ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на открытии того, что белки оболочки HCV индуцируют благотворный иммунный ответ у шимпанзе, которые хронически инфицированы гетерологичным штаммом HCV подтипа 1а или подтипа 1b. Более конкретно настоящее изобретение относится к открытию того, что белки оболочки являются высоко иммуногенными и приводят к стимуляции как клеточного, так и гуморального иммунного ответа. Кроме того, настоящее изобретение относится к открытию того, что блокирование цистеинов алкилированием имеет результатом даже более иммуногенные белки. Кроме того, белки оболочки по настоящему изобретению можно включать в частицы, которые проявляют высокую иммуногенность и иммунную реактивность. Кроме того, было продемонстрировано, что такие частицы могут включать в себя другие белки.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Инфекция вирусом гепатита С (HCV) является наиболее важной проблемой здравоохранения как в развитых, так и в развивающихся странах. Подсчитано, что примерно от 1 до 5% населения мира поражено этим вирусом. Оказалось, что HCV инфекция является наиболее важной причиной гепатита, связанного с переливанием крови, и часто прогрессирует до хронического поражения печени. Кроме того, имеется свидетельство вовлечения HCV в индукцию печеночно-клеточного рака. Следовательно, высока потребность в надежных диагностических способах и эффективных терапевтических агентах. Необходимы также чувствительные и специфические способы скрининга зараженных HCV продуктов крови и усовершенствованные способы культивирования HCV.
HCV представляет собой вирус с однонитевой позитивной РНК из примерно 9600 оснований, которая кодирует по меньшей мере три структурных белка и шесть неструктурных белков. На основании гомологии последовательности эти структурные белки функционально определили как один отдельный ядерный белок и два белка оболочки: Е1 и Е2. Белок Е1 состоит из 192 аминокислот и содержит от 5 до 6 сайтов N-гликозилирования в зависимости от генотипа HCV. Белок Е2 состоит из 363-370 аминокислот и содержит 9-11 сайтов N-гликозилирования в зависимости от генотипа HCV (в качестве обзоров смотри: Major and Feinstone, 1997; Maertens and Stuyver, 1997). Белок Е1 содержит различные вариабельные домены (Maertens and Stuyver, 1997), тогда как белок Е2 содержит три гипервариабельных домена, из которых главный домен локализован на N-конце этого белка (Maertens and Stuyver, 1997). Последние белки оболочки получены с помощью рекомбинантных методик в Escherichia соli, клетках насекомых, дрожжевых клетках и клетках млекопитающих. Использование системы экспрессии в культуре клеток высших эукариот и особенно в культуре клеток млекопитающих имеет результатом белки оболочки, которые эффективно распознаются антителами в образцах от пациентов (Maertens et al., 1994).
Было выдвинуто предположение, что белок оболочки Е1 нуждается в белке оболочки Е2, чтобы достичь состояния правильной укладки субъединиц (Deleersnyder et al., 1997). Кроме того, предположили, что Е1 и Е2 образуют гетеродимеры, которые могут образовать основную единицу вирусной оболочки (Yi et al., 1997). В WO 98/21338 Liang et al. использовали эти предположения, чтобы сконструировать частицы HCV, которые состоят из Е1 и Е2, а также из Core и Р7. Иными словами, в уровне техники не было предложено использовать Е1 или Е2 по отдельности для иммунизации и других целей. Однако Houghton (1997) сообщал, что повторные иммунизации рекомбинантным gрЕ1Е2 (4×25 мкг) 3 хронически инфицированных HCV шимпанзе не индуцировали значимого иммунного ответа. Авторы настоящей заявки убедились, что индукция иммунного ответа против оболочки у пациентов с хроническим гепатитом С действительно будет желательной и благотворной для пациента, поскольку высокие уровни таких антител, по-видимому, коррелируют с хорошим ответом на терапию интерфероном и могут, следовательно, способствовать выведению вируса у пациента (РСТ/ЕР 95/03031, Maertens et al.). Авторы настоящего изобретения, кроме того, убедились, что, поскольку уровни антител против Е1 у хронических носителей HCV находятся в числе низших среди всех HCV антител, то поднятие этих уровней антител и, возможно, клеточного ответа может быть благотворным для индукции борьбы с этой инфекцией или даже клиренса этой инфекции у хозяина. По-видимому, высокие уровни клеточного иммунитета против Е1 также коррелируют с хорошим ответом на терапию интерфероном (Leroux-Roels et al., 1996).
Помимо важности иммунитета против Е1 в отношении терапии интерфероном, другие показатели указывают на то, что некоторые другие участки генома HCV могут быть значимыми для индукции специфического иммунного ответа, который может дать возможность бороться с этой инфекцией. У пациентов, отвечающих на терапию интерфероном, также чаще наблюдали реактивность Т-клеток против С-концевого района ядерного белка (Leroux-Roels et al., 1996). Потенциально нейтрализующие антитела против белка NS4B были продемонстрированы у пациентов, выводящих HCV после трансплантации печени (Villa et al., 1998). В пределах NS3 картировали также несколько Т-клеточных эпитопов, которые, по-видимому, коррелируют с выведением HCV в период острой фазы (смотри: РСТ/ЕР 94/03555, Leroux-Roels et al.; Leroux-Roels et al., 1996; Rehermann et al., 1996 и 1997; Diepolder et al., 1995 и 1997). Кроме того, антитела к NS5A, подобно антителам к Е1, демонстрируют высокие уровни на базовом уровне перед терапией интерфероном-альфа в долговременных отвечающих организмах (long term responders, LTR) по сравнению с неотвечающими организмами.
В настоящее время терапевтическая вакцинация против HCV не является успешной. Показано также, что профилактическая вакцинация эффективна только против гомологичного штамма HCV (Сhоо et al., 1994). Настоящее изобретение относится к удивительному открытию того, что введение антигена оболочки HCV может кардинально улучшить состояние при хроническом активном гепатите у индивидуума, инфицированного гетерологичным штаммом или изолятом, как при инфекции гетерологичным субтипом 1а, так и при инфекции гетерологичным субтипом 1b. Действительно, хронически инфицированные шимпанзе, которым вводили шесть доз по 50 мкг E1s (то есть аминокислоты 192-326 из Е1), неожиданно продемонстрировали сильный гуморальный и клеточный иммунные ответы, которые не повышались в течение всего периода хронической инфекции до последней вакцинации. Более того, вирусный антиген становился не обнаружимым в печени в течение периода от двух до пяти месяцев и оставался не обнаружимым в течение по меньшей мере 5 месяцев после вакцинации. Хотя титры HCV РНК в печени не снижались, уровни ферментов печени в сыворотке показывали явную тенденцию к нормализации. Наиболее важно, что при обеих вакцинах кардинально улучшалась гистология печени. Настоящее изобретение далее относится к удивительному открытию того, что белок Е1, используемый для вакцинации, который экспрессировали в виде отдельного HCV белка без его гидрофобного якоря, образует стабильные частицы. Следует также отметить, что для того, чтобы избежать индукции иммунного ответа против ненужных эпитопов, белок Е1, используемый для вакцинации, конструировали в виде согласованной последовательности индивидуальных клонов, имеющих происхождение от одного образца сыворотки от одного хронического носителя. Кроме того, настоящая заявка относится к открытию того, что индукцию таких ответов против Е1 можно усилить с помощью использования антигенов иного генотипа, чем генотипы инфекции, присутствующей у хозяина. Кроме того, настоящая заявка относится к открытию того, что, когда цистеины белков оболочки HCV алкилированы, например, N-(иодэтил)-трифторацетамидом, этиленимином или активными галогенами, такими как иодацетамид, олигомерные частицы, как описано выше, демонстрируют даже более высокую иммуногенность. Наконец, настоящее изобретение относится также к открытию того, что мутация цистеинов белков оболочки HCV в любую другую природную аминокислоту, предпочтительно метионин, глутаминовую кислоту, глутамин или лизин, в олигомерных частицах, как описано выше, также приводит к более высокой иммуногенности по сравнению с исходными белками оболочки.
ЗАДАЧА ИЗОБРЕТЕНИЯ
Из литературы ясно, что существует настоятельная необходимость в разработке надежных вакцин и эффективных терапевтических агентов для HCV. Следовательно, задачей настоящего изобретения является разработка антигенного препарата, который способен индуцировать специфический гуморальный и клеточный иммунитет к белкам оболочки HCV даже (но не исключительно) у хронических HCV носителей. Те же самые антигены можно применять для диагностики иммунного ответа.
Более конкретно задачей настоящего изобретения является разработка антигенного препарата, как определено выше, который состоит из стабильных частиц отдельных белков оболочки HCV. Должно быть ясно, что в настоящее время такие частицы или способ получения таких частиц в данной области техники неизвестны. Кроме того, в уровне техники отсутствуют указания на то, что какой-либо антигенный препарат, включающий в себя такие стабильные частицы или такие очищенные отдельные белки оболочки HCV, можно успешно применять в качестве (гетерологичной) профилактической или терапевтической вакцины против HCV. Таким образом, задачей настоящего изобретения также является разработка способа получения стабильных частиц, которые можно успешно применять в качестве профилактического или терапевтического агента против HCV, и, кроме того, разработка ДНК вакцин, кодирующих HCV антигены. Более конкретно задачей настоящего изобретения является разработка способа получения последних частиц на основе образования частиц в присутствии детергента (смотри далее). Кроме того, задачей настоящего изобретения является разработка способов получения частиц, состоящих из антигенов, полученных от различных HCV генотипов.
Кроме того, задачей настоящего изобретения является разработка антигена, который представляет собой согласованную последовательность из индивидуальных клонов, которая может обеспечить возможность более правильной укладки этих белков. Это делается, чтобы избежать стимуляции иммунитета против ненужных эпитопов.
Кроме того, задачей настоящего изобретения является разработка антигенного препарата, в частности, для терапевтической вакцинации, на основе генотипа HCV, которым инфицирован хронический носитель. В этом отношении задачей настоящего изобретения является разработка белка оболочки либо гомологичного, либо отличного генотипа или подтипа по сравнению с генотипом или субтипом этого хронического носителя.
Следующей задачей настоящего изобретения является разработка способа лечения или терапевтической вакцинации хронически инфицированных пациентов при использовании указанных выше антигенов или ДНК вакцин, возможно в сочетании с другими соединениями. Задачей настоящего изобретения также является разработка способа профилактической вакцинации людей против HCV.
Другой задачей настоящего изобретения является разработка олигомерных частиц, которые обладают лучшей иммуногенностью вследствие мутации по меньшей мере одного цистеинового остатка белка оболочки HCV в природную аминокислоту, предпочтительно метионин, глутаминовую кислоту, глутамин или лизин. Альтернативно можно осуществлять алкилирование по меньшей мере одного цистеинового остатка белка оболочки HCV. В частности, последний белок можно алкилировать с помощью этиленимина, N-(иодэтил)трифторацетамида или активных галогенов. В этом отношении задачей настоящего изобретения является разработка дополнительного применения олигомерных частиц в качестве носителей для эффективного представления не-HCV эпитопов.
В задачу настоящего изобретения также входила разработка способа лечения остро или хронически инфицированных пациентов антителом против оболочки, таким как антитело против Е1, например антителом против V2 района Е1, либо отдельно, либо в сочетании с другими терапиями.
В задачу настоящего изобретения также входила разработка антигена, стимулирующего Т-клетки, такого как Core, Е1, Е2, Р7, NS2, NS3, NS4A, NS4B, NS5A или NS5B, вместе с белками оболочки по изобретению.
Считается, что все аспекты задачи настоящего изобретения встречаются в воплощениях, как указано ниже.
КРАТКОЕ ОПИСАНИЕ ТАБЛИЦ И ГРАФИЧЕСКИХ МАТЕРИАЛОВ
В таблице 1 представлены последовательности клонов Е1, полученных от одного хронического носителя, конструкция Е1, использованная для продуцирования вакцины, представляет собой согласованную последовательность всех этих индивидуальных клонов. V1-V5, вариабельные районы 1-5; С4, константный домен 4; HR, гидрофобный район; HCV-B con, согласованная последовательность в положениях, которые являются вариабельными между клонами и HCV-J.
В таблице 2 представлены последовательности белка Е1 вакцины и белка Е1, как он обнаружен у инфицированных шимпанзе Phil и Ton. Изолят субтипа 1b от Phil отличается на 5,92% от вакцинного штамма. Различие между вакциной и изолятом субтипа 1а от Ton составило 20,74%.
В таблице 3 представлен схематический обзор изменений, индуцированных терапевтической вакцинацией у двух хронически инфицированных шимпанзе (Ton и Phil). Анализ проводили, как описано для Фиг.8 и 11. Кроме того, оценивали гистологию и воспаление на основании биопсий печени.
В таблице 4 представлены последовательности пептидов, использованных для картирования эпитопов В-клеток. Отметим, что HR перекрывается с V4V5.
В таблице 5 показана перестройка NS3 для получения более короткого белка, несущего все главные эпитопы, коррелирующие с клиренсом вируса.
В таблице 6 показана реактивность в LIA (иммунолюминесцентный анализ) E1s-ацетамида против E1s-малеимида с сыворотками хронических носителей HCV. Белки иммобилизовали на LIA мембранах. E1s-ацетамид сам по себе распыляли на полосках LIA, тогда как E1s-малеимид (также содержащий биотин-малеимид) перед распылением подвергали комплексованию со стрептавидином. Антигены связывали с LIA-мембранами и полоски обрабатывали по существу, как описано в Zrein et al. (1998). Человеческие антитела, направленные против этих антигенов, визуализировали, используя человеческий анти-IgG, конъюгированный с щелочной фосфатазой. NBT (нитросиний тетразолий) и BCIP (5-бром-4-хлор-3-индолилфосфат) использовали для цветового проявления полоски. Окрашивание оценивали от 0,5 до 4, как указано в Zrein et al. (1998). Число (#поз.) и процент (%поз.) позитивных образцов указаны в нижней части таблицы с использованием отсечки для данного анализа, равной 0,5.
Фиг.1. Наложенные друг на друга профили гель-хроматографии в 3ФР (забуференный фосфатом физиологический раствор)/3% Empigen-BB белков E1s и E2s, экспрессированных и очищенных согласно Maertens et al. (РСТ/ЕР95/03031).
Фиг.2. Наложенные друг на друга профили гель-хроматографии (size exclusion chromatography, SEC) белков E1s и E2s, экспрессированных и очищенных согласно Maertens et al. (PCT/EP95/03031) и подвергнутых второму пробегу на той же SEC колонке в ЗФР/0,2% CHAPS (3-[(3-холамидопропил)диметил-аммонио]-1-пропансульфонат) для получения конкретных олигомерных структур с установленной примерной молекулярной массой порядка 250-300 кДа. Подобные степени связывания можно получить с помощью использования 3%-ного бетаина.
Фиг.3. Наложенные друг на друга профили гель-хроматографии белков E1s и E2s, экспрессированных и очищенных согласно Maertens et al. (PCT/EP95/03031), подвергнутых второму пробегу в 0,2%-ном CHAPS или в 3%-ном бетаине, чтобы получить конкретные олигомерные структуры, как показано на Фиг.2, и подвергнутых третьему пробегу на той же SEC колонке в 0,05% CHAPS для получения специфичных гомо-олигомерных структур с установленной примерной молекулярной массой порядка 250-300 кДа (E2s) и свыше 600 кДа (E1s). Подобные степени связывания можно получить путем использования 0,1 или 0,5%-ного бетаина.
Фиг.4. Анализ динамического светорассеяния, выраженного в виде процента числа частиц по отношению к наблюдаемому диаметру в нм, для Е1s в ЗФР/0,05% CHAPS.
Фиг.5. Анализ динамического светорассеяния, выраженного в виде процента числа частиц по отношению к наблюдаемому диаметру в нм, для Е1s в ЗФР/0,1% бетаина (сверху) или 0,5% бетаина (снизу).
Фиг.6. Электронно-микроскопическое окрашивание (А) Е1s в ЗФР/0,05% CHAPS и (Б) E1s в ЗФР/3% бетаина.
Фиг.7. Распределение по размеру частиц E1s в ЗФР/0,05% CHAPS.
Фиг.8. Эволюция антител анти-Е1, индуцированных шестью последовательными и 3 повторными иммунизациями (указаны маленькими стрелками) у инфицированного 1b шимпанзе (Phil), и эволюция АЛТ (ALT, аланинаминотрансферазы), HCV РНК и анти-Е1 антител. Анти-Е1 антитела, связывающиеся с твердой фазой Е1, обнаруживали, используя специфическую вторичную антисыворотку против IgG человека, конъюгированную с пероксидазой. В качестве субстрата для цветового проявления использовали ТМВ (тетраметилбензидин). Результаты выражены в виде конечного титра. АЛТ уровни определяли клиническим анализатором и выражали в виде ед./л. HCV РНК в сыворотке определяли, используя HCV Monitor (Roche, Basel, Switzerland). Вирусную нагрузку в печени определяли с помощью полуколичественного определения количества Е2 антигена, окрашенного в печеночной биопсии, используя специфичное моноклональное антитело (ЕСАСС, номер по каталогу 98031215, как описано в заявке ЕР №98870060.5).
Фиг.9. Эпитопное картирование гуморальных ответов, индуцированных иммунизацией Е1 у шимпанзе Phil. Реактивность антител по отношению к различным пептидам измеряли с помощью непрямого ELISA (твердофазного иммуноферментного анализа), при котором биотинилированные пептиды (смотри также Таблицу 4) адсорбировали на титрационных микропланшетах с помощью стрептавидина. Специфические антитела обнаруживали, используя специфическую вторичную антисыворотку против IgG человека, конъюгированную с пероксидазой. В качестве субстрата для цветового проявления использовали ТМВ.
Фиг.10. Результаты анализа пролиферации лимфоцитов до и после вакцинации у шимпанзе Phil. Замороженные РВМС (мононуклеарные клетки периферической крови) оттаивали и стимулировали в трех повторах различными антигенами. Негативный контроль составляла чистая среда, тогда как в качестве позитивного контроля использовали конканавалин А в концентрации 5 мкг/мл. РВМС в концентрации 2-4×105 клеток/лунку в конечном объеме 150 мкл культивировали в среде RPMI 1640, в которую добавляли 10% инактивированной нагреванием ФТС (фетальной телячьей сыворотки), в U-образных 96-луночных титрационных микропланшетах вместе с контролями или 1 мкг/мл Е1 в течение 90 ч при 37°С в увлажненной атмосфере, содержащей 5% CO2. В течение последних 18 ч клетки подвергали импульсному мечению 0,5 мкКи (3H) тимидина на лунку. Затем эти культуры собирали на фильтрах из стекловолокна и определяли включение метки.
Результаты выражены в виде индексов стимуляции (ИС): среднее число импульсов в минуту (срm) антигена/среднее число импульсов в минуту (срm) чистой среды в трех определениях.
Фиг.11. Эволюция антител анти-Е1, индуцированных шестью последовательными и 3 повторными иммунизациями (указаны маленькими стрелками) у шимпанзе Ton, инфицированного HCV подтипа 1а. Показаны эволюция АЛТ, HCV РНК в сыворотке и определение HCV антигена в печени. Анти-Е1 антитела определяли с помощью непрямого ELISA: специфические антитела, связывающиеся с твердой фазой, покрытой Е1, обнаруживали, используя специфическую вторичную антисыворотку против IgG человека, конъюгированную с пероксидазой. В качестве субстрата для цветового проявления использовали ТМВ. Результаты выражены в виде конечных титров. АЛТ уровни определяли клиническим анализатором и выражали в виде ед./л. HCV РНК в сыворотке определяли, используя HCV Monitor (Roche, Basel, Switzerland). E2 антиген окрашивали в печеночной биопсии, используя специфическое моноклональное антитело (ЕСАСС, номер по каталогу 98031215, как описано в заявке ЕР №98870060.5). Полуколичественная оценка указана черными квадратами для явного позитивного окрашивания в большинстве клеток, серыми квадратами - для явного окрашивания в большинстве клеток и белыми квадратами - для биопсий, не выявляющих обнаружимого окрашивания. HCV РНК указана маленькими черными прямоугольниками. Окрашивание E2 можно подтвердить окрашиванием Core и Е1 (данные не представлены).
Фиг.12. Эпитопное картирование гуморального ответа, индуцированного иммунизацией Е1 у Ton. Реактивность антител по отношению к различным пептидам измеряли с помощью непрямого ELISA, при котором биотинилированные пептиды (смотри также Таблицу 4) адсорбировали на титрационных микропланшетах с помощью стрептавидина. Специфические антитела обнаруживали, используя специфическую вторичную антисыворотку против IgG человека, конъюгированную с пероксидазой. В качестве субстрата для цветового проявления использовали ТМВ.
Фиг.13. Анализ Е1 гуморальных ответов на Е1 белки субтипа 1а и субтипа 1b у шимпанзе Ton. Для этой цели Е1 генотип 1а получали из HCV-H последовательности; получали рекомбинантный вирус коровьей оспы, экспрессирующий тот же участок Е1, что и для генотипа 1b (смотри ниже). Е1 получали из сырых лизатов инфицированных вирусом коровьей оспы клеток RK13 (полученных, как описано в Maertens et al. (PCT/EP95/03031)). Реактивность антител измеряли с помощью непрямого ELISA, при котором Е1 адсорбировали на титрационных микропланшетах посредством высоко связывающего маннозу агглютинина Galathus nivalis (GNA). Специфические антитела обнаруживали, используя специфическую вторичную антисыворотку против IgG человека, конъюгированную с пероксидазой. В качестве субстрата для цветового проявления использовали ТМВ. Результаты выражены в виде дифференциальной ОП (оптической плотности) (ОП лунки с адсорбированным Е1 минус ОП лунки без адсорбированного Е1).
Фиг.14. Результаты анализа пролиферации лимфоцитов до и после вакцинации у шимпанзе Ton. Замороженные РВМС оттаивали и стимулировали в трех повторах различными антигенами. Негативный контроль составляла чистая среда, тогда как в качестве позитивного контроля использовали конканавалин А в концентрации 5 мкг/мл. РВМС в концентрации 2-4×105 клеток/лунку в конечном объеме 150 мкл культивировали в среде RPMI 1640, в которую добавляли 10% инактивированной нагреванием ФТС, в U-образных 96-луночных титрационных микропланшетах вместе с контролями или 1 мкг/мл Е1 в течение 90 ч при 37°С в увлажненной атмосфере, содержащей 5% СO2. В течение последних 18 ч клетки подвергали импульсному мечению 0,5 мкКи (3H) тимидина на лунку. Затем эти культуры собирали на фильтрах из стекловолокна и определяли включение метки. Результаты выражены в виде индексов стимуляции (ИС): среднее число импульсов в минуту антигена/среднее число импульсов в минуту чистой среды в трехкратных определениях.
Фиг.15 (а, б, в, г). Карты конструкций, использованных для получения экспрессии белка Е2 с делегированным N-концевым гипервариабельным районом. Конструкции pvHCV-92 и pvHCV-99 являются промежуточными конструкциями, использованными для конструирования делеционных мутантов pvHCV-100 и pvHCV-101.
