20%) растворителя B в течение 40 минут с последующим линейным повышением уровня растворителя В от 20 до 100% в течение 5 минут. Растворитель А представляет собой 0,2 М NaOH в бидистиллированной воде, растворитель В состоит из 0,6 М NaOAc в растворителе А. В случае нативных олигосахаридов колонку уравновешивают 100% растворителем С (0,1 М NaOH в бидистиллированной воде) и элюцию (скорость течения: 1 мл/мин) проводят с использованием линейного градиента (0-35%) растворителя D в течение 48 минут с последующим линейным повышением уровня растворителя D от 35 до 100% в течение 10 минут. Растворитель D представляет собой 0,6 М NaAc в растворителе С.
Пример 20.10. Анализ моносахаридного состава N-гликанов, HES-модифицированных N-гликанов и белка EPO методом GC-MS
Моносахариды анализируют в виде соответствующих метилгликозидов после проведения метанолиза, N-реацетилирования и триметилсиалилирования по методу GC-MS [Chaplin, M. F. (1982) A rapid and sensitive method for analysis of carbohydrate. Anal. Biochem. 123, 336-341]. Анализы проводят в масс-спектрометре, действующем по механизму захвата ионов, Finnigan GCQ (Finnigan MAT corp., San Jose, CA), работающем в режиме положительных ионов EI, который снабжен капиллярной колонкой DB5 длиной 30 м. Прибор работает по следующей температурной программе: 2 минуты изотерма при 80°С, затем температуру повышают со скоростью 10 градусов/мин до 300°С.
Моносахариды идентифицируют по значению времени удерживания и показателям характерных фрагментов. Для количественного определения используют некорректированные результаты интеграции электронных пиков. Моносахарид, дающий более чем один пик в связи с аномерностью и/или присутствием фураноидных и пираноидных форм, количественно определяют путем добавления всех основных пиков. Используют 0,5 мкг миоинозита в качесте внутреннего стандартного соединения.
Пример 20.11 Результаты
Пример 29.11(а) Характеристика N-гликанов в EPO-GT-1 после его обработки кислотой в мягких условиях (частично десиалилированного)
Препараты EPO-GT-1 подвергают обработке кислотой в мягких условиях в течение 5, 10 или 60 минут и затем анализируют в SDS-PAGE до и после высвобождения N-связанных олигосахаридов при инкубации с N-гликозидазой, как показано на фиг.9. N-связанные олигосахариды подвергают картированию олигосахаридов по методу HPAEC-PAD (фиг.10). Необработанный ЕРО-GT-1 содержит более 90% N-связанных олигосахаридов с 3 или 4 остатками сиаловой кислоты, тогда как после 5-минутной инкубации в присутствии слабой кислоты менее 40% углеводных цепей содержат 3 или 4 остатка сиаловой кислоты. Анализ по методу HPAEC-PAD десиалилированных N-гликанов показал, что соотношение нейтральных олигосахаридов, которые были обнаружены в необработанном ЕРО-GT-1, остается стабильным в препаратах, подвергнутых кислой обработке в течение 5, 10 или 60 минут. Анализ методом MALDI/TOF-MS десиалилированных гликанов показал, что менее 90% проксимальной фукозы присутствует после обработки слабой кислотой данного белка.
Пример 20.11(b) Характеристика ЕРО-GT-1, обработанного перйодатом
На фиг.12 приведены данные, показывающие мобильность в SDS-PAGE обработанных в мягких условиях перйодатом форм EPO, которые были предварительно подвергнуты обработке кислотой в течение 5 и 10 минут или не обрабатывались. Условия, используемые для перйодатного окисления сиаловых кислот, не влияют на поведение препаратов EPO в SDS-PAGE (в сравнении с фиг.9). Окисление сиаловых кислот приводит к смещению олигосахаридов в рамках анализа HPAEC-PAD, к более ранним показателям по времени элюирования (для сравнения см. фиг.10 и 13).
Пример 20.11(с) Характеристика HES-модифицированных производных EPO
(аа) HES-модификация с течением времени ЕРО-GT-1-А с использованием модифицированного гидроксиламином HES-производного Х, полученного по методике примера 14.4
400 мкг модифицированного гидроксиламином HES-производного Х добавляют к 20 мкг ЕРО-GT-1-А (EPO, окисленный в мягких условиях перйодатом, но не подвергнутый кислотному гидролизу перед указанным окислением перйодатом в мягких условиях) в 20 мкл 0,5 М NaOAc буфера, pH 5,5, и далее реакцию останавливают через 30 минут, 2, 4, и 17 часов соответственно путем замораживания образцов в жидком азоте. Впоследствии образцы хранят при -20°С до проведения дальнейшего анализа.
Добавляют буфер для проведения анализа образцов в SDS-PAGE и нагревают указанные образцы до 90°С, после чего наносят на SDS-содержащие гели. Как показано на фиг.14, увеличение времени инкубации приводит к более сильному смещению образцов в сторону более высокой молекулярной массы белка. После 17 часов инкубации в присутствии модифицированного гидроксиламином HES-производного Х обнаруживается диффузная окрашиваемая кумасси белковая полоса, мигрирующая в зоне между 60 и 11 кДа относительно положения стандартных препаратов для определения молекулярной массы (см. правую часть фиг.14). При обработке N-гликозидазой бульшая часть белков мигрирует в сторону положения де-N-гликозилированного EPO (см. фиг.14, правый гель; стрелка А указывает на положение перемещения N-гликозидазы, стрелка В указывает на миграцию де-N-гликозилированного EPO; диффузная белковая полоса, видимая на участке между положениями стандартных препаратов для определения молекулярной массы в диапазоне между 28 кДа и 36 кДа, преимущественно отражает те EPO-формы, которые включают модификации, определяемые HES и сайтом О-гликозилирования молекулы. В связи со специфичностью действия N-гликозидазы авторы на основании данного эксперимента пришли к выводу, что фактически HES-модификация происходит на окисленных перйодатом остатках сиаловой кислоты гликанов в белке EPO.
(bb) Характеристика конъюгатов HES-EPO
Конъюгаты HES-EPO I (получаемые из ЕРО-GT-1 после окисления перйодатом в мягких условиях, например, на основе ЕРО-GT-1-А), II (получаемые из ЕРО-GT-1, подвергнутого 5-минутному кислотному гидролизу и окислению перйодатом в мягких условиях) и III (получаемые из ЕРО-GT-1, подвергнутого 10-минутному кислотному гидролизу и окислению перйодатом в мягких условиях) синтезируют, как было описано ранее. Методика включает вариант контрольной инкубации (К), содержащий немодифицированный ЕРО-GT-1 в том же буфере, к которому добавляют эквивалентное количество немодифицированного HES. Инкубационные смеси подвергают дальнейшей очистке для проведения последующего биохимического анализа производных HES-EPO.
Полученные при инкубации конъюгаты HES-EPO I, II и III, а также продукт контрольной инкубации К подвергают очистке на сефарозе (Q-Sepharose), как было описано в разделе "Материалы и методы" (пример 20.8) с целью удаления избытка непрореагировавшего HES-реагента, который, как ожидается, будет выходить с потоком из ионообменной колонки. В связи с большими количествами основных EPO-форм, содержащихся в предварительно обработанных кислотой образцах II и III, авторы ожидали появление в потоке значительных количеств модифицированного EPO продукта как результат указанных вариантов инкубации. Как видно на фиг.15, практически весь EPO материал из образцов I удерживается на колонке с сефарозой (Q-Sepharose), тогда как лишь примерно 20-30% образцов II и III выходят во фракции, элюируемой при высокой концентрации соли. Весь белковый материал, взятый из инкубационных сред, включающих HES-производное Х как в потоке, так и во фракциях, элюируемых при высокой концентрации соли, характеризуется явно более высокой молекулярной массой при анализе в SDS-PAGE в сравнении с контрольным препаратом EPO.
С целью более детальной характеристики HES-модифицированного EPO в образцах А и К (см. фиг.13) проводят сравнение с окисленной перйодатом формой ЕРО-GT-1-А. Образцы подвергают обработке N-гликозидазой, и, как видно из фиг.16а и 16b, высвобождение N-гликанов приводит к получению двух полос с низкой молекулярной массой в положении О-гликозилированной и негликозилированной форм EPO стандартного препарата EPO. В случае образца А обнаруживается другая полоса, мигрирующая в положение, соответствующее М.м. 28 кДа, что предположительно указывает на наличие HES-модификации по О-гликану в данном варианте EPO (см. пример 20.11 (с)(аа)). Указанная полоса (а также высокомолекулярная форма N-гликозилированного EPO с высокой степенью HES-модификации, см. фиг.16а и 16b) исчезает после проведения мягкого гидролиза образцов, что согласуется с точкой зрения, согласно которой HES-модификация осуществляется по окисленным перйодатом остаткам сиаловой кислоты в эритропоэтине.
