Область техники
Изобретение относится к биотехнологии, генной инженерии и медицине и может быть использовано для создания растений - суперпродуцентов антител, в частности против HER2/neu-позитивных раковых клеток. Такие антитела могут быть использованы для диагностики и лечения рака молочной железы.
Предшествующий уровень техники
Рак молочной железы поражает сейчас каждую девятую женщину, и его распространение с каждым годом увеличивается. Лечение на ранней стадии заболевания - это хирургическое воздействие. Однако коварство и опасность рака молочной железы состоит в быстром метастазировании, и тогда назначается химио- и радиотерапия. Несколько лет назад в медицинскую практику был введен трастузумаб, созданный фирмой Дженентек (Genentech, США) и производимый фирмой Хоффман-Ля Рош (Hoffman La Roche, Швейцария). Это лекарство создано на основе гуманизированных моноклональных антител (MA) (huMab4D5-8 или rhuMabHer2), производимых мышиной гибридомой АТСС CRL-10463 из коллекции штаммов микроорганизмов и клеточных культур США. Эти получившие широкое распространение антитела - всего лишь один из типов известных антител, взаимодействующих с онкобелком HER2 (от Human Epidermal growth factor Receptor 2). Онкоген HER2/neu представляет собой трансмембранную тирозиновую протеинкиназу ErbB2 с молекулярной массой 185 кДа. Примерно у 30% больных раком молочной железы, страдающих так называемой HER2/neu-позитивной и наиболее опасной формой рака, наблюдается суперпродукция этого онкобелка, что вызывает ускоренное метастазирование и устойчивость к терапевтическому воздействию. Антитело, введенное в кровоток пациента, взаимодействует с внеклеточный частью HER2/neu и угнетает деление раковых клеток, редко сопровождающееся разрушением раковых клеток. В комбинации с химиотерапией антитела huMab4D5-8 оказывают выраженный лечебный эффект. Фирма Дженентек свою заявку на использование МА против раковых клеток SK-BR-3 молочной железы, суперэкспрессирующих онкоген Her2/neu, впервые подала в Патентное Ведомство США 5 августа 1994 года и получила 14 октября 1997 года патент за номером US 5,677,171 (изобретатели Hudziak R.M., Shepard Н.М., Ullrich A., Fendly В.М.). Суммарно формула изобретения Дженентек декларирует (а) создание МА против рецептора Her2/neu, ингибирующих рост опухолевых клеток; (б) МА, связывающие внеклеточный домен Her2/neu и сенсибилизирующие клетки к действию цитотоксического фактора. В последующем на протяжении первого полугодия 1998 были поданы заявки на использование huMab4D5-8 в клинике (патент US 6,165,464, WO9704801, WO9817797). Чуть позже использование МА 454С11, продуцируемых гибридомой НВ 8484, для этой же цели предложила компания Хирон (Chiron Corp., США) (заявка от 7 июня 1995 года), и ей 25 апреля 2000 года был выдан патент US 6,054,561.
В России успешно используется гуманизированное МА huMab4D5-8, известное как трастузумаб или под торговой маркой - «герцептин». Показано, что использование герцептина снижает риск развития отдаленных метастазов и увеличивает продолжительность жизни пациентов (Ганьшина и Личиницер. 2005. Фарматека №18, 8-11). Стоимость лечения Her2/neu-позитивного рака молочной железы этим препаратом очень высока и составляет 70000 долларов США [Fleck L. "The costs of caring: Who pays? Who profits? Who panders?" Hastings Cent Rep 36 (3):13-17]. Вообще, стоимость продукции любого целевого белка в культуре клеток животных достаточно высока и превышает 10000 долларов США за 1 г белка (Danielli et al., 2001. TRENDS in Plant Science 6, 219-226). Это в несколько раз выше, чем стоимость продукции аналогичного белка в дрожжах, клетках насекомых и тем более в бактериях. Обычно МА получают, используя культуру клеток животных, клетки яичника китайского хомячка (Chinese hamster ovary, СНО) и клетки миеломы мыши. Уровень продукции МА в настоящее время достаточно высок и может достигать 3 г МА на один литр культуры животных клеток (Zhang et al., 2006. Biotechnol Bioeng. 95, 1188-1197). Однако его высокая стоимость складывается из затрат на оборудование, систему поддержания и стерильности, а также системы сертификации и контроля сред, использующих продукты животного происхождения и увеличивающих риск загрязнения препарата МА прионами и вирусами.
Растения имеют ряд преимуществ по сравнению с клетками бактерий, дрожжей и млекопитающих для продукции чужеродных белков. Растения как «фабрики» (а) не содержат патогенных для человека вирусов и прионов, и (б) не требуют применения дорогостоящей аппаратуры (ферментеры), культуральных сред и системы стерильности. Стоимость выращивания исходных опытных растений несравнимо ниже стоимости культивирования клеток бактерий, дрожжей или животных. Технология «временной» продукции (transient expression) белков в растении позволяет наработать чужеродный белок в количестве, достигающем 10-30% от общего растворимого белка растения в короткое время (5-10 дней), когда целевой ген включается в бинарный вектор с последующей доставкой его в клетки инфицируемого растения методом агроинокуляции или агроинфильтрации (Kapila et al., 1997. Plant Science 122, 101-108). В этих условиях уровень накопления целевого белка определяется эффективностью (а) транскрипции бинарного вектора (Rombauts et al., 2003. Plant Physiol. 132, 1162-1176), (б) репликации (Asurmendi et al., 2004. Proc. Natl. Acad. Sci. USA 101, 1415-1420; Marrillonet et al., 2005. Nature Biotechn. 23, 718-723), (в) защиты транскриптома от деградации (Voinnet et al, 2003. Plant J.33, 949-956), (г) стабильностью целевого белка.
В литературе имеются два сообщения о продукции полного антиракового антитела в растении. В трансгенных растениях табака удалось накопить небольшое количество (0,02% от растворимых белков) антител против рака прямой кишки (Ко et al., 2006, Proc. Natl. Acad. Sci. USA 102, 7026-7030). Использование вирусных векторов и системы транзиентной (временной) экспрессии позволило обеспечить накопление до 3-5% от растворимого белка антираковых антител А5 (Giritch et al., Proc.Natl. Acad. Sci. USA 103, 14701-14706). Однако пока никому не удалось экспрессировать в растении антитело против онкогена HER2/neu.
