Введение
Область техники, к которой относится изобретение
Настоящее изобретение относится к области получения рекомбинантного генетически сконструированного антитела, более конкретно к получению линейного одноцепочечного рекомбинантного триспецифического анти-СЕА/СD3/CD28-антитела (scTsAb), к способам, используемым для конструирования, экспрессии и очистки scTsAb, к векторам и клеткам-хозяевам E.coli, которые содержат указанное scTsAb.
Уровень техники
Для активации Т-лимфоцитов требуются два вида сигналов in vivo: 1) взаимодействие между комплексом МНС/антигенный пептид, находящимся на антигенпрезентирующих клетках (АРС), и комплексом TCR/CD3 на Т-лимфоцитах создает первый сигнал; и 2) взаимодействие между костимулирующим рецептором на АРС и ко стимулирующей молекулой на Т-лимфоцитах создает второй костимулирующий сигнал. В общем, считается, что существование только первого сигнала не может полностью стимулировать Т-лимфоциты (Baxter and Hodgkin, 2002; Bernard et al., 2002).
Существуют два типа Т-лимфоцитов, включая цитотоксические Т-лимфоциты (CTL) и Т-хелперные клетки (ТН). CTL является основной эффекторной клеткой для клеточного иммунологического ответа, в то время как ТН принимает участие в клеточном иммунологическом ответе косвенно, посредством секреции цитокинов (таких как интерлейкин-2 (IL-2)). Так как противоопухолевый иммунитет главным образом является клеточным иммунитетом, то конструирование противоопухолевых лекарственных средств для специфичной активации CTL является чрезвычайно важным для противоопухолевой иммунотерапии (Foss, 2002).
На практике сконструированы некоторые рекомбинантные биспецифические (BsAb) анти-ТАА/СD3-антитела для обеспечения первого сигнала для активации CTL и нацеливания активированных CTL на окружение опухолевых клеток, чтобы вызвать их специфичную гибель. Из указанных антител некоторые биспецифические антитела уже поступили в клинику (Daniel, Kroidl et al., 1998; Holliger, Manzke et al., 1999; Loffler, Kufer et al., 2000; Manzke, Tesch et al., 2001; Manzke, Tesch et al., 2001; Dreier, Lorenczewski et al., 2002; Fang M., 2002; Dreier, Baeuerle et al., 2003; Loffler, Gruen et al., 2003; Fan, Zhao et al., 2004). Было доказано, что они специфично активируют Т-лимфоциты и индуцируют заметный специфичный цитолиз опухолевых клеток. Однако большинство таких антител не могут полностью активировать Т-лимфоциты и иногда могут приводить к активации индуцированной клеточной гибели (AICD) Т-лимфоцитов (Daniel, Kroidl et al., 1998).
Для преодоления указанных выше недостатков сконструированы BsAb другого вида, анти-TAA/CD28-BsAb. В присутствии анти-TAA/CD3-BsAb клетки CTL получают двойные активирующие сигналы, которые индуцируют более эффективный специфичный цитолиз опухолевых клеток. Однако существует несколько недостатков при комбинированном применении двух BsAb. К недостаткам относится в два раза большее количество стадий экспрессии и очистки антител и, как следствие, увеличение себестоимости производства, и проблема «партнерства» двух BsAb при клиническом применении. Триспецифическое антитело (TsAb) с тремя специфичностями связывания (ТАА, CD3 и CD28) может заменить два указанных выше BsAb в обеспечении двойных активирующих сигналов (Jung, Brandl et al., 2001; Kodama, Suzuki et al., 2002) и может иметь преимущество перед ними в процессе экспрессии, очистки и при клиническом применении.
До настоящего времени имеются три типа TsAb. Такие TsAb включают 1) химически конъюгированные TsAb (Jung, Freimann et al. 1991; Tutt, Stevenson et al., 1991; French, 1998; Wong, Vakis et al., 2000); 2) рекомбинантные полимерные TsAb (Atwell, Breheney et al., 1999; Dolezal, Pearce et al., 2000; Schoonjans, Willems et al., 2000; Schoonjans, Willems et al., 2000; Kortt, Dolezal et al., 2001; Schoonjans, Willems et al., 2001; Willems, Leoen et al., 2003) и 3) одноцепочечные рекомбинантные TsAb (scTsAb) (Li-ping, Ju-long et al., 2003). Считается, что третий тип TsAb имеет преимущества по сравнению с другими в отношении простоты конструирования, экспрессии и очистки. Так как раковый эмбриональный антиген (карциноэмбриональный антиген) (СЕА) является ТАА широкого спектра действия (Ganjei, Nadji et al., 1988; Horie, Miura et al., 1996; Kuo, Tsai et al., 1996; Feil, Wechsel et al., 1999; Kammerer, Thanner et al., 2003; Shang-zhi, Chang-cheng et al., 2004), то scTsAb, содержащие анти-СЕА-антитела, могут быть использованы для профилактики или лечения различных опухолей в клинике.
Сущность изобретения
В настоящем изобретении введение анти-СЕА-антитела в анти-CEA/CD3/CD28-scTsAb обеспечивает возможность отличать опухолевые клетки от нормальных клеток in vivo и избегать или уменьшать неспецифическую гибель клеток, вызываемую активированными Т-лимфоцитами.
Так как СЕА широко экспрессируется во многих опухолевых клетках, другой аспект настоящего изобретения относится к широкому применению указанных антител для лечения или профилактики различных опухолей в будущем.
Другой аспект настоящего изобретения относится к способу конструирования scTsAb.
Аминокислотная последовательность (SEQ ID NO: 1) мышиного одноцепочечного анти-СЕА-фрагмента вариабельной области, находящейся в CEA-scTsAb, указана ниже:




Другой аспект настоящего изобретения относится к вектору для экспрессии CEA-scTsAb: CEA-scTsAb/pTRI.
Дополнительные аспекты настоящего изобретения относятся к способу стимуляции цитоплазматической экспрессии указанных выше растворимых scTsAb при более низкой температуре и относятся к способу очистки указанных выше scTsAb с использованием DEAE-анионообменной хроматографии.
Однако в контексте настоящего изобретения другие аспекты и преимущества изобретения будут очевидны для специалистов в данной области, особенно на основе описания в разделе «Примеры».
Краткое описание чертежей
Фиг.1. Диаграмма способа конструирования рамки ДНК для множественного клонирования с использованием ПЦР с перекрыванием. Номера со 2 по 11 означают различные синтетические фрагменты полимерных нуклеиновых кислот. Символы «А, В, С, D, Е, I, II, III, IV, UP, DOWN» означают различные промежуточные продукты конструирования. Обозначение «WHOLE» означает конечный продукт.
Фиг.2. Выявление продуктов ПЦР с перекрыванием электрофорезом в агарозном геле. Дорожка 1: продукт ПЦР с перекрыванием; дорожка 2: маркер ДНК DL2000 (Dalian Takara Biothech.).
Фиг.3. Последовательность рамки ДНК для множественного клонирования.
Фиг.4. Способ конструирования СЕА-scTsAb.
Фиг.5. Схематичные карты векторов для конструирования и экспрессии CEA-scTsAb.
Фиг.6. Идентификация способа конструирования электрофорезом в агарозном геле. Дорожка 1: продукт ПЦР, амплифицированный с пустого вектора pTRI; дорожка 2: продукт ПЦР, амплифицированный с вектора CD28 VH/pTRI; дорожка 3: продукт ПЦР, амплифицированный с вектора CD3scFv/CD28 VH/pTRI; дорожка 4: продукт ПЦР, амплифицированный с вектора CEA-scTsAb/pTRI; дорожка 5: маркер ДНК DL2000 (Dalian Takara Biotech.).
Фиг.7. Показана схема процесса конструирования мышиного анти-CEA-scFv посредством ПЦР с перекрыванием. Номера 1-22 означают различные синтетические фрагменты полимерных нуклеиновых кислот; Символы «А, В, С, D, Е, F, G, H, I, J, К, а, b, с, d, e, f, g, I, II, III, IV, UP, DOWN» означают промежуточные продукты конструирования. Символ «WHOLE» означает конечный продукт.
Фиг.8. Выявление продукта ПЦР с перекрыванием с использованием электрофореза в агарозном геле. Дорожки 1 и 9: маркеры ДНК DL2000 (Dalian Takara Biotech.); дорожки 2-5: промежуточные продукты I, II, III, IV; дорожки 6 и 7: промежуточные продукты UP и DOWN; дорожка 8: конечный продукт WHOLE.
Фиг.9. SDS-ПААГ экспрессированного растворимого CEA-scTsAb. Дорожка 1: осадок после ультразвуковой обработки, содержащий экспрессированный CEA-scTsAb/pTRI; дорожка 2: надосадок после ультразвуковой обработки, содержащий продукты экспрессии CEA-scTsAb/pTRI; дорожка 3: стандарты молекулярной массы белков (Shanghai Biochemistry Institute); дорожка 4: осадок после ультразвуковой обработки, содержащий продукты экспрессии пустого вектора pTRI; дорожка 5: надосадок после ультразвуковой обработки, содержащий продукты экспрессии пустого вектора pTRI. Полосы CEA-scTsAb указаны стрелками на соответствующих дорожках.
