РОДСТВЕННЫЕ ЗАЯВКИ НА ПАТЕНТЫ
Данная заявка притязает на приоритет предварительной заявки на патент США №62/689,053, поданной 22 июня 2018 г., полностью включенной сюда посредством ссылки.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Многие цитокины изучались как потенциальные терапевтические агенты для лечения заболеваний. Тем не менее, их избыточная системная стимуляция или избыточное подавление иммунной системы организма сильно ограничивали их разработку и клиническое применение.
У интерлейкина-2 (IL-2) и интерлейкина-15 (IL-15) есть общие рецепторные компоненты (γс и IL-2Rβ) и сигнальные пути, и они выполняют несколько сходных функций. Оба цитокина стимулируют пролиферацию Т-клеток, индуцируют развитие цитотоксических Т-лимфоцитов (CTL), способствуют пролиферации В-клеток и синтезу ими иммуноглобулинов и индуцируют развитие и персистенцию натуральных клеток-киллеров (NK-клеток). Исходя из многочисленных доклинических исследований, а также множества клинических исследований, оба цитокина рассматриваются как потенциально ценные терапевтические агенты при раке, аутоиммунных расстройствах, воспалительных расстройствах, трансплантации и различных других расстройствах. Рекомбинантный IL-2 был одобрен для применения у пациентов с метастатическим почечно-клеточным раком и злокачественной меланомой. Применительно к IL-15 в настоящее время проводится несколько онкологических клинических исследований, однако его применение пока не одобрено. Кроме того, как IL-2, так и IL-15 имеют третью, уникальную, несигнальную рецепторную α-субъединицу, IL-2Rα (также известную как CD25) или IL-15Rα, соответственно, которая может способствовать различиям в их рецепторной специфичности и биологических функциях.
Рекомбинантный человеческий IL-2 является эффективной иммунотерапией, применяемой при метастатической меланоме и раке почки с продолжительными ответами приблизительно у 10% пациентов. Тем не менее, оптимальное дозирование IL-2 ограничено коротким периодом полувыведения и выраженной токсичностью. Кроме того, IL-2 также связывается с его гетеротримерным рецептором IL-2Rαβγ с большей аффинностью, что приводит к преимущественному росту числа иммуносупрессивных регуляторных Т-клеток (Treg) с высокими уровнями конститутивной экспрессии IL-2Rα. Рост числа Treg может быть нежелательным эффектом IL-2 при иммунотерапии рака. Тем не менее, способность IL-2 стимулировать Treg-клетки даже в низких дозах может быть использована для лечения аутоиммунных и хронических воспалительных расстройств. Недавно было обнаружено, что IL-2 может быть модифицирован для селективной стимуляции цитотоксических эффекторных Т-клеток или Treg-клеток. Различные методики позволили получить варианты IL-2 с улучшенной и селективной иммуномодулирующей активностью.
Как IL-2, так и IL-15 являются сильными агонистами эффекторных иммунных клеток, и очень важно, чтобы цитотоксические иммунные клетки полностью активировались только во время их нахождении в очаге заболевания или в непосредственной близости к нему, например в раковом очаге для специфичного уничтожения только опухолевых клеток или в очаге воспаления только для действия, направленного против аутоиммунных или хронических воспалительных расстройств. Повышение специфичности и селективности в отношении мишеней и оставление здоровых клеток и тканей интактными и неповрежденными представляют большой интерес применительно ко всем цитокинам, хемокинам и факторам роста.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В одном аспекте согласно настоящему изобретению предложена биологически активируемая лекарственная платформа на основе цитокинов («VitoKine»), направленная на снижение системной токсичности, основанной на механизме действия, и обеспечение более широкого терапевтического применения цитокинов, хемокнов, гормонов и факторов роста, таких как IL-15 и IL-2, для лечения рака, аутоиммунных расстройств, воспалительных расстройств и различных других расстройств. Платформа VitoKine определена конструкциями, показанными на ФИГ. 1, и предложенными способами активации, показанными на ФИГ. 2. Со ссылкой на ФИГ. 1, новые конструкции VitoKine по настоящему изобретению содержат 3 домена: 1) домен D1 («D1»), выбранный из группы, состоящей из тканенаправленного домена, домена, увеличивающего период полувыведения, или домена группировки с двойной функцией; (2) домен D2 («D2»), представляющий собой «домен активной группировки»; и (3) домен D3 («D3»), представляющий собой «домен маскирующей группировки». Важно, что домен D2 конструкции VitoKine остается почти инертным или минимально активным до его местной активации протеазами, регуляция которых повышена в тканях, пораженных заболеванием, или гидролизом в очагах заболевания, что будет ограничивать связывание активной группировки с ее рецепторами на периферии или на поверхности клеток, не пораженных заболеванием, или нормальных тканей, предотвращая избыточную активацию ее пути и уменьшая нежелательную токсичность, обусловленную связыванием с мишенью вне целевой ткани, и нежелательный клиренс, обусловленный связыванием с мишенью.
В различных воплощениях конструкции VitoKine по настоящему изобретению содержат D1, представляющий собой направленную группировку, такую как антитело или фрагмент антитела, связывающиеся с опухоль-ассоциированным антигеном (ТАА) или тканеспецифичным антигеном, молекулой клеточной поверхности, или белком внеклеточного матрикса, или протеазой(ами), или любым(и) остатком(ами), подверженным(и) посттрансляционной модификации. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат D1, представляющий собой направленную группировку, такую как белок или пептид, демонстрирующий аффинность связывания в отношении клетки или ткани, пораженной заболеванием. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат D1, представляющий собой модифицированный белок или пептид, такой как гликан-модифицированный белок или пептид, демонстрирующий аффинность связывания в отношении определенного рецептора, такого как рецептор лектинов типа с, экспрессированного на клетке или ткани, пораженной заболеванием. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой антитело к модулятору иммунных контрольных точек. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат D1, функция которого состоит в удержании цитокина в тканевом очаге. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат D1, являющийся бифункциональным, например, тканенаправленным и обеспечивающим удержание в ткани. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой полимер. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой группировку, увеличивающую период полувыведения. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой Fc-домен (или его функциональный фрагмент).
«Fc-домен» относится к димеру двух Fc-доменных мономеров, который обычно содержит полноразмерную шарнирную область или ее часть. В различных воплощениях Fc-домен выбран из группы, состоящей из Fc-домена человеческого IgG1 (иммуноглобулин G1), Fc-домена человеческого IgG2, Fc-домена человеческого IgG3, Fc-домена человеческого IgG4, Fc-домена IgA, Fc-домена IgD, Fc-домена IgE, Fc-домена IgG и Fc-домена IgM, или любой их комбинации. В различных воплощениях Fc-домен содержит аминокислотное изменение, приводящее к Fc-домену, имеющему измененные свойства связывания с комплементом или Fc-рецепторами. Аминокислотные изменения, приводящие к получению Fc-домена с измененными свойствами связывания с комплементом или Fc-рецепторами, известны в данной области. В различных воплощениях последовательность Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность Fc-домена человеческого IgG1, изложенную в SEQ ID NO:13. В различных воплощениях последовательность Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность, изложенную в SEQ ID NO:14, содержащую аминокислотные замены, устраняющие связывание с FcγR и C1q. В различных воплощениях Fc-домен содержит аминокислотные изменения, приводящие к дополнительному увеличению периода полувыведения in vivo. Аминокислотные изменения, приводящие к получению Fc-домена с дополнительно увеличенным периодом полувыведения, известны в данной области. В различных воплощениях последовательность Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность, изложенную в SEQ ID NO:156 или 166, обе из которых содержат аминокислотные замены, устраняющие связывание с FcγR и C1q и увеличивающие период полувыведения in vivo. В различных воплощениях последовательность гетеродимерного Fc-домена, используемая для получения конструкций VitoKine, имеет происхождение от последовательности Knob-Fc-домена, изложенной в SEQ ID NO:15. В различных воплощениях последовательность гетеродимерного Fc-домена, используемая для получения конструкций VitoKine, имеет происхождение от последовательности Hole-Fc-домена, изложенной в SEQ ID NO:16. В различных воплощениях последовательность гетеродимерного Fc-домена, используемая для получения конструкций VitoKine, имеет происхождение от последовательности Knob-Fc-домена с увеличенным периодом полувыведения in vivo, изложенной в SEQ ID NO:167. В различных воплощениях последовательность гетеродимерного Fc-домена, используемая для получения конструкций VitoKine, имеет происхождение от последовательности Hole-Fc-домена с увеличенным периодом полувыведения in vivo, изложенной в SEQ ID NO:168.
В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D2, представляющий собой белок. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D2, представляющий собой цитокин, выбранный из группы, включающей, без ограничения, IL-1, IL-2, IL-4, IL-5, IL-6, IL-7, IL-9, IL-10, IL-12, IL-15, IL-23 и лиганды суперсемейства трансформирующего фактора роста β (TGFβ), например TGFβ (SEQ ID NO:24). В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D2, представляющий собой IL-15. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D2, представляющий собой вариант (или мутант) IL-15, содержащий одну или более чем одну аминокислотную замену, делецию или вставку в полипептиде IL-15. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D2, представляющий собой IL-2. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D2, представляющий собой вариант (или мутант) IL-2, содержащий одну или более чем одну аминокислотную замену, делецию или вставку по сравнению с полипептидом IL-2.
В различных воплощениях домен D2 конструкции VitoKine представляет собой домен IL-15, содержащий последовательность полипептида зрелого человеческого IL-15 (также называемого здесь huIL-15 или IL-15 дикого типа (wt)), изложенную в SEQ ID NO:2. В различных воплощениях домен IL-15 будет вариантом (или мутантом) IL-15, содержащим последовательность, имеющую происхождение от последовательности полипептида зрелого человеческого IL-15, изложенной в SEQ ID NO:2. В различных воплощениях домен IL-15 будет вариантом (или мутантом) IL-15, содержащим последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% гомологичную SEQ ID NO:2. Варианты (или мутанты) IL-15 названы здесь с использованием нативной аминокислоты, ее положения в зрелой последовательности и аминокислоты варианта. Например, huIL-15 «S58D» относится к человеческому IL-15, содержащему замену S на D в положении 58 SEQ ID NO:2. В различных воплощениях вариант IL-15 функционирует как агонист IL-15, что продемонстрировано, например, повышенной связывающей активностью в отношении рецепторов IL-15Rβγc по сравнению с нативным полипептидом IL-15. В различных воплощениях вариант IL-15 функционирует как антагонист IL-15, что продемонстрировано, например, сниженной связывающей активностью в отношении рецепторов IL-15Rβγc или сходной или повышенной связывающей активностью в отношении рецепторов IL-15Rβγc, но сниженной сигнальной активностью по сравнению с нативным полипептидом IL-15 или ее устранением. В различных воплощениях вариант IL-15 имеет повышенную аффинность связывания или сниженную связывающую активность в отношении рецепторов IL-15Rβγc по сравнению с нативным полипептидом IL-15. В различных воплощениях последовательность варианта IL-15 имеет по меньшей мере одно (то есть 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более) аминокислотное изменение по сравнению с нативной последовательностью IL-15. Аминокислотное изменение может включать одну или более чем одну аминокислотную замену, делецию или вставку в полипептиде IL-15, такую как замена, делеция или вставка в домене IL-15, взаимодействующем с IL-15Rβ и/или IL-15Rβγc. В различных воплощениях аминокислотное изменение представляет собой одну или более чем одну аминокислотную замену или делецию в положении 30, 31, 32, 58, 62, 63, 67, 68 или 108 SEQ ID NO:2. В различных воплощениях аминокислотное изменение представляет собой замену D на Т в положении 30, V на Y в положении 31, Н на Е в положении 32, S на D в положении 58, Т на D в положении 61, V на F в положении 63, I на V в положении 67, I на F, или Н, или D, или К в положении 68 или Q на А, или М, или S в положении 108 последовательности зрелого человеческого IL-15 или любую комбинацию этих замен. В различных воплощениях аминокислотное изменение представляет собой замену S на D в положении 58 последовательности зрелого человеческого IL-15. В различных воплощениях полипептид IL-15 содержит вариант IL-15 по SEQ ID NO:3. В различных воплощениях домен IL-15 имеет любые комбинации аминокислотных замен, делеций и вставок.
В различных воплощениях домен D2 конструкций VitoKine по настоящему изобретению содержит полипептид IL-2. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D2, представляющий собой вариант (или мутант) IL-2, содержащий одну или более чем одну аминокислотную замену, делецию или вставку. В различных воплощениях конструкция VitoKine содержит домен D2, где домен IL-2 содержит последовательность полипептида зрелого человеческого IL-2 (также называемого здесь huIL-2 или IL-2 дикого типа (wt)), изложенную в SEQ ID NO:8. В различных воплощениях домен IL-2 будет вариантом (или мутантом) IL-2, содержащим последовательность, имеющую происхождение от последовательности полипептида зрелого человеческого IL-2, изложенной в SEQ ID NO:8. В различных воплощениях домен IL-2 будет вариантом (или мутантом) IL-2, содержащим последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% гомологичную SEQ ID NO:8. В различных воплощениях вариант IL-2 функционирует как агонист IL-2. В различных воплощениях вариант IL-2 функционирует как антагонист IL-2. В различных воплощениях аминокислотное изменение представляет собой одну или более чем одну аминокислотную замену в положении 19, 20, 38, 41, 42, 44, 88, 107, 125 или 126 SEQ ID NO:8. В различных воплощениях аминокислотное изменение представляет собой замену L на D, или Н, или N, или Р, или Q, или R, или S, или Y в положении 19, D на Е, или I, или N, или Q, или S, или Т, или Y в положении 20, R на Е, или А в положении 38, Т на А, или G, или V в положении 41, F на А в положении 42, F на G или V в положении 44, N на D, Е, или G, или I, или М, или Q, или Т, или R в положении 88, Y на G, или Н, или L, или V в положении 107, S на Е, Н, K, I или W в положении 125, Q на D, или Е, или K, или L, или М, или N в положении 126 последовательности зрелого человеческого IL-2 или любую комбинацию этих замен.
В различных воплощениях конструкции VitoKine по настоящему изобретению содержат «домен маскирующей группировки» (D3), являющийся когнатным рецептором / партнером по связыванию или любым партнером по связыванию, идентифицированным для белка или цитокина D2. В различных воплощениях домен D3 представляет собой вариант когнатного рецептора / партнера по связыванию для домена D2. В различных воплощениях домен D3 имеет усиленное связывание с доменом D2 по сравнению с когнатным рецептором / партнером по связыванию дикого типа. В различных воплощениях домен D3 имеет сниженное связывание с доменом D2 по сравнению с когнатным рецептором / партнером по связыванию дикого типа или его связывание с доменом D2 устранено. В различных воплощениях домен D3 представляет собой белок, или пептид, или антитело, или фрагмент антитела, способные маскировать активность D2. В различных воплощениях домен D3 представляет собой ДНК, фрагмент РНК или полимер, такой как PEG (полиэтиленгликоль). В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой внеклеточный домен IL-15Rα или его функциональный фрагмент. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой домен IL-15RαSushi. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой внеклеточный домен IL-2Rα или его функциональный фрагмент. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой домен IL-15RαSushi. В различных воплощениях домен D3 способен маскировать функциональную активность D2 до активации в намеченном месте терапии.
В различных воплощениях домены D1, D2 и D3 конструкции VitoKine соединены последовательностью полипептидного линкера, расщепляемого протеазой. В различных воплощениях домены D1, D2 и D3 конструкции VitoKine соединены последовательностью нерасщепляемого полипептидного линкера. В различных воплощениях оба из L1 и L2 конструкций VitoKine по настоящему изобретению представляют собой пептидные линкеры, расщепляемые протеазами. В различных воплощениях L1 конструкций VitoKine по настоящему изобретению представляет собой пептидный линкер, расщепляемый протеазой, и L2 представляет собой нерасщепляемый пептидный линкер. В различных воплощениях L1 конструкций VitoKine по настоящему изобретению представляет собой нерасщепляемый пептидный линкер и L2 представляет собой пептидный линкер, расщепляемый протеазой. В различных воплощениях оба из L1 и L2 конструкций VitoKine по настоящему изобретению представляют собой нерасщепляемые пептидные линкеры. В различных воплощениях линкер имеет высокое содержание G/S (например, по меньшей мере приблизительно 60%, 70%, 80%, 90% или более аминокислот линкера представляют собой G или S). Последовательность каждого пептидного линкера может быть выбрана независимо. В различных воплощениях линкер, расщепляемый протеазой, выбран из группы последовательностей, изложенных в SEQ ID NO:71-96 и 157-161. В различных воплощениях линкер, расщепляемый протеазой, может иметь дополнительный пептидный спейсер вариабельной длины на N-конце расщепляемого линкера, или на С-конце расщепляемого линкера, или на обоих концах расщепляемого линкера. В различных воплощениях нерасщепляемый линкер выбран из группы последовательностей, изложенных в SEQ ID NO:107-127. В различных воплощениях линкер представляет собой гибкий или ригидный линкер вариабельной длины.
В различных воплощениях домены D2 и D3 конструкции VitoKine расположены на N-конце домена D1, как показано на ФИГ. 1. В различных воплощениях домены D2 и D3 конструкции VitoKine расположены на С-конце домена D1, как показано на ФИГ. 1.
В различных воплощениях домены D1, D2 и D3 конструкции VitoKine могут представлять собой мономеры или димеры или комбинацию димера и мономера, такую как комбинация, где D1 представляет собой димер, a D2 и D3 представляют собой мономеры.
В другом аспекте согласно настоящему изобретению предложен способ лечения рака или метастазирования рака у субъекта, включающий введение терапевтически эффективного количества фармацевтических композиций по изобретению субъекту, нуждающемуся в этом. В одном воплощении субъект представляет собой субъекта-человека. В различных воплощениях рак выбран из рака поджелудочной железы, рака желудка, рака печени, рака молочной железы, рака яичника, колоректального рака, меланомы, лейкоза, миелодиспластического синдрома, рака легкого, рака предстательной железы, рака мозга, рака мочевого пузыря, рака головы и шеи, или рабдомиосаркомы, или любого рака.
В другом аспекте согласно настоящему изобретению предложен способ лечения рака или метастазирования рака у субъекта, включающий введение терапевтически эффективного количества фармацевтических композиций по изобретению в комбинации со второй терапией, выбранной из группы, состоящей из цитотоксической химиотерапии, иммунотерапии, направленной терапии низкомолекулярными ингибиторами киназ, хирургического вмешательства, лучевой терапии, трансплантации стволовых клеток, клеточных терапий, включая CAR-T, CAR-NK, iPS-индуцированные CAR-T или iPS-индуцированные CAR-NK, и вакцины, такой как бацилла Кальметта-Герена (BCG). В различных воплощениях комбинированная терапия может включать проведение у субъекта терапевтически эффективного количества иммунотерапии, включая, без ограничения: лечение с использованием истощающих антител к определенным опухолевым антигенам; лечение с использованием конъюгатов «антитело - лекарственное средство»; лечение с использованием агонистических, антагонистических или блокирующих антител к костимулирующим или коингибирующим молекулам (иммунным контрольным точкам), таким как CTLA-4, PD-1, PD-L1, CD40, ОХ-40, CD137, GITR, LAG3, TIM-3, Siglec 7, Siglec 8, Siglec 9, Siglec 15 и VISTA; лечение с использованием биспецифичных антител, привлекающих Т-клетки (BiTE®), таких как блинатумомаб; лечение, включающее введение модификаторов биологического ответа, таких как IL-12, IL-21, GM-CSF, IFN-α, IFN-β и IFN-γ; лечение с использованием терапевтических вакцин, таких как сипулейцел-Т; лечение с использованием дендритноклеточных вакцин или вакцин на основе опухолевых антигенных пептидов; лечение с использованием Т-клеток с химерными антигенными рецепторами (CAR-T-клеток); лечение с использованием CAR-NK-клеток; лечение с использованием опухоль-инфильтрирующих лимфоцитов (TIL); лечение с использованием адоптивно перенесенных противоопухолевых Т-клеток (культивированных ex vivo и/или трансгенных по TCR); лечение с использованием клеток TALL-104; и лечение с использованием иммуностимулирующих агентов, таких как агонисты Toll-подобных рецепторов (TLR) CpG и имихимод; и лечение с использованием вакцины, такой как BCG; где комбинированная терапия обеспечивает усиленное уничтожение опухолевых клеток эффекторными клетками, то есть при совместном применении конструкций VitoKine и иммунотерапии возникает синергия.
В другом аспекте согласно настоящему изобретению предложен способ лечения вирусной инфекции у субъекта, включающий введение терапевтически эффективного количества фармацевтических композиций по изобретению субъекту, нуждающемуся в этом. В одном воплощении субъект представляет собой субъекта-человека. В различных воплощениях вирус представляет собой HIV (вирус иммунодефицита человека).
В другом аспекте согласно настоящему изобретению предложен способ лечения вирусной инфекции у субъекта, включающий введение терапевтически эффективного количества фармацевтических композиций по изобретению в комбинации со второй терапией, включая, без ограничения, ацикловир, эпклусу, мавирет, зидовудин и энфувиртид.
В другом аспекте согласно настоящему изобретению предложен способ лечения аутоиммунного заболевания у субъекта, включающий введение терапевтически эффективного количества фармацевтических композиций по изобретению субъекту, нуждающемуся в этом. В одном воплощении субъект представляет собой субъекта-человека. В различных воплощениях аутоиммунное заболевание выбрано из группы, состоящей из системной красной волчанки (SLE), обыкновенной пузырчатки, злокачественной миастении, гемолитической анемии, тромбоцитопенической пурпуры, болезни Грейвса, болезни Шегрена, дерматомиозита, болезни Хашимото, полимиозита, воспалительного заболевания кишечника, рассеянного склероза (MS), сахарного диабета, ревматоидного артрита и склеродермии.
В другом аспекте согласно настоящему изобретению предложен способ лечения воспалительного заболевания у субъекта, включающий введение терапевтически эффективного количества фармацевтических композиций по изобретению субъекту, нуждающемуся в этом. В одном воплощении субъект представляет собой субъекта-человека. В различных воплощениях воспалительное заболевание выбрано из группы, состоящей из болезни Крона, язвенного колита, коллагенозного колита, лимфоцитарного колита, ишемического колита, воспаления в отключенной кишке, синдрома Бехчета и неуточненного колита.
В различных воплощениях воспалительное заболевание выбрано из группы, состоящей из других аутоиммунных и воспалительных заболеваний, таких как: ахалазия, болезнь Стилла у взрослых, агаммаглобулинемия, амилоидоз, анти-GBM/анти-ТВМ нефрит, антифосфолипидный синдром, аутоимунный ангионевротический отек, аутоиммунная вегетативная невропатия и ганглиопатия, аутоиммунный энцефаломиелит, аутоиммунное заболевание внутреннего уха, аутоиммунный оофорит, аутоиммунный орхит, аутоиммунный панкреатит, аутоиммунная ретинопатия, аутоиммунная крапивница, аксональная и нейрональная нейропатия, болезнь Бало, болезнь Бехчета, доброкачественный пемфигоид слизистых оболочек, болезнь Каслмана, болезнь Шагаса, хроническая воспалительная демиелинизирующая полинейропатия, хронический рецидивирующий мультифокальный остеомиелит, синдром Чарджа-Стросс, рубцовый пемфигоид, синдром Когана, миокардит Коксаки, CREST-синдром, герпетиформный дерматит, болезнь Девика/ нейромиелит зрительного нерва, дискоидная волчанка, синдром Дресслера, эозинофильный эзофагит, эозинофильный фасциит, узловатая эритема, эссенциальная смешанная криоглобулинемия, фиброзирующий альвеолит, гигантоклеточный артериит, гигантоклеточный миокардит, пурпура Шенлейна-Геноха, герпес беременных или пемфигоид беременных, IgA-нефропатия, IgG4-связанная склерозирующая болезнь, иммуноопосредованные нежелательные явления, миозит с включениями, интерстициальный цистит, ювенильный артрит, ювенильный миозит, синдром Ламберта-Итона, лейкоцитокластический васкулит, плоский лишай, склерозирующий лишай, деревянистый конъюнктивит, линейная IgA-болезнь, хроническая болезнь Лайма, болезнь Меньера, микроскопический полиангиит, смешанное заболевание соединительной ткани, язва Морена, болезнь Муха-Габермана, мультифокальная моторная нейропатия, неврит зрительного нерва, палиндромный ревматизм, PANDAS, паранеопластическая мозжечковая дегенерация, синдром Парри-Ромберга, промежуточный увеит, синдром Персонейджа-Тернера, перивенозный энцефаломиелит, POEMS-синдром, узелковый полиартериит, полигландулярные синдромы, ревматическая полимиалгия, постинфарктный синдром, постперикардиотомический синдром, первичный склерозирующий холангит, прогестероновый дерматит, псориатический артрит, истинная эритроцитарная аплазия, гангренозная пиодермия, феномен Рейно, рефлекторная симпатическая дистрофия, рецидивирующий полихондрит, ретроперитонеальный фиброз, склерит, аутоиммунные заболевания семенной жидкости и яичек, синдром мышечной скованности, подострый бактериальный эндокардит, синдром Сусака, симпатическая офтальмия, артериит Такаясу, тромбоцитопеническая пурпура, синдром Толоса-Ханта, поперечный миелит, недифференцированное заболевание соединительной ткани, болезнь Фогта-Коянаги-Харада.
В другом аспекте согласно изобретению предложено применение конструкций VitoKine для изготовления лекарственного средства для лечения рака.
В другом аспекте согласно изобретению предложено применение конструкций VitoKine для изготовления лекарственного средства для лечения вирусной инфекции.
В другом аспекте согласно изобретению предложено применение конструкций VitoKine для изготовления лекарственного средства для лечения аутоиммунного заболевания.
В другом аспекте согласно изобретению предложено применение конструкций VitoKine для изготовления лекарственного средства для лечения воспаления.
В другом аспекте согласно изобретению предложено применение конструкций VitoKine по изобретению в комбинации со вторым терапевтическим агентом или клеточной терапией, позволяющими лечить рак, вирусную инфекцию, или аутоиммунное заболевание, или воспаление.
В другом аспекте согласно настоящему изобретению предложены выделенные молекулы нуклеиновых кислот, содержащие полинуклеотид, кодирующий конструкцию VitoKine по настоящему изобретению. В другом аспекте согласно настоящему изобретению предложены векторы, содержащие нуклеиновые кислоты, описанные здесь. В различных воплощениях вектор представляет собой вектор экспрессии. В другом аспекте согласно настоящему изобретению предложены выделенные клетки, содержащие нуклеиновые кислоты по изобретению. В различных воплощениях клетка представляет собой клетку-хозяина, содержащую вектор экспрессии по изобретению. В другом аспекте предложены способы получения конструкций VitoKine, включающие культивирование клеток-хозяев в условиях, способствующих экспрессии белков или полипептидов.
В другом аспекте согласно настоящему изобретению предложена фармацевтическая композиция, содержащая выделенные конструкции VitoKine в смеси с фармацевтически приемлемым носителем.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На ФИГ. 1 показаны типичные форматы конструкций VitoKine по настоящему изобретению.
На ФИГ. 2 показан предложенный механизм активации конструкций VitoKine. Типичная конструкция VitoKine содержит два линкера, расщепляемые протеазами; активация протеазой 1 в результате расщепления линкера L1 приводит к образованию активной формы 1; активация протеазой 2 в результате расщепления линкера L2 приводит к образованию активной формы 2; активация обеими протеазами в результате расщепления линкеров L1 и L2 приводит к образованию активной формы 3.
На ФИГ. 3 показан белковый профиль при (A) SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), в отсутствие (NR) и в присутствии (R) восстанавливающего агента, и (В) эксклюзионной хроматографии типичной IL-15 VitoKine Р-0315 после очистки на белке А.
На ФИГ. 4 показаны связывание и функциональная активность IL-15 VitoKine Р-0172 в сравнении с высокоактивным слитым белком IL-15 Р-0198. (А) Связывающая активность в отношении IL-2Rβ, измеренная посредством ELISA-анализа (твердофазный иммуноферментный анализ). (В-С) Индуцирование экспрессии CD69 на человеческих CD8+ Т-клетках (В) и NK-клетках (С) свежих человеческих РВМС (мононуклеарные клетки периферической крови) по результатам FACS-анализа.
На ФИГ. 5 показана функциональная активность мономерной Fc IL-15 VitoKine Р-0170 в сравнении с высокоактивным слитым белком IL-15 Р-0166. Индуцирование экспрессии CD69 на человеческих CD8+ Т-клетках свежих человеческих РВМС измеряли и анализировали посредством FACS (сортировка клеток с активированной флуоресценцией).
На ФИГ. 6 показано индуцирование экспрессии CD69 на (A) CD8+ Т-клетках и (В) NK-клетках (CD56+) человеческих РВМС типичными конструкциями VitoKine (Р-0204, Р-0205 и Р-0206) с различной длиной линкеров в сравнении с высокоактивным Fc-слитым белком IL-15/IL-15Rα Р-0165.
На ФИГ. 7 показана пролиферация NK-клеток (CD56+) человеческих РВМС под действием типичных конструкций VitoKine с различными линкерами L1 и L2 (Р-0202, Р-0203 и Р-0204) в сравнении с полностью активными Fc-слитыми белками IL-15/IL-15Rα Р-0207 и Р-0217.
На ФИГ. 8 показана пролиферация (А) NK-клеток (CD56+) и (В) CD8+ Т-клеток человеческих РВМС под действием типичных конструкций Fc IL-15 VitoKine (Р-0351, Р-0488 и Р-0489) с различным составом последовательности линкера L2, измеренная посредством FACS, в сравнении с Fc-слитым белком IL-15/IL-15Rα Р-0156.
На ФИГ. 9 показан SDS-PAGE-анализ протеолиза Fc IL-15 VitoKine Р-0315 с использованием различных количеств ММР-2.
На ФИГ. 10 показан SDS-PAGE-анализ протеолиза Fc IL-15 VitoKine Р-0203 с использованием uPA в различных условиях для определения подходящих условий взаимодействия для полного расщепления.
На ФИГ. 11 показаны: (А) SDS-PAGE-анализ Fc IL-15 VitoKine Р-0203 до и после протеолиза под действием uPA; (В) белковый профиль активированной VitoKine Р-0203 после расщепления под действием uPA и очистки на белке А для удаления отщепленного Fc-фрагмента.
На ФИГ. 12 показаны: (А) SDS-PAGE-анализ Fc IL-15 VitoKine Р-0315 до и после протеолиза под действием ММР-2, на геле также показан профиль Р-0315, расщепленной под действием ММР-2 и очищенной на белке А; (В) белковый профиль активной формы 2 VitoKine Р-0315, полученной в результате расщепления под действием ММР-2 и последующей очистки на белке А; (С) белковый профиль активной формы 3 VitoKine Р-0315, полученной в результате двойного протеолиза под действием как ММР-2, так и uPA и последующей очистки на белке А в проточном режиме.
На ФИГ. 13 показана оценка активности активированной протеазой (uPA) Fc IL-15 VitoKine P-0203 посредством анализа индуцирования маркера активации CD69 на (A) CD56+ NK-клетках и (В) CD8+ Т-клетках. В качестве положительного контроля был включен Р-0165, высокоактивный слитый белок IL-15.
На ФИГ. 14 показана оценка активности двух активированных протеазой форм Fc IL-15 VitoKine Р-0315 посредством анализа индуцирования маркера активации CD69 на (A) CD56+ NK-клетках и (В) CD8+ Т-клетках. Активная форма 2 Р-0315 была получена в результате расщепления под действием ММР-2, а активная форма 3 Р-0315 была получена в результате двойного протеолиза под действием как ММР-2, так и uPA. В качестве положительного контроля был включен Р-0313, высокоактивный слитый белок IL-15, структурно сходный с активной формой 2 Р-0315.
На ФИГ. 15 показана оценка активности ММР-2-активированной IL-15 VitoKine Р-0315 (активная форма 2) посредством анализа индуцирования маркера пролиферации Ki67 на (A) CD56+ NK-клетках и (В) CD8+ Т-клетках. Для сравнения был включен Р-0351, где оба линкера, L1 и L2, нерасщепляемые, а длина линкера L2 такая же, как в Р-0315.
На ФИГ. 16 показаны дозо- и времязависимые эффекты расщепляемой Fc IL-15 VitoKine Р-0315 и нерасщепляемой Fc IL-15 VitoKine Р-0351 на рост числа CD8+ Т-клеток (А), NK-клеток (В) и лейкоцитов (С) в периферической крови после однократной инъекции мышам Balb/C. Для сравнения был включен полностью активный Fc-слитый IL-15 Р-0313. Сбор крови проводили в -1, 3, 5 и 7 сутки для фенотипирования лимфоцитов посредством FACS-анализа. Данные представлены как среднее плюс/минус SEM. Статистический анализ проводили двусторонним ANOVA с последующим применением апостериорного критерия Тьюки. ****р менее 0,0001, ***р менее 0,001, *р менее 0,05 по сравнению с группой PBS в соответствующей временной точке.
На ФИГ. 17 показано ингибирование легочных метастатических узлов в модели легочного метастазирования СТ26 у мышей через одни сутки после 4-кратного введения Р-0315, Р-0351, Р-0313 или контрольного PBS 1 раз в 5 суток. Первое введение проводили через одни сутки после инъекции клеток СТ26. Если не указано иное, все сравнения против группы PBS; ****р менее 0,0001; **р менее 0,01; *р менее 0,05.
На ФИГ. 18 показаны (А) % CD8+ Т-клеток и (В) % NK-клеток в общих лимфоцитах крови у мышей с метастазами СТ26. Число клеток определяли проточной цитометрией через 4 суток после трех инъекций Р-0315, Р-0351, Р-0313 или контрольного PBS, в/б, 1 раз в 5 суток. Все сравнения против группы PBS; ****р менее 0,0001; **р менее 0,01; *р менее 0,05.
На ФИГ. 19 показана противоопухолевая эффективность Fc IL-15 VitoKine Р-0315 в сравнении с полностью активным Fc-слитым IL-15 Р-0313 в модели развившегося колоректального рака СТ26 у мышей. Кривая роста п/к опухолей СТ26 у отдельных мышей после двух введений Q5D (1 раз в 5 суток) показана для (А) группы наполнителя PBS, (В) группы 0,1 мг/кг Р-0315 или (С) группы 0,1 мг/кг Р-0313. (D) Средний объем опухоли плюс/минус SEM с течением времени для каждой группы введения. Все сравнения против введения наполнителя; n равно 11 на группу; ****Р менее 0,0001.
На ФИГ. 20 показано иммунофармакодинамическое профилирование периферической крови мышей после введения VitoKine Р-0315 или высокоактивного Fc-слитого IL-15 Р-0313 в модели колоректального рака СТ26 у мышей. После двух введений Q5D, начиная с 11 суток после имплантации опухоли, процентное повышение маркера пролиферации Ki67 в (А) NK-клетках и (В) CD8+ Т-клетках в 19 сутки определяли проточной цитометрией. ****р менее 0,0001 против PBS.
На ФИГ. 21 показано иммунофармакодинамическое профилирование периферической крови мышей после введения Р-0315 или Р-0313 в модели колоректального рака СТ26 у мышей. После двух введений, 1 раз в 5 суток, начиная с 11 суток после имплантации опухоли, увеличение числа циркулирующих (на мкл цельной крови) (А) общих лейкоцитов, (В) NK-клеток и (С) CD8+ Т-клеток в 19 сутки определяли проточной цитометрией. ****р менее 0,0001 против PBS.
На ФИГ. 22 показано иммунофармакодинамическое профилирование селезенки после введения Р-0315 или Р-0313 в модели колоректального рака СТ26 у мышей. После двух введений Q5D, начиная с 11 суток после имплантации опухоли, увеличение числа (А) общих лейкоцитов, (В) NK-клеток и (С) CD8+ Т-клеток в селезенке в 19 сутки определяли проточной цитометрией. ****, Р менее 0,0001, *Р менее 0,05, против PBS.
На ФИГ. 23 показано сравнение активности нерасщепляемой Fc IL-15 VitoKine Р-0351 и Benchmark посредством анализа индуцирования маркера пролиферации Ki67 на (A) CD56+ NK-клетках и (В) CD8+ Т-клетках.
На ФИГ. 24 показан белковый профиль при (A) SDS-PAGE, в отсутствие и в присутствии восстанавливающего агента, и (В) эксклюзионной хроматографии типичной IL-2 VitoKine Р-0320 после очистки на белке А.
На ФИГ. 25 показана оценка активности двух Fc IL-2 VitoKine, Р-0320 (IL-2, слитый с С-концом Fc) и Р-0329 (IL-2, слитый с N-концом Fc), посредством анализа уровней pStat5 в субпопуляциях (A) CD4+Foxp3+/CD25high Treg-клеток и (В) CD4+Foxp3-/D25lowCD4 обычных Т-клеток свежих человеческих РВМС. В качестве положительного контроля был включен Р-0250, Fc-слитый белок IL-2 с высокой активностью.
На ФИГ. 26 показан вариант механизма активации VitoKine, отличный от показанного на ФИГ. 2, когда желательны отделение и диффузия D3 от D2 после расщепления протеазой.
На ФИГ. 27 показаны: (А) SDS-PAGE-анализ IL-2 VitoKine Р-0382 и ее активация расщеплением под действием ММР-2 с последующей очисткой на Ni-Excel; (В) белковый профиль ММР-2-активированной Р-0382, очищенной на белке А в режиме связывания и элюирования.
На ФИГ. 28 показана оценка активности активированных протеазой IL-2 VitoKine Р-0382 посредством анализа уровней pStat5 в субпопуляциях (А) CD4+Foxp3+/CD25high Treg-клеток и (В) CD4+Foxp3-/D25lowCD4 обычных Т-клеток (Tconv) свежих человеческих РВМС.Два активированных образца очищали либо на смоле Ni-Excel с удалением протеазы (activ. 1), либо на белке А с удалением как протеазы, так и домена IL-2RαSushi, отщепленного в результате протеолиза (activ. 2). В качестве положительного контроля был включен Р-0250, Fc-слитый белок IL-2 с высокой активностью.
На ФИГ. 29 показана оценка активности IL-2 VitoKine Р-0398 до и после протеолиза под действием ММР-2 посредством анализа уровней pStat5 в субпопуляциях (A) CD4+Foxp3+/CD25high Treg-клеток и (В) CD4+Foxp3-/D25lowCD4 Tconv-клеток свежих человеческих РВМС. Для сравнения были включены Р-0382, отличающаяся от Р-0398 только длиной линкера L2, и Р-0250, Fc-слитый белок IL-2 с высокой активностью.
На ФИГ. 30 показана оценка активности Fc IL-15 VitoKine Р-0315 в сравнении с антитело IL-15 VitoKine Р-0485 посредством анализа индуцирования маркера пролиферации Ki67 на (A) CD56+ NK-клетках и (В) CD8+ Т-клетках, определенной проточной цитометрией.
На ФИГ. 31 показано связывание блокирующих пептидов (L01, L02, L03, L04 и L05) с IL-15 в формате ELISA.
На ФИГ. 32 показано связывание слитых белков IL-15 (Р-0153, Р-0159, Р-0160 и Р-0161), содержащих блокирующий пептид на основе IL-2Rβ, с IL-2Rβ, сорбированным на планшет.
На ФИГ. 33 показана эксклюзионная хроматограмма четырех IL-2 VitoKine (Р-0320, Р-0382, Р-0362 и Р-0379) (B-F) и одного аналога Fc-слитого белка Р-0250 с одиночной аминокислотной заменой S125I в IL-2 в сравнении с хроматограммой Fc-слитого белка IL-2 Р-0250 (А).
На ФИГ. 34 показан SDS-PAGE-гель Fc IL-15 VitoKine Р-0389 (А) в сравнении с Р-0315 (В).
ВАРИАНТ(Ы) ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Согласно настоящему изобретению предложены новые конструкции «VitoKine» в качестве технологической платформы для снижения системной токсичности, обусловленной связыванием с мишенью, и повышения терапевтического индекса цитокинов, предназначенных для применения в лечении рака, вирусной инфекции, аутоиммунных заболеваний или воспалительных заболеваний. Со ссылкой на ФИГ. 1, платформа VitoKine определена конструкциями, показанными на ФИГ. 1, и предложенными способами активации, показанными на ФИГ. 2. Со ссылкой на ФИГ. 1, новые конструкции VitoKine по настоящему изобретению содержат 3 домена: (1) домен D1 («D1»), выбранный из группы, состоящей из тканенаправленного домена, домена, увеличивающего период полувыведения, или домена группировки с двойной функцией; (2) домен D2 («D2»), представляющий собой «домен активной группировки»; и (3) домен D3 («D3»), представляющий собой «домен маскирующей группировки». Важно, что домен D3 способен маскировать или снижать функциональную активность D2 до активации в намеченном месте терапии.
Три домена соединены с использованием ликеров, имеющих вариабельные длину и ригидность, в сочетании с последовательностями, расщепляемыми протеазами, представляющими собой пептидные субстраты протеаз определенных подтипов, экспрессия которых в очагах заболевания повышена или неправильно отрегулирована, что позволяет раскрывать или высвобождать функциональный домен D2 в очаге заболевания. Длина и состав линкера были оптимизированы для наилучшей маскировки доступности домена D2 для его рецепторов для снижения его системного действия, сохраняя в то же время стабильность конструкций VitoKine в циркулирующей крови и обеспечивая возможность их эффективного расщепления после контакта с определенными протеазами в намеченном очаге заболевания. При разработке «VitoKine» также учитывались рациональные соображения, основанные на имеющихся данных о молекулярном взаимодействии цитокинов с их когнатными рецепторами. Рецепторы цитокинов обычно функционируют как олигомерные комплексы, состоящие из двух-четырех рецепторных субъединиц. Разные субъединицы выполняют специализированные функции, такие как связывание с лигандом или трансдукция сигнала. Альфа-субъединица рецепторов цитокинов представляет собой связывающий рецептор, придающий специфичность в отношении лиганда, усиливающий взаимодействие лиганда с сигнальными рецепторами и превращающий низкоаффинные сигнальные рецепторы в высокоаффинные. Таким образом, домен D3 VitoKine предпочтительно представляет собой когнатный связывающий рецептор домена D2. После расщепления возможна диссоциация или повторная ассоциация домена D3 с D2 с полным восстановлением связывающей и сигнальной активности домена D2 на месте. Таким образом, домен D3 может играть двойную роль в регуляции функции домена D2. Он сохраняет инертность домена D2, когда VitoKine инактивирована, и может участвовать в функционировании D2 после расщепления и активации VitoKine. Тем не менее, домен D3 может представлять собой любой белок, пептид, антитело, фрагмент антитела, или полимер, или нуклеотиды, способные маскировать активность D2.
В другом аспекте добавление домена D3 может также существенно расширять возможности разработки конструкции VitoKine с большим выходом экспрессии и меньшей склонностью к агрегации.
Домен D1 может представлять собой домен, увеличивающий период полувыведения, продлевая период полувыведения VitoKine из циркуляции, помимо того, что он служит дополнительным доменом, маскирующим функциональную активность домена D2. Домен D1 также может представлять собой направленный на болезнь или ткань мотив, направляющий VitoKine специфически к месту, представляющему интерес, и ограничивающий местную активацию VitoKine, дополнительно улучшая терапевтический индекс. Поэтому платформа «VitoKine» обеспечивает возможность селективной активации цитокинов в намеченном месте и обладает преимуществами снижения системной токсичности с одновременным увеличением терапевтического эффекта в очагах заболевания, улучшая таким образом их терапевтический индекс.
Домен D2 конструкции VitoKine представляет собой активную группировку, но остается инертным до местной активации протеазами, регуляция которых повышена в тканях, пораженных заболеванием; это будет ограничивать связывание активной группировки с ее рецепторами на периферии или на поверхности клеток или тканей, не пораженных заболеванием, предотвращая избыточную активацию ее пути и уменьшая нежелательную токсичность, обусловленную связыванием с мишенью вне целевой ткани. Кроме того, инертность активной группировки VitoKine до активации протеазами будет существенно снижать возможный клиренс, обусловленный связыванием с антигеном, и, таким образом, увеличивать период полувыведения in vivo и улучшать биораспределение, биодоступность и эффективность в намеченных местах терапии. Кроме того, исходя из настоящего изобретения, платформа VitoKine может расширять возможности разработки белков, включая, без ограничения, повышение уровня экспрессии и уменьшение склонности к агрегации, например при использовании когнатного рецептора альфа в качестве домена D3.
Несмотря на то что расщепляемые линкеры предпочтительны для большинства конструкций VitoKine, ограничивая их системную активацию и высвобождая активный домен в намеченном месте после введения, нерасщепляемые линкеры могут быть желательны для обеспечения продолжительного системного действия фармакологически активной конструкции VitoKine и повышения терапевтической эффективности.
В типичных воплощениях конструкции VitoKine содержат активную группировку (D2) на основе IL-15, варианта IL-15, IL-2 или варианта IL-2. В случае этих конструкций VitoKine на основе IL-15 и/или IL-2, уникальную и несигнальную α-субъединицу рецепторов каждого цитокина используют в качестве субъединицы домена маскирующей группировки (D3) через линкер, расщепляемый протеазой, для обратимой маскировки активности цитокина. В зависимости от избирательных свойств каждого рецепторного комплекса и различных требований при разных заболеваниях, являющихся предполагаемыми показаниями для лечения молекулами VitoKine, может быть предпочтительным, чтобы после расщепления линкера протеазой маскирующая α-субъединица образовывала комплекс с активированным цитокином посредством нековалентных связей (например, в случае IL-15) или диссоциировала от него (например, в случае IL-2 для селективного увеличения числа Treg-клеток). В результате может возникнуть необходимость в аминокислотных модификациях α-рецептора для модуляции аффинности связывания с его когнатным цитокином.
Эта концепция сочетания когнатного рецептора, белка, антитела, фрагмента антитела, связывающего пептида с цитокином через активируемый линкер для маскировки его функциональной активности до активации в намеченных местах терапии может быть адаптирована для различных цитокинов, включая, без ограничения, IL-4, IL-7, IL-9, IL-10, IL-12, IL-22, IL-23 и TGFβ, хемокинов, таких как CXCR3, или различных факторов роста, таких как семейство TNF, TGFα и TGFβ, и гормонов. Такая же концепция может также быть применена к другим белкам для получения пробелков для достижения усиленного направленного действия на очаг заболевания и расширения возможностей их терапевтического применения.
Определения
Термины «полипептид», «пептид» и «белок» использованы здесь взаимозаменяемо для обозначения полимера из аминокислотных остатков. В различных воплощениях «пептиды», «полипептиды» и «белки» представляют собой цепи из аминокислот, альфа-углероды которых связаны пептидными связями. Таким образом, концевая аминокислота на одном конце цепи (N-концевая) имеет свободную аминогруппу, в то время как концевая аминокислота на другом конце цепи (С-концевая) имеет свободную карбоксильную группу. При использовании здесь термин «N-конец» относится к свободной α-аминогруппе аминокислоты на N-конце пептида или к α-аминогруппе (аминогруппе при ее участии в пептидной связи) аминокислоты в любом другом месте в пределах рассматриваемого пептида. Сходным образом, термин «С-конец» относится к свободной карбоксильной группе на С-конце пептида или карбоксильной группе аминокислоты в любом другом месте в пределах пептида. Пептиды также включают практически любую полиаминокислоту, включая, без ограничения, миметики пептидов, такие как аминокислоты, соединенные эфирными, а не амидными связями.
Полипептиды по изобретению включают полипептиды, которые были модифицированы любым образом и по любой причине, например, для: (1) снижения чувствительности к протеолизу; (2) снижения чувствительности к окислению; (3) изменения аффинности связывания для образования белковых комплексов; (4) изменения аффинности связывания; и (5) придания или модификации других физико-химических или функциональных свойств.
При использовании здесь «аминокислотная замена» относится к замене одной аминокислоты полипептида в определенном положении исходной полипептидной последовательности на другую аминокислоту. Аминокислотные замены могут быть проведены с применением генетических или химических методов, хорошо известных в данной области. Например, в последовательности, встречающейся в природе (например, в части полипептида вне домена(ов), образующего(их) межмолекулярные контакты), могут быть проведены одна аминокислотная замена или множество аминокислотных замен (например, консервативных аминокислотных замен). «Консервативная аминокислотная замена» относится к замене аминокислоты полипептида на функционально сходную аминокислоту. Каждая из следующих шести групп включает аминокислоты, являющиеся консервативными заменами друг друга:
1) аланин (А), серии (S) и треонин (Т);
2) аспарагиновая кислота (D) и глутаминовая кислота (Е);
3) аспарагин (N) и глутамин (Q);
4) аргинин (R) и лизин (K);
5) изолейцин (I), лейцин (L), метионин (М) и валин (V);
6) фенилаланин (F), тирозин (Y) и триптофан (W).
«Неконсервативная аминокислотная замена» относится к замене представителя одного из этих классов на представителя другого класса. При проведении таких изменений, согласно различным воплощениям, можно учитывать индекс гидрофобности аминокислот. Каждой аминокислоте индекс гидрофобности был присвоен, исходя из ее гидрофобности и зарядных характеристик. Он составляет: изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серии (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин (-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9); и аргинин (-4,5).
Важность индекса гидрофобности аминокислот для придания белку функции биологического взаимодействия известна в данной области (см., например, Kyte etal, 1982, J. Mol. Biol. 157:105-131). Известно, что определенные аминокислоты могут быть заменены на другие аминокислоты, имеющие сходный индекс или показательгидрофобности, с сохранением сходной биологической активности. В различных воплощениях изменения, основанные на индексе гидрофобности, включают замены аминокислот, индексы гидрофобности которых различаются не более чем на 2. В различных воплощениях включены замены аминокислот, индексы гидрофобности которых различаются не более чем на 1, и, в различных воплощениях, замены аминокислот, индексы гидрофобности которых различаются не более чем на 0,5.
В данной области также известно, что замена аналогичных аминокислот может быть эффективно проведена, исходя из гидрофильности, особенно там, где полученный таким образом биологически функциональный белок или пептид предназначен для использования в иммунологических воплощениях, как раскрыто здесь. В различных воплощениях наибольшая местная средняя гидрофильность белка, определяемая гидрофильностью его смежных аминокислот, коррелирует с его иммуногенностью и антигенностью, то есть с биологическими свойствами белка.
Указанным аминокислотным остаткам были присвоены следующие значения гидрофильности: аргинин (+3,0); лизин (+3,0); аспартат (+3,0 плюс/минус 0,1); глутамат (+3,0 плюс/минус 0,1); серии (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5 плюс/минус 0,1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5); и триптофан (-3,4). В различных воплощениях изменения, основанные на сходных значениях гидрофильности, включают замены аминокислот, значения гидрофильности которых различаются не более чем на 2, в различных воплощениях включены замены аминокислот, значения гидрофильности которых различаются не более чем на 1, и, в различных воплощениях, замены аминокислот, значения гидрофильности которых различаются не более чем на 0,5.
Примеры аминокислотных замен приведены в Таблице 1.