Раскрытие изобретения
Настоящее изобретение относится к способу продукции в клетке растения антитела, специфически связывающего онкобелок HER2/neu. Способ настоящего изобретения включает в себя:
(а) обеспечение одного или более экспрессионных векторов, направляющих в клетке растения синтез тяжелой, легкой цепи антитела или его антигенсвязывающих фрагментов (dsFv, scFv, scFv-Fc);
(б) введение указанных одного или более векторов в клетку растения;
(в) культивирование клетки растения в условиях, обеспечивающих совместную экспрессию в клетке указанных векторов.
В одном из воплощений клетка растения является изолированной клеткой или находится в культуре клеток растения, культуре ткани растения, культуре органа растения, в целом растении или его части.
В одном из воплощений способ предусматривает использование одного или более экспрессионных векторов, которые обеспечивают синтез неамплифицирующейся РНК. В одном из предпочтительных воплощений указанные один или более экспрессионных векторов обеспечивают синтез неамплифицирующейся РНК под контролем индуцибельного промотора.
В альтернативном воплощении способ предусматривает использование одного или более экспрессионных векторов, которые являются вирусными экспрессионными векторами. В одном из предпочтительных воплощений используемые вирусные экспрессионные векторы являются векторами на основе генома ДНК-содержащего вируса. В еще одном из предпочтительных воплощений указанные вирусные экспрессионные векторы являются векторами на основе генома РНК-содержащего вируса. Наиболее предпочтительно, когда вирусные экспрессионные векторы на основе генома ДНК- или РНК-содержащего вируса обеспечивают синтез вирусной РНК с помощью индуцибельного промотора. Предпочтительно вирусные экспрессионные векторы подбирают таким образом, чтобы исключить конкуренцию друг с другом в одной и той же клетке.
В одном из предпочтительных воплощений экспрессионный вектор обеспечивает синтез РНК вируса табачной мозаики, кодирующей тяжелую цепь антитела. В еще одном из предпочтительных воплощений экспрессионный вектор обеспечивает синтез РНК вируса табачной мозаики, кодирующей легкую цепь антитела.
В следующем предпочтительном воплощении экспрессионный вектор обеспечивает синтез РНК Х-вируса картофеля, кодирующей тяжелую цепь антитела. В еще одном из предпочтительных воплощений экспрессионный вектор обеспечивает синтез РНК Х-вируса картофеля, кодирующей легкую цепь антитела.
В еще одном воплощении экспрессионные векторы обеспечивают синтез легкой и тяжелой цепи антитела в одной и той же клетке.
В следующем воплощении продуцируемое антитело содержит сигнальный пептид, обеспечивающий адресацию антитела в клеточный компартмент или секрецию за пределы клетки.
В одном из воплощений антитело принадлежит к иммуноглобулину класса G или А, или М, или D, или Е.
В еще одном из воплощений растительная клетка является клеткой двудольного или однодольного растения. В предпочтительном воплощении двудольное растение принадлежит к семейству пасленовых. Еще более предпочтительно, когда растением семейства пасленовых является растение рода Nicotiana. Наиболее предпочтительно растением рода Nicotiana является N. tabacum или N. benthamiana.
В еще одном из воплощений двудольным растением является растение семейств Brassicacea, Legume или Chenopodiaceae.
В следующем воплощении экспрессионные векторы вводят в клетку растения путем агроинъекции.
В еще одном из воплощений экспрессионные векторы обеспечивают стабильную трансформацию. В альтернативном воплощении экспрессионные векторы обеспечивают транзиентную трансформацию.
В своем следующем аспекте настоящее изобретение относится к растительной клетке, которая продуцирует антитело, взаимодействующее с онкобелком HER2/neu. В одном из воплощений клетка стабильно трансформирована генетической конструкцией, направляющей синтез антитела. В альтернативном воплощении растительная клетка транзиентно трансформирована генетической конструкцией, направляющей синтез антитела.
Краткое описание фигур
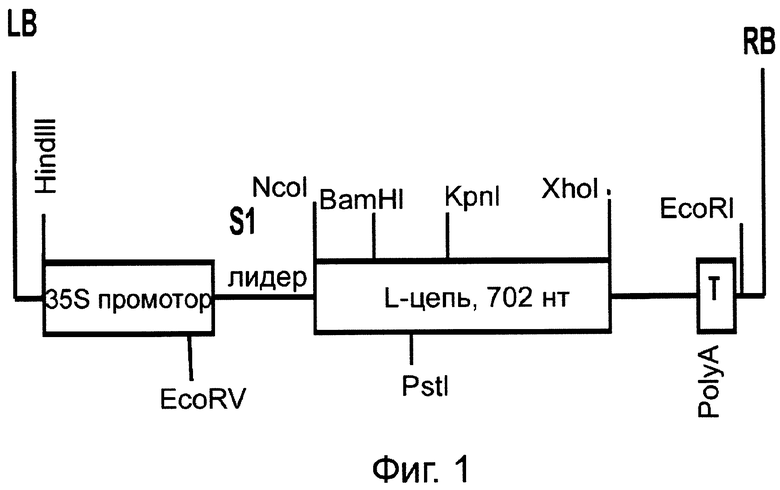
Фиг.1. Схема строения бинарного вектора рА11866, кодирующего легкую (L) цепь HER2/neu-специфического антитела, где RB и LB - соответственно правая и левая часть участка встраивания в геном растения бинарного вектора; 35S - транскрипционный промотор вируса мозаики цветной капусты (ВМЦК); PolyA/T - полиаденилирующий сигнал ВМЦК/терминатор транскрипции геномной РНК ВМЦК. S1-лидер - 5'-нетранслируемая область геномной РНК ВМЦК.
