ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам получения липидов. В частности, настоящее изобретение относится к способам увеличения уровня одного или нескольких неполярных липидов и/или общего содержания неполярных липидов в трансгенном организме или его части. В одном определенном варианте осуществления настоящее изобретение относится к применению ацилтрансферазы, например, моноацилглицерол-ацилтрансферазы (MGAT), для увеличения уровня одного или нескольких неполярных липидов и/или общего содержания неполярных липидов в растениях, семени и/или листьях растения, водорослях и грибах.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Растительные липиды, такие как триацилглицеролы масла семян (TAG) находят многочисленные применения, например, применения в кулинарии (шортенинг, консистенция, вкус), промышленные применения (в мылах, свечах, духах, косметических средствах, подходящие в качестве влагопоглотителей, изолирующих средств, смазочных средств) и обеспечивают питательную ценность. Имеет место также растущий интерес в применении растительных липидов для получения биотоплива.
Биотопливо
Растущая потребность в альтернативных источниках энергии может быть удовлетворена по меньшей мере частично с помощью возобновляемых источников снабжения полученным из растений биотопливом. Чтобы являться обоснованной альтернативой ископаемым видам топлива, биотопливо должно обеспечивать прирост объема производства полезной энергии, иметь преимущества применения для окружающей среды, быть экономически конкурентным и производимым в больших количествах без снижения продовольственных ресурсов, имеющегося непредусмотренного побочного продукта существующего получения биотоплива.
Растения представляют собой значительный источник липидов, поскольку многие виды накапливают липиды в качестве основных запасных компонентов в семенах. Основной формой вегетативных запасных липидов в семенах, которые представляют собой, в зависимости от вида, 15-50% веса семени, является триацилглицерол (TAG). Однако, первичным субстратом для синтеза липидов являются углеводы, образованные в зеленых фотосинтетических тканях (листьях и стеблях), которые впоследствии метаболизируются в хлоропластах для получения элементарных звеньев свободных жирных кислот и ацетил-коэнзима A (ацетил-СоА), основных строительных элементов для TAG. Следовательно, листья растений являются основным местом синтеза строительных элементов для TAG. Количество TAG, накопленное в семенах масличных культур, может быть отчасти определено с помощью количества жирной кислоты, образованной в пластидах (Bao and Ohlrogge, 1999). Конечное запасание TAG происходит в семенах в маленьких сферических органоидах, имеющих название масляные тельца. Только около 0,2-0,3% биомассы листьев представляют собой TAG.
Растения с высоким содержанием биомассы, особенно широколистные растения с высоким содержанием биомассы, имеют большой биотопливный потенциал. Растения, которые могут давать на выходе 100-400 тонн на акр материалов с низкой стоимостью с высоким показателем биомассы являются особенно применимыми, особенно, если отсутствуют большие расходы, потребности в рабочей силе, химических средствах или географические ограничения, связанные с получением растений с низкой биомассой.
Моноацилглицерол-ацилтрансферазы
Фермент моноацилглицерол-ацилтрансфераза (MGAT) ассоциирован с млекопитающими, в основном с кишечником млекопитающих, где он катализирует синтез диацилглицерола (DAG) непосредственно из моноацилглицерола (MAG) и ацил-coA жирной кислоты. В отличие от этого, первичный путь синтеза TAG, обнаруженный в растениях, представляет собой путь Кеннеди или глицеролфосфатный путь (фигура 1), который не включает этап MGAT. В пути Кеннеди DAG образуется из ацилированного скелета глицерола в двухэтапной реакции, состоящей из начального ацилирования с помощью ацилтрансферазы лизофосфатидной кислоты (LPAAT), которая добавляет ацил-coA жирной кислоты к субстрату лизофосфатидной кислоты (LysoPA; LPA), и последующего удаления фосфатной группы из продукта, фосфатидной кислоты (РА), с выходом неорганического фосфата (Pi) и DAG. В отличие от этого, MGAT катализирует образование DAG непосредственно путем ацилирования MAG с помощью ацильной группы, происходящей от ацил-coA жирной кислоты. После синтеза DAG другой фермент, диацилглицерол-ацилтрансфераза (DGAT), ацилирует DAG для образования TAG.
Первый ген MGAT был выделен у мыши (MGAT1), и этот ген кодировал мембраносвязанный нерастворимый фермент (Yen et al., 2002). Другие сходные гены MGAT были охарактеризованы у животных, включая второй ген MGAT у мыши (MGAT2) и три гена человека, но не для одного из генов, кодирующих MGAT, не было подтверждено, что они были клонированы из растений (Cao et al., 2003; Cheng et al., 2003).
Диацилглицерол-ацилтрансферазы
DGAT представляет собой интегральный мембранный белок, который катализирует конечный ферментативный этап в образовании TAG у растений, грибов и млекопитающих. Этот фермент отвечает за перенос ацильной группы с ацил-коэнзима А (ацил-СоА) на DAG для образования TAG. DGAT ассоциирована с мембраной и фракциями липидных телец у растений и грибов, в частности, в семенах масличных культур, где она вносит вклад в запасание углерода, используемого в качестве энергетических резервов. Известно, что DGAT контролирует структуру TAG и направляет синтез TAG. Кроме того, известно, что реакция DGAT является специфической для синтеза липидов. Сверхэкспрессия ацил-СоА-зависимой DGAT специфическим для семян образом приводит к увеличению депонирования масла семян и среднего веса семян у растений дикого типа (Jako et al., 2001).
Для того чтобы сделать выходы для коммерческого получения липидов максимальными, существует необходимость в дополнительных средствах для увеличения уровней липидов, в частности, неполярных липидов, таких как DAG и TAG, в трансгенных организмах или их частях, таких как растения, семена, листья, водоросли и грибы.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретатели настоящего изобретения неожиданно обнаружили, что трансгенная экспрессия гена MGAT или родственного гена приводит к значительным увеличениям выхода липидов в клетках, таких как растительные клетки. Изобретатели настоящего изобретения также определили новый путь синтеза DAG и TAG в трансгенных организмах, таких как растения, который отличается от хорошо известного пути Кеннеди.
Соответственно, настоящее изобретение обеспечивает способ получения экстрагированного липида, причем способ включает этапы:
i) получения трансгенного, не являющегося человеком организма или его части, содержащей один или несколько экзогенных полинуклеотидов, где трансгенный, не являющийся человеком организм или его часть имеет повышенный уровень одного или нескольких неполярных липидов по сравнению с соответствующим организмом или его частью, где отсутствует один или несколько экзогенных полинуклеотидов, и
ii) экстрагирования липида из трансгенного, не являющегося человеком организма или его части, таким образом получая экстрагированный липид.
В одном варианте осуществления общее содержание неполярных липидов трансгенного, не являющегося человеком организма или его части увеличивается по сравнению с соответствующим организмом или его частью.
Трансгенный, не являющийся человеком организм или его часть дополнительно можно охарактеризовать с помощью признаков (i), (ii), (iii), отдельно или в комбинации: признак (i) количественно определяет степень повышенного уровня одного или нескольких неполярных липидов или общего содержания неполярных липидов, которую можно выразить как степень увеличения на основе веса или как относительное увеличение по сравнению с уровнем в соответствующем не являющемся человеком организме или его части, и/или признак (ii) устанавливает род или вид растений, или вид грибов или водорослей, или другой тип клеток, и признак (iii) устанавливает один или несколько специфических липидов, которые увеличиваются.
Для признака (i) в варианте осуществления степень увеличения одного или нескольких неполярных липидов составляет по меньшей мере на 0,5%, по меньшей мере на 1%, по меньшей мере на 2%, по меньшей мере на 3%, по меньшей мере на 4%, по меньшей мере на 5%, по меньшей мере на 6%, по меньшей мере на 7%, по меньшей мере на 8%, по меньшей мере на 9%, по меньшей мере на 10%, по меньшей мере на 11%, по меньшей мере на 12%, по меньшей мере на 13%, по меньшей мере на 14%, по меньшей мере на 15%, по меньшей мере на 16%, по меньшей мере на 17%, по меньшей мере на 18%, по меньшей мере на 19%, по меньшей мере на 20%, по меньшей мере на 21%, по меньшей мере на 22%, по меньшей мере на 23% или по меньшей мере на 24% (вес/вес.) больше на основе веса, чем у соответствующего не являющегося человеком организма или его части, предпочтительно до максимального увеличения на около 25% (вес/вес.) на основе веса.
Также для признака (i) в предпочтительном варианте осуществления общее содержание неполярных липидов трансгенного, не являющегося человеком организма или его части увеличивается по сравнению с соответствующим организмом или его частью. В варианте осуществления общее содержание липидов увеличивается по меньшей мере на 0,5%, по меньшей мере на 1%, по меньшей мере на 2%, по меньшей мере на 3%, по меньшей мере на 4%, по меньшей мере на 5%, по меньшей мере на 6%, по меньшей мере на 7%, по меньшей мере на 8%, по меньшей мере на 9%, по меньшей мере на 10%, по меньшей мере на 11%, по меньшей мере на 12%, по меньшей мере на 13%, по меньшей мере на 14%, по меньшей мере на 15%, по меньшей мере на 16%, по меньшей мере на 17%, по меньшей мере на 18%, по меньшей мере на 19%, по меньшей мере на 20%, по меньшей мере на 21%, по меньшей мере на 22%, по меньшей мере на 23% или по меньшей мере на 24% (вес/вес.) больше на основе веса, чем у соответствующего не являющегося человеком организма или его части, предпочтительно до максимального увеличения на около 25% (вес/вес.) на основе веса.
Дополнительно, для признака (i) в варианте осуществления уровень одного или нескольких неполярных липидов и/или общее содержание неполярных липидов составляет по меньшей мере на 1%, по меньшей мере на 2%, по меньшей мере на 3%, по меньшей мере на 4%, по меньшей мере на 5%, по меньшей мере на 6%, по меньшей мере на 7%, по меньшей мере на 8%, по меньшей мере на 9%, по меньшей мере на 10%, по меньшей мере на 11%, по меньшей мере на 12%, по меньшей мере на 13%, по меньшей мере на 14%, по меньшей мере на 15%, по меньшей мере на 16%, по меньшей мере на 17%, по меньшей мере на 18%), по меньшей мере на 19%), по меньшей мере на 20%, по меньшей мере на 21%, по меньшей мере на 22%, по меньшей мере на 23%, по меньшей мере на 24%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 60%>, по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 90%>больше на относительной основе, чем у соответствующего не являющегося человеком организма или его части.
Также для признака (i) степень увеличения в уровне одного или нескольких неполярных липидов и/или общего содержания неполярных липидов может быть по меньшей мере в 2 раза, по меньшей мере в 3 раза, по меньшей мере в 4 раза, по меньшей мере в 5 раз, по меньшей мере в 6 раз, по меньшей мере в 7 раз, по меньшей мере в 8 раз, по меньшей мере в 9 раз или по меньшей мере в 10 раз, предпочтительно до максимума около в 12 раз больше на относительной основе, чем у соответствующего не являющегося человеком организма или его части.
Для признака (ii) в варианте осуществления трансгенный, не являющийся человеком организм представляет собой растение, водоросль или организм, подходящий для ферментации, такой как дрожжи или другой гриб, предпочтительно жировые дрожжи или другой гриб. Растение может быть, например, Brassica sp., Gossypium hirsutum, Linum usitatissimum, Helianthus sp., Carthamus tinctorius, Glycine max, Zea mays, Arabidopsis thaliana, Sorghum bicolor, Sorghum vulgare, Arena sativa, Trifolium sp., Elaesis guineenis, Nicotiana benthamiana, Hordeum vulgare, Lupinus angustifolius, Oryza sativa, Oryza glaberrima, Camelina sativa, Miscanthus x giganteus или Miscanthus sinensis.
Для признака (iii) увеличивается(ются) TAG, DAG, TAG и DAG, MAG, полиненасыщенная жирная кислота (PUFA) или специфическая PUFA (такая как эйкозадиеновая кислота (EDA), арахидоновая кислота (ARA), альфа-линоленовая кислота (ALA), стеаридоновая кислота (SDA), эйкозатриеновая кислота (ЕТЕ), эйкозатетраеновая кислота (ЕТА), эйкозапентаеновая кислота (ЕРА), докозапентаеновая кислота (DPA), докозагексаеновая кислота (DHA)) или комбинация двух или более из них. Степень увеличения TAG, DAG, TAG и DAG, MAG, PUFA или специфической PUFA может быть такой, как определено в признаке (i) выше. В предпочтительном варианте осуществления MAG представляет собой 2-MAG. Предпочтительно, DAG и/или TAG, более предпочтительно общие содержания DAG и TAG увеличиваются.
В предпочтительных вариантах осуществления один или несколько неполярных липидов и/или общее содержание неполярных липидов определяется с помощью комбинации признаков (i), (ii) и (iii), или признаков (i) и (ii), или признаков (i) и (iii).
В одном варианте осуществления часть представляет собой семя, плод, клубень, корень или вегетативную часть растения. Вегетативная часть растения может быть надземной частью растения или зеленой частью, такой как лист или стебель. В другом варианте осуществления часть представляет собой клетку многоклеточного организма. Степень увеличения уровня одного или нескольких неполярных липидов и/или общего содержания неполярных липидов конкретной части растения в данном варианте осуществления может быть такой, как определено в признаке (i) выше. Часть растения для Brassica sp., Gossypium hirsutum, Linum usitatissimum, Helianthus sp., Carthamus tinctorius, Oryza sativa, Oryza glaberrima, Camelina saliva, Glycine max или Zea mays представляет собой предпочтительно семя, тогда как предпочтительная часть для Sorghum bicolor, Sorghum vulgare, Avena sativa, Trifolium sp., Elaesis guineenis, Nicotiana benthamiana, Hordeum vulgare, Lupinus angustifolius, Miscanthus x giganteus или Miscanthus sinensis представляет собой вегетативную часть, в частности, листья и стебли.
В одном варианте осуществления часть представляет собой семя растения, и экстрагированный липид представляет собой масло семян. Способ по настоящему изобретению дополнительно может включать сбор семян из трансгенного растения, отжим масла семян из семян и/или очищение масла семян в один или несколько этапов. Семя может быть, например, из растения канолы, растения кукурузы, растения сои, растения люпина, растения арахиса, растения подсолнечника, растения хлопка, растения сафлора или растения льна.
В одном варианте осуществления общее содержание масла или общее содержание жирных кислот семени составляет по меньшей мере на 0,5% (вес./вес.) - 25% (вес./вес.) больше на основе веса, чем у соответствующего семени, где отсутствует один или несколько экзогенных полинуклеотидов.
В одном варианте осуществления относительное содержание DAG масла семян составляет по меньшей мере на 10%, по меньшей мере на 10,5%, по меньшей мере на 11%, по меньшей мере на 11,5%, по меньшей мере на 12%, по меньшей мере на 12,5%, по меньшей мере на 13%, по меньшей мере на 13,5%, по меньшей мере на 14%, по меньшей мере на 14,5%, по меньшей мере на 15%, по меньшей мере на 15,5%, по меньшей мере на 16%, по меньшей мере на 16,5%, по меньшей мере на 17%, по меньшей мере на 17,5%, по меньшей мере на 18%, по меньшей мере на 18,5%, по меньшей мере на 19%, по меньшей мере на 19,5%, по меньшей мере на 20% (вес./вес.) больше, чем у соответствующего семени. В варианте осуществления содержание DAG семени увеличивается на количество, как определено в признаке (i), и семя принадлежит к роду и/или виду, как определено в признаке (ii).
В одном варианте осуществления относительное содержание TAG масла семян составляет по меньшей мере на 5%, по меньшей мере на 5,5%, по меньшей мере на 6%, по меньшей мере на 6,5%, по меньшей мере на 7%, по меньшей мере на 7,5%, по меньшей мере на 8%, по меньшей мере на 8,5%, по меньшей мере на 9%, по меньшей мере на 9,5% или по меньшей мере на 10% (вес./вес.) больше, чем у соответствующего семени. В варианте осуществления содержание TAG семени увеличивается на количество, как определено в признаке (i), и семя принадлежит к роду и/или виду, как определено в признаке (ii). В одном варианте осуществления семя представляет собой семя канолы, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 45%, по меньшей мере 46%, по меньшей мере 47%, по меньшей мере 48%, по меньшей мере 49%), по меньшей мере 50%, по меньшей мере 51%, по меньшей мере 52%, по меньшей мере 53%, по меньшей мере 54% или по меньшей мере 55%.
В одном варианте осуществления семя представляет собой семя кукурузы, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 5%, по меньшей мере 6%, по меньшей мере 7%, по меньшей мере 8%, по меньшей мере 9% или по меньшей мере 10%.
В одном варианте осуществления семя представляет собой семя сои, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 20%, по меньшей мере 21%, по меньшей мере 22%, по меньшей мере 23%, по меньшей мере 24%, по меньшей мере 25%, по меньшей мере 26%, по меньшей мере 27%, по меньшей мере 28%, по меньшей мере 29% или по меньшей мере 30%.
В одном варианте осуществления семя представляет собой семя люпина, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 10%, по меньшей мере 11%, по меньшей мере 12%, по меньшей мере 13%, по меньшей мере 14%, по меньшей мере 15% или по меньшей мере 16%.
В одном варианте осуществления семя представляет собой семя арахиса, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 50%, по меньшей мере 51%, по меньшей мере 52%, по меньшей мере 53%, по меньшей мере 54% или по меньшей мере 55%.
В одном варианте осуществления семя представляет собой семя подсолнечника, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 50%, по меньшей мере 51%, по меньшей мере 52%, по меньшей мере 53%, по меньшей мере 54% или по меньшей мере 55%.
В одном варианте осуществления семя представляет собой семя хлопка, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 41%, по меньшей мере 42%, по меньшей мере 43%, по меньшей мере 44%, по меньшей мере 45%, по меньшей мере 46%, по меньшей мере 47%, по меньшей мере 48%, по меньшей мере 49% или по меньшей мере 50%.
В одном варианте осуществления семя представляет собой семя сафлора, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 35%, по меньшей мере 36%, по меньшей мере 37%, по меньшей мере 38%, по меньшей мере 39%, по меньшей мере 40%, по меньшей мере 41%, по меньшей мере 42%, по меньшей мере 43%, по меньшей мере 44% или по меньшей мере 45%.
В одном варианте осуществления семя представляет собой семя льна, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 36%, по меньшей мере 37%, по меньшей мере 38%, по меньшей мере 39% или по меньшей мере 40%.
В одном варианте осуществления семя представляет собой семя Camelina saliva, имеющее содержание масла на основе веса (вес./вес.) по меньшей мере 36%, по меньшей мере 37%, по меньшей мере 38%, по меньшей мере 39%, по меньшей мере 40%, по меньшей мере 41%, по меньшей мере 42%, по меньшей мере 43%, по меньшей мере 44% или по меньшей мере 45%.
В другом варианте осуществления часть организма представляет собой вегетативную часть растения, и содержание TAG, DAG, TAG и DAG или MAG вегетативной части растения составляет по меньшей мере на 10%, по меньшей мере на 11%, по меньшей мере на 12%, по меньшей мере на 13%, по меньшей мере на 14%, по меньшей мере на 15%, по меньшей мере на 16%, по меньшей мере на 17%, по меньшей мере на 18%, по меньшей мере на 19%, по меньшей мере на 20%, по меньшей мере на 21%, по меньшей мере на 22%, по меньшей мере на 23%, по меньшей мере на 24%, по меньшей мере на 25%, по меньшей мере на 30% по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 90% (вес/вес.) больше на относительной основе, чем содержание TAG, DAG, TAG и DAG или MAG соответствующей вегетативной части растения, где отсутствует один или несколько экзогенных полинуклеотидов. В предпочтительном варианте осуществления MAG представляет собой 2-MAG. В варианте осуществления содержание TAG, DAG, TAG и DAG или MAG вегетативной части растения определяется из количества этих липидных компонентов в экстрагируемом липиде вегетативной части растения. В дополнительном варианте осуществления содержание TAG, DAG, TAG и DAG или MAG трансгенной вегетативной части растения увеличивается на количество, как определено в признаке (i).
В одном варианте осуществления по меньшей мере 60% (моль%) содержания жирной кислоты общего содержания неполярных липидов организма или его части, или липида, экстрагированного из них, представляет собой олеиновую кислоту.
В другом варианте осуществления содержание PUFA организма или его части увеличивается по сравнению с соответствующим организмом или его частью. В этом контексте содержание PUFA включает как эстерифицированную PUFA (включая TAG, DAG и т.д.), так и неэстерифицированную PUFA. В варианте осуществления содержание PUFA организма или его части предпочтительно определяется из количества PUFA в экстрагируемом липиде организма или его части. Степень увеличения в содержании PUFA может быть такой, как определено в признаке (i). Содержание PUFA может включать EDA, ARA, ALA, SDA, ETE, ETA, ЕРА, DPA, DHA или комбинацию двух или более из них.
В другом варианте осуществления уровень PUFA в организме или его части или липиде, экстрагированном из них, увеличивается по сравнению с соответствующим организмом или его частью, или липидом, экстрагированным из них. PUFA может быть EDA, ARA, ALA, SDA, ETE, ETA, EPA, DPA, DHA или комбинацией двух или более из них. Степень увеличения PUFA может быть такой, как определено в признаке (i).
В варианте осуществления уровень одного или нескольких неполярных липидов (таких как TAG, DAG, TAG и DAG, MAG, PUFA или специфическая PUFA) и/или общее содержание неполярных липидов можно определить с помощью анализа путем применения газовой хроматографии метиловых сложных эфиров жирных кислот, полученных из экстрагированного липида. Альтернативные способы для определения любого из этих содержаний известны в уровне техники и включают способы, которые не требуют экстракции липида из организма или его части, например, анализ с помощью ближнего инфракрасного диапазона (NIR) или ядерного магнитного резонанса (ЯМР).
В одном варианте осуществления один или несколько экзогенных полинуклеотидов кодируют:
i) моноацилглицерол-ацилтрансферазу (MGAT),
ii) диацилглицерол-ацилтрансферазу 2 (DGAT2),
iii) MGAT и глицерол-3-фосфат-ацилтрансферазу (GPAT) или
iv) MGAT и DGAT, или
v) MGAT, GPAT и DGAT.
В одном варианте осуществления экзогенный полинуклеотид кодирует MGAT, которая катализирует ацилирование либо sn-1 MAG, либо sn-2 MAG для образования sn-1,3-D AG или sn-l,2/2,3-DAG, соответственно. В предпочтительном варианте осуществления MGAT катализирует ацилирование sn-2 MAG для образования sn-1,2/2,3-DAG. Экзогенный полинуклеотид, кодирующий MGAT, может включать одно или несколько из следующего:
i) последовательности нуклеотидов, выбранной из любой из SEQ ID NO:1-44,
ii) последовательности нуклеотидов, кодирующей полипептид, содержащий аминокислоты, с последовательностью, как представлено в любой из SEQ ID NO:45-82, или ее биологически активным фрагментом,
iii) последовательности нуклеотидов, которая по меньшей мере на 50% идентична i) или ii), или
iv) последовательности нуклеотидов, которая гибридизируется с любой из i)-iii) при жестких условиях.
В одном варианте осуществления экзогенный полинуклеотид кодирует MGAT1, причем включает одно или несколько из следующего:
i) последовательности нуклеотидов, выбранной из любой из SEQ ID NO. 1, 3-5 или 7-23,
ii) последовательности нуклеотидов, кодирующей полипептид, содержащий аминокислоты, с последовательностью, как представлено в любой из SEQ ID NO:45-61, или ее биологически активным фрагментом,
iii) последовательности нуклеотидов, которая по меньшей мере на 50% идентична i) или ii), или
iv) последовательности нуклеотидов, которая гибридизируется с любой из i)-iii) при жестких условиях.
В другом варианте осуществления экзогенный полинуклеотид кодирует MGAT2, причем включает одно или несколько из следующего:
i) последовательности нуклеотидов, выбранной из любой из SEQ ID NO:2, 6 или 24-37,
ii) последовательности нуклеотидов, кодирующей полипептид, содержащий аминокислоты, с последовательностью, как представлено в любой из SEQ ID NO:62-75, или ее биологически активным фрагментом,
iii) последовательности нуклеотидов, которая по меньшей мере на 50% идентична i) или ii), или
iv) последовательности нуклеотидов, которая гибридизируется с любой из i)-iii) при жестких условиях.
В другом варианте осуществления экзогенный полинуклеотид кодирует MGAT3, причем включает одно или несколько из следующего:
i) последовательности нуклеотидов, выбранной из любой из SEQ ID NO:38-44,
ii) последовательности нуклеотидов, кодирующей полипептид, содержащий аминокислоты, с последовательностью, как представлено в любой из SEQ ID NO:76-82, или ее биологически активным фрагментом,
iii) последовательности нуклеотидов, которая по меньшей мере на 50% идентична i) или ii), или
iv) последовательности нуклеотидов, которая гибридизируется с любой из i)-iii) при жестких условиях.
В другом варианте осуществления экзогенный полинуклеотид кодирует DGAT, которая катализирует ацилирование либо sn-1,3-DAG, либо sn-1,2/2,3-DAG, предпочтительно sn-1,2/2,3-DAG для образования TAG. В варианте осуществления экзогенный полинуклеотид кодирует DGAT2, причем включает одно или несколько из следующего:
i) последовательности нуклеотидов, выбранной из любой из SEQ ID NO:204-211,
ii) последовательности нуклеотидов, кодирующей полипептид, содержащий аминокислоты, с последовательностью, как представлено в любой из SEQ ID NO:212-219, или ее биологически активным фрагментом,
iii) последовательности нуклеотидов, которая по меньшей мере на 50% идентична i) или ii), или
iv) последовательности нуклеотидов, которая гибридизируется с любой из i)-iii) при жестких условиях. В предпочтительном варианте осуществления DGAT2 включает последовательность нуклеотидов SEQ ID NO:204 и/или последовательность нуклеотидов, кодирующую полипептид, содержащий аминокислоты, с последовательностью, как представлено в SEQ ID NO:212.
В другом варианте осуществления экзогенный полинуклеотид кодирует глицерол-3-фосфат-ацилтрансферазу (GPAT). В предпочтительном варианте осуществления GPAT также имеет фосфатазную активность и образует MAG (т.е. GPAT, которая ацилирует G-3-P для образования либо sw-1-LPA, либо лл-2-LPA и удаляет фосфатную группу из LPA для образования MAG). В дополнительном предпочтительном варианте осуществления GPAT представляет собой sn-2-GPAT (т.е., имеет преимущество для образования sn-2-LPA из G-3-P) и имеет фосфатазную активность для образования 2-MAG, например, GPAT4 или GPAT6 Arabidopsis. Экзогенный полинуклеотид, кодирующий GPAT, может включать одно или несколько из следующего:
i) последовательности нуклеотидов, выбранной из любой из SEQ ID NO:84-141,
ii) последовательности нуклеотидов, кодирующей полипептид, содержащий аминокислоты, с последовательностью, как представлено в любой из SEQ ID NO: 144-201, или ее биологически активным фрагментом,
iii) последовательности нуклеотидов, которая по меньшей мере на 50% идентична i) или ii), или
iv) последовательности нуклеотидов, которая гибридизируется с любой из i)-iii) при жестких условиях.
В другом или дополнительном варианте осуществления экзогенный полинуклеотид кодирует GPAT с фосфатазной активностью, включающую одну или несколько консервативных аминокислотных последовательностей, как представлено в SEQ ID NO:225, 226 и 227, или последовательность аминокислот, которая по меньшей мере на 50%, предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 65% идентична им.
В одном варианте осуществления один или несколько экзогенных полинуклеотидов кодируют мутантный MGAT и/или DGAT, и/или GPAT. Например, один или несколько экзогенных полинуклеотидов могут кодировать MGAT и/или DGAT, и/или GPAT, имеющую консервативную аминокислотную замену, как проиллюстрировано в Таблице 1, относительно MGAT и/или DGAT, и/или GPAT дикого типа.
В одном варианте осуществления трансгенный, не являющийся человеком организм или его часть содержит первый экзогенный полинуклеотид, который кодирует MGAT, и второй экзогенный полинуклеотид, который кодирует GPAT. Первый и второй полинуклеотиды могут быть обеспечены как отдельные молекулы или могут быть обеспечены как одна непрерывная молекула. В предпочтительном варианте осуществления GPAT представляет собой GPAT с фосфатазной активностью, такую как GPAT4 или GPAT6 Arabidopsis. GPAT с фосфатазной активностью действует, чтобы катализировать образование MAG из G-3-P (т.е. ацилирует G-3-P для образования LPA и впоследствии удаляет фосфатную группу для образования MAG) в трансгенном, не являющемся человеком организме или его части. MGAT затем действует, чтобы катализировать образование DAG в трансгенном, не являющемся человеком организме или его части путем ацилирования MAG с помощью ацильной группы, полученной из ацил-coA жирной кислоты. MGAT, такая как MGAT1 А. thaliana, также может действовать, чтобы катализировать образование TAG в трансгенном, не являющемся человеком организме или его части, если она также имеет DGAT активность.
Трансгенный, не являющийся человеком организм или его часть может содержать третий экзогенный полинуклеотид, кодирующий, например, DGAT. Первый, второй и третий полинуклеотиды могут быть обеспечены как отдельные молекулы или могут быть обеспечены как одна непрерывная молекула. DGAT действует, чтобы катализировать образование TAG в трансгенном, не являющемся человеком организме или его части путем ацилирования DAG (предпочтительно полученного с помощью MGAT пути) с помощью ацильной группы, полученной из ацил-coA жирной кислоты.
В другом варианте осуществления трансгенный, не являющийся человеком организм или его часть содержит первый экзогенный полинуклеотид, который кодирует MGAT, и второй экзогенный полинуклеотид, который кодирует DGAT. Первый и второй полинуклеотиды могут быть обеспечены как отдельные молекулы или могут быть обеспечены как одна непрерывная молекула. Трансгенный, не являющийся человеком организм может содержать третий экзогенный полинуклеотид, кодирующий, например, GPAT, предпочтительно GPAT с фосфатазной активностью, такую как GPAT4 или GPAT6 Arabidopsis. Первый, второй и третий полинуклеотиды могут быть обеспечены как отдельные молекулы или могут быть обеспечены как одна непрерывная молекула.
В еще одном варианте осуществления уровень одного или нескольких неполярных липидов и/или общее содержание неполярных липидов трансгенного организма или его части составляет по меньшей мере на 0,5% (вес./вес.) больше на основе веса и/или по меньшей мере на 1% (вес./вес.) больше на относительной основе, чем у соответствующего организма или его части, где отсутствует один или несколько экзогенных полинуклеотидов, но который содержит экзогенный полинуклеотид, кодирующий DGAT1 Arabidopsis thaliana (SEQ ID NO: 83).
В еще дополнительном варианте осуществления трансгенный, не являющийся человеком организм или его часть дополнительно содержит одну или несколько введенных мутаций и/или экзогенный полинуклеотид, который отрицательно регулирует образование и/или активность эндогенного фермента трансгенного, не являющегося человеком организма или его части, выбранного из DGAT, sn-1 GPAT, 1-ацил-глицерол-3-фосфат-ацилтрансферазы (LPAAT), ацил-CoA:лизофосфатидилхолин-ацилтрансферазы (LPCAT), фосфатазы фосфатидной кислоты (РАР) или комбинации двух или более из них. Sn-1 GPAT может быть GPAT, которая в состоянии своего дикого типа не обладает обнаруживаемой фосфатазной активностью, например, GPAT1 или GPAT3. GPAT1 может иметь аминокислотную последовательность, как представлено в SEQ ID NO:202, или ее гомолог.GPAT3 может иметь аминокислотную последовательность, как показано в SEQ ID NO:203, или ее гомолог.