Фиг.10. Вестерн-блоттинг растворимого экспрессированного CEA-scTsAb. Дорожка 1: стандарты молекулярной массы белков (NEB); дорожка 2: осадок после ультразвуковой обработки, содержащий продукты экспрессии CEA-scTsAb/pTRI; дорожка 3: осадок после ультразвуковой обработки, содержащий продукты экспрессии пустого вектора pTRI; дорожка 4: надосадок после ультразвуковой обработки, содержащий продукты экспрессии CEA-scTsAb/pTRI; дорожка 5: надосадок после ультразвуковой обработки, содержащий продукты экспрессии пустого вектора pTRI.
Фиг.11. SDS-ПААГ CEA-scTsAb, очищенного анионообменной хроматографией на DEAE. Дорожка 1: надосадок после ультразвуковой обработки, содержащий продукты экспрессии пустого вектора pTRI; дорожка 2: надосадок после ультразвуковой обработки, содержащий продукты экспрессии CEA-scTsAb/pTRI; дорожка 3: поток, проходящий через колонку при анионообменной хроматографии на DEAE; дорожка 4: элюирование NaCl при анионообменной хроматографии на DEAE; дорожка 5: элюирование NaOH при анионообменной хроматографии на DEAE; дорожка 6: стандарты молекулярной массы белков (Shanghai Biochemistry Institute). Полосы CEA-scTsAb указаны стрелками на соответствующих дорожках.
Фиг.12. Результаты ELISA (твердофазного иммуноферментного анализа) CEA-scTsAb. На фигуре сверху вниз первая кривая: 10 мкг/мл мембранного антигена Jurkat; вторая кривая: 1 мкг/мл очищенного СЕА (R&D); третья кривая: 1 мкг/мл химеры CD28-Fc (R&D); четвертая кривая: без антигенов.
Фиг.13. FCM связывания CEA-scTsAb с различными опухолевыми клетками. Заштрихованный пик является негативным контролем без добавления CEA-scTsAb.
Фиг.14. FCM связывания CEA-scTsAb с клетками Jurkat и РВМС мононуклеарными клетками периферической крови (РВМС). Заштрихованный пик является негативным контролем без добавления CEA-scTsAb.
Фиг.15. Влияние соотношения Е/Т (эффекторные клетки/клетки-мишени) на специфичный цитолиз опухолевых клеток, индуцированный CEA-scTsAb, который определяли в анализе МТТ. На фигуре сверху вниз первая кривая: Е/Т = 10; вторая кривая: Е/Т = 5, третья кривая: Е/Т = 1. Эффекторные клетки: РВМС; клетки-мишени: опухолевые клетки SW1116.
Фиг.16. Влияние концентрации CEA-scTsAb на специфичный цитолиз опухолевых клеток в анализе МТТ. Существует четыре ступенчатых фазы специфичного цитолиза опухолевых клеток. В первой фазе, от 6 мкг/мл до 12 мкг/мл, наблюдается отрицательная корреляция эффективности специфичного цитолиза опухолевых клеток и концентрации CEA-scTsAb, и пик достигается при 6 мкг/мл. Во второй фазе, от 750 нг/мл до 6 мкг/мл, наблюдается прямая корреляция, и минимальная эффективность наблюдается при 750 нг/мл. В третьей фазе, от 24 нг/мл до 750 нг/мл, наблюдается возврат к отрицательной корреляции. В четвертой фазе, от 24 нг/мл до нуля, снова проявляется прямая корреляция.
Фиг.17. Влияние концентрации CEA-scTsAb на пролиферацию эффекторных клеток, определяемое в анализе МТТ. Существуют 3 ступенчатые фазы в изменении индекса стимуляции (SI). В первой фазе, от 750 нг/мл до 12 мкг/мл, наблюдается прямая корреляция SI и концентрации CEA-scTsAb, и наименьшее значение достигается при 750 нг/мл. Во второй фазе, от 50 нг/мл до 750 нг/мл, наблюдается отрицательная корреляция, и пик достигается при 50 нг/мл. В третьей фазе, от 50 нг/мл до нуля, наблюдается возврат к прямой корреляции.
Фиг.18. Морфологические изменения различных клеток в процессе специфичного цитолиза опухолевых клеток, индуцированного CEA-scTsAb. (А) Опухолевые клетки SW1116 после 20 часов культивирования; (В)-(I) Смесь SW1116 (клетки-мишени) и РВМС (эффекторные клетки) при добавлении CEA-scTsAb. Е/Т=5. (В) Прикрепленные клетки-мишени начинают открепляться; (С) Эффекторные клетки агрегируют на поверхности клеток-мишеней; (D) TsAb появлялись на поверхности клеток-мишеней; (Е) Оболочки клеток-мишеней частично разрушаются; (F) Оболочки клеток-мишеней полностью разрушаются; (G)-(I) Клетки-мишени распадаются на фрагменты.
Фиг.19. Схема механизма специфичного цитолиза опухолевых клеток, индуцированного CEA-scTsAb. Верхняя схема: структура CEA-scTsAb; нижняя схема: схема механизма специфичного цитолиза опухолевых клеток, индуцированного CEA-scTsAb; CEA-scTsAb одновременно связывается как с клеткой-мишенью, так и с эффекторной клеткой, таким образом приводя к активации эффекторной клетки, которая обеспечивает двойные сигналы, которые вызывают специфичную гибель клеток-мишеней.
Фиг.20. Микрофотографии убитых клеток-мишеней (SW1116). Дорожки А, В и С представляют три разных состояния гибели опухолевых клеток соответственно: некроз, апоптоз на поздней стадии и апоптоз на ранней стадии.
Фиг.21. FCM (PI/аннексин V-ФИТЦ (изотиоцианат флуоресцеина)) специфичного цитолиза опухолевых клеток. Четыре сектора представляют разные состояния опухолевых клеток: живые клетки в нижнем левом секторе (LL); клетки на ранней стадии апоптоза в нижнем правом секторе (LR); клетки на поздней стали апоптоза в верхнем правом секторе (UR); некротические клетки в верхнем левом секторе (UL). Образец без добавления CEA-scTsAb: LL (90,17%), LR (1,66%), UL (5,94%), UR (2,33%). Образец с добавлением 50 нг/мл CEA-scTsAb: LL (52,83%), LR (16,12%), UL (9,80%), UR (21,25%).
Подробное описание изобретения
В описании настоящего изобретения все термины легко понятны специалистам в данной области, за исключением тех, которые объясняются подробно. Некоторые термины, используемые в описании, указаны ниже:
Линейное одноцепочечное рекомбинантное триспецифическое антитело представляет собой одну линейную молекулу, сконструированную с тремя разными специфичностями связывания антигенов способом генетической инженерии. В частности, линейное одноцепочечное рекомбинантное триспецифическое анти-СЕА/СD3/СD28-антитело представляет собой единую линейную молекулу, сконструированную слиянием фрагментов трех разных антител (анти-СЕА-антитела, анти-CD3-антитела, анти-CD-28-антитела), которые разделены двумя линкерами (Fc-линкером и HSA-линкером) (Min Fang, 2003). Может быть добавлена с-mус-метка и (His) 6-метка к С-концу молекулы для выявления активности или дополнительной очистки (Hengen, 1995; Fan, Villegas et al., 1988). Фрагменты антител, указанные в данном описании, могут представлять собой одноцепочечные фрагменты вариабельной области (scFv) антител, Fab-фрагменты антител или однодоменные фрагменты антител (VH или VL). Более конкретно СЕА-scTsAb конструируют последовательным слиянием анти-CEA-scFv, Fc-линкера, анти-СD3-sсFv, HSA-линкера и анти-CD28-VН с c-myc-меткой и His-6-меткой на С-конце. Указанный блок имеет два преимущества:
1. Антитело основано на модели активации Т-клеток двумя сигналами и обладает способностью полностью активировать Т-клетки.
2. СЕА является ТАА широкого спектра действия, поэтому СЕА имеет широкие перспективы для применения при лечении и профилактике многих опухолей.
Способ индукции цитоплазматической экспрессии растворимого CEA-scTsAb при пониженной температуре, предлагаемый в настоящем изобретении, требует индукции бактерий-хозяев 0,4 мМ IPTG при 30°С. В случае такого способа может быть заметно снижено образование телец включений, и примерно 50% экспрессированного CEA-scTsAb является растворимым. Растворимый экспрессированный CEA-scTsAb может быть использован на следующей стадии очистки непосредственно без денатурации или ренатурации, что может снизить себестоимость продукции и повысить выход.
Способ одностадийной очистки, при котором собирают проходящий поток при анионообменной хроматографии на DEAE, требует нанесения растворимых продуктов экспрессии на колонку с анионообменной смолой DEAE при рН 8,0. Затем почти все белки, не представляющие интерес, могут быть абсорбированы, тогда как большая часть CEA-scTsAb выходит в проходящем потоке с чистотой примерно 75%.