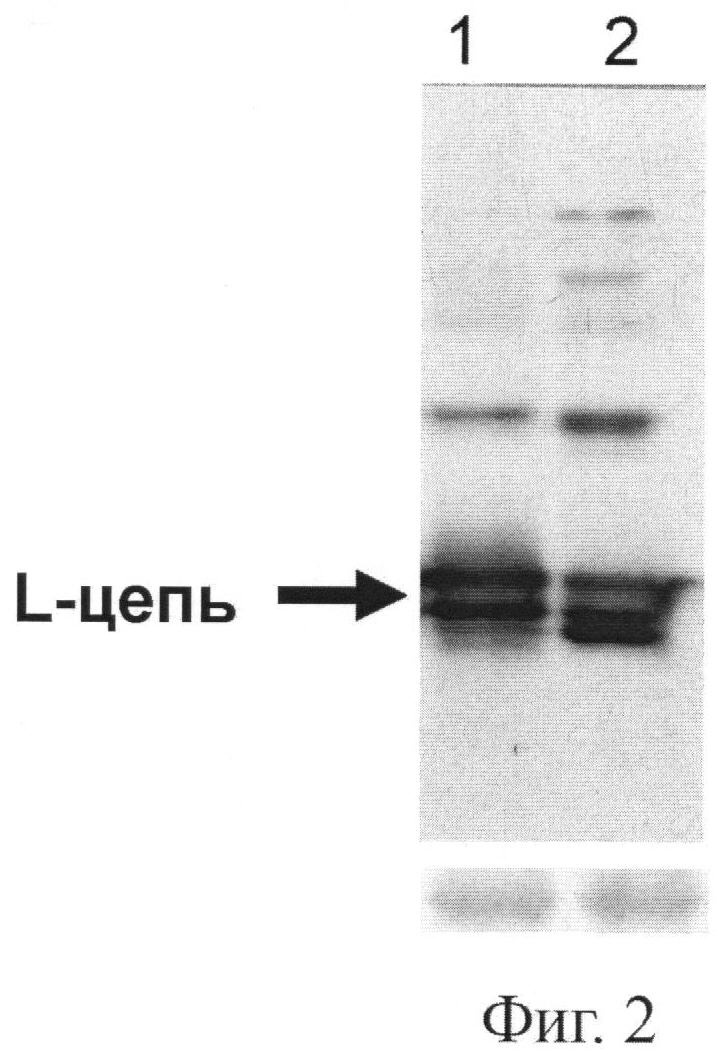
Фиг.2. Накопление легкой цепи (L-цепь) в листьях растений, агроинъецированных вектором рА11866, описанным на Фиг.1. Листья растения N.benthamiana агроинъецировали рА11866 в присутствии бинарного вектора, кодирующего супрессор сайленсинга р19 томбусвируса кустистой карликовости томатов (дорожка 1), или вектора рА11691, направляющего в клетке синтез искусственной тРНК под контролем промотора тРНК (дорожка 2). Белки анализировали в вестерн-блоттинге с помощью антител, специфичных к легкой цепи Her2/neu-специфического антитела человека.
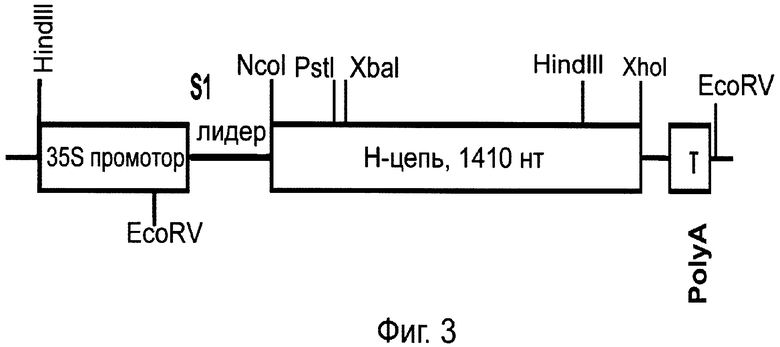
Фиг.3. Схема строения бинарного вектора рА11903, кодирующего тяжелую (Н) цепь Her2/neu-специфического антитела. Условные обозначения те же, что и на Фиг.1.
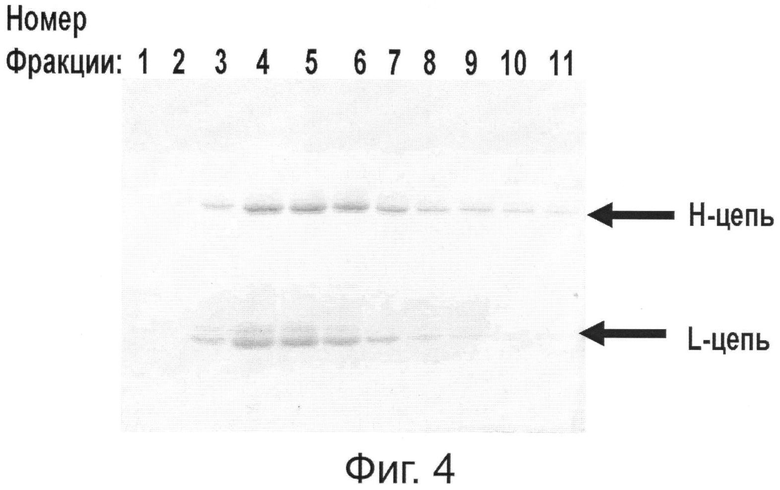
Фиг.4. Накопление Her2/neu-специфических антител в растении, агроинъецированном неамплифицирующимися бинарными векторами рА11866 и рА11903, и их очистка на колонке с протеин G-сефарозой.
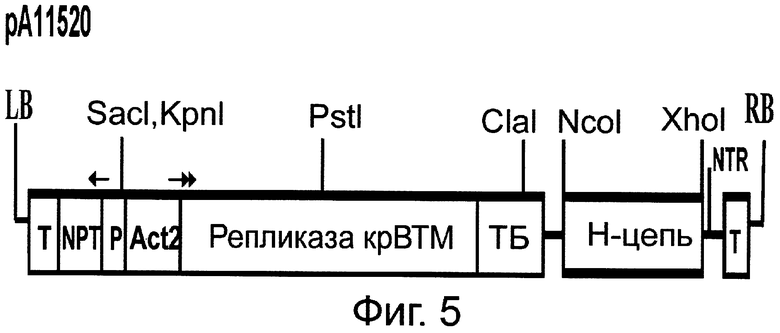
Фиг.5. Схема строения бинарного вектора рА11520, основанного на кДНК крВТМ и кодирующего тяжелую (Н) цепь Her2/neu-специфического антитела, где (слева направо) LB - левая часть участка встраивания в геном растения бинарного вектора; Т - нопалинсинтазный терминатор; NPT - ген неомицинфосфотрансферазы; Р - нопалинсинтазный транскрипционный промотор; Act2 - актиновый транскрипционный промотор 2 из арабидопсиса; ген репликазы крВТМ; ТБ - транспортный белок крВТМ, Н-цепь - тяжелая цепь; NTR - 3'нетранслируемая область геномной РНК крВТМ; Т - нопалинсинтазный терминатор; RB - правая часть участка встраивания в геном растения бинарного вектора. Стрелки показывают направление транскрипции.