Экзогенный полинуклеотид, например, может быть выбран из следующего: антисмыслового полинуклеотида, смыслового полинуклеотида, каталитического полинуклеотида, микроРНК, полинуклеотида, который кодирует полипептид, который связывает эндогенный фермент, и двухцепочечной РНК.
В другом аспекте настоящее изобретение обеспечивает способ получения экстрагированного липида, где способ включает этапы:
i) получения трансгенного фототрофного организма или его части, содержащей экзогенную моноацилглицерол-ацилтрансферазу (MGAT), где трансгенный фототрофный организм или его часть имеет повышенный уровень неполярного липида по сравнению с таковым у организма или его части, где отсутствует экзогенная MGAT. и
ii) экстрагирования липида из трансгенного фототрофного организма или его части, таким образом получая экстрагированный липид.
В другом аспекте настоящее изобретение обеспечивает трансгенный, не являющийся человеком организм или его часть, содержащую один или несколько экзогенных полинуклеотидов, где трансгенный, не являющийся человеком организм или его часть имеет повышенный уровень одного или нескольких неполярных липидов по сравнению с соответствующим организмом или его частью, где отсутствует один или несколько экзогенных полинуклеотидов.
В предпочтительном варианте осуществления общее содержание липидов трансгенного, не являющегося человеком организма или его части увеличивается по сравнению с соответствующим организмом или его частью. Степень увеличения одного или нескольких липидов и/или общего содержания неполярных липидов может быть такой, как определено в признаке (i). Трансгенный, не являющийся человеком организм или его часть могут быть такими, как определено в признаке (ii). Неполярный липид может быть определен, как в признаке (iii). Трансгенный, не являющийся человеком организм или его часть могут быть определены с помощью комбинации признаков (i), (ii) и (iii) или признаков (i) и (ii), или признаков (i) и (iii).
В одном варианте осуществления уровень одного или нескольких неполярных липидов трансгенного, не являющегося человеком организма или его части по меньшей мере на 0,5% (вес./вес.) больше на основе веса и/или по меньшей мере на 1% (вес./вес.) больше на относительной основе, чем у соответствующего организма или его части, или как дополнительно определено в признаке (i).
В еще одном варианте осуществления общее содержание неполярных липидов трансгенного, не являющегося человеком организма или его части по меньшей мере на 0,5% (вес./вес.) больше на основе веса и/или по меньшей мере на 1% (вес./вес.) больше на относительной основе, чем у соответствующего не являющегося человеком организма или его части, или как дополнительно определено в признаке (i).
В одном варианте осуществления содержание PUFA трансгенного, не являющегося человеком организма или его части или липида, экстрагированного из них, увеличивается по сравнению с соответствующим организмом или его частью, или липидом, экстрагированным из них. Степень увеличения содержания PUFA может быть такой, как определено в признаке (i).
В другом варианте осуществления уровень PUFA в трансгенном, не являющемся человеком организме или его части, или липиде, экстрагированном из них, увеличивается по сравнению с соответствующим организмом или его частью, или липидом, экстрагированным из них. Степень увеличения PUFA может быть такой, как определено в признаке (i).
В еще одном варианте осуществления уровень одного или нескольких неполярных липидов и/или общее содержание липидов трансгенного организма или его части по меньшей мере на 0,5% (вес./вес.) больше на основе веса и/или по меньшей мере на 1% (вес./вес.) больше на относительной основе, чем у соответствующего организма или его части, где отсутствует один или несколько экзогенных полинуклеотидов, но который содержит экзогенный полинуклеотид, кодирующий DGAT1 Arabidopsis thaliana.
В одном варианте осуществления трансгенный, не являющийся человеком организм представляет собой растение, водоросль или организм, подходящий для ферментации, такой как дрожжи или гриб. Растение может быть таким, как определено в признаке (ii).
Настоящее изобретение также обеспечивает трансгенный, не являющийся человеком организм, содержащий один или несколько экзогенных полинуклеотидов, кодирующих:
i) MGAT,
ii) DGAT2,
iii) MGAT и GPAT,
iv) MGAT и DGAT, или
v) MGAT, GPAT и DGAT.
Трансгенный, не являющийся человеком организм дополнительно можно охарактеризовать с помощью одного или нескольких признаков, определенных в данном документе. Один или несколько экзогенных полинуклеотидов могут содержать последовательность, как определено выше.
В другом аспекте настоящее изобретение обеспечивает способ получения клетки с усиленной способностью к образованию одного или нескольких неполярных липидов, где способ включает следующее:
i) введение в клетку одного или нескольких экзогенных полинуклеотидов, кодирующих
a) MGAT,
b) DGAT2,
c) MGAT и GPAT,
d) MGAT и DGAT, или
e) MGAT, GPAT и DGAT,
где один или несколько экзогенных полинуклеотидов функционально связаны с одним или несколькими промоторами, которые способны управлять экспрессией одного или нескольких экзогенных полинуклеотидов в клетке,
ii) экспрессирование одного или нескольких экзогенных полинуклеотидов в клетке,
iii) анализ содержания липидов клетки, и
iv) выбор клетки, имеющей повышенный уровень одного или нескольких неполярных липидов по сравнению с соответствующей клеткой, где отсутствуют экзогенные полинуклеотиды.
В предпочтительном варианте осуществления общее содержание липидов выбранной клетки увеличивается по сравнению с соответствующей клеткой.
В одном варианте осуществления один или несколько неполярных липидов и/или общее содержание неполярных липидов выбранной клетки составляет по меньшей мере на 0,5% (вес./вес.) больше на основе веса и/или по меньшей мере на 1% (вес./вес.) больше на относительной основе, чем у соответствующей клетки, или как дополнительно определено в признаке (i).
В другом варианте осуществления содержание PUFA выбранной клетки увеличивается по сравнению с соответствующей клеткой. Степень увеличения содержания PUFA может быть такой, как определено в признаке (i).
В другом варианте осуществления уровень PUFA в выбранной клетке увеличивается по сравнению с соответствующей клеткой. Степень увеличения PUFA может быть такой, как определено в признаке (i).
Один или несколько экзогенных полинуклеотидов в этом аспекте могут включать последовательность, как определено выше. Дополнительно, один или несколько экзогенных полинуклеотидов могут быть неизвестными до данного способа в отношении кодирования MGAT, DGAT2, MGAT и GPAT, MGAT и DGAT, или MGAT, GPAT и DGAT, но скорее могут быть кандидатами для этого. Способ, следовательно, можно применять в качестве анализа для определения или выбора полинуклеотидов, кодирующих MGAT, DGAT2, MGAT и GPAT (предпочтительно GPAT, также имеющую фосфатазную активность), MGAT и DGAT, или MGAT, GPAT (предпочтительно GPAT, также имеющую фосфатазную активность) и DGAT.
Экспрессия GPAT может приводить к увеличению в уровнях MAG в клетке, если GPAT также имеет фосфатазную активность, экспрессия MGAT может приводить к увеличению в уровнях DAG в клетке, тогда как экспрессия DGAT может приводить к увеличению в уровнях TAG в клетке. Экспрессия MGAT также может приводить к увеличению в уровнях TAG в клетке, если MGAT также имеет DGAT активность. Экспрессия MGAT и DGAT может приводить к увеличению в уровнях DAG и/или TAG в клетке. Экспрессия MGAT и GPAT может приводить к увеличению в уровнях MAG (если GPAT также имеет фосфатазную активность) и/или DAG в клетке. Экспрессия MGAT и GPAT также может приводить к увеличению в уровнях TAG в клетке, если MGAT также имеет DGAT активность. Экспрессия MGAT, GPAT и DGAT может приводить к увеличению в уровнях MAG (если GPAT также имеет фосфатазную активность) и/или DAG, и/или TAG в клетке.
В одном варианте осуществления выбранная клетка представляет собой клетку согласно настоящему изобретению.
В одном варианте осуществления экзогенные полинуклеотиды являются стабильно интегрированными в геном клетки.
Способ может дополнительно включать этапы регенерации трансгенного организма из клетки и/или получения потомства из клетки, например, получения семени, причем этапы могут иметь место в любой момент после этапа i).
В альтернативном варианте осуществления экзогенные полинуклеотиды временно экспрессируются в клетке.
В другом аспекте настоящее изобретение обеспечивает трансгенную клетку или растение, полученные с применением способа по настоящему изобретению, или их потомство.
В другом аспекте настоящее изобретение обеспечивает применение одного или нескольких экзогенных полинуклеотидов, кодирующих
i) MGAT,
ii) DGAT2,
iii) MGAT и GPAT,
iv) MGAT и DGAT, или
v) MGAT, GPAT и DGAT,
для получения трансгенного, не являющегося человеком организма или его части с усиленной способностью к образованию одного или нескольких неполярных липидов по сравнению с соответствующим организмом или его частью, где отсутствует один или несколько экзогенных полинуклеотидов. Степень усиленной способности может быть такой, как определено в признаке (i). Трансгенный, не являющийся человеком организм или его часть могут быть определены, как в признаке (ii), или могут быть определены с помощью комбинации признаков (i), (ii) и (iii) или признаков (i) и (ii), или признаков (i) и (iii). Один или несколько экзогенных полинуклеотидов могут содержать последовательность, как определено выше.
В другом аспекте настоящее изобретение обеспечивает трансгенное семя растения, причем семя содержит один или несколько экзогенных полинуклеотидов и имеет повышенный уровень одного или нескольких неполярных липидов и/или общего содержания неполярных липидов по сравнению с соответствующим семенем, где отсутствуют экзогенные полинуклеотиды. Степень увеличения может быть такой, как определено в признаке (i). Трансгенное семя может быть из рода и/или вида, как определено в признаке (ii). Трансгенное семя может быть определено с помощью комбинации признаков (i), (ii) и (iii) или признаков (i) и (ii), или признаков (i) и (iii). Один или несколько экзогенных полинуклеотидов могут содержать последовательность, как определено выше.
В другом аспекте настоящее изобретение обеспечивает трансгенное растение, которое образует семя по настоящему изобретению. Трансгенное растение, например, может быть Brassica sp., Gossypium hirsutum, Linum usitatissimum, Helianthus sp., Carthamus tinctorius, Glycine max, Zea mays, Arabidopsis thaliana, Sorghum bicolor. Sorghum vulgare, Avena sativa, Trifolium sp., Elaesis guineenis, Nicotiana benthamiana, Hordeum vulgare, Lupinus angustifolius, Oryza sativa, Oryza glaberrima, Camelina sativa, Miscanthus x giganteus или Miscanthus sinensis.
В другом аспекте настоящее изобретение обеспечивает способ получения семени, причем способ включает следующее:
i) выращивание трансгенного растения по настоящему изобретению, и
ii) сбор семян.
В другом аспекте настоящее изобретение обеспечивает процесс ферментации, включающий этапы:
i) обеспечения сосуда, содержащего жидкую композицию, содержащую трансгенный, не являющийся человеком организм по настоящему изобретению, который является подходящим для ферментации, и составляющих, необходимых для ферментации и биосинтеза жирной кислоты, и
ii) обеспечения условий, способствующих ферментации жидкой композиции, содержащейся в указанном сосуде.
В другом аспекте настоящее изобретение обеспечивает экстрагированный липид, получаемый из способа по настоящему изобретению, трансгенного, не являющегося человеком организма по настоящему изобретению, трансгенной клетки по настоящему изобретению, трансгенного растения по настоящему изобретению или трансгенного семени по настоящему изобретению. Экстрагируемый липид может иметь усиленное содержание TAG, содержание DAG, содержание TAG и DAG, содержание MAG, содержание PUFA или содержание специфической PUFA и/или общее содержание неполярных липидов. В предпочтительном варианте осуществления MAG представляет собой 2-MAG. Степень увеличенного содержания TAG, содержания DAG, содержания TAG и DAG, содержания MAG, содержания PUFA, содержания специфической PUFA и/или общего содержания неполярных липидов может быть такой, как определено в признаке (i).
Настоящее изобретение также обеспечивает экстрагированный липид, включающий содержание DAG, которое составляет по меньшей мере 1% (вес./вес.) или по меньшей мере 2% (вес./вес.) на основе веса от общего содержания экстрагированного липида. Предпочтительно, экстрагированный липид также содержит MAG на обнаруживаемых уровнях. В одном варианте осуществления экстрагированный липид представляет собой масло канолы, кукурузы, сои, люпина, арахиса, подсолнечника, хлопка, сафлора или льна. Липид может включать эруковую кислоту, циклопропеноидные жирные кислоты (CPFA) и/или глюкозинолаты на обнаруживаемых уровнях.
Настоящее изобретение также обеспечивает применение трансгенного, не являющегося человеком организма или его части по настоящему изобретению, трансгенной клетки по настоящему изобретению, трансгенного растения по настоящему изобретению, трансгенного семени по настоящему изобретению или экстрагированного липида по настоящему изобретению для производства промышленного продукта. Промышленный продукт, например, может быть топливом. Топливо может содержать минимальный уровень экстрагированного липида согласно настоящему изобретению, такой как по меньшей мере 10%, по меньшей мере 20% или по меньшей мере 30% (вес/вес).
В другом аспекте настоящее изобретение обеспечивает способ получения топлива, причем способ включает следующее:
i) реакцию липида по настоящему изобретению со спиртом, необязательно, в присутствии катализатора, для образования алкиловых сложных эфиров, и
ii) необязательно, смешивание алкиловых сложных эфиров с топливом на нефтяной основе.
В одном варианте осуществления алкиловые сложные эфиры представляют метиловые сложные эфиры. Топливо, полученное с помощью данного способа, может содержать минимальный уровень липида по настоящему изобретению, такой как по меньшей мере 10%, по меньшей мере 20% или по меньшей мере 30% (вес./вес).
В другом аспекте настоящее изобретение обеспечивает способ получения пищевого продукта, причем способ включает примешивание трансгенного, не являющегося человеком организма или его части по настоящему изобретению, трансгенной клетки по настоящему изобретению, трансгенного растения по настоящему изобретению, трансгенного семени по настоящему изобретению или экстрагированного липида по настоящему изобретению, или их экстракта или части, к по меньшей мере одному другому пищевому ингредиенту.
В другом аспекте настоящее изобретение обеспечивает пищевые продукты, косметические средства или химические средства, содержащие трансгенный, не являющийся человеком организм или его часть по настоящему изобретению, трансгенную клетку по настоящему изобретению, трансгенное растение по настоящему изобретению, трансгенное семя по настоящему изобретению или экстрагированный липид по настоящему изобретению, или их экстракт или часть.
В другом аспекте настоящее изобретение обеспечивает способ для определения молекулы нуклеиновой кислоты, кодирующей ацилтрансферазу с увеличенной способностью к образованию MAG, DAG и/или TAG в клетке, причем способ включает следующее:
i) получение клетки, содержащей молекулу нуклеиновой кислоты, функционально связанную с промотором, который является активным в клетке, где молекула нуклеиновой кислоты включает последовательность нуклеотидов, как определено в данном документе, и/или последовательность нуклеотидов, кодирующую полипептид, имеющий одну или несколько аминокислотных последовательностей, как представлено в SEQ ID NO:220, 221, 222, 223, 224, 225, 226 и 227, или последовательность аминокислот, которая по меньшей мере на 50%, предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 65% идентична им,
ii) определение того, увеличивается ли уровень MAG, DAG и/или TAG, образованных в клетке, по сравнению с соответствующей клеткой, где отсутствует нуклеиновая кислота, и
iii) определение молекулы нуклеиновой кислоты, кодирующей ацилтрансферазу, имеющую повышенную способность к образованию MAG, DAG и/или TAG в клетке.
В одном варианте осуществления молекула нуклеиновой кислоты включает последовательность нуклеотидов, кодирующую одну или несколько консервативных аминокислотных последовательностей DGAT2 и/или MGAT 1/2, как представлено в SEQ ID NO:220, 221, 222, 223 и 224. В предпочтительном варианте осуществления молекула нуклеиновой кислоты включает последовательность нуклеотидов, кодирующую консервативные аминокислотные последовательности, представленные в SEQ ID NO:220 и/или SEQ ID NO:224. В другом или дополнительном варианте осуществления молекула нуклеиновой кислоты включает последовательность нуклеотидов, кодирующую одну или несколько консервативных аминокислотных последовательностей GPAT, как представлено в SEQ ID NO:225, 226 и 227, или последовательность аминокислот, которая по меньшей мере на 50%, предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 65% идентична им.
В одном варианте осуществления способ предназначен для определения MGAT, имеющей повышенную способность к образованию DAG и/или TAG.
В другом варианте осуществления способ предназначен для определения GPAT с фосфатазной активностью и повышенной способностью к образованию MAG. В предпочтительном варианте осуществления способ предназначен для определения sn-2-GPAT с фосфатазной активностью и повышенной способностью к образованию 2-MAG.
В другом варианте осуществления способ предназначен для определения MGAT и GPAT, которые вместе имеют повышенную способность к образованию MAG и/или DAG, и/или TAG, предпочтительно TAG. В предпочтительном варианте осуществления способ предназначен для определения 5w-2-GPAT с фосфатазной активностью для образования 2-MAG и, вместе с MGAT, повышенной способности к образованию sn-1,2/2,3-DAG и/или TAG.
В другом варианте осуществления способ предназначен для определения DGAT, имеющей повышенную способность к образованию TAG.
В другом варианте осуществления способ предназначен для определения MGAT и DGAT, которые вместе имеют повышенную способность к образованию DAG и/или TAG, предпочтительно TAG.
В другом варианте осуществления способ предназначен для определения MGAT, GPAT и DGAT, которые вместе имеют повышенную способность к образованию MAG и/или DAG, и/или TAG, предпочтительно TAG. В предпочтительном варианте осуществления GPAT имеет фосфатазную активность для образования MAG. В дополнительном предпочтительном варианте осуществления GPAT представляет собой sn-2-GPAT с фосфатазной активностью для образования 2-MAG.
В одном варианте осуществления способ дополнительно включает введение молекулы нуклеиновой кислоты в клетку перед этапом i).
В другом варианте осуществления способ дополнительно включает этап регенерации трансгенного растения из клетки этапа i) и, необязательно, получение семени из трансгенного растения. В одном варианте осуществления клетка представляет собой растительную клетку. Степень повышенной способности к образованию MAG и/или DAG, и/или TAG может быть такой, как определено в признаке (i). В предпочтительном варианте осуществления MAG представляет собой 2-MAG. Клетка может быть из рода и/или вида, как определено в признаке (ii). Клетка может быть определена с помощью комбинации признаков (i), (ii) и (iii), или признаков (i) и (ii), или признаков (i) и (iii).
В другом варианте осуществления MGAT и/или GPAT, и/или DGAT увеличивает образование TAG в клетке на большее количество, чем DGAT1 Arabidopsis thaliana (SEQ ID NO:83).
В другом аспекте настоящее изобретение обеспечивает выделенный и/или рекомбинантный полинуклеотид, включающий следующее:
i) последовательность нуклеотидов, выбранную из любой из SEQ ID NO: 1-6, или
ii) последовательность нуклеотидов, которая по меньшей мере на 80% идентична i).
В другом аспекте настоящее изобретение обеспечивает вектор, содержащий полинуклеотид по настоящему изобретению.
В другом аспекте настоящее изобретение обеспечивает трансгенную клетку, содержащую полинуклеотид по настоящему изобретению или вектор по настоящему изобретению.
В другом аспекте настоящее изобретение обеспечивает трансгенный, не являющийся человеком организм или его часть, содержащую полинуклеотид по настоящему изобретению, вектор по настоящему изобретению или трансгенную клетку по настоящему изобретению. В одном варианте осуществления трансгенный организм представляет собой растение, водоросль или организм, подходящий для ферментации, такой как дрожжи или гриб. В другом варианте осуществления часть представляет собой семя, плод, клубень, корень или вегетативную часть растения.
Специалистам в данной области техники будет понятно, что многочисленные изменения и/или модификации могут быть сделаны в настоящем изобретении, как показано в конкретных вариантах осуществления, без отступления от объема настоящего изобретения, как описано в широком смысле. Настоящие варианты осуществления, следовательно, должны рассматриваться во всех отношениях как иллюстративные и неограничивающие. Каждый вариант осуществления, описанный в данном документе, подлежит применению mutatis mutandis ко всем и каждому другому варианту осуществления, если особо не указано иное.
По всему данному описанию слово "содержать" или варианты, такие как "содержит" или "содержащий", будет пониматься как подразумевающее включение указанного элемента, целого числа или этапа, или группы элементов, целых чисел или этапов, но не исключение какого-либо другого элемента, целого числа или этапа, или группы элементов, целых чисел или этапов.
Выражение "и/или", например, "X и/или Y" должно пониматься как означающее либо "X и Y", либо "X или Y" и должно использоваться для обеспечения недвусмысленного обоснования для обоих значений или для одного из значений.
КРАТКОЕ ОПИСАНИЕ ПРИЛАГАЕМЫХ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1. Представление различных путей синтеза липидов, большинство из которых сходятся на DAG, центральной молекуле в синтезе липидов. Эта модель включает один возможный путь для образования sn-2-MAG, который можно использовать с помощью бифункциональной MGAT/DGAT для образования DAG из глицерол-3-фосфата (G-3-Р). Сокращения являются следующими:
G-3-P; глицерол-3-фосфат
LysoPA; лизофосфатидная кислота
РА; фосфатидная кислота
MAG; моноацилглицерол
DAG; диацилглицерол
TAG; триацилглицерол
Ацил-СоА и FA-CoA; ацил-коэнзим A и ацил-коэнзим A жирной кислоты PC; фосфатидилхолин
GPAT; глицерол-3-фосфат-ацилтрансфераза; глицерол-3-фосфат-О-ацилтрансфераза; ацил-СоА: от-глицерол-3-фосфат-1-O-ацилтрансфераза; ЕС 2.3.1.15 GPAT4; глицерол-3-фосфат-ацилтрансфераза 4 GPAT6; глицерол-3-фосфат-ацилтрансфераза 6
LPAAT; 1-ацил-глицерол-3-фосфат-ацилтрансфераза; 1-ацилглицерол-3-фосфат-(9-ацилтрансфераза; ацил-CoA: 1-ацил-sn-глицерол-3-фосфат-2-O-ацилтрансфераза; EC 2.3.1.51
PAP; фосфатаза фосфатидной кислоты; фосфатидат-фосфатаза; фосфогидролаза фосфатной кислоты; фосфатаза фосфатидной кислоты; ЕС 3.1.3.4
MGAT; ацилтрансфераза, имеющая моноацилглицерол-ацилтрансферазную активность (MGAT; 2-ацилглицерол-(9-ацилтрансфераза, ацил-CoA: 2-ацилглицерол-O-ацилтрансфераза; ЕС 2.3.1.22)
M/DGAT; ацилтрансфераза, имеющая моноацилглицерол-ацилтрансферазную (MGAT; 2-ацил глицерол-О-ацилтрансфераза; ацил-СоА:2-ацилглицецерол-О-ацилтрансфераза; ЕС 2.3.1.22) и/или диацилглицерол-ацилтрансферазную активность (DGAT; диацилглицерол-О-ацилтрансфераза; ацил-СоА: 1,2-диацил-sn-глицерол-sn-ацилтрансфераза; ЕС 2.3.1.20)
LPCAT; ацил-СоА:лизофосфатидилхолин-ацилтрансфераза; 1-ацилглицерофосфохолин-O-ацилтрансфераза; ацил-CoA.1-ацил-sn-глицеро-3-фосфохолин-О-ацилтрансфераза; EC 2.3.1.23
PLD-Z; Фосфолипаза D зета; холиновая фосфатаза; лецитиназа D; липофосфодиэстераза II; EC 3.1.4.4
СРТ; СВР-холин:диацилглицерол-холинфосфотрансфераза; 1-алкил-2-ацетилглицерол-холинфосфотрансфераза; алкилацилглицерол-холинфосфотрансфераза; холинфосфотрансфераза; фосфорилхолин-глицерид-трансфераза; ЕС 2.7.8.2
PDCT; фосфатидилхолин:диацилглицерол-холинфосфотрансфераза
PLC; фосфолипаза C; ЕС 3.1.4.3
PDAT; фосфолипид:диацилглицерол-ацилтрансфераза; фосфолипид: 1,2-диацил-sn-глицерол-O-ацилтрансфераза; EC 2.3.1.158
Pi; неорганический фосфат
Фигура 2. Относительные увеличения DAG и TAG в ткани листа Nicotiana benthamiana, трансформированного при помощи конструктов, кодирующих p 19 (отрицательный контроль), DGAT1 Arabidopsis thaliana, MGAT1 Mus "musculus и комбинацией DGAT1 и MGAT1, каждый из которых экспрессируется с промотора 35S. Фермент MGAT1 был значительно более активным, чем фермент DGAT1 в стимуляции накопления как DAG, так и TAG в ткани листа. Экспрессия гена MGAT1 приводила к в два раза большему накоплению DAG и TAG в ткани листа по сравнению с экспрессией только DGAT1.
Фигура 3. Относительные увеличения TAG в листе N.benthamiana, трансформированном при помощи конструктов, кодирующих р19 (отрицательный контроль), DGAT1 A.thaliana, MGAT2 М.musculus и комбинацией MGAT2 и DGAT1. Усы погрешностей обозначают стандартную ошибку от образцов в трех повторностях.
Фигура 4. Радиоактивность (DPM) в фракциях MAG, DAG и TAG, выделенных из лизатов листьев временно трансформированного N. benthamiana, с введенными sn-2-MAG[14C] и немеченой олеиновой кислотой, в зависимости от времени. Примененные конструкты были такими, как для фигуры 3.
Фигура 5. Как для фигуры 4, но с введенными [I4C]G-3-P и немеченой олеиновой кислотой.