Аликвоты инкубационных смесей для N-гликозидазной обработки гидролизуют с использованием условий, позволяющих полностью удалить остатки сиаловой кислоты (а также производное сиаловой кислоты с присоединенным HES) из олигосахаридов; после нейтрализации смеси абсорбируют на небольшие колонки Hypercarb для обессоливания. Колонки тщательно промывают водой и затем проводят элюцию связанных нейтральных олигосахаридов 40% ацетонитрилом в воде, содержащей 0,1% трифторуксусной кислоты. Полученные олигосахариды подвергают анализу по методу MALDI/TOF-MS. Спектры десиалилированных фракций олигосахаридов из образца А, ЕРО-GT-1-А и образца К демонстрируют идентичную массу относительно олигосахаридов комплексного типа со значениями m/z = 1810 Да (диантенный), 2175 = триантенный, 2540 = тетраантенный, 2906 = татраантенный плюс 1 N-ацетиллактозаминовый повтор и 3271 = татраантенный плюс 2 N-ацетиллактозаминовых повтора; обнаруживаются небольшие сигналы, соответствующие отсутствию фукозы (-146) и галактозы (минус 162), которые определяются условиями кислотного гидролиза, применяемого для удаления сиаловой кислоты (см. MALDI-фиг.19, 20 и 21).
В параллельном эксперименте смесь после расщепления N-галактозидазой абсорбируют на картридже RP-C18 на 1 мл (без предварительного кислотного гидролиза олигосахаридов) и проводят элюцию 5% ацетонитрилом в воде, содержащей 0,1% TFA; в указанных условиях белок EPO полностью удерживается на RP-материале и олигосахариды вымываются из колонки 5% ацетонитрилом в воде, содержащей 0,1% TFA. Белок в де-N-гликозилированном EPO элюируют 70% ацетонитрилом в воде, содержащей 0,1% TFA. Олигосахаридные фракции на стадии N-гликозидазной обработки в RP-C18 образца А, ЕРО-GT-1-А и образца К нейтрализуют и подвергают обессоливанию с использованием картриджей Hypercarb, как было описано ранее. Выделенные олигосахариды подвергают картированию по методу анализа HPAEC-PAD до (см. фиг.13) и после обработки кислотой в мягких условиях, позволяющей осуществить количественное удаление сиаловых кислот из гликанов (см. фиг.18).
Профиль, выявляемый при анализе по методу HPAEC-PAD нативного материала, полученного из HES-модифицированного образца А, демонстрирует лишь ничтожные сигналы, характерные для олигосахаридов, тогда как олигосахариды из ЕРО-GT-1-А демонстрирует тот же профиль гликанов, что и вариант, показанный на фиг.13 (образец, обозначенный как ЕРО-GT-1 после обработки перйодатом в мягких условиях). Профиль элюции олигосахаридов, полученных из контрольного образца EPO (К), соответствует ожидаемым характеристикам (в сравнении с профилем, показанным на фиг.10). Для сравнения приведен профиль нативного олигосахарида в международном стандартном препарате BRP-EPO, выполняющем функцию эталонного стандарта.
После проведения кислотного гидролиза в мягких условиях все олигосахаридные препараты демонстрируют идентичный профиль элюции нейтральных олигосахаридных структур (см. фиг.18) с ожидаемым количественным и качественным составом ди-, три- и тетраантенных углеводных цепей комплексного типа, как описано в методическом разделе относительно препарата EPO, который использовался в качестве исходного материала для настоящего исследования. Указанный результат показывает, что HES-модификация образца EPO приводит к ковалентному связыванию HES-производного, которое отделяется от EPO белка с помощью N-гликозидазы и которое является кислотолабильным, поскольку удаляется из N-гликанов в условиях мягкого кислотного гидролиза, приводящего, как известно, к десиалилированию углеводов (см. фиг.16а и 16b).
(сс) Анализ моносахаридного состава HES-EPO и N-гликанов HES-EPO по методу GC-MS
Для того чтобы дополнительно подтвердить наличие HES-модификации EPO по N-гликанам в молекуле, образцы EPO расщепляют N-гликозидазой и белок EPO адсорбируют на картриджах RP-C18, тогда как олигосахаридный материал вымывают, как было описано выше. Как показано в таблице 3, глюкоза и гидроксиэтилированные производные глюкозы обнаруживаются только в EPO-белке, который был подвергнут HES-модификации по цистеиновым остаткам, и в олигосахаридных фракциях EPO в образце А2.
Пример 20.11(d) Тестирование in vivo биологической активности HES-модифицированного EPO
Тестирование EPO в системе нормоцитных мышей проводят в соответствии с процедурами, описанными в Европейской Фармакопее; в лаборатории, проводящей EPO-тестирование, используется международный стандартный препарат BRP EPO. В случае препарата HES-модифицированного EPO А2 было определено среднее значение удельной активности, равное 294600 единиц на мг белка EPO, указывающее на наличие примерно в 3 раза более высокой удельной активности, чем в международном стандартном препарате BRP EPO, который был включен для оценки активности вместе с исследуемыми образцами.
Результаты исследования приведены в таблице 4.
Ссылки к примерам 13-20:
Nimtz M, Noll G, Paques EP, Conradt HS.
Carbohydrate structures of a human tissue plasminogen activator expressed in recombinant Chinese hamster ovary cells.
FEBS Lett. 1990 Oct. 1; 271(1-2):14-8.
Domer AJ, Wasley LC, Kaufman RJ.
Increased synthesis of secreted proteins induced expression of glucose-regulated proteins in butyrate-treated Chinese hamster ovary cells.
J. Biol. Chem. 1989 Dec 5; 264 (34): 20602-7.
Mueller PP, Schlenke P, Nimtz M, Conradt HS, Hauser H
Recombinant glycoprotein quality in proliferation-controlled BHK-21 cells.
Biotehnol Bioeng. 1999 Dec 5; 65(5): 529-36.
Nimtz M, Martin W, Wray V, Kloppel KD, Augustin J, Conradt HS.
Structures of sialylated oligosaccharides of human erythropoietin expressed in recombinant BHK-21 cells.
Eur J Biochem. 1993 Apr. 1; 213(1): 39-56.
Hermentin P, Witzel R, Vliegenthart JF, Kamerling JP, Nimtz M, Conradt HS.
A strategy for the mapping of N-glycans by high-ph anion-exchange chromatography with pulsed amperometric detection.
Anal Biochem. 1992 Jun; 203(2): 281-9.
Schroter S, Derr P, Conradt HS, Nimtz M, Hale G, Kirchhoff C.
Male specific modification of human CD52.
