Изобретение относится к физиологическим наукам и может быть использовано для предотвращения роста утомляемости и атрофии мышечных волокон «медленного» типа при функциональной разгрузке постуральных мышц.
Отсутствие опорной стимуляции при длительной гипокинезии, космическом полете, а также при моделировании гравитационной разгрузки на Земле приводит к функциональной разгрузке скелетных мышц, то есть к отсутствию механической нагрузки на мышцы. В данных условиях происходит атрофия постуральных (позно-тонических) и, в меньшей степени, локомоторных мышц и трансформация медленных мышечных волокон в быстрые, что приводит к снижению мышечной силы и повышению утомляемости.
Постуральные мышцы служат для поддержания позы, поэтому в них преобладают медленные мышечные волокна, которым свойственна низкая утомляемость, синтез АТФ с помощью окислительного фосфорилирования, относительно низкая максимальная сила и скорость сокращения, но повышенная устойчивость к утомлению. Однако основное отличие медленных мышечных волокон от быстрых - экспрессия медленной изоформы тяжелой цепи миозина ТЦМ I(β), в то время как в быстрых мышечных волокнах экспрессируются быстрые изоформы ТЦМ II (Schiaffino, Reggiani, 2011).
Во время функциональной разгрузки постуральной мышцы из-за отсутствия сократительной активности происходит изменение миозинового фенотипа - снижается экспрессия медленной изоформы миозина ТЦМ I и повышается экспрессия быстрых ТЦМ II изоформ. Уже после суток антиортостатического вывешивания (модулируемой функциональной разгрузки) происходит снижение экспрессии MyHCI (Vilchinskaya et al., 2017). Помимо этого, происходит количественное снижение митохондриальных белков и ДНК (Liu et al., 2012). Содержание ключевого регулятора биогенеза митохондрий PGC-1α, регулирующего транскрипцию и репликацию митохондриальной ДНК, в камбаловидной мышце грызунов снижается уже на третьи сутки функциональной разгрузки (Tyganov et al., 2019).
Кроме того, показано, что во время функциональной разгрузки происходит атрофия скелетной мышцы, мышца теряет свою массу за счет нарушения баланса между синтезом и деградацией белка. Данный процесс приводит к снижению площади поперечного сечения медленных мышечных волокон m. soleus (Allen et al., 1996). Важную роль в атрофии скелетных мышц, вызванной функциональной разгрузкой, играет Е3 компонент убиквитин-протеасомной системы - убиквитин лигазы MuRF1 и MAFbx/atrogin 1. Экспрессия убиквитин лигаз MuRF1 и MAFbx значительно повышается во всех типах мышц в модели антиортостатического вывешивания на 3, 7, и 14 сутки, в постуральной камбаловидной мышце отмечается усиление их протеасомной активности (Baehr et al., 2017). Также во время функциональной разгрузки снижается интенсивность синтеза белка. На 3, 7 и 14 сутки антиортостатического вывешивания в камбаловидной мышце крысы уменьшается емкость трансляции (количество рибосом на единицу ткани), что ведет к снижению интенсивности синтеза белка (Rozhkov et al., 2021).
В совокупности, вышеописанные изменения во время функциональной разгрузки влияют на функцию и механические характеристики постуральных мышц: уменьшается сила сокращения мышц и нарастает мышечная утомляемость (Sharlo et al., 2022).
Митокин MOTS-c - это короткий биоактивный пептид митохондриального происхождения, который кодируется последовательностью, заключенной в гене 12S рибосомальной РНК и обладает цитопротекторным действием. MOTS-c действует как транскрипционный фактор, он связывается с хроматином и регулирует широкий спектр генов, в том числе гены, отвечающие за антиоксидантный ответ (ARE) (Kim et al., 2018), гены субъединиц mTOR (Jacinto et al., 2006), гены ферментов цикла фолиевой кислоты (Lee et al., 2015). Ингибируя цикл фолиевой кислоты, MOTS-c тормозит также связанный с ним биосинтез пуринов de-novo, что активирует АМПК (АМФ-зависимую протеинкиназу) (Lee et al., 2015). АМПК частично опосредует метаболические эффекты MOTS-c - увеличивает клиренс глюкозы и инсулин-зависимое поглощение глюкозы скелетными мышцами, предотвращает ожирение и инсулинорезистентность у животных на высокожировой диете (high-fat diet (HFD)) (Lee et al., 2015).
Во время тренировок на выносливость, то есть при аэробной нагрузке, у людей повышается экспрессия MOTS-c в скелетных мышцах, а также уровень данного пептида в плазме (Reynolds et al., 2021).
В результате проведенных исследований было обнаружено, что введение митокина MOTS-c на фоне 7 суток функциональной разгрузки (в модели антиортостатического вывешивания по Ильину-Новикову в модификации Morey-Holton) приводит к полному предотвращению повышенной утомляемости и частичному предотвращению снижения площади поперечного сечения «медленных» волокон m. soleus; предотвращению трансформации медленных мышечных волокон в быстрые; повышению уровня фосфорилирования и активности АМПК (АМФ-зависимой протеинкиназы) на 45%; к увеличению экспрессии мРНК основного регулятора биогенеза митохондрий PGC1α на 10%, по сравнению с группой вывешивания, росту уровня ингибирующего фосфорилирования (Ser9) GSK3β на 97% по сравнению с группой вывешивания и на 48% по сравнению с группой сидячего контроля, а также предотвращению роста экспрессии мРНК убиквитин лигаз MuRf-1 и Atrogin1.
Таким образом, заявляемое изобретение основано на применении короткого биоактивного пептида MOTS-c (16 аминокислотных остатков) на фоне 7-суточной функциональной разгрузки для предотвращения роста утомляемости, предотвращения атрофии медленных мышечных волокон и их трансформации в быстрые при функциональной разгрузке.
Наиболее близким способом (прототипом) является работа Kumagai Н, (Kumagai et al., 2024) В данной работе показано предотвращение атрофии скелетных мышц мышей при 8-дневной гипсовой иммобилизации мышцы, предотвращение снижения фосфорилированной формы киназы Akt, а также ряда параметров распада белка, однако авторы не исследовали влияние MOTS-С на экспрессию миозиновых генов и мышечную утомляемость, а также на содержание рибосомальной РНК. Также модель иммобилизации мышцы отличается от модели вывешивания по эффектам на скелетную мышцу. Также близким способом предотвращения снижения устойчивости к утомлению мышц крыс в условиях функциональной разгрузки является применение активатора SERCA CDN1163 (Sharlo et al., 2023b), однако данный активатор оказывает схожее действие на мышечную утомляемость за счет другого механизма - в частности, CDN1163 не оказывает влияния на экспрессию Е3 убиквитинлигаз и рибосомальных РНК 18S и 28S, в отличие от MOTSC, и влияет на содержание кальция в миоплазме мышечных волокон, чего в случае MOTSC не показано.
Техническим результатом заявленного изобретения является предотвращение роста утомляемости, атрофии и трансформации медленных волокон в быстрые в m. soleus во время функциональной разгрузки при введении MOTS-c.
Этот технический результат достигается тем, что препарат вводили ежедневно на протяжении 7 суток эксперимента, внутрибрюшинно, в дозировке 40 мг/кг в 0,9% растворе NaCl.
Животным из контрольной группы и группы «чистой» 7-суточной функциональной разгрузки вводили плацебо (0,9% раствор NaCl).
Гипотеза была проверена с помощью внутрибрюшинного введения MOTS-c (40 мг/кг) на фоне 7-суточного вывешивания крыс.
Возможные механизмы действия метода.