Фигура 6. Радиоаутограмма TLC планшета, показывающая образование TAG с помощью DGAT1 A. thaliana и MGAT1 М. musculus, но не MGAT2 М. musculus в анализе на дрожжах. Анализ описан в примере 5
КЛЮЧ К ПЕРЕЧНЮ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:l кодон-оптимизированный MGAT1 Mus musculus
SEQ ID NO:2 кодон-оптимизированный MGAT2 Mus musculus
SEQ ID NO:3 кодон-оптимизированный предсказанный MGAT1 Ciona intestinalis
SEQ ID NO:4 кодон-оптимизированный предсказанный MGAT1 Tribolium castaneum
SEQ ID NO:5 кодон-оптимизированный MGAT1 Danio rerio
SEQ ID NO:6 кодон-оптимизированный MGAT2 Danio rerio
SEQ ID NO:7 полинуклеотид MGAT1 Mus musculus (AF384162)
SEQ ID NO:8 полинуклеотид MGAT1 Homo sapiens (AF384163)
SEQ ID NO:9 предсказанный полинуклеотид MGAT1 Pan troglodytes, вариант
транскрипта (XM001166055)
SEQ ID NO:10 предсказанный полинуклеотид MGAT1 Pan troglodytes, вариант 2 транскрипта (XM_0526044.2)
SEQ ID NO:l1 предсказанный полинуклеотид MGAT1 Canis familiaris (XM545667.2)
SEQ ID NO:12 полинуклеотид MGAT1 Bos taurus (NMOO1001153.2)
SEQ ID NO:13 полинуклеотид MGAT1 Rattus norvegicus (NM 001108803.1)
SEQ ID NO:14 полинуклеотид MGAT1 Danio rerio (NM001122623.1)
SEQ ID NO:15 предсказанный полинуклеотид MGAT1 Caenorhabditis elegans (NM_073012.4)
SEQ ID NO:16 предсказанный полинуклеотид MGAT1 Caenorhabditis elegans (NMl 82380.5)
SEQ ID NO:17 предсказанный полинуклеотид MGAT1 Caenorhabditis elegans (NM_065258.3)
SEQ ID NO:18 предсказанный полинуклеотид MGAT1 Caenorhabditis elegans (NM_075068.3)
SEQ ID NO:19 предсказанный полинуклеотид MGAT1 Caenorhabditis elegans (NM_072248.3)
SEQ ID NO:20 предсказанный полинуклеотид MGAT1 Kluyveromyces lactis (XM_455588.1)
SEQ ID NO:21 предсказанный полинуклеотид MGAT 1 Ashbya gossypii (NM_208895.1)
SEQ ID NO:22 предсказанный полинуклеотид MGAT1 Magnaporthe oryzae (XM_36874I.l)
SEQ ID NO:23 предсказанный полинуклеотид MGAT1 Ciona intestinalis (XM_002120843.1)
SEQ ID NO:24 полинуклеотид MGAT2 Mus musculus (AY 157609)
SEQ ID NO:25 полинуклеотид MGAT2 Homo sapiens (AY 157608)
SEQ ID NO:26 предсказанный полинуклеотид MGAT2 Pan troglodytes (XM522112.2)
SEQ ID NO:27 предсказанный полинуклеотид MGAT2 Canis familiaris (XM 542304.1)
SEQ ID NO:28 полинуклеотид MGAT2 Bos taurus (NM_001099136.1)
SEQ ID NO:29 полинуклеотид MGAT2 Rattus norvegicus (NM_001109436.2)
SEQ ID NO:30 предсказанный полинуклеотид MGAT2 Gallus gallus (XM 424082.2)
SEQ ID NO:31 полинуклеотид MGAT2 Danio rerio (NM_001006083.1)
SEQ ID NO:32 полинуклеотид MGAT2 Drosophila melanogaster (NM_136474.2)
SEQ ID NO:33 полинуклеотид MGAT2 Drosophila melanogaster (NM 136473.2)
SEQ ID NO:34 полинуклеотид MGAT2 Drosophila melanogaster (NM 136475.2)
SEQ ID NO:35 полинуклеотид MGAT2 Anopheles gambiae (XM_001688709.1)
SEQ ID NO:36 полинуклеотид MGAT2 Anopheles gambiae (XM_3 15985)
SEQ ID NO:37 предсказанный полинуклеотид MGAT2 Tribolium castaneum (XM 970053.1)
SEQ ID NO:38 полинуклеотид MGAT3 Homo sapiens (AY229854)
SEQ ID NO:39 предсказанный полинуклеотид MGAT3 Pan troglodytes, вариант 1 транскрипта (XM_001154107.1)
SEQ ID NO:40 предсказанный полинуклеотид MGAT3 Pan troglodytes, вариант 2 транскрипта (XM_001154171.1)
SEQ ID NO:41 предсказанный полинуклеотид MGAT3 Pan troglodytes, вариант 3 транскрипта (XM527842.2)
SEQ ID NO:42 предсказанный полинуклеотид MGAT3 Canis familiaris (XM 845212.1)
SEQ ID NO:43 предсказанный полинуклеотид MGAT3 Bos taurus (XM 870406.4)
SEQ ID NO:44 предсказанный полинуклеотид MGAT3 Danio rerio (XM 688413.4)
SEQ ID NO:45 полипептид MGAT1 Mm musculus (AAK84177.1)
SEQ ID NO:46 полипептид MGAT1 Homo sapiens (AAK84178.1)
SEQ ID NO:47 предсказанная изоформа 1 полипептида MGAT1 Pan troglodytes (XP_001166055.1)
SEQ ID NO:48 предсказанная изоформа 2 полипептида MGAT1 Pan troglodytes (XP 526044.2)
SEQ ID NO:49 предсказанный полипептид MGAT1 Canis familiaris (XP545667.2)
SEQ ID NO:50 полипептид MGAT1 Bos taurus (NP_001001 153.1)
SEQ ID NO:51 полипептид MGAT1 Rattus norvegicus (NPOOl 102273.1)
SEQ ID NO:52 полипептид MGAT1 Danio rerio (NPOOl 116095.1)
SEQ ID NO:53 предсказанный полипептид MGAT1 Caenorhabditis elegans (NP 505413.1)
SEQ ID NO:54 предсказанный полипептид MGAT1 Caenorhabditis elegans (NP_872180.1)
SEQ ID NO:55 предсказанный полипептид MGAT1 Caenorhabditis elegans (NP 497659.1)
SEQ ID NO:56 предсказанный полипептид MGAT1 Caenorhabditis elegans (NP_507469.1)
SEQ Ю NO:57 предсказанный полипептид MGAT1 Caenorhabditis elegans (NP_504649.1)
SEQ ID NO:58 предсказанный полипептид MGAT1 Kluyveromyces lactis (XP455588.1)
SEQ ID NO:59 предсказанный полипептид MGAT1 Ashbya gossypii (NP 983542.1)
SEQ ID NO:60 предсказанный полипептид MGAT1 Magnaporthe oryzae (XP 368741.1)
SEQ ID NO:61 предсказанный полипептид MGAT1 Ciona intestinalis (XP002120879)
SEQ ID NO:62 полипептид MGAT2 Mus musculus (AA023673.1)
SEQ ID NO:63 полипептид MGAT2 Homo sapiens (AA023672.1)
SEQ ID NO:64 предсказанный полипептид MGAT2 Pan troglodytes (XP 522112.2)
SEQ ID NO:65 предсказанный полипептид MGAT2 Canis familiaris (XP 542304.1)
SEQ ID NO:66 полипептид MGAT2 Bos taurus (NPOO 1092606.1)
SEQ ID NO:67 полипептид MGAT2 Rattus norvegicus (NPOOl 102906.2)
SEQ ID NO:68 предсказанный полипептид MGAT2 Callus gallus (XP424082.2)
SEQ ID NO:69 полипептид MGAT2 Danio rerio (NPOO 1006083.1)
SEQ ID NO:70 полипептид MGAT2 Drosophila melanogaster (NP610318.1)
SEQ ID NO:71 полипептид MGAT2 Drosophila melanogaster (NP610317.1)
SEQ ID NO:72 полипептид MGAT2 Drosophila melanogaster (NP610319.2)
SEQ ID NO:73 полипептид MGAT2 Anopheles gambiae (XP_001688761)
SEQ ID NO:74 полипептид MGAT2 Anopheles gambiae (XPJ 15985.3)
SEQ ID NO:75 предсказанный полипептид MGAT2 Tribolium castaneum (XP 975146)
SEQ ID NO:76 полипептид MGAT3 Homo sapiens (AA063579.1)
SEQ ID NO:77 предсказанная изоформа 1 полипептида MGAT3 Pan troglodytes (XP_001154107.1)
SEQ ID NO:78 предсказанная изоформа 2 полипептида MGAT3 Pan troglodytes (XP_001154171.1)
SEQ ID NO:79 предсказанная изоформа 3 MGAT3 Pan troglodytes (XP527842.2)
SEQ ID NO:80 предсказанный полипептид MGAT3 Canis familiaris (XP 850305.1)
SEQ ID NO:81 предсказанный полипептид MGAT3 Bos taurus (XP 875499.3)
SEQ ID NO:82 предсказанный полипептид MGAT3 Danio rerio (XP 693505.1)
SEQ ID NO:83 полипептид DGAT1 Arabidopsis thaliana (CAB44774.1)
SEQ ID NO:84 полинуклеотид GPAT4 Arabidopsis thaliana (NMl 00043.4)
SEQ ID NO:85 полинуклеотид GPAT6 Arabidopsis thaliana (NMJ29367.3)
SEQ ID NO:86 полинуклеотид ВАС F'5110 Arabidopsis thaliana (AF195115.1)
SEQ ID NO: 87 полинуклеотид неизвестного белка Arabidopsis thaliana (AY062466.1)
SEQ ID NO:88 полинуклеотид из хромосомы 3 Oryza sativa (AC118133.4)
SEQ ID NO:89 полинуклеотид из клона WS0276_F13 Picea sitchensis (EF086095.1)
SEQ ID NO:90 полинуклеотид из клона ZMBFcOl 10A1 Zea mays (BT067649.1)
SEQ ID NO:91 полинуклеотид из клона RAFL16-19-H05 Arabidopsis thaliana (AK228870.1)
SEQ ID NO:92 полинуклеотид из клона J065058J01 Oryza sativa (AK241033.1)
SEQ ID NO:93 полинуклеотид из хромосомы 2 Oryza sativa (CM000127.1)
SEQ ID NO:94 полинуклеотид из хромосомы 5 Oryza sativa (CM000130.1)
SEQ ID NO:95 полинуклеотид из хромосомы 2 Oryza sativa (CM000139.1)
SEQ ID NO:96 полинуклеотид из хромосомы 1 Oryza sativa (CM000126.1)
SEQ ID NO:97 полинуклеотид из хромосомы 3 Oryza sativa (CM000128.1)
SEQ ID NO:98 полинуклеотид из хромосомы 3 Oryza sativa (CM000140.1)
SEQ ID NO:99 полинуклеотид SELMOscaffold_102 Selaginella moellendorffu (GL377667.1)
SEQ ID NO:100 полинуклеотид SELMOscaffold_102 Selaginella moellendorffu (GL377667.1)
SEQ ID NO:101 полинуклеотид SELMOscaffold_83 Selaginella moellendorffu (GL377648.1)
SEQ ID NO:102 полинуклеотид SELMOscaffold_57 Selaginella moellendorffu (GL3 77622.1)
SEQ ID NO:103 полинуклеотид SELMOscaffold_25 Selaginella moellendorffu (GL377590.1)
SEQ ID NO:104 полинуклеотид SELMOscaffold_11 Selaginella moellendorffu (GL377576.1)
SEQ ID NO:105 полинуклеотид SELMOscaffold_11 Selaginella moellendorffu (GL377576.1)
SEQ ID NO:106 полинуклеотид Os01g0855000 Oryza sativa (NM_001051374.2)
SEQ ID NO: 107 полинуклеотид Os02g0114400 Oryza sativa (NM_00O1052203.1)
SEQ ID NO:108: полинуклеотид GPAT8 Zea mays (NM_001153970.1)
SEQ ID NO:109: полинуклеотид LOC100282930 Zea mays (NM_001155835.1)
SEQ ID NO:110: полинуклеотид LOC100382119 Zea mays (NM 001174880.1)
SEQ ID NO: 111 полинуклеотид GPAT6 Arabidopsis thaliana (NM_129367.3)
SEQ ID NO:112 полинуклеотид GPAT8 Arabidopsis thaliana (NM_116264.5)
SEQ ID NO:113 полинуклеотид предсказанного белка (PHYPADRAFTJ 28739) Physcomitrella patens (XM_001764949.1)
SEQ ID NO:114 полинуклеотид предсказанного белка (PHYPADRAFT 83824) Physcomitrella patens (XM_001769619.1)
SEQ ID NO:115 полинуклеотид предсказанного белка (PHYPADRAFTJ 88308) Physcomitrella patens (XM_001769672.1)
SEQ ID NO:116 полинуклеотид предсказанного белка (PHYPADRAFT 189499) Physcomitrella patens (XM_001771134.1)
SEQ ID NO:117 полинуклеотид предсказанного белка (PHYPADRAFT 95487) Physcomitrella patens (XM_001780481.1)
SEQ ID NO:118 полинуклеотид предсказанного белка LOCI00243321 Vitis Vinifera (XM_002268477.1)
SEQ ID NO:119 полинуклеотид предсказанного белка LOCI00243093 Vitis Vinifera (XM_002275312.1)
SEQ ID NO:120 полинуклеотид предсказанного белка LOC100259433 Vitis Vinifera (XM 002275996.1)
SEQ IDNO:121 полинуклеотид предсказанного белка LOCI00264832 Vitis Vinifera (XM_002279055.1)
SEQ ID NO:122 полинуклеотид предсказанного белка Populus trichocarpa (XM 002309088.1)
SEQ ID NO:123 полинуклеотид предсказанного белка Populus trichocarpa (XM_0023 09240.1)
SEQ ID NO:124 полинуклеотид предсказанного белка Populus trichocarpa (XM_002322716.1)
SEQ ID NO:125 полинуклеотид предсказанного белка Populus trichocarpa (XM_002323527.1)
SEQ ID NO:126 полинуклеотид белка Sorghum bicolor (XM 002439842.1)
SEQ ID NO:127 полинуклеотид белка Sorghum bicolor (XM 002458741.1)
SEQ ID NO:128 полинуклеотид белка Sorghum bicolor (XM 002463 871.1)
SEQ ID NO:129 полинуклеотид белка Sorghum bicolor (XM 002464585.1)
SEQ ID NO:130 полинуклеотид ER глицерол-фосфат-ацилтрансферазы Ricinus communis (XM_002511827.1)
SEQ ID NO:131 полинуклеотид ER глицерол-фосфат-ацилтрансферазы Ricinus communis (XM_002517392.1)
SEQ ID NO:132 полинуклеотид ER глицерол-фосфат-ацилтрансферазы Ricinus communis (XM_002520125.1)
SEQ IDNO:133 полинуклеотид белка семейства фосфолипид/глицерол-ацилтрансферазы Arabidopsis lyrata (ХМ_002872909.1)
SEQ ID NO:134 полинуклеотид GPAT6 Arabidopsis lyrata (XM_002881518.1)
SEQ ID NO:135 предполагаемый полинуклеотид GPAT8 Verniciafordii (FJ479753.1)
SEQ ID NO:136 полинуклеотид Os03g0735900 Oryza sativa (NM_001057724.1)
SEQ ID NO:137 полинуклеотид GPAT4 Arabidopsis thaliana (NM 100043.4)
SEQ ID NO: 138 полинуклеотид предсказанного белка Populus trichocarpa (XM_002320102.1)
SEQ ID NO:139 полинуклеотид GenKaSorghum bicolor (XM 002451332.1)
SEQ ID NO:140 полинуклеотид ER глицерол-фосфат-ацилтрансферазы Ricinus communis (XM_002531304.1)
SEQ ID NO:141 полинуклеотид GPAT4 Arabidopsis lyrata (XM_002889315.1)
SEQ ID NO:142 полинуклеотид GPAT1 Arabidopsis thaliana (NM_100531.2)
SEQ ID NO:143 полинуклеотид GPAT3 Arabidopsis thaliana (NM_116426.2)
SEQ ID NO:144 полипептид GPAT4 Arabidopsis thaliana (NP 171667.1)
SEQ ID NO:145 полипептид GPAT6 Arabidopsis thaliana (NP 181346.1)
SEQ ID NO:146 полипептид продукта гена F5I10.4 Arabidopsis thaliana (AAF02784.1)
SEQ ID NO:147 полипептид неизвестного белка Arabidopsis thaliana (AAL32544.1)
SEQ ID NO:148 полипептид белка Oryza sativa (AAP03413.1)
SEQ ID NO:149 неизвестный полипептид Picea sitchensis (ABK25381.1)
SEQ ID NO:150 неизвестный полипептид Zea mays (ACN34546.1)
SEQ NO ID:151 полипептид предсказанного белка Arabidopsis thaliana (BAF00762.1)
SEQ ID NO:152 безымянный полипептид белкового продукта Oryza sativa (BAH00933.1)
SEQ ID NO: 153 полипептид предсказанного белка Osl_05566 Oryza sativa (EAY84189.1)
SEQ ID NO: 154 полипептид предсказанного белка Osl_20155 Oryza sativa (EAY98245.1)
SEQ ID NO: 155 полипептид предсказанного белка OsJ_05094 Oryza sativa (EAZ21484.1)
SEQ ID NO: 156 полипептид предсказанного белка Osl_04478 Oryza sativa (EEC71826.1)
SEQ ID NO: 157 полипептид предсказанного белка Osl_13423 Oryza sativa (EEC76137.1)
SEQ ID NO: 158 полипептид предсказанного белка OsJ_12482 Oryza sativa (EEE59882.1)
SEQ ID NO:159 полипептид предсказанного белка SELMODRAFT_269600 Selaginella moellendorffu (EFJ08963.1)
SEQ ID NO:160 полипептид предсказанного белка SELMODRAFT_184962 Selaginella moellendorffu (EFJ08964.1)
SEQ ID NO: 161 полипептид предсказанного белка SELMODRAFT_23533 1 Selaginella moellendorffii (EFJ11200.1)
SEQ ID NO:162 полипептид предсказанного белка SELMODRAFT_118155 Selaginella moellendorffu (EFJ15664.1)
SEQ ID NO:163 полипептид предсказанного белка SELMODRAFT_102257 Selaginella moellendorffu (EFJ24086.1)
SEQ ID NO:164 полипептид предсказанного белка SELMODRAFT_170164 Selaginella moellendorffii (EFJ29 816.1)
SEQ ID NO:165 полипептид предсказанного белка SELMODRAFT_170165 Selaginella moellendorffii (EFJ29817.1)
SEQ ID NO:166 полипептид Os01g0855000 Oryza sativa (NP_001044839.1)
SEQ ID NO:167 полипептид Os02g0114400 Oryza sativa (NP 001045668.1)
SEQ ID NO:168 полипептид GPAT8 Zea mays (NP_001147442.1)
SEQ ID NO:169 полипептид LOCI 00282930 Zea mays (NP_001149307.1)
SEQ ID NO:170 полипептид белка LOC1003 82119 Zea mays (NP_001168351.1)
SEQ ID NO:171 полипептид GPAT6 Arabidopsis thaliana (NP_181346.1)
SEQ ID NO:172 полипептид GPAT8 Arabidopsis thaliana (NP_191950.2)
SEQ ID NO:173 полипептид белка Physcomitrellapatens (XP_001765001.1)
SEQ ID NO:174 полипептид белка Physcomitrella patens (XP_001769671.1)
SEQ ID NO:175 полипептид белка Physcomitrella patens (XP_001769724.1)
SEQ ID NO:176 полипептид белка Physcomitrella patens (XP_001771186.1)
SEQ ID NO:177 полипептид белка Physcomitrella patens (XP_001780533.1)
SEQ ID NO:178 полипептид белка Vitis vinifera (XP_002268513.1)
SEQ ID NO:179 полипептид белка Vitis vinifera (XP_002275348.1)
SEQ ID NO:180 полипептид белка Vitis Vinifera (XP_002276032,1)
SEQ ID NO:181 полипептид белка Vitis Vinifera (XP_002279091.1)
SEQ ID NO:182 полипептид белка Populus trichocarpa (XP_002309124.1)
SEQ ID NO:183 полипептид белка Populus trichocarpa (XP_002309276.1)
SEQ ID NO.184 полипептид белка Populus trichocarpa (XP_002322752.1)
SEQ ID NO:185 полипептид белка Populus trichocarpa (XP_002323563.1)
SEQ ID NO:186 полипептид белка SORBIDRAFT_09g022020 Sorghum bicolor (XP_002439887.1)
SEQ ID NO:187 полипептид белка SORBIDRAFT_03g040260 Sorghum bicolor (XP_002458786.1)
SEQ ID NO:188 полипептид белка SORBIDRAFT_01g008880 Sorghum bicolor (XP_002463916.1)
SEQ ID NO:189 полипептид белка SORBIDRAFT_01g022140 Sorghum bicolor (XP_002464630.1)
SEQ ID NO:190 полипептид ER глицерол-фосфат-ацилтрансферазы Ricinus communis (XP_002511873.1)
SEQ ID NO:191 полипептид ER глицерол-фосфат-ацилтрансферазы Ricinus communis (XP_002 517438.1)
SEQ ID NO:192 полипептид ER глицерол-фосфат-ацилтрансферазы Ricinus communis (XP_002520171.1)
SEQ ID NO:193 полипептид белка семейства фосфолипид/глицерол-ацилтрансферазы Arabidopsis lyrata (XP_002872955.1)
SEQ ID NO:194 полипептид GPAT6 Arabidopsis lyrata (XP_002881564.1)
SEQ ID NO:195 предполагаемый полипептид GPAT Vernicia fordii (ACT32032.1)
SEQ ID NO:196 полипептид Os03g0735900 Oryza sativa (NPOO1051189.1)
SEQ ID NO:197 полипептид GPAT4 Arabidopsis thaliana (NP_171667.1)
SEQ ID NO:198 полипептид белка Populus trichocarpa (XP_002320138.1)
SEQ ID NO:199 полипептид белка SORBIDRAFT_04g001060 Sorghum bicolor (XP_002451377.1)
SEQ ID NO:200 полипептид ER глицерол-фосфат-ацилтрансферазы Ricinus communis (XP_002531350.1)
SEQ ID NO:201 полипептид GPAT4 Arabidopsis lyrata (XP_002889361.1)
SEQ ID NO:202 полипептид GPAT1 Arabidopsis thaliana (NP_563768.1)
SEQ ID NO:203 полипептид GPAT3 Arabidopsis thaliana (NP_192104.1)
SEQ ID NO:204 полинуклеотид DGAT2 Arabidopsis thaliana (NM_115011.3)
SEQ ID NO:205 полинуклеотид DGAT2 Ricinus communis (AY916129.1)
SEQ ID NO:206 полинуклеотид DGAT2 Vernicia fordii (DQ356682.1)
SEQ ID NO:207 полинуклеотид DGAT2 Mortierella ramanniana (AF391089.1)
SEQ ID NO:208 полинуклеотид DGAT2 Homo sapiens (NM_032564.])
SEQ ID NO:209 полинуклеотид DGAT2 Homo sapiens (NM_001013579.2)
SEQ ID NO:210 полинуклеотид DGAT2 Bos taurus (NM_205793.2)
SEQ ID NO:211 полинуклеотид DGAT2 Mus musculus (AF384160.1)
SEQ ID NO:212 полипептид DGAT2 Arabidopsis thaliana (NP_566952.1)
SEQ ID NO:213 полипептид DGAT2 Ricinus communis (AAY 16324.1)
SEQ ID NO:214 полипептид DGAT2 Vernicia fordii (ABC94474.1)
SEQ ID NO:215 полипептид DGAT2 Mortierella ramanniana (AAK84179.1)
SEQ ID NO:216 полипептид DGAT2 Homo sapiens (Q96PD7.2)
SEQ ID NO:217 полипептид DGAT2 Homo sapiens (Q58HT5.1)
SEQ ID NO:218 полипептид DGAT2 Bos taurus (Q70VZ8.1)
SEQ ID NO:219 полипептид DGAT2 Mus musculus (AAK84175.1)
SEQ ID NO:220 трипептид YFP - консервативный мотив последовательности DGAT2 и/или MGAT 1/2
SEQ ID NO:221 тетрапептид HPHG - консервативный мотив последовательности DGAT2 и/или MGAT 1/2
SEQ ID NO:222 тетрапептид EPHS - консервативный мотив последовательности растительного DGAT2
SEQ ID NO:223 RXGFX(K/R)XAXXXGXXX(L/V)VPXXXFG(E/Q) - длинный консервативный мотив последовательности DGAT2, который является частью предполагаемого глицерол-фосфолипидного домена
SEQ ID NO:224 FLXLXXXN - консервативный мотив последовательности мышиных DGAT2 и MGAT 1/2, который представляет собой предполагаемый домен связывания нейтральных липидов
SEQ ID NO:225 домен plsC ацилтрансферазы (PF01553) GPAT
SEQ ID NO:226 домен GPAT суперсемейства HAD-подобных гидролаз (PF12710)
SEQ ID NO:227 Домен фосфосерин-фосфатазы (PF00702). GPAT4-8 содержат N-концевой участок, гомологичный этому домену
SEQ ID NO:228 Консервативная аминокислотная последовательность GDLVICPEGTTCREP GPAT
SEQ ID NO:229 Консервативная аминокислотная последовательность GPAT/фосфатазы (мотив I)
SEQ ID NO:230 Консервативная аминокислотная последовательность GPAT/фосфатазы (мотив III)
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Общие методики и определения
Если конкретно не определено иное, все технические и научные выражения, используемые в данном документе, должны использоваться с такими же значениями, как обычно понятно специалисту в данной области техники (например, в культивировании клеток, молекулярной генетике, иммунологии, иммуногистохимии, химии белков, химии липидов и жирных кислот, получении биотоплива и биохимии).
Если не указано иное, рекомбинантный белок, культура клеток и иммунологические методики, используемые в настоящем изобретении, являются стандартными процедурами, хорошо известными специалистам в данной области техники. Такие методики описаны и объяснены в литературе в источниках, таких как J. Perbat., A Practical Guide to Molecular Cloning, John Wiley and Sons (1984), J. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbour Laboratory Press (1989), T.A. Brown (редакторы), Essential Molecular Biology: A Practical Approach, Volumes 1 and 2, IRL Press (1991), D.M. Glover and B.D. Hames (редакторы), DNA Cloning: A Practical Approach, Volumes 1-4, IRL Press (1995 and 1996), F.M. Ausubel et al. (редакторы), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988, включая все обновленные редакции до настоящего времени), Ed Harlow and David Lane (редакторы) Antibodies: A Laboratory Manual, Cold Spring Harbour Laboratory, (1988), и J.E. Coligan et al. (редакторы) Current Protocols in Immunology, John Wiley & Sons (включая все обновленные редакции до настоящего времени).
Выбранные определения
Выражение "трансгенный, не являющийся человеком организм" относится, например, к целому растению, водоросли, не являющемуся человеком животному или организму, подходящему для ферментации, такому как дрожжи или гриб, содержащему экзогенный полинуклеотид (трансген) или экзогенный полипептид. В варианте осуществления трансгенный, не являющийся человеком организм не является животным или его частью. В одном варианте осуществления трансгенный, не являющийся человеком организм представляет собой фототрофный организм (например, растение или водоросль), способный получать энергию из солнечного света для синтеза органических соединений для питания. В другом варианте осуществления трансгенный, не являющийся человеком организм представляет собой фотосинтетическую бактерию. Выражение "экзогенный" в контексте полинуклеотида или полипептида относится к полинуклеотиду или полипептиду, присутствующему в клетке в измененном количестве по сравнению с его нативным состоянием. В одном варианте осуществления клетка представляет собой клетку, которая в естественном состоянии не содержит данный полинуклеотид или полипептид. В другом варианте осуществления экзогенный полинуклеотид или полипептид принадлежит к другому роду. В другом варианте осуществления экзогенный полинуклеотид или полипептид принадлежит к другому виду. В одном варианте осуществления экзогенный полинуклеотид или полипептид экспрессируется в растении-хозяине или растительной клетке, и экзогенный полинуклеотид или полипептид принадлежит к другому виду или роду. В одном варианте осуществления экзогенный полипептид представляет собой экзогенную MGAT. Как используется в данном документе, выражение "экстрагированный липид" относится к композиции, экстрагированной из трансгенного организма или его части, которая содержит по меньшей мере 60% (вес./вес.) липида.
Как используется в данном документе, выражение "неполярный липид" относится к жирным кислотам и их производным, которые являются растворимыми в органических растворителях, но нерастворимы в воде. Жирные кислоты могут быть свободными жирными кислотами и/или могут находиться в эстерифицированной форме. Примеры эстерифицированных форм включают, но без ограничения, триацилглицерол (TAG), диацилглицерол (DAG), моноацилглицерол (MAG). Неполярные липиды также включают стеролы, сложные эфиры стерола и сложные эфиры воска. Неполярные липиды также известны как "нейтральные липиды". Неполярный липид типично является жидкостью при комнатной температуре. Предпочтительно, неполярный липид преимущественно (>50%) содержит жирные кислоты, которые составляют по меньшей мере 16 углеродов в длину. Более предпочтительно, по меньшей мере 50% общего содержания жирных кислот в неполярном липиде являются C18 жирными кислотами, например, олеиновой кислотой. В варианте осуществления по меньшей мере 50%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 91%, более предпочтительно по меньшей мере 92%, более предпочтительно по меньшей мере 93%, более предпочтительно по меньшей мере 94%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, более предпочтительно по меньшей мере 97%, более предпочтительно по меньшей мере 98%, более предпочтительно по меньшей мере 99% жирных кислот в неполярном липиде по настоящему изобретению могут быть обнаружены в виде TAG. Неполярный липид можно дополнительно очистить или обработать, например, путем гидролиза с помощью сильного основания для высвобождения свободной жирной кислоты или путем фракционирования, дистилляции или подобного. Неполярный липид по настоящему изобретению может образовывать часть "масла семян", если его получают из семян. Неполярный липид может присутствовать в или быть полученным из других частей растения, включая листья или плод, из рекомбинантных клеток или не являющихся человеком организмов, в таком случае липид не является маслом семян, как определено в данном документе.
Концентрации свободного и эстерифицированного стерола (например, ситостерола, кампестерола, стигмастерола, брассикастерола, 05-авенастерола. ситостанола, кампестанола и холестерола) в экстрагированном липиде могут быть такими, как описано в Phillips et al., 2002.
Как используется в данном документе, выражение "масло семян" относится к композиции, полученной из семени/зерна растения, которое содержит по меньшей мере 60% (вес./вес.) липида, или получаемой из семени/зерна, если масло семян все еще присутствует в семени/зерне. Иными словами, масло семян по настоящему изобретению включает масло семян, которое присутствует в семени/зерне или их части, а также масло семян, которое было экстрагировано из семени/зерна. Масло семян предпочтительно представляет собой экстрагированное масло семян. Масло семян типично является жидкостью при комнатной температуре. Предпочтительно, содержание общее содержание жирных кислот (TFA) в масле семян преимущественно (>50%) включает жирные кислоты, которые составляют по меньшей мере 16 углеродов в длину. Более предпочтительно, по меньшей мере 50% общего содержания жирных кислот в масле семян являются C18 жирными кислотами, например, олеиновой кислотой. Жирные кислоты типично находятся в эстерифицированной форме, такой как, например, TAG, DAG, ацил-coA или фосфолипид. Жирные кислоты могут быть свободными жирными кислотами и/или могут находиться в эстерифицированной форме. В варианте осуществления по меньшей мере 50%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 91%, более предпочтительно по меньшей мере 92%, более предпочтительно по меньшей мере 93%, более предпочтительно по меньшей мере 94%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%), более предпочтительно по меньшей мере 97%, более предпочтительно по меньшей мере 98%, более предпочтительно по меньшей мере 99% жирных кислот в масле семян по настоящему изобретению могут быть обнаружены в виде TAG. В варианте осуществления масло семян по настоящему изобретению является "по сути очищенным" или "очищенным" маслом, которое было отделено от одного или нескольких других липидов, нуклеиновых кислот, полипептидов или других загрязняющих молекул, с которыми оно связано в семени или в сыром экстракте. Предпочтительно, что по сути очищенное масло семян является по меньшей мере на 60% свободным, более предпочтительно по меньшей мере на 75% свободным и более предпочтительно по меньшей мере на 90% свободным от других компонентов, с которыми оно связано в семени или экстракте. Масло семян по настоящему изобретению может дополнительно содержать молекулы, не являющиеся жирными кислотами, такие как, без ограничения, стеролы. В варианте осуществления масло семян представляет собой масло канолы (Brassica napus, Brassica rapa ssp.J, масло горчицы (Brassica juncea), масло другого вида Brassica (например, Brassica napobrassica, Brassica camelina), масло подсолнечника (Helianthus annus), масло семян льна (Linum usitatissimum), соевое масло (Glycine max), масло сафлора (Carthamus tinctorius), кукурузное масло (Zea mays), масло табака (Nicotiana tabacum), арахисовое масло (Arachis hypogaea), пальмовое масло (Elaeis guineensis), масло семян хлопка (Gossypium hirsutum), кокосовое масло (Cocos nucifera), масло авокадо (Persea americana), оливковое масло (Olea europaea), масло кешью (Anacardium occidentale), масло макадамии (Macadamia intergrifolia), миндальное масло (Prunus amygdalus), масло семян овса (Avena sativa), рисовое масло (Oryza sativa или Oryza glaberrima) или масло семян Arabidopsis (Arabidopsis thaliana). Масло семян можно экстрагировать из семени/зерна с помощью любого способа, известного в уровне техники. Способ типично включает экстракцию при помощи неполярных растворителей, таких как диэтиловый эфир, петролейный эфир, смеси хлороформа/метанола или бутанола, как правило, связанную с предварительным дроблением семян. Липиды, связанные с крахмалом в зерне, можно экстрагировать с помощью насыщенного водой бутанола. Масло семян можно "дегуммировать" с помощью способов, известных в уровне техники, для удаления полисахаридов или обработать по-другому для удаления загрязнителей или улучшения чистоты, стабильности или цвета. TAG и другие сложные эфиры в масле семян можно гидролизовать для высвобождения свободных жирных кислот, или масло семян гидрогенизировано, обработано химически или ферментативно, как известно в уровне техники.
Как используется в данном документе, выражение "жирная кислота" относится к карбоновой кислоте с длинным алифатическим хвостом по меньшей мере из 8 углеродных атомов в длину, либо насыщенной, либо ненасыщенной. Типично, жирные кислоты имеют цепь с углерод-углеродными связями по меньшей мере из 12 углеродов в длину. Наиболее часто встречающиеся в природе жирные кислоты имеют четное число углеродных атомов, поскольку их биосинтез включает ацетат, который имеет два углеродных атома. Жирные кислоты могут быть в свободном состоянии (неэстерифицированные) или в эстерифицированной форме, такой как часть TAG, DAG, MAG, ацил-coA (тио-сложноэфирной) связи, или в другой ковалентно связанной форме. Будучи ковалентно связанной в эстерифицированной форме, жирная кислота называется в данном документе "ацильной" группой. Жирная кислота может быть эстерифицированной как фосфолипид, такой как фосфатидилхолин (PC), фосфатидилэтаноламин, фосфатидилсерин, фосфатидилглицерол, фосфатидилинозитол или дифосфатидилглицерол. Насыщенные жирные кислоты не содержат каких-либо двойных связей или других функциональных групп вдоль цепи. Выражение "насыщенный" относится к водороду, в котором все углероды (кроме группы карбоновой кислоты [-COOH]) содержат столько водородов, сколько возможно. Другими словами, омега (ω) конец содержит 3 водорода (СН3-), и каждый углерод в пределах цепи содержит 2 водорода (-СН2-). Ненасыщенные жирные кислоты являются формой, подобной насыщенным жирным кислотам, за исключением того, что вдоль цепи существуют одна или несколько алкеновых функциональных групп, причем каждый алкен замещает связанную одинарной связью часть "-СН2-СН2-" цепи связанной двойной связью частью "-CH=СН-" (иными словами, углерод, связан двойной связью с другим углеродом). Два следующих углеродных атома в цепи, которые связаны с обеих сторон от двойной связи, могут находится в цис или транс конфигурации.