фиг.16 (а, б). Последовательность (нуклеотиды: А; трансляция: Б), соответствующая конструкциями, изображенным на Фиг.15 (смотри выше).
Фиг.17. Титры антител, полученные у мышей при иммунизации различными препаратами Е1, как описано в примере 9. Титры определяли с помощью ELISA: мышиные сыворотки разбавляли 1/20 и далее (0,5 log10) и инкубировали на E1s (модифицированном либо ацетамидом, либо малеимидом), нанесенном на титрационные микропланшеты. После отмывки связанные антитела обнаруживали, используя специфическую вторичную антисыворотку против IgG мыши, конъюгированную с пероксидазой. В качестве субстрата для цветового проявления использовали ТМВ. Результаты выражены в виде конечного титра, показаны стандартные отклонения (n=6).
Фиг.18. Эпитопное картирование гуморального иммунного ответа, индуцированного иммунизацией различными препаратами E1s у мышей. Реактивность антител по отношению к различным пептидам измеряли с помощью непрямого ELISA, при котором биотинилированные пептиды (перечисленные в таблице 4) адсорбировали на титрационных микропланшетах посредством стрептавидина. Мышиные сыворотки разбавляли 1/20, и специфические антитела обнаруживали, используя специфическую вторичную антисыворотку против IgG человека, конъюгированную с пероксидазой. В качестве субстрата для цветового проявления использовали ТМВ.
Фиг.19. Профиль изотипирования иммуноглобулина мышей, иммунизированных различными препаратами E1s. Специфические антитела класса и подкласса Ig адсорбировали на титрационном микропланшете. После захвата мышиного Ig из иммунных сывороток, разбавленных 1/500, E1s инкубировали при 1 мкг/мл. Образовавшиеся иммунокомплексы далее инкубировали с поликлональной антисывороткой кролика, направленной против Е1. Наконец, антитела кролика обнаруживали, используя вторичную антисыворотку Ig коза против кролика, конъюгированную с пероксидазой. В качестве субстрата для цветового проявления использовали ТМВ. Результаты нормировали по IgG (то есть IgG сигнал для каждого животного отдельно считали равным 1 и все результаты для других изотипов выражали относительно этого результата IgG1).
Фиг.20. Титры антител, индуцированных двумя иммунизациями приблизительно на 1000-е сутки E1s-ацетамидом у шимпанзе Phil. Антитела анти-Е1 определяли с помощью непрямого ELISA: специфические антитела, связывающиеся с твердой фазой Е1, обнаруживали, используя специфическую вторичную антисыворотку против IgG человека, конъюгированную с пероксидазой. Титр выражен в единицах/мл, эти единицы относятся к местному стандарту, который основан на человеческих сыворотках.
Фиг.21. Титры антител, индуцированных двумя иммунизациями приблизительно на 900-е сутки E1s-ацетамидом у шимпанзе Ton. Антитела анти-Е1 определяли с помощью непрямого ELISA: специфические антитела, связанные с твердой фазой Е1, обнаруживали, используя специфическую вторичную антисыворотку против IgG человека, конъюгированную с пероксидазой. Титр выражен в единицах/мл, эти единицы относятся к местному стандарту, который основан на человеческих сыворотках.
Фиг.22 (А, Б, В). SEC профиль конечной стадии снижения детергента (от 0,2 до 0,5% CHAPS): частица одного Е1 (А), частица одного Е2 (Б) или эквимолярная смесь Е1 и Е2; смешанная частица (В). На этой фигуре также показано перекрывание значений ОП ELISA, специфически обнаруживающего только Е1 (сверху), только Е2 (в середине) и ELISA, обнаруживающего только смешанные частицы (снизу).
Фиг.23. SEC профиль конечной стадии снижения детергента (от 0,2 до 0,5% CHAPS): частица одного Е1 генотипа 1b (сверху), частица одного Е1 генотипа 4 (в середине) или эквимолярная смесь Е1 генотипа 1b и 4; смешанная частица (снизу). На этой фигуре также показано перекрывание значений ОП ELISA, специфически обнаруживающего только смешанные частицы (смотри также Фиг.22).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение, описанное здесь, основано на ранее опубликованной работе и заявках на патент, находящихся на рассмотрении. К примеру, такая работа состоит из научных работ, патентов или заявок на патент, находящихся на рассмотрении. Все эти публикации и заявки, цитированные ранее или ниже, включены здесь путем ссылки.
Настоящее изобретение относится к HCV вакцинации. Впервые успешная иммунотерапия шимпанзе с тяжелым хроническим активным гепатитом С смогла быть достигнута с помощью вакцинации HCV антигеном. Эта вакцина не только индуцировала высокие иммунные ответы, но также индуцировала клиренс вирусного антигена из печени и значительное улучшение гистологической активности и степени заболевания печени. Кроме того, настоящее изобретение относится к очищенным отдельным белкам оболочки HCV и, в частности, к олигомерным частицам. Эти олигомерные частицы состоят по существу из белков оболочки HCV и имеют диаметр от 1 до 100 нм при измерении с помощью динамического светорассеяния или, возможно, электронной микроскопии. В этом отношении следует подчеркнуть, что эти частицы могут быть образованы только Е1 и/или Е2 белками, либо их частями (смотри ниже). Следовательно, олигомерные частицы по настоящему изобретению фундаментально отличаются от HCV-подобных частиц, описанных в WO 98/21338, которые обязательно состоят из Е1 и Е2, а также Core и Р7. Термин “олигомерные частицы, состоящие по существу из белков оболочки HCV”, здесь определен как структуры специфической природы и формы, содержащие несколько основных единиц HCV Е1 и/или Е2 белков оболочки, которые сами по себе считают состоящими из одного или двух Е1 и/или Е2 мономеров соответственно. Должно быть ясно, что частицы по настоящему изобретению определяют как свободные от инфекционных HCV РНК геномов. Частицы по настоящему изобретению могут представлять собой частицы высшего порядка сферической природы, которые могут быть пустыми, состоящими из оболочки из оболочечных белков, в которую могут быть включены липиды, детергенты, ядерный белок HCV или молекулы адъювантов. Последние частицы также можно инкапсулировать липосомами или аполипопротеинами, такими как, например, аполипопротеин В или липопротеины низкой плотности, либо с помощью любых других средств, обеспечивающих направленную доставку указанных частиц к конкретному органу или ткани. В этом случае на такие пустые сферические частицы часто ссылаются как на “вирусоподобные частицы” или ВПЧ. Альтернативно эти частицы высшего порядка могут представлять собой твердые сферические структуры, в которых заполненная сфера состоит из олигомеров белков оболочки Е1 или Е2 HCV, в которые могут быть дополнительно включены липиды, детергенты, ядерный белок HCV или молекулы адъювантов, или которые в свою очередь сами могут быть инкапсулированы липосомами или аполипопротеинами, такими как, например, аполипопротеин В, липопротеины низкой плотности, либо с помощью других средств, обеспечивающих направленную доставку указанных частиц к конкретному органу или ткани, например асиалогликопротеинов. Эти частицы могут также состоять из более мелких структур (по сравнению с пустыми или твердыми сферическими структурами, указанными выше), которые обычно имеют округлую (смотри далее) форму и которые обычно содержат не более одного слоя белков оболочки HCV. Типичным примером таких более мелких частиц являются розетко-подобные структуры, которые состоят из меньшего числа белков оболочки HCV, обычно между 4 и 16. Конкретный пример последних включает в себя более мелкие частицы, полученные с E1s в 0,2%-ном CHAPS, пример которых приведен здесь, которые содержат примерно 8-10 мономеров E1s. Такие розетко-подобные структуры имеют плоскостную организацию и характеризуются круглой формой, например формой колеса. Опять же, дополнительно могут быть включены липиды, детергенты, капсидный белок HCV или молекулы адъюванта, либо эти более мелкие частицы могут быть инкапсулированы липосомами или аполипопротеинами, такими как, например, аполипопротеин В или липопротеины низкой плотности, либо с помощью любых других средств, обеспечивающих направленную доставку указанных частиц к конкретному органу или ткани. Более мелкие частицы могут также образовывать небольшие сферические или глобулярные структуры, состоящие из подобного меньшего числа белков оболочки Е1 или Е2 HCV, в которые могут быть дополнительно включены липиды, детергенты, ядерный белок HCV или молекулы адъюванта, или которые в свою очередь могут быть инкапсулированы липосомами или аполипопротеинами, такими как, например, аполипопротеин В или липопротеины низкой плотности, либо с помощью любых других средств, обеспечивающих направленную доставку указанных частиц к конкретному органу или ткани. Размер (то есть диаметр) этих определенных выше частиц, измеренный с помощью известных в данной области методик динамического светорассеяния (смотри далее в разделе примеров), составляет обычно между 1 и 100 нм, более предпочтительно между 2 и 70 нм, еще более предпочтительно между 2 и 40 нм, между 3 и 20 нм, между 5 и 16 нм, между 7 и 14 нм или между 8 и 12 нм.
Далее изобретение относится к олигомерной частице, как определено выше, где указанные белки оболочки выбраны из группы, состоящей из HCV Е1, HCV E1s, HCV E2 белков, SEQ ID No 13 или SEQ ID No 14, либо их частей. Белки HCV E1 и HCV E2, а также детальное описание того, как очистить последние белки, подробно представлены и охарактеризованы в РСТ/ЕР 95/03031, Maertens et al. HCV E1s, SEQ ID No 13 или SEQ ID No 14, либо их части можно очистить подобно тому, как описано для HCV E1 или HCV E1s в РСТ/ЕР 95/03031, Maertens et al. Следует подчеркнуть, что все содержание, включая все определения, последнего документа включено в настоящую заявку посредством ссылки. Белок HCV E1s относится к аминокислотам 192-326 E1 и представляет собой E1 белок без его С-концевого гидрофобного якоря. Термин “или его части” относится к любой части указанных здесь белков, которая является иммуногенной, если она представляет собой часть частицы по настоящему изобретению.
Изобретение далее относится к олигомерным частицам, как описано здесь, где по меньшей мере один цистеиновый остаток белка оболочки HCV, как описано выше, алкилирован, предпочтительно алкилирован с помощью алкилирующих агентов, таких как, например, активные галогены, этиленимин или N-(иодэтил)трифторацетамид. В этом отношении следует понимать, что алкилирование цистеинов относится к цистеинам, в которых атом водорода при атоме серы заменен на (СН2)nR, в котором n составляет 0, 1, 2, 3 или 4, a R=Н, СООН, NH2, CONH2, фенил или любое их производное. Алкилирование можно осуществлять любым способом, известным в данной области, таким как, например, использование активных галогенов Х(СН2)nR, в которых Х представляет собой галоген, такой как I, Br, Cl или F. Примерами активных галогенов являются метилиодид, иодуксусная кислота, иодацетамид и 2-бромэтиламин. Другие способы алкилирования включают в себя использование этиленимина или N-(иодэтил)трифторацетамида, причем оба приводят к замещению Н на -CH2-CH2-NH2 (Hermanson, 1996). Термин “алкилирующие агенты”, как его используют здесь, относится к соединениям, которые способны осуществлять алкилирование, как описано здесь. Такие алкилирования в конечном итоге приводят к получению модифицированного цистеина, который может имитировать другие аминокислоты. Алкилирование этиленимином приводит к структуре, имеющей сходство с лизином, таким образом, что вводятся новые сайты расщепления для трипсина (HerNanson, 1996). Подобным же образом использование метилиодида приводит к получению аминокислоты, имеющей сходство с метионином, тогда как использование иодацетата и иодацетамида приводит к получению аминокислот, имеющих сходство с глутаминовой кислотой и глутамином соответственно. По аналогии эти аминокислоты предпочтительно используют при прямой мутации цистеина. Следовательно, настоящее изобретение относится к олигомерным частицам, как описано здесь, где по меньшей мере один цистеиновый остаток белка оболочки HCV, как описано здесь, мутирован в природную аминокислоту, предпочтительно метионин, глутаминовую кислоту, глутамин или лизин. Термин “мутирован” относится к сайт-направленному мутагенезу нуклеиновых кислот, кодирующих эти аминокислоты, то есть к хорошо известным в данной области способам, таким как, например, сайт-направленный мутагенез посредством ПЦР или опосредованный олигонуклеотидами мутагенез, как описано в Sambrook et al. (1989).
Термин “очищенный”, как его применяют здесь, относится к композиции, где желаемые компоненты, такие как, например, белки оболочки HCV или олигомерные частицы, составляют по меньшей мере 35% совокупных компонентов в этой композиции. Желаемые компоненты предпочтительно составляют по меньшей мере примерно 40%, более предпочтительно по меньшей мере примерно 50%, еще более предпочтительно по меньшей мере примерно 60%, еще более предпочтительно по меньшей мере примерно 70%, даже более предпочтительно по меньшей мере примерно 80%, даже более предпочтительно по меньшей мере примерно 90%, даже более предпочтительно по меньшей мере примерно 95% и наиболее предпочтительно по меньшей мере примерно 98% фракции совокупных компонентов в этой композиции. Эта композиция может содержать другие соединения, такие как, например, углеводы, соли, липиды, растворители и тому подобное, без воздействия на определение процента чистоты, как используют здесь. “Выделенная” олигомерная частица HCV подразумевает композицию олигомерных частиц HCV, которая является по меньшей мере на 35% чистой. В этом отношении должно быть ясно, что термин “очищенный отдельный белок оболочки HCV”, как его используют здесь, относится к выделенным белкам оболочки HCV в по существу чистой форме. Термины “по существу очищенные олигомерные частицы” и “отдельные белки оболочки HCV”, как их используют здесь, относятся к олигомерным частицам HCV или отдельным белкам оболочки HCV, таким, которые можно применять для in vitro способов диагностики и терапии. Эти олигомерные частицы HCV являются по существу свободными от клеточных белков, белков векторного происхождения или других HCV вирусных компонентов. Обычно эти частицы или белки очищены до гомогенности (по меньшей мере 80% чистоты, предпочтительно 85%, более предпочтительно 90%, более предпочтительно 95%, более предпочтительно 97%, более предпочтительно 98%, более предпочтительно 99%, даже более предпочтительно 99,5%, и наиболее предпочтительно загрязняющие белки должны быть не обнаружимы общепринятыми способами, такими как ДДС-ПААГ (электрофорез в полиакриламидном геле с додецилсульфатом натрия) и окрашивание серебром).
Настоящее изобретение также относится к олигомерной частице, как определено выше, где указанные белки оболочки составляют любую возможную смесь HCV E1, HCV E1s, HCV E2, SEQ ID No 13 и/или SEQ ID No 14 либо их частей, так, например, частица по настоящему изобретению может по существу состоять из белков HCV E1 и HCV E2, белков HCV E1 и HCV E1s, белков HCV E1s и HCV E2 и белков HCV E1, HCV E1s и HCV E2. Кроме того, настоящее изобретение также относится к олигомерной частице, как определено выше, где указанные белки получены из различных штаммов, субтипов или генотипов HCV, так, например, указанные белки получены из генотипа 1b и геноти па 4 либо представляют собой смесь, состоящую из белков оболочки HCV из одного штамма или генотипа HCV и по меньшей мере одного иного штамма или генотипа HCV. Различные штаммы или генотипы HCV хорошо определены и охарактеризованы в РСТ/ЕР95/04155, Naertens et al. Еще раз подчеркнем, что полное содержание, включая все определения, последнего документа включено путем ссылки в настоящую заявку. Таким образом, настоящее изобретение относится к олигомерным частицам, содержащим белки оболочки, имеющие происхождение от любого штамма или генотипа HCV, известного в данной области, либо к частицам, содержащим смесь белков, имеющих происхождение от любого штамма или генотипа HCV, известного в данной области. В этом отношении настоящее изобретение также относится к согласованным последовательностям, имеющим происхождение от индивидуальных клонов, пример которых приведен ниже и в разделе примеров (смотри далее).
Далее настоящее изобретение относится как к олигомерной частице, как описано выше, которую можно получить с помощью способа, так и к указанному способу получения указанной олигомерной частицы. Указанный способ характеризуется следующими стадиями:
(I) Очистка белков оболочки HCV, возможно включая использование первого детергента. По существу методика очистки стадии (I) подробно описана в РСТ ЕР 95/03031, Naertens et al. Важно, что согласно настоящему изобретению стадию блокирования в этой методике очистки, как описано в РСТ ЕР 95/03031, например NEN ((N-этилмалеимид)-биотин), осуществляют вместе со стадией алкилирования, как описано в настоящей заявке, предпочтительно путем использования иодацетамида. Кроме того, методика очистки стадии (I) может, возможно, включать в себя использование агента, расщепляющего дисульфидную связь, а также, возможно, включать в себя использование алкилирующего агента. В конечном итоге методика стадии (I) дает в результате очищенные белки оболочки HCV в растворе.
(II) Замещение раствора указанных очищенных белков оболочки HCV детергентом или солью, приводящее к образованию олигомерных частиц.
(III) Извлечение или очистка указанных олигомерных частиц, возможно, включая дополнительное снижение концентрации детергента или соли стадии (II), что дополнительно способствует образованию или стабилизации указанных олигомерных частиц, образованных после указанной замены.
Более предпочтительно настоящее изобретение относится как к олигомерной частице, как определено здесь, так и к способу получения указанной частицы, где указанный первый детергент представляет собой Empigen-BB. Более предпочтительно настоящее изобретение относится как к олигомерной частице, как определено здесь, так и к способу получения указанной частицы, где детергент стадии (II) представляет собой CHAPS, октилглюказид или Твин, более предпочтительно Твин-20 или Твин-80, или любой другой детергент. Более предпочтительно настоящее изобретение относится как к олигомерной частице, как определено здесь, так и к способу получения указанной частицы, где указанная соль представляет собой бетаин. Еще более предпочтительно настоящее изобретение относится как к олигомерной частице, как определено здесь, так и к способу получения указанной частицы, где указанный Empigen-BB используют в концентрации от 1% до 10% и где указанный CHAPS или Твин используют в концентрации от 0,01% до 10%, или указанный бетаин используют в концентрации от 0,01% до 10%. Еще более предпочтительно настоящее изобретение относится как к олигомерной частице, как определено здесь, так и к способу получения указанной частицы, где указанный Empigen-BB используют в концентрации 3% и где указанный CHAPS или бетаин используют в концентрациях 0,2% или 0,3% соответственно, после чего буфер меняют и указанный CHAPS или бетаин используют в концентрациях 0,05% или 0,1-0,5% соответственно. Следует понимать, что все проценты, используемые в вышеописанном способе, даны в виде мас./об. Должно быть ясно, что вышеописанный способ (смотри также РСТ/ЕР 95/03031 и раздел примеров настоящей заявки) представляет собой пример того, как получать частицы по настоящему изобретению. В этом отношении настоящее изобретение также касается любого другого способа, известного в данной области, который можно использовать для получения олигомерных частиц по настоящему изобретению, такого как, например, исключение восстанавливающего агента, как описано в РСТ/ЕР 95/03031 и в разделе примеров (ниже), и вместо этого использование для восстановления цистеиновых мостиков клеток-хозяев, которые имеют оптимизированное окислительно-восстановительное состояние в эндоплазматическом ретикулуме. Кроме того, должно быть ясно, что можно использовать целый ряд алкилбетаинов, таких как, например, с Сn хвостом, в котором n - положительное целое число, находящееся в диапазоне от 1 до 20, а также производные бетаина, такие как, например, сульфобетаины.
Поскольку впервые была достигнута успешная иммунотерапия шимпанзе с тяжелым хроническим активным гепатитом С путем вакцинации очищенным HCV антигеном, настоящее изобретение также относится к очищенным отдельным белкам оболочки HCV, в частности Е1 или E1s. Кроме того, настоящее изобретение относится к композиции, содержащей указанные отдельные белки оболочки HCV, и к ее применению в качестве HCV вакцины или для производства HCV вакцины.
Чтобы избежать индукции иммунного ответа против ненужных эпитопов, белок оболочки HCV, применяемый для вакцинации, предпочтительно конструируют в виде согласованной последовательности индивидуальных подтипов, штаммов или клонов. Следовательно, настоящее изобретение также относится к применению HCV антигена (в форме пептида, белка или полинуклеотида) для вакцинации или диагностики. Кроме того, настоящее изобретение также относится к олигомерной частице, как определено здесь, и ее применению, в которой белок оболочки HCV кодируется согласованными последовательностями, основанными на квазивидовой вариабельности в пределах изолята (согласованной последовательности изолята), или основанными на согласованной последовательности различных изолятов в пределах подтипа (согласованной последовательности подтипа), типа или вида (согласованной последовательности типа или вида), либо целого рода HCV (согласованной последовательности рода). Поэтому аминокислотная последовательность этого согласованного белка оболочки HCV представляет собой согласованную последовательность, имеющую происхождение от согласованной последовательности изолята, субтипа, штамма или рода. Для коннотации термина “согласованный”, в частности, ссылаются на Naertens and Stuyver (1997), а также ссылки, используемые здесь.
Олигомерная частица по настоящему изобретению отображает эпитопы крайне эффективно (смотри ниже). Следовательно, эта олигомерная частица является средством представления эпитопов таким образом, что они могут вызывать четкий иммунный ответ. В этом контексте понятно, что нет необходимости в том, чтобы белки оболочки HCV, как определено здесь, содержали HCV эпитопы исключительно. Белки оболочки HCV, которые образуют олигомерные частицы, могут содержать эпитопы, которые имеют происхождение только от HCV, и, возможно, содержат эпитопы, которые имеют происхождение от других экзогенных агентов, таких как, например, HBV (вирус гепатита В) или HIV (вирус иммунодефицита человека). Иными словами, олигомерную частицу с остовом из белков оболочки HCV можно использовать в качестве носителя для представления не-HCV эпитопов, возможно, в дополнение к HCV эпитопам. Следовательно, настоящее изобретение также охватывает олигомерную частицу, как определено здесь, но, возможно, без HCV эпитопов, возможно содержащую не-HCV эпитопы, а также ее применения и производство. Термин “экзогенный агент”, как его используют здесь, относится к любому агенту, либо живому, либо неживому, способному вызывать иммунный ответ, причем этот агент не является эндогенным по отношению к хозяину и не является HCV. Конкретно последний термин относится к группе, состоящей из патогенных агентов, аллергенов и гаптенов. Патогенные агенты включают в себя прионы, вирусы, прокариоты и эукариоты. Более конкретно, вирус включает в себя, в частности, HBV, HIV или вирус герпеса, но не HCV. Аллергены включают в себя вещества или молекулы, способные провоцировать собственный иммунный ответ у хозяина, когда хозяин контактирует с указанными аллергенами. Гаптены ведут себя подобно аллергенам в отношении способности к провокации иммунного ответа, но, в противоположность аллергенам, гаптены нуждаются в молекуле-носителе.
