Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT для повышения уровня экспрессии этих целевых генов, способ его получения и применения, штамм Escherichia coli SCS110-AF/VTvaf17-SOD1 или Escherichia coli SCS110-AF/VTvaf17-SOD2 или Escherichia coli SCS110-AF/VTvaf17-SOD3 или Escherichia coli SCS110-AF/VTvaf17-CAT, несущий генотерапевтический ДНК-вектор, способ его получения, способ производства в промышленных масштабах генотерапевтического ДНК-вектора.
Область техники
Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии.
Уровень техники
Генная терапия - это современный медицинский подход, направленный на лечение наследственных и приобретенных заболеваний путем введения нового генетического материала в клетки пациента с целью компенсации или подавления функции мутантного гена и/или исправления генетического дефекта. Конечным продуктом экспрессии гена может являться молекула РНК или белка. Однако осуществление большей части физиологических процессов в организме связано с функциональной активностью белковых молекул, тогда как молекулы РНК являются либо промежуточным продуктом в синтезе белков, либо осуществляют регуляторные функции. Таким образом целью генной терапии является, в большинстве случаев, введение в организм генов, обеспечивающих транскрипцию и последующую трансляцию белковых молекул, кодируемых этими генами. В рамках описания настоящего изобретения под экспрессией гена подразумевается продукция белковой молекулы, аминокислотная последовательность которой кодируется этим геном.
Гены SOD1, SOD2, SOD3, CAT входящие в группу генов, играют ключевую роль в ряде процессов в организме человека и животных. Показана связь низких/недостаточных концентраций этих белков с различными неблагоприятными состояниями человека, которая, в ряде случаев, подтверждена нарушениями в нормальной экспрессии генов, кодирующих эти белки. Таким образом, генотерапевтическое повышение экспрессии гена, выбранного из группы генов SOD1, SOD2, SOD3, CAT обладает потенциалом для коррекции различных состояний человека и животных.
Активные формы кислорода необходимы для энергетического баланса организма, а также для борьбы с инфекционными агентами, детоксикации ксенобиотиков, регуляции структурных процессов (пролиферации, дифференцировки и апоптоза). Вместе с тем высокая реакционная способность кислорода, особенно его активных форм, участвующих в разнообразных патологических процессах (воспаление, лихорадка, гиперемия, ишемия и другие нарушения работы организма), определяет целесообразность существования многоуровневой системы антиоксидантной защиты. В нормальных физиологических условиях небольшие количества кислорода постоянно конвертируются в супероксид-анионы, перекись водорода и гидроксильные радикалы. Избыточная продукция этих радикалов выступает в роли фактора повреждения, компенсаторным механизмом которого является антиоксидантная система. Главный компонент этой системы - сеть ферментов: супероксиддисмутаз (SOD), глютатионпероксидаз (GPX), каталаза (CAT) и параоксоназа (PON). При этом активность ферментов эволюционно и генетически запрограммирована для оптимизации баланса окислительных процессов и активности систем антиокислительной защиты.
Среди этих ферментов в первую очередь следует выделить SOD - супероксиддисмутазы. Эти ферменты активны во всех клетках, потребляющих кислород. Роль SOD заключается в ускорении реакции превращения токсичного для организма кислородного радикала - супероксида - в перекись водорода и молекулярный кислород. У млекопитающих известно три типа SOD: цитозольная (Cu/Zn-SOD; SOD1), митохондриальная (Mn-SOD; SOD2) и внеклеточная (EC-SOD; SOD3). Ген SOD1 кодирует супероксиддисмутазу I, которая локализуется в ядре, цитоплазме и митохондриях клеток. В настоящее время известно 113 мутаций гена SOD1 у больных с боковым амиотрофическим склерозом, которые в основном затрагивают экзоны и, гораздо реже, некодирующие области гена и связаны с повреждением нейронов (Kaur SJ et al. // Gene. 2016 Feb 15;577(2):109-18). При этом подход с использованием модифицированных генноинженерными методами аллогенных клеток, экспрессирующих нормальный вариант гена SOD1, рассматривается как один из перспективных подходов для терапии бокового амиотрофического склероза (Wang L et al. // Protein Cell. 2017 May;8(5):365-378). В тоже время генотерапевтический подход с использованием герперсвирусного вектора, экспрессирующего ген SOD1, позволил на более чем 50% увеличить выживаемость нейронов после индуцированного инсульта у лабораторных животных (Davis et al. // Neurosci Lett. 2007 Jan 3;411(1):32-6). В 1993 г. рекомбинантный белок SOD1, полученный в дрожжах, был успешно использован для терапии 4 пациентов с анемией Фанкони, однако дальнейшего продолжения этих клинических испытаний не последовало (Liu JM et al. // Br J Haematol. 1993 Oct;85(2):406-8).
Ген SOD2 кодирует марганецзависимую супероксиддисмутазу (Mn-SOD), которая локализуется в митохондриях клеток. Описано около 60 однонуклеотидных мутаций. Широко изучен в разных популяциях полиморфизм Ala16Val, замена аланина на валин в 16-м положении последовательности пептида. Показано, что данный полиморфизм влияет на вторичную структуру сигнального пептида и приводит к дестабилизации его альфа-спирального участка, что влияет на перенос фермента из цитоплазмы в митохондриальный матрикс. В результате данный полиморфизм, по мнению ряда авторов, может приводить к абсолютному или относительному локальному дефициту фермента (Ergen H.A et al. Anticancer Res. 2007; 27 (2): 1227-1230). Наибольшее количество исследований посвящено ассоциации данного полиморфизма с онкологической патологией: раком молочной железы, яичников, предстательной железы и др. (Kang D. et al. // Cancer Epidemiol Biomarkers Prev. 2007; 16: 1581-1586). L. Han и соавт., изучив ассоциацию полиморфизма с риском развития злокачественных образований в корейской и китайской популяциях, указывают на триггерную функцию аллеля Val данного полиморфизма в отношении рака желудка и кишечника (Han L et al. // APMIS. 2012; 10: 463-466). Н.А. Семёнова и соавт. указывают на превышение частоты встречаемости аллеля Val полиморфизма Ala16Val у больных хроническим вирусным гепатитом С из русской популяции Томской области Semenova N.A et al. // Bulletin of Siberian medicine. 2009; 3: 64-69). Согласно результатам исследования роли полиморфизма Ala16Val гена SOD2 в реализации ишемической болезни сердца (ИБС), у пациентов, перенесших острый инфаркт миокарда, зарегистрировано статистически значимое увеличение частоты аллеля Val (р=0,008). При этом расчет показателей относительного риска показывает, что носители аллели Val (ОШ=1,89, 95%) в 2 раза больше подвержены риску развития инфаркта миокарда при ИБС (Maikopova E.V. et al. // Dnepropetrovsk. 2011; 4: 6-9).
В экспериментальной работе было показано, что субретинальное введение аденоассоциированного вирусного вектора, экспрессирующего ген SOD2, предотвращало дегенеративное изменение сетчатки глаза у мышей и может быть использовано для разработки подходов к терапии акулодистрофии (Biswal MR et al. // Invest Ophthalmol Vis Sci. 2017 Feb 1;58(2):1237-1245). Аналогичные положительные результаты с использованием аденоассоциированного вектора были получены и для других офтальмологических заболеваний (Qi X et al. // Ann Neurol. 2004 Aug;56(2):182-91).
Генотерапевтический подход с использованием ДНК-векторов, экспрессирующих ген SOD2, может повышать чувствительность опухолевых клеток к радиотерапии, при этом оказывая протективный эффект на нормальные ткани организма (Epperly MW et al. // Radiat Res. 2007 Mar;167(3):289-97).
Ген SOD3 кодирует внеклеточную супероксиддизмутазу (EC-SOD), которая представляет собой тетрамономер, содержащий в каждой субъединице по одному атому меди и цинка. В кровеносных сосудах SOD3 связана с поверхностью эндотелиальных клеток и внеклеточным матриксом (Nozik-Grayck E., Suliman H.B., Piantadosi C.A. // Int J Biochem Cell Biol. 2005; 37: 2466-2471). Наиболее изучен полиморфизм Arg213Gly гена SOD3, замена аргинина на глицин в 213-м положении полипептидной цепи. Этот полиморфизм объясняют не сниженной экспрессией или ферментативной активностью, а ухудшенным связыванием SOD3 с поверхностью клеток. K. Juul и соавт. доказали увеличение в 1,5 раза риск ишемической болезни сердца и ишемических цереброваскулярных заболеваний у жителей Дании - носителей полиморфизма Arg213Gly гена SOD3 (Juul K et al. // Circulation. 2004; 109: 59-65). И.А. Строковым и соавт. была показана ассоциация полиморфизма Arg213Gly с диабетической полинейропатией при сахарном диабете 1-го типа (Strokov I et al. // Genetics and pathogenesis of effects. 2003; 2: 3-5). При этом гиперэкспрессия SOD3 может положительно влиять на липидный метаболизм и быть протективным фактором в развитии инсулин резистентности (Cui R et al. // Gene Ther. 2014 Sep;21(9):840-8).
Было показано, что в тканях рака простаты экспрессия гена SOD3 значительно снижена, а введение рекомбинантного белка SOD3 способно снизить онкогенный потенциал раковых клеток (Kim J et al. // Anticancer Res. 2014 Jun;34(6):2821-31).
SOD3 способна ингибировать опосредованное нейтрофилами воспаление и может использоваться в качестве нового терапевтического агента против ROS-зависимого повреждения тканей, вызванного нейтрофилами. Доклинические исследования с бычьим SOD3 показали обнадеживающие результаты для его использования в качестве терапевтического агента при острых и хронических воспалительных состояниях, включая дерматозы и раны (Younus H. // International Journal of Health Sciences. 2018;12(3):88-93).
Также было обнаружено, что активность SOD3 у пациентов с ревматоидным артритом низкая, а введение SOD3 положительно влияет на лечение экспериментального артрита (Younus H. // International Journal of Health Sciences. 2018;12(3):88-93).
Подкожное введение генномодифицированных мезенхимальных стволовых клеток, экспрессирующих ген SOD3, предотвращало развитие химически-индуцированного псориаза у лабораторных мышей и оказывало иммуномодулирующее действие (Sah SK et al. // Antioxid Redox Signal. 2016 Feb 10;24(5):233-48).
Следующим биохимическим звеном антиоксидативной системы организма является разложение пероксида водорода до нетоксичных метаболитов и воды с помощью каталазы и ферментов семейства глутатионпероксидаз. Ген CAT кодирует белок каталазу - фермент, участвующий в защите организма от оксидативного стресса и токсического воздействия активных форм кислорода, путем расщепления пероксида водорода. Каталаза присутствует в составе пероксисом практически всех аэробных клеток. Известно, что оксидативный стресс играет роль в патогенезе различных заболеваний, таких как диабет, астма, болезнь Альцгеймера, системная красная волчанка, ревматоидный артрит, рак. Однако единственным наследственным заболеванием, доказанная причина которого заключается в полиморфизме гена CAT, является акаталаземия (синдром Такахары). Тем не менее снижение экспрессии гена CAT может усугублять течение ряда заболеваний, например, бронхиальной астмы у детей (Babusikova E et al. // Arch Bronconeumol. 2013 Dec;49(12):507-12).
В исследованиях полиморфизма С262T гена САТ наблюдалось повышение риска развития ИБС при замене цитозина на тимидин в позиции 262, при этом риск стенокардии у индивидуумов с данным генотипом увеличивается в 7 раз (Maikopova et al. // Dnepropetrovsk. 2011; 4: 6-9). Другой полиморфизм гена САТ был ассоциирован с развитием инсульта у мужчин на фоне гипертонической болезни. Стратифицированный анализ пациентов-носителей этого варианта гена САТ с учетом средовых факторов риска показал повышение риска развития инсульта у курящих и злоупотребляющих алкоголем мужчин (Ahn J et al. // Cancer Epidemiol Biomarkers Prev. 2006; 15 (6): 1217-1222).
Использование генотерапевтического подхода с применением аденовирусного вектора, экспрессирующего ген САТ, приводило к значительному улучшению физиологических параметров, связанных с почечной ишемией у лабораторных животных (Yang CC et al. // Chin J Physiol. 2015 Dec 31;58(6):420-30).
Также использование аденоассоциированных векторов, экспрессирующих ген САТ, способно предотвращать старение организма и связанные с этим дегенеративные процессы у мышей (Li D, Duan D. // Methods Mol Biol. 2013;1048:161-80).
Предотвращение развития атеросклероза также является областью, в которой генотерапевтический подход для повышения экспрессии каталазы признан одним из многообещающих направлений развития биомедицины (Van-Assche T, Huygelen V, Crabtree MJ, Antoniades C. // Curr Pharm Des. 2011 Dec;17(37):4210-23. Review).
Раннее поседение волос также связывают с нарушением активности каталазы или ее сниженной экспрессией в волосяных фолликулах, механизм этого процесса связан с разложением в клетках перекиси водорода, которая обесцвечивает волосы (Seiberg M. // Int J Cosmet Sci. 2013 Dec;35(6):532-8).
Также известно, что EUK-207 - миметик SOD / каталазы, оказывает терапевтическое действие и предотвращает прогрессию амилоидов и tau-патологии, а также предупреждает снижение когнитивной функции на мышиной модели болезни Альцгеймера (Younus H. // International Journal of Health Sciences. 2018;12(3):88-93).
Тем не менее основными ограничениями для разработки терапевтических средств на основе супероксиддисмутаз и каталазы является их низкая способность проникать в клетки организма и небольшое время полужизни в организме. Таким образом генотерапевтический подход, обеспечивающий in vivo экспрессию этих ферментов является приоритетным направлением в этой области (Younus H. // International Journal of Health Sciences. 2018;12(3):88-93).
Таким образом предшествующий уровень техники свидетельствует о том, что мутации в генах SOD1, SOD2, SOD3, CAT или недостаточная экспрессия белков, кодируемых этими генами, связаны с развитием спектра заболеваний, включающих в себя, но не ограничивающихся, отклонениями, в которых оксидативный стресс играет патогенетическую роль, сердечно-сосудистыми, дегенеративными и аутоиммунными заболеваниями, раком, наследственными и приобретенными патологическими процессами, такими как повреждения тканей, и другими неблагоприятными для организма состояниями. Этим обусловлено объединение генов SOD1, SOD2, SOD3, CAT в рамках данного патента в группу генов. Генетические конструкции, обеспечивающие экспрессию белков, кодируемых генами из группы SOD1, SOD2, SOD3, CAT, могут быть использованы для разработки лекарственных препаратов для предотвращения и терапии различных заболеваний и патологических состояний.
Более того, приведенные данные свидетельствуют о том, что недостаточная экспрессия белков, кодируемых генами SOD1, SOD2, SOD3, CAT, входящими в группу генов, связана не только с патологическими состояниями, но и с предрасположенность к их развитию. Также приведенные данные свидетельствуют о том, что недостаточная экспрессия данных белков может не проявляться в явном виде в форме патологии, которая может быть однозначно описана в рамках существующих стандартов клинической практики (например, с применением кода МКБ), однако при этом вызывать состояния, которые неблагоприятны для человека и животных и связанны с ухудшением качества жизни.
Анализ подходов для повышения экспрессии целевых генов подразумевает возможность использования различных генотерапевтических векторов.
Генотерапевтические векторы разделяют на вирусные, клеточные и ДНК-векторы (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal Products EMA/CAT/80183/2014). В последнее время в генной терапии всё большее внимание уделяется разработке невирусных систем доставки генетического материала, среди которых лидируют плазмидные векторы. Плазмидные векторы лишены недостатков, присущих клеточным и вирусным векторам. В клетке-мишени они существуют в эписомальной форме, не интегрируют в геном, производство их достаточно дешево, отсутствие иммунного ответа и побочных реакций на введение плазмидного вектора делают их удобным инструментом генной терапии и генетической профилактики (ДНК-вакцины) (Li L, Petrovsky N. // Expert Rev Vaccines. 2016;15(3):313-29).
Тем не менее, ограничениями для использования плазмидных векторов для генной терапии являются: 1) наличие генов устойчивости к антибиотикам для наработки в бактериальных штаммах, 2) наличие различных регуляторных элементов, представленных последовательностями вирусных геномов 3) размер терапевтического плазмидного вектора, определяющий эффективность проникновения вектора в клетку-мишень.
Известно, что Европейское агентство по лекарственным средствам считает необходимым избегать введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies). Данная рекомендация связана, в первую очередь, с потенциальной опасностью проникновения ДНК-вектора или горизонтального переноса генов антибиотикорезистентности в клетки бактерий, представленных в организме в составе нормальной или оппортунистической микрофлоры. Помимо этого, наличие генов антибиотикорезистентности значительно увеличивает размер ДНК-вектора, что приводит к снижению эффективности его проникновения в эукариотические клетки.
Необходимо отметить, что гены антибиотикорезистентности также вносят принципиальный вклад в способ получения ДНК-векторов. В случае наличия генов антибиотикорезистентности штаммы для наработки ДНК-векторов обычно культивируются в среде, содержащей селективный антибиотик, что создает риск наличия следовых количеств антибиотика в недостаточно очищенных препаратах ДНК-векторов. Таким образом, получение ДНК-векторов для генной терапии, в которых отсутствуют гены антибиотикорезистентности, связано с получением штаммов, обладающих такой отличительной особенностью как способность к стабильной амплификации целевых ДНК-векторов в среде без содержания антибиотиков.
Кроме того, Европейское Медицинское Агентство рекомендует избегать наличия в составе терапевтических плазмидных векторов регуляторных элементов для повышения экспрессии целевых генов (промоторов, энхансеров, посттрансляционных регуляторных элементов), являющихся нуклеотидными последовательностями геномов различных вирусов (Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products,http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf). Данные последовательности, хотя и могут увеличивать уровень экспрессии целевого трансгена, однако создают риск рекомбинации с генетическим материалом вирусов дикого типа и интеграции в геном эукариотической клетки. Более того, целесообразность гиперэкспрессии того или иного гена в целях терапии остается нерешенным вопросом.
Также, существенным моментом является размер терапевтического вектора. Известно, что современные плазмидные векторы зачастую перегружены нефункциональными участками, серьезно увеличивающими размер вектора (Mairhofer J, Grabherr R. // Mol Biotechnol. 2008.39(2):97-104). Например, ген устойчивости к ампициллину в векторах серии pBR322, как правило, состоит из не менее чем 1000 п.н., что составляет более 20% от размера самого вектора. При этом наблюдается обратная зависимость между размером вектора и его способностью проникать в эукариотические клетки - ДНК-векторы с небольшим размером эффективней проникаю в клетки человека и животных. Так, например, в серии экспериментов по трансфекции клеток HELA ДНК-векторами с размером от 383 до 4548 п.н. было показано, что разница в эффективности проникновения может достигать двух порядков (отличаться в 100 раз) (Hornstein BD et al. // PLoS ONE. 2016;11(12): e0167537.).
Таким образом при выборе ДНК-вектора в целях безопасности и наибольшей эффективности следует отдавать предпочтение тем конструкциям, в которых не содержатся гены устойчивости к антибиотикам, последовательности вирусного происхождения и размер которых позволяет эффективно проникать в эукариотические клетки. Штамм для получения такого ДНК-вектора в количествах, достаточных для целей генной терапии, должен обеспечивать возможность стабильной амплификации ДНК-вектора с использованием питательных сред, не содержащих антибиотики.
Примером использования рекомбинантных ДНК-векторов для генной терапии является способ получения рекомбинантного вектора для генетической иммунизации по патенту US 9550998 В2. Плазмидный вектор представляет собой суперскученный плазмидный ДНК-вектор и предназначен для экспрессии клонированных генов в клетках животных и человека. Вектор состоит из ориджина репликации, регуляторных элементов, включающих промотор и энхансер цитомегаловируса человека, регуляторные элементы из Т-лимфотропного вируса человека.
Накопление вектора проводят в специальном штамме E. coli без использования антибиотиков за счет антисенс-комплементации гена sacB, введенного в штамм посредством бактериофага. Недостатком данного изобретения является наличие в составе ДНК-вектора регуляторных элементов, представляющих собой последовательности вирусных геномов.
Прототипами настоящего изобретения в части использования генотерапевтических подходов для повышения уровня экспрессии генов из группы SOD1, SOD2, SOD3, CAT являются следующие заявки.
В заявке JP2009501009A описывается изобретение, в основе которого лежит аденоассоциированный вирусный вектор, экспрессирующий ген SOD1, используемый для терапии ряда заболеваний и неблагоприятных состояний человека, связанных с недостаточной экспрессией этого гена. Недостатком данного изобретения является использование вирусного вектора, интегрирующего в геном человека.