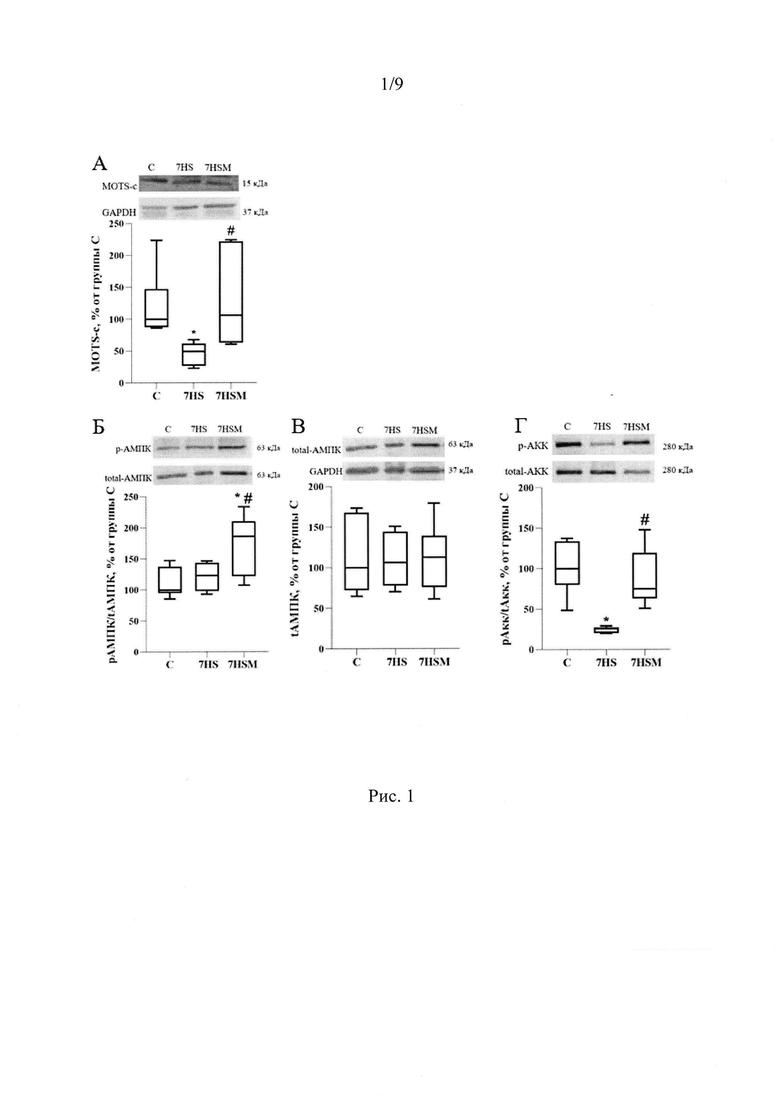
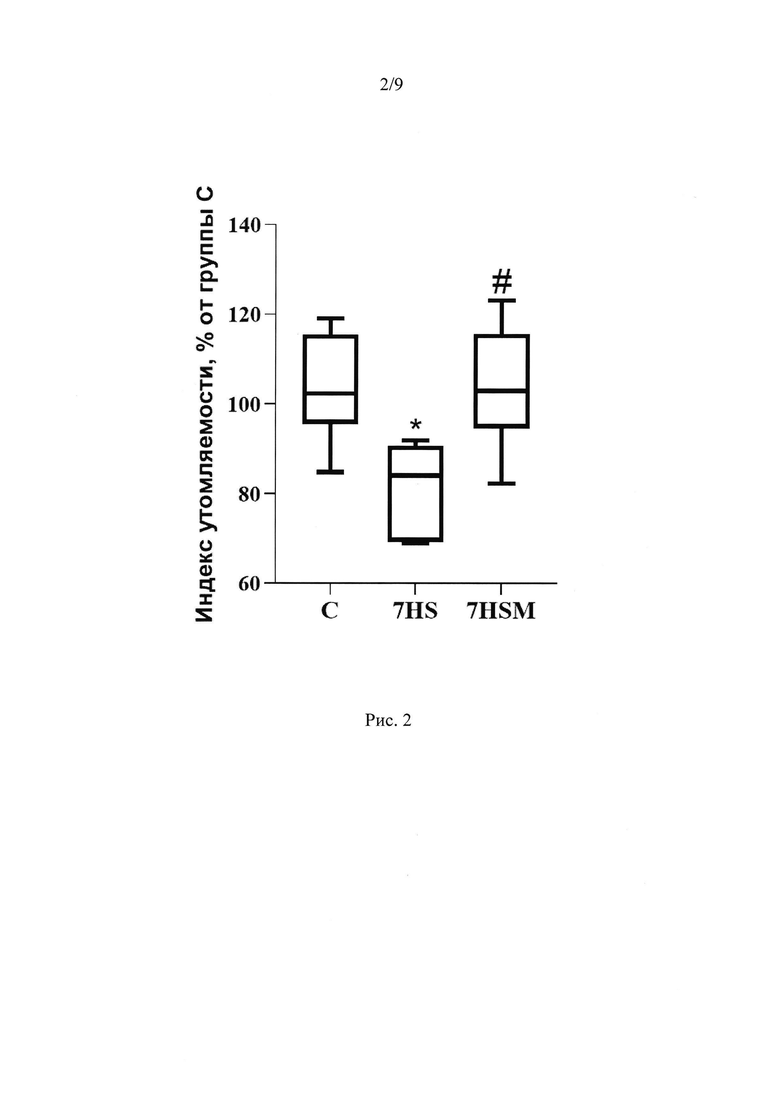
Мы выяснили, что в m. soleus крысы на 7 сутки функциональной разгрузки снижается содержание пептида MOTS-c на 60% по сравнению с виварным контролем. Введение экзогенного MOTS-C предотвращает данное снижение (Рис 1А). Также на 7 сутки функциональной разгрузки растет падает индекс утомляемости камбаловидной мышцы (что означает рост утомляемости мышцы), а введение MOTS-C предотвращает данное снижение (Рис. 2).
Известно, что MOTS-c действует как транскрипционный фактор, влияя на экспрессию ряда ядерных генов, в том числе на гены ферментов, вовлеченных в фолат-метиониновый цикл, приводя к активации аденозин монофосфат-зависимую протеинкиназы (АМПК). MOTS-c ингибирует цикл фолиевой кислоты на уровне 5-метил-тетрагидрофолата, что также тормозит связанный с ним биосинтез пуринов de novo, приводя к накоплению 5-аминоимидазол-4-карбоксамид рибонуклеотида (AICAR). AICAR активирует АМПК, которая частично опосредует метаболические эффекты MOTS-c, такие как активация гликолиза, пентозофосфатного пути, окисления жирных кислот (Lee et al., 2015). АМПК является ключевой чувствительной к уровням энергии киназой и основным регулятором метаболизма, который активируется метаболическим стрессом (Hardie et al., 2016). Помимо этих эффектов, АМПК способствует экспрессии «медленной» устойчивой к утомлению изоформы тяжелых цепей миозина, а также играет важную роль в обеспечении устойчивости мышц к утомлению (Sharlo et al., 2023, Holmes et al., 2005) Известно, что на ранних стадиях функциональной разгрузки происходит накопление макроэргических фосфатов, повышается соотношение АТФ/АДФ, что приводит к снижению фосфорилирования АМПК и снижению ее активности (Vilchinskaya et al., 2015).