Способ согласно изобретению описан ниже:
Сначала конструируют исходный вектор pTRI с помощью введения новых специфичных сайтов для множественного клонирования (MCS). Затем фрагмент ДНК, кодирующий анти-CD28 VH, амплифицируют в ПЦР с вектора CD28 VH/pTMF, на обоих концах которого добавляют специфичные пары сайтов рестрикции NdeI/KpnI. Таким же способом готовят фрагмент ДНК, кодирующий анти-СD3-scFv, с сайтами рестрикции ScaI/SalI. Фрагмент ДНК, кодирующий анти-CEA-scFv, с сайтами рестрикции XhoI/EcoRI вырезают из CEA-scFv/pTMF. Наконец все три фрагмента последовательно вводят в pTRI, получая конечный вектор CEA-scTsAb/pTRI. Фрагменты ДНК, кодирующие анти-CEA-scFv, анти-CD3-scFv и анти-CD28-Ун, располагают последовательно от N-конца к С-концу.
После трансформации Е.coli BL21(DE3) и индукции с использованием IPTG при низкой температуре (30°С) растворимое CEA-scTsAb экспрессируется в цитоплазме. Антитело сначала очищают, используя одну стадию анионообменной хроматографии на DEAE. Специфичности связывания с тремя антигенами (СЕА, CD3, CD28) выявляют в ELISA. Специфичности связывания с опухолевыми клетками выявляют с использованием FCM после конъюгирования CEA-scTsAb с FITC. Цитолиз опухолевых клеток и пролиферацию Т-лимфоцитов, индуцированные CEA-scTsAb, анализируют в МТТ-анализе. Морфологические изменения опухолевых клеток регистрируют на микрофотографиях в инвертированном микроскопе. Некроз и апоптоз опухолевых клеток, индуцированные CEA-scTsAb, визуализируют с помощью FCM с двойным окрашиванием, РI/аннексин-V-FIТС, и флуоресцентной микроскопии.
Примеры
Пример 1. Получение фрагмента ДНК, содержащего сайты множественного клонирования с использованием ПЦР с перекрыванием.
Схема способа показана на фиг.1. Все синтезированные фрагменты, используемые согласно изобретению, указаны ниже:


Стадии обработки:
Стадия 1. Как показано на фиг.1, попарно смешивают фрагменты (2-11) и осуществляют реакцию элонгации, как указано ниже.
Реакционная смесь: синтетические фрагменты, 1 мкл (каждого); 10 х буфер для ПЦР, 2 мкл; dNTP (по 2 ммоль/мл каждого) (Dalian Takara Biotechnology Co. Ltd.) 2 мкл; Taq (1 ед.) (Dalian Takara Biotechnology Co. Ltd.) 0,5 мкл; дистиллированная вода, 14 мкл.
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 30 секунд; 10 циклов.
Все продукты собирали без очистки и непосредственно использовали для следующей стадии.
Стадия 2. Как показано на фиг.1, попарно смешивают продукты (А, В, С, D, Е) со стадии 1 и осуществляют реакцию элонгации, как указано ниже.
Реакционная смесь: продукты со стадии 1, 10 мкл (каждого).
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 30 секунд; 10 циклов.
Все продукты наносили на агарозный гель для электрофореза (1%) и очищали, используя набор для очистки ДНК из геля (Watson Biotech. Inc.).
Стадия 3. Как показано на фиг.1, попарно смешивают продукты (I, II, III, IV) со стадии 2 и осуществляют реакцию элонгации, как указано ниже.
Реакционная смесь: продукты со стадии 2, по 1 мкл (каждого); 10 х буфер для ПЦР, 2 мкл; dNTP (по 2 ммоль/мл каждого) (Dalian Takara Biotechnology Co. Ltd.), 2 мкл; Taq (1 ед.) (Dalian Таkara Biotechnology Co. Ltd.), 0,5 мкл; дистиллированная вода, 13 мкл.
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 30 секунд; 10 циклов.
Все продукты наносили на агарозный гель для электрофореза (1%) и очищали, используя набор для очистки ДНК из геля (Watson Biotech. Inc.).
Стадия 4. Как показано на фиг.1, попарно смешивают продукты (UP, DOWN) со стадии 3 и осуществляют реакцию амплификации, как указано ниже.
Реакционная смесь: продукты со стадии 3, по 1 мкл (каждого); праймеры (синтетические фрагменты 1 и 12), 1 мкл; 10 х буфер для ПЦР, 2 мкл; dNTP (по 2 ммоль/мл каждого) (Dalian Takara Biotechnology Со. Ltd.), 2 мкл; Taq (1 ед.) (Dalian Takara Biotechnology Со. Ltd.), 0,5 мкл; дистиллированная вода, 12 мкл.
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 50 секунд; 25 циклов.
Конечные продукты (493 п.о.) разделяли электрофорезом в агарозном геле (1%) (фиг.2) и очищали с использованием набора для очистки ДНК из геля (Watson Biotech. Inc.). Последовательность, сайты рестрикции и компоненты показаны на фиг.3.
Пример 2. Конструирование CEA-scTsAb
Способ конструирования показан на фиг.4 и схематичные карты всех векторов, используемых в способе, приведены на фиг.5.
(1) Конструирование вектора pTRI:
Фрагмент ДНК, содержащий сайты множественного клонирования, и пустой вектор pTMF (Zhang et al., 2002) разрезали NcoI/BamHI и лигировали вместе. Продуктами лигирования трансформировали Е.coli штамма ТОР10 (Invitrogen). Плазмиды, выделенные из трансформированных бактериальных клеток, названы pTRI.
Расщепление ферментами рестрикции, лигирование, получение и трансформацию компетентных клеток ТОР10 осуществляли, как указано ниже:
Реакция расщепления ферментами рестрикции: в объеме 20 мкл 1 мкг pTMF или фрагмента ДНК, содержащего сайты множественного клонирования, расщепляли согласно инструкции Promega. Продукты разделяли электрофорезом в агарозном геле (1%) и очищали, используя набор для очистки ДНК из геля (Watson Biotech. Inc.).
Реакция лигирования: 50-100 нг расщепленного вектора и в 3-10 большее количество (молярное соотношение) расщепленных фрагментов ДНК смешивали в 20 мкл раствора, который содержал 2 мкл 10 х буфера для ДНК-лигазы Т4; 1 ед. ДНК-лигазы Т4 (Dalian Takara Biotechnology Co. Ltd.) и необходимое количество дистиллированной воды. Реакцию лигирования осуществляли при 16°С в течение ночи.
Подготовка компетентных клеток ТОР10: бактерии ТОР10 (Invitrogen Co.) инокулировали в 2 мл среды LB (10 г/л триптона (GIBCO Co.), 5 г/л дрожжевого экстракта (GIBCO Co.), 5 г/л NaCl, рН 7,5) и инкубировали в течение ночи при 37°С на качалке. Переносили в 20-40 мл среды LB в соотношении 1:100, инкубировали при 37°С при встряхивании до достижения А600 0,3-0,4 (примерно 2,5 часа). Охлаждали на льду в течение 15 минут и центрифугировали при 4°С со скоростью 4000 об./мин в течение 10 минут. Осадок ресуспендировали в 10 мл предварительно охлажденного раствора 0,1 моль/мл CaCl2 (Sigma Co.) и охлаждали на льду в течение 20 минут. После повторного центрифугирования при 4°С со скоростью 4000 об./мин в течение 10 минут осадок осторожно ресуспендировали в 1-2 мл предварительно охлажденного раствора, содержащего 0,1 моль/л CaCl2 и 12% глицерина. Делили на аликвоты по 200 мкл в ЕТ-пробирки, хранили при -80°С.
Трансформирование: Смесь для лигирования добавляли к 200 мкл компетентных клеток. После осторожного перемешивания и охлаждения на льду в течение 30 минут клетки помещали на водяную баню при температуре 42°С на 100 секунд, затем охлаждали на льду в течение 2 минут. После добавления к смеси 0,8 мл среды LB смесь встряхивали при 37°С (<150 об./мин) в течение 45 минут для восстановления клеток. Клетки центрифугировали при 10000 об./мин в течение 1 минуты, ресуспендировали в 50-100 мл среды LB, высевали на чашку LB-K (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, 15 г/л агара (SIGMA Co.), 50 мкг/мл канамицина (SIGMA Со.), рН 7,5) и инкубировали при 37°С в течение ночи.
Отбор позитивных клонов: Собирали отдельные клоны на чашке LB-K и затем по отдельности переносили в 2 мл среды LB-K (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, 15 г/л агара (SIGMA Co.), 50 мкг/мл канамицина (SIGMA Co.), рН 7,5). После встряхивания при 37°С в течение ночи выделяли плазмиды, используя набор для выделения плазмид (Watson Biotechnologies, Inc) согласно стандартной инструкции. Позитивные клоны идентифицировали в ПЦР, используя выделенные плазмиды в качестве матриц. Реакционная смесь для ПЦР: 0,1-1 мкл плазмидной ДНК (примерно 20-200 нг), 10 пмоль выше расположенного праймера (Т7-up: 5'-TAATACGACTCACTATAGGGGA-3') (SEQ ID NO: 17), 10 пмоль ниже расположенного праймера (T7-down: 5'-GCTAGTTATTGCTCAGCGG-3')(SEQ ID NO: 18), 2 мкл 10 х буфера Taq; 2 мкл раствора, содержащего 2 ммоль/мл dNTP; 1 ед. фермента Taq, 12 мкл дистиллированной воды. Условия реакции ПЦР: предварительная денатурация при 94°С в течение 5 минут, денатурация при 94°С в течение 40 секунд, отжиг при 53°С в течение 40 секунд; элонгация при 72°С в течение 40 секунд; 25 циклов. В конце 5 мкл продукта ПЦР разделяли электрофорезом в агарозном геле (1%). Как показано на фиг.6, продукт ПЦР имеет размер примерно 500 п.о.