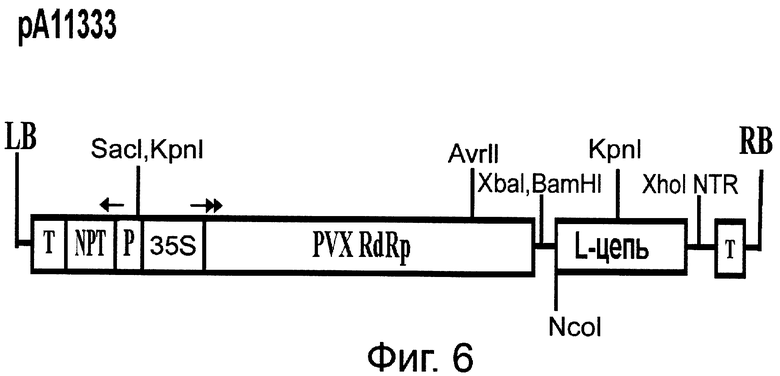
Фиг.6. Схема строения бинарного вектора рА11333, основанного на кДНК ХВК и кодирующего легкую (L) цепь Her2/neu-специфического антитела, где (слева направо) LB - левая часть участка встраивания в геном растения бинарного вектора; Т - нопалинсинтазный терминатор; NPT - ген неомицинфосфотрансферазы; Р - нопалинсинтазный транскрипционный промотор; 35S - транскрипционный промотор ВМЦК; РVХ RdRp-репликаза ХВК, L-цепь - легкая цепь; NTR - 3'нетранслируемая область геномной РНК ХВК; Т - 35S терминатор ВМЦК; RB - правая часть участка встраивания в геном растения бинарного вектора. Стрелки показывают направление транскрипции.
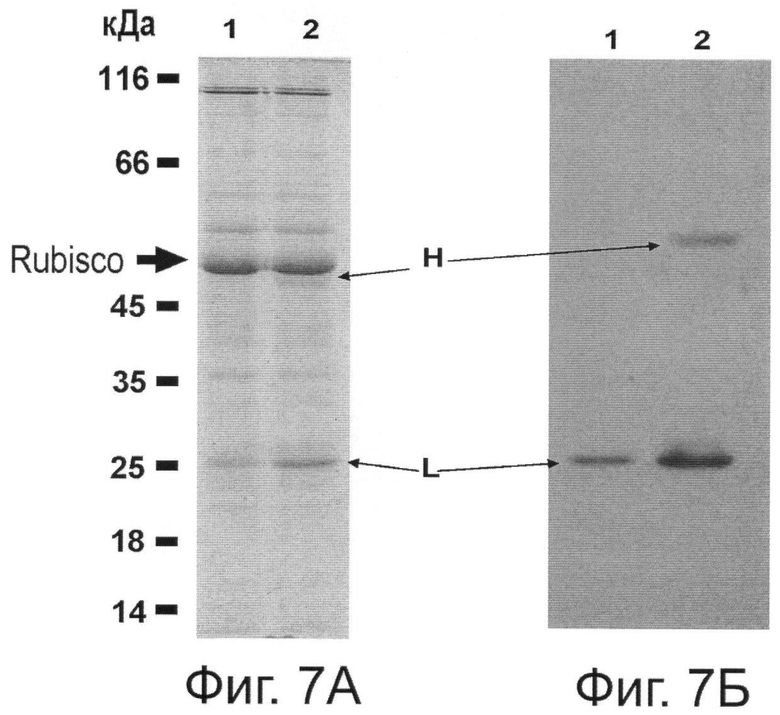
Фиг.7. Накопление Her2/neu-специфических антител в растении, агроинъецированном бинарными векторами рА11520 и рА11333. Фиг.7А. Белковые экстракты растений через 3 (дорожка 1) и 7 (дорожка 2) дней после агроинъекции векторами рА11520 и рА11333 фракционировали электрофорезом, гели окрашивали кумасси. Фиг.7Б. Выявление Her2/neu-специфического антитела Вестерн-блоттингом с антителами к альфа- и каппа-цепям антител человека. Стрелками показано: Rubisco - тяжелая субъединица рибулозобисфосфаткарбоксилазы хлоропластов.
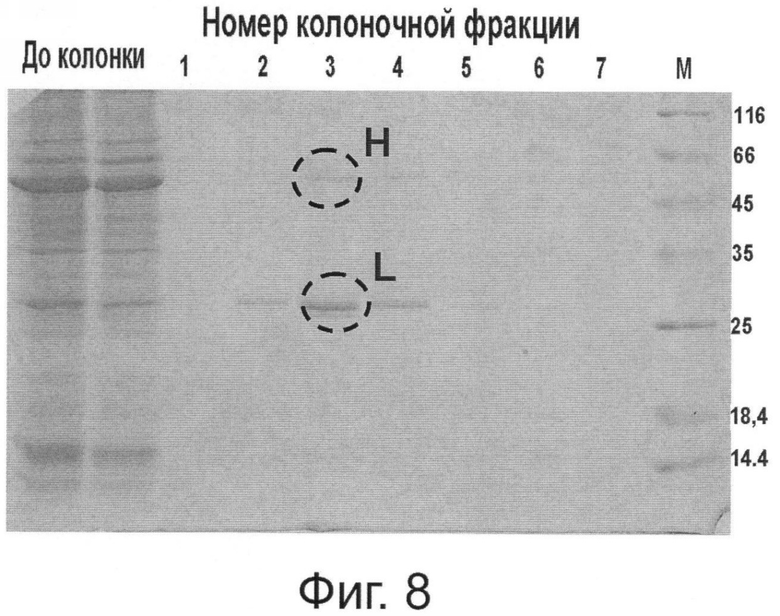
Фиг.8. Накопление Her2/neu-специфических антител в растении, агроинокулированном вирусными бинарными векторами рА11520 и рА11333, и их очистка на колонке с протеин G-сефарозой. Отмечены белковые зоны, соответствующие тяжелой (Н) и легкой цепи (L) антитела.