В патенте RU2651757C2 описан способ компенсации недостаточной экспрессии гена SOD2, в том числе с использованием генотерапевтического подхода на основе плазмидных векторов. Недостатком данного изобретения являются неустановленные требования к безопасности используемых векторов.
В патенте CA2273173A1 описан способ увеличения продолжительности жизни за счет введения в организм вектора, экспрессирующего ген SOD3. Недостатком данного изобретения являются неустановленные требования к безопасности используемых векторов, а также ограниченность использования подхода только увеличением продолжительности жизни.
В заявке CN102080102A описан генотерапевтический вектор на основе аденовирусного вектора, экспрессирующего ген CAT, и способ его получения. Недостатком данного изобретения является использование вирусного вектора, а также неопределенная область применения данного изобретения для человека и животных.
Раскрытие изобретения
Задачей изобретения является конструирование генотерапевтических ДНК-векторов для повышения уровня экспрессии генов, выбранных из группы генов: SOD1, SOD2, SOD3, CAT в организме человека и животных, сочетающих в себе следующие свойства:
(I) Эффективность генотерапевтического ДНК-вектора для повышения уровня экспрессии целевых генов в эукариотических клетках.
(II) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов.
(III) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора генов антибиотикорезистентности.
(IV) Технологичность получения и возможность наработки генотерапевтического ДНК-вектора в промышленных масштабах.
Пункты II и III предусмотрены в данном техническом решении в соответствии с рекомендациями государственных регуляторов к лекарственным средствам для генной терапии, в частности, Европейского Агентства по лекарственным средствам касательно отказа от введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011, EMA/CAT/GTWP/44236/2009 Committee for advanced therapies) и касательно отказа от введения в разрабатываемые плазмидные векторы для генной терапии элементов вирусных геномов (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products / 23 March 2015, EMA/CAT/80183/2014, Committee for Advanced Therapies).
Задачей изобретения также является конструирование штаммов, несущих эти генотерапевтические ДНК-вектора, для наработки и производства в промышленных масштабах генотерапевтических ДНК-векторов.
Поставленная задача решается за счет того, что создан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT, для повышения уровня экспрессии этого целевого гена в организме человека и животных, при этом генотерапевтический ДНК-вектор VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT имеет нуклеотидную последовательность SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 соответственно. При этом каждый из созданных генотерапевтических ДНК-векторов: VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT за счет ограниченного размера векторной части VTvaf17, не превышающей 3200 п.н., обладает способностью эффективно проникать в клетки и экспрессировать клонированный в него целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT соответственно. В составе генотерапевтического ДНК-вектора отсутствуют нуклеотидные последовательности вирусного происхождения и отсутствуют гены антибиотикорезистентности, обеспечивая возможность его безопасного применения для генетической терапии человека и животных.
Создан также способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: ген SOD1, ген SOD2, ген SOD3, ген CAT, который заключается в том, что каждый из генотерапевтических ДНК-векторов: VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT получают следующим образом: кодирующую часть целевого гена из группы SOD1, или SOD2, или SOD3, или CAT клонируют в ДНК-вектор VTvaf17 и получают генотерапевтический ДНК-вектор VTvaf17-SOD1, SEQ ID №1, или VTvaf17-SOD2, SEQ ID №2 или VTvaf17-SOD3, SEQ ID №3, или VTvaf17-CAT, SEQ ID №4 соответственно.
Способ применения созданного генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: ген SOD1, ген SOD2, ген SOD3, ген CAT для повышения уровня экспрессии этих целевых генов, заключается во введении выбранного генотерапевтического ДНК-вектора или нескольких выбранных генотерапевтических ДНК-векторов в клетки, органы и ткани человека или животного, и/или во введении в органы и ткани человека или животного аутологичных клеток человека или животного, трансфицированных выбранным генотерапевтическим ДНК-вектором или несколькими выбранными генотерапевтическими ДНК-векторами, или в сочетании обозначенных способов.
Способ получения штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT заключается в электропорации компетентных клеток штамма Escherichia coli SCS110-AF созданным генотерапевтическим ДНК-вектором и последующей селекцией стабильных клонов штамма с использованием селективной среды.
Заявлен штамм Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамм Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамм Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамм Escherichia coli SCS110-AF/VTvaf17-CAT, несущий генотерапевтический ДНК-вектор для его наработки с возможностью культивирования штамма без использования антибиотиков.
Способ производства в промышленных масштабах генотерапевтического ДНК-вектора заключается в масштабировании бактериальной культуры штамма до количеств, необходимых для наращивания бактериальной биомассы в промышленном ферментере, после чего биомассу используют для выделения фракции, содержащей целевой ДНК-продукт - генотерапевтический ДНК-вектор VTvaf17-SOD1 или VTvaf17-SOD2 или VTvaf17-SOD3 или VTvaf17-CAT, многостадийно фильтруют и очищают хроматографическими методами. Изобретение поясняется чертежами, где:
На фиг.1
приведена схема генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT, который представляет собой кольцевую двуцепочечную молекулу ДНК, способную к автономной репликации в клетках бактерии Escherichia coli.
На фиг.1 приведены схемы, соответствующие:
A - генотерапевтический ДНК-вектор VTvaf17-SOD1,
B - генотерапевтический ДНК-вектор VTvaf17-SOD2,
C - генотерапевтический ДНК-вектор VTvaf17-SOD3,
D - генотерапевтический ДНК-вектор VTvaf17-CAT.
На схемах отмечены следующие структурные элементы вектора:
EF1a - промоторная область гена человеческого фактора элонгации EF1Aс собственным энхансером, содержащимся в первом интроне гена. Служит для обеспечения высокого уровня транскрипции рекомбинантного гена в большинстве тканей человека;
Рамка считывания целевого гена, соответствующая кодирующей части гена SOD1 (фиг. 1A), или SOD2 (фиг. 1B), или SOD3 (фиг. 1C), или CAT (фиг. 1D) соответственно;
hGH-TA - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
ori - ориджин репликации, служащий для автономной репликации с однонуклеотидной заменой для повышения копийности плазмиды в клетках большинства штаммов Escherichia coli;
RNA-out - регуляторный элемент РНК-out транспозона Tn 10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Eshcerichia coli SCS 110.
Отмечены уникальные сайты рестрикции.
На фиг.2
показаны графики накопления ампликонов кДНК целевого гена, а именно, гена SOD1, в первичной культуре клеток фибробластов кожи человека HDFa (ATCC PCS-201-012) до их трансфекции и через 48 часов после трансфекции этих клеток генотерапевтическим ДНК-вектором VTvaf17-SOD1 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.2 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена SOD1 в первичной культуре клеток фибробластов кожи человека HDFa до трансфекции ДНК-вектором VTvaf17-SOD1
2 - кДНК гена SOD1 в первичной культуре клеток фибробластов кожи человека HDFa после трансфекции ДНК-вектором VTvaf17-SOD1
3 - кДНК гена B2M в первичной культуре клеток фибробластов кожи человека HDFa до трансфекции ДНК-вектором VTvaf17-SOD1
4 - кДНК гена B2M в первичной культуре клеток фибробластов кожи человека HDFa после трансфекции ДНК-вектором VTvaf17-SOD1.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.3
показаны графики накопления ампликонов кДНК целевого гена, а именно гена SOD2, в первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa (ATCC PCS-200-011) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-SOD2 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.3 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена SOD2 в первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa до трансфекции ДНК-вектором VTvaf17-SOD2;
2 - кДНК гена SOD2 первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa после трансфекции ДНК-вектором VTvaf17-SOD2;
3 - кДНК гена B2M в первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa до трансфекции ДНК-вектором VTvaf17-SOD2;
4 - кДНК гена B2M первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa после трансфекции ДНК-вектором VTvaf17-SOD2.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.4
показаны графики накопления ампликонов кДНК целевого гена, а именно гена SOD3 в клетках скелетных миобластов человека Human Skeletal Myoblasts (HSKM) (GIBCO® Кат. A12555) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-SOD3 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.4 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена SOD3 в клетках скелетных миобластов человека линии HSKM до трансфекции ДНК-вектором VTvaf17-SOD3;
2 - кДНК гена SOD3 клетках скелетных миобластов человека линии HSKM после трансфекции ДНК-вектором VTvaf17-SOD3;
3 - кДНК гена B2M клетках скелетных миобластов человека линии HSKM до трансфекции ДНК-вектором VTvaf17-SOD3;
4 - кДНК гена B2M клетках скелетных миобластов человека линии HSKM после трансфекции ДНК-вектором VTvaf17-SOD3.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.5
показаны графики накопления ампликонов кДНК целевого гена, а именно гена CAT, в первичной культуре клеток эндотелия человека линии Primary Umbilical Vein Endothelial Cells; Normal, Human (HUVEC) (ATCC® PCS-100-010™) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-CAT с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.5 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена CAT в первичной культуре клеток эндотелия человека линии HUVEC до трансфекции ДНК-вектором VTvaf17-CAT;
2 - кДНК гена CAT в первичной культуре клеток эндотелия человека линии HUVEC после трансфекции ДНК-вектором VTvaf17-CAT;
3 - кДНК гена B2M в первичной культуре клеток эндотелия человека линии HUVEC до трансфекции ДНК-вектором VTvaf17-CAT;
4 - кДНК гена B2M в первичной культуре клеток эндотелия человека линии HUVEC после трансфекции ДНК-вектором VTvaf17-CAT.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.6
показана диаграмма концентрации белка SOD1 в клеточном лизате первичной культуры фибробластов кожи человека HDFa (ATCC PCS-201-01) после трансфекции этих клеток ДНК-вектором VTvaf17- SOD1 с целью оценки функциональной активности, то есть экспрессии на уровне белка, по изменению количества белка SOD1 в лизате клеток.
На фиг.6 отмечены следующие элементы:
культура А - культура клеток первичных фибробластов кожи человека HDFa, трансфицированная водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток первичных фибробластов кожи человека HDFa, трансфицированных ДНК-вектором VTvaf17;
культура C - культура клеток первичных фибробластов кожи человека HDFa, трансфицированных ДНК-вектором VTvaf17-SOD1.
На фиг. 7
показана диаграмма концентрации белка SOD2 в лизате клеток первичной культуры кератиноцитов эпидермиса человека HEKa (ATCC PCS-200-01), после трансфекции этих клеток ДНК-вектором VTvaf17-SOD2 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SOD2.
На фиг.7 отмечены следующие элементы:
культура А - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированная водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированная ДНК-вектором VTvaf17;
культура C - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированных ДНК-вектором VTvaf17-SOD2.
На фиг.8
показана диаграмма концентрации белка SOD3 в кондиционированной среде с клеток скелетных миобластов человека Human Skeletal Myoblasts (HSKM) (GIBCO® Кат. A12555) после трансфекции этих клеток ДНК-вектором VTvaf17-SOD3 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SOD3.
На фиг.8 отмечены следующие элементы:
культура А - среда с клеток скелетных миобластов человека линии HSKM, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - среда с клеток скелетных миобластов человека линии HSKM, трансфицированных ДНК-вектором VTvaf17;
культура C - среда с клеток скелетных миобластов человека линии HSKM, трансфицированных ДНК-вектором VTvaf17-SOD3.
На фиг. 9
показана диаграмма концентрации белка CAT в лизате клеток первичной культуры эндотелия человека линии Primary Umbilical Vein Endothelial Cells; Normal, Human (HUVEC) (ATCC® PCS-100-010™) после трансфекции этих клеток ДНК-вектором VTvaf17-CAT с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген CAT.
На фиг.9 отмечены следующие элементы:
культура А - культура клеток эндотелия человека линии HUVEC, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток эндотелия человека линии HUVEC, трансфицированных ДНК-вектором VTvaf17;
культура C - культура клеток эндотелия человека линии HUVEC, трансфицированных ДНК-вектором VTvaf17-CAT.
На фиг. 10
показана диаграмма концентрации белка CAT в биоптатах кожи трех пациентов после введения в кожу этих пациентов генотерапевтического ДНК-вектора VTvaf17-CAT с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген CAT.
На фиг.10 отмечены следующие элементы:
П1I - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17-CAT;
П1II - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П1III - биоптат кожи пациента П1 из интактного участка;
П2I - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17-CAT;
П2II - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П2III - биоптат кожи пациента П2 из интактного участка;
П3I - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17-CAT;
П3II - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П3III - биоптат кожи пациента П3 из интактного участка.
На фиг. 11
показана диаграмма концентрации белка SOD3 в биоптатах икроножной мышцы трех пациентов после введения в икроножную мышцу этих пациентов генотерапевтического ДНК-вектора VTvaf17- SOD3, с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SOD3.
На фиг.11 отмечены следующие элементы:
П1I - биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17- SOD3;
П1II - биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П1III - биоптат интактного участка икроножной мышцы пациента П1;
П2I - биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17- SOD3;
П2II - биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П2III - биоптат интактного участка икроножной мышцы пациента П2;
П3I - биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17- SOD3;
П3II - биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П3III - биоптат интактного участка икроножной мышцы пациента П3.
На фиг.12
показана диаграмма концентрации белка SOD2 в биоптатах кожи трех пациентов после введения в кожу этих пациентов генотерапевтического ДНК-вектора VTvaf17-SOD2 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SOD2.
На фиг.12 отмечены следующие элементы:
П1I - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17-SOD2;
П1II - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П1III - биоптат кожи пациента П1 из интактного участка;
П2I - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17-SOD2;
П2II - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П2III - биоптат кожи пациента П2 из интактного участка;
П3I - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17-SOD2;
П3II - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
П3III - биоптат кожи пациента П3 из интактного участка.
На фиг. 13
показана диаграмма концентрации белка SOD2 в биоптатах кожи человека после введения в кожу культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2 с целью демонстрации способа применения путем введения аутологичных клеток, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2.
На фиг.13 отмечены следующие элементы:
П1C - биоптат кожи пациента П1 в зоне введения культуры аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2;
П1B - биоптат кожи пациента П1 в зоне введения аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17;
П1A - биоптат кожи пациента П1 из интактного участка.
На фиг. 14
показана диаграмма концентраций белков: белка SOD1 человека, белка SOD2 человека, белка SOD3 человека, белка CAT человека в биоптатах кожи трех крыс после инъекционного введения смеси генотерапевтических векторов: генотерапевтического ДНК-вектора VTvaf17-SOD1, генотерапевтического ДНК-вектора VTvaf17-SOD2, генотерапевтического ДНК-вектора VTvaf17-SOD3, генотерапевтического ДНК-вектора VTvaf17-CAT с целью демонстрации способа применения смеси генотерапевтических ДНК-векторов.
На фиг.14 отмечены следующие элементы:
К1I - биоптат кожи крысы К1 в зоне введения смеси генотерапевтических ДНК векторов: VTvaf17- SOD1, VTvaf17- SOD2, VTvaf17- SOD3 и VTvaf17- CAT
К1II - биоптат кожи крысы К1 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
К1III - биоптат контрольного интактного участка кожи крысы К1,
К2I - биоптат кожи крысы К2 в зоне введения смеси генотерапевтических ДНК векторов: VTvaf17-SOD1, VTvaf17-SOD2, VTvaf17-SOD3 и VTvaf17-CAT;
К2II - биоптат кожи крысы К2 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
К2III - биоптат контрольного интактного участка кожи крысы К2;
К3I - биоптат кожи крысы К3 в зоне введения смеси генотерапевтических ДНК векторов: VTvaf17- SOD1, VTvaf17- SOD2, VTvaf17- SOD3 и VTvaf17- CAT;
К3II - биоптат кожи крысы К3 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо);
К3III - биоптат контрольного интактного участка кожи крысы К3.
На фиг. 15
показаны графики накопления ампликонов кДНК целевого гена SOD3 в клетках эпителия эндометрия коровы Bovine Endometrial Epithelial Cells (BEnEpC) (Cell Applications, Inc. Кат. B932-05) до и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17- SOD3 с целью демонстрации способа применения путем введения генотерапевтического ДНК-вектора животным.
На фиг.15 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена SOD3 в клетках эпителия эндометрия коровы BEnEpC до трансфекции генотерапевтическим ДНК-вектором VTvaf17- SOD3;
2 - кДНК гена SOD3 в клетках эпителия эндометрия коровы BEnEpC после трансфекции генотерапевтическим ДНК-вектором VTvaf17- SOD3;
3 - кДНК гена ACT в клетках эпителия эндометрия коровы BEnEpC до трансфекции генотерапевтическим ДНК-вектором VTvaf17- SOD3;
4 - кДНК гена ACT в клетках эпителия эндометрия коровы BEnEpC после трансфекции генотерапевтическим ДНК-вектором VTvaf17- SOD3.
В качестве референтного гена использовали ген актина быка/ коровы (АСТ), приведенного в базе данных GenBank под номером AH001130.2.
Реализация изобретения
На основе ДНК-вектора VTvaf17 размером 3165 п.н. созданы генотерапевтические ДНК-векторы, несущие целевые гены человека, предназначенные для повышения уровня экспрессии этих целевых генов в тканях человека и животных. При этом способ получения каждого генотерапевтического ДНК-вектора, несущего целевые гены заключается в том, что в полилинкер генотерапевтического ДНК-вектора VTvaf17 клонируют белок-кодирующую последовательность целевого гена, выбранного из группы генов: ген SOD1 (кодирует белок SOD1), ген SOD2 (кодирует белок SOD2), ген SOD3 (кодирует белок SOD3), ген CAT (кодирует белок CAT) человека. Известно, что способность ДНК-векторов проникать в эукариотические клетки обусловлена, главным образом, размером вектора. При этом ДНК-вектора с наименьшим размером обладают более высокой проникающей способностью. Таким образом, предпочтительным является отсутствие в составе вектора элементов, которые не несут функциональной нагрузки, но при этом увеличивают размер ДНК-вектора. Данные особенности ДНК-векторов были учтены при получении генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT путем отсутствия в составе вектора крупных нефункциональных последовательностей и генов антибиотикорезистентности, что позволило, помимо технологических преимуществ и преимуществ в плане безопасности применения, значительно уменьшить размер полученного генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT. Таким образом, способность проникать в эукариотические клетки полученного генотерапевтического ДНК-вектора обусловлена его небольшими размерами.
Каждый из генотерапевтических ДНК-векторов: ДНК-вектор VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT получали следующим образом: кодирующую часть целевого гена SOD1, или SOD2, или SOD3, или CAT клонировали в генотерапевтический ДНК-вектор VTvaf17 и получали генотерапевтический ДНК-вектор VTvaf17-SOD1, SEQ ID №1, или VTvaf17-SOD2, SEQ ID №2 или VTvaf17-SOD3, SEQ ID №3, или VTvaf17-CAT, SEQ ID №4 соответственно. Кодирующую часть гена SOD1 размером 466 п.н., или гена SOD2 размером 671 п.н., или гена SOD3 размером 725 п.н., или гена CAT размером 1590 п.н. получали путем выделения суммарной РНК из биологического образца ткани здорового человека. Для получения первой цепи кДНК генов SOD1, SOD2, SOD3, CAT человека использовали реакцию обратной транскрипции. Амплификацию проводили с использованием созданных для этого методом химического синтеза олигонуклеотидов. Расщепление продукта амплификации специфическими эндонуклеазами рестрикции проводили с учетом оптимальной процедуры дальнейшего клонирования, причем клонирование в генотерапевтический ДНК-вектор VTvaf17 проводили по сайтам рестрикции BamHI, SalI, EcoRI, HindIII расположенными в полилинкере вектора VTvaf17. Выбор сайтов рестрикции проводили таким образом, чтобы клонированный фрагмент попадал в рамку считывания экспрессионной кассеты вектора VTvaf17, при этом белок-кодирующая последовательность не содержала сайты рестрикции для выбранных эндонуклеаз. При этом специалистам в данной области техники понятно, что методическая реализация получения генотерапевтического ДНК-вектора VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT может варьировать в рамках выбора известных методов молекулярного клонирования генов, при этом эти способы подпадают под объем настоящего изобретения. Так, например, могут быть использованы различные последовательности олигонуклеотидов для амплификации гена SOD1, или SOD2, или SOD3, или CAT различные эндонуклеазы рестрикции или такие лабораторные техники как безлигазное клонирование генов.