Как используется в данном документе, выражения "полиненасыщенная жирная кислота" или "PUFA" относятся к жирной кислоте, которая содержит по меньшей мере 12 углеродных атомов в своей углеродной цепи и по меньшей мере две алкеновые группы (углерод-углеродные двойные связи).
"Моноацилглицерид" или "MAG" представляет собой глицерид, в котором глицерол эстерифицирован одной жирной кислотой. Как используется в данном документе, MAG содержит гидроксильную группу в sn-1/3 (также называемый в данном документе sn-1 MAG или 1-MAG, или 1/3-MAG) или sn-2 положении (также называемый в данном документе 2-MAG) и, следовательно, MAG не включает фосфорилированные молекулы, такие как PA или PC. MAG, таким образом, является компонентом нейтральных липидов в клетке.
"Диацилглицерид" или "DAG" представляет собой глицерид, в котором глицерол эстерифицирован двумя жирными кислотами. Как используется в данном документе, DAG содержит гидроксильную группу в sn-l,3 или от-1,2/2,3 положении и, следовательно, DAG не включает фосфорилированные молекулы, такие как РА или PC. DAG, таким образом, является компонентом нейтральных липидов в клетке. В пути Кеннеди синтеза DAG (фигура 1), предшественник от-глицерол-3-фосфат (G-3-P) эстерифицируется двумя ацильными группами, каждая из которых происходит из сложного эфира жирной кислоты и коэнзима A, в первой реакции, катализированной глицерол-3-фосфат-ацилтрансферазой (GPAT), в положении sn-1 для образования LysoPA, с последующим вторым ацилированием в положении sn-2, катализированной ацилтрансферазой лизофосфатидной кислоты (LPAAT) для образования фосфатидной кислоты (РА). Это промежуточное соединение затем дефосфорилируется для образования DAG. В альтернативном анаболическом пути (фигура 1) DAG может быть образован путем ацилирования либо sn-1 MAG, либо предпочтительно sn-2 MAG, катализированного MGAT. DAG также может быть образован из TAG путем удаления ацильной группы с помощью липазы, или из PC по сути путем удаления холиновой головки с помощью любого из ферментов СРТ, PDCT или PLC (фигура 1).
"Триацилглицерид" или "TAG" представляет собой глицерид, в котором глицерол эстерифицирован тремя жирными кислотами. В пути Кеннеди синтеза TAG DAG образуется, как описано выше, и затем третья ацильная группа эстерифицируется с глицерольным скелетом с помощью активности DGAT. Альтернативные пути для образования TAG включают таковой, катализируемый с помощью фермента PDAT, и MGAT путь, описанный в данном документе.
Как используется в данном документе, выражение "ацилтрансфераза" относится к белку, который способен переносить ацильную группу с ацил-coA на субстрат и включает MGAT, GPAT и DGAT.
Как используется в данном документе, выражение "моноацилглицерол-ацилтрансфераза" или "MGAT" относится к белку, который переносит ацильную группу жирной кислоты от ацил-coA на субстрат MAG для получения DAG. Таким образом, выражение "моноацилглицерол-ацилтрансферазная активность" по меньшей мере относится к переносу ацильной группы от ацил-coA на MAG для получения DAG. MGAT является наиболее известным в отношении его роли во всасывании жира в кишечнике млекопитающих, где жирные кислоты и лтг-2-MAG, образованные в результате расщепления пищевого жира, ресинтезируются до TAG в энтероцитах для синтеза и секреции хиломикронов. MGAT катализирует первый этап этого процесса, в котором ацильная группа из ацил-coA жирной кислоты, образованного из жирных кислот и coA, и sn-2-MAG связываются ковалентно. Выражение "MGAT", как используется в данном документе, включает ферменты, которые действуют на субстраты sn-l/3-MAG и/или sn-2-MAG для образования 5«-l,3-DAG и/или sn-1,2/2,3-DAG, соответственно. В предпочтительном варианте осуществления MGAT является предпочтительным для субстрата 5W-2-MAG по сравнению с sn-1-MAG, или по сути использует только sn-2 MAG в качестве субстрата (примеры включают MGAT, описанные в Cao et al., 2003 (специфичность мышиного MGAT1 для sn2-18:1-MAG>sn1/3-18:l-MAG (фигура 5)); Yen and Farese, 2003 (общие активности мышиного MGAT 1 и MGAT2 человека выше на 2-MAG, чем на ацил-акцепторных субстратах 1-MAG (фигура 5); и Cheng et al., 2003 (активность MGAT3 человека на 2-MAG намного выше, чем на субстратах 1/3-MAG (фигура 2D)).
Как используется в данном документе, MGAT не включает ферменты, которые переносят ацильную группу предпочтительно на LysoPA по сравнению с MAG, такие ферменты известны как LPAAT. Иными словами, MGAT предпочтительно использует нефосфорилированные моноацильные субстраты, несмотря на то, что они могут иметь низкую каталитическую активность на LysoPA. Предпочтительная MGAT не имеет обнаруживаемой активности в ацилировании LysoPA. Как показано в данном документе, MGAT (т.е., MGAT2 М.musculus) также может иметь функцию DGAT, но преимущественно функционирует как MGAT, т.е., она имеет большую каталитическую активность в качестве MGAT, чем в качестве DGAT, если активность фермента выражается в единицах нмоль продукта в минуту на мг белка (также смотри Yen et al., 2002).
Известно три класса MGAT, которые называются MGAT1, MGAT2 и MGAT3, соответственно. Гомологи гена MGAT1 человека (AF384163) присутствуют (т.е. известны последовательности) по меньшей мере у шимпанзе, собаки, коровы, мыши, крысы, данио, Caenorhabditis elegans, Schizosaccharomyces pombe, Saccharomyces cerevisiae, Kluyveromyces lactis, Eremothecium gossypii, Magnaporthe grisea и Neurospora crassa. Гомологи гена MGAT2 человека (AY 157608) присутствуют по меньшей мере у шимпанзе, собаки, коровы, мыши, крысы, курицы, данио, дрозофилы и комара. Гомологи гена MGAT3 человека (AY229854) присутствуют по меньшей мере у шимпанзе, собаки, коровы и данио. Однако гомологи из других организмов могут быть легко определены с помощью способов, известных в уровне техники, для определения гомологичных последовательностей.
Примеры полипептидов MGAT1 включают белки, кодируемые генами MGAT1 из Homo sapiens (AF384163), Mus musculus (AF384162), Pan troglodytes (XM_001166055, XM_0526044.2), Canis familiaris (XM 545667.2), Bos taurus (NM_001001153.2), Rattus norvegicus (NM_001108803.1), MGAT1 Danio rerio (NM_001122623.1), Caenorhabditis elegans (NM_073012.4, NM_182380.5, NM_065258.3, NM_075068.3, NM_072248.3), Kluyveromyces lactis (XM_455588.1), Ashbya gossypii (NM 208895.1), Magnaporthe oryzae (XM_3 68741.1), предсказанным Ciona intestinalis (XM_002120843,1). Примеры полипептидов MGAT2 включают белки, кодируемые генами MGAT2 из Homo sapiens (AY157608), Mus musculus (AY157609), Pan troglodytes (XM_522112.2), Canis familiaris (XM_542304.1), Bos taurus (NM 001099136.1), Rattus norvegicus, Gallus gallus (XM 424082.2), Danio rerio (NM 001006083.1), Drosophila melanogaster (NM_ 136474.2, NMJ36473.2, NM_136475.2), Anopheles gambiae (XM_001688709.1, XM_315985), Tribolium castaneum (XM 970053.1). Примеры полипептидов MGAT3 включают белки, кодируемые генами MGAT3 из Homo sapiens (AY229854), Pan troglodytes (XM 001154107.1, XM_001154171.1, XM_527842.2), Canis familiaris (XM_845212.1), Bos taurus (XM 870406.4), Danio rerio (XM_688413.4).
Как используется в данном документе "MGAT путь" относится к анаболическому пути, отличному от пути Кеннеди, для образования TAG, в котором DAG образуется путем ацилирования либо.«7-1-MAG, либо предпочтительно sn-2-MAG, катализированного MGAT. DAG впоследствии может быть использован для образования TAG или других липидов. MGAT путь проиллюстрирован на Фигуре 1.
Как используется в данном документе, выражение "диацилглицерол-ацилтрансфераза" (DGAT) относится к белку, который переносит ацильную группу жирной кислоты от ацил-coA на субстрат DAG для получения TAG. Таким образом, выражение "диацилглицерол-ацилтрансферазная активность" относится к переносу ацильной группы от ацил-coA на DAG для получения TAG. DGAT также может иметь функцию MGAT, но преимущественно функционирует в качестве DGAT, т.е., она имеет большую каталитическую активность в качестве DGAT, чем в качестве MGAT, если активность фермента выражается в единицах нмоль продукта в минуту на мг белка (смотри, например, Yen et al., 2005).
Известно три типа DGAT, которые имеют название DGAT1, DGAT2 и DGAT3, соответственно. Полипептиды DGAT1 типично имеют 10 трансмембранных доменов, полипептиды DGAT2 типично имеют 2 трансмембранных домена, тогда как полипептиды DGAT3 типично не имеют ни одного и считаются растворимыми в цитоплазме, не будучи интегрированными в мембранах. Примеры полипептидов DGAT1 включают белки, кодируемые генами DGAT1 из Aspergillus fumigatus (№доступа ХР_755172), Arabidopsis thaliana (САВ44774), Ricinus communis (AAR11479), Vernicia fordii (ABC94472), Vernonia galamensis (ABV21945, ABV21946), Euonymus alatus (AAV31083), Caenorhabditis elegans (AAF82410), Rattus norvegicus (NP445889), Homo sapiens (NP_036211), а также их варианты и/или мутанты. Примеры полипептидов DGAT2 включают белки, кодируемые генами DGAT2 из Arabidopsis thaliana (NP 566952.1; SEQ ID NO:212), Ricinus communis (AAY 16324.1; SEQ ID NO:213), Vernicia fordii (ABC94474.1; SEQ ID NO:214), Mortierella ramanniana (AAK84179.1; SEQ ID NO:215), Homo sapiens (Q96PD7.2; SEQ ID NO:216) (Q58HT5.1; SEQ ID NO:217), Bos taurus (Q70VZ8.1; SEQ ID NO:218), Mus musculus (AAK84175.1; SEQ ID NO:219), а также их варианты и/или мутанты.
Примеры полипептидов DGAT3 включают белки, кодируемые генами DGAT3 из арахиса (Arachis hypogaea, Saha, et al., 2006), а также их варианты и/или мутанты. DGAT имеет небольшую или не обнаруживаемую MGAT активность, например, менее 300 пмоль в минуту на мг белка, предпочтительно менее 200 пмоль в минуту на мг белка, более предпочтительно 100 пмоль в минуту на мг белка.
DGAT2, но не DGAT1 имеет высокую гомологию последовательностей с ферментами MGAT, позволяя предположить, что гены DGAT2 и MGAT, вероятно, имеют общее генетическое происхождение. Хотя многочисленные изоформы включены в катализ одного и того же этапа в синтезе TAG, они могут играть различные функциональные роли, что было предположено на основании неравномерного распределения в тканях и субклеточной локализации семейства ферментов DGAT/MGAT. У млекопитающих MGAT1 в основном экспрессируется в желудке, почке, жировой ткани, тогда как MGAT2 и MGAT3 демонстрируют самую высокую экспрессию в тонком кишечнике. У млекопитающих DGAT1 повсеместно экспрессируется во многих тканях, с самой высокой экспрессией в тонком кишечнике, тогда как DGAT2 наиболее распространена в печени. Из биоинформатического анализа известно, что MGAT3 существует только у высших млекопитающих и людей, но не у грызунов. MGAT3 имеет более высокую гомологию последовательности с DGAT2, чем с MGAT1 и MGAT3. MGAT3 проявляет существенно более высокую DGAT активность, чем ферменты MGAT1 и MGAT2 (MGAT3>MGAT1>MGAT2), если либо MAG, либо DAG используются в качестве субстратов, позволяя предположить, что MGAT3 функционирует как предполагаемая TAG-синтаза.
Как MGAT1, так и MGAT2 принадлежат к одному и тому же классу ацилтрансфераз, что и DGAT2. Некоторые из мотивов, которые, как было показано, являются важными для каталитической активности DGAT2 в некоторых DGAT2, также являются консервативными в ацилтрансферазах MGAT. Особый интерес представляет предполагаемый связывающий нейтральные липиды домен с консенсусной последовательностью FLXLXXXN (SEQ ID NO:224), где каждый X независимо представляет собой любую аминокислоту, отличную от пролина, и N представляет собой любую неполярную аминокислоту, расположенную в пределах N-концевого трансмембранного участка, за которым следует предполагаемый домен глицерол/фосфолипид-ацилтрансферазы. Мотив FLXLXXXN обнаруживается в мышиной DGAT2 (аминокислоты 81-88) и MGAT 1/2, но не в DGAT2 дрожжей или растений. Он является важным для активности мышиной DGAT2. Другие мотивы последовательностей DGAT2 и/или MGAT 1/2 включают:
1. Высоко консервативный трипептид YFP (SEQ ID NO:220) в большинстве полипептидов DGAT2, а также в MGAT1 и MGAT2, например, присутствующий как аминокислоты 139-141 в мышиной DGAT2. Мутирование этого мотива в DGAT2 дрожжей при помощи неконсервативных замен делало фермент нефункциональным.
2. Тетрапептид HPHG (SEQ ID NO:221), высоко консервативный в MGAT, а также в последовательностях DGAT2 из животных и грибов, например, присутствующий как аминокислоты 161-164 в мышиной DGAT2, и являющийся важным для каталитической активности по меньшей мере в DGAT2 дрожжей и мышиной DGAT2. Растительные ацилтрансферазы DGAT2 вместо этого имеют консервативную последовательность EPHS (SEQ ID NO:222), поэтому консервативные замены первой и четвертой аминокислот могут быть допустимыми.
3. Более длинный консервативный мотив, который является частью предполагаемого глицерол-фосфолипидного домена. Пример этого мотива представляет собой RXGFX(K/R)XAXXXGXXX(L/V)VPXXXFG(E/Q) (SEQ ID NO:223), который присутствует как аминокислоты 304-327 в мышиной DGAT2. Этот мотив является менее консервативным в аминокислотной последовательности, чем другие, как можно было бы предположить из его длины, но гомологи могут быть распознаны с помощью поиска мотивов. Интервал между более консервативными аминокислотами может варьировать, т.е., могут быть добавочные X аминокислоты в пределах мотива или может быть меньше X аминокислот по сравнению с последовательностью выше.
Как используется в данном документе, выражение "глицерол-3-фосфат-ацилтрансфераза" или "GPAT" относится к белку, который ацилирует глицерол-3-фосфат (G-3-P) для образования LysoPA и/или MAG, причем последний продукт образуется, если GPAT также имеет фосфатазную активность на LysoPA. Ацильная группа, которая переносится, типично происходит от ацил-СоА. Таким образом, выражение "глицерол-3-фосфат-ацилтрансферазная активность" относится к ацилированию G-3-P для образования LysoPA и/или MAG. Выражение "GPAT" включает ферменты, которые ацилируют G-3-P для образования sn-1-LPA и/или sn-2-LPA, предпочтительно sn-2-LPA. В предпочтительном варианте осуществления GPAT имеет фосфатазную активность. В наиболее предпочтительном варианте осуществления GPAT представляет собой sn-2-GPAT с фосфатазной активностью, которая образует ли-2-MAG.
Как используется в данном документе, выражение "sn-1-глицерол-3-фосфат-ацилтрансфераза" (sn-1 GPAT) относится к белку, который ацилирует sn-1-гицерол-3-фосфат (G-3-P) для предпочтительного образования 1-ацил-sn-глицерол-3-фосфата (sn-1-LPA). Таким образом, выражение "sn-1-глицерол-3-фосфат-ацилтрансферазная активность" относится к ацилированию sn-глицерол-3-фосфата для образования 1-ацил- sn-глицерол-3-фосфата (sn-1-LPA).
Как используется в данном документе, выражение "sn-2-глицерол-3-фосфат-ацилтрансфераза" sn-2-GPAT) относится к белку, который ацилирует sn-глицерол-3-фосфат (G-3-P) для предпочтительного образования 2-ацил-sn-глицерол-3-фосфата (sn-2-LPA). Таким образом, выражение "sn-2-глицерол-3-фосфат-ацилтрансферазная активность" относится к ацилированию от-глицерол-3-фосфата для образования 2-ацил-sn-глицерол-3-фосфата (sn-2-LPA).
Семейство GPAT является большим, и все известные представители содержат два консервативных домена, домен plsC ацилтрансферазы (PF01553; SEQ ID NO:225) и домен суперсемейства HAD-подобной гидролазы (PF12710; SEQ ID NO:226). В дополнение к этому в Arabidopsis thaliana, все GPAT4-8 содержат N-концевой участок, гомологичный домену фосфосерин-фосфатазы (PF00702; SEQ ID NO:227). Как GPAT4, так и GPAT6 содержат консервативные остатки, которые, как известно, являются ключевыми для фосфатазной активности, в частности, консервативные аминокислоты (показанные жирным шрифтом) в мотиве I (DXDX[T/V][L/V]; SEQ ID NO:229) и мотиве III (K-[G/S][D/S]XXX[D/N]; SEQ ID NO:330), расположенном на N-конце (Yang et al., 2010). Предпочтительно, GPAT имеет предпочтение sn-2 и фосфатазную активность для получения sn-2-MAG (также называемого в данном документе "2-MAG") из глицерол-3-фосфата (G-3-P) (фигура 1), например, (GPAT4 (NP 171667.1) и GPAT6 (NP_181346.1)) из Arabidopsis. Более предпочтительно, GPAT использует ацил-coAв качестве субстрата жирной кислоты.
Гомологи GPAT4 (NP_171667) и GPAT6 (NPJ 81346) включают AAF02784, AAL32544, ААР03413, АВК25381, ACN34546, BAF00762, ВАН00933, EAY84189, EAY98245, EAZ21484, ЕЕС71826, ЕЕС76137, ЕЕЕ59882, EFJ08963, EFJ08964, EFJ11200, EFJ15664, EFJ24086, EFJ29816, EFJ29817, NP_001044839, NP_001045668, NP_01147442, NP_001149307, NP_001168351, NP_81346, NP_191950, XP_001765001, XP_001769671, XP_001769724, XP_00771186, XP_001780533, XP_002268513, XP_002275348, XP_002276032, XP_002279091, XP_002309124, XP_002309276, XP_002322752, XP_002323563, XP_002439887, XP_002458786, XP_002463916, XP_002464630, XP_002511873, XP_002517438, XP_002520171, XP_002872955, XP_002881564, ACT32032, ACT32032, NP_001051189, NP_171667, XP_002320138, XP_002451377, XP_002451377, XP_002531350, XP_002872955 и XP_002889361.
Консервативные мотивы и/или остатки могут использоваться как диагностические средства на основе последовательности для определения бифункциональных GPAT/фосфатазных ферментов. Альтернативно, можно использовать более строгий анализ на основе функции. Такой анализ включает, например, введение меченого глицерол-3-фосфата в клетки или микросомы и количественное определение уровней меченых продуктов с помощью тонкослойной хроматографии или подобной методики. GPAT активность приводит к образованию меченого LPA, тогда как GPAT/фосфатазная активность приводит к образованию меченого MAG.
Как используется в данном документе, выражение "дикий тип" или его варианты относится к клетке или не являющемуся человеком организму или его части, которая не была генетически модифицирована.
Выражение "соответствующий" относится к клетке или не являющемуся человеком организму или его части, которая имеет одинаковую или сходную генетическую среду, как и клетка или не являющийся человеком организм или его часть по настоящему изобретению, но которая не была модифицирована, как описано в данном документе (например, клетка или не являющийся человеком организм или его часть, в которой отсутствует экзогенный полинуклеотид, кодирующий MGAT или экзогенный MGAT). Соответствующая клетка или не являющийся человеком организм или его часть может использоваться как контроль для сравнения уровней нуклеиновой кислоты или экспрессии белка, или степени и природы модификации признака, например, образования и/или содержания неполярного липида, с клеткой или не являющимся человеком организмом или его частью, модифицированной, как описано в данном документе.
Как используется в данном документе, "по сравнению с" относится к сравнению уровней неполярного липида или общего содержания неполярных липидов трансгенного, не являющегося человеком организма или его части, экспрессирующей один или несколько экзогенных полинуклеотидов или экзогенных полипептидов, с трансгенным, не являющимся человеком организмом или его частью, где отсутствует один или несколько экзогенных полинуклеотидов или полипептидов.
Как используется в данном документе, "усиленная способность к образованию неполярного липида" представляет собой относительное выражение, которое относится к общему количеству неполярного липид, образованного клеткой или не являющимся человеком организмом или его частью по настоящему изобретению, которое увеличено по сравнению с соответствующей клеткой или не являющимся человеком организмом или его частью. В одном варианте осуществления содержание TAG и/или полиненасыщенной жирной кислоты неполярного липида увеличивается.
Как используется в данном документе, выражение "выделенный или рекомбинантный полинуклеотид, который отрицательно регулирует образование и/или активность эндогенного фермента" или его варианты относится к полинуклеотиду, который кодирует молекулу РНК, которая отрицательно регулирует образование и/или активность (например, кодирует миРНК) или сам по себе отрицательно регулирует образование и/или активность (например, представляет собой миРНК, которая может быть доставлена прямо, например, к клетке) эндогенного фермента, например, DGAT, sn-1-глицерол-3-фосфат-ацилтрансферазы (GPAT), 1-ацил-глицерол-3-фосфат-ацилтрансферазы (LPAAT), ацил-СоА:лизофосфатидилхолин-ацилтрансферазы (LPCAT), фосфатазы фосфатидной кислоты (PAP) или комбинации двух или более из них.
Как используется в данном документе, выражение "на основе веса" относится к весу вещества (например, TAG, DAG, жирной кислоты) в виде процентного отношения от веса композиции, содержащей вещество (например, семя, лист). Например, если трансгенное семя имеет 25 мкг общего содержания жирной кислоты на 120 мкг веса семени, то процентное отношение общего содержания жирной кислоты на основе веса составляет 20,8%.
Как используется в данном документе, выражение "на относительной основе" относится к количеству вещества в композиции, содержащей вещество, по сравнению с соответствующей композицией в виде процентного отношения.
Как используется в данном документе, выражение "относительное нелипидное содержание" относится к выражению содержания неполярных липидов клетки, организма или его части, или экстрагированного из них липида, по сравнению с соответствующей клеткой, организмом или его частью, или липидом, экстрагированным из соответствующей клетки, организма или его части, в виде процентного отношения. Например, если трансгенное семя имеет 25 мкг общего содержания жирной кислоты, тогда как соответствующее семя имело 20 мкг общего содержания жирной кислоты, то увеличение в содержании неполярных липидов на относительной основе равняется 25%.
Образование диацилглицеролов и триацилглицеролов
В одном варианте осуществления трансгенный, не являющийся человеком организм или его часть по настоящему изобретению образует более высокие уровни неполярных липидов, таких как DAG или TAG, предпочтительно обоих, чем у соответствующего не являющегося человеком организма или его части. В одном примере трансгенные растения по настоящему изобретению образуют семена и/или листья с повышенным содержанием неполярных липидов, таких как DAG или TAG, предпочтительно обоих, по сравнению с соответствующими семенами и/или листьями. Содержание неполярных липидов не являющегося человеком организма или его части составляет на 0,5% больше на основе веса по сравнению с соответствующим не являющимся человеком организмом или его частью.
В другом варианте осуществления трансгенный, не являющийся человеком организм или его часть, предпочтительно растение или семя, образуют DAG и/или TAG, которые обогащены одной или несколькими конкретными жирными кислотами. Широкий спектр жирных кислот может быть включен в DAG и/или TAG, включая насыщенные и ненасыщенные жирные кислоты, а также короткоцепочечные и длинноцепочечные жирные кислоты. Некоторые неограничивающие примеры жирных кислот, которые могут быть включены в DAG и/или TAG, включают следующее: каприновую (10:0), лауриновую (12:0), миристиновую (14:0), пальмитиновую (16:0), пальмитолеиновую (16:1), стеариновую (18:0), олеиновую (18:1), вакценовую (18:1), линоленовую (18:2), элеостеариновую (18:3), γ-линоленовую (18:3), α-линоленовую (18:3ω3), стеаридоновую (18:4ω3), арахиновую (20:0), эйкозадиеновую (20:2), дигомо-γ-линолевую (20:3), эйкозатриеновую (20:3), арахидоновую (20:4), эйкозатетраеновую (20:4), эйкозапентаеновую (20:5ω3), бегеновую (22:0), докозапентаеновую (22:5ω), докозагексаеновую (22:6ω3), лигноцериновую (24:0), нервоновую (24:1), церотиновую (26:0) и монтановую (28:0) жирные кислоты. В одном варианте осуществления настоящего изобретения трансгенный организм или его части обогащены DAG и/или TAG, содержащими полиненасыщенные жирные кислоты.
В одном варианте осуществления настоящего изобретения трансгенный, не являющийся человеком организм или его часть, предпочтительно растение или семя, трансформировано при помощи химерной ДНК, которая кодирует MGAT, как определено в данном документе, которая может иметь или не иметь DGAT активность. Экспрессия MGAT предпочтительно приводит к более высоким уровням неполярных липидов, таких как DAG или TAG и/или увеличенному выходу неполярных липидов в указанном трансгенном, не являющемся человеком организме или его части. В предпочтительном варианте осуществления трансгенный, не являющийся человеком организм представляет собой растение.
В еще одном варианте осуществления трансгенный, не являющийся человеком организм или его часть трансформирован при помощи химерной ДНК, которая кодирует GPAT или DGAT. Предпочтительно, организм трансформирован при помощи обеих химерных ДНК, которые предпочтительно ковалентно связаны с одной молекулой ДНК, такой как, например, отдельная молекула Т-ДНК.
Yang et al. (2010) описывают две глицерол-3-фосфат-ацилтрансферазы (GPAT4 и GPAT6) из Arabidopsis с предпочтением sn-2 и фосфатазной активностью, которые способны к образованию sn-2-MAG из глицерол-3-фосфата (G-3-P) (фигура 1). Эти ферменты, предположительно, являются частью пути синтеза кутина. Было показано, что GPAT4 и GPAT6 Arabidopsis, используют ацил-coAв качестве субстрата жирной кислоты (Zheng et al., 2003).
Комбинирование бифункциональной GPAT/фосфатазы с MGAT дает на выходе новый путь синтеза DAG с использованием G-3-P в качестве одного субстрата и двух ацильных групп, полученных из ацил-CoA, в качестве других субстратов. Аналогично, комбинирование такой бифункциональной GPAT/фосфатазы с MGAT, которая имеет DGAT активность, дает на выходе новый путь синтеза TAG с использованием глицерол-3-фосфата в качестве одного субстрата и трех ацильных групп, полученные из ацил-СоА, в качестве других субстратов.
Соответственно, в одном варианте осуществления настоящего изобретения трансгенный, не являющийся человеком организм или его часть является котрансформированной при помощи бифункциональной GPAT/фосфатазы и при помощи MGAT, которая не имеет DGAT активности. Это будет приводить к образованию MAG с помощью бифункциональной GPAT/фосфатазы, которая затем будет превращаться в DAG с помощью MGAT и затем в TAG с помощью нативной DGAT или другой активности. Образование DAG могло подтвердиться и быть выбранным с помощью, например, осуществления такой котрансформации в штамме дрожжей, содержащем летальные нокауты SLC1+SLC4, такие как те, которые описаны Benghezal et al. (2007; фигура 2). На фигуре 2 Benghezal et al. (2007) показано, что нокаутирование двух LPATS дрожжей (SLC1 и SLC4) является летальным. Двойной мутант дрожжей SLC1+SLC4 может поддерживаться только благодаря дополнительной плазмиде, которая обеспечивает один из этих генов sic (SLC1 в их случае) в транс-положении. Отрицательная селекция путем добавления FOA в среду приводит к потере этой дополнительной плазмиды (контрселекция маркера селекции Ura) и делает клетки нежизнеспособными.
В другом варианте осуществления настоящего изобретения трансгенный, не являющийся человеком организм или его часть, предпочтительно растение или семя, является ^трансформированным при помощи химерных ДНК, кодирующих бифункциональную GPAT/фосфатазу и MGAT, которая имеет DGAT активность. Это будет приводить к образованию MAG с помощью бифункциональной GPAT/фосфатазы, который затем будет превращаться в DAG и затем в TAG с помощью MGAT.
В еще одном варианте осуществления одна или несколько эндогенных GPAT без обнаруживаемой фосфатазной активности подвергаются сайленсингу, например, один или несколько генов, кодирующих GPAT, которые ацилируют глицерол-3-фосфат для образования LPA в пути Кеннеди (например, GPAT1 Arabidopsis) подвергается сайленсингу.
Субстратные предпочтения могут быть сконструированы в новые пути синтеза DAG и TAG с помощью, например, обеспечения трансгенных штаммов дрожжей Н1246, экспрессирующих варианты MGAT с концентрацией конкретной свободной жирной кислоты (например, DHA), что предотвращает комплементацию с помощью гена MGAT дикого типа. Только варианты, которые способны использовать обеспеченную свободную жирную кислоту, будут расти. Несколько циклов инженерии MGAT будут приводить к получению MGAT с увеличенным предпочтением в отношении конкретных жирных кислот.
Различные дополнения и добавления к пути Кеннеди, описанные выше, могут быть осуществлены в любом типе клеток благодаря повсеместно распространенной природе начального субстрата глицерол-3-фосфата. В одном варианте осуществления применение трансгенов приводит к увеличенным выходам масла.
Полинуклеотиды
Выражения "полинуклеотид" и "нуклеиновая кислота" используют взаимозаменяемо. Они относятся к полимерной форме нуклеотидов любой длины, либо дезоксирибонуклеотидам, либо рибонуклеотидам или их аналогам. Полинуклеотид по настоящему изобретению может быть геномного, кДНК, полусинтетического или синтетического происхождения, двухцепочечным или одноцепочечным и посредством своего происхождения или манипуляции с ним: (1) не связанным со всем или частью полинуклеотида, с которым он связан в природе, (2) соединенным с полинуклеотидом, отличным от того, с которым он связан в природе, или (3) не встречаться в природе. Следующие неограничивающие примеры полинуклеотидов: кодирующие или некодирующие участки гена или фрагмента гена, локусы (локус), определенный с помощью анализа сцепления, экзоны, интроны, матричная РНК (мРНК), транспортная РНК (тРНК), рибосомальная РНК (рРНК), рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенная ДНК любой последовательности, выделенная РНК любой последовательности, химерная ДНК любой последовательности, зонды нуклеиновой кислоты и праймеры. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и аналоги нуклеотидов. Если они присутствуют, модификации нуклеотидной структуры могут быть обеспечены до или после сборки полимера. Последовательность нуклеотидов может быть прервана ненуклеотидными компонентами. Полинуклеотид может быть дополнительно модифицирован после полимеризации, например, с помощью конъюгации с маркирующим компонентом.
Под "выделенным полинуклеотидом" подразумевают полинуклеотид, который был, как правило, отделен от полинуклеотидных последовательностей, с которыми он связан или соединен в его нативном состоянии. Предпочтительно, выделенный полинуклеотид по меньшей мере на 60% свободен, более предпочтительно по меньшей мере на 75% свободен и более предпочтительно по меньшей мере на 90% свободен от полинуклеотидных последовательностей, с которыми он связан или соединен в естественном состоянии.