(2) Конструирование CD28 VH/pTRI:
Фрагмент ДНК, кодирующий анти-CD28-VН, амплифицировали с плазмиды CD28 VH/pTMF (Cheng, Wang et al., 2002), используя P1 (P1: 5'-TCACATATGCAGGTACAGCTACAG-3') (SEQ ID NO: 19) в качестве расположенного выше праймера и Р2 (Р2: 5'-TTCGCTAGCGGAAGATACGGTACCA-3') (SEQ ID NO: 20) в качестве расположенного ниже праймера. Сайты рестрикции NdeI/NheI вводили в 5'- и 3'-конец соответственно в ходе ПЦР.
Реакционная смесь для ПЦР: по 1 мкл каждого из праймеров; 2 мкл dNTP (каждый 2 ммоль/мл); 2 мкл 10 x буфера pfu; 100 нг плазмиды CD28 VH/pTRI; 0,3 мкл фермента pfu (Promega Co.); дистиллированную воду добавляли до получения объема 20 мкл. Условия реакции ПЦР: предварительная денатурация при 94°С в течение 3 минут; денатурация при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд; элонгация при 72°С в течение 50 секунд; 25 циклов. Продукты ПЦР очищали электрофорезом в агарозном геле (1%), используя набор для очистки ДНК в геле (Watson Biotech. Inc.).
Указанный выше продукт ПЦР и плазмиду pTRI одновременно расщепляли NdeI/NheI (Promega Co.). Расщепленный продукт ПЦР (примерно 350 п.о.) и pTRI (примерно 5300 п.о.) лигировали вместе и трансформировали штамм Е.coli TOP10. Плазмиды, выделенные из позитивных клонов, названы CD28 VH/pTRI и идентифицированы в ПЦР с продуктом размером примерно 750 п.о. (как показано на фиг.6). Все рабочие операции, необходимые в данном случае, описаны в разделе (1).
(3) Конструирование CD3-scFv/CD28 VH/pTRI:
Фрагмент ДНК, кодирующий анти-CD3-scFv амплифицировали с плазмиды CD3-scFv/pTMF (Liu X.F.1996) с использованием P1 (P1: 5'-AAGAGTACTGAGGTGAAGCTGGTGG-3) (SEQ ID NO: 21) в качестве расположенного выше праймера и Р2 (Р2: 5'-GAAGTCGACAGCGCGCT TCAGTTCCAG-3) (SEQ ID NO: 22) в качестве расположенного ниже праймера. Сайты рестрикции. ScaI и SalI, вводили в 5'-конец и 3'-конец соответственно в ходе ПЦР.
Реакционная смесь для ПЦР: по 1 мкл каждого из праймеров; 2 мкл dNTP (каждый 2 ммоль/мл); 2 мкл 10 х буфера pfu; 100 нг плазмиды CD28 VH/pTRI; 0,3 мкл фермента pfu (Promega Co.); дистиллированную воду добавляли до получения объема 20 мкл. Условия реакции ПЦР: предварительная денатурация при 94°С в течение 3 минут; денатурация при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд; элонгация при 72°С в течение 50 секунд; 25 циклов. Продукты ПЦР очищали электрофорезом в агарозном геле (1%), используя набор для очистки ДНК в геле (Watson Biotech. Inc.).
Указанный выше продукт ПЦР и плазмиду CD28 VH/pTRI одновременно расщепляли ScaI/ScaII (Promega Co.). Расщепленный продукт ПЦР (примерно 750 п.о.) и CD28 VH/pTRI (примерно 5700 п.о.) лигировали вместе и трансформировали штамм Е.coli TOP10. Плазмиды, выделенные из позитивных клонов, названы
CD3-scFv/CD28-VH/pTRI и идентифицированы в ПЦР с продуктом размером примерно 1400 п.о. (как показано на фиг.6). Все рабочие операции, необходимые в данном случае, описаны в разделе (1).
(4) Конструирование CEA-scTsAb/pTRI:
Конструирование анти-CEA-scFv в ПЦР с перекрыванием:
Анти-CEA-scFv конструировали связыванием VH (вариабельной области тяжелой цепи) и VL (вариабельной области легкой цепи) моноклонального анти-СЕА-антитела (Koga, Kanda et al., 1990) с полипептидом GGGGSGGGGSGGGGS (SEQ ID NO: 23). Полную аминокислотную последовательность анти-CEA-scFv обратно переводили в последовательность ДНК согласно таблице предпочтительного использования кодонов Е.coli (Nakamura, Gojobori et al., 2000), которую разделяли на 22 комплементарных олигонуклеотида. Синтезировали 22 олигонуклеотида, которые указаны ниже, и собирали в целый фрагмент ДНК, кодирующий анти-CEA-scFv, используя ПЦР с перекрыванием.


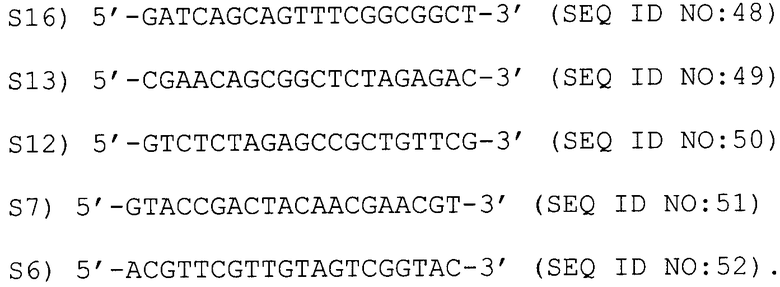
Стадии обработки:
Стадия 1. Как показано на фиг.7, попарно смешивали фрагменты (1-22) и осуществляли реакцию элонгации, как указано ниже.
Реакционная смесь: синтетические фрагменты, 1 мкл (каждого);
10 х буфер для ПЦР, 2 мкл; dNTP (каждый 2 ммоль/мл) (Dalian Takara Biotechnology Co. Ltd.) 2 мкл; Taq (1 ед.) (Dalian Takara Biotechnology Co. Ltd.) 0,5 мкл; дистиллированная вода, 14 мкл.
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 30 секунд; 10 циклов.
Все продукты собирали без очистки и непосредственно использовали для следующей стадии.
Стадия 2. Как показано на фиг.7, попарно смешивали продукты (А, В, D, Е, G, H, J, К) со стадии 1 и осуществляли реакцию элонгации, как указано ниже.
Реакционная смесь: продукты со стадии 1, 10 мкл (каждого).
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 30 секунд; 10 циклов.
Все продукты наносили на агарозный гель для электрофореза (1%) и очищали, используя набор для очистки ДНК из геля (Watson Biotech. Inc.). Фрагменты «а» и «g» имели размер примерно 120 п.о., фрагменты «с» и «е» имели размер примерно 170 п.о., фрагменты «d» и «f» имели размер примерно 100 п.о.
Стадия 3. Как показано на фиг.7, попарно смешивали продукты (а, b, с, d, е, f, g) со стадии 2 и осуществляли реакцию амплификации, как указано ниже. Праймеры S1 и S6 соответствовали паре «а» и «b»; праймеры S7 и S12 соответствовали паре «с» и «d»; праймеры S13 и S16 соответствовали «е», праймеры S17 и 22 соответствовали паре «f» и «g».
Реакционная смесь: продукты со стадии 2, по 1 мкл (каждого); праймеры, 1 мкг (каждого); 10 х буфер для ПЦР, 2 мкл; dNTP (2 ммоль/мл каждого) (Dalian Takara Biotechnology Co. Ltd.), 2 мкл; Taq (1 ед.) (Dalian Takara Biotechnology Co. Ltd.), 0,5 мкл; дистиллированная вода, 12 мкл.
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 30 секунд; 25 циклов.
Все продукты наносили на агарозный гель для электрофореза (1%) и очищали, используя набор для очистки ДНК из геля (Watson Biotech. Inc.). Фрагмент I имел размер примерно 200 п.о.; фрагмент II имел размер примерно 250 п.о.; фрагмент III имел размер примерно 140 п.о., фрагмент IV имел размер примерно 230 п.о.
Стадия 4. Как показано на фиг.7, попарно смешивали продукты (I, II, III, IV) со стадии 3 и осуществляли реакцию амплификации, как указано ниже. Праймеры S1 и S12 соответствовали паре I и II; праймеры S13 и 22 соответствовали паре III и IV.
