Предлагаемое изобретение относится к медицине, более точно к биофизике, биомеханике, и может быть использовано для определения жесткости костамера мышечных волокон.
Мышечное волокно является трехмерной механической конструкцией, в которой передача усилия осуществляется как в продольном, так и в поперечном направлении (Ervasti J.M. Costameres: the Achilles' Heel of Herculean muscle // Journal of Biological Chemistry. 2003. Vol.278. P.13591-13594). Эффективность такой передачи обусловлена структурными особенностями мышечного волокна, которое представляет собой вытянутую в продольном направлении клеточную структуру, большую часть объема которой занимает сократительный аппарат, образованный миофибриллами. В целом, механические характеристики мышечного волокна определяются структурно-функциональным взаимодействием трех компартментов с принципиально различными механическими свойствами: миофибриллярный аппарат, внесаркомерный цитоскелет (костамеры) и сарколемма.
Особый интерес вызывает определение жесткости костамера в связи с сигнальной и структурной ролью формирующих его различных белков в механотрансдукции. Так, показано, что мутации в некоторых белках костамера ведут к развитию миопатий и миодистрофий, таких как конечностно-поясная дистрофия типов 2С, 2D, 2Е, 2F, связанная с изменением саркогликанового комплекса (Bonneman С.G., McNally Е.М., Kunkel L.M. Beyond dystrophin: current progress in the muscular dystrophies // Curr. Opin. Pediatr. 1996. Vol.8. P.569-582); врожденная (конгенитальная) мышечная дистрофия, при которой изменен ген ламинина (Pegoraro Е., Marks H., Garcia С.А., Crawford Т., Mancias P., Connolly A.M., Fanin M, Martinello F., Trevisan C.P., Angelini C, Stella A., Scavina M., Munk R.L., Servidei S., Bonnemann C.C., Bertorini Т., Acsadi G., Thompson C.E., Gagnon D., Hoganson G., Carver V., Zimmerman R.A., Hoffman E.P. Laminin alpha2 muscular dystrophy: genotype/phenotype studies of 22 patients // Neurolgy. 1998. Vol.51. P.101-110); миодистрофии Беккера и Дюшенна, ассоциированные с мутацией в гене дистрофина, которая приводит к отсутствию стабильной молекулы дистрофина в клетках и, как результат, к нестабильности сарколеммы и индуцируемому нагрузками некрозу мышечных волокон (Hoffman Е.Р., Gorospe J.R.M. The animal models of Ducehnne muscular dystrophy: windows on the pathophysiologic consequences of dystrophin deficiency // Curr. Top. Membr. Transp.1991. Vol.38. P.113-154).
Полагая, что изменение механических свойств костамера под действием внешних факторов, может приводить к запуску различных сигнальных путей, представляется особенно важным дифференцированно и корректно определять жесткость костамера. Результат таких измерений, возможно, позволит глубже проникнуть в малопонятный к настоящему времени механизм связи между мутациями в генах, кодирующих различные белки внесаркомерного цитоскелета, и механическими свойствами мышечных волокон, что может дать возможность в дальнейшем разработать методы коррекции этих патологических явлений.
В настоящее время наиболее информативным методом, применяемым для получения информации о поперечной жесткости как миофибриллярного аппарата, так и сарколеммы со связанными с ней костамерами является атомная силовая микроскопия. Определение механических характеристик обычно проводят в контактном режиме в жидкости, снимая силовые кривые, и, затем, используя решение контактной задачи Герца, вычисляют модуль Юнга и поперечную жесткость.
