ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области медицины и фармацевтической промышленности, в частности, к разработке химерных антигенов против вируса гепатита C (HCV) и вакцинных композиций, содержащих их. В этом изобретении используют минимальное количество компонентов, поскольку точный выбор конкретных областей антигенов HCV и включение искусственных эпитопов, специфичных для CD4+ T-лимфоцитов, позволяет химерным антигенам индуцировать сильный и широкий иммунный ответ против HCV.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Вирусом HCV инфицировано примерно 3% мирового населения (Williams R. Hepatology 2006; 44: 521-526). У большинства инфицированных людей инфекция переходит в хроническую форму (Amoroso P., et al., J Hepatol 1998; 28: 939-944). Инфекция гепатита C является одной из основных причин хронического повреждения печени, цирроза печени, печеночной недостаточности и рака печени (Hoofnagle J.H. Hepatology 2002; 36 (5 Suppl 1): S21-S29). В настоящее время HCV-инфекция является основной причиной трансплантации печени в странах первого мира. Кроме того, HCV-инфекция связана с внепеченочными проявлениями, такими как криоглобулинемия II типа, мембранопролиферативный гломерулонефрит, поздняя кожная порфирия, среди прочих. В настоящее время профилактическая вакцина против этого вируса отсутствует и используемые обычные методы противовирусного лечения, основанные на комбинации пегилированный интерферон (IFN)+рибавирин, эффективны менее чем в 50% случаев. Более того, вышеупомянутые методы лечения вызывают множество побочных эффектов (Ghany M.G. et al., Hepatology. 2009; 49 (4): 1335-74).
HCV принадлежит к роду Hepacivirus семейства Flaviviridae. Это оболочечный вирус, вирусные частицы которого имеют диаметр примерно 50-70 нм и связаны с липопротеинами очень низкой плотности (ЛПОНП) (VLDL) (Popescu C.I. and Dubuisson J. Biol Cell. 2009; 102 (1): 63-74). Вирусный геном представляет собой положительную цепь рибонуклеиновой кислоты (РНК) размером примерно 9,6 т.н. Геном кодирует вирусный полибелок, который процессируется ко- и посттрансляционно в по меньшей мере 10 вирусных белков: Core, E1, E2, p7 (структурные белки) и неструктурные белки: NS2, NS3, NS4A, NS4B, NS5A, NS5B (Bartenschlager R. and Lohmann V. 2000. J Gen Virol. 81: 1631-48).
Существует несколько серьезных препятствий для разработки эффективной вакцины против HCV. Этот патоген представляет собой РНК вирус, который может быстро мутировать, приспосабливаясь к среде хозяина. Это способствует широкому разнообразию многочисленных вирусных изолятов, идентифицированных по всему миру. Определено шесть основных генотипов HCV, различия нуклеотидных последовательностей которых могут составлять до 30% (Simmonds P. J Hepatol. 1999; 31 Suppl 1: 54-60). Наибольшая гетерогенность наблюдается в гипервариабельной области белка E2 HCV, где обнаружен эпитоп, потенциально являющийся мишенью для нейтрализующих антител. Фактически, HCV циркулирует в организме в виде гетерогенной популяции вирусных молекул, это явление известно как квазивиды (Simmonds P. J Gen Virol. 2004; 85 (Pt 11): 3173-88). Показано, что мутации являются способом, за счет которого вирусам удается избегать специфического гуморального и клеточного иммунного ответа, вырабатываемого хозяином.
Следует подчеркнуть, что HCV вызывает хроническую инфекцию у иммунокомпетентных индивидуумов, несмотря на наличие активного иммунного ответа (Lechmann et al., Semin Liver Dis 2000, 20, 211-226). В настоящее время выяснено несколько вирусных эффектов, которые способствуют сохранению инфекции за счет стимулирования нерелевантных иммунных ответов и предотвращения эффективного иммунного ответа. Эти эффекты обнаружены как в случае врожденного, так и приобретенного иммунитета (Grakoui A. et al. Science 2003, 302 (5645): 659-62). Существуют доказательства того, что неэффективный иммунный ответ против HCV не только не устраняет этот патоген, но также способствует повреждению печени.
На сегодняшний день иммунологические параметры, коррелирующие с защитой и избавлением от HCV, полностью не определены. Однако считается, что особенно большое значение имеет индукция мощного и устойчивого клеточного иммунного ответа против различных антигенов HCV (Lechmann et al., Semin Liver Dis 2000, 20, 211-226). У хронически инфицированных HCV пациентов нарушение специфического ответа T-лимфоцитов является особенно важным. Судя по всему, в данный эффект вносят вклад несколько механизмов; одним из них является действие регуляторных Т-клеток.
Почти все стратегии иммунизации были использованы в попытках разработать вакцину против HCV. Некоторые из этих стратегий включают: рекомбинантные белки, синтетические пептиды, вирусоподобные частицы, «голую» дезоксирибонуклеиновую кислоту (ДНК) и рекомбинантные вирусы. Все вирусные антигены были оценены в качестве мишеней для потенциальных вакцин против HCV. Большинство вакцин-кандидатов находятся на стадии изучения иммуногенности в животных моделях. Тем не менее, в настоящее время некоторые кандидаты достигли стадии клинической оценки, показано, что они являются безопасными и иммуногенными, однако четкий клинический эффект пока не был продемонстрирован (Alvarez-Lajonchere L., Dueñas-Carrera S. Int Rev Immunol. 2012; 31(3): 223-42).
Разработка субъединичной белковой вакцины-кандидата была одной из первых стратегий, изученных с целью получения вакцины против HCV. Некоторые из этих кандидатов на основе структурных антигенов позволяют добиться ограниченной защиты против заражения вирусами в животных моделях. Это относится к ситуации с шимпанзе, иммунизированными олигомером E1 и E2. Были иммунизированы семь шимпанзе, у пятерых из них развилась защита и две стали инфицированы, но затем избавились от вируса без достижения хронической стадии (Choo et al., Proc Natl Acad Sci USA 1994, 91, 1294-1298). Эта защита коррелировала с присутствием антител, способных ингибировать взаимодействие между белком E2 и человеческими клетками (Rosa et al., Proc Natl Acad Sci USA 1996, 93, 1759-1763).
Рекомбинантный белок E1 из изолята генотипа 1b очищали в виде гомодимеров (Maertens et al., Acta Gastroenterol Belg 2000, 63, 203). Два шимпанзе, хронически инфицированных HCV, получали 9 доз по 50 мкг этого рекомбинантного белка E1. Вакцинация приводила к улучшению гистологии печени, клиренсу вирусных антигенов из печени и снижению уровней аланинаминотрансферазы. Однако сывороточные уровни РНК не менялись в процессе лечения, и воспаление печени и вирусные антигены вновь появлялись после завершения лечения. Наблюдали связь между высокими уровнями анти-E1 антител и уменьшением повреждения печени (Maertens et al., Acta Gastroenterol Belg 2000, 63, 203). Действие варианта белка E1, сформулированного в квасцах, изучали на людях. Этот кандидат был безопасным и иммуногенным, индуцировал продукцию специфических антител и лимфопролиферативные ответы (Nevens F., et al., Hepatology. 2003; 38 (5): 1289-96). Однако введение этого кандидата не влияло на клиническое течение HCV-инфекции, поскольку пациенты не избавлялись от вируса и не наблюдалось улучшения гистологии печени. Недостаток подхода с белковыми субъединицами заключается в том, что в некоторых случаях он не приводил к индукции сильного клеточного иммунного ответа. Этот подход может иметь еще один недостаток: введение областей, вовлеченных в различные механизмы нарушения HCV-специфического иммунного ответа, вызываемого патогеном на разных уровнях (Grakoui A et al., Science 2003, 302 (5645): 659-62).
Две вакцины-кандидата на основе смесей синтетических пептидов, включая эпитопы T-лимфоцитов, также достигли стадии клинических испытаний (Yutani et al., Cancer Sci 2009, 100(10): 1935-42, Klade et al., Gastroenterology 2008, 134(5): 1385-95). Оба кандидата индуцировали специфические иммунные ответы и имели низкую реактогенность у хронически инфицированных HCV пациентов во время фаз I и II клинических испытаний, проведенных до настоящего времени (Alvarez-Lajonchere L, Dueñas-Carrera S. Int Rev Immunol. 2012; 31 (3): 223-42). Однако эти вакцины-кандидаты не оказывали существенного влияния на вирусную нагрузку или оказывали временное влияние. Принимая во внимание, что эти кандидаты не вызывали никакого улучшения в гистологии печени, их клинический эффект еще предстоит продемонстрировать. Различные эпитопы для CD4+ и CD8+ T-клеток, которые могут быть важны для клиренса вируса, были идентифицированы на всем протяжении полибелка HCV. Эти данные свидетельствуют в пользу стратегии получения вакцин на основе синтетических пептидов. Различные пептиды, включая эпитопы Core, NS4 и NS5, самостоятельно или с липидными фрагментами, индуцировали сильные ответы цитотоксических T-лимфоцитов у мышей (Shirai et al., J Infect Dis 1996, 173, 24-31; Hiranuma et al., J Gen Virol 1999, 80, 187-193; Oseroff et al., Vaccine 1998, 16, 823-833). Основной недостаток этого подхода состоит в том, что эти пептиды в отсутствие функции T-хелперов могут быть плохими иммуногенами. Кроме того, эффективность вакцины часто зависит от индукции мультивалентного и с широким спектром иммунного ответа против нескольких антигенов. При увеличении количества пептидов, включенных в вакцину, сложность препарата возрастает со всех точек зрения. Эти ограничения являются недостатками данного подхода.
С другой стороны, различные рекомбинантные вирусные векторы прошли оценку в качестве вакцин-кандидатов против HCV. Дефектные рекомбинантные аденовирусы являются привлекательными кандидатами вследствие их высокого тропизма к печени, их способности индуцировать гуморальный и клеточный иммунные ответы, а также возможности их введения пероральным и парентеральным путями. Рекомбинантные аденовирусы, экспрессирующие структурные белки HCV, индуцируют продукцию антител против каждого из этих белков (Makimura et al., Vaccine 1996, 14, 28-36). Кроме того, после иммунизации мышей рекомбинантным аденовирусом с Core и E1, был обнаружен специфический иммунный ответ цитотоксических T-лимфоцитов против этих антигенов (Bruna-Romero et al., Hepatology 1997, 25, 470-477). Хотя эти результаты являются обнадеживающими, некоторые проблемы, связанные с использованием рекомбинантных аденовирусов в генной терапии, вызывают сомнения по поводу их безопасности для людей. В настоящее время вакцины-кандидаты против HCV на основе рекомбинантного аденовируса проходят оценку в клинических испытаниях с хорошими результатами по иммуногенности, но без признаков клинического эффекта (Barnes et al., Sci Transl Med. 2012; 4(115): 115). Использование других рекомбинантных вирусных векторов, таких как вирус осповакцины, вирус оспы кур и вирус канарипокс, содержащих различные гены HCV, приводило к индукции сильных ответов цитотоксических T-лимфоцитов и T-хелперов у мышей (Shirai et al., J Virol 1994, 68, 3334-3342; Large et al., J Immunol 1999, 162, 931-938). В частности, модифицированный вирус осповакцины Ankara, рекомбинантный для неструктурных антигенов NS3-NS5 HCV, прошел оценку в клинических испытаниях на людях (Fournillier et al., Vaccine, 2007; 25 (42): 7339-53). Этот кандидат был иммуногенным и отличался хорошей переносимостью в фазе I клинических испытаний с участием хронически инфицированных HCV пациентов. Аналогично подходу с использованием пептидов, эффект в отношении вирусной нагрузки был временным и наблюдался только у части вакцин; таким образом, клинический эффект по-прежнему не был продемонстрирован. Как правило, вакцины-кандидаты на основе рекомбинантных вирусов встречают препятствия с точки зрения безопасности и регулирующих положений для их применения.