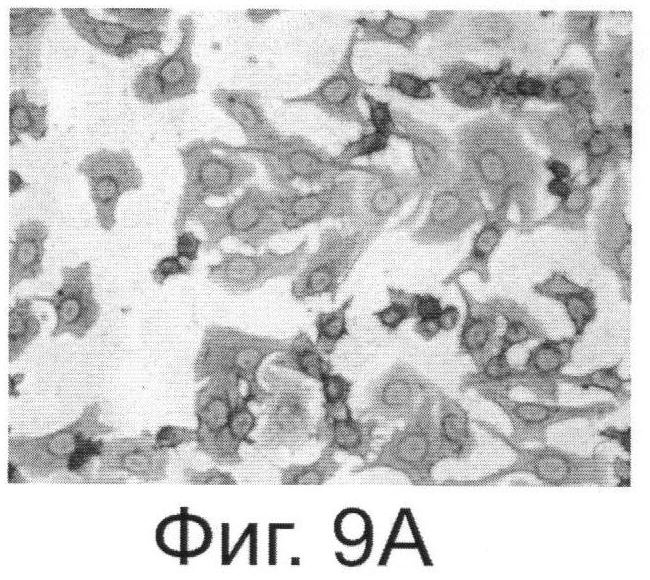
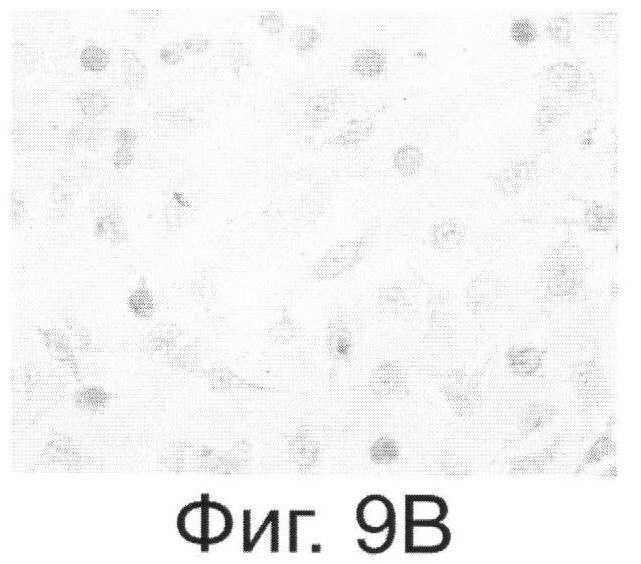
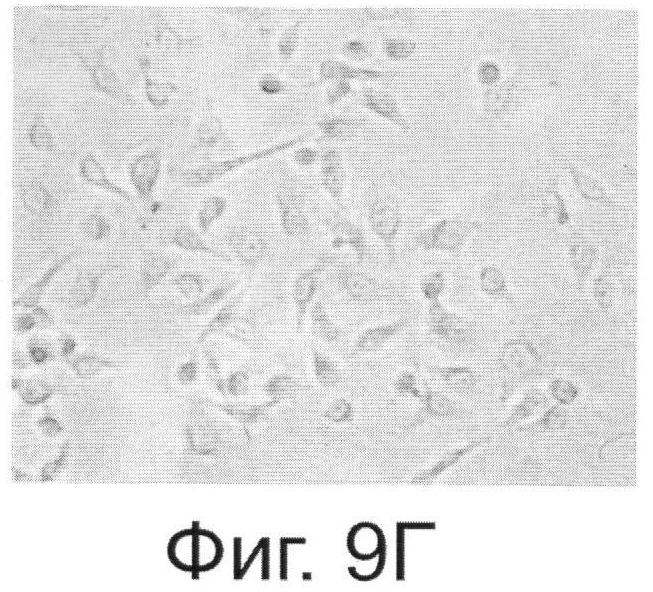
Фиг.9. Антитела, выделенные из растений, реагируют с внеклеточной частью онкогена Her2/neu. Фиг.9А. - Цитологический анализ клеток SKBR-3, экспрессирующих онкоген Her2/neu и обработанных антителами, выделенными из растений. Специфичность связывания тестировали с помощью кроличьих антител, конъюгированных с пероксидазой из хрена и реагирующих с антителами человека. Фиг.9Б. - Клетки SKBR-3 после обработки диагностическими антителами, специфически реагирующими с Her2/neu фирмы Dako (позитивный контроль). Фиг.9В и Фиг.9Г - Негативные контроли, соответственно к А и Б, полученные путем обработки клеток SKBR-3 только кроличьим антителами.
Варианты осуществления изобретения
В основе настоящего изобретения лежит тот обнаруженный автором факт, что гены, кодирующие тяжелую и легкую цепи МА против онкогена HER2/neu, могут быть экспрессированы в клетке растения с получением полностью функционального антитела, специфически связывающегося со «своим» антигеном. Предложенный способ предполагает создание экспрессионных векторов, направляющих в клетке растения синтез тяжелой (Н) и легкой (L) цепи антитела, введение указанных векторов в клетку растения и культивирование клетки растения в условиях, обеспечивающих совместную экспрессию в клетке указанных векторов. Вообще, предложенный способ может быть использован для экспрессии любых антираковых антител, аминокислотная последовательность которых известна и опубликована, например бевацизумаба (авастин) (Kohen et al., 2007. Oncologist 12, 713-718; патент US 7,241,444), панитумумаба (Easley and Kirkpatrick, 2006. Nature Review 5, 987-988; Патентная заявка США №20070179086). В настоящее время известен целый ряд МА против внеклеточного домена онкогена HER2/neu, которые могут быть использованы для лечения рака молочной железы, и кодирующие их гены. В частности, гены, кодирующие антитела против внеклеточного домена онкогена HER2/neu, могут быть выделены из гибридомы ICO85, синтезирующей мышиные МА, направленные против онкогена HER2/neu (см. Пример 1). мРНК, кодирующая легкую и тяжелую цепь антитела, может образовываться как с реплицирующихся векторов, созданных на основе вирусных геномов (см. Пример 3), так и с неамплифицирующихся моно- (см. Пример 2) или полицистронных мРНК с помощью участка внутренней посадки рибосом (Dorokhov et al., 2002. Proc. Natl. Acad. Sci.USA 99, 5301-5306). В контексте настоящего изобретения экспрессионные векторы, кодирующие легкую и тяжелую цепи антитела против онкогена HER2/neu, могут быть созданы на основе бинарного вектора Bin19 (см. Фиг.1 и 3 Примера 2). Эти векторы обеспечивают синтез и накопление в растении антитела в количестве, достаточном для выделения и очистки (см. Фиг.4 Примера 2).
