Изобретение относится к медицине, более точно к спортивной медицине, авиакосмической медицине, физиологии спорта, спорту, и может быть использовано в практике подготовки спортсменов или же других лиц, подвергающихся эксцентрическим нагрузкам.
Эксцентрические нагрузки представляют собой такие физические нагрузки, при которых происходит сокращение мышцы на фоне ее растяжения. При этом часто наступает деструкция белков скелетных мышц. В особенности это относится к цитоскелетным и мембранным белкам.
Аналогичные разрушения возникают при гипокинезии и гравитационной разгрузке, при которых цитоскелетные и сократительные белки скелетных мышц подвергаются деструкции [1-4]. Эти разрушения были обнаружены как в мышцах человека после различных видов нагрузок (что приводит к снижению работоспособности и появлению отставленной мышечной боли), так и в мышцах животных. Физиологические и клеточные факторы, приводящие к деструкции цитоскелетных белков, остаются неизвестными.
Известно, что эксцентрическое сокращение обладает наиболее повреждающим эффектом на цитоскелетные белки мышечного волокна (десмин, дистрофии, спектрин, актинин) [1]. После единичной эксцентрической нагрузки (например, бег под уклон вниз на тредбане, спуск по лестнице) как у людей, так и у экспериментальных животных насчитывалось до 30 процентов мышечных волокон, негативных по десмину [2, 3, 4]. У них также отмечено существенное количество волокон, имеющих разрывы в дистрофиновом и спектриновом слое мембран [5, 6], в некоторых работах обнаружили разрушения титина и небулина. При сокращении растянутой мышцы (эксцентрическая тренировка) восстановление мышечной работоспособности происходит только через 24 часа или позже [7, 8, 9, 10]. Это может быть связано с разрушением цитоскелетных белков при такой нагрузке.
Интересно отметить, что таких разрушительных последствий для мышцы не наблюдалось после короткой спринтерской тренировки или после циклических тренировок выносливости у тренированных людей [11, 12], а восстановление мышечной работоспособности в этом случае происходило в течение часа.
Известны работы, в которых показана взаимосвязь между изменением кальция при эксцентрической тренировке и активацией кальпаинов, разрушающих цитоскелетные белки [9, 11, 12, 13]. При блокировании L-кальциевых каналов нифедипином эти разрушения удается предотвратить [14, 15].
Так, согласно наблюдениям Beaton Louise J. и соавторов [16] для предотвращения повреждений мышц и мышечных белков при эксцентрической нагрузке в течение 7 дней перед однократной эксцентрической нагрузкой вводили аналог нифедипина амлодипин. Такое превентивное введение этого блокатора кальциевых каналов приводило к достоверному уменьшению процессов деструкции при эксцентрических нагрузках.
Этот способ предупреждения разрушения цитоскелетных белков при эксцентрической нагрузке был выбран нами в качестве прототипа заявляемого изобретения. Однако механизм действия этого препарата в корне отличается от предложенного нами.
Недостатком этого способа, на наш взгляд, является то, что нифедипин и амлодипин, являясь блокатором кальциевых каналов, может приводить к побочным эффектам воздействия на сердце, так как происходит блокада L-кальциевых каналов кардиомиоцитов.
Техническим результатом способа профилактики разрушения цитоскелетных белков при эксцентрической нагрузке является повышение эффективности способа в виде практически полного предупреждения разрушений цитоскелетных белков скелетных мышц, а также устранение побочных негативных эффектов применяемого ранее профилактического средства.
Этот технический результат достигается тем, что в известном способе профилактики разрушения цитоскелетных белков при эксцентрической нагрузке, включающем предварительное (до эксцентрической нагрузки) введение в организм химических соединений, в качестве химического соединения в организм вводят соединения, обеспечивающие увеличение оксида азота в скелетных мышцах.
В качестве соединения, обеспечивающего увеличение оксида азота в скелетных мышцах, вводят донор оксида азота - L аргинин.
Введение в организм донора азота осуществляют в течение 1-2 дней.
Способ осуществляют следующим образом.
В организм за 1-2 дня до эксцентрической нагрузки вводят соединения, которые обеспечивают увеличение оксида азота в мышцах. Такими соединениями могут быть доноры оксида азота в мышцах, например L-аргинин.
Количество вводимого соединения подбирают экспериментальным путем. Как правило, доза L -аргинина составляет 500 мг на кг веса тела.