В качестве стратегии для разработки вакцины против HCV была всесторонне изучена ДНК-иммунизация. Исследования на животных моделях показали способность этих кандидатов индуцировать клеточный и гуморальный иммунные ответы против почти всех антигенов HCV (Alvarez-Lajonchere L., Dueñas-Carrera S., Hum Vaccin. 2009; 5 (8): 568-71). Две вакцины-кандидата, включающие плазмиды для ДНК-иммунизации, которые содержат последовательности, кодирующие антигены HCV, находятся в стадии клинических испытаний на людях (Alvarez-Lajonchere L., Dueñas-Carrera S., Int Rev Immunol. 2012; 31 (3): 223-42). В одном случае это вакцина на основе ДНК, экспрессирующей белки NS3-NS5, которую вводят электропорацией (Sällberg M, et al., Expert Opin Biol Ther. 2009; 9 (7): 805-15). В другом случае это вакцинная композиция на основе смеси рекомбинантного белка core и ДНК плазмиды, которая экспрессирует структурные антигены HCV (Castellanos M, et al., J Gene Med. 2010; 12 (1): 107-16). Показано, что оба кандидата безопасны, хорошо переносимы и индуцируют специфические иммунные ответы у иммунизированных субъектов (Alvarez-Lajonchere L., Dueñas-Carrera S., Int Rev Immunol. 2012; 31 (3): 223-42). Ни в одном из этих двух случаев не было продемонстрировано влияние на течение инфекции HCV или устойчивое гистологическое улучшение. ДНК-вакцины, несмотря на их потенциальные преимущества, связанные с их простотой и стабильностью, встречают серьезные возражения со стороны регулирующих органов. Их принципиальное ограничение, судя по всему, связано с их недостаточной иммуногенностью у человека, явлением, до сих пор до конца не изученным, что в значительной степени отличается от результатов, полученных на животных моделях.
Учитывая вышесказанное, разработка профилактической или терапевтической вакцины против HCV является нерешенной проблемой. Настоящее изобретение направлено на достижение именно этой цели.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение решает вышеупомянутую проблему за счет получения химерного антигена для вакцины против вируса гепатита C (HCV), содержащего: a) первый сегмент, соответствующий области E2 (аминокислоты 408-540) полибелка HCV, b) второй сегмент, соответствующий области E1 (аминокислоты 190-222) полибелка HCV, и c) третий сегмент, соответствующий области Core (аминокислоты 1-50) полибелка HCV, в таком порядке.
Новизна изобретения заключается в выборе конкретных эпитопов и порядке, в котором они размещены на полученных вариантах белка. Такая конструкция вакцинного антигена позволяет уменьшить количество компонентов, необходимых для усиления и расширения спектра иммунного ответа против различных антигенов HCV. В настоящем изобретении впервые описан слитый белок, содержащий в одном химерном антигене области полибелка HCV, соответствующие E2 (аминокислоты 408-540), E1 (аминокислоты 190-222) и Core (аминокислоты 1-50), именно в таком порядке. Возникающий в результате иммунный ответ является релевантным и направлен, таким образом, против широкого спектра вирусных антигенов.
Выбор конкретных областей HCV позволяет избегать использования таких областей вирусных белков, которые могут проявлять подавляющие иммунитет эффекты. Точно так же это позволяет избегать использования других областей, которые могут быть иммунодоминантными в сравнении с теми, которые выбраны по настоящему изобретению, и которые при включении в сконструированный антиген ограничивали бы индукцию специфического иммунного ответа против областей, выбранных по настоящему изобретению. С другой стороны, изобретение включает порядок, в котором выбранные области размещены в искусственном белковом антигене, учитывая тот факт, что этот элемент существенно влияет на индукцию иммунного ответа против HCV, вследствие различий в экспозиции и процессинге/представлении эпитопов для иммунной системы. Действительно, в химерных антигенах по настоящему изобретению области из белков core, E1 и E2 размещены в обратном порядке по сравнению с порядком в природном вирусном полибелке.
В варианте осуществления изобретения последовательность химерного вакцинного антигена выбрана из группы, состоящей из SEQ ID No. 10 (антиген Eq1) и SEQ ID No. 16 (антиген Eq1b).
Кроме того, химерные антигены по изобретению могут содержать в своей последовательности по меньшей мере один специфичный для хелперных T-лимфоцитов эпитоп. В варианте осуществления изобретения специфичный для хелперных T-лимфоцитов эпитоп, включенный в химерные антигены, представляет собой один эпитоп из неструктурных белков HCV. В конкретном варианте осуществления неструктурный белок представляет собой NS3. Более конкретно, изобретение относится к химерному вакцинному антигену, характеризующемуся аминокислотной последовательностью, обозначенной SEQ ID No. 14 (антиген EqNS3).
В другом аспекте изобретения специфичный для хелперных T-лимфоцитов эпитоп, включенный в химерные антигены, представляет собой искусственный эпитоп для CD4+ T-лимфоцитов. В контексте изобретения термин «искусственный эпитоп» означает эпитоп, аминокислотная последовательность которого не существует в естественном виде; вместо этого он разработан при помощи биоинформатики. Выбор и включение искусственных эпитопов, которые узнаются хелперными T-лимфоцитами, способствует индукции специфического иммунного ответа против эпитопов HCV, включенных в химерные антигенные варианты. В данном изобретении впервые описаны в качестве искусственных эпитопов для хелперных T-лимфоцитов эпитопы P1M (SEQ ID No. 17) и P2B (SEQ ID No. 18). Эти эпитопы были разработаны для содержания мотива связывания HLA-DR13 и HLA-DR11. Таким образом, изобретение относится к специфичному для хелперных T-лимфоцитов эпитопу, аминокислотная последовательность которого соответствует последовательности SEQ ID No. 17 или SEQ ID No. 18.
В варианте осуществления изобретения химерные антигены содержат аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 12 (антиген NSEq2), SEQ ID No. 13 (антиген EqNSb) и SEQ ID No. 15 (антиген EqP1), и они содержат по меньшей мере один из этих специфичных для хелперных T-лимфоцитов эпитопов.
Следующие химерные антигены имеют характеристики, приведенные ниже: химерный антиген Eq1 (SEQ ID No. 10) содержит области E2 (аминокислоты 408-540), E1 (аминокислоты 190-222) и Core (аминокислоты 1-50) полибелка HCV в этом конкретном порядке. Химерный антиген NSEq2 (SEQ ID No. 12) содержит эпитопы P2B и P1M, в таком порядке, включенные в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). Химерный антиген EqNSb (SEQ ID No. 13) содержит эпитоп P2B, включенный в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). Химерный антиген EqNS3 (SEQ ID No. 14) содержит области аминокислот 1242-1415 полибелка HCV, включенные в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). Химерный антиген EqP1 (SEQ ID No. 15) содержит эпитоп P1M, включенный в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). Химерный антиген Eq1b (SEQ ID No. 16) содержит области E2 (аминокислоты 408-540), E1 (аминокислоты 190-222) и Core (аминокислоты 1-50) полибелка HCV, в этом конкретном порядке, но в этом случае аминокислотная последовательность соответствует варианту H77 HCV генотипа 1a (справочная последовательность NCBI: NC_004102.1).
В варианте осуществления изобретения химерные антигены получены с помощью технологии рекомбинантной ДНК из бактерий, трансформированных плазмидами, описанными в примере 1. Тем не менее, экспертам в данной технологии известно, что такие антигены могут быть получены из других хозяев и могут быть очищены широко известными методами для использования в иммунизации.
В другом аспекте изобретение относится к вакцинной композиции, содержащей химерный антиген для вакцины против HCV, который состоит из a) первого сегмента, состоящего из области E2 (аминокислоты 408-540) полибелка HCV, b) второго сегмента, состоящего из области E1 (аминокислоты 190-222) полибелка HCV, и c) третьего сегмента, состоящего из области Core (аминокислоты 1-50) полибелка HCV, в таком порядке; и фармацевтически приемлемые эксципиенты и/или адъюванты.
В варианте осуществления изобретения вакцинная композиция содержит антигены, обозначенные SEQ ID No. 10 (антиген Eq1), SEQ ID No. 16 (антиген Eq1b), SEQ ID No. 14 (антиген EqNS3), SEQ ID No. 12 (антиген NSEq2), SEQ ID No. 13 (антиген EqNSb) или SEQ ID No. 15 (антиген EqP1).
Для целей изобретения можно использовать широкий спектр фармацевтически приемлемых адъювантов, коммерчески доступных или тех, которые находятся на стадии разработки, для потенцирования иммунного ответа против химерных антигенов, содержащихся в вакцинных композициях, являющихся объектами изобретения.
Вакцинные композиции по изобретению могут также содержать рекомбинантный белковый вариант структурных или NS3 антигенов HCV. Кроме того, вакцинные композиции по изобретению могут содержать плазмиду для ДНК-иммунизации, которая экспрессирует структурные антигены HCV. В другом варианте осуществления изобретения химерные антигены можно формулировать с плазмидой для ДНК-иммунизации, которая одновременно экспрессирует структурные антигены HCV и рекомбинантный капсидный белок HCV.
В другом аспекте изобретение включает, что вакцинную композицию, содержащую химерный антиген для вакцины против HCV, который содержит: a) первый сегмент, состоящий из области E2 (аминокислоты 408-540) полибелка HCV, b) второй сегмент, состоящий из области E1 (аминокислоты 190-222) полибелка HCV, и c) третий сегмент, состоящий из области Core (аминокислоты 1-50) полибелка HCV, в таком порядке, можно вводить по схемам первичного/бустерного введения, наряду с препаратами на основе плазмид для ДНК-иммунизации, рекомбинантных вариантов структурных белков HCV или смеси обоих.
Вакцинные композиции, являющиеся объектами настоящего изобретения, имеют преимущества индукции как гуморального, так и клеточного иммунных ответов против нескольких антигенов HCV, таким образом, они активны против широкого спектра вирусных изолятов и способны создавать защиту в модели суррогатного вирусного заражения.
По изобретению, вакцинные композиции можно вводить внутримышечным, внутрикожным, внутрибрюшинным, подкожным, подслизистым, внутривенным или подъязычным путями введения, или любым другим путем введения, известным специалистам в данной области. С другой стороны, введение можно осуществлять с использованием шприцев, распылителей или любых других устройств для введения.
Объектом изобретения также является использование химерных вакцинных антигенов, которые состоят из первого сегмента, состоящего из области E2 (аминокислоты 408-540) полибелка HCV, второго сегмента, состоящего из области E1 (аминокислоты 190-222) полибелка HCV, и третьего сегмента, состоящего из области Core (аминокислоты 1-50) полибелка HCV, в таком порядке, для изготовления вакцины, предназначенной для индукции иммунного ответа против HCV. В варианте осуществления изобретения вышеуказанная вакцина способна создавать защиту в модели суррогатного вирусного заражения.
С другой стороны, вакцины по изобретению способны индуцировать ответы у здоровых индивидуумов или у инфицированных HCV пациентов. Таким образом, один из аспектов изобретения также относится к способу индукции иммунного ответа против HCV. Данный способ отличается введением химерного антигена, который состоит из первого сегмента, состоящего из области E2 (аминокислоты 408-540) полибелка HCV, второго сегмента, состоящего из области E1 (аминокислоты 190-222) полибелка HCV, и третьего сегмента, состоящего из области Core (аминокислоты 1-50) полибелка HCV, в таком порядке, или вакцинной композиции, содержащей вышеуказанный антиген, здоровому или инфицированному HCV индивидууму.