J Biol Chem. 1999 Oct. 15; 274(42): 29862-73
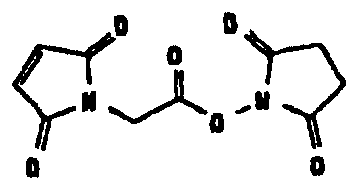

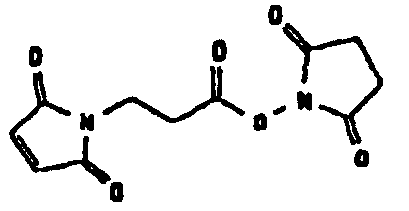
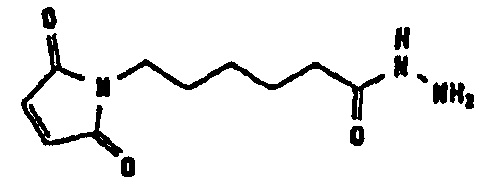
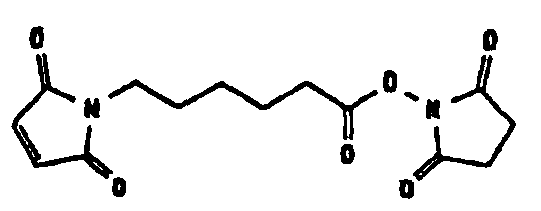
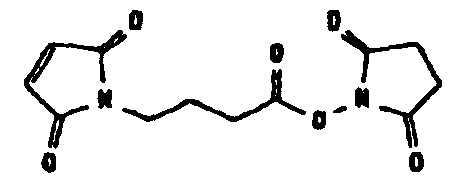

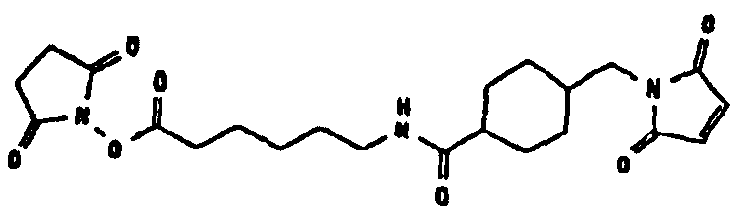
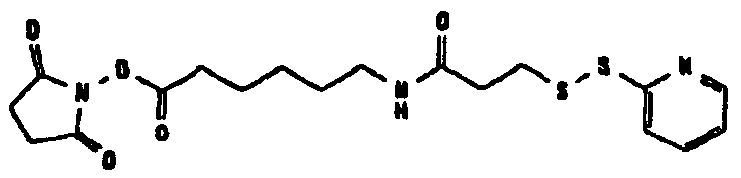
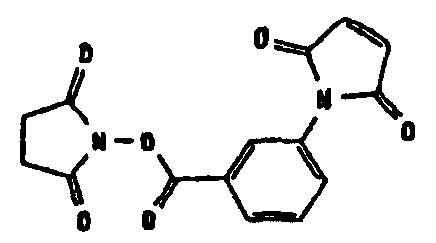
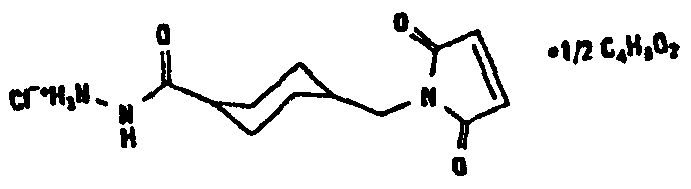

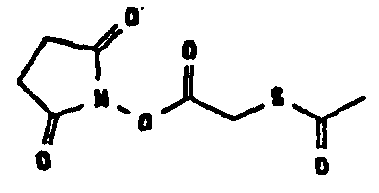
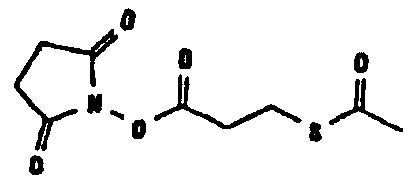
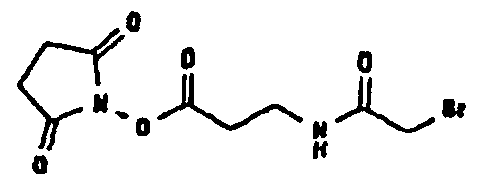
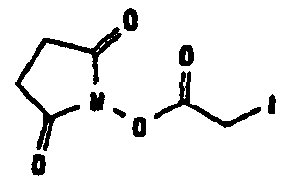
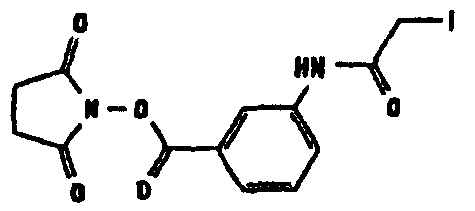
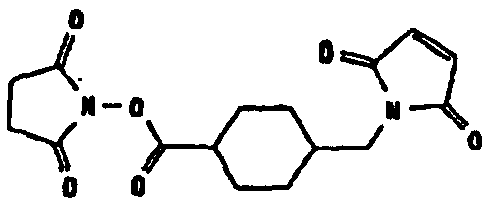
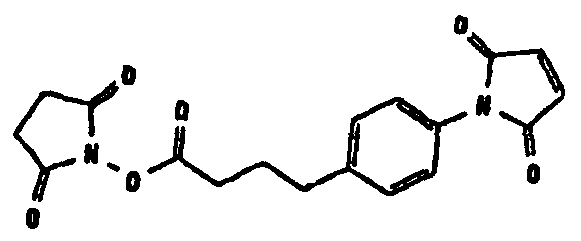
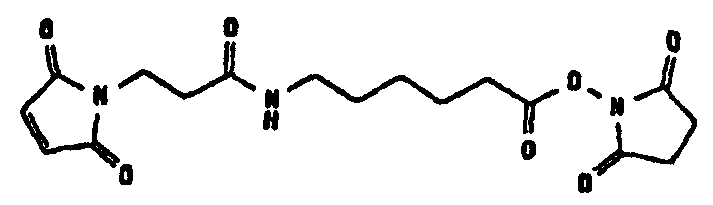
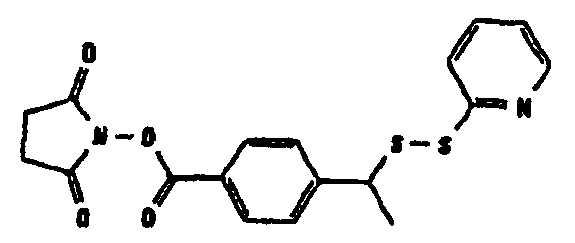
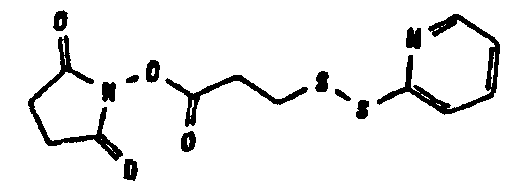
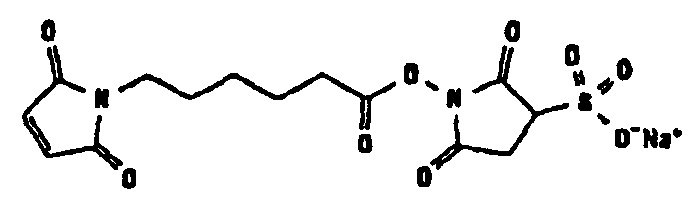
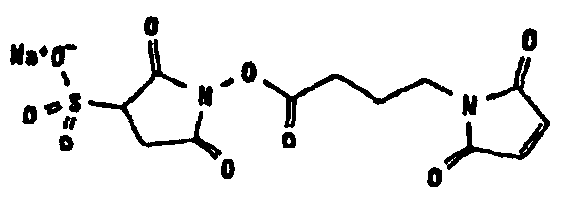
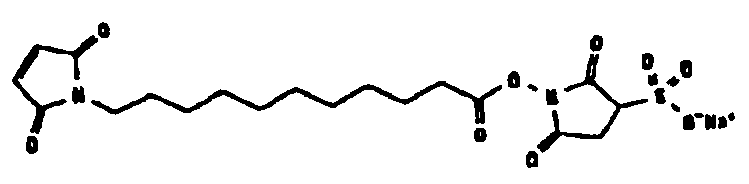
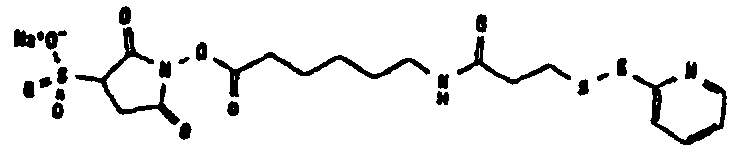
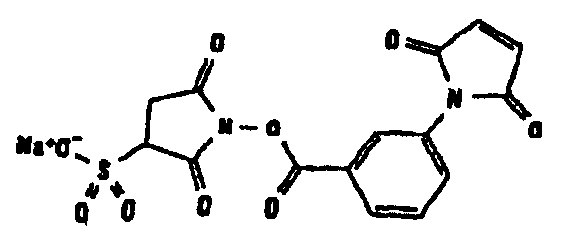