Генотерапевтический ДНК-вектор VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT обладает нуклеотидной последовательностью SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 соответственно. При этом специалистам в данной области техники известно свойство вырожденности генетического кода, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей, отличающихся инсерцией, делецией или заменой нуклеотидов, которые не приводят к изменению полипептидной последовательности, кодируемой целевым геном, и/или не приводят к потере функциональной активности регуляторных элементов вектора VTvaf17. При этом специалистам в данной области техники известно явление генетического полиморфизма, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей генов из группы генов SOD1, SOD2, SOD3, CAT, которые при этом кодируют различные варианты аминокислотных последовательностей белков SOD1, SOD2, SOD3, CAT, не отличающихся от приведенных по своей функциональной активности при физиологических условиях.
Способность проникать в эукариотические клетки и функциональную активность, то есть способность экспрессировать целевой ген, полученного генотерапевтического ДНК-вектора VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT подтверждают путем введения в эукариотические клетки полученного вектора и последующим анализом экспрессии специфической мРНК и/или белкового продукта целевого гена. Наличие специфической мРНК в клетках, в которые был введен генотерапевтический ДНК-вектор VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT свидетельствует как о способности полученного вектора проникать в эукариотические клетки, так и о его способности экспрессировать мРНК целевого гена. При этом, как известно специалистам в данной области техники, наличие мРНК гена является обязательным условием, но не доказательством трансляции белка, кодируемого целевым геном. Поэтому для подтверждения свойства генотерапевтического ДНК-вектора VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT экспрессировать целевой ген на уровне белка в эукариотических клетках, в которые был введен генотерапевтический ДНК-вектор, проводят анализ концентрации белков, кодируемых целевыми генами, с использованием иммунологических методов. Наличие белка SOD1, или SOD2, или SOD3, или CAT подтверждает эффективность экспрессии целевых генов в эукариотических клетках и возможность повышения уровня концентрации белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT. Таким образом для подтверждения эффективности экспрессии созданного генотерапевтического ДНК-вектора VTvaf17-SOD1, несущего целевой ген, а именно, ген SOD1, генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего целевой ген, а именно, ген SOD2, генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего целевой ген, а именно, ген SOD3, генотерапевтического ДНК-вектора VTvaf17-CAT, несущего целевой ген, а именно, ген CAT использовали следующие методы:
А) ПЦР в реальном времени - изменение накопления ампликонов кДНК целевых генов в лизате клеток человека и животного, после трансфекции различных клеточных линий человека и животного генотерапевтическим ДНК-векторами;
B) Иммуноферментный анализ - изменение количественного уровня целевых белков в лизате или среде с клеток человека, после трансфекции различных клеточных линий человека генотерапевтическими ДНК-векторами;
C) Иммуноферментный анализ - изменение количественного уровня целевых белков в супернатанте биоптатов тканей человека и животного, после введения в эти ткани генотерапевтических ДНК-векторов;
D) Иммуноферментный анализ - изменение количественного уровня целевых белков в супернатанте биоптатов тканей человека, после введения в эти ткани аутологичных клеток этого человека, трансфицированных генотерапевтическими ДНК-векторами.
Для подтверждения реализуемости способа применения созданного генотерапевтического ДНК-вектора VTvaf17-SOD1, несущего целевой ген, а именно, ген SOD1, генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего целевой ген, а именно, ген SOD2, генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего целевой ген, а именно, ген SOD3, генотерапевтического ДНК-вектора VTvaf17-CAT, несущего целевой ген, а именно, ген CAT выполняли:
А) трансфекцию генотерапевтическими ДНК-векторами различных клеточных линий человека и животного;
B) введение генотерапевтических ДНК-векторов в различные ткани человека и животного;
С) введение в ткани животного смеси генотерапевтических ДНК-векторов;
D) введение в ткани человека аутологичных клеток, трансфицированных генотерапевтическими ДНК-векторами.
Указанные способы применения характеризуются отсутствием потенциальных рисков для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов, и за счет отсутствия в составе генотерапевтического ДНК-вектора генов устойчивости к антибиотикам, что подтверждается отсутствием участков, гомологичных вирусным геномам и генам антибиотикорезистентности в нуклеотидных последовательностях генотерапевтического ДНК-вектора VTvaf17-SOD1, или генотерапевтического ДНК-вектора VTvaf17-SOD2, или генотерапевтического ДНК-вектора VTvaf17-SOD3, или генотерапевтического ДНК-вектора VTvaf17-CAT (SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 соответственно).
Как известно специалистам в данной области техники, гены антибиотикорезистентности в составе генотерапевтических ДНК-векторов используются с целью получения этих векторов в препаративных количествах путем наращивания бактериальной биомассы в питательной среде, содержащей селективный антибиотик. В рамках настоящего изобретения в целях возможности безопасного применения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SOD1 или SOD2, или SOD3 или CAT, использование селективных питательных сред, содержащих антибиотик, не представляется возможным. В качестве технологического решения для получения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT для возможности масштабирования до промышленных масштабов получения генотерапевтических векторов предлагается способ получения штаммов для наработки указанных генотерапевтических векторов на основе бактерии Escherichia coli SCS110-AF. Способ получения штамма Escherichia coli SCS110-AF/VTvaf17-SOD1 или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT заключается в получении компетентных клеток штамма Escherichia coli SCS110-AF с введением в эти клетки генотерапевтического ДНК-вектора VTvaf17-SOD1, или ДНК-вектора VTvaf17-SOD2, или ДНК-вектора VTvaf17-SOD3, или ДНК-вектора VTvaf17-CAT соответственно с помощью методов трансформации (электропорации), общеизвестных специалистам в данной области техники. Полученный штамм Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамм Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамм Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамм Escherichia coli SCS110-AF/VTvaf17-CAT используется для наработки генотерапевтического ДНК-вектора VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT соответственно с возможностью использования сред без содержания антибиотиков.
Для подтверждения получения штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT проводили трансформацию, селекцию и последующее наращивание с выделением плазмидной ДНК.
Для подтверждения технологичности получения и возможности масштабирования до промышленного производства генотерапевтического ДНК-вектора VTvaf17-SOD1, несущего целевой ген, а именно, ген SOD1, генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего целевой ген, а именно, ген SOD2, генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего целевой ген, а именно, ген SOD3 генотерапевтического ДНК-вектора VTvaf17-CAT, несущего целевой ген, а именно, ген CAT, выполняли ферментацию в промышленном масштабе штамма Escherichia coli SCS110-AF/VTvaf17-SOD1 или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2 или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT, каждый из которых содержит генотерапевтический ДНК-вектор VTvaf17, несущий целевой ген, а именно SOD1, или SOD2, или SOD3, или CAT.
Способ масштабирования получения бактериальной массы до промышленных масштабов для выделения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT, заключается в том, что затравочную культуру штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT инкубируют в объеме питательной среды без содержания антибиотика обеспечивающим подходящую динамику накопления биомассы, по достижению достаточного количества биомассы в логарифмической фазе роста, бактериальную культуру переносят в промышленный ферментер, после чего растят до достижения стационарной фазы роста, затем выделяют фракцию, содержащую целевой ДНК-продукт - генотерапевтический ДНК-вектор VTvaf17-SOD1, или генотерапевтический ДНК-вектор VTvaf17-SOD2, или генотерапевтический ДНК-вектор VTvaf17-SOD3, или генотерапевтический ДНК-вектор VTvaf17-CAT, многостадийно фильтруют и очищают хроматографическими методами. При этом специалистам в данной области техники понятно, что условия культивирования штаммов, состав питательных сред (за исключением содержания антибиотиков), используемое оборудование, методы очистки ДНК могут варьировать в рамках стандартных операционных процедур в зависимости от отдельно взятой производственной линии, но известные подходы к масштабированию, промышленному получению и очистке ДНК-векторов с использованием штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT, или подпадают под объем настоящего изобретения.
Описанное раскрытие изобретения подтверждается примерами реализации настоящего изобретения.
Изобретение поясняется следующими примерами.
Пример 1.
Получение генотерапевтического ДНК-вектора VTvaf17-SOD1, несущего целевой ген, а именно, гена SOD1.
Генотерапевтический ДНК-вектор VTvaf17-SOD1 конструировали клонированием кодирующей части гена SOD1 размером 466 п.н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции BamHI и EcoRI. Кодирующую часть гена SOD1 размером 466 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
SOD1_F AGGATCCACCATGGCGACGAAGGCCGTG
SOD1_R TATGAATTCATTGGGCGATCCCAATTACAC
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США).
Генотерапевтический ДНК-вектор VTvaf17 конструировали объединением шести фрагментов ДНК, полученных из разных источников:
(а) ориджин репликации получали путем ПЦР-амплификации участка коммерческой плазмиды pBR322 с внесением точечной мутации;
(б) промоторный регион EF1а получали путем ПЦР-амплификации участка геномной ДНК человека;
(в) терминатор транскрипции hGH-TA получали путем ПЦР-амплификации участка геномной ДНК человека;
(г) регуляторный участок транспозона Tn10 РНК-out получали путем синтеза из олигонуклеотидов;
(д) ген устойчивости к канамицину получали путем ПЦР-амплификации участка коммерческой плазмиды pET-28 человека;
(е) полилинкер получали отжигом двух синтетических олигонуклеотидов.
ПЦР-амплификацию проводили с использованием коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США) в соответствии с инструкцией производителя. Фрагменты имеют перекрывающиеся области для возможности их объединения с последующей ПЦР-амплификацией. Объединяли фрагменты (а) и (б) с использованием олигонуклеотидов Ori-F и EF1-R, а также фрагменты (в), (г) и (д) с использованием олигонуклеотидов hGH-F и Kan-R. Далее, полученные участки объединяли путем рестрикции с последующим лигированием по сайтам BamHI и NcoI. В результате получали плазмиду, пока еще не содержащую полилинкер. Для его введения проводили расщепление плазмиды по сайтам BamHI и EcoRI, и лигирование с фрагментом (е). Таким образом, получали вектор размером 3165 п.н., несущий ген устойчивости к канамицину, который фланкирован сайтами рестрикции SpeI. Далее этот участок выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор VTvaf17 размером 3165 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков.
Расщепление продукта амплификации кодирующей части гена SOD1 и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции BamHI и EcoRI (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-SOD1 размером 3607 п.н. с нуклеотидной последовательностью SEQ ID №1 и общей структурой изображенной на фиг.1A.
Пример 2.
Получение генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего целевой ген, а именно, гена SOD2.
Генотерапевтический ДНК-вектор VTvaf17-SOD2 конструировали клонированием кодирующей части гена SOD2 размером 671 п.н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции SalI и EcoRI. Кодирующую часть гена SOD2 размером 671 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
SOD2_F ATCGTCGACCACCATGTTGAGCCGGGCAGTGTG
SOD2_R TATGAATTCACTTTTTGCAAGCCATGTATCT
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции SalI и EcoRI (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-SOD2 размером 3824 п.н. с нуклеотидной последовательностью SEQ ID №2 и общей структурой изображенной на фиг.1B.
При этом генотерапевтический ДНК-вектор VTvaf17 конструировали по Примеру 1.
Пример 3
Получение ДНК-вектора VTvaf17-SOD3, несущего целевой ген, а именно, гена SOD3 человека.
Генотерапевтический ДНК-вектор VTvaf17-SOD3 конструировали клонированием кодирующей части гена SOD3 размером 725 п.н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции SalI и EcoRI. Кодирующую часть гена SOD3 размером 725 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
SOD3_F ATCGTCGACCACCATGCTGGCGCTACTGTGTTC
SOD3_R TATGAATTCAGGCGGCCTTGCACTCGCT
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции SalI и EcoRI (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-SOD3 размером 3878 п.н. с нуклеотидной последовательностью SEQ ID №3 и общей структурой изображенной на фиг.1C.
При этом генотерапевтический ДНК-вектор VTvaf17 конструировали по Примеру 1.
Пример 4.
Получение генотерапевтического ДНК-вектора VTvaf17-CAT, несущего целевой ген, а именно, гена CAT.
Генотерапевтический ДНК-вектор VTvaf17-CAT конструировали клонированием кодирующей части гена CAT размером 1590 п.н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции BamHII и HindIII. Кодирующую часть гена CAT размером 1590 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
CAT_F ATAGATCTGCCACCATGGCTGACAGCCGGGATC
CAT_R TATAAGCTTCACAGATTTGCCTTCTCCCT
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции BamHII и HindIII (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-CAT размером 4743 п.н. с нуклеотидной последовательностью SEQ ID №4 и общей структурой изображенной на фиг.1D.
При этом генотерапевтический ДНК-вектор VTvaf17 конструировали по Примеру 1.
Пример 5.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-SOD1, несущего целевой ген, а именно, ген SOD1, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена SOD1, в первичной культуре фибробластов кожи человека HDFa (ATCC PCS-201-01) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-SOD1, несущим ген SOD1 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Для оценки изменения накопления мРНК целевого гена SOD1, использовалась первичная культура фибробластов кожи человека HDFa. Клеточную культуру HDFa выращивали в стандартных условиях (37 °С, 5% СО2) с использованием питательной среды Fibroblast Growth Kit-Serum-Free (ATCC® PCS-201-040). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-SOD1, экспрессирующим ген SOD1 человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) согласно рекомендациям производителя. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора ДНК-вектора VTvaf17-SOD1 (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивали легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора Lipofectamin 3000. Аккуратно перемешивали легким встряхиванием. Добавляли содержимое пробирки №1 к содержимому пробирки №2, инкубировали 5 мин при комнатной температуре. Полученный раствор по каплям добавляли к клеткам в объеме 40 мкл.
В качестве контроля использовали клетки HDFa, трансфицированные генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена (кДНК гена SOD1 до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена, на фигурах не показано). Подготовку контрольного вектора VTvaf17 для трансфекции проводили как описано выше.
Суммарную РНК из клеток HDFa выделяли с использованием Trizol Reagent (Invitrogen, США) согласно рекомендациям производителя. В лунку с клетками добавляли 1 мл Trizol Reagent и гомогенизировали с последующим прогреванием в течении 5 мин при 65 °С. Далее образец центрифугировали при 14000g в течении 10 мин и снова прогревали в течении 10 мин при 65 °С. Далее добавляли 200 мкл хлороформа, плавно перемешивали и центрифугировали при 14000g в течении 10 мин. Затем отбирали водную фазу, добавляли к ней 1/10 объема 3М ацетата натрия, рН 5.2 и равный объем изопропилового спирта. Инкубировали образец при -20 °С в течении 10 мин с последующим центрифугированием при 14000g в течении 10 мин. Осадок промывали 1 мл 70% этилового спирта, высушивали на воздухе и растворяли в 10 мкл воды, свободной от РНКаз. Определение уровня экспрессии мРНК гена SOD1 после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени. Для получения и амплификации кДНК, специфичной для гена SOD1 человека, использовали олигонуклеотиды SOD1_SF и SOD1_SR:
SOD1_SF GACTGAAGGCCTGCATGGAT
SOD1_SR ATCGGCCACACCATCTTTGT
Длина продукта амплификации - 175 п.н.
Реакцию обратной транскрипции и ПЦР-амплификацию проводили с помощью набора реагентов SYBR GreenQuantitect RT-PCR Kit (Qiagen, США) для ПЦР в режиме реального времени. Реакцию проводили в объеме 20 мкл, содержащих: 25 мкл QuantiTect SYBR Green RT-PCR MasterMix, 2,5 мM хлорида магния, по 0,5 мкМ каждого праймера, 5 мкл РНК. Реакцию осуществляли на амплификаторе CFX96 (Bio-Rad, США) при следующих условиях: 1 цикл обратной транскрипции при 42°С - 30 минут, денатурация 98°С - 15 мин, затем 40 циклов, включающих денатурацию 94оС - 15 сек, отжиг праймеров 60°C - 30 сек и элонгацию 72°С - 30 сек. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов SOD1 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество динамику накопления ампликонов кДНК генов SOD1 и B2M оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 2.
Из фигуры 2 следует, что в результате трансфекции первичной культуры клеток фибробластов человека HDFa генотерапевтическим ДНК-вектором VTvaf17-SOD1, уровень специфической мРНК гена SOD1 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SOD1 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-SOD1 для повышения уровня экспрессии гена SOD1 в эукариотических клетках.
Пример 6.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего целевой ген, а именно, ген SOD2, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена SOD2, в первичной культуре кератиноцитов эпидермиса человека HEKa (ATCC PCS-200-011) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-SOD2, несущим ген SOD2 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Первичную культуру клеток кератиноцитов эпидермиса человека HEKa выращивали в среде Keratinocyte Growth Kit (ATCC® PCS-200-040™) в стандартных условиях (37°С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-SOD2, экспрессирующим ген SOD2 человека, проводили как описано в примере 5. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток HEKa, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген (кДНК гена SOD2 до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена, на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 5, за исключением олигонуклеотидов с отличающимися от примера 5 последовательностями. Для амплификации кДНК, специфичной для гена SOD2 человека, использовали олигонуклеотиды SOD2_SF и SOD2_SR:
SOD2_SF GTGGAGAACCCAAAGGGGAG
SOD2_SR GCCTGTTGTTCCTTGCAGTG
Длина продукта амплификации - 197 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов SOD2 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов - кДНК генов SOD2 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 3.
Из фигуры 3 следует, что в результате трансфекции культуры клеток HEKa генотерапевтическим ДНК-вектором VTvaf17-SOD2, уровень специфической мРНК гена SOD2 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SOD2 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-SOD2 для повышения уровня экспрессии гена SOD2 в эукариотических клетках.
Пример 7.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего целевой ген, а именно, ген SOD3, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена SOD3 в культуре клеток скелетных миобластов человека Human Skeletal Myoblasts (HSKM) (GIBCO® Кат. A12555) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-SOD3, несущим ген SOD3 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Культуру клеток скелетных миобластов человека линии HSKM выращивали в среде Dulbecco's Modified Eagle's Medium (DMEM) (ATCC® 30-2002™) с добавлением 2 % лошадиной сыворотки (Gibco Кат. 16050130) в стандартных условиях (37°С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-SOD3, экспрессирующим ген SOD3 человека, проводили как описано в примере 5. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток HSKM, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген (кДНК гена SOD3 до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена, на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 5, за исключением олигонуклеотидов с отличающимися от примера 5 последовательностями. Для амплификации кДНК, специфичной для гена SOD3 человека, использовали олигонуклеотиды SOD3_SF и SOD3_SR:
SOD3_SF GATCCGAGACATGTACGCCA
SOD3_SR GAAGAAGGCGTCGAGCTTGG
Длина продукта амплификации - 190 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов SOD3 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов - кДНК генов SOD3 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 4.
Из фигуры 4 следует, что в результате трансфекции культуры клеток скелетных миобластов человека линии HSKM генотерапевтическим ДНК-вектором VTvaf17-SOD3 уровень специфической мРНК гена SOD3 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SOD3 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-SOD3 для повышения уровня экспрессии гена SOD3 в эукариотических клетках.
Пример 8.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-CAT, несущего целевой ген, а именно, ген CAT, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена CAT, в первичной культуре клеток эндотелия человека линии Primary Umbilical Vein Endothelial Cells; Normal, Human (HUVEC) (ATCC® PCS-100-010™) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-CAT, несущим ген CAT человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Культуру клеток эндотелия человека линии HUVEC выращивали в среде Vascular Cell Basal Medium (ATCC PCS-100-030) с добавлением Endothelial Cell Growth Kit-BBE (ATCC® PCS-100-040™) в стандартных условиях (37°С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-CAT, экспрессирующим ген CAT человека, проводили как описано в примере 5. В качестве контроля использовали культуру клеток эндотелия человека линии HUVEC, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген (кДНК гена CAT до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена, на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 5, за исключением олигонуклеотидов с отличающимися от примера 5 последовательностями. Для амплификации кДНК, специфичной для гена CAT человека, использовали олигонуклеотиды CAT_SF и CAT_SR:
CAT_SF ACCAAGGTTTGGCCTCACAA
CAT_SR CGGTGAGTGTCAGGATAGGC
Длина продукта амплификации - 191 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов CAT и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов - кДНК генов CAT и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 5.
Из фигуры 5 следует, что в результате трансфекции культуры клеток эндотелия человека линии HUVEC генотерапевтическим ДНК-вектором VTvaf17-CAT, уровень специфической мРНК гена CAT человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген CAT на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором VTvaf17-CAT для повышения уровня экспрессии гена CAT в эукариотических клетках.
Пример 9.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-SOD1, несущего ген SOD1, для повышения экспрессии белка SOD1 в клетках млекопитающих.
Оценивали изменение количества белка SOD1 в лизате фибробластов кожи человека HDFa (ATCC PCS-201-01) после трансфекции этих клеток ДНК-вектором VTvaf17-SOD1, несущим ген SOD1 человека.