Как используется в данном документе, выражение "ген" следует использовать в его самом широком контексте, и оно включает дезоксирибонуклеотидные последовательности, содержащие транскрибирующийся участок и, если он транслируется, кодирующий белок участок структурного гена, и включает последовательности, расположенные рядом с кодирующим участком как на 5'-, так и 3'-концах на расстоянии по меньшей мере около 2 т.п.о. на любом конце и вовлеченные в экспрессию гена. В этом отношении, ген включает контрольные сигналы, такие как промоторы, энхансеры, сигналы терминации и/или полиаденилирования, которые в естественном состоянии связаны с данным геном, или гетерологичные контрольные сигналы, причем в этом случае ген называется "химерным геном". Последовательности, которые расположены 5' от кодирующего белок участка и которые присутствуют на мРНК, называются 5'-нетранслируемыми последовательностями. Последовательности, которые расположены 3' или ниже кодирующего белок участка, и которые присутствуют на мРНК, называются 3'-нетранслируемыми последовательностями. Выражение "ген" охватывает как кДНК, так и геномные формы гена. Геномная форма или клон гена содержит кодирующий участок, который может прерываться некодирующими последовательностями, называемыми "интроны", "вставочные участки" или "вставочные последовательности". Интроны представляют собой сегменты гена, которые транскрибируются в ядерную РНК (яРНК). Интроны могут содержать регуляторные элементы, такие как энхансеры. Интроны удаляются или "сплайсируются" из ядерного или первичного транскрипта; интроны, следовательно, отсутствуют в транскрипте мРНК. Во время трансляции мРНК функционирует в отношении определения последовательности или порядка аминокислот в образующемся полипептиде. Выражение "ген" включает синтетическую или слитую молекулу, кодирующую все или часть белков по настоящему изобретению, описанных в данном документе, и комплементарную нуклеотидную последовательность к любой из перечисленных выше.
Как используется в данном документе, "химерная ДНК" относится к любой молекуле ДНК, которая в естественном состоянии не обнаруживается в природе; также называемая в данном документе "ДНК-конструкт". Типично, химерная ДНК содержит регуляторные и транскрибирующиеся или кодирующие белок последовательности, которые в естественном состоянии не обнаруживаются вместе в природе. Соответственно, химерная ДНК может содержать регуляторные последовательности и кодирующие последовательности, которые получены из различных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но расположенные отличным от обнаруживаемого в природе образом. Открытая рамка считывания может быть или не быть связанной с ее естественными регуляторными элементами выше или ниже. Открытая рамка считывания может быть встроена, например, в геном растения, в не встречающейся в природе локализации, или в репликон или вектор, где она не обнаруживается в естественном состоянии, такой как бактериальная плазмида или вирусный вектор. Выражение "химерная ДНК" не ограничивается молекулами ДНК, которые реплицируются в хозяине, но включает ДНК, способную быть лигированной в репликон с помощью, например, специфических адапторных последовательностей.
"Трансген" представляет собой ген, который был введен в геном с помощью процедуры трансформации. Выражения "генетически модифицированный", "трансгенный" и их варианты включают введение гена в клетку с помощью трансформации или трансдукции, мутирования гена в клетке и генетического изменения или модуляции регуляции гена в клетке или потомстве любой клетки, модифицированной, как описано выше.
"Геномный участок", как используется в данном документе, относится к положению в геноме, где трансген или группа трансгенов (также называемая в данном документе кластер), был введен в клетку или ее предшественник. Такие участки содержат только нуклеотиды, которые были встроены с помощью вмешательства человека, например, с помощью способов, описанных в данном документе.
"Рекомбинантный полинуклеотид" по настоящему изобретению относится к молекуле нуклеиновой кислоты, которая была сконструирована или модифицирована с помощью искусственных рекомбинантных способов. Рекомбинантный полинуклеотид может присутствовать в клетке в измененном количестве или экспрессироваться на измененном уровне (например, в случае мРНК) по сравнению с его нативным состоянием. В одном варианте осуществления полинуклеотид вводят в клетку, которая в естественном состоянии не содержит данный полинуклеотид. Типично экзогенная ДНК используется в качестве матрицы для транскрипции мРНК, которая затем транслируется в непрерывную последовательность аминокислотных остатков, кодирующих полипептид по настоящему изобретению в трансформированной клетке. В другом варианте осуществления полинуклеотид является эндогенным по отношению к клетке, и его экспрессия изменяется с помощью рекомбинантных средств, например, экзогенную контрольную последовательность вводят выше эндогенного гена интереса, чтобы обеспечить экспрессию трансформированной клеткой полипептида, кодируемого геном.
Рекомбинантный полинуклеотид по настоящему изобретению включает полинуклеотиды, которые не были отделены от других компонентов клеточной или бесклеточной системы экспрессии, в которой он присутствует, и полинуклеотиды, полученные в указанных клеточных или бесклеточных системах, которые впоследствии очищаются от по меньшей мере некоторых других компонентов. Полинуклеотид может представлять собой непрерывный фрагмент из нуклеотидов, существующий в природе, или содержать два или более непрерывных фрагмента из нуклеотидов из различных источников (встречающиеся в природе и/или синтетические), соединенные для образования одного полинуклеотида. Типично, такие химерные полинуклеотиды содержат по меньшей мере открытую рамку считывания, кодирующую полипептид по настоящему изобретению, функционально связанную с промотором, подходящим для управления транскрипцией открытой рамки считывания в клетке интереса.
В отношении определенных полинуклеотидов следует понимать, что количественные показатели % идентичности, более высокие чем те, которые обеспечены выше, будут включать предпочтительные варианты осуществления. Таким образом, где применимо, с учетом минимальных количественных показателей % идентичности, предпочтительным является, что полинуклеотид включает полинуклеотидную последовательность, которая по меньшей мере на 60%, более предпочтительно по меньшей мере на 65%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 91%, более предпочтительно по меньшей мере на 92%, более предпочтительно по меньшей мере на 93%, более предпочтительно по меньшей мере на 94%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 96%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%, более предпочтительно по меньшей мере на 99,1%, более предпочтительно по меньшей мере на 99,2%, более предпочтительно по меньшей мере на 99,3%, более предпочтительно по меньшей мере на 99,4%, более предпочтительно по меньшей мере на 99,5%, более предпочтительно по меньшей мере на 99,6%, более предпочтительно по меньшей мере на 99,7%, более предпочтительно по меньшей мере на 99,8% и даже более предпочтительно по меньшей мере на 99,9% идентична соответственной предлагаемой SEQ ID NO.
Полинуклеотид по настоящему изобретению или применимый для настоящего изобретения может селективно гибридизоваться, при жестких условиях, с полинуклеотидом, определенным в данном документе. Как используется в данном документе, жесткие условия являются такими, при которых: (1) используют во время гибридизации денатурирующее средство, такое как формамид, например, 50% (об./об.) формамид с 0,1% (вес./об.) бычьим сывороточным альбумином, 0,1% Ficoll, 0,1% поливинилпирролидоном, 50 мМ натрий-фосфатным буфером при рН 6,5 с 750 мМ NaCl, 75 мМ цитрата натрия при 42°C; или (2) используют 50% формамид, 5×SSC (0,75 М NaCl, 0,075 М цитрата натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5× раствор Денхардта, обработанную ультразвуком ДНК из молок лососевых (50 г/мл), 0,1% SDS и 10% сульфат декстрана при 42°C в 0,2×SSC и 0,1% SDS, и/или (3) используют низкую ионную силу и высокую температуру для промывки, например, 0,015 М NaCl/0,0015 М цитрат натрия/0,1% SDS при 50°C.
Полинуклеотиды по настоящему изобретению могут обладать, по сравнению с встречающимися в природе молекулами, одной или несколькими мутациями, которые представляют собой делеции, инсерции или замены нуклеотидных остатков. Полинуклеотиды, которые имеют мутации по сравнению с эталонной последовательностью, могут быть либо встречающимися в природе (иными словами, выделенными из естественного источника), либо синтетическими (например, путем осуществления сайт-направленного мутагенеза или ДНК-шаффлинга на нуклеиновой кислоте, как описано выше).
Рекомбинантные векторы
Один вариант осуществления настоящего изобретения включает рекомбинантный вектор, который содержит по меньшей мере один полинуклеотид, определенный в данном документе, и способен доставлять полинуклеотид в клетку-хозяина. Рекомбинантные векторы включают векторы экспрессии. Рекомбинантные векторы содержат гетерологичные полинуклеотидные последовательности, иными словами, полинуклеотидные последовательности, которые не обнаруживаются в природе рядом с полинуклеотидом, определенным в данном документе, которые предпочтительно получены из различных видов. Вектор может представлять собой либо РНК, либо ДНК, либо для прокариот, либо для эукариот, и типично представляет собой вирусный вектор, полученный из вируса, или плазмиду. Плазмидные векторы типично включают дополнительные последовательности нуклеиновой кислоты, которые обеспечивают легкую селекцию, амплификацию и трансформацию кассеты экспрессии в прокариотических клетках, например, полученные из pUC векторы, полученные из pSK векторы, полученные из pGEM векторы, полученные из pSP векторы, полученные из pBS векторы или бинарные векторы, содержащие один или несколько участков Т-ДНК. Дополнительные последовательности нуклеиновой кислоты включают точки начала репликации, чтобы обеспечить автономную репликацию вектора, гены селектируемых маркеров, предпочтительно кодирующие устойчивость к антибиотику или гербициду, уникальные множественные сайты клонирования, обеспечивающие в отношении множественных сайтов вставку последовательностей нуклеиновой кислоты или генов, кодируемых в конструкте нуклеиновой кислоты, и последовательности, которые усиливают трансформацию прокариотических и эукариотических (в особенности, растительных) клеток.
"Функционально связанный", как используется в данном документе, относится к функциональному взаимоотношению между двумя или более сегментами нуклеиновой кислоты (например, ДНК). Типично, оно относится к функциональному взаимоотношению транскрипционного регуляторного элемента (промотора) с транскрибирующейся последовательностью. Например, промотор функционально связан с кодирующей последовательностью полинуклеотида, определенного в данном документе, если он стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке. Как правило, промоторные транскрипционные регуляторные элементы, которые функционально связаны с транскрибирующейся последовательностью, являются физически смежными с транскрибирующейся последовательностью, т.е., они являются г/иодействующими. Однако, некоторые транскрипционные регуляторные элементы, такие как энхансеры, не нуждаются в том, чтобы быть физически смежными или расположенными в непосредственной близости от кодирующих последовательностей, транскрипцию которых они усиливают.
Когда присутствуют множественные промоторы, то каждый промотор независимо может быть одинаковым или различным.
Рекомбинантные векторы также могут содержать: (a) один или несколько секреторных сигналов, которые кодируют сигнальные пептидные последовательности, чтобы обеспечить секрецию из клетки, которая образует полипептид, экспрессированного полипептида, определенного в данном документе, или чтобы обеспечить локализацию экспрессированного полипептида, например, для удержания полипептида в эндоплазматическом ретикулуме (ER) в клетке, или переноса в пластиду, и/или (b) содержать слитые последовательности, которые приводят к экспрессии молекул нуклеиновой кислоты в виде слитых белков. Примеры подходящих сигнальных сегментов включают любой сигнальный сегмент, способный управлять секрецией или локализацией полипептида, определенного в данном документе. Предпочтительные сигнальные сегменты включают, но без ограничения, сигнальный пептид Nicotiana nectarin (патент США №5939288), сигнал экстенсина табака или белковый сигнал, связывающийся с олеозином масляных телец сои. Рекомбинантные векторы могут также включать вставочные и/или нетранслируемые последовательности, окружающие и/или находящиеся в пределах последовательности нуклеиновой кислоты полинуклеотида, определенного в данном документе.
Для облегчения определения трансформантов рекомбинантный вектор желательно содержит селектируемый или поддающийся скринингу маркерный ген в виде, или в дополнение к, последовательности нуклеиновой кислоты полинуклеотида, определенного в данном документе. Под "маркерным геном" подразумевают ген, который обеспечивает отличающийся фенотип клеток, экспрессирующих маркерный ген и, таким образом, позволяет отличить такие трансформированные клетки от клеток, которые не имеют маркер. Селектируемый маркерный ген обеспечивает признак, по которому его можно "выбрать" на основе устойчивость к селективному средству (например, гербициду, антибиотику, облучению, нагреванию или другой обработке, повреждающей нетрансформированные клетки). Поддающийся скринингу маркерный ген (или репортерный ген) обеспечивает признак, по которому его можно определить посредством наблюдения или тестирования, иными словами, путем "скрининга" (например, β-глюкуронидазная, люциферазная, GFP или другая ферментативная активность, не присутствующая в нетрансформированных клетках). Маркерный ген и нуклеотидная последовательность интереса не должны быть связаны, поскольку котрансформация несвязанных генов, как, например, описано в патенте США №4399216, также является эффективным способом, например, в трансформации растений. Фактический выбор маркера не является принципиальным, поскольку он является функциональным (т.е., селективным) в комбинации с клетками выбора, такими как растительная клетка.
Примеры бактериальных селектируемых маркеров представляют собой маркеры, которые обеспечивают устойчивость к антибиотику, такую как устойчивость к ампицилину, эритромицину, хлорамфениколу или тетрациклину, предпочтительно устойчивость к канамицину. Иллюстративные селектируемые маркеры для селекции растительных трансформантов включают, но без ограничения, ген hyg, который кодирует устойчивость к гигромицину B; ген неомицин-фострансферазы (nptll), обеспечивающий устойчивость к канамицину, паромицину, G418; ген глутатион-S-трансферазы из печени крыс, обеспечивающий устойчивость к гербицидам, производным от глутатиона, как, например, описано в европейском патенте №256223; ген глутамин-синтетазы, обеспечивающий при сверхэкспрессии устойчивость к ингибиторам глутамин-синтетазы, таким как фосфинотрицин, как, например, описано в публикации международной заявки WO 87/05327; ген ацетилтрансферазы из Streptomyces viridochromogenes, обеспечивающий устойчивость к селективному средству фосфинотрицину, как, например, описано в европейском патенте №275957; ген, кодирующий 5-енолшикимат-3-фосфатсинтазу (EPSPS), обеспечивающий устойчивость к N-фосфонометилглицину, как, например, описано Hinchee et al. (1988); ген bar, обеспечивающий устойчивость к биалафосу, как, например, описано в публикации международной заявки WO 91/02071; ген нитрилазы, такой как Ъхп из Klebsiella ozaenae, который обеспечивает устойчивость к бромоксинилу (Stalker et al., 1988); ген дигидрофолат-редуктазы (DHFR), обеспечивающий устойчивость к метотрексату (Thillet et al., 1988); мутантный ген ацетолактат-синтазы (ALS), который обеспечивает устойчивость к имидазолинону, сульфонилмочевине или другим ALS-ингибирующим химическим средствам (европейский патент №154204); мутированный ген антранилат-синтазы, который обеспечивает устойчивость к 5-метилтриптофану; или ген далапон-дегалогеназы, который обеспечивает устойчивость к гербициду.
Предпочтительные поддающиеся скринингу маркеры включают, но без ограничения, ген uidA, кодирующий фермент β-глюкуронидазу (GUS), для которой известны различные хромогенные субстраты; ген β-галактозидазы, кодирующий фермент, для которого известны хромогенные субстраты; ген экворина (Prasher et al., 1985), который может быть использован в кальций-чувствительном биолюминисцентном обнаружении; ген зеленого флуоресцентного белка (Niedz et al., 1995) или его производных; или ген люциферазы (luc) (Ow et al., 1986), который обеспечивает биолюминисцентное обнаружение. Под "репортерной молекулой" подразумевают молекулу, которая по своей химической природе обеспечивает аналитически распознаваемый сигнал, который облегчает определение промоторной активности в соответствии с белковым продуктом.
Предпочтительно, рекомбинантный вектор стабильно встроен в геном клетки, такой как растительная клетка. Соответственно, рекомбинантный вектор может содержать необходимые элементы, которые позволяют встроить вектор в геном или в хромосому клетки.
Вектор экспрессии
Как используется в данном документе, "вектор экспрессии" представляет собой ДНК- или РНК-вектор, который способен трансформировать клетку-хозяина и воздействовать на экспрессию одного или нескольких определенных полинуклеотидов. Предпочтительно, вектор экспрессии также способен реплицироваться в клетке-хозяине. Векторы экспрессии могут быть либо для прокариот, либо для эукариот, и типично представляют собой вирусы или плазмиды. Векторы экспрессии по настоящему изобретению включают любые векторы, которые функционируют (т.е., управляют экспрессией гена) в клетках-хозяевах по настоящему изобретению, включая клетки бактерий, грибов, эндопаразитов, членистоногих, животных, водорослей и растений. Особенно предпочтительные векторы экспрессии по настоящему изобретению могут управлять экспрессией генов в дрожжах, водорослях и/или растительных клетках.
Векторы экспрессии по настоящему изобретению содержат регуляторные последовательности, такие как последовательности контроля транскрипции, последовательности контроля трансляции, точки начала репликации и другие регуляторные последовательности, которые совместимы с клеткой-хозяином и которые контролируют экспрессию полинуклеотидов по настоящему изобретению. В частности, векторы экспрессии по настоящему изобретению включают последовательности контроля транскрипции. Последовательности контроля транскрипции представляют собой последовательности, которые контролируют инициацию, элонгацию и терминацию транскрипции. Особенно важными последовательностями контроля транскрипции являются те, которые контролируют инициацию транскрипции, такие как промоторные, энхансерные, операторные и репрессорные последовательности. Подходящие последовательности контроля транскрипции включают любую последовательность контроля транскрипции, которая может функционировать по меньшей мере в одной из рекомбинантных клеток по настоящему изобретению. Выбор применяемых регуляторных последовательностей зависит от целевого организма, такого как растение, и/или целевого органа или ткани интереса. Такие регуляторные последовательности могут быть получены из любого эукариотического организма, такого как растения или вирусы растений, или могут быть химически синтезированы. Различные такие последовательности контроля транскрипции известны специалистам в данной области техники. Особенно предпочтительные последовательности контроля транскрипции представляют собой промоторы, активные в отношении управления транскрипцией в растениях, либо конститутивно, либо специфически для стадии развития и/или ткани, в зависимости от применения растения или его части(ей).
Ряд векторов, подходящих для стабильной трансфекции растительных клеток или для разработки трансгенных растений, были описаны, например, в Pouwels et al., Cloning Vectors: A Laboratory Manual, 1985, supp.1987, Weissbach and Weissbach, Methods for Plant Molecular Biology, Academic Press, 1989, и Gelvin et al., Plant Molecular Biology Manual, Kluwer Academic Publishers, 1990. Типично, векторы экспрессии для растений включают, например, один или несколько клонированных растительных генов под транскрипционным контролем 5'- и 3'-регуляторных последовательностей и доминантный селектируемый маркер. Такие векторы экспрессии для растений также могут содержать промоторный регуляторный участок (например, регуляторный участок, контролирующий индуцибельную или конститутивную, регулируемую условиями среды или стадией развития или клеточно-, или ткане-специфическую экспрессию), стартовый сайт инициации транскрипции, сайт связывания рибосомы, сигнал процессинга РНК, сайт терминации транскрипции и/или сигнал полиаденилирования.
Был описан ряд конститутивных промоторов, которые являются активными в растительных клетках. Подходящие промоторы для конститутивной экспрессии в растениях включают, но без ограничения, промотор 35S вируса мозаики цветной капусты (CaMV), 35S вируса мозаики норичника (FMV), промотор бациллярного вируса сахарного тростника, промотор вируса желтой крапчатости коммелины, индуцируемый светом промотор из малой субъединицы рибулоза-1,5-бис-фосфат-карбоксилазы, промотор цитозольной триозофосфат-изомеразы риса, промотор аденинфосфорибозилтрансферазы Arabidopsis, промотор гена актина 1 риса, промоторы маннопин-синтазы и октопин-синтазы, промотор Adh, промотор синтазы сахарозы, промотор генного комплекса R и промотор гена хлорофилл α/β-связывающего белка. Эти промоторы были использованы для создания ДНК-векторов, которые экспрессировались в растениях, смотри, например, публикацию международной заявки WO 84/02913. Все эти промоторы были использованы для создания различных типов экспрессируемых в растениях рекомбинантных ДНК-векторов.
Для цели экспрессии в тканях-источниках растения, таких как лист, семя, корень или стебель, предпочтительно, чтобы промоторы, используемые в настоящем изобретении имели относительно высокую экспрессию в этих специфических тканях. Для этой цели можно выбрать из ряда промоторов генов с ткане- или клеточно-специфической, или - усиленной экспрессией. Примеры таких промоторов, о которых сообщалось в литературе, включают промотор глутаминсинтетазы GS2 хлоропластов из гороха, промотор фруктоза-1,6-бифосфатазы хлоропластов из пшеницы, ядерный промотор ST-LS1 фотосинтетических клеток из картофеля, промотор серин/треонинкиназы и промотор глюкоамилазы (CHS) из Arabidopsis thaliana. Также сообщалось, что активными в фотосинтетически активных тканях являются промотор рибулозо-1,5-бисфосфаткарбоксилазы из восточной лиственницы (Larix laricina), промотор гена Cab, СаЬб из сосны, промотор гена Cab-1 из пшеницы, промотор гена Cab-1 из шпината, промотор гена Cab 1R из риса, промотор пируват, ортофосфат-дикиназы (PPDK) из Zea mays, промотор гена Lhcbl*2 табака, промотор Suc2 сахароза-Н30 симпортера Arabidopsis thaliana и промотор генов белков тилакоидной мембраны из шпината (PsaD, PsaF, PsaE, PC, FNR, AtpC, AtpD, Cab, RbcS). Другие промоторы для хлорофилл а/р-связывающих белков также могут быть использованы в настоящем изобретении, такие как промоторы гена LhcB и гена PsbP из белой горчицы (Sinapis alba).
Различные промоторы генов растений, которые регулируются в ответ на сигналы окружающей среды, гормональные, химические сигналы и/или сигналы стадий развития, также могут использоваться для экспрессии генов РНК-связывающих белков в растительных клетках, включая промоторы, регулируемые с помощью (1) нагревания, (2) света (например, промотор RbcS-3A гороха, промотор RbcS маиса), (3) гормонов, таких как абсцизовая кислота, (4) ранения (например, WunI) или (5) химических соединений, таких как метилжасмонат, салициловая кислота, стероидные гормоны, спирт, сафенеры (WO 97/06269) или также может быть преимущественным использование (6) орган-специфических промоторов.
Как используется в данном документе, выражение "специфический для запасающих органов растения промотор" относится к промотору, который предпочтительно, по сравнению с другими тканями растения, управляет транскрипцией генов в запасающем органе растения. Предпочтительно, промотор управляет экспрессией только гена интереса в запасающем органе, и/или экспрессией гена интереса в других частях растения, таких как листья, не обнаруживаемой с помощью нозерн-блот анализа и/или RT-ПЦР. Типично, промотор управляет экспрессией генов во время роста и развития запасающего органа, в частности, во время фазы синтеза и накопления запасных соединений в запасающем органе. Такие промоторы могут управлять экспрессией генов во всем запасающем органе растения или только его части, такой как оболочка семени, зародыш или семядоля(и) в семенах двудольных растений или эндосперм или алейроновый слой семени однодольных растений.
Для цели экспрессии в накопительных тканях растения, таких как клубень картофеля, плод томата или семя сои, канолы, хлопка, Zea mays, пшеницы, риса и ячменя, предпочтительно, чтобы промоторы, используемые в настоящем изобретении, имели относительно высокую экспрессию в этих специфических тканях. Известен ряд промоторов для генов с клубеньспецифической или - усиленной экспрессией, включая промотор пататина класса I, промотор генов ADPGPP картофельных клубней, как больших, так и малых субъединиц, промотор сахароза-синтазы, промотор для основных белков клубней, включая белковые комплексы 22 кДа и ингибиторы протеиназы, промотор для гена связывающейся гранулами крахмала синтазы (GBSS) и другие промоторы пататинов класса I и II. Другие промоторы также могут быть использованы для экспрессии белка в специфических тканях, таких как семена или плоды. Могуг использоваться промоторы (3-конглицинина или другие семя-специфические промоторы, такие как промоторы напина, зеина, линина и фазеолина. Также могут быть использованы корень-специфические промоторы. Пример такого промотора представляет собой промотор для гена кислотной хитиназы. Экспрессия в ткани корня также может осуществляться путем использования промотора 35S корень-специфических субдоменов CaMV, который был определен.
В особенно предпочтительном варианте осуществления промотор управляет экспрессией в тканях и органах, в которых имеет место биосинтез липидов. Такие промоторы действуют в развитии семени в подходящее время для модификации липидного состава в семенах.
В особенно предпочтительном дополнительном варианте осуществления промотор представляет собой специфический для запасающих органов растения промотор. В одном варианте осуществления специфический для запасающих органов растения промотор представляет собой семя-специфический промотор. В более предпочтительном варианте осуществления промотор предпочтительно управляет экспрессией в семядолях двудольного растения или в эндосперме однодольного растения, по сравнению с экспрессией в зародыше семени или относительно других органов в растении, таких как листья. Предпочтительные промоторы для семя-специфической экспрессии включают: 1) промоторы из генов, кодирующих ферменты, вовлеченные в биосинтез и накопление липидов в семенах, такие как десатуразы и элонгазы, 2) промоторы из генов, кодирующих запасные белки семени и 3) промоторы из генов, кодирующих ферменты, вовлеченные в биосинтез и накопление углеводов в семенах. Семя-специфические промоторы, которые являются подходящими, представляют собой промотор гена напина масличного рапса (патент США №5608152), промотор USP Vicia faba (Baumlein et al., 1991), промотор олеозина Arabidopsis (WO 98/45461), промотор фазеолина Phaseolus vulgaris (патент США №5504200), промотор Все4 Brassica (WO 91/13980) или промотор легумина В4 (Baumlein et al., 1992) и промоторы, которые приводят к семя-специфической экспрессии в однодольных, таких как маис, ячмень, пшеница, рожь, рис и т.п. Особо следует отметить промоторы, которые являются подходящими, такие как промотор гена lpt2 или lptl ячменя (WO 95/15389 и WO 95/23230) или промоторы, описанные в WO 99/16890 (промоторы из гена гордеина ячменя, гена глутелина риса, гена оризина риса, гена проламина риса, гена глиадина пшеницы, гена глутелина пшеницы, гена зеина маиса, гена глутелина овса, гена касирина сорго, гена секалина ржи). Другие промоторы включают таковые, описанные Broun et al. (1998), Potenza et al. (2004), патентной заявке США №20070192902 и патентной заявке США №20030159173. В варианте осуществления семя-специфический промотор предпочтительно экспрессируется в определенных частях семени, таких как семядоля(и) или эндосперм. Примеры семядоля-специфических промоторов включают, но без ограничения, промотор FP1 (Ellerstrom et al., 1996), промотор легумина гороха (Perrin et al., 2000) и промотор фитогемагглютинина бобов (Perrin et at., 2000). Примеры эндосперм-специфических промоторов включают, но без ограничения, промотор зеина-1 маиса (Chikwamba et al., 2003), промотор глутелина-1 риса (Yang et al., 2003), промотор D-гордеина ячменя (Horvath et at., 2000) и промоторы глутенина HMW пшеницы (Alvarez et al., 2000). В еще одном варианте осуществления семя-специфический промотор не экспрессируется или экспрессируется только на низком уровне в зародыше и/или после прорастания семени.
В другом варианте осуществления специфический для запасающих органов растения промотор представляет собой клубень-специфический промотор. Примеры включают, но без ограничения, промоторы ВЗЗ, РАТ21 и GBSS пататина картофеля, а также промотор спорамина сладкого картофеля (для обзора, смотри Potenza et at., 2004). В предпочтительном варианте осуществления промотор управляет экспрессией предпочтительно в сердцевине клубня по сравнению с внешними слоями (кожурой, корой) или зародыше клубня.
В другом варианте осуществления специфический для запасающих органов растения промотор представляет собой плод-специфический промотор. Примеры включают, но без ограничения, промоторы Е8 и Pds полигалактуроназы томата, а также промотор АСС-оксидазы яблока (для обзора, смотри Potenza et at., 2004). В предпочтительном варианте осуществления промотор предпочтительно управляет экспрессией в съедобных частях плода, например, мякоти плода, по сравнению с кожурой плода или семенах внутри плода.
5'-нетранслируемая лидерная последовательность может быть получена из промотора, выбранного для экспрессии гетерологичной генной последовательности полинуклеотида по настоящему изобретению, или может быть гетерологичной по отношению к кодирующему участку фермента, который необходимо получить, и может быть, в частности, модифицирована при необходимости так, чтобы повышать трансляцию мРНК. Для обзора оптимизированной экспрессии трансгенов, смотри Koziel et al. (1996). 5'-нетранслируемые участки могут также быть получены из РНК вирусов растений (вирус табачной мозаики, вирус гравировки табака, вирус карликовой мозаики маиса, вирус мозаики люцерны, среди прочих), из подходящих эукариотических генов, генов растений (лидер гена хлорофилл α/β-связывающего белка пшеницы и маиса) или из синтетической генной последовательности. Настоящее изобретение не ограничивается конструктами, где нетранслируемый участок получен из 5'-нетранслируемой последовательности, которая сопровождает промоторную последовательность. Лидерная последовательность также может быть получена из несвязанной промоторной или кодирующей последовательности. Лидерные последовательности, применимые в контексте настоящего изобретения, содержат лидер Hsp70 маиса (патент США №5362865 и патент США №5859347) и элемент TMV омега.
Терминация транскрипции осуществляется с помощью 3'-нетранслируемой последовательности ДНК, функционально связанной в векторе экспрессии с полинуклеотидом интереса. 3'-нетранслируемый участок рекомбинантной молекулы ДНК содержит сигнал полиаденилирования, который функционирует в растениях, чтобы вызвать добавление аденилатных нуклеотидов к 3'-концу РНК. 3'-нетранслируемый участок может быть получен из различных генов, которые экспрессируются в растительных клетках. 3'-нетранслируемый участок нопалинсинтазы, 3'-нетранслируемый участок из гена малой субъединицы Rubisco гороха, 3'-нетранслируемый участок из гена запасного белка 7S семени сои широко используются для этой функции. 3'-транскрибированные нетранслируемые участки, содержащие полиаденилатный сигнал генов опухоль-индуцирующей (Ti) плазмиды Agrobacterium, также являются подходящими.
Рекомбинантные ДНК технологии могут использоваться для улучшения экспрессии трансформированного полинуклеотида путем манипуляции, например, с числом копий полинуклеотида внутри клетки хозяина, эффективностью, с которой эти полинуклеотиды транскрибируются, эффективностью, с которой полученые транскрипты транслируются и эффективностью посттрансляционных модификаций. Рекомбинантные методики, применимые для увеличения экспрессии полинуклеотидов, определенных в данном документе, включают, но без ограничения, функциональное связывание полинуклеотида с плазмидой с высокой копийностью, интеграцию полинуклеотидной молекулы в одну или несколько хромосом клетки-хозяина, добавление последовательностей для стабилизации вектора к плазмиде, замен или модификаций сигналов контроля транскрипции (например, промоторов, операторов, энхансеров), замен или модификаций сигналов контроля трансляции (например, сайтов связывания рибосомы, последовательностей Шайна-Дальгарно), модификацию полинуклеотида для соответствия использованию кодонов клетки-хозяина и делецию последовательностей, которые дестабилизируют транскрипты.