Преимущество настоящего изобретения состоит в том, что оно обеспечивает возможность продуцирования в растении не только полноразмерных антител, но их антигенсвязывающих фрагментов, например dsFv, scFv, scFv-Fc, а также верблюжьих антител, которые содержат только тяжелые цепи (см. обзор Jain et al., 2007. Trends in Biotechnology 25, 307-316).
Одновременная экспрессия цепей антитела в растительной клетке с помощью вирусных векторов возможна при условии отсутствия конкуренции между вирусами. Наши эксперименты показывают, что ВТМ и ХВК не только не конкурируют между собой, но ХВК даже стимулирует репродукцию ВТМ. Накопление тяжелой цепи с ВТМ-вектора увеличивается в присутствии ХВК, кодирующего легкую цепь антитела (см. Пример 3). Анализ механизмов репродукции синдбис-подобных фитовирусов показывает, что конкуренция у них происходит на стадии формирования так называемых вирусных «фабрик», когда вирусная репликаза взаимодействует (прикрепляется) с мембранами эндоплазматического ретикулума. ВТМ и ХВК не конкурируют между собой на стадии формирования репликативного комплекса и вирусной «фабрики». Вирус желтой мозаики турнепса (ВЖМТ), формирующий вирусную «фабрику» на мембранах хлоропластов, также не конкурирует с ВТМ и ХВК и поэтому может быть использован для синтеза в одной клетке цепей антитела. Этот подход может быть использован для подбора других неконкурирующих вирусных пар.
Настоящее изобретение предусматривает использование невирусных векторов для синтеза в растительной клетке антитела. Эти векторы не конкурируют между собой в цитоплазме. Однако их синтез и стабильность в цитоплазме находится под контролем механизма «умолкания» (сайленсинга) генов, что может приводить к снижению уровня продукции целевого антитела. В том случае, когда действительно имеет место снижение продукции целевого белка в результате сайленсинга генов, настоящим изобретением предусмотрено использование антисайленсинговых белков. В качестве примера антисайленсингового белка может быть упомянут, например, белок Р19 вируса кустистой карликовости томатов, ген которого легко синтезировать (см. пример 2). Такой же антисайленсинговый эффект могут оказывать короткие некодирующие РНК, синтезируемые под контролем РНК-полимеразы III (см Фиг.2).
Растительная клетка, экспрессирующая антитело, может быть в виде изолированной клетки, лишенной клеточной стенки (протопласт), или в составе культивируемого эксплантата (ткань, орган) или целого растения. Культивирование клеток в искусственных условиях происходит в стерильной среде известного состава и в контролируемых условиях при постоянной температуре (26-28°С) и освещенности. Настоящее изобретение предполагает возможность использования некодирующих РНК для стимулирования продукции антител в культивируемых клетках как в виде суспензионной культуры, так и растительных эксплантатах, культивируемых на твердой среде, или же в целом растении, выращиваемом в подходящей среде, такой как природный субстрат (например, почва, заменители почвы), или в гидропонной культуре. Антитело может накапливаться внутри растительной клетки или с помощью введения в его состав сигнальной последовательности может быть экспортировано в культуральную среду. Извлечение антитела может происходить из внутриклеточного содержимого путем разрушения клеток культуры или непосредственно из культуральной среды в том случае, когда за счет включения в состав антитела сигнальной последовательности обеспечивается секреция в культуральную среду.
ДНК, кодирующая цепи антитела, может быть введена в растительную клетку известными для специалиста способами, например с помощью электропорации, бомбардирования микроснарядами, микроинъекций и посредством инфицирования вирусом. Кроме того, в растительную клетку ДНК может быть доставлена с помощью Agrobacterium tumefaciens. Для транзиентной экспрессии чужеродного гена в листовой ткани чаще используют метод агроинфильтрации и агроинокуляции (Kapila et al., 1997. Plant Science 122, 101-108). Трансформация растительных клеток может быть стабильной, сопровождающейся интеграцией вводимой ДНК в клеточный геном. Желаемый эффект также может быть достигнут при введении в ядро ДНК и транзиентном (временном) синтезе РНК без стабильной интеграции ДНК в хозяйский геном (см. Примеры 2 и 3).
Важным и решающим преимуществом настоящего изобретения является то, что система синтеза антитела против онкогена HER2/neu в растении может достигать высокого уровня продукции дешевого белка. Наши эксперименты показывают, что из 1 кг листьев N.benthamiana можно выделить до 300-500 мг очищенного антитела (3-5% от растворимого белка). С помощью хроматографии на колонке с протеином G или А можно получить чистый препарат антитела против онкогена Her2/neu (Фиг.8). Этот препарат обладает способностью связывать внеклеточный домен онкогена Her2/neu, как это показано в цитохимическом тесте (Фиг.9). Наши оценки показывают, что стоимость препарата МА, полученного при экспрессии в клетке растения, может быть в 10-20 раз ниже стоимости аналогичного препарата, полученного из животных клеток СНО.
Примеры
Следующие далее примеры раскрывают наиболее предпочтительные воплощения данного изобретения и приведены исключительно с целью лучшего пояснения его сущности. Специалисту в данной области будет понятно, что можно осуществить множество модификаций как в отношении используемых средств и материалов, так и в отношении используемых способов без отступления за рамки изобретения.
Пример 1. Получение генов, кодирующих тяжелую и легкую цепь МА против онкогена Her2/neu.
Из культуры клеток гибридомы ICO85, полученной в лаборатории экспериментальной диагностики и биотерапии опухолей НИИ ЭДиТО ГУ РОНЦ им. Н.Н.Блохина РАМН под руководством проф. А.Ю.Барышникова, генно-инженерными методами были выделены последовательности, кодирующие вариабельные участки антитела, афинные к онкогену Her2/neu. Полученные фрагменты были встроены в конструкции, содержащие последовательности, кодирующие конститутивные участки антител класса G.
Пример 2. Экспрессия легкой и тяжелой цепи антитела против онкогена Her2/neu в растении с помощью невирусного вектора.