Подтверждением решения поставленной задачи являются следующие исследования.
28 3-месячных самцов крыс Wistar были разделены на 4 группы: одна группа животных служила контролем (n=7, вес тела 276,0±29,5 г; масса m.soleus 139,2±31,1 мг; гр. С); крыс второй группы подвергали бегу на тредбане вниз по беговой дорожке со скоростью 20 м/мин в течение 40 минут. Дорожка располагалась под углом 16° к поверхности пола (n=7, вес тела 275,5±39,6 г; масса m.soleus 98,7±4,6 мг; гр. R). Третьей группе за 2 дня до физической нагрузки давали L-аргинин с водой в дозе 500 мг на кг веса тела в день, а затем подвергали бегу на тредбане по предыдущей программе (n=7, вес тела 272±6, масса m.soleus 114±13; гр. RA). Четвертой группе животных за 2 дня до бега вводили L-NAME с водой в дозе 90 мг на кг веса тела в день, а затем подвергали бегу на тредбане (n=7, вес тела 254±15; масса m.soleus 100±5, гр. RN). Через 24 часа после бега крыс забивали сверхдозой нембутала (75 мг/кг веса), m.soleus немедленно замораживали в жидком азоте. Пробы затем хранили при -85°С. Определяли содержание десмина методом иммуноблота, дистрофин - на поперечных срезах мышцы, содержание nNOS (нейрогенной NO-синтазы) методом ПЦР в реальном времени. Для выяснения относительных изменений содержания оксида азота в мышцах брали 6 контрольных крыс (С) и 6 крыс с введением L-аргинина по предыдущей схеме (СА). Относительное содержание оксида азота в мышцах крыс обеих групп определялось по содержанию в них парамагнитных центров, количество которых пропорционально концентрации оксида азота в тканях.
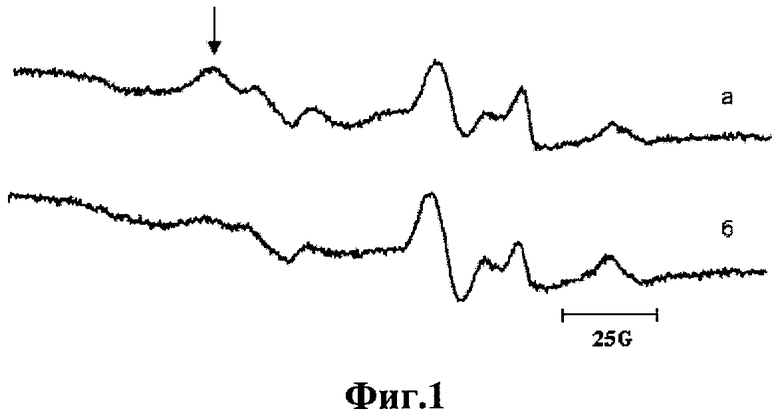
Электронно-парамагнитный резонанс (ЭПР). Крысе вводилась спиновая ловушка - диэтилдитиокарбамат (ДЭТК) из расчета 500 мг/кг веса, которая равномерно распределялась в органах и тканях. ДЭТК с высокой константой связывалась с оксидом азота, образуя парамагнитные комплексы, которые находятся в равновесии со стационарной концентрацией оксида азота в ткани. Сразу после введения ДЭТК животному внутримышечно вводили водный раствор смеси 29 мМ FeSO4 и 116 мМ цитрата натрия (2,0 мл/кг веса). Через 30 мин животных декапитировали, m.soleus замораживали в жидком азоте и регистрировали сигнал ЭПР при температуре жидкого азота на ЭПР-спектрометре фирмы "Bruker EMX-8". Сигнал ЭПР представляет собой суперпозицию сигнала комплекса NO- Fe2+ (ДЭТК)2, а также других парамагнитных центров, которые обнаруживаются в тканях. Однако в регистрируемом сигнале видны компоненты, которые по форме и положению сигнала совпадают с сигналом ЭПР мононитрозильных комплексов железа с дитиокарбаматом. Это позволяет оценивать относительную концентрацию оксида азота по первой компоненте триплетной сверхтонкой структуры этого комплекса [19, 20]. (На фиг.1 указано стрелкой.)