Клеточную культуру фибробластов кожи человека выращивали как это описано в примере 5.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена SOD1 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-SOD1, несущий ген SOD1 человека (С). Приготовление комплекса ДНК/дендример проводили по методике производителя (QIAGEN, SuperFect Transfection Reagent Handbook, 2002) с некоторыми изменениями. Для трансфекции клеток в одной лунке 24-луночного планшета к 1 мкг ДНК-вектора, растворенного в буфере ТЕ, добавляли культуральную среду до конечного объема 60 мкл, затем добавляли 5 мкл SuperFect Transfection Reagent и осторожно перемешивали пятикратным пипетированием. Комплекс инкубировали при комнатной температуре в течении 10-15 мин. Далее отбирали из лунок культуральную среду, лунки промывали 1 мл буфера PBS. К образовавшемуся комплексу добавляли 350 мкл среды, содержащей 10 мкг/мл гентамицина, осторожно перемешивали и добавляли к клеткам. Клетки инкубировали с комплексом 2-3 часа при 37°С в атмосфере, содержащей 5% СО2.
Затем осторожно удаляли среду, промывали клеточный монослой 1 мл буфера PBS. Затем добавляли среду, содержащую 10 мкг/мл гентамицина, и инкубировали 24-48 часов при 37°С в атмосфере 5% СО2.
После трансфекции клетки промывали три раза PBS, далее к клеткам добавляли 1 мл PBS и подвергали трехкратному замораживанию/оттаиванию. Далее суспензию центрифугировали при 15 000 об/мин в течение 15 мин, отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка SOD1 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human SOD1 / Cu-Zn SOD ELISA Kit (Sandwich ELISA) (LifeSpan BioSciences Кат. LS-F5770, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка SOD1, входящим в состав набора. Чувствительность метода составляла не менее 26,7 пг/мл, диапазон измерения - от 62.50 пг/мл до 4000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 6.
Из фигуры 6 следует, что трансфекция клеток фибробластов кожи человека HDFa генотерапевтическим ДНК-вектором VTvaf17-SOD1 приводит к увеличению количества белка SOD1 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SOD1 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-SOD1 для повышения уровня экспрессии SOD1 в эукариотических клетках.
Пример 10.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего ген SOD2, для повышения экспрессии белка SOD2 в клетках млекопитающих.
Оценивали изменение количества белка SOD2 в лизате клеток первичной культуры кератиноцитов эпидермиса человека HEKa (ATCC PCS-200-01) после трансфекции этих клеток ДНК-вектором VTvaf17-SOD2, несущим ген SOD2 человека. Клетки выращивали как описано в примере 6.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена SOD2 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-SOD2, несущий ген SOD2 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток HEKa проводили как описано в примере 9.
После трансфекции клетки промывали три раза PBS, далее к клеткам добавляли 1 мл PBS и подвергали трехкратному замораживанию/оттаиванию. Далее суспензию центрифугировали при 15 000 об/мин в течение 15 мин, отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка SOD2 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human Superoxide dismutase [Mn], mitochondrial ELISA Kit (Signalway Antibody LLC Кат. EK4403, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка SOD2, входящим в состав набора. Чувствительность метода составляла не менее 12 пг/мл, диапазон измерения - от 31.2 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 7.
Из фигуры 7 следует, что трансфекция первичной культуры клеток эндометрия матки человека генотерапевтическим ДНК-вектором VTvaf17-SOD2 приводит к увеличению количества белка SOD2 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SOD2 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-SOD2 для повышения уровня экспрессии SOD2 в эукариотических клетках.
Пример 11.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего ген SOD3, для повышения экспрессии белка SOD3 в клетках млекопитающих.
Оценивали изменение количества белка SOD3 в кондиционированной среде с клеток скелетных миобластов человека Human Skeletal Myoblasts (HSKM) (GIBCO® Кат. A12555) после трансфекции этих клеток ДНК-вектором VTvaf17-SOD3, несущим ген SOD3 человека. Клетки культивировали как описано в примере 7.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена SOD3 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-SOD3, несущий ген SOD3 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток HSKM проводили как описано в примере 9.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка SOD3 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human SOD3 ELISA Kit (Sandwich ELISA) (LifeSpan BioSciences Кат. LS-F5969-1, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка SOD3, входящим в состав набора. Чувствительность метода составляла не менее 125 пг/мл, диапазон измерения - от 125 пг/мл до 8000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 8.
Из фигуры 8 следует, что трансфекция культуры клеток скелетных миобластов человека линии HSKM генотерапевтическим ДНК-вектором VTvaf17-SOD3 приводит к увеличению количества белка SOD3 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SOD3 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-SOD3 для повышения уровня экспрессии SOD3 в эукариотических клетках.
Пример 12.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-CAT, несущего ген CAT, для повышения экспрессии белка CAT в клетках млекопитающих.
Оценивали изменение количества белка CAT в лизате культуры клеток первичной культуры эндотелия человека линии Primary Umbilical Vein Endothelial Cells; Normal, Human (HUVEC) (ATCC® PCS-100-010™) после трансфекции этих клеток ДНК-вектором VTvaf17-CAT, несущим ген CAT человека. Клетки культивировали как описано в примере 8.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена CAT (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-CAT, несущий ген CAT человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию клеток HUVEC проводили как описано в примере 9.
После трансфекции клетки промывали три раза PBS, далее к клеткам добавляли 1 мл PBS и подвергали трехкратному замораживанию/оттаиванию. Далее суспензию центрифугировали при 15000 об/мин в течение 15 мин, отбирали супернатант и использовали его для количественного определения целевого белка. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка CAT проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human CAT / Catalase ELISA Kit (Sandwich ELISA) (LifeSpan BioSciences Кат. LS-F23006-1, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка CAT, входящим в состав набора. Чувствительность метода составляла не менее 9,375 пг/мл, диапазон измерения - от 15,625 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 9.
Из фигуры 9 следует, что трансфекция клеток первичной культуры эндотелия человека линии HUVEC генотерапевтическим ДНК-вектором VTvaf17-CAT приводит к увеличению количества белка CAT по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген CAT на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-CAT для повышения уровня экспрессии CAT в эукариотических клетках.
Пример 13.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-CAT, несущего ген CAT, для повышения экспрессии белка CAT в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-CAT, несущего целевой ген, а именно, ген CAT и реализуемости способа его применения оценивали изменения количества белка CAT в коже человека при введении в кожу человека генотерапевтического ДНК-вектора VTvaf17-CAT, несущего ген человека CAT.
С целью анализа изменения количества белка CAT трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор VTvaf17-CAT, несущий ген CAT и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор VTvaf17, не содержащий кДНК гена CAT.
Пациент 1, мужчина 58 лет, (П1); пациент 2, женщина 60 лет, П2); пациент 3, мужчина, 51 года, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор VTvaf17-CAT, который содержит кДНК гена CAT и генотерапевтический ДНК-вектор VTvaf17, используемый в качестве плацебо, который не содержит кДНК гена CAT, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор VTvaf17 (плацебо) и генотерапевтический ДНК вектор VTvaf17-CAT, несущий ген CAT вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора VTvaf17 (плацебо) и генотерапевтического ДНК вектора VTvaf17-CAT, несущего ген CAT - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора VTvaf17-CAT, несущего ген CAT (I), генотерапевтического ДНК-вектора VTvaf17 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа (ELISA). Количественное определение белка CAT проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human CAT / Catalase ELISA Kit (Sandwich ELISA) (LifeSpan BioSciences Кат. LS-F23006-1, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка CAT, входящим в состав набора. Чувствительность метода составляла не менее 9,375 пг/мл, диапазон измерения - от 15,625 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США). Графики, полученные в результате анализа, представлены на фиг. 10.
Из фигуры 10 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора VTvaf17- CAT, несущего целевой ген CAT человека, произошло увеличение количества белка CAT в сравнении с количеством белка CAT в области введения генотерапевтического ДНК-вектора VTvaf17 (плацебо), не содержащего ген CAT человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-CAT и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 14.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего ген SOD3, для повышения экспрессии белка SOD3 в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего целевой ген SOD3 и реализуемости способа его применения оценивали изменение количества белка SOD3 в мышечной ткани человека при введении генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего целевой ген, а именно, ген SOD3 человека.
С целью анализа изменения количества белка SOD3 трем пациентам вводили в ткань икроножной мышцы генотерапевтический ДНК-вектор VTvaf17- SOD3, несущий ген SOD3 с транспортной молекулой, и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор VTvaf17, не содержащий кДНК гена SOD3 с транспортной молекулой.
Пациент 1, женщина 50 лет, (П1); пациент 2, мужчина 43 года (П2); пациент 3, мужчина 62 года, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция), пробоподготовку проводили в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор VTvaf17 (плацебо) и генотерапевтический ДНК вектор VTvaf17-SOD3, несущий ген SOD3 вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину около 10 мм. Объем вводимого раствора генотерапевтического ДНК-вектора VTvaf17 (плацебо) и генотерапевтического ДНК вектора VTvaf17- SOD3, несущего ген SOD3 - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагали медиально на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков мышечной ткани пациентов в зоне введения генотерапевтического ДНК вектора VTvaf17-SOD3, несущего ген SOD3 (I), генотерапевтического ДНК-вектора VTvaf17 (плацебо) (II), а также из интактного участка икроножной мышцы (III), используя устройство для взятия биопсии MAGNUM (BARD, США). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 20 куб. мм, масса - около 22 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка SOD3 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human SOD3 ELISA Kit (Sandwich ELISA) - LS-F5969 (LifeSpan BioSciences Кат. LS-F5969, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка SOD3, входящим в состав набора. Чувствительность метода составляла не менее 125 пг/мл, диапазон измерения - от 125 пг/мл до 8000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 11.
Из фигуры 11 следует, что в икроножной мышце всех трех пациентов в области введения генотерапевтического ДНК вектора VTvaf17-SOD3, несущего целевой ген, а именно, ген SOD3 человека, произошло увеличение количества белка SOD3 в сравнении с количеством белка SOD3 в области введения генотерапевтического ДНК-вектора VTvaf17 (плацебо), не содержащего ген SOD3 человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-SOD3 и подтверждает реализуемость способа его применения, в частности при внутримышечном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 15.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего ген SOD2, для повышения экспрессии белка SOD2 в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего целевой ген, а именно, ген SOD2 и реализуемости способа его применения оценивали изменения количества белка SOD2 в коже человека при введении в кожу человека генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего ген человека SOD2.
С целью анализа изменения количества белка SOD2 трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор VTvaf17-SOD2, несущий ген SOD2 и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор VTvaf17, не содержащий кДНК гена SOD2.
Пациент 1, женщина 45 лет (П1); пациент 2, мужчина 50 лет (П2); пациент 3, мужчина, 49 лет (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор VTvaf17-SOD2, который содержит кДНК гена SOD2 и генотерапевтический ДНК-вектор VTvaf17, используемый в качестве плацебо, который не содержит кДНК гена SOD2, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор VTvaf17 (плацебо) и генотерапевтический ДНК вектор VTvaf17-SOD2, несущий ген SOD2 вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора VTvaf17 (плацебо) и генотерапевтического ДНК вектора VTvaf17-SOD2, несущего ген SOD2 - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора VTvaf17-SOD2, несущего ген SOD2 (I), генотерапевтического ДНК-вектора VTvaf17 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа (ELISA), используя набор используя набор Human Superoxide dismutase [Mn], mitochondrial ELISA Kit (Signalway Antibody LLC Кат. EK4403, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка SOD2, входящим в состав набора. Чувствительность метода составляла не менее 12 пг/мл, диапазон измерения - от 31.2 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 12.
Из фигуры 12 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора VTvaf17- SOD2, несущего целевой ген SOD2 человека, произошло увеличение количества белка SOD2 в сравнении с количеством белка SOD2 в области введения генотерапевтического ДНК-вектора VTvaf17 (плацебо), не содержащего ген SOD2 человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-SOD2 и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 16.
Подтверждение эффективности генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего ген SOD2 и реализуемости способа его применения для повышения уровня экспрессии белка SOD2 в тканях человека путем введения аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего ген SOD2 и реализуемости способа его применения оценивали изменения количества белка SOD2 в коже человека при введении в кожу пациента культуры аутологичных фибробластов этого пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2.
Пациенту вводили в кожу предплечья соответствующую культуру аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2, несущим ген SOD2, и параллельно вводили плацебо, представляющее собой культуру аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген SOD2.
Первичную культуру фибробластов человека выделяли из биоптатов кожи пациента. Используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL, Италия) брали образец биоптата кожи из зоны, защищенной от действия ультрафиолета, а именно за ушной раковиной или с боковой внутренней поверхности в зоне локтевого сустава, размером около 10 кв. мм, массой - около 11 мг. Кожу пациента предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Культивирование первичной культуры клеток осуществляли при 37°С в атмосфере, содержащей 5 % СО2 в среде DMEM с 10 % фетальной телячьей сывороткой и ампициллином 100 Ед/мл. Пересев культуры и смену культуральной среды осуществляли каждые 2 дня. Общая продолжительность роста культуры не превышала 25-30 дней. Из культуры клеток отбирали аликвоту, содержащую 5×104 клеток. Культуру фибробластов пациента трансфицировали генотерапевтическим ДНК-вектором VTvaf17-SOD2, несущим ген SOD2 или плацебо - вектор VTvaf17, не несущий целевой ген SOD2.
Трансфекцию осуществляли с помощью катионного полимера полиэтиленимина JETPEI (Polyplus Transfection, Франция), согласно инструкции фирмы-производителя. Клетки культивировали 72 часа и затем вводили пациенту. Введение пациенту культуры аутологичных фибробластов этого пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2, и плацебо, представляющее собой культуру аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, осуществляли туннельным методом в область предплечья иглой 30G длиной 13 мм на глубину около 3 мм. Концентрация модифицированных аутологичных фибробластов в вводимой суспензии составляла примерно 5 млн клеток на 1 мл суспензии, количество вводимых клеток не превышало 15 млн. Очаги введения культуры аутологичных фибробластов располагались на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 4-е сутки после введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2, несущим целевой ген, а именно, ген SOD2 и плацебо. Взятие биопсии осуществляли из участков кожи пациента в зоне введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2, несущим целевой ген, а именно, ген SOD2 (C), культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген SOD2 (плацебо) (B), а также из участков интактной кожи (A), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу в зоне отбора биоптата пациентов предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка как это описано в примере 10.
Графики, полученные в результате анализа, представлены на фиг. 13.
Из фигуры 13 следует, что в коже пациента в области введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2, несущим ген SOD2, произошло увеличение количества белка SOD2 в сравнении с количеством белка SOD2 в области введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген SOD2 (плацебо), что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-SOD2 и подтверждает реализуемость способа его применения для повышения уровня экспрессии SOD2 в тканях человека, в частности при введении аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-SOD2.
Пример 17.
Подтверждение эффективности и реализуемости способа сочетанного применения генотерапевтического ДНК-вектора VTvaf17-SOD1, несущего ген SOD1, генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего ген SOD2, генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего ген SOD3, генотерапевтического ДНК-вектора VTvaf17-CAT, несущего ген CAT, для повышения уровня экспрессии белков SOD1, SOD2, SOD3 и CAT в тканях млекопитающих.
Оценивали изменение количества белков SOD1, SOD2, SOD3 и CAT в зоне кожи крысы линии Вистар при введении в эту зону смеси генотерапевтических векторов.
В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Эквимолярную смесь генотерапевтических ДНК-векторов растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя. Объем вводимого раствора составлял 0,3 мл с суммарным количеством ДНК 100 мкг. Введение раствора осуществляли тоннельным методом иглой 33G на глубину 1 мм в зону кожи крысы.
Биопсийные образцы отбирали на 2 сутки после введения генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков на коже животных в зоне введения смеси четырех генотерапевтических ДНК-векторов, несущих гены SOD1, SOD2, SOD3, CAT (зона I), генотерапевтического ДНК-вектора VTvaf17 (плацебо) (зона II), а также из участков аналогичного участка кожи, не подвергавшегося никаким манипуляциям ( зона III), используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL). Зону отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер каждого биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Каждый образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевых белков как это описано в примере 9 (количественное определение белка SOD1), примере 10 (количественное определение белка SOD2), примере 11 (количественное определение белка SOD3), примере 12 (количественное определение белка CAT). Графики, полученные в результате анализа, представлены на фиг. 14.
Из фигуры 14 следует, что в зоне кожи крыс (зона I), в которую вводилась смесь генотерапевтических векторов: генотерапевтического ДНК-вектора VTvaf17-SOD1, несущего целевой ген SOD1, генотерапевтического ДНК-вектора VTvaf17-SOD2, несущего целевой ген SOD2, генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего целевой ген SOD3, генотерапевтического ДНК-вектора VTvaf17-CAT, несущего целевой ген CAT, произошло увеличение количества белков SOD1, SOD2, SOD3 и CAT по сравнению с зоной II (зона плацебо) и зоной III (интактная зона). Полученные результаты свидетельствуют об эффективности сочетанного применения генотерапевтических ДНК-векторов и подтверждает реализуемость способа применения для повышения уровня экспрессии целевых белков в тканях млекопитающих.
Пример 18.
Подтверждение эффективности генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего ген SOD3 и реализуемости способа его применения для повышения уровня экспрессии белка SOD3 в клетках млекопитающих.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-SOD3, несущего ген SOD3 оценивали изменение накопления мРНК целевого гена SOD3 в клетках эпителия эндометрия коровы Bovine Endometrial Epithelial Cells (BEnEpC) (Cell Applications, Inc. Кат. B932-05) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17- SOD3, несущим ген SOD3 человека.
Клетки клетках эпителия эндометрия коровы Bovine Endometrial Epithelial Cells (BEnEpC) (Cell Applications, Inc. Кат. B932-05) выращивали в среде Bovine Endometrial Cell Growth Medium (Cell Applications, Inc. Кат. B911-500). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17- SOD3, несущим ген SOD3 человека и ДНК-вектором VTvaf17, не несущим ген SOD3 человека (контроль), выделение РНК, реакцию обратной транскрипции, ПЦР амплификации и анализ данных проводили как описано в примере 7. В качестве референтного гена использовали ген актина быка/коровы (АСТ), приведенного в базе данных GenBank под номером AH001130.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности генов SOD3 и ACT. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов - кДНК генов SOD3 и ACT, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1. (Bio-Rad, USA).
Графики, полученные в результате анализа представлены на фиг. 15.
Из фигуры 15 следует, что в результате трансфекции клеток эпителия эндометрия коровы BEnEpC генотерапевтическим ДНК-вектором VTvaf17-SOD3, уровень специфической мРНК гена SOD3 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SOD3 на уровне мРНК. Представленные результаты подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором VTvaf17-SOD3 для повышения уровня экспрессии гена SOD3 в клетках млекопитающих.
Пример 19.
Штамм Escherichia coli SCS110-AF/VTvaf17-SOD1 или штамм Escherichia coli SCS110-AF/VTvaf17-SOD2 или штамм Escherichia coli SCS110-AF/VTvaf17-SOD3 или штамм Escherichia coli SCS110-AF/VTvaf17-CAT, несущий генотерапевтический ДНК-вектор, и способ его получения.
Конструирование штамма для наработки в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: SOD1, SOD2, SOD3, CAT, а именно штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT, несущего генотерапевтический ДНК-вектор VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT соответственно для его наработки с возможностью селекции без использования антибиотиков заключается в получении электрокомпетентных клеток штамма Escherichia coli SCS110-AF с проведением электропорации этих клеток генотерапевтическим ДНК-вектором VTvaf17-SOD1, или ДНК-вектором VTvaf17-SOD2, или ДНК-вектором VTvaf17-SOD3, или ДНК-вектором VTvaf17-CAT, после чего клетки высеваются на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола. При этом штамм Escherichia coli SCS110-AF для наработки генотерапевтического ДНК-вектора VTvaf17 или генотерапевтических ДНК-векторов на его основе с возможностью положительной селекции без использования антибиотиков получали путем конструирования линейного фрагмента ДНК, содержащего регуляторный элемент RNA-in транспозона Tn10 для селекции без применения антибиотиков размером 64 п.н., ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде размером 1422 п.н., ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация размером 763 п.н. и две гомологичные последовательности, обеспечивающие процесс гомологичной рекомбинации в области гена recA с одновременной его инактивацией размером 329 п.н. и 233 п.н., после чего проводили трансформацию клеток Escherichia coli путем электропорации и отбирали клоны, выжившие на среде, содержащей 10 мкг/мл хлорамфеникола.