Специалист сможет определить подходящие варианты полипептидов, как изложено здесь, с использованием хорошо известных методик. В различных воплощениях специалист в данной области сможет определить подходящие области молекулы, которые могут быть изменены без нарушения активности посредством направленного изменения областей, которые, как предполагается, не важны для активности. В других воплощениях специалист сможет определить остатки и части молекул, консервативные у сходных полипептидов. В других воплощениях даже те области, которые могут быть важны для биологической активности или для структуры, могут быть подвержены консервативным аминокислотным заменам без нарушения биологической активности или без нежелательного влияния на структуру полипептида.
Кроме того, специалист в данной области сможет ознакомиться со структурно-функциональными исследованиями, в которых определены остатки сходных полипептидов, важные для их активности или структуры. Исходя из такого сравнения, специалист сможет спрогнозировать важность аминокислотных остатков полипептида, соответствующих аминокислотным остаткам, важным для активности или структуры сходных полипептидов. Специалист в данной области может предпочесть замены таких спрогнозированных важных аминокислотных остатков на химически сходные аминокислотные остатки.
Специалист в данной области сможет также проанализировать трехмерную структуру и аминокислотную последовательность в сравнении с такой структурой сходных полипептидов. Исходя из такой информации, специалист в данной области сможет спрогнозировать выравнивание аминокислотных остатков полипептида в связи с его трехмерной структурой. В различных воплощениях специалист в данной области может отказаться от радикальных изменений аминокислотных остатков, предположительно расположенных на поверхности полипептида, поскольку такие остатки могут быть вовлечены в важные взаимодействия с другими молекулами. Более того, специалист в данной области может получить тестовые варианты, содержащие одиночную аминокислотную замену каждого желаемого аминокислотного остатка. Затем может быть проведен скрининг этих вариантов с применением анализов активности, известных специалистам в данной области. Такие варианты могут быть использованы для получения информации о подходящих вариантах. Например, при обнаружении того, что изменение определенного аминокислотного остатка привело к нарушению, нежелательному снижению активности или к неподходящей активности, можно избегать вариантов с таким изменением. Иными словами, исходя из информации, полученной в таких рутинных экспериментах, специалист в данной области сможет легко определить аминокислоты, где дальнейших замен, самих по себе или в комбинации с другими мутациями, следует избегать.
При использовании здесь термины «фрагмент полипептида» и «усеченный полипептид» относятся к полипептиду, имеющему N-концевую и/или С-концевую делецию по сравнению с соответствующим полноразмерным белком. В различных воплощениях длина фрагментов может составлять, например, по меньшей мере 5, по меньшей мере 10, по меньшей мере 25, по меньшей мере 50, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 300, по меньшей мере 350, по меньшей мере 400, по меньшей мере 450, по меньшей мере 500, по меньшей мере 600, по меньшей мере 700, по меньшей мере 800, по меньшей мере 900 или по меньшей мере 1000 аминокислот. В различных воплощениях длина фрагментов может также составлять, например, не более 1000, не более 900, не более 800, не более 700, не более 600, не более 500, не более 450, не более 400, не более 350, не более 300, не более 250, не более 200, не более 150, не более 100, не более 50, не более 25, не более 10 или не более 5 аминокислот. Фрагмент может дополнительно содержать, на одном или обоих его концах, одну или более чем одну дополнительную аминокислоту, например последовательность аминокислот из другого встречающегося в природе белка (например, Fc-домен или домен с лейциновой молнией) или искусственную аминокислотную последовательность (например, последовательность искусственного линкера).
При использовании здесь термины «вариант полипептида», «гибридный полипептид» и «мутант полипептида» относятся к полипептиду, содержащему аминокислотную последовательность с вставкой, делецией и/или заменой одного или более чем одного аминокислотного остатка аминокислотной последовательности относительно другой последовательности полипептида. В различных воплощениях число аминокислотных остатков, подлежащих вставке, делеции или замене, может составлять, например, по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 10, по меньшей мере 25, по меньшей мере 50, по меньшей мере 75, по меньшей мере 100, по меньшей мере 125, по меньшей мере 150, по меньшей мере 175, по меньшей мере 200, по меньшей мере 225, по меньшей мере 250, по меньшей мере 275, по меньшей мере 300, по меньшей мере 350, по меньшей мере 400, по меньшей мере 450 или по меньшей мере 500 аминокислот в длину. Гибриды по настоящему изобретению включают слитые белки.
«Производное» полипептида представляет собой полипептид, который был химически модифицирован, например посредством конъюгирования с другой химической группировкой, такой как, например, полиэтиленгликоль, альбумин (например, человеческий сывороточный альбумин), фосфорилирования и гликозилирования.
Термин «% идентичности последовательностей» использован здесь взаимозаменяемо с термином «% идентичности» и относится к уровню идентичности аминокислотных последовательностей между двумя или более пептидными последовательностями или уровню идентичности нуклеотидных последовательностей между двумя или более нуклеотидными последовательностями при выравнивании с использованием программы для выравнивания последовательностей. Например, при использовании здесь 80%-я идентичность обозначает то же, что 80%-я идентичность последовательностей, определенная заданным алгоритмом, и означает, что заданная последовательность по меньшей мере на 80% идентична другому участку другой последовательности. В различных воплощениях % идентичности выбран из, например, по меньшей мере 60%-й, по меньшей мере 65%-й, по меньшей мере 70%-й, по меньшей мере 75%-й, по меньшей мере 80%-й, по меньшей мере 85%-й, по меньшей мере 90%-й, по меньшей мере 95%-й, или по меньшей мере 99%-й, или большей идентичности последовательности заданной последовательности. В различных воплощениях % идентичности входит в диапазон, например, от приблизительно 60% до приблизительно 70%, от приблизительно 70% до приблизительно 80%, от приблизительно 80% до приблизительно 85%, от приблизительно 85% до приблизительно 90%, от приблизительно 90% до приблизительно 95% или от приблизительно 95% до приблизительно 99%.
Термин «% гомологии последовательностей» использован здесь взаимозаменяемо с термином «% гомологии» и относится к уровню гомологии аминокислотных последовательностей между двумя или более пептидными последовательностями или уровню гомологии нуклеотидных последовательностей между двумя или более нуклеотидными последовательностями при выравнивании с использованием программы для выравнивания последовательностей. Например, при использовании здесь 80%-я гомология обозначает то же, что 80%-я гомология последовательностей, определенная заданным алгоритмом, и, соответственно, гомолог заданной последовательности имеет более чем 80%-ю гомологию последовательности на участке заданной последовательности. В различных воплощениях % гомологии выбран из, например, по меньшей мере 60%-й, по меньшей мере 65%-й, по меньшей мере 70%-й, по меньшей мере 75%-й, по меньшей мере 80%-й, по меньшей мере 85%-й, по меньшей мере 90%-й, по меньшей мере 95%-й, или по меньшей мере 99%-й, или большей гомологии последовательности заданной последовательности. В различных воплощениях % гомологии входит в диапазон, например, от приблизительно 60% до приблизительно 70%, от приблизительно 70% до приблизительно 80%, от приблизительно 80% до приблизительно 85%, от приблизительно 85% до приблизительно 90%, от приблизительно 90% до приблизительно 95% или от приблизительно 95% до приблизительно 99%.
Типичные компьютерные программы, которые могут быть использованы для определения идентичности двух последовательностей, включают, без ограничения, набор программ BLAST, например, BLASTN, BLASTX, TBLASTX, BLASTP и TBLASTN, общедоступный в интернете на сайте NCBI. См. также Altschul et at, J. Mol. Biol. 215:403-10, 1990 (с особой ссылкой на опубликованные настройки по умолчанию, то есть параметры w, равный 4, и t, равный 17) и Altschul et at, Nucleic Acids Res., 25:3389-3402, 1997. При оценке заданной аминокислотной последовательности в сравнении с аминокислотными последовательностями из GenBank Protein Sequences и других общедоступных баз данных поиск последовательностей обычно проводят с использованием программы BLASTP. Программа BLASTX предпочтительна для поиска последовательностей нуклеиновых кислот, транслированных во всех рамках считывания, против аминокислотных последовательностей из GenBank Protein Sequences и других общедоступных баз данных. Как BLASTP, так и BLASTX запускают с использованием штрафа за открытие разрыва 11,0 и штрафа за продолжение разрыва 1,0, являющихся параметрами по умолчанию, и матрицы BLOSUM-62.
Помимо расчета процента идентичности алгоритм BLAST также позволяет проводить статистический анализ сходства двух последовательностей (см., например, Karlin & Altschul, Proc. Nat'l. Acad. Sci. USA, 90:5873-5787, 1993). Одной мерой сходства при применении алгоритма BLAST является наименьшая суммарная вероятность (P(N)), которая предоставляет собой показатель вероятности случайного совпадения между двумя нуклеотидными или аминокислотными последовательностями. Например, нуклеиновую кислоту считают сходной с эталонной последовательностью, если наименьшая суммарная вероятность при сравнении анализируемой нуклеиновой кислоты с эталонной нуклеиновой кислотой составляет, например, менее чем приблизительно 0,1, менее чем приблизительно 0,01 или менее чем приблизительно 0,001.
При использовании здесь термин «модификация» относится к любой манипуляции с остовом пептида (например, аминокислотной последовательностью) или к посттрансляционным модификациям (например, гликозилированию) полипептида.
При использовании здесь термин «модификация типа «выступ во впадину» («knob-into-hole»)» относится к модификации обращенных друг к другу поверхностей двух тяжелых цепей иммуноглобулина в домене СН3. В одном воплощении «модификация типа «выступ во впадину»» включает аминокислотную замену T366W и, возможно, аминокислотную замену S354C в одной тяжелой цепи антитела и аминокислотные замены T366S, L368A, Y407V и, возможно, Y349C в другой тяжелой цепи антитела. Технология «выступ во впадину» описана, например, в патенте США №5,731,168; патенте США №7,695,936; Ridgway et al, Prot Eng 9, 617-621 (1996); и Carter, J Immunol Meth 248, 7-15 (2001).
При использовании здесь термин «биологически активируемое лекарственное средство» или «VitoKine» обозначает соединение, являющееся предшественником лекарственного средства, который, после его введения субъекту, высвобождает лекарственное средство in vivo посредством некоторого химического или физиологического процесса таким образом, что биологически активируемое лекарственное средство превращается в продукт, активный в целевых тканях. Биологически активируемое лекарственное средство представляет собой любое соединение, проходящее биологическую активацию до оказания его фармакологических эффектов. Таким образом, биологически активируемые лекарственные средства можно рассматривать как лекарственные средства, содержащие специализированные нетоксичные защитные группы, используемые временно для изменения или устранения нежелательных свойств исходной молекулы.
При использовании здесь термин «слитый белок» относится к слитой полипептидной молекуле, содержащей два или более генов, исходно кодирующих раздельные белки, где компоненты слитого белка соединены друг с другом пептидными связями, непосредственно или через пептидные линкеры. При использовании здесь термин «слитый» относится к компонентам, соединенным пептидными связями, непосредственно или через один или более чем один пептидный линкер.
«Линкер» относится к молекуле, соединяющей две другие молекулы, ковалентно или через ионные, ван-дер-Ваальсовы или водородные связи, например, к молекуле нуклеиновой кислоты, гибридизующейся с одной комплементарной последовательностью на 5'-конце и с другой комплементарной последовательностью на 3'-конце, соединяя таким образом две некомплементарные последовательности. «Расщепляемый линкер» относится к линкеру, который может быть разрушен, расщеплен или иным образом разъединен для разделения двух компонентов, соединенных этим расщепляемым линкером. Расщепляемые линкеры в большинстве случаев расщепляются ферментами, обычно пептидазами, протеазами, нуклеазами, липазами и тому подобными. Также возможно расщепление расщепляемых линкеров под действием стимулов окружающей среды, таких как, например, изменения температуры, рН, концентрации солей и так далее.
При использовании здесь термин «пептидный линкер» относится к пептиду, содержащему одну или более чем одну аминокислоту, обычно приблизительно 1-30 аминокислот. Пептидные линкеры представляют собой линкеры, известные в данной области или описанные здесь. Подходящие неиммуногенные линкерные пептиды включают, например, пептидные линкеры (G4S)n, (SG4)n или G4(SG4)n. В большинстве случаев «п» представляет собой число от 1 до 10, обычно от 2 до 4.
«Фармацевтическая композиция» относится к композиции, подходящей для фармацевтического применения у животного. Фармацевтическая композиция содержит фармакологически эффективное количество активного агента и фармацевтически приемлемый носитель. «Фармакологически эффективное количество» относится к такому количеству агента, которое эффективно для получения намеченного фармакологического результата. «Фармацевтически приемлемый носитель» относится к любому из стандартных фармацевтических носителей, наполнителей, буферов и эксципиентов, таких как забуференный фосфатом физиологический раствор, 5%-й водный раствор декстрозы, эмульсии, такие как масляная/водная или водная/масляная эмульсия, и различные типы смачивающих агентов и/или адъювантов. Подходящие фармацевтические носители и композиции описаны в Remington's Pharmaceutical Sciences, 21st Ed. 2005, Mack Publishing Co, Easton. «Фармацевтически приемлемая соль» представляет собой соль, которая может быть включена в соединение для фармацевтического применения, включая, например, соли металлов (натрия, калия, магния, кальция и так далее) и соли аммиака или органических аминов.
При использовании здесь «лечение» (и его грамматические варианты, такие как «лечить») относится к клиническому вмешательству в попытке изменить естественный ход заболевания у индивида, проходящего лечение, и может быть проведено для профилактики или в процессе развития клинической патологии. Желаемые эффекты лечения включают, без ограничения, предотвращение развития или рецидива заболевания, облегчение симптомов, уменьшение любых прямых или непрямых патологических последствий заболевания, предотвращение метастазирования, снижение скорости прогрессирования заболевания, уменьшение степени выраженности или временное облегчение болезненного состояния и ремиссию или улучшение прогноза. При использовании здесь «облегчение» заболевания, расстройства или состояния обозначает снижение степени тяжести и/или частоты возникновения симптомов заболевания, расстройства или состояния. Кроме того, ссылки на «лечение» включают здесь ссылки на радикальное, паллиативное и профилактическое лечение.
При использовании здесь термин «эффективное количество» или «терапевтически эффективное количество» относится к количеству соединения или композиции, достаточному для лечения указанного расстройства, состояния или заболевания, такому как уменьшение степени выраженности, временное облегчение, уменьшение и/или задержка одного или более чем одного из его симптомов. Применительно к раку или другой нежелательной пролиферации клеток эффективное количество включает количество, достаточное для: (1) уменьшения числа раковых клеток; (2) уменьшения размера опухоли; (3) ингибирования, торможения, некоторого замедления и, предпочтительно, остановки инфильтрации периферических органов раковыми клетками; (4) ингибирования, (то есть некоторого замедления и, предпочтительно, остановки) метастазирования опухоли; (5) ингибирования роста опухоли; (6) предотвращения или отсрочки развития и/или рецидива опухоли; и/или (7) некоторого облегчения одного или более чем одного из симптомов, связанных с раком. Эффективное количество может быть введено за одно или более чем одно введение.
Фраза «введение» или «действия, приводящие к введению» относится к действиям, предпринимаемым медицинским работником (например, врачом) или лицом, контролирующим оказание медицинской помощи пациенту, которое контролирует и/или разрешает вводить агент(ы)/соединение(я) пациенту. Действия, приводящие к введению, могут включать диагностику, и/или определение подходящей терапевтической схемы, и/или назначение определенного агента(ов)/соединений пациенту. Такое назначение может включать, например, заполнение рецептурного бланка, запись в истории болезни и тому подобное. При описании введения здесь также подразумевают «действия, приводящие к введению».
Термины «пациент», «индивид» и «субъект» могут быть использованы взаимозаменяемо и относятся к млекопитающему, предпочтительно человеку или примату, не являющемуся человеком, но также к одомашненным млекопитающим (например, кошкам или собакам), лабораторным млекопитающим (например, мышам, крысам, кроликам, хомякам, морским свинкам) и сельскохозяйственным млекопитающим (например, лошадям, быкам, свиньям, овцам). В различных воплощениях пациент может представлять собой человека (например, взрослого мужчину, взрослую женщину, подростка мужского пола, подростка женского пола, ребенка мужского пола, ребенка женского пола), которому врач или другой медицинский работник оказывает помощь в стационаре, в учреждении для оказания психиатрической помощи, амбулаторно или в ином клиническом контексте. В различных воплощениях пациент может представлять собой пациента со сниженным иммунитетом или пациента с ослабленной иммунной системой, включая, без ограничения, пациентов с первичным иммунодефицитом, СПИД, онкологических пациентов и пациентов после трансплантации, принимающих определенные иммуносупрессивные лекарственные средства, и пациентов с наследственными заболеваниями, влияющими на иммунную систему (например, врожденной агаммаглобулинемией, врожденным дефицитом IgA). В различных воплощениях пациент представляет собой пациента с иммуногенным раком, включая, без ограничения, рак мочевого пузыря, рак легкого, меланому и другие виды рака, при которых, согласно сообщениям, повышена частота мутаций (Lawrence et al., Nature, 499(7457): 214-218, 2013).
Термин «иммунотерапия» относится к лечению рака, включающему, без ограничения: лечение с использованием истощающих антител к определенным опухолевым антигенам; лечение с использованием конъюгатов «антитело лекарственное средство»; лечение с использованием агонистических, антагонистических или блокирующих антител к костимулирующим или коингибирующим молекулам (иммунным контрольным точкам), таким как CTLA-4, PD-1, PDL-1, CD40, ОХ-40, CD137, GITR, LAG3, TIM-3, SIRPa, CD47, GITR, ICOS, CD27, Siglec 7, Siglec 8, Siglec 9, Siglec 15 и VISTA, CD276, CD272, TIM-3, B7-H4; лечение с использованием биспецифичных антител, привлекающих Т-клетки (BiTE®), таких как блинатумомаб; лечение, включающее введение модификаторов биологического ответа, таких как IL-2, IL-4, IL-7, IL-10, IL-12, IL-15, IL-21, IL-22, GM-CSF, IFN-α, IFN-βи IFN-γ, антагонист TGF-βили ловушка TGF-β; лечение с использованием терапевтических вакцин, таких как сипулейцел-Т; лечение с использованием терапевтических вирусов, включая, без ограничения, онколитические вирусы, такие как T-vec; лечение с использованием дендритноклеточных вакцин или вакцин на основе опухолевых антигенных пептидов или неоантигенов; лечение с использованием NK-клеток; лечение с использованием Т-клеток с химерными антигенными рецепторами (CAR-T-клеток); лечение с использованием CAR-NK-клеток; лечение с использованием DC (дендритных клеток) или Т-клеток; лечение с использованием iPS-индуцированных NK-клеток; лечение с использованием iPS-индуцированных Т-клеток; лечение с использованием вакцины, такой как бацилла Кальметта-Герена (BCG); лечение с использованием опухоль-инфильтрирующих лимфоцитов (TIL); лечение с использованием адоптивно перенесенных противоопухолевых Т-клеток (культивированных ex vivo и/или TCR-T-клеток); лечение с использованием клеток TALL-104; и лечение с использованием иммуностимулирующих агентов, таких как агонисты Toll-подобных рецепторов (TLR) CpG, TLR7, TLR8, TLR9 и имихимод.
«Резистентный или рефрактерный рак» относится к опухолевым клеткам или раку, не отвечающим на предшествующую противораковую терапию, включая, например, химиотерапию, хирургические вмешательства, лучевую терапию, трансплантацию стволовых клеток и иммунотерапию. Опухолевые клетки могут быть резистентными или рефрактерными в начале лечения, или они могут становиться резистентными или рефрактерными во время лечения. Рефрактерные опухолевые клетки включают опухоли, не отвечающие на момент начала лечения или изначально отвечающие на протяжении короткого периода, но в конечном счете не отвечающие на лечение. Рефрактерные опухолевые клетки также включают опухоли, отвечающие на проведение противораковой терапии, но не отвечающие на последующие раунды терапии. Для целей данного изобретения рефрактерные опухолевые клетки также охватывают опухоли, подавляемые проведением противораковой терапии, но рецидивирующие в течение пяти лет, иногда в течение десяти лет или более после прекращения терапии. При противораковой терапии возможно применение химиотерапевтических агентов самих по себе, облучения самого по себе, направленной терапии самой по себе, хирургического вмешательства самого по себе или их комбинаций. Для простоты описания и не для ограничения следует понимать, что «рефрактерные опухолевые клетки» взаимозаменяемы с «резистентной опухолью».
Термин «опухоль-ассоциированный антиген» (ТАА) относится, например, к антигенам клеточной поверхности, селективно экспрессированным раковыми клетками или сверхэкспрессированным у раковых клеток по сравнению с большинством нормальных клеток. При использовании здесь термины «вариант ТАА» и «мутант ТАА» относятся к ТАА, содержащему аминокислотную последовательность с вставкой, делецией и/или заменой одного или более чем одного аминокислотного остатка аминокислотной последовательности относительно другой последовательности ТАА. В различных воплощениях число аминокислотных остатков, подлежащих вставке, делеции или замене, может составлять, например, по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 10, по меньшей мере 25, по меньшей мере 50, по меньшей мере 75, по меньшей мере 100, по меньшей мере 125, по меньшей мере 150, по меньшей мере 175, по меньшей мере 200, по меньшей мере 225, по меньшей мере 250, по меньшей мере 275, по меньшей мере 300, по меньшей мере 350, по меньшей мере 400, по меньшей мере 450 или по меньшей мере 500 аминокислот в длину.
Термин «неоантиген» относится, например, к антигенам клеточной поверхности, не известным ранее иммунной системе, особенно к антигенам, возникшим при изменении антигенов хозяина в результате облучения, химиотерапии, вирусной инфекции, опухолевой трансформации/мутации, метаболизма лекарственного средства и так далее, селективно экспрессированным раковыми клетками или сверхэкспрессированным у раковых клеток по сравнению с большинством нормальных клеток.
При использовании здесь термин «антитело» использован в наиболее широком смысле и охватывает различные структуры антител (IgG1, 2, 3 или 4, IgM, IgA, IgE), включая, без ограничения, моноклональные антитела, поликлональные антитела, мультиспецифичные антитела (например, биспецифичные или бифункциональные антитела) и фрагменты антител при условии, что они демонстрируют желаемую антигенсвязывающую активность.
При использовании здесь термин «фрагмент антитела» относится к молекуле, отличной от интактного антитела, содержащей часть интактного антитела, связывающуюся с антигеном, с которым связывается интактное антитело. Примеры фрагментов антител включают, без ограничения, Fv, Fab, Fab', Fab' SH, F(ab')2, диатела, линейные антитела, одноцепочечные молекулы антител (например, scFv) и однодоменные антитела.
При использовании здесь термин «Fab фрагмент» относится к фрагменту иммуноглобулина, содержащему домен VL и константный домен (CL) легкой цепи с доменом VH и первым константным доменом (СН1) тяжелой цепи.
При использовании здесь термины «вариабельная область» или «вариабельный домен» относятся к домену тяжелой или легкой цепи иммуноглобулина или антитела, который обычно вовлечен в связывание иммуноглобулина или антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) иммуноглобулина или антитела обычно имеют сходную структуру, при этом каждый домен содержит четыре консервативные каркасные области (FR) и три участка, определяющих комплементарность (CDR).
При использовании здесь «человеческий иммуноглобулин» представляет собой иммуноглобулин, обладающий аминокислотной последовательностью, соответствующей аминокислотной последовательности иммуноглобулина, продуцированного человеком или клеткой человека или имеющего происхождение из источника, не являющегося человеком, в котором использованы репертуары человеческих иммуноглобулинов или другие последовательности, кодирующие человеческие иммуноглобулины. Это определение человеческого иммуноглобулина прямо исключает гуманизированный иммуноглобулин, содержащий антигенсвязывающие остатки, не являющиеся человеческими.
При использовании здесь термин «Fc домен» или «Fc-область» использован для определения С-концевой области тяжелой цепи иммуноглобулина, содержащей по меньшей мере часть константной области. Данный термин включает Fc-области с нативной последовательностью и варианты Fc-областей. Fc-область IgG содержит домен СН2 IgG и домен СН3 IgG. Указанная СН3 область может представлять собой домен СНЗ3 с нативной последовательностью или вариант домена СН3 (например, домен СН3, в одну цепь которого введена «выпуклость» («выступ»), а в другую цепь - соответствующая «полость» («впадина»); см. патент США №5,821,333, прямо включенный сюда посредством ссылки). Такие варианты доменов СН3 могут быть использованы для стимулирования гетеродимеризации двух неидентичных тяжелых цепей иммуноглобулина, как описано здесь. Если здесь не указано иное, нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU.
При использовании здесь термин «эффекторные функции» относится к биологической активности, свойственной Fc-области иммуноглобулина, которая варьирует у иммуноглобулинов разных изотипов. Примеры эффекторных функций иммуноглобулинов включают связывание с C1q и комплемент-зависимую цитотоксичность (CDC), связывание с Fc-рецепторами, антителозависимую клеточно-опосредованную цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз (ADCP), секрецию цитокинов, захват антигенов антигенпредставляющими клетками, опосредованный иммунными комплексами, понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточных рецепторов) и активацию В-клеток.
При использовании здесь термин «регуляторная Т-клетка» или «Treg-клетка» обозначает специализированный тип CD4+ Т-клеток, которые могут подавлять ответы других Т-клеток (эффекторных Т-клеток). Treg-клетки характеризуются экспрессией CD4, α субъединицы рецептора IL-2 (CD25) и транскрипционного фактора forkhead box Р3 (FOXP3) (Sakaguchi, Annu Rev Immunol 22, 531-62 (2004)) и играют критическую роль в индуцировании и поддержании периферической самотолерантности к антигенам, включая антигены, экспрессируемые опухолями.
При использовании здесь термин «обычные CD4+ Т-клетки» обозначает CD4+ Т-клетки, отличные от регуляторных Т-клеток.
При использовании здесь термин «селективная активация Treg-клеток» обозначает активацию Treg-клеток практически без сопутствующей активации других субпопуляций Т-клеток (таких как CD4+ Т-клетки-хелперы, CD8+ цитотоксические Т-клетки, NK Т-клетки) или натуральных клеток-киллеров (NK-клеток). Методы идентификации и различения этих типов клеток описаны в Примерах. Активация может включать индуцирование сигналов рецептора IL-2 (оцениваемую, например, посредством выявления фосфорилированного STAT5a), индуцирование пролиферации (оцениваемую, например, посредством выявления Ki-67) и/или повышающую регуляцию экспрессии маркеров активации (таких как, например, CD25).
При использовании здесь «специфичное связывание» означает, что связывание селективно в отношении антигена и его можно отличить от нежелательных или неспецифических взаимодействий. Способность иммуноглобулина к связыванию с определенным антигеном может быть измерена твердофазным иммуноферментным анализом (ELISA) или другими методиками, знакомыми специалисту в данной области, например, методикой поверхностного плазмонного резонанса (SPR).
При использовании здесь термины «аффинность» или «аффинность связывания» относятся к силе общей суммы нековалентных взаимодействий между одним связывающим сайтом молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Аффинность молекулы X в отношении ее партнера Y может обычно быть представлена как константа диссоциации (KD), представляющая собой отношение констант скорости диссоциации и ассоциации (koff и kon, соответственно). Особым методом измерения аффинности является поверхностный плазмонный резонанс (SPR).
При использовании здесь термин «сниженное связывание» относится к снижению аффинности соответствующего взаимодействия, измеренному, например, посредством SPR. Наоборот, «повышенное связывание» относится к повышению аффинности связывания при соответствующем взаимодействии.
При использовании здесь термин «полимер» обычно включает, без ограничения: гомополимеры; сополимеры, такие как, например, блок-сополимеры, привитые, случайные и чередующиеся сополимеры; и тройные полимеры; и их смеси и модификации. Кроме того, в отсутствие прямых ограничений, термин «полимер» включает все возможные геометрические конфигурации вещества. Эти конфигурации включают, без ограничения, изотактическую, синдиотактическую и случайную симметрию.
«Полинуклеотид» относится к полимеру, состоящему из нуклеотидных единиц. Полинуклеотиды включают встречающиеся в природе нуклеиновые кислоты, такие как дезоксирибонуклеиновая кислота («ДНК») и рибонуклеиновая кислота («РНК»), а также аналоги нуклеиновых кислот. Аналоги нуклеиновых кислот включают аналоги, содержащие основания, не встречающиеся в природе, нуклеотиды, образующие связи с другими нуклеотидами, отличные от встречающейся в природе фосфодиэфирной связи, или содержащие основания, присоединенные через связи, отличные от фосфодиэфирных связей. Таким образом, аналоги нуклеотидов включают, например и без ограничения, фосфоротиоаты, фосфородитиоаты, фосфоротриэфиры, фосфорамидаты, боранфосфонаты, метилфосфонаты, хиральные метилфосфонаты, 2-O-метилрибонуклеотиды, пептидные нуклеиновые кислоты (ПНК) и тому подобное. Такие полинуклеотиды могут быть синтезированы, например, с использованием автоматического синтезатора ДНК. Термин «нуклеиновая кислота» обычно относится к большим полинуклеотидам. Термин «олигонуклеотид» обычно относится к коротким полинуклеотидам, обычно из не более чем приблизительно 50 нуклеотидов. Следует понимать, что представление нуклеотидной последовательности как последовательности ДНК (то есть А, Т, G, С) также включает последовательность РНК (то есть A, U, G, С), в которой «Т» заменен на «U».
Для описания полинуклеотидных последовательностей здесь применена традиционная система записи: левый конец одноцепочечной полинуклеотидной последовательности представляет собой 5'-конец; левое направление двуцепочечной полинуклеотидной последовательности обозначают как 5'-направление. Направление 5'-3' присоединения нуклеотидов к синтезируемым РНК-транскриптам обозначают как направление транскрипции. Цепь ДНК, имеющую ту же последовательность, что мРНК, называют «кодирующей цепью»; последовательности цепи ДНК, имеющие ту же последовательность, что мРНК, транскрибированная с этой ДНК, и расположенные ближе к 5'-концу от 5'-конца РНК транскрипта, называют «последовательностями, расположенными выше»; последовательности цепи ДНК, имеющие ту же последовательность, что мРНК, и расположенные ближе к 3'-концу от 3'-конца кодирующего РНК транскрипта, называют «последовательностями, расположенными ниже».
«Комплементарность» относится к топологической совместимости или взаимной сопоставимости взаимодействующих поверхностей двух полинуклеотидов. Таким образом, две молекулы могут быть описаны как комплементарные и, кроме того, характеристики их контактных поверхностей комплементарны друг другу. Первый полинуклеотид комплементарен второму полинуклеотиду, если нуклеотидная последовательность первого полинуклеотида практически идентична нуклеотидной последовательности полинуклеотидного партнера второго полинуклеотида по связыванию или если первый полинуклеотид может гибридизоваться со вторым полинуклеотидом в жестких условиях гибридизации.
«Специфично гибридизуется с», «специфичная гибридизация» или «селективная гибридизация» относятся к преимущественному связыванию, образованию дуплексов или гибридизации молекулы нуклеиновой кислоты с определенной нуклеотидной последовательностью в жестких условиях, когда эта последовательность присутствует в сложной смеси (например, общей клеточной) ДНК или РНК. Термин «жесткие условия» относится к условиям, при которых зонд будет гибридизоваться преимущественно с его подпоследовательностью-мишенью и в меньшей степени с другими последовательностями или вообще не будет гибридизоваться с другими последовательностями. «Жесткие условия гибридизации» и «жесткие условия промывки при гибридизации» в контексте экспериментов по гибридизации нуклеиновых кислот, таких как Саузерн и нозерн-гибридизация, зависят от последовательности и различаются при разных параметрах внешней среды. Всеобъемлющие руководства по гибридизации нуклеиновых кислот представлены в Tijssen, 1993, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, part I, chapter 2, "Overview of principles of hybridization and the strategy of nucleic acid probe assays", Elsevier, N.Y.; Sambrook et al., 2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, 3.sup.rd ed., NY; и Ausubel et al., eds., Current Edition, Current Protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, NY.
Обычно условия гибридизации и промывки высокой жесткости выбирают так, чтобы они были примерно на 5°С ниже температуры плавления (Tm) конкретной последовательности при заданных ионной силе и рН. Tm представляет собой температуру (при заданных ионной силе и рН), при которой 50% целевой последовательности гибридизуется с идеально соответствующим ей зондом. Очень жесткие условия выбирают так, чтобы они были равны Tm для соответствующего зонда. Примером жестких условий гибридизации для гибридизации комплементарных нуклеиновых кислот, имеющих более чем приблизительно 100 комплементарных остатков, на фильтре при Саузерн- или нозерн-блоттинге является 50%-й формалин с 1 мг гепарина при 42°С, при этом гибридизацию проводят в течение ночи. Примером условий промывки высокой жесткости является 0,15 М NaCl при 72°С на протяжении приблизительно 15 минут. Примером жестких условий промывки является промывка с использованием 0,2х SSC при 65°С на протяжении 15 минут. SSC-буфер описан в Sambrook et al. Перед промывкой в условиях высокой жесткости может быть проведена промывка в условиях низкой жесткости для удаления фонового сигнала зонда. Типичная промывка средней жесткости для дуплекса из, например, более чем приблизительно 100 нуклеотидов представляет собой 1x SSC при 45°С на протяжении 15 минут. Типичная промывка низкой жесткости для дуплекса из, например, более чем приблизительно 100 нуклеотидов представляет собой 4-6х SSC при 40°С на протяжении 15 минут. Обычно отношение сигнала к шуму, в 2 (или более) раза превышающее наблюдаемое для неродственного зонда в соответствующем гибридизационном анализе, указывает на выявление специфичной гибридизации.
«Праймер» относится к полинуклеотиду, способному специфично гибридизоваться с обозначенной полинуклеотидной матрицей и обеспечивать точку начала синтеза комплементарного полинуклеотида. Такой синтез происходит при помещении полинуклеотидного праймера в условия, в которых происходит запуск синтеза, то есть в присутствии нуклеотидов, комплементарной полинуклеотидной матрицы и агента для полимеризации, такого как ДНК-полимераза. Праймер обычно является одноцепочечным, но может быть двуцепочечным. Праймеры обычно представляют собой дезоксирибонуклеиновые кислоты, но во многих случаях применим широкий спектр синтетических и встречающихся в природе праймеров. Праймер комплементарен матрице, для гибридизации с которой он разработан, обеспечивая место начала синтеза, но он не обязательно отражает точную последовательность матрицы. В таком случае специфичная гибридизация праймера с матрицей зависит от строгости условий гибридизации. Праймеры могут быть мечены, например, хромогенными, радиоактивными или флуоресцентными группировками и использованы в качестве выявляемых группировок.
При использовании в связи с полинуклеотидом «зонд» относится к полинуклеотиду, способному специфично гибридизоваться с обозначенной последовательностью другого полинуклеотида. Зонд специфично гибридизуется с целевым комплементарным полинуклеотидом, но не обязательно отражает точную комплементарную последовательность матрицы. В таком случае специфичная гибридизация зонда с целевым полинуклеотидом зависит от строгости условий гибридизации. Зонды могут быть мечены, например, хромогенными, радиоактивными или флуоресцентными группировками и использованы в качестве выявляемых группировок. В случаях, когда зонд обеспечивает точку начала синтеза комплементарного полинуклеотида, зонд может также быть праймером.
«Вектор» представляет собой полинуклеотид, который может быть использован для введения другой нуклеиновой кислоты, связанной с ним, в клетку. Одним типом вектора является «плазмида», что относится к линейной или кольцевой молекуле двуцепочечной ДНК, в которую могут быть лигированы дополнительные сегменты нуклеиновой кислоты. Другим типом вектора является вирусный вектор (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), где в геном вируса могут быть введены дополнительные сегменты ДНК. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, содержащие бактериальный репликатор, и эписомные векторы для клеток млекопитающих). Другие векторы (например, неэписомные векторы для клеток млекопитающих) проходят интеграцию в геном клетки-хозяина при введении в клетку-хозяина и, благодаря этому, проходят репликацию вместе с геномом хозяина. «Вектор экспрессии» представляет собой тип вектора, способный управлять экспрессией выбранного полинуклеотида.
«Регуляторная последовательность» представляет собой нуклеиновую кислоту, влияющую на экспрессию (например, уровень, время или место экспрессии) нуклеиновой кислоты, с которой она функционально связана. Регуляторная последовательность может, например, оказывать свои эффекты на регулируемую нуклеиновую кислоту непосредственно или через действие одной или более чем одной другой молекулы (например, полипептидов, связанных с регуляторной последовательностью и/или нуклеиновой кислотой). Примеры регуляторных последовательностей включают промоторы, энхансеры и другие элементы контроля экспрессии (например, сигналы полиаденилирования). Другие примеры регуляторных последовательностей описаны, например, в Goeddel, 1990, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Calif, and Baron et al., 1995, Nucleic Acids Res. 23:3605-06. Нуклеотидная последовательность «функционально связана» с регуляторной последовательностью, если регуляторная последовательность влияет на экспрессию (например, уровень, время или место экспрессии) нуклеотидной последовательности.
«Клетка-хозяин» представляет собой клетку, которая может быть использована для экспрессии полинуклеотида по изобретению. Клетка-хозяин может представлять собой прокариота, например, Е. coli, или она может представлять собой эукариота, например, одноклеточного эукариота (например, дрожжи или другой гриб), растительную клетку (например, растительную клетку табака или томата), животную клетку (например, человеческую клетку, клетку обезьяны, клетку хомяка, крысиную клетку, мышиную клетку или клетку насекомого) или гибридому. Обычно клетка-хозяин представляет собой культивируемую клетку, которая может быть трансформирована или трансфицирована нуклеиновой кислотой, кодирующей полипептид, которая может затем быть экспрессирована в клетке-хозяине. Фраза «рекомбинантная клетка-хозяин» может быть использована для обозначения клетки-хозяина, трансформированной или трансфицированной нуклеиновой кислотой, которую следует экспрессировать. Клетка-хозяин может также представлять собой клетку, содержащую нуклеиновую кислоту, но не экспрессирующую ее на желаемом уровне, если в клетку-хозяина не введена регуляторная последовательность таким образом, что она становится функционально связанной с этой нуклеиновой кислотой. Следует понимать, что термин «клетка-хозяин» относится не только к конкретной рассматриваемой клетке, но также к потомству или возможному потомству такой клетки. Поскольку в последующих поколениях возможны определенные модификации, обусловленные, например, мутациями или влиянием внешней среды, такое потомство может фактически не быть идентичным исходной клетке, но объем данного термина, при его использовании здесь, все равно включает такое потомство.
Термин «выделенная молекула» (где молекула представляет собой, например, полипептид или полинуклеотид) представляет собой молекулу, которая, в силу источника ее происхождения: (1) не связана с компонентами, с которыми она связана естественным образом в ее нативном состоянии; (2) по существу свободна от других молекул того же вида; (3) экспрессирована клеткой другого вида; и (4) не встречается в природе. Таким образом, химически синтезированная молекула или молекула, экспрессированная в клеточной системе, отличной от клетки, являющейся ее естественным источником, будет «выделенной» из компонентов, с которыми она связана естественным образом. Молекула может также стать практически свободной от компонентов, с которыми она связана естественным образом, в результате ее выделения с применением методик очистки, хорошо известных в данной области. Чистоту или однородность молекулы можно анализировать с применением ряда методов, хорошо известных в данной области. Например, чистоту образца полипептида можно анализировать с применением электрофореза в полиакриламидном геле и окрашивания геля для визуализации полипептида с применением методик очистки, хорошо известных в данной области. Для определенных задач более высокое разрешение можно обеспечить с применением HPLC (высокоэффективной жидкостной хроматографии) или других методов очистки, хорошо известных в данной области.
Белок или полипептид является «практически чистым», «практически однородным» или «практически очищенным», когда по меньшей мере приблизительно от 60% до 75% образца представлены одной разновидностью полипептида. Полипептид или белок может быть мономерным или мультимерным. По существу чистый полипептид или белок будет обычно составлять приблизительно 50%, 60%, 70%, 80% или 90%, масс/масс, образца белка, чаще приблизительно 95%, и предпочтительно будет чистым более чем на 99%. Чистота или однородность белка может быть продемонстрирована рядом методов, хорошо известных в данной области, таких как электрофорез образца белка в полиакриламидном геле с последующей визуализацией единственной полосы полипептида после окрашивания геля красителем, хорошо известным в данной области. Для определенных задач более высокое разрешение можно обеспечить с применением HPLC или других методов очистки, хорошо известных в данной области.
При использовании здесь термины «метка» или «меченый»/«меченный» относятся к включению другой молекулы в антитело. В одном воплощении метка представляет собой выявляемый маркер, например, при включении радиоактивно меченной аминокислоты или присоединении к полипептиду биотинильных группировок, которые могут быть выявлены авидином, содержащим маркер (например, стрептавидином, содержащим флуоресцентный маркер или имеющим ферментативную активность, которая может быть выявлена оптическими или калориметрическими методами). В другом воплощении метка или маркер могут быть терапевтическими, например, конъюгатом лекарственного средства или токсином. Могут быть применены различные методы мечения полипептидов и гликопротеинов, хорошо известные в данной области. Примеры меток для полипептидов включают, без ограничения, следующее: радиоизотопы или радионуклиды (например, 3Н, 14С, 15N, 35S, 90Y, 99Тс, 111In, 125I, 131I), флуоресцентные метки (например, FITC, родамин, лантаноидные люминофоры), ферментные метки (например, пероксидазу хрена, β-галактозидазу, люциферазу, щелочную фосфатазу), хемилюминесцентные маркеры, биотинильные группы, предопределенные полипептидные эпитопы, распознаваемые вторичным репортером (последовательности пар лейциновых застежек-молний, сайты связывания для вторичных антител, металлосвязывающие домены, эпитопные метки), магнитные агенты, такие как хелаты гадолиния, токсины, такие как коклюшный токсин, таксол, цитохалазин В, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенипозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин, и их аналоги и гомологи. В различных воплощениях метки присоединены через спейсеры различной длины для уменьшения возможного стерического несоответствия.
При использовании здесь термин «гетерологичный» относится к композиции или структуре, не являющимся нативными или не представленными в природе, например к композиции или структуре, которые могут быть получены посредством замены существующей естественной композиции или структуры композицией или структурой, имеющей происхождение от другого источника. Сходным образом, экспрессия белка в организме, отличном от организма, экспрессирующего этот белок естественным образом, составляет гетерологичную систему экспрессии и гетерологичный белок.
Следует понимать, что аспекты и воплощения изобретения, описанные здесь, включают аспекты и воплощения, «состоящие» или «состоящие по существу из» описанных аспектов и воплощений.
Ссылка на «приблизительно» в приведенном здесь описании значения или параметра включает (и описывает) варианты, направленные непосредственно на это значение или параметр. Например, описание со ссылкой на «приблизительно X» включает описание «X».
Если контекстом ясно не продиктовано иное, при использовании здесь и в приложенной формуле изобретения формы единственного числа включают множественное число. Следует понимать, что аспекты и варианты изобретения, описанные здесь, включают аспекты и воплощения, «состоящие» или «состоящие по существу из» описанных аспектов и вариантов.
Описание платформы VitoKine
Согласно настоящему изобретению предложена биологически активируемая лекарственная платформа на основе цитокинов («VitoKine»), направленная на снижение системной токсичности, основанной на механизме действия, и обеспечение более широкого терапевтического применения белков, например, цитокинов. Со ссылкой на ФИГ. 1, новые конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой направленный домен, домен, увеличивающий период полувыведения, или домен группировки с двойной функцией или многофункциональной группировки, «домен активной группировки» (D2) и «домен маскирующей группировки» (D3). Предложенные способы активации домена D2 VitoKine показаны на ФИГ. 2. Важно, что сохранение инертности или сниженной активности D2 конструкции VitoKine до ее местной активации протеазами, регуляция которых повышена в тканях, пораженных заболеванием, будет ограничивать связывание активной группировки с ее рецепторами на периферии или на поверхности клеток, не пораженных заболеванием, предотвращая избыточную активацию ее пути и уменьшая нежелательную токсичность, обусловленную связыванием с мишенью вне целевой ткани. Кроме того, инертность активной группировки VitoKine до активации протеазами будет существенно снижать возможный клиренс, обусловленный связыванием с антигеном или мишенью, и, таким образом, увеличивать период полувыведения in vivo и улучшать биораспределение и биодоступность в намеченных местах терапии.
Домен D1 («направленный домен, домен, увеличивающий период полувыведения, и. in домен группировки с двойной функцией или многофункциональной группировки»)
В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой направленную группировку в форме антитела, или фрагмента антитела, или белка, или пептида к опухоль-ассоциированному антигену. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой антитело, фрагмент антитела, белок или пептид к модулятору иммунных контрольных точек. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой антитело, или фрагмент антитела, или белок, или пептид как аутоиммунный модулятор. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат D1, функция которого состоит в удержании домена D2 в тканевом очаге, таком как опухолевое микроокружение (ТМЕ) или тканевые очаги воспаления. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат D1, являющийся бифункциональным, например тканенаправленным и обеспечивающим удержание в ткани. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой полимер. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой группировку, увеличивающую период полувыведения. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D1, представляющий собой Fc-домен.
Fc-домены
Иммуноглобулины класса IgG являются одними из наиболее распространенных белков в человеческой крови. Период их полувыведения из циркуляции может достигать 21 суток. Были сообщения о слитых белках, содержавших комбинации Fc-областей IgG с доменами других белков, таких как различные цитокины и рецепторы (см., например, Capon et al., Nature, 337:525-531, 1989; Chamow et al., Trends Biotechnol., 14:52-60, 1996; патенты США №№5,116,964 и 5,541,087). Прототипом слитого белка является гомодимерный белок, связанный через цистеиновые остатки в шарнирной области Fc IgG с получением молекулы, сходной с молекулой IgG, без вариабельного домена и домена СН1 тяжелой цепи и легких цепей. Димерная природа слитых белков, содержащих Fc-домен, может быть предпочтительной с точки зрения обеспечения взаимодействий более высокого порядка (то есть бивалентного или биспецифичного связывания) с другими молекулами. В силу структурной гомологии профиль фармакокинетики Fc-слитых белков in vivo сопоставим с соответствующим профилем человеческого IgG сходного изотипа.
Термин «Fc» относится к молекуле или последовательности, содержащим последовательность фрагмента целого антитела, не являющегося антигенсвязывающим фрагментом, в мономерной или мультимерной форме. Иммуноглобулин, являющийся источником нативного Fc, предпочтительно имеет происхождение из человека и может представлять собой любой из иммуноглобулинов, однако предпочтительны IgG1 и IgG2. Нативные Fc составлены из мономерных полипептидов, которые могут быть связаны с образованием димерных или мультимерных форм ковалентными (то есть дисульфидными) и нековалентными связями. Число межмолекулярных дисульфидных связей между мономерными субъединицами нативных молекул Fc варьирует от 1 до 4 в зависимости от класса (например, IgG, IgM, IgA, IgE) или подкласса (например, IgG1, IgG2, IgG3, IgG4, IgA1, IgGA2). Одним примером нативного Fc является дисульфидно связанный димер, полученный в результате расщепления IgG папаином (см. Ellison et al. (1982), Nucleic Acids Res. 10: 4071-9). При использовании здесь термин «нативный Fc» является общим для мономерных, димерных и мультимерных форм. Fc-домены содержат сайты связывания для белка А, белка G, различных Fc-рецепторов и белков комплемента.
В различных воплощениях термин «вариант Fc» относится к молекуле или последовательности, модифицированным относительно нативного Fc, но все еще содержащим сайт связывания для реутилизационного рецептора FcRn. Типичные варианты Fc, а также взаимодействие с реутилизационным рецептором описаны в международных заявках WO 97/34631 (опубликована 25 сентября 1997 г. ) и WO 96/32478, включенных сюда посредством ссылки. Кроме того, нативный Fc содержит сайты, которые могут быть удалены, поскольку они придают структурные характеристики или биологическую активность, не являющиеся необходимыми для слитых молекул по настоящему изобретению. Таким образом, в различных воплощениях термин «вариант Fc» включает молекулу или последовательность, в которых нет одного или более чем одного сайта или остатка нативного Fc, влияющих на или вовлеченных в (1) образование дисульфидных связей, (2) несовместимость с выбранной клеткой-хозяином, (3) N-концевую неоднородность при экспрессии в выбранной клетке-хозяине, (4) гликозилирование, (5) взаимодействие с комплементом, такое как CDC, (6) связывание с Fc-рецептором, отличным от реутилизационного рецептора, или (7) антителозависимую клеточную цитотоксичность (ADCC).
Термин «Fc-домен» охватывает молекулы и последовательности нативных Fc и вариантов Fc, определенные выше. Как в случае вариантов Fc и нативных Fc, термин «Fc-домен» включает молекулы в мономерной или мультимерной форме, полученные расщеплением целого антитела, рекомбинантной экспрессией генов или иным образом. В различных воплощениях «Fc-домен» относится к димеру двух Fc-доменных мономеров (SEQ ID NO: 13), который обычно содержит полноразмерную шарнирную область или ее часть. В различных воплощениях возможны мутации Fc-домена для устранения эффекторных функций. В различных воплощениях каждый из Fc-доменных мономеров в Fc-домене содержит аминокислотные замены в константном домене СН2 антитела для снижения взаимодействия или связывания между Fc-доменом и Fcγ-рецептором. В различных воплощениях каждая субъединица Fc-домена содержит две аминокислотные замены, снижающие связывание с активирующим Fc-рецептором и/или эффекторную функцию, где указанные аминокислотные замены представляют собой L234A и L235A. В различных воплощениях каждая субъединица Fc-домена содержит три аминокислотные замены, снижающие связывание с активирующим Fc-рецептором и/или эффекторную функцию, где указанные аминокислотные замены представляют собой L234A, L235A и G237A (SEQ ID NO: 14).
В различных воплощениях возможны мутации Fc-домена для дополнительного увеличения периода полувыведения in vivo. В различных воплощениях каждая субъединица Fc-домена содержит три аминокислотные замены, усиливающие связывание с человеческим FcRn, где указанные аминокислотные замены представляют собой M252Y, S254T и Т256Е, раскрытые в публикации патента США №7,658,921 (SEQ ID NO: 156). В различных воплощениях каждая субъединица Fc-домена содержит одну аминокислотную замену, усиливающую связывание с человеческим FcRn, где указанная аминокислотная замена представляет собой N434A (SEQ ID NO: 166), раскрытую в публикации патента США №7,371,826. В различных воплощениях каждая субъединица Fc-домена содержит одну аминокислотную замену, усиливающую связывание с человеческим FcRn, где указанные аминокислотные замены представляют собой M428L и N434S, раскрытые в публикации патента США №8,546,543. В различных воплощениях возможны комбинации мутаций, увеличивающих период полувыведения, с аминокислотными заменами, снижающими связывание с активирующим Fc-рецептором и/или эффекторную функцию.
В различных воплощениях каждый из двух Fc-доменных мономеров в Fc-домене содержит аминокислотные замены, способствующие гетер од имеризации двух мономеров. В различных воплощениях гетеродимеризации Fc-доменных мономеров может способствовать введение разных, но совместимых замен в два Fc-доменных мономера, таких как пары остатков типа «выступ во впадину» («knob-into-hole»). Методика «выступ во впадину» также раскрыта в публикации патента США №8,216,805. В еще одном воплощении один Fc-доменный мономер содержит мутацию типа «выступ» T366W, а другой Fc-доменный мономер содержит мутации типа «впадина» T366S, L358A и Y407V. В различных воплощениях проводят введение двух остатков Cys (S354C на стороне «выступа» и Y349C на стороне «впадины»), образующие стабилизирующую дисульфидную связь (SEQ ID NO: 15 и 16). Использование гетеродимерного Fc может приводить к получению моновалентной конструкции VitoKine.
В различных воплощениях последовательность Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность Fc-домена человеческого IgG1, изложенную в SEQ ID NO: 14:

где SEQ ID NO: 14 содержит аминокислотные замены (подчеркнуты), устраняющие связывание с FcγR и C1q.
В различных воплощениях последовательность гетеродимерного Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность Knob-Fc-домена, изложенную в SEQ ID NO: 15:

где SEQ ID NO: 15 содержит аминокислотные замены (подчеркнуты), устраняющие связывание с FcγR и C1q.
В различных воплощениях последовательность гетеродимерного Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность Hole-Fc-домена, изложенную в SEQ ID NO: 16:

где SEQ ID NO: 16 содержит аминокислотные замены (подчеркнуты), устраняющие связывание с FcγR и C1q.
В различных воплощениях последовательность Fc-домена, используемая для получения конструкций VitoKine, представляет собой Fc-домен IgG1 со сниженной/устраненной эффекторной функцией и увеличенным периодом полувыведения, имеющий аминокислотную последовательность, изложенную в SEQ ID NO: 156:

где SEQ ID NO: 156 содержит аминокислотные замены (подчеркнуты), устраняющие связывание с FcγR и C1q, и аминокислотные замены (выделены полужирным шрифтом) для увеличения периода полувыведения.
В различных воплощениях последовательность Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность Fc-домена человеческого IgG1, изложенную в SEQ ID NO: 166:

где SEQ ID NO: 166 содержит аминокислотные замены (подчеркнуты), устраняющие связывание с FcγR и C1q, и аминокислотную замену (выделена полужирным шрифтом) для увеличения периода полувыведения.
В различных воплощениях последовательность гетеродимерного Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность Knob-Fc-домена с увеличенным периодом полувыведения, изложенную в SEQ ID NO: 167:

где SEQ ID NO:1 67 содержит аминокислотные замены (подчеркнуты), устраняющие связывание с FcγR и C1q, и аминокислотную замену (выделена полужирным шрифтом) для увеличения периода полувыведения.
В различных воплощениях последовательность гетеродимерного Fc-домена, используемая для получения конструкций VitoKine, представляет собой последовательность Hole-Fc-домена с увеличенным периодом полувыведения, изложенную в SEQ ID NO: 168:

где SEQ ID NO: 168 содержит аминокислотные замены (подчеркнуты), устраняющие связывание с FcγR и C1q, и аминокислотную замену (выделена полужирным шрифтом) для увеличения периода полувыведения.
Антитела и белки/пептиды, связывающиеся с ассоциированными с болезнью мишенями или опухоль-ассоциированными антигенами
В различных воплощениях D1 может представлять собой направленную группировку в форме антитела к опухоль-ассоциированному антигену (ТАА) или другого белка или пептида, демонстрирующего аффинность связывания в отношении клетки, пораженной заболеванием, или ткани, пораженной заболеванием. ТАА может представлять собой любую молекулу, макромолекулу, комбинацию молекул и так далее, против которых желателен иммунный ответ. ТАА может представлять собой белок, содержащий более чем одну полипептидную субъединицу. Например, белок может представлять собой димер, тример или мультимер более высокого порядка. В различных воплощениях две или более субъединицы белка могут быть соединены ковалентной связью, такой как, например, дисульфидная связь. В различных воплощениях субъединицы белка могут удерживаться вместе за счет нековалентных взаимодействий. Таким образом, ТАА может представлять собой любые пептид, полипептид, белок, нуклеиновую кислоту, липид, углевод или малую органическую молекулу или любую их комбинацию, против которых специалист в данной области желает индуцировать иммунный ответ. В различных воплощениях ТАА представляет собой пептид, содержащий приблизительно 5, приблизительно 6, приблизительно 7, приблизительно 8, приблизительно 9, приблизительно 10, приблизительно 11, приблизительно 12, приблизительно 13, приблизительно 14, приблизительно 15, приблизительно 16, приблизительно 17, приблизительно 18, приблизительно 19, приблизительно 20, приблизительно 25, приблизительно 30, приблизительно 35, приблизительно 40, приблизительно 45, приблизительно 50, приблизительно 55, приблизительно 60, приблизительно 65, приблизительно 70, приблизительно 75, приблизительно 80, приблизительно 85, приблизительно 90, приблизительно 95, приблизительно 100, приблизительно 150, приблизительно 200, приблизительно 250, приблизительно 300, приблизительно 400, приблизительно 500, приблизительно 600, приблизительно 700, приблизительно 800, приблизительно 900 или приблизительно 1000 аминокислот. В различных воплощениях пептид, полипептид или белок представляет собой молекулу, обычно вводимую субъектам посредством инъекции. В различных воплощениях, поел* введения, антитело или связывающий белок, специфичные в отношении опухоли действуют как направленная группировка, направляющая VitoKine в очаг заболевания такой как раковый очаг, где возможно высвобождение активного домена и его взаимодействие с его когнатными рецепторами на клетках, пораженных заболеванием, или ткани, пораженной заболеванием.
Любой из указанных выше маркеров может быть использован в качестве ассоциированной с болезнью мишени или ТАА-мишени для конструкций VitoKine по данному изобретению. В различных воплощениях одна или более чем одна ассоциированная с болезнью мишень, или ее вариант, или ТАА, вариант ТАА, или мутант ТАА, которые предполагается использовать в конструкциях VitoKine и способах по настоящему изобретению, выбраны из списка, представленного в Таблице 2, или имеют происхождение от антигенов, представленных в этом списке.
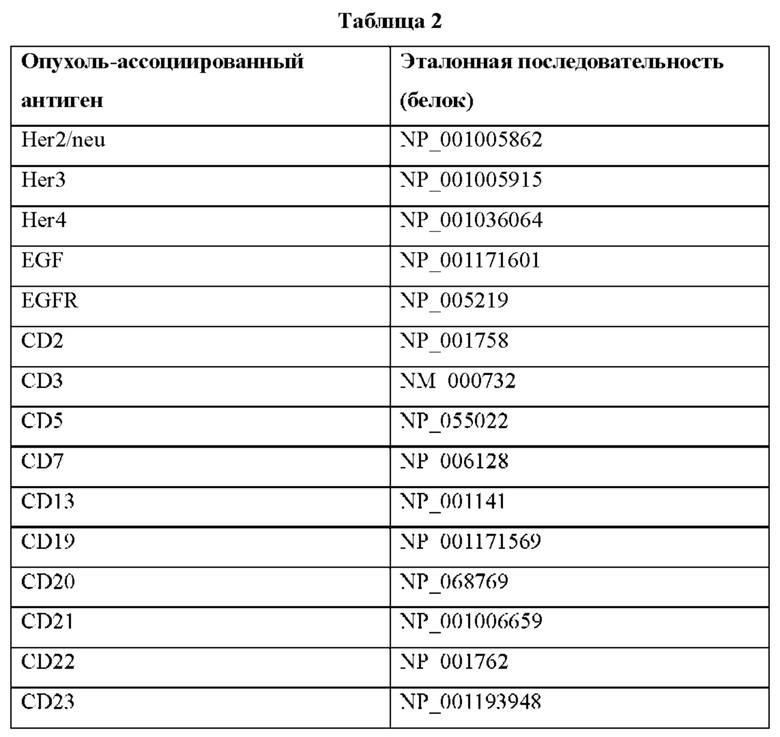



Другие примеры опухоль-ассоциированных антигенов включают TRP-1, TRP-2, MAG-1, MAGE-3, BAGE, GAGE-1, GAGE-2, р15(58), СЕА, RAGE, NY-BSO (LAGE), SCP-1, Hom/Mel-40, H-Ras, BCR-ABL, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, антигены E6 и E7 вируса папилломы человека (HPV), TSP-180, MAGE-4, MAGE-5, MAGE-6, p185erbB2, p180erbB-3, c-met, nm-23H1, PSA, TAG-72-4, CA 19-9, CA 72-4, CAM 17.1, Numa, K-ras, β-катенин, CDK4, Muni-1, p16, TAGE, PSCA, CT7, теломеразу, 43-9F, 5T4, 791Tgp72, β-HCG, BCA225, BTAA, CA 15-3 (CA 27.29/BCAA), CA 195, CA 242, CA-50, CAM43, CD68/KF1, CO-029, FGF-5, G250, Ga733 (EpCAM), HTgp-175, M344, MA-50, MG7-Ag, MOV18, NB/70K, NY-CO-1, RCAS1, SDCCAG16, TA-90 (белок, связывающий Мас-2/белок, ассоциированный с циклофилином С), TAAL6, TAG72, TLP и TPS.
Модуляторы иммунных контрольных точек
Согласно сообщениям, различные иммунные клетки экспрессируют ряд белковых антигенов иммунных контрольных точек, включая, например, CD152 (экспрессируемый активированными CD8+ Т-клетками, CD4+ Т-клетками и регуляторными Т-клетками), CD279 (экспрессированный на опухоль-инфильтрирующих лимфоцитах, экспрессируемый активированными Т-клетками (как CD4, так и CD8), регуляторными Т-клетками, активированными В-клетками, активированными NK-клетками, анергичными Т-клетками, моноцитами, дендритными клетками), CD274 (экспрессированный на Т-клетках, Вклетках, дендритных клетках, клетках эндотелия сосудов, островковых клетках поджелудочной железы) и CD223 (экспрессируемый активированными Т-клетками, регуляторными Т-клетками, анергичными Т-клетками, NK-клетками, NKT-клетками и плазмоцитоидными дендритными клетками) (см., например, Pardoll, D., Nature Reviews Cancer, 12:252-264, 2012). Антитела, связывающиеся с антигеном, определенным как белок иммунных контрольных точек, известны специалистам в данной области. Например: в данной области описаны различные антитела против CD276 (см., например, публикацию заявки на патент США №20120294796 (Johnson et al.) и указанные в ней источники); в данной области описаны различные антитела против CD272 (см., например, публикацию заявки на патент США №20140017255 (Mataraza et al.) и указанные в ней источники); в данной области описаны различные антитела против CD152/CTLA-4 (см., например, публикацию заявки на патент США №20130136749 (Korman et al.) и указанные в ней источники); в данной области описаны различные антитела против LAG-3/CD223 (см., например, публикацию заявки на патент США №20110150892 (Thudium et al.) и указанные в ней источники); в данной области описаны различные антитела против CD279/PD-1 (см., например, патент США №7,488,802 (Collins et al.) и указанные в нем источники); в данной области описаны различные антитела против PD-L1 (см., например, публикацию заявки на патент США №20130122014 (Korman et al.) и указанные в ней источники); в данной области описаны различные антитела против TIM-3 (см., например, публикацию заявки на патент США №20140044728 (Takayanagi et al.) и указанные в ней источники); и в данной области описаны различные антитела против В7-Н4 (см., например, публикацию заявки на патент США №20110085970 (Terrett et al.) и указанные в ней источники). Каждый из этих источников полностью включен сюда посредством ссылки для специфических антител и последовательностей, изложенных здесь.
В различных воплощениях D1 может содержать антитело, фрагмент антитела, или белок, или пептид, демонстрирующие связывание с белковым антигеном иммунных контрольных точек, присутствующим на поверхности иммунной клетки. В различных воплощениях белковый антиген иммунных контрольных точек выбран из группы, состоящей из, без ограничения, CD276, CD272, CD 152, CD223, CD279, CD274, CD40, SIRPa, CD47, ОХ-40, GITR, ICOS, CD27, 4-1ВВ, TIM-3, В7-Н4, Siglec 7, Siglec 8, Siglec 9, Siglec 15 и VISTA.
В различных воплощениях Dl может содержать антитело к белковому антигену иммунных контрольных точек, присутствующему на поверхности опухолевой клетки, выбранному из группы, состоящей из, без ограничения, PD-L1, В7-Н3 и В7-Н4.
Модуляторы для аутоиммунных и воспалительных расстройств
Любой из указанных выше белков, экспрессия которых высока в различных воспаленных тканях или на иммунных клетках, может быть использован в качестве мишени, связанной с аутоиммунными/воспалительными заболеваниями, для конструкций VitoKine по данному изобретению. В различных воплощениях одна или более чем одна мишень, связанная с аутоиммунными/воспалительными заболеваниями, ее вариант или ее мутант/изоформа, которые предполагается использовать в конструкциях VitoKine и способах по настоящему изобретению, выбраны из списка, представленного в Таблице 3, или имеют происхождение от мишеней, представленных в этом списке. Эти мишени можно также использовать для направленного воздействия на рак.


В различных воплощениях направленная группировка D1 может представлять собой антитело, фрагмент антитела, другой белок или пептид, специфичные в отношении воспаленной ткани, демонстрирующие связывание с клеткой, пораженной заболеванием, или микроокружением заболевания, таким как TNF, TNFR, интегрин А4β7, IL-6Rα, BLYS, TSLP.
Полимеры
В различных воплощениях D1 может представлять собой полимер, например полиэтиленгликоль (PEG). В различных воплощениях полимер, например PEG, может быть ковалентно присоединен к N- или С-концу или во внутреннем положении с использованием обычных химических методов, например химической конъюгации. В различных воплощениях полимер, например PEG, может быть ковалентно присоединен к N-концу домена D2 сайт-специфической конъюгацией или заменой аминокислот цитокина на другие аминокислоты или специальные конструированные аминокислоты.
Группировки, увеличивающие период полувыведения
В различных воплощениях в качестве доменов D1 в настоящем изобретении могут быть использованы другие группировки, увеличивающие период полувыведения, для увеличения периода полувыведения VitoKine из сыворотки. Группировки, увеличивающие период полувыведения, включают, без ограничения, Fc-домен, вариант Fc, фрагмент антитела (Fab, scFv), EXTEN (Schellenberger et al., Nat. Biotechnol. 27:1 186-1 192, 2009) и человеческий сывороточный альбумин.
Домен D2 («домен активной группировки»)
D2 представляет собой активную группировку конструкции VitoKine, активность которой обратимо замаскирована в конструкции и может быть восстановлена при расщеплении протеазами в очаге заболевания. Активная группировка может представлять собой любой белок, включая, без ограничения, полипептид любого нативного или вариантного интерлейкина или цитокина. Важно, что сохранение инертности или сниженной активности «активной группировки» конструкции VitoKine до ее местной активации протеазами, регуляция которых повышена в тканях, пораженных заболеванием, будет ограничивать связывание активной группировки с рецепторами на периферии или на поверхности клеток, не пораженных заболеванием, предотвращая избыточную активацию ее пути и уменьшая нежелательную токсичность, обусловленную связыванием с мишенью вне целевой ткани. Кроме того, инертность активной группировки VitoKine до активации протеазами будет существенно снижать возможный клиренс, обусловленный связыванием с антигеном или мишенью, и, таким образом, увеличивать период полувыведения in vivo и улучшать биораспределение и действие в намеченных местах терапии.
IL-15
Интерлейкин-15 (IL-15) представляет собой цитокин, идентифицированный двумя независимыми группами на основании его способности стимулировать пролиферацию IL-2-зависимой Т-клеточной линии CTLL-2 в присутствии нейтрализующих антител против IL-2 (Steel et al., Trends in Pharmcological Sciences, 33(1):35-41, 2012). IL-15 и интерлейкин-2 (IL-2) имеют сходные биологические свойства in vitro в соответствии с их общими рецепторными (R) сигнальными компонентами (IL-2/15Rβγc). Тем не менее, специфичность в отношении IL-15 против IL-2 обеспечивают уникальные собственные рецепторы из α-цепей, завершающие гетеротримерные высокоаффинные рецепторные комплексы IL-15Rαβγ и IL-2Rαβγ и таким образом позволяющие клеткам отвечать по-разному в зависимости от лиганда и экспрессируемого высокоаффинного рецептора. Примечательно, что как транскрипты IL-15, так и транскрипты IL-15Rα имеют намного более широкое распространение в тканях, чем IL-2/IL-2Rα. Кроме того, тщательный контроль экспрессии IL-15 обеспечивают множество сложных посттранскрипционных регуляторных механизмов. Таким образом, исходя из сложной регуляции, а также различий паттернов экспрессии IL-15 и IL-15Rα, вероятно, что критически важные функции этой пары рецептор/лиганд in vivo отличаются от функций IL-2 и IL-2Rα. В проведенных до настоящего времени исследованиях по изучению биологии IL-15 были определены несколько ключевых неисчерпывающих функций, такие как важность IL-15 при развитии и функционировании натуральных клеток-киллеров (NK-клеток), NK-T-клеток и кишечных интраэпителиальных лимфоцитов. Роль IL-15 при аутоиммунных процессах, таких как ревматоидный артрит, и злокачественных новообразованиях, таких как Т-клеточная лимфома взрослых, указывает на то, что нарушения регуляции IL-15 могут оказывать пагубное действие на хозяина (Fehniger et al., Bllod, 97:14-32, 2001).
При использовании здесь в контексте белков и полипептидов термины «нативный IL-15» и «нативный интерлейкин-15» относятся к любым встречающимся в природе аминокислотным последовательностям интерлейкина-15 млекопитающих, включая как незрелые формы или формы-предшественники, так и зрелые формы. Неограничивающие примеры регистрационных номеров аминокислотных последовательностей различных разновидностей нативного интерлейкина-15 млекопитающих в GenBank включают NP_032383 (Mus musculus, незрелая форма), ААВ60398 (Масаса mulatta, незрелая форма), NP_000576 (человеческий, незрелая форма), САА62616 (человеческий, незрелая форма), AAI00964 (человеческий, незрелая форма) и ААН18149 (человеческий). В различных воплощениях настоящего изобретения нативный IL-15 представляет собой незрелую форму или форму-предшественника встречающегося в природе IL-15 млекопитающего. В других воплощениях нативный IL-15 представляет собой зрелую форму встречающегося в природе IL-15 млекопитающего. В различных воплощениях нативный IL-15 представляет собой форму-предшественника встречающегося в природе человеческого IL-15. В различных воплощениях нативный IL-15 представляет собой зрелую форму встречающегося в природе человеческого IL-15. В различных воплощениях белок/полипептид нативного IL-15 является выделенным или очищенным. В различных воплощениях домен D2 на основе IL-15 имеет происхождение от аминокислотной последовательности предшественника человеческого IL-15, изложенной в SEQ ID NO: 1:

В различных воплощениях домен D2 на основе IL-15 содержит аминокислотную последовательность зрелой формы человеческого IL-15, изложенную в SEQ ID NO: 2:

В различных воплощениях домен D2 на основе IL-15 будет вариантом (или мутантом) IL-15, содержащим последовательность, имеющую происхождение от последовательности полипептида зрелого человеческого IL-15, изложенной в SEQ ID NO: 2. Варианты (или мутанты) IL-15 названы здесь с использованием нативной аминокислоты, ее положения в зрелой последовательности и вариантной аминокислоты. Например, «huIL-15 S58D» относится к человеческому IL-15, содержащему замену S на D в положении 58 SEQ ID NO: 2. В различных воплощениях домен D2 по настоящему изобретению содержит домен IL-15, представляющий собой вариант IL-15 (также называемый здесь мутантным доменом IL-15). В различных воплощениях вариант IL-15 содержит аминокислотную последовательность, отличную от нативного белка IL-15 (или IL-15 дикого типа). В различных воплощениях вариант IL-15 связывается с полипептидом IL-15Rα и функционирует как агонист или антагонист IL-15. В различных воплощениях варианты IL-15 с агонистической активностью имеют суперагонистическую активность. В различных воплощениях вариант IL-15 может функционировать как агонист или антагонист IL-15 независимо от его связывания с IL-15Rα. Для агонистов IL-15 характерна сопоставимая или повышенная биологическая активность по сравнению с IL-15 дикого типа. Для антагонистов IL-15 характерны сниженная биологическая активность по сравнению с IL-15 дикого типа или способность ингибировать ответы, опосредованные IL-15. В различных воплощениях вариант IL-15 связывается с рецепторами IL-15Rβγc с повышенной или сниженной активностью. В различных воплощениях последовательность варианта IL-15 имеет по меньшей мере одно аминокислотное изменение, например замену или делецию, по сравнению с последовательностью нативного IL-15, где такие изменения приводят к агонистической или антагонистической активности относительно IL-15. В различных воплощениях аминокислотные замены/делеции присутствуют в доменах IL-15, взаимодействующих с IL-15RP и/или γс. В различных воплощениях аминокислотные замены/делеции не влияют на связывание с полипептидом IL-15Rα или способность продуцировать вариант IL-15. Подходящие аминокислотные замены/делеции для получения вариантов IL-15 могут быть определены, исходя из известных структур IL-15, сравнений IL-15 с гомологичными молекулами, такими как IL-2, с известной структурой, посредством рационального или случайного мутагенеза и функциональных анализов, предложенных здесь, или другими эмпирическими методами. Кроме того, подходящие аминокислотные замены могут представлять собой консервативные или неконсервативные изменения и вставки дополнительных аминокислот. В различных воплощениях варианты IL-15 по изобретению содержат одну или более чем одну аминокислотную замену или делецию в положении 30, 31, 32, 58, 62, 63, 67, 68 или 108 последовательности зрелого человеческого IL-15, изложенной в SEQ ID NO: 2. В различных воплощениях замены D30T («D30» относится к аминокислоте и положению остатка в нативной последовательности зрелого человеческого IL-15, и «Т» относится к заменяющему аминокислотному остатку в указанном положении варианта IL-15), V31Y, Н32Е, D62T, I68F или Q108M приводят к получению вариантов IL-15 с антагонистической активностью, а замены S58D приводят к получению вариантов IL-15 с агонистической активностью. В некоторых воплощениях вариант IL-15 содержит аминокислотную последовательность, изложенную в SEQ ID NO: 3:
Типичные конструкции Fc IL-15 VitoKine представлены в Таблице 4.


В различных воплощениях антитело IL-15 VitoKine или Fc-слитые молекулы IL-15 будут содержать две или более чем две гетеродимерные цепи, изложенные в Таблице 5.
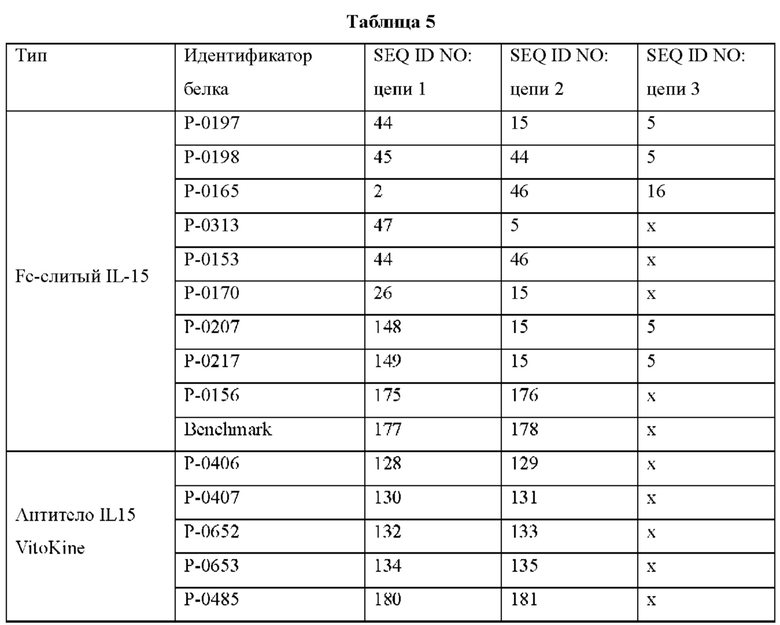
В различных воплощениях домен D2 на основе IL-15 будет содержать конструкцию IL-15, содержащую блокирующий пептид на основе IL-2Rβ, выбранную из конструкций, имеющих аминокислотные последовательности, изложенные в SEQ ID NO: 66-70.
В различных воплощениях домен D2 на основе IL-15 будет содержать конструкцию IL-15, содержащую блокирующий пептид на основе IL-2Rβ и имеющую две или более чем две гетеродимерные цепи, изложенные в Таблице 6.