Настоящее изобретение также относится к композиции, содержащей олигомерную частицу, как определено выше. Более конкретно настоящее изобретение относится к вакцинной композиции. Термин “вакцинная композиция” относится к иммуногенной композиции, способной элисировать защиту против HCV, либо частичную, либо полную. Она, следовательно, включает в себя пептиды, белки или полинуклеотиды HCV. Защита против HCV относится, в частности, к людям, но также относится к приматам, кроме человека, trimera мыши (Zauberman et al., 1999) или другим млекопитающим.
Частицы по настоящему изобретению можно применять сами по себе, в биотинилированной форме (как показано в WO 93/18054) и/или в комплексе с Neutrality Avidin (Nolecular Probes Inc., Eugene, OR, USA). Следует также отметить, что “вакцинная композиция” содержит, в дополнение к активному веществу, подходящий эксципиент, разбавитель, носитель и/или адъювант, которые сами по себе не индуцируют продуцирование антител, вредных для индивидуума, получающего эту композицию, и не элисируют защиту. Подходящими носителями являются обычно большие медленно метаболизируемые макромолекулы, такие как белки, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, сополимеры аминокислот и неактивные вирусные частицы. Такие носители хорошо известны специалистам в данной области. Предпочтительные адъюванты для усиления эффективности композиции включают в себя: гидроксид алюминия, алюминий в комбинации с 3-O-деацилированным монофосфориллипидом А, как описано в WO 93/19780, фосфат алюминия, как описано в WO 93/24148, N-ацетил-мурамил-L-треонил-D-изоглутамин, как описано в Патенте США №4606918, N-ацетил-нормурамил-L-аланил-D-изоглутамин, N-ацетилмурамил-L-аланил-D-изоглутамил-L-аланин 2-(1'2'-дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)этиламин и RIBI (ImmunoChem Research Inc., Hamilton, NT, USA), который содержит монофосфориллипид А, обезвреженный эндотоксин, трегалозо-6,6-димиколат и остов клеточных стенок (MPL+TDM+CWS) в эмульсии 2% сквален/Tвин 80, но не ограничиваются ими. Любой из этих трех компонентов, MPL, TDM или CWS, можно также использовать отдельно или комбинировать 2 на 2. Кроме того, можно использовать такие адъюванты, как Stimulon (Cambridge Bioscience, Worcester, MA, USA) или SAF-1 (Syntex), а также такие адъюванты, как комбинации между QS21 и 3-де-O-ацетилированным монофосфориллипидом А (WO 94/00153) или MF-59 (Chiron), либо адъюванты на основе поли[ди(карбоксилатофенокси)фосфазена] (Virus Research Institute), либо адъюванты на основе блокполимеров, такие как Optivax (Vaxcel, Cythx), либо адъюванты на основе инулина, такие как Algammulin и Gammalnulin (Anutech), неполный адъювант Фрейнда (IFA) или препараты Gerbu (Gerbu Biotechnik), Следует понимать, что полный адъювант Фрейнда (CFA) можно также использовать для применений не для человека и для исследовательских целей. “Вакцинная композиция” дополнительно содержит эксципиенты и разбавители, которые являются по своей сути являются нетоксичными и нетерапевтическими, такие как вода, физиологический раствор, глицерин, этанол, увлажняющие или эмульгирующие агенты, вещества, забуферивающие рН, консерванты и тому подобное. Обычно вакцинную композицию готовят в инъекционной форме, либо в виде жидкого раствора, либо в виде суспензии. Можно также готовить твердые формы, подходящие для растворения или для суспендирования в жидких носителях перед инъекцией. Препарат можно также эмульгировать или инкапсулировать в липосомы для усиления эффекта адъюванта. Полипептиды можно также включать в иммуностимулирующие комплексы вместе с сапонинами, например Quit A (ISCONS). Вакцинные композиции содержат иммунологически эффективное количество полипептидов по настоящему изобретению, а также любой другой из вышеупомянутых компонентов. “Иммунологически эффективное количество” означает, что введение этого количества индивидууму, либо в однократной дозе, либо в виде части серии, является эффективным для предупреждения или лечения. Это количество варьирует в зависимости от состояния здоровья и физического состояния индивидуума, которого нужно лечить, от таксономической группы индивидуума, которого нужно лечить (например, человек, примат, кроме человека, примат и так далее), от способности иммунной системы индивидуума поддерживать эффективный иммунный ответ, от степени желаемой защиты, от препарата вакцины, от назначения лечащего врача, от штамма инфицирующего HCV и других относящихся к делу факторов. Полагают, что это количество находится в пределах относительно широкого диапазона, который можно определить посредством традиционных испытаний. Обычно это количество варьирует от 0,01 до 1000 мкг/дозу, более конкретно от 0,1 до 100 мкг/дозу. Как правило, вакцинные композиции вводят парентерально, обычно посредством инъекции, например подкожно или внутримышечно. Дополнительные препараты, подходящие для других способов введения, включают в себя пероральные препараты и суппозитории. Лечебная система доз может представлять собой режим однократной дозировки или режим многократной дозировки. Вакцину можно вводить в сочетании с другими иммунорегулирующими агентами. Следовательно, настоящее изобретение относится к применению олигомерной частицы, как определено здесь, для профилактической индукции иммунитета против HCV. Следует отметить, что вакцина также может быть полезной для лечения индивидуума, как указано выше, причем в этом случае ее называют “терапевтической вакциной”.
Настоящее изобретение также относится к композиции, как определено выше, которая также содержит ядерный белок (core) HCV, Е1, Е2, Р7, NS2, NS3, NS4A, NS4B, NS5A и/или белок NS5B или их части. Е1, Е2 и/или Е1Е2 частицы можно, например, комбинировать с антигенами, стимулирующими Т-клетки, такими как, например, ядерный белок (core), P7, NS3, NS4A, NS4B, NS5A и/или NS5B. В частности, настоящее изобретение относится к композиции, как определено выше, где указанный белок NS3 или его части имеют аминокислотную последовательность, представленную SEQ ID 1 или SEQ ID 2 (смотри далее в разделе примеров).
Очистка этих NS3 белков будет предпочтительно включать в себя обратимую модификацию цистеиновых остатков и даже более предпочтительно сульфирование цистеинов. Способы получения такой обратимой модификации, включая сульфирование, описаны для NS3 белка в Maertens et al. (PCT/EP99/02547). Следует подчеркнуть, что полное содержание, включая все определения, последнего документа включено путем ссылки в настоящую заявку.
Из вышеописанного ясно, что настоящее изобретение также относится к применению олигомерной частицы, как определено выше, или к композиции, как определено выше, для производства HCV вакцинной композиции. В частности, настоящее изобретение относится к применению олигомерной частицы, как определено выше, для индукции иммунитета против HCV у хронических носителей HCV. Более конкретно настоящее изобретение относится к применению олигомерной частицы, как определено выше, для индукции иммунитета против HCV у хронических носителей HCV до, во время или после любой другой терапии, такой как, например, хорошо известная терапия интерфероном, либо в сочетании, либо не в сочетании с введением малых лекарств, лечащих HCV, таких как, например, рибавирин. Такую композицию можно также применять до или после трансплантации печени или после предполагаемой инфекции, такой как, например, повреждение уколом иглы. Кроме того, настоящее изобретение относится к набору, содержащему олигомерные частицы или отдельные белки оболочки HCV по настоящему изобретению, для определения HCV антител, присутствующих в биологическом образце. Термин “биологический образец”, как его используют здесь, относится к образцу ткани или жидкости, выделенной от индивидуума, включая, но не ограничиваясь ими, сыворотку, плазму, лимфатическую жидкость, поверхностные срезы кожи, респираторного, кишечного и мочеполового трактов, ооциты, слезы, слюну, молоко, клетки крови, опухоли, органы, желудочные секреты, слизь, спинномозговую жидкость, наружные секреты, такие как, например, экскременты, моча, сперма и тому подобное. Поскольку олигомерные частицы и отдельные белки оболочки HCV по настоящему изобретению являются высоко иммуногенными и стимулируют как гуморальный, так и клеточный иммунный ответ, настоящее изобретение относится также к набору для обнаружения связанной с HCV Т-клеточной иммунной реакции, содержащему олигомерную частицу или очищенный отдельный белок оболочки HCV по настоящему изобретению. HCV Т-клеточную иммунную реакцию можно, например, измерять, как описано в разделе примеров, или как описано в РСТ/ЕР 94/03555, Leroux-Roels et al.
Следует подчеркнуть, что полное содержание, включая все определения, данного документа включено путем ссылки в настоящую заявку.
Должно быть ясно, что настоящее изобретение также относится к применению специфических HCV иммуноглобулинов для лечения и предупреждения HCV инфекции. Именно здесь впервые продемонстрировано, что достаточные уровни HCV антител, особенно антител к HCV оболочке, индуцируют уменьшение интенсивности заболевания гепатитом С. Также впервые продемонстрировано, что достаточные уровни антител могут связывать циркулирующий вирус и что присутствие комплекса Аb с вирусом совпадает с исчезновением HCV антигена из печени и с уменьшением интенсивности заболевания печени. Антитела к HCV оболочке можно индуцировать с помощью вакцинации, либо их можно пассивно перенести с помощью инъекции после того, как эти антитела были очищены из пулов HCV инфицированной крови или из крови, полученной от вакцинированных HCV. Следовательно, настоящее изобретение далее относится к специфическим антителам, генерируемым против олигомерной частицы, как описано выше, или против композиции, как описано выше, либо против отдельного белка оболочки HCV. В частности, настоящее изобретение относится к набору, содержащему указанные антитела для обнаружения HCV антигенов. Термин “специфические антитела”, как его используют здесь, относится к антителам, которые индуцируются против эпитопов, которые являются специфическими для олигомерной частицы, как описано в настоящем изобретении. Иными словами, специфические антитела индуцируются против эпитопов, которые являются результатом образования олигомерных частиц и присутствуют только на олигомерных частицах. Кроме того, существуют различные методики, известные для получения HCV пептидов. Результатом осуществления этих методик могут быть HCV пептиды, способные к представлению эпитопов. Предположительно эти HCV пептиды, полученные с помощью этих разнообразных и различных методик, способны к представлению сходных эпитопов. Сходные эпитопы представляют собой эпитопы, являющиеся результатом осуществления различных методик получения или очистки, но распознаваемые одним и тем же антителом. Однако, олигомерные частицы по настоящему изобретению представляют эпитопы чрезвычайно эффективно. Поэтому эпитопы на этих олигомерных частицах являются высоко иммуногенными. Следовательно, настоящее изобретение также относится к эпитопам на олигомерных частицах, причем указанные эпитопы являются по меньшей мере в 10 раз, предпочтительно по меньшей мере в 20 раз, предпочтительно по меньшей мере в 50 раз, предпочтительно по меньшей мере в 100 раз, предпочтительно по меньшей мере в 500 раз и наиболее предпочтительно по меньшей мере в 1000 раз более иммуногенными, чем эпитопы на HCV пептидах, которые не продуцируются согласно настоящему изобретению, то есть не продуцируются путем образования частицы в присутствии детергента. Специалистам следует понимать, что указанную иммуногенность можно, например, обнаружить, а затем сравнить путем иммунизации млекопитающих посредством введения сравнимых количеств пептидов, полученных любым способом. Кроме того, термин “специфическое антитело” относится также к антителам, которые индуцируются против очищенного отдельного белка оболочки HCV. Как его используют здесь, термин “антитело” относится к поликлональным или моноклональным антителам. Термин “моноклональное антитело” относится к композиции антител, имеющей однородную популяцию антител. Термин “антитело” не ограничен в отношении вида или источника этого антитела и не предназначен для ограничения способом, которым оно получено. Кроме того, термин “антитело” также относится к гуманизированным антителам, в которых по меньшей мере часть районов рамки считывания иммуноглобулина имеет происхождение от последовательностей человеческого иммуноглобулина, и к одноцепочечным антителам, таким как, например, описанные в патенте США №4946778, к фрагментам антител, таким как Fab, F(аь)2, FV, и другим фрагментам, которые сохраняют функцию связывания антигена и специфичность родительского антитела.
Кроме того, признаком настоящего изобретения также является применение олигомерной частицы, как описано выше, или композиции, как описано выше, для обнаружения антител против белков оболочки HCV. Как его используют здесь, термин “обнаруживать” относится к любому анализу, известному в данной области, подходящему для обнаружения. В частности, этот термин относится к любому иммунологическому анализу, как описано в WO 96/13590.
Термины “пептид”, “полипептид” и “белок” в настоящем изобретении используют взаимозаменяемо. “Полипептид” относится к полимеру из аминокислот (аминокислотной последовательности) и не касается конкретной длины молекулы. Таким образом, олигопептиды включены в определение полипептида. Следует понимать, что пептидомиметики включены в термины “полипептид” и “белок”.
Настоящее изобретение также относится к применению олигомерной частицы, как описано здесь, для индукции иммунитета против HCV, отличающемуся тем, что указанную олигомерную частицу применяют как часть системы времени и соединений. В этом отношении следует понимать, что термин “система времени и соединений” относится к введению индивидууму с временными интервалами соединений, применяемых для элисирования иммунного ответа. Последние соединения могут содержать любые из следующих компонентов: олигомерные частицы, вакцинная композиция HCV ДНК, HCV полипептиды.
В этом отношении система включает в себя введение любого из:
(I) HCV антиген, такой как, например, олигомерная частица, с временными интервалами, или
(II) HCV антиген, такой как, например, олигомерная частица, в комбинации с вакцинной композицией HCV ДНК, где указанные олигомерные частицы и указанную вакцинную композицию HCV ДНК можно вводить одновременно или с различными временными интервалами, включая чередующиеся временные интервалы, или
(III) либо (I), либо (II), возможно в комбинации с другими HCV пептидами, с временными интервалами.
В этом отношении должно быть ясно, что вакцинная композиция HCV ДНК содержит нуклеиновые кислоты, кодирующие пептид оболочки HCV, включая Е1-, Е2-, Е1/Е2-пептиды, E1s пептид, SEQ ID No 13, SEQ ID No 14, NS3 пептид, другие HCV пептиды или части указанных пептидов. Кроме того, cледует понимать, что указанные HCV пептиды включают в себя пептиды оболочки HCV, включая Е1-, Е2-, Е1/Е2-пептиды, E1s пептид, SEQ ID No 13, SEQ ID No 14, NS3 пептид, другие HCV пептиды или их частии. Термин “другие HCV пептиды” относится к любому HCV пептиду или его фрагменту с условием, что указанный HCV пептид не представляет собой Е1, Е2, E1s, SEQ ID No 13, SEQ ID No 14 или NS3. В пункте II вышеуказанной схемы вакцинная композиция HCV ДНК содержит предпочтительно нуклеиновые кислоты, кодирующие пептиды оболочки HCV. В пункте II вышеуказанной схемы вакцинная композиция HCV ДНК состоит даже более предпочтительно из нуклеиновых кислот, кодирующих пептиды оболочки HCV, возможно в комбинации с вакцинной композицией ДНК HCV-NS3. В этом отношении должно быть ясно, что вакцинная композиция HCV ДНК содержит плазмидный вектор, содержащий полинуклеотидную последовательность, кодирующую HCV пептид, как описано выше, связанный с возможностью функционирования с транскрипционными регуляторными элементами. Как его используют здесь, термин “плазмидный вектор” относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она была сшита. Предпочтительными векторами являются вектора, способные к автономной репликации и/или экспрессии нуклеиновых кислот, с которыми они были сшиты. Как правило, но не ограничиваясь этим, плазмидные вектора представляют собой кольцевые петли двунитевой ДНК, которые в своей векторной форме не связаны с хромосомой. Как его используют здесь, термин “полинуклеотидная последовательность” относится к полинуклеотидам, таким как дезоксирибонуклеиновая кислота (ДНК) и, где это является подходящим, рибонуклеиновая кислота (РНК). Следует также понимать, что в этот термин включают в качестве эквивалентов аналоги либо РНК, либо ДНК, полученные с использованием аналогов нуклеотидов, а также однонитевые (смысловые или антисмысловые) и двунитевые полинуклеотиды. Как его используют здесь, термин “транскрипционные регуляторные элементы” относится к нуклеотидной последовательности, которая содержит существенные регуляторные элементы, такие как те, которые при введении в живую клетку позвоночного способны направлять клеточные механизмы на продуцирование продуктов трансляции, кодируемых этим полинуклеотидом. Термин “связанный с возможностью функционирования” относится к непосредственному соседству, где компоненты находятся в такой конфигурации, при которой они осуществляют свою обычную функцию. Так, транскрипционные регуляторные элементы, связанные с возможностью функционирования с нуклеотидной последовательностью, способны осуществлять экспрессию указанной нуклеотидной последовательности. Специалисты в данной области могут понять, что можно успешно использовать различные транскрипционные промоторы, терминаторы, векторы-носители или специфические генные последовательности.
Наконец, настоящее изобретение относится к иммунологическому анализу для обнаружения HCV антитела, при котором: (1) берут олигомерную частицу или очищенный отдельный белок оболочки HCV, как определено здесь, или его функциональный эквивалент, (2) инкубируют биологический образец с указанной олигомерной частицей или указанным белком оболочки HCV в условиях, которые дают возможность образования комплекса антиген-антитело, (3) определяют, образуется ли указанный комплекс антиген-антитело, содержащий указанную олигомерную частицу или указанный белок оболочки HCV.
Далее настоящее изобретение будет проиллюстрировано ссылкой на следующие примеры, в которых изложены особенно предпочтительные воплощения. Однако следует отметить, что эти воплощения являются только иллюстративными, и их нельзя истолковывать как каким-либо образом ограничивающие изобретение.
ПРИМЕРЫ
Пример 1: Экспрессия, очистка и гомоолигомеризация в присутствии детергента HCV белка Е1
HCV белок E1s (аминокислоты 192-326) экспрессировали и очищали из клеток RK13, используя рекомбинантный вирус коровьей оспы pvHCV-11A, согласно протоколу, описанному в Maertens et al. (PCT/EP 95/03031). Кроме того, очищенный Е1 белок в 3%-ном Empigen-BB, который демонстрирует видимую молекулярную массу, соответствующую Е1 гомодимеру (примерно около 60 кДа; Фиг.1), пулировали, и пулированные фракции снова наносили на колонку для гель-хроматографии (согласно РСТ/ЕР 95/03031) и пропускали в присутствии 0,2% CHAPS или 3% бетаина. Удивительно, что, хотя E1s белок лишен своего района мембранного якоря, с обоими детергентами можно получить гомогенную популяцию специфически ассоциированных Е1 гомоолигомеров с видимой молекулярной массой 260-280 кДа (Фиг.2). Такая гомоолигомерная структура может содержать примерно 9 мономеров E1s. Должно быть ясно, что последнее является грубой оценкой, поскольку форма этого олигомера может коренным образом влиять на его видимую молекулярную массу при ее измерении с помощью гель-хроматографии. При переходе от 0,2% CHAPS к 0,05% CHAPS и повторении той же процедуры видимая молекулярная масса далее смещалась за пределы разрешающей способности колонки (пустое пространство колонки, свыше 600 кДа, Фиг.3), что позволяет предположить образование частиц. Переход от 3% бетаина к 0,1% бетаина давал популяцию E1s олигомеров со сходным поведением (данные не представлены). Можно выбрать другие детергенты, посредством которых можно достичь подобной олигомеризации в присутствии детергента. Олигомеризация, приводящая к образованию частиц, не является уникальной для CHAPS или бетаина, поскольку сходные результаты можно получить путем использования Твина-20 или Твина-80 либо октилглюказида. Кроме того, может быть возможно дальнейшее удаление детергента, что может дать возможность образования даже больших частиц. Поэтому для получения частиц присутствие детергента далее может не требоваться. Эти частицы можно получить с помощью, например, SCC без какого-либо детергента. Исключительно Е1 мономер составляет примерно 31 кДа, тогда как Е2 мономер составляет примерно 70 кДа. Эти значения, однако, могут отличаться в зависимости от состояния гликозилирования белка.
Пример 2: Анализ олигомерных структур E1s высшего порядка с помощью динамического светорассеяния
Чтобы подтвердить тот неожиданный результат, что образовались частицы, препараты E1s в 0,05%-ном CHAPS и 0,1%-ном бетаине, полученные согласно примеру 1, или в 0,1%-ном бетаине, полученные с помощью разведения препаратов в 0,5%-ном бетаине, подвергали анализу с помощью динамического светорассеяния (DLS, dynamic light scattering).
Методика динамического светорассеяния дает возможность измерять броуновское движение и соотносить его с размером частиц. Чем больше частица, тем медленнее будет броуновское движение. Скорость броуновского движения определяют по свойству, известному как коэффициент диффузии (обычно его обозначают символом D). Размер частицы вычисляют из коэффициента диффузии с помощью использования уравнения Стокса-Эйнштейна (Stokes-Einstein): d(H)=kT/3πηD, в котором d(H) представляет собой гидродинамический диаметр, k представляет собой постоянную Больцмана, Т представляет собой абсолютную температуру, η представляет собой вязкость. Отметим, что измеренный диаметр представляет собой величину, которая относится к тому, как частица диффундирует внутри жидкости. Поэтому на него ссылаются как на гидродинамический диаметр. Коэффициент диффузии выводят из функций автокорреляции (изменение интенсивности флуктуации света по времени). В этом приборе для автоматического вычисления интенсивности функции автокорреляции использован коррелятор с компьютерным управлением.
Для измерения распределений по размеру вышеуказанная функция автокорреляции скорректирована так, чтобы получить линейные кривые, и прибор снабжен компьютерной программой для анализа распределения по размеру. Однако эта методика имеет ограничивающие допущения, подобные допущениям методики, названной многоугольным лазерным светорассеянием (MALLS, multi angle laser light scattering), и ни один способ нельзя считать дающим абсолютные данные. Результаты распределений по размеру согласно DLS скорее следует интерпретировать как полуколичественные показатели полидисперсности, нежели как истинное представление этого распределения.
Образцы, содержащие E1s частицы (80-400 мкг E1s/мл 3ФР (забуференный фосфатом физиологический раствор) - 0,05% CHAPS, 0,1% или 0,5% бетаин) переносили пипеткой в измерительную ячейку прибора LSP 3.53 DLS, оборудованного 10 мВ лазером HeNe (PolymerLabs). Результат этого анализа представлен на Фиг.4 (E1s в 0,05%-ном CHAPS) и 5 (E1s в 0,1%-ном или 0,5%-ном бетаине).
Эти анализы действительно подтвердили неожиданный результат, что полученные Е1 s структуры представляют собой сферические монодисперсные частицы. E1s частицы в ЗФР/0,1% бетаин показали среднее распределение размеров 21,3±4 нм, в ЗФР/0,5% бетаин: 27,9±5 нм, тогда как в ЗФР/0,05% CHAPS для E1s получили диаметр, составляющий 12,5.
Пример 3: Анализ размера и формы с помощью электронной микроскопии
Десять мкл E1s (226 мкг/мл в ЗФР/0,05% CHAPS и 143 мкг/мл в ЗФР/3% бетаин) визуализировали стандартным негативным окрашиванием 1%-ным уранилацетатом на стабилизированных углеродом формвар (formvar)-решетках. Образец наносили на 30 секунд, а затем перед окрашиванием в течение 5 секунд и фотографированием промывали dН2О (Фиг.6).