Реакционная смесь: продукты со стадии 3, по 1 мкл (каждого); праймеры, 1 мкл (каждого); 10 х буфер для ПЦР, 2 мкл; dNTP (2 ммоль/мл каждого) (Dalian Takara Biotechnology Co. Ltd.), 2 мкл; Taq (1 ед.) (Dalian Takara Biotechnology Co. Ltd.), 0,5 мкл; дистиллированная вода, 12 мкл.
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 30 секунд; 25 циклов.
Все продукты наносили на агарозный гель для электрофореза (1%) и очищали, используя набор для очистки ДНК из геля (Watson Biotech. Inc.). Фрагмент UP имел размер примерно 430 п.о.; фрагмент DOWN имел размер примерно 340 п.о.
Стадия 5. Как показано на фиг.7, попарно смешивали продукты (UP и DOWN) со стадии 4 и осуществляли реакцию амплификации, как указано ниже. Праймеры S1 и 22 соответствовали паре UP и DOWN.
Реакционная смесь: продукт со стадии 4, по 1 мкл (каждого); праймеры, 1 мкл (каждого); 10 х буфер для ПЦР, 2 мкл; dNTP (2 ммоль/мл каждого) (Dalian Takara Biotechnology Co. Ltd.), 2 мкл; Taq (1 ед.) (Dalian Takara Biotechnology Co. Ltd.), 0,5 мкл; дистиллированная вода, 12 мкл.
Условия реакции: предварительная денатурация при 94°С в течение 1 минуты; денатурация при 94°С в течение 30 секунд; отжиг при 45°С в течение 30 секунд; элонгация при 72°С в течение 30 секунд; 25 циклов.
Все продукты наносили на агарозный гель для электрофореза (1%) и очищали, используя набор для очистки ДНК из геля (Watson Biotech. Inc.). Фрагмент WHOLE имел размер примерно 750 п.о.
Схема указанных выше стадий способа показана на фигуре 7, результаты ПЦР-идентификации показаны на фиг.8.
Указанный выше продукт ПЦР и плазмиду pTMF одновременно расщепляли XhoI/EcoRI (Promega Co.). Расщепленный продукт ПЦР (примерно 750 п.о.) и расщепленный продукт pTMF (примерно 5200 п.о.) лигировали вместе и трансформировали штамм Е.coli TOP10. Плазмиды, выделенные из позитивных клонов, названы CEA-scFv/pTMF и идентифицированы в ПЦР с продуктом размером примерно 750 п.о. Все рабочие операции, необходимые в данном случае, описаны в разделе (1).
Плазмиду CEA-scFv/pTMF и плазмиду CD3-scFv/CD28-Vн/pTRI одновременно расщепляли XhoI/EcoRI (Promega Co.). Продукт расщепления первой плазмиды (примерно 750 п.о.) и продукт расщепления второй плазмиды (примерно 6000 п.о.) лигировали вместе и трансформировали штамм Е.coli ТОР10. Плазмиды, выделенные из позитивных клонов, были названы CEA-scTsAb/pTRI и идентифицированы в ПЦР с продуктом размером примерно 2100 п.о. (как показано на фиг.6). Все рабочие операции, необходимые в данном случае, описаны в разделе (1).
Пример 3. Цитоплазматическая экспрессия растворимого СЕА-scTsAb, индуцированная при пониженной температуре.
(1) Трансформация Е.coli штамма линии BL21 (DE3) (Novagen) плазмидой СЕА-scTsAb/pTRI
Компетентные клетки Е.coli BL21 (DE3) готовили согласно способу, описанному в примере 2. Плазмиду CEA-scTsAb/pTRI выделяли, используя набор для выделения плазмид (Watson Biotech. Inc.). Последующие процедуры трансформации и идентификации позитивных клонов осуществляли в соответствии с примером 2.
(2) Индуцированная экспрессия при пониженной температуре
Одиночный клон BL21 (DE3), содержащий плазмиду CEA-scTsAb/pTRI, собирали из чашки LB-K и инокулировали на 5 мл среды LB-K. После культивирования при 37°С в течение ночи на качалке культуру переносили в 250 мл среды LB-K в соотношении 1/100, встряхивали при 37°С вплоть до достижения А600=0,6. Добавляли IPTG (Takara Biotech. (Dalian)) до конечной концентрации примерно 0,4 ммоль/л для индукции экспрессии растворимого продукта при 30°С в течение 4 часов. Бактериальные клетки собирали центрифугированием при 12000 об./мин в течение 10 минут и затем ресуспендировали в фосфатно-солевом буфере (PBS: 8 г NaCl, 0,2 г КСl, 1,44 г Na2HPO4, 0,24 г KH2PO4, pH 7,4, 1 литр) (1/5 объема культуральной среды). Таким образом цитоплазматическое растворимое CEA-scTsAb высвобождалось в надосадок, получаемый при центрифугировании лизированных ультразвуком клеток. Кроме того, экспрессию CEA-scTsAb в виде растворимого продукта и экспрессию CEA-scTsAb в виде телец включений выявляли в восстанавливающем SDS-ПААГ и на Вестерн-блотах согласно «Molecular cloning: a laboratory manual» (Sambrook and Russel, 2001). Результаты фотографировали, используя систему для регистрации и анализа Alpha-Image 2200 (Amerikan Alpha Innotech Company). Как показано на фиг.10, экспрессия растворимого СЕА-scTsAb составляет примерно 70% от общей экспрессии. Так как полученный после обработки ультразвуком надосадок можно непосредственно использовать на следующих стадиях очистки и непосредственно в анализах активности in vitro без необходимости в денатурации или ренатурации, то себестоимость и время получения может быть существенно сокращены.
Пример 4. Очистка CEA-scTsAb путем одностадийной анионообменной хроматографии на DEAE
250 мл культуральной среды, содержащей бактериальные клетки, экспрессирующие CEA-scTsAb, центрифугировали при 12000 об./мин при 4°С в течение 10 минут. Осадок ресуспендировали в 50 мл буфера для уравновешивания колонки с DEAE при анионообменной хроматографии (20 ммоль/л NaCl, 20 ммоль/л трис-HCl, рН 8,0) для дальнейшей ультразвуковой обработки. После второго центрифугирования при 12000 об./мин при 4°С в течение 10 минут надосадок, содержащий экспрессированное растворимое CEA-scTsAb, непосредственно использовали на стадии очистки.
20 мл анионообменной смолы DEAE (Amersham Bioscience) суспендировали в 100 мл буфера для уравновешивания и заполняли колонку 16×20 см (Shanghai Hua-mei). Колонку уравновешивали 5 объемами буфера для уравновешивания со скоростью 1 мл/мин. Затеем указанный выше надосадок наносили со скоростью 0,25 мл/мин. Очищенное CEA-scTsAb находилось в проходящем потоке. Колонку промывали или элюировали 2 объемами элюирующего буфера (500 ммоль/л NaCl, 20 ммоль/л трис-HCl, рН 8,0) со скоростью 0,25 мл/мин и очищали 2 объемами 500 ммоль/л NaOH со скоростью 0,5 мл/мин. Во время стадии регенерации колонку уравновешивали 2 объемами буфера для уравновешивания со скоростью 1 мл/мин.
Результат очистки, выявляемый при электрофорезе проходящей через колонку фракции в восстанавливающем SDS-ПААГ, показан на фиг.11. В результате большую часть бактериального белка в надосадке можно было удалить в результате одной стадии анионообменной хроматографии на DEAE. CEA-scTsAb составляло 70% проходящей через колонку фракции.
Затем очищенный образец диализовали против PBS при 4°С в течение ночи. Концентрацию белка определяли способом Бредфорда (Ausubel 1999). После добавления азида натрия (0,05% мас./об.) и фукозы (0,15 моль/л) диализованный раствор делили на аликвоты по 1 мл и хранили при -80°С.
Пример 5. Определение специфичности связывания по отношению к трем антигенам (СЕА, CD3, CD28) в ELISA
Подготовка антигена клеточной оболочки Jurkat: 5×106 клеток Jurkat (Американская коллекция типов культур, АТСС, TIB-152) собирали центрифугированием при 1000 g в течение 10 минут. Осадок клеток суспендировали в 0,5 мл PBS и лизировали обработкой ультразвуком. К надосадку, полученному центрифугированием обработанных ультразвуком клеток при 12000 об./мин в течение 10 минут, добавляли азид натрия 0,05% (мас./об.) и фукозу (0,15 моль/л), делили на аликвоты по 100 мкл и хранили при -80°С.
ELISA:
(1) Покрывание: очищенный СЕА (R&D), химеру rhCD28-Fc (R&D) и описанный выше очищенный мембранный антиген Jurkat разбавляли в буфере для покрывания (1,36 г Na2CO3, 7,35 г NaHCO3, 1 литр, рН 9,2) до концентрации 1 мкг/мл (СЕА и химера rhCD28-Fc) или 10 мкг/мл (мембранный антиген Jurkat) и вносили по 100 мкл/лунку в 96-луночный планшет для ELISA (Nunc). Планшет инкубировали при 37°С в течение 2 часов или при 4°С в течение ночи.