IL-2
Интерлейкин-2 (IL-2), классический Th1-цитокин, продуцируют Т-клетки после активации через Т-клеточный антигенный рецептор и костимулирующую молекулу CD28. Регуляция IL-2 осуществляется посредством активации сигнальных путей и транскрипционных факторов, действующих на промотор IL-2, приводя к новой транскрипции гена, но также включает модуляцию стабильности мРНК IL-2. IL-2 связывается с рецептором, содержащим несколько цепей, включая высоко регулируемую α-цепь и β- и γ-цепи, опосредующие передачу сигналов по пути Jak-STAT. IL-2 передает сигналы активации, роста и дифференцировки Т-клеткам, В-клеткам и NK-клеткам. IL-2 также важен для опосредования гибели Т-клеток, индуцированной их активацией, и эта функция является важнейшим механизмом прекращения иммунных ответов. Имеющийся в продаже препарат негликозилированного человеческого рекомбинантного IL-2, алдеслейкин (доступный под торговым названием Пролейкин® как дезаланил-1-серии-125 человеческий интерлейкин-2 от Prometheus Laboratories Inc., Сан-Диего, Калифорния) одобрен для введения пациентам с метастатическим почечно-клеточным раком и метастатической меланомой. Также было предложено вводить IL-2 пациентам, инфицированным вирусом гепатита С (HCV), вирусом иммунодефицита человека (HIV), пациентам с острым миелолейкозом, неходжкинской лимфомой, Т-клеточной лимфомой кожи, ювенильным ревматоидным артритом, атопическим дерматитом, раком молочной железы и раком мочевого пузыря. К сожалению, оптимальное дозирование IL-2 ограничено коротким периодом полувыведения и выраженной токсичностью.
При использовании здесь в контексте белков или полипептидов термины «нативный IL-2» и «нативный интерлейкин-2» относятся к любым встречающимся в природе аминокислотным последовательностям интерлейкина-2 млекопитающих, включая как незрелые формы или формы-предшественники, так и зрелые формы. Неограничивающие примеры регистрационных номеров аминокислотных последовательностей различных разновидностей нативного интерлейкина-2 млекопитающих в GenBank включают NP_032392.1 (Mus musculus, незрелая форма), NP_001040595.1 (Macaco mulatto, незрелая форма), NP_000577.2 (человеческий, форма-предшественник), САА01199.1 (человеческий, незрелая форма), AAD48509.1 (человеческий, незрелая форма) и ААВ20900.1 (человеческий). В различных воплощениях настоящего изобретения нативный IL-2 представляет собой незрелую форму или форму-предшественника встречающегося в природе IL-2 млекопитающего. В других воплощениях нативный IL-2 представляет собой зрелую форму встречающегося в природе IL-2 млекопитающего. В различных воплощениях нативный IL-2 представляет собой форму-предшественника встречающегося в природе человеческого IL-2. В различных воплощениях нативный IL-2 представляет собой зрелую форму встречающегося в природе человеческого IL-2. В различных воплощениях домен D2 на основе IL-2 имеет происхождение от аминокислотной последовательности предшественника человеческого IL-2, изложенной в SEQ ID NO: 6:

В различных воплощениях домен D2 на основе IL-2 имеет происхождение от аминокислотной последовательности зрелой формы человеческого IL-2 дикого типа, изложенной в SEQ ID NO: 8, содержащей замену цистеина в положении 125 на серии, но не изменяющей связывание с рецептором IL-2 по сравнению с IL-2, встречающимся в природе:

В различных воплощениях домен D2 на основе IL-2 будет вариантом (или мутантом) IL-2, содержащим последовательность, имеющую происхождение от последовательности полипептида зрелого человеческого IL-2, изложенной в SEQ ID NO: 8.
В различных воплощениях вариант IL-2 содержит аминокислотную последовательность, отличную от нативного белка IL-2 (или IL-2 дикого типа). В различных воплощениях вариант IL-2 связывается с полипептидом IL-2Rα и функционирует как агонист или антагонист IL-2. В различных воплощениях варианты IL-2 с агонистической активностью имеют суперагонистическую активность. В различных воплощениях вариант IL-2 может функционировать как агонист или антагонист IL-2 независимо от его связывания с IL-2Rα. Для агонистов IL-2 характерна сопоставимая или повышенная биологическая активность по сравнению с IL-2 дикого типа. Для антагонистов IL-2 характерны сниженная биологическая активность по сравнению с IL-2 дикого типа или способность ингибировать ответы, опосредованные IL-2. В различных воплощениях последовательность варианта IL-2 имеет по меньшей мере одно аминокислотное изменение, например замену или делецию, по сравнению с последовательностью нативного IL-2, где такие изменения приводят к агонистической или антагонистической активности относительно IL-2. В различных воплощениях вариант IL-2 имеет аминокислотную последовательность, имеющую происхождение от SEQ ID NO: 8, со сниженным/устраненным связыванием с IL-2Rα для селективной активации и пролиферации эффективных Т-клеток (Teff) для лечения рака; типичные аминокислотные замены приведены в Таблице 7. В различных воплощениях вариант IL-2 имеет аминокислотную последовательность, имеющую происхождение от SEQ ID NO: 8, со сниженным связыванием с IL-2Rβ и/или γс и повышенной селективностью активации и пролиферации регуляторных Т-клеток (Treg) для лечения аутоиммунных заболеваний; типичные аминокислотные замены приведены в Таблице 7. Как будет ясно специалистам в данной области, все мутации можно, необязательно и независимо, комбинировать любым образом для достижения оптимальной аффинности и модуляции активности.

Типичные конструкции VitoKine на основе IL-2 представлены в Таблице 8.


В различных воплощениях активная группировка выбрана из группы последовательностей, состоящей из, без ограничения, аминокислотных последовательностей интерлейкина-4 (IL-4) (SEQ ID NO: 17), интерлейкина-7 (IL-7) (SEQ ID NO: 18), интерлейкина-9 (IL-9) (SEQ ID NO:19), интерлейкина-10 (IL-10) (SEQ ID NO: 20), интерлейкина-12 альфа (IL-12α) (SEQ ID NO: 21), интерлейкина-12 бета (IL-12β) (SEQ ID NO: 22), интерлейкина-23 альфа (IL-23α) (SEQ ID NO: 23) и TGFβ (SEQ ID NO: 24). В различных воплощениях активная группировка представляет собой гетеродимерный человеческий цитокин IL-12, содержащий SEQ ID NO: 21 в качестве цепи 1 и SEQ ID NO: 22 в качестве цепи 2. В различных воплощениях активная группировка представляет собой гетеродимерный человеческий цитокин IL-23, содержащий SEQ ID NO: 23 в качестве цепи 1 и SEQ ID NO: 22 в качестве цепи 2.
Домен D3 («домен маскирующей группировки»)
Домен D3 представляет собой «домен маскирующей группировки», и его используют главным образом для обратимой маскировки активности домена D2 в конкретной конструкции VitoKine. Домен D3 способен маскировать функциональную активность D2 до активации в намеченном месте терапии. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат «домен маскирующей группировки» (D3), являющийся когнатным рецептором / партнером по связыванию для белка или цитокина D2. В различных воплощениях домен D3 представляет собой вариант когнатного рецептора / партнера по связыванию или специфического связывающего агента, такого как пептид или фрагмент антитела, для домена D2. В различных воплощениях домен D3 имеет усиленное связывание с доменом D2 по сравнению с когнатным рецептором / партнером по связыванию дикого типа. В различных воплощениях домен D3 имеет сниженное связывание с доменом D2 по сравнению с когнатным рецептором / партнером по связыванию дикого типа или его связывание с доменом D2 устранено. В различных воплощениях домен D3 представляет собой белок, или пептид, или антитело, или фрагмент антитела, способные маскировать активность D2. В различных воплощениях домен D3 представляет собой ДНК, фрагмент РНК или полимер, такой как PEG, посредством расщепляемого линкера. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой внеклеточный домен IL-15Rα или его функциональный фрагмент или вариант. В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой домен IL-15RαSushi (аминокислоты 1-65 SEQ ID NO: 5). В различных предпочтительных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой домен IL-15RαSushi+, содержащий 1-30 дополнительных остатков IL-15Rα на С-конце домена Sushi (например, SEQIDNO:5). В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой внеклеточный домен IL-2Rα или его функциональный фрагмент. В различных предпочтительных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой домен IL-2RαSushi. В различных воплощениях домен D3 способен маскировать функциональную активность D2 до активации в намеченном месте терапии.
Альфа-рецептор IL-15
Рецептор IL-15 представляет собой рецептор цитокинов 1-го типа, состоящий из бета(β)- и гамма(γ)-субъединиц, общих для него и рецептора IL-2, и альфа(α)-субъединицы, связывающейся с IL-15 с высокой аффинностью. Полноразмерный человеческий IL-15Rα представляет собой трансмембранный белок 1-го типа с сигнальным пептидом из 32 аминокислот, внеклеточным доменом из 173 аминокислот, транс мембранным доменом из 21 аминокислоты, цитоплазматическим концевым сегментом из 37 аминокислот и множеством сайтов N- или О-гликозилирования (Anderson et al., J. Biol Chem, 270:29862 - 29869, 1995). Ранее было продемонстрировано, что естественная растворимая форма альфа-цепи IL-15R, соответствующая полноразмерному внеклеточному домену IL-15R альфа, действует как высокоаффинный антагонист IL-15. Тем не менее, в резком противоречии с этими данными, было продемонстрировано, что рекомбинантный растворимый sushi-домен IL-15R альфа, обладающий большей частью аффинности связывания в отношении IL-15, действует как сильный агонист IL-15, усиливая его связывание с гетеродимером IL-15R бета/гамма и оказываемые через него биологические эффекты, не влияя в то же время на связывание трехкомпонентного мембранного рецептора IL-15R альфа/бета/гамма с IL-15 и его функцию. Эти результаты позволяют предположить, что в естественных условиях такие sushi-домены могут быть вовлечены в механизм транспрезентации IL-15 (Mortier et al., J. Biol Chem, 281(3): 1612-1619, 2006).
При использовании здесь в контексте белков или полипептидов термины «нативный IL-15Rα» и «нативный альфа-рецептор интерлейкина-15» относятся к любой встречающейся в природе аминокислотной последовательности альфа-рецептора интерлейкина-15 («IL-15Rα») млекопитающих, включая как незрелые формы или формы-предшественники, так и зрелые формы, и встречающиеся в природе изоформы. Неограничивающие примеры регистрационных номеров аминокислотных последовательностей различных нативных IL-15Rα млекопитающих в GenBank включают NP_002180 (человек), АВК41438 (Масаса mulatto), NP_032384 (Mus musculus), Q60819 (Mus musculus), CA141082 (человек). В различных воплощениях нативный IL-15Rα представляет собой незрелую форму встречающегося в природе полипептида IL-15Rα млекопитающего. В различных воплощениях нативный IL-15Rα представляет собой зрелую форму встречающегося в природе полипептида IL-15Rα млекопитающего. В различных воплощениях нативный IL-15Rα представляет собой форму встречающегося в природе полипептида IL-15Rα млекопитающего. В различных воплощениях нативный IL-15Rα представляет собой полноразмерную форму встречающегося в природе полипептида IL-15Rα млекопитающего. В различных воплощениях нативный IL-15Rα представляет собой незрелую форму встречающегося в природе полипептида человеческого IL-15Rα. В различных воплощениях нативный IL-15Rα представляет собой зрелую форму встречающегося в природе полипептида человеческого IL-15Rα. В различных воплощениях нативный IL-15Rα представляет собой полноразмерную форму встречающегося в природе полипептида человеческого IL-15Rα. В различных воплощениях белок или полипептид нативного IL-15Rα является выделенным или очищенным. В различных воплощениях домен IL-15Rα имеет происхождение от аминокислотной последовательности человеческого IL-15Rα, изложенной в SEQ ID NO:4:

В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой домен IL-15RαSushi+, содержащий аминокислотную последовательность полипептида зрелого человеческого IL-15Rα, изложенную в SEQ ID NO: 5:
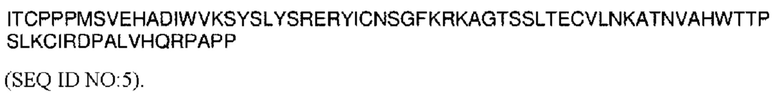
В различных воплощениях IL-15RαSushi+(SEQ ID NO: 5), усеченный когнатный корецептор IL-15, воспроизводящий большую часть аффинности связывания полноразмерного IL-15Rα (SEQ ID NO: 4), использовали в качестве домена D3 для маскировки активности IL-15, подбирая расщепляемый или нерасщепляемый линкер, соединяющий IL-15 и IL-15RαShushi+, с получением IL-15 VitoKine. Специалисту в данной области будет ясно, что длина домена D3 может отличаться от последовательности, изложенной в SEQIDNO:5, при условии, что он может воспроизводить большую часть связывающей активности полноразмерного IL-15Rα (SEQ ID NO: 4), то есть является его функциональным фрагментом. Особенность конструкции IL-15 VitoKine состоит в полном использовании уникальных свойств пути IL-15, включая исключительно высокую аффинность между IL-15 и IL-15α (30 пМ) и то, что образование комплексов с IL-15α усиливает активность IL-15 in vivo. После расщепления линкера, соединяющего IL-15 и IL-15αSushi+, протеазами, регуляция которых повышена в очаге заболевания, IL-15RαShushi+ или любой его функциональный фрагмент, имеющий происхождение от IL-15RαECD, будет, предположительно, сохранять нековалентную связь с IL-15 и усиливать активность IL-15.
Рецептор IL-2
Рецептор IL-2 (IL-2R) представляет собой гетеротримерный белок, экспрессируемый на поверхности определенных иммунных клеток, таких как лимфоциты, связывающий и отвечающий на цитокин, называемый IL-2. IL-2R имеет три субъединицы: α (CD25), β (CD122) и γс (CD132, общая цепь, входящая в состав пяти других цитокиновых рецепторов: IL-4R, IL-7R, IL-9R, IL-15R и IL-21R). Альфа-цепь (также называемая Тас-антигеном или р55) человеческого рецептора кодирована на хромосоме 10р14-15 геном IL-2RA. Ген человеческой β-цепи (IL-2RB, CD 122) данного рецептора расположен на хромосоме 22q11.2-12, в то время как ген общей цепи IL-2Rγc (IL-2RG) расположен на хромосоме Xq13. Сборка всех трех субъединиц рецептора важна для трансдукции сигналов в В- и Т-клетках. IL-2R присутствует (временно или постоянно) на поверхности почти всех кроветворных клеток, включая лимфоидные линии Т-, В- и NK-клеток, а также миелоидные линии, такие как макрофаги, моноциты и нейтрофилы. Передача сигналов в клетки происходит через Янус-киназы Jak1 и Jak3. Фосфорилирование внутрицитозольной части β-цепи рецептора приводит к возможности образования гомодимеров факторов STAT-3 и STAT-5. Гомодимеры STAT-3 и STAT-5 демонстрируют повышенную аффинность в отношении ядра, где они связываются с определенными ДНК-элементами, усиливающими транскрипцию IL-2-зависимых генов.
При использовании здесь в контексте белков или полипептидов термины «нативный IL-2Rα» и «нативный альфа-рецептор интерлейкина-2» относятся к любой встречающейся в природе аминокислотной последовательности альфа-рецептора интерлейкина-2 («IL-2Rα») млекопитающих, включая как незрелые формы или формы-предшественники, так и зрелые формы, и встречающиеся в природе изоформы. Неограничивающие примеры регистрационных номеров аминокислотных последовательностей различных нативных IL-2Rα млекопитающих в GenBank включают NP_032393.3 (Мня musculus), САК26553.1 (человек) и NP_000408.1 (человек). В различных воплощениях нативный IL-2Rα представляет собой незрелую форму встречающегося в природе полипептида IL-2Rα млекопитающего. В различных воплощениях нативный IL-2Rα представляет собой зрелую форму встречающегося в природе полипептида IL-2Rα млекопитающего. В различных воплощениях нативный IL-2Rα представляет собой форму встречающегося в природе полипептида IL-2Rα млекопитающего. В различных воплощениях нативный IL-2Rα представляет собой полноразмерную форму встречающегося в природе полипептида IL-2Rα млекопитающего. В различных воплощениях нативный IL-2Rα представляет собой незрелую форму встречающегося в природе полипептида человеческого IL-2Rα. В различных воплощениях нативный IL-2Rα представляет собой зрелую форму встречающегося в природе полипептида человеческого IL-2Rα. В различных воплощениях нативный IL-2Rα представляет собой полноразмерную форму встречающегося в природе полипептида человеческого IL-2Rα. В различных воплощениях белок или полипептид нативного IL-2Rα является выделенным или очищенным. В различных воплощениях домен IL-2Rα имеет происхождение от аминокислотной последовательности человеческого IL-2Rα, изложенной в SEQ ID NO: 9:

В различных воплощениях конструкции VitoKine по настоящему изобретению содержат домен D3, представляющий собой домен IL-2RαSushi, содержащий аминокислотную последовательность полипептида зрелого человеческого IL-2Rα, изложенную в SEQ ID NO: 10:

В различных воплощениях IL-2RαSushi (SEQ ID NO: 10) был использован для маскировки активности IL-2 с получением IL-2 VitoKine. В отличие от IL-15Rα, содержащего один sushi-домен, IL-2Rα содержит два sushi-домена, разделенные линкерной областью. В различных воплощениях IL-2 VitoKine содержит вариант IL-2RαSushi, содержащий аминокислотные замены для нарушения специфичных нековалентных взаимодействий между IL-2Rα и IL-2, снижая посредством этого аффинность связывания IL-2Rα с IL-2. Несмотря на то, что нативный IL-2Rα связывается с IL-2 с умеренной аффинностью, составляющей 30 нМ, есть вероятность, что IL-2Rα может не диссоциировать после расщепления линкера. Связывание IL-2Rα с IL-2 может снижать активность IL-2 и/или смещать баланс субпопуляций Т-клеток в нежелательном направлении. В случае введения в IL-2RαSushi мутации (мутаций), снижающей аффинность, например, К38Е или Y43A, или комбинации этих двух замен высока вероятность диссоциации sushi-доменов IL-2Rα от IL-2 после расщепления линкера протеазой.
Линкеры L1 и L2
Расщепляемые линкеры
Расщепляемый линкер или линкер, чувствительный к ассоциированному с болезнью ферменту, может содержать группировку, например белковый субстрат, которая может быть специфично расщеплена протеазой, уровни которой в очаге заболевания повышены по сравнению с тканями, не пораженными заболеванием. В литературе есть сообщения о повышении уровней ферментов, имеющих известные субстраты, при различных типах рака, например при солидных опухолях. См., например, La Rocca etal, Brit. J. Cancer 90:1414-1421 и Ducry et al., Bioconjug. Chem. 21:5-13, 2010, каждая из которых полностью включена сюда посредством ссылки. В различных воплощениях протеаза, способная расщеплять линкер, расщепляемый протеазой, выбрана из группы, состоящей из металл о протеиназ, например матриксных металлопротеиназ (ММР) 1-28, сериновых протеаз, например активатора плазминогена урокиназного типа (иРА) и матриптазы, цистеиновых протеаз, например легумаина, аспартатных протеаз и катепсиновых протеаз. Пептидные последовательности типичных субстратов протеаз представлены в Таблице 9.


Пептидные последовательности типичных субстратов протеаз, которые могут быть использованы в качестве линкеров, расщепляемых протеазами, с пептидными спейсерами различной длины или без них на С-конце, или на N-конце, или на обоих концах домена D2, представлены в Таблице 10.

В различных воплощениях протеаза представляет собой ММР-9 или ММР-2. В другом конкретном воплощении протеаза представляет собой uPA. В другом конкретном воплощении протеаза представляет собой ММР-14. В другом конкретном воплощении протеаза представляет собой легумаин. В различных воплощениях одна молекула VitoKine содержит две разные протеазы. В различных воплощениях линкер, расщепляемый протеазой, содержит последовательность распознавания протеазы «GPLGMLSQ» (SEQ ID NO: 77). В различных воплощениях линкер, расщепляемый протеазой, содержит последовательность распознавания протеазы «LGGSGRSANAILE» (SEQ ID NO: 80). В различных воплощениях линкер, расщепляемый протеазой, содержит последовательность распознавания протеазы «SGRSENIRTA» (SEQ ID NO: 157). В различных воплощениях линкер, расщепляемый протеазой, содержит последовательность распознавания протеазы «GPTNKVR» (SEQ ID NO: 158). В различных воплощениях возможно расщепление линкера (например, расщепляемого линкера) опухоль-ассоциированными протеазами. В различных воплощениях, при заболеваниях, отличных от рака, таких как воспалительные заболевания, возможно расщепление расщепляемого линкера другими специфичными в отношении болезни протеазами.
В различных воплощениях возможно включение пептидных спейсеров с любой стороны от последовательности, расщепляемой протеазой, или для фланкирования последовательности, расщепляемой протеазой, с обеих сторон, или в качестве нерасщепляемого линкера без сайта субстрата протеазы. Пептидный спейсер используют для расположения расщепляемого линкера, повышающего его доступность для фермента, обеспечивающего расщепление. Длина спейсеров может быть изменена или оптимизирована для соблюдения баланса доступности для ферментативного расщепления и пространственных ограничений, необходимых для обратимой маскировки биологической активности домена D2. Спейсер может содержать 1-100 аминокислот. Подходящие пептидные спейсеры известны в данной области и включают, без ограничения, пептидные линкеры, содержащие гибкие аминокислотные остатки, такие как глицин и серии. В различных воплощениях спейсер может содержать мотивы GS, GGS, GGGGS, GGSG или SGGG. В различных воплощениях спейсер может содержать от 1 до 12 аминокислот, включая мотивы G, S, GS (SEQ ID NO: 116), GGS (SEQ ID NO: 117), GSGS (SEQ ID NO: 121), GSGSGS (SEQ ID NO: 122), GSGSGSGS (SEQ ID NO: 123), GSGSGSGSGS (SEQ ID NO: 124) или GSGSGSGSGSGS (SEQ ID NO: 125). В других воплощениях спейсер может содержать мотивы (GGGGS)n, где n представляет собой целое число от 1 до 10. В других воплощениях спейсер может также содержать аминокислоты, отличные от глицина и серина.
Типичные линкеры, расщепляемые протеазами, со спейсерными пептидами, фланкирующими пептид субстрата протеазы (подчеркнуты), представлены в Таблице 11.

В различных воплощениях возможна активация расщепляемого линкера механизмами, отличными от протеолиза, включая, без ограничения, гидролиз, такой как в случае пегилированного полимера, обеспечивающего высвобождение посредством контролируемого механизма при различных значениях рН.
Нерасщепляемые линкеры
Нерасщепляемый линкер обеспечивает ковалентное связывание и дополнительную структурную и/или пространственную гибкость между белковыми доменами. Как известно в данной области, в качестве нерасщепляемых линкеров могут быть использованы пептидные линкеры, содержащие гибкие аминокислотные остатки, такие как глицин и серии. В различных воплощениях нерасщепляемый линкер может содержать 1-100 аминокислот. В различных воплощениях спейсер может содержать мотивы GS (SEQ ID NO: 116), GGS (SEQ ID NO: 117), GGGGS (SEQ ID NO: 118), GGSG (SEQ ID NO: 119) или SGGG (SEQ ID NO: 120). В других воплощениях линкер может содержать мотивы (GGGGS)n, где n представляет собой целое число от 1 до 10. В других воплощениях линкер может также содержать аминокислоты, отличные от глицина и серина. В другом воплощении нерасщепляемый линкер может представлять собой простую химическую связь, например, амидную связь (например, при химической конъюгации PEG). Нерасщепляемый линкер стабилен в физиологических условиях, а также в очаге заболевания, таком как раковый очаг или очаги воспалительных заболеваний.
Типичные нерасщепляемые линкеры представлены в Таблице 12.