Изобретение также включает, что в вышеуказанном способе химерный вакцинный антиген или вакцинную композицию, содержащую его, вводят по схемам первичного/бустерного введения, наряду с препаратами на основе плазмид для ДНК-иммунизации, рекомбинантных вариантов структурных белков HCV или смеси обоих.
Для лечения инфицированных HCV пациентов антигены по изобретению или вакцинные композиции, содержащие их, можно вводить одновременно с лекарственными средствами, входящими в стандарт медицинской помощи для этого типа пациентов.
КРАТКОЕ ОПИСАНИЕ ФИГУР
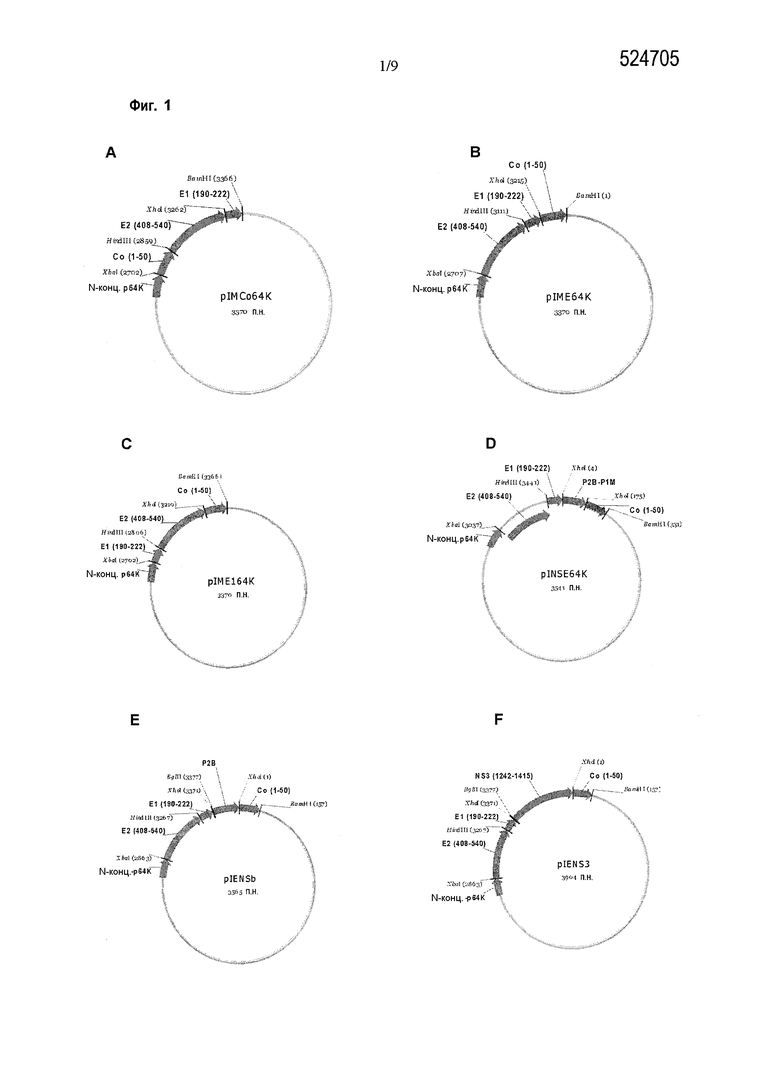
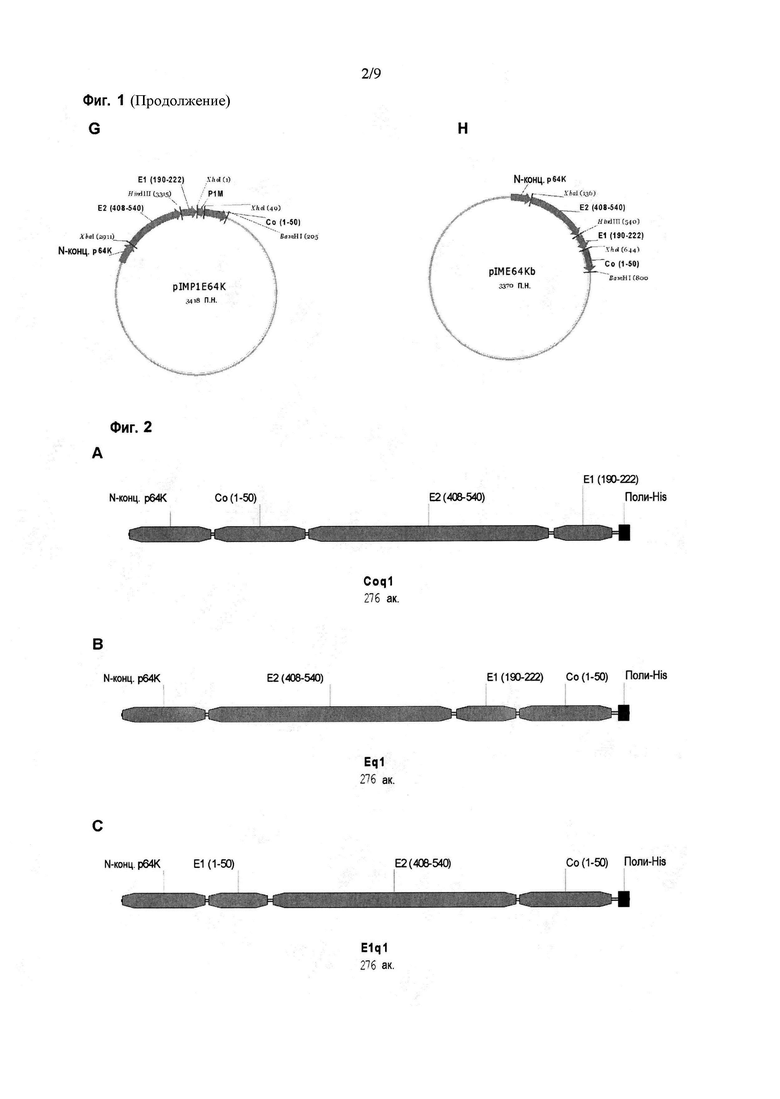
Фигура 1. Карты плазмид, содержащих последовательности, кодирующие различные химерные антигены. A: pIMCo64K, плазмида для экспрессии антигена Coq1. B: pIME64K, плазмида для экспрессии антигена Eq1. C: pIME164K, плазмида для экспрессии антигена E1q1. D: pINSE64K, плазмида для экспрессии антигена NSEq2. E: pIENSb, плазмида для экспрессии антигена EqNSb. F: pIENS3, плазмида для экспрессии антигена EqNS3. G: pIMP1E64K, плазмида для экспрессии антигена EqP1. H: pIME64Kb, плазмида для экспрессии антигена Eq1b.
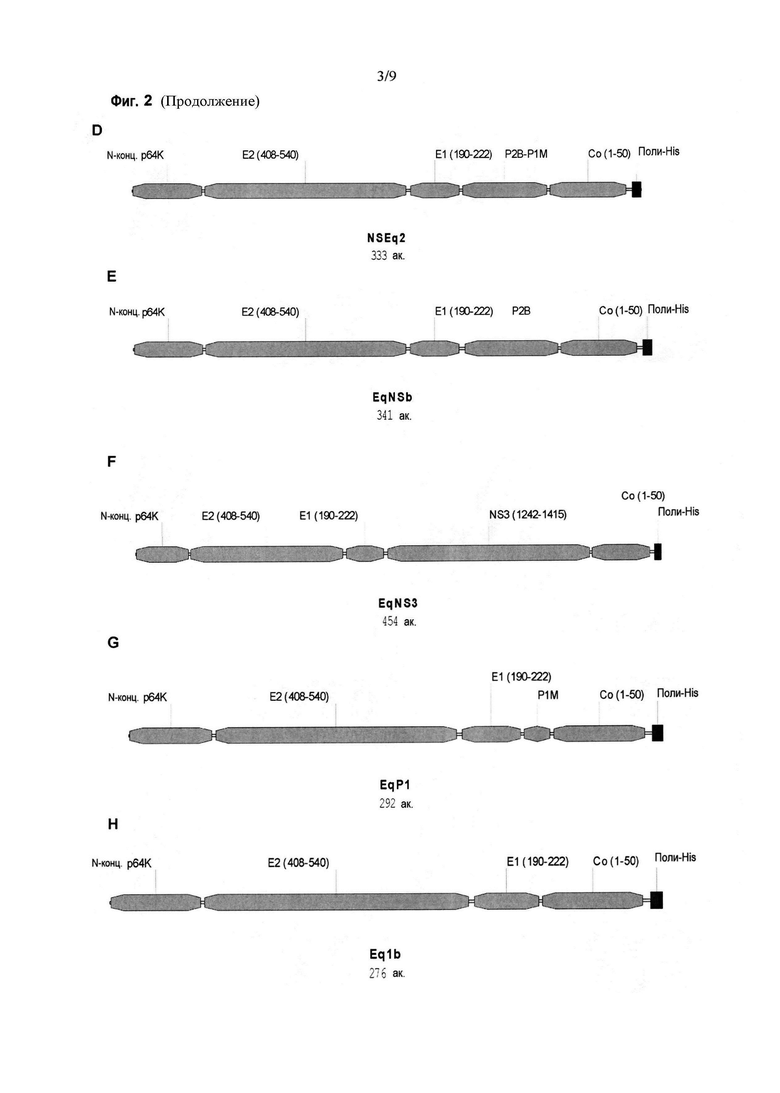
Фигура 2. Схематическое изображение различных химерных антигенов. A: антиген Coq1, содержит области Core (аминокислоты 1-50), E1 (аминокислоты 190-222) и E2 (аминокислоты 408-540) полибелка HCV. B: антиген Eq1, содержит области E2 (аминокислоты 408-540), E1 (аминокислоты 190-222) и Core (аминокислоты 1-50) полибелка HCV. C: антиген E1q1, содержит области E1 (аминокислоты 190-222), E2 (аминокислоты 408-540) и Core (аминокислоты 1-50) полибелка HCV. D: антиген NSEq2, содержит эпитопы P2B и P1M, включенные в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). E: антиген EqNSb, содержит эпитоп P2B, включенный в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). F: антиген EqNS3, содержит область аминокислот 1242-1415 полибелка HCV, включенную между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50) в Eq1. G: антиген EqP1, содержит эпитоп P1M, включенный в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). H: антиген Eq1b, содержит области E2 (аминокислоты 408-540), E1 (аминокислоты 190-222) и Core (аминокислоты 1-50) варианта полибелка H77 HCV генотипа 1a.