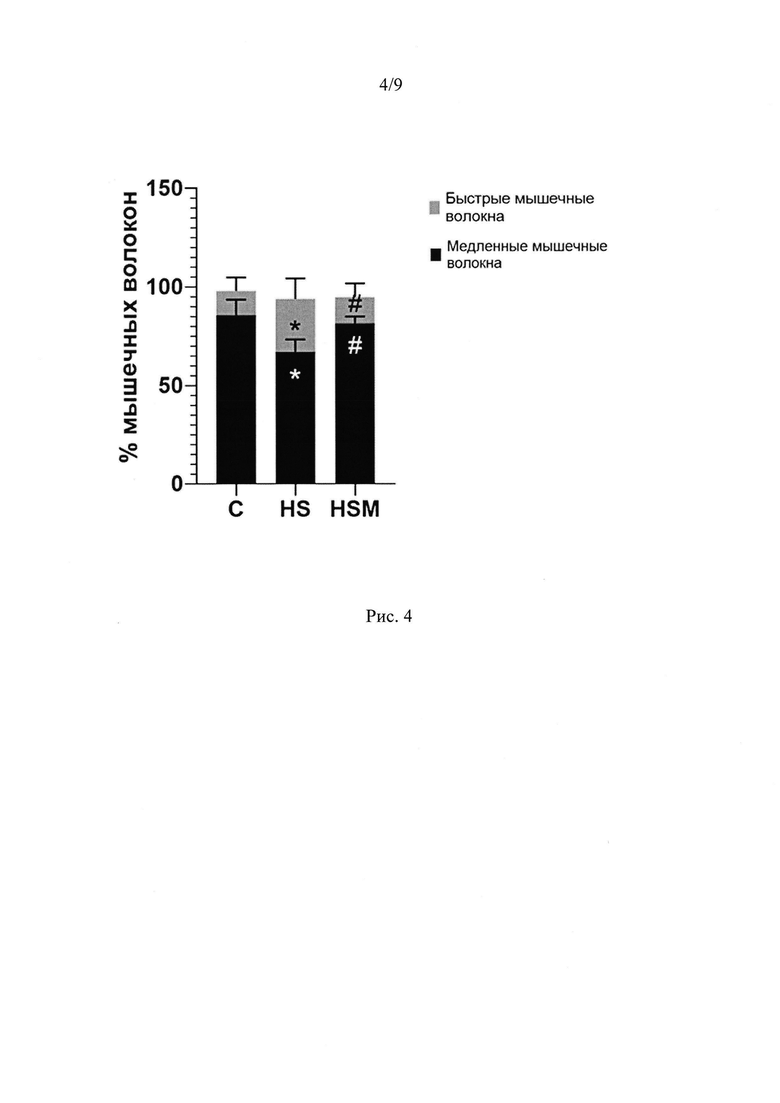
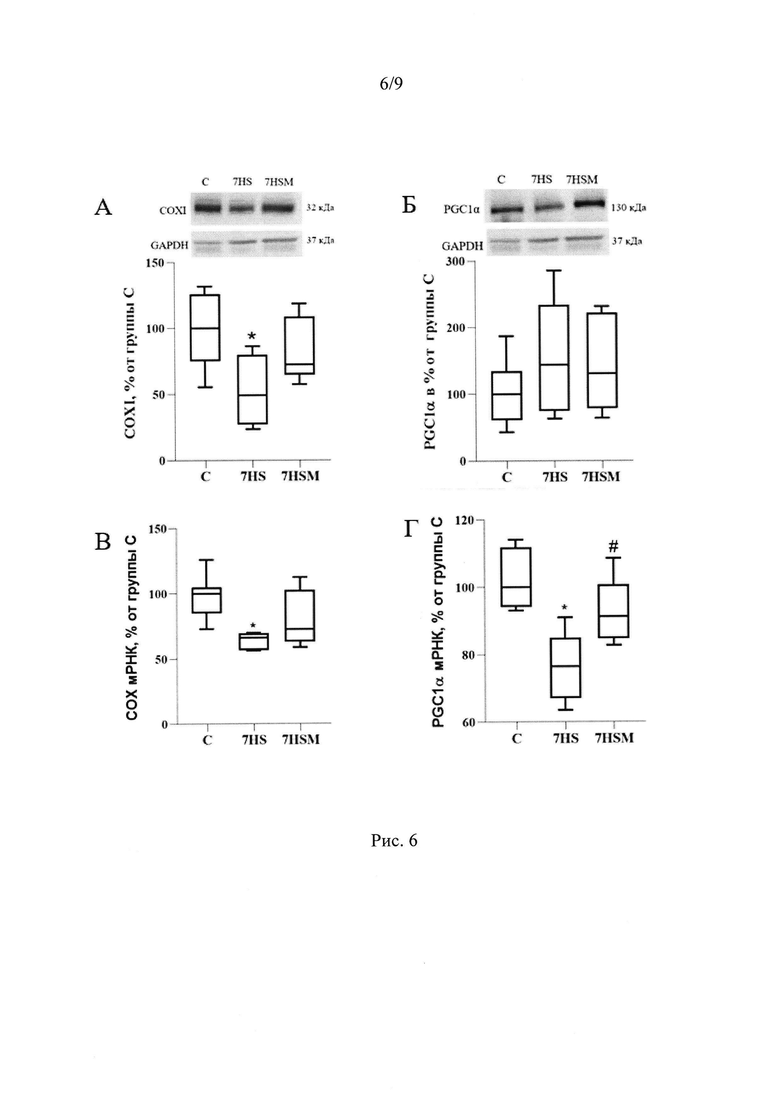
Мы показали, что при введении MOTS-c на фоне функциональной разгрузки достоверно повышается уровень фосфорилирования АМПК на 24% по сравнению с группой вывешивания и на 28% по сравнению с группой контроля, а также растет активность данной киназы (фосфорилирование мишени АМПК Ацетил-Коа карбоксилазы повышается на 55% по сравнению с группой вывешивания) (Рис. 1Б-Г). Известно, что АМПК может влиять на миозиновый фенотип мышечных волокон (Vilchinskaya et al., 2018, Sharlo et al., 2023). Мы показали, что у животных, получавших MOTS-c на фоне вывешивания предотвращалась трансформация медленных мышечных волокон в быстрые (Рис. 4). Инъекции MOTS-c во время функциональной разгрузки не только предотвращали снижение экспрессии мРНК медленной изоформы тяжелой цепи миозина ТЦМ-I, но также угнетали рост экспрессии мРНК быстрых изоформ миозина ТЦМ-IIB и ТЦМ-IId/x (Рис. 5А-Г). Снижение утомляемости мышцы во время функциональной разгрузки после введения MOTS-c можно объяснить влиянием митокина на митохондриальные параметры. Так, в группе, получавшей MOTS-c на фоне вывешивания, частично предотвращается снижение экспрессии мРНК активатора биогенеза митохондрий PGC1α в камбаловидной мышце крысы. Экспрессия PGC1α в группе 7HS достоверно ниже, чем в группе С, в то время как группа 7HSM не отличается от группы С.PGC1α (Peroxisome proliferator-activated receptor gamma coactivator 1-alpha) это ключевой регулятор биогенеза митохондрий, регулирующий транскрипцию и репликацию