Статистический анализ дал следующие результаты: E1s частица в CHAPS имеет средний диаметр 8,7±0,27 нм (диапазон 4,3-29,0; 95% Cl 5,4), а E1s частица в бетаине являлась менее гомогенной со средним диаметром 9,7±0,55 нм (диапазон 4,3-40,5; 95% Cl 11,0). Удивительно, что препарат в 3% бетаине, который исходно показывал молекулярную массу (ММ) 250-300 кДа при анализе с помощью SEC, показал даже большие частицы, чем препарат в 0,05% CHAPS, который исходно показывал ММ свыше 600 кДа. Поэтому авторы изобретения выдвинули гипотезу, что промежуточные гомоолигомерные формы E1s, полученные с помощью 3%-ного бетаина, могли образовать частицы высшего порядка с течением времени. Этот неожиданный эффект указывает на другие возможности получения частиц высшего порядка. Распределение частиц по размеру (Фиг.7) показывает, что препарат в CHAPS является монодисперсным, хотя у частиц большего размера наблюдается образование хвостов (вплоть до 29 нм для 0,05% CHAPS). Поскольку большие структуры переоцениваются в DLS анализах, именно присутствием этих больших частиц, хотя и в меньшем числе, можно объяснить больший диаметр, полученный с помощью DLS анализа (пример 2). Различие в диаметре можно также объяснить тем фактом, что DLS измеряет частицу в движении, тогда как электронная микроскопия измеряет статичные частицы. Должно быть ясно, что иммуногенность этих препаратов, как показано в примерах ниже, является результатом суммарного препарата и может быть результатом средних, меньших или больших частиц либо их смеси.
Пример 4: Иммунизация шимпанзе, хронически инфицированных HCV субтипа 1b
Шимпанзе (Phil), уже инфицированного в течение более 13 лет (5015 суток до иммунизации) штаммом HCV субтипа 1b, вакцинировали Е1 (аминокислоты 192-326), который имел происхождение от другого штамма генотипа 1b с 95,1% идентичностью на аминокислотном уровне (смотри также таблицу 2) и который получали, как описано в примерах 1-3. Шимпанзе получил в сумме 6 внутримышечных иммунизации, каждая по 50 мкг Е1 в ЗФР/0,05% CHAPS, смешанного с RIBI R-730 (MPLA+TDM+CWS) согласно протоколу производителя (Ribi Inc. Hamilton, MT). Эти 6 иммунизаций давали в двух сериях из трех иммунизаций с трехнедельным интервалом и с лаг-периодом 6 недель между двумя сериями. Начиная со 150 суток до иммунизации в течение периода иммунизации и до 1 года после иммунизации (но смотри ниже) проводили непрерывный мониторинг этого шимпанзе по различным параметрам, показательным для активности индуцированного HCV заболевания. Эти параметры включали в себя химический анализ крови, АЛТ, ACT (аспартатаминотрансфераза, AST), гаммаGТ, химический анализ крови, вирусную нагрузку в сыворотке, вирусную нагрузку в печени и гистологию печени. Кроме того, мониторинг иммунного ответа на иммунизацию проводили как на гуморальном, так и на клеточном уровне. В течение этого периода проводили также мониторинг животного на какие-либо вредные эффекты иммунизации, такие как изменение в поведении, клинические симптомы, масса тела, температура и местные реакции (краснота, опухание, уплотнения). Такие эффекты не были обнаружены.
Очевидно, что уровни АЛТ (и особенно гаммаGТ, данные не представлены) снижались, как только уровень антител против Е1 достигал своего максимума (Фиг.8). АЛТ восстанавливалась, как только уровни антител начинали снижаться, но гаммаGТ оставался на низком уровне, пока анти-Е1 оставались обнаружимыми.
Е2 антиген в печени снижался до почти необнаружимых уровней в течение периода, во время которого анти-Е1 были обнаружимы, и Е2 антиген восстанавливался вскоре после исчезновения этих антител. Вместе с тем, как антигены Core и Е2 становились не обнаружимыми в печени, воспаление печени значительно уменьшалось (смотри также таблицу 3). Это является основным доказательством того, что вакцина индуцирует уменьшение поражения печени, вероятно, посредством выведения, по меньшей мере частично, вирусных антигенов из их главного органа-мишени, печени.
Уровень вирусемии при измерении с помощью Amplicor HCV Monitor (Roche, Basel, Switzerland) оставался примерно неизменным в сыворотке в течение всего исследуемого периода.
Более детальные анализы гуморального ответа выявили, что максимальный конечный титр достигал 14,5×103 (после шестой иммунизации) и что этот титр падал до необнаружимого уровня через 1 год после иммунизации (Фиг.8). На Фиг.9 показано, что главные эпитопы, которые можно имитировать пептидами, распознаваемые В-клетками, расположены в N-концевом районе Е2 (пептиды V1V2 и V2V3, подробности по использованным пептидам смотри в Таблице 4). Поскольку реактивность против рекомбинантного Е1 является более высокой и продолжительной, на основании этой фигуры можно вывести заключение, что антитела, распознающие эти пептиды, представляют собой только часть общей популяции антител против Е1. Оставшаяся часть направлена против эпитопов, которые нельзя имитировать пептидами, то есть против прерывистых эпитопов. Такие эпитопы присутствуют только на полноразмерной молекуле Е1 или даже только на структуре, подобной частице. Такой иммунный ответ против Е1 является уникальным, по меньшей мере по сравнению с тем, что обычно наблюдают у людей - хронических носителей HCV (WO 96/13590 Maertens et al.) и у шимпанзе (van Doorn et al., 1996), у которых образуются анти-Е1 антитела при естественном ходе инфекции. У этих пациентов анти-Е1 частично также направлен против прерывистых эпитопов, но большая доля направлена против С4 эпитопа (±50% сыворотки пациента), а минорная доля - против V1V2 (варьируя от 2 до 70% в зависимости от генотипа), и отмечалась исключительно реактивность только против V2V3 (Maertens et al., 1997).
Анализ Т-клеточной реактивности указывал на то, что этот отдел иммунной системы также специфичным образом стимулируется вакциной, поскольку индекс стимуляции этих Т-клеток повышается от 1 до 2,5 и остается некоторым образом повышенным в течение последующего периода (Фиг.10). Именно эта Т-клеточная реактивность наблюдается только в долгосрочных клетках-респондерах при терапии интерфероном (смотри: РСТ/ЕР 94/03555, Leroux-Roels et al.; Leroux-Roels et al., 1996).
Пример 5: Иммунизация хронического носителя HCV с другим субтипом
Шимпанзе (Ton), уже инфицированного в течение более 10 лет (3809 суток до иммунизации) штаммом HCV от генотипа 1а, вакцинировали Е1 от генотипа 1b с только 79,3% идентичности на аминокислотном уровне (смотри также Таблицу 2), полученным, как описано в предыдущих примерах. Шимпанзе получал всего 6 внутримышечных иммунизаций по 50 мкг Е1 в ЗФР/0,05% CHAPS, в каждом случае смешанного с RIBI R-730 согласно протоколу производителя (Ribi Inc. Hamilton, MT). Эти 6 иммунизаций давали в двух сериях из трех иммунизаций с трехнедельным интервалом и с лаг-периодом 4 недели между двумя сериями. Начиная с 250 суток до иммунизации, в течение периода иммунизации и до 9 месяцев после иммунизации (но смотри ниже) проводили непрерывный мониторинг этого шимпанзе по различным параметрам, показательным для активности индуцированного HCV заболевания. Эти параметры включали в себя химический анализ крови, АЛТ, ACT, гаммаGТ, вирусную нагрузку в сыворотке, вирусную нагрузку в печени и гистологию печени. Кроме того, мониторинг иммунного ответа на иммунизацию проводили как на гуморальном, так и на клеточном уровне. В течение этого периода проводили также мониторинг животного на какие-либо вредные эффекты иммунизации, такие как изменение в поведении, клинические симптомы, масса тела, температура и местные реакции (краснота, опухание, уплотнения). Такие эффекты не были обнаружены.
Очевидно, что уровни АЛТ (и уровни гаммаGТ, данные не представлены) снижались, как только уровень антител против Е1 достигал своего максимума (Фиг.11). АЛТ и гаммаGТ восстанавливались, как только уровни антител начинали снижаться, однако АЛТ и гаммаGТ оставались на более низком уровне в течение всего последующего периода. Уровни АЛТ даже значительно снижались после вакцинации (62±6 ед./л) по сравнению с периодом до вакцинации (85±11 ед./л). Поскольку меньше маркеров поражения ткани выделялись из сыворотки, эти открытия были первым указанием на то, что вакцинация индуцировала улучшение в заболевании печени.
Уровни Е2 антигена становились необнаружимыми в течение периода, на протяжении которого анти-Е1 сохранял титр выше 1,0×103, но снова становились обнаружимыми при более низких уровнях Е1 антитела. Вместе с исчезновением HCV антигенов воспаление печени значительно уменьшалось от умеренного хронического активного гепатита до минимальных форм хронического персистентного гепатита (Таблица 3). Это является другим основным доказательством того, что вакцина индуцирует уменьшение поражения печени, вероятно, посредством выведения, по меньшей мере частично, вирусных антигенов из их главного органа-мишени, печени.
Уровень вирусемии при измерении с помощью Amplicor HCV Monitor (Roche, Basel, Switzerland) в сыворотке оставался на примерно одинаковых уровнях в течение всего исследуемого периода. Более детальный анализ гуморального ответа показал, что максимальный достигнутый конечный титр составлял 30×103 (после шестой иммунизации) и что этот титр падал до 0,5×103 через 9 месяцев после иммунизации (Фиг.11). На Фиг.12 видно, что главные эпитопы, которые можно имитировать пептидами и которые распознаются В-клетками, расположены в N-концевом районе Е2 (пептиды V1V2 и V2V3, подробности по использованным пептидам смотри в таблице 4). Поскольку реактивность против рекомбинантного Е1 является более высокой и продолжительной, то на основании этой фигуры можно вывести заключение, что антитела, распознающие эти пептиды, представляют собой только часть общей популяции антител против Е1. Оставшаяся часть наиболее вероятно направлена против эпитопов, которые нельзя имитировать пептидами, то есть против прерывистых эпитопов. Такие эпитопы, вероятно, присутствуют только на полноразмерной молекуле Е1 или даже только на структуре, подобной частице. Такой иммунный ответ против Е1 является уникальным, по меньшей мере по сравнению с тем, что обычно наблюдают у людей - хронических носителей HCV, которые имеют обнаружимые анти-Е1. У этих пациентов анти-Е1 частично также является прерывистым, но большая его доля направлена против С4 эпитопа (50% сыворотки пациента), а минорная доля - против V1V2 (варьируя от 2 до 70% в зависимости от генотипа), и отмечалась исключительная реактивность против V2V3 (Maertens et al., 1997). Поскольку этот шимпанзе инфицирован изолятом 1а, гуморальный иммунный ответ также оценивали на перекрестную реактивность по отношению к Е1-1а антигену. Как можно видеть на Фиг.13, такие перекрестно-реактивные антитела действительно образуются, хотя они составляют только часть общей популяции антител. Значительной является корреляция между исчезновением вирусного антигена в печени и исчезновением обнаружимых анти-1а Е1 антител в сыворотке.
Анализ Т-клеточной реактивности указывал на то, что этот отдел иммунной системы также специфично стимулируется вакциной, поскольку индекс стимуляции этих Т-клеток увеличивается от 0,5 до 5 и остается повышенным в течение последующего периода (Фиг.14).
Пример 6: Повторная ревакцинация Е1 хронических носителей HCV
Поскольку титры Е1 антител, как наблюдали в примерах 4 и 5, были нестабильными и снижались с течением времени даже до необнаружимых уровней для шимпанзе, инфицированных 1b, исследовали, можно ли снова усилить этот гуморальный иммунный ответ с помощью дополнительной ревакцинации. Обоих шимпанзе снова иммунизировали тремя последовательными внутримышечными иммунизациями с трехнедельным интервалом (50 мкг Е1, смешанного с адъювантом RIBI). Как можно судить на основании Фиг.8 и 11, анти-Е1 ответ действительно можно усилить, вирусный антиген в печени снова снижался ниже обнаружимого уровня. Вирусная нагрузка в печени оставалась постоянной даже у Ton (Фиг.11). В первое время в течение последующего периода уровень вирусемии составлял менее 105 геномных эквивалентов на мл.
Важным является открытие того, что, как уже было в случае первых серий иммунизации, шимпанзе, инфицированный штаммом HCV субтипа 1b (Phil), отвечает более низкими титрами анти-Е1, чем шимпанзе, инфицированный штаммом HCV субтипа 1а (максимальный титр в первом раунде 14,5×103 против 30×103 для Ton и после дополнительной ревакцинации только 1,2×103 для Phil против 40×103 для Ton). Хотя для обоих животных благоприятный эффект оказывается сходным, из этого эксперимента можно сделать вывод, что иммунизация хронического носителя белком Е1, имеющим происхождение от другого подтипа или генотипа, может быть особенно благоприятной для достижения более высоких титров, возможно, в обход предсуществующей и специфической иммуносупрессии, существующей у хозяина и индуцированной инфицирующим субтипом или генотипом. Альтернативно более низкие титры, наблюдаемые при гомологичном состоянии (вакцина 1b + инфекция 1b), могут указывать на связывание основной массы антител с вирусом. Следовательно, индуцированные антитела могут обладать нейтрализующей способностью.
Пример 7а: Конструирование NS3 белка путем комбинирования главных эпитопов, коррелирующих, как известно, с контролем инфекции
Кроме эпитопов в Е1, другие эпитопы также могут быть связаны с выведением HCV в течение острой фазы или с помощью терапии интерфероном. Несколько таких эпитопов локализовано в NS3 (Leroux-Roels et al., 1996; Rehermann et al., 1996 и 1997; Diepolder et al., 1995 и 1997). Два из этих главных эпитопов представляют собой CTL эпитоп, картированный Rehermann и соавторами (аминокислоты 1073-1081) и Т-клеточный (CD4) эпитоп, картированный Diepolder и соавторами (аминокислоты 1248-1261). К сожалению, эти эпитопы разбросаны по всему NS3 белку. Чтобы иметь в распоряжении по меньшей мере эти эпитопы, будет необходим относительно большой белок (аминокислоты 1073-1454). Продуцирование такого большого белка обычно идет с низкими выходами и при вакцинации может привести к ответу, очень небольшая часть которого имеет мишенью эти важные эпитопы.
Поэтому продуцирование меньшего белка было бы более подходящим решением этой проблемы. Чтобы это осуществить, некоторые из этих эпитопов нужно перерасположить в пределах такого меньшего белка. Учитывая существование другого CTL эпитопа (аминокислоты 1169-1177), который не связан с клиренсом HCV (Rehermann et al., 1996, 1997), сконструировали молекулу NS3 таким образом, что она начинается с аминокислоты 1166 и заканчивается на аминокислоте 1468 (Таблица 5). Эта конструкция уже включает в себя эпитопы, описанные Weiner и соавторами и Diepolder и соавторами. Путем мутирования района 1167-1180 в. последовательность района 1071-1084 ненужный CTL эпитоп изменили на эпитоп, который, как открыли Rehermann и соавторы, связан с клиренсом вируса. Эту конструкцию дополнительно модифицировали так, что она содержит метионин в положении 1166 для обеспечения возможности инициации трансляции. Этот метионин будет отщепляться в E.coli, поскольку за ним следует аланин. Таким образом, введение новых эпитопов, которые не присутствуют в природном NS3, ограничили до минимума. Альтернативно, если экспрессия этого белка будет затруднительна, CTL эпитоп можно сшить с С-концом на аминокислоте 1468, как детально изображено в Таблице 5.
Кодирующую последовательность HCV NS-3 фрагмента выделяли и экспрессировали, как описано в Naertens et al. (PCT/EP99/02547; клон 19b; HCV аминокислоты 1188-1468 использовали в качестве исходного материала). CTL эпитоп, как описано Rehermann, который не присутствует в фрагменте 19b NS-3, слили с этим фрагментом. Сконструировали как С-концевые, так и N-концевые слияния, поскольку на эффекты слияния по отношению к уровням экспрессии, чувствительности к протеолитическому разрушению и функциональности может влиять положение эпитопа.
Используя плазмиду pIGR12NS-3, которая представляет собой плазмиду экспрессии E.coli, экспрессирующую фрагмент 19b NS-3 под контролем левого промотора фага лямбда, в качестве матрицы для ПЦР, кодирующие последовательности NS3 19b, слитые соответственно на N- и С-конце с CTL эпитопом Rehermann (названные NS-3 19bTn и NS-3 19bТс соответственно), сначала субклонировали в вектор клонирования pGEM-T (Promega) с образованием векторов pGEN-TNS-319bTn и pGEN-TNS-319bTc. ПЦР-амплифицированные последовательности проверяли путем сиквенса ДНК.
В случае слияния последовательности Т-клеточного эпитопа с N-концевым районом NS-3 ПЦР осуществляли с длинным смысловым праймером, несущим CTL эпитоп, и коротким антисмысловым праймером, гомологичным последовательностям 3' NS-3 19b стоп-кодона. Последовательности праймеров изображены ниже.
Праймер 9038 (смысловой)
5'-GCCATGGCGACCTGCATCAACGGTGTTTGCTGGACCGTTTACCACGGTCGTGC GGCTGTTTGCACCCGTGGGGTTGCGAAGGCGGTGG-3' (SEQ ID NO 5)
Праймер 1901 (антисмысловой)
5'-TTTTATCAGACCGCTTCTGCG-3' (SEQ ID NO 6)
В случае слитого на С-конце NS-3 ПЦР осуществляли с коротким смысловым праймером, гомологичным последовательностям 5' NS-3 19b стартового кодона, и длинным антисмысловым праймером, несущим CTL эпитоп, за которым следуют стоп-кодоны в рамке считывания. Последовательности праймеров изображены ниже.
Праймер 1052 (смысловой)
5'-AGCAAACCACCAAGTGGA-3' (SEQ ID NO 7)
Праймер 9039 (антисмысловой)
5'-CTCTAGACTATTAACCGTGGTAAACGGTCCAGCAAACACCGTTGATGCAGGTC GCCAGGCTGAAGTCGACTGTCTGG-3' (SEQ ID NO 8)
Начиная с кодирующих последовательностей, клонированных в pGEM-T векторы, последовательности NS-3 19bТ встраивали в вектор экспрессии E.coli pIGRI2. Для слитого на N-конце NS-3 19bТ кодирующую последовательность NS-3 19bТ выделяли в виде 379 п.о. фрагмента Ncol/SnaBI и лигировали с фрагментами SnaBI/AllwNI и AlwNI/Ncol из вектора pIGRI2NS-3, что давало в результате вектор pIGRI2NS-3Tn. Для слитого на С-конце NS-3 19bТ кодирующую последовательность NS-3 19bТ выделяли в виде 585 п.о. фрагмента SnaBI/Spel и встраивали в открытый SnaBI/Spel вектор pIGRI2NS-3, что давало в результате вектор pIGRI2NS-3Tc.
Оба вектора pIGRI2NS-3Tn и pIGRI2NS-3Tc последовательно трансформировали в экспрессирующий штамм E.coli MC1061 (pAcl) и после температурной индукции промотора pL лямбда уровни экспрессии анализировали на ДДС-ПААГ (электрофорез в полиакриламидном геле с додецилсульфатом натрия) и Вестерн-блоте, используя поликлональную крольчью сыворотку против NS-3.