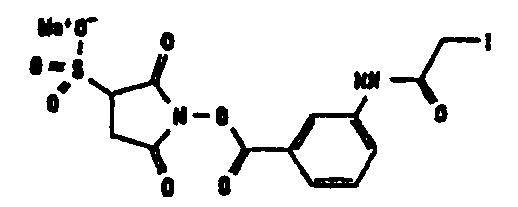
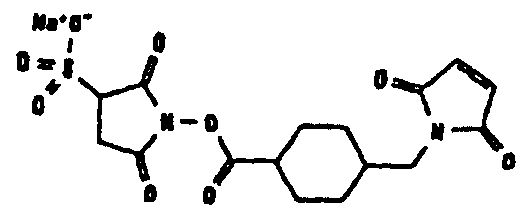
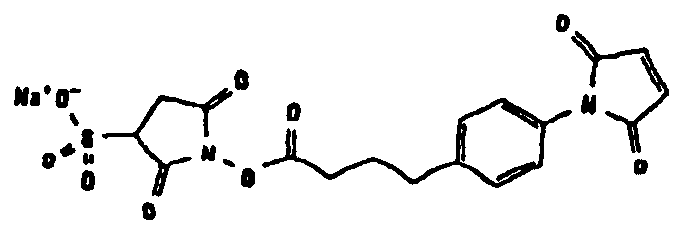

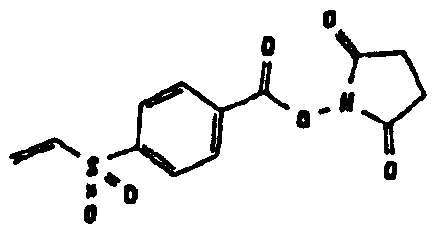
Анализ моносахаридного состава гликанов из HES-модифицированного EPO и контрольных образцов
** Определение количества моносахаридов проводят при одном пропускании через GC пертриметилсиалилированных метилгликозидов; значения пиков при их электронной интеграции приводятся без коррекции на потери в ходе процедур дериватизации и восстановления каждого соединения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ПРОИЗВОДНЫХ ГИДРОКСИАЛКИЛКРАХМАЛА | 2003 |
|
RU2329274C2 |
| ПРОИЗВОДНЫЕ ГИДРОКСИАЛКИЛКРАХМАЛА | 2003 |
|
RU2326891C2 |
| КОНЪЮГАТЫ ГИДРОКСИАЛКИЛКРАХМАЛА И G-CSF | 2004 |
|
RU2370281C2 |
| КОНЪЮГАТЫ ГИДРОКСИАЛКИЛКРАХМАЛА И АЛЛЕРГЕНА | 2003 |
|
RU2325925C2 |
| ГЛИКОПОЛИСИАЛИРОВАНИЕ БЕЛКОВ, НЕ ЯВЛЯЮЩИХСЯ БЕЛКАМИ СВЕРТЫВАНИЯ КРОВИ | 2010 |
|
RU2533619C2 |
| ГЛИКОПОЛИСИАЛИРОВАНИЕ БЕЛКОВ, НЕ ЯВЛЯЮЩИХСЯ БЕЛКАМИ СВЕРТЫВАНИЯ КРОВИ | 2014 |
|
RU2662807C2 |