Комбинация расщепляемого и нерасщепляемого линкера
В различных воплощениях оба линкера, L1 и L2, могут быть расщепляемыми или нерасщепляемыми или представлять собой комбинацию расщепляемого и нерасщепляемого линкера с получением различных форм активной группировки домена D2 для решения различных терапевтических задач или для обеспечения сбалансированного соотношения пользы и риска или различных свойств цитокинов. Типичные активные формы, высвобождаемые при расщеплении линкеров, показаны на ФИГ. 2. Активные формы 1 и 3, полученные в результате расщепления L1 и как L1, так и L2, соответственно, являются короткодействующими цитокинами с различной степенью функциональной активности в зависимости от конформации D3. Отщепление и отделение от группировки D1, увеличивающей период полувыведения или направленной на ткань, пораженную заболеванием, будет повышать местные концентрации активированного домена D2. После оказания местного действия короткодействующие активные формы могут быть быстро выведены из системной циркуляции для снижения токсичности. В отличие от этого активная форма 2, полученная в результате расщепления L2, представляет собой длительнодействующий цитокин с полностью восстановленной функцией и сохранной тканенаправленностью, устойчиво удерживаемый в очаге заболевания, обеспечивая более продолжительную и высокую эффективность.
Полинуклеотиды
В другом аспекте согласно настоящему изобретению предложены выделенные молекулы нуклеиновых кислот, содержащие полинуклеотид, кодирующий IL-15, вариант IL-15, IL-15Rα, вариант IL-15Rα, Fc, вариант Fc, слитый белок IL-15-Fc, слитый белок IL-15RαSushi-Fc или конструкцию VitoKine по настоящему изобретению. Рассматриваемые нуклеиновые кислоты могут быть одноцепочечными или двуцепочечными. Такие нуклеиновые кислоты могут представлять собой молекулы ДНК или РНК. ДНК включает, например, кДНК (комплементарная ДНК), геномную ДНК, синтетическую ДНК, ДНК, амплифицированную посредством PCR (полимеразная цепная реакция), и их комбинации. Геномную ДНК, кодирующую конструкции VitoKine, получают из геномных библиотек, доступных для ряда видов. Синтетическая ДНК может быть получена химическим синтезом перекрывающихся олигонуклеотидных фрагментов с последующей сборкой фрагментов для восстановления части или всех кодирующих областей и фланкирующих последовательностей. РНК может быть получена с использованием прокариотических векторов экспрессии, приводящих к высоким уровням синтеза мРНК, таких как векторы с промоторами и РНК-полимеразой Т7. Молекулы ДНК по изобретению включают полноразмерные гены, а также полинуклеотиды и их фрагменты. Полноразмерный ген может также содержать последовательности, кодирующие N-концевую сигнальную последовательность. Такие нуклеиновые кислоты могут быть использованы, например, в способах получения новых конструкций VitoKine.
В различных воплощениях выделенные молекулы нуклеиновых кислот содержат полинуклеотиды, описанные здесь, и дополнительно содержат полинуклеотид, кодирующий по меньшей мере один гетерологичный белок, описанный здесь. В различных воплощениях молекулы нуклеиновых кислот дополнительно содержат полинуклеотиды, кодирующие линкеры или шарнирные линкеры, описанные здесь.
В различных воплощениях рекомбинантные нуклеиновые кислоты по настоящему изобретению могут быть функционально связаны с одной или более чем одной регуляторной нуклеотидной последовательностью в экспрессионной конструкции. Регуляторные последовательности известны в данной области, и их выбирают для управления экспрессией конструкции VitoKine. Соответственно, термин регуляторная последовательность включает промоторы, энхансеры и другие элементы контроля экспрессии. Типичные регуляторные последовательности описаны в Goeddel; Gene Expression Technology: Methods in Enzymology, Academic Press, San Diego, Calif. (1990). Обычно указанная одна или более чем одна регуляторная нуклеотидная последовательность может включать, без ограничения, промоторные последовательности, лидерные или сигнальные последовательности, сайты связывания рибосом, последовательности начала и терминации транскрипции, последовательности начала и терминации трансляции и энхансерные или активаторные последовательности. Настоящее изобретение предполагает конститутивные или индуцируемые промоторы, известные в данной области. Промоторы могут представлять собой промоторы, встречающиеся в природе, или гибридные промоторы, в которых совмещены элементы более чем одного промотора. Экспрессионная конструкция может присутствовать в клетке на эписоме, такой как плазмида, или экспрессионная конструкция может быть введена в хромосому. В различных воплощениях вектор экспрессии содержит селективный маркерный ген, позволяющий проводить селекцию трансформированных клеток-хозяев. Селектируемые маркерные гены хорошо известны в данной области и будут варьировать в зависимости от используемой клетки-хозяина.
В другом аспекте настоящего изобретения рассматриваемая нуклеиновая кислота представлена в векторе, содержащем нуклеотидную последовательность, кодирующую конструкцию VitoKine и функционально связанную по меньшей мере с одной регуляторной последовательностью. Термин «вектор экспрессии» относится к плазмиде, фагу, вирусу или вектору для экспрессии полипептида с полинуклеотидной последовательности. Векторы, подходящие для экспрессии в клетке-хозяине, легкодоступны, и молекулы нуклеиновых кислот вводят в векторы с применением стандартных методик рекомбинантных ДНК. Такие векторы могут содержать широкий спектр последовательностей контроля экспрессии, которые, будучи функционально связанными с последовательностью ДНК, контролируют ее экспрессию и могут быть использованы в этих векторах для экспрессии последовательностей ДНК, кодирующих конструкцию VitoKine. Такие полезные последовательности контроля экспрессии включают, например, ранний и поздний промоторы SV40, промотор tet, предранний промотор аденовируса или цитомегаловируса, промоторы RSV, систему lac, систему trp, систему ТАС или TRC, промотор Т7, экспрессию которого обеспечивает РНК-полимераза Т7, основные операторные и промоторные области фага лямбда, контрольные области белка оболочки fd, промотор 3-фосфоглицераткиназы или других гликолитических ферментов, промоторы кислой фосфатазы, например, PhoS, промоторы а-факторов спаривания дрожжей, полиэдриновый промотор бакуловирусной системы и другие известные последовательности, контролирующие экспрессию генов прокариотических или эукариотических клеток или их вирусов, и различные их комбинации. Следует понимать, что структура вектора экспрессии может зависеть от таких факторов, как выбор клетки-хозяина, которую следует трансформировать, и/или тип белка, который желательно экспрессировать. Более того, следует учитывать число копий вектора, возможность контролировать это число копий и экспрессию любых других белков, кодируемых вектором, таких как антибиотические маркеры. Типичными векторами экспрессии, подходящими для экспрессии VitoKine, являются pDSRa и его производные, содержащие полинуклеотиды VitoKine, а также любые дополнительные подходящие векторы, известные в данной области или описанные ниже.
Рекомбинантная нуклеиновая кислота по настоящему изобретению может быть получена лигированием клонированного гена или его части в вектор, подходящий для экспрессии в прокариотических клетках и/или эукариотических клетках (клетках дрожжей, птиц, насекомых или млекопитающих). Экспрессионные носители для получения рекомбинантной конструкции VitoKine включают плазмиды и другие векторы. Например, подходящие векторы включают плазмиды следующих типов: плазмиды, имеющие происхождение от pBR322, плазмиды, имеющие происхождение от pEMBL, плазмиды, имеющие происхождение от рЕХ, плазмиды, имеющие происхождение от рВТас, и плазмиды, имеющие происхождение от pUC, для экспрессии в прокариотических клетках, таких как Е. coli.
Некоторые векторы экспрессии для клеток млекопитающих содержат как прокариотические последовательности, способствующие воспроизведению вектора у бактерий, так и одну или более чем одну эукариотическую транскрипционную единицу, экспрессируемую в эукариотических клетках. Примерами векторов экспрессии для клеток млекопитающих, подходящих для трансфекции эукариотических клеток, являются векторы, имеющие происхождение от pcDNAI/amp, pcDNAI/neo, pRc/CMV, pSV2gpt, pSV2neo, pSV2-dhfr, pTk2, pRSVneo, pMSG, pSVT7, pko-neo и pHyg. Некоторые из этих векторов модифицируют, используя последовательности из бактериальных плазмид, таких как pBR322, для облегчения репликации и отбора по лекарственной устойчивости как в прокариотических, так и в эукариотических клетках. Альтернативно, для транзиторной экспрессии белков в эукариотических клетках могут быть использованы производные вирусов, таких как вирус папилломы крупного рогатого скота (BPV-1) или вирус Эпштейна-Барр (рНЕВо, имеющий происхождение от pREP и р205). Примеры вирусных (включая ретровирусные) систем экспрессии приведены ниже при описании систем доставки для генной терапии. Различные методы, применяемые при получении плазмид и трансформации организмов-хозяев, хорошо известны в данной области. Другие подходящие системы экспрессии как для прокариотических, так и для эукариотических клеток, а также общие рекомбинантные методики приведены в Molecular Cloning А Laboratory Manual, 2nd Ed., ed. by Sambrook, Fritsch and Maniatis (Cold Spring Harbor Laboratory Press, 1989) Chapters 16 and 17. В некоторых случаях может быть желательным экспрессировать рекомбинантные полипептиды с использованием бакуловирусной системы экспрессии. Примеры таких бакул о вирусных систем экспрессии включают векторы, имеющие происхождение от pVL (такие как pVL1392, pVL1393 и pVL941), векторы, имеющие происхождение от pAcUW (такие как pAcUW1), и векторы, имеющие происхождение от pBlueBac (такие как pBlueBac III, содержащий B-gal).
В различных воплощениях будет разработан вектор для получения рассматриваемых конструкций VitoKine в клетках СНО, такой как вектор Pcmv-Script (Stratagene, Ла-Холья, штат Калифорния), векторы pcDNA4 (bivitrogen, Карлсбад, штат Калифорния) и векторы pCI-neo (Promega, Мэдисон, штат Висконсин). Как будет ясно, рассматриваемые генные конструкции могут быть использованы для экспрессии рассматриваемых конструкций VitoKine в культивированных клетках, например для получения белков, включая слитые белки или варианты белков, для очистки.
Настоящее изобретение также относится к клетке-хозяину, трансфицированной рекомбинантным геном, содержащим нуклеотидную последовательность, кодирующую аминокислотную последовательность одной или более чем одной рассматриваемой конструкции VitoKine. Клетка-хозяин может представлять собой любую прокариотическую или эукариотическую клетку. Например, конструкция VitoKine по настоящему изобретению может быть экспрессирована в бактериальных клетках, таких как Е. coli, клетках насекомых (например, с использованием бакуловирусной системы экспрессии), клетках дрожжей или клетках млекопитающих. Специалистам в данной области известны другие подходящие клетки, такие как клетки яичника китайского хомяка (Chinese Hamster Ovary, СНО) или клетки эмбриональной почки человека 293 (Human Embryonic Kidney 293, НЕК293).
Соответственно, настоящее изобретение также относится к способам получения рассматриваемых конструкций VitoKine. Например, клетку-хозяина, трансфицированную вектором экспрессии, кодирующим конструкцию VitoKine, можно культивировать в подходящих условиях, обеспечивающих возможность экспрессии конструкции VitoKine. Конструкция VitoKine может быть секретирована и выделена из смеси клеток и среды, содержащей конструкцию VitoKine. Альтернативно, конструкция VitoKine может оставаться в цитоплазматической или мембранной фракции, и может быть проведен сбор и лизис клеток с последующим выделением белка. Клеточная культура содержит клетки-хозяева, среды и другие побочные продукты. Подходящие среды для культивирования клеток хорошо известны в данной области.
Полипептиды и белки по настоящему изобретению могут быть очищены в соответствии с методиками очистки белка, хорошо известными специалистам в данной области. Эти методики включают, на одном уровне, разделение неочищенных белковых и небелковых фракций. После отделения пептидов и полипептидов от других белков может быть проведена дальнейшая очистка интересующего пептида или полипептида с применением хроматографических и электрофоретических методик с достижением частичной или полной очистки (или очистки до однородности). При использовании здесь термин «выделенный полипептид» или «очищенный полипептид» предназначен для обозначения композиции, которая может быть отделена от других компонентов, где полипептид очищен в любой степени по сравнению его естественным состоянием. Таким образом, очищенный полипептид также относится к полипептиду, свободному от среды, в которой он может встречаться в природе. Обычно «очищенный» будет относиться к полипептидной композиции, которая была разделена на фракции для удаления различных других компонентов и практически сохраняет свою выраженную биологическую активность. Там, где использован термин «практически очищенный», это обозначение будет относиться к пептидной или полипептидной композиции, где полипептид или пептид является основным компонентом композиции, составляя приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 85%, или приблизительно 90%, или более от белков композиции.
Различные методики, подходяще для применения при очистке, будут хорошо известны специалистам в данной области. Они включают, например: преципитацию с сульфатом аммония, PEG, антителами (иммунопреципитация) и тому подобным или посредством термической денатурации с последующим центрифугированием; хроматографию, такую как аффинная хроматография (колонки с белком А), ионообменная хроматография, гель-фильтрация, хроматография с обращенной фазой, хроматография на гидроксиапатите, хроматография гидрофобного взаимодействия; изоэлектрическое фокусирование; гель-электрофорез; и комбинации этих методик. Как общеизвестно в данной области, полагают, что порядок проведения различных стадий очистки можно менять, или что определенные стадии можно опустить, и при этом метод все еще будет подходящим для получения практически очищенного полипептида.
Фармацевтические композиции
В другом аспекте согласно настоящему изобретению предложена фармацевтическая композиция, содержащая конструкции VitoKine в смеси с фармацевтически приемлемым носителем. Такие фармацевтически приемлемые носители хорошо известны и понятны специалистам в данной области и описаны во многих источниках (см., например, Remington's Pharmaceutical Sciences, 18th Edition, A. R. Gennaro, ed., Mack Publishing Company, 1990). Фармацевтически приемлемые носители могут быть включены для модификации, поддержания или сохранения, например, рН, осмолярности, вязкости, прозрачности, цвета, изотоничности, запаха, стерильности, стабильности, скорости растворения или высвобождения, всасывания или проникновения композиции. Такие фармацевтические композиции могут влиять на физическое состояние, стабильность, скорость высвобождения полипептида in vivo и скорость клиренса полипептида in vivo. Подходящие фармацевтически приемлемые носители включают, без ограничения: аминокислоты (такие как глицин, глутамин, аспарагин, аргинин или лизин); антимикробные агенты; антиоксиданты (такие как аскорбиновая кислота, сульфит натрия или гидросульфит натрия); буферы (такие как борат, бикарбонат, трис-HCl, цитраты, фосфаты, другие органические кислоты); объемообразующие агенты (такие как маннит или глицин); хелатирующие агенты (такие как этилендиаминтетрауксусная кислота (EDTA)); комплексообразующие агенты (такие как кофеин, поливинилпирролидон, бета-циклодекстрин или гидроксипропил-бета-циклодекстрин); наполнители; моносахариды, дисахариды и другие углеводы (такие как глюкоза, манноза или декстрины); белки (такие как сывороточный альбумин, желатин или иммуноглобулины); красители; корригенты и разбавители; эмульгаторы; гидрофильные полимеры (такие как поливинилпирролидон); полипептиды с малой молекулярной массой; солеобразующие противоионы (такие как натрий); консерванты (такие как хлорид бензалкония, бензойная кислота, салициловая кислота, тимеросал, фенилэтиловый спирт, метилпарабен, пропил пар абен, циклодекстрин, сорбиновая кислота или перекись водорода); растворители (такие как глицерин, пропиленгликоль или полиэтиленгликоль); сахарные спирты (такие как маннит или сорбит); суспендирующие агенты; поверхностно-активные вещества или смачивающие агенты (такие как плюроники, PEG, эфиры сорбита, полисорбаты, такие как полисорбат 20, полисорбат 80, тритон, трометамин, лецитин, холестерин, тилоксапал); агенты, повышающие стабильность (сахароза или сорбит); агенты, усиливающие тоничность (такие как галогениды щелочных металлов (предпочтительно хлорид натрия или калия), маннит, сорбит); наполнители для доставки; разбавители; эксципиенты; и/или фармацевтические адъюванты.
Основной наполнитель или носитель в фармацевтической композиции может иметь водную или неводную природу. Например, подходящий наполнитель или носитель может представлять собой воду для инъекций, физиологический раствор или искусственную спинномозговую жидкость, возможно, дополненные другими веществами, часто используемыми в композициях для парентерального введения. Другими типичными наполнителями являются нейтральный забуференный физиологический раствор или физиологический раствор, смешанный с сывороточным альбумином. Другие типичные фармацевтические композиции содержат трис-буфер с рН приблизительно 7,0-8,5 или ацетатный буфер с рН приблизительно 4,0-5,5, которые могут дополнительно содержать сорбит или его подходящий заменитель. В одном воплощении настоящего изобретения композиции могут быть подготовлены к хранению смешиванием выбранной композиции, имеющей желаемую степень чистоты, с дополнительными агентами (Remington's Pharmaceutical Sciences, supra) в форме лиофилизированной массы или водного раствора. Кроме того, терапевтическая композиция может быть изготовлена в форме лиофилизата с использованием подходящих эксципиентов, таких как сахароза. Оптимальная фармацевтическая композиция будет определена специалистом в данной области в зависимости от, например, намеченного пути введения, формата доставки и желаемой дозы.
Когда предполагается парентеральное введение, терапевтические фармацевтические композиции могут иметь форму апирогенного водного раствора, приемлемого для парентерального введения, содержащего желаемую конструкцию VitoKine в фармацевтически приемлемом наполнителе. Особенно подходящим наполнителем для парентеральной инъекции является стерильная дистиллированная вода, где полипептид включен в композицию в форме стерильного изотонического раствора с нужным консервантом (консервантами). В различных воплощениях фармацевтические композиции, подходящие для инъекционного введения, могут быть изготовлены в водных растворах, предпочтительно в физиологически совместимых буферах, таких как раствор Хэнкса, раствор Рингера или забуференный физиологический раствор. Водные суспензии для инъекций могут содержать вещества, повышающие вязкость суспензии, такие как карбоксиметилцеллюлоза натрия, сорбит или декстран. Кроме того, суспензии активных соединений могут быть получены в форме подходящих масляных суспензий для инъекций. Возможно, суспензия может также содержать подходящие стабилизаторы или агенты для повышения растворимости соединений, позволяя получать высококонцентрированные растворы.
В различных воплощениях терапевтические фармацевтические композиции могут быть изготовлены для направленной доставки с использованием коллоидной дисперсионной системы. Коллоидные дисперсионные системы включают макромолекулярные комплексы, нанокапсулы, микросферы, гранулы и системы на основе липидов, включая эмульсии типа «масло в воде», мицеллы, смешанные мицеллы и липосомы. Примеры липидов, полезных при получении липосом, включают фосфатидильные соединения, такие как фосфатидилглицерин, фосфатидилхолин, фосфатидилсерин, фосфатидилэтаноламин, сфинголипиды, цереброзиды и ганглиозиды. Типичные фосфолипиды включают фосфатидилхолин яйца, дипальмитоилфосфатидилхолин и дистеароилфосфатидилхолин. Также возможно направленное действие липосом на основе, например, их специфичности в отношении органов, клеток и органелл, известное в данной области.
В различных воплощениях предполагается пероральное введение фармацевтических композиций. Фармацевтические композиции, вводимые таким образом, могут быть изготовлены с теми носителями, которые обычно используют при изготовлении твердых лекарственных форм, таких как таблетки и капсулы, или без таких носителей. В твердых лекарственных формах для перорального введения (капсулах, таблетках, пилюлях, драже, порошках, гранулах и тому подобных) одно или более чем одно терапевтическое соединение по настоящему изобретению может быть смешано с одним или более чем одним фармацевтически приемлемым носителем, таким как цитрат натрия или гидрофосфат кальция, и/или любым из следующего: (1) наполнителей или заполнителей, таких как крахмалы, лактоза, сахароза, глюкоза, маннит и/или кремниевая кислота; (2) связывающих агентов, таких как, например, карбоксиметилцеллюлоза, альгинаты, желатин, поливинилпирролидон, сахароза и/или аравийская камедь; (3) увлажнителей, таких как глицерин; (4) разрыхлителей, таких как агар-агар, карбонат кальция, картофельный или тапиоковый крахмал, альгиновая кислота, определенные силикаты и карбонат натрия; (5) агентов, замедляющих растворение, таких как парафин; (6) агентов, ускоряющих всасывание, таких как четвертичные аммониевые соединения; (7) смачивающих агентов, таких как, например, цетиловый спирт и моностеарат глицерина; (8) абсорбентов, таких как каолин и бентонитовая глина; (9) смазывающих агентов, таких как тальк, стеарат кальция, стеарат магния, твердые полиэтиленгликоли, лаурилсульфат натрия и их смеси; и (10) красителей. В случае капсул, таблеток и пилюль фармацевтические композиции могут также содержать буферные агенты. Твердые композиции сходного типа могут также быть использованы в качестве наполнителей в мягких и твердых желатиновых капсулах с использованием таких эксципиентов, как лактоза или молочные сахара, а также полиэтиленгликоли с высокой молекулярной массой и тому подобное. Жидкие лекарственные формы для перорального введения включают фармацевтически приемлемые эмульсии, микроэмульсии, растворы, суспензии, сиропы и эликсиры. В дополнение к активному ингредиенту, жидкие лекарственные формы могут содержать инертные разбавители, часто используемые в данной области, такие как водные или иные растворители, солюбилизирующие агенты и эмульгаторы, такие как этиловый спирт, изопропиловый спирт, этилкарбонат, этилацетат, бензиловый спирт, бензилбензоат, пропиленгликоль, 1,3-бутиленгликоль, масла (в частности хлопковое, арахисовое, кукурузное, зародышевое, оливковое, касторовое и кунжутное масло), глицерин, тетрагидрофуриловый спирт, полиэтиленгликоли, сорбитановые эфиры жирных кислот и их смеси. Помимо инертных разбавителей, композиции для перорального введения могут также содержать адъюванты, такие как смачивающие агенты, эмульгаторы и суспендирующие агенты, подсластители, корригенты, красители, отдушки и консерванты.
В различных воплощениях предполагается местное введение фармацевтических композиций на кожу или на слизистые оболочки. Композиции для местного введения могут дополнительно содержать один или более чем один из широкого спектра известных агентов, эффективно усиливающих проникновение через кожу или роговой слой. Их примерами являются 2-пирролидон, М-метил-2-пирролидон, диметилацетамид, диметилформамид, пропиленгликоль, метиловый или изопропиловый спирт, диметилсульфоксид и азон. Кроме того, могут быть включены дополнительные агенты для обеспечения косметической приемлемости композиции. Их примерами являются жиры, воски, масла, краски, ароматизаторы, консерванты, стабилизаторы и поверхностно-активные агенты. Также возможно включение кератолитических агентов, таких как кератолитические агенты, известные в данной области. Примерами являются салициловая кислота и сера. Лекарственные формы для местного или трансдермального введения включают порошки, спреи, мази, пасты, кремы, лосьоны, гели, растворы, пластыри и лекарственные формы для ингаляционного введения. Активное соединение может быть смешано в стерильных условиях с фармацевтически приемлемым носителем и с любыми консервантами, буферами или пропеллентами, которые могут быть необходимы. Мази, пасты, кремы и гели могут содержать, в дополнение к рассматриваемому соединению по изобретению (например, конструкции VitoKine), эксципиенты, такие как животные и растительные жиры, масла, воски, парафины, крахмал, трагакантовая камедь, производные целлюлозы, полиэтиленгликоли, силиконы, бентониты, кремниевая кислота, тальк и оксид цинка, или их смеси.
Дополнительные фармацевтические композиции, которые предполагается применять здесь, включают композиции, содержащие полипептиды в композициях для длительной или контролируемой доставки. Методики изготовления множества других средств для длительной или контролируемой доставки, таких как липосомальные носители, биоразлагаемые микрочастицы или пористые гранулы, и депо-инъекций также известны специалистам в данной области.
Эффективное количество фармацевтической композиции для терапевтического применения будет зависеть, например, от терапевтического контекста и поставленных задач. Специалисту в данной области будет ясно, что подходящие уровни дозы для лечения будут, таким образом, варьировать в зависимости, отчасти, от доставляемой молекулы, показания к применению полипептида, пути введения и биометрических данных (массы тела, площади поверхности тела или размеров органов) и состояния (возраста и общего состояния здоровья) пациента. Соответственно, клинический специалист может корректировать дозу и модифицировать путь введения для получения оптимального терапевтического эффекта. Типичная доза может входить в диапазон от приблизительно 0,0001 мг/кг до приблизительно 100 мг/кг или более, в зависимости от факторов, указанных выше. Предпочтительно, инъекцию или введение полипептидных композиций проводят внутривенно. Длительнодействующие фармацевтические композиции можно вводить один раз в трое или четверо суток, один раз в неделю или один раз в две недели, в зависимости от периода полувыведения и скорости клиренса конкретной композиции. Частота введения будет зависеть от фармакокинетических параметров полипептида в применяемой композиции. Обычно композицию вводят до достижения дозы, обеспечивающей желаемый эффект. Таким образом, композицию можно вводить однократно или многократно (в одинаковых или разных концентрациях/дозах) с течением времени или в форме непрерывной инфузии. Кроме того, обычно проводят подбор подходящей дозы. Подходящие дозы могут быть определены с использованием доступных данных о зависимости ответа от дозы.
Путь введения фармацевтической композиции соответствует известным способам, например, пероральное введение, введение посредством внутривенной, внутрибрюшинной, внутриопухолевой, интрацеребральной (интрапаренхимальной), интрацеребровентрикулярной, внутримышечной, внутриглазной, внутриартериальной, интрапортальной, внутриочаговой инъекции, интрамедуллярное, интратекальное, интравентрикулярное, интравезикальное, трансдермальное, подкожное или внутрибрюшинное введение, а также введение интраназальным, энтеральным, местным, сублингвальным, уретральным, вагинальным или ректальным способом, с использованием систем для длительной доставки или имплантируемых устройств. Там, где это желательно, композиции могут быть введены болюсной инъекцией или непрерывной инфузией или с использованием имплантируемого устройства. Альтернативно или дополнительно, композиция может быть введена местно посредством имплантации мембраны, губки или другого подходящего материала, на котором абсорбирована или инкапсулирована желаемая молекула. При использовании имплантируемого устройства оно может быть имплантировано в любые подходящие ткань или орган и доставка желаемой молекулы может происходить посредством диффузии, времязависимого болюсного или непрерывного высвобождения.
Терапевтическое применение
Согласно настоящему изобретению предложен способ лечения раковых клеток у субъекта, включающий введение указанному субъекту терапевтически эффективного количества (в виде монотерапии или в схеме комбинированной терапии) конструкции VitoKine по настоящему изобретению в фармацевтически приемлемом носителе, где такое введение ингибирует рост и/или пролиферацию раковой клетки. Конкретно, конструкция VitoKine по настоящему изобретению полезна при лечении расстройств, характеризуемых как рак. Такие расстройства включают, без ограничения, солидные опухоли, такие как рак молочной железы, дыхательной системы, головного мозга, репродуктивных органов, пищеварительной системы, мочевыделительной системы, глаза, печени, кожи, головы и шеи, щитовидной железы, паращитовидных желез и их отдаленные метастазы, лимфомы, саркомы, множественную миелому и лейкоз. Примеры рака молочной железы включают, без ограничения, инвазивный протоковый рак, инвазивный дольковый рак, протоковый рак in situ и дольковый рак in situ. Примеры рака дыхательной системы включают, без ограничения, мелкоклеточный и немелкоклеточный рак легкого, а также бронхиальную аденому и плевропульмональную бластому. Примеры рака головного мозга включают, без ограничения, глиому ствола головного мозга и гипоталамуса, астроцитому мозжечка и головного мозга, нейробластому, медуллобластому, эпендимому, а также нейроэктодермальные опухоли и опухоли шишковидной железы. Опухоли мужских репродуктивных органов включают, без ограничения, рак предстательной железы и яичка. Опухоли женских репродуктивных органов включают, без ограничения, рак эндометрия, шейки матки, яичника, влагалища и вульвы, а также саркому матки. Опухоли пищеварительной системы включают, без ограничения, анальный рак, рак ободочной кишки, колоректальный рак, рак пищевода, желчного пузыря, желудка, печени, молочной железы, поджелудочной железы, прямой кишки, тонкой кишки и слюнной железы. Опухоли моче выделитель ной системы включают, без ограничения, рак мочевого пузыря, полового члена, почки, почечной лоханки, мочеточника и мочеиспускательного канала. Рак глаза включает, без ограничения, внутриглазную меланому и ретинобластому. Примеры рака печени включают, без ограничения, печеночно-клеточный рак (печеночно-клеточный рак с или без фиброламеллярного варианта), холангиокарциному (рак внутрипеченочных желчных протоков) и смешанную печеночно-клеточную холангиокарциному. Рак кожи включает, без ограничения, плоскоклеточный рак, саркому Капоши, злокачественную меланому, рак кожи из клеток Меркеля и немеланомный рак кожи. Рак головы и шеи включает, без ограничения, назофарингеальный рак и рак губы и полости рта. Лимфомы включают, без ограничения, ADIS (синдром приобретенного иммунодефицита)-ассоциированную лимфому, неходжкинскую лимфому, Т-клеточную лимфому кожи, болезнь Ходжкина и лимфому центральной нервной системы. Саркомы включают, без ограничения, саркому мягких тканей, остеосаркому, злокачественную фиброзную гистиоцитому, лимфосаркому и рабдомиосаркому. Лейкозы включают, без ограничения, острый миелолейкоз, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, хронический миелогенный лейкоз и волосатоклеточный лейкоз. В различных воплощениях рак будет представлять собой рак с высокой экспрессией представителя семейства TGF-β, такого как активин А, миостатин, TGF-β и GDF15, например рак поджелудочной железы, рак желудка, рак печени, рак молочной железы, рак яичника, колоректальный рак, меланому, лейкоз, рак легкого, рак предстательной железы, рак головного мозга, рак мочевого пузыря и рак головы и шеи.
В различных воплощениях конструкция VitoKine может быть использована в виде монотерапии для лечения всех видов рака, включая, без ограничения, немелкоклеточный рак легкого, мелкоклеточный рак легкого, меланому, почечно-клеточный рак, уротелиальный рак, рак печени, рак молочной железы, рак поджелудочной железы, колоректальный рак, рак желудка, рак предстательной железы и саркому.
В другом аспекте согласно настоящему изобретению предложен способ лечения аутоиммунного заболевания у субъекта, включающий введение указанному субъекту терапевтически эффективного количества (в виде монотерапии или в схеме комбинированной терапии) конструкции VitoKine по настоящему изобретению в фармацевтически приемлемом носителе. «Аутоиммунное заболевание» относится к незлокачественному заболеванию или расстройству, происходящему из и направленному против собственных тканей индивида. Примеры аутоиммунных заболеваний или расстройств включают, без ограничения: воспалительные ответы, такие как воспалительные заболевания кожи, включая псориаз и дерматит (например, атопический дерматит); ответы, связанные с воспалительным заболеванием кишечника (таким как болезнь Крона и язвенный колит); дерматит; аллергические состояния, такие как экзема и астма; ревматоидный артрит; системную красную волчанку (SLE) (включая, без ограничения, волчаночный нефрит, кожную волчанку); сахарный диабет (например, сахарный диабет 1-го типа или инсулинозависимый сахарный диабет); рассеянный склероз и ювенильный диабет.
В другом аспекте согласно настоящему изобретению предложен способ лечения воспалительного заболевания у субъекта, включающий введение указанному субъекту терапевтически эффективного количества (в виде монотерапии или в схеме комбинированной терапии) конструкции VitoKine по настоящему изобретению в фармацевтически приемлемом носителе. «Воспалительные заболевания» включают все заболевания, связанные с острым или хроническим воспалением. Острое воспаление является начальным ответом организма на вредные стимулы и результатом усиленного перемещения плазмы и лейкоцитов (таких как, например, гранулоциты) из крови в поврежденные ткани. Ряд биохимических явлений приводит к распространению и созреванию воспалительного ответа, затрагивающего местную сосудистую систему, иммунную систему и различные клетки поврежденной ткани. Продолжительное воспаление называют хроническим воспалением, которое приводит к прогрессивной смене типов клеток, присутствующих в очаге воспаления, и характеризуется одновременным разрушением и заживлением ткани под действием воспалительного процесса. Примеры воспалительных заболеваний хорошо известны в данной области. В различных воплощениях воспалительное заболевание выбрано из группы, состоящей из воспалительного заболевания кишечника, псориаза и бактериального сепсиса. При использовании здесь термин «воспалительное заболевание кишечника» относится к группе воспалительных состояний ободочной кишки и тонкой кишки, включающей, например, болезнь Крона, язвенный колит, коллагенозный колит, лимфоцитарный колит, ишемический колит, воспаление в отключенной кишке, синдром Бехчета и неуточненный колит.
В другом аспекте согласно настоящему изобретению предложен способ лечения вирусной инфекции у субъекта, включающий введение указанному субъекту терапевтически эффективного количества (в виде монотерапии или в схеме комбинированной терапии) конструкции VitoKine по настоящему изобретению в фармацевтически приемлемом носителе. В различных воплощениях вирусная инфекция, подлежащая лечению, может быть вызвана инфекционными агентами, включая, без ограничения, бактерии, грибы, простейшие и вирусы. Вирусные заболевания, которые можно предотвращать, лечить и/или контролировать в соответствии со способами, описанными здесь, включают, без ограничения, заболевания, вызванные гепатитом типа А, гепатитом типа В, гепатитом типа С, гриппом, ветряной оспой, аденовирусом, простым герпесом 1-го типа (HSY-I), простым герпесом II-го типа (HSY-II), чумой рогатого скота, риновирусом, эховирусом, ротавирусом, респираторно-синцитиальным вирусом, папилломавирусом, паповавирусом, цитомегаловирусом, эхиновирусом, арбовирусом, хантавирусом, вирусом Коксаки, вирусом эпидемического паротита, вирусом кори, вирусом краснухи, вирусом полиомиелита, натуральной оспой, вирусом Эпштейна-Барр, вирусом иммунодефицита человека 1-го типа (HIV-1), вирусом иммунодефицита человека 2-го типа (HIV-2) и возбудителями таких вирусных заболеваний, как вирусный менингит, энцефалит, лихорадка денге или натуральная оспа.
Бактериальные заболевания, вызванные бактериями (например, Escherichia coli, Klebsiella pneumoniae, Staphylococcus aureus, Enterococcus faecalis, Candida albicans, Proteus vulgaris, Staphylococcus viridians и Pseudomonas aeruginosa), которые можно предотвращать, лечить и/или контролировать в соответствии со способами, описанными здесь, включают, без ограничения, микобактерий, риккетсий, микоплазм, нейссерий, S. pneumonia, Borrelia burgdorferi (болезнь Лайма), Bacillus antracis (сибирскую язву), столбняк, стрептококков, стафилококков, микобактерий, коклюш, холеру, чуму, дифтерию, хламидий, S. aureus и легионелл.
Протозойные заболевания, вызванные простейшими, которые можно предотвращать, лечить и/или контролировать в соответствии со способами, описанными здесь, включают, без ограничения, лейшманий, кокцидий, трипаносом и малярию.
Паразитарные заболевания, вызванные паразитами, которые можно предотвращать, лечить и/или контролировать в соответствии со способами, описанными здесь, включают, без ограничения, хламидий и риккетсий.
«Терапевтически эффективное количество» или «терапевтически эффективная доза» относятся к такому количеству вводимого терапевтического агента, которое будет в некоторой степени облегчать один или более чем один симптом расстройства, по поводу которого проводят лечение.
Исходная оценка терапевтически эффективной дозы может быть проведена в анализах клеточных культур посредством определения IC50. Затем в моделях на животных может быть подобрана доза, позволяющая достичь диапазона концентрации в циркулирующей плазме, включающего IC50, определенную в клеточной культуре. Такая информация может быть использована для более точного определения доз, применимых у человека. Уровни в плазме можно измерять, например, посредством HPLC. Точные композиция, путь введения и доза могут быть выбраны конкретным врачом с учетом состояния субъекта.
Схемы введения можно корректировать для обеспечения оптимального желаемого ответа (например, терапевтического или профилактического ответа). Например, может быть проведено однократное болюсное введение, могут быть введены несколько раздельных доз (многократных, повторных или поддерживающих) на протяжении некоторого периода времени, и дозу можно пропорционально снижать или повышать в зависимости от особенностей терапевтической ситуации. Особенно предпочтительно изготавливать композиции для парентерального введения в стандартной лекарственной форме для простоты введения и единообразия доз. При использовании здесь стандартная лекарственная форма относится к физически дискретным единицам, подходящим для использования в качестве единичных доз у субъектов-млекопитающих, для которых предназначено лечение, где каждая единица содержит предопределенное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта, вместе с необходимым фармацевтическим носителем. Характеристики стандартных лекарственных форм по настоящему изобретению будут продиктованы в первую очередь уникальными свойствами антитела и конкретным терапевтическим или профилактическим эффектом, подлежащим достижению.
Таким образом, исходя из представленного здесь описания, специалисту в данной области будет ясно, что дозу и схему введения корректируют в соответствии с методами, хорошо известными в терапевтической области. То есть, может быть легко определена максимальная переносимая доза, и также может быть определено эффективное количество, обеспечивающее выявляемый терапевтический полезный эффект у субъекта, как и временные требования для введения каждого агента для обеспечения выявляемого терапевтического полезного эффекта у субъекта. Соответственно, несмотря на то, что здесь приведены примеры определенных доз и схем введения, эти примеры никоим образом не ограничивают дозу и схему введения, которые могут быть использованы у субъекта при практическом применении настоящего изобретения.
Следует отметить, что значения доз могут варьировать в зависимости от типа и тяжести состояния, подлежащего облегчению, и могут включать однократные или многократные дозы. Следует также понимать, что для любого конкретного субъекта указанные схемы введения следует корректировать с течением времени в соответствии с индивидуальными потребностями и профессиональным суждением лица, поводящего или контролирующего введение композиций, и что диапазоны доз, приведенные здесь, являются лишь примерами, и подразумевают, что они не ограничивают объем или практическое применение заявленной композиции. Кроме того, схема введения композиций по данному изобретению может быть основана на множестве факторов, включая тип заболевания, возраст, массу тела, пол и состояние здоровья субъекта, тяжесть состояния, путь введения и конкретное используемое антитело. Таким образом, схема введения может варьировать в широких пределах, но может быть определена рутинным образом с применением стандартных методов. Например, дозы можно корректировать, исходя из фармакокинетических или фармакодинамических параметров, которые могут включать клинические эффекты, такие как токсические эффекты, и/или лабораторные показатели. Таким образом, настоящее изобретение охватывает повышение дозы, применяемой у субъекта, по усмотрению специалиста. Определение подходящих доз и схем введения хорошо известно в соответствующей области и будет ясно специалисту, располагающему информацией, раскрытой здесь.
Неограничивающими примерами диапазона суточных доз для терапевтически или профилактически эффективного количества VitoKine или варианта VitoKine по изобретению могут быть диапазоны, составляющие от 0,0001 до 100 мг/кг, от 0,0001 до 90 мг/кг, от 0,0001 до 80 мг/кг, от 0,0001 до 70 мг/кг, от 0,0001 до 60 мг/кг, от 0,0001 до 50 мг/кг, от 0,0001 до 40 мг/кг, от 0,0001 до 30 мг/кг, от 0,0001 до 20 мг/кг, от 0,0001 до 10 мг/кг, от 0,0001 до 5 мг/кг, от 0,0001 до 4 мг/кг, от 0,0001 до 3 мг/кг, от 0,0001 до 2 мг/кг, от 0,0001 до 1 мг/кг, от 0,001 до 50 мг/кг, от 0,001 до 40 мг/кг, от 0,001 до 30 мг/кг, от 0,001 до 20 мг/кг, от 0,001 до 10 мг/кг, от 0,001 до 5 мг/кг, от 0,001 до 4 мг/кг, от 0,001 до 3 мг/кг, от 0,001 до 2 мг/кг, от 0,001 до 1 мг/кг, от 0,010 до 50 мг/кг, от 0,010 до 40 мг/кг, от 0,010 до 30 мг/кг, от 0,010 до 20 мг/кг, от 0,010 до 10 мг/кг, от 0,010 до 5 мг/кг, от 0,010 до 4 мг/кг, от 0,010 до 3 мг/кг, от 0,010 до 2 мг/кг, от 0,010 до 1 мг/кг, от 0,1 до 50 мг/кг, от 0,1 до 40 мг/кг, от 0,1 до 30 мг/кг, от 0,1 до 20 мг/кг, от 0,1 до 10 мг/кг, от 0,1 до 5 мг/кг, от 0,1 до 4 мг/кг, от 0,1 до 3 мг/кг, от 0,1 до 2 мг/кг, от 0,1 до 1 мг/кг, от 1 до 50 мг/кг, от 1 до 40 мг/кг, от 1 до 30 мг/кг, от 1 до 20 мг/кг, от 1 до 10 мг/кг, от 1 до 5 мг/кг, от 1 до 4 мг/кг, от 1 до 3 мг/кг, от 1 до 2 мг/кг или от 1 до 1 мг/кг массы тела, Следует отметить, что значения доз могут варьировать в зависимости от типа и тяжести состояний, подлежащих облегчению. Следует также понимать, что для любого конкретного субъекта конкретные схемы введения следует корректировать с течением времени в соответствии с индивидуальными потребностями и профессиональным суждением лица, поводящего или контролирующего введение композиций, и что диапазоны доз, приведенные здесь, являются лишь примерами, и подразумевают, что они не ограничивают объем или практическое применение заявленной композиции.
Токсичность и терапевтический индекс фармацевтических композиций по изобретению могут быть определены стандартными фармацевтическими методиками в клеточных культурах или у экспериментальных животных, например, методиками определения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной для 50% популяции). Отношение токсичной и терапевтически эффективной дозы представляет собой терапевтический индекс, и он может быть выражен как отношение LD50/ED50. Обычно предпочтительны композиции, демонстрирующие большие терапевтические индексы.
Частота введения фармацевтической композиции конструкции VitoKine зависит от вида терапии и конкретного заболевания, подлежащего лечению. Композицию можно вводить субъекту через равные интервалы, такие как неделя или месяц, до достижения желаемого терапевтического результата. Примеры частоты введения включают, без ограничения: один раз в неделю без перерывов; один раз в неделю; один раз в две недели; один раз в 2 недели; один раз в 3 недели; один раз в неделю без перерыва на протяжении 2 недель, затем один раз в месяц; один раз в неделю без перерыва на протяжении 3 недель, затем один раз в месяц; один раз в месяц; один раз в два месяца; один раз в три месяца; один раз в четыре месяца; один раз в пять месяцев; или один раз в шесть месяцев; или один раз в год.
Комбинированная терапия
При использовании здесь термины «совместное введение», «совместно вводимые» и «в комбинации с», относящиеся к конструкции VitoKine по изобретению и одному или более чем одному другому терапевтическому агенту, предназначены для обозначения и включения следующего: одновременного введения такой комбинации конструкции VitoKine по изобретению и терапевтического(их) агента(ов) субъекту, нуждающемуся в лечении, когда такие компоненты включены вместе в одну лекарственную форму, высвобождающую указанные компоненты у указанного субъекта по существу в одно и то же время; по существу одновременного введения такой комбинации конструкции VitoKine по изобретению и терапевтического(их) агента(ов) субъекту, нуждающемуся в лечении, когда такие компоненты включены отдельно друг от друга в раздельные лекарственные формы, принимаемые указанным субъектом по существу одновременно, вследствие чего высвобождение указанных компонентов у указанного субъекта происходит по существу в одно и то же время; последовательного введения такой комбинации конструкции VitoKine по изобретению и терапевтического(их) агента(ов) субъекту, нуждающемуся в лечении, когда такие компоненты включены отдельно друг от друга в раздельные лекарственные формы, принимаемые указанным субъектом последовательно с существенным временным интервалом между каждым приемом, вследствие чего высвобождение указанных компонентов у указанного субъекта происходит по существу в разное время; и последовательного введения такой комбинации конструкции VitoKine по изобретению и терапевтического(их) агента(ов) субъекту, нуждающемуся в лечении, когда такие компоненты включены вместе в одну лекарственную форму, высвобождающую указанные компоненты контролируемым образом, вследствие чего у указанного пациента происходит их одновременное, последовательное и/или перекрывающееся высвобождение, в одно и то же и/или в разное время, где все части могут быть введены одним и тем же способом введения или разными способами введения.
В другом аспекте согласно настоящему изобретению предложен способ лечения рака или метастазирования рака у субъекта, включающий введение терапевтически эффективного количества фармацевтических композиций по изобретению в комбинации со второй терапией, включая, без ограничения, иммунотерапию, цитотоксическую химиотерапию, направленную терапию низкомолекулярными ингибиторами киназ, хирургическое вмешательство, лучевую терапию и трансплантацию стволовых клеток. Например, такие способы могут быть применены в профилактическом предотвращении рака, предотвращении рецидивов и метастазов рака после хирургического вмешательства и в качестве дополнения к другой стандартной терапии рака. Согласно настоящему изобретению, применение комбинированных способов, описанных здесь, может повысить эффективность стандартной терапии рака (например, химиотерапии, лучевой терапии, фототерапии, иммунотерапии и хирургических вмешательств).
Показано, что широкий спектр стандартных соединений обладает противоопухолевой активностью. Эти соединения применяют в качестве фармацевтических агентов при химиотерапии для уменьшения размеров солидных опухолей, предотвращения метастазов и дальнейшего роста или уменьшения числа злокачественных Т-клеток при лейкозах или злокачественных новообразованиях костномозгового происхождения. Несмотря на эффективность химиотерапии при лечении различных типов злокачественных новообразований, многие противоопухолевые соединения вызывают нежелательные побочные эффекты. Было показано, что при сочетании двух или более разных видов лечения они могут действовать синергично, позволяя снижать дозу каждого из них, что приводит к уменьшению вредных побочных эффектов, оказываемых каждым соединением в более высоких дозах. В других случаях злокачественные новообразования, рефрактерные к лечению, могут отвечать на комбинированную терапию из двух или более разных видов лечения.
В различных воплощениях пациенту будут вводить второй противораковый агент, такой как химиотерапевтический агент. Перечень типичных химиотерапевтических агентов включает, без ограничения, даунорубицин, дактиномицин, доксорубицин, блеомицин, митомицин, азотистый иприт, хлорамбуцил, мелфалан, циклофосфамид, 6-меркаптопурин, 6-тиогуанин, бендамустин, цитарабин (СА), 5-фторурацил (5-FU), флоксуридин (5-FUdR), метотрексат (МТХ), колхицин, винкристин, винбластин, этопозид, тенипозид, цисплатин, карбоплатин, оксалиплатин, пентостатин, кладрибин, цитарабин, гемцитабин, паклитаксел, митоксантрон, диэтилстилбестрол (DES), флударабин, ифосфамид, гидроксикарбамидтаксаны (такие как паклитаксел и доцетаксел) и/или антрациклиновые антибиотики, а также комбинации агентов, такие как, без ограничения, DA-EPOCH, CHOP, CVP или FOLFOX. В различных воплощениях дозы таких химиотерапевтических агентов включают, без ограничения, любое из приблизительно 10 мг/м2, 20 мг/м2, 30 мг/м2, 40 мг/м2, 50 мг/м2, 60 мг/м2, 75 мг/м2, 80 мг/м2, 90 мг/м2, 100 мг/м2, 120 мг/м2, 150 мг/м2, 175 мг/м2, 200 мг/м2, 210 мг/м2, 220 мг/м2, 230 мг/м2, 240 мг/м2, 250 мг/м2, 260 мг/м2 и 300 мг/м2.
В различных воплощениях способы комбинированной терапии по настоящему изобретению могут дополнительно включать проведение у субъекта терапевтически эффективного количества иммунотерапии, включая, без ограничения: лечение с использованием истощающих антител к определенным опухолевым антигенам; лечение с использованием конъюгатов «антитело - лекарственное средство»; лечение с использованием агонистических, антагонистических или блокирующих антител к костимулирующим или коингибирующим молекулам (иммунным контрольным точкам), таких как, включая, без ограничения, антитела к CTLA-4, PD-1, PDL-1, CD40, ОХ-40, CD 137, GITR, LAG3, TIM-3, SIRPa, CD47, GITR, ICOS, CD27, Siglec 7, Siglec 8, Siglec 9, Siglec 15 и VISTA, CD276, CD272, TIM-3, B7-H4; лечение с использованием биспецифичных антител, привлекающих Т-клетки (BiTE®), таких как блинатумомаб; лечение, включающее введение модификаторов биологического ответа, таких как IL-2, IL-7, IL-10, IL-12, IL-15, IL-21, IL-22, GM-CSF, IFN-α, IFN-β, IFN-γ, антагонист TGF-β или ловушка TGF-β; лечение с использованием терапевтических вакцин, включая, без ограничения, онколитические вирусы, такие как T-vec, или терапевтические вакцины, такие как сипулейцел-Т; лечение с использованием дендритноклеточных вакцин или вакцин на основе опухолевых антигенных пептидов или неоантигенов; лечение с использованием Т-клеток с химерными антигенными рецепторами (CAR-T-клеток); лечение с использованием CAR-NK-клеток; лечение с использованием NK-клеток; лечение с использованием iPS-индуцированных NK-клеток; лечение с использованием iPS-индуцированных Т-клеток; лечение с использованием iPS-индуцированных CAR-T-клеток или iPS-индуцированных NK-клеток; лечение с использованием опухоль-инфильтрирующих лимфоцитов (TIL); лечение с использованием адоптивно перенесенных противоопухолевых Т-клеток (культивированных ex vivo и/или TCR-Т-клеток); лечение с использованием клеток TALL-104; и лечение с использованием иммуностимулирующих агентов, таких как агонисты Toll-подобных рецепторов (TLR) CpG, TLR7, TLR8, TLR9, и вакцины, такой как бацилла Кальметта-Герена (BCG), и имихимода; где комбинированная терапия обеспечивает усиленное уничтожение опухолевых клеток эффекторными клетками, то есть при совместном применении конструкций VitoKine и иммунотерапии возникает синергия.
В различных воплощениях способы комбинированной терапии по настоящему изобретению могут дополнительно включать введение субъекту терапевтически эффективного количества противовоспалительных агентов для лечения аутоиммунных заболеваний, воспалительных заболеваний и других иммунных расстройств, включая, без ограничения: лечение с использованием истощающих антител к определенным иммунным клеткам; лечение с использованием модулирующих антител (агонистических, антагонистических или блокирующих) в качестве направленных модификаторов иммунного ответа, действующих на мишени (лиганды или их рецепторы), включающие, без ограничения, IL-1α, IL-1β или IL-1R, IL-4 или IL-4R, IL-5 или IL-5R, IL-6 или IL-6R, IL-8 или IL-8R, IL-7 или IL-7R, IL-10 или IL-10R, IL-11 или IL-11R, IL-12 или IL-12R, IL-17 или IL-17R, IL-18 или IL-18R, IL-21 или IL-18R, IL-22 или IL-22R, IL-23 или IL-23R, MCSF или MCSF-R, GM-CSF или GM-CSFR, IFN-α, IFN-β, IFN-γ, TGF-α, TGF-β или TGF-β, семейство TNF или их соответствующие рецепторы, семейство интегринов (например, α4p7), TSLP, компонент комплемента 5 (С5) или С5а, IgE, APRIL, TACI, ВСМА, CD20, CD22, CD40/CD40L, В7Н1, В7Н2, ICOS, BAFF, BCR, BLys, B7RP1, TLR7, TLR8, TLR9; лечение с использованием малых молекул (агонистических или антагонистических) в качестве направленных модификаторов иммунного ответа, действующих на мишени, включающие, без ограничения, NFkB, Jak1, Jak2, Jak3, Tyk2, Syk, ВТК, PIK3, циклооксигеназу-2 и NMDA-рецепторы; где комбинированная терапия повышает эффективность модуляции иммунных ответов, то есть при совместном применении конструкций VitoKine и противовоспалительной терапии возникает синергия.
В различных воплощениях комбинированная терапия включает введение конструкции VitoKine и композиции второго агента одновременно, в одной и той же фармацевтической композиции или в раздельных фармацевтических композициях. В различных воплощениях композицию конструкции VitoKine и композицию второго агента вводят последовательно, то есть композицию конструкции VitoKine вводят до или после введения композиции второго агента. В различных воплощениях композицию конструкции VitoKine и композицию второго агента вводят одновременно, то есть периоды введения композиции конструкции VitoKine и композиции второго агента перекрываются друг с другом. В различных воплощениях композицию конструкции VitoKine и композицию второго агента вводят неодновременно. Например, в различных воплощениях введение композиции конструкции VitoKine прекращают до введения композиции второго агента. В различных воплощениях введение композиции второго агента прекращают до введения композиции конструкции VitoKine.
Следующие примеры предложены для более полного пояснения изобретения, но не ограничивают его объем.
Пример 1
Конструирование и получение конструкций IL-15 VitoKine
Задача состояла в разработке конструкций IL-15 VitoKine, которые будут сохранять инертность до их местной активации протеазами, регуляция которых повышена в раковой ткани или ткани, пораженной заболеванием. Здесь описаны конструкции VitoKine с IL-15 дикого типа (SEQ ID NO: 2) или мутеином IL-15 (например, SEQ ID NO: 3) в качестве активной группировки, обратимо замаскированной между Fc-доменом и IL-15RαSushi+ (SEQ ID NO: 5). Эти конструкции содержат один или два расщепляемых линкера, распознаваемые опухоль-специфичными протеазами. В присутствии опухолевых клеток, экспрессирующих протеазы, будет происходить расщепление линкера, соединяющего Fc и мутеин IL-15, и/или линкера, соединяющего IL-15 и IL-15αSushi+, с восстановлением посредством этого активности IL-15. Следует отметить, что после протеолиза IL-15αSushi+ предположительно останется нековалентно связанным с IL-15 ввиду исключительно высокой аффинности между IL-15 и IL-15Rα (30 пМ). Были получены конструкции Fc IL-15 VitoKine с различными комбинациями линкеров и пептидных спейсеров, которые схематически показаны на ФИГ. 1, а их соответствующие последовательности приведены как SEQ ID NO: 25-43,162-165 и 169-174.
Все гены, синтезированные и субклонированные в реципиентный вектор экспрессии для клеток млекопитающих (GenScript), прошли оптимизацию кодонов для экспрессии в клетках млекопитающих. Экспрессией белка управляет промотор CMV, а на 3'-конце CDS присутствует синтетическая поли-А сигнальная последовательность SV40. На N-конце конструкций была сконструирована лидерная последовательность для обеспечения необходимых сигналов и процессинга для секреции.
Конструкции получали котрансфекцией клеток HEK293-F, растущих в суспензии, векторами экспрессии для клеток млекопитающих с использованием полиэтиленимина (PEI, линейный, молекулярная масса 25000, Polysciences). При наличии двух или более векторов экспрессии их использовали для трасфекции в соотношении 1:1. Для трансфекции клетки НЕК293 культивировали в бессывороточной экспрессионной среде FreeStyle™ 293 (ThermoFisher). Для продукции в шейкерных колбах объемом 1000 мл (рабочий объем 330 мл) клетки НЕК293 в плотности 0,8×106 клеток/мл высевали за 24 часа до трансфекции. Векторы экспрессии, до общего количества ДНК 330 мкг, смешивали с 16,7 мл среды Opti-MEM (ThermoFisher). После добавления 0,33 мг PEI, разведенного в 16,7 мл среды Opti-MEM, смесь перемешивали на вихревой мешалке и затем инкубировали на протяжении 10 мин при комнатной температуре. После этого раствор ДНК/PEI добавляли к клеткам и проводили инкубацию при 37°С в инкубаторе с атмосферой с 8% СО2. В 4 сутки для содействия устойчивой экспрессии белка в клеточную культуру добавляли бутират натрия (Millipore Sigma) в конечной концентрации 2 мг/л. После 6 суток культивирования получали супернатант для очистки центрифугированием на протяжении 20 мин при 2200 об/мин. Полученный раствор стерилизовали фильтрацией (0,22 мкм фильтр, Corning). Секретированный белок очищали из супернатантов клеточных культур с применением аффинной хроматографии на белке А.
Для аффинной хроматографии супернатант загружали на колонку HiTrap MabSelectSure Protein A FF (CV равен 5 мл, GE Healthcare), уравновешенную с использованием 25 мл забуференного фосфатом физиологического раствора, рН 7,2 (ThermoFisher). Несвязанный белок удаляли промывкой 5 объемами колонки PBS, рН 7,2, и целевой белок элюировали с использованием 25 мМ цитрата натрия, 25 мМ хлорида натрия, рН 3,2. Раствор белка нейтрализовали, добавляя 3% 1 М трис, рН 10,2. Целевой белок концентрировали с использованием концентратора Amicon®Ultra-15 с отсечкой по молекулярной массе 10 кДа (Merck Millipore Ltd.).
Чистоту и молекулярную массу очищенных конструкций анализировали посредством SDS-PAGE в присутствии или в отсутствии восстанавливающего агента и окрашиванием кумасси (hnperialR Stain). Использовали гель-систему NuPAGE® Pre-Cast (4-12% или 8-16% бис-трис, ThermoFisher), следуя инструкциям изготовителя. Концентрацию белка в очищенных белковых образцах определяли, измеряя поглощение UV при 280 им (спектрофотометр Nanodrop, Thermo Fisher), разделенное на молярный коэффициент экстинкции, рассчитанный, исходя из аминокислотной последовательности. Содержание агрегатов конструкций анализировали на высокоэффективной жидкостно-хроматографической (HPLC) системе Agilent 1200. Образцы наносили на эксклюзионную колонку AdvanceBio (300Ǻ, 4,6×150 мм, 2,7 мкм, колонка для ЖХ, Agilent) с использованием 150 мкМ фосфата натрия, рН 7,0, в качестве подвижной фазы при 25°С.
Р-0315 представляет собой димерную С-концевую Fc-слитую IL-15 VitoKine, содержащую последовательности, расщепляемые uPA и ММР, в линкерах L1 и L2, соответственно. IL-15 представляет собой вариант белка с S58D. В качестве примера, демонстрирующего белковый профиль конструкций Fc IL-15 VitoKine, на ФИГ. 3А показан SDS-PAGE-анализ Р-0315 (SEQ ID NO: 33). Эксклюзионная хроматограмма представлена на ФИГ. 3В.
Пример 2
Активность IL-15 in vitro была эффективно замаскирована в формате VitoKine
IL-15 VitoKine Р-0172 (SEQIDNO:27) содержит слитый полипептид IL-15/IL-15RαSushi+, соединенный коротким пептидным линкером GS (SEQ ID NO: 116), соединяющим С-конец гомодимерного Fc-домена через линкер, расщепляемый иРА, в гомодимерном слитом формате. Р-0198 представляет собой димерный С-концевой Fc-слитый белок IL-15 в нековалентном комплексе с IL-15RαSushi. Две молекулы имеют сходную конфигурацию слитой части Fc и IL-15 и существенно различаются тем, как в них включен IL-15RαSushi. С одной из них он слит через короткий линкер GS (Р-0172), а с другой связан нековалентно (Р-0198). Связывающую активность Р-0172 в отношении IL-2Rp определяли твердофазным иммуноферментным анализом (ELISA) в сравнении с Р-0198 (содержащим SEQ ID NO: 45, 44 и 5), высокоактивным слитым белком IL-15/IL-15Rα-Fc.