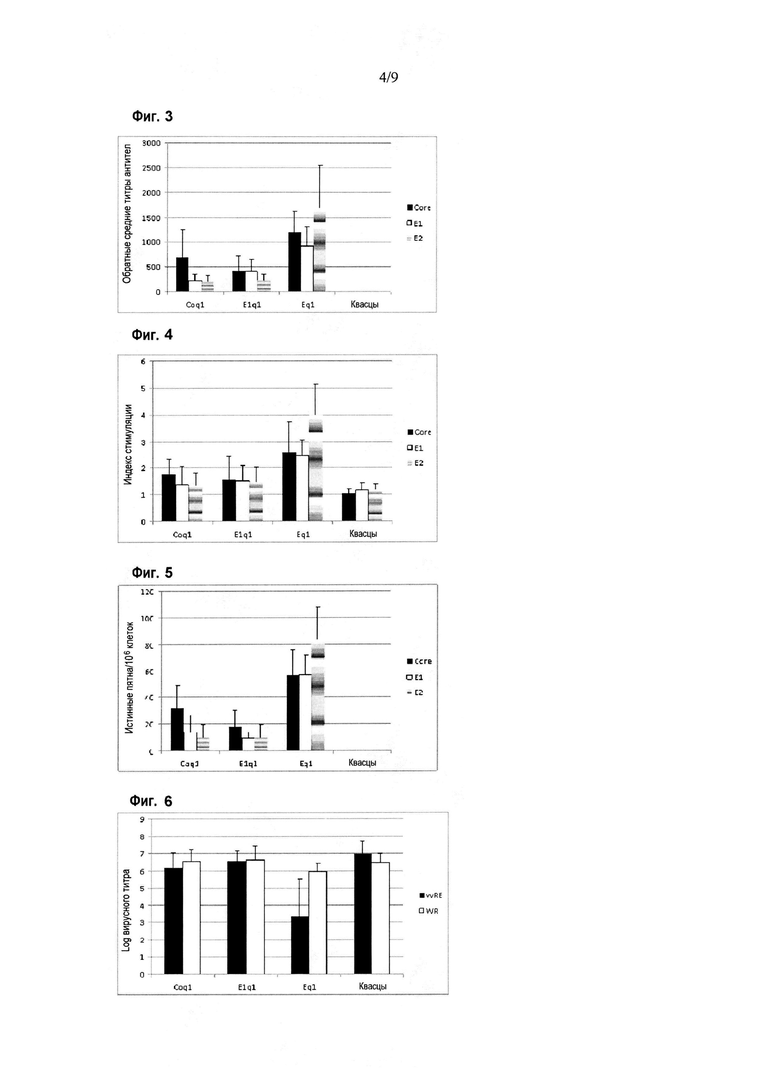
Фигура 3. Ответ в виде продукции антител против белков HCV в схемах иммунизации различными химерными антигенами. Результаты приведены по оси Y в виде величины, обратной титру антител, определяемому как максимальное разведение, при котором сыворотки демонстрируют оптическую плотность при 492 нм, по меньшей мере в два раза превышающую среднюю оптическую плотность сывороток группы отрицательного контроля, при определении методом ELISA. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве покрывающих антигенов в анализе ELISA.
Фигура 4. Пролиферативный ответ против белков HCV в схеме иммунизации различными химерными антигенами. Результаты приведены по оси Y в виде индекса стимуляции, определяемого как отношение количества клеток, пролиферирующих при стимуляции, к количеству клеток, пролиферирующих без стимуляции, при определении в цитометрическом анализе с окрашиванием CFSE. Индекс стимуляции, превышающий два, считается положительным. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 5. Ответ в виде секреции IFN-гамма против белков HCV в схеме иммунизации различными химерными антигенами. По оси Y приведено число истинных пятен на миллион клеток, определяемое как число обнаруженных пятен в условиях стимулирования минус число обнаруженных пятен в условиях без стимулирования, при определении секреции IFN-гамма в анализе ELISPOT. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 6. Ответ на заражение вирусом в схеме иммунизации различными химерными антигенами. Результаты приведены по оси Y в виде логарифма вирусного титра, определяемого как логарифм количества бляшкообразующих единиц на мл, обнаруживаемых в яичниках самок мышей после заражения вирусом. Вирусами осповакцины, используемыми для вирусного заражения, были вирус vvRE, вирус осповакцины, экспрессирующий антигены Core, E1 и E2 (область аминокислот 1-650 в полибелке HCV), и вирус осповакцины WR, не экспрессирующий антигены HCV. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы.
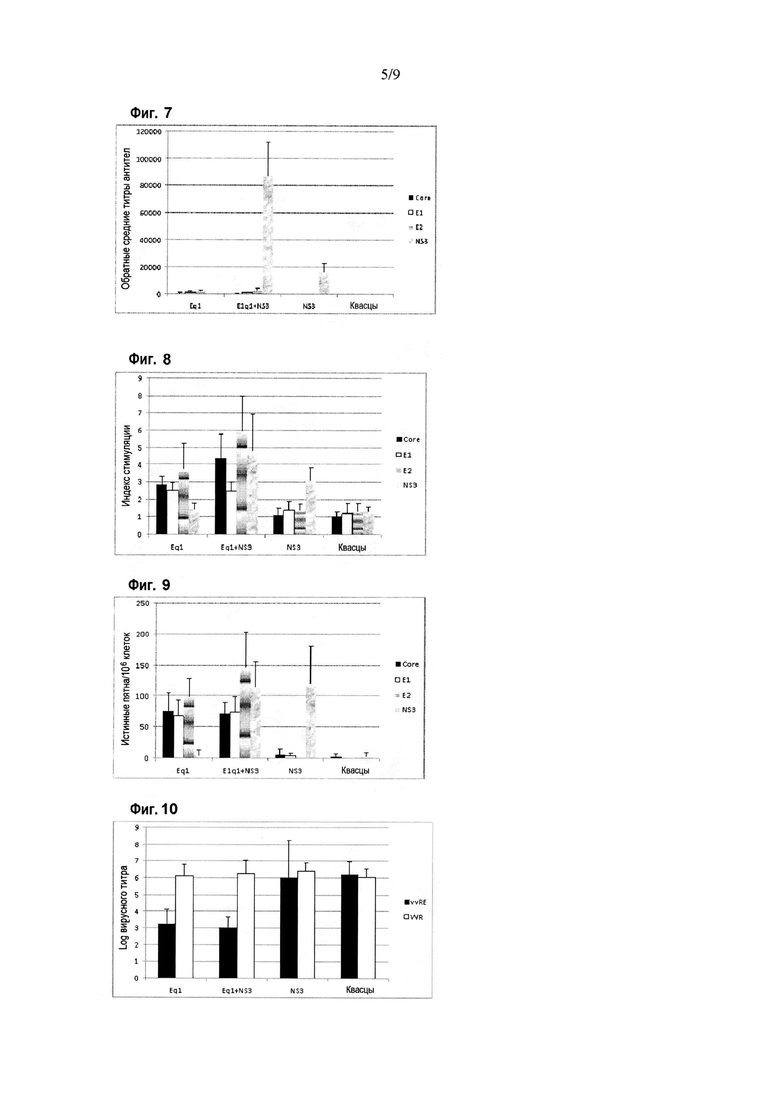
Фигура 7. Ответ в виде продукции антител против белков HCV в схеме иммунизации Eq1 в смеси с NS3. Результаты приведены по оси Y в виде величины, обратной титру антител, при определении методом ELISA. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Ответ оценивали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV), E2 (аминокислоты 384-680 полибелка HCV) и NS3 (аминокислоты 1192-1457 полибелка HCV) в качестве покрывающих антигенов в анализе ELISA.
Фигура 8. Пролиферативный ответ против белков HCV в схеме иммунизации Eq1 в смеси с NS3. Результаты приведены по оси Y в виде индекса стимуляции при определении в цитометрическом анализе с окрашиванием CFSE. Индекс стимуляции, превышающий два, считается положительным. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Ответ оценивали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и NS3 (аминокислоты 1192-1457 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 9. Ответ в виде секреции IFN-гамма против белков HCV в схеме иммунизации Eq1 в смеси с NS3. По оси Y приведено число истинных пятен на миллион клеток при определении секреции IFN-гамма в анализе ELISPOT. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Ответ оценивали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 384-680 полибелка HCV) и NS3 (аминокислоты 1192-1457 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 10. Ответ на заражение вирусом в схеме иммунизации Eq1 в смеси с NS3. Результаты приведены по оси Y в виде логарифма вирусного титра. Вирусами осповакцины, используемыми для вирусного заражения, были vvRE и WR. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы.
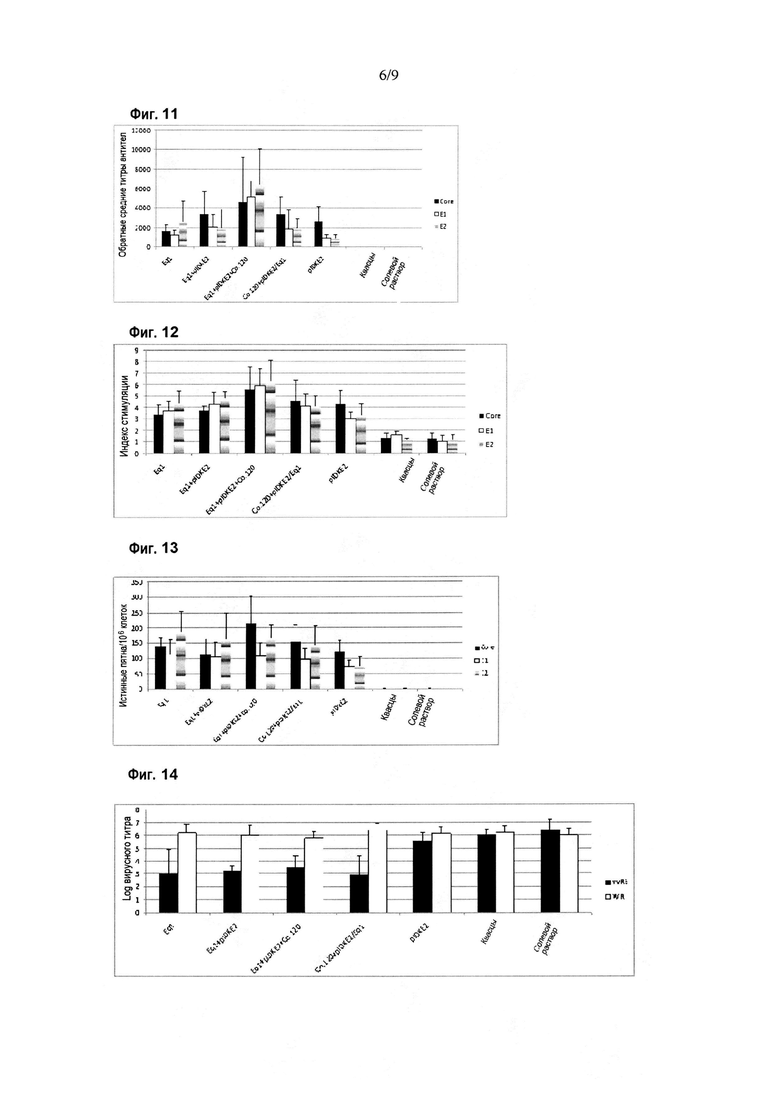
Фигура 11. Ответ в виде продукции антител против белков HCV в схеме иммунизации Eq1 в сочетании с плазмидой для ДНК-иммунизации. Результаты приведены по оси Y в виде величины, обратной титру антител, при определении методом ELISA. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Ответ оценивали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве покрывающих антигенов в анализе ELISA.
Фигура 12. Пролиферативный ответ против белков HCV в схеме иммунизации Eq1 в сочетании с плазмидой для ДНК-иммунизации. Результаты приведены по оси Y в виде индекса стимуляции при определении в цитометрическом анализе с окрашиванием CFSE. Индекс стимуляции, превышающий два, считается положительным. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 13. Ответ в виде секреции IFN-гамма против белков HCV в схеме иммунизации Eq1 в сочетании с плазмидой для ДНК-иммунизации. По оси Y приведено число истинных пятен на миллион клеток при определении секреции IFN-гамма в анализе ELISPOT. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Ответ оценивали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 14. Ответ на заражение вирусом в схеме иммунизации Eq1 в сочетании с плазмидой для ДНК-иммунизации. Результаты приведены по оси Y в виде логарифма вирусного титра, определяемого в яичниках самок мышей после заражения вирусом. Для вирусного заражения использовали вирусы vvRE и WR. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы.