В современной литературе представлено относительно небольшое количество работ, посвященных исследованию поперечной жесткости как целых мышечных волокон, так и одиночных миофибрилл. Большинство результатов получено на интактных мышцах кроликов и мышей различных линий, а также на культурах клеток и частично дифференцированных миобластах. Так, Collinsworth A.M. et al. (Collinsworth A.M., Zhang S., Kraus W.E., Truskey G.A. Apparent elastic modulus and hysteresis of skeletal muscle cells throughout differentiation // Am J. Physiol Cell Physiology. 2002. Vol.283. P.1219-1227) на недифференцированных мышечных клетках мышей показали, что модуль Юнга составляет 11,5±1,3 кПа, а на 8-ой день дифференцировки повышается в 4 раза и достигает значения 45,3±4,0 кПа. Mathur А.В. et al. (Mathur А.В., Collinsworth A.M., Reichert W.M., Kraus W.E., Truskey G.A. Endothelial, cardiac muscle and skeletal muscle exhibit different viscous and elastic properties as determined by atomic force microscopy // Journal of Biomechanics. 2001. Vol.34. P.1545-1553) определили модуль Юнга C2C12 миобластов мышей линии С3Н равным 24,7±3,5 кПа. Этот же показатель для дифференцированных мышечных волокон мышей линии CD1 составляет 61±5 (Defranchi Е., Bonaccurso Е., Tedesco М., Canato М., Pavan Е., Raiteri R., Reggiani С. Imaging and elasticity measurements of the sarcolemma of fully differentiated skeletal muscle fibres // Microscopy Research and Technique. 2005. Vol.67. P.27-35).
Однако в представленных работах исследователи не дифференцировали участки, на которых были определены механические характеристики, в связи с чем остается неясным вопрос о причинах существенных различий в данных авторов. Можно лишь предполагать, что это обусловлено как неравномерностью отногенетической экспрессии белков внесаркомерного цитоскелета, так и видовыми различиями, но наиболее вероятным кажется то, что исследователи проводили измерения на различных участках мышечного волокна, не дифференцируя их.
Кроме того, в большинстве представленных работах абсолютные значения поперечной жесткости и модуля Юнга получены при фиксированном значении приложенной силы. Однако методика атомной силовой микроскопии при снятии силовых кривых такова, что начальное отклонение кантилевера вносит существенный вклад в приложенную силу, что необходимо учитывать, определяя механические свойства образцов. В качестве прототипа нами выбран способ определения поперечной жесткости мышечных волокон, описанный Mathur А.В., Collinsworth A.M., Reichert W.M., Kraus W.E., Truskey G.A. (Endothelial, cardiac muscle and skeletal muscle exhibit different viscous and elastic properties as determined by atomic force microscopy // Journal of Biomechanics. 2001. Vol.34. P.1545-1553). В известном способе определение поперечной жесткости мышечных волокон проводят путем выделения мышечных волокон, снятия силовых кривых и проведения математических расчетов. Этому известному способу присущи все недостатки ранее цитированных аналогичных способов определения жесткости.
Техническим результатом заявленного изобретения является повышение точности определения жесткости костамера мышечных волокон за счет предварительного сканирования поверхности образца. Нами показано, что учет начального отклонения кантилевера и фиксирование глубины продавливания позволяет более точно определить жесткость и модуль Юнга.
Этот технический результат достигается тем, что в известном способе определения поперечной жесткости мышечных волокон путем выделения их, снятия силовых кривых и проведения математичесих расчетов, проводят выделение одиночного мышечного волокна, прикрепляют его к дну жидкостной ячейки атомного силового микроскопа, определяют на воздухе резонансную частоту колебаний, подводят кантилевер к дну жидкостной ячейки, получают в контактном режиме калибровочную силовую кривую и определяют по ней калибровочный коэффициент а (в м/А), далее определяют локализацию костамера путем сканирования поверхности волокна, переводят систему в контактный режим и получают силовые кривые на выпуклостях, соответствующих костамеру, по которым находят отклонение кантилевера (в А), обобщенную глубину продавливания (в м) с последующим вычислением реальной глубины продавливания волокна (hs) и реальной приложенной к нему силы (Fs) по формулам
hs=x-y·a,
Fs=y·a·kc,
где hs - реальная глубина продавливания (в м),
x - обобщенная глубина продавливания (в м),
y - измеряемое отклонение кантилевера (в А),
a - калибровочный коэффициент (в м/А),
Fs - реальная приложенная к волокну сила (в Н),
kc - коэффициент жесткости кантилевера (в Н/м);
после чего определяют поперечную жесткость костамера ks (в Н/м) по формуле 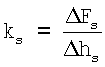 ,
,
где Δhs - изменение глубины продавливания волокна (в м),
ΔFs - соответствующее Δhs изменение приложенной к волокну силы (в Н),
и затем строят кривую Fs(hs) и аппроксимируют ее следующей зависимостью, отражающей решение контактной задачи Герца для скругленного кончика кантилевера:
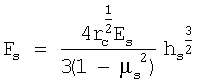 ,
,
где µs - коэффициент Пуассона, который, считая клетку несжимаемой, обычно принимают равным 0,5; rc - радиус скругления кончика кантилевера. Исходя из значения параметра, связывающего приложенную силу и глубину продавливания, а также зная радиус скругления кончика, вычисляют модуль Юнга костамера, [Es]=Па.