(2) Блокирование: покрытый планшет 1-2 раза промывали PBS и добавляли блокирующий буфер (РВS-1%БСА (бычий сывороточный альбумин, мас./об.) по 200 мкл/лунку. Планшет инкубировали 2 часа при 37°С.
(3) Добавление образцов: блокированный планшет 3 раза промывали PBS и добавляли разбавленные в PBS образцы по 100 мкл/лунку. Образцы CEA-scTsAb 6 раз серийно разводили из исходного раствора с концентрацией 10 мкг/мл. Планшет инкубировали при 37°С на 2 часа.
(4) Добавление первого антитела: планшет после добавления образца 3 раза промывали PBS-T (PBS-0,05%, твин-20 (мас./об.) и добавляли мышиное моноклональное антитело против cmyc-метки в разбавлении 1/1000 (Santa Crutz) no 100 мкл/лунку. Планшет инкубировали при 37°С в течение 2 часов.
(5) Добавление второго антитела: планшет после добавления первого антитела 3 раза промывали PBS-T и добавляли конъюгированное с HRP (пероксидазой хрена) антитело козы против IgG мыши (Santa Crutz) в разведении 1/1000 по 100 мкл/лунку. Планшет инкубировали при 37°С в течение 2 часов.
(6) Визуализация: планшет после добавления второго антитела 5 раз промывали PBS-T и добавляли раствор для визуализации, содержащий 10 мл буфера для субстрата (36,6 г моногидрата лимонной кислоты, 113,5 г двухосновного фосфата калия, 1 литр, рН 6,0) и 4 мг OPD (ортофенилендиамин-2 НСl, Sigma) no 100 мкг/лунку. Планшет помещали в темноту при комнатной температуре на 20 минут.
(7) Остановка реакции: чтобы остановить реакцию, добавляли раствор 1 моль/л НСl по 100 мкл/лунку.
(8) Измерение: результаты оптической плотности регистрировали при 490 нм.
Как показано на фиг.12, CEA-scTsAb очень специфично связывается с тремя антигенами: СЕА, экспрессируемым на высоком уровне в клетках Jurkat мембранным антигеном CD3 и химерой rhCD28-Fc.
Пример 6. Определение специфичности связывания с опухолевыми клетками посредством FCM
Непрямой способ FCM использовали для определения связывания с различными опухолевыми клетками. Происхождение таких опухолевых клеток указано ниже
Последовательность операций
(1) Культивирование и сбор опухолевых клеток: Три типа опухолевых клеток (А549, MCF-7, SK-OV-3) культивировали в среде RPMI 1640 (Gibco), содержащей 10% фетальную сыворотку теленка (FCS) с добавлением антибиотика (100 ед. пенициллина) в инкубаторе во влажной среде с 5% СO2 при 37°С. SW1116 культивировали в среде L15 (Gibco), содержащей 10% фетальной сыворотки теленка (FCS) с добавлением антибиотика (100 ед. пенициллина) в инкубаторе во влажной среде с 5% СO2 при 37°С. 5×105 опухолевых клеток в экспоненциальной фазе роста собирали центрифугированием при 1000 g в течение 10 минут и суспендировали в 100 мкл PBS.
(2) Инкубация CEA-scTsAb с опухолевыми клетками: CEA-scTsAb добавляли к раствору PBS, содержащему указанные выше опухолевые клетки в конечной концентрации 10 мкг/мл. Для каждого образца опухолевых клеток использовали контроль изотипа. Суспензии клеток инкубировали при 4°С в течение 30 минут.
(3) Инкубация первого антитела, мышиного моноклонального антитела против cmyc-метки (Santa Crutz), с опухолевыми клетками:
Несвязанное CEA-scTsAb удаляли, сливая надосадок после центрифугирования при 1000 g в течение 10 минут. Осадок клеток суспендировали в 100 мкл PBS, содержащего первое антитело в разбавлении 1/1000. Суспензии клетки инкубировали при 4°С в течение 30 минут.
(4) Инкубация второго антитела, конъюгированного с FITC антитела козы против IgG мыши, с опухолевыми клетками: Несвязанные первые антитела удаляли, сливая надосадок после центрифугирования при 1000 g в течение 10 минут. Осадок клеток суспендировали в 100 мкл PBS, содержащего второе антитело в разведении 1/1000. Суспензии клеток инкубировали при 4°С в течение 30 минут.
(5) FMC-анализ: Несвязанные вторые антитела удаляли, сливая надосадок после центрифугирования при 1000 g в течение 10 минут.
Осадок клеток суспендировали в 400 мкл PBS и анализировали, используя прибор FACS Colibur (BD). Длина волны возбуждения 488 нм. Каждый раз собирали 10000 клеток.
Как показано на фиг.13, CEA-scTsAb лучше всего связывается с SW1116 и SK-OV-3; связывание CEA-scTsAb с А549 является средним; CEA-scTsAb не связывается с MCF-7.
Пример 7. FMC-анализ специфичности связывания CEA-scTsAb с РВМС и клетками Jurkat
Способ прямого FMC-анализа использовали для тестирования специфичности связывания CEA-scTsAb с РВМС и клетками Jurkat.
Последовательность операций:
(1) Конъюгирование FITC (Sigma) с CEA-scTsAb: FITC конъюгировали с CEA-scTsAb, используя способ Кларка (Qui Fa-zhu et al., 2002).
(2) Получение РВМС: РВМС получали центрифугированием в градиенте фикола (Qui Fa-zhu et al., 2002) и культивировали в среде RPMI 1640 (Gibco), содержащей 10% фетальной сыворотки теленка с добавлением антибиотика (100 ед. пенициллина) в инкубаторе во влажной среде с 5% СO2 при 37°С. После инкубации в течение 4 часов суспендированные клетки, главным образом лимфоциты, переносили в новую колбу. Таким образом удаляли прикрепленные клетки.
(3) Культивирование клеток Jurkat: клетки Jurkat культивировали в среде RPMI 1640 (Gibco), содержащей 10% фетальной сыворотки теленка с добавлением антибиотика (100 ед. пенициллина) в инкубаторе во влажной среде с 5% СО2 при 37°С.
(4) Анализ FMC: 5×105 РВМС или клеток Jurkat в экспоненциальной фазе роста собирали центрифугированием при 1000 g в течение 10 минут и суспендировали в 100 мкл PBS, содержащего 10 мкг/мл конъюгированного с FITC CEA-scTsAb. Суспензии клеток инкубировали при 4°С в течение 30 минут и затем центрифугировали при 1000 g в течение 10 минут. Клеточный осадок ресуспендировали в 400 мкл PBS и анализировали, используя прибор FACS Colibur (BD). Длина волны возбуждения 488 нм. Каждый раз собирали 10000 клеток.
Как показано на фиг.14, CEA-scTsAb специфично связывается с РВМС и клетками Jurkat.
Суммируя результаты, описанные в примере 6 и примере 7, видно, что CEA-scTsAb может специфично связываться с РВМС, Jurkat и некоторыми опухолевыми клетками, что важно для индукции специфичного цитолиза опухолевых клеток.
Пример 8. Выявление специфичного цитолиза опухолевых клеток карциномы прямой и ободочной кишки, SW1116, индуцированного CEA-scTsAb, в присутствии лимфоцитов.
В системе анализа in vitro специфичного цитолиза опухолевых клеток использовали линию СЕА-экспрессирующих опухолевых клеток SW1116 в качестве клеток-мишеней (Т) и лимфоциты из РВМС (мононуклеарные клетки периферической крови) в качестве эффекторных клеток (Е). После смешивания клеток-мишеней и эффекторных клеток в определенном соотношении Е/Т и добавления CEA-scTsAb специфичный цитолиз опухолевых клеток индуцировали инкубацией при 37°С в течение 48 часов. Результаты оценивали в МТТ-анализе.
(1) РВМС получали, как описано в примере 7.
(2) Клетки SW1116 культивировали и собирали согласно примеру 6.
(3) Клетки SW1116 (1×105/мл) сначала помещали в 96-луночные планшеты (Nunc) по 100 мкл/лунку. Затем добавляли эффекторные клетки (РВМС) при разных соотношениях Е/Т (1, 5, 10) по 100 мкл/лунку. Добавляли концентрированные CEA-scTsAb по 50 мкл/лунку, чтобы достичь конечной концентрации в 1 мкг/мл. Смесь инкубировали в инкубаторе с 5% СO2 при 37°С в течение примерно 48 часов. Для каждой концентрации использовали по 4 лунки. Негативные контроли: лунки без CEA-scTsAb для каждого соотношения Е/Т; лунки, содержащие только эффекторные клетки; лунки, содержащие только клетки-мишени; лунки, не содержащие клеток.
(4) МТТ-анализ: Надосадки среды удаляли аспирацией и прикрепленные клетки один раз промывали PBS. В каждую лунку добавляли 200 мкл раствора МТТ (МТТ: бромид (3-(4,5-диметилтиазолил-2)-2,5-дифенилтетразолия, 500 мкг/мл, Sigma) и инкубировали в течение 4 часов при 37°С. Планшет один раз промывали PBS и в каждую лунку добавляли 200 мкл ДМСО (Sigma). Продолжали инкубировать при 37°С в течение 30 минут. Оптическую плотность в каждой лунке измеряли при длине волны 570 нм, вычитая фон при 620 нм.