митохондриальной ДНК, снижение его содержания описано уже на третьи сутки функциональной разгрузки (Tyganov et al., 2019). Также инъекции MOTS-c частично предотвращают снижение экспрессии мРНК и содержания белка субъединицы IIV комплекса дыхательной цепи (COXI), мишени PGC1α (Рис. 6А-Г).
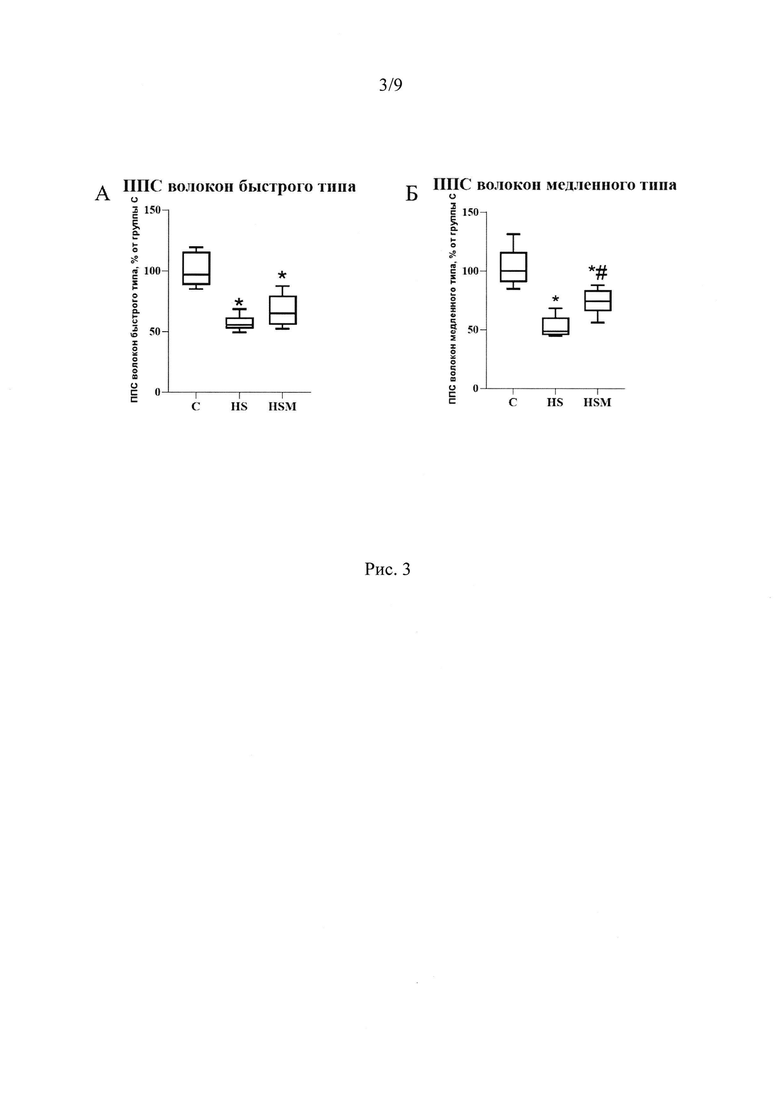
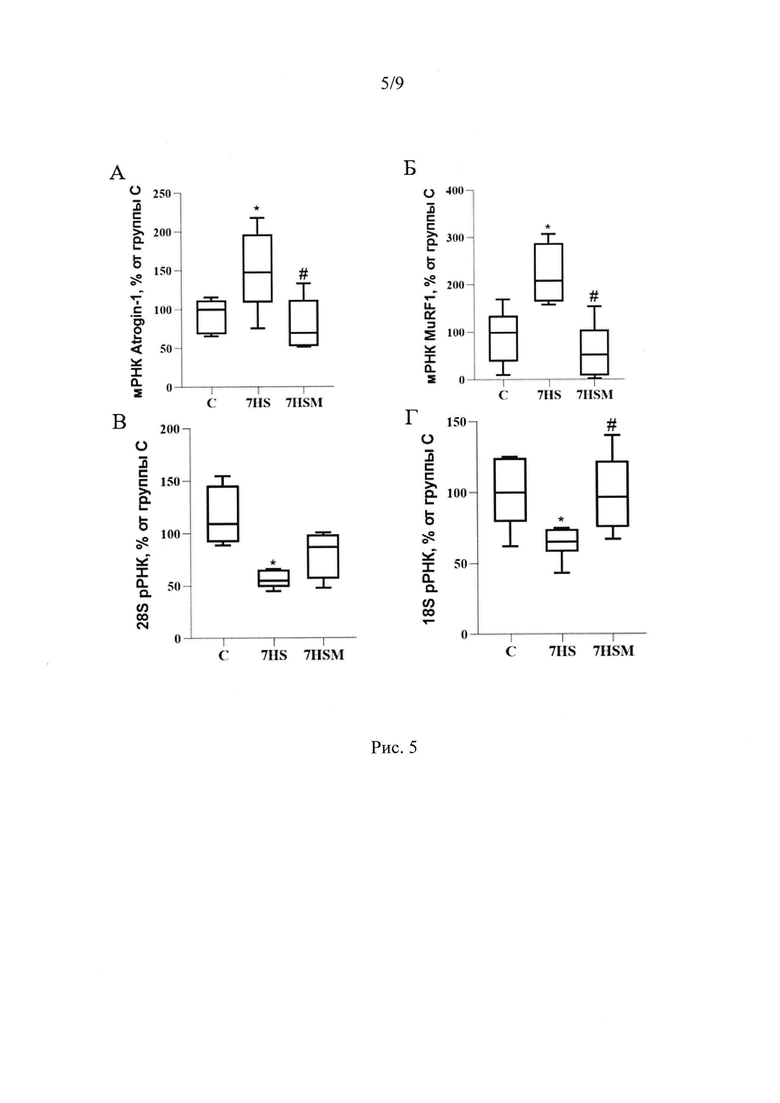
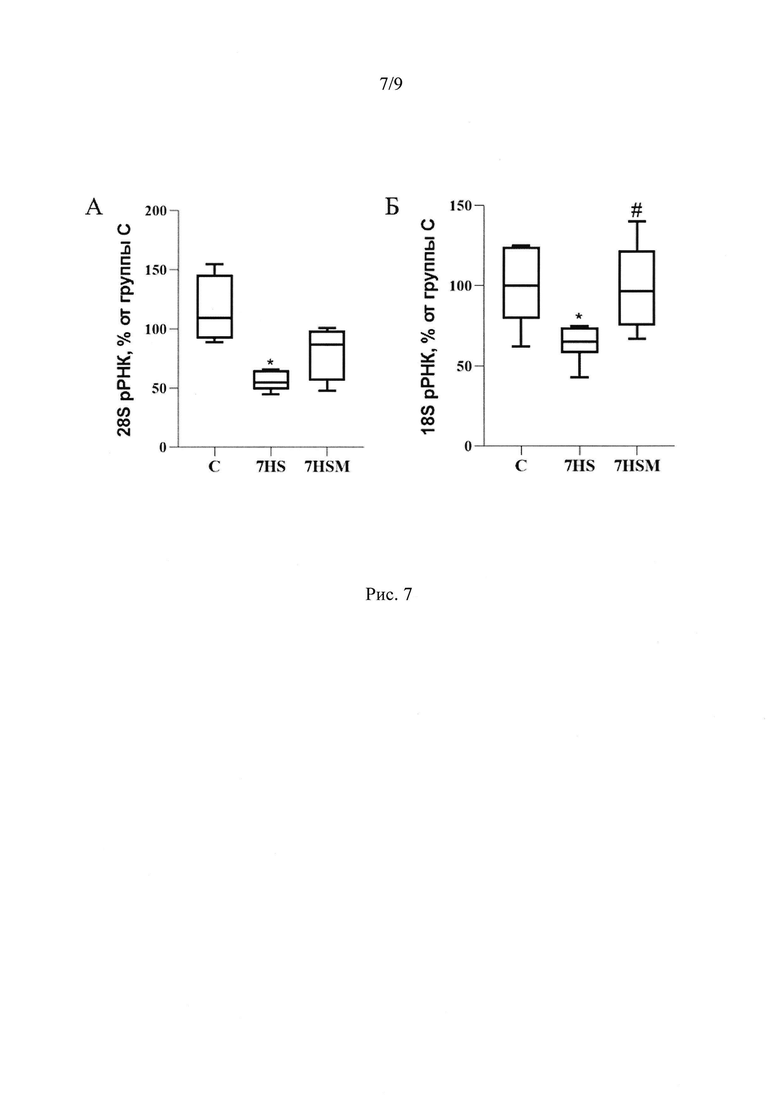
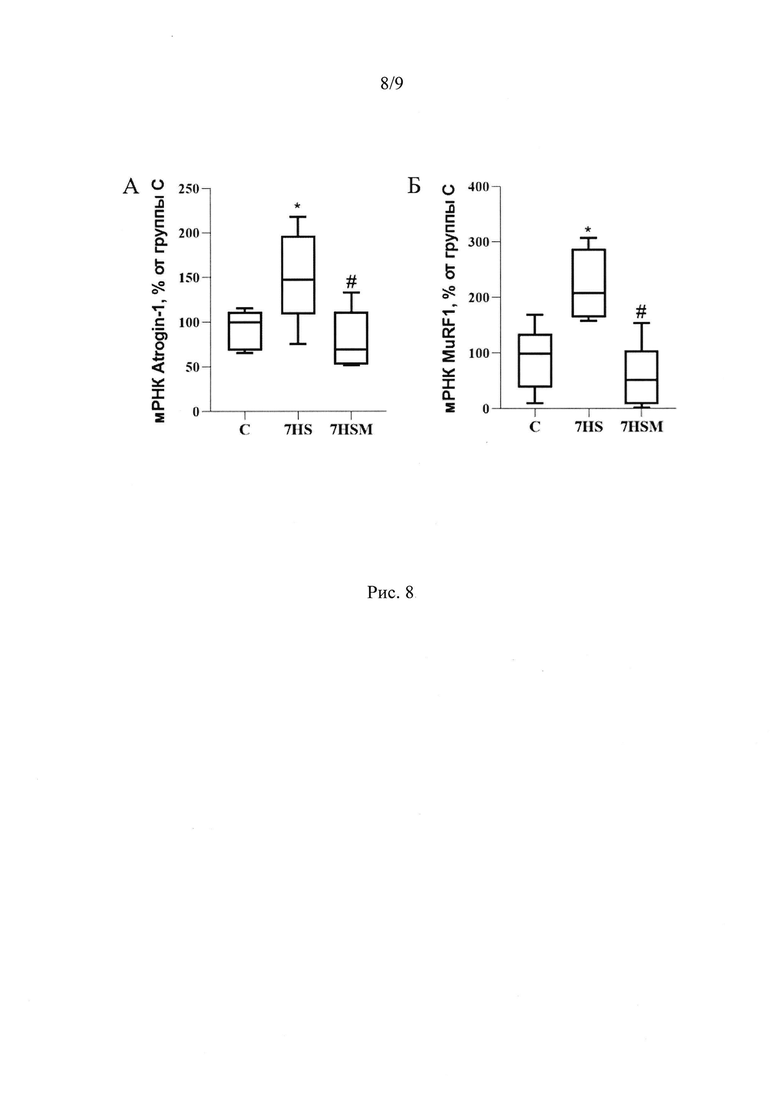
Помимо этих эффектов, введение MOTS-c предотвратило атрофию медленных мышечных волокон в условиях 7-суточной функциональной разгрузки (Рис 3А-Б). Атрофия - это нарушение баланса между синтезом и деградацией белка. Интенсивность синтеза белка зависит, в частности, от количества рибосом в ткани. Мы показали, что количество 28S и 18S рРНК в группе вывешивания достоверно снижается на 59% и 38% соответственно по сравнению с группой виварного контроля. MOTS-c полностью предотвращает снижение содержания 18S рРНК и частично предотвращает снижение количества 28S рРНК (Рис 7А, Б). Одним из механизмов предотвращения атрофии медленных мышечных волокон m. soleus в группе, получавшей MOTS-c на фоне 7-суточной функциональной разгрузки может быть влияние митокина на экспрессию мРНК убиквитин-лигаз MuRF-1 и Atrogin-1. Мы показали, что введение митокина MOTS-c на фоне 7 суток функциональной разгрузки предотвращает рост экспрессии мРНК маркеров атрофии MuRF1 и Atrogin-1. В группе 7HSM содержание мРНК MuRF1 на 160%, а мРНК Atrogin-1 на 70% ниже, чем в группе 7HS, и находится на уровне группы С (Рис 8).