Нуклеиновые кислоты переноса
Нуклеиновые кислоты переноса могут использоваться для доставки экзогенного полинуклеотида к клетке и содержат одну, предпочтительно две, пограничные последовательности и полинуклеотид интереса. Нуклеиновая кислота переноса может кодировать или не кодировать селектируемый маркер. Предпочтительно, нуклеиновая кислота переноса образует часть бинарного вектора в бактерии, где бинарный вектор дополнительно содержит элементы, которые обеспечивают репликацию вектора в бактерии, селекцию или поддержание бактериальных клеток, содержащих бинарный вектор. При переносе к эукариотической клетке компонент нуклеиновой кислоты переноса бинарного вектора способен к интеграции в геном эукариотической клетки.
Как используется в данном документе, выражение "нуклеиновая кислота внехромосомного переноса" относится к молекуле нуклеиновой кислоты, которая способна переноситься из бактерии, такой как Agrobacterium sp., к эукариотической клетке, такой как клетка листа растения. Нуклеиновая кислота внехромосомного переноса представляет собой генетический элемент, который хорошо известен как элемент, способный переноситься с последующей интеграцией нуклеотидной последовательности, содержащейся в пределах пограничных последовательностей, в геном клетки-реципиента. В этом отношении нуклеиновая кислота переноса фланкируется типично с помощью двух "пограничных" последовательностей, хотя в некоторых случаях может применяться одна пограничная последовательность на одном конце, а второй конец перенесенной нуклеиновой кислоты создается случайно в процессе переноса. Полинуклеотид интереса типично помещается между левой подобной пограничной последовательностью и правой подобной пограничной последовательностью нуклеиновой кислоты переноса. Полинуклеотид, содержащийся внутри нуклеиновой кислоты переноса, может быть функционально связан с разнообразием различных промоторных и терминаторных регуляторных элементов, которые облегчают его экспрессию, иными словами, транскрипцию и/или трансляцию полинуклеотида. ДНК переноса (Т-ДНК) из Agrobacterium sp., таких как Agrobacterium tumefaciens или Agrobacterium rhizogenes, и ее искусственные варианты/мутанты, вероятно, являются наилучшим образом охарактеризованными примерами нуклеиновых кислот переноса. Другой пример представляет собой Р-ДНК ("ДНК растения"), которая содержит подобные пограничным последовательности Т-ДНК из растений.
Как используется в данном документе, "Т-ДНК" относится, например, к Т-ДНК из плазмиды Ti Agrobacterium tumefaciens или из плазмиды Ri Agrobacterium rhizogenes, или их искусственным вариантам, которые функционируют как Т-ДНК. Т-ДНК могут содержать целую Т-ДНК, включая как правую, так и левую пограничные последовательности, но необходимо, чтобы они содержали лишь минимальные последовательности, необходимые в цис-положении для переноса, иными словами, правую и пограничную последовательность Т-ДНК. Т-ДНК по настоящему изобретению были введены в них, где-либо между правой и левой пограничными последовательностями (в случае, когда они присутствуют) полинуклеотида интереса, фланкированного целевыми сайтами для сайт-специфической рекомбиназы. Последовательности, кодирующие факторы, необходимые в транс-положении для переноса Т-ДНК в растительную клетку, такие как гены vir, могут быть введены в Т-ДНК или могут присутствовать на одном и том же репликоне, что и Т-ДНК, или предпочтительно находятся в транс-положении на совместимом репликоне в хозяине Agrobacterium. Такие "бинарные векторные системы" хорошо известны в уровне техники.
Как используется в данном документе, "Р-ДНК" относится к нуклеиновой кислоте переноса, выделенной из генома растения, или ее искусственных вариантов/мутантов, и содержит на каждом конце или только на одном конце подобную пограничной последовательность Т-ДНК. Подобная пограничной последовательность предпочтительно имеет по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95%, но менее чем 100% идентичности последовательности с пограничной последовательностью Т-ДНК из Agrobacterium sp., таких как Agrobacterium tumefaciens или Agrobacterium rhizogenes. Таким образом, Р-ДНК могут использоваться вместо Т-ДНК для переноса нуклеотидной последовательности, содержащейся внутри Р-ДНК из, например, Agrobacterium, к другой клетке. Р-ДНК, до вставки экзогенного полинуклеотида, который необходимо перенести, может быть модифицирована для облегчения клонирования и предпочтительно не должна кодировать какие-либо белки. Р-ДНК характеризуется тем, что содержит по меньшей мере правую пограничную последовательность и предпочтительно также левую пограничную последовательность.
Как используется в данном документе, "пограничная" последовательность нуклеиновой кислоты переноса может быть выделена из выбранного организма, такого как растение или бактерия, или быть ее вариантом/мутантом. Пограничная последовательность стимулирует и облегчает перенос полинуклеотида, с которым она связана и может облегчать его интеграцию в геном клетки-реципиента. В варианте осуществления пограничная последовательность составляет 5-100 пар оснований (п.о.) в длину, 10-80 п.о. в длину, 15-75 п.о. в длину, 15-60 п.о. в длину, 15-50 п.о. в длину, 15-40 п.о. в длину, 15-30 п.о. в длину, 16-30 п.о. в длину, 20-30 п.о. в длину, 21-30 п.о. в длину, 22-30 п.о. в длину, 23-30 п.о. в длину, 24-30 п.о. в длину, 25-30 п.о. в длину или 26-30 п.о. в длину. Пограничные последовательности из Т-ДНК из Agrobacterium sp.хорошо известны в уровне техники и включают последовательности, описанные в Lacroix et al. (2008), Tzfira and Citovsky (2006) и Glevin (2003).
Тогда как традиционно только Agrobacterium sp.применялись для переноса генов в растительные клетки, в настоящее время существует большое количество систем, которые были определены/разработаны, чтобы действовать подобно Agrobacterium sp.Некоторые не относящиеся к Agrobacterium виды недавно были генетически модифицированы, чтобы быть способными переносить гены (Chung et at., 2006; Broothaerts et at., 2005). Они включают Rhizobium sp.NGR234, Sinorhizobium meliloti и Mezorhizobium loti. Бактерии созданы со способностью к переносу генов путем обеспечения бактерий с механизмом, необходимым для процесса трансформации, иными словами, набора вирулентных генов, кодируемых Ti-плазмидой Agrobacterium и сегментом Т-ДНК, находящимся на отдельной небольшой бинарной плазмиде. Бактерии, сконструированные таким образом, способны трансформировать различные ткани растений (листовые диски, каллюсы и овальную ткань), однодольных или двудольных, и различные виды растений (например, табак, рис).
Прямой перенос плазмид экспрессии для эукариот из бактерий к эукариотическим хозяевам был впервые достигнут несколько десятилетий назад путем слияния клеток млекопитающих и протопластов, несущих плазмиду Escherichia coli (Schaffner, 1980). С того времени число бактерий, способных доставлять гены в клетки млекопитающих, непрерывно увеличивалось (Weiss, 2003), причем они были независимо открыты четырьмя группами (Sizemore et al. 1995; Courvalin et at., 1995; Powell et al., 1996; Darji et at., 1997).
Аттенуированные Shigella flexneri, Salmonella typhimurium или E.coli, которые стали инвазивными с помощью плазмиды вирулентности (pWR100) S. flexneri, как было показано, являлись способными переносить плазмиды экспрессии после инвазии клеток-хозяев и внутриклеточной гибели вследствие метаболического аттенуирования. Мукозальное введение, либо назально, либо перорально, таких рекомбинантных Shigella или Salmonella вызвало иммунные реакции на антиген, который кодировался плазмидами экспрессии. Между тем, перечень бактерий, которые, как было показано, были способны переносить плазмиды экспрессии к клеткам-хозяевам млекопитающих in vitro и in vivo был более чем удвоен и был документально подтвержден для S. typhi, S. choleraesuis, Listeria monocytogenes, Yersinia pseudotuberculosis и Y. enterocolitica (Fennelly et at., 1999; Shiau et at., 2001; Dietrich et al., 1998; Hense et at., 2001; Al-Mariri et at., 2002).
В основном, можно допустить, что все бактерии, которые способны проникать в цитозоль клетки-хозяина (подобно S. flexneri или L. monocytogenes) и лизироваться внутри этого клеточного компартмента, должны быть способными к переносу ДНК. Это известно как 'абортивная' или 'суицидальная' инвазия, так как бактерии должны лизироваться для того, чтобы произошел перенос ДНК (Grillot-Courvalin et at., 1999). К тому же, даже многие из бактерий, которые остаются в фагоцитарной вакуоле (подобно S. typhimurium) также могут быть способны к этому. Таким образом, рекомбинантные лабораторные штаммы Е. coli, которые были сконструированы, чтобы быть инвазивными, но не способными покидать фагосому, все же могут доставлять их плазмидный груз к ядру инфицированной клетки млекопитающего (Grillot-Courvalin et at., 1998). Кроме того, недавно было показано, что Agrobacterium tumefaciens способны вводить трансгены в клетки млекопитающих (Kunik et al., 2001).
Как используется в данном документе, выражения "трансфекция", "трансформация" и их варианты, как правило, используются взаимозаменяемо. "Трансфицированные" или "трансформированные" клетки могли быть подвергнуты манипуляции для введения полинуклеотида(ов) интереса или могут быть клетками потомства, полученными из них.
Рекомбинантные клетки
Настоящее изобретение также обеспечивает рекомбинантную клетку, например, рекомбинантную растительную клетку, которая представляет собой клетку-хозяина, трансформированную при помощи одного или нескольких полинуклеотидов или векторов, определенных в данном документе, или их комбинацией. Выражение "рекомбинантная клетка" в данном документе используется взаимозаменяемо с выражением "трансгенная клетка". Подходящие клетки по настоящему изобретению включают любую клетку, которая может быть трансформирована при помощи полинуклеотида или рекомбинантного вектора по настоящему изобретению, кодирующего, например, полипептид или фермент, описанный в данном документе. Клетка предпочтительно представляет собой клетку, которую, таким образом, можно использовать для получения липида. Рекомбинантная клетка может быть клеткой в культуре, клеткой in vitro или в организме, таком как, например, растение, или в органе, таком как, например, семя или лист.Предпочтительно, клетка находится в растении, более предпочтительно в семени растения.
Клетки-хозяева, в которые вводят полинуклеотид(ы), могут быть либо нетрансформированными клетками, либо клетками, которые уже трансформированы при помощи по меньшей мере одной нуклеиновой кислоты. Такие нуклеиновые кислоты могут быть связаны с синтезом липидов или не связаны с ним. Клетки-хозяева по настоящему изобретению либо могут быть эндогенно (т.е., в естественном состоянии) способны образовывать полипептид(ы), определенные в данном документе, в этом случае рекомбинантная клетка, полученная из них, имеет усиленную способность образовывать полипептид(ы), либо могут быть способны образовывать указанный полипептид(ы) только после трансформации при помощи по меньшей мере одного полинуклеотида по настоящему изобретению. В варианте осуществления рекомбинантная клетка по настоящему изобретению имеет усиленную способность образовывать неполярный липид.
Клетки-хозяева по настоящему изобретению могут быть любой клеткой, способной образовывать по меньшей мере один белок, описанный в данном документе, и включают клетки бактерий, грибов (включая дрожжи), паразитов, членистоногих, животных, водорослей и растений. Клетки могут быть прокариотическими или эукариотическими. Предпочтительные клетки-хозяева представляют собой клетки дрожжей, водорослей и растений. В предпочтительном варианте осуществления растительная клетка представляет собой клетку семени, в частности, клетку в семядоле или эндосперме семени. В одном варианте осуществления клетка представляет собой клетку животного. Клетка животного может принадлежать любому типу животного, такому как, например, клетка не являющегося человеком животного, клетка не являющегося человеком позвоночного, клетка не являющегося человеком млекопитающего или клетки водных животных, таких как рыбы или ракообразные, беспозвоночных, насекомых и т.д. Неограничивающие примеры клеток членистоногих включают клетки насекомых, такие как клетки (Sf) Spodoptera frugiperda, например, Sf9, Sf21, клетки Trichoplusia ni и клетки S2 Drosophila. Пример бактериальной клетки, применимой в качестве клетки-хозяина по настоящему изобретению, представляют собой Synechococcus spp.(также известные как Synechocystis spp.), например Synechococcus elongatus. Примеры клеток водорослей, применимых в качестве клеток-хозяев по настоящему изобретению, включают, например, Chlamydomonas sp.(например, Chlamydomonas reinhardtii), Dunaliella sp., Haematococcus sp., Chlorella sp., Thraustochytrium sp., Schizochytrium sp., и Volvox sp.
Клетки-хозяева для экспрессии нуклеиновых кислот по настоящему изобретению могут включать микроорганизмы-хозяева, которые растут на разнообразных видах сырья, включая простые или сложные углеводы, органические кислоты и спирты и/или углеводороды, в широком диапазоне температуры и значений рН. Предпочтительные микроорганизмы-хозяева представляют собой жировые организмы, которые в естественном состоянии способны к синтезу неполярных липидов.
Клетки-хозяева могут быть клетками организма, подходящего для процесса ферментации, такие как, например, Yarrowia lipolytica или другие дрожжи.
Трансгенные растения
Настоящее изобретение также обеспечивает растение, содержащее экзогенный полинуклеотид или полипептид по настоящему изобретению, клетку по настоящему изобретению, вектор по настоящему изобретению или их комбинацию. Выражение "растение" относится к целым растениям, тогда как выражение "его часть" относится к органам растения (например, листьям, стеблям, корням, цветкам, плодам), отдельным клеткам (например, пыльце), семени, частям семени, таким как зародыш, эндосперм, щиток или оболочка семени, растительной ткани, такой как проводящая ткань, растительным клеткам и потомству перечисленного. Как используется в данном документе, части растения включают растительные клетки.
Как используется в данном документе, выражение "растение" используют в его самом широком смысле. Оно включает, но без ограничения, любой вид травы, орнаментального или декоративного растения, сельскохозяйственную культуру или злак (например, масличную культуру, маис, сою), кормовое или фуражное, плодовое или овощное растение, травянистое растение, древесное растение, цветковое растение или дерево. Не подразумевается ограничение растения любой конкретной структурой. Оно также относится к одноклеточному растению (например, микроводоросли). Выражение "его часть" в отношении растения относится к растительной клетке и ее потомству, множеству растительных клеток, которые в значительной степени дифференцированы в колонию (например, вольвокс), структуре, которая присутствует на любой стадии развития растения или ткани растения. Такие структуры включают, но без ограничения, листья, стебли, цветки, плоды, орехи, корни, семя, оболочку семян, зародыши. Выражение "ткань растения" включает дифференцированные и недифференцированные ткани растений, включая те, которые присутствуют в листьях, стеблях, цветках, плодах, орехах, корнях, семени, например, зародышевая ткань, эндосперм, дермальная ткань (например, эпидермис, перидерма), проводящая ткань (например, ксилема, флоэма) или покровная ткань (включающая клетки паренхимы, колленхимы и/или склеренхимы), а также клетки в культуре (например, отдельные клетки, протопласты, каллюс, зародыши и т.д.). Ткань растения может быть in planta, в культуре органа, культуре тканей или культуре клеток.
"Трансгенное растение", "генетически модифицированное растение" или их варианты относятся к растению, которое содержит трансген, не обнаруживаемый в растении дикого типа одного и того же вида, сорта или культивара. Трансгенные растения, как определено в контексте настоящего изобретения, включают растения и их потомство, которые были генетически модифицированы с использованием рекомбинантных методик, чтобы вызвать образование по меньшей мере одного полипептида, определенного в данном документе, в необходимом растении или его части. Части трансгенного растения имеют соответствующее значение.
Выражения "семя" и "зерно" используются в данном документе взаимозаменяемо. "Зерно" относится к зрелому зерну, такому как собранное зерно или зерно, которое все еще находится на растении, но готово к сбору, но также может относиться к зерну после набухания или прорастания, согласно контексту. Зрелое зерно, как правило, имеет содержание влаги менее около 18-20%. "Развивающееся семя", как используется в данном документе, относится к семени перед зрелостью, типично, обнаруживаемом в репродуктивных структурах растения после опыления или цветения, но также может относиться к таким семенам перед зрелостью, которые выделены из растения.
Как используется в данном документе, выражение "запасающий орган растения" относится к части растения, специализирующейся на запасании энегрии в форме, например, белков, углеводов, липида. Примеры запасающих органов растения представляют собой семя, плод, клубневидные корни и клубни. Предпочтительный запасающий орган растения по настоящему изобретению представляет собой семя.
Как используется в данном документе, выражение "фенотипически нормальный" относится к генетически модифицированному растению или его части, в частности, запасающему органу, такому как семя, клубень или плод по настоящему изобретению, не имеющему значительно сниженной способности к росту и репродукции по сравнению с немодифицированным растением или его частью. В варианте осуществления генетически модифицированное растение или его часть, которое является фенотипически нормальным, содержит рекомбинантный полинуклеотид, кодирующий супрессор сайленсинга, функционально связанный со специфическим для запасающих органов растения промотором, и имеет способность расти или репродуцироваться, которая по сути одинакова с таковой у соответствующего растения или его части, не содержащей указанный полинуклеотид. Предпочтительно, биомасса, скорость роста, скорость прорастания, размер запасающего органа, размер семени и/или количество образованных жизнеспособных семян, составляет не менее 90% от таковой у растения, где отсутствует указанный рекомбинантный полинуклеотид, при выращивании при идентичных условиях. Это выражение не включает признаки растения, которые могут быть отличными от растения дикого типа, но которые не влияют на пригодность растения для коммерческих целей, такие как, например, фенотип балерины листьев саженцев.
Растения, обеспеченные или предусмотренные для применения на практике по настоящему изобретению, включают как однодольные, так и двудольные. В предпочтительных вариантах осуществления растения по настоящему изобретению представляют собой сельскохозяйственные растения (например, злаковые и зернобобовые, маис, пшеница, разновидности картофеля, тапиока, рис, сорго, просо, маниок, ячмень или горох) или другие бобовые. Растения могут выращиваться для получения съедобных корней, клубней, листьев, стеблей, цветков или плодов. Растения могут быть овощными или орнаментальными растениями. Растения по настоящему изобретению могут быть следующими: кукуруза (Zea mays), канола (Brassica napus, Brassica rapa ssp.), другие виды Brassica, такие как, например, брюква (Brassica napobrassica) или Brassica camelina, сахарная свекла (Beta vulgaris), клевер (Trifolium sp.), лен (Linum usitatissimum), люцерна (Medicago sativa), рис (Oryza sativa), рожь (Secale cerale), сорго (Sorghum bicolor, Sorghum vulgare), подсолнечник (Helianthus annus), пшеница (Tritium aestivum), соя (Glycine max), табак (Nicotiana tabacum), картофель (Solatium tuberosum), разновидности арахиса (Arachis hypogaea), хлопок (Gossypium hirsutum), сладкий картофель (Lopmoea batatus), маниока (Manihot esculenta), кофе (Cofea spp.), кокос (Cocos nucifera), ананас (Anana comosus), лимонное дерево {Citrus spp.), какао (Theobroma cacao), чай (Camellia senensis), банан (Musa spp.), авокадо (Persea americana), инжир (Ficus casica), гуаява (Psidium guajava), манго (Mangifer indica), маслина (Olea europaea), папайя (Carica papaya), кешью (Anacardium occidentale), макадамия (Macadamia intergrifolia), миндаль (Prunus amygdalus), разновидности сахарной свеклы (Beta vulgaris), разновидности овса или ячмень.
Другие предпочтительные растения включают С4-травы, такие как Andropogon gerardi, Bouteloua curtipendula, В. gracilis, Buchloe dactyloides, Panicum virgalum, Schizachyrium scoparium, виды Miscanthus, например, Miscanthus x giganteus и Miscanthus sinensis, Sorghastrum nutans, Sporobolus cryptandrus, просо прутьевидное (Panicum virgatum); С3-травы, такие как Elymus canadensis, бобовые Lespedeza capitata и Petalostemum villosum, полукустарник Aster azureus; и древесные растения, такие как Quercus ellipsoidalis и Q. macrocarpa.
В предпочтительном варианте осуществления растение является покрытосеменным.
В варианте осуществления растение представляет собой масличное растение, предпочтительно масличное сельскохозяйственное растение. Как используется в данном документе, "масличное растение" представляет собой вид растения, используемый для коммерческого получения липида из семян растения. Масличное растение может быть масличным рапсом (таким как канола), маисом, подсолнечником, сафлором, соей, сорго, льном (семенем льна) или сахарной свеклой. Кроме того, масличное растение может представлять собой следующее: другие виды Brassica, хлопок, арахис, мак, брюкву, горчицу, касторовый боб, кунжут, сафлор или образующие орехи растения. Растение может образовывать высокие уровни липида в своих плодах, например, маслина, масличная пальма или кокос. Плодовые растения, к которым может быть применено настоящее изобретение, представляют собой латук, эндивий или овощные виды Brassica, включая капусту, брокколи или цветную капусту. Настоящее изобретение может быть применено к табаку, тыкве, моркови, клубнике, томату или перцу.
В предпочтительном варианте осуществления трансгенное растение является гомозиготным по каждому гену, который был введен (трансген), так чтобы его потомство не сегрегировалось в отношении желаемого фенотипа. Трансгенное растение также может быть гетерозиготным в отношении введенного трансгена(ов), предпочтительно однородно гетерозиготного в отношении трансгена, как, например, в потомстве F1, которое было выращено из гибридного семени. Такие растения могут обеспечить преимущества, такие как гибридная сила, хорошо известные в уровне техники.
Там, где это уместно, трансгенные растения также могут содержать дополнительные трансгены, кодирующие ферменты, вовлеченные в образование неполярного липида, такие как, без ограничения, LPAAT, LPCAT, РАР или фосфолипид:диацилглицерол-ацилтрансфераза (PDAT1, PDAT2 или PDAT3; смотри, например, Ghosal et at., 2007) или комбинацию двух или более из них. Трансгенные растения по настоящему изобретению также могут экспрессировать олеозин из экзогенного полинуклеотида.
Трансформация растений
Трансгенные растения могут быть получены с использованием методик, известных в уровне техники, таких как те, которые в общем описаны в Slater et al., Plant Biotechnology - The Genetic Manipulation of Plants, Oxford University Press (2003) и Christou and Klee, Handbook of Plant Biotechnology, John Wiley and Sons (2004).
Как используется в данном документе, выражения "стабильная трансформация", "стабильно трансформированный" и их вариации относятся к интеграции полинуклеотида в геном клетки так, чтобы они передавались потомству клетки во время клеточного деления без необходимости положительной селекции на их присутствие. Стабильные трансформанты или их потомство, могут быть выбраны с помощью любых средств, известных в уровне техники, таких как саузерн-блот анализы на хромосомной ДНК, или in situ гибридизация геномной ДНК.
Agrobacterium-опосредованный перенос представляет собой широко применяемую систему для введения генов в растительные клетки, так как ДНК может быть введена в клетки в тканях целого растения, органах растения или эксплантатах в культуре тканей, либо для временной экспрессии, либо для стабильной интеграции ДНК в геном растительной клетки. Применение Agrobacterium-опосредованных интегрирующихся в растения векторов для введения ДНК в растительные клетки является хорошо известным в уровне техники (смотри, например, патент США №5177010, патент США №5104310, патент США №5004863 или патент США №5159135). Участок ДНК, подлежащий переносу, определяют с помощью пограничных последовательностей, и вставочную ДНК (Т-ДНК), как правило, вводят в геном растения. Дополнительно, интеграция Т-ДНК представляет собой относительно точный процесс, приводящий к немногочисленным перестановкам. В тех сортах растений, где Agrobacterium-опосредованная трансформация является эффективной, она является способом выбора из-за легкости и определенной природы переноса генов. Предпочтительные векторы для трансформации на основе Agrobacterium способны к репликации в Е. coli, также как и в Agrobacterium, обеспечивая удобство манипуляций, как описано (Klee et al., В: Plant DNA Infectious Agents, Hohn and Schell, eds., Springer-Verlag, New York, pp.179-203 (1985)).
Способы ускорения, которые могут быть использованы, включают, например, бомбардировку микрочастицами и т.п.Один пример способа для доставки трансформирующих молекул нуклеиновой кислоты к растительным клеткам представляет собой бомбардировку микрочастицами. Этот способ был описан в обзоре Yang et al., Particle Bombardment Technology for Gene Transfer, Oxford Press, Oxford, England (1994). Небиологические частицы (микрочастицы), которые могут быть покрыты нуклеиновыми кислотами и доставлены в клетки с помощью реактивной силы. Иллюстративные частицы включают те, которые состоят из вольфрама, золота, платины и т.п. Конкретное преимущество бомбардировки микрочастицами, кроме того, что она является эффективным средством воспроизводимой трансформации однодольных, состоит в том, что ни выделение протопластов, ни чувствительность к инфицированию Agrobacterium не являются необходимыми. Иллюстративный вариант осуществления способа для доставки ДНК в клетки Zea mays с помощью ускорения представляет собой систему доставки а-частиц с помощью биобаллистики, которая может использоваться для проталкивания частиц, покрытых ДНК, через сито, такое как сито из нержавеющей стали или сито Nytex, на поверхность фильтра, покрытую клетками кукурузы, культивируемыми в суспензии. Система доставки частиц, подходящая для применения в настоящем изобретении, представляет собой гелиевую пушку для ускорения PDS-1000/Не, доступную от Bio-Rad Laboratories.
Для бомбардировки клетки в суспензии могут концентрироваться на фильтре. Фильтры, содержащие клетки, подлежащие бомбардировке, располагают на необходимом расстоянии ниже тормозного листа для микрочастиц. При необходимости, одно или несколько сит также располагают между пушкой и клетками, подлежащими бомбардировке.
Альтернативно, незрелые зародыши или другие целевые клетки могут быть расположены на твердой культуральной среде. Клетки, подлежащие бомбардировке, располагают на необходимом расстоянии ниже тормозного листа для микрочастиц. При необходимости, одно или несколько сит также располагают между устройством ускорения и клетками, подлежащими бомбардировке. Посредством применения методик, изложенных в данном документе, можно получить до 1 ООО или более очагов клеток, временно экспрессирующих маркерный ген. Количество клеток в очаге, которые экспрессируют продукт гена через 48 часов после бомбардировки часто находится в диапазоне от одного до десяти и в среднем от одного до трех.
В трансформации бомбардировкой можно оптимизировать условия культивирования перед бомбардировкой и параметры бомбардировки для получения максимальных количеств стабильных трансформантов. Как физические, гак и биологические параметры для бомбардировки являются важными в этой технологии. Физические факторы представляют собой те, которые включают манипуляцию с ДНК/преципитатом микрочастиц или те, которые воздействуют на полет и скорость либо макро-, либо микрочастиц. Биологические факторы включают все этапы, вовлеченные в манипуляцию с клетками перед и непосредственно после бомбардировки, осмотическое регулирование целевых клеток, чтобы помочь ослабить травмирование, связанное с бомбардировкой, а также природа трансформирующей ДНК, такой как линеаризированная ДНК или интактные сверхспиральные плазмиды. Считается, что манипуляции перед бомбардировкой являются особенно важными для успешной трансформации незрелых зародышей.
В другом альтернативном варианте осуществления пластиды могут быть стабильно трансформированы. Способы, раскрытые для трансформации пластид у высших растений, включают биобаллистическую доставку ДНК, содержащей селектируемый маркер, и направленное воздействие ДНК на геном пластиды посредством гомологичной рекомбинации (патент США №5451513, патент США №5545818, патент США №5877402, патент США №5932479 и публикация международной заявки WO 99/05265).
Соответственно, предусматривается, что может возникнуть желание отрегулировать различные аспекты параметров бомбардировки в исследованиях малого масштаба для того, чтобы полностью оптимизировать условия. Может возникнуть желание, в частности, отрегулировать физические параметры, такие как расстояние зазора, расстояние полета, расстояние до ткани и давление гелия. Можно также минимизировать снижающие факторы травмирования путем модификации условий, которые влияют на физиологическое состояние клеток-реципиентов и которые могут, следовательно, влиять на эффективности трансформации и интеграции. Например, осмотическое состояние, гидратация ткани и стадия субкультуры или клеточного цикла клеток-реципиентов, могут быть отрегулированы для оптимальной трансформации. Выполнение других рутинных регулировок будет известно специалистам в данной области техники с учетом настоящего раскрытия.
Трансформация растительных протопластов может быть достигнута с использованием способов, основанных на преципитации фосфатом кальция, обработке полиэтиленгликолем, электропорации и комбинации этих обработок. Применение этих систем к различным сортам растений зависит от способности к регенерации растений этой конкретной разновидности из протопластов. Описаны иллюстративные способы для регенерации злаковых из протопластов (Fujimura et at., 1985; Toriyama et at., 1986; Abdullah et at., 1986).
Также можно применять другие способы трансформации клеток, и они включают, но без ограничения, введение ДНК в растения с помощью прямого переноса ДНК в пыльцу, прямой инъекции ДНК в репродуктивные органы растения или прямой инъекции ДНК в клетки незрелых зародышей с последующей регидратацией высушенных зародышей.
Регенерация, развитие и культивирование растений из отдельных трансформантов растительных протопластов или из различных трансформированных эксплантатов является хорошо известным в уровне техники (Weissbach et al., В: Methods for Plant Molecular Biology, Academic Press, San Diego, Calif, (1988)). Этот процесс регенерации и роста типично включает этапы селекции трансформированных клеток, культивирования этих точно определенных клеток через обычные стадии развития зародышей, через стадию укорененного ростка. Трансгенные зародыши и семена регенерируют аналогично. Полученные трансгенные укорененные побеги затем выращивают в соответствующей среде для роста растения, такой как почва.
Развитие или регенерация растений, содержащих инородный экзогенный ген, хорошо известна в уровне техники. Предпочтительно, регенерированные растения являются самоопыляющимися для обеспечения гомозиготных трансгенных растений. В ином случае, пыльцой, полученной из регенерированных растений, перекрестно опыляют выращенные из семян растения агротехнически важных линий. В свою очередь, пыльцу из растений этих важных линий используют для опыления регенерированных растений. Трансгенное растение по настоящему изобретению, содержащее желаемый полинуклеотид, культивируют с использованием способов, хорошо известных специалисту в данной области техники.
Способы для трансформации двудольных растений, в основном путем применения Agrobacterium tumefaciens, и получения трансгенных растений были опубликованы для хлопка (патент США №5004863, патент США №5159135, патент США №5518908), сои (патент США №5569834, патент США №5416011), Brassica (патент США №5463174), арахиса (Cheng et al., 1996) и гороха (Grant et al., 1995).
Способы для трансформации злаковых растений, таких как пшеница и ячмень, для введения генетического изменения в растение путем введения экзогенной нуклеиновой кислоты и для регенерации растений из протопластов или незрелых зародышей растения хорошо известны в уровне техники, смотри, например, патент Канады №2092588, патент Австралии №61781/94, патент Австралии №667939, патент США №6100447, РСТ заявку на патент США №97/10621, патент США №5589617, патент США №6541257, и другие способы приведены в публикации международной заявки WO 99/14314. Предпочтительно, трансгенные растения пшеницы или ячменя получают с помощью опосредованных Agrobacterium tumefaciens процедур трансформации. Векторы, несущие желаемый полинуклеотид, могут быть введены в способные к регенерации клетки пшеницы растений из культуры тканей или эксплантов, или подходящие растительные системы, такие как протопласты.