Отдельные гены легкой и тяжелой цепи антитела против онкогена Her2/neu были клонированы в субклон pFF19, содержащий 35S-промотор по сайтам NcoI и XhoI. На следующем этапе гены по отдельности клонировали в вектор Bin 19 (Bevan, 1984. Nucleic Acids Res. 12, 8711-8721) по сайтам HindIII-EcoRI с получением векторов соответственно рА11860 и рА11903 (Фиг.1 и 3). Agrobacterium tumefaciens с бинарными векторами, экспрессирующими тяжелую или легкую цепь антитела против онкогена Her2/neu, засевали в жидкую среду 2-YT и выращивали ночь при температуре 28°С. Ночную культуру осаждали центрифугированием при 5 тыс. об/мин в течение 3 мин. Затем осадок ресуспендировали в буфере для агроинфильтрации, содержащем 10 мМ MgCl2 и 10 мМ MES рН 5,0, и вводили в лист в соответствующем разведении. Фиг.2 показывает накопление легкой цепи (L-цепь) антитела против онкогена Her2/neu в листьях N. benthamiana через 3 дня после агроинъекции бинарным вектором рА11866. В этих экспериментах использовали конструкцию, кодирующую антисайленсинговый белок Р19 вируса кустистой карликовости томатов, ген которого синтезирован нами ранее (Dorokhov et al., 2006. FEBS Letters 580, 3872-3878).
При одновременной ко-инъекции векторов рА11903 и рА11860 в листья бентамианы в них накапливаются легкая и тяжелая цепи антитела, которые можно выделить и очистить аффинной хроматографией на колонке с протеин G-сефарозой (Фиг.4).
Пример 3. Экспрессия легкой и тяжелой цепи антитела против онкогена Her2/neu в растении с помощью вирусных векторов.
Для создания вектора на основе генома ВТМ (Фиг.5) использована кДНК крВТМ (Dorokhov et al. 1994, FEBS Lett. 350, 5-8), в котором ген репликазы был из близкородственного штамма, вируса просветления жилок турнепса (Lartey et al., 1994, Arch. Virol., 138, 287-298). В качестве эукариотического транскрипционного промотора послужил промотор актина 2 из Arabidopsis thaliana (EMBL AF308778, An et al., 1996, Plant J., 10, 107-121). Для создания вектора на основе генома ХВК (Фиг.6) использована кДНК, описанная ранее (Комарова и др., 2006, Биохимия 71, 1043-1049). Agrobacterium tumefaciens с бинарными векторами рА11520 и рА11333, экспрессирующими соответственно тяжелую и легкую цепь антитела против онкогена Her2/neu, засевали в жидкую среду 2-YT и выращивали ночь при температуре 28°С. Ночную культуру осаждали центрифугированием при 5 тыс. об/мин в течение 3 мин. Затем осадок ресуспендировали в буфере для агроинфильтрации, содержащем 10 мМ MgCl2 и 10 мМ MES рН 5,0, и вводили в лист в соответствующем разведении. Фиг.7 показывает накопление антитела против онкогена Her2/neu в листьях N. benthamiana через 4 дня после агроинъекции смесью бинарных векторов. Можно видеть (Фиг.7) существенное накопление легкой и тяжелой цепей антитела, выявляемое в геле с помощью окрашивания кумасси (Фиг.7А) и вестерн-анализом (Фиг.7Б). С помощью хроматографии на колонке с протеин G-сефарозой можно получить чистый препарат антитела против онкогена Her2/neu (Фиг.8). Наши эксперименты показывают, что из 1 кг листьев N. benthamiana можно выделить до 300-500 мг очищенного антитела.
Пример 4. Синтезированное в растительной клетке антитело способно связываться с онкогеном Her2/neu на поверхности раковых клеток SKBR-3.
Для доказательства активности синтезированного в растительной клетке антитела использовали иммуноцитохимический (ИЦХ) тест. Этот тест наряду с радиолигандным и иммуноферментным методом широко используется в клинической практике. Иммуноцитохимический анализ не требует больших временных затрат, выполняется быстро в течение 2-3 часов и широко описан в литературе (см., например, Глузман Д.Ф., Скляренко Л.М., Надгорная В.А., Крячок И.А. Диагностическая иммуноцитохимия опухолей. Киев, Морион; 2003, с.28-31). Мы использовали наше антитело в ИЦХ раковых клеток SKBR-3, имеющих на своей поверхности онкоген Her2/neu. Из Фиг.9 видно, что антитело обладает способностью связывать внеклеточный домен онкогена Her2/neu. Таким образом, антитело, полученное способом согласно настоящему изобретению, является полностью функциональным и способно специфически узнавать и связываться с онкогеном Her2/neu. Это открывает широкие возможности для применения способа настоящего изобретения для широкомасштабного производства противораковых антител, которые могут быть использованы в терапевтических и диагностических целях. Себестоимость диагностического препарата антител, полученных в растении, в 12 раз ниже имеющихся в продаже антител фирмы Dako.
Все патенты, публикации, научные статьи и другие документы и материалы, цитируемые или упоминаемые здесь, включены в настоящее описание путем отсылки в такой степени, как если бы каждый из этих документов был включен путем отсылки индивидуально или приведен здесь в его полном виде.
Конкретные гены, векторы, виды растений, применения, используемые материалы и методы, описанные здесь, относятся к предпочтительным вариантам осуществления, приведены в качестве примеров и не предназначены для ограничения объема данного изобретения. При изучении этого описания другие объекты, аспекты и воплощения будут приходить в голову специалистам в данной области, и они охватываются рамками данного изобретения. Специалистам в данной области будет понятно, что различные замены и модификации могут быть произведены в отношении описанного здесь изобретения без отклонения от его объема и рамок, которые определяются прилагаемой формулой изобретения.