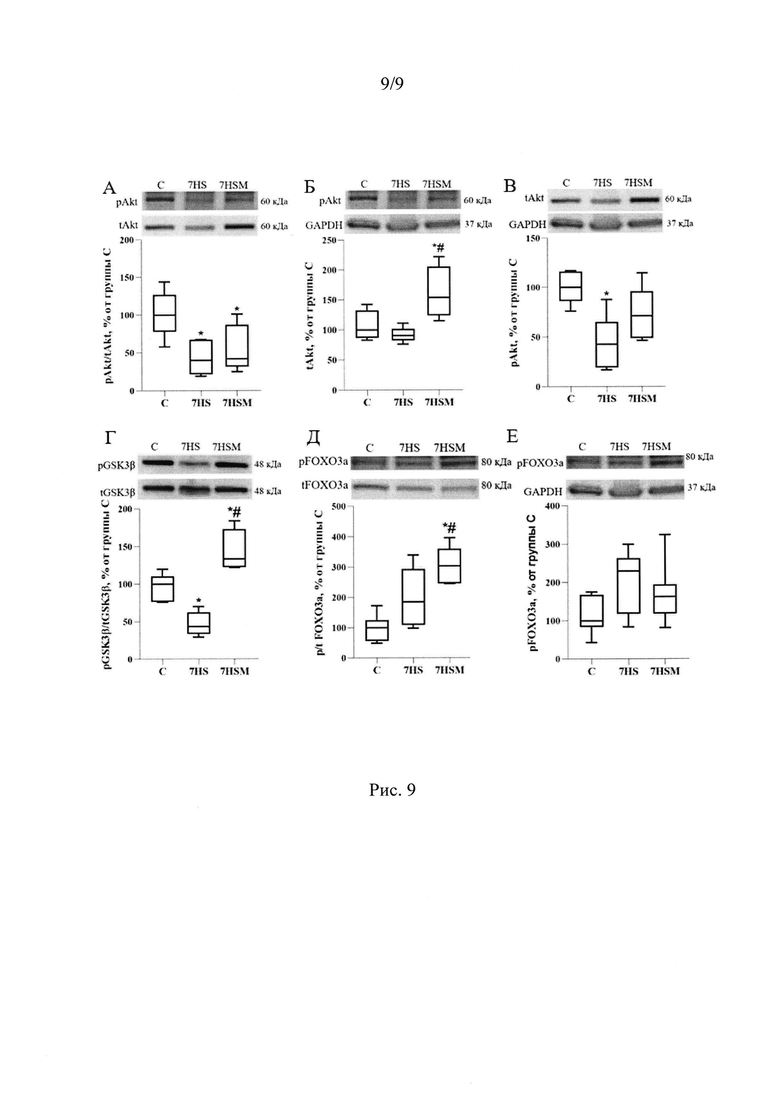
Кроме того, MOTS-c мог повлиять на эффективность синтеза белка. В группе, получавшей MOTS-c, уровень тотального белка киназы Akt (протеинкиназы В) повышался на 70% по сравнению с группой чистого вывешивания, однако, уровень фосфорилирования Akt (по Ser 473) не изменялся (Рис 9 А). Киназа Akt является ключевым ферментом сигнального пути PI3K/AKT и вовлечена в регуляцию пролиферации, роста и выживания клеток, фосфорилирует mTOR (Chen et al., 2001), а также способна блокировать экспрессию убиквитин лигаз MuRF-1 и атрогин-1 за счет увеличения уровня фосфорилирования транскрипционного фактора FOXO3a (Sandri et al., 2004). 7-суточная функциональная разгрузка в описываемом эксперименте не повлияла на уровень фосфорилирования FOXO3a, однако в группе с введением MOTS-C уровень фосфорилирования FOXO3a был достоверно более высок, чем в группе контроля (Рис 9Д, Е).
Киназа Akt инактивирующе фосфорилирует GSK3β (Ser9), мы показали, что в группе чистого вывешивания снижается уровень инактивирующего фосфорилирования GSK3β по сравнению с группой контроля на 50%. В то время, как в группе, получавшей MOTS-c, не только полностью предотвращается это снижение, но и уровень инактивирующего фосфорилирования GSK3β (Ser9) растет на 48% по сравнению с группой контроля. Таким образом митокин может влиять на скорость синтеза белка (Рис 9Б).
Все перечисленные сигнальные эффекты MOTS-C, включая активацию параметров синтеза белка и снижение параметров протеолиза, частичное предотвращение снижения параметров протеолиза и трансформации миозинового фенотипа, могли способствовать предотвращению повышения утомляемости камбаловидной мышцы на фоне 7-суточного антиортостатического вывешивания.
В качестве модели функциональной разгрузки мы использовали антиортостатическое вывешивание. Животное вывешивалось за хвост в специально оборудованной клетке таким образом, чтобы задние конечности были приподняты и не касались пола клетки (и не получали опорную стимуляцию), а передние - опирались на пол. Тело животного при этом было наклонено под углом 30°. С помощью передних лап животное свободно передвигалось по решетчатому полу клетки в любом направлении, имело доступ к воде и корму ad libitum (Е.А. Ильин, Новиков, 1980).
Животные были поделены на три группы: С-виварный контроль; 7HS - 7-суточное вывешивание с внутрибрюшинным введением физиологического раствора; 7HSM - 7-суточное вывешивание с ежедневным внутрибрюшинным введением MOTS-c (40 мг/кг).
После экспериментов у животных под бромэтанольным наркозом из задних конечностей изымались камбаловидные мышцы, а затем замораживались в жидком азоте. Затем крыс эвтаназировали методом цервикальной дислокации. Из замороженной ткани выделяли тотальную белковую фракцию, ДНК и мРНК, после чего проводили синтез кДНК. Содержание белков было проанализировано в тотальной белковой фракции с помощью Вестерн-блота. Экспрессию мРНК и содержание митохондриальной ДНК оценивали с помощью ПЦР-анализа в реальном времени. С помощью иммуногистохимического окрашивания оценивалась степень атрофии постуральной мышцы и трансформации медленных волокон в быстрые.
Все эксперименты проводились с соблюдением биоэтических норм и одобрены Комиссией по биомедицинской этике ГНЦ РФ-ИМБП РАН (№628 от 6 декабря 2022 года).
Анализ содержания белков-маркеров в m. soleus проводился с помощью электрофореза с последующим Вестерн-блоттингом.
Для этого образцы m. soleus были прогомогенизированы в шариковом гомогенизаторе TissueLyser LT (QIAGEN, Германия) в течение 25 минут в 100 мкл лизирующего буфера RIPA (Santa-Cruz, USA). При этом дополнительно использовались ингибиторы протеаз Complete Protease Inhibitor Coctail («Santa Cruz», США), 10 мкг/мл апротинина («SIGMA», США), 10 мкг/мл леупептина («SIGMA», США), 10 мкг/мл пепстатина («SIGMA», США), «complete Mini Protease Inhibitor Cocktail» (Roche, Швейцария) 20 мкл, фосфоингибиторный коктейль (Santa Cruz, США), 40 мкл.
Часть мышечных лизатов отбирали для определения концентрации общего белка с помощью реактива Бредфорда (Bio-Rad Laboratories, США). Мы готовили стандарты БСА в концентрации 0 мг/мкл, 0,25 мг/мкл, 0,5 мг/мкл, 0,7 мг/мкл, 1 мг/мкл. Затем мы разводили пробы 1:25 (3 мкл пробы к 72 мкл H2O). После добавляли к стандартам и пробам реактив Бредфорда, из расчета к 6 мкл пробы 300 мкл реактива. Наносили пробы и стандарты на специальный планшет. Затем измеряли концентрацию на спектрофотометре Epoch (BioTek Instruments, США) при длине волны 595 нм.
Электрофорез проводили в полиакриламидном геле (ПААГ), для этого образцы разводились в двукратном буфере для образцов (5,4 мМ Tris-HC1 (рН 6,8), 4%-ный Ds-Na, 20%-ный глицерин, 10%-ный β-меркаптоэтанол, 0,02%-ный бромфеноловый синий). Для проведения электрофореза использовали 10%-ный разделяющий ПААГ. Образцы каждой группы загружались на один гель с контрольными образцами. Сила тока, при которой проводился электрофорез составляла 12 мА на гель первые 30 минут и 17 мА на гель последующие 45 минут в мини-системе («Bio-Rad Laboratories») при комнатной температуре.
