Изобретение относится к области молекулярной биологии и медицины, а именно к разработке методов генотипирования полиморфизма генов ферментов антиоксидантной защиты у человека.
В последние годы отмечается значительный интерес исследователей к изучению полиморфизма генов системы антиоксидантной защиты при различной патологии у человека (Chu Y., Alwahdani A., Iida S. et al. Vascular effects of the human extracellular superoxide dismutase R213G variant. // Circulation. - 2005. - Vol.112. - P.1047-1053; Young R.P., Hopkins R., Black P.N. et al. Functional variants of antioxidant genes in smokers with COPD and in those with normal lung function. // Thorax. - 2006. - Vol.61. - P.394-399). Внеклеточная супероксиддисмутаза (EC-SOD или SOD3) является одним из ключевых антиоксидантных ферментов, широко представленных в различных тканях, который осуществляет контроль за избыточным образованием свободных радикалов и продуктов перекисного окисления липидов (Petersen S.V., Enghild J.J. Extracellular superoxide dismutase: structural and functional considerations of a protein shaped by two different disulfide bridge patterns. // Biomed. Pharmacother. - 2005. - Vol.59. - P.175-182).
К настоящему времени нуклеотидная структура гена SOD3 расшифрована полностью (NCBI GenelD: 6649; OMIM: 185490), установлены однонуклеотидные полиморфизмы (ОНП) в кодирующей части гена. Среди методов генотииирования наиболее подробно описаны способы идентификации полиморфизма R213G SOD3. В то же самое время методов рутинного обнаружения другого распространенного ОНП А40Т (Ala40Thr) (референтная последовательность NCI dbSNP rs2536512), расположенного также в кодирующей части 3 экзона гена SOD3, на настоящее время не разработано.
Наиболее близким к заявляемому способу идентификации А40Т полиморфизма гена SOD3 является метод прямого секвенирования кодирующей части гена, примененный Tamai M. et al. (Tamai M., Furuta H., Kawashima H. et al. Extracellular superoxide dismutase gene polymorphism is associated with insulin resistance and the susceptibility to type 2 diabetes. // Diabetes Res. Clin. Pract. - 2006. - Vol.71. - P.140-145).
Метод прямого секвенирования ДНК осуществляется на специальном приборе - генетическом анализаторе (автоматический секвенатор нуклеотидных последовательностей ДНК) и основан на детекции флуоресцентно-меченых нуклеотидных последовательностей, разделяющихся с помощью капиллярного гель-электрофореза. Для специфического связывания с каждым из 4 нуклеотидов (А, Т, С, G) используются 4 различных флуоресцентных красителя. Во время электрофореза луч лазера фокусируется на определенном месте в геле. При прохождении фрагмента ДНК через эту точку лазер вызывает флуоресценцию красителя. Максимальная степень флуоресценции возникает при различных длинах волн для каждого из 4 красителей. Информация о последовательной флуоресценции регистрируется автоматически специальным детектором, а нуклеотидная последовательность сохраняется в базе данных компьютера.
Данный метод, несмотря на свою высокую чувствительность и специфичность, требует специального дорогостоящего оборудования - автоматического секвенатора, в частности ABI PRISM 310 Genetic Analyzer, a также специальных реагентов, необходимых для проведения анализа.
Кроме прямого секвенирования, на сегодняшний день не разработано других менее дорогостоящих методов обнаружения полиморфизма Ala40Thr в 3 экзоне гена SOD3. Существует потребность в создании быстрого, недорогого способа идентификации полиморфизма А40Т гена SOD3, который мог бы использоваться в качестве рутинного метода генотипирования в ПЦР-лаборатории со стандартным набором оборудования и реактивов.
Задачей изобретения является расширение арсенала методов генотипирования полиморфизма А40Т гена SOD3 и уменьшение материальных затрат путем разработки способа его идентификации относительно недорогими и легко выполнимыми в любой ПЦР-лаборатории методами ПЦР-ПДРФ.
Изобретение поясняется чертежом:
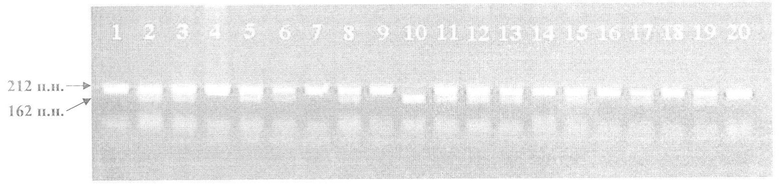
На чертеже изображено электрофоретическое разделение продуктов ПЦР-рестрикции гена SOD3. Слева обозначены размеры (п.н.) фракционированных фрагментов ДНК. Сверху обозначены порядковые номера проанализированных образцов 20 человек. Образец 10 (фрагмент 162 п.н.) - гомозиготный генотип по аллелю "дикого" типа 40АА SOD3; образцы 1, 4, 7, 9, 14, 16, 18 и 20 (фрагмент 212 п.н.) - гомозиготные генотипы по вариантному аллелю 40ТТ SOD3; образцы 2, 3, 5, 6, 8, 11, 12, 13, 15, 17 и 19 (фрагменты 212 и 162 п.н.) - гетерозиготные генотипы 40АТ SOD3 (фрагменты размерами 48, 39 и 2 п.н. на геле не визуализируются).