Выявление дистрофина. Поперечные криостатные срезы мышечной ткани толщиной 10 мкм инкубировали с первичными моноклональными антителами против дистрофина (1:20, Novocastra, NCL-DYSI) и вторичными поликлональными антителами, конъюгированными с флуоресцентной меткой (GAM, 1:1000, Alexa546, Molecular probes). Все инкубации проводили в течение часа при комнатной температуре. Подсчитывали процентное содержание волокон с поврежденным дистрофиновым слоем и вычисляли отношение длины разрыва дистрофинового слоя ко всему периметру волокна. Анализировали не менее 100 волокон с каждого среза.
Выявление десмина. С каждой пробы m.soleus были сделаны срезы толщиной 20 мкм, помещены в лизирующий буфер (Tris-HCl, pH 6,8, 62,5 mM, SDS 2,0%, Glicerin 10%, b-mercaptoethanol 5,0%, Bromphenol blue 0,004%) из расчета 25 мкл буфера на 5 мг ткани, гомогенизированы, нагреты 10-15 мин при 95°С, центрифугированы 10 мин при 13000 об/мин. Концентрацию белка в надосадочной жидкости определяли методом Шафнера и Вайсмана. Электрофорез проводили в 12% разделяющем акриламидном геле (1% SDS; 1,5 М Tris-HCl, pH 8,8, 0,8% метилбисакриламид, APS, TEMED 0,1%) и 6% концентрирующем акриламидном геле (0,1% SDS; 0,5 М Tris-HCl, pH 6,8, 0,4% метилбисакриламид, APS, TEMED 0,1%). Использовали анодный буфер (0,25 М Трис-HCl, pH 8,6) и катодный (1,92 М трис-глицин, рН 8,6; 0,1% SDS). Образцы загружались из расчета 20 мкг общего белка. Электрофорез проводили при 15 мА на гель в мини-системе Bio-Rad при комнатной температуре. Вестернблоттинг белков проводили при 300 мА 3 часа при температуре +4°С в системе mini Trans-Blot (Bio-Rad). Буфер для электропереноса Towbin pH 8,3 (Tris-Base 25 mM, glycine 192 mM, метанол 20%). Мембрану блокировали в растворе 4% сухого молока в TBST (трис-ОН 4 mM, NaCl 100 mM, 0,1% Tween 20), pH 7,4, 30 мин при 37°С. Для выявления десмина были использованы первичные моноклональные антитела (Novocastra, NCL-L-DES-DERII, 1:500) и вторичные антитела (GAM, Имтек, 1:100), конъюгированные с пероксидазой хрена. Инкубации проводили 1 час при комнатной температуре. В качестве субстрата и хромогена брали перекись водорода и диаминобензидин. Сигнал анализировали сканером GS-800 (Bio-Rad) и программой ImageJ. Вестернблоттинг был повторен 2-3 раза. Оптическую плотность полосы контрольной группы на анализируемой мембране принимали за 100%, а полосы групп других групп сравнивали контрольными крысами, расположенными на одной и той же мембране.
Проведение РТ-ПЦР. Тотальная РНК была экстрагирована из 10 мг замороженной m.soleus при помощи RNeasy Micro Kit (QIAGEN, Germany). Все образцы РНК были обработаны протеиназой К и ДНКазой I. Концентрация РНК была определена при 260 нм. Затем водный раствор выделенной РНК был заморожен на -84°С для последующего проведения обратной транскрипции. Для обратной транскрипции использовали водный раствор 1 мкг РНК, олиго(dT)15, случайные гексануклеотиды d(N)6, обратную транкскриптазу MMLV. Обратную транскрипцию проводили при 37°С в течение 60 мин согласно стандартному протоколу. Полученные образцы кДНК были помещены на хранение при -84°С для проведения ПНР в реальном времени. При помощи ПЦР-РВ была определена экспрессия мРНК nNOS. β-актин использовали в качестве внутреннего контроля. Для проведения ПЦР-РВ применяли следующие праймеры nNOS: прямой AGTCCCCTGCTTCGTGAGAG, обратный CACCCGAAGACCAGAACCAT. Для анализа полученных данных мы использовали относительное количественное определение целевого гена nNOS, нормализованное к референсному β-актину, метод 2-ΔΔCt (Ливака метод). За нулевой уровень был взят ΔCt в контроле.