Электроперенос белков проводили на нитроцеллюлозную мембрану при 100 V при температуре +4°С в течение 120 мин в системе mini Trans-Blot («Bio-Rad Laboratories»). После электропереноса НЦ-мембраны инкубировали в растворе 5%-ого сухого молока («Bio-Rad Laboratories») в PBST (PBS + 0,1% Tween 20) в течение 1 часа при комнатной температуре. Для выявления белковых полос использовали первичные поликлональные антитела против MOTS-C (MBS542112, Mybiosource, США, 1:1000)), GAPDH Cell Signaling Technology, США, 1:10 000), COX1/MT-CO1 (1: 1000, Cell Signaling Technology, США), p-AMPK (Cell Signaling Technology, США, 1:1000), AMPK (Cell Signaling Technology, США, 1:1000), p-ACC (Cell Signaling Technology, США, 1:1000), ACC (Cell Signaling Technology, США, 1:1000) p-AKT (Ser473) (1:1000, Cell Signaling Technology, USA, #4058), AKT (1:2000, Cell Signaling Technology, USA, #9272), p-GSK-3β (Ser 9) (1:1000, Cell Signaling Technology, USA, #9322), GSK-3β (1:1000, Cell Signaling Technology, USA, #12456).
В качестве вторичных антител использовали антитела goat anti-rabbit или goat-anti-mouse, конъюгированные с пероксидазой хрена (Santa Cruz, США) в разведении 1:30 000. Определение тотальной формы белка проводилось на той же мембране, что и определение фосфорилированной формы: комплекс антител к фосфорилированной форме белка снимали с помощью инкубации в Restore buffer (Thermo Fisher, США), после чего проводили инкубацию с антителами против тотальной формы белка.
Инкубация блотов с первичными антителами проводилась в течение ночи при 4°С, со вторичными антителами 1 час при комнатной температуре. Выявление белковых полос осуществляли с помощью набора Immun Star Substrate Kit («BioRad Laboratories», США). Анализ белковых полос проводили с использованием C-DiGit Blot Scanner (LI-COR Biotechnology, США). Все измерения плотности изображений проводились в линейном диапазоне проявляющего реагента и сканера. Нормирование содержания белков проводили по содержанию GAPDH, уровень фосфорилирования определяли по соотношению содержания фосфорилированной и тотальной форм белка.
ПЦР в реальном времени
Для исследования экспрессии генов и содержания митохондриальной ДНК методом ПНР в реальном времени проводили выделение РНК и ДНК из образцов мышечной ткани.
Обратную транскрипцию тотальной РНК и синтез кДНК проводили с помощью набора для обратной транскрипции ОТ-1 (Синтол, Россия) согласно рекомендациям производителя.
Для проведения ПЦР в реальном времени использовали праймеры с концентрацией 10 мкМ мастер-микс (0,3 мМ дНТФ, 3 мМ MgCl2, 2,5 мкл 10-кратного ПЦР-буфера Б (рН 8,8), 0,06 ед./мкл Taq ДНК-полимеразы, «Синтол»). Для анализа содержания нуклеиновых кислот в пробах использовали метод анализа данных ПЦР по Pffafl (Pfaffl, 2001). Нормирование уровня экспрессии проводили по экспрессии гена «домашнего хозяйства» Sumo, так как продукт этого гена не вовлечен в процессы, происходящие при функциональной разгрузке, экспрессия мРНК Sumo не имеет достоверных различий между группами.
Используемые в работе праймеры:


Иммуногистохимический анализ мышечных волокон
С помощью криостата с микротомом Leica СМ 1900 при 20°С готовили поперечные срезы камбаловидной мышцы крысы толщиной 7 мкм. Затем срезы высушивали при комнатной температуре в течение 10 мин и инкубировали в PBST в течение 15 мин.
Затем готовили раствор первичных антител на PBST. Использовали первичные антитела МуНС fast, 1:60 (DSMZ, Германия), в растворе которых инкубировали срезы в течение ночи при +4°С. В качестве вторичных антител использовались Alexa Fluor 546 (1:1000) и Alexa Fluor 488 (1:1000) (Waltham, Massachusetts, USA). Срезы исследовали и фотографировали с помощью флуоресцентного микроскопа Leica Q500MC со встроенной цифровой камерой (ТСМ 300F, Leica, Брауншвейг, Германия) при 20-кратном увеличении объектива. Анализ площади поперечного сечения (ППС) быстрого типа проводили с помощью программного обеспечения ImageJ 1.52а. Было проанализировано не менее 10 поперечных сечений на образец для определения процентного содержания типов мышечных волокон в образце (n=8), и в каждом образце оценивалось не менее 100 волокон каждого типа. Процентное содержание медленных волокон вычисляли делением числа медленных волокон на сумму быстрых и медленных, полученное значение умножали на 100%. Для подсчета количества медленных мышечных волокон мы проводили иммуногистохимическое окрашивание на быструю изоформу тяжелой цепи миозина, чтобы исключить ошибку, связанную с окраской смешанных типов мышечных волокон.
Статистическая обработка данных
Статистический анализ проводили в программе GraphPad Prism 8.0.1, для проверки выборки на принадлежность нормальному распределению использовали критерий Колмогорова-Смирнова, достоверность отличий между группами определяли с помощью one-way ANOWA с апостериорным критерием Тьюки (для нормально распределенных выборок), или с помощью критерия Краскелла-Уоллеса, для распределений отличных от нормального. Статистически достоверными различия считали при уровне значимости р<0,05.
Содержание пептида MOTS-c в постуральной мышце на 7 сутки функциональной разгрузки
Мы показали, что на 7 сутки функциональной разгрузки в m. soleus крысы содержание MOTS-c снижается на 60% по сравнению с контрольной группой. Внутрибрюшинное введение MOTS-C вывешенным животным предотвращает снижение содержания MOTS-C в миоплазме камбаловидных мышц (Рис. 1А).
Активные механические характеристики m. soleus при введении MOTS-c
Индекс утомления достоверно снижался в группе 7HS, по сравнению с группой контроля. В группе 7HSM частично предотвращалось это снижение (Рис. 2).
Влияние MOTS-c на атрофию и синтез белка
Вес m. soleus в группах вывешивания 7HS и 7HSM не различался и был достоверно ниже, чем в группе виварного контроля (66,80±9,1 мг и 71,81±3,6 мг соответственно против 111,3±13,5 мг в группе контроля, р<0,05). Однако, по данным иммуногистохимии, MOTS-c частично предотвращает атрофию медленных мышечных волокон - площадь поперечного сечения (ППС) этих волокон в группе 7HS на 51% ниже (р<0,05), чем в группе С. Но в группе, получавшей MOTS-c, ППС достоверно отличается как от группы 7HS (на 22% больше), так и от группы С (на 29% меньше), р<0,05 (Рис 3 А, Б).
Количество 28S и 18S рРНК в группе 7HS достоверно снижается на 59% и 38% соответственно (р<0,05), по сравнению с группой виварного контроля, что свидетельствует об уменьшении количества рибосом и, следовательно, снижении интенсивности синтеза белка. Но в группе 7HSM нет значимых отличий содержания 28S рРНК от группы С, а содержание 18S рРНК в группе 7HSM достоверно больше (на 36%), р<0,05, чем в группе 7HS и не отличается от группы С (Рис 7А, Б).
Кроме того, в группе 7HS достоверно повышается экспрессия мРНК убиквитинлигазы MuRFl (на 130%) и белка Atrogin-1 (на 58%), р<0,05, повышение мРНК которого является маркером атрофии в скелетной мышце (Gomes et al., 2001). Но введение митокина MOTS-c на фоне 7 суток функциональной разгрузки предотвратило рост экспрессии мРНК маркеров атрофии MuRF1 и Atrogin-1 (в группе 7HSM содержание мРНК на 160% и на 70% соответственно ниже (р<0,05), чем в группе 7HS, и находится на уровне группы С) (Рис 8А, Б).
Влияние MOTS-c на миозиновый фенотип
После 7 суток вывешивания в группе 7HS по сравнению с группой С, процент мышечных волокон медленного типа был ниже на 23%, а процент «быстрых» мышечных волокон существенно выше на 14% (р<0.05). Введение MOTS-c предотвратило эти изменения. В группе 7HSM процент «быстрых» мышечных волокон на 13% ниже (р<0.05), чем в группе 7HS и не отличается достоверно от группы С, а процент «медленных» мышечных волокон на 14% выше, чем в группе 7HS (р<0.05) и не отличался от процента контрольной группы (р<0.05) (Рис 4).
Кроме того, в группе 7HS, по сравнению с группой контроля, содержание мРНК быстрых изоформ МуНС-IIB и MyHC-IId/x достоверно растет, снижается содержание быстрой изоформы тяжелой цепи миозина МуНС-IIa. В группе, получавшей MOTS-c эти изменения полностью предотвращаются. Содержание мРНК быстрых изоформ МуНС-IIВ и МуНС-IId/x в группе 7HSM достоверно ниже, чем в группе 7HS, и не отличается от контроля, а содержание мРНК быстрой изоформы МуНС-IIа выше на 35%, чем в группе 7HS.Также в группе 7HSM показано достоверное увеличение экспрессии мРНК медленной изоформы тяжелой цепи миозина МуНС-I на 23% и 40%, по сравнению с группами 7HS и С (Рис 5А-Г).