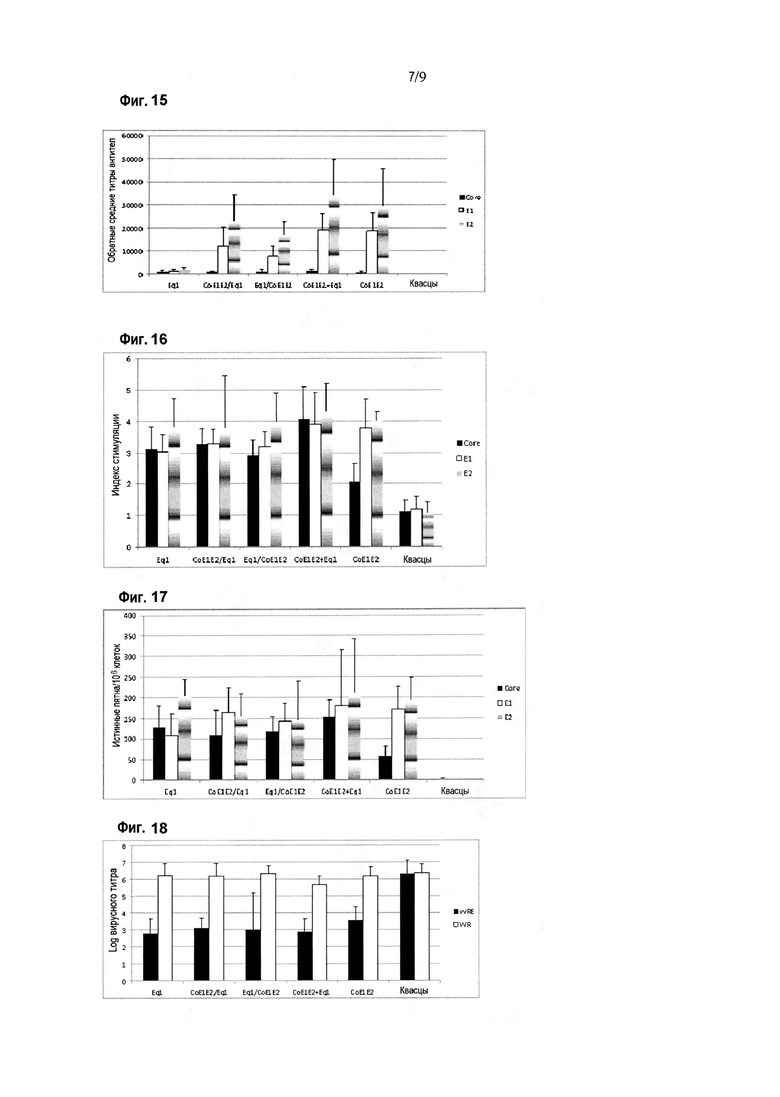
Фигура 15. Ответ в виде продукции антител против белков HCV в схеме иммунизации Eq1 в смеси с рекомбинантными вариантами структурных белков HCV. Результаты приведены по оси Y в виде величины, обратной титру антител, при определении методом ELISA. По оси X показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Ответ оценивали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120 капсидного белка), E1 (192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве покрывающих антигенов в анализе ELISA.
Фигура 16. Пролиферативный ответ против белков HCV в схеме иммунизации Eq1 в смеси с рекомбинантными вариантами структурных белков HCV. Результаты приведены в виде индекса стимуляции при определении в цитометрическом анализе с окрашиванием CFSE. Индекс стимуляции, превышающий два, считается положительным. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 17. Ответ в виде секреции IFN-гамма против белков HCV в схеме иммунизации Eq1 в смеси с рекомбинантными вариантами структурных белков HCV. Результаты приведены в виде числа истинных пятен на миллион клеток при определении в анализе ELISPOT на IFN-гамма. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 18. Ответ на заражение вирусом в схеме иммунизации Eq1 в смеси с рекомбинантными вариантами структурных белков HCV. Результаты приведены в виде логарифма вирусного титра, определяемого в яичниках самок мышей после заражения вирусом. Для вирусного заражения использовали вирусы осповакцины vvRE и WR. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы.
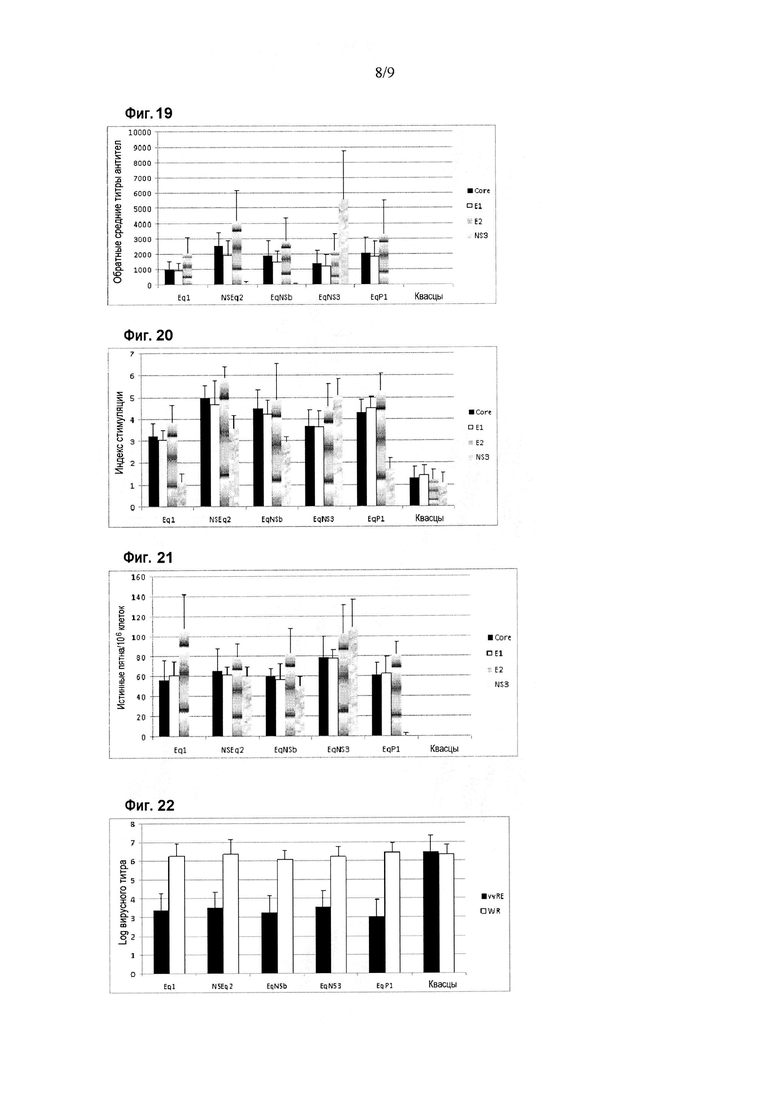
Фигура 19. Ответ в виде продукции антител против белков HCV в схеме иммунизации различными химерными антигенами, содержащими искусственные эпитопы и эпитопы белка NS3. Результаты приведены в виде величины, обратной среднему титру антител, при определении методом ELISA. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV), а также NS3 (аминокислоты 1192-1457 полибелка HCV) в качестве покрывающих антигенов в анализе ELISA.
Фигура 20. Пролиферативный ответ против белков HCV в схеме иммунизации различными химерными антигенами, содержащими искусственные эпитопы и эпитопы белка NS3. Результаты приведены в виде индекса стимуляции при определении в цитометрическом анализе с окрашиванием CFSE. Индекс стимуляции, превышающий два, считается положительным. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV), а также NS3 (аминокислоты 1192-1457 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 21. Ответ в виде секреции IFN-гамма против белков HCV в схеме иммунизации различными химерными антигенами, содержащими искусственные эпитопы и эпитопы белка NS3. Результаты приведены в виде числа истинных пятен на миллион клеток при определении в анализе ELISPOT на IFN-гамма. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV), а также NS3 (аминокислоты 1192-1457 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 22. Ответ на заражение вирусом в схеме иммунизации различными химерными антигенами, содержащими искусственные эпитопы и эпитопы белка NS3. Результаты приведены в виде логарифма вирусного титра, определяемого в яичниках самок мышей после заражения вирусом. Для вирусного заражения использовали вирусы осповакцины vvRE и WR. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы.
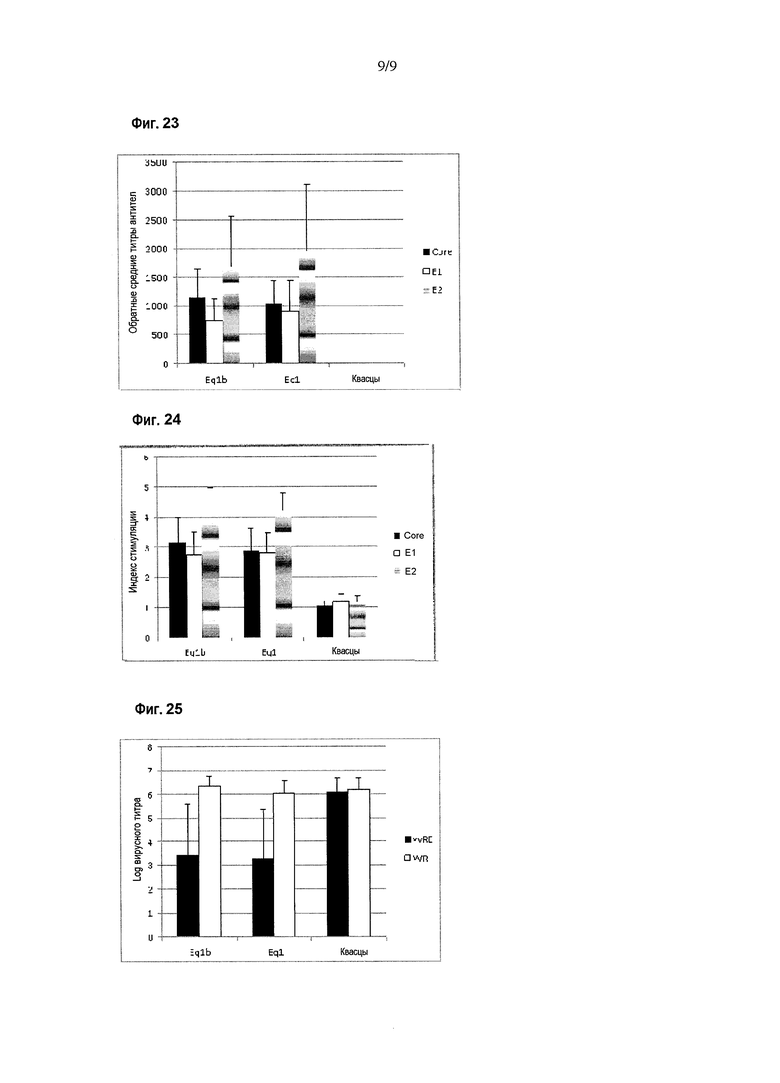
Фигура 23. Ответ в виде продукции антител против белков HCV в схеме иммунизации химерными антигенами Eq1 и Eq1b. Результаты приведены в виде величины, обратной среднему титру антител, при определении методом ELISA. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве покрывающих антигенов в анализе ELISA.
Фигура 24. Пролиферативный ответ против белков HCV в схеме иммунизации химерными антигенами Eq1 и Eq1b. Результаты приведены в виде индекса стимуляции при определении в цитометрическом анализе с окрашиванием CFSE. Индекс стимуляции, превышающий два, считается положительным. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы. Иммунный ответ оценивали с использованием рекомбинантных вариантов Core (аминокислоты 1-120 капсидного белка), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве стимулирующих клетки антигенов.