(5) Специфичный цитолиз опухолевых клеток в процентах рассчитывали согласно формуле:
Процент специфичного цитолиза опухолевых клеток (%) = [А600(ЕТ)-А600(ЕТА)]/[А600(ЕТ)-А600(М)]×100%
А600 (ЕТ): оптическая плотность в лунках с негативным контролем без добавления CEA-scTsAb;
А600 (ЕТА): оптическая плотность лунок с образцом;
А600 (М): оптическая плотность лунок с негативным контролем, не содержащих клеток.
Влияние отношения Е/Т на специфичный цитолиз опухолевых клеток, индуцированный CEA-scTsAb, показано на фиг.15. Можно сделать вывод, что Е/Т=5 является оптимальным соотношением, при котором специфичный цитолиз опухолевых клеток достигает 85%. Это свидетельствует о том, что кроме соотношения Е/Т существуют другие факторы, влияющие на специфичный цитолиз опухолевых клеток. Влияние концентрации CEA-scTsAb, возрастающей от 0,4 нг/мл до 12 мкг/мл, на специфичный цитолиз опухолевых клеток при постоянном соотношении Е/Т, равном 5, показано на фиг.16. На кривой видно четыре ступенчатых фазы специфичного цитолиза опухолевых клеток. В первой фазе от 6 мкг/мл до 12 мкг/мл имеет место отрицательная корреляция эффективности специфичного цитолиза опухолевых клеток и концентрации CEA-scTsAb, и эффективность достигает пика при 6 мкг/мл. Во второй фазе, от 750 нг/мл до 6 мкг/мл, наблюдается прямая корреляция, и минимальная эффективность наблюдается при 750 нг/мл. В третьей фазе, от 24 нг/мл до 750 нг/мл, наблюдается возврат к отрицательной корреляции. В четвертой фазе, от 24 нг/мл до нуля, снова проявляется прямая корреляция. Во всяком случае существуют два пика специфичного цитолиза опухолевых клеток: 85% при 6 мкг/мл и 70% при 24 нг/мл. На основании приведенных выше результатов можно сделать вывод, что очень эффективный специфичный цитолиз опухолевых клеток может быть индуцирован даже при более низком соотношении Е/Т или более низкой концентрации CEA-scTsAb.
Пример 9. Морфологическое исследование опухолевых клеток в процессе специфичного цитолиза опухолевых клеток
После смешивания РВМС (эффекторных клеток) с клетками SW1116 (клетки-мишени) в среде L15 (10% FBS) при соотношении Е/Т, равном 5, и добавления очищенных CEA-scTsAb в концентрации 1 мкг/мл смесь инкубировали при 37°С в течение 20-40 часов в инкубаторе с 5% СO2. Морфологические изменения опухолевых клеток и РВМС наблюдали, используя 40 х линзы объектива в инвертированном микроскопе OLYMPUS IMT-2 и регистрировали на микрофотографиях. Как показано на фиг.18 (А)-(G), существуют четыре стадии морфологических изменений.
Пример 10. Определение пролиферации эффекторных клеток, инкубируемых с клетками-мишенями и CEA-scTsAb.
Пролиферацию эффекторных клеток (главным образом Т-лимфоцитов), выявляемую в МТТ-анализе, можно использовать для оценки активации Т-лимфоцитов, индуцированных совместной инкубацией опухолевых клеток и CEA-scTsAb.
Последовательность операций:
(1) РВМС получали согласно примеру 6.
(2) Опухолевые клетки SW1116 культивировали и собирали согласно примеру 6.
(3) Концентрацию клеток SW1116 доводили примерно до 106/мл в среде L15, содержащей митомицин С (25 мкг/мл, Sigma) и клеточную смесь инкубировали при 37°С в инкубаторе с 5% СO2 в течение 20 минут. После трехкратной промывки опухолевых клеток в PBS оставшийся митомицин С удаляли из культуральной среды.
(4) Концентрацию клеток SW1116 доводили до 105/мл и концентрацию РВМС доводили до 5×105/мл в среде L15. По 100 мкл клеток высевали в 96-луночный планшет и клетки инкубировали при 37°С в инкубаторе с 5% СO2 в течение 4 суток. Для каждой концентрации CEA-scTsAb использовали по 4 лунки. Отрицательные контроли: лунки без добавления CEA-scTsAb для каждого соотношения Е/Т; лунки, содержащие только эффекторные клетки; лунки, содержащие только клетки-мишени; лунки, не содержащие клеток.
(5) ММТ-анализ: Надосадок среды удаляли аспирацией и прикрепленные клетки один раз промывали PBS. В каждую лунку добавляли 200 мкл раствора МТТ (МТТ: бромид (3-(4,5-диметилтиазолил-2)-2,5-дифенилтетразолия, 500 мкг/мл, Sigma) и инкубировали в течение 4 часов при 37°С. Планшет один раз промывали PBS и в каждую лунку добавляли 200 мкл ДМСО (Sigma). Продолжали инкубировать при 37°С в течение 30 минут. Оптическую плотность в каждой лунке измеряли при длине волны 570 нм, вычитая фон при 620 нм.
(6) Индекс стимуляции (SI) рассчитывали согласно указанной ниже формуле:
SI=[А600(ЕТА)/А600(ЕТ)]
A600 (ET): оптическая плотность лунок с негативным контролем без добавления CEA-scTsAb;
А600 (ЕТА): оптическая плотность в лунках с образцами.
Как показано фиг.17, имеется три ступенчатых фазы изменения индекса стимуляции (SI). В первой фазе, от 750 нг/мл до 12 мкг/мл, наблюдается прямая корреляция SI и концентрации CEA-scTsAb, и наименьшее значение достигается при 750 нг/мл. Во второй фазе, от 50 нг/мл до 6 мкг/мл, наблюдается отрицательная корреляция, и пик достигается при 50 нг/мл. В третьей фазе, от 50 нг/мл до нуля, наблюдается возврат к прямой корреляции. Можно сделать вывод, что CEA-scTsAb обладает способностью стимулировать Т-лимфоциты даже при очень низких концентрациях. Также обнаружено, что специфичный цитолиз опухолевых клеток, индуцированный CEA-scTsAb, соответствовал активирующему состоянию совместно инкубируемых Т-лимфоцитов. Суммируя результаты примеров 4-9, авторы считают, что функция CEA-scTsAb сосредоточена на двух аспектах: (1) нацеливание эффекторных клеток на окружение опухолевых клеток и (2) стимуляция эффекторных клеток, вызывающих специфичную гибель клеток-мишеней. Как показано на фиг.19, нацеленные цитотоксические Т-лимфоциты (CTL) активируются для того, чтобы непосредственно уничтожать опухолевые клетки-мишени; Т-хелперные клетки секретируют цитокины, такие как Il-2, IFN-γ и TNF-α, косвенно помогая CTL или природным клеткам-киллерам (NK-клеткам) уничтожать опухолевые клетки-мишени.
Пример 11: Механизм специфичного цитолиза опухолевых клеток, индуцированного CEA-scTsAb
Существуют три пути, посредством которых активированные CTL уничтожают опухолевые клетки in vivo (Kagi, Vignaux et al., 1994; Lowin, Hahne et al., 1994): активированные CTL секретируют перфорины, делающие отверстия на поверхности мембран опухолевых клеток, которые затем разрушаются и подвергаются некрозу; гразимы, секретируемые активированными CTL, могут проникать в опухолевые клетки через указанные выше отверстия и индуцировать апоптоз; активированные CTL могут быть индуцированы к экспрессии Fas-лигандов на поверхности, которые взаимодействуют с молекулами Fas на опухолевых клетках и индуцируют апоптоз. В настоящем изобретении использовали FMC (флуоресцентную цитометрию) с двойным окрашиванием РI/аннексин-V-FITC и последующее флуоресцентное микрофотографирование, чтобы отличать некроз от апоптоза опухолевых клеток в анализе in vitro специфичного цитолиза опухолевых клеток.
(1) РВМС получали в соответствии с примером 7.
(2) Клетки SW1116 культивировали и собирали в соответствии с примером 6.
(3) Сначала высевали в каждую лунку 48-луночного планшета (Nunc) по 104 клеток SW1116 в среде L15. Затем добавляли эффекторные клетки (РВМС) в соотношении Е/Т, равном 5. Добавляли концентрированное CEA-scTsAb в среде L15 до конечной концентрации 1 мкг/мл. Смесь инкубировали при 37°С в инкубаторе с 5% СO2 в течение примерно 20 часов. Для каждой концентрации использовали по 4 лунки. Негативные контроли: лунки без CEA-scTsAb для каждого соотношения Е/Т; лунки, содержащие только эффекторные клетки; лунки, содержащие только клетки-мишени.
(4) суспендированные эффекторные клетки и некоторые погибшие опухолевые клетки собирали аспирацией среды из каждой лунки. Прикрепленные клетки обрабатывали трипсином и также собирали. Объединяли указанные клетки во флаконах объемом 1,5 мл.