Результаты исследования
Содержание NO в ткани. На Фиг.1а, б представлены типичные спектры ЭПР мышц после введения Fe-ДЭТК. В среднем, в относительных единицах, содержание оксида азота увеличивалось от 1,0±0,1 в группе контроля до 1,64±0,28 в группе после введения в рацион L-аргинина (p<0.05).
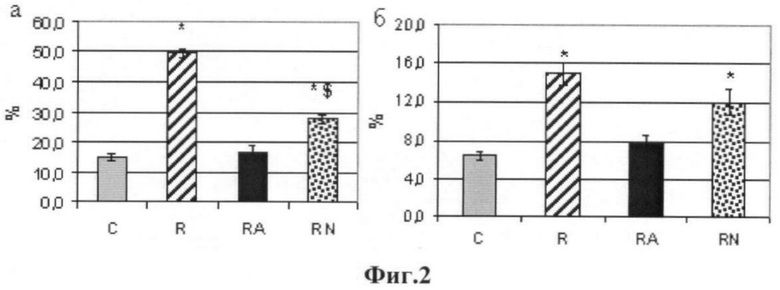
Процент мышечных волокон с разрывами в дистрофиновом слое. В группе «Бег» процент мышечных волокон, имеющих разрывы в дистрофиновом слое, был на 327%, а в группе RN - на 187% выше, чем в группе контроля (p<0.05). Введение L-аргинина полностью предотвратило разрушение дистрофинового слоя при беге (фиг.1). При подсчете длины разрывов в дистрофиновом слое мы получили аналогичные результаты (фиг.2, 3).
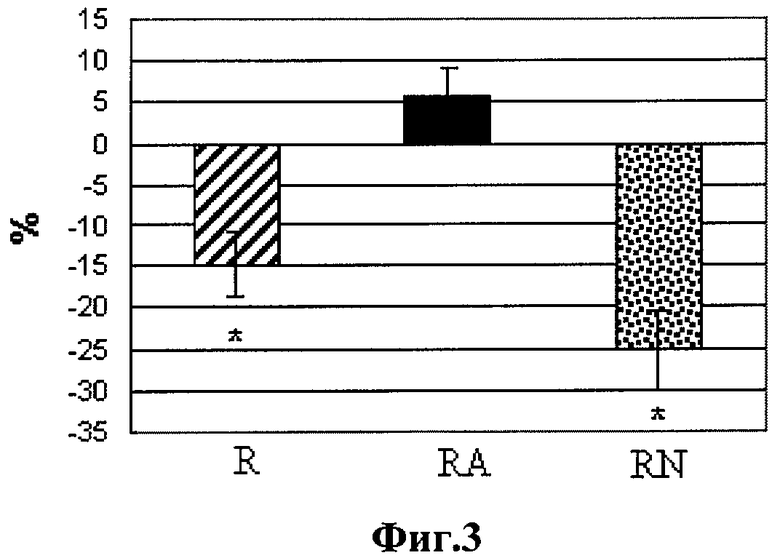
Содержание десмина было снижено в R и RN группах на 15 и 25% соответственно относительно уровня контроля. Введение донора NO L-аргинина при беге предотвратило эти изменения (фиг.4).
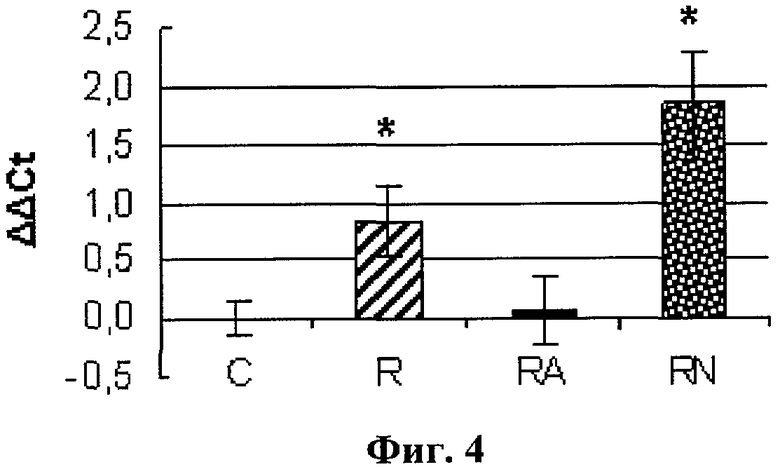
Концентрация мРНК nNOS после бега в группе R была существенно выше (p<0,05), чем в группе контроля. В группе RN концентрация мРНК nNOS превышала уровень группы R в два раза и была достоверно выше как группы С, так и RA (p<0.05).