О том, что используемые показатели дают более точное определение поперечной жесткости свидетельствует наблюдения Defranchi Е. et al. (Defranchi Е., Bonaccurso Е., Tedesco М., Canato М., Pavan Е., Raiteri R., Reggiani С. Imaging and elasticity measurements of the sarcolemma of fully differentiated skeletal muscle fibres // Microscopy Research and Technique. 2005. Vol.67. P.27-35), где показано наличие регулярных выпуклостей поверхности, с характерным периодом, равным длине саркомера у мышечных волокон. Представляется совершенно очевидным, что эти выпуклости сформированы белками внесаркомерного цитоскелета, которые ориентированы перпендикулярно сократительному аппарату. Наиболее насыщенным регионом такого рода является проекция Z-диска, а именно костамер. Таким образом, можно утверждать, что различимые при сканировании поверхности волокна выпуклости сформированы костамером.
Длина костамера, то есть расстояние от поверхности волокна до миофибриллярного аппарата, ориентировочно составляет по разным данным от 100 нм до 500-700 нм. При этом жесткость, определенная при глубинах продавливания до 100 нм, отражает скорее жесткость мембраны, а более 500-700 нм - жесткость сократительного аппарата.
Сканирование поверхности в полуконтактном режиме заявляемого способа можно осуществлять с помощью более мягкого контактного кантилевера с тем, чтобы затем провести снятие силовых кривых в районе выпуклостей поверхности, не прибегая к замене датчика и, тем самым, не рискуя сместить образец.
Способ осуществляется следующим образом.
Одиночные мышечные волокна, выделенные либо из целой мышцы животного, либо из биоптатов мышцы человека, прикрепляют к дну жидкостной ячейки атомного силового микроскопа, каким-либо образом фиксируя их концы (например, с помощью специального клея Fluka shellac wax-free фирмы Sigma, Германия), и заполняют ячейку жидкостью (например, релаксирующим раствором).
Сначала на воздухе определяют резонансную частоту колебаний каждого используемого кантилевера, и затем вычисляют его жесткость kc (в Н/м).
Далее подводят зонд к дну жидкостной ячейки и в контактном режиме снимают калибровочную силовую кривую, отражающую зависимость отклонения кантилевера (в нА) от смещения головки (в нм) и вычисляют коэффициент пересчета а (в м/А).
Затем в полуконтактном режиме подводят зонд к мышечному волокну и сканируют поверхность. Далее переводят систему в контактный режим и снимают силовые кривые в интересующих точках поверхности волокна, а именно на выпуклостях, соответствующих костамеру, получая зависимость y(x), где [y]=нА, [x]=нм. Затем пересчитывают эти зависимости с учетом калибровочной кривой следующим образом. Реальная глубина продавливания волокна и приложенная к нему сила вычисляются так:
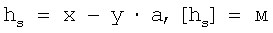
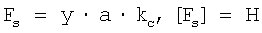
Затем строят кривую Fs(hs) и аппроксимируют ее следующей зависимостью, отражающей решение контактной задачи Герца для скругленного кончика кантилевера:
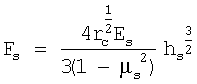 ,
,
где µs - коэффициент Пуассона, который, считая клетку несжимаемой, обычно принимают равным 0,5; rc - радиус скругления кончика кантилевера. Исходя из значения параметра, связывающего приложенную силу и глубину продавливания, а также зная радиус скругления кончика, вычисляют модуль Юнга костамера, [Es]=Па.