Кратко IL-2Rβ-ECD (SEQ ID NO: 12) сорбировали на лунки 96-луночных микропланшетов Nunc Maxisorp, по 1 мкг на лунку. После инкубации в течение ночи при 4°С и блокировки суперблокирующим раствором (superblock) (ThermoFisher), в каждую лунку вносили по 100 мкл 3-кратных серийных разведений соединений IL-15, начиная со 100 нМ. После инкубации на протяжении одного часа при комнатной температуре в каждую лунку вносили по 100 мкл козьего антитела против Fc человеческого IgG, конъюгированного с HRP (разведение 1:5000 в разбавителе), и проводили инкубацию при комнатной температуре на протяжении 1 часа. После каждой стадии содержимое лунок тщательно отбирали и проводили их трехкратную промывку с использованием PBS/0,05% Tween-20. В завершение, в каждую лунку вносили по 100 мкл ТМВ-субстрата, планшет проявляли при комнатной температуре в температуре в темноте на протяжении 10 минут и вносили останавливающий раствор (2 Н серная кислота, Ricca Chemical), по 100 мкл на лунку. Оптическую плотность определяли при 450 им и кривые строили с использованием программного обеспечения Prism (GraphPad). Как показано на ФИГ. 4А, VitoKine Р-0172 связывается с IL-2Rp со значительно меньшей активностью, чем Р-0198 (12,2 нМ против 0,21 нМ), вероятно, вследствие пространственных ограничений в результате короткого ковалентного связывания IL-15 и IL-15RαSushi, указывая на то, что IL-15RαSushi в платформе VitoKine эффективно препятствовал связыванию домена IL-15 с его рецептором.
Дальнейшую оценку функциональной активности IL-15 VitoKine Р-0172 в сравнении с Р-0198 проводили, изучая опосредованное IL-15 индуцирование экспрессии CD69 на человеческих NK-клетках и CD8+ Т-клетках из свежих мононуклеарных клеток периферической крови (РВМС) посредством FACS-анализа. CD69 является гликопротеином клеточной поверхности, индуцируемым на ранних стадиях лимфоидой активации, включая NK- и Т-клетки.
Кратко, человеческие РВМС выделяли центрифугированием с использованием Ficoll-Hypaque из лейкоцитарной пленки, приобретенной у Института крови Оклахомы (Oklahoma Blood Institute). Очищенные человеческие РВМС обрабатывали серийными разведениями каждого анализируемого соединения IL-15 и инкубировали при 37°С на протяжении 48 часов. Клетки собирали центрифугированием при 300 g и ресуспендировали в FACS-буфере. После блокировки Fc-рецепторов добавлением Human TruStain FcX (разведение 1:50) клетки окрашивали антителами против человеческого CD56 с FITC, против человеческого CD69 с РЕ и против человеческого CD8 с АРС (разведение 1:50). После 30 минут инкубации с антителами при комнатной температуре клетки собирали и промывали, ресуспендировали в FACS-буфере и анализировали проточной цитометрией. Экспрессию CD69 определяли, анализируя CD56+ NK-клетки и CD8+ Т-клетки, и данные выражены как % СВ69-положительных клеток в анализируемой популяции.
Как продемонстрировано на ФИГ. 4В и 4С, активация CD69 на CD8+ Т- и NK-клетках под действием VitoKine Р-0172 была значительно снижена и поддавалась измерению только при наибольшей проанализированной концентрации, при этом активность была по меньшей мере на 2-3 логарифма ниже, чем у Р-0198. Это указывает на эффективную маскировку активности IL-15 в формате VitoKine. Этот маскирующий эффект в анализе активации CD69 на РВМС был более выраженным, чем в ELISA-анализе связывания с IL-2Rp, указывая на более выраженное снижение активности IL-15 в физиологических условиях по сравнению с условиями in vitro при ELISA. Ввиду наличия пространственных ограничений, VitoKine значительно снижает связывание IL-15 с комплексом IL-2RJ3 и ус, экспрессированным на лимфоцитах, и, в результате, приводит к неэффективной активации пути и значительному снижению активности.
Также проводили анализ биологической активности мономерной IL-15 VitoKine. Р-0170 (SEQ ID NO:26 и 15) является мономерным аналогом Р-0172, имеющим такие же линкеры и слитую конфигурацию. По сравнению с высокоактивным Fc-слитым белком IL-15 Р-0166, Р-0172 показал значительно меньшую способность к активации CD69 на CD8+Т-клетках (ФИГ. 5), указывая на то, что мономерная платформа VitoKine эффективно маскировала биологическую активность IL-15 в домене D2.
Пример 3
Сравнение эффективности маскировки в конструкциях Fc IL-15 VitoKine с линкерами разной длины и состава между IL-15 и IL-15RαSushi+и между Fc и IL-15
IL-15 VitoKine конструировали посредством слияния человеческого IL-15 между двумя разными доменами, такими как Fc-домен, увеличивающий период полувыведения, и альфа-домен его когнатного высокоаффинного корецептора, через линкеры L1 и L2, как показано на ФИГ. 1. Дифференциальный эффект двух линкеров, соединяющих Fc с IL-15 и IL-15 с доменом IL-15RaSushi, а также длины и состава этих линкеров на биологическую активность IL-15 анализировали применительно к желаемому снижению активности.
Для оценки конструкций IL-15 VitoKine с разной длиной нерасщепляемого линкера между IL-15 и IL-15RαSushi+(L2) проводили FACS-анализ маркера активации CD69 в субпопуляциях иммунных клеток свежих человеческих РВМС. Следовали тому же протоколу, что в Примере 2.
Р-0204 (SEQ ID NO:30), Р-0205 (SEQIDNO:31) и Р-0206 (SEQIDNO:32) представляют собой конструкции Fc IL-15 VitoKine, имеющие одинаковую последовательность линкера, расщепляемого uPA, между Fc и IL-15 (L1), но линкеры, соединяющие домены IL-15 и IL-15RαSushi+(L2), в этих трех конструкциях VitoKine имеют разную длину и представляют собой (GGGGS)3 (SEQ ID NO: 127), (GGGGS)2 (SEQ ID NO:126) и GGGGS (SEQ ID NO: 118), соответственно.
Как показано на ФИГ. 6, все конструкции Fc IL-15 VitoKine с разными линкерами длиной от 5 до 15 аминокислот между IL-15 и IL- 15RαSushi+приводили к значительному снижению активности по активации CD8+Т-клеток (ФИГ. 6А) или NK-клеток (ФИГ. 6 В). При сравнении активности Р-0206, Р-0205 и Р-0204, было очевидно, что чем меньше длина линкера, соединяющего домены IL-15 и IL- 15RαSushi+, тем более неактивной становится VitoKine, указывая на то, что степень снижения активности можно дополнительно корректировать длиной линкера L2. В заключение, было продемонстрировано почти полное устранение активности IL-15 посредством его маскировки между Fc-доменом и его когнатным высокоаффинным корецептором альфа и корректировки длины линкера между IL-15 и IL- 15R((L2) с получением подходящего уровня пространственного ограничения.
Также анализировали эффект линкера, соединяющего Fc и IL-15 (L1), на биологическую активность конструкций VitoKine, как показано на ФИГ. 7. Р-0204 и Р-0203 (SEQ ID N0:29) имеют одинаковый гибкий линкер из 15 аминокислот (G4S)3 между IL-15 и IL- 15R((L2), но различаются по длине линкера L1, в то время как Р-0203 содержит более длинный, на 7 остатков GS длиннее, чем в Р-0204, пептидный спейсер, фланкирующий пептид-субстрат uPA, соединяющий Fc и IL-15. Несмотря на различия в длине линкера L1, соединяющего Fc и IL-15, биологическая активность Р-0204 и Р-0203 была сходной (ФИГ. 7), указывая на то, что линкер L1, соединяющий Fc и IL-15, оказывал минимальное влияние на снижение активности IL-15 при его длине от 13 до 35 аминокислотных остатков. Тем не менее, при длине линкера L1 менее 13 аминокислотных остатков или более 35 аминокислотных остатков или в контексте других цитокинов он может влиять на маскировку активности домена D2. В то же исследование был включен Р-0202, имеющий, по сравнению с Р-0203, такой же линкер L1, соединяющий Fc и IL-15, но на 13 аминокислот более короткий линкер L2, соединяющий IL-15 и домен IL- 15RαSushi. Р-0202 показал меньшую биологическую активность, чем Р-0203, подтвердив, что для маскировки активности VitoKine линкер L2 важнее линкера L1.
Эффект состава линкера или пептидной последовательности линкера на активность VitoKine оценивали, измеряя экспрессию Ki67 в ядре NK-клеток и CD8 Т-клеток после обработки IL-15 VitoKine. Ki67 является маркером пролиферации клеток, и был разработан ех v/vo-анализ с человеческими РВМС. Кратко, очищенные человеческие РВМС обрабатывали серийными разведениями соединений IL-15 VitoKine и инкубировали при 37°С на протяжении 5 суток. В 5 сутки клетки промывали один раз FACS-буфером (1% FBS/PBS) и сначала окрашивали Fc-блокирующим раствором и антителами к поверхностным маркерам, включая антитела против человеческого CD56 с FITC и антитела против человеческого CD8 с АРС (разведение 1:50). После 30 минут инкубации и промывки клеточные осадки полностью ресуспендировали, внося 1х фиксирующий и пермеабилизирующий рабочий раствор Foxp3, 200 мкл на лунку, и проводили инкубацию на протяжении 30 минут при комнатной температуре в темноте. После центрифугирования в каждую лунку вносили по 200 мкл 1х пермеабилизирующего буфера для еще одной промывки. Клеточные осадки ресуспендировали в пермеабилизирующем буфере с антителом против человеческого Ki67 с РЕ (разведение 1:25). После 30 минут инкубации при комнатной температуре клетки собирали и промывали, ресуспендировали в FACS-буфере и анализировали проточной цитометрией. Данные выражены как % К167-положительных клеток в анализируемой популяции.
Поскольку линкер L2 оказывает более существенное, чем линкер L1, влияние на активность IL-15 VitoKine, был изучен эффект разного состава последовательности линкера L2 на биологическую активность IL-15 VitoKine. Р-0351 (SEQ ID NO:25), Р-0488 (SEQ ID N0:163) и Р-0489 (SEQ ID NO: 164) имеют одинаковый линкер (G4S)3, соединяющий Fc и IL-15 (L1). Все линкеры, соединяющие IL-15 и IL- 15Rα, имеют длину 10 аминокислот, но разные последовательности. Этот линкер представляет собой (G4S)2 в Р-0351, пептид-субстрат ММР-14 (SEQ ID NO:157) в Р-0488 или пептид-субстрат легумаина (SEQ ID NO: 160) в Р-0489.
Как показано на ФИГ. 8, все три конструкции IL-15 VitoKine имели значительно сниженную активность по пролиферации NK-клеток (ФИГ. 8А) или CD8+Т-клеток (ФИГ. 8В) по сравнению с высокоактивным Fc-слитым белком IL-15/IL- 15R(Р-0156 (SEQ ID NO: 175+176). Разные последовательности пептидных линкеров оказывали несущественное влияние на биологическую активность соответствующих конструкций VitoKine (ФИГ. 8А и 8В), вероятно, вследствие структурной гибкости каждого линкерного пептида. Чем более ригидным является ликер L2, тем к большим структурным ограничениям в молекулах VitoKine он приводит, результатом чего может быть большее снижение активности. Тем не менее, влияние последовательности линкера L2 на активность VitoKine было минимальным, и полученные данные указывают на возможность включения разных расщепляемых линкеров в качестве линкера L2 для эффективной маскировки активности домена D2, что расширяет возможности разработки и применения VitoKine.
В заключение, полученные данные, вместе взятые, продемонстрировали, что линкер L2, соединяющий домены IL-15 (D2) и IL- 15RαSushi+ (D3), играл фундаментальную роль в сокрытии активности D2 для обеспечения инертности VitoKine. Степень снижения активности можно дополнительно корректировать, изменяя длину и последовательность/гибкость линкера L2. При выборе длины и последовательности расщепляемого линкера L2 следует учитывать присутствие специфических протеаз в очаге намеченного заболевания, доступность пептида-субстрата для протеаз и желаемую скорость протеолиза.
Пример 4
Определение подходящих условий взаимодействия для полного расщепления протеазами
Начальные эксперименты по расщеплению протеазами in vitro проводили с использованием конструкций IL-15 VitoKine Р-0315 и Р-0203 для определения возможности расщепления протеазами и оптимальных условий для ММР-2 и uPA, соответственно. Р-0315 (SEQ ID NO:33) содержит линкер, расщепляемый uPA, соединяющий домены Fc и IL-15, и линкер, расщепляемый ММР-2/9, соединяющий домены IL-15 и IL- 15RαSushi+. Р-0203 (SEQ ID NO:29) содержит один линкер, расщепляемый протеазой (uPA), соединяющий домены Fc и IL-15. Линкер между доменами IL-15 и IL- 15RαSushi+в Р-0203 представляет собой гибкий линкер (G4S)3. Рекомбинантные человеческие uPA и ММР-2 были приобретены у BioLegend. ММР-2 поставляли в латентной форме и активировали n-аминофенилацетатом ртути (АРМА, Millipore Sigma), следуя инструкциям изготовителя.
Для протеолитического расщепления под действием ММР-2 4 мкг Р-0315 инкубировали с 30 нг, 100 нг или 300 нг АРМА-активированной ММР-2 в аналитическом буфере, рекомендованном изготовителем (100 мМ трис, 20 мМ CaCl2, 300 мМ NaCl, 0,1% (масс./об.) Brij 35, рН 7,5), при 37°С на протяжении 3 часов. Для остановки взаимодействия в реакционную смесь добавляли краситель для внесения для SDS-PAGE и полученную смесь прогревали при 95°С на протяжении 5 минут. Для оценки расщепления расщепленные образцы разделяли на SDS-PAGE-геле с 4-12% бис-трис. Сравнение обработанных и необработанных образцов продемонстрировало полное расщепление IL-15 VitoKine после обработки ММР-2 во всех проанализированных концентрациях. На это указывало изменение размера и появление отчетливой полосы, приблизительно 9 кДа, на SDS-PAGE-геле (ФИГ. 9), которая представляла собой домен IL- 15RαSushi+, отщепленный от Р-0315.
Способность к расщеплению под действием uPA оценивали с использованием Р-0203. Сначала различные количества uPA добавляли к 2 мкг Р-0203 в 20 мкл PBS-буфере, рН 7,2, и реакционную смесь инкубировали при 37°С на протяжении 2 часов. Расщепление проводили с использованием 0,25 нг, 50 нг, 100 нг и 300 нг uPA; результаты показаны на ФИГ. 10А. Три стрелки на ФИГ. 10А относятся к невосстановленным (NR) образцам и указывают на изменение Fc-цепи при uPA-протеолизе. При «частичном расщеплении» слитый полипептид IL-15/IL- 15RαSushi+ был отщеплен только от одной из двух Fc-цепей, в то время как при «полном расщеплении» слитый полипептид IL-15/IL- 15RαSushi+ был отщеплен от обеих Fc-цепей. Смазанная полоса, обведенная на ФИГ. 10А, представляла собой слитый полипептид IL-15/IL- 15RαSushi+, отщепленный от Fc, а ее смазанный вид был, вероятнее всего, обусловлен гликозилированием. В восстановленных образцах (R) верхняя полоса представляла собой Fc-цепь, связанную со слитым полипептидом IL-15/IL- 15RαSushi+, а отчетливая нижняя полоса представляла собой Fc-цепь после отщепления слитого полипептида IL-15/IL- 15RαSushi+.
SDS-PAGE-гель отчетливо демонстрирует, что при увеличении количества uPA происходило постепенное увеличение количества полностью расщепленного белка в невосстановленных образцах. Сходным образом, происходило увеличение количества расщепленной Fc-цепи в восстановленном образце, что указывает на более высокий уровень расщепления. Тем не менее, ни одно из условий не привело к полному расщеплению. Для достижения полного расщепления сходные реакционные смеси для uPA-расщепления инкубировали на протяжении более продолжительного времени. На ФИГ. 10В показано расщепление 2 мкг Р-0203 с использованием 50 нг, 100 нг и 300 нг uPA при 37°С на протяжении 24 часов. Полученные данные показывают, что 100 нг uPA при инкубации продолжительностью 24 часа приводили к почти полному расщеплению.
Пример 5
Протеазное расщепление Fc IL-15 VitoKine Р-0203 с получением активированных продуктов IL-15
VitoKine Р-0203 (SEQ ID NO:29) содержит пептидный линкер, являющийся субстратом uPA, со спейсерным пептидом, фланкирующим оба конца (SEQ ID NO:90), соединяющий Fc и IL-15, и второй гибкий линкер из 15 аминокислот (GGGGS)3 (SEQ ID NO: 127), соединяющий домены IL-15 и IL- 15RαSushi+. Расщепление протеазой in vitro осуществляли посредством инкубации 100 мкг VitoKine Р-0203 с 5 мкг рекомбинантной человеческой uPA (BioLegend) в 500 мкл PBS-буфера, рН 7,2, на протяжении 24 часов при 37°С. Для остановки взаимодействия вносили 25 мкл смолы Ni-Excel (50%-я суспензия, уравновешенная в PBS, GE Healthcare) для удаления uPA, меченной 6-His, из раствора. В то же время, в реакционную смесь вносили также 50 мкл смолы MabSelect SuRe с белком А (50%-я суспензия, уравновешенная в PBS, GE Healthcare) для удаления фракции отщепленного Fc и нерасщепленной или неполностью расщепленной Р-0203. После инкубации с обеими аффинными смолами при комнатной температуре продолжительностью 15 мин смолы удаляли центрифугированием и получали элюат, содержавший Р-0203, активированную протеазой, а именно слитый полипептид IL-15/IL-15aSushi+ (схематически показанный как активная форма 1 на ФИГ. 2). Как видно на ФИГ. 11А и 11В, активированный фрагмент Р-0203 перемещается в виде смазанной полосы, что, наиболее вероятно, обусловлено гликозилированием.
Пример 6
Протеазное расщепление Fc IL-15 VitoKine Р-0315 с получением активированных продуктов IL-15
VitoKine Р-0315 (SEQ ID NO:33) содержит пептидный линкер, являющийся субстратом uPA (SEQ ID NO:92), соединяющий Fc и IL-15, и второй линкер из 10 аминокислот, расщепляемый ММР-2/9 (SEQ ID NO:95), между доменами IL-15 и IL- 15RαSushi+. Домен IL-15 в Р-0315 содержит замену S58D для усиления связывания с (3-субъединицей рецептора. Расщепление протеазой приводило к образованию двух активированных форм Р-0315.
Одна активированная форма Р-0315 (схематически показанная как активная форма 2 на ФИГ. 2) была получена протеазным расщеплением in vitro с использованием ММР-2. Кратко, 660 нг латентной ММР-2 (BioLegend) активировали с использованием АРМА (Millipore Sigma), следуя инструкциям изготовителя, проводили замену буфера и добавляли к Р-0315 (80 мкг) в 0,4 мл аналитического буфера, рекомендованного изготовителем (100 мМ трис, 20 мМ СаС12, 300 мМ NaCl, 0,1% (масс./об.) Brij 35, рН 7,5). После инкубации при 37°С на протяжении 3 часов в реакционную смесь вносили 50 мкл смолы MabSelect SuRe с белком А (50%-я суспензия, уравновешенная в PBS, GE Healthcare). Желаемую активированную форму 1 элюировали с использованием 25 мМ цитрата натрия, 25 мМ хлорида натрия, рН 3,2. Белок нейтрализовали, добавляя 3% 1 М трис, рН 10,2. Для оценки расщепления образцы разделяли на SDS-PAGE-геле с 4-12% бис-трис (ФИГ. 12А). Р-0315 до расщепления под действием ММР-2 в присутствии восстанавливающего агента показана на дорожке 1, а на дорожках 2 и 3 представлена невосстановленная и восстановленная Р-0315 после протеолиза под действием ММР-2, но до очистки на белке А. Появление на геле отчетливой полосы домена IL- 15Rα-sushi+, 9 кДа, подтвердило эффективное расщепление под действием ММР-2 по пептидному линкеру, являющемуся субстратом ММР-2/9. После очистки на белке А образцы (дорожки 4 и 5) демонстрируют идентичный паттерн перемещения. Эти данные показывают, что домен IL- 15RαSushi+, освобожденный от ковалентного связывания, остается нековалентно связанным с IL-15, слитым с Fc, что показано как активная форма 2 на ФИГ. 2; такая связь была достаточно сильной, чтобы выдержать условия низкого-рН при элюировании от белка А. На ФИГ. 12В дополнительно показаны два нековалентно связанных компонента этой активированной формы.
Другая активированная форма Р-0315 (схематически показанная как активная форма 3 на ФИГ. 2) была получена протеазным расщеплением Р-0315 с использованием как uPA, так и ММР-2. Кратко, 100 мкг Р-0315 инкубировали с 5 мкг в 400 мкл PBS-буфера, рН 7,2, на протяжении 20 часов. Затем в реакционную смесь добавляли равный объем буфера, содержащего 200 мМ трис, 40 мМ CaCl2, 450 мМ NaCl, 0,2% (масс/об.) Brij 35, рН 7,5, для коррекции буфера с получением буфера, близкого к аналитическому буферу, рекомендованному изготовителем для ММР-2 (100 мМ трис, 20 мМ CaCl2, 300 мМ NaCl, 0,1% (масс/об.) Brij 35, рН 7,5). Латентную ММР-2 (660 нг) активировали с использованием АРМА, проводили замену буфера на аналитический буфер, добавляли ее к реакционной смеси и проводили инкубацию при 37°С на протяжении 3 часов. Вносили смолу Ni-Excel (50 мкл 50%-й суспензии, уравновешенной в PBS, GE Healthcare) для удаления His-меченной ММР-2 и uPA из раствора. В то же время, в реакционную смесь вносили 100 мкл смолы MabSelect SuRe с белком А (50%-я суспензия, уравновешенная в PBS, GE Healthcare) для удаления фракции отщепленного Fc и оставшейся нерасщепленной или неполностью расщепленной Р-0315. После инкубации с обеими аффинными смолами при комнатной температуре продолжительностью 15 мин смолы удаляли центрифугированием и получали элюат, содержавший активную форму 3 Р-0315, схематически показанную на ФИГ. 2. Как показано на ФИГ. 12С, активная форма 3 Р-0315 содержит нековалентный комплекс IL-15/IL-15aSushi+, как и следовало ожидать после двойного протеолиза; IL-15 перемещается в виде смазанной полосы, в то время как IL- 15RαSushi+ представляет собой отчетливую полосу, приблизительно 9 кДа, как видно на примере активной формы 2 (ФИГ. 12В).
Пример 7
Оценка активности протеазоактивированных конструкций Fc IL-15 VitoKine в анализе с человеческими РВМС
Для оценки активности протеазоактивированных конструкций IL-15 VitoKine проводили FACS-анализ маркера активации CD69 в субпопуляциях иммунных клеток из свежих человеческих РВМС, как подробно описано в Примере 2. Сравнение Р-0203 и ее соответствующей активированной формы (Р-0203 Activ.; схематически показанной как активная форма 1 на ФИГ. 2), полученной расщеплением под действием uPA, показано на ФИГ. 13. Активность VitoKine до активации протеазой была приблизительно на 3 логарифма ниже, чем у высокоактивного Fc-слитого белка IL-15/IL- 15Rα Р-0165, что согласуется с активностью VitoKine, описанной в Примере 3. После расщепления под действием uPA активность по активации как CD56+ NK-клеток (ФИГ. 13А), так и CD8+Т-клеток (ФИГ. 13В) существенно возрастала, но все равно была заметно ниже, чем у Р-0165, возможно, вследствие ковалентного связывания IL-15 и домена IL- 15Rα. Предположительно, увеличение длины гибкого линкера, соединяющего IL-15 и IL- 15Rα, повысит активность активированной формы. Парадоксально, увеличение длины гибкого линкера, вероятно, также снизит эффективность маскировки активности доменом D3 и, вследствие этого, приведет к получению конструкций VitoKine с более высокой базальной активностью.
Биологическую активность другой IL-15 Fc VitoKine Р-0315 и ее двух активированных форм оценивали, измеряя активацию CD69 при активации субпопуляций иммунных клеток свежих человеческих РВМС. Как видно на ФИГ. 14, активность нерасщепленной Р-0315 почти не поддавалась измерению, подтверждая эффективную маскировку активной группировки в формате VitoKine. Активная форма 2 Р-0315 содержит Fc-слитый IL-15 в нековалентном комплексе с доменом IL- 15RαSushi+, высвобождаемым при расщеплении под действием ММР-2, как показано на ФИГ. 2; он структурно сходен с Р-0313 положительного контроля, являющимся высокоактивным Fc-слитым белком IL-15 IL-15Rα. Активная форма 3 Р-0315 содержит свободный домен IL-15, отщепленный от Fc-домена под действием uPA, и домен IL- 15RαSushi+, высвобожденный при расщеплении под действием ММР-2, два из которых образуют нековалентные комплексы, как показано на ФИГ. 2. Обе активированные формы Р-0315 продемонстрировали полное или почти полное восстановление активности по активации как CD56+ NK-клеток (ФИГ. 14А), так и CCD8+ Т-клеток (ФИГ. 14В); при этом активная форма 3 была несколько активнее активной формы 2. Отсутствие Fc-домена в активной форме 3 может быть полезным, когда желательна временная активация намеченного пути в опухолевом микроокружении.
Активность Р-0315 до и после протеолиза под действием ММР-2 также исследовали, измеряя экспрессию Ki67 в ядрах NK-клеток (ФИГ. 15А) и CD8+ Т-клеток (ФИГ. 15В) после обработки. Для сравнения была включена Р-0351, содержащая два нерасщепляемых гибких линкера. Полученные данные также продемонстрировали инертность данной VitoKine и рост активности в отношении как NK-клеток, так и CD8+ Т-клеток приблизительно на 3 логарифма после протеолитической активации in vitro. Наблюдаемая идентичная активность Р-0351 и Р-0315 указывает на то, что два расщепляемых линкера в Р-0315 оставались интактными при получении, экспрессии и хранении, и были специфичны в отношении соответствующих протеаз.
В заключение, расщепление IL-15 VitoKine Р-0315 под действием ММР-2/9 и/или uPA приводит к активации молекулы, и активность цитокина восстанавливалась до уровней, сходных с высокоактивным соединением IL-15 Р-0313 с ЕС50 в субнаномолярном диапазоне.
Пример 8
Минимальное системное действие цитокина при использовании конструкций Fc IL-15 VitoKine у здоровых мышей
Задача технологической платформы VitoKine состоит в снижении системной токсичности, обусловленной связыванием с мишенью, и расширении терапевтического окна. VitoKine маскирует активный цитокин в инертном состоянии и предотвращает его связывание с рецепторами на периферии или на поверхности клеток, не пораженных заболеванием. Вследствие этого платформа VitoKine ограничивает избыточную активацию пути цитокина и уменьшает нежелательную токсичность, обусловленную связыванием с мишенью вне целевой ткани. VitoKine предназначена для местной активации протеазами, регуляция которых повышена в тканях, пораженных заболеванием. Для проверки этой гипотезы конструкцию VitoKine, расщепляемую протеазами, и нерасщепляемую конструкцию VitoKine вводили здоровым мышам и оценивали их системное цитокиновое действие в сравнении с высокоактивным Fc-слитым белком IL-15.
Р-0313 (SEQ ID NO:47 и 5) представляет собой полностью активную Fc-слитую молекулу IL-15/IL-15Rα, использованную в качестве положительного контроля. Р-0315 (SEQ ID NO:33) представляет собой Fc IL-15 VitoKine, содержащую два линкера, расщепляемые протеазами. Р-0351 (SEQ ID NO:25) представляет собой Fc IL-15 VitoKine, содержащую два нерасщепляемых линкера. В качестве отрицательного контроля был включен наполнитель (PBS). Соединения вводили однократной в/б инъекцией здоровым мышам BALB/c (возраст 8-10 недель, n равно 6 на группу) в дозах 0,1 и 0,3 мг/кг. Образцы крови для иммунофенотипирования получали перед введением (-1 сутки) или в 3, 5 и 7 сутки после введения.
После лизиса эритроцитов лизирующим буфером BD Pharm рассчитывали общее число жизнеспособных мононуклеарных клеток крови методом исключения нежизнеспособных клеток с использованием трипанового синего. После блокировки Fc-рецепторов очищенным антителом против мышиного CD16/CD32 (разведение 1:50), клетки окрашивали антителом против мышиного CD3 с FITC, антителом против мышиного CD49b с АРС и антителом против мышиного CD8 с Регсрсу5.5 (разведение 1:50). После 30 минут инкубации клетки собирали и промывали, ресуспендировали в FACS-буфере и анализировали проточной цитометрией.
Как показано на ФИГ. 16, Р-0313, полностью активный Fc-слитый белок IL-15, в обеих проанализированных дозах приводил к значительному дозозависимому росту числа цитотоксических CD8+ Т-клеток (ФИГ. 16А), NK-клеток (ФИГ. 16В) и общего числа лейкоцитов (ФИГ. 16С) в периферической крови. Число клеток возрастало в 3 сутки, достигало максимума в 5 сутки и почти возвращалось к исходным значениям в 7 сутки. В отличие от этого, как расщепляемая конструкция VitoKine (Р-0315), так и нерасщепляемая конструкция VitoKine (Р-0351) продемонстрировали отсутствие повышения CD8 Т-клеток на всем протяжении исследования (7 суток). Незначительный и поздний рост числа NK-клеток был отмечен у мышей, получавших расщепляемую VitoKine Р-0315 в высокой дозе. Р-0351 и низкая доза Р-0315 не продемонстрировали признаков повышения ни в одной из проанализированных целевых клеточных популяций. В целом, по сравнению с активной молекулой Р-0313, две проанализированные конструкции VitoKine продемонстрировали минимальные системную активацию и рост численности целевых популяций лимфоцитов, успешно маскируя и задерживая активность IL-15 на периферии.
Пример 9
Ингибирование легочного метастазирования клеток рака ободочной кишки конструкциями Fc IL-15 VitoKine у мышей
Антиметастатическую эффективность и иммунологические ответы молекул IL-15 Fc VitoKine изучали в модели легочного метастазирования СТ26 у мышей. Кратко, 1 х 105 клеток рака ободочной кишки мыши, CT26-WT (АТСС CRL-2638), вводили внутривенной инъекцией самкам мышей Balb/C (возраст 9-11 недель). В следующие сутки (1 сутки) начинали введение исследуемых конструкций, внутрибрюшинно инъекционно, четырехкратно, 1 раз в 5 суток. Группы введения (в общей сложности 6, п равно 7 на группу) включали 0,3 мг/кг Р-0315, 0,3 мг/кг Р-0351 и 0,1 мг/кг Р-0313. Р-0315 (SEQ ID NO:33) представляет собой Fc IL-15 VitoKine, содержащую два линкера, расщепляемые протеазами. Р-0351 (SEQ ID NO:25) представляет собой нерасщепляемую Fc IL-15 VitoKine. Р-0313 (SEQID NO:47 и 5) представляет собой полностью активную Fc-слитую молекулу IL-15/IL-15Rα. В качестве отрицательного контроля был включен наполнитель (PBS). В 17 сутки всех мышей умерщвляли для забора ткани. Легкие раздували 15%-й тушью и обесцвечивали в растворе Фекете (10% формальдегида, 5% ледяной уксусной кислоты и 60% этанола). Проводили подсчет опухолевых узлов в легких и антиметастатический эффект представляли как разное число опухолевых узлов в разных группах введения и в группе контроля наполнителем.
Как показано на ФИГ. 17, Р-0313 оказывала значительный эффект, подавляя образование и рост легочных метастазов. При дозе 0,1 мг/кг введение Р-0313 приводило к почти полному ингибированию легочного метастазирования. Расщепляемая VitoKine Р-0315 продемонстрировала 70%-е ингибирование развития узлов в легких; ее антиметастатическая эффективность была сопоставима при всех трех дозах (0,3, 1 или 3 мг/кг). Нерасщепляемая VitoKine Р-0351 продемонстрировала относительно слабый, но значимый эффект по уменьшению развития метастазов, указывая на некоторую собственную базальную активность в высокой дозе. Тем не менее, по сравнению с Р-0351, Р-0315 продемонстрировала заметно лучшую антиметастатическую эффективность (р менее 0,05; ФИГ. 17), указывая на то, что протеолитическое расщепление одного или обоих линкеров Р-0315 и последующее высвобождение активной формы IL-15, вероятно, способствовали большей, чем у Р-0351, эффективности Р-0315 in vivo. Развитие опухолевых метастазов может приводить к повышению протеолитической активности вблизи опухолевого микроокружения.
Иммунологический ответ после введения соединений IL-15 исследовали проточно-цитометрическим анализом периферической крови мышей в 15 сутки (через 4 суток после третьего введения). По сравнению с контролем, рост числа CD8+ Т-клеток был отмечен у мышей, получавших активный Fc-слитый IL-15 Р-0313, но не расщепляемую VitoKine Р-0315 или нерасщепляемую VitoKine Р-0351, указывая на то, что антиметастатическая эффективность в отношении рака ободочной кишки не сопровождалась системным повышением CD8+Т-клеток под действием использованных конструкций VitoKine (ФИГ. 17 и 18А). Тем не менее, во всех трех группах введения соединений IL-15 было отмечено повышение NK-клеток периферической крови, наиболее выраженное в группе нерасщепляемой VitoKine после многократного введения (ФИГ. 18В). Усиленный системный рост числа NK-клеток, но не CD8+ Т-клеток в группах введения VitoKine указывает на то, что NK-клетки более чувствительны к введению IL-15, чем CD8+ Т-клетки, и собственная базальная активность VitoKine может приводить к росту числа NK-клеток. Поэтому критически важно корректировать концентрацию вводимых конструкций IL-15 VitoKine для снижения остаточного системного эффекта. Выраженное повышение NK-клеток в группе Р-0351 также указывает на то, что низкая активность нерасщепляемой VitoKine может приводить к слабой, но стойкой активации ее пути и продолжительным иммунным ответам.
Пример 10
Fc IL-15 VitoKine Р-0315 ингибировала рост развившейся опухоли СТ26 у мышей при минимальной системной цитокиновой активации
Противоопухолевую эффективность и иммунологические ответы Fc IL-15 VitoKine Р-0315 изучали в модели колоректального рака СТ26 у мышей в сравнении с полностью активным Fc-слитым белком IL-15/IL- 15Rα Р-0313. Кратко, самкам мышей Balb/С (возраст 10-12 недель) проводили подкожную инъекцию 1 х 105 клеток СТ26 в правый бок. На 11 сутки, когда средний объем опухоли составлял приблизительно 70 мм3, мышей рандомизировали в три группы (п равно 11 на группу) и в тот же день проводили им внутрибрюшинную инъекцию наполнителя (PBS) или 0,1 мг/кг Р-0315 или Р-0313. Одну дополнительную внутрибрюшинную инъекцию соответствующих исследуемых агентов проводили в 16 сутки (2 x 1 раз в 5 суток). Опухоли измеряли три раза в неделю с использованием циркуля и объем опухоли рассчитывали следующим образом: объем равен 0,5 х (ширина)2 х (длина). Для исследования иммунологического ответа в 19 сутки проводили сбор нетерминальной периферической крови в пробирки, обработанные гепарином. В 21 сутки всех мышей умерщвляли для забора ткани.
Как показано на ФИГ. 19А, у мышей, получавших PBS, быстро развивались большие подкожные опухоли, а введение мышам Р-0315 или Р-0313 задерживало рост опухоли в приблизительно равной степени (ФИГ. 19В и 19С). В 21 сутки после введения опухолевых клеток средний объем опухоли у мышей, получавших контроль, составлял приблизительно 1000 мм3 против приблизительно 450 мм3 у мышей, получавших Р-0315 или Р-0313 (****, р менее 0,0001; односторонний ANOVA с апостериорным критерием Тьюки) (ФИГ. 19D). Следует отметить, что сначала Р-0313 продемонстрировала большее снижение опухолевой нагрузки, чем Р-0315, но эта разница уменьшалась по мере продолжения их введения. Отсроченный противоопухолевый эффект Р-0315 был, вероятно, обусловлен тем временем, которое было необходимо для накопления количества протеаз(ы), достаточного для достижения и расщепления ими пептидных линкеров, являющихся их субстратами, и активации VitoKine.
Затем эффект Р-0315 на пролиферацию CD8+ Т-клеток и NK-клеток периферической крови исследовали в сравнении с Р-0313 и наполнителем посредством проточной цитометрии. Кроме того, сходным образом оценивали эффект Р-0315 на общие периферические и селезеночные популяции лейкоцитов и субпопуляции лимфоцитов (CD8+ Т- и NK-клеток).
Инъекция полностью активного Fc-слитого IL-15/IL- 15Rα Р-0313 мышам с опухолями приводила к выраженной пролиферации и росту числа лимфоцитов как в периферической крови, так и в селезенке (ФИГ. 20-22). По сравнению с группой PBS, после введения Р-0313 пролиферация, оцениваемая по Ki67, была усилена в 4 раза у NK-клеток (61% против 15%; ФИГ. 20А) и 5,3 раза у CD8+клеток (46% против 8,6%; ФИГ. 20В). Сходным образом, введение Р-0313 приводило к выраженному росту общего числа лейкоцитов, NK-клеток и CD8+ Т-клеток как в периферической крови (ФИГ. 21А-21С), так и в селезенке (ФИГ. 22А-22С). Например, общее число периферических лейкоцитов возрастало в 6 раз, а число CD8+ Т-клеток увеличивалось в 5 раз; был отмечен резкий рост числа NK-клеток в 85 раз. В селезенке наиболее выраженный рост числа клеток был отмечен у NK-клеток (в 10 раз) и CD8+ Т-клеток, число которых возрастало в 2,9 раза. Общее количество лейкоцитов в селезенке возрастало умеренно, в 1,7 раза. Выраженная активация цитотоксических CD8+ Т-клеток и NK-клеток согласуется с общими иммуномодулирующими свойствами IL-15, и сильные иммунные ответы, вероятно, были основным фактором, способствовавшим противоопухолевой активности Р-0313 in vivo. Тем не менее, резкие изменения субпопуляций лимфоцитов в крови могут приводить к токсичности и уменьшать терапевтический индекс.
Разительно отличаясь от Р-0313, введение Fc IL-15 VitoKine Р-0315 приводило к минимальным изменениям гомеостаза субпопуляций лимфоцитов в крови. Данные наблюдений представлены на ФИГ. 20 применительно к пролиферации периферических NK- и CD8+ Т-клеток, оцениваемой по Ki67, и ФИГ. 21 применительно к росту общего числа лейкоцитов, NK-клеток CD8+ Т-клеток в периферической крови. Единственным заметным фармакодинамическим эффектом после введения Р-0315 было 4-кратное увеличение числа NK-клеток в селезенке (ФИГ. 22В). Поскольку Р-0315 и Р-0313 задерживали рост развившейся опухоли СТ26 в приблизительно равной степени (ФИГ. 19A-19D), противоопухолевая активность Р-0315 in vivo была, вероятно, результатом протеолиза расщепляемого(ых) линкера(ов) и последующей активации VitoKine вблизи опухолевого микроокружения. Поскольку активированная VitoKine присутствовала только вблизи опухоли, ответ периферических лимфоцитов на введение инертной молекулы VitoKine был намного менее выраженным, чем при введении полностью активного Р-0313.
В целом, IL-15 Fc VitoKine, примером которой была Р-0315, могла эффективно задерживать рост опухоли без существенных изменений пролиферации и численности субпопуляций лимфоцитов в крови и селезенке. Благодаря этому, формат VitoKine позволял предотвращать избыточную активацию пути, нежелательную токсичность, обусловленную связыванием с мишенью вне целевой ткани, и нежелательный клиренс, обусловленный связыванием с мишенью, обычно возникающие при введении полностью активного цитокина, без снижения противоопухолевого эффекта.
Пример 11
Нерасщепляемая конструкция VitoKine как форма цитокина со ослабленной активностью
В данной области известно, что цитокин, активный in vitro, может не приводить к наиболее сильному лимфоцитарному ответу in vivo. Высокоактивные цитокины часто связаны с более сильной стимуляцией, интернализацией и десенситизацией рецепторов, ослаблением сигналов, пролиферации и функции, а также усиленной гибелью клеток или клональным истощением. Поэтому цитокин с ослабленной активностью может быть крайне желателен для предотвращения избыточно сильной активации лимфоцитов и достижения стойкого и усиленного фармакодинамического эффекта и противоопухолевой эффективности in vivo.
Нерасщепляемая Fc IL-15 VitoKine Р-0351 продемонстрировала сильное снижение активности по сравнению с полностью активными соединениями IL-15 in vitro; тем не менее, она показала антиметастатическую эффективность и выраженные NK-клеточные ответы в модели легочного метастазирования СТ26 у мышей (Пример 8). Таким образом, нерасщепляемые конструкции VitoKine могут быть использованы в качестве цитокинов с ослабленной и продолжительной активностью для оптимизации фармакодинамики in vivo.
Активность Р-0351, включая пролиферацию NK-клеток и CD8+ Т-клеток, оцениваемую по Ki67 (ФИГ. 23А и 23В), была такой же, как у эталонной (Benchmark) молекулы (SEQ ID NO: 177 и 178), что эквивалентно XENP024306 в заявке на патент WO 2018071919 A1. XENP024306 представляет собой Fc-слитую молекулу IL-15/IL-15Rα, содержащую аминокислотные замены (D30N/E64Q/N65D) в IL-15 и мутации, увеличивающие период полувыведения, в Fc. Сообщается, что эти три мутации в IL-15-цепи XENP024306 приводили к снижению активности in vitro в 200 раз, но было продемонстрировано, что in vivo XENP024306 была активнее, вероятно, вследствие оптимизированной фармакодинамики in vivo.
Сходным образом, Р-0351 с ослабленной активностью, предположительно, приведет к более продолжительному действию и улучшенной фармакодинамике (PD), благодаря отсутствию или уменьшению избыточной активации и нежелательного клиренса, обусловленного связыванием с мишенью, обычно возникающих при введении полностью активного цитокина. Таким образом, аналог Р-0351 с увеличенным периодом полувыведения, Р-0651 (SEQ ID NO:170), будет способствовать большему периоду полувыведения и дополнительно продлевать PD in vivo.
Пример 12
Конструирование и получение конструкций Fc IL-2 VitoKine для селективного увеличения числа регуляторных Т-клеток (Treg IL-2 VitoKine) для лечения аутоиммунных заболеваний, воспалительных расстройств, трансплантации и других расстройств
Задача состоит в разработке конструкций IL-2 VitoKine, которые будут сохранять инертность до их местной активации протеазами, регуляция которых повышена в очагах воспаления. Низкие дозы IL-2 дикого типа стимулируют преимущественно Treg, но не эффекторные Т-клетки, и, согласно сообщениям, мутеины IL-2 со сниженной аффинностью связывания с IL-2Rβ расширяют окно такой селективности. Эти молекулы можно разрабатывать как терапевтические средства для профилактики аутоиммунных заболеваний. Другие мутации, препятствующие связыванию с IL-2Rβ или γС и не влияющие на взаимодействие с IL-2Rα, могут также увеличивать окно селективности применительно к преимущественной активации Treg в сравнении с Teff.
Использовали IL-2 Fc VitoKine, содержащую IL-2 дикого типа или мутеин IL-2 с повышенной селективностью стимуляции Treg в сравнении с эффекторными Т-клетками в качестве активной группировки, обратимо замаскированной между Fc-доменом и IL-2RaSushi (SEQ ID NO:10). IL-2Rα (SEQ ID NO:9) содержит два sushi-домена, разделенные естественной пептидной линкерной областью. Конструкции IL-2 VitoKine содержат один или два расщепляемых линкера, распознаваемых протеазами, регуляция которых, согласно сообщениям, повышена в очагах воспалительных расстройств. В то время как линкер, соединяющий Fc и IL-2/мутеин, может быть как расщепляемым, так и нерасщепляемым, предпочтительно, чтобы линкер, соединяющий IL-2 и IL-2αSushi, мог быть подвержен специфичному расщеплению протеазой.
Активность мутеина IL-2 по селективной стимуляции Treg будет, предположительно, восстановлена при высвобождении и диффузии IL-2Rα от IL-2 после расщепления протеазой. Ввиду наномолярной аффинности связывания IL-2Rα и IL-2 есть вероятность того, что после расщепления линкера IL-2RαSushi останется нековалентно связанным с IL-2, в результате чего взаимодействие IL-2 с IL-2Rα на Treg-клетках останется заблокированным. Для решения этой возможной проблемы были разработаны мутеины IL-2Rα с аминокислотными заменами на его поверхности, взаимодействующей с IL-2, для ослабления связывания с IL-2. Таким образом, после расщепления линкера протеазой будет происходить диссоциация и диффузия мутанта IL-2RαSushi от IL-2; этот механизм активации (схематически показанный на ФИГ. 26) несколько отличается от показанного на ФИГ. 2.
Типичные аминокислотные замены были сделаны в положениях 38 (то есть К38Е) и 43 (то есть Y43A) домена IL-2Rα. Другие варианты IL-2Rα с заменами остатков, взаимодействующих с IL-2, будут, предположительно, приводить к нарушению взаимодействий IL-2 и IL-2Rα и могут также быть включены. Как будет ясно специалистам в данной области, все мутации можно, необязательно и независимо, комбинировать любым образом для достижения оптимальной модуляции аффинности. Были получены молекулы IL-2 VitoKine, содержащие различные комбинации линкеров, IL-2 дикого типа или варианты IL-2 и IL-2RαSushi дикого типа или варианты IL-2RaSushi, и их соответствующие последовательности представлены как SEQ ID NO:49-65.
Синтез генов, конструирование векторов экспрессии и получение, очистку и описание белков проводили, следуя тем же методикам, что подробно описаны в Примере 1. В качестве примера, демонстрирующего белковый профиль конструкций IL-2 VitoKine, на ФИГ. 24А показан SDS-PAGE-анализ Р-0320. Эксклюзионная хроматограмма, представленная на ФИГ. 24В, демонстрирует, что после исходной стадии захвата на белке А без стадии завершающей очистки содержание агрегатов составляло менее 5%. Малая склонность к агрегации указывает на широкие возможности разработки конструкций IL-2 VitoKine.
Пример 13
Конструирование и получение конструкций Fc IL-2 VitoKine для селективного увеличения числа эффекторных Т-клеток (Teff IL-2 VitoKine) для лечения рака и других расстройств
Задача состоит в разработке конструкций IL-2 VitoKine, которые будут сохранять инертность до их местной активации протеазами, которые присутствуют или регуляция которых повышена только в опухолевых очагах. Преимущественное увеличение числа Treg под действием IL-2 является нежелательным эффектом IL-2 с точки зрения иммунотерапии рака, поскольку Treg могут сдерживать эффекторные Т-клеточные ответы. Для преодоления этих ограничений были разработаны аминокислотные замены на поверхности IL-2, связывающейся с IL-2Rα, включая F42A и R38E (PNAS, 1991. 88: 4636-4640), для снижения/устранения связывания с IL-2Rα. Могут также быть включены другие мутации, препятствующие только связыванию с IL-2Rα, и не влияющие на взаимодействие с IL-2Rβγ, например, R38A, Т41А, T41G, T41V, Y107G, Y107H, Y107L или Y107V. Как будет ясно специалистам в данной области, все мутации можно, необязательно и независимо, комбинировать любым образом для достижения оптимальной модуляции аффинности.
Конструкции Fc IL-2 VitoKine содержат IL-2 дикого типа или вариант IL-2 со сниженным/устраненным связыванием с IL-2Rα в качестве активной группировки, обратимо замаскированной между Fc-доменом и IL-2RαSushi (SEQ ID N0:10). Эти конструкции содержат один или два расщепляемых линкера, распознаваемых протеазами, регуляция которых, согласно сообщениям, повышена при различных типах рака, например, при солидных опухолях. В то время как линкер, соединяющий Fc и мутеин IL-2, может быть как расщепляемым, так и нерасщепляемым, линкер, соединяющий IL-2 и IL-2αSushi, предпочтительно может быть подвержен специфичному расщеплению протеазой. Предпочтительно, IL-2Rα может оставаться связанным с IL-2 после расщепления для повышения селективности в отношении функции Teff. Активность мутеина IL-2 восстанавливается при высвобождении и диффузии IL-2Rα от IL-2 после расщепления протеазой. Молекулы IL-2 VitoKine, содержащие различные мутеины IL-2 в качестве активной группировки, схематически показаны на ФИГ. 1. Были сконструированы и получены типичные молекулы Fc IL-2 VitoKine для селективного увеличения числа Teff-клеток, и их соответствующие последовательности представлены как SEQ ID NO:59-61.
Синтез генов, конструирование векторов экспрессии и получение, очистку и описание белков проводили, следуя тем же методикам, что подробно описаны в Примере 1.
Пример 14
Оценка активности Fc IL-2 VitoKiiie in vitro
Биологическую активность конструкций IL-2 VitoKine в отношении Т-клеток определяли, измеряя уровни фосфорилированного STAT5 (pStat5), в определенных субпопуляциях Т-клеток свежих человеческих РВМС. Известно, что Stat5 вовлечен в последующую внутриклеточную передачу сигналов, индуцированных связыванием IL-2 с трансмембранным комплексом IL-12RβγC. Уровни pStatS измеряли проточной цитометрией в фиксированных и пермеабилизированных клетках с использованием антитела к пептиду pStat5. Кратко, человеческие РВМС выделяли центрифугированием с использованием Ficoll-Hypaque из лейкоцитарной пленки здорового донора, приобретенной у Института крови Оклахомы (Oklahoma Blood Institute). РВМС, 2 х 105, обрабатывали серийными разведениями исследуемых соединений на протяжении 30 минут при 37°С. Затем проводили обработку клеток с использованием набора окрашивающих буферов Foxp3/Transcription Factor Staining Buffer Set (EBIO), следуя инструкциям изготовителя. После этого клетки фиксировали буфером Cytofix, пермеабилизировали буфером Perm Buffer III (BD Biosciences) и затем промывали. После блокировки Fc-рецепторов добавлением Human TruStain FcX (разведение 1:50) клетки окрашивали антителами против CD25 с РЕ, против FOXP3 с АРС, против pSTATS с FITC и против CD4 с PerCP-Су5.5 в концентрациях, рекомендованных изготовителем, на протяжении 60 минут при комнатной температуре. Затем клетки собирали и промывали, ресуспендировали в FACS-буфере и анализировали проточной цитометрией. Данные проточной цитометрии субпопуляций Treg и CD4 эффекторных Т-клеток анализировали по группам Foxp3+/CD25high и Foxp3-/D25low, соответственно. Данные выражены как процент pStat5-положительных клеток в анализируемой популяции.
Конструкции IL-2 VitoKine Р-0320 (SEQ ID NO:49) и Р-0329 (SEQ ID NO:62) оценивали на предмет активации pStat5 в сравнении с Р-0250 (SEQ ID NO:48). Р-0320 содержит домен IL-2 дикого типа, N-конец которого слит с Fc-доменом через линкер, расщепляемый uPA, а С-конец - с доменом IL-2RαSushi через гибкий линкер (GGGGS)3 (SEQ ID NO: 127). Р-0329 содержит домен IL-2 дикого типа, С-конец которого слит с Fc-доменом через линкер, расщепляемый uPA, а N-конец - с доменом IL-2RαSushi через гибкий линкер (GGGGS)3. Р-0250 является высокоактивным Fc-слитым белком IL-2. Процент pStat5-положительных клеток в субпопуляциях Treg и CD4+ обычных Т-клеток (Tconv) при использовании исследуемых соединений показан на ФИГ. 25. Отчетливо видно, что при использовании обеих конструкций IL-2 VitoKine pStat5-активация Treg значительно меньше, чем при использовании полностью активного слитого белка IL-2, а pStat5-активация CD4+Tconv-клеток почти не поддавалась измерению. Эти данные отчетливо демонстрируют эффективную маскировку активности IL-2 в формате VitoKine.
Пример 15
Протеазная активация IL-2 VitoKine и оценка активности in vitro
IL-2 VitoKine P-0382 (SEQ ID NO:51) содержит гибкий линкер GGGSGGGS (SEQ ID NO: 115), соединяющий Fc и IL-2, и линкер из 10 аминокислот, расщепляемый ММР-2/9 (SEQ ID NO:77), между доменами IL-2 и IL-2RαSushi. Домен IL-2RαSushi в Р-0382 содержит аминокислотную замену (К38Е), разработанную для снижения аффинности его связывания с IL-2 и обеспечения диссоциации и последующей диффузии от IL-2 после расщепления линкера протеазой.
Р-0382 активировали протеазным расщеплением in vitro с использованием ММР-2. Кратко, сначала 3,3 мкг латентной ММР-2 (BioLegend) активировали с использованием АРМА (Millipore Sigma), следуя инструкциям изготовителя, затем проводили замену буфера и добавляли к 120 мкг Р-0382 в 0,4 мл аналитического буфера, рекомендованного изготовителем (100 мМ трис, 20 мМ СаС12, 300 мМ NaCl, 0,1% (масс./об.) Brij 35, рН 7,5). После инкубации при 37°С на протяжении 20 часов половину реакционной смеси очищали на смоле MabSelect SuRe с белком А и активированную VitoKine элюировали с использованием 25 мМ цитрата натрия, 25 мМ хлорида натрия, рН 3,2. Белок нейтрализовали, добавляя 3% 1 М трис, рН 10,2. Другую половину образца инкубировали со смолой Ni-Excel для остановки взаимодействия посредством удаления His-меченного белка ММР-2 и активированную VitoKine получали, удаляя Ni-смолу центрифугированием. Очистку на белке А проводили для подтверждения отсутствия ковалентного связывания домена IL-2RαSushi с IL-2 после расщепления полипептидной цепи, как схематически показано на ФИГ. 26. Анализ образцов посредством 4-12% бис-трис SDS-PAGE показан на ФИГ. 27. Несмотря на большее количество протеазы и более продолжительное время взаимодействия по сравнению со структурно сходными конструкциями IL-15 VitoKine (например, Р-0315), взаимодействие не привело к полному расщеплению. Сравнение с образцами, обработанными ММР-2 с и без очистки на белке А [ФИГ. 27А и 27В), подтвердило освобождение домена IL-2RαSushi от ковалентного связывания и его отсутствие в очищенном Fc-слитом полипептиде IL-2.
Несмотря на неполное расщепление, была проведена оценка этих двух ММР-2-активированных образцов, одного в форме Ni-Excel-элюата (activ. 1), а другого в форме элюата после белка A (activ. 2), в анализе активации pStat5, описанном в Примере 13, и полученные данные показаны на ФИГ. 28. Активность Р-0382 была очень низкой у Treg и почти не поддавалась измерению у CD4+ Tconv-клеток, еще раз подтвердив эффективную маскировку активной группировки в формате IL-2 VitoKine. Оба активированных образца продемонстрировали почти полное восстановление активности. Умеренное снижение активности по сравнению с Р-0250 было, вероятно, обусловлено неполным протеолизом.
Присутствие домена IL-2RαSushi, отщепленного под действием ММР-2, в образце activ. 1, по-видимому, не влияло на активность активированной IL-2 VitoKine, поскольку activ. 1 и activ. 2 имели сопоставимую активность при индуцировании фосфорилирования pStat5 как в Treg-, так и в Tconv-клетках (ФИГ. 28А и 28В). Эти данные указывают на то, что домен IL-2RαSushi, полученный в результате расщепления под действием ММР-2, не был связан с IL-2 и не должен препятствовать связыванию IL-2 с рецепторными комплексами, экспрессированными на лимфоцитах.
Протеолиз Р-0382 под действием ММР-2 не приводил к полному расщеплению, и было сделано предположение, что увеличение длины расщепляемого линкера может повысить доступность пептида-субстрата для протеазы, обеспечивающей расщепление. Линкер из 10 аминокислот (SEQ ID NO:95) в Р-0382 был заменен на линкер из 15 аминокислот, расщепляемый ММР-2/9 (SEQ ID NO:94), содержащий дополнительные фланкирующие остатки, с получением новой конструкции VitoKine Р-0398 (SEQ ID NO:52). Р-0398 активировали протеазным расщеплением in vitro с использованием ММР-2, следуя тому же протоколу, что подробно описан выше. В три раза меньшее количество ММР-2 (1,5 мкг ММР-2 на 180 мкг Р-0398 против 3,3 мкг ММР-2 на 120 мкг Р-0382) привело к полному расщеплению Р-0398, на которое указывало присутствие только полосы «полного расщепления» на SDS-PAGE-геле (данные не показаны).
Биологическую активность активированной Р-0398 с удалением домена IL-2RαSushi посредством очистки на белке А определяли в анализе pStat5 (ФИГ. 29А и 29В). Активированная Р-0398 сходна с Fc-слитой молекулой IL-2 Р-0250 по последовательности и структуре, и они имеют почти идентичную активность при индуцировании фосфорилирования Stat5 как в Treg-, так и в Tconv-клетках. Несмотря на то, что обе конструкции VitoKine, Р-0382 и Р-0398, имели значимо сниженную (на 4 логарифма) биологическую активность вследствие ковалентного связывания с доменом IL-2RαSushi, Р-0398, содержащий линкер L2 большей длины, по-видимому, имел тенденцию к большей активности. Аналогично IL-15 Fc VitoKine, степень снижения активности конструкций IL-2 VitoKine можно было дополнительно корректировать, изменяя длину линкера L2. Сходным образом, при выборе длины и последовательности расщепляемого линкера L2 следует учитывать присутствие специфических протеаз в очаге намеченного заболевания, доступность пептида-субстрата для протеаз и желаемую скорость протеолиза.
В заключение, по сравнению с IL-15 VitoKine, в случае IL-2 VitoKine был необходим линкер L2 большей длины для обеспечения его оптимальной доступности для фермента и достижения полного протеолиза. Расщепление типичных конструкций IL-2 VitoKine Р-0382 и Р-0398 под действием ММР-2 приводило к полной активации этих молекул. Активированные конструкции IL-2 VitoKine достигали биологической активности, сходной с высокоактивным Fc-слитым соединением IL-2 Р-0250.
Пример 16
Конструирование VitoKine с антителами
Применение рекомбинантных слитых белков антител с цитокинами (иммуноцитокинов), возможно, повысит терапевтический индекс цитокинов, направляя их в очаг заболевания. Тем не менее, слияние полностью активного цитокина с антителом может привести к периферической активации и недостаточному направленному действию на опухоль. Снижение активности VitoKine перед активацией в намеченном месте терапии делает VitoKine с антителами новой и инновационной формой иммуноцитокинов. Помимо опухоленаправленных антител, для конструирования VitoKine с антителами могут также быть использованы антитела, блокирующие иммунные контрольные точки, позволяющие обойти иммуносупрессивные эффекты опухолевого микроокружения, или иммуностимулирующие антитела для усиления существующих ответов, что может дополнительно усилить противоопухолевую активность иммунной системы. Кроме того, конструкции VitoKine с антителами, направленными на воспаленную ткань, могут быть использованы для лечения аутоиммунных и хронических воспалительных расстройств.
Исходя из этой концепции, были сконструированы белки VitoKine с антителами, содержащие IL-15 или IL-2 в качестве домена D2. Примеры антител включают PD-1-блокирующее антитело JS-001, PD-Ll-блокирующее антитело Тецентрик, антитело против CTLA4 ипилимумаб, агонистическое антитело к CD40 RO7009789, антитела, направленные на опухолевые антигены, включая L19, направленное против экстра-домена фибронектина, ритуксимаб, направленный против CD20, Герцептин, направленный против Her-2, цетуксимаб, направленный против EGFR, и противовоспалительные антитела ведолизумаб против интегрина α4β7 и Хумира против TNFa. Последовательности типичных конструкций VitoKine с антителами представлены как SEQ ID NO:128-143.
Синтез генов, конструирование векторов экспрессии и получение, очистку и описание белков проводили, следуя тем же методикам, что подробно описаны в Примере 1. Биологическую активность типичной IL-15 VitoKine с антителом против PD-L1 Р-0485 (SEQ ID NO:180 и 181) анализировали, измеряя экспрессию Ki67 в NK-клетках (ФИГ. 30А) и CD8+ Т-клетках (ФИГ. 30В) после обработки человеческих РВМС соединениями IL-15 VitoKine. Р-0485 имеет те же линкеры L1 и L2 и домены D2 и D3, что и аналогичная ей Fc VitoKine Р-0315. Согласно данным, представленным на ФИГ. 29, обе конструкции VitoKine имели сопоставимую и сильно сниженную биологическую активность по сравнению с активированной Р-0315, активность которой показана на ФИГ. 15. Р-0485 имела несколько более высокую активность, чему могла способствовать активация лимфоцитов в результате блокады PD-L1.
Пример 17
Блокирующие пептиды на основе IL-15RP для полученных протеазоактивируемых инертных слитых белков IL-15 или IL-2
Другим способом получения протеазоактивируемых инертных слитых белков IL-15 или IL-2 является генетическое слияние блокирующих пептидов (например, блокирующего пептида на основе IL-2Rβ с IL-15 или IL-2 посредством расщепляемого линкера. Исследованные блокирующие пептиды основаны на двух петлях IL-2Rβ (SEQ ID NO:97 и 98), содержащих ключевые остатки, непосредственно контактирующие с IL-15. Пептиды, изложенные в Таблице 13, основаны на последовательностях этих двух петель.
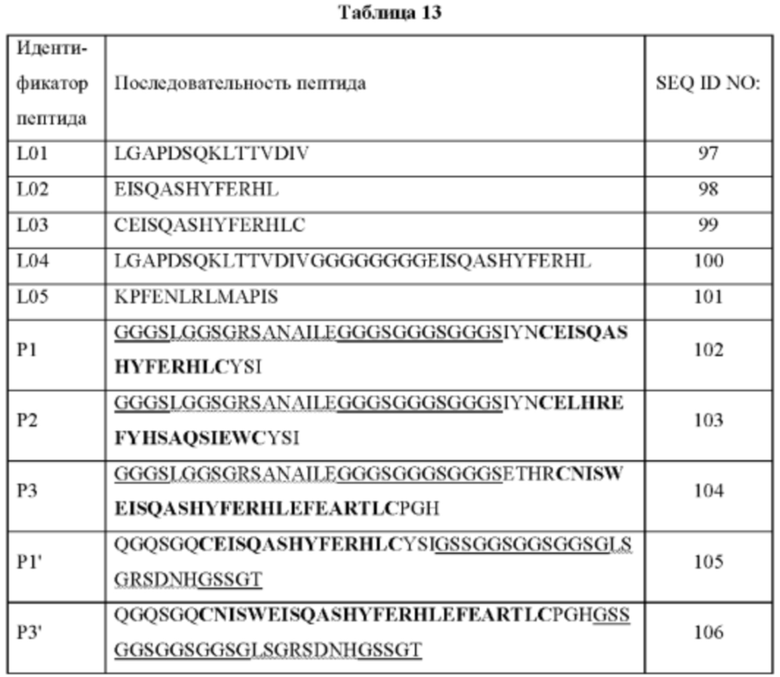
Пять пептидов, L01-L05 (SEQ ID NO:97-101) из Таблицы 13, были синтезированы и проанализированы на предмет их связывания с IL-15 в формате ELISA. Кратко, Р-0153 (SEQ ID NO:44 и 46), Fc-слитый белок IL-15/IL- 15RαSushi+, сорбировали на лунки 96-луночных микропланшетов Nunc Maxisorp, 1 мкг на лунку, и в каждую лунку вносили 3-кратные серийные разведения биотинилированных пептидов, начиная со 100 мкМ. Вносили комплекс стрептавидина с HRP в концентрации, рекомендованной изготовителем, и сигнал проявляли ТМВ-субстратом. Как показано на ФИГ. 30, было отмечено специфичное связывание L03 (SEQ ID NO:99), представлявшего собой циклизованную петлю 2 (SEQ ID NO:98).
Последовательность на основе петли 2 была использована в качестве блокирующих пептидов и включена в слитый белок IL-15. Типичные последовательности слитых белков, содержащих блокирующий пептид на основе IL-2RJ3, слитый с IL-15 посредством расщепляемого линкера и пептидных спейсеров (SEQ ID NO:102-106), показаны в Таблице 13, где полужирным шрифтом выделен блокирующий пептид на основе IL-15Rp, волнистым подчеркиванием выделен расщепляемый линкер, а прямым подчеркиванием выделены спейсерные пептиды. IL- 15RαSushi+ (SEQIDNO:5) коэкспрессировали и получали их нековалентные комплексы со слитым белком IL-15, содержащим блокирующий пептид.
Синтез генов, конструирование векторов экспрессии и получение, очистку и описание белков проводили, следуя тем же методикам, что подробно описаны в Примере 1. Эти слитые белки IL-15, содержащие блокирующие пептиды, сначала изучали в ELISA-анализе для оценки их способности к связыванию с IL-2Rβ. Как показано на ФИГ. 32, аффинность связывания, по сравнению с Р-0153, была умеренно снижена вследствие связи с различными блокирующими пептидами. Тем не менее, анализ с использованием РВМС, в котором была оценена выраженность активации иммунных клеток, включая CD56+ NK-клетки или CD8+ Т-клетки, этими слитыми белками IL-15, содержащими блокирующие пептиды, не продемонстрировал заметного снижения активности (данные не показаны), указывая на недостаточную маскирующую эффективность анализируемых блокирующих пептидов. Блокирующие пептиды разной длины, включая полноразмерный внеклеточный домен IL-2Rp, могут быть изучены на предмет эффективности маскировки активности. Такой же способ может быть сходным образом применен к IL-2.
Пример 18
Формат VitoKine расширяет возможности разработки слитых белков
В данной области известно, что встречающийся в природе белок IL-2 имеет тенденцию к не очень высокой стабильности и склонен к агрегации. В экспериментах, проведенных авторами изобретения, было продемонстрировано, что Fc-слитый белок IL-2 дикого типа (Р-0250), экспрессированный на низком уровне (примерно 3 мг/л, транзиторно в клетках HEK-293F), имеет высокую склонность к агрегации, как показано на SEC-хроматограмме, представленной на ФИГ. 33А. Четыре молекулы IL-2 VitoKine, Р-0320, Р-0382, Р-0362 и Р-0379, сравнивали с Р-0250. Р-0320 (SEQ ID NO:49) содержит домен IL-2 дикого типа, N-конец которого слит с Fc-доменом, а С-конец связан с доменом IL-2RαSushi. Линкер L1, соединяющий Fc и IL-2, представляет собой расщепляемый линкер, содержащий пептид-субстрат uPA и фланкирующие спейсерные пептиды (SEQ ID NO:92), а линкер L2 между IL-2 и IL-22RαSushi представляет собой гибкий линкер (GGGGS)3 (SEQ ID NO:127). P-0382 (SEQ IDNO:51) отличается от Р-0320 только последовательностями линкеров; линкер L1 Р-0382 представляет собой гибкий линкер (G3S)2 (SEQ ID NO: 115), а линкер L2 представляет собой линкер, расщепляемый ММР-2/9 (SEQ ID NO:95). Р-0362 (SEQ ID NO:) и Р-0379 (SEQ ID N0:59) отличаются от Р-382 одной точковой мутацией. Р-0362 содержит мутацию К38Е в домене IL-2RαSushi, в то время как Р-0379 содержит замену F42A в домене IL-2. Р-0250 (SEQ ID NO:48) представляет собой Fc-слитый белок IL-2, где IL-2 слит с С-концом Fc с использованием гибкого линкера (G3S)2 (SEQ ID NO: 115).
Эксклюзионные диаграммы этих 5 молекул показаны на ФИГ. 33А-33Е. Из представленных хроматограмм очевидно, что все четыре конструкции IL-2 VitoKine имеют значительно лучший профиль чистоты по сравнению с Fc-слитым белком IL-2. Р-0250 содержит более 25% нежелательных высокомолекулярных частиц. В отличие от этого, все четыре молекулы VitoKine демонстрируют острые пики мономера с содержанием мономера более 96%. Изменения линкеров, мутации в IL-2 или IL-2RαSushi не оказывали влияния на качество. Такое значимое повышение качества белка было, вероятно, обусловлено слиянием домена IL-2αSushi с VitoKine.
Помимо качества белка был также повышен уровень экспрессии конструкций IL-2 VitoKine, особенно в формате VitoKine с линкером GS между Fc и IL-2 и линкером из 10 аминокислот, активируемым ММР-2/9, между IL-2 и IL-2RαSushi. Несмотря на то, что уровни экспрессии белка в разных партиях могут варьировать из-за условий роста клеток, очевидно, что уровень экспрессии конструкций VitoKines стабильно превышает уровень экспрессии Fc-слитого белка IL-2 в несколько раз. В Таблице 14 приведены титры экспрессии белка в мг/л и процент мономера белка.