Изобретение относится к биотехнологии. Описан способ продукции в клетке растения антитела, специфически связывающего онкобелок HER2/neu, который включает в себя: (а) обеспечение одного или более экспрессионных векторов, направляющих в клетке растения синтез тяжелой, легкой цепи антитела или его антигенсвязывающих фрагментов (dsFv, scFv, scFv-Fc); (б) введение указанных одного или более векторов в клетку растения; (в) культивирование клетки растения в условиях, обеспечивающих совместную экспрессию в клетке указанных векторов. Раскрыта растительная клетка, продуцирующая антитело, взаимодействующее с онкобелком HER2/neu, трансформированная одним или более экспрессионных векторов, направляющих в клетке растения синтез тяжелой, легкой цепи антитела или его антигенсвязывающих фрагментов (dsFv, scFv, scFv-Fc). Изобретение может быть использовано для диагностики и лечения рака молочной железы. 2 н. и 25 з.п. ф-лы, 9 ил.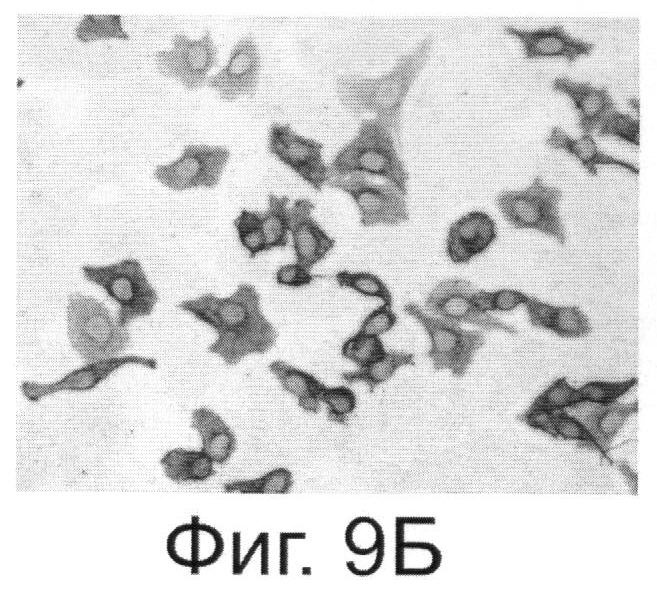
1. Способ продукции в клетке растения антитела, специфически связывающего онкобелок HER2/neu, который включает в себя
(а) обеспечение одного или более экспрессионных векторов, направляющих в клетке растения синтез тяжелой, легкой цепи антитела или его антигенсвязывающих фрагментов (dsFv, scFv, scFv-Fc);
(б) введение указанных одного или более векторов в клетку растения;
(в) культивирование клетки растения в условиях, обеспечивающих совместную экспрессию в клетке указанных векторов.
2. Способ по п.1, в котором клетка растения является изолированной клеткой или находится в составе культивируемого эксплантата (ткани, органа) или в составе целого растения.
3. Способ по п.1, характеризующийся тем, что указанный один или более экспрессионных векторов обеспечивают синтез неамплифицирующейся РНК.
4. Способ по п.3, характеризующийся тем, что указанный один или более экспрессионных векторов обеспечивают синтез неамплифицирующейся РНК с помощью индуцибельного промотора.
5. Способ по п.1, характеризующийся тем, что указанный один или более экспрессионных векторов являются вирусными экспрессионными векторами.
6. Способ по п.5, характеризующийся тем, что указанные вирусные экспрессионные векторы являются векторами на основе генома ДНК-содержащего вируса.
7. Способ по п.5, характеризующийся тем, что указанные вирусные экспрессионные векторы являются векторами на основе генома РНК-содержащего вируса.
8. Способ по любому из пп.5-7, характеризующийся тем, что указанные вирусные экспрессионные векторы обеспечивают синтез вирусной РНК с помощью индуцибельного промотора.
9. Способ по п.7, характеризующийся тем, что экспрессионный вектор обеспечивает синтез РНК вируса табачной мозаики, кодирующей тяжелую цепь антитела.
10. Способ по п.7, характеризующийся тем, что экспрессионный вектор обеспечивает синтез РНК вируса табачной мозаики, кодирующей легкую цепь антитела.
11. Способ по п.7, характеризующийся тем, что экспрессионный вектор обеспечивает синтез РНК Х-вируса картофеля, кодирующей тяжелую цепь антитела.
12. Способ по п.7, характеризующийся тем, что экспрессионный вектор обеспечивает синтез РНК Х-вируса картофеля, кодирующей легкую цепь антитела.
13. Способ по п.1, характеризующийся тем, что экспрессионные векторы обеспечивают синтез легкой и тяжелой цепи антитела в одной и той же клетке.
14. Способ по п.5, характеризующийся тем, что используют не конкурирующие друг с другом в одной и той же клетке вирусные векторы.
15. Способ по п.1, характеризующийся тем, что антитело содержит сигнальный пептид, обеспечивающий адресацию антитела в клеточный компартмент или секрецию за пределы клетки.
16. Способ по п.1, в котором антитело принадлежит к иммуноглобулину класса G или А, или М, или D, или Е.
17. Способ по п.1, характеризующийся тем, что растительная клетка является клеткой двудольного или однодольного растения.
18. Способ по п.17, характеризующийся тем, что двудольное растение принадлежит к семейству пасленовых.
19. Способ по п.18, характеризующийся тем, что растением семейства пасленовых является растение рода Nicotiana.
20. Способ по п.19, характеризующийся тем, что растением рода Nicotiana является N. tabacum или N. benthamiana.
21. Способ по п.17, характеризующийся тем, что двудольным растением является растение семейств Brassicacea, Legume, или Chenopodiaceae.
22. Способ по п.1, характеризующийся тем, что указанные векторы вводят в клетку растения путем агроинъекции.
23. Способ по п.1, характеризующийся тем, что в клетку растения вводят экспрессионные векторы, обеспечивающие стабильную трансформацию.
24. Способ по п.1, характеризующийся тем, что в клетку растения вводят экспрессионные векторы, обеспечивающие транзиентную трансформацию.
25. Растительная клетка, продуцирующая антитело, взаимодействующее с онкобелком HER2/neu, трансформированная одним или более экспрессионных векторов, направляющих в клетке растения синтез тяжелой, легкой цепи антитела или его антигенсвязывающих фрагментов (dsFv, scFv, scFv-Fc).
26. Растительная клетка по п.25, стабильно-трансформированная одним или более экспрессионных векторов, направляющих синтез антитела.
27. Растительная клетка по п.25, транзиентно-трансформированная одним или более экспрессионных векторов, направляющих синтез антитела.
| ANATOLI GIRITCH et al | |||
| Rapid high-yield expression of full-size IgG antibodies in plants coinfected with noncompeting viral vectors | |||
| Proc | |||
| Natl ACAD SCI USA, 103(40):14701-6, 2006 | |||
| MA J.K., LEHNER T., STABILA P., FUX C.I., HIАТТ A.: Assembly of monoclonal antibodies with IgGI and IgA heavy chain domains in transgenic tobacco plants | |||
| Eur | |||
| J. |