Полученные штаммы для наработки были внесены в коллекции Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и NCIMB Patent Deposit Service, UK с присвоением регистрационных номеров:
штамм Escherichia coli SCS110-AF/VTvaf17-SOD1 - регистрационный № ВКПМ: B-13321, дата депонирования: 12.11.2018, accession № NCIMB: 43251, дата депонирования: 08.11.2018.
штамм Escherichia coli SCS110-AF/VTvaf17-SOD2 - регистрационный № ВКПМ: B-13320, дата депонирования: 12.11.2018, accession № NCIMB: 43248, дата депонирования: 08.11.2018.
штамм Escherichia coli SCS110-AF/VTvaf17-SOD3 - регистрационный № ВКПМ: B-13343, дата депонирования: 22.11.2018, accession № NCIMB: 43288, дата депонирования: 22.11.2018.
штамм Escherichia coli SCS110-AF/VTvaf17-CAT - регистрационный № ВКПМ: B-13330, дата депонирования: 12.11.2018, accession № NCIMB: 43246, дата депонирования: 08.11.2018.
Пример 20.
Способ масштабирования получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: SOD1, SOD2, SOD3, CAT до промышленного масштаба.
Для подтверждения технологичности получения и возможности производства в промышленных масштабах генотерапевтического ДНК-вектора VTvaf17-SOD1 (SEQ ID №1), или VTvaf17-SOD2 (SEQ ID №2), или VTvaf17-SOD3 (SEQ ID №3), или VTvaf17-CAT (SEQ ID №4), или проводили масштабную ферментацию штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT, каждый из которых содержит генотерапевтический ДНК-вектор VTvaf17, несущий целевой ген, а именно SOD1, или SOD2, или SOD3, или CAT. Каждый штамм Escherichia coli SCS110-AF/VTvaf17-SOD1, или Escherichia coli SCS110-AF/VTvaf17-SOD2, или Escherichia coli SCS110-AF/VTvaf17-SOD3, или Escherichia coli SCS110-AF/VTvaf17-CAT получали на базе штамма Escherichia coli SCS110-AF (Cell and Gene Therapy LLC, Великобритания) по примеру 19 путем электропорации компетентных клеток этого штамма генотерапевтическим ДНК-вектором VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT, несущим целевой ген, а именно, SOD1, или SOD2, или SOD3, или CAT с последующим высевом трансформированных клеток на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы и отбором отдельных клонов.
Ферментацию полученного штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, несущего генотерапевтический ДНК-вектор VTvaf17-SOD1 проводили в ферментере объемом 10 л с последующим выделением генотерапевтического ДНК-вектора VTvaf17-SOD1.
Для ферментации штамма Escherichia coli SCS110-AF/VTvaf17-SOD1 готовили среду, содержащую на 10 л: 100 г триптон, 50 г дрожжевой экстракт (Becton Dickinson, США), доводили водой до 8800 мл и автоклавировали при 121 0С 20 мин, затем добавляли 1200 мл 50% (вес/объем) сахарозы. Далее засевали в колбу затравочную культуру штамма Escherichia coli SCS110-AF/VTvaf17-SOD1 в объеме 100 мл. Инкубировали в шейкере-инкубаторе 16 ч при 30°С. Переносили затравочную культуру в ферментер Techfors S (Infors HT, Швейцария), растили до достижения стационарной фазы. Контроль осуществляли измерением оптической плотности культуры при длине волны 600 нм. Клетки осаждали центрифугированием 30 мин при 5000-10000 g. Супернатант удаляли, клеточную массу ресуспендировали в 10% по объему фосфатно-солевого буфера. Повторно центрифугировали 30 мин при 5000-10000g. Супернатант удаляли, к клеточной массе добавляли раствор 20 мМ TrisCl, 1 мМ ЭДТА, 200 г/л сахароза, рН 8,0 в объеме 1000 мл, тщательно перемешивали до образования гомогенной суспензии. Добавляли раствор яичного лизоцима до конечной концентрации 100мкг/мл. Инкубировали 20 мин на льду при бережном перемешивании. Далее, добавляли 2500 мл раствора 0,2 М NaOH, 10 г/л додецилсульфат натрия, инкубировали 10 мин на льду при бережном перемешивании, затем добавляли 3500 мл раствора 3M ацетат натрия, 2М уксусная кислота, рН 5-5,5, инкубировали 10 мин на льду при бережном перемешивании. Полученный образец центрифугировали 20-30 мин при 15000 g или более. Раствор аккуратно декантировали, от остатков осадка избавлялись фильтрацией через грубый фильтр (фильтровальную бумагу). Добавляли РНКазу А (Sigma, США) до конечной концентрации 20 нг/мл, инкубировали ночь 16 ч при комнатной температуре. Раствор центрифугировали 20-30 мин при 15000g, затем фильтровали через мембранный фильтр с порами 0,45 мкм (Millipore, США). Далее проводили ультрафильтрацию через мембрану размером отсечения 100кДа (Millipore, США) и разбавляли до начального объема буфером 25 мМ TrisCl, pH 7.0. Операцию повторяли три - четыре раза. Наносили раствор, на колонку с 250 мл сорбента DEAE sepharose HP (GE, США), уравновешенную раствором 25 мМ TrisCl, pH 7.0. После нанесения образца колонку промывали тремя объемами этого же раствора, а затем проводили элюцию генотерапевтического ДНК-вектора VTvaf17-SOD1 линейным градиентом от раствора 25 мМ TrisCl, pH 7.0 до раствора 25 мМ TrisCl, pH 7.0, 1М NaCl в объеме пяти объемов колонки. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм. Хроматографические фракции, содержащие генотерапевтический ДНК-вектор VTvaf17-SOD1, объединяли и проводили гель-фильтрацию на сорбенте Superdex 200 (GE, США). Колонку уравновешивали фосфатно-солевым буфером. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм, фракции анализировали электрофорезом в агарозном геле. Фракции, содержащие генотерапевтический ДНК-вектор VTvaf17-SOD1 объединяли и хранили при -20°С. Для оценки воспроизводимости техпроцесса обозначенные технологические операции повторяли пятикратно. Все технологические операции для штаммов Escherichia coli SCS110-AF/VTvaf17-SOD2, или Escherichia coli SCS110-AF/VTvaf17-SOD3, или Escherichia coli SCS110-AF/VTvaf17-CAT проводили аналогичным образом.
Воспроизводимость техпроцесса и количественные характеристики выхода конечного продукта подтверждают технологичность получения и возможность производства в промышленных масштабах генотерапевтического ДНК-вектора VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT.
Таким образом, созданный генотерапевтический ДНК-вектор с целевым геном может быть использован для введения в клетки организма животных и человека, характеризующихся сниженной или недостаточной экспрессией белка, кодируемого данным геном, обеспечивая таким образом достижение терапевтического эффекта.
Задача, поставленная в данном изобретении, а именно: конструирование генотерапевтических ДНК-векторов для повышения уровня экспрессии генов SOD1, SOD2, SOD3, CAT, сочетающих в себе следующие свойства:
I) Эффективность повышения экспрессии целевых генов в эукариотических клетках за счет полученных генотерапевтических векторов с минимальным размером;
II) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов и генов антибиотикорезистентности;
III) Технологичность получения и возможность наработки в штаммах в промышленных масштабах;
IV) также задача по конструированию штаммов, несущих эти генотерапевтические ДНК-вектора для производства этих генотерапевтических ДНК-векторов решена, что подтверждается примерами:
для п. I - пример 1, 2, 3, 4, 5; 6; 7; 8; 9; 10; 11; 12; 13; 14; 15; 16; 17; 18
для п. II - пример 1, 2, 3, 4
для п. III и п. IV - пример 19, 20.
Промышленная применимость:
Все представленные выше примеры подтверждают промышленную применимость предлагаемого генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: ген SOD1, ген SOD2, ген SOD3, ген CAT для повышения уровня экспрессии этих целевых генов, штамма Escherichia coli SCS110-AF/VTvaf17-SOD1 или Escherichia coli SCS110-AF/VTvaf17-SOD2 или Escherichia coli SCS110-AF/VTvaf17-SOD3 или Escherichia coli SCS110-AF/VTvaf17-CAT, несущего генотерапевтический ДНК-вектор, способа получения генотерапевтического ДНК-вектора, способа производства в промышленных масштабах генотерапевтического ДНК-вектора.
Перечень сокращений
VTvaf17 - вектор генотерапевтический, не содержащий последовательностей вирусных геномов и маркеров антибиотико-резистентности (vector therapeutic virus-antibiotic-free)
ДНК - дезоксирибонуклеиновая кислота
кДНК - комплементарная дезоксирибонуклеиновая кислота
РНК - рибонуклеиновая кислота
мРНК - матричная рибонуклеиновая кислота
п.н. - пар нуклеотидов
ПЦР - полимеразная цепная реакция
мл - миллилитр, мкл - микролитр
куб. мм - кубический миллиметр
л - литр
мкг - микрограмм
мг - миллиграмм
г - грамм
мкМ - микромоль
мМ - миллимоль
мин - минута
сек - секунда
об/мин - обороты в минуту
нм - нанометр
см - сантиметр
мВт - милливатт
о.е. ф-относительная единица флуоресценции
РВС - фосфатно-солевой буфер.
Список литературы
1. Ahn J., Nowell S., McCann S.E., Yu J., Carter L., Lang N.P., Kadlubar F.F., Ratnasinghe L.D., Ambrosone C.B. Associations between catalase phenotype and genotype: modification by epidemiologic factors. Cancer Epidemiol Biomarkers Prev. 2006; 15 (6): 1217-1222.
2. Babusikova E, Jesenak M, Evinova A, Banovcin P, Dobrota D. Frequency of polymorphism -262 c/t in catalase gene and oxidative damage in Slovak children with bronchial asthma. Arch Bronconeumol. 2013 Dec;49(12):507-12.
3. Biswal MR, Han P, Zhu P, Wang Z, Li H, Ildefonso CJ, Lewin AS. Timing of Antioxidant Gene Therapy: Implications for Treating Dry AMD. Invest Ophthalmol Vis Sci. 2017 Feb 1;58(2):1237-1245.
4. Cells Transduced with Superoxide Dismutase on Imiquimod-Induced Psoriasis Like Skin Inflammation in Mice. Antioxid Redox Signal. 2016 Feb 10;24(5):233-48.
5. Cui R, Gao M, Qu S, Liu D. Overexpression of superoxide dismutase 3 gene blocks high-fat diet-induced obesity, fatty liver and insulin resistance. Gene Ther. 2014 Sep;21(9):840-8.
6. Davis AS, Zhao H, Sun GH, Sapolsky RM, Steinberg GK. Gene therapy using SOD1 protects striatal neurons from experimental stroke. Neurosci Lett. 2007 Jan 3;411(1):32-6. Epub 2006 Nov 15.
7. Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products, http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf.
8. Epperly MW, Wegner R, Kanai AJ, Kagan V, Greenberger EE, Nie S, Greenberger JS. Effects of MnSOD-plasmid liposome gene therapy on antioxidant levels in irradiated murine oral cavity orthotopic tumors. Radiat Res. 2007 Mar;167(3):289-97. PubMed PMID: 17316075.
9. Ergen H.A., Narter F., Timirci O., Isbir T. Effects of manganase superoxide dismutase Ala-9Val polymorphism on prostate cancer: a case-control study. Anticancer Res. 2007; 27 (2): 1227-1230.
10. Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal Products EMA/CAT/80183/2014.
11. Han L., Lee S.W., Yoon J.H., Park Y.G., Choi Y.J., Nam S.W., Lee J.Y., Wang Y.P., Park W.S. Association of SOD1 and SOD2 single nucleotide polymorphisms with susceptibility to gastric cancer in a Korean population. APMIS. 2012; 10: 463-466.
12. Hornstein BD, Roman D, Arévalo-Soliz LM, Engevik MA, Zechiedrich L. Effects of Circular DNA Length on Transfection Efficiency by Electroporation into HeLa Cells. Ceña V, ed. PLoS ONE. 2016;11(12).
13. Juul K., Tybjaerg-Hansen A., Marklund S., Heegaard N.H., Steffensen R., Sillesen H., Jensen G., Nordestgaard B.G., Genetically reduced antioxidative protection and increased ischemic heart disease risk: The Copenhagen City Heart Study. Circulation. 2004; 109: 59-65.
14. Kang D., Lee K.M., Park S.K., Berndt S.I., Peters U., Reding D., Chatterjee N., Welch R., Chanock S., Huang W.Y., Hayes R.B. Functional variant of manganese superoxide dismutase (SOD2 V16A) polymorphism is associated with prostate cancer risk in the prostate, lung, colorectal, and ovarian cancer study. Cancer Epidemiol Biomarkers Prev. 2007; 16: 1581-1586.
15. Kaur SJ, McKeown SR, Rashid S. Mutant SOD1 mediated pathogenesis of Amyotrophic Lateral Sclerosis. Gene. 2016 Feb 15;577(2):109-18.
16. Kim J, Mizokami A, Shin M, Izumi K, Konaka H, Kadono Y, Kitagawa Y, Keller ET, Zhang J, Namiki M. SOD3 acts as a tumor suppressor in PC-3 prostate cancer cells via hydrogen peroxide accumulation. Anticancer Res. 2014 Jun;34(6):2821-31.
17. Li D, Duan D. Mitochondria-targeted antiaging gene therapy with adeno-associated viral vectors. Methods Mol Biol. 2013; 1048:161-80.
18. Li L, Petrovsky N. Molecular mechanisms for enhanced DNA vaccine immunogenicity. Expert Rev Vaccines. 2016;15(3):313-29
19. Liu G., Zhou W., Wang L.I., Park S., Miller D.P., Xu L.L., Wain J.C., Lynch T.J., Su L., Christiani D.C. MPO and SOD2 polymorphisms, gender, and the risk of non-small cell lung carcinoma. Cancer Lett. 2004; 214: 69-79.
20. Liu JM, Auerbach AD, Anderson SM, Green SW, Young NS. A trial of recombinant human superoxide dismutase in patients with Fanconi anaemia. Br J Haematol. 1993 Oct;85(2):406-8.
21. Maikopova E.V., Alimova F.K., Podol'skaya A.A., Kravtsova O.A. Dnepropetrovsk - Dnepropetrovsk. 2011; 4: 6-9.
22. Maikopova E.V., Alimova F.K., Podol'skaya A.A., Kravtsova O.A. Dnepropetrovsk - Dnepropetrovsk. 2011; 4: 6-9.
23. Mairhofer J, Grabherr R. Rational vector design for efficient non-viral gene delivery: challenges facing the use of plasmid DNA. Mol Biotechnol. 2008.39(2):97-104.
24. Nozik-Grayck E., Suliman H.B., Piantadosi C.A. Extracellular superoxide dismutase. Int J Biochem Cell Biol. 2005; 37: 2466-2471.
25. Olson S.H., Carlson M.D., Ostrer H., Harlap S., Stone A., Winters M., Ambrosone C.B. Genetic variants in SOD2, MPO, and NQO1, and risk of ovarian cancer. Gynecol Oncol. 2004; 93: 615-620.
26. Qi X, Lewin AS, Sun L, Hauswirth WW, Guy J. SOD2 gene transfer protects against optic neuropathy induced by deficiency of complex I. Ann Neurol. 2004 Aug;56(2):182-91.
27. Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies.
28. Sah SK, Park KH, Yun C-O, Kang K-S, Kim T-Y. Effects of Human Mesenchymal Stem Cells Transduced with Superoxide Dismutase on Imiquimod-Induced Psoriasis-Like Skin Inflammation in Mice. Antioxidants & Redox Signaling. 2016;24(5):233-248.
29. Seiberg M. Age-induced hair greying - the multiple effects of oxidative stress. Int J Cosmet Sci. 2013 Dec;35(6):532-8.
30. Semenova N.A., Ryazantseva N.V., Novitskii V.V., Dmitrieva A.I., Chechina O.E., Bychkov V.A., Moiseenko I.P. Byulleten' sibirskoi meditsiny - Bulletin of Siberian medicine. 2009; 3: 64-69.
31. Sobkowiak A., Lianeri M., Wudarski M., Lacki J.K., Jagodzinski P.P. Manganese superoxide dismutase Ala-9Val mitochondrial targeting sequence polymorphism in systemic lupus erythematosus in Poland. Clin. Rheumatol. 2008; 27: 827-831.
32. Strokov I.A., Bursa T.R., Zotova E.V., Ametov A.S. Genetika i patogenez oslozhnenii - Genetics and pathogenesis of effects. 2003; 2: 3-5.
33. Van-Assche T, Huygelen V, Crabtree MJ, Antoniades C. Gene therapy targeting inflammation in atherosclerosis. Curr Pharm Des. 2011 Dec;17(37):4210-23. Review.
34. Wang L, Yi F, Fu L, Yang J, Wang S, Wang Z, Suzuki K, Sun L, Xu X, Yu Y, Qiao J, Belmonte JCI, Yang Z, Yuan Y, Qu J, Liu GH. CRISPR/Cas9-mediated targeted gene correction in amyotrophic lateral sclerosis patient iPSCs. Protein Cell. 2017 May;8(5):365-378.
35. Yang CC, Hsu SP, Chen KH, Chien CT. Effect of Adenoviral Catalase Gene Transfer on Renal Ischemia/Reperfusion Injury in Rats. Chin J Physiol. 2015 Dec 31;58(6):420-30.
36. Younus H. Therapeutic potentials of superoxide dismutase. International Journal of Health Sciences. 2018;12(3):88-93.
37. Молекулярная биология, 2011, том 45, № 1, с. 44-55.
38. Заявка JP2009501009A.
39. Патент RU2651757C2.
40. Патент CA2273173A1.
41. Заявка CN102080102A.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Селл энд Джин Терапи Лтд (CELL and GENE THERAPY Ltd), Общество с ограниченной ответственностью «Прорывные Инновационные Технологии».
<120> Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT для повышения уровня экспрессии этих целевых генов, способ его получения и применения, штамм Escherichia coli SCS110-AF/VTvaf17-SOD1 или Escherichia coli SCS110-AF/VTvaf17-SOD2 или Escherichia coli SCS110-AF/VTvaf17-SOD3 или Escherichia coli SCS110-AF/VTvaf17-CAT, несущий генотерапевтический ДНК-вектор, способ его получения, способ производства в промышленных масштабах генотерапевтического ДНК-вектора.