Далее определяют поперечную жесткость на фиксированной глубине продавливания (в нашем случае - 150 нм) по формуле
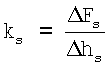 ,
, 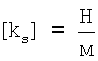 .
.
Пример реализации способа.
В эксперименте использовали семь половозрелых самцов крыс линии Вистар в возрасте 2 месяцев, выращенных в питомнике ГНЦ РФ ИМБП РАН. Животных содержали в стандартных условиях. Они получали корм в соответствии с рационом для лабораторных животных и воду ad libitum. Все процедуры с животными были одобрены Комиссией по биомедицинской этике ГНЦ РФ ИМБП РАН.
Камбаловидную мышцу вырезали от сухожилия до сухожилия и, с целью частичного разрушения клеточной мембраны, обрабатывали согласно методике химического сканирования, которая ранее была описана L.Stevens et al. (Stevens L., Mounier Y., Holy X. Functional adaptation of different rat skeletal muscles to weightlessness // Am. J. Physiol. 264 (4, pt 2): 770-776. 1993). До момента проведения экспериментов пробы хранили при температуре -20°С в буфере, содержащем в равных долях по объему расслабляющего раствора R (20 мМ MOPS, 170 мМ пропионат калия, 2,5 мМ ацетата магния, 5 мМ K2EGTA, 2,5 мМ АТФ) и глицерола.
В день эксперимента пробы переносили в раствор R, в котором выделяли одиночные мышечные волокна. Для измерения жесткости костамера выделенные волокна прикрепляли к дну жидкостной ячейки атомно-силового микроскопа, фиксируя их кончики с помощью специального клея Fluka shellac wax-free (Sigma).
Измерения жесткости костамера мышечных волокон проводили, используя инвертированный микроскоп Olympus XI, соединенный с атомной силовой головкой SMENA™ (NT-MDT, Россия). Жесткость (Н/м) для каждого кантилевера корректировалась по положению резонанса.
Для работы в жидкости использовали самые мягкие кантилеверы с коэффициентом жесткости на уровне 0,05 Н/м. Применялся полуконтактный режим для получения изображения и контактный для измерения поперечной жесткости. Радиус rc кривизны кончика всех использованных кантилеверов считался равным 10 нм.
Положение костамера определяли по изображению, формируемому при сканировании образца, принимая за искомую структуру периодические выпуклости поверхности волокна.
Для определения глубины продавливания hs мягкого волокна использовалось две силовые кривые: измеренная в точке интереса на волокне и измеренная на стекле при тех же значениях приложенной силы Fs.
Поперечную жесткость образца ks (пН/нм) находили по формуле
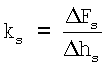 ,
,
при этом максимальная глубина продавливания составляла 300 нм.
Модуль Юнга образца Es (кПа) вычисляли, используя решение контактной задачи Герца по формуле
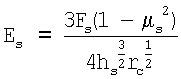 ,
,
где µs=0,5.
Полученные в ходе эксперимента результаты статистически обрабатывали с помощью стандартных методов, реализованных в Microsoft Excel. Данные представляли в виде М±m, где М - среднее значение оцениваемой величины, m - стандартная ошибка среднего значения.
В полуконтакном режиме сканирования получено изображение поверхности мышечного волокна крысы, на котором отчетливо видны выпуклости поверхности, ассоциированные с наличием подмембранной структуры - костамера.