Аминокислотная последовательность белка NS-3 19bTn
MATCINGVCWTVYHGRAAVCTRGVAKAVDFVPVESMETTMRSPVFTDNSSPPAVPQTF
QVAHLHAPTGSGKSTKVPAAYAAQGYKVLVLNPSVAATLGFGAYMSKAHGVDPNIRTG
VRTTTTGAPITYSTYGKFLADGGCSGGAYDIIICDECHSIDSTSILGIGTVLDQAETAGARL
VVLATATPPGSVTVPHPNTEEVALSSTGEIPFYGKAIPIEVIKGGRHLIFCHSKKKCDELAA
KLSGFGINAVAYYRGLDVSVTPTSGDVVVVATDALMTGFTGDFDSVIDCNTCVTQTVDF
S(SEQ ID NO 1)
Аминокислотная последовательность белка NS-3 19bТс
MGVAKAVDFVPVESMETTMRSPVFTDNSSPPAVPQTFQVAHLHAPTGSGKSTKVPAAY
AAQGYKVLVLNPSVAATLGFGAYMSKAHGVDPNIRTGVRTTTTGAPITYSTYGKFLADG
GCSGGAYDIIICDECHSIDSTSILGIGTVLDQAETAGARLVVLATATPPGSVTVPHPNIEEV
ALSSTGEIPFYGCAIPIEVIKGGRHLIFCHSKKKCDELAAKLSGFGINAVAYYRGLDVSVIP
TSGDVVVVATDALMTGFTGDFDSVIDCNTCVTQTVDFSLATCINGVCWTVYHG (SEQ ID NO 2)
Нуклеотидная последовательность кодирующего района NS-3 19bТn
ATGGCGACCTGCATCAACGGTGTTTGCTGGACCGTTTACCACGGTCGTGCGGCTGTT
TGCACCCGTGGGGTTGCGAAGGCGGTGGACTTTGTACCCGTAGAGTCTATGGAAACC
ACCATGCGGTCCCCGGTCTTTACGGATAACTCATCTCCTCCGGCCGTACCGCAGACA
TTCCAAGTGGCCCATCTACACGCCCCCACTGGTAGTGGCAAGAGCACTAAGGTGCCG
GCTGCATATGCAGCCCAAGGGTACAAGGTACTTGTCCTGAACCCATCCGTTGCCGCC
ACCTTAGGATTCGGGGCGTATATGTCTAAAGCACATGGTGTCGACCCTAACATTAGA
ACTGGGGTAAGGACCATCACCACGGGCGCCCCCATTACGTACTCCACCTACGGCAA
GTTTCTTGCCGACGGTGGTTGCTCTGGGGGCGCTTACGACATCATAATATGTGATGA
GTGCCACTCGATTGACTCAACCTCCATCTTGGGCATCGGCACCGTCCTGGATCAGGC
GGAGACGGCTGGAGCGCGGCTTGTCGTGCTCGCCACTGCTACACCTCCGGGGTCGGT
CACCGTGCCACATCCCAACATCGAGGAGGTGGCTCTGTCCAGCACTGGAGAGATCCC
CTTTTATGGCAAAGCCATCCCCATCGAGGTCATCAAAGGGGGGAGGCACCTCATTTT
CTGCCATTCCAAGAAGAAATGTGACGAGCTCGCCGCAAAGCTATCGGGCTTCGGAA
TCAACGCTGTAGCGTATTACCGAGGCCTTGATGTGTCCGTCATACCGACTAGCGGAG
ACGTCGTTGTTGTGGCAACAGACGCTCTAATGACGGGCTTTACCGGCGACTTTGACT
CAGTGATCGACTGTAACACATGCGTCACCCAGACAGTCGACTTCAGCTAA (SEQ ID NO 3)
Нуклеотидная последовательность кодирующего района NS-3 19bТс
ATGGGGGTTGCGAAGGCGGTGGACTTTGTACCCGTAGAGTCTATGGAAACCACC
ATGCGGTCCCCGGTCTTTACGGATAACTCATCTCCTCCGGCCGTACCGCAGACAT
TCCAAGTGGCCCATCTACACGCCCCCACTGGTAGTGGCAAGAGCACTAAGGTGC
CGGCTGCATATGCAGCCCAAGGGTACAAGGTACTTGTCCTGAACCCATCCGTTGC
CGCCACCTTAGGATTCGGGGCGTATATGTCTAAAGCACATGGTGTCGACCCTAAC
ATTAGAACTGGGGTAAGGACCATCACCACGGGCGCCCCCATTACGTACTCCACCT
ACGGCAAGTTTCTTGCCGACGGTGGTTGCTCTGGGGGCGCTTACGACATCATAAT
ATGTGATGAGTGCCACTCGATTGACTCAACCTCCATCTTGGGCATCGGCACCGTC
CTGGATCAGGCGGAGACGGCTGGAGCGCGGCTTGTCGTGCTCGCCACTGCTACA
CCTCCGGGGTCGGTCACCGTGCCACATCCCAACATCGAGGAGGTGGCTCTGTCCA
GCACTGGAGAGATCCCCTTTTATGGCAAAGCCATCCCCATCGAGGTCATCAAAGG
GGGGAGGCACCTCATTTTCTGCCATTCCAAGAAGAAATGTGACGAGCTCGCCGC
AAAGCTATCGGGCTTCGGAATCAACGCTGTAGCGTATTACCGAGGCCTTGATGTG
TCCGTCATACCGACTAGCGGAGACGTCGTTGTTGTGGCAACAGACGCTCTAATGA
CGGGCTTTACCGGCGACTTTGACTCAGTGATCGACTGTAACACATGCGTCACCCA
GACAGTCGACTTCAGCCTGGCGACCTGCATCAACGGTGTTTGCTGGACCGTTTAC
CACGGTTAA (SEQ ID NO 4)
Пример 7б: Очистка белков NS-3 19bTn и NS-3 19bТс
Клеточные массы Е.соli из культур Эрленмейера разрушали с помощью клеточного дезинтегратора (CSL, модель В) при 1,4 кбар (1,4·105 кПа) в 50 мМ Трис, рН 8. Этот лизат осветляли с помощью центрифугирования (15000 g, 30 мин, 4°С). Супернатант отбрасывали, поскольку N- и С-концевую конструкцию NS3 выделяли из осадка. Этот осадок делали высоко стабильным для N-концевой конструкции, производя тщательную отмывку (первая отмывка: 2% саркозил, 0,5 М гуанидинийхлорид и 10 мМ ДТТ (дитиотреитол), вторая и третья отмывка: 1% Тритон Х-100, 0,5 М гуанидинийхлорид и 10 мМ ЭДТА) перед солюбилизацией. В случае С-концевой конструкции этого не делали. Очистку N-концевой конструкции продолжали далее. Отмытый осадок в конце растворяли в 6 М гуанидинийхлорид/50 мМ Na2HPO4 при рН 7,2 и сульфонировали, как описано в Maertens et al. (PCT/EP99/02547). Сульфонированный осадок сначала высаливали на колонке Sephadex G25 до 6 М мочевинf/50 мМ триэтаноламин, рН 7,5 и наконец очищали с помощью двух последовательных анионообменных хроматографий в том же составе буфера. Первый анионный обмен проводили на колонке Hyper DQ (50 мкМ) (BioSpra Inc.Marlborough, Ma. USA) и выделяли NS3 между 0,11 и 0,19 М NaCl. После разведения эти фракции наносили на вторую колонку Hyper DQ (20 мкМ) (BioSpra Inc. Narlborough, Na. USA) и выделяли NS3 во фракциях, содержащих 0,125 М NaCl. Эти фракции высаливали до 6 М мочевины в ЗФР, рН 7,5. На основании ДДС-ПААГ с последующим окрашиванием серебром установили, что конечная чистота составляла свыше 90%. N-концевое секвенирование с помощью деградации EDMAN показало, что этот NS3 имеет интактный N-конец, в котором желаемый эпитоп присутствует в правильной последовательности. Подтвердили также, что метионин, используемый для старта трансляции, отщеплен, как предсказывали.
Пример 8: Конструирование белка Е2 без гипервариабельного района I
Был отмечен иммунодоминантный гомологичный ответ на HVR I район Е2. Этот ответ не может найти широкого применения при разработке вакцины, поскольку вакцина представляет собой гетерологичное состояние (вакцинный штамм всегда отличается от полевых штаммов). Следовательно, делеция этого района будет необходима, чтобы иметь белок Е2, индуцирующий антитела против более консервативных, но менее иммуногенных районов Е2. Путем тщательного анализа лидерной последовательности Е2 и гипервариабельного района Е2 разработали идеальную конструкцию для экспрессии белка Е2 без HVR I. Эта конструкция дает возможность экспрессии пептида Е2, начиная с положения аминокислоты 409 вместо аминокислоты 384. В качестве лидерной последовательности использовали 20 С-концевых аминокислот Е1. Однако, поскольку описание этого HVR не является однозначным, то получили вторую версию (начиная с аминокислоты 412), которая также имеет высокую вероятность расщепления в правильном положении.
Промежуточная конструкция pvHCV-99 (смотри также Фиг.15 и 16)
В экспрессирующей кассете кодирующей последовательности Е2-715 должен предшествовать лидерный сигнальный пептид Е1, начинающийся с Met364. Поэтому в плазмиде pvHCV-92 (Фиг.15), которая содержит кодирующую последовательность Е2-715 HCV типа 1b с длинной версией сигнального пептида Е1 (начиная с Met347), получали делецию путем двойного ферментативного гидролиза EcoRI и Ncol с последующей достройкой 5' выступающего конца реакцией с Т4 ДНК полимеразой. Лигирование полученных тупых концов (рециркуляризация 6621 п.о. фрагмента) в результате давало плазмиду pvHCV-99, которая кодирует тот же белок (Е2-715) с более коротким лидерным сигнальным пептидом Е1 (начиная с Net364), эта плазмида pvHCV-99 была депонирована в списке штаммов как ICCG 3635. Должно быть ясно, что можно использовать HCV или гетерологичные сигнальные последовательности варьирующей длины.
Плазмиды pvHCV-100 и -101 должны содержать делецию в последовательности Е2, то есть делецию гипервариабельного района I (HVR-I). В плазмиде pvHCV-100 делетировали аминокислоты 384 (His)-408 (Ala), тогда как в плазмиде pvHCV-101 делетировали аминокислоты 384 (His)-411 (IIe).
Конструирование pvHCV-100
Для конструирования pvHCV-100 сконструировали два олигонуклеотида:
HCV-рr 409 [8749]:
5'-СТT TGC CGG CGT CGA CGG GCA GAA AAT CCA GCT CGT AA - 3' (SEQ ID NO 9)
HCV-рr 408 [8750]:
5'-TTA CGA GCT GGA TTT TCT GCC CGT CGA CGC CGG CAA AG - 3' (SEQ ID NO 10)
ПЦР амплификация (денатурация 5 мин 95°С, 30 циклов амплификации, состоящих из отжига при 55°С, полимеризации при 72°С и денатурации при 95°С в течение 1 мин каждого, элонгация в течение 10 мин при 72°С) матрицы pvHCV-99 с Gpt-рr [3757] и HCV-рr 408 [8750] давала в результате 221 п.о. фрагмент, тогда как амплификация с HCV-рг 409 [8749] и ТКr-рr [3756] давала в результате 1006 п.о. фрагмент. Оба ПЦР фрагмента перекрываются друг с другом на 19 нуклеотидов. Эти два фрагмента объединяли и амплифицировали с помощью ПЦР с праймерами Gpt-рr [3757] и ТКr-рr [3756]. Полученный 1200 п.о. фрагмент подвергали ферментативному гидролизу по EcoRI и HinDIII и лигировали в вектор pgsATA18 [ICCG 1998] (5558 п.о.), подвергнутый ферментативному гидролизу по EcoRI/HinDIII. Эту конструкцию, pvHCV-100, проанализировали с помощью рестрикционного анализа и сиквенс-анализа и депонировали в перечне штаммов как ICCG 3636.
Конструирование pvHCV-101
Для конструирования pvHCV-101 сконструировали два олигонуклеотида:
HCV-рr 411 [8747]:
5'-CTT TGC CGG CGT CGA CGG GCA GCT CGT AAA CAC CAA CG - 3' (SEQ ID NO 11)
HCV-рr 410 [8748]:
5'-CGT TGG TGT TTA CGA GCT GCC CGT CGA CGC CGG CAA AG - 3' (SEQ ID NO 12)
ПЦР амплификация матрицы pvHCV-99 с Gpt-рr [3757] и HCV-рr 410 [8748] давала в результате 221 п.о. фрагмент, тогда как амплификация с HCV-рr 411 [8747] и ТКr-рr [3756] давала в результате 997 п.о. фрагмент. Оба ПЦР фрагмента перекрываются друг с другом на 19 нуклеотидов. Эти два фрагмента объединяли и амплифицировали с помощью ПЦР с праймерами Gpt-рr [3757] и ТКr-рr [3756]. Полученный 1200 п.о. фрагмент подвергали ферментативному гидролизу по EcoRI и HinDIII и лигировали в вектор pgsATA18 [ICCG 1998] (5558 п.о.), подвергнутый ферментативному гидролизу по EcoRI/HinDIII. Эту конструкцию, pvHCV-100, анализировали с помощью рестрикционного анализа и сиквенс-анализа и депонировали в перечне штаммов как ICCG 3637.
Все плазмиды проверяли сиквенс-анализом и депонировали в перечне штаммов Innogenetics. Для каждой плазмиды получали два мини-ДНК препарата (PLASmix) в стерильных условиях и пулировали. Определяли концентрацию ДНК и осуществляли QA с помощью рестрикционного анализа. Очищенную ДНК использовали для создания рекомбинантного вируса коровьей оспы, как описано в Maertens et al. (PCT/EP95/03031). Рекомбинантные вирусы vvHCV-100 и vvHCV-101 создавали, однако, на проверенных WHO (Всемирная организация здравоохранения) клетках Vero. После двух раундов очистки бляшек продукт экспрессии анализировали с помощью Вестерн-блот-анализа, как описано в Naertens et al. (PCT/EP95/03031). Белки визуализировали с помощью специфического моноклонального анти-Е2 антитела (IGH 212, которое можно получить от авторов изобретения в Innogenetics N.V., Zwijnaarde, Belgium) с установленной молекулярной массой 69 и 37 кДа для vvHCV-100 и 68 и 35 кДа для vvHCV-101. Эти молекулярные массы указывают на присутствие как гликозилированного, так и негликозилированного белка Е2, что подтвердили с помощью обработки образцов РNGазойF перед Вестерн-блот-анализом. Эта обработка приводит к обнаружению только одного отдельного белка 37 кДа и 35 кДа для vvHCV-100 и vvHCV-101 соответственно.
Аминокислотная последовательность зрелого Е2. полученного из pvHCV-100
QKIQLVNTNGSWHINRTALNCNDSLQTGFFAALFYKHKFNSSGCPERLASCRSIDKFAQG
WGPLTYTEPNSSDQRPYCWHYAPRPCGIVPASQVCGPVYCFTPSPVVVGTTDRFGVPTY
NWGANDSDVLILNNTRPPRGNWFGCTWMNGTGFTKTCGGPPCNIGGAGNNTLTCPTDC
FRKHPEATYARCGSGPWLTPRCMVHYPYRLWHYPCTVNFTIFKVRMYVGGVEHRFEAA
CNWTRGERCDLEDRDRSELSPLLLSTTEWQILPCSFTTLPALSTGLIHLHQNTVDVQYLYG
VGSAVVSLVIK (SEQ ID NO 13)
Аминокислотная последовательность зрелого Е2, полученного из pvHCV-101
QLVNTNGSWHINRTALNCNDSLQTGFFAALFYKHKFNSSGCPERLASCRSIDKFAQGWG
PLTYTEPNSSDQRPYCWHYAPRPCGIVPASQVCGPVYCFTPSPVWGTTDRFGVPTYNW
GANDSDVLILNNTRPPRGNWFGCTWMNGTGFTKTCGGPPCNIGGAGNNTLTCPTDCFR
KHPEATYARCGS GPWLTPRCMVHYPYRLWHYPCTVNFTIFKVRMYVGGVEHRFEAAC
NWTRGERCDLEDRDRSELSPLLLSTTEWQILPCSFTTLPALSTGLIHLHQNIVDVQYLYGV
GSAWSLVIK(SEQ ID NO 14)
Пример 9: Частица Е1 с дополнительно улучшенной иммуногенностью
Как изложено в примере 1, белок E1s очищали согласно протоколу, описанному в РСТ/ЕР 95/03031 Naertens et al. Этот протокол включает в себя ковалентную модификацию цистеинов (свободные цистеины и цистеины, вовлеченные в образование межмолекулярных мостиков, в последнем случае после восстановления этих цистеиновых мостиков с использованием DTT (дитиотреитол)), используя производные малеимида (N-этилмалеимид и биотин-малеимид, оба получены от Sigma). В качестве альтернативного способа блокирования малеимидом также оценивали активные галогены. Эти соединения, то есть активные галогены, блокируют свободные цистеины посредством алкилирования. В качестве примера оценивали активный галоген (иодацетамид, Merck). Тот же протокол использовали для очистки Е1, как описано в Naertens et al. (РСТ/ЕР 95/03031), но вместо малеимидных соединений использовали иодацетамид. Белок E1s, полученный с помощью этой методики, в течение всей очистки вел себя подобно белкам, блокированным малеимидом. При конечном снижении концентрации детергента до 0,05% CHAPS или при переходе к 0,5%-ному бетаину, как описано в примере 1, получали сходные частицы, как определяли с помощью DLS. Однако при иммунизации мышей E1s, модифицированным ацетамидом, был обнаружен неожиданный эффект.
В сумме три серии по 6 мышей в каждой иммунизировали E1s, используя три инъекции с трехнедельным интервалом, причем каждая инъекция состояла из 5 мкг E1s в концентрации 100 мкг/мл в 3ФР, смешивая с равным объемом адъюванта RIBI (R-700). Первая серия получала Е1-малеимид в виде препарата в 0,05%-ном CHAPS, вторая серия получала Е1-ацетамид также в виде препарата в 0,05%-ном CHAPS, тогда как третья серия получала Е1-ацетамид в виде препарата в 0,12%-ном бетаине. Затем у всех мышей брали кровь через 10 суток после третьей иммунизации. Конечные титры (определяемые как разведение сыворотки, все еще дающее в результате ОП в 2 раза выше, чем фоновые значения) для каждого животного индивидуально определяли против Е1-малеимида и Е1-ацетамида. На Фиг.17 показаны эти конечные титры, представленные в виде среднего со стандартными отклонениями. У мышей, которые получали Е1-малеимид, происходило только образование антител, которые были способны распознавать малеимидсодержащие эпитопы (на Е1-ацетамид реактивности вообще не было), у мышей, которые получали Е1-ацетамид, с очевидностью происходило образование антител против действительных Е1 эпитопов, поскольку эти антитела являются реактивными как против Е1-ацетамида, так и против Е1-малеимида. Это ясно продемонстрировали в дополнительном эксперименте, в котором антитела на специфические районы Е1 определяли, используя пептиды, которые не были модифицированы ни ацетамидом, ни малеимидом. Эти результаты, как показано на Фиг.18, демонстрируют, что именно у мышей, иммунизированных Е1-ацетамидом (в виде препаратов в CHAPS и бетаине), происходило образование антител, которые способны распознавать пептиды V1V2, V2V3, V3V4, V5C4, C4V6. Поскольку V6 не является частью E1s, то авторы изобретения заключили, что антитела продуцировались против С4, V3 (V3V4 является позитивным, тогда как V4V5 не является позитивным) и V1V2. У мышей, иммунизированных E1s-малеимидом, развивался только очень низкий ответ против пептидов V1V2 и V2V3. Это еще больше подтверждает тот факт, что умеренно высокий титр, измеренный для этих мышиных антител против малеимид-E1s, главным образом направлен против малеимид-зависимых эпитопов. Кроме того, авторы изобретения смогли доказать, что ответ, индуцированный E1s-ацетамидом, отчасти представляет собой ответ Th1 типа, поскольку существенное количество индуцированных антител принадлежат к субтипу IgG2(a+b). Количество IgG2 даже выше для бетаинового препарата по сравнению с препаратом CHAPS (Фиг.19). Из этих результатов заключили, что белки оболочки HCV, в которых по меньшей мере один цистеин (но возможно более чем один цистеин) алкилирован, являются в высшей степени иммуногенными белками.
В результате модифицированный ацетамидом Е1 в виде препарата в бетаине также использовали для ревакцинации шимпанзе Phil и Ton. Обоих шимпанзе иммунизировали повторно двумя последовательными внутримышечными иммунизациями с трехнедельным временным интервалом (50 мкг Е1, смешанного с адъювантом RIBI, как в примерах 4 и 5). Как можно судить из Фиг.20 и 21, анти-Е1 ответ действительно можно снова усилить и усилить до более высоких уровней, чем те, что были получены при предшествующих иммунизациях после двух инъекций. Это титрование проводили против стандарта, который представляет собой смесь из трех человеческих анти-Е1 сывороток высокого титра (полученных от хронических носителей HCV). Титр анти-Е1 этих сывороток определяли как одна единица/мл. У шимпанзе Phil (Фиг.20) только после двух иммунизаций индуцировались титры в два раза выше, чем у людей-носителей. У шимпанзе Ton (Фиг.21) индуцировались титры, более высокие вплоть до 140-кратного увеличения. Это еще раз подтверждает высокую иммуногенность этих E1-частиц.
Пример 10: Алкилированный Е1 обладает превосходными качествами для диагностического применения
Е1 s-ацетамид, как описано в примере 9, далее оценивали в качестве антигена для обнаружения анти-Е1 антител в образцах сыворотки от людей - хронических носителей HCV. В качестве примера эти антигены связывали с LIA-мембранами и полоски обрабатывали по существу так, как описано в Zrein et al. (1998). Образцы сыворотки от 72 доноров крови сначала оценивали, чтобы определить оптимальную концентрацию Е1 антигена, которую можно использовать в анализе, чтобы исключить “ложные” позитивы. Для E1s-малеимида эта концентрация оказалась равной 8 мкг/мл, тогда как для E1s-ацетамида концентрация вплоть до 50 мкг/мл не приводила к ложным позитивным результатам (ни один из образцов не показывал относительного цветного окрашивания выше 0,5). Используя 8 и 50 мкг/мл соответственно для E1s-малеимида и Е1s-ацетамида, 24 сыворотки хронических носителей HCV подвергали скринингу на антитела против E1s. Как показано в Таблице 6, E1s-ацетамид в результате дает больше образцов, оцениваемых позитивно (67% против 38% для E1s-малеимида). Не обнаружили ни одного образца, который оценивали позитивно только для E1s-малеимида. Для образцов, оцениваемых позитивно как для E1s-малеимида, так и для E1s-ацетамида, реактивность последнего была более высокой. Из этого примера можно заключить, что алкилированные белки оболочки HCV являются лучшими антигенами для обнаружения человеческих антител, чем модифицированные малеимидом белки оболочки.
Пример 11: Получение смешанных частиц, содержащих Е1 и Е2
E1s и E2s (vvHCV-44) продуцировали и очищали, как описано в Maertens et al., PCT/EP95/03031, за исключением того факта, что модификацию малеимидом заменили алкилированием при использовании иодацетамида. E1s и E2s в 3%-ном Empigen отдельно или в эквимолярной смеси впрыскивали в колонку Superdex-200 PC 3.2/30, уравновешенную ЗФР/0,2% CHAPS. Эта колонка предназначена для использования с ВЭЖХ оборудованием SNART™ от Pharmacia LKB (Sweden). Фракции подвергали скринингу с помощью трех различных сандвич-ELISA (твердофазных иммуноферментных анализов). Для этих ELISA специфические моноклональные антитела E1-(IGH 207) и E2-(IGH 223) наносили в концентрации 2 мкг/мл. Фракции из гель-фильтрации инкубировали в разведении 1/2500. Два других моноклональных антитела Е1 (IGH 200) и Е2 (IGH 212), конъюгированные с биотином, использовали для обнаружения связанного антигена. Систему стрептавидин-HRP (пероксидаза из хрена)/ТМВ (3,3',5,5'-тетраметилбензидин) использовали для проявления связанного биотина в желтом цвете, который измеряли при 450 нм.
Эту систему ELISA использовали при гомологичной (анти-Е1 покрытие/анти-Е1 обнаружение или анти-Е2 покрытие/анти-Е2 обнаружение) и гетерологичной постановке (анти-Е1 покрытие/анти-Е2 обнаружение). Последняя теоретически обнаруживает только частицы, в которые включены и Е1, и Е2. Реактивные фракции пулировали, концентрировали на 10 кДа фильтре и снова подвергали хроматографии на Superdex-200 в ЗФР/0,05% CHAPS. Все эти фракции тестировали на реактивность с помощью использования различных постановок ELISA. Как можно судить из Фиг.22, добавление Е2 к Е1 не приводит к большому сдвигу во времени удерживания по сравнению с одним Е1, что указывает на то, что частицы действительно все еще присутствуют. Эти частицы содержат и Е1, и Е2, поскольку только в этой постановке гетерологичную ELISA оценивают позитивно.
Пример 12: Очистка смешанных частиц, содержащих Е1 от 2 различных генотипов
E1s генотипа 1b и генотипа 4 (vvHCV-72) продуцировали и очищали, как описано в Maertens et al., PCT/EP95/03031, за исключением того факта, что модификацию малеимидом заменили алкилированием при использовании иодацетамида для генотипа 1b. E1s-1b и E1s-4 в 3% Empigen отдельно или в эквимолярной смеси впрыскивали в колонку Superdex-200 PC 3.2/30, уравновешенную ЗФР/0,2% CHAPS. Эта колонка предназначена для использования с ВЭЖХ оборудованием SNART™ от Pharmacia LKB (Sweden). Основные фракции, содержащие белок, пулировали, концентрировали на 10 кДа фильтре и снова подвергали хроматографии на Superdex-200 в ЗФР/0,05% CHAPS. Все эти фракции тестировали на реактивность с помощью использования постановки ELISA, которая должна обнаруживать только частицы, содержащие Е1 от обоих генотипов. Для этого ELISA-стрептавидин наносили в концентрации 2 мкг/мл. Фракции из гель-фильтрации инкубировали в разведении 1/2500. Для обнаружения связанного антигена использовали моноклональное Е1 антитело (IGH 200), которое распознает только Е1 от генотипа 1 и 10. Систему коза-против-мыши-HRP/TMB использовали для проявления этого анализа в желтом цвете, который измеряли при 450 нм. Как можно судить из Фиг.23, добавление Е1-4 к Е1-1b не приводит к большому сдвигу во времени удерживания белков по сравнению с одним Е1, что указывает на то, что частицы дейcтвительно все еще присутствуют. Эти частицы содержат оба белка Е1, то есть E1s генотипа 1b и генотипа 4, поскольку только в этой постановке гетерологичную ELISA оценивают позитивно.