Способные к регенерации клетки пшеницы предпочтительно происходят из щитка незрелых зародышей, зрелых зародышей, каллюса, полученного из них или меристематической ткани.
Для подтверждения присутствия трансгенов в трансгенных клетках и растениях можно осуществить амплификацию с помощью полимеразной ценой реакции (ПЦР) или саузерн-блот анализ с использованием способов, известных специалистам в данной области техники. Продукты экспрессии трансгенов могут быть обнаружены с помощью любого из разнообразных путей в зависимости от природы продукта, и включающих вестерн-блоттинг и ферментный анализ. Одним особенно применимым путем количественного определения экспрессии белка и обнаружения репликации в различных тканях растения является применение репортерного гена, такого как GUS. Как только были получены трансгенные растения, их можно выращивать, чтобы получить ткани растения или части, имеющие желаемый фенотип.Ткань растения или части растения могут быть собраны и/или семена могут быть собраны. Семя может служить источником для выращивания дополнительных растений с тканями или частями, имеющими желаемые характеристики.
Трансгенное растение, образованное с применением Agrobacterium или других способов трансформации, типично содержит один генетический локус на одной хромосоме. Такие трансгенные растения могут быть названы гемизиготными по добавленному гену(ам). Более предпочтительным является трансгенное растение, которое является гомозиготным по добавленному гену(ам), иными словами, трансгенное растение, которое содержит два добавленных гена, один ген в одном и том же локусе на каждой хромосоме из пары хромосом. Гомозиготное трансгенное растение может быть получено путем самоопыления гемизиготного трансгенного растения, проращивания какого-либо из полученных семян и анализа полученных растений в отношении гена интереса.
Следует также понимать, что два различных трансгенных растения, которые содержат два независимо сегрегирующих экзогенных гена или локуса, могут также быть скрещены (спарены), чтобы получить потомков, которое содержат оба набора генов или локусов. Самоопыление соответствующего потомства F1 может приводить к получению растений, которые являются гомозиготными по обоим экзогенным генам или локусам. Возвратное скрещивание с родительским растением и ауткроссинг с нетрансгенным растением, также предусматриваются, как и вегетативное размножение. Описания других способов выведения, которые широко применяются для различных признаков и сельскохозяйственных культур, можно найти в Fehr, В. Breeding Methods for Cultivar Development, Wilcox J. ed., American Society of Agronomy, Madison Wis. (1987).
TILLING
В одном варианте осуществления TILLING (направленное воздействие индуцированных местных повреждений в геномах) можно использовать для получения растений, в которых эндогенные гены нокаутированы, например, гены, кодирующие DGAT, от-1-глицерол-3-фосфат-ацилтрансферазу (GPAT), 1-ацил-глицерол-3-фосфат-ацилтрансферазу (LPAAT), ацил-СоА: лизофосфатидилхол ин-ацилтрансферазу (LPCAT), фосфатазу фосфатидной кислоты (РАР) или комбинацию двух или более из них.
На первом этапе введенные мутации, такие как новые изменения одной пары оснований, индуцируют в популяции растений путем обработки семян (или пыльцы) с помощью химического мутагена, а затем выращивания растений до поколения, в котором мутации будут стабильно наследоваться. ДНК экстрагируют, и хранят семена от всех представителей популяции для создания ресурса, который может быть повторно оценен с течением времени.
Для анализа TILLING, праймеры для ПЦР конструируют для специфической амплификации одного целевого гена интереса. Специфичность является особенно важной, если мишень представляет собой представителя семейства генов или часть полиплоидного генома. Далее, меченные красителем праймеры можно использовать для амплификации продуктов ПЦР из объединенной ДНК от многочисленных особей. Эти продукты ПЦР денатурируются и повторно отжигаются, чтобы обеспечить образование некомплементарных пар оснований. Некомплементарности или гетеродуплексы, представляют как встречающийся в природе однонуклеотидный полиморфизм (SNP) (т.е., некоторые растения из популяции, вероятно, несут одинаковый полиморфизм), так и введенный SNP (т.е., только у редких отдельных растений, вероятно, проявляется мутация). После образования гетеродуплекса применение эндонуклеазы, такой как Cell, которая распознает и расщепляет некомплементарную ДНК, является ключом для открытия новых SNP внутри популяции при помощи TILLING.
С применением этого подхода, многие тысячи растений могут подвергаться скринингу для определения любой особи с изменением одного основания, а также небольших инсерций или делеций (1-30 п.о.) в любом гене или специфическом участке генома. Геномные фрагменты, подлежащие анализу, могут находиться где-либо в диапазоне размеров от 0,3 до 1,6 т.п.о. При 8-кратном объединении, фрагменты 1,4 т.п.о. (не учитывая концы фрагментов, где определение SNP является проблематичным из-за шума) и 96 дорожек на анализ, эта комбинация позволяет провести скрининг до миллиона пар оснований геномной ДНК за один анализ, что делает TILLING методикой с высокой производительностью.
TILLING дополнительно описано в Slade and Knauf (2005) и Henikoff et al. (2004).
В дополнение к обеспечению эффективного обнаружения мутаций, технология TILLING с высокой производительностью является идеальной для обнаружения природных полиморфизмов. Следовательно, детальное исследование неизвестной гомологичной ДНК с помощью образования гетеродуплексов с известной последовательностью выявляет количество и положение полиморфных сайтов. Определяются как нуклеотидные изменения, так и небольшие инсерций и делеций, включая по меньшей мере некоторые полиморфизмы по количеству повторов. Это было названо Ecotilling (Comai et al., 2004).
Каждый SNP регистрируется с помощью его приблизительного положения в пределах нескольких нуклеотидов. Таким образом, каждый гаплотип может быть заархивирован на основе его мобильности. Данные последовательностей могут быть получены с относительно небольшой пошаговой работой с использованием аликвот такой же амплифицированной ДНК, которую применяют для анализа расщепления некомплементарностей. Левый или правый секвенирующий праймер для одной реакции выбирают по его близости к полиморфизму. Программное обеспечение Sequencher осуществляет множественное выравнивание и находит изменение основания, которое в каждом случае подтверждается полосой геля.
Ecotilling может быть осуществлен более дешево, чем полное секвенирование, способ, который в настоящее время используется для нахождения большинства SNP. Планшеты, содержащие упорядоченные ДНК типа для Ecotilling, могут быть подвергнуты скринингу вместо объединений ДНК из мутировавших растений. Из-за того, что обнаружение происходит на гелях с разрешением близким к паре оснований и фоновые паттерны являются однородными для всех дорожек, полосы, которые имеют идентичный размер, могут быть выровнены, таким образом, обнаруживая и генотипируя SNP за один этап.Таким путем, окончательное секвенирование SNP является простым и эффективным, что в большей мере обусловлено тем фактом, что аликвоты одинаковых продуктов ПЦР, используемых для скрининга, могут быть подвергнуты секвенированию ДНК.
Усиление уровней экзогенной РНК и стабилизированная экспрессия
Посттранскрипционный сайленсинг гена (PTGS) представляет собой защитный механизм, специфический для нуклеотидной последовательности, который может направлять как клеточные, так и вирусные мРНК на деградацию. PTGS имеет место в растениях или грибах, стабильно или временно трансформированных при помощи рекомбинантного(ых) полинуклеотида(ов), и приводит к сниженному накоплению молекул РНК с последовательностью, сходной с введенным полинуклеотидом.
Уровни молекул РНК могут быть увеличены, и/или уровни РНК молекул стабилизируются через несколько поколений путем ограниченной экспрессии супрессора сайленсинга в запасающем органе растения или его части. Как используется в данном документе, "супрессор сайленсинга" представляет собой любой полинуклеотид или полипептид, который может экспрессироваться в растительной клетке, который усиливает уровень продукта экспрессии различного трансгена в растительной клетке, в частности, через много поколений от изначально трансформированного растения. В варианте осуществления супрессор сайленсинга представляет собой вирусный супрессор сайленсинга или его мутант.Большое количество вирусных супрессоров сайленсинга, известны в уровне техники и включают, но без ограничения, PI9, V2, Р38, Ре-Ро и RPV-P0. Примеры подходящих вирусных супрессоров сайленсинга включают те, которые описаны в публикации международной заявки WO 2010/057246. Супрессор сайленсинга может стабильно экспрессироваться в растении или его части по настоящему изобретению.
Как используется в данном документе, выражение "стабильно экспрессируется" или его варианты относится к уровню молекулы РНК, который по сути является таким же или более высоким в потомстве растений через много поколений, например, по меньшей мере три, по меньшей мере пять или по меньшей мере десять поколений, по сравнению с соответствующими растениями, где отсутствует экзогенный полинуклеотид, кодирующий супрессор сайленсинга. Однако, это выражение(я) не исключает возможность того, что через много поколений происходит некоторая потеря уровней молекулы РНК по сравнению с предыдущим поколением, например, не менее чем 10% потеря за поколение.
Супрессор можно выбрать из любого источника, например, растения, вируса, млекопитающего и т.д. Супрессор может представлять собой, например, следующее:
домашний вирус стада В2,
латентный вирус сциндапсуса Р14,
латентный вирус сциндапсуса AC2,
африканский вирус мозаики маниоки АС4,
возбудитель заболевания желтой прожилковой мозаики бамии C2,
возбудитель заболевания желтой прожилковой мозаики бамии C4,
возбудитель заболевания желтой прожилковой мозаики бамии DC 1,
вирус хлороза томата р22,
вирус хлороза томата CP,
вирус хлороза томата CPm,
вирус золотой мозаики томата AL2,
яванский вирус курчавости листьев томата βC1,
вирус желтой курчавости листьев томата V2,
китайский вирус желтой курчавости листьев томата С2,
китайский вирус желтой курчавости листьев томата Y10 изолят βC1,
израильский изолят желтой курчавости листьев томата V2,
вирус желтой мозаики золотистой фасоли - вигны AC2,
вирус хлоротической кольцевой пятнистости гибискуса CP,
вирус складчатости репы Р38,
вирус складчатости репы CP,
вирус мозаики цветной капусты P6,
вирус желтухи свеклы р21,
вирус тристецы цитрусовых p20,
вирус тристецы цитрусовых p23,
вирус тристецы цитрусовых CP, вирус мозаики китайской вигны SCP,
вирус хлоротической карликовости сладкого картофеля p22,
вирус мозаики огурца 2b,
вирус бессемянности томата НС-Pro,
вирус курчавости верхушки свеклы L2,
передающийся через почву вирус мозаики пшеницы 19К,
вирус Gammab полосатой мозаики ячменя,
полулатентный вирус Gammab мятлика,
очаговый пеклувирус арахиса Р15,
вирус карликовости риса Pns 10,
переносимый тлей вирус желтухи тыквы P0
вирус западной желтухи свеклы P0,
вирус картофеля X Р25,
вирус пожелтения жилок огурца P1b,
покс-вирус сливы НС-Рго,
вирус мозаики сахарного тростника НС-Pro,
вирус картофеля Y штамм НС-Pro,
вирус гравировки табака Р1/НС-Pro,
вирус мозаики репы Р1/НС-Pro,
вирус крапчатости ежи сборной P1,
вирус крапчатости ежи сборной - норвежский изолят P1,
вирус желтой крапчатости риса P1,
вирус желтой крапчатости риса - нигерийский изолят P1,
вирус белолистности риса NS3, вирус полосатости риса NS3,
инфицирующий крестоцветных вирус мозаики табака 126К,
инфицирующий крестоцветных вирус мозаики табака р122,
вирус мозаики табака р122,
вирус мозаики табака 126,
вирус мозаики табака 130К,
вирус погремковости табака 16К,
вирус кустистой карликовости томата Р19,
вирус бронзовости томата NSs,
вирус хлоротической пятнистости листьев яблока Р50,
вирус винограда A p10,
гомолог вируса скручивания листьев винограда-2 BYV р21, а также их варианты/мутанты. Список выше обеспечивает вирус, из которого может быть получен супрессор и белок (например, B2, P14 и т.д.), или обозначение кодирующего участка для супрессора из каждого конкретного вируса.
Можно использовать множественные копии супрессора. Различные супрессоры можно использовать вместе (например, в тандеме).
По сути, любая молекула РНК, которую необходимо экспрессировать в запасающем органе растения, может коэкспрессироваться с супрессором сайленсинга. Молекула РНК может влиять на агротехнический признак, устойчивость к насекомым, устойчивость к заболеваниям, устойчивость к гербицидам, стерильность, зерновые характеристики и т.п. Кодируемые полипептиды могут быть вовлечены в метаболизм липида, крахмала, углеводов, питательных веществ и т.д., или могут отвечать за синтез белков, пептидов, липидов, восков, крахмалов, сахаров, углеводов, вкусовых веществ, ароматических веществ, токсинов, каротиноидов, гормонов, полимеров, флавоноидов, запасных белков, фенольных кислот, алкалоидов, лигнинов, таннинов, целлюлоз, гликопротеинов, гликолипидов и т.д.
В конкретном примере растения, образующие повышенные уровни ферментов для образования липидов в растениях, таких как виды Brassica, например, масличный рапс, или подсолнечник, сафлор, лен, хлопок, соевые бобы или маис.
Получение неполярных липидов
Методики, которые обычно применяют на практике в уровне техники, могут использоваться для экстрагирования, обработки, очистки и анализа неполярных липидов, образованных клетками, организмами или их частями по данному изобретению. Такие методики описываются и объясняются в литературе в таких источниках, как Fereidoon Shahidi, Current Protocols in Food Analytical Chemistry, John Wiley & Sons, Inc. (2001) D1.1.1-D 1.1.11, и Perez-Vichetal (1998).
Получение масла семян
Типично, семена растений подвергают действию тепла, прессуют и/или экстрагируют для получения сырого масла семян, которое затем гидратируют, рафинируют, отбеливают и дезодорируют.Как правило, методики дробления семян известны в уровне техники. Например, семена масличных культур могут быть доведены до нужного состояния путем распыления на них воды для повышения содержания влаги до, например, 8,5%, и отвальцованы с помощью гладкого ролика с настройкой зазора 0,23-0,27 мм. В зависимости от типа семени воду перед дроблением можно не добавлять. Применение тепла деактивирует ферменты, облегчает дальнейший разрыв клеток, объединяет липидные капли и собирает белковые частицы, каждая из которых облегчает процесс экстракции.
Большая часть масла семян высвобождается при прохождении через винтовой пресс. Жмыхи, выталкиваемые из винтового пресса, затем экстрагируются растворителем, например, гексаном с использованием колонны с сопровождающим подогревом. Альтернативно, сырое масло семян, полученное посредством операции отжима, может пропускаться через бак-отстойник с дренажом из проволочных щелей в верхней части для удаления твердых частиц, которые отжимаются с маслом семян во время операции отжима. Осветленное масло семян может пропускаться через плиточно-рамный фильтр-пресс для удаления любых оставшихся мелких твердых частиц. При необходимости, масло семян, восстановленное в процессе экстракции, можно объединять с осветленным маслом семян для получения смешанного сырого масла семян.
Как только растворитель отгоняют из сырого масла семян, отпрессованную и экстрагированную части объединяют и подвергают обычным способам обработки липидов (т.е., гидратации, каустическому рафинированию, отбеливанию и дезодорации). Гидратация может выполняться путем добавления концентрированной фосфорной кислоты к сырому маслу семян для преобразования негидратируемых фосфатидов в гидратируемую форму и для хелатирования присутствующих побочных металлов. Осадок отделяют от масла семян путем центрифугирования. Масло семян можно рафинировать добавлением достаточного количества раствора гидроксида натрия, чтобы оттитровать все жирные кислоты и удалить образованные таким образом мыла.
Дезодорация может выполняться путем нагревания масла семян до 260°C под вакуумом и медленного введения пара в масло семян со скоростью около 0,1 мл/мин/100 мл масла семян. После около 30 минут барботирования маслу семян дают охладиться под вакуумом. Масло семян, как правило, переносят в стеклянный контейнер и продувают аргоном перед хранением при замораживании. Если количество масла семян ограничено, масло семян можно поместить под вакуум, например в реактор Parr, и нагреть до 260°C в течение того же периода времени, при котором его бы дезодорировали. Эта обработка улучшает цвет масла семян и удаляет большинство летучих веществ.
Растительная биомасса для получения липидов
Части растений, вовлеченные в фотосинтез (например, стебли и листья высших растений и водные растения, такие как водоросли), также могут использоваться для получения липидов. Независимо от типа растения есть несколько способов экстрагирования липидов из зеленой биомассы. Одним способом является физическая экстракция, при которой часто не применяется экстракция растворителем. Это "традиционный" способ с использованием нескольких различных типов механической экстракции. Экстракция прессованием на шнековом прессе является распространенным типом, как и способы экстракции на винтовом прессе и гидравлическом прессе. Количество липидов, экстрагированных с использованием этих способов, широко варьирует в зависимости от растительного материала и использованного механического процесса. Механическая экстракция, как правило, менее эффективна, чем экстракция растворителем, описанная ниже.
При экстракции растворителем органический растворитель (например, гексан) смешивают с по меньшей мере генетически модифицированной растительной зеленой биомассой, предпочтительно после того, как зеленая биомассу высушили и размололи. Конечно, другие части растения помимо зеленой биомассы (например, липид-содержащие семена) также могут размалываться и примешиваться. Растворитель растворяет липиды в биомассе и тому подобном, этот раствор затем отделяют от биомассы путем механического воздействия (например, с помощью процессов прессования выше). Этот этап разделения также может выполняться путем фильтрации (например, с помощью фильтр-пресса или аналогичного устройства) или центрифугирования и т.д. Органический растворитель может затем отделяться от неполярных липидов (например, дистилляцией). Этот второй этап разделения дает выход неполярных липидов из растения и может давать выход растворителя многократного использования, если используется обычное улавливание паров.
Получение водорослевого топлива
Культура водорослей представляет собой форму аквакультуры, включающую выращивание видов водорослей (включая микроводоросли, также называемые фитопланктоном, микрофиты, или планктонные водоросли, и макроводоросли, общеизвестные как морская водоросль). Виды водорослей, применимые в настоящем изобретении, включают, например, Chlamydomonas sp.(например, Chlamydomonas reinhardtii), Dunaliella sp., Haematococcus sp., Chlorella sp., Thraustochytrium sp., Schizochytrium sp.и Volvox sp.
Коммерческое и промышленное культивирование водорослей имеет множество применений, включая получение ингредиентов пищи, продуктов питания и водорослевого топлива.
Моно или смешанные водорослевые культуры могут культивироваться в открытых водоемах (таких как пруды и озера типа подводящих каналов) или фотобиореакторах.
Водоросли можно собирать с использованием микросит, центрифугированием, флокуляцией (с использованием, например, хитозана, квасцов и хлорида железа) и посредством пенной флотации. Прерывание подвода диоксида углерода может приводить к тому, что водоросли флокулируют сами по себе, что называется "автофлокуляция". При пенной флотации человек, который проводит культивирование, аэрирует воду в пену и затем снимает водоросли с поверхности. Ультразвук и другие способы сбора находятся в настоящее время в стадии разработки.
Липиды можно выделять из водорослей путем механического дробления. Когда водоросль сушат, она сохраняет свое липидное содержание, которое затем может "отжиматься" маслоотжимным прессом. Поскольку различные штаммы водорослей широко варьируют по своим физическим свойствам, различные конфигурации пресса (винтовой, шнековый, поршневый и т.д.) лучше работают для конкретных типов водорослей.
Осмотический шок иногда используется для высвобождения клеточных компонентов, таких как липиды, из водорослей. Осмотический шок представляет собой резкое снижение осмотического давления и может вызывать разрывы клеток в растворе.
Ультразвуковая экстракция может ускорять процессы экстракции, в частности, процессы ферментативной экстракции, использованные для экстрагирования липидов из водорослей. Ультразвуковые волны применяются для создания кавитационных пузырьков в материале растворителя. Когда эти пузырьки схлопываются вблизи клеточных стенок, полученные в результате ударные волны и струи жидкости вызывают разламывание этих клеточных стенок и высвобождение их содержимого в растворитель.
Химические растворители (например, гексан, бензол, петролейный эфир) часто применяют при экстракции липидов из водорослей. Экстракцию Сокслета можно применять для экстрагирования липидов из водорослей путем многократного промывания, или перколяции, органическим растворителем при нагревании с обратным холодильником в специальной стеклянной посуде.
Для экстрагирования ферментов из водорослей может применяться ферментативная экстракция. В ферментативной экстракции применяются ферменты для деградации клеточных стенок с помощью воды, действующей в качестве растворителя. Ферментативную экстракцию можно поддерживать обработкой ультразвуком.
Сверхкритический CO2 также может быть использован в качестве растворителя. В этом способе С02 сжижается под давлением и нагревается до точки, при которой он становится сверхкритическим (имеющим свойства как жидкости, так и газа), что позволяет ему действовать как растворитель.
Процессы ферментации для получения липидов
Как используется в данном документе, выражение "процесс ферментации" относится к любому процессу ферментации или любому процессу, включающему этап ферментации. Процесс ферментации включает без ограничения процессы ферментации, применяемые для получения спиртов (например, этанола, метанола, бутанола), органических кислот (например, лимонной кислоты, уксусной кислоты, итаконовой кислоты, молочной кислоты, глюконовой кислоты), кетонов (например, ацетона), аминокислот (например, глутаминовой кислоты), газов (например, H2 и CO2), антибиотиков (например, пенициллина и тетрациклина), ферментов, витаминов (например, рибофлавина, бета-каротина) и гормонов. Процессы ферментации также включают процессы ферментации, применяемые в производстве потребляемых спиртных напитков (например, пива и вина), молочной промышленности (например, ферментированных молочных продуктов), кожевенной промышленности и табачной промышленности. Предпочтительные процессы ферментации включают процессы ферментации для получения спирта, как хорошо известные из уровня техники. Предпочтительные процессы ферментации представляют собой процессы анаэробной ферментации, как хорошо известной из уровня техники. Подходящие ферментирующие клетки, как правило микроорганизмы, которые способны ферментировать, то есть преобразовывать, сахара, такие как глюкоза или мальтоза, прямо или опосредовано, в желаемый продукт ферментации. Примеры ферментирующих микроорганизмов включают грибковые организмы, такие как дрожжи, предпочтительно, жировой организм. Как используется в данном документе, "жировой организм" представляет собой организм, накапливающий по меньшей мере 25% от его сухого веса в качестве триглицеридов. Как используется в данном документе, "дрожжи" включает Saccharomyces spp., Saccharomyces cerevisiae, Saccharomyces carl'bergensis, Candida spp., Kluveromyces spp., Pichia spp., Hansenula spp., Trichoderma spp., Lipomyces starkey и Yarrowia lipolytica. Предпочтительные дрожжи включают Yarrowia lipolytica или другие жировые дрожжи и штаммы Saccharomyces spp., и в частности Saccharomyces cerevisiae.
Трансгенный микроорганизм предпочтительно выращивается в условиях, оптимизирующих активность генов биосинтеза жирных кислот и генов ацилтрансферазы. Это приводит к образованию наибольшего и самого экономически выгодного выхода липидов. В основном, условия среды, которые могут оптимизироваться, включают тип и количество источника углерода, тип и количество источника азота, соотношение углерод-к-азоту, уровень кислорода, рост температуры, рН, длину фазы образования биомассы, длину фазы накопления липидов и время сбора урожая клеток.
Среда ферментации должна содержать подходящий источник углерода. Подходящие источники углерода могут включать без ограничения: моносахариды (например, глюкозу, фруктозу), дисахариды (например, лактозу, сахарозу), олигосахариды, полисахариды (например, крахмал, целлюлозу или их смеси), сахарные спирты (например, глицерин) или смеси из возобновляемого сырья (например, фильтрата сырной сыворотки, кукурузного экстракта, свеклосахарной мелассы, ячменного солода). Кроме того, источники углерода могут включать алканы, жирные кислоты, сложные эфиры жирных кислот, моноглицериды, диглицериды, триглицериды, фосфолипиды и различные коммерческие источники жирных кислот, в том числе растительные масла (например, соевое масло) и животные жиры. Кроме того, углеродный субстрат может включать одноуглеродные субстраты (например, диоксид углерода, метанол, формальдегид, формиат, углеродсодержащие амины), для которых было показано метаболическое преобразование в ключевые биохимические промежуточные соединения. Следовательно, предполагается, что источник углерода, используемый в настоящем изобретении, может охватывать широкое разнообразие углеродсодержащих субстратов и будет ограничиваться только выбором микроорганизма-хозяина. Хотя все из вышеупомянутых углеродных субстратов и их смесей, как ожидается, будут подходящими в настоящем изобретении, предпочтительными углеродными субстратами являются сахара и/или жирные кислоты. Наиболее предпочтительной является глюкоза и/или жирные кислоты, содержащие 10-22 атомов углерода.
Азот может поставляться из неорганического (например, (NH4)2SO4) или органического источника (например, мочевина, глутамат). В дополнение к соответственным источникам углерода и азота среда ферментации может также содержать подходящие минералы, соли, кофакторы, буферы, витамины и другие компоненты, известные специалистам в данной области техники, подходящие для роста микроорганизмов и стимуляции ферментативных путей, необходимых для образования липидов.
Подходящий диапазон рН для ферментации находится, как правило, от около pH 4,0 до pH 8,0, где pH 5,5-pH 7,0 предпочтителен в качестве диапазона для начальных условий роста. Ферментация можно осуществлять в аэробных или анаэробных условиях, где микроаэробные условия являются предпочтительными.
Как правило, накопление высоких уровней липидов в клетках жировых микроорганизмов требует двухстадийного процесса, поскольку метаболическое состояние должно быть "сбалансировано" между ростом и синтезом/накопление жиров. Таким образом, наиболее предпочтительно для получения липидов в микроорганизмах необходим двухстадийный процесс ферментации. В этом подходе первая стадия ферментации предназначается для образования и накопления клеточной массы и характеризуется быстрым клеточным ростом и делением клеток. На второй стадии ферментации предпочтительно создать условия азотного голодания в культуре для содействия высоким уровням накопления липидов. Эффектом этого азотного голодания является снижение эффективной концентрации AMP в клетках, таким образом, снижается активность NAD-зависимой изоцитратдегидрогеназы митохондрий. В этом случае будет накапливаться лимонная кислота, при этом образуя обильные запасы ацетил-CoA в цитоплазме и служа затравкой для синтеза жирных кислот. Таким образом, эта фаза характеризуется прекращением клеточного деления с последующим синтезом жирных кислот и накоплением TAG.
Хотя клетки, как правило, выращивают при около 30°C, некоторые исследования показали повышенный синтез ненасыщенных жирных кислот при более низких температурах. На основании экономики процесса этот температурный сдвиг должен происходить, вероятно, после первой фазы двухстадийной ферментации, когда основная часть роста микроорганизмов уже произошла.
Предполагается, что множество схем процессов ферментации могут применяться, если желательным является коммерческое получение липидов с использованием нуклеиновых кислот настоящего изобретения. Например, коммерческое получение липидов из рекомбинантного микробного хозяина может производиться в периодическом, периодическом с подпиткой или непрерывном процессе ферментации.
Периодический процесс ферментации представляет собой закрытую систему, где состав среды устанавливается в начале процесса и не подвергается дополнительным добавлениям помимо тех, которые требуются для поддержания рН и уровня кислорода во время процесса. Таким образом, в начале процесса культивирования среду засевают желаемым организмом, и росту или метаболической активности дают возможность происходить без добавления дополнительных субстратов (т.е. источников углерода и азота) к среде. В периодических процессах составы метаболитов и биомассы системы постоянно меняются вплоть до времени, когда культивирование завершают. В типичном периодическом процессе клетки сдерживаются в течение статической лаг-фазы до лог-фазы с высоким ростом и, наконец, в стационарной фазе, где скорость роста уменьшается или останавливается. Оставленные без обработки клетки в стационарной фазе в конечном итоге погибнут. Разновидностью стандартного периодического процесса является периодический процесс с подпиткой, где субстрат постоянно добавляют в ферментер в течение процесса ферментации. Периодический процесс с подпиткой также подходит для настоящего изобретения. Периодические процессы с подпиткой применимы, когда катаболитная репрессия склонна ингибировать метаболизм клеток, или если желательно иметь ограниченное количество субстрата в среде в любой момент времени. Измерение концентрации субстрата в периодических системах с подпиткой является сложным и, следовательно, она может оцениваться на основании изменений измеримых факторов, таких как рН, растворенный кислород и парциальное давление газообразных отходов (например, СО2). Периодический и периодический с подпиткой способы культивирования являются общими и хорошо известны из уровня техники, и примеры можно найти в Brock, In Biotechnology: A Textbook of Industrial Microbiology, 2.sup.nd ed., Sinauer Associates, Sunderland, Mass., (1989) или Deshpande and Mukund (1992).
Коммерческое получение липидов с использованием клеток настоящего изобретения также может осуществляться посредством непрерывного процесса ферментации, где определенную среду непрерывно добавляют в биореактор, и в то же время равное количество объема культуры одновременно удаляют для восстановления продукта. В непрерывных культурах клетки преимущественно поддерживаются в лог-фазе роста при постоянной плотности клеток. Способы непрерывных или полунепрерывных культур дают возможность модулирования одного фактора или любого числа факторов, которые влияют на клеточный рост или концентрацию конечного продукта. Например, в одном подходе можно ограничивать источник углерода и позволить всем остальным параметрам уменьшить метаболизм. В других системах ряд факторов, влияющих на рост, можно менять непрерывно, в то время как концентрация клеток, измеряемая по мутности среды, поддерживается постоянной. Непрерывные системы стремятся поддерживать рост в стационарном состоянии, и таким образом скорость роста клеток должна быть в равновесии с потерей клеток из-за отведения среды из культуры. Способы модулирования питательных веществ и факторов роста для процессов непрерывной культуры, а также методики максимизации скорости образования продукта хорошо известны из уровня техники промышленной микробиологии, и многочисленные способы подробно приводятся в Brock, выше.
Жирные кислоты, включая PUFA, могут обнаруживаться в микроорганизме-хозяине в виде свободных жирных кислот или в этерифицированных формах, таких как ацилглицеролы, фосфолипиды, сульфолипиды или гликолипиды, и могут экстрагироваться из клетки-хозяина посредством множества способов, хорошо известных из уровня техники.
В основном, способы очистки жирных кислот, включая PUFA, могут включать экстракцию органическими растворителями, обработку ультразвуком, экстракцию сверхкритической жидкостью (например, с использованием диоксида углерода), омыление и физические способы, такие как отжимы, или их комбинации. Особый интерес вызывает экстракция с метанолом и хлороформом в присутствии воды (Bligh and Dyer, 1959). При желании водный слой можно окислить, чтобы протонировать отрицательно заряженные фрагменты, и таким образом увеличить распределение желаемых продуктов в органический слой. После экстракции органические растворители можно удалять путем выпаривания под струей азота. При выделении в конъюгированных формах продукты могут расщепляться ферментативно или химически для высвобождения свободной жирной кислоты или менее комплексного конъюгата интереса, и могут затем подвергаться дополнительным манипуляциям для получения желаемого конечного продукта. Желательным является, что конъюгированные формы жирных кислот расщепляют гидроксидом калия.