Кроме того, усилия авторов изобретения по получению конструкций IL-2 также привели к обнаружению одиночной аминокислотной замены серина на изолейцин в положении 125, которая приводила к универсальному расширению возможностей разработки Fc-слитых конструкций IL-2 с полным сохранением биологической активности. Замена на Не в положении 125 IL-2 дикого типа и вариантах IL-2 с различным мутационным контекстом в Fc-слитом формате приводила к повышению уровня экспрессии в 4-11 раз и одинаково низкой склонности к агрегации. Сводные данные по уровню экспрессии в мг/л и чистоте вещества, очищенного на белке А, оцененной SEC-хроматографией, в виде процента агрегации для типичных молекул представлены в Таблице 15. Две молекулы в одной строке Таблицы 15 имеют одинаковую(ые) другую(ие) аминокислотную(ые) замену(ы) и различаются только остатком 125, представляющим собой серии или изолейцин. В качестве примера на ФИГ. 33F показана SEC-хроматограмма молекулы-аналога Р-250 с S125I в IL-2.

В заключение, платформа VitoKine значительно расширяла возможности разработки белков, что было продемонстрировано повышением экспрессии белка и существенным снижением склонности конструкций Fc IL-2 VitoKine к агрегации. Кроме того, конструкции IL-2 VitoKine (с IL-2 дикого типа или вариантами IL-2), содержащие полезную аминокислоту S125I в IL-2, обеспечат еще более широкие возможности разработки.
Пример 19
Выбор домена D3 VitoKine может сильно влиять на экспрессию белка
Платформа VitoKine была также изучена с доменами D3, представляющими собой вариант когнатного рецептора домена D2 или домена неродственного белка. Исходя из результатов кристаллографического анализа (Wang et al., Science 310: 1159-1163, 2005), sushi-домены 1 и 2 IL-2Rα участвуют в обмене цепей, в результате чего остатки 1-19 IL-2Rα являются частью sushi-домена 2, а остатки 102-122 являются частью sushi-домена 1. Такое структурное расположение было воспроизведено в варианте IL-2RαSushi (SEQ ID NO: 147), содержащем остатки 102-122 IL-2Rα (SEQ ID NO:10) на N-конце и остатки 20-68 IL-2Rα на С-конце. Такой вариант IL-2RαSushi содержит большинство остатков, взаимодействующих с IL-2, и, предположительно, будет иметь большую часть активности, сохраняя структурную целостность. Замена домена IL-2RαSushi в Р-0320 (SEQ ID NO:49) на вариант IL-2RαSushi приводила к получению IL-2 VitoKine Р-0321 (SEQ ID NO:179). Неожиданно, Р-0321, содержавшая вариант IL-2RαSushi в качестве домена D3, не поддавалась экспрессии или экспрессировалась на таком низком уровне, который не позволял провести ее выделение и очистку.
Сходным образом, домен IL-15aSushi+в VitoKine Р-0315 (SEQ ID NO:33) был заменен на IL-2RαSushi (SEQ ID NO:10), и полученный белок представляет собой Р-0389 (SEQ ID NO:42). Уровень экспрессии Р-0389 был значительно ниже уровня экспрессии Р-0315. Еще примечательнее то, что Р-0389 представляла собой, главным образом, высокомолекулярные агрегаты, что продемонстрировано на изображении SDS-PAGE-геля, показанном на ФИГ. 34А. Для сравнения на ФИГ. 34В показано изображение SDS-PAGE-геля с аналогичной молекулой Р-0315. Кроме того, очищенная Р-0389 была устойчива к расщеплению под действием ММР-2, несмотря на присутствие пептида-субстрата ММР-2/9 в ее последовательности, что указывает на неправильный фолдинг молекулы или ограничение доступности для протеазы вследствие агрегации.
В заключение, D3 является критически важным компонентом конструкций VitoKine. Помимо выполнения функций маскирующей группировки, он может оказывать сильное влияние на возможности разработки белка, как положительным, так и отрицательным образом.
В свете настоящего изобретения, все изделия и способы, раскрытые и заявленные здесь, могут быть изготовлены и осуществлены без ненужных экспериментов. Несмотря на то, что изделия и способы по данному изобретению были описаны посредством предпочтительных воплощений, специалистам в данной области будет ясно, что указанные изделия и способы могут быть изменены без выхода за рамки сущности и объема изобретения. Подразумевается, что все такие измененные варианты и эквиваленты, очевидные специалистам в данной области, существовавшие на момент подачи данной заявки или разработанные позднее, включены в сущность и объем изобретения, определенные приложенной формулой изобретения. Все патенты, заявки на патенты и публикации, упомянутые в данном описании, указывают на уровень специалистов в области, к которой относится изобретение. Все патенты, заявки на патенты и публикации полностью включены сюда посредством ссылки для любых задач и в такой же степени, как если бы было конкретно и по отдельности указано, что каждая отдельная публикация полностью включена посредством ссылки для любых задач. Изобретение, наглядно описанное здесь, может быть подходящим образом применено в отсутствие любого(ых) элемента(ов), не раскрытого(ых) здесь конкретно. Таким образом, следует понимать, что, несмотря на то, что настоящее изобретение было конкретно раскрыто посредством предпочтительных воплощений и возможных признаков, специалисты в данной области могут прибегнуть к модификациям и вариантам концепций, раскрытых здесь, и что такие модификации и варианты рассматриваются как входящие в объем данного изобретения, определенный приложенной формулой изобретения.
Перечень последовательностей
Последовательности нуклеиновых кислот и аминокислотные последовательности, приведенные в приложенном перечне последовательностей, показаны с применением стандартных буквенных сокращений нуклеотидных оснований и трехбуквенных кодов аминокислот, как определено в части 1.822 раздела 37 Свода федеральных нормативных актов США (37 C.F.R. 1.822).
SEQ ID NO: 1 представляет собой аминокислотную последовательность предшественника человеческого IL-15.
SEQ ID NO: 2 представляет собой аминокислотную последовательность зрелой формы человеческого IL-15.
SEQ ID NO: 3 представляет собой аминокислотную последовательность полипептида варианта IL-15.
SEQ ID NO: 4 представляет собой аминокислотную последовательность человеческого IL- 15Rα.
SEQ ID NO: 5 представляет собой аминокислотную последовательность sushi-flOMeHa+человеческого IL- 15Rα.
SEQ ID NO: 6 представляет собой аминокислотную последовательность предшественника человеческого IL-2.
SEQ ID NO: 7 представляет собой аминокислотную последовательность зрелой формы встречающегося в природе человеческого IL-2.
SEQ ID NO: 8 представляет собой аминокислотную последовательность зрелой формы человеческого IL-2 дикого типа.
SEQ ID NO: 9 представляет собой аминокислотную последовательность предшественника человеческого IL-2Rα (CD25).
SEQ ID NO: 10 представляет собой аминокислотную последовательность sushi-домена человеческого IL-2Rα.
SEQ ID NO: 11 представляет собой аминокислотную последовательность предшественника человеческого IL-2Rβ
SEQ ID NO: 12 представляет собой аминокислотную последовательность внеклеточного домена человеческого IL-2Rβ.
SEQIDNO: 13 представляет собой аминокислотную последовательность Fc человеческого IgGl.
SEQ ID NO: 14 представляет собой последовательность Fc человеческого IgGl со сниженной/устраненной эффекторной функцией.
SEQ ID NO: 15 представляет собой аминокислотную последовательность Knob-Fc.
SEQ ID NO: 16 представляет собой аминокислотную последовательность Hole-Fc.
SEQ ID NO: 17 представляет собой аминокислотную последовательность зрелой формы человеческого IL-4.
SEQ ID NO: 18 представляет собой аминокислотную последовательность зрелой формы человеческого IL-7.
SEQ ID NO: 19 представляет собой аминокислотную последовательность зрелой формы человеческого IL-9.
SEQ ID NO: 20 представляет собой аминокислотную последовательность зрелой формы человеческого IL-10.
SEQ ID NO: 21 представляет собой последовательность альфа-субъединицы зрелой формы человеческого IL-12.
SEQ ID NO: 22 представляет собой последовательность бета-субъединицы зрелой формы человеческого IL-12.
SEQ ID NO: 23 представляет собой последовательность альфа-субъединицы зрелой формы человеческого IL-23.
SEQ ID NO: 24 представляет собой последовательность бета-субъединицы зрелой формы человеческого IL-27.
SEQ ID NO: 25-43 представляют собой аминокислотные последовательности различных конструкций Fc IL-15 VitoKine.
SEQ ID NO: 44 представляет собой аминокислотную последовательность слитого белка Hole-Fc-IL-15.
SEQ ID NO: 45 представляет собой аминокислотную последовательность слитого белка Knob-Fc-IL-15.
SEQ ID NO: 46 представляет собой аминокислотную последовательность слитого белка Knob-Fc-IL- 15Rα-Sushi+.
SEQ ID NO: 47 представляет собой аминокислотную последовательность слитого белка Fc-IL-15 S58D.
SEQ ID NO: 48 представляет собой аминокислотную последовательность слитого белка IL-2.
SEQ ID NO: 49-65 представляют собой аминокислотные последовательности различных конструкций Fc IL-2 VitoKine.
SEQ ID NO: 66-70 представляют собой аминокислотные последовательности различных конструкций IL-15, содержащих блокирующий пептид.
SEQ ID NO: 71-87 и 157-159 представляют собой аминокислотные последовательности различных пептидов-субстратов протеаз.
SEQ ID NO: 88-96 и 160-161 представляют собой аминокислотные последовательности различных линкеров, расщепляемых протеазами, содержащих различные спейсерные пептиды, фланкирующие пептиды-субстраты протеаз.
SEQ ID NO: 97-106 представляют собой аминокислотные последовательности различных блокирующих пептидных последовательностей.
SEQ ID NO: 107-127 представляют собой аминокислотные последовательности различных нерасщепляемых линкерных последовательностей.
SEQ ID NO: 128-146 представляют собой аминокислотные последовательности различных конструкций VitoKine с антителами.
SEQ ID NO: 147 представляет собой последовательность варианта человеческого IL-2Rα.
SEQ ID NO: 148-149 представляют собой аминокислотные последовательности слитых конструкций Hole-Fc-IL-15.
SEQ ID NO: 150-155 представляют собой аминокислотные последовательности различных конструкций Fc IL-2 VitoKine.
SEQ ID NO: 156 представляет собой последовательность Fc человеческого IgGl со сниженной/устраненной эффекторной функцией и увеличенным периодом полувыведения.
SEQ ID NO: 162-165 представляют собой аминокислотные последовательности различных конструкций Fc IL-15 VitoKine.
SEQ ID NO: 166 представляет собой последовательность Fc человеческого IgGl со сниженной/устраненной эффекторной функцией и увеличенным периодом полувыведения.
SEQ ID NO: 167 представляет собой аминокислотную последовательность Knob-Fc с увеличенным периодом полувыведения.
SEQ ID NO: 168 представляет собой аминокислотную последовательность Hole-Fc с увеличенным периодом полувыведения.
SEQ ID NO: 169-174 представляют собой аминокислотные последовательности различных конструкций Fc IL-15 VitoKine.
SEQ ID NO: 175-178 представляют собой аминокислотные последовательности различных Fc-слитых конструкций IL-15.
SEQ ID NO: 179 представляет собой аминокислотную последовательность конструкции Fc IL-2 VitoKine.
SEQ ID NO: 180-181 представляют собой аминокислотные последовательности различных конструкций антитело IL-15 VitoKine.
SEQ ID NO: 182-192 представляют собой нуклеотидные последовательности различных конструкций Fc IL-15 VitoKine.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Последовательность предшественника человеческого IL-15

Последовательность зрелой формы человеческого IL-15

Мутеин человеческого IL-15 S58D

Последовательность предшественника человеческого IL- 15Rα

Человеческий IL- 15Rα, sushi-домен+
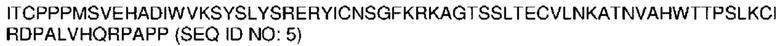
Последовательность предшественника человеческого IL-2

Последовательность зрелой формы встречающегося в природе человеческого IL-2

Последовательность зрелой формы человеческого IL-2 дикого типа

Последовательность предшественника человеческого IL-2Rα (CD25)

Человеческий IL-2RαSushi

Последовательность предшественника человеческого IL-2RP

Последовательность внеклеточного домена человеческого IL-2RP

Fc человеческого IgGl

Fc человеческого IgGl со сниженной/устраненной эффекторной функцией

Knob-Fc

Hole-Fc

Последовательность зрелой формы человеческого IL-4

Последовательность зрелой формы человеческого IL-7

Последовательность зрелой формы человеческого IL-9

Последовательность зрелой формы человеческого IL-10

Последовательность альфа-субъединицы зрелой формы человеческого IL-12

Последовательность бета-субъединицы зрелой формы человеческого IL-12

Последовательность альфа-субъединицы зрелой формы человеческого IL-23

Последовательность зрелой формы человеческого TGF-бета

Р-0351

Hole-цепь Р-0170

Р-0172

Р-0202

Р-0203

P-0204

Р-0205

Р-0206

Р-0315

Р-0316

Р-0350

Р-0354

Р-0355

Р-0385

Р-0386

Р-0387

Р-0388

Р-0389

Р-0397

Hole-Fc-IL-15

Knob-Fc-IL-15

Knob-Fc-IL-15Rα-Sushi+

Fc-IL-15 S58D

P-0250

P-0320

P-0352

Р-0382

Р-0398

Р-0362

Р-0380

Р-0384

Р-0400

Р-0404

Р-0399

Р-0379

Р-0381

Р-0383

Р-0329

Р-0401

Р-0402

Р-0403

Hole-Fc-15p1

Hole-Fc-15p2

Hole-Fc-15p3

p1'-15-Fc

p3'-15-Fc

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы
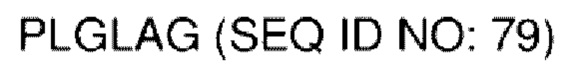
Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы
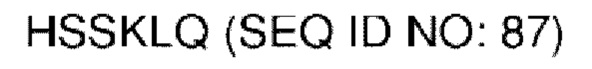
Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Последовательность пептида

Последовательность пептида

Последовательность пептида

Последовательность пептида

Последовательность пептида
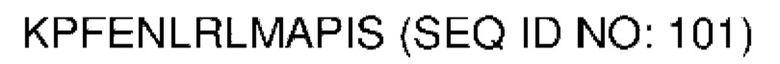
Последовательность пептида

Последовательность пептида

Последовательность пептида

Последовательность пептида

Последовательность пептида

Последовательность нерасщепляемого линкера
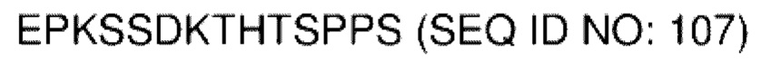
Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера
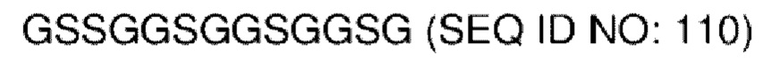
Последовательность нерасщепляемого линкера
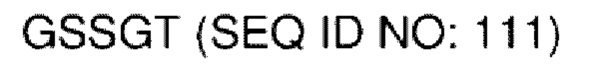
Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера
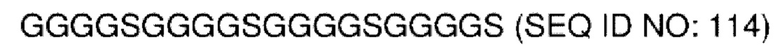
Последовательность нерасщепляемого линкера
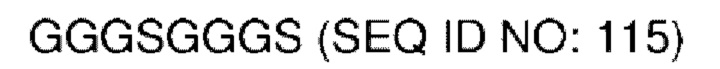
Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера
Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера

Последовательность нерасщепляемого линкера
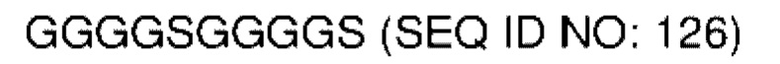
Последовательность нерасщепляемого линкера