Т.е. проведенные исследования показали, что увеличение содержания оксида азота (NO) в скелетных мышцах предупреждает в них деструкцию цитоскелетных белков при эксцентрических нагрузках.
Краткое описание чертежей
На фиг.1 изображен типичный спектр ЭПР m.soleus крысы после введения Fe-ДЭТК:
а - в рацион крысы вводился L-аргинин в питьевую воду;
б - контрольное животное.
Спектр ЭПР регистрировался при температуре жидкого азота, при частоте 9,61 GHz, мощности 2 mW и амплитуде модуляции 5 G. Компонента спектра, по которой оценивалась относительная концентрация оксида азота, указана стрелкой.
На фиг.2 представлены данные:
- процент волокон с разрывами в дистрофиновом слое;
- отношение (в процентах) общей длины разрывов дистрофинового слоя к периметру всего волокна, в %;
* - достоверные отличия от Контроля, p<0,05;
$ - отличия от группы R, p<0,05;
С - контроль, R - бег, RA-бег + L-аргинин, RN-бег + L-NAME).
На фиг.3 представлены данные содержания десмина через 24 часа после тренировки.
Нулевой уровень-содержание десмина в контроле:
* - отличия от контроля существенны при p<0,05;
R-бег, КА-бег + L-аргинин, RN-бег + L-NAME.
На фиг.4 представлены данные - содержание мРНК nNOS в m.soleus
* - отличия от контроля, p<0,05;
С-контроль, R-бег, КА-бег + L-аргинин, RN-бег + L-NAME).
Список литературы
1. Fridén J, Lieber R. «Eccentric exercise-induced injuries to contractile and cytoskeletal muscle fibre components». Acta Physiol Scand. 2001 Mar; 171(3): 321-6.
2. Komulainen J, Koskinen SO, Kalliokoski R, Takala ТЕ, Vihko V. «Gender differences in skeletal muscle fibre damage after eccentrically biased downhill running in rats». Acta Physiol Scand. 1999 Jan; 165(1): 57-63.
3. Lieber RL, Schmitz MC, Mishra DK, Fridén J. «Contractile and cellular remodeling in rabbit skeletal muscle after cyclic eccentric contractions». J Appl Physiol. 1994 Oct; 77(4): 1926-34.
4. Lieber RL, Thomell LE, Fridén J. «Muscle cytoskeletal disruption occurs within the first 15 min of cyclic eccentric contraction». J Appl Physiol. 1996 Jan; 80(1): 278-84.
5. Germinario E, Esposito A, Megighian A, Midrio M, Biral D, Betto R, Danieli-Betto D. «Early changes of type 2B fibers after denervation of rat EDL skeletal muscle». J Appl Physiol. 2002 May; 92(5): 2045-52.
6. Lovering RM, De Deyne PG. «Contractile function, sarcolemma integrity, and the loss of dystrophin after skeletal muscle eccentric contraction-induced injury». Am J Physiol Cell Physiol. 2004 Feb; 286(2): C230-8.
7. Rey MA, Davies PL. «The protease core of the muscle-specific calpain, p94, undergoes Ca2+-dependent intramolecular autolysis». FEBS Lett. 2002 Dec 18; 532(3): 401-6.
8. Duguez S, Bartoli M, Richard I. «Calpain 3: a key regulator of the sarcomere?». FEBS J. 2006 Aug; 273(15):3427-36.
9. Murphy RM, Goodman CA, McKenna MJ, Bennie J, Leikis M, Lamb GD. «Calpain-3 is autolyzed and hence activated in human skeletal muscle 24 h following a single bout of eccentric exercise». J Appl Physiol. 2007 Sep; 103(3): 926-31.
10. Ingalls CP, Warren GL, Armstrong RB. «Intracellular Ca2+ transients in mouse soleus muscle after hindlimb unloading and reloading». J Appl Physiol. 1999 Jul; 87(1): 386-90.
11. Lynch GS, Fary CJ, Williams DA. «Quantitative measurement of resting skeletal muscle [Ca2+]i following acute and long-term downhill running exercise in mice». Cell Calcium. 1997 Nov; 22(5): 373-83.