<160> 4
<170> BiSSAP 1.3.6
<210> 1
<211> 4743
<212> DNA
<213> Homo Sapiens
<400> 1
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaaa gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcagtagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacgcccctg gctgcagtac gtgattcttg atcccgagct tcgggttgga 360
agtgggtggg agagttcgag gccttgcgct taaggagccc cttcgcctcg tgcttgagtt 420
gaggcctggc ttgggcgctg gggccgccgc gtgcgaatct ggtggcacct tcgcgcctgt 480
ctcgctgctt tcgataagtc tctagccatt taaaattttt gatgacctgc tgcgacgctt 540
tttttctggc aagatagtct tgtaaatgcg ggccaagatc tgcacactgg tatttcggtt 600
tttggggccg cgggcggcga cggggcccgt gcgtcccagc gcacatgttc ggcgaggcgg 660
ggcctgcgag cgcggccacc gagaatcgga cgggggtagt ctcaagctgg ccggcctgct 720
ctggtgcctg gcctcgcgcc gccgtgtatc gccccgccct gggcggcaag gctggcccgg 780
tcggcaccag ttgcgtgagc ggaaagatgg ccgcttcccg gccctgctgc agggagctca 840
aaatggagga cgcggcgctc gggagagcgg gcgggtgagt cacccacaca aaggaaaagg 900
gcctttccgt cctcagccgt cgcttcatgt gactccacgg agtaccgggc gccgtccagg 960
cacctcgatt agttctcgag cttttggagt acgtcgtctt taggttgggg ggaggggttt 1020
tatgcgatgg agtttcccca cactgagtgg gtggagactg aagttaggcc agcttggcac 1080
ttgatgtaat tctccttgga atttgccctt tttgagtttg gatcttggtt cattctcaag 1140
cctcagacag tggttcaaag tttttttctt ccatttcagg tgtcgtgaaa actaccccta 1200
aaagccagga tctgccacca tggctgacag ccgggatccc gccagcgacc agatgcagca 1260
ctggaaggag cagcgggccg cgcagaaagc tgatgtcctg accactggag ctggtaaccc 1320
agtaggagac aaacttaatg ttattacagt agggccccgt gggccccttc ttgttcagga 1380
tgtggttttc actgatgaaa tggctcattt tgaccgagag agaattcctg agagagttgt 1440
gcatgctaaa ggagcagggg cctttggcta ctttgaggtc acacatgaca ttaccaaata 1500
ctccaaggca aaggtatttg agcatattgg aaagaagact cccatcgcag ttcggttctc 1560
cactgttgct ggagaatcgg gttcagctga cacagttcgg gaccctcgtg ggtttgcagt 1620
gaaattttac acagaagatg gtaactggga tctcgttgga aataacaccc ccattttctt 1680
catcagggat cccatattgt ttccatcttt tatccacagc caaaagagaa atcctcagac 1740
acatctgaag gatccggaca tggtctggga cttctggagc ctacgtcctg agtctctgca 1800
tcaggtttct ttcttgttca gtgatcgggg gattccagat ggacatcgcc acatgaatgg 1860
atatggatca catactttca agctggttaa tgcaaatggg gaggcagttt attgcaaatt 1920
ccattataag actgaccagg gcatcaaaaa cctttctgtt gaagatgcgg cgagactttc 1980
ccaggaagat cctgactatg gcatccggga tctttttaac gccattgcca caggaaagta 2040
cccctcctgg actttttaca tccaggtcat gacatttaat caggcagaaa cttttccatt 2100
taatccattc gatctcacca aggtttggcc tcacaaggac taccctctca tcccagttgg 2160
taaactggtc ttaaaccgga atccagttaa ttactttgct gaggttgaac agatagcctt 2220
cgacccaagc aacatgccac ctggcattga ggccagtcct gacaaaatgc ttcagggccg 2280
cctttttgcc tatcctgaca ctcaccgcca tcgcctggga cccaattatc ttcatatacc 2340
tgtgaactgt ccctaccgtg ctcgagtggc caactaccag cgtgacggcc cgatgtgcat 2400
gcaggacaat cagggtggtg ctccaaatta ctaccccaac agctttggtg ctccggaaca 2460
acagccttct gccctggagc acagcatcca atattctgga gaagtgcgga gattcaacac 2520
tgccaatgat gataacgtta ctcaggtgcg ggcattctat gtgaacgtgc tgaatgagga 2580
acagaggaaa cgtctgtgtg agaacattgc cggccacctg aaggatgcac aaattttcat 2640
ccagaagaaa gcggtcaaga acttcactga ggtccaccct gactacggga gccacatcca 2700
ggctcttctg gacaagtaca atgctgagaa gcctaagaat gcgattcaca cctttgtgca 2760
gtccggatct cacttggcgg caagggagaa ggcaaatctg tgaaagcttg gtaccgaatt 2820
ccctgtgacc cctccccagt gcctctcctg gccctggaag ttgccactcc agtgcccacc 2880
agccttgtcc taataaaatt aagttgcatc attttgtctg actaggtgtc cttctataat 2940
attatggggt ggaggggggt ggtatggagc aaggggcaag ttgggaagac aacctgtagg 3000
gcctgcgggg tctattggga accaagctgg agtgcagtgg cacaatcttg gctcactgca 3060
atctccgcct cctgggttca agcgattctc ctgcctcagc ctcccgagtt gttgggattc 3120
caggcatgca tgaccaggct cagctaattt ttgttttttt ggtagagacg gggtttcacc 3180
atattggcca ggctggtctc caactcctaa tctcaggtga tctacccacc ttggcctccc 3240
aaattgctgg gattacaggc gtgaaccact gctcccttcc ctgtccttac gcgtagaatt 3300
ggtaaagaga gtcgtgtaaa atatcgagtt cgcacatctt gttgtctgat tattgatttt 3360
tggcgaaacc atttgatcat atgacaagat gtgtatctac cttaacttaa tgattttgat 3420
aaaaatcatt aactagtcca tggctgcctc gcgcgtttcg gtgatgacgg tgaaaacctc 3480
tgacacatgc agctcccgga gacggtcaca gcttgtctgt aagcggatgc cgggagcaga 3540
caagcccgtc agggcgcgtc agcgggtgtt ggcgggtgtc ggggcgcagc catgacccag 3600
tcacgtagcg atagcggagt gtatactggc ttaactatgc ggcatcagag cagattgtac 3660
tgagagtgca ccatatgcgg tgtgaaatac cgcacagatg cgtaaggaga aaataccgca 3720
tcaggcgctc ttccgcttcc tcgctcactg actcgctgcg ctcggtcgtt cggctgcggc 3780
gagcggtatc agctcactca aaggcggtaa tacggttatc cacagaatca ggggataacg 3840
caggaaagaa catgtgagca aaaggccagc aaaaggccag gaaccgtaaa aaggccgcgt 3900
tgctggcgtt tttccatagg ctccgccccc ctgacgagca tcacaaaaat cgacgctcaa 3960
gtcagaggtg gcgaaacccg acaggactat aaagatacca ggcgtttccc cctggaagct 4020
ccctcgtgcg ctctcctgtt ccgaccctgc cgcttaccgg atacctgtcc gcctttctcc 4080
cttcgggaag cgtggcgctt tctcatagct cacgctgtag gtatctcagt tcggtgtagg 4140
tcgttcgctc caagctgggc tgtgtgcacg aaccccccgt tcagcccgac cgctgcgcct 4200
tatccggtaa ctatcgtctt gagtccaacc cggtaagaca cgacttatcg ccactggcag 4260
cagccactgg taacaggatt agcagagcga ggtatgtagg cggtgctaca gagttcttga 4320
agtggtggcc taactacggc tacactagaa gaacagtatt tggtatctgc gctctgctga 4380
agccagttac cttcggaaaa agagttggta gctcttgatc cggcaaacaa accaccgctg 4440
gtagcggtgg tttttttgtt tgcaagcagc agattacgcg cagaaaaaaa ggatctcaag 4500
aagatccttt gatcttttct acggggtctg acgctcagtg gaacgaaaac tcacgttaag 4560
ggattttggt catgagatta tcaaaaagga tcttcaccta gatcctttta aattaaaaat 4620
gaagttttaa atcaatctaa agtatatatg agtaaacttg gtctgacagt taccaatgct 4680
taatcagtga ggcacctatc tcagcgatct gtctatttcg ttcatccata gttgcctgac 4740
tcc 4743
<210> 2
<211> 3607
<212> DNA
<213> Homo Sapiens
<400> 2
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaaa gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcagtagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacgcccctg gctgcagtac gtgattcttg atcccgagct tcgggttgga 360
agtgggtggg agagttcgag gccttgcgct taaggagccc cttcgcctcg tgcttgagtt 420
gaggcctggc ttgggcgctg gggccgccgc gtgcgaatct ggtggcacct tcgcgcctgt 480
ctcgctgctt tcgataagtc tctagccatt taaaattttt gatgacctgc tgcgacgctt 540
tttttctggc aagatagtct tgtaaatgcg ggccaagatc tgcacactgg tatttcggtt 600
tttggggccg cgggcggcga cggggcccgt gcgtcccagc gcacatgttc ggcgaggcgg 660
ggcctgcgag cgcggccacc gagaatcgga cgggggtagt ctcaagctgg ccggcctgct 720
ctggtgcctg gcctcgcgcc gccgtgtatc gccccgccct gggcggcaag gctggcccgg 780
tcggcaccag ttgcgtgagc ggaaagatgg ccgcttcccg gccctgctgc agggagctca 840
aaatggagga cgcggcgctc gggagagcgg gcgggtgagt cacccacaca aaggaaaagg 900
gcctttccgt cctcagccgt cgcttcatgt gactccacgg agtaccgggc gccgtccagg 960
cacctcgatt agttctcgag cttttggagt acgtcgtctt taggttgggg ggaggggttt 1020
tatgcgatgg agtttcccca cactgagtgg gtggagactg aagttaggcc agcttggcac 1080
ttgatgtaat tctccttgga atttgccctt tttgagtttg gatcttggtt cattctcaag 1140
cctcagacag tggttcaaag tttttttctt ccatttcagg tgtcgtgaaa actaccccta 1200
aaagccagga tccaccatgg cgacgaaggc cgtgtgcgtg ctgaagggcg acggcccagt 1260
gcagggcatc atcaatttcg agcagaagga aagtaatgga ccagtgaagg tgtggggaag 1320
cattaaagga ctgactgaag gcctgcatgg attccatgtt catgagtttg gagataatac 1380
agcaggctgt accagtgcag gtcctcactt taatcctcta tccagaaaac acggtgggcc 1440
aaaggatgaa gagaggcatg ttggagactt gggcaatgtg actgctgaca aagatggtgt 1500
ggccgatgtg tctattgaag attctgtgat ctcactctca ggagaccatt gcatcattgg 1560
ccgcacactg gtggtccatg aaaaagcaga tgacttgggc aaaggtggaa atgaagaaag 1620
tacaaagaca ggaaacgctg gaagtcgttt ggcttgtggt gtaattggga tcgcccaatg 1680
aattccctgt gacccctccc cagtgcctct cctggccctg gaagttgcca ctccagtgcc 1740
caccagcctt gtcctaataa aattaagttg catcattttg tctgactagg tgtccttcta 1800
taatattatg gggtggaggg gggtggtatg gagcaagggg caagttggga agacaacctg 1860
tagggcctgc ggggtctatt gggaaccaag ctggagtgca gtggcacaat cttggctcac 1920
tgcaatctcc gcctcctggg ttcaagcgat tctcctgcct cagcctcccg agttgttggg 1980
attccaggca tgcatgacca ggctcagcta atttttgttt ttttggtaga gacggggttt 2040
caccatattg gccaggctgg tctccaactc ctaatctcag gtgatctacc caccttggcc 2100
tcccaaattg ctgggattac aggcgtgaac cactgctccc ttccctgtcc ttacgcgtag 2160
aattggtaaa gagagtcgtg taaaatatcg agttcgcaca tcttgttgtc tgattattga 2220
tttttggcga aaccatttga tcatatgaca agatgtgtat ctaccttaac ttaatgattt 2280
tgataaaaat cattaactag tccatggctg cctcgcgcgt ttcggtgatg acggtgaaaa 2340
cctctgacac atgcagctcc cggagacggt cacagcttgt ctgtaagcgg atgccgggag 2400
cagacaagcc cgtcagggcg cgtcagcggg tgttggcggg tgtcggggcg cagccatgac 2460
ccagtcacgt agcgatagcg gagtgtatac tggcttaact atgcggcatc agagcagatt 2520
gtactgagag tgcaccatat gcggtgtgaa ataccgcaca gatgcgtaag gagaaaatac 2580
cgcatcaggc gctcttccgc ttcctcgctc actgactcgc tgcgctcggt cgttcggctg 2640
cggcgagcgg tatcagctca ctcaaaggcg gtaatacggt tatccacaga atcaggggat 2700
aacgcaggaa agaacatgtg agcaaaaggc cagcaaaagg ccaggaaccg taaaaaggcc 2760
gcgttgctgg cgtttttcca taggctccgc ccccctgacg agcatcacaa aaatcgacgc 2820
tcaagtcaga ggtggcgaaa cccgacagga ctataaagat accaggcgtt tccccctgga 2880
agctccctcg tgcgctctcc tgttccgacc ctgccgctta ccggatacct gtccgccttt 2940
ctcccttcgg gaagcgtggc gctttctcat agctcacgct gtaggtatct cagttcggtg 3000
taggtcgttc gctccaagct gggctgtgtg cacgaacccc ccgttcagcc cgaccgctgc 3060
gccttatccg gtaactatcg tcttgagtcc aacccggtaa gacacgactt atcgccactg 3120
gcagcagcca ctggtaacag gattagcaga gcgaggtatg taggcggtgc tacagagttc 3180
ttgaagtggt ggcctaacta cggctacact agaagaacag tatttggtat ctgcgctctg 3240
ctgaagccag ttaccttcgg aaaaagagtt ggtagctctt gatccggcaa acaaaccacc 3300