JS001-IL-15-VitoKine-HC

JS001-Lк

Ипилимумаб-IL-15-VitoKine-HC

Ипилимумаб-Lк

RO7009789-IL-15-VitoKine-HC

RO7009789-Lk

L19-IL-15-VitoKine-HC

L19-Lк

Ритуксимаб-IL-2-VitoKine-HC

Ритуксимаб-Lк

Герцептин-IL-2-VitoKine-НС

Герцептин-Lк

Цетуксимаб-IL-2-VitoKine-НС

Цетуксимаб-Lк

JS001-IL-2-VitoKine-HC

Ведолизумаб-IL-2-VitoKine-HC

Ведолизумаб-Lк

Хумира-IL-2-VitoKine-HC

Хумира-Lк

Измененный домен IL-2RαSushi
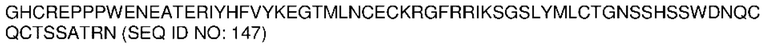
Hole-Fc-IL-15-2

Hole-Fc-IL-15-3

Р-0420

Р-0421

Р-0423

Р-0424

Р-0425

Р-0426

Fc человеческого IgG1 со сниженной/устраненной эффекторной функцией н увеличенным периодом полувыведения

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность пептида-субстрата протеазы

Последовательность линкера, расщепляемого протеазой

Последовательность линкера, расщепляемого протеазой

Р-0660

Р-0488

Р-0489

Р-0661

Fc человеческого IgG1 со сниженной/устраненной эффекторной функцией и увеличенным периодом полувыведения in vivo

Knob-Fc с увеличенным периодом полувыведения in vivo

Hole-Fc с увеличенным периодом полувыведения in vivo

Р-0650

Р-0651

Hole-цепь Р-0662

Hole-цепь Р-0663 с увеличенным периодом полувыведения

Hole-цепь Р-0664 с увеличенным периодом полувыведения

Hole-цепь Р-0665 с увеличенным периодом полувыведения

Knob-цепь Р-0156

Hole-цепь Р-0156

Benchmark, цепь 1

Benchmark, цепь 2

Р-0321

Тецентрик-IL-15-VitoKine-HC

Тецентрик-Lк

Р-0315

Р-0350

Р-0351

Р-0650

Р-0651

Цепь 1 Р-0662

Цепь 2 Р-0662

Цепь 1 Р-0663

Цепь 1 Р-0664

Цепь 1 Р-0665

Цепь 2 Р-0663/Р-0664/Р-0665

| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОЦИТОКИНЫ НА ОСНОВЕ IL-15 И IL-R[альфа] ДОМЕНА SUSHI | 2012 |
|
RU2644671C2 |
| Новые варианты интерлейкина-2 для лечения рака | 2020 |
|
RU2833537C2 |
| ГЕТЕРОДИМЕРНЫЙ БЕЛОК IL-15 И ЕГО ПРИМЕНЕНИЯ | 2014 |
|
RU2689717C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2642310C2 |
| ОДНОЦЕПОЧЕЧНЫЕ БЕЛКИ-АГОНИСТЫ РЕЦЕПТОРА CD40 | 2016 |
|
RU2745801C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2727452C1 |
| СЛИТЫЕ ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ДОМЕН WAP, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2639526C2 |
| БЕЛКОВЫЙ КОМПЛЕКС ИНТЕРЛЕЙКИНА 15 И ЕГО ПРИМЕНЕНИЕ | 2015 |
|
RU2711979C2 |
| СЛИТЫЕ БЕЛКИ IL-7 | 2004 |
|
RU2369616C2 |
| ИММУНОЦИТОКИН, ВКЛЮЧАЮЩИЙ ГЕТЕРОДИМЕРНЫЙ БЕЛКОВЫЙ КОМПЛЕКС НА ОСНОВЕ IL-15/IL-15Rα, И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2753282C2 |

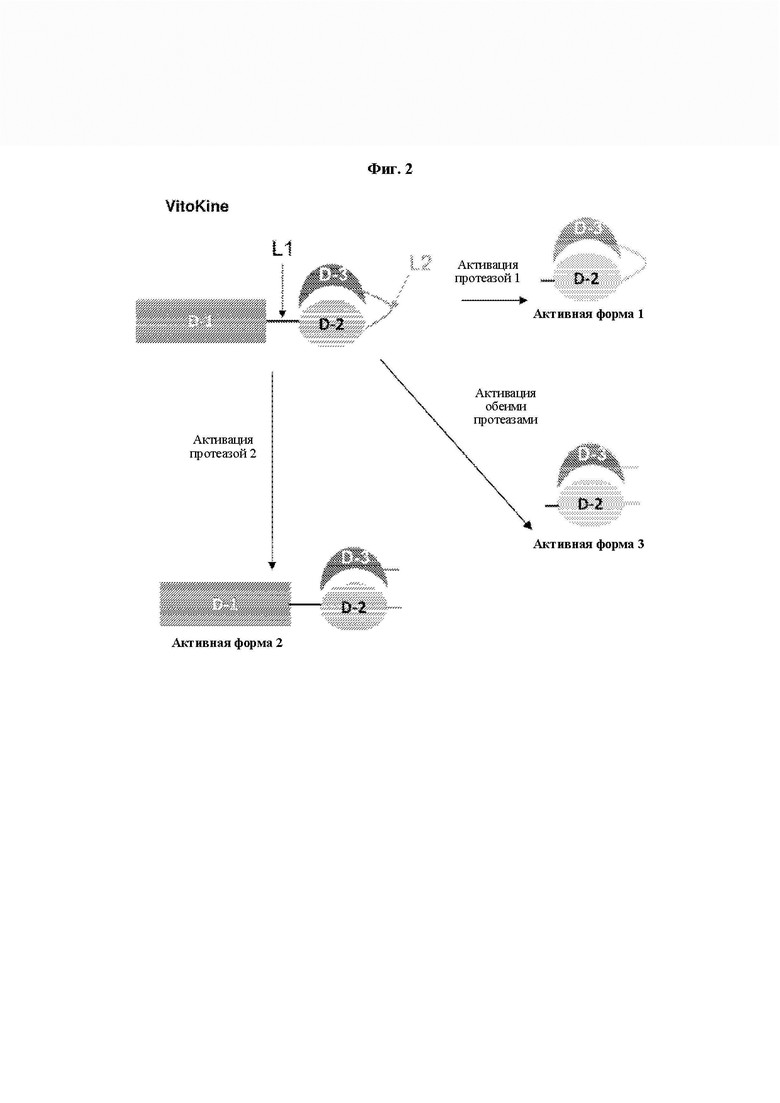

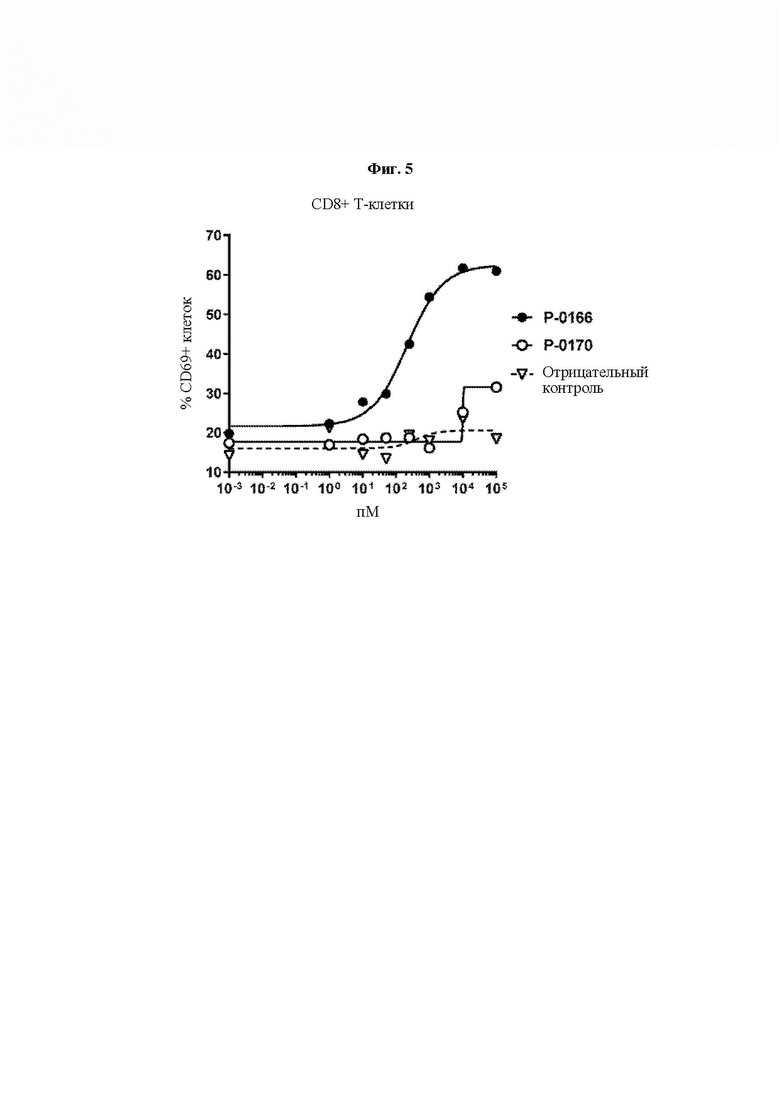

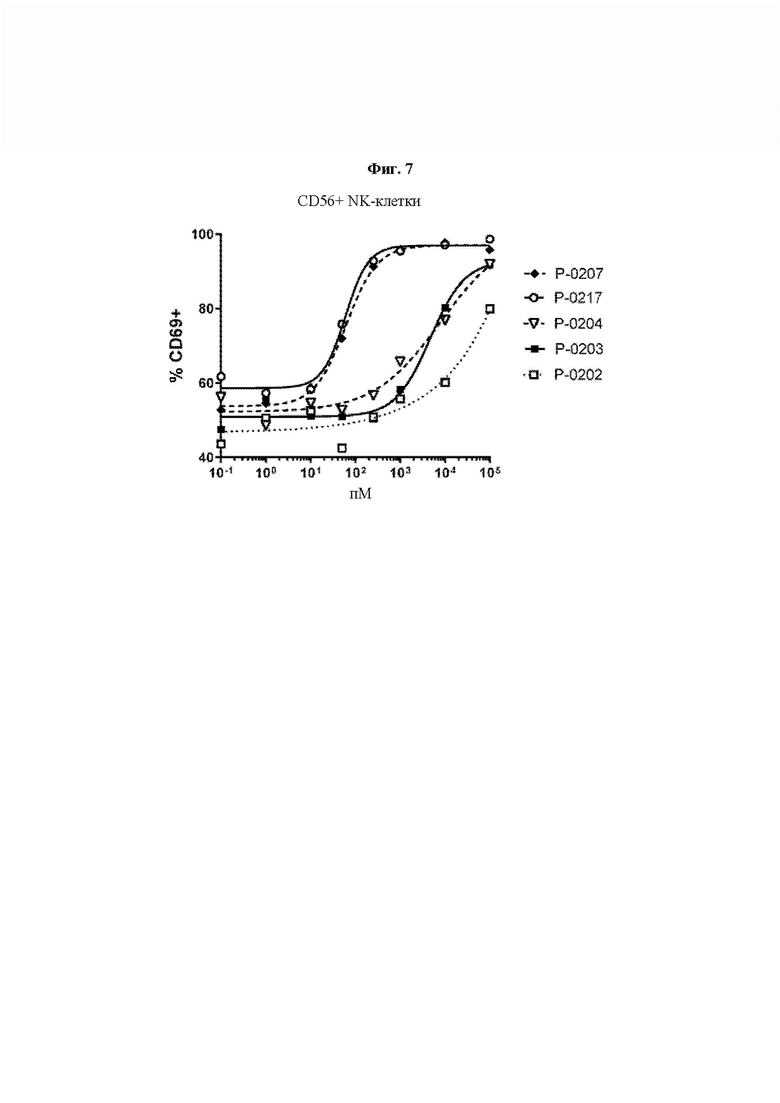
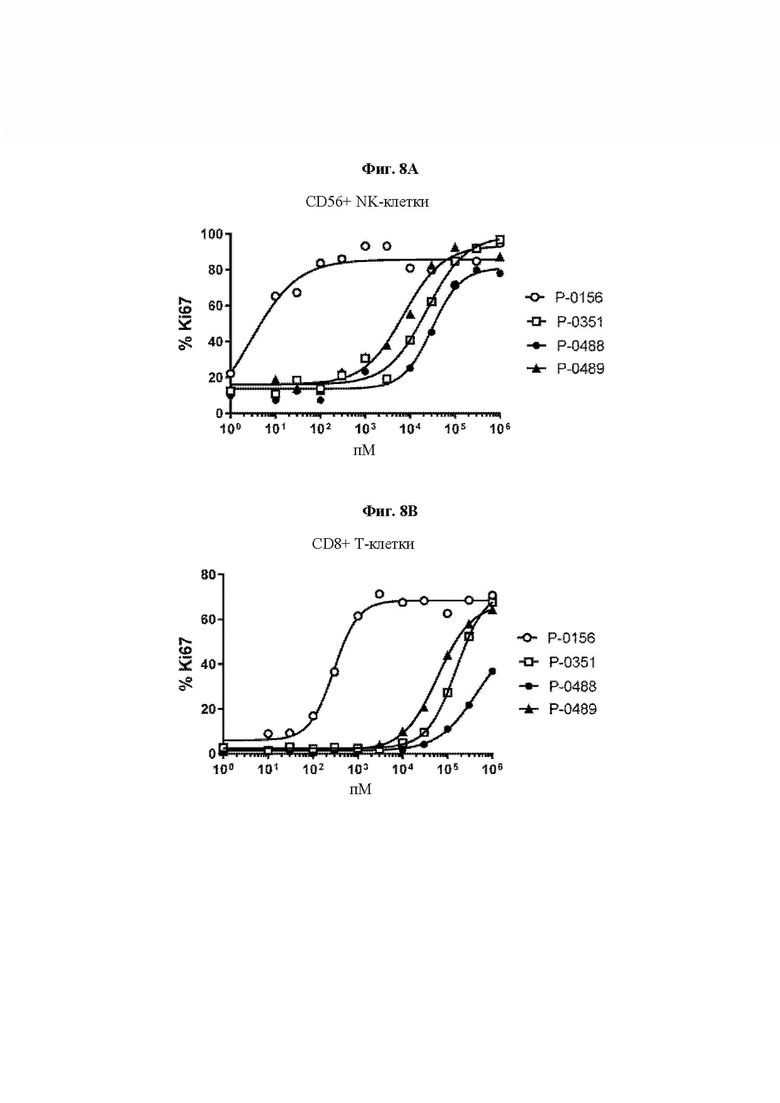

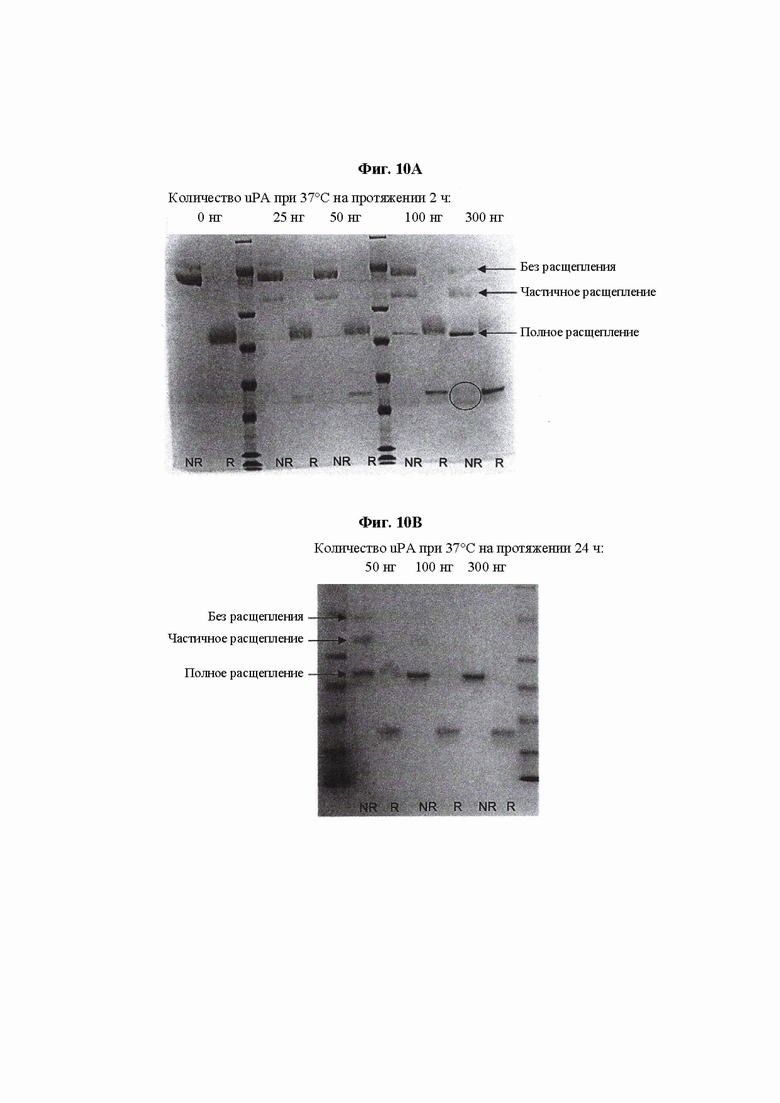
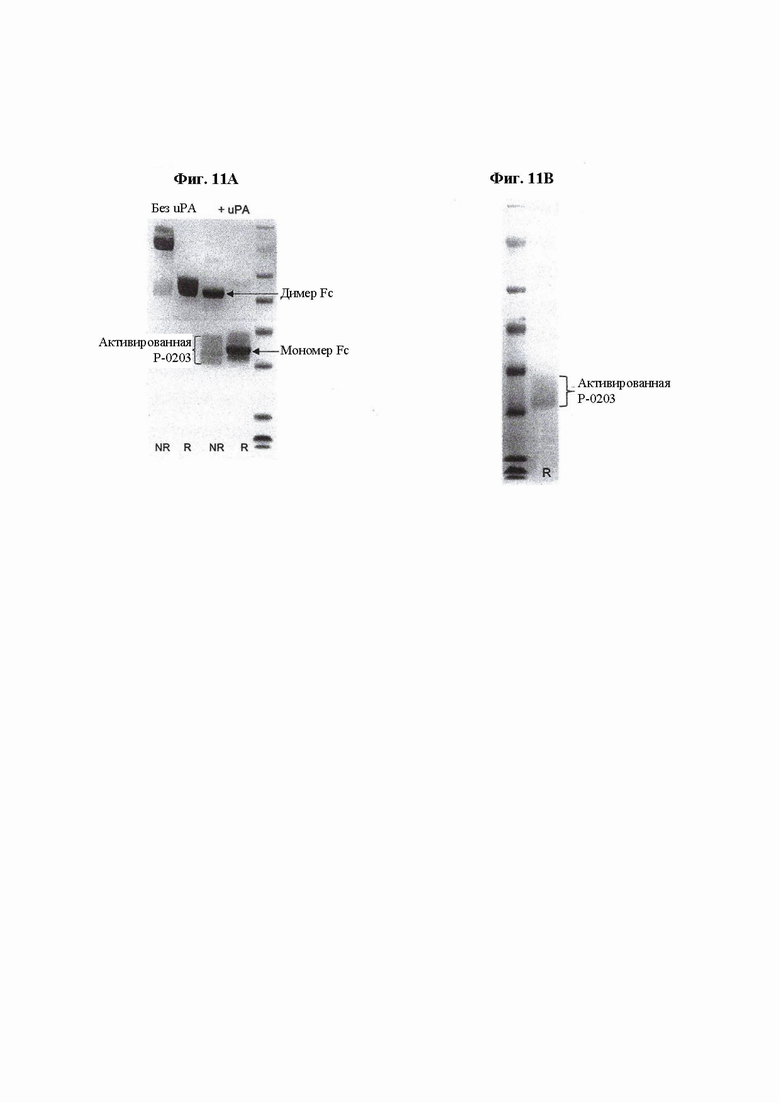










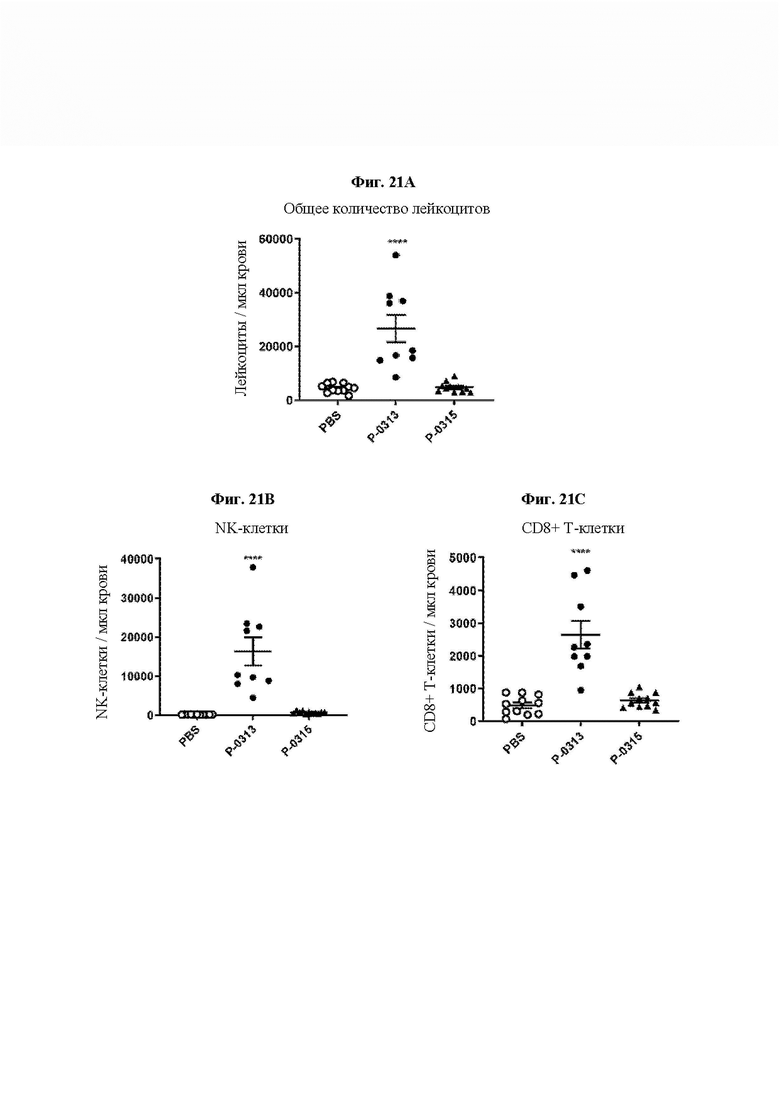

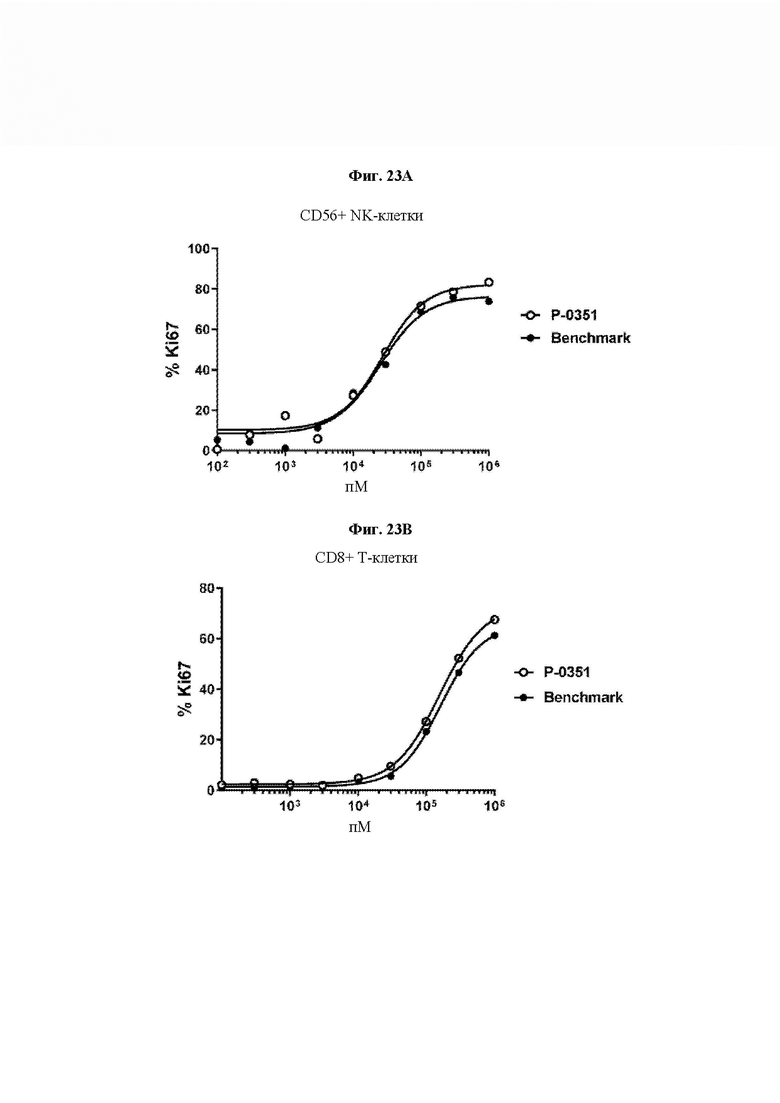

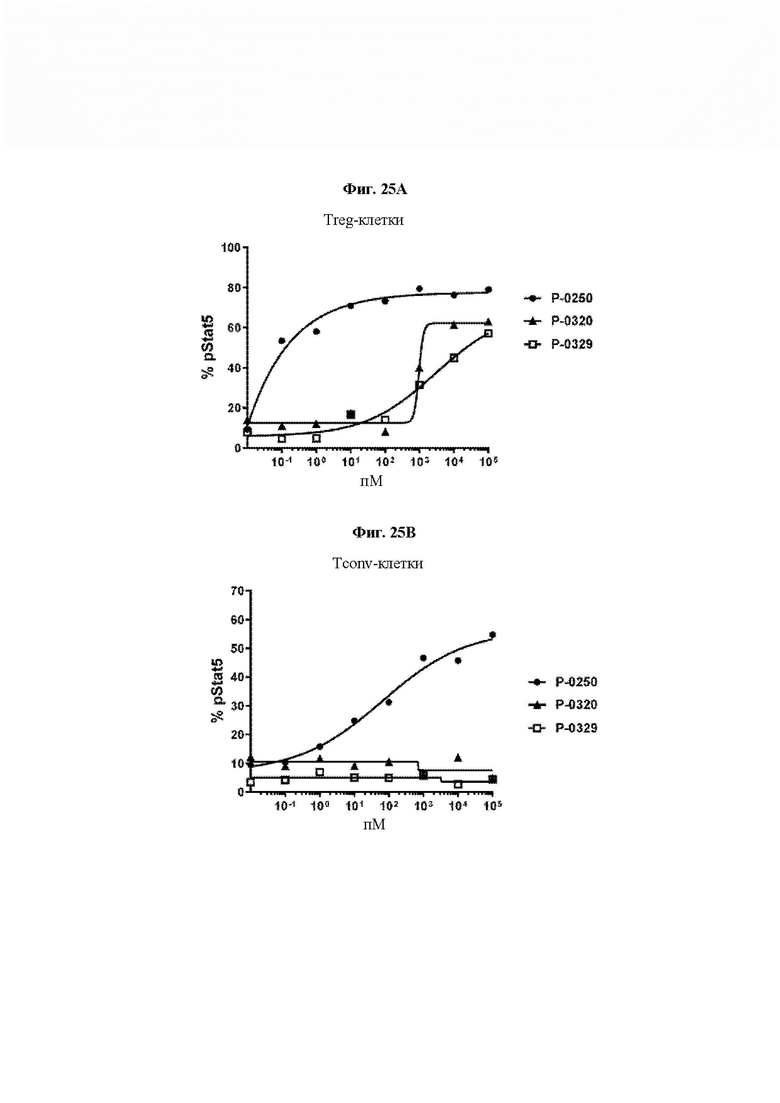


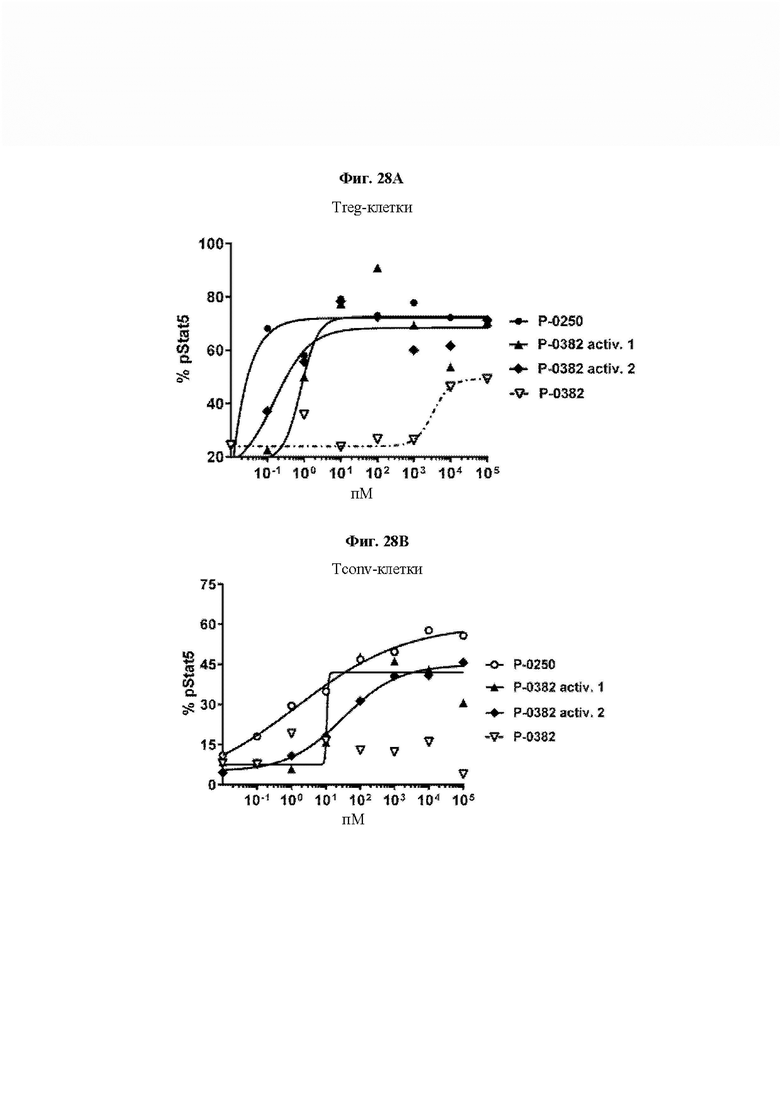


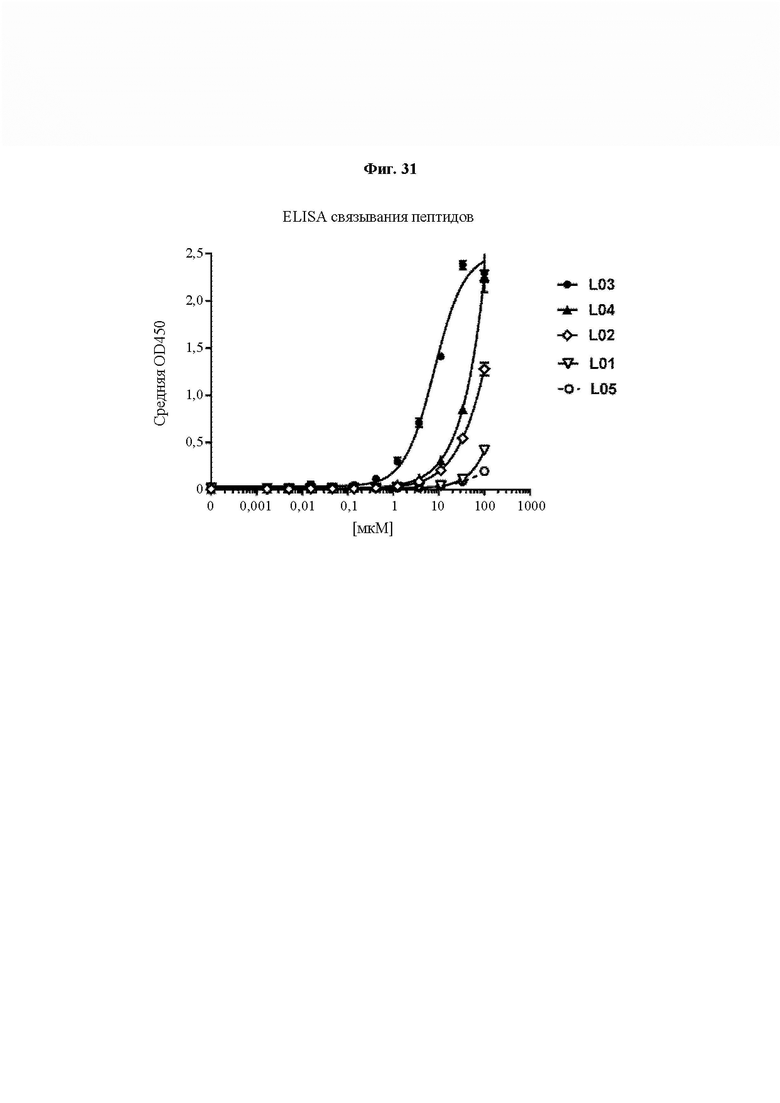
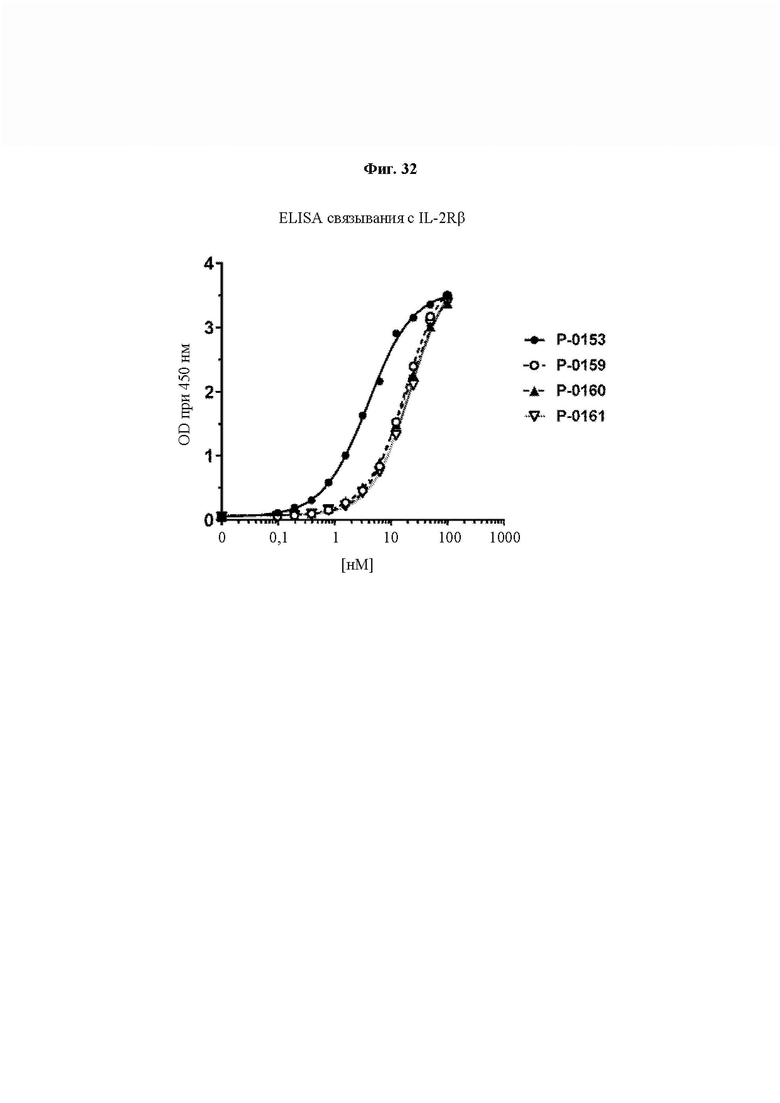


Настоящее изобретение относится к области биотехнологии, в частности к новым пептидным конструкциям на основе интерлейкинов, и может быть использовано в медицине. Согласно настоящему изобретению предложена платформа биологически активируемой лекарственной конструкции на основе цитокинов (VitoKine), направленная на снижение системной токсичности и обеспечение более широкого терапевтического применения интерлейкина-15 (IL-15) для лечения рака, аутоиммунных заболеваний, воспалительных заболеваний, вирусной инфекции, трансплантации и различных других расстройств. Новые конструкции VitoKine по настоящему изобретению содержат: (1) домен D1 группировки, направленной на ткань или очаг заболевания; (2) домен D2 биологически активируемой группировки; и (3) домен D3 маскирующей группировки. Важно, что сохранение инертности «активной группировки» конструкции VitoKine до ее местной активации протеазами, регуляция которых повышена в тканях, пораженных заболеванием, будет ограничивать связывание активной группировки с ее рецепторами или мишенями на периферии или на поверхности клеток и тканей, не пораженных заболеванием, предотвращая избыточную активацию ее пути и уменьшая нежелательную токсичность, обусловленную связыванием с мишенью вне целевой ткани. Кроме того, инертность активной группировки VitoKine до активации протеазами будет существенно снижать возможный клиренс, обусловленный связыванием с антигеном или мишенью, и, таким образом, увеличивать период полувыведения in vivo и улучшать биораспределение, биодоступность и терапевтическую эффективность. 2 н. и 13 з.п. ф-лы, 19 пр., 15 табл., 34 ил.
1. Иммуномодулирующая биологически активируемая полипептидная лекарственная конструкция, содержащая, в направлении от N-конца к C-концу (D1-D2-D3):
(1) домен D1 (D1), выбранный из группировки, направленной на ткань или очаг заболевания, где функция D1 состоит в направлении биологически активируемой группировки к намеченному месту терапии;
группировки, увеличивающей период полувыведения, где функция D1 состоит в направлении биологически активируемой группировки к намеченному месту терапии и увеличении периода полувыведения D2, и
группировки с двойной функцией, где функция D1 состоит в направлении и удержании биологически активируемой группировки в намеченном месте терапии;
(2) домен D2 биологически активируемой группировки (D2); и
(3) домен D3 маскирующей группировки (D3);
где D3 способен маскировать функциональную активность D2 до активации в намеченном месте терапии;
где D1 представляет собой Fc-домен, имеющий аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, изложенных в SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 156 и SEQ ID NO: 166-168;
где домен D2 представляет собой полипептид IL-15, имеющий аминокислотную последовательность, изложенную в SEQ ID NO: 2, полипептид варианта IL-15, содержащий одну или более чем одну аминокислотную замену или делецию в положении 30, 31, 32, 58, 62, 63, 67, 68 или 108 SEQ ID NO: 2, или полипептид варианта IL-15, имеющий аминокислотную последовательность, изложенную в SEQ ID NO: 3; и
где домен D3 является когнатным рецептором/партнером по связыванию или его вариантом для IL-15 и содержит аминокислотную последовательность, изложенную в SEQ ID NO: 4, или ее любой функциональный фрагмент или содержит аминокислотную последовательность, изложенную в SEQ ID NO: 5.
2. Конструкция по п. 1, где каждый из доменов D1, D2 и D3 конструкции имеет форму мономера, форму димера или они взятые вместе имеют форму комбинации димера и мономера.
3. Конструкция по п. 1 или 2, где D2 соединен с D1 пептидным линкером (L1), выбранным из группы, состоящей из пептидного линкера, расщепляемого протеазой, и нерасщепляемого пептидного линкера.
4. Конструкция по п. 3, где пептидный линкер, расщепляемый протеазой, выбран из группы последовательностей, изложенных в SEQ ID NO: 71-96 и 157-161.
5. Конструкция по п. 3, где нерасщепляемый пептидный линкер выбран из группы последовательностей, изложенных в SEQ ID NO: 107-127.
6. Конструкция по любому из пп. 1-5, где D2 соединен с D3 пептидным линкером (L2), выбранным из группы, состоящей из пептидного линкера, расщепляемого протеазой, и нерасщепляемого пептидного линкера.
7. Конструкция по п. 6, где пептидный линкер, расщепляемый протеазой, выбран из группы последовательностей, изложенных в SEQ ID NO: 71-96 и 157-161.
8. Конструкция по п. 6, где нерасщепляемый пептидный линкер выбран из группы последовательностей, изложенных в SEQ ID NO: 107-127.
9. Конструкция по любому из пп. 1-8, где оба из L1 и L2 представляют собой пептидные линкеры, расщепляемые протеазами.
10. Конструкция по любому из пп. 1-8, где оба из L1 и L2 представляют собой нерасщепляемые пептидные линкеры.
11. Конструкция по любому из пп. 1-8, где L1 представляет собой пептидный линкер, расщепляемый протеазой, и L2 представляет собой нерасщепляемый пептидный линкер.
12. Конструкция по любому из пп. 1-8, где L1 представляет собой нерасщепляемый пептидный линкер и L2 представляет собой пептидный линкер, расщепляемый протеазой.
13. Конструкция по любому из пп. 1-12, где указанная конструкция выбрана из группы конструкций, содержащих аминокислотные последовательности, изложенные в SEQ ID NO: 25-43, 162-165 и 169-175, 180-181, 49-65, 150-155, 179 или 128-146.
14. Иммуномодулирующая фармацевтическая композиция, содержащая терапевтически эффективное количество конструкции по любому из пп. 1-13 в смеси с фармацевтически приемлемым носителем.
15. Фармацевтическая композиция по п. 14 для использования в лечении рака или метастазирования рака у субъекта, включающего введение субъекту терапевтически эффективного количества указанной фармацевтической композиции.
| WO 2017156178 A1, 14.09.2017 | |||
| WO 2008096158 A1, 14.08.2008 | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| ПЕПТИД-АНТАГОНИСТ АКТИВНОСТИ ИНТЕРЛЕЙКИНА-15 | 2009 |
|
RU2506270C2 |
| WO 2017156178 A1 (MAVERICK THERAPEUTICS INC.), 14.09.2017, реферат, параграфы [0005], [0009], [0012], [0015], [0237], | |||