| КОНЪЮГАТЫ БЕЛКОВ СВЕРТЫВАНИЯ КРОВИ | 2010 |
|
RU2595442C2 |
| КОНЪЮГАТЫ БЕЛКОВ СВЕРТЫВАНИЯ КРОВИ | 2010 |
|
RU2744370C2 |
| СПОСОБ ПОЛУЧЕНИЯ КОНЪЮГАТА НОНА-β-(1→3)-ГЛЮКОЗИДА С БЫЧЬИМ СЫВОРОТОЧНЫМ АЛЬБУМИНОМ СКВАРАТНЫМ МЕТОДОМ | 2011 |
|
RU2476444C1 |
| КОНЪЮГАТ ГИДРОКСИАЛКИЛКРАХМАЛА И НИЗКОМОЛЕКУЛЯРНОГО ВЕЩЕСТВА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2003 |
|
RU2327702C2 |
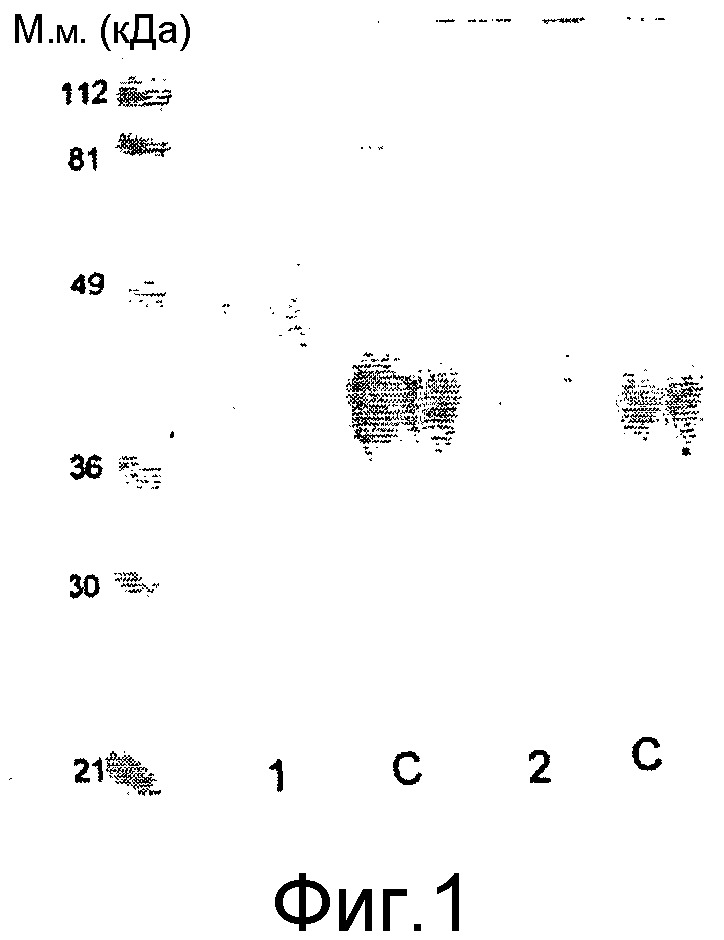
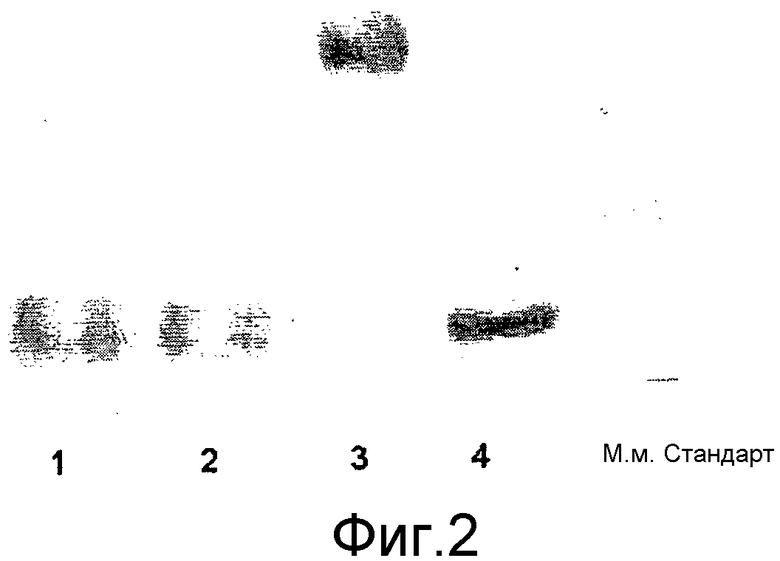


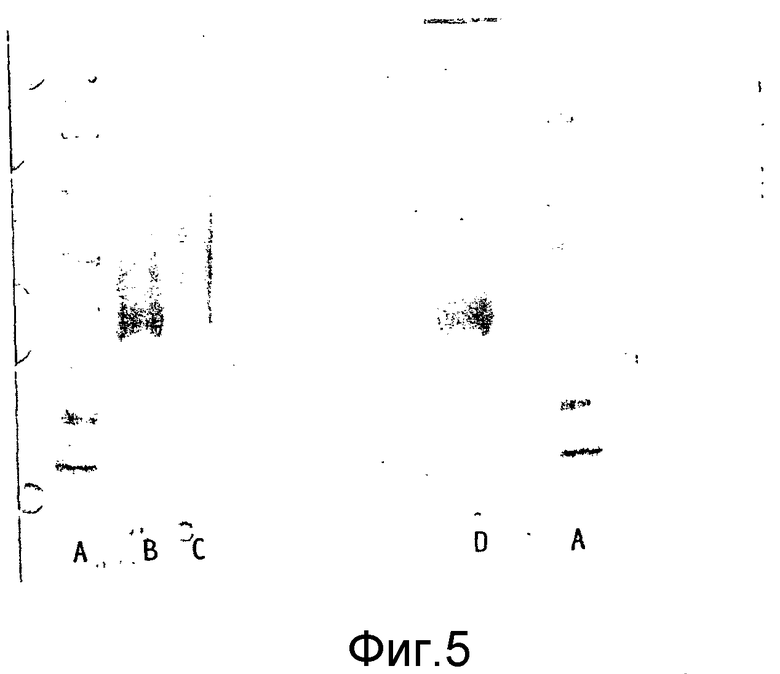
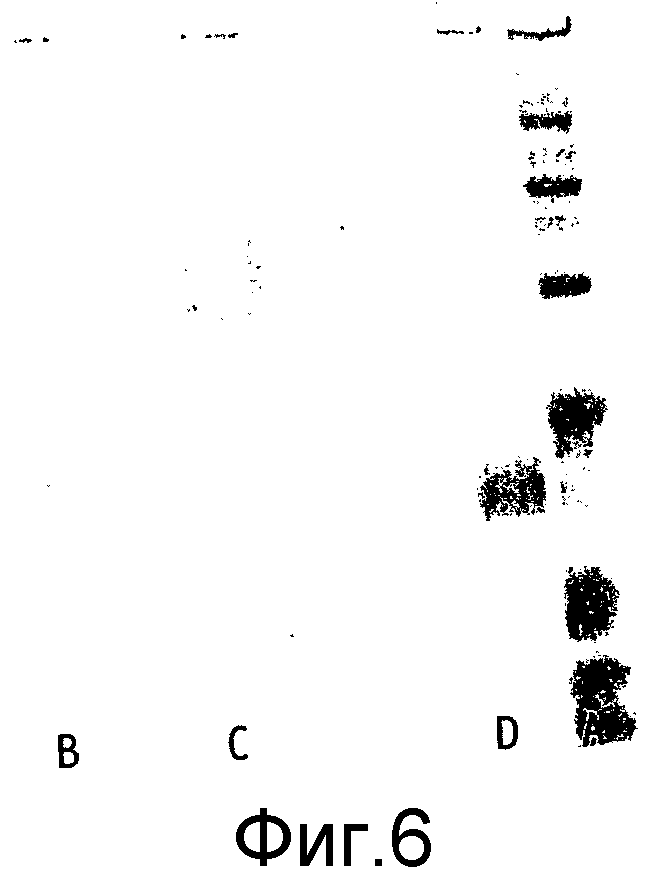
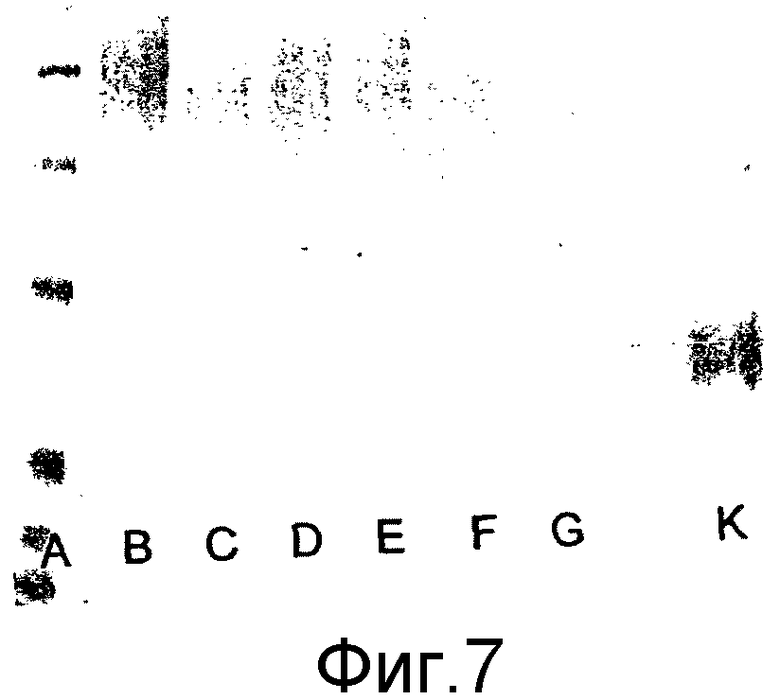
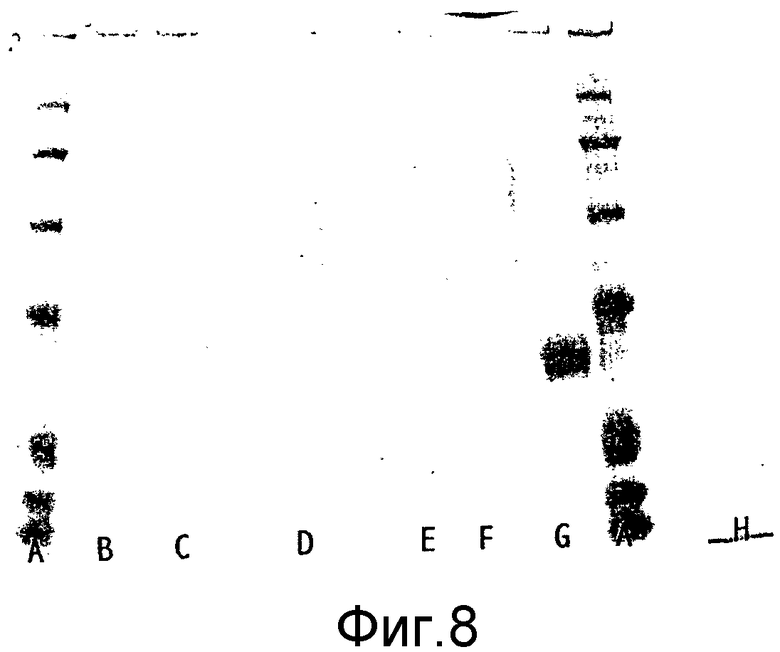
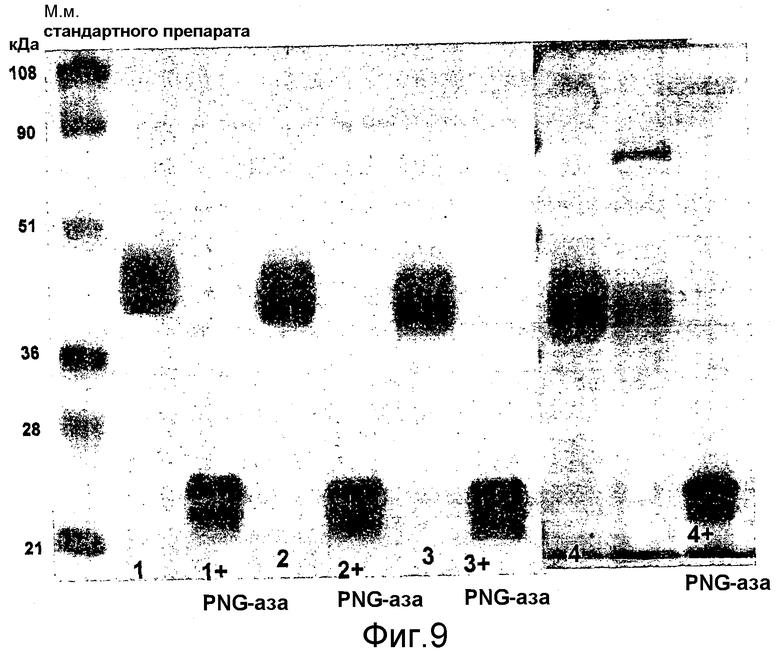
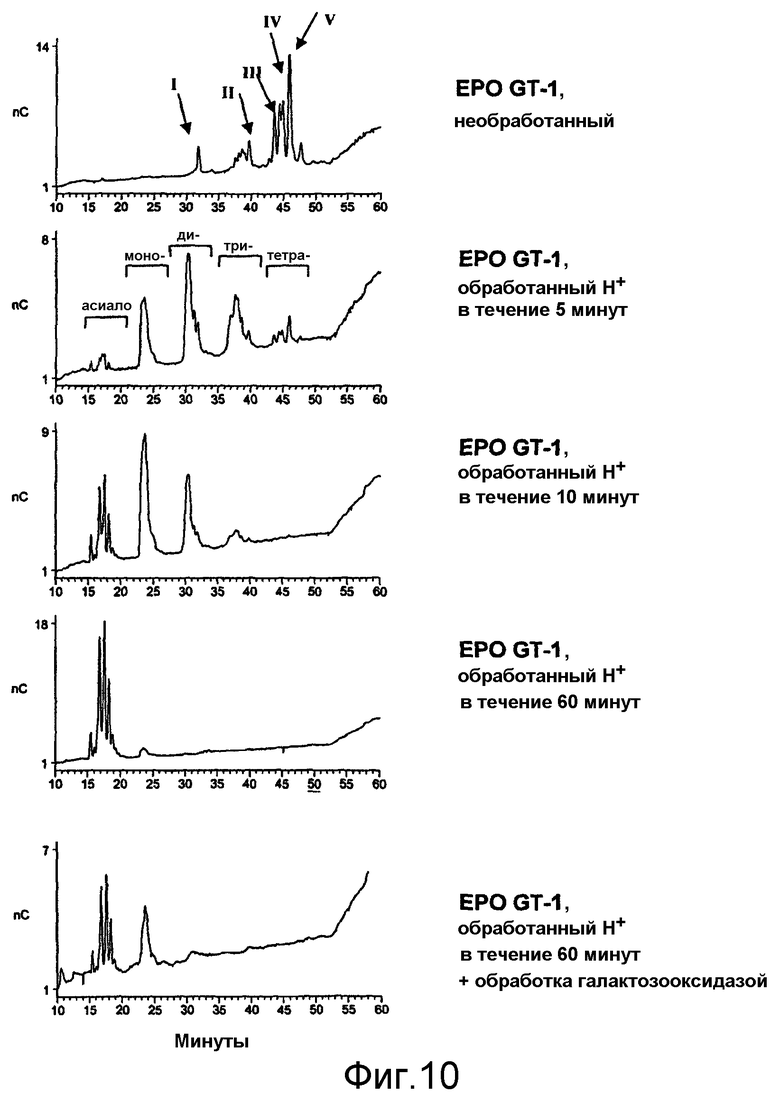
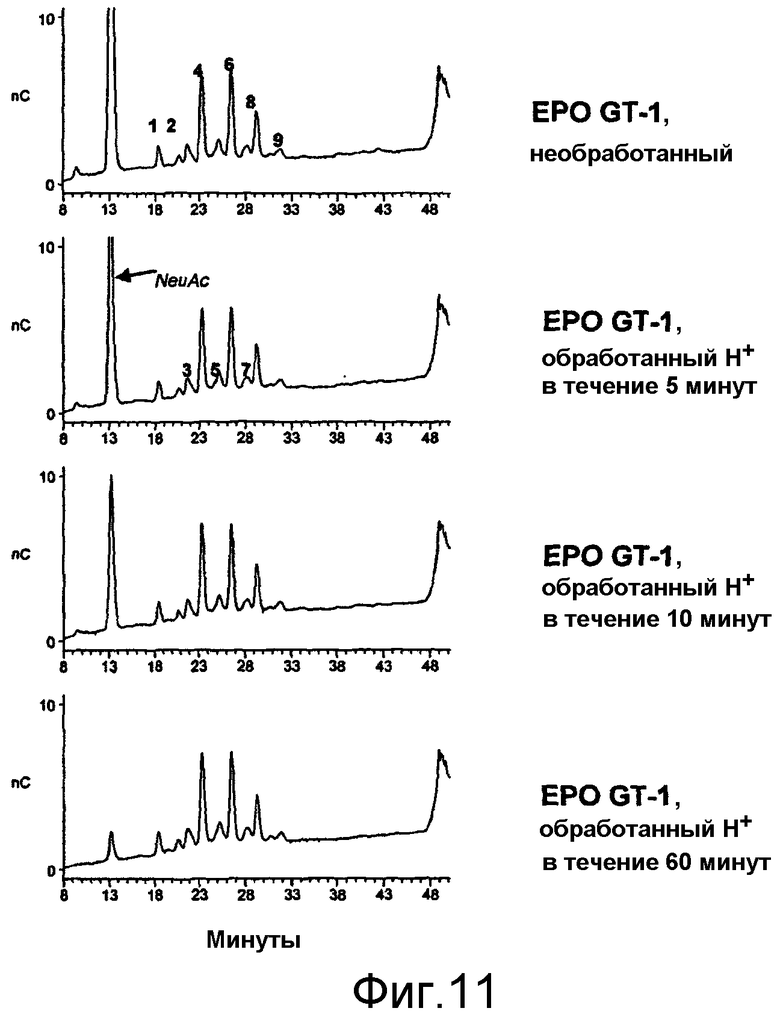
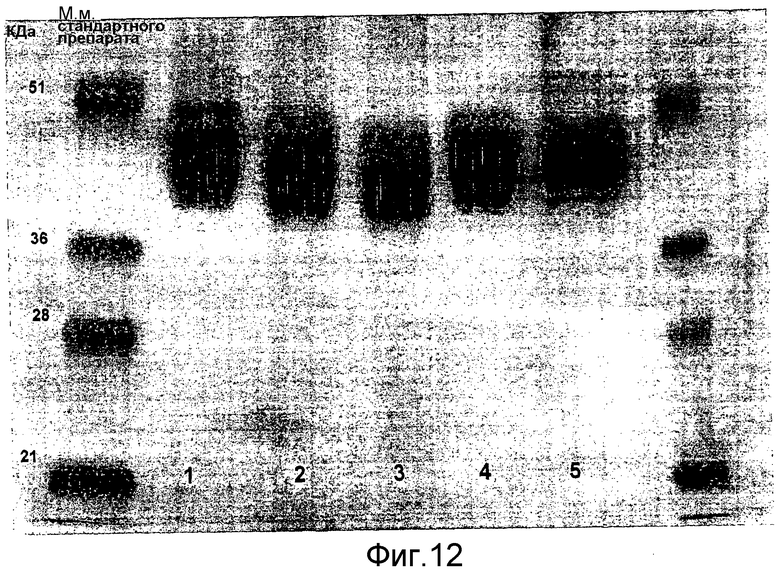
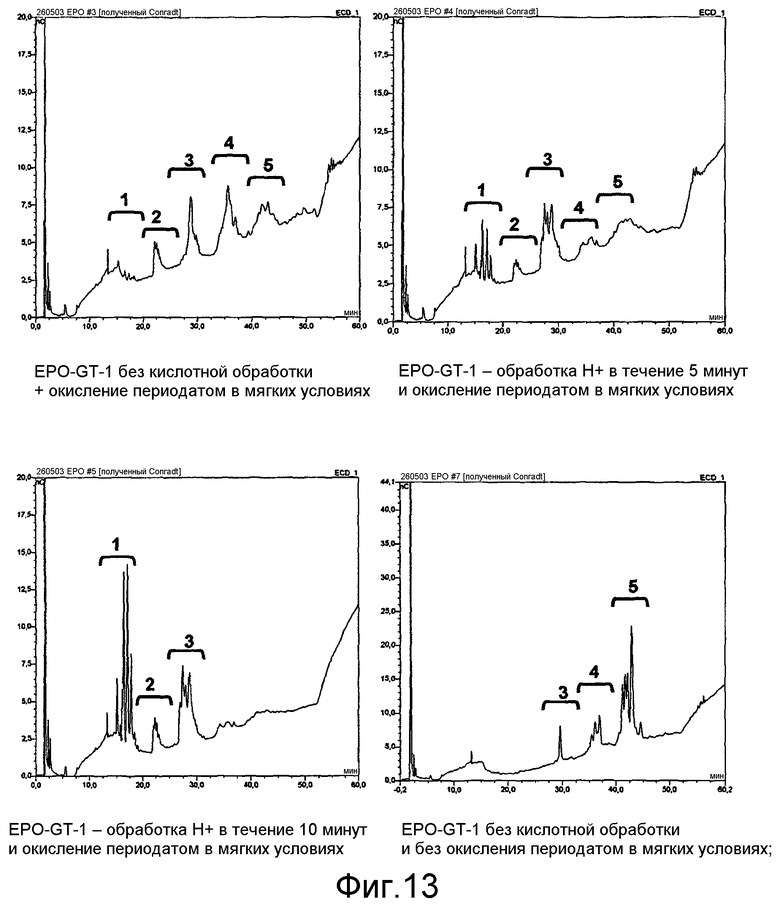
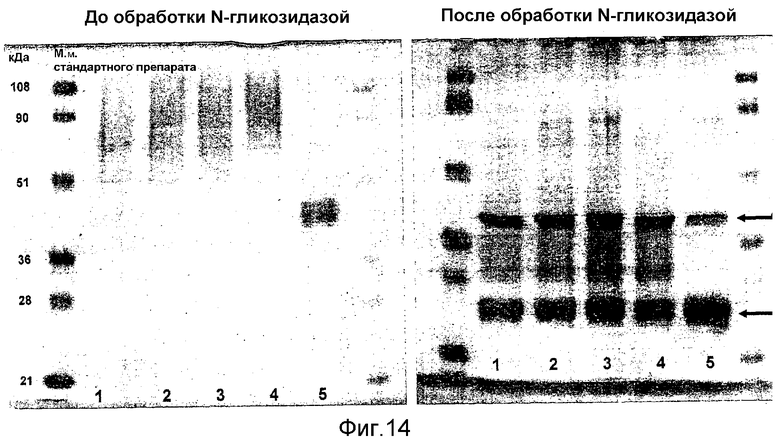
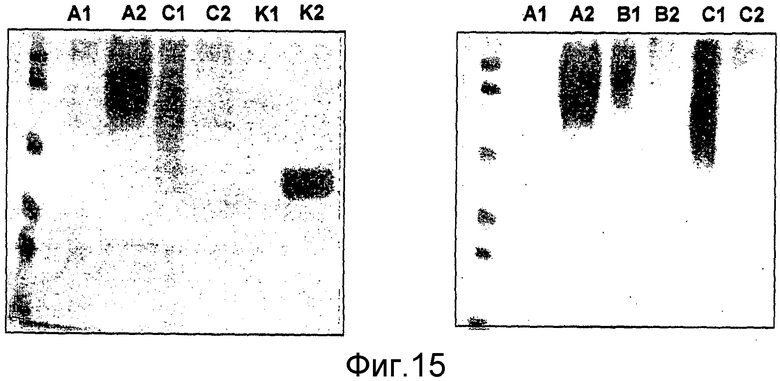

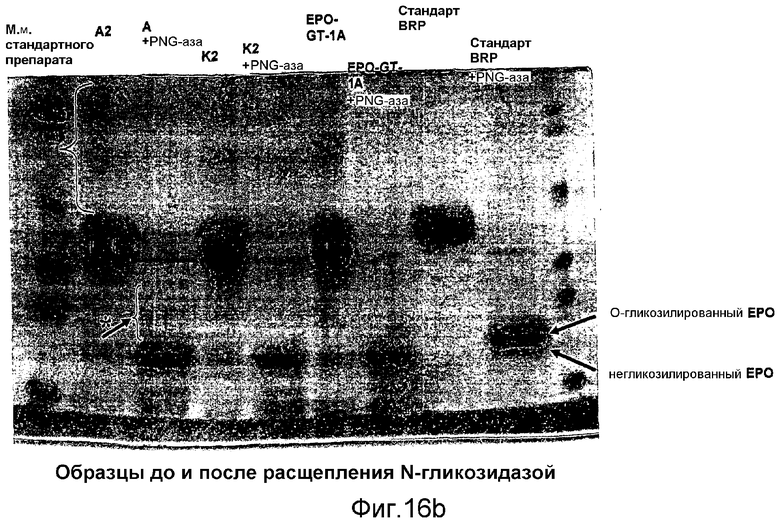
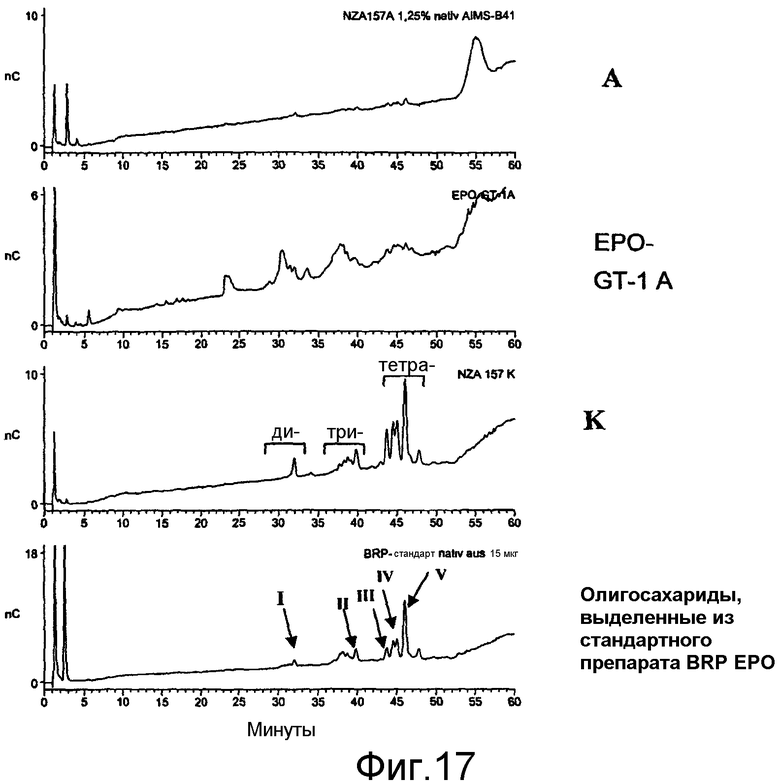
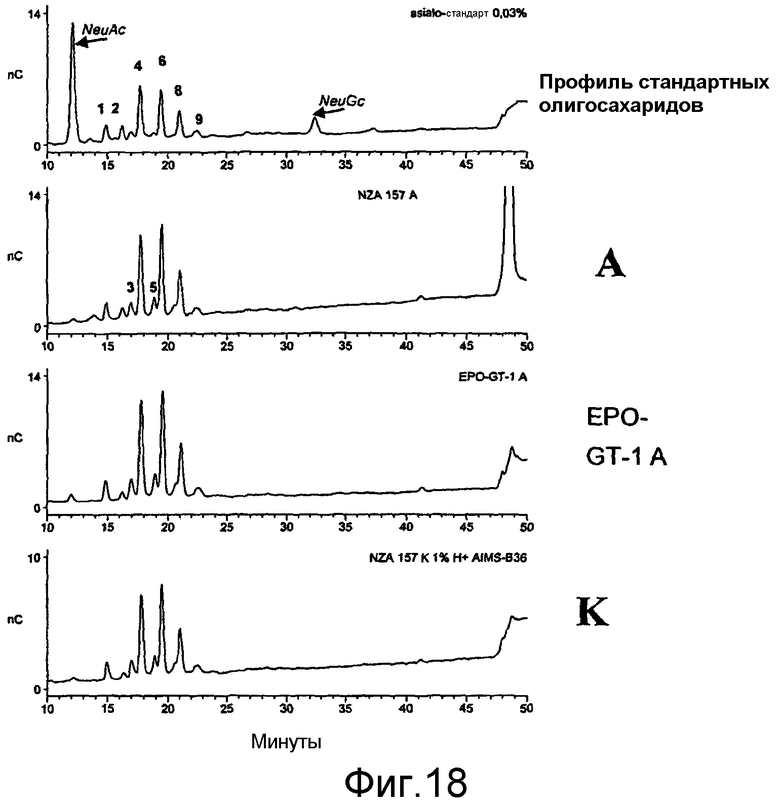