gctggtagcg gtggtttttt tgtttgcaag cagcagatta cgcgcagaaa aaaaggatct 3360
caagaagatc ctttgatctt ttctacgggg tctgacgctc agtggaacga aaactcacgt 3420
taagggattt tggtcatgag attatcaaaa aggatcttca cctagatcct tttaaattaa 3480
aaatgaagtt ttaaatcaat ctaaagtata tatgagtaaa cttggtctga cagttaccaa 3540
tgcttaatca gtgaggcacc tatctcagcg atctgtctat ttcgttcatc catagttgcc 3600
tgactcc 3607
<210> 3
<211> 3824
<212> DNA
<213> Homo Sapiens
<400> 3
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaaa gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcagtagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacgcccctg gctgcagtac gtgattcttg atcccgagct tcgggttgga 360
agtgggtggg agagttcgag gccttgcgct taaggagccc cttcgcctcg tgcttgagtt 420
gaggcctggc ttgggcgctg gggccgccgc gtgcgaatct ggtggcacct tcgcgcctgt 480
ctcgctgctt tcgataagtc tctagccatt taaaattttt gatgacctgc tgcgacgctt 540
tttttctggc aagatagtct tgtaaatgcg ggccaagatc tgcacactgg tatttcggtt 600
tttggggccg cgggcggcga cggggcccgt gcgtcccagc gcacatgttc ggcgaggcgg 660
ggcctgcgag cgcggccacc gagaatcgga cgggggtagt ctcaagctgg ccggcctgct 720
ctggtgcctg gcctcgcgcc gccgtgtatc gccccgccct gggcggcaag gctggcccgg 780
tcggcaccag ttgcgtgagc ggaaagatgg ccgcttcccg gccctgctgc agggagctca 840
aaatggagga cgcggcgctc gggagagcgg gcgggtgagt cacccacaca aaggaaaagg 900
gcctttccgt cctcagccgt cgcttcatgt gactccacgg agtaccgggc gccgtccagg 960
cacctcgatt agttctcgag cttttggagt acgtcgtctt taggttgggg ggaggggttt 1020
tatgcgatgg agtttcccca cactgagtgg gtggagactg aagttaggcc agcttggcac 1080
ttgatgtaat tctccttgga atttgccctt tttgagtttg gatcttggtt cattctcaag 1140
cctcagacag tggttcaaag tttttttctt ccatttcagg tgtcgtgaaa actaccccta 1200
aaagccagga tccgatatcg tcgaccacca tgttgagccg ggcagtgtgc ggcaccagca 1260
ggcagctggc tccggttttg gggtatctgg gctccaggca gaagcacagc ctccccgacc 1320
tgccctacga ctacggcgcc ctggaacctc acatcaacgc gcagatcatg cagctgcacc 1380
acagcaagca ccacgcggcc tacgtgaaca acctgaacgt caccgaggag aagtaccagg 1440
aggcgttggc caagggagat gttacagccc agatagctct tcagcctgca ctgaagttca 1500
atggtggtgg tcatatcaat catagcattt tctggacaaa cctcagccct aacggtggtg 1560
gagaacccaa aggggagttg ctggaagcca tcaaacgtga ctttggttcc tttgacaagt 1620
ttaaggagaa gctgacggct gcatctgttg gtgtccaagg ctcaggttgg ggttggcttg 1680
gtttcaataa ggaacgggga cacttacaaa ttgctgcttg tccaaatcag gatccactgc 1740
aaggaacaac aggccttatt ccactgctgg ggattgatgt gtgggagcac gcttactacc 1800
ttcagtataa aaatgtcagg cctgattatc taaaagctat ttggaatgta atcaactggg 1860
agaatgtaac tgaaagatac atggcttgca aaaagtgaat tccctgtgac ccctccccag 1920
tgcctctcct ggccctggaa gttgccactc cagtgcccac cagccttgtc ctaataaaat 1980
taagttgcat cattttgtct gactaggtgt ccttctataa tattatgggg tggagggggg 2040
tggtatggag caaggggcaa gttgggaaga caacctgtag ggcctgcggg gtctattggg 2100
aaccaagctg gagtgcagtg gcacaatctt ggctcactgc aatctccgcc tcctgggttc 2160
aagcgattct cctgcctcag cctcccgagt tgttgggatt ccaggcatgc atgaccaggc 2220
tcagctaatt tttgtttttt tggtagagac ggggtttcac catattggcc aggctggtct 2280
ccaactccta atctcaggtg atctacccac cttggcctcc caaattgctg ggattacagg 2340
cgtgaaccac tgctcccttc cctgtcctta cgcgtagaat tggtaaagag agtcgtgtaa 2400
aatatcgagt tcgcacatct tgttgtctga ttattgattt ttggcgaaac catttgatca 2460
tatgacaaga tgtgtatcta ccttaactta atgattttga taaaaatcat taactagtcc 2520
atggctgcct cgcgcgtttc ggtgatgacg gtgaaaacct ctgacacatg cagctcccgg 2580
agacggtcac agcttgtctg taagcggatg ccgggagcag acaagcccgt cagggcgcgt 2640
cagcgggtgt tggcgggtgt cggggcgcag ccatgaccca gtcacgtagc gatagcggag 2700
tgtatactgg cttaactatg cggcatcaga gcagattgta ctgagagtgc accatatgcg 2760
gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgct cttccgcttc 2820
ctcgctcact gactcgctgc gctcggtcgt tcggctgcgg cgagcggtat cagctcactc 2880
aaaggcggta atacggttat ccacagaatc aggggataac gcaggaaaga acatgtgagc 2940
aaaaggccag caaaaggcca ggaaccgtaa aaaggccgcg ttgctggcgt ttttccatag 3000
gctccgcccc cctgacgagc atcacaaaaa tcgacgctca agtcagaggt ggcgaaaccc 3060
gacaggacta taaagatacc aggcgtttcc ccctggaagc tccctcgtgc gctctcctgt 3120
tccgaccctg ccgcttaccg gatacctgtc cgcctttctc ccttcgggaa gcgtggcgct 3180
ttctcatagc tcacgctgta ggtatctcag ttcggtgtag gtcgttcgct ccaagctggg 3240
ctgtgtgcac gaaccccccg ttcagcccga ccgctgcgcc ttatccggta actatcgtct 3300
tgagtccaac ccggtaagac acgacttatc gccactggca gcagccactg gtaacaggat 3360
tagcagagcg aggtatgtag gcggtgctac agagttcttg aagtggtggc ctaactacgg 3420
ctacactaga agaacagtat ttggtatctg cgctctgctg aagccagtta ccttcggaaa 3480
aagagttggt agctcttgat ccggcaaaca aaccaccgct ggtagcggtg gtttttttgt 3540
ttgcaagcag cagattacgc gcagaaaaaa aggatctcaa gaagatcctt tgatcttttc 3600
tacggggtct gacgctcagt ggaacgaaaa ctcacgttaa gggattttgg tcatgagatt 3660
atcaaaaagg atcttcacct agatcctttt aaattaaaaa tgaagtttta aatcaatcta 3720
aagtatatat gagtaaactt ggtctgacag ttaccaatgc ttaatcagtg aggcacctat 3780
ctcagcgatc tgtctatttc gttcatccat agttgcctga ctcc 3824
<210> 4
<211> 3194
<212> DNA
<213> Homo Sapiens
<400> 4
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaaa gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcagtagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacgcccctg gctgcagtac gtgattcttg atcccgagct tcgggttgga 360
agtgggtggg agagttcgag gccttgcgct taaggagccc cttcgcctcg tgcttgagtt 420
gaggcctggc ttgggcgctg gggccgccgc gtgcgaatct ggtggcacct tcgcgcctgt 480
ctcgctgctt tcgataagtc tctagccatt taaaattttt gatgacctgc tgcgacgctt 540
tttttctggc aagatagtct tgtaaatgcg ggccaagatc tgcacactgg tatttcggtt 600
tttggggccg cgggcggcga cggggcccgt gcgtcccagc gcacatgttc ggcgaggcgg 660
ggcctgcgag cgcggccacc gagaatcgga cgggggtagt ctcaagctgg ccggcctgct 720
ctggtgcctg gcctcgcgcc gccgtgtatc gccccgccct gggcggcaag gctggcccgg 780
tcggcaccag ttgcgtgagc ggaaagatgg ccgcttcccg gccctgctgc agggagctca 840
aaatggagga cgcggcgctc gggagagcgg gcgggtgagt cacccacaca aaggaaaagg 900
gcctttccgt cctcagccgt cgcttcatgt gactccacgg agtaccgggc gccgtccagg 960
cacctcgatt agttctcgag cttttggagt acgtcgtctt taggttgggg ggaggggttt 1020
tatgcgatgg agtttcccca cactgagtgg gtggagactg aagttaggcc agcttggcac 1080
ttgatgtaat tctccttgga atttgccctt tttgagtttg gatcttggtt cattctcaag 1140
cctcagacag tggttcaaag tttttttctt ccatttcagg tgtcgtgaaa actaccccta 1200
aaagccagga tccgatatcg tcgaccacca tgctggcgct actgtgttcc tgcctgctcc 1260
tggcagccgg tgcctcggac gcctggacgg gcgaggactc ggcggagccc aactctgact 1320
cggcggagtg gatccgagac atgtacgcca aggtcacgga gatctggcag gaggtcatgc 1380
agcggcggga cgacgacggc acgctccacg ccgcctgcca ggtgcagccg tcggccacgc 1440
tggacgccgc gcagccccgg gtgaccggcg tcgtcctctt ccggcagctt gcgccccgcg 1500
ccaagctcga cgccttcttc gccctggagg gcttcccgac cgagccgaac agctccagcc 1560
gcgccatcca cgtgcaccag ttcggggacc tgagccaggg ctgcgagtcc accgggcccc 1620
actacaaccc gctggccgtg ccgcacccgc agcacccggg cgacttcggc aacttcgcgg 1680
tccgcgacgg cagcctctgg aggtaccgcg ccggcctggc cgcctcgctc gcgggcccgc 1740
actccatcgt gggccgggcc gtggtcgtcc acgctggcga ggacgacctg ggccgcggcg 1800
gcaaccaggc cagcgtggag aacgggaacg cgggccggcg gctggcctgc tgcgtggtgg 1860
gcgtgtgcgg gcccgggctc tgggagcgcc aggcgcggga gcactcagag cgcaagaagc 1920
ggcggcgcga gagcgagtgc aaggccgcct gaattccctg tgacccctcc ccagtgcctc 1980
tcctggccct ggaagttgcc actccagtgc ccaccagcct tgtcctaata aaattaagtt 2040
gcatcatttt gtctgactag gtgtccttct ataatattat ggggtggagg ggggtggtat 2100
ggagcaaggg gcaagttggg aagacaacct gtagggcctg cggggtctat tgggaaccaa 2160
gctggagtgc agtggcacaa tcttggctca ctgcaatctc cgcctcctgg gttcaagcga 2220
ttctcctgcc tcagcctccc gagttgttgg gattccaggc atgcatgacc aggctcagct 2280
aatttttgtt tttttggtag agacggggtt tcaccatatt ggccaggctg gtctccaact 2340
cctaatctca ggtgatctac ccaccttggc ctcccaaatt gctgggatta caggcgtgaa 2400
ccactgctcc cttccctgtc cttacgcgta gaattggtaa agagagtcgt gtaaaatatc 2460
gagttcgcac atcttgttgt ctgattattg atttttggcg aaaccatttg atcatatgac 2520
aagatgtgta tctaccttaa cttaatgatt ttgataaaaa tcattaacta gtccatggct 2580
gcctcgcgcg tttcggtgat gacggtgaaa acctctgaca catgcagctc ccggagacgg 2640
tcacagcttg tctgtaagcg gatgccggga gcagacaagc ccgtcagggc gcgtcagcgg 2700
gtgttggcgg gtgtcggggc gcagccatga cccagtcacg tagcgatagc ggagtgtata 2760
ctggcttaac tatgcggcat cagagcagat tgtactgaga gtgcaccata tgcggtgtga 2820
aataccgcac agatgcgtaa ggagaaaata ccgcatcagg cgctcttccg cttcctcgct 2880
cactgactcg ctgcgctcgg tcgttcggct gcggcgagcg gtatcagctc actcaaaggc 2940
ggtaatacgg ttatccacag aatcagggga taacgcagga aagaacatgt gagcaaaagg 3000
ccagcaaaag gccaggaacc gtaaaaaggc cgcgttgctg gcgtttttcc ataggctccg 3060
cccccctgac gagcatcaca aaaatcgacg ctcaagtcag aggtggcgaa acccgacagg 3120
actataaaga taccaggcgt ttccccctgg aagctccctc gtgcgctctc ctgttccgac 3180
cctgccgctt accg 3194
<---
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ОЛИГОНУКЛЕОТИДОВ
(1) CAT_F ATAGATCTGCCACCATGGCTGACAGCCGGGATC
(2) CAT_R TATAAGCTTCACAGATTTGCCTTCTCCCT
(3) CAT_SF ACCAAGGTTTGGCCTCACAA
(4) CAT_SR CGGTGAGTGTCAGGATAGGC
(5) SOD1_F AGGATCCACCATGGCGACGAAGGCCGTG
(6) SOD1_R TATGAATTCATTGGGCGATCCCAATTACAC
(7) SOD1_SF GACTGAAGGCCTGCATGGAT
(8) SOD1_SR ATCGGCCACACCATCTTTGT
(9) SOD2_F ATCGTCGACCACCATGTTGAGCCGGGCAGTGTG
(10) SOD2_R TATGAATTCACTTTTTGCAAGCCATGTATCT
(11) SOD2_SF GTGGAGAACCCAAAGGGGAG
(12) SOD2_SR GCCTGTTGTTCCTTGCAGTG
(13) SOD3_F ATCGTCGACCACCATGCTGGCGCTACTGTGTTC
(14) SOD3_R TATGAATTCAGGCGGCCTTGCACTCGCT
(15) SOD3_SF GATCCGAGACATGTACGCCA
(16) SOD3_SR GAAGAAGGCGTCGAGCTTGG
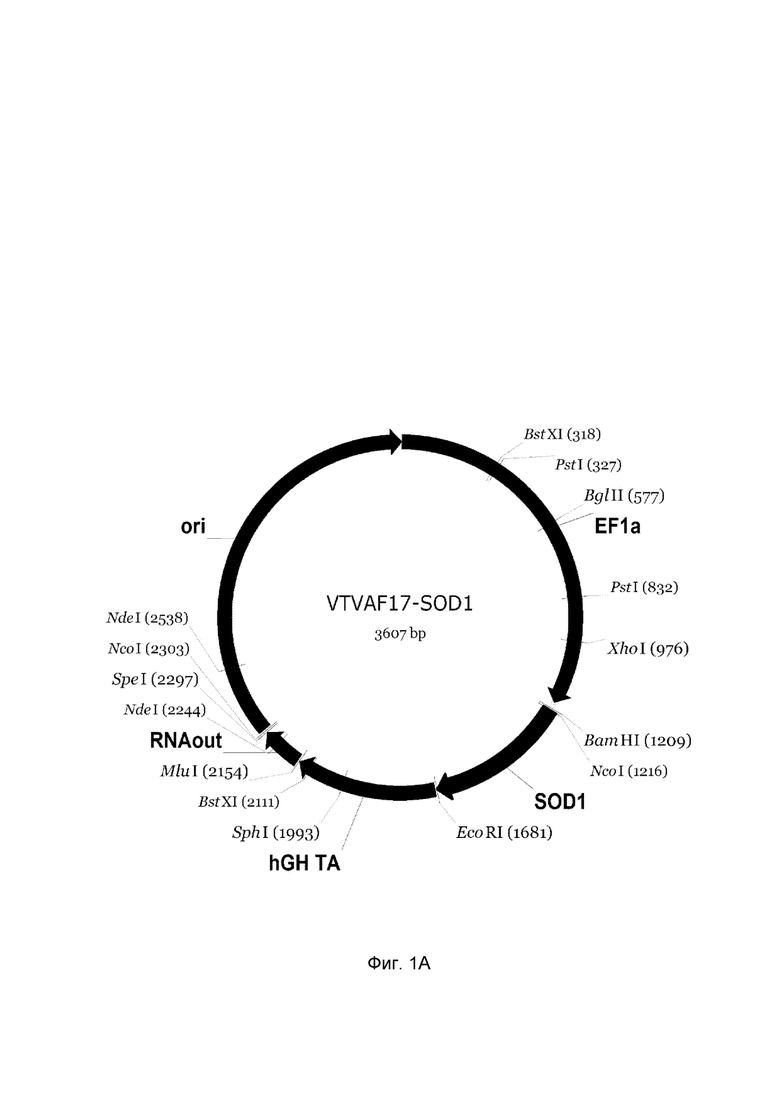

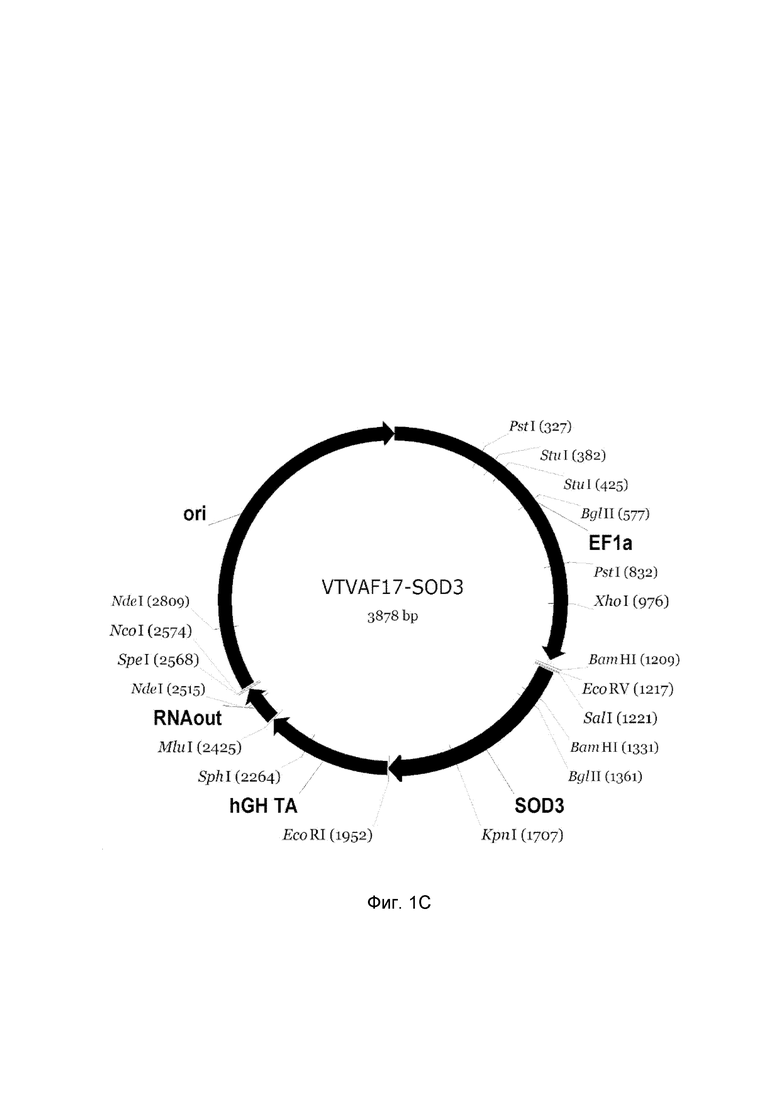

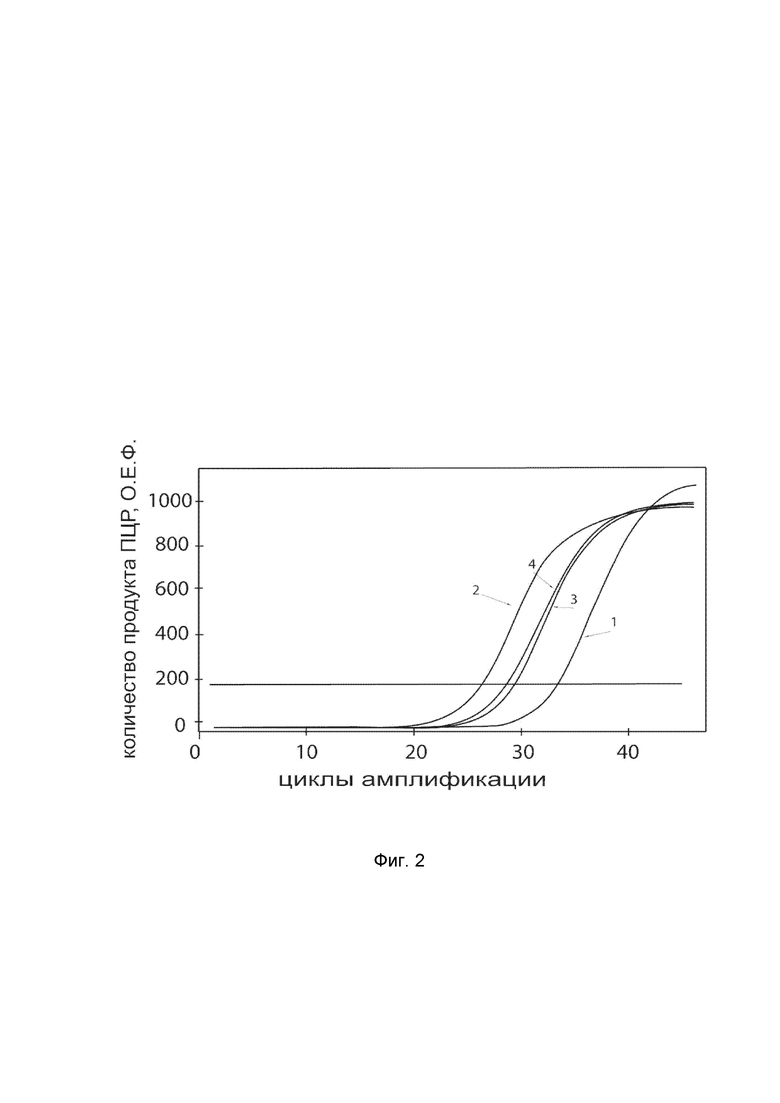