Фигура 25. Ответ на заражение вирусом в схеме иммунизации химерными антигенами Eq1 и Eq1b. Результаты приведены в виде логарифма вирусного титра, определяемого в яичниках самок мышей после заражения вирусом. Для вирусного заражения использовали вирусы осповакцины vvRE и WR. Показаны различные иммуногены, введенные мышам BALB/c. Планки погрешностей показывают стандартное отклонение для средних значений каждой группы.
ПОДРОБНОЕ ОПИСАНИЕ ЭКСПЕРИМЕНТАЛЬНЫХ МЕТОДОВ/ПРИМЕРЫ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Пример 1. Получение различных химерных антигенов, содержащих эпитопы HCV
Как показано на фигуре 1, были получены плазмиды pIMCo64K (SEQ ID No. 1), pIME64K (SEQ ID No. 2), pIME164K (SEQ ID No. 3), pINSE64K (SEQ ID No. 4), pIENSb (SEQ ID No. 5), pIENS3 (SEQ ID No. 6), pIMP1E64K (SEQ ID No. 7), pIME64Kb (SEQ ID No. 8). Эти плазмиды позволяли экспрессировать в Escherichia coli химерные антигены Coq1 (SEQ ID No. 9), Eq1 (SEQ ID No. 10), E1q1 (SEQ ID No. 11), NSEq2 (SEQ ID No. 12), EqNSb (SEQ ID No. 13), EqNS3 (SEQ ID No. 14), EqP1 (SEQ ID No. 15) и Eq1b (SEQ ID No. 16), соответственно, приведенные на фигуре 2. Во всех случаях, за исключением антигена Eq1b (последовательность которого происходит из штамма H77 HCV генотипа 1a), аминокислотная последовательность происходит из изолята HCV генотипа 1b (González-Horta E.E., Eur Rev Med Pharmacol Sci. 2011; 15 (11): 1320-7).
Как показано на фигуре 2, химерный антиген Coq1 (SEQ ID No. 9) содержит области Core (аминокислоты 1-50), E1 (аминокислоты 190-222) и E2 (аминокислоты 408-540) полибелка HCV, расположенные именно в таком порядке. Химерный антиген Eq1 (SEQ ID No. 10) содержит, точно так же, области E2 (аминокислоты 408-540, E1 (аминокислоты 190-222) и Core (аминокислоты 1-50) полибелка HCV, но именно в этом другом порядке. Химерный антиген E1q1 (SEQ ID No. 11) содержит те же области, но в следующем порядке: E1 (аминокислоты 190-222), E2 (аминокислоты 408-540) и Core (аминокислоты 1-50) полибелка HCV. Химерный антиген NSEq2 (SEQ ID No. 12) содержит эпитопы P2B и P1M, в таком порядке, включенные в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). С другой стороны, химерный антиген EqNSb (SEQ ID No. 13) содержит эпитоп P2B, включенный в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). Химерный антиген EqNS3 (SEQ ID No. 14) содержит область аминокислот 1242-1415 полибелка HCV, включенную между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50) химерного антигена Eq1. Химерный антиген EqP1 (SEQ ID No. 15) содержит эпитоп P1M, включенный в Eq1 между областями E1 (аминокислоты 190-222) и Core (аминокислоты 1-50). Химерный антиген Eq1b (SEQ ID No. 16) содержит области E2 (аминокислоты 408-540), E1 (аминокислоты 190-222) и Core (аминокислоты 1-50) полибелка HCV, именно в таком порядке, но в этом случае аминокислотная последовательность соответствует штамму H77 HCV генотипа 1a (справочная последовательность NCBI: NC_004102.1).
Искусственные эпитопы P1M и P2B, включенные в некоторые из химерных антигенов, были разработаны методами биоинформатики так, чтобы они узнавались хелперными T-лимфоцитами человека. Связывающиеся с HLA-DR13 и HLA-DR11 мотивы были изучены с использованием программ Rankpep, SYFPETHI и ProPred с целью предложения аминокислотных вариантов на каждое положение в искусственных эпитопах в соответствии с частотой появления. В качестве искусственных эпитопов, специфичных для хелперных T-лимфоцитов, описаны P1M, из 14 аминокислот с последовательностью LPEYVIMVKLPSRA (SEQ ID No. 17), и P2B, из 15 аминокислот с последовательностью GYKVIVLNPRVASTL (SEQ ID No. 18).
Для экспрессии рекомбинантных белковых антигенов компетентные клетки бактериального штамма E. coli GC-366 трансформировали соответствующими плазмидами. Экспрессия рекомбинантных белков происходила в течение 12 ч при 37 градусах Цельсия, с использованием минимальной среды для культивирования клеток. Все белковые антигены содержали для белковой экспрессии на N-конце фрагмент, происходящий из белка P64K из Neisseria meningitidis, ранее известный для этой функции (Yero D et al., Biotechnol Appl Biochem. 2006; 44 (Pt 1): 27-34). С другой стороны, белковые варианты содержали на C-конце метку из шести остатков гистидина для облегчения очистки белка. Фактически, белки очищали путем солюбилизации нерастворимой фракции, образующейся в результате разрушения клеток, с использованием карбонат-бикарбонатного буфера, pH 9,6, 8 M мочевины, с последующей металл-хелатной аффинной хроматографией.
Пример 2. Изучение на мышах иммуногенности различных химерных антигенов, содержащих эпитопы HCV
Иммунизировали 8-недельных самок мышей BALB/c массой 16-18 г, по 17 животных на группу. Группы иммунизации были следующими: группа 1, химерный антиген Coq1, сформулированный в квасцах; группа 2, антиген E1q1, сформулированный в квасцах; группа 3, антиген Eq1, сформулированный в квасцах; группа 4, квасцы (контрольная группа). Во всех случаях вводили 20 мкг рекомбинантных антигенов. Иммунизации проводили в недели 0, 2 и 4 путем внутримышечной инъекции.
Кровь собирали в недели 0 и 6 для изучения продукции антител против антигенов HCV. Кроме того, 5 мышей на группу умерщвляли в неделю 6 для изучения специфического клеточного ответа. Дополнительно, 5 животных на группу заражали рекомбинантным вирусом осповакцины vvRE (Alvarez-Lajonchere et al., Biotecnología Aplicada 2007; 24 (3-4): 246-253), экспрессирующим структурные белки HCV, и другие 5 животных заражали контрольным вирусом осповакцины WR в неделю 6. Через пять дней после заражения мышей умерщвляли и вирусный титр определяли в яичниках, как описано ранее (Alvarez-Lajonchere et al., Biotechnol Appl Biochem. 2008; 51 (Pt 2): 97-105).
Специфический иммунный ответ против антигенов HCV показан на фигурах 3-5. Оцениваемый ответ обнаруживали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120, Alvarez-Obregon J.C. et al. Vaccine 2001; 19: 3940-3946), E1 (аминокислоты 192-340 полибелка HCV, Lorenzo L.J. et al., Biotechnol Appl Biochem 2000; 32(2): 137-143) и E2 (аминокислоты 384-680 полибелка HCV, Martinez-Donato et al., Mol Biotechnol. 2007; 35(3): 225-36) в качестве захватывающих антигенов в ELISA или в качестве антигенов для стимуляции в анализах для определения клеточного иммунного ответа. Как показано на фигуре 3, продукция антител против структурных белков HCV была выше в группе животных, иммунизированных Eq1 (p<0,0001; критерии множественного сравнения Крускала-Уоллиса и Данна), для всех оцениваемых антигенов. Аналогичную ситуацию наблюдали для пролиферативного ответа (фигура 4) и для секреции IFN-гамма (фигура 5), соответственно. Кроме того, как показано на фигуре 6, группа животных, иммунизированных белком Eq1, была единственной, где наблюдали способность в значительной степени контролировать специфическую вирусемию (после заражения vvRE) в модели суррогатного заражения (p=0,0069, критерии множественного сравнения Крускала-Уоллиса и Данна).
Эти результаты свидетельствуют о способности химерного антигена Eq1 индуцировать специфический иммунный ответ, как гуморальный, так и клеточный, против нескольких антигенов HCV, с функциональной активностью in vivo, поскольку он способен создавать защиту в модели суррогатного заражения. Кроме того, это свидетельствует, что порядок, в котором выбранные области структурных белков HCV расположены в химерных антигенах, является критическим для индукции специфического иммунного ответа и для развития функционального защитного иммунного ответа в модели заражения, поскольку варианты Coq1 и E1q1 не смогли индуцировать иммунный ответ такого типа. Таким образом, недостаточно просто иметь эпитопы в антигене, но нужно иметь их в правильном контексте.
Пример 3. Изучение на мышах иммуногенности антигена Eq1 в смеси с NS3
Иммунизировали 8-недельных самок мышей BALB/c массой 16-18 г, по 17 животных на группу. Группы иммунизации были следующими: группа 1, антиген Eq1, сформулированный в квасцах; группа 2, антиген Eq1 в смеси с рекомбинантным белком NS3 (Palenzuela D et al., Biotecnología Aplicada 2006; 23: 94-98), сформулированные в квасцах; группа 3, рекомбинантный белок NS3, сформулированный в квасцах; группа 4, квасцы (контрольная группа). Во всех случаях 20 мкг рекомбинантного антигена Eq1 и 10 мкг белка NS3 вводили в соответствующих группах. Иммунизации проводили в недели 0, 2 и 4 путем внутримышечной инъекции.
Кровь собирали в недели 0 и 6 для изучения продукции антител против антигенов HCV. Кроме того, 5 мышей на группу умерщвляли в неделю 6 для изучения специфического клеточного ответа. Дополнительно, 5 животных на группу заражали рекомбинантным вирусом осповакцины vvRE (Alvarez-Lajonchere et al., Biotecnología Aplicada 2007; 24 (3-4): 246-253), экспрессирующим структурные белки HCV, и другие 5 животных заражали контрольным вирусом осповакцины WR в неделю 6. Через пять дней после заражения мышей умерщвляли и вирусный титр определяли в яичниках, как описано ранее.
Специфический иммунный ответ против антигенов HCV показан на фигурах 7-9. Оцениваемый иммунный ответ обнаруживали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120), E1 (аминокислоты 192-340 полибелка HCV), E2 (аминокислоты 384-680 полибелка HCV) и NS3 (аминокислоты 1192-1457 в полибелке HCV) в качестве захватывающих антигенов в ELISA или в качестве антигенов для стимуляции в анализах для определения клеточного иммунного ответа. Как показано на фигуре 7, продукция антител была индуцирована против структурных антигенов HCV в группах животных, иммунизированных Eq1 отдельно или в смеси с NS3, без статистически значимых различий между ними. Однако продукция антител против NS3 была значительно выше в группе смеси NS3 и Eq1 (p=0,0001, критерий Манна-Уитни).
С другой стороны, анализ пролиферативного ответа, представленный на фигуре 8, свидетельствовал о значительно более сильном ответе против антигенов Core, E2 и NS3 в группе животных, которым вводили смесь Eq1 с NS3, чем при введении отдельных антигенов (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса). Статистически значимые различия между группами животных, иммунизированных Eq1 отдельно или в смеси с NS3, не наблюдали для ответа против E1.
Что касается специфического ответа в виде секреции IFN-гамма, который показан на фигуре 9, он был индуцирован со статистически значимыми различиями между вариантами, иммунизированными рекомбинантными белками, только в отношении антигена E2, в случае которого наблюдали значительно более сильный ответ в группе животных, иммунизированных смесью Eq1 и NS3 (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса).
Кроме того, как показано на фигуре 10, в группах животных, иммунизированных как белком Eq1 отдельно, так и в смеси с NS3, наблюдали значительный контроль специфической вирусемии (заражение vvRE) в модели суррогатного заражения (p<0,05, критерии множественного сравнения Крускала-Уоллиса и Данна).