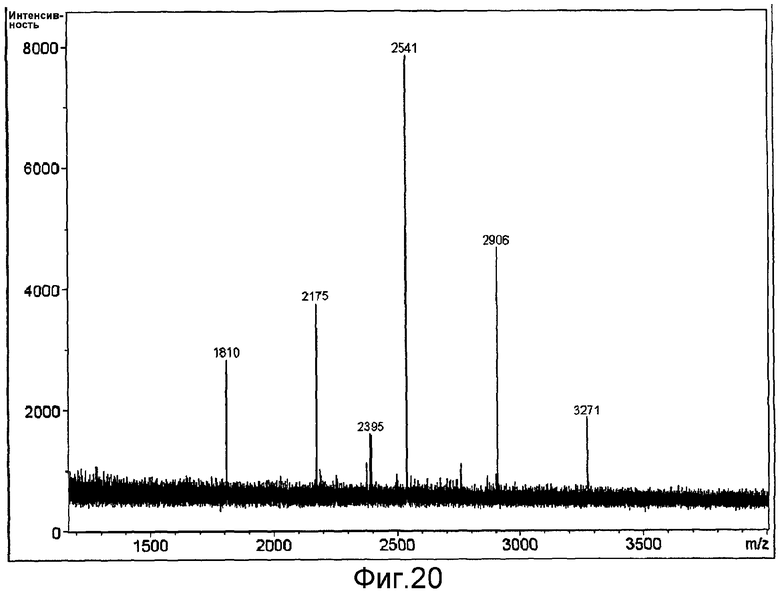
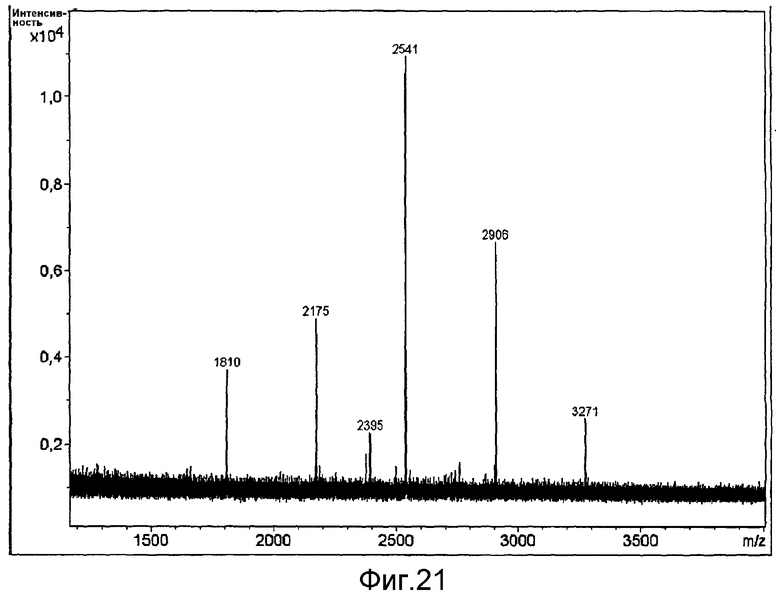
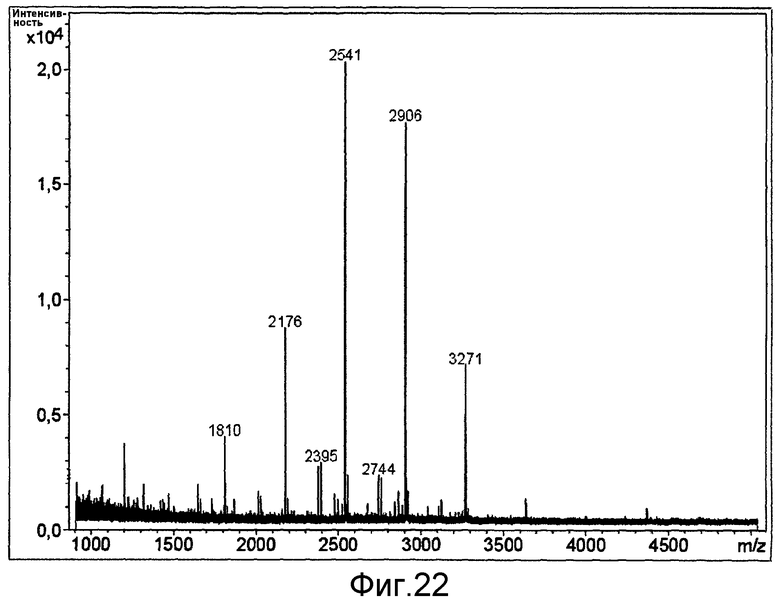
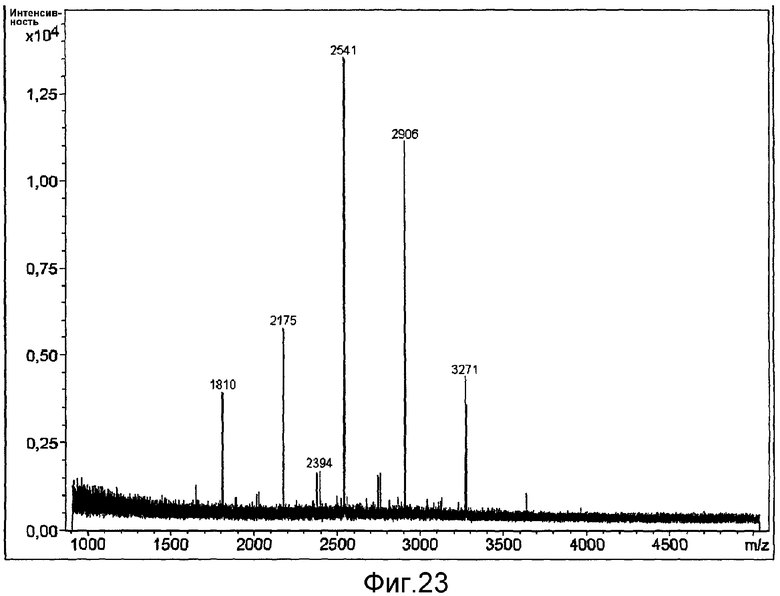
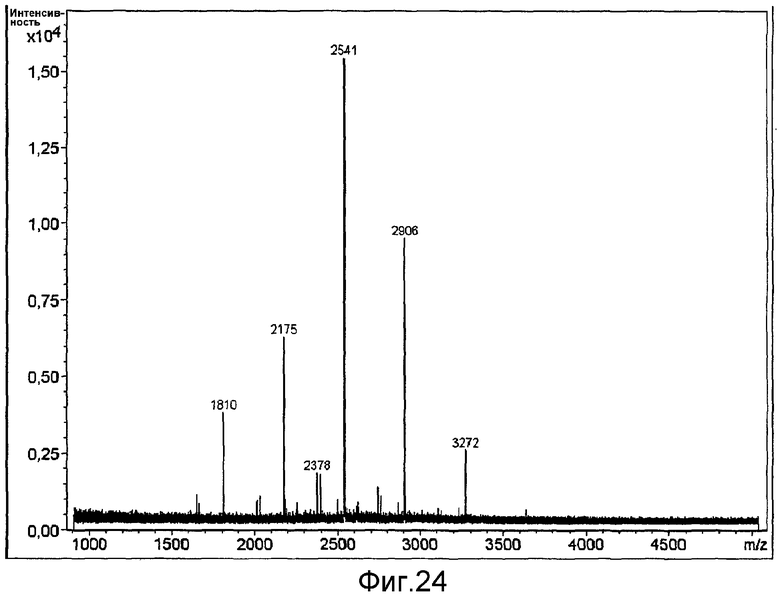
Изобретение относится к биоорганической химии и касается нового конъюгата эритропоэтина, способа его получения и фармацевтической композиции на его основе. Предложен конъюгат гидроксиэтилкрахмала и эритропоэтина, содержащий одну или несколько молекул гидроксиэтилкрахмала. Каждая молекула гидроксиэтилкрахмала конъюгирована с эритропоэтином, через углеводный фрагмент. Указанный гидроксиэтилкрахмал имеет молекулярную массу от 1 до 300 кДа и проявляет соотношение С2:С6-замещения в диапазоне 2-20 по гидроксиэтильным группам. Предложен способ получения конъюгата гидроксиэтилкрахмала и эритропоэтина, в соответствии с которым необязательно частично или полностью ферментативно и/или химически удаляют концевую сиаловую кислоту и необязательно частично или полностью окисляют концевое сахаридное звено эритропоэтина с получением