ПЕРЕЧЕНЬ ССЫЛОК
Deleersnyder V., Pillez A., Wychowski С., Blight К., Xu J., Hahn Y.S., Rice C.M., Dubuisson J. Formation of native hepatitis С virus glycoprotein complexes. J. Virol. 1997: 71:697-704.
Diepolder H.M., Zachoval R., Hoffmann R.M., Wierenga E.A., Santantonio Т., Jung M.C., Eichenlaub D., Pape G.R. Possible mechanism involving T-lymphocyte response to non-structural protein 3 in viral clearance in acute hepatitis С virus infection. Lancet 1995: 346: 1006-1007.
Diepolder H.M., Gerlach J.T., Zachoval R., HoffNann R.M., Jung M.C., Wierenga E.A., Scholz S., Santantonio Т., Houghton M., Southwood S., Sette A., Pape G.R. Immunodominant CD4+ T-cell epitope within non-structural protein 3 in acute hepatitis С virus infection. J. Virol., 1997: 71: 6011-6019.
Fancy D.A., Melcher K., Johnston S.T. and Kodadek T. New chemistry for the study of multiprotein complexes: the six-histidine tag as a receptor for a protein crosslinking reagent. Chem. Biol. (1996) 3: 551-559.
G.T.Hermanson in Bioconjugate Techniques (1996) Part I section 1.43 and section 2.2.1, Academic Press San Diego CA, USA.
Houghton M. Immunity to HCV: The case for vaccine development. 4th International meeting on hepatitis С Virus and related viruses. Satellite Symposium: New approach to prevention and therapy of HCV infection. March 7 1997, Kyoto, Japan.
Leroux-Roels G., Esquivel C.A., DeLeys R., Stuyver L, Elewaut A., Philippe J., Desombere I., Paradijs J., Maertens G. Lymphoproliterative responses to hepatitis С virus core, E1, E2, and NS3 in patients with chronic hepatitis С infection treated with interferon alfa. Hepatology 1996: 23: 8-16.
Maertens G. and Stuyver L. Genotypes and genetic variation of hepatitis virus. In: The molecular medicine of viral hepatitis. Ed: Harrison T.J. and Zuckerman A.J. 1997.
Major M.E. and Feinstone S.M. The molecular virology of hepatitis C. Hepatology 1997: 25: 1527-1538.
Maertens G., Depla E., Ducatteeuw A., Vandeponseele P., Bosman F., Venneman A., de Martynoff G., Stuyver L, Dekeyser F., Vandeperre В., Zrein M. and Buyse M. - A. Hepatology 1997: 26: 186A.
Rehermann В., Chang K.M., McHutchinson J., Kokka R., Houghton M., Rice C.M., Chisari F.V. Differential cytotoxic T-lymphocyte responsiveness to the hepatitis В and С viruses in chronically infected patients. J. Virol. 1996 70: 7092-7102.
Rehermann В., Takaki A., Liebetrau A., Luda S., Seifert U., Salha K., Manns M., Wiese M. Characterization of the cytotoxic and helper Т cell response in patients 18 years after a single source outbreak of HCV infection. Hepatology, 1997: 26: 406A.
Sambrook J., Fritsch E.F., and Maniatis T. (1989) Nolecular Cloning, a laboratory manual, second edition. Cold Spring Harbor University Press, Cold Spring Harbor, NY USA.
Van Doorn L.J., Kleter В., Pike I., Quint W. Analysis of hepatitis С virus isolates by serotyping and genotyping. J. Clin. Microbiol. 1996: 34: 1784-1787.
Villa E., Buttafoco P., Grottola A., Scarcelli A., Giannini F., Manerti F. Neutralizing antibodies against HCV: liver transplant as an experimental model. J. Hepatol. 1998:28:
Weiner A.J., Erickson A.L, Kansopon J., Crawford K., Muchmore E., Houghton M., Walker C.M. Association of cytotoxic T lymphocyte (CTL) escapemutations with persistent hepatitis С virus (HCV) infection. Princess Takamatsu Symp, 1995:25:227-235.
Yi M., Nakamoto Y., Kaneko S., Yamashita Т., Murakami S. Delineation of regions important for heteromeric association of hepatitis С virus E1 and E2. Virol. 1997a 231: 119-129.