В основе предлагаемого способа обнаружения полиморфизма А40Т гена SOD3 лежит методология ПЦР-опосредованного сайт-направленного мутагенеза, предложенная Eiken H.G. et al. (Eiken H.G., Odland E., Ramos С. et al. Application of natural and amplification created restriction sites for the diagnosis of PKU mutations. // Nucleic Acids Res. - 1991. - Vol.19. - P.1427-1430).
Предлагаемый способ осуществляется следующим образом.
На основе нуклеотидной последовательности гена внеклеточной супероксиддисмутазы-3 с использованием программы "GeneFisher" 1.3 (http://bibiserv.techfak.uni-bielefeld.de) была подобрана пара праймеров для специфической амплификации интересуемого участка гена SOD3:
F: 5'-TACTGTGTTCCTGCCTGCTC-3' (SEQ ID NO: 1)
R: 5'-AAGCTGCCGGAAGAGGACG-3' (SEQ ID NO: 2)
Праймеры были синтезированы в ЗАО "Синтол" (г.Москва). Амплификацию проводили на многоканальном термоциклере "Терцик" (ЗАО НПФ "ДНК-Технология", г.Москва). Параметры ПНР были следующими: "горячий старт" в течение 5 мин при 94°С, затем 37 циклов амплификации в режиме 94°С - 1 мин; 57°С - 1 мин; 72°С - 1 мин, заключительный цикл элонгации ДНК - 10 мин при 72°С. Размер амплифицированного в ходе ПЦР фрагмента гена SOD3 составил 251 пара нуклеотидов (п.н.). Нуклеотидную последовательность продукта амплификации гена SOD3 определяли на генетическом анализаторе ABI PRISM 310 Genetic Analyzer с использованием набора реактивов BigDye Termination Kit 1.1 ("Applied Biosystems", США).
При приготовлении растворов для проведения ПЦР использовали импортные реагенты высокой степени очистки (Ultra pure и Biotechnology Grade). ПЦР проводили в 12 мкл реакционной смеси, содержащей 0,5 мкл образца геномной ДНК. Смесь для амплификации включала: 67 мМ Трис-HCl рН 8,8, 16,6 мМ сульфата аммония, 6,7 мкМ ЭДТА, 1,5 mM MgCl2, 1 мМ β-меркаптоэтанола, 1 мМ каждого dNTPs (dATP, dCTP, dTTP и dGTP), 15 нМ каждого из праймеров и 1U (1 единица активности) Taq-полимеразы. Для предотвращения испарения амплификационной смеси и образования конденсата на реакционную смесь наслаивали по 30 мкл минерального масла.
Обнаружение А40Т полиморфизма гена SOD3 проводилось путем обработки ампликона 5U эндонуклеазы BstHHI (прототип HhaI, сайт узнавания GCG↑C) согласно протоколу, описанному производителем фермента (ООО "Сибэнзим", г.Новосибирск). Рестрикционную смесь термостатировали 16 часов при температуре 50°С. После инкубации фрагменты ДНК фракционировали с помощью электрофореза в 2% агарозном геле, приготовленном на основе ТВЕ буфера (0,089М Трис-HCl, 2 мМ ЭДТА, 0,089М борная кислота) с 0,01% бромистым этидием. Фракционирование фрагментов ДНК проводили в камере для горизонтального электрофореза SH-2 (ООО "Хеликон", г.Москва) в течение 30 мин при напряжении 180V. В качестве маркера молекулярного веса использовали ДНК фага λ, гидролизированную эндонуклеазой PstI. Разделенные фрагменты ДНК визуализировали на трансиллюминаторе с помощью прибора компьютерной видеосъемки GDS-8000 ("UVP", США). Документирование и обработка изображений электрофоретических гелей проводилась с помощью аналитического пакета Labworks.
Для демонстрации эффективности разработанного способа идентификации А40Т полиморфизма гена SOD3 было проведено генотипирование у 20 добровольцев. Результаты электрофоретического разделения продуктов ПЦР-рестрикции А40Т полиморфизма гена SOD3 представлены на чертеже. Аллель "дикого" типа 40А SOD3 (GCG) формирует сайт рестрикции для эндонуклеазы BstHHI, которая расщепляет ампликон размером 251 и.н. на следующие фрагменты: 162, 48, 39 и 2 п.н. (вне зависимости от аллельного варианта полиморфизма А40Т гена SOD3 имеется дополнительный контрольный сайт рестрикции для эндонуклеазы BstHHI). В случае наличия вариантного аллеля 40Т SOD3 (ACG) один сайт рестрикции для эндонуклеазы BstHHI исчезает и при электрофоретическом разделении идентифицируют только фрагмент размером 212 п.н. (фрагмент 39 п.н. не визуализируется). У гетерозиготных носителей 40АТ SOD3 благодаря наличию обоих аллельных вариантов гена присутствуют оба дифференцирующих фрагмента размерами 212 и 162 п.н. Как видно из фигуры, высокое качество визуализации фрагментов ДНК позволило эффективно провести детекцию трех вариантов генотипов А40Т полиморфизма гена SOD3 у всех 20 проанализированных образцов.