12. Немировская Т.Л., Китина Ю.Н., Железнякова А.В., Вихляндцев И.М. «Цитоскелетные белки и белки теплового шока 27 при эксцентрической нагрузке: эффект блокады 1-кальциевых каналов».Физиологический журнал им. Сеченова, 2008, №3, т.94, стр.293-300.
13. Murphy RM, Snow RJ, Lamb GD. «mu-Calpain and calpain-3 are not autolyzed with exhaustive exercise in humans». Am J Physiol Cell Physiol. 2006 Jan; 290(1): C116-22.
14. Stupka N, Tarnopolsky MA, Yardley NJ, Phillips SM. «Cellular adaptation to repeated eccentric exercise-induced muscle damage». J Appl Physiol. 2001 Oct; 91(4): 1669-78.
15. Murphy RM, Verburg E, Lamb GD. «Ca2+ activation of diffusible and bound pools of mu-calpain in rat skeletal muscle». J Physiol. 2006 Oct 15; 576 (Pt2): 595-612.
16. Beaton Louise J., Tarnopolsky Mark A. and Phillips Stuart M.. «Contraction-induced muscle damage in humans following calcium channel blocker administration». Journal of Physiology (2002), 544.3, pp.849-859.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНГИБИТОР ПРОТЕОЛИЗА ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ФИЗИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2389486C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2010 |
|
RU2444354C2 |
| СРЕДСТВО ДЛЯ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517576C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517259C1 |
| ПРИМЕНЕНИЕ VX 745 ДЛЯ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2019 |
|
RU2714923C1 |
| Способ предотвращения атрофии скелетных мышц при их функциональной разгрузке | 2022 |
|
RU2797216C1 |
| СПОСОБ УСКОРЕНИЯ ВОССТАНОВЛЕНИЯ СКЕЛЕТНЫХ МЫШЦ ОТ АТРОФИИ ПОСЛЕ ДЛИТЕЛЬНОЙ АЛКОГОЛЬНОЙ ИНТОКСИКАЦИИ | 2014 |
|
RU2578463C1 |
| Способ поддержания силы и жесткости постуральных мышц млекопитающих при их функциональной разгрузке | 2022 |
|
RU2800317C1 |
| СПОСОБ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2012 |
|
RU2481105C1 |
| СПОСОБ ВОССТАНОВЛЕНИЯ МАССЫ ПОСТУРАЛЬНЫХ МЫШЦ ГОЛЕНИ ПОСЛЕ ВОЗДЕЙСТВИЯ ГИПОКИНЕЗИИ И/ИЛИ ГИПОГРАВИТАЦИИ | 2008 |
|
RU2368398C1 |
Изобретение относится к медицине, а именно к спортивной медицине авиакосмической медицине и может быть использовано при подготовке спортсменов. Для этого в организм вводят соединения, обеспечивающие увеличение оксида азота (NO) в скелетных мышцах. В качестве такого соединения вводят L-аргинин. Способ позволяет предупредить деструкцию цитоскелетных белков при эксцентрических нагрузках при отсутствии негативного воздействия на сердечно-сосудистую систему. 1 з.п. ф-лы, 4 ил.
1. Способ профилактики разрушения цитоскелетных белков при эксцентрической нагрузке, включающий предварительное, до эксцентрической нагрузки, введение в организм химических соединений, отличающийся тем, что в качестве химического соединения в организм вводят соединения, обеспечивающие увеличение оксида азота (NO) в скелетных мышцах.
2. Способ по п.1, отличающийся тем, что в качестве соединения, обеспечивающего увеличение оксида азота в скелетных мышцах, вводят донор оксида азота L-аргинин.
| BEATON L.J | |||
| et al | |||
| Contraction-induced muscle damage in humans following calcium channel blocker administration | |||
| J | |||
| Physiol | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ПРОДУКТ "СПОРТАМИН" ДЛЯ ПИТАНИЯ СПОРТСМЕНОВ | 2006 |
|
RU2306720C1 |
| US 6967102 B1, 22.11.2005, реферат | |||
| Устройство для записи простоев текстильных и тому подобных машин | 1937 |
|
SU53191A1 |
| WO 9602241 A1, 01.02.1996, реферат | |||
| НЕМИРОВСКАЯ Т.Л | |||
| и др | |||
| Цитоскелетные белки и | |||