Для определения механических характеристик были сняты силовые кривые в трех типах точек и определены поперечная жесткость и модуль Юнга различных типов структур мышечных волокон (таблица 1): на утолщении поверхности волокна (костамер), на мембране посередине между костамерами (проекция М-линии) и между проекцией М-линии и костамером (собственно поперечная жесткость сарколеммы).

Полученные результаты свидетельствуют о неоднородности механических характеристик вдоль мышечного волокна, которая обусловлена, по-видимому, различной структурой подмембранных участков мышечного волокна.
Среди различных регионов мышечного волокна наиболее жестким является костамер, поскольку его формирует большое количество филаментных белков, собственная жесткость которых весьма существенна.
Наиболее податливым к внешним нагрузкам является мембрана мышечного волокна между костамером и проекцией М-линии, что, вероятнее всего, связано с незначительностью подмембранного каркаса из цитоскелетных внесаркомерных белков.
Следует отметить, что полученные нами результаты впервые отражают механические характеристики мышечного волокна в поперечном направлении с дифференциацией различных областей. Кроме того, в своих измерениях мы ориентировались на глубину продавливания волокна, что позволило определить отдельно жесткость мембраны и внесаркомерного цитоскелета.
Нами показано, что:
- костамер формирует выпуклости поверхности мышечного волокна;
- жесткость костамера и его модуль Юнга составляют 9,10±0,14 пН/нм и 90,1±1,3 кПа соответственно;
- жесткость и модуль Юнга в проекции М-линии составляют 7,26±0,08 пН/нм и 69,2±0,6 кПа соответственно;
- жесткость и модуль Юнга сарколеммы мышечного волокна крысы составляют 6,36±0,13 пН/нм и 54,9±0,8 кПа соответственно.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ поддержания силы и жесткости постуральных мышц млекопитающих при их функциональной разгрузке | 2022 |
|
RU2800317C1 |
| ИНГИБИТОР ПРОТЕОЛИЗА ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ФИЗИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2389486C1 |
| СПОСОБ ПРОФИЛАКТИКИ РАЗРУШЕНИЯ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ЭКСЦЕНТРИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2391968C1 |
| СПОСОБ ИССЛЕДОВАНИЯ БИОСОВМЕСТИМОСТИ СИНТЕТИЧЕСКОГО МАТЕРИАЛА МЕДИКО-БИОЛОГИЧЕСКОГО НАЗНАЧЕНИЯ С БИОЛОГИЧЕСКИМИ ТКАНЯМИ | 2007 |
|
RU2381485C2 |
| Способ предотвращения атрофии скелетных мышц при их функциональной разгрузке | 2022 |
|
RU2797216C1 |
| СПОСОБ УСКОРЕНИЯ ВОССТАНОВЛЕНИЯ СКЕЛЕТНЫХ МЫШЦ ОТ АТРОФИИ ПОСЛЕ ДЛИТЕЛЬНОЙ АЛКОГОЛЬНОЙ ИНТОКСИКАЦИИ | 2014 |
|
RU2578463C1 |
| Способ изготовления дисковых секторов для захвата, удержания и анализа магнитных микрочастиц и меченных ими биологических объектов на поверхности спиновых вентилей с помощью фемтосекундного лазерного облучения | 2019 |
|
RU2704972C1 |
| СПОСОБ ОЦЕНКИ АНАБОЛИЧЕСКОГО ДЕЙСТВИЯ ЛЕКАРСТВЕННЫХ ПРЕПАРАТОВ | 2014 |
|
RU2568903C2 |
| Способ предотвращения атрофии, повышенной утомляемости медленных мышечных волокон и их трансформации в быстрые мышечные волокна в камбаловидной мышце | 2024 |
|
RU2834246C1 |
| СРЕДСТВО ДЛЯ СТИМУЛЯЦИИ СИНТЕЗА ГЛИКОГЕНА В СЕРДЕЧНОЙ МЫШЦЕ И СПОСОБ СТИМУЛЯЦИИ СИНТЕЗА ГЛИКОГЕНА В СЕРДЕЧНОЙ МЫШЦЕ | 2010 |
|
RU2438679C1 |