эритропоэтина, способного к реакции с модифицированным гидроксиэтилкрахмалом. В гидроксиэтилкрахмал вводят свободную гидразидную, гидроксиламиновую, тиольную или семикарбазидную функциональную группу с получением модифицированного гидроксиэтилкрахмала, который конъюгируют с эритропоэтином, способным к реакции с модифицированным гидроксиэтилкрахмалом, и получают конъюгат гидроксиэтилкрахмала и эритропоэтина, содержащий одну или несколько молекул гидроксиэтилкрахмала. Фармацевтическая композиция содержит конъюгат гидроксиэтилкрахмала и эритропоэтина, который обладает активностью эритропоэтина, и фармацевтически приемлемый разбавитель, адъювант и/или носитель. Технический результат - конъюгат гидроксиэтилкрахмала и эритропоэтина, обладающий активностью эритропоэтина. 3 н. и 16 з.п. ф-лы, 4 табл., 25 ил.
(1) получение ЕРО, способного к реакции с модифицированным HES, предусматривающее необязательное частичное или полное окисление концевого сахаридного звена необязательно после частичного или полного (ферментативного и/или химического) удаления концевой сиаловой кислоты,
(2) модификация HES путем введения свободной гидразидной, гидроксиламиновой, тиольной или семикарбазидной функциональной группы, и
(3) конъюгирование модифицированного HES со стадии (2) с модифицированным ЕРО со стадии (1), посредством чего получают HES-EPO, содержащий одну или несколько молекул HES.
| WO 9428024 А, 12.08.1994 | |||
| DE 19628705 A, 15.01.1998 | |||
| US 5281698 A, 25.01.1994 | |||
| ЕР 0809996 А, 03.12.1997 | |||
| US 4847325 А, 11.07.1989 | |||
| ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ ЭРИТРОПОЭТИН ЧЕЛОВЕКА, РЕКОМБИНАНТНЫЕ ДНК, ЛИНИЯ КЛЕТОК, РЕКОМБИНАНТНЫЙ О-ГЛИКОЗИЛИРОВАННЫЙ ЭРИТРОПОЭТИН ЧЕЛОВЕКА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1991 |
|
RU2120996C1 |
| LIN F.-Q | |||
| et al | |||
| Cloning and expression of the human erythropoietin gene | |||
| Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| U.S.A., 1985, 82:7580-4 | |||
| TAKEUCHI M | |||
| et al | |||
| Relationships between sugar chain structure and | |||