Представленные результаты показывают, что предлагаемый способ позволяет эффективно идентифицировать аллельные варианты полиморфизма А40Т в гене внеклеточной супероксиддисмутазы методами ПЦР-ПДРФ. В связи с низкой себестоимостью анализа, обусловленной низкой ценой на эндонуклеазу BstHHI OOO "Сибэнзим", предлагаемый способ генотипирования может быть применен в любой ПЦР-лаборатории с минимальными материальными затратами.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ идентификации полиморфизмов Cys1079Gly и Cys1079Phe медь-транспортной АТФ-азы Вильсона | 2020 |
|
RU2756112C1 |
| СПОСОБ ГЕНОТИПИРОВАНИЯ ПОЛИМОРФИЗМА rs2551715 ГЕНА ГЛУТАТИОНРЕДУКТАЗЫ У ЧЕЛОВЕКА | 2013 |
|
RU2549688C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ПОЛИМОРФИЗМА R287Q В 8 ЭКЗОНЕ ГЕНА ЦИТОЗОЛЬНОЙ ЭПОКСИДГИДРОЛАЗЫ У ЧЕЛОВЕКА | 2006 |
|
RU2346053C2 |
| СПОСОБ ГЕНОТИПИРОВАНИЯ ПОЛИМОРФИЗМА rs17522918 ГЕНА ПЕРОКСИРЕДОКСИНА-1 У ЧЕЛОВЕКА | 2011 |
|
RU2458145C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ПОЛИМОРФИЗМА ГЕНЕТИЧЕСКИХ МАРКЕРОВ МОЛОЧНОЙ ПРОДУКТИВНОСТИ КРУПНОГО РОГАТОГО СКОТА | 2022 |
|
RU2782833C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ПОЛИМОРФИЗМА rs1128446 ГЕНА ТИОРЕДОКСИНРЕДУКТАЗЫ-1 У ЧЕЛОВЕКА | 2011 |
|
RU2458146C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ПОЛИМОРФИЗМА ГЕНА HSP70.1, ОБУСЛАВЛИВАЮЩЕГО ТЕПЛОВОЙ СТРЕСС КРУПНОГО РОГАТОГО СКОТА | 2023 |
|
RU2826316C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АЛЛЕЛЕЙ ГЕНА ХЕМОКИНОВОГО РЕЦЕПТОРА CCR2 ПО ПОЛИМОРФНОМУ САЙТУ V64I | 2001 |
|
RU2180922C1 |
| Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT для повышения уровня экспрессии этих целевых генов, способ его получения и применения, штамм Escherichia coli SCS110-AF/VTvaf17-SOD1, или Escherichia coli SCS110-AF/VTvaf17-SOD2, или Escherichia coli SCS110-AF/VTvaf17-SOD3, или Escherichia coli SCS110-AF/VTvaf17-CAT, несущий генотерапевтический ДНК-вектор, способ его получения, способ производства в промышленных масштабах генотерапевтического ДНК-вектора | 2018 |
|
RU2731515C2 |
| Способ выявления мутации p.L265P в гене MYD88 | 2020 |
|
RU2756909C1 |
Изобретение относится к области молекулярной биологии и медицины. Предложенный способ заключается в детекции полиморфизма А40Т в 3 экзоне гена SOD3 методом ПЦР-ПДРФ с использованием пары праймеров F: 5'-TAC-TGT-GTT-CCT-GCC-TGC-TC-3' и R: 5'-AAG-CTG-CCG-GAA-GAG-GAC-G-3'. Способ позволяет осуществлять генотипирование полиморфизма генов ферментов антиоксидантной защиты у человека и расширяет арсенал методов генотипирования. 1 ил.
Способ обнаружения мутации А40Т в гене внеклеточной супероксиддисмутазы, отличающийся тем, что обнаружение мутации проводят методами ПЦР-ПДРФ с использованием специально подобранных праймеров SEQ ID NO 1-2 и эндонуклеазы BstHHI для дифференцировки аллелей 40А и 40Т гена SOD3, причем генотипу 40АА соответствуют длины рестрикционных фрагментов размерами 162 пары нуклеотидов, генотипу 40ТТ - длины рестрикционных фрагментов размерами 212 пар нуклеотидов, генотипу 40АТ - длины рестрикционных фрагментов размерами 162 и 212 пар нуклеотидов, а рестрикционные фрагменты 48, 39 и 2 на геле не визуализируются.
| EIKEN H.G., ODLAND E., BOMAN H., SKJELKVALE L., ENGEBRETSEN L.F., APOLD J | |||
| Application of natural and amplification created restriction sites for the diagnosis of PKU mutations | |||
| Nucleic Acids Res | |||
| Циркуль-угломер | 1920 |
|
SU1991A1 |
| TAMAI M., FURUTA H., KAWASHIMA H., DOI A., HAMANISHI T., SHIMOMURA H., et al | |||
| Extracellular superoxide dismutase gene | |||