Влияние MOTS-C на параметры биогенеза митохондрий
Вывешивание задних конечностей крыс в течение 7 суток привело к достоверному снижению снижению уровня экспрессии PGC1 и его мишени, I субъединицы IV комплекса дыхательной цепи митохондрий СОХ I. В группе с введением MOTS-C отличия содержания митохондриальной ДНК, а также уровня экспрессии PGC1 и СОХ I не были достоверными (Рис 6А-Г).
7-суточная функциональная разгрузка m. soleus привела к достоверному снижению ППС медленных мышечных волокон, трансформации миозинового фенотипа, и росту утомляемости мышцы. В группе, получавшей MOTS-c, мы обнаружили частичное предотвращение атрофии медленных мышечных волокон, снижение утомляемости и сохранение миозинового фенотипа в m. soleus крысы.
Причины, предотвращающие атрофию медленных мышечных волокон, трансформацию миозинового фенотипа и снижении утомляемости m. soleus в нашем эксперименте, могут быть следующие.
1. Наблюдаемые эффекты 7-суточного вывешивания на изменение процентов «быстрых» и «медленных» мышечных волокон в группе 7HS согласуются с ранее полученными данными в нашей лаборатории (Sharlo et al., 2021). Вклад в увеличение устойчивости m. soleus к утомлению в группе 7HSM может вносить предотвращение снижения «медленных» мышечных волокон и увеличение количества «быстрых» мышечных волокон в этой группе, а также предотвращение снижения содержания митохондриальной ДНК.
2. Данные о частичном предотвращении атрофии медленных мышечных волокон в группе 7HSM, полученные с помощью иммуногистохимического анализа согласуются с данными ПЦР о снижении экспрессии мРНК маркеров атрофии убиквитинлигаз MuRF-1 и Atrogin-1, а также с данными о повышении фосфорилирования GSK3beta по ингибирующему сайту, для которой характерно фосфорилирование elF2Be, который обычно тормозит общий синтез белка (Welsh et al., 1998)
Также о повышении интенсивности синтеза белка в группе 7HSM по сравнению с группой 7HS свидетельствует повышение содержания 18S и 28S рРНК.
3. Введение митокина MOTS-c предотвратило трансформацию миозинового фенотипа на фоне 7-суточной функциональной разгрузки. Похожие результаты были получены Kumagai на мышах, в этих исследованиях MOTS-c оказывал стимулирующее влияние на экспрессию медленных изоформ миозина (Kumagai et al., 2022). Этот эффект может быть связан с влиянием MOTS-c на экспрессию ферментов цикла фолиевой кислоты, что в конечном итоге приводит к активации АМПК. Данные о повышении фосфорилирования и активности АМПК в группе 7HSM, по сравнению с группами С и 7HS согласуются с литературными данными (Lee et al., 2015). Ранее в нашей лаборатории было показано, что АМПК может влиять на миозиновый фенотип мышечных волокон (Vilchinskaya et al., 2018, Sharlo et al., 2023).
Краткое описание рисунков
Рис. 1. Содержание MOTS-c (А), АМПК (В), уровень фосфорилирования АМПК (Б), АКК (Г) в тотальной белковой фракции m. soleus крысы. С -контрольная группа (n=8), 7HS - группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде диаграмм размаха (на графике присутствуют медиана, первая и третья квартили, максимум и минимум), значения представлены в процентах от медианы контрольной группы. * - отличия от группы С, # -отличия от группы 7HS
Рис. 2. Индекс утомляемости m. soleus. С - контрольная группа (n=8), 7HS - группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде диаграмм размаха (на графике присутствуют медиана, первая и третья квартили, максимум и минимум), значения представлены в процентах от медианы контрольной группы. * - отличия от группы С, # - отличия от группы 7HS
Рис. 3. Площадь поперечного сечения волокон быстрого (А) и медленного (Б) типа m. soleus крысы. С - контрольная группа (n=8), 7HS -группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде диаграмм размаха (на графике присутствуют медиана, первая и третья квартили, максимум и минимум), значения представлены в процентах от медианы контрольной группы. * - отличия от группы С, # - отличия от группы 7HS
Рис. 4. Процентное соотношение мышечных волокон медленного и быстрого типа m. soleus крысы. С - контрольная группа (n=8), 7HS - группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде среднего и стандартного отклонения. * - отличия от группы С, # - отличия от группы 7HS
Рис. 5. Содержание мРНК МуНС I (А), МуНС IIa (Б), МуНС IIB (В), МуНС II d/x (Г) в m. soleus крысы. С - контрольная группа (n=8), 7HS - группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде диаграмм размаха (на графике присутствуют медиана, первая и третья квартили, максимум и минимум), значения представлены в процентах от медианы контрольной группы. * - отличия от группы С, # - отличия от группы 7HS
Рис. 6. Содержание COXI (A), PGC1α (Б), в тотальной белковой фракции m. soleus крысы, содержание мРНК COXI (В), PGC1α (Г) в m. soleus крысы. С - контрольная группа (n=8), 7HS - группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде диаграмм размаха (на графике присутствуют медиана, первая и третья квартили, максимум и минимум), значения представлены в процентах от медианы контрольной группы. * - отличия от группы С, # -отличия от группы 7HS
Рис. 7. Содержание 28S (А) и 18S (Б) РНК в m. soleus крысы. С -контрольная группа (n=8), 7HS - группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде диаграмм размаха (на графике присутствуют медиана, первая и третья квартили, максимум и минимум), значения представлены в процентах от медианы контрольной группы. * - отличия от группы С, # -отличия от группы 7HS
Рис. 8. Содержание мРНК Atrogin (А) и мРНК MuRF1 (Б) в m. soleus крысы. С - контрольная группа (n=8), 7HS - группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде диаграмм размаха (на графике присутствуют медиана, первая и третья квартили, максимум и минимум), значения представлены в процентах от медианы контрольной группы. * - отличия от группы С, # - отличия от группы 7HS
Рис. 9. Уровень фосфорилирования Akt (A), GSK3β (Г) и FOXO3a (Д), содержание фосфорилированной формы Akt (Б), FOXO3a(E) и тотального белка Akt (В) в m. soleus крысы. С - контрольная группа (n=8), 7HS - группа 7-суточного вывешивания (n=8), 7HSM - группа 7-суточного вывешивания с введением MOTS-c (n=8). Данные представлены в виде диаграмм размаха (на графике присутствуют медиана, первая и третья квартили, максимум и минимум), значения представлены в процентах от медианы контрольной группы. * - отличия от группы С, # - отличия от группы 7HS
Литература
1. Allen D.L., Yasui W., Тапака Т., Ohira Y, Nagaoka S., Sekiguchi C, Hinds W.E., Roy R.R., Edgerton V.R. Myonuclear number and myosin heavy chain expression in rat soleus single muscle fibers after spaceflight // https://doi.org/10.1152/jappl.1996.81.l.145. 1996. V. 81, №1. - P. 145-151.
2. Baehr L.M., West D. W.D., Marshall A.G., Marcotte G.R., Baar K, Bodine S.C. Muscle-specific and age-related changes in protein synthesis and protein degradation in response to hindlimb unloading in rats // J. Appl. Physiol. 2017. V. 122, №5.- P. 1336-1350.
3. Chen W.S., Xu P.Z., Gottlob K, Chen M.L., Sokol K, Shiyanova Т., Roninson I., Weng W., Suzuki R., TobeK., Kadowaki Т., Hay N. Growth retardation and increased apoptosis in mice with homozygous disruption of the akt1 gene // Genes Dev. 2001. V. 15, №17.- P. 2203.
4. Gomes M.D., Lecker S.H., Jagoe R.T., Navon A., Goldberg A.L. Atrogin-1, a muscle-specific F-box protein highly expressedduring muscle atrophy // Proc. Natl. Acad. Sci. U. S. A. 2001. V. 98, №25. - P. 14440.
5. Holmes B.F., Sparling D.P., Olson A.L., Winder W.W., Dohm G.L. Regulation of muscle GLUT4 enhancer factor and myocyte enhancer factor 2 by AMP-activated protein kinase // Am. J. Physiol. Endocrinol. Metab. 2005. V. 289, №6.