Zauberman A., Nussbaum O., llan E., Eren R,, Ben-Moshe O., Arazi Y., Berre S., Lubin I., Shouval D., Galun E., Reisner Y. and Dagan S. The trimera mouse system: a mouse model for hepatitis С infection and evaluation of therapeutic agents. June 6-9, 1999; Oral 4.3. In: 6th International Symposium on Hepatitis С & Related Viruses. Bethesda USA.
Zrein M., Louwagie J., Boeykens H., Covers L., Hendrickx G., Bosman F., Sablon E., Demarquilly C., Boniface M. and Saman E. (1998). Assessment of a new immunoassay for serological confirmation and discrimination of human T-cell lymphotropic virus infections. Clin. Diagn. Lab. Imm. 5: 45-49.
Пример 13 Получение олигомерных частиц, состоящих из частей белков оболочки HCV
E1s белок HCV (аминокислоты 192-326) был экспрессирован в клетках Vero с использованием рекомбинантного вируса HCV11B. Белок был очищен по существу, как описано в примере 9. После очистки 3% empigen-BB был заменен на 3% бетаин с помощью хроматографии с исключением по размеру, как описано в Примере 1. В результате этого процесса получают E1s в виде частиц.
E2deltaHVRI (аминокислоты 412-715) экспрессировали и очищали из Vero по существу также, как описано для Е1, с использованием рекомбинантного вакцинного вируса HCV101, который был получен рекомбинантным путем из pvHCV-101, описанной в Примере 8, и вакцинного вируса дикого типа. E2deltaHVRI также ведет себя как частица (что измерено динамически светорассеянием) после замены empigen на бетаин.
У шимпанзе, иммунизированных такими частицами, был зарегистрирован иммунный ответ.
Были получены также частицы, состоящие из другой части белка Е2, а именно из аминокислот 384-673. Сконструированной соответствующим образом плазмидой были трансформированы клетки Hansenula polymorpha. Из экспрессированных белков с помощью хроматографии с исключением по размеру были получены частицы, размер которых, определенный динамическим светорассеянием, составлял 20-56 нм.
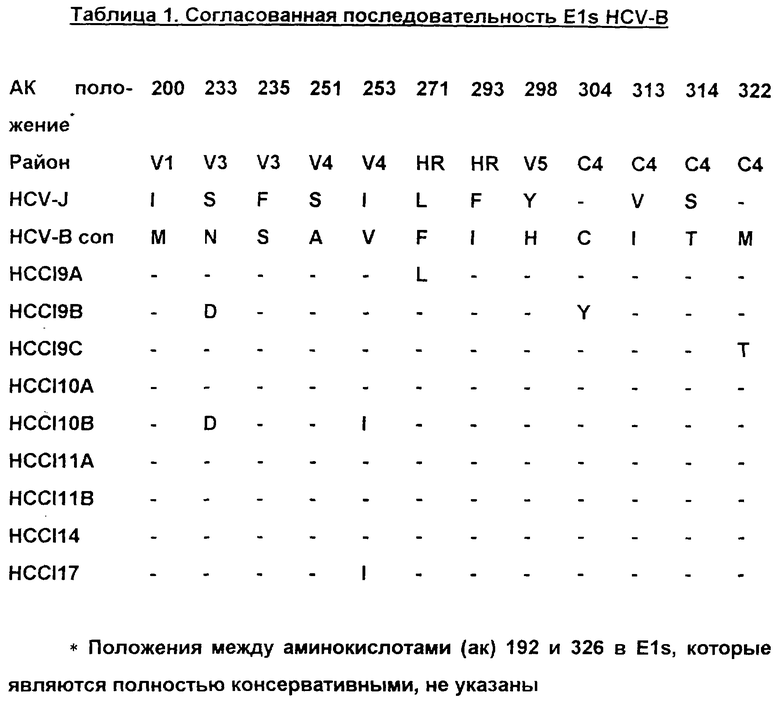

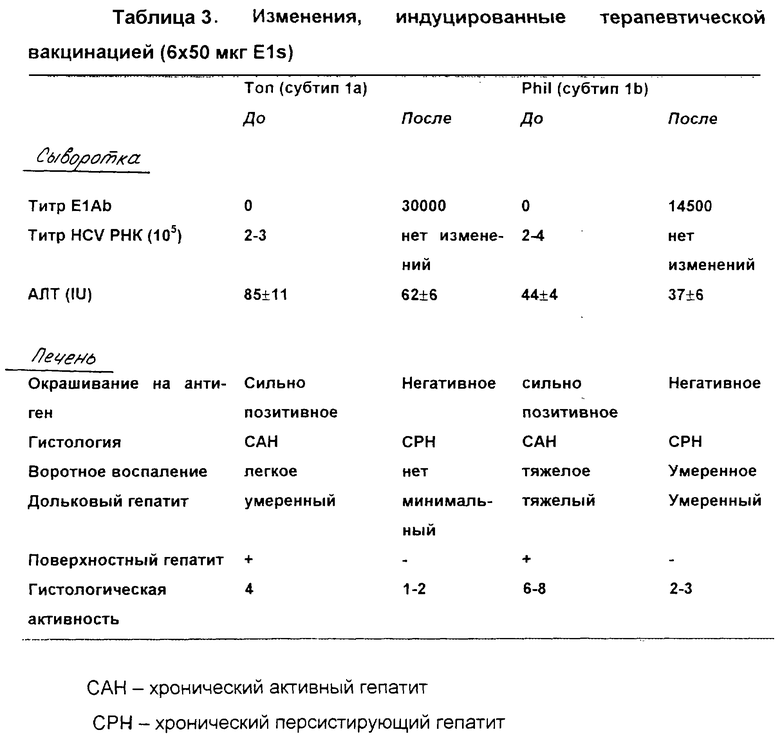
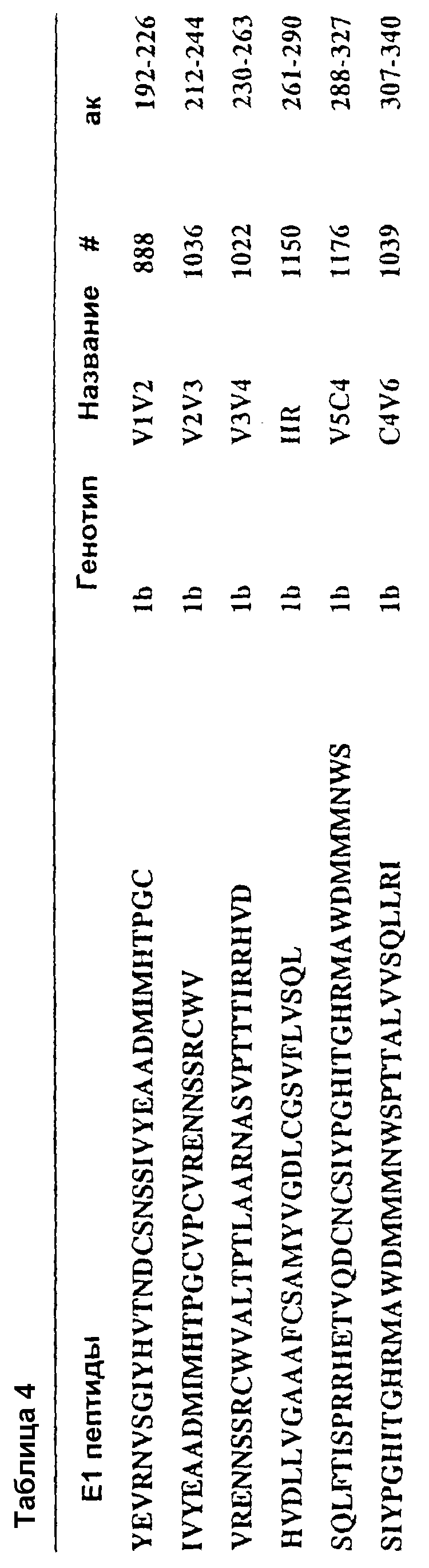

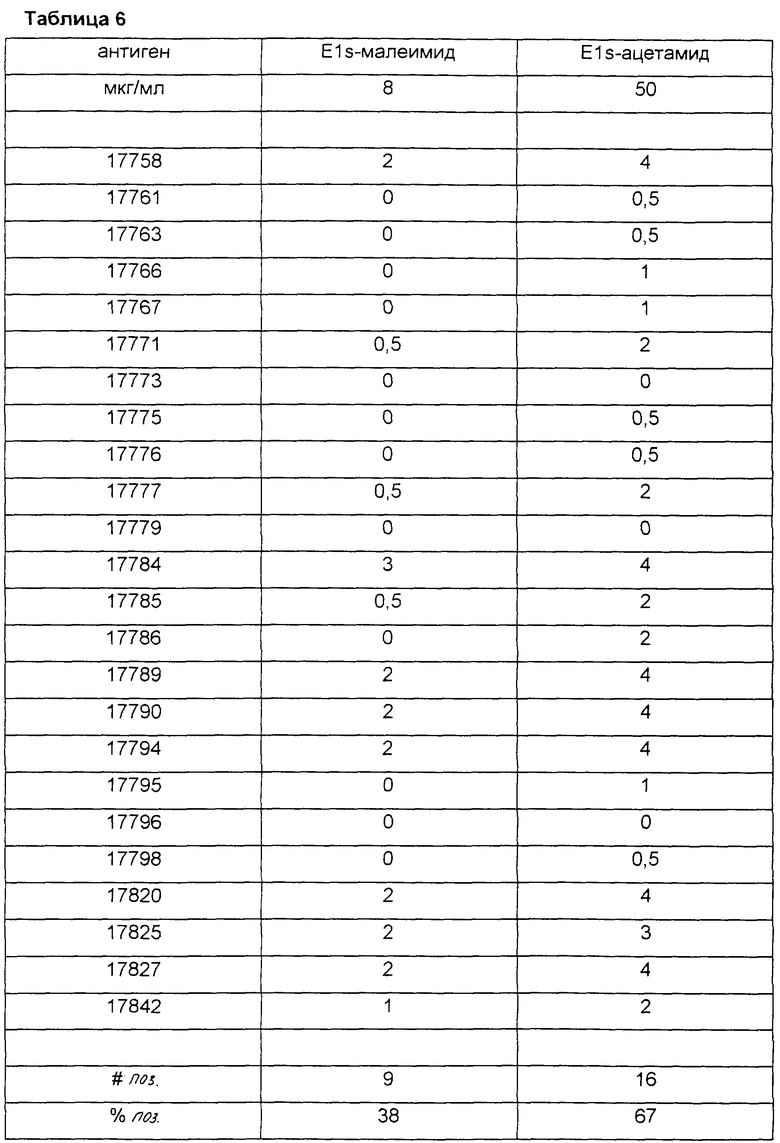
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЕ АНТИГЕНЫ ДЛЯ ВАКЦИНЫ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2013 |
|
RU2639504C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2351363C2 |
| ВЫДЕЛЕННЫЙ ОБОЛОЧЕЧНЫЙ БЕЛОК HCV, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЛЕКАРСТВО, ВАКЦИНА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) ЕГО СОДЕРЖАЩИЕ | 2002 |
|
RU2274643C2 |
| РЕКОМБИНАНТНЫЙ МОДИФИЦИРОВАННЫЙ ВИРУС ВАКЦИНЫ АНКАРА, СПОСОБНЫЙ ЭКСПРЕССИРОВАТЬ СТРУКТУРНЫЕ АНТИГЕНЫ ВИРУСА ГЕПАТИТА С | 2002 |
|
RU2270860C2 |
| ВАКЦИННЫЙ СОСТАВ, ПОТЕНЦИРОВАННЫЙ КОМБИНАЦИЕЙ ДНК И АНТИГЕНА | 2002 |
|
RU2294212C2 |
| СПОСОБ НЕЙТРАЛИЗАЦИИ ВИРУСА ГЕПАТИТА С, ПОЛНОСТЬЮ ЧЕЛОВЕЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ ВИРУСА ГЕПАТИТА С (ВАРИАНТЫ), КОМПОЗИЦИЯ ПОЛНОСТЬЮ ЧЕЛОВЕЧЕСКИХ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ ПРОТИВ ВИРУСА ГЕПАТИТА С И ГИБРИДНАЯ МЫШЬ/ЧЕЛОВЕК КЛЕТОЧНАЯ ЛИНИЯ - ПРОДУЦЕНТ ПОЛНОСТЬЮ ЧЕЛОВЕЧЕСКИХ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ ПРОТИВ ВИРУСА ГЕПАТИТА С (ВАРИАНТЫ) | 2013 |
|
RU2539770C1 |
| Иммунобиологическое средство для повышения клеточного ответа против вируса гепатита С | 2023 |
|
RU2811136C1 |
| ВАКЦИНА НА ОСНОВЕ ВИРУСОПОДОБНЫХ ЧАСТИЦ, СОДЕРЖАЩИХ ВСЕ СТРУКТУРНЫЕ АНТИГЕНЫ ВИРУСА ГЕПАТИТА С, И СПОСОБ ЕЕ ПОЛУЧЕНИЯ В ДРОЖЖАХ Hansenula polymorpha | 2012 |
|
RU2510998C2 |
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ НА ОСНОВЕ "ЖИВОГО" ВИРУСА ОСПЫ ДОМАШНЕЙ ПТИЦЫ И ИХ ПРИМЕНЕНИЕ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2353651C2 |