Эти результаты свидетельствовали о том, что препарат на основе смеси антигена Eq1 с NS3 способен индуцировать усиленный специфический иммунный ответ, как гуморальный, так и клеточный, против структурных антигенов HCV и NS3 с функциональной активностью in vivo, поскольку он способен создавать защиту в модели суррогатного заражения.
Пример 4. Изучение на мышах иммуногенности химерного антигена Eq1 в смеси с плазмидой для ДНК-иммунизации
Иммунизировали 8-недельных самок мышей BALB/c массой 16-18 г, по 17 животных на группу. Группы иммунизации были следующими: группа 1, антиген Eq1, сформулированный в квасцах; группа 2, антиген Eq1 в смеси с плазмидой для ДНК-иммунизации pIDKE2 (Dueñas-Carrera et al., Biotechnol Appl Biochem. 2004; 39: 249-55) в солевом растворе; группа 3, антиген Eq1 в смеси с плазмидой pIDKE2 и с белком Co.120 (Dueñas-Carrera et al., Biotecnología Aplicada 1999; 16(4), 226-231) в солевом растворе; группа 4, белок Co.120 в смеси с плазмидой pIDKE2 в солевом растворе в недели 0 и 3, с дозой антигена Eq1, сформулированного в квасцах, в неделю 6; группа 5, плазмида pIDKE2 в солевом растворе в недели 0, 3 и 6; группа 6, квасцы (контроль); группа 7, солевой раствор (контроль). Мыши в соответствующих группах получали 20 мкг химерного антигена Eq1 и 10 мкг рекомбинантного белка Co.120. В случае плазмиды pIDKE2 в каждой дозе вводили 100 мкг. Иммунизации проводили в недели 0, 3 и 6 путем внутримышечной инъекции.
Кровь собирали в недели 0 и 8 для изучения продукции антител против антигенов HCV. Кроме того, 5 мышей на группу умерщвляли в неделю 8 для изучения специфического клеточного ответа. Дополнительно, 5 животных на группу заражали рекомбинантным вирусом осповакцины vvRE, экспрессирующим структурные белки HCV, и другие 5 животных заражали контрольным вирусом осповакцины WR в неделю 8. Через пять дней после заражения мышей умерщвляли и вирусный титр определяли в яичниках, как описано ранее.
Специфический иммунный ответ против антигенов HCV показан на фигурах 11-13. Оцениваемый ответ обнаруживали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве захватывающих антигенов в ELISA или в качестве антигенов для стимуляции в анализах для определения клеточного иммунного ответа. Как показано на фигуре 11, продукция антител против структурных антигенов HCV была индуцирована во всех группах иммунизации, за исключением контроля. Значительно более высокая продукция антител против E1 и E2 была обнаружена в группе животных, иммунизированных смесью белка Eq1 и Co.120 с плазмидой pIDKE2, по сравнению с группой животных, иммунизированных только плазмидой pIDKE2 (p<0,05, критерии множественного сравнения Крускала-Уоллиса и Данна). Точно так же, статистически значимые различия между этими двумя группами наблюдали для пролиферативного ответа (фигура 12) против Core, E1 и E2 (p<0,05, критерии множественного сравнения Крускала-Уоллиса и Данна). Фактически, в группе животных, иммунизированных смесью белка Eq1 и Co.120 с плазмидой pIDKE2, был индуцирован пролиферативный ответ против антигенов E1 и E2, который значительно превосходил тот, который был индуцирован в остальных группах (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса), за исключением группы животных, иммунизированных по схеме первичное/бустерное введение (группа 4).
Что касается специфического ответа в виде секреции IFN-гамма, у животных во всех группах был индуцирован поддающийся обнаружению ответ (фигура 13) против структурных белков HCV (за исключением контроля). Статистически значимые различия в ответах против E1 и E2 не наблюдали между группами животных, вакцинированных иммуногенными вариантами, за исключением наблюдаемого превосходящего ответа (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса), индуцированного против E2, в группе животных, иммунизированных отдельно Eq1 (группа 1), в сравнении с группой животных, иммунизированных только плазмидой pIDKE2 (группа 5). Однако ответ в виде секреции IFN-гамма против антигена Core был значительно выше в группе животных, иммунизированных смесью белков Eq1 и Co.120 с плазмидой pIDKE2, по сравнению с остальными группами (p<0,05; критерии множественного сравнения ANOVA и Ньюмена-Кейлса), за исключением группы животных, иммунизированных по схеме первичное/бустерное введение (группа 4).
Кроме того, все группы животных, которым вводили химерный антиген Eq1 (группы 1-4), отличались способностью в значительной степени контролировать специфическую вирусемию (заражение vvRE) в модели суррогатного заражения (фигура 14) (p<0,05, критерии множественного сравнения Крускала-Уоллиса и Данна), но не остальные группы.
Результаты свидетельствовали о том, что вакцинная композиция на основе Eq1 в смеси с плазмидой pIDKE2 и белком Co.120, или введенная по схеме первичное/бустерное введение, делает возможной индукцию усиленного специфического иммунного ответа, как гуморального, так и клеточного, против структурных антигенов HCV с функциональной активностью in vivo, поскольку она способна создавать защиту в модели суррогатного заражения.
Пример 5. Изучение на мышах иммуногенности химерного антигена Eq1 в смеси с рекомбинантными белковыми вариантами структурных белков HCV
Иммунизировали 8-недельных самок мышей BALB/c массой 16-18 г, по 17 животных на группу. Группы иммунизации были следующими: группа 1, антиген Eq1, сформулированный в квасцах; группа 2, смесь белков Co.120, E1.340 (Lorenzo L.J. et al., Biotechnol Appl Biochem 2000; 32(2): 137-143) и E2.680 (Martinez-Donato et al., Mol Biotechnol. 2007; 35(3): 225-36), сформулированная в квасцах, в недели 0 и 2 с дозами химерного антигена Eq1, сформулированного в квасцах, в неделю 4; группа 3, химерный антиген Eq1, сформулированный в квасцах, в неделю 0 и дозы смеси белков Co.120, E1.340 и E2.680, сформулированной в квасцах, в недели 2 и 4; группа 4, смесь белков Co.120, E1.340, E2.680 и Eq1, сформулированная в квасцах; группа 5, смесь белков Co.120, E1.340 и E2.680, сформулированная в квасцах; группа 6, квасцы (контроль). Мыши в соответствующих группах получали 20 мкг химерного антигена Eq1, 16,7 мкг белков E1 и E2, а также 0,1 мкг белка Co.120. Иммунизации проводили в недели 0, 2 и 4 путем внутримышечной инъекции.
Кровь собирали в недели 0 и 6 для изучения продукции антител против антигенов HCV. Кроме того, 5 мышей на группу умерщвляли в неделю 6 для изучения специфического клеточного ответа. Дополнительно, 5 животных на группу заражали рекомбинантным вирусом осповакцины vvRE, экспрессирующим структурные белки HCV, и другие 5 животных заражали контрольным вирусом осповакцины WR в неделю 8. Через пять дней после заражения мышей умерщвляли и вирусный титр определяли в яичниках, как описано ранее.
Специфический иммунный ответ против HCV показан на фигурах 15-17. Оцениваемый ответ обнаруживали с использованием рекомбинантных вариантов белка Core (аминокислоты 1-120), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве захватывающих антигенов в ELISA или в качестве антигенов для стимуляции в анализах для определения клеточного иммунного ответа. Как показано на фигуре 15, у иммунизированных животных во всех группах, за исключением контроля, была индуцирована продукция специфических антител против структурных антигенов HCV. Никаких статистически значимых различий не наблюдали для продукции антител против белка Core среди групп. Напротив, в группах животных, иммунизированных смесью структурных белков (группа 5), и в группе животных, иммунизированных смесью структурных белков плюс Eq1 (группа 4), наблюдали значительно более высокую продукцию антител против E1 и E2, чем в группах животных, иммунизированных отдельно Eq1 (p<0,05, критерии множественного сравнения Крускала-Уоллиса и Данна).
С другой стороны, пролиферативный ответ против антигенов HCV был индуцирован во всех группах, за исключением контроля, как показано на фигуре 16. Никаких статистически значимых различий между группами не наблюдали с точки зрения пролиферативного ответа против E1 и E2. Однако во всех группах животных, получавших химерный антиген Eq1 в любом из сочетаний (группы 1-4), был индуцирован пролиферативный ответ против Core, значительно превосходящий тот, который был индуцирован смесью структурных белков HCV (группа 5) (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса).
Как показано на фигуре 17, ответ в виде секреции IFN-гамма имел такую же закономерность, которую наблюдали для пролиферативного ответа. В этом случае, в группе животных, иммунизированных смесью структурных белков HCV (группа 5), ответ в виде секреции IFN-гамма против Core был значительно ниже, чем ответ, индуцированный в группах животных, иммунизированных химерным антигеном Eq1 отдельно (группа 1) и смесью Eq1 со структурными белками HCV (группа 4) (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса).
Кроме того, животные во всех группах, за исключением контроля, были способны в значительной степени контролировать специфическую вирусемию (заражение vvRE) в модели суррогатного заражения вирусом (фигура 18) (p<0,05, критерии множественного сравнения Крускала-Уоллиса и Данна).
Результаты свидетельствовали о том, что вакцинная композиция на основе химерного антигена Eq1 в смеси с препаратом, содержащим рекомбинантные белковые варианты структурных антигенов HCV Core, E1 и E2, делает возможной индукцию усиленного специфического иммунного ответа, как гуморального, так и клеточного, против структурных белков HCV с функциональной активностью in vivo, поскольку она способна создавать защиту в модели суррогатного заражения.
Пример 6. Изучение на мышах иммуногенности различных химерных антигенов, содержащих эпитопы HCV и искусственные эпитопы, специфичные для хелперных T-лимфоцитов
Иммунизировали 8-недельных самок мышей BALB/c массой 16-18 г, по 17 животных на группу. Группы иммунизации были следующими: группа 1, химерный антиген Eq1, сформулированный в квасцах; группа 2, химерный антиген NSEq2, сформулированный в квасцах; группа 3, химерный антиген EqNSb, сформулированный в квасцах; группа 4, химерный антиген EqNS3, сформулированный в квасцах; группа 5, химерный антиген EqP1, сформулированный в квасцах; группа 6, квасцы (контроль). Мыши в соответствующих группах получали 20 мкг рекомбинантных химерных антигенов. Иммунизации проводили в недели 0, 2 и 4 путем внутримышечной инъекции.
Кровь собирали в недели 0 и 6 для изучения продукции антител против антигенов HCV. Кроме того, 5 мышей на группу умерщвляли в неделю 6 для изучения специфического клеточного ответа. Дополнительно, 5 животных на группу заражали рекомбинантным вирусом осповакцины vvRE, экспрессирующим структурные белки HCV, и другие 5 животных заражали контрольным вирусом осповакцины WR в неделю 8. Через пять дней после заражения мышей умерщвляли и вирусный титр определяли в яичниках, как описано ранее.
Специфический иммунный ответ против антигенов HCV показан на фигурах 19-21. Оцениваемый ответ обнаруживали с использованием рекомбинантных вариантов белков Core (аминокислоты 1-120), E1 (аминокислоты 192-340 полибелка HCV), E2 (аминокислоты 384-680 полибелка HCV) и NS3 (аминокислоты 1192-1457 в полибелке HCV) в качестве захватывающих антигенов в ELISA или в качестве антигенов для стимуляции в анализах для определения клеточного иммунного ответа. Как показано на фигуре 19, во всех группах, за исключением контроля, у животных была индуцирована продукция специфических антител против структурных белков HCV без статистически значимых различий между группами животных, иммунизированных различными химерными антигенами. Только в группах 2, 3 и 4, для которых использовали области NS3 HCV, была индуцирована продукция антител против этого вирусного антигена, которая была значительно более сильной в группе 4 по сравнению с группами 2 и 3 (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса).