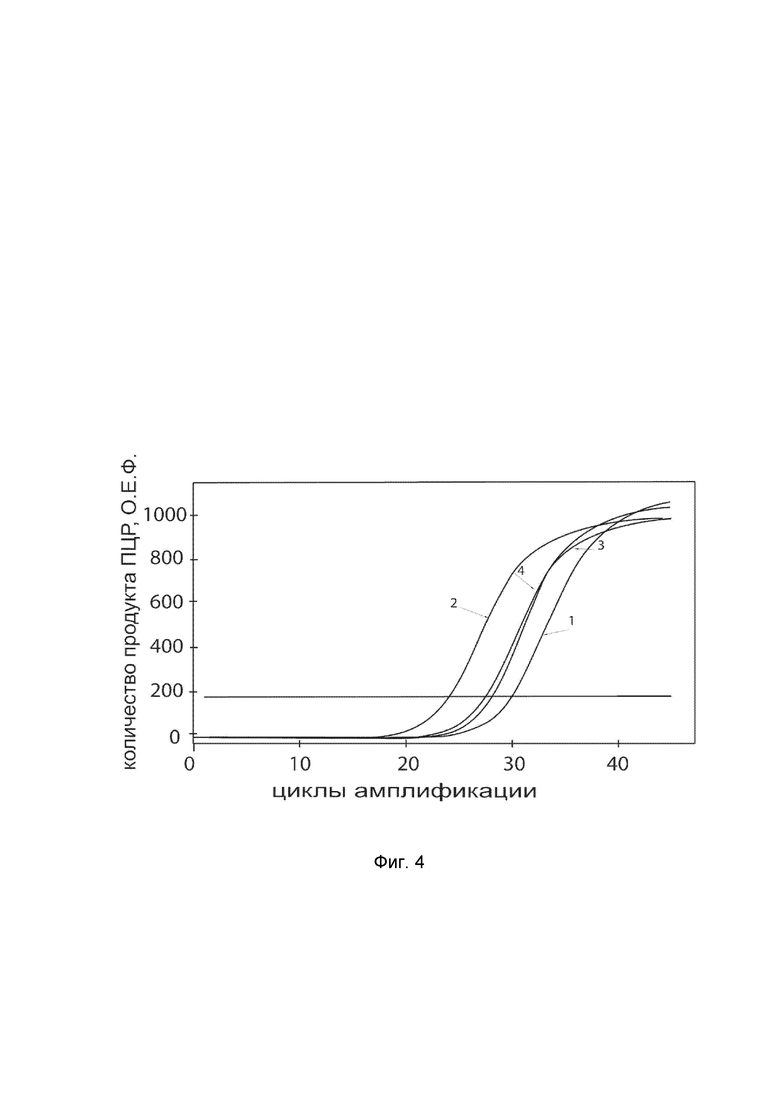

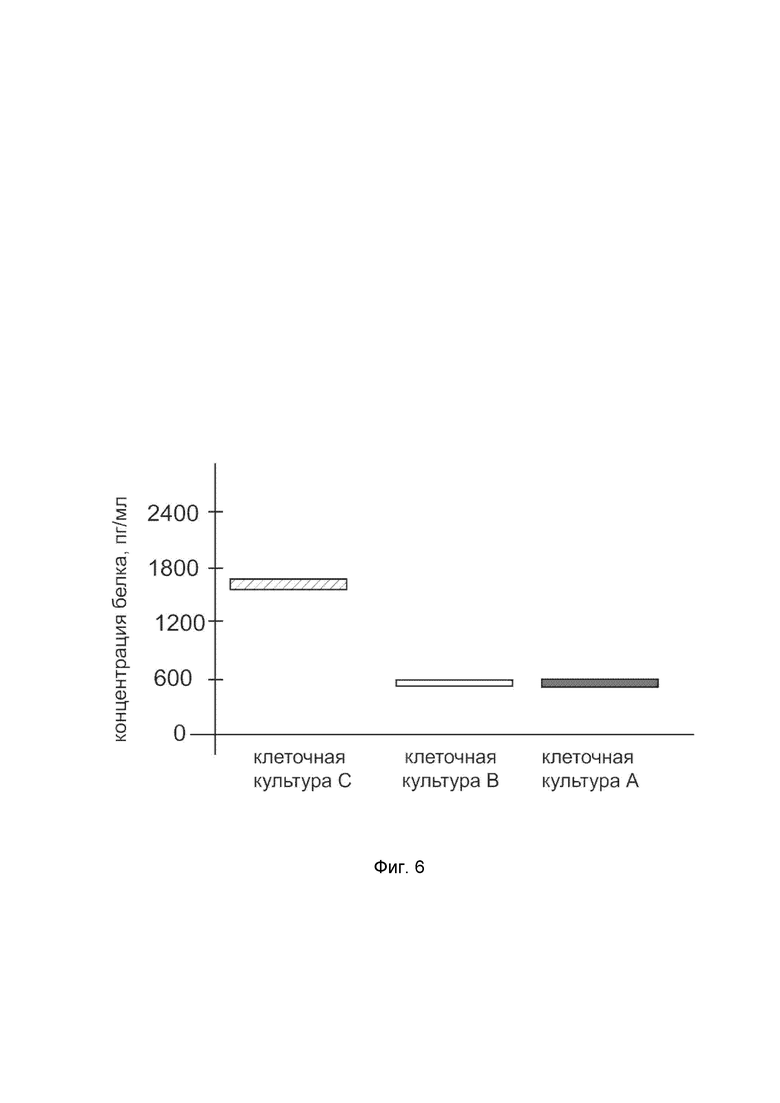
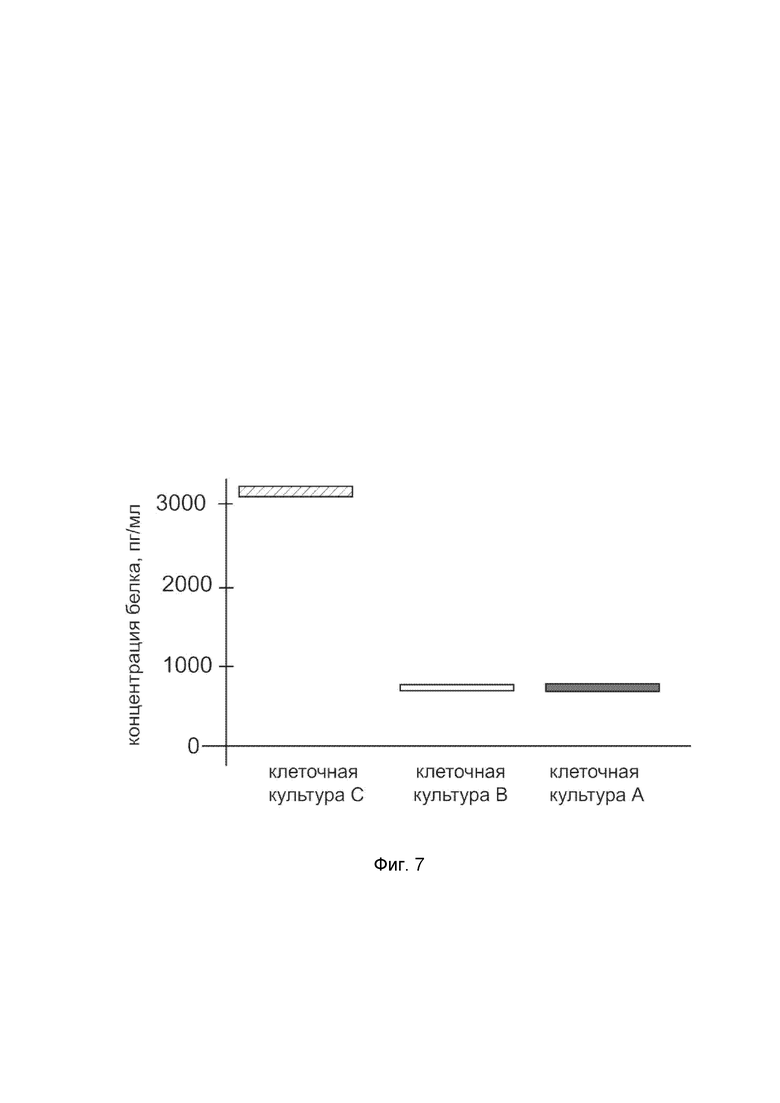

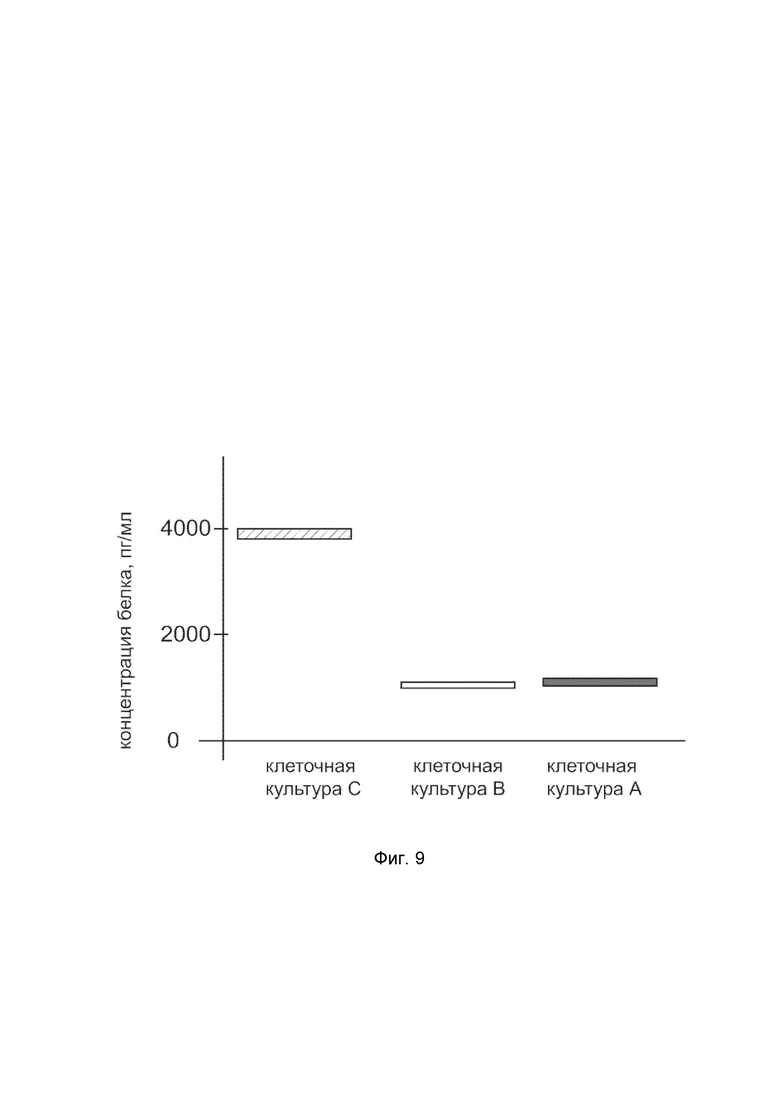

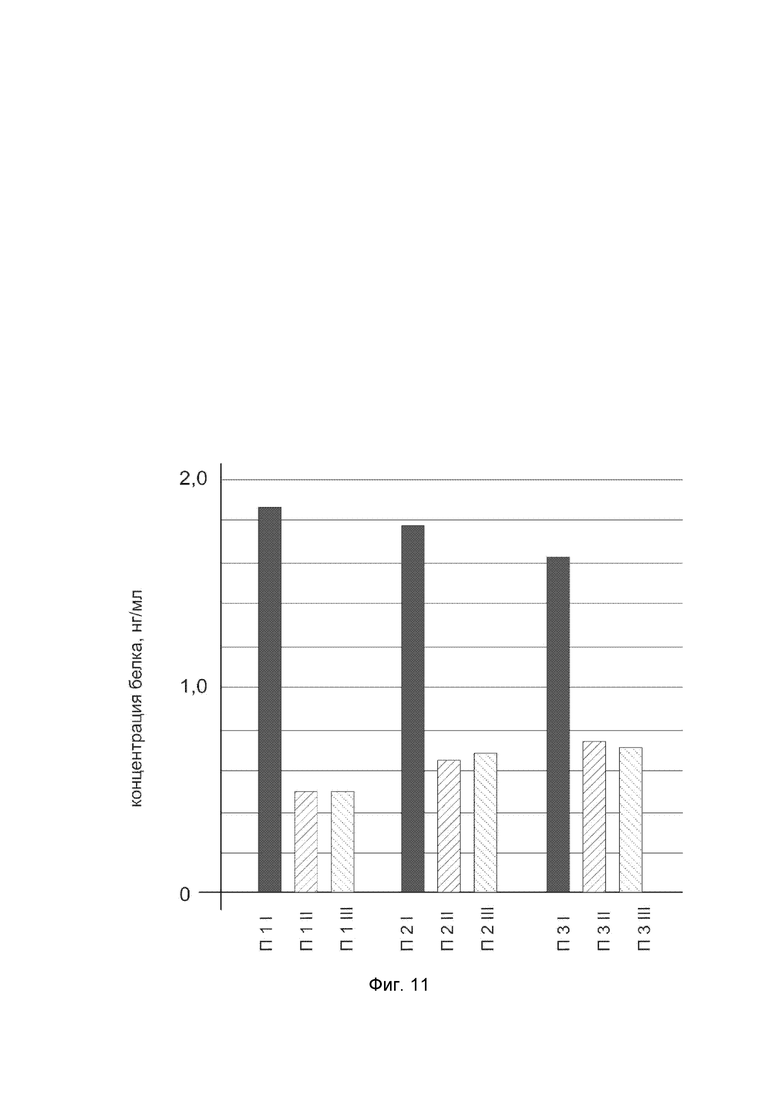

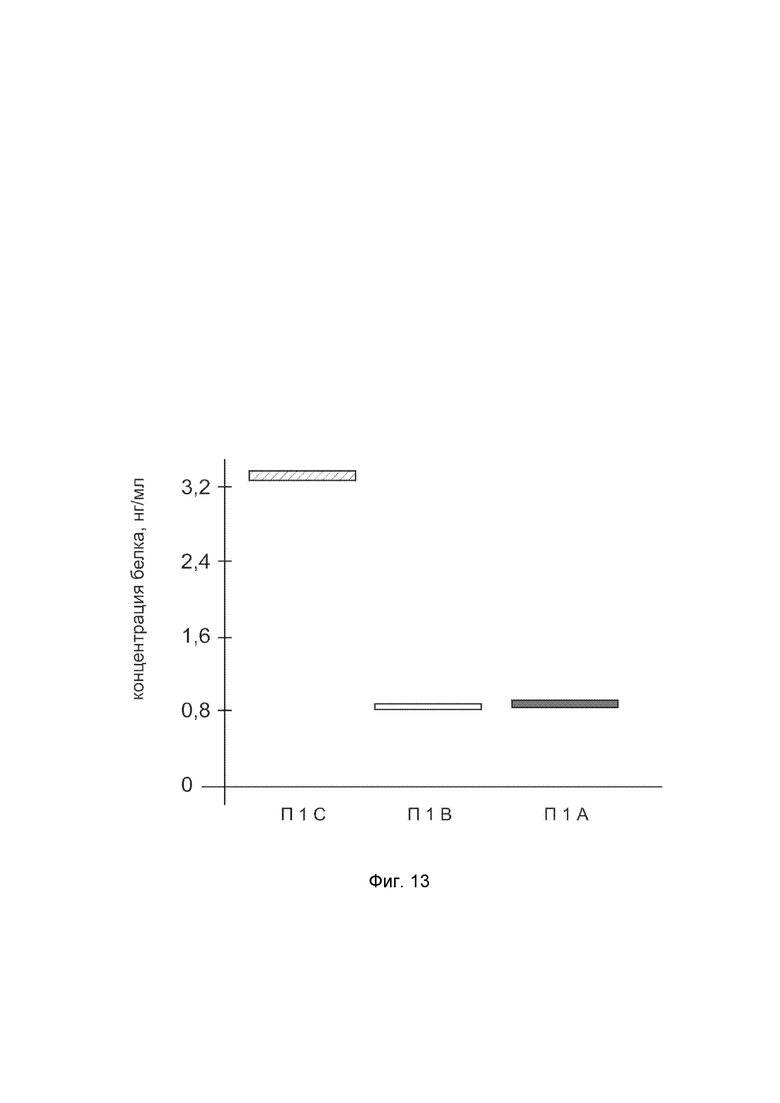
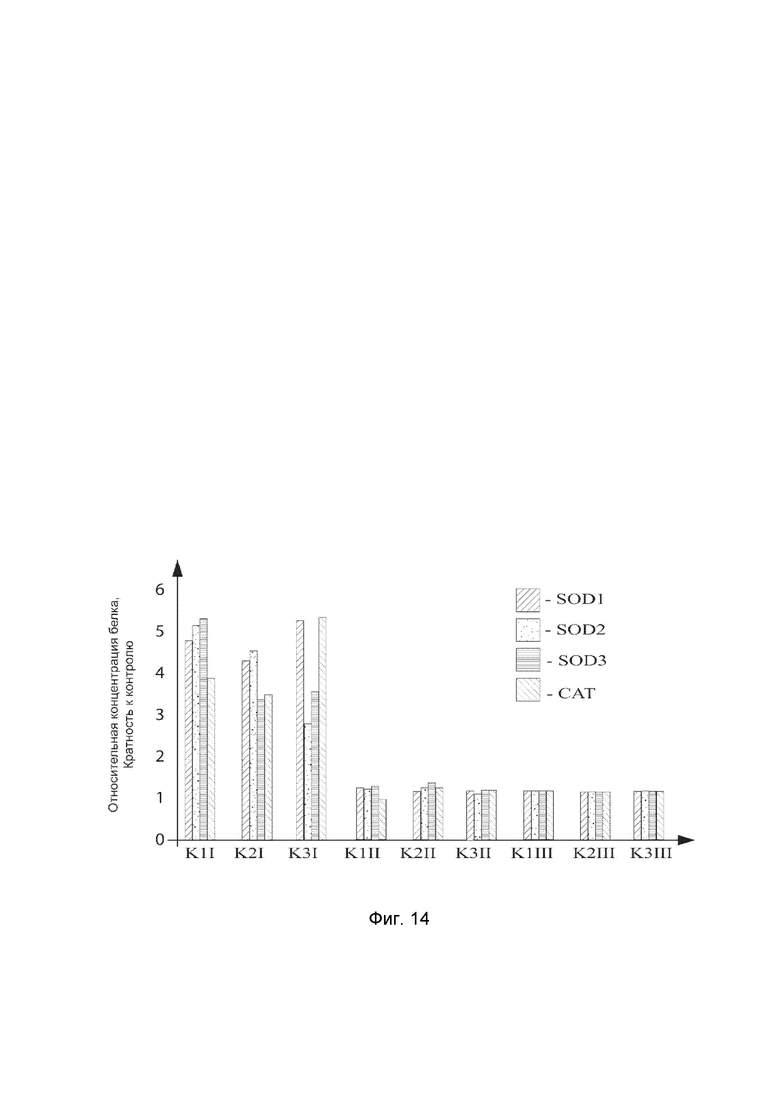

Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии. Предложен генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген, выбранный из группы генов SOD1, SOD2, SOD3, CAT для повышения уровня экспрессии этого целевого гена в организме человека и животных. При этом генотерапевтический ДНК-вектор VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT имеет нуклеотидную последовательность SEQ ID №1, или SEQ ID №2, или SEQ ID №3, или SEQ ID №4 соответственно. Предложены также способ получения указанного вектора, применение вектора, штамм Escherichia coli, несущий указанный вектор, а также способ производства в промышленных масштабах указанного вектора. Изобретение обеспечивает возможность его безопасного применения для генетической терапии человека и животных. 12 н. и 2 з.п. ф-лы, 15 ил., 20 пр.
1. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена SOD1, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-SOD1, с нуклеотидной последовательностью SEQ ID №1.
2. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена SOD2, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-SOD2, с нуклеотидной последовательностью SEQ ID №2.
3. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена SOD3, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-SOD3, с нуклеотидной последовательностью SEQ ID №3.
4. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена CAT, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-CAT, с нуклеотидной последовательностью SEQ ID № 4.
5. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген SOD1, SOD2, SOD3, CAT по пп.1-3 или 4, отличающийся тем, что каждый из созданных генотерапевтических ДНК-векторов: VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT по пп.1-3 или 4 за счет ограниченного размера векторной части VTvaf17, не превышающей 3200 п.н., обладает способностью эффективно проникать в клетки человека и животных и экспрессировать клонированный в него целевой ген SOD1, или SOD2, или SOD3, или CAT.
6. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген SOD1, SOD2, SOD3, CAT по пп.1-3 или 4, отличающийся тем, что в составе каждого из созданных генотерапевтических ДНК-векторов: VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT по пп.1-3 или 4 в качестве структурных элементов используются нуклеотидные последовательности, которые не являются генами антибиотикорезистентности, вирусными генами и регуляторными элементами вирусных геномов, обеспечивая возможность его безопасного применения для генетической терапии человека и животных.
7. Способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SOD1, SOD2, SOD3, CAT по пп.1-3 или 4, заключающийся в том, что каждый из генотерапевтических ДНК-векторов: VTvaf17-SOD1, или VTvaf17-SOD2, или VTvaf17-SOD3, или VTvaf17-CAT получают следующим образом: кодирующую часть целевого гена SOD1, SOD2, SOD3, CAT по пп.1-3 или 4 клонируют в генотерапевтический ДНК-вектор VTvaf17 и получают генотерапевтический ДНК-вектор VTvaf17-SOD1, SEQ ID №1, или VTvaf17-SOD2, SEQ ID №2, или VTvaf17-SOD3, SEQ ID №3, или VTvaf17-CAT, SEQ ID №4, соответственно, при этом кодирующую часть целевого гена SOD1, или SOD2, или SOD3, или CAT получают путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции и ПЦР-амплификации с использованием созданных олигонуклеотидов и расщеплением продукта амплификации соответствующими эндонуклеазами рестрикции, причем клонирование в генотерапевтический ДНК-вектор VTvaf17 проводят по сайтам рестрикции NheI и HindIII, причем селекцию проводят без антибиотиков,
при этом при получении генотерапевтического ДНК-вектора VTvaf17-SOD1, SEQ ID №1 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
SOD1_F AGGATCCACCATGGCGACGAAGGCCGTG
SOD1_R TATGAATTCATTGGGCGATCCCAATTACAC,
а расщепление продукта амплификации и клонирование кодирующей части гена SOD1 в генотерапевтический ДНК-вектор VTvaf17 проводят с использованием эндонуклеаз рестрикции BamHI и EcoRI,
причем при получении генотерапевтического ДНК-вектора VTvaf17-SOD2, SEQ ID №2 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
SOD2_F ATCGTCGACCACCATGTTGAGCCGGGCAGTGTG
SOD2_R TATGAATTCACTTTTTGCAAGCCATGTATCT,
а расщепление продукта амплификации и клонирование кодирующей части гена SOD2 в генотерапевтический ДНК-вектор VTvaf17 проводят с использованием эндонуклеаз рестрикции SalI и EcoRI,
причем при получении генотерапевтического ДНК-вектора VTvaf17-SOD3, SEQ ID №3 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
SOD3_F ATCGTCGACCACCATGCTGGCGCTACTGTGTTC
SOD3_R TATGAATTCAGGCGGCCTTGCACTCGCT,
а расщепление продукта амплификации и клонирование кодирующей части гена SOD3 в генотерапевтический ДНК-вектор VTvaf17 проводят с использованием эндонуклеаз рестрикции SalI и EcoRI,
причем при получении генотерапевтического ДНК-вектора VTvaf17-CAT, SEQ ID №4 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
CAT_F ATAGATCTGCCACCATGGCTGACAGCCGGGATC
CAT_R TATAAGCTTCACAGATTTGCCTTCTCCCT,
а расщепление продукта амплификации и клонирование кодирующей части гена CAT в генотерапевтический ДНК-вектор VTvaf17 проводят с использованием эндонуклеаз рестрикции BamHII и HindIII.
8. Способ использования генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SOD1, SOD2, SOD3, CAT по пп.1-3 или 4, для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения, заключающийся в трансфекции выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора VTvaf17, или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, клеток органов и тканей пациента или животного, и/или во введении в органы и ткани пациента или животного аутологичных клеток этого пациента или животного, трансфицированных выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора VTvaf17 или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, и/или во введении в органы и ткани пациента или животного выбранного генотерапевтического ДНК-вектора, несущего целевой ген на основе генотерапевтического ДНК-вектора VTvaf17 или нескольких выбранных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, или в сочетании обозначенных способов.
9. Способ получения штамма для производства генотерапевтического ДНК-вектора по пп.1-3 или 4 для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения, заключающийся в получении электрокомпетентных клеток штамма Escherichia coli SCS110-AF с последующим проведением электропорации этих клеток генотерапевтическим ДНК-вектором VTvaf17-SOD1, или ДНК-вектором VTvaf17-SOD2, или ДНК-вектором VTvaf17-SOD3, или ДНК-вектором VTvaf17-CAT, после чего клетки высеивают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола с получением в результате штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT.
10. Штамм Escherichia coli SCS110-AF/VTvaf17-SOD1, полученный способом по п.9, несущий генотерапевтический ДНК-вектор VTvaf17-SOD1, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения.
11. Штамм Escherichia coli SCS110-AF/VTvaf17-SOD2, полученный способом по п.9, несущий генотерапевтический ДНК-вектор VTvaf17-SOD2, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения.
12. Штамм Escherichia coli SCS110-AF/VTvaf17-SOD3, полученный способом по п.9, несущий генотерапевтический ДНК-вектор VTvaf17-SOD3, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся, отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения.
13. Штамм Escherichia coli SCS110-AF/VTvaf17-CAT, полученный способом по п.9, несущий генотерапевтический ДНК-вектор VTvaf17-CAT, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения.
14. Способ производства в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SOD1, SOD2, SOD3, CAT по пп.1-3 или 4, для лечения заболеваний, связанных с недостаточной экспрессией целевого гена и характеризующихся дефицитом ферментов семейства супероксиддисмутаз и каталазы, для предотвращения развития атеросклероза, защиты организма от оксидативного стресса и токсического воздействия активных форм кислорода, для предотвращения развития спектра заболеваний, включающих в себя, но не ограничивающихся отклонениями, в которых оксидативный стресс играет патогенетическую роль в развитии наследственных сердечно-сосудистых, онкологических, дегенеративных и аутоиммунных заболеваний, когнитивных дисфункций и старения, заключающийся в том, что генотерапевтический ДНК-вектор VTvaf17-SOD1, или генотерапевтический ДНК-вектор VTvaf17-SOD2, или генотерапевтический ДНК-вектор VTvaf17-SOD3, или генотерапевтический ДНК-вектор VTvaf17-CAT получают путем того, что засевают в колбу с приготовленной средой затравочную культуру, выбранную из штамма Escherichia coli SCS110-AF/VTvaf17-SOD1, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD2, или штамма Escherichia coli SCS110-AF/VTvaf17-SOD3, или штамма Escherichia coli SCS110-AF/VTvaf17-CAT, затем инкубируют затравочную среду в шейкере-инкубаторе и переносят ее в промышленный ферментер, после чего растят до достижения стационарной фазы, затем выделяют фракцию, содержащую целевой ДНК-продукт, многостадийно фильтруют и очищают хроматографическими методами.