6. Jacinto E., Facchinetti V., Liu D., Soto N., Wei S., Jung S.Y., Huang Q., Qin J., Su B. SIN1/MIP1 maintains rictor-mTOR complex integrity and regulates Akt phosphorylation and substrate specificity // Cell. 2006. V. 127, №1. - P. 125-137.
7. Kim K.Н., Son J.M., Benayoun B.A., Lee C. The Mitochondrial-Encoded Peptide MOTS-c Translocates to the Nucleus to Regulate Nuclear Gene Expression in Response to Metabolic Stress // Cell Metab. 2018. V. 28, №3. - P. 516- 524.e7.
8. Kumagai H, Kim S.-J., Miller В., Natsume Т., Wan J., Kumagai M.E., II R.R., Lee S.H., Sato A., Mehta H.H, Yen K, Cohen P. Mitochondrial-derived microprotein MOTS-c attenuates immobilization-induced skeletal muscle atrophy by suppressing lipid infiltration // https://doi.org/10.1152/ajpendo.00285.2023. 2024.
9. Kumagai H., Natsume Т., Kim S.J., et al. The MOTS-c K14Q polymorphism in the mtDNA is associated with muscle fiber composition and muscular performance // Biochim. Biophys. acta. Gen. Subj. 2022. V. 1866, №2.
10. Lee С., Zeng J., Drew B.G., Sallam Т., Martin-Montalvo A., Wan J., Kim S.J., Mehta H., Hevener A.L., Cabo R. De, Cohen P. The mitochondrial-derived peptide MOTS-c promotes metabolic homeostasis and reduces obesity and insulin resistance // Cell Metab. 2015. V. 21, №3. - P. 443-454.
11. Liu J., Peng Y, Cui Z., Wu Z., Qian A., Shang P., Qu L., Li Y, Liu J., Long J. Depressed mitochondrial biogenesis and dynamic remodeling in mouse tibialis anterior and gastrocnemius induced by 4-week hindlimb unloading // IUBMB Life. 2012. V. 64, №11.- P. 901-910.
12. Reynolds J.C., Lai R.W., Woodhead J.S.T., Joly J.H., Mitchell C.J., Cameron-Smith D., LuR., Cohen P., Graham N.A., BenayounB.A., Merry T.L., Lee С.MOTS-c is an exercise-induced mitochondrial-encoded regulator of age-dependent physical decline and muscle homeostasis // Nat. Commun. 2021 121. 2021. V. 12, №1. - P. 1-11.
13. Rozhkov S. V., Sharlo K.A., Mirzoev T.M., Shenkman B.S. Temporal changes in the markers of ribosome biogenesis in rat soleus muscle under simulated microgravity // Acta Astronaut. 2021. V. 186, - P. 252-258.
14. Sandri M., Sandri C., Gilbert A., Skurk C., Calabria E., Picard A., Walsh K., Schiaffino S., Lecker S.H., Goldberg A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy // Cell. 2004. V. 117, №3. - P. 399-412.
15. Schiaffino S., Reggiani C. Fiber types in mammalian skeletal muscles // Physiol. Rev. 2011. V.91, №4. - P. 1447-1531.
16. Sharlo K., Tyganov S.A., Tomilovskaya E., Popov D.V., Saveko A.A., Shenkman B.S. Effects of Various Muscle Disuse States and Countermeasures on Muscle Molecular Signaling // Int. J. Mol. Sci. 2022. V. 23, №1.
17. Sharlo K.A., Lvova I.D., Belovq S.P., Zaripova K.A., Shenkman B.S., Nemirovskaya T.L. Metformin Attenuates Slow-to-Fast Fiber Shift and Proteolysis Markers Increase in Rat Soleus after 7 Days of Rat Hindlimb Unloading // Int. J. Mol. Sci. 2023a. V. 24, №1.
18. Sharlo K.A., Lvova I.D., Tyganov S.A., Zaripova K.A., Belova S.P., Kostrominova T.Y., Shenkman B.S., Nemirovskaya T.L. The Effect of SERCA Activation on Functional Characteristics and Signaling of Rat Soleus Muscle upon 7 Days of Unloading // Biomolecules. 2023b. V. 13, №9.
19. Sharlo K.A., Paramonova Lvova I.D., Mochalova E.P., Kalashnikov V.E., Vilchinskaya N.A., Tyganov S.A., Konstantinova T.S., Shevchenko T.F., Kalamkarov G.R., Shenkman B.S. Plantar Mechanical Stimulation Maintains Slow Myosin Expression in Disused Rat Soleus Muscle via NO-Dependent Signaling // Int. J. Mol. Sci. 2021. V. 22, №3. - P. 1-23.
20. Tyganov S.A., Mochalova E.P., Belova S.P., Sharlo K.A., Rozhkov S.V., Vilchinskaya N.A., Paramonova I.I., Mirzoev T.M., Shenkman B.S. Effects of Plantar Mechanical Stimulation on Anabolic and Catabolic Signaling in Rat Postural Muscle Under Short-Term Simulated Gravitational Unloading // Front. Physiol. 2019. V. 10, №SEP.
21. Vilchinskaya N.A., Krivoi I.I., Shenkman B.S. AMP-Activated Protein Kinase as a Key Trigger for the Disuse-Induced Skeletal Muscle Remodeling // Int. J. Mol. Sci. 2018. V. 19.,. №11.
22. Vilchinskaya N.A., Mochalova E.P., Nemirovskaya T.L., Mirzoev T.M., Turtikova О. V, Shenkman B.S. Rapid decline in MyHC I(β) mRNA expression in rat soleus during hindlimb unloading is associated with AMPK dephosphorylation // J. Physiol. 2017. V. 595, №23. - P. 7123-7134.
23. Welsh G.I., Miller C.M., Loughlin A.J., Price N.T., Proud C.G. Regulation of eukaryotic initiation factor eIF2B: glycogen synthase kinase-3 phosphorylates a conserved serine which undergoes dephosphorylation in response to insulin // FEBS Lett. 1998. V. 421.,. №2. - P. 125-130.
24. E.А. Ильин, Новиков B.E. Стенд для моделирования физиологических эффектов невесомости в лабораторных условиях // Косм. биол. мед. 1980. V. 14, №3.- Р. 79-80.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ предотвращения развития утомления скелетной мышцы | 2023 |
|
RU2814077C1 |
| Способ поддержания силы и жесткости постуральных мышц млекопитающих при их функциональной разгрузке | 2022 |
|
RU2800317C1 |
| Способ предотвращения атрофии скелетных мышц при их функциональной разгрузке | 2022 |
|
RU2797216C1 |
| СРЕДСТВО ДЛЯ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517576C1 |
| СПОСОБ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2012 |
|
RU2481105C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517259C1 |
| ПРИМЕНЕНИЕ VX 745 ДЛЯ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2019 |
|
RU2714923C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2010 |
|
RU2444354C2 |
| ФИТОЭКДИЗОНЫ И ИХ ПРОИЗВОДНЫЕ ДЛЯ ПРИМЕНЕНИЯ В ЛЕЧЕНИИ НЕРВНО-МЫШЕЧНЫХ ЗАБОЛЕВАНИЙ | 2020 |
|
RU2820936C2 |
| СПОСОБ УСКОРЕНИЯ ВОССТАНОВЛЕНИЯ СКЕЛЕТНЫХ МЫШЦ ОТ АТРОФИИ ПОСЛЕ ДЛИТЕЛЬНОЙ АЛКОГОЛЬНОЙ ИНТОКСИКАЦИИ | 2014 |
|
RU2578463C1 |
Изобретение относится к медицине, а именно к физиологии. Предотвращают рост экспрессии мРНК маркеров атрофии MuRF1 и Atrogin-1, а также снижение содержания 18S рРНК и 28S рРНК. Для этого внутрибрюшинно в дозировке 40 мг/кг в 0,9% растворе NaCl вводят МИТОКИН MOTS-C на фоне 7-суточной функциональной разгрузки. Способ позволяет предотвратить атрофию, повышенную утомляемость медленных мышечных волокон и их трансформацию в быстрые мышечные волокна в камбаловидной мышце в эксперименте во время функциональной разгрузки при введении MOTS-c. 9 ил.
Способ предотвращения атрофии, повышенной утомляемости медленных мышечных волокон и их трансформации в быстрые мышечные волокна в камбаловидной мышце, характеризующийся тем, что предотвращают рост экспрессии мРНК маркеров атрофии MuRF1 и Atrogin-1, а также снижение содержания 18S рРНК и 28S рРНК, для чего внутрибрюшинно в дозировке 40 мг/кг в 0,9% растворе NaCl вводят МИТОКИН MOTS-C на фоне 7-суточной функциональной разгрузки.
| ШАРЛО К.А | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ПРИМЕНЕНИЕ VX 745 ДЛЯ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2019 |
|
RU2714923C1 |