(5) после однократной промывки в PBS готовили осадок клеток из каждой лунки центрифугированием при 1000 g в течение 10 минут и затем суспендировали в 100 мкл буфера для связывания (BD) и инкубировали с 5 мкл конъюгата FITC и аннексина V (BD) и 5 мкл раствора PI (Sigma, 50 мкг/мл). Смесь инкубировали при комнатной температуре в темноте в течение 10 минут.
(6) Результаты после добавления 300 мкг буфера для связывания и фотографирования в флуоресцентном микроскопе (Leica DMRA2) показаны на фиг 20.
(7) Разбавленную смесь клеток непосредственно анализировали, используя FCM (FACS Calibur, BD). Длина волны возбуждения 488 нм. Для каждого флакона собирали 20000 клеток.
Как показано на фиг.20, с помощью двух красителей можно было отличить ранний апоптоз, поздний апоптоз и некроз: клетки на ранней стадии апоптоза окрашены только зеленой флуоресценцией (конъюгат FITC и аннексина V); некротические клетки окрашены только красной флуоресценцией (PI); клетки на поздней стадии апоптоза имеют и зеленую, и красную флуоресценцию.
На фиг.21 четыре сектора представляют 4 состояния опухолевых клеток: верхний левый сектор (UL) - некротические клетки; верхний правый сектор (UR) - клетки на поздней стадии апоптоза; нижний левый сектор - живые клетки; нижний правый сектор - клетки на ранней стадии апоптоза. Типичные результаты показаны на фиг.21. Негативные контроли без добавления CEA-scTsAb: UL -5,94%, UR - 2,33%, LL - 90,17%, LR - 1,66%. Лунки с образцами: UL - 9,80%, UR - 21,25%, LL - 52,83%, LR - 16,12%. Можно сделать вывод, что индуцированы оба типа специфичного цитолиза опухолевых клеток: некроз и апоптоз. По сравнению с негативным контролем ранний апоптоз и поздний апоптоз возрастают в 9 раз, тогда как некроз возрастает в 2 раза.
Ссылки:


















| название | год | авторы | номер документа |
|---|---|---|---|
| ОДНОЦЕПОЧЕЧНОЕ ЦИКЛИЧНОЕ ТРИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2002 |
|
RU2355705C2 |
| ОПУХОЛЕСПЕЦИФИЧНОЕ АНТИТЕЛО ПРОТИВ EGFR И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2730605C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ С УСТОЙЧИВОСТЬЮ К РАСТВОРИМОМУ СЕА | 2010 |
|
RU2573893C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ С УСТОЙЧИВОСТЬЮ К РАСТВОРИМОМУ СЕА | 2006 |
|
RU2426743C2 |
| АНТИТЕЛО ПРОТИВ РЕЦЕПТОРА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 2013 |
|
RU2652880C2 |
| ФРАГМЕНТЫ АНТИТЕЛА, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ ЧЕЛОВЕЧЕСКОГО РАКОВО-ЭМБРИОНАЛЬНОГО АНТИГЕНА (CEA) | 2003 |
|
RU2294939C2 |
| НОВЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1(PD-1) | 2016 |
|
RU2757316C2 |
| МОЛЧАЩИЕ АНТИ-CD28-АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2001 |
|
RU2261723C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И ОДНОЦЕПОЧЕЧНЫЕ ФРАГМЕНТЫ АНТИТЕЛ ПРОТИВ КЛЕТОЧНО-ПОВЕРХНОСТНОГО СПЕЦИФИЧЕСКОГО ДЛЯ ПРОСТАТЫ МЕМБРАННОГО АНТИГЕНА | 2006 |
|
RU2458073C2 |
| АНТИ-VEGF-АНТИТЕЛО И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРЕДУПРЕЖДЕНИЯ, ДИАГНОСТИКИ ИЛИ ЛЕЧЕНИЯ РАКА ИЛИ СВЯЗАННОГО С АНГИОГЕНЕЗОМ ЗАБОЛЕВАНИЯ | 2014 |
|
RU2644245C2 |
Настоящее изобретение относится к области биотехнологии и иммунологии. Предложено рекомбинантное одноцепочечное триспецифическое антитело для лечения опухолей, экспрессирующих СЕА. Указанное антитело состоит из трех последовательно расположенных фрагментов антител: анти-CEA-scFv, анти-CD3-scFv и VH СD28-антитела, связанных двумя промежуточными линкерами (промежуточным линкером Fc и промежуточным линкером HSA). При необходимости к антителу может быть добавлена с-mус-метка или (Нis)6-метка на С-конце. Описана ДНК, кодирующая антитело, экспрессирующий вектор на ее основе и клетка Е.coli, содержащая вектор. Использование изобретения приводит к преимуществам при клиническом применении по сравнению с биспецифическими антителами и известными триспецифическими антителами, облегчает очистку и экспрессию антитела, что может найти дальнейшее применение при лечении опухолей, опосредованных СЕА. 4 н. и 6 з.п. ф-лы, 21 ил., 1 табл.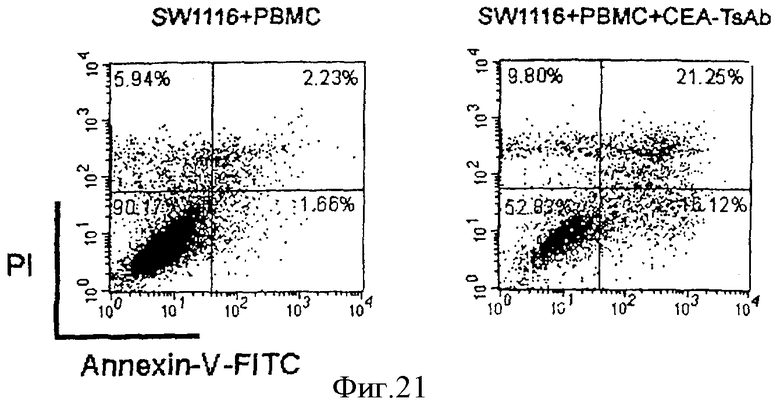
1. Линейное одноцепочечное триспецифическое антитело (scTsAb) для лечения опухолей, экспрессирующих СЕА, состоящее из антитела против раково-эмбрионального антигена (СЕА), соединяющего пептида Fc, анти-CD3-антитела, соединяющего пептида сывороточного альбумина человека (HSA) и VH анти-СD28-антитела.
2. scTsAb по п.1, в котором с-mус-метка и (His)6-метка добавлены в качестве альтернативы или одновременно к его С-концу.
3. scTsAb no п.1, в котором анти-СЕА-антитело, анти-СD3-антитело и анти-СD28-антитело являются одноцепочечным фрагментом вариабельной области (scFv), Fab-фрагментом или тяжелой вариабельной цепью (VH) антитела.
4. scTsAb по п.1, которое последовательно слито из одноцепочечного фрагмента вариабельной области (scFv) анти-СЕА-антитела, соединяющего пептида Fc, aнти-CD3-scFv, соединяющего пептида HSA и тяжелой вариабельной цепи (VH) анти-СD28-антитела и содержит аминокислотную последовательность SEQ ID NO: 4.
5. scTsAb по п.4, в котором анти-CEA-scFv содержит аминокислотную последовательность SEQ ID NO: 1.
6. scTsAb по п.4, в котором анти-CD3-scFv содержит аминокислотную последовательность SEQ ID NO: 2.
7. Последовательность ДНК, кодирующая scTsAb по п.4, содержащая последовательность нуклеиновой кислоты SEQ ID NO.3.
8. Экспрессирующий вектор, содержащий последовательность ДНК по п.7.
9. Экспрессирующий вектор по п.8, сконструированный клонированием анти-СЕА одноцепочечного триспецифического антитела (CEA-scTsAb) в pTRI.
10. Клетка-хозяин E.coli, содержащая экспрессирующий вектор по п.8.
| SONG Li-Ping et al., "A new model of trispecific antibody resulting the cytotoxity directed against tumor cells", Acta biochimica et biophysica sinica, 2003, 35(6), с.503-510 | |||
| Behr et al., "Variables influencing tumor dosimetry in radioimmunotherapy of CEA-expressing cancers with anti-cea and antimucin monoclonal antibodies", J.Nucl Med., 1997, 38, с.409-418 | |||
| US 5855886 A, 05.01.1999 | |||
| МУЛЬТИСПЕЦИФИЧЕСКАЯ ИЛИ БИСПЕЦИФИЧЕСКАЯ МОЛЕКУЛА (ВАРИАНТЫ), СПОСОБ УНИЧТОЖЕНИЯ ИЛИ УМЕНЬШЕНИЯ КОЛИЧЕСТВА КЛЕТОК-МИШЕНЕЙ В ОРГАНИЗМЕ СУБЪЕКТА, СПОСОБ ЛЕЧЕНИЯ СУБЪЕКТА, ИНФИЦИРОВАННОГО ПАТОГЕНОМ И СПОСОБ ВАКЦИНАЦИИ СУБЪЕКТА | 1997 |
|
RU2201766C2 |