Изобретение относится к медицине, более точно к биофизике, биомеханике, и может быть использовано для определения жесткости костамера мышечных волокон. Проводят выделение одиночного мышечного волокна, прикрепляют его к дну жидкостной ячейки атомного силового микроскопа, определяют на воздухе резонансную частоту колебаний. Подводят кантилевер к дну жидкостной ячейки, получают в контактном режиме калибровочную силовую кривую и определяют по ней калибровочный коэффициент а (в м/А). Определяют локализацию костамера путем сканирования поверхности волокна. Переводят систему в контактный режим и получают силовые кривые на выпуклостях, соответствующих костамеру, по которым находят отклонение кантилевера (в А), обобщенную глубину продавливания (в м) с последующим вычислением реальной глубины продавливания волокна (hs) и реальной приложенной к нему силы (Fs) по формулам hs=x-y·a, Fs=y·a·kc, где hs - реальная глубина продавливания (в м), x - обобщенная глубина продавливания (в м), y - измеряемое отклонение кантилевера (в А), а - калибровочный коэффициент (в м/А), Fs - реальная приложенная к волокну сила (в Н), kc - коэффициент жесткости кантилевера (в Н/м). Способ позволяет повысить точность определения жесткости костамера мышечных волокон. 1 табл.
Способ определения жесткости костамера мышечных волокон путем выделения их, снятия силовых кривых и проведения математических расчетов, отличающийся тем, что проводят выделение одиночного мышечного волокна, прикрепляют его к дну жидкостной ячейки атомного силового микроскопа, определяют на воздухе резонансную частоту колебаний, подводят кантилевер к дну жидкостной ячейки, получают в контактном режиме калибровочную силовую кривую и определяют по ней калибровочный коэффициент а (м/А), далее определяют локализацию костамера путем сканирования поверхности волокна, переводят систему в контактный режим и получают силовые кривые на выпуклостях, соответствующих костамеру, по которым находят отклонение кантилевера (А), обобщенную глубину продавливания (м) с последующим вычислением реальной глубины продавливания волокна (hs) и реальной приложенной к нему силы (Fs) по формулам
hs=x-y·а,
Fs=y·a·kc,
где hs - реальная глубина продавливания, м,
x - обобщенная глубина продавливания, м,
y - измеряемое отклонение кантилевера, А, a - калибровочный коэффициент, м/А),
Fs - реальная приложенная к волокну сила, Н,
kc - коэффициент жесткости кантилевера, Н/м;
после чего определяют поперечную жесткость костамера ks, (Н/м) по формуле:
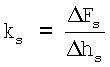 ,
,
где Δhs - изменение глубины продавливания волокна, м,
ΔFs - соответствующее Δhs изменение приложенной к волокну силы (Н) и затем строят кривую Fs(hs) и аппроксимируют ее следующей зависимостью, отражающей решение контактной задачи Герца для скругленного кончика кантилевера
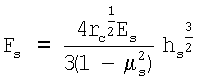 ,
,
где µs - коэффициент Пуассона, который, считая клетку несжимаемой, обычно принимают равным 0,5; rc - радиус скругления кончика кантилевера,
исходя из значения параметра, связывающего приложенную силу и глубину продавливания, а также зная радиус скругления кончика вычисляют модуль Юнга костамера [Еs]=Па.
| Mathur АВ et al | |||
| Endothelial, cardiac muscle and skeletal muscle exhibit different viscous and elastic properties as determined by atomic force microscopy | |||
| J Biomech | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| Способ определения жесткости и демпфирования мышцы человека | 1988 |
|
SU1586680A1 |
| Гевлич Г.И., Григорьева Л.С, Бойко М.И., Козловская И.Б | |||
| Оценка тонуса скелетных мышц методом регистрации поперечной | |||