Как показано на фигуре 20, во всех группах, за исключением контроля, у животных был индуцирован специфический пролиферативный ответ против структурных антигенов HCV. В этом случае пролиферативный ответ против антигенов Core и E1 был значительно выше в группах 2, 3 и 5 по сравнению с группой 1 (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса). Никаких статистически значимых различий не было обнаружено среди групп, иммунизированных различными химерными антигенами, с точки зрения пролиферативного ответа против E2. Только в группах 2, 3 и 4, для которых использовали области NS3 HCV, был индуцирован пролиферативный ответ против этого вирусного антигена, который был значительно сильнее в группе Group 4 по сравнению с группами 2 и 3 (p<0,05, критерии множественного сравнения ANOVA и Ньюмена-Кейлса).
Анализ специфического ответа в виде секреции IFN-гамма (фигура 21) показал его индукцию против структурных антигенов HCV во всех группах, за исключением контроля, без статистически значимых различий между группами. Только в группах 2, 3 и 4, для которых использовали области NS3 HCV, был индуцирован ответ в виде секреции IFN-гамма против этого вирусного антигена без статистически значимых различий между группами.
Кроме того, животные во всех группах, за исключением контроля, были способны в значительной степени контролировать специфическую вирусемию (заражение vvRE) в модели суррогатного заражения вирусом (фигура 22) (p<0,05, критерии множественного сравнения Крускала-Уоллиса и Данна).
Результаты свидетельствуют, что включение эпитопов или областей NS3 HCV в последовательность Eq1 делает возможной индукцию специфического иммунного ответа против этого вирусного антигена без оказания влияния на иммунный ответ, индуцированный против структурных антигенов HCV. Более того, включение искусственных эпитопов P1M и P2B в последовательность белка Eq1 способствует значительному возрастанию пролиферативного ответа против антигенов Core и E1 без оказания влияния на способность индуцировать специфический гуморальный или клеточный иммунный ответ против этих антигенов или E2, и при этом также сохраняется функциональная активность in vivo, поскольку это может создавать защиту в модели суррогатного заражения вирусом.
Пример 7. Сравнительное изучение на мышах иммуногенности химерных антигенов Eq1 и Eq1b
Иммунизировали 8-недельных самок мышей BALB/c массой 16-18 г, по 17 животных на группу. Группы иммунизации были следующими: группа 1, химерный антиген Eq1b, сформулированный в квасцах; группа 2, химерный антиген Eq1, сформулированный в квасцах; группа 3, квасцы (контроль). Мыши в соответствующих группах получали 20 мкг химерных антигенов. Иммунизации проводили в недели 0, 2 и 4 путем внутримышечной инъекции.
Кровь собирали в недели 0 и 6 для изучения продукции антител против антигенов HCV. Кроме того, 5 мышей на группу умерщвляли в неделю 6 для изучения специфического клеточного ответа. Дополнительно, 5 животных на группу заражали рекомбинантным вирусом осповакцины vvRE, экспрессирующим структурные белки HCV, и другие 5 животных заражали контрольным вирусом осповакцины WR в неделю 8. Через пять дней после заражения мышей умерщвляли и вирусный титр определяли в яичниках, как описано ранее.
Специфический иммунный ответ против антигенов HCV показан на фигурах 23 и 24. Оцениваемый ответ обнаруживали с использованием рекомбинантных вариантов белков Core (аминокислоты 1-120), E1 (аминокислоты 192-340 полибелка HCV) и E2 (аминокислоты 384-680 полибелка HCV) в качестве захватывающих антигенов в ELISA или в качестве антигенов для стимуляции в анализах для определения клеточного иммунного ответа. Как показано на вышеупомянутых фигурах, в группах животных, иммунизированных белками, но не в контроле, были индуцированы специфические ответы в виде продукции антител и пролиферативного ответа против структурных антигенов HCV без статистически значимых различий между группами животных, иммунизированных рекомбинантными белками.
С другой стороны, группы животных, иммунизированных химерными антигенами, но не контроль, проявляли способность в значительной степени контролировать специфическую вирусемию (заражение vvRE) в модели суррогатного заражения вирусом (фигура 25) (p<0,05; критерии множественного сравнения Крускала-Уоллиса и Данна) без различий между группами животных, иммунизированных рекомбинантными белками.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИННЫЙ СОСТАВ, ПОТЕНЦИРОВАННЫЙ КОМБИНАЦИЕЙ ДНК И АНТИГЕНА | 2002 |
|
RU2294212C2 |
| ОЛИГОМЕРНАЯ ЧАСТИЦА, ИНДУЦИРУЮЩАЯ ИММУНИТЕТ ПРОТИВ ВИРУСА ГЕПАТИТА С, СПОСОБ ПОЛУЧЕНИЯ ОЛИГОМЕРНОЙ ЧАСТИЦЫ, КОМПОЗИЦИЯ, СПЕЦИФИЧЕСКОЕ АНТИТЕЛО, НАБОР (ВАРИАНТЫ), ИММУНОЛОГИЧЕСКИЙ АНАЛИЗ И ВАКЦИНА ПРОТИВ ВИРУСА ГЕПАТИТА С | 1999 |
|
RU2247729C2 |
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ НА ОСНОВЕ "ЖИВОГО" ВИРУСА ОСПЫ ДОМАШНЕЙ ПТИЦЫ И ИХ ПРИМЕНЕНИЕ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2353651C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2351363C2 |
| ИММУНОСТИМУЛИРУЮЩАЯ КОМБИНАЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ГЕПАТИТА С | 2006 |
|
RU2431499C2 |
| АГЕНТЫ И СПОСОБЫ, ОСНОВАННЫЕ НА ПРИМЕНЕНИИ ДОМЕНА EDA ФИБРОНЕКТИНА | 2006 |
|
RU2430738C2 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛЯЦИИ ИММУННОГО ОТВЕТА (ВАРИАНТЫ), СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ, ПРИМЕНЕНИЯ И СПОСОБЫ СТИМУЛЯЦИИ ИММУННОГО ОТВЕТА С ИСПОЛЬЗОВАНИЕМ ЭТИХ КОМПОЗИЦИЙ | 2002 |
|
RU2316347C2 |
| ВАКЦИНА ПРОТИВ HCV | 2003 |
|
RU2323744C2 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ КАТИОННЫЕ МИКРОЧАСТИЦЫ И ДНК HCV Е1Е2, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2364419C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ДЛЯ БОРЬБЫ С ЗАРАЖЕНИЯМИ ЭКТОПАРАЗИТАМИ | 2011 |
|
RU2585226C2 |




















Представленные изобретения касаются химерного антигена, вакцинной композиции, содержащей такой химерный антиген, и применения химерного антигена для получения вакцин против вируса гепатита C (HCV). Охарактеризованный химерный анетиген состоит из а) первого сегмента, состоящего из области Е2 (аминокислоты 408-540) полибелка HCV, b) второго сегмента, состоящего из области Е1 (аминокислоты 190-222) полибелка HCV, и с) третьего сегмента, состоящего из области Core (аминоксилоты 1-50) этого белка, в таком порядке. Изобретения обеспечивают сильный иммунный ответ широкого спектра против различных антигенов вируса и могут быть использованы в медицине и фармацевтической промышленности, а также для профилактического и/или терапевтического применения против HCV. 3 н. и 11 з.п. ф-лы, 25 ил., 7 пр.
1. Химерный антиген для вакцины против вируса гепатита С (HCV), состоящий из а) первого сегмента, состоящего из области Е2 (аминокислоты 408-540) полибелка HCV, b) второго сегмента, состоящего из области Е1 (аминокислоты 190-222) полибелка HCV, и с) третьего сегмента, состоящего из области Core (аминокислоты 1-50) этого белка, в таком порядке.
2. Химерный вакцинный антиген по п. 1, в котором его аминокислотная последовательность выбрана из группы, состоящей из SEQ ID No. 10 (антиген Eq1) и SEQ ID No. 16 (антиген Eq1b).
3. Вакцинный химерный антиген по п. 1, связанный с неструктурными белками HCV NS3 и состоящий из SEQ ID No. 14 (антиген EqNS3).
4. Химерный вакцинный антиген по п. 1, связанный с искусственным эпитопом SEQ ID No. 17 (Р1М) или SEQ ID No. 18 (P2B).
5. Химерный вакцинный антиген по п. 4, в котором его аминокислотная последовательность выбрана из группы, состоящей из SEQ ID No. 12 (антиген NSEq2), SEQ ID No. 13 (антиген EqNSb) и SEQ ID No. 15 (антиген EqP1).
6. Вакцинная композиция, содержащая 20 мкг химерного антигена для вакцины против HCV, который содержит: а) первый сегмент, состоящий из области Е2 (аминокислоты 408-540) полибелка HCV, b) второй сегмент, состоящий из области Е1 (аминокислоты 190-222) полибелка HCV, и с) третий сегмент, состоящий из области Core (аминокислоты 1-50) полибелка HCV, именно в таком порядке; и эксципиенты и/или фармацевтически приемлемые адъюванты, для индукции специфического иммунного ответа против HCV.
7. Вакцинная композиция по п. 6, содержащая химерный вакцинный антиген по любому из пп. 2-5.
8. Вакцинная композиция по п. 6 или 7, дополнительно содержащая плазмиду для ДНК-иммунизации, экспрессирующую структурные антигены HCV.
9. Вакцинная композиция по п. 6 или 7, дополнительно содержащая плазмиду для ДНК-иммунизации, экспрессирующую структурные антигены HCV и рекомбинантный капсидный белок HCV.
10. Вакцинная композиция по п. 6 или 7, которую можно вводить по схемам первичного/бустерного введения с препаратами на основе плазмид для ДНК-иммунизации, рекомбинантных белков структурных антигенов HCV или смеси обоих.
11. Применение химерных вакцинных антигенов по любому из пп. 1-5 для получения вакцины, предназначенной для индукции специфического иммунного ответа против HCV.
12. Применение по п. 11, в котором такая вакцина способна создавать защиту in vivo в модели суррогатного заражения вирусом.
13. Применение по п. 11, в котором такая вакцина способна индуцировать иммунный ответ у здоровых индивидуумов или у пациентов, инфицированных HCV.
14. Применение по п. 11, в котором такую вакцину вводят по схемам первичного/бустерного введения с препаратами на основе плазмид для ДНК-иммунизации, рекомбинантных белков структурных антигенов HCV или смеси обоих.
| ARASH ARASHKIA et al | |||
| Construction of HCV-polytope vaccine candidates harbouring immune-enhancer sequences and primary evaluation of their immunogenicity in BALB/c mice, Virus Genes, 2010, No | |||
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| Приспособление для плетения проволочного каркаса для железобетонных пустотелых камней | 1920 |
|
SU44A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| HUI LI et al | |||
| Genetic Diversity of Near Genome-Wide Hepatitis C Virus Sequences during Chronic Infection: Evidence for Protein Structural Conservation Over Time, PLoS ONE, 2011, Vol | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| А.В | |||
| ИВАНОВ и др | |||
| МОЛЕКУЛЯРНАЯ БИОЛОГИЯ ВИРУСА ГЕПАТИТА С | |||
| Успехи биологической химии, 2005, т | |||
| Железобетонный фасонный камень для кладки стен | 1920 |
|
SU45A1 |
| Пишущая машина | 1922 |
|
SU37A1 |
| Устройство для усиления микрофонного тока с применением самоиндукции | 1920 |
|
SU42A1 |