| ВАКЦИНЫ ПРОТИВ ВИРУСОВ ГЕПАТИТА | 1995 |
|
RU2189254C2 |
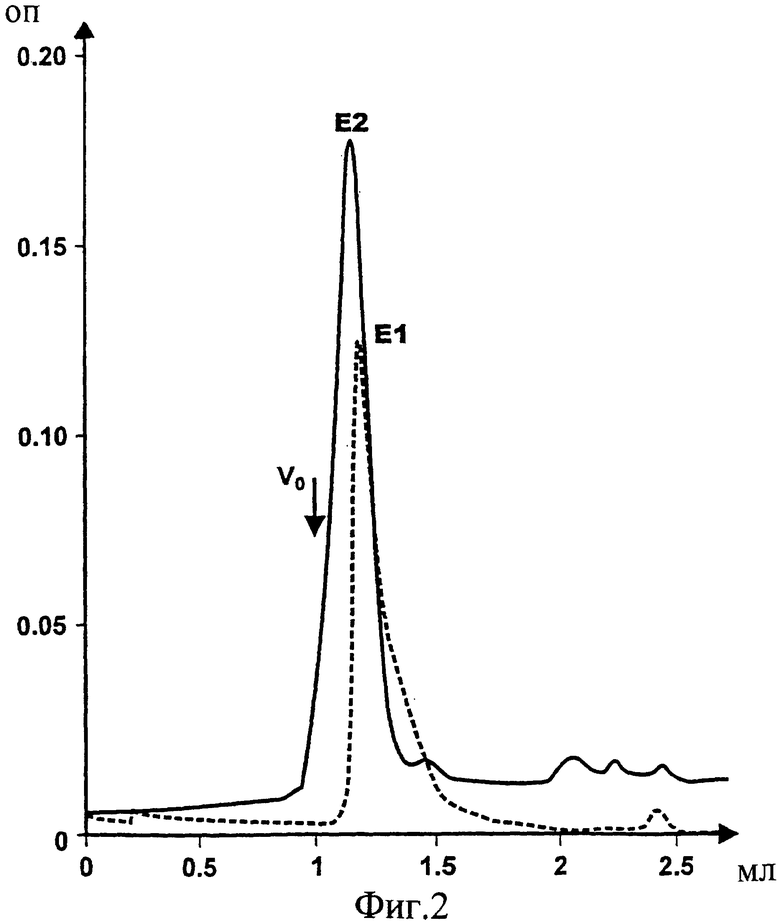
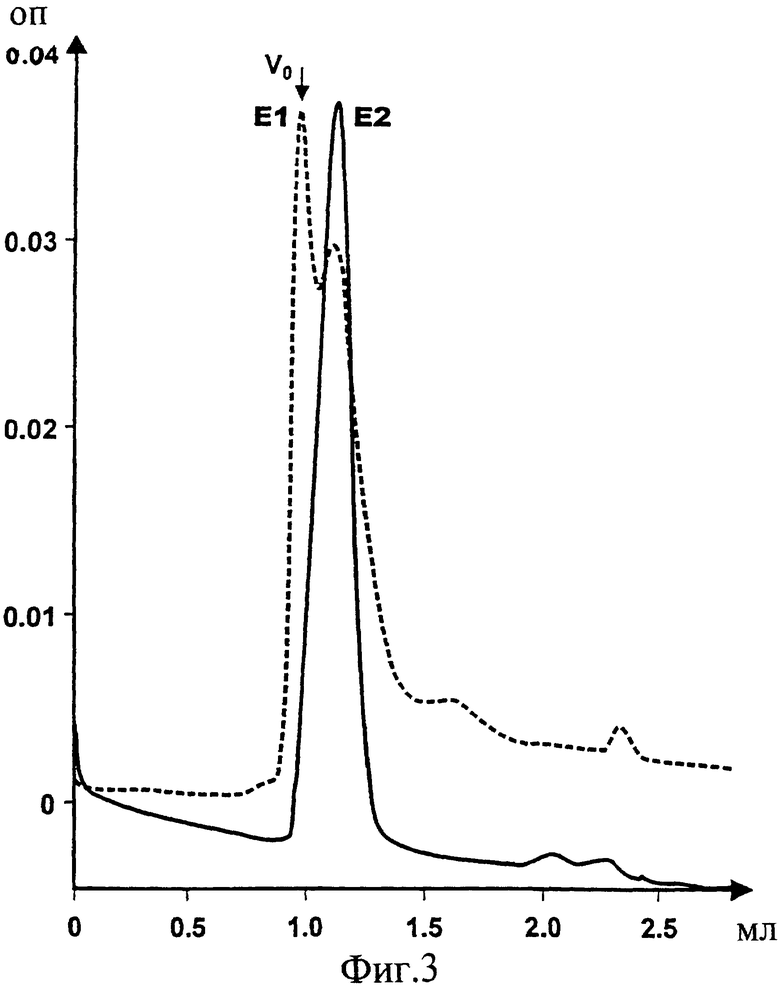
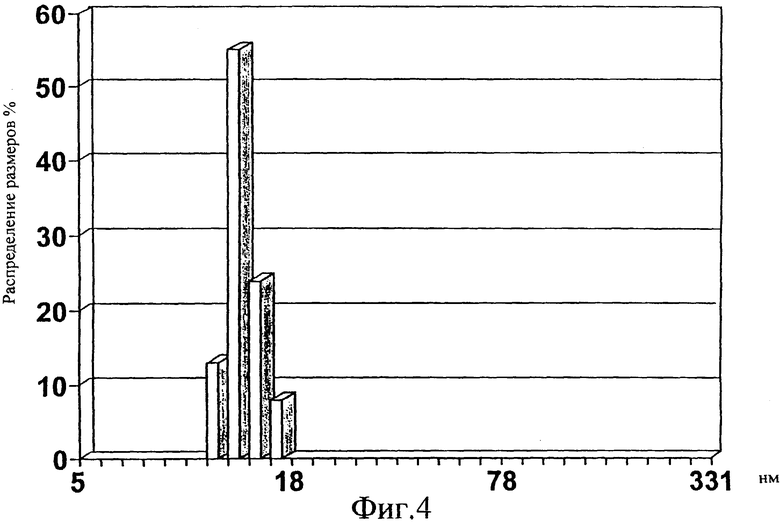


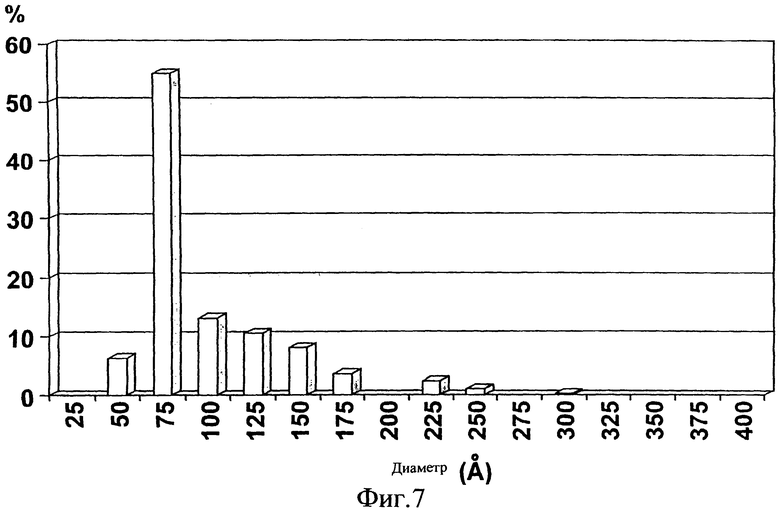
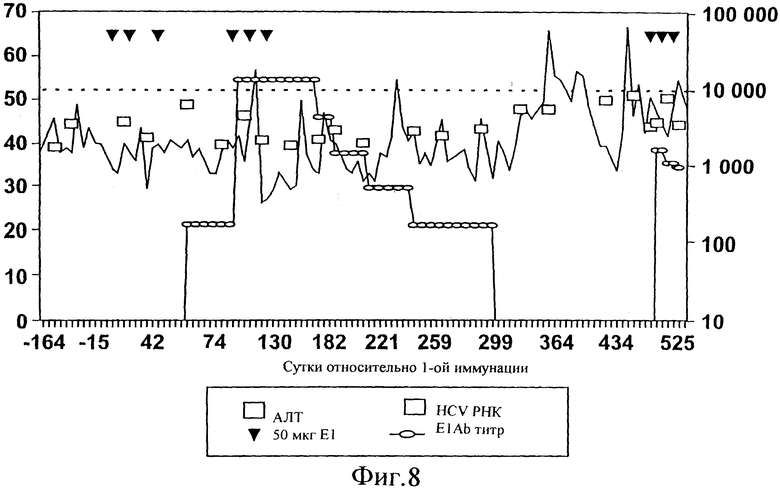
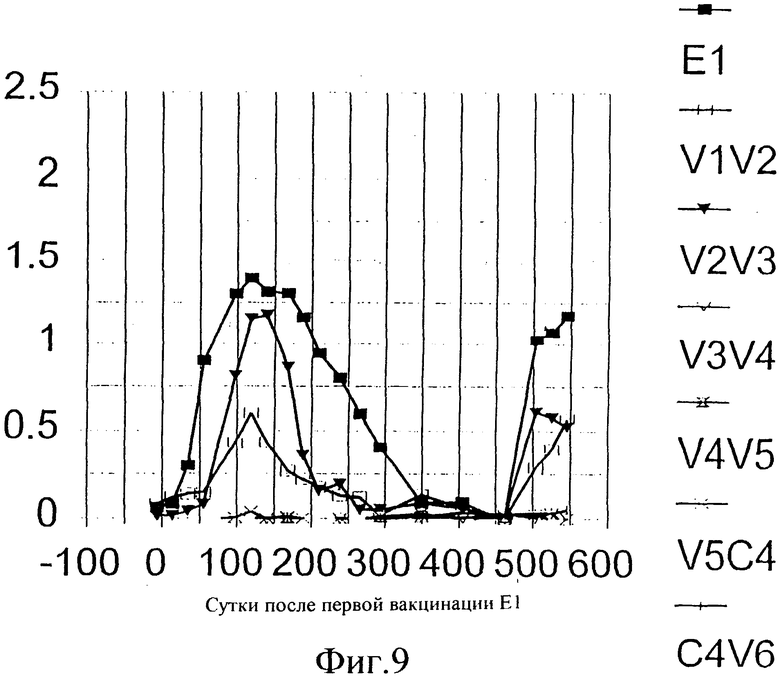

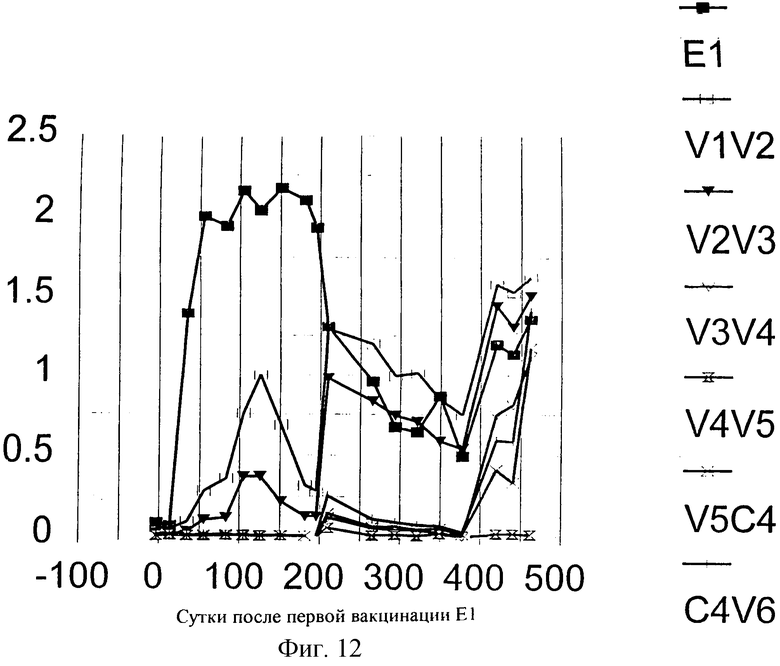

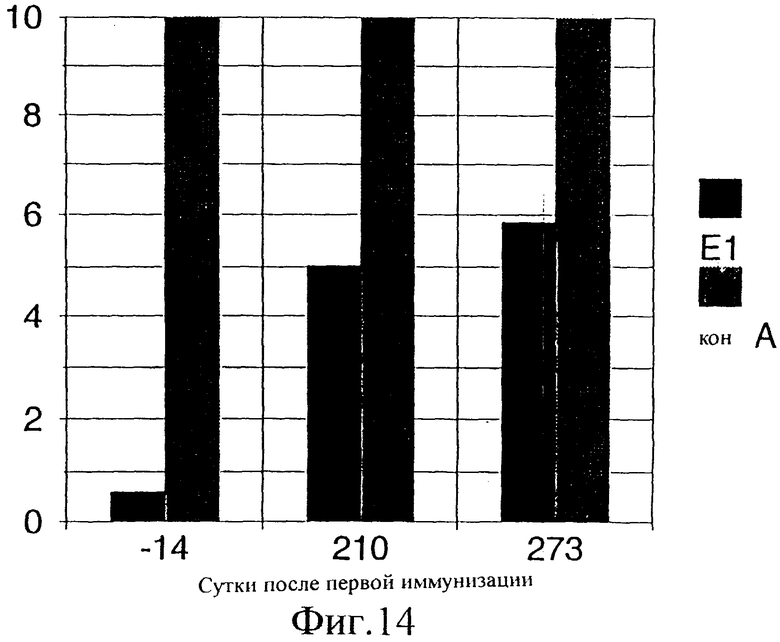
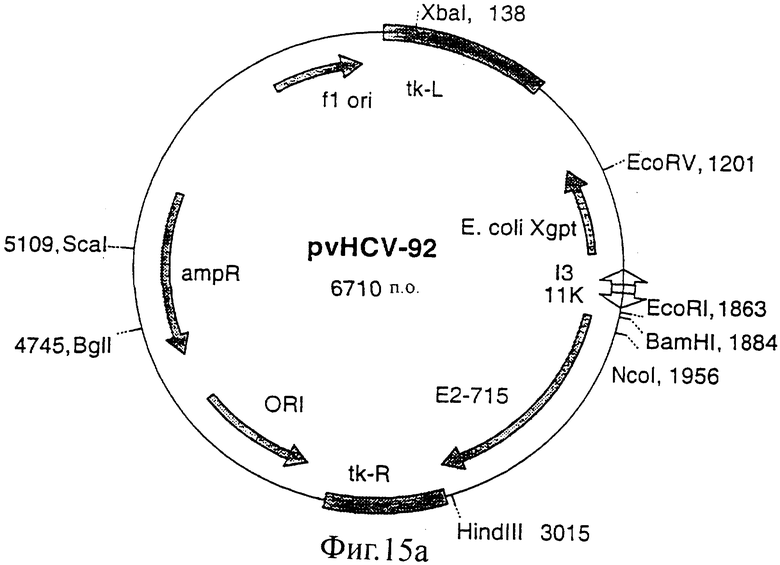
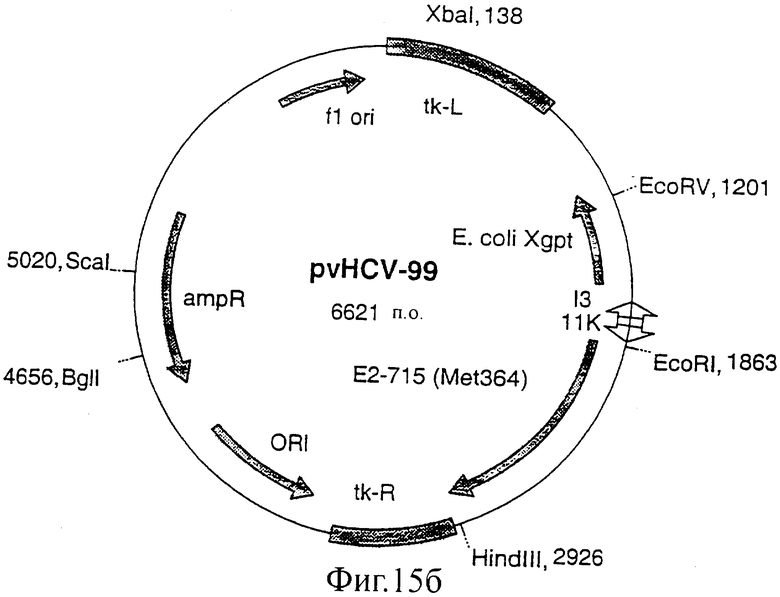
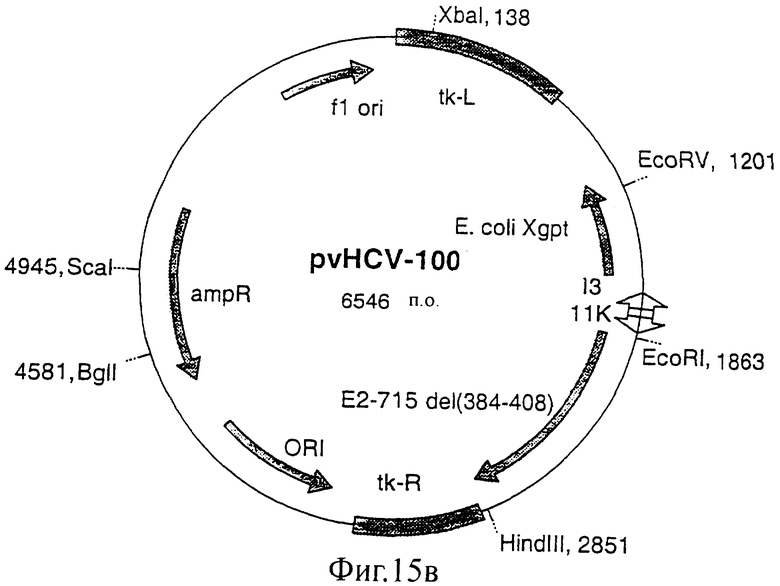
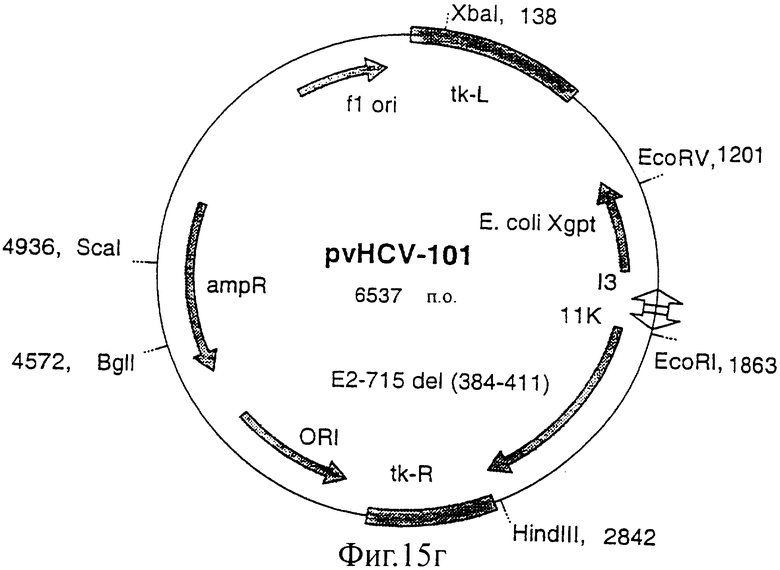

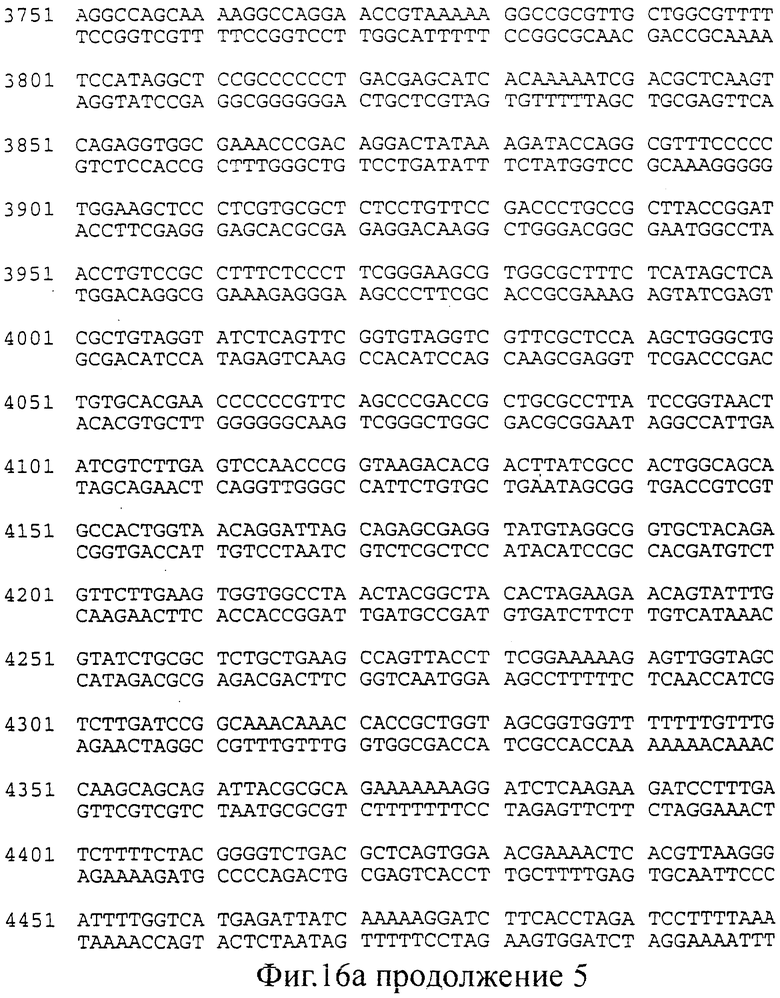
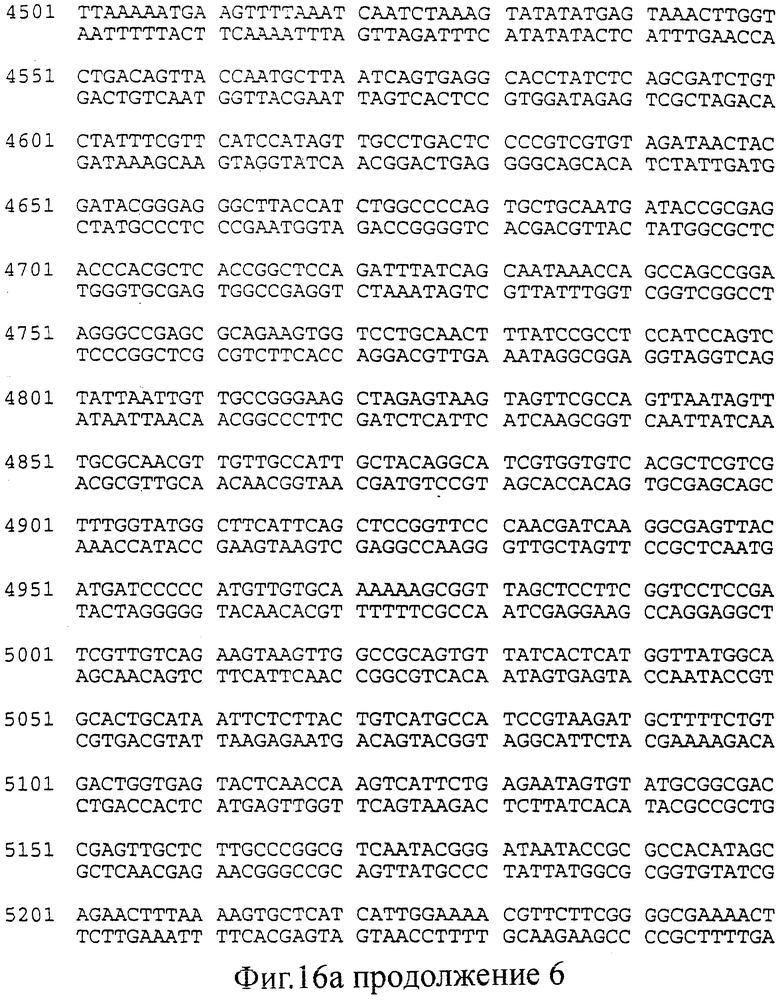
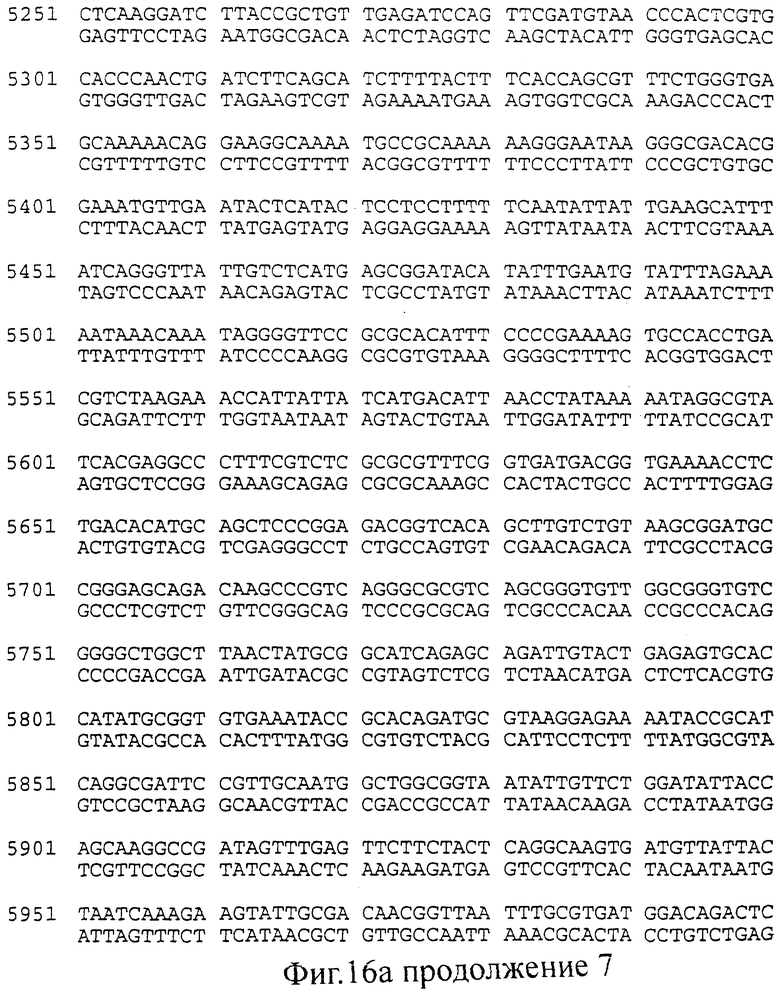
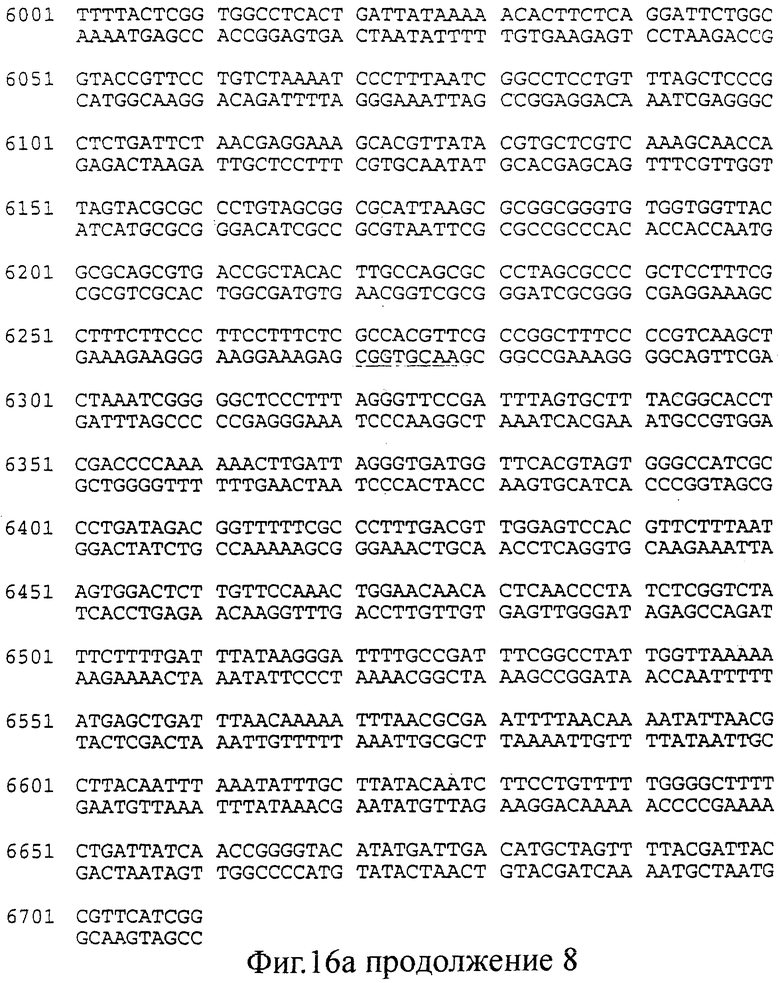


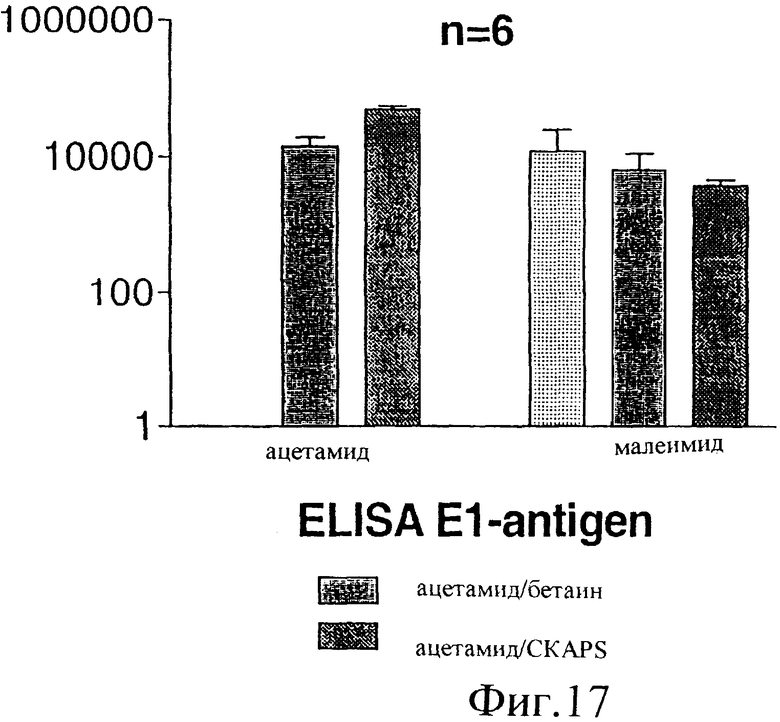
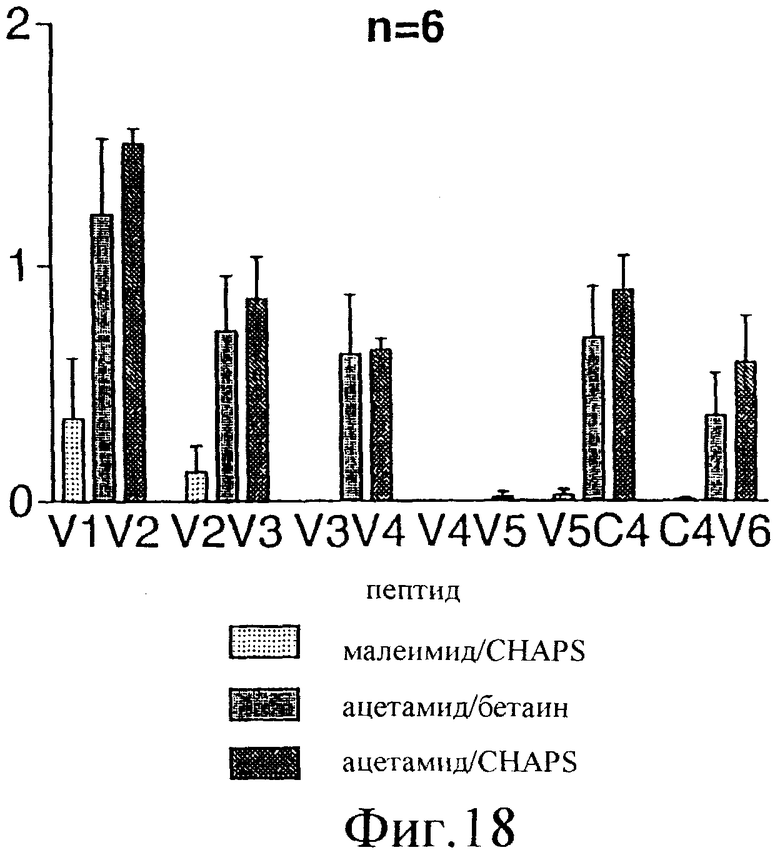
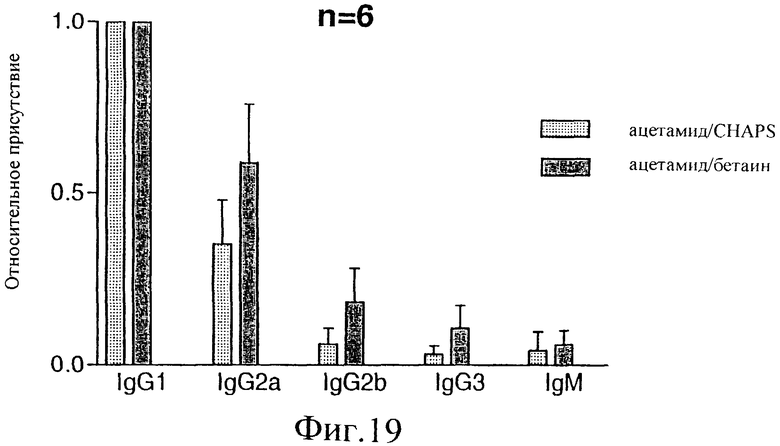
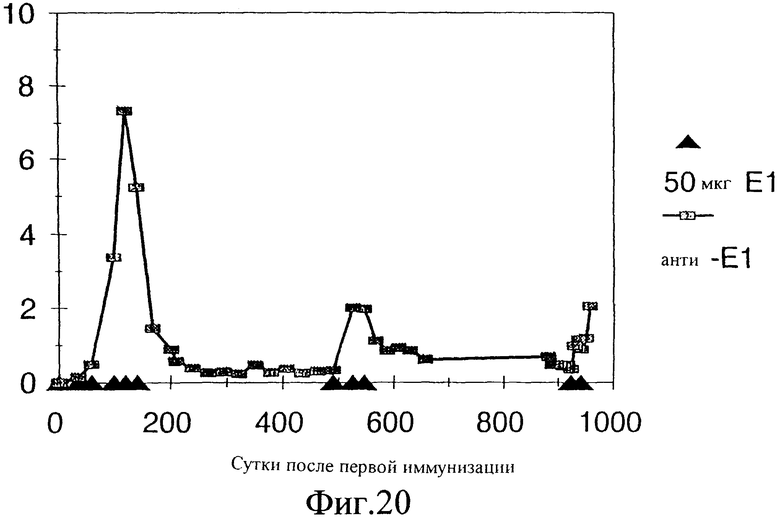
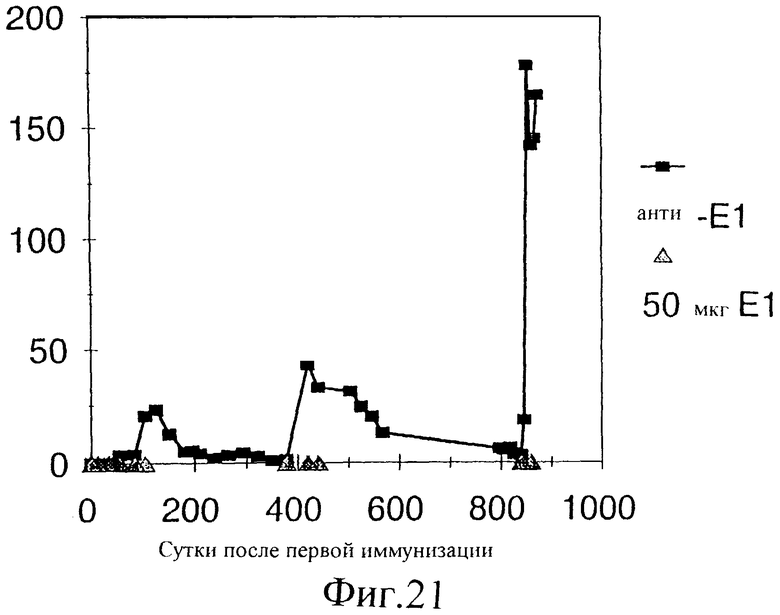
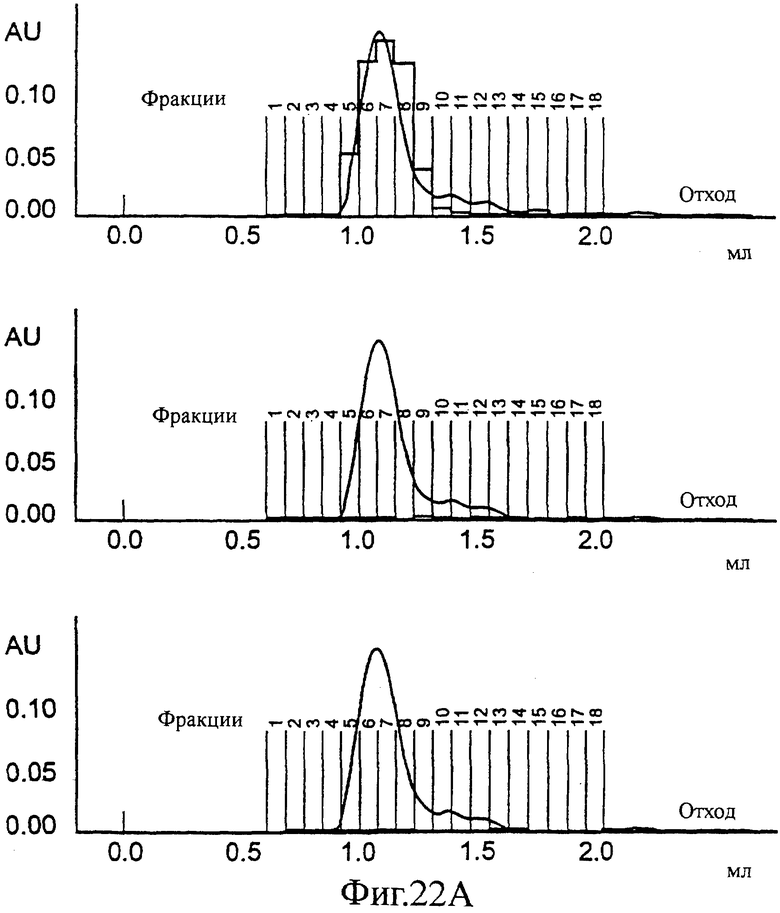
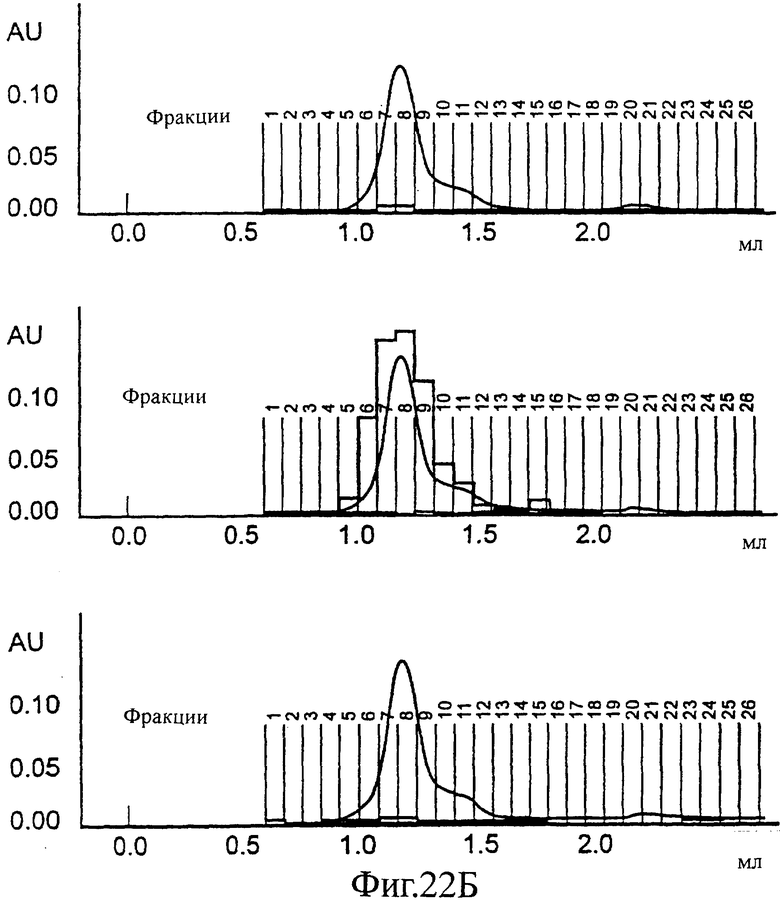
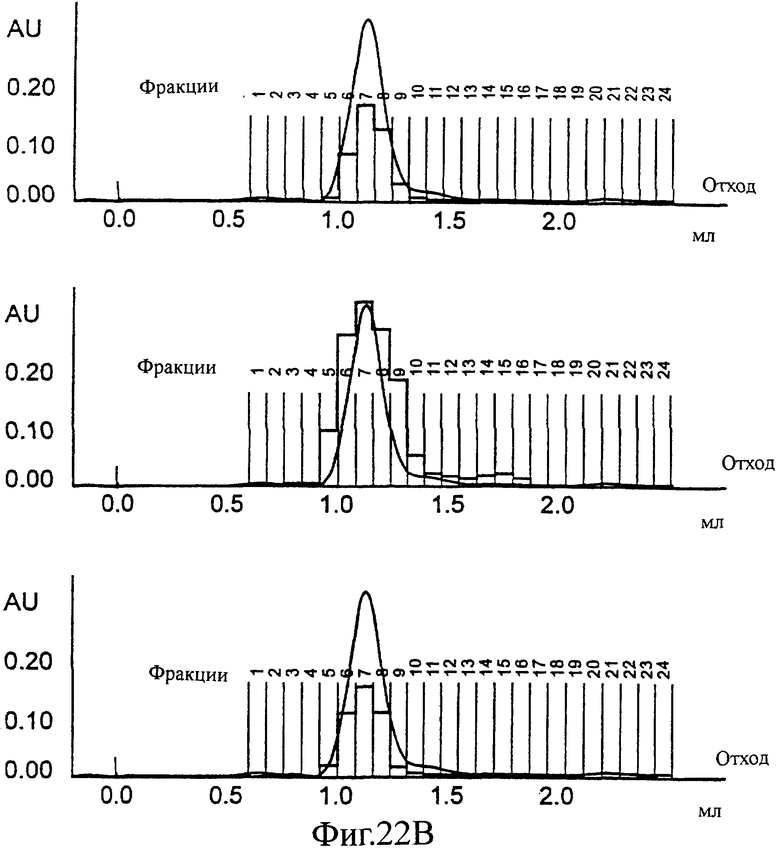
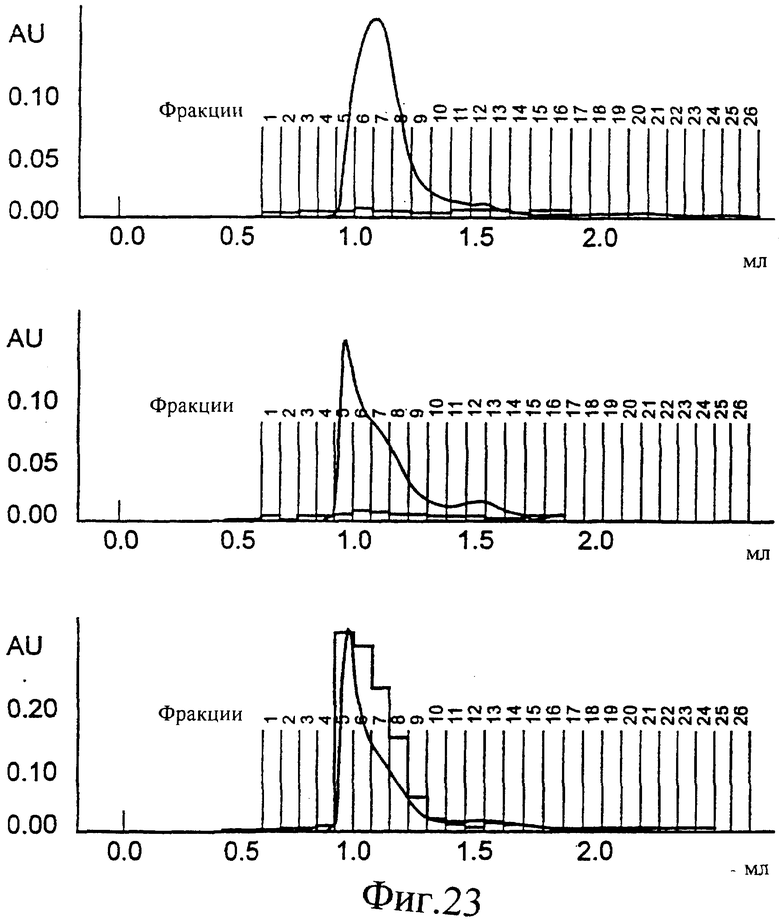
Изобретение относится к области биотехнологии. Предложена олигомерная частица, способная индуцировать иммунитет против вируса гепатита С. Частица состоит из очищенных белков или их частей, оболочки вируса гепатита С (HCV) E1, E1s и Е2. Предложенная частица имеет диаметр от 5 до 100 нанометров. Предложен также способ получения такой частицы. Способ предусматривает очистку белков HCV в растворе, образование олигомерных частиц путем замены очищенных белков раствором детергента или раствором соли и очистку олигомерных частиц. Предложены также композиция для индукции иммунитета против HCV, содержащая предложенную частицу, специфическое к частице антитело, наборы для обнаружения HCV антигенов, иммунологический анализ для обнаружения антител к HCV и вакцина против вируса гепатита С (HCV), которая содержит олигомерную частицу. Предложенные изобретения позволяют индуцировать специфический гуморальный и клеточный иммунитет к белкам оболочки HCV, а также проводить диагностику HCV. Изобретение может быть использовано в медицине. 9 н. и 62 з.п. ф-лы, 23 ил., 6 табл.
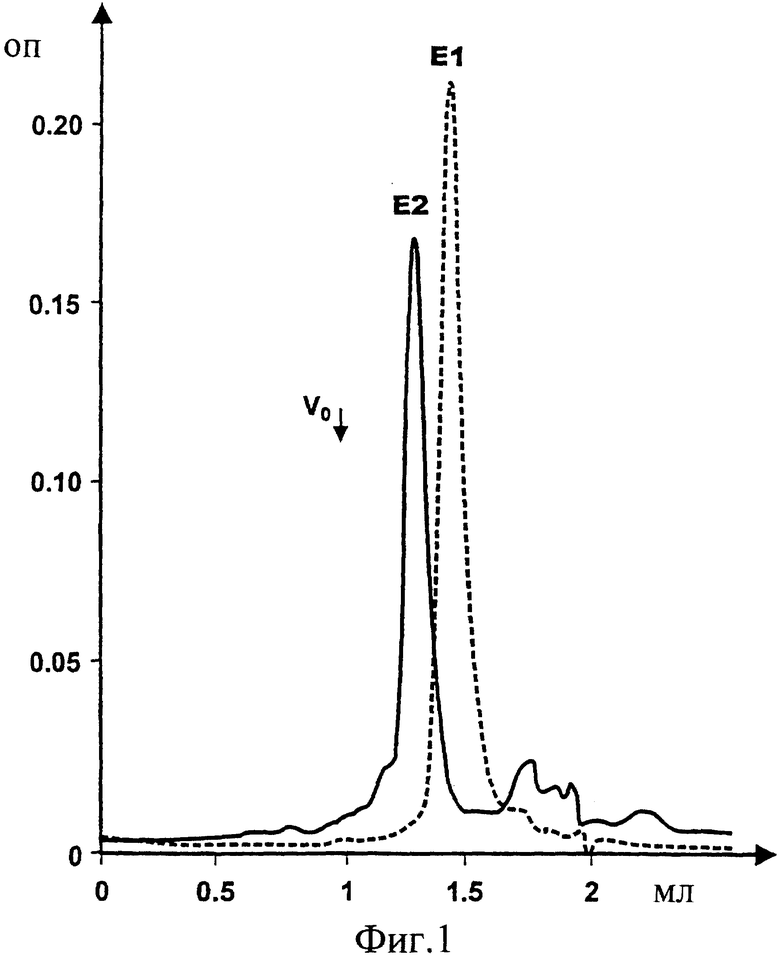
(I) очистка белков оболочки HCV в растворе,
(II) образование олигомерных частиц путем замены очищенных белков оболочки HCV со стадии (I) раствором детергента или раствором соли,
(III) очистка олигомерных частиц, образованных на стадии (II).
(I) очистка белков оболочки HCV в растворе,
(II) образование олигомерных частиц путем замены очищенных белков оболочки HCV со стадии (I) раствором детергента или раствором соли,
(III) очистка олигомерных частиц, образованных на стадии (II).
(I) берут олигомерную частицу по любому из п.п.1-23,
(II) инкубируют указанный биологический образец с олигомерной частицей со стадии (I) в условиях, которые дают возможность образования комплекса между указанной олигомерной частицей и указанными антителами к HCV,
(III) определяют после стадии (II), образовался ли указанный комплекс и, отсюда, присутствие антител к HCV в указанном биологическом образце.
Приоритет по пунктам формулы изобретения
| WO 9604385, 15.02.1996 | |||
| WO 9613590, 09.05.1996 | |||
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКОВ HCV, ПРИГОДНЫХ ДЛЯ ИСПОЛЬЗОВАНИЯ В ВАКЦИНЕ ИЛИ ИММУНОАНАЛИЗЕ, АСИАЛОГЛИКОПРОТЕИН (ВАРИАНТЫ), КОМПОЗИЦИЯ ДЛЯ ИСПОЛЬЗОВАНИЯ В ВАКЦИНЕ ИЛИ В ИММУНОАНАЛИЗЕ (ВАРИАНТЫ), СПОСОБ ОЧИСТКИ АСИАЛОГЛИКОПРОТЕИНА И СПОСОБ ПОНИЖЕНИЯ СОДЕРЖАНИЯ ИЛИ ЭЛИМИНАЦИИ HCV | 1991 |
|
RU2123528C1 |