Если необходима дополнительная очистка, могут использоваться стандартные способы. Такие способы могут включать экстракцию, обработку мочевиной, фракционированную кристаллизацию, ВЭЖХ, фракционную перегонку, хроматографию на силикагеле, высокоскоростное центрифугирование или дистилляцию, или комбинацию этих методик. Защита реакционноспособных групп, таких как кислотная или алкенильная группы, может выполняться на любом этапе посредством известных методик (например, алкилирования, йодирования). Применяемые способы включают метилирование жирных кислот для получения метиловых сложных эфиров. Подобным образом, защитные группы могут удаляться на любом этапе. Желательно, очистка фракций, содержащих GLA, STA, ARA, DHA и ЕРА, может выполняться с помощью обработки мочевиной и/или фракционной перегонкой.
Применения липидов
Липиды, полученные описанными способами, имеют множество применений. В некоторых вариантах осуществления липиды применяют в качестве пищевых масел. В других вариантах осуществления липиды рафинируют и применяют в качестве смазочных средств или для других промышленных применений, таких как синтез пластмасс.В некоторых предпочтительных вариантах осуществления липиды рафинируют для получения биодизеля.
Биодизель
Получение биодизеля, или алкиловых сложных эфиров, хорошо известно. Существует три основных пути получения сложных эфиров из липидов: 1) катализированная щелочью трансэтерификация липида спиртом; 2) катализированная кислотой прямая этерификация липида метанолом и 3) преобразование липидов в жирные кислоты, и затем в алкиловые сложные эфиры с помощью кислотного катализа.
В некоторых предпочтительных вариантах осуществления липиды трансэтерифицируют для получения метиловых сложных эфиров и глицерина. В некоторых предпочтительных вариантах осуществления липиды реагируют со спиртом (таким как метанол или этанол) в присутствии катализатора (гидроксида калия или натрия) для получения алкиловых сложных эфиров. Алкиловые сложные эфиры можно применять для биодизеля или смешивать с топливом на основе бензина.
Пищевые продукты
Настоящее изобретение включает композиции, которые можно применять в качестве пищевых продуктов. Для целей настоящего изобретения "пищевые продукты" включают любую пищу или препарат для потребления человеком или животным (в том числе для энтерального и/или парентерального потребления), которые при приеме в организм: (1) служат для питания или построения ткани или снабжения энергией, и/или (2) сохраняют, восстанавливают или поддерживают адекватное состояние питания или метаболическую функцию. Пищевые продукты настоящего изобретения включают пищевые композиции для младенцев и/или маленьких детей.
Пищевые продукты по настоящему изобретению включают, например, клетку настоящего изобретения, растение настоящего изобретения, часть растения настоящего изобретения, семя настоящего изобретения, экстракт настоящего изобретения, продукт способа настоящего изобретения, продукт процесса ферментации настоящего изобретения или композицию вместе с подходящим носителем(ями). Выражение "носитель" применяется в его самом широком смысле, чтобы охватить любой компонент, который может иметь или не иметь питательную ценность. Как будет понятно специалисту в данной области техники, носитель должен подходить для применения (или применяться в достаточно низкой концентрации) в пищевом продукте, чтобы он не имел вредного эффекта на организм, который потребляет пищевой продукт.
Пищевой продукт настоящего изобретения включает липид, полученный прямо или опосредованно с помощью применения способов, клеток или организмов, раскрытых в данном документе. Композиция может находиться или в твердой, или в жидкой форме. Дополнительно, композиция может включать съедобные макроэлементы, витамины и/или минералы в количествах, желаемых для конкретного применения. Количества этих ингредиентов будет меняться в зависимости от того, предназначается ли композиция к применению у обычных лиц, или к применению у лиц, имеющих определенные потребности, таких как лица, страдающие от метаболических расстройств и т.п.
Примеры подходящих носителей с питательной ценностью включают без ограничений макроэлементы, такие как съедобные жиры, углеводы и белки. Примеры таких съедобных жиров включают без ограничений кокосовое масло, масло бурачника, грибное масло, масло черной смородины, соевое масло, а также моно- и диглицериды. Примеры таких углеводов включают без ограничений глюкозу, съедобную лактозу и гидролизованный крахмал. Дополнительно, примеры белков, которые можно использовать в пищевой композиции настоящего изобретения включают без ограничений соевые белки, электродиализную сыворотку, электродиализное обезжиренное молоко, молочную сыворотку или гидролизаты этих белков.
Что касается витаминов и минералов, к композициям пищевого продукта настоящего изобретения можно добавлять следующее: кальций, фосфор, калий, натрий, хлорид, магний, марганец, железо, медь, цинк, селен, йод, витамины A, Е, D, C и комплекса B. Также можно добавлять другие такие витамины и минералы.
Компоненты, используемые в композициях пищевого продукта настоящего изобретения, могут быть полуочищенного или очищенного происхождения. Под полуочищенным или очищенным подразумевается материал, который был получен с помощью очистки природного материала или путем синтеза de novo.
Композиции пищевого продукта настоящего изобретения также можно добавлять в пищу, даже если дополнение диеты не требуется. Например, композицию можно добавлять в пище любого типа, включая без ограничений маргарин, модифицированное сливочное масло, сыры, молоко, йогурт, шоколад, конфеты, закуски, салатные масла, кулинарные масла, кулинарные жиры, мясо, рыбу и напитки.
Род Saccharomyces spp.применяется как в пивоварении и виноделии, так и в качестве средства при выпечке, в частности хлеба. Дрожжи представляют собой одну из основных составляющих растительных экстрактов. Дрожжи также применяются в качестве добавки в корм для животных. Будет очевидно, что можно обеспечивать генетически модифицированные штаммы дрожжей, которые адаптированы к синтезу липидов, как описано в данном документе. Эти штаммы дрожжей можно применять в производстве продуктов питания, вина и пива для обеспечения продуктов с повышенным содержанием липидов.
Дополнительно, липиды, полученные в соответствии с настоящим изобретением, или клетки-хозяева, трансформированные так, чтобы содержать и экспрессировать генов-объектов, также могут применяться в качестве добавок пищи животных для изменения состава жирных кислот тканей или молока животного на более желательные для потребления человеком или животными. Примеры таких животных включают овец, крупный рогатый скот, лошадей и тому подобное.
Кроме того, пищевые продукты настоящего изобретения могут применяться в аквакультуре для повышения уровней жирных кислот в рыбе для потребления человеком или животными.
Предпочтительные пищевые продукты настоящего изобретения представляют собой растения, семена и другие части растений, такие как листья, плоды и стебли, которые могут применяться непосредственно в качестве пищи или корма для людей или других животных. Например, животные могут непосредственно пастись на таких растениях, выращенных в поле, или их кормят более рассчитанными количествами при контролируемом кормлении. Изобретение включает в себя применение таких растений и частей растений в качестве корма для увеличения уровней полиненасыщенных жирных кислот у человека и других животных.
Композиции
Настоящее изобретение также охватывает композиции, в частности, фармацевтические композиции, включающие один или несколько липидов, полученных с использованием способов настоящего изобретения.
Фармацевтическая композиция может включать один или несколько липидов в комбинации со стандартным известным нетоксичным фармацевтически приемлемым носителем, вспомогательным веществом или наполнителем, таким как фосфатно-буферный солевой раствор, вода, этанол, полиолы, растительные масла, смачивающим средством или эмульсией, такой как эмульсия вода/масло. Композиция может находиться в жидкой или твердой форме. Например, композиция может быть в форме таблетки, капсулы, жидкости для приема внутрь, порошка, местной мази или крема. Надлежащая текучесть может поддерживаться, например, с помощью поддержания требуемого размера частиц в случае дисперсий, и с применением поверхностно-активных веществ. Желательным может быть также включение изотонических средств, например, Сахаров, хлорида натрия и т.п.Кроме таких инертных разбавителей композиция может также включать вспомогательные вещества, такие как смачивающие средства, эмульгирующие и суспендирующие средства, подсластители, вкусовые вещества и ароматизирующие средства.
Суспензии в дополнение к активным соединениям могут включать суспендирующие средства, такие как этоксилированные изостеариловые спирты, полиоксиэтиленсорбит и сложные эфиры сорбита, микрокристаллическую целлюлозу, метагидроксид алюминия, бентонит, агар-агар и трагакант или смеси этих веществ.
Твердые лекарственные формы, такие как таблетки и капсулы, можно получать с использованием методик, хорошо известных из уровня техники. Например, липиды, полученные в соответствии с настоящим изобретением, могут таблетировать с традиционными таблеточными основами, такими как лактоза, сахароза и кукурузный крахмал, в сочетании со связующими веществами, такими как аравийская камедь, кукурузный крахмал или желатин, разрыхлителями, такими как картофельный крахмал или альгиновая кислота, и смазками, такими как стеариновая кислота или стеарат магния. Капсулы можно получать включением этих наполнителей в желатиновую капсулу вместе с антиоксидантами и соответствующим липидом(ами).
Для внутривенного введения липиды, полученные в соответствии с настоящим изобретением, или их производные могут включаться в коммерческие составы.
Типичная дозировка для определенной жирной кислоты составляет от 0,1 мг до 20 г, принимаемая от одного до пяти раз в день (до 100 г ежедневно), и предпочтительно в диапазоне от около 10 мг до около 1, 2, 5, или 10 г ежедневно (принимаемая в одной или многократных дозах). Как известно из уровня техники, желательным является как минимум около 300 мг/день жирной кислоты, главным образом, полиненасыщенной жирной кислоты. Однако следует принимать во внимание, что любое количество жирной кислоты будет полезным для субъекта.
Возможные пути введения фармацевтических композиций настоящего изобретения включают, например, энтеральный и парентеральный. Например, жидкий препарат можно вводить перорально. Дополнительно, гомогенную смесь можно полностью диспергировать в воде, смешать в стерильных условиях с физиологически приемлемыми разбавителями, консервантами, буферами или газами-вытеснителями для образованием аэрозоля или средства для ингаляций.
Дозировка композиции, которая будет вводиться субъекту, может определяться специалистом в данной области техники и зависит от различных факторов, таких как вес, возраст, общее состояние здоровья, анамнез, иммунный статус и т.д. субъекта.
Дополнительно, композиции настоящего изобретения можно использовать для косметических целей. Композиции можно добавлять к уже существующим косметическим композициям, так что образуется смесь, или жирная кислота, полученная в соответствии с настоящим изобретением, может применяться в качестве единственного "активного" ингредиента в косметической композиции.
Полипептиды
Выражения "полипептид" и "белок", как правило, применяются взаимозаменяемо.
Полипептид или класс полипептидов может определяться по степени идентичности (% идентичности) его аминокислотной последовательности с эталонной аминокислотной последовательностью, или по наличию большего % идентичности с одной эталонной аминокислотной последовательностью, чем с другой. % идентичности полипептида с эталонной аминокислотной последовательностью, как правило, определяется с помощью GAP-анализа (Needleman and Wunsch, 1970; программа GCG) с параметрами штраф за внесение гэпа=5 и штраф за продолжение гэпа=0,3. Запрашиваемая последовательность составляет по меньшей мере 100 аминокислот в длину, и GAP-анализ выравнивает две последовательности по области по меньшей мере 100 аминокислот. Еще более предпочтительно, запрашиваемая последовательность составляет по меньшей мере 250 аминокислот в длину, и GAP-анализ выравнивает две последовательности по области по меньшей мере 250 аминокислот. Еще более предпочтительно, GAP-анализ выравнивает две последовательности по их полной длине. Полипептид или класс полипептидов может иметь такую же ферментативную активность как, или иную активность чем, или отсутствие активности эталонного полипептида. Предпочтительно, полипептид имеет ферментативную активность по меньшей мере 10% активности эталонного полипептида.
Как используется в данном документе, "биологически активный фрагмент" представляет собой часть полипептида настоящего изобретения, который сохраняет определенную активность эталонного полипептида полной длины, например, MGAT активность. Биологически активные фрагменты, как используется в данном документе, исключают полипептид. Биологически активные фрагменты могут быть частью любого размера до тех пор, пока они сохраняют определенную активность. Предпочтительно, биологически активный фрагмент сохраняет, по меньшей мере, 10% активности полипептида полной длины.
В отношении определенного полипептида или фермента следует понимать, что количественные данные % идентичности выше, чем предоставленные в данном документе, будут содержать предпочтительные варианты осуществления. Таким образом, когда это применимо, в свете минимальных количественных данных % идентичности, предпочтительным является, что полипептид/фермент включает аминокислотную последовательность, которая по меньшей мере на 60%, более предпочтительно по меньшей мере на 65%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 91%, более предпочтительно по меньшей мере на 92%, более предпочтительно по меньшей мере на 93%, более предпочтительно по меньшей мере на 94%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 96%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%, более предпочтительно по меньшей мере на 99,1%, более предпочтительно по меньшей мере на 99,2%, более предпочтительно по меньшей мере на 99,3%, более предпочтительно по меньшей мере на 99,4%, более предпочтительно по меньшей мере на 99,5%, более предпочтительно по меньшей мере на 99,6%, более предпочтительно по меньшей мере на 99,7%, более предпочтительно по меньшей мере на 99,8% и еще более предпочтительно по меньшей мере на 99,9% идентична соответственной предлагаемой SEQ ID NO.
Мутанты аминокислотных последовательностей полипептидов, определяемых в данном документе, можно получать введением подходящих нуклеотидных замен в нуклеиновую кислоту, определенную в данном документе, или путем синтеза in vitro желаемого полипептида. Такие мутанты имеют в своем составе, например, делении, вставки или замены остатков в пределах аминокислотной последовательности. Можно создавать комбинацию делеций, вставок и замен, чтобы получить конечный конструкт, при условии, что конечный полипептидный продукт обладает желаемыми характеристиками.
Мутантные (измененные) полипептиды можно получать с использованием любой методики, известной из уровня техники, например, с использованием стратегий направленной эволюции или рационального конструирования (см. ниже). Продукты, полученные из мутантной/измененной ДНК, можно легко подвергнуть скринингу с применением методик, описанных в данном документе, для определения обладают ли они ацилтрансферазной активностью, например, MGAT, DGAT, или GPAT/фосфатазной активностью.
При конструировании мутантов аминокислотных последовательностей местоположение сайта мутации и природа мутации будут зависеть от характеристики(характеристик), которую следует модифицировать. Сайты для мутации можно модифицировать по отдельности или последовательно, например, путем (1) замены вначале вариантами консервативных аминокислот и затем более радикальными выборами в зависимости от достигнутых результатов, (2) делеции целевого остатка или (3) вставки других остатков рядом с определенным сайтом.
Делеции аминокислотной последовательности обычно варьируют от около 1 до 15 остатков, более предпочтительно от около 1 до 10 остатков и типично от около 1 до 5 последовательных остатков.
Мутанты с заменами имеют по меньшей мере один удаленный аминокислотный остаток в полипептиде и другой остаток, вставленный на его место. Сайты, представляющие наибольший интерес для замещающего мутагенеза, включают сайты, определенные как активный сайт(ы). Другими сайтами интереса являются те, в которых определенные остатки, полученные из различных штаммов или видов, являются идентичными. Эти положения могут быть важны для биологической активности. Эти сайты, особенно входящие в последовательность из по меньшей мере трех других одинаково консервативных сайтов, предпочтительно замещают относительно консервативным образом. Такие консервативные замены показаны в таблице 1 с заголовком "Иллюстративные замены".
В предпочтительном варианте осуществления мутантный/вариантный полипептид имеет только, или не больше одной или двух, или трех, или четырех консервативных аминокислотных замен по сравнению с встречающимся в природе полипептидом. Подробная информация о консервативных аминокислотных заменах предоставлена в таблице 1. Как известно специалисту, можно обоснованно предположить, что такие незначительные замены не меняют активность полипептида при экспрессии в рекомбинантной клетке.

Направленная эволюция
В направленной эволюции к белку применяется случайный мутагенез, и применяется режим отбора для выбора вариантов, которые имеют желаемые качества, например, увеличенную ацилтрансферазную активность. Затем применяются дополнительные раунды мутации и отбора. Типичная стратегия направленной эволюции состоит из трех этапов:
1) Диверсификация: Ген, кодирующий белок интереса, мутируют и/или рекомбинируют случайным образом для создания большой библиотеки вариантов гена. Библиотеки вариантных генов можно конструировать с помощью ПЦР с внесением ошибок (см., например, Leung, 1989; Cadweli and Joyce, 1992), из пулов фрагментов, расщепленных ДНКазой 1, полученных из родительских матриц (Stemmer, 1994а; Stemmer, 1994b; Crameri et at., 1998; Coco et al., 2001), из вырожденных олигонуклеотидов (Ness et al., 2002, Coco, 2002) или из смесей обоих, или даже из нерасщепленных родительских матриц (Zhao et al., 1998; Eggert et at., 2005; Jezequek et al., 2008), и как правило собранных с помощью ПЦР. Библиотеки также можно получить из родительских последовательностей, рекомбинированных in vivo или in vitro путем гомологичной или негомологичной рекомбинации (Ostermeier et al., 1999; Volkov et at., 1999; Sieber et at., 2001). Библиотеки вариантных генов можно также конструировать путем субклонирования гена интереса в подходящий вектор, трансформирования вектора в "мутаторный" штамм, такой как Е. coli XL-1 red (Stratagene), и размножения трансформированной бактерии в течение подходящего числа поколений. Библиотеки вариантных генов можно также конструировать, подвергая ген интереса ДНК-шаффлингу (т.е., in vitro гомологичная рекомбинация пулов отобранных мутантных генов путем случайной фрагментации и повторной сборки), как в целом описано Harayama (1998).
2) Отбор: Библиотеку тестируют на присутствие мутантов (вариантов), обладающих желательными свойствами, используя сортировку или отбор. Сортировки дают возможность идентификации и выделения высокопроизводительных мутантов вручную, в то время как отбор автоматически удаляет все нефункциональные мутанты. Сортировка может включать скрининг на присутствие известных консервативных аминокислотных мотивов. Альтернативно, или дополнительно, сортировка может включать экспрессирование мутированного полинуклеотида в организме-хозяине или его части и анализ уровня ацилтрансферазной активности с помощью, например, количественного определения уровня полученного в результате продукта в липиде, экстрагированном из организма или его части, и определения уровня продукта в экстрагированном липиде из организма или его части по сравнению с соответствующим организмов или его частью, где отсутствует мутированный полинуклеотид, и необязательно, экспрессирования родительского (немутированного) полинуклеотида. Альтернативно, сортировка может включать кормление организма или его части меченым субстратом и определение уровня субстрата или продукта в организме или его части по сравнению с соответствующим организмом или его частью, где отсутствует мутированный полинуклеотид, и необязательно, экспрессирование родительского (немутированного) полинуклеотида.
3) Амплификация: Варианты, идентифицированные при отборе или сортировке, многократно реплицируют, давая возможность исследователям секвенировать их ДНК для того, чтобы понять, какие мутации произошли.
Вместе эти три этапа называются "раундом" направленной эволюции. Большинство экспериментов будет включать более одного раунда. В этих экспериментах "победителей" предыдущего раунда диверсифицируют в следующем раунде для создания новой библиотеки. В конце эксперимента все превращенные белковые или полинуклеотидные мутанты характеризуются с использованием биохимических способов.
Рациональное конструирование
Белок может конструироваться рационально на основе известной информации о структуре и укладке белка. Это можно выполнять путем конструирования с нуля (de novo конструирование) или путем реконструирования на основе природных каркасов (см., например, Hallinga, 1997; Lu and Berry, Protein Structure Design and Engineering, Handbook of Proteins 2, 1153-1157 (2007)). Конструирование белка, как правило, включает идентификацию последовательностей, которые складываются в данную или целевую структуру, и может выполняться с применением компьютерных моделей. Вычислительные алгоритмы конструирования белка отыскивают последовательность-конформационное пространство для последовательностей, которые обладают наименьшей энергией при укладке в целевую структуру. Вычислительные алгоритмы конструирования белка применяют модели энергетики белка для оценки того, как мутации будут влиять на структуру и функцию белка. Эти функции энергии, как правило, включают комбинацию молекулярной механики, статистические (т.е. основанные на знаниях) и другие эмпирические показатели. Подходящее доступное программное обеспечение включает IPRO (Interative Protein Redesign and Optimization), EGAD (A Genetic Algorithm for Protein Design), Rosetta Design, Sharpen и Abalone.
Также в объем настоящего изобретения включены полипептиды, определенные в данном документе, которые неодинаково модифицированы во время или после синтеза, например, с помощью биотинилирования, бензилирования, гликозилирования, ацетилирования, фосфорилирования, амидирования, дериватизации известныхми защитными/блокирующими группами, протеолитического расщепления, связывания с молекулой антитела или другими клеточными лигандами и т.д. Эти модификации могут служить для повышения стабильности и/или биоактивности полипептида настоящего изобретения.
Идентификация ацилтрансфераз
В одном аспекте настоящее изобретение обеспечивает способ идентификации молекулы нуклеиновой кислоты, кодирующей ацилтрансферазу с повышенной способностью к образованию MAG, DAG и/или TAG в клетке.
Способ включает получение клетки, содержащей молекулу нуклеиновой кислоты, кодирующую ацилтрансферазу, функционально связанную с промотором, который активен в клетке. Молекула нуклеиновой кислоты может кодировать встречающуюся в природе ацилтрансферазу, такую как MGAT, GPAT и/или DGAT, или их мутанта(ов). Мутанты могут разрабатываться с использованием стандартных процедур из уровня техники (см. выше), как например, осуществляя случайный мутагенез, нацеленный мутагенез или насыщающий мутагенез известных генов интереса или подвергая различные гены ДНК-шаффлингу. Например, полинуклеотид, содержащий последовательность, выбранную из любой из SEQ ID NO: 1-44, кодирующей MGAT, можно мутировать и/или рекомбинировать случайным образом для создания большой библиотеки вариантов гена (мутантов) с использованием, например, ПЦР с внесением ошибок и/или ДНК-шаффлинга. Мутанты могут отбираться для дальнейшего исследования на основании того, что они содержат консервативный аминокислотный мотив. Например, в случае кандидатной нуклеиновой кислоты, кодирующей MGAT, специалист может определить, содержит ли она последовательность, предоставляемую в SEQ ID NO:220, 221, 222, 223 и/или 224, перед тестированием того, кодирует ли нуклеиновая кислота функциональный MGAT-мутант (с помощью, например, трансфекции в клетку-хозяина, такую как растительная клетка, и анализа на ацилтрансферазную (т.е. MGAT) активность, как описано в данном документе). Прямое ПЦР секвенирование нуклеиновой кислоты или ее фрагмента может применяться для определения точной нуклеотидной последовательности и выведения соответствующей аминокислотной последовательности, и тем самым идентификации консервативных аминокислотных последовательностей. Вырожденные праймеры исходя из консервативных аминокислотных последовательностей могут применяться для прямой ПЦР-амплификации. Вырожденные праймеры также могут применяться в качестве зондов в анализах ДНК-гибридизации. Альтернативно, консервативную аминокислотную последовательность(и) можно выявлять в анализах гибридизации белка, в которых применяется, например, антитело, которое специфично связывается с консервативной аминокислотной последовательностыо(ями), или субстрат, который специфически связывается с консервативной аминокислотной последовательностью(ями), как например, липид, который связывает FLXLXXXN (предполагаемый домен связывания нейтральных липидов; SEQ ID NO: 224).
В одном варианте осуществления молекула нуклеиновой кислоты содержит последовательность нуклеотидов, кодирующую MGAT. Последовательность нуклеотидов может i) содержать последовательность, выбранную из любой из SEQ ID NO:l-44, ii) кодировать полипептид, содержащий аминокислоты с последовательностью, как представлено в любой из SEQ ID NO:45-82, или их биологически активный фрагмент, iii) быть по меньшей мере на 50% идентичной с i) или ii), или iv) гибридизироваться с любой из i) до iii) при жестких условиях. В другом или дополнительном варианте осуществления молекула нуклеиновой кислоты содержит последовательность нуклеотидов, кодирующую одну или несколько консервативных DGAT2 и/или MGAT 1/2 аминокислотных последовательностей, как представлено в SEQ ID NO:220, 221, 222, 223 и 224. В предпочтительном варианте осуществления молекула нуклеиновой кислоты содержит последовательность нуклеотидов, кодирующую консервативные аминокислотные последовательности, предоставленные в SEQ ID NO:220 и/или SEQ ID NO:224.
В другом варианте осуществления молекула нуклеиновой кислоты содержит последовательность нуклеотидов, кодирующую GPAT, предпочтительно GPAT, которая имеет фосфатазную активность. Последовательность нуклеотидов может
i) содержать последовательность, выбранную из любой из SEQ ID NO:84-141,
ii) кодировать полипептид, содержащий аминокислоты с последовательностью, как представлено в любой из SEQ ID NO:144-201, или их биологически активный фрагмент,
iii) быть по меньшей мере на 50% идентичной с i) или ii), или
iv) гибридизироваться с любой из i) до iii) при жестких условиях.
В другом или дополнительном варианте осуществления молекула нуклеиновой кислоты содержит последовательность нуклеотидов, кодирующую одну или несколько консервативных аминокислотных последовательностей GPAT, как представлено в SEQ ID NO:225, 226 и 227, или последовательность аминокислот, которая по меньшей мере на 50%, предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 65% идентична с ней.
В другом варианте осуществления молекула нуклеиновой кислоты содержит последовательность нуклеотидов, кодирующую DGAT2. Последовательность нуклеотидов может i) содержать последовательность нуклеотидов, выбранную из любой из SEQ ГО N0:204-211,
ii) кодировать полипептид, содержащий аминокислоты с последовательностью, как представлено в любой из SEQ ID NO:212-219, или их биологически активный фрагмент,
iii) быть по меньшей мере на 50% идентичной с i) или ii), или
iv) гибридизироваться с любой из i) до iii) при жестких условиях.
В предпочтительном варианте осуществления DGAT2 содержит последовательность нуклеотидов из SEQ ID NO:204 и/или последовательность нуклеотидов, кодирующую полипептид, содержащий аминокислоты с последовательностью, как представлено в SEQ ID NO:212.
Клетку, содержащую молекулу нуклеиновой кислоты, кодирующую ацилтрансферазу, функционально связанную с промотором, который активен в клетке, можно получить с использованием стандартных процедур из уровня техники, как например, введением молекулы нуклеиновой кислоты в клетку с помощью, например, осаждения фосфатом кальция, обработкой полиэтиленгликолем, электропорацией и комбинациями этих обработок. Другие способы трансформации клеток также могут применяться, и они включают без ограничений введение ДНК в растения путем прямого переноса или инъекции ДНК. Трансформированные растительные клетки также можно получить с использованием способов переноса и стимуляции, опосредованного Agrobacterium, как описано в данном документе.
Способ дополнительно включает определение того, увеличивается ли уровень образуемых в клетке MAG, DAG и/или TAG при сравнении с соответствующей клеткой, где отсутствует эта нуклеиновая кислота, с использованием известных методик из уровня техники, подобных тем, которые проиллюстрированы в примере 1.
Например, липиды можно экстрагировать в растворе хлороформа/метанола, высушивать и разделять с помощью тонкослойной хроматографии (TCL). Идентификацию TAG, DAG, MAG, свободной жирной кислоты и других липидов можно подтверждать с помощью внутренних стандартов липидов после окрашивания парами йода. Полученные в результате хроматограммы можно анализировать с помощью Phosphorlmager, и количественно определять количество MAG, DAG и TAG на основе известного количества внутренних стандартов, или альтернативно, клетки можно снабжать sn-2 моноолеилглицеролом[14С] или [bС]глицерол-3-фосфатом, и связанная радиоактивность определяется количественно с помощью жидкостного сцинтилляционного счетчика (т.е. количественно определяется количество меченого MAG, DAG и TAG).
Способ дополнительно включает идентификацию молекулы нуклеиновой кислоты, кодирующей ацилтрансферазу с повышенной способностью производить MAG, DAG и/или TAG в клетке. В предпочтительном варианте осуществления ацилтрансфераза катализирует ферментативную реакцию в MGAT пути. В еще одном предпочтительном варианте осуществления DAG увеличивается с помощью MGAT пути (например, MGAT катализирует ацилирование MAG ацил-coAc образованием DAG). В другом или дополнительном варианте осуществления субстрат MAG образуется с помощью GPAT, которая также имеет фосфатазную активность, и/или DAG ацилируется ацил-coAc помощью DGAT и/или MGAT с DGAT активностью с образованием TAG.
ПРИМЕРЫ
Пример 1. Общие материалы и способы
Экспрессия генов в растительных клетках в системе временной экспрессии
Гены были экспрессированы в растительных клетках с использованием системы временной экспрессии в целом как описано Voinnet et al. (2003) и Wood et al. (2009). Бинарные векторы, содержащие кодирующую область, подлежащую экспрессии с помощью сильного конститутивного промотора e35S, содержащего дуплицированную энхансерную область, вводили в штамм AGL1 Agrobacterium tumefaciens. Химерный бинарный вектор 35S:pl9 для экспрессии вирусного молчащего супрессора р 19 отдельно вводили в AGL1, как описано в WO2010/057246. Рекомбинантные клетки выращивали до стационарной фазы при 28°C в жидкой среде LB с добавлением 50 мг/л канамицина и 50 мг/л рифампицина. Затем бактерии осадили с помощью центрифугирования при 5000 g в течение 5 мин. при комнатной температуре перед тем, как ресуспендировать до OD600=1,0 в инфильтрационном буфере, содержащем 10 мМ MES pH 5,7, 10 мМ MgCl2 и 100 мкМ ацетосирингона. Затем клетки инкубировали при 28°C с встряхиванием в течение 3 часов, после чего измеряли OD600 и объем каждой культуры, включая 35S:pl9, требуемый для достижения конечной концентрации OD600=0,125, вносили в чистую пробирку. Конечный объем приготавливали с помощью вышеупомянутого буфера. Затем культуральной смесью инфильтровали листья, и растения, как правило, выращивали в течение дополнительных трех-пяти дней после инфильтрации до того, как собирали листовые диски либо для получения очищенных клеточных лизатов, либо выделения общего липида. Контрольные инфильтрации производили только со штаммом 35S:pl9.
Анализ очищенных листовых лизатов
Ткани листа Nicotiana benthamiana, ранее инфильтрованного, как описано выше, измельчали в растворе, содержащем 0,1 М калий-фосфатного буфера (рН 7,2) и 0,33 М сахарозы, с использованием стеклянного гомогенизатора. Гомогенат листьев центрифугировали при 20000 g в течение 45 минут при 4°C, после чего собирали каждую надосадочную жидкость. Содержание белка в каждой надосадочной жидкости измеряли согласно Bradford (1976) с использованием многофункционального счетчика Wallac 1420 и красящего реагента от Bio-Rad Protein Assay (Bio-Rad Laboratories, Геркулес, Калифорния, США). В анализах на ацилтрансферазу использовали 100 мкг белка согласно Cao et al. (2007) с некоторыми модификациями. Реакционная среда содержала 100 мМ Tris-HCl (pH 7,0), 5 мМ MgCl2, 1 мг/мл BSA (без жирных кислот), 200 мМ сахарозы, 40 мМ холодного олеил-СоА, 16,4 мкМ sn-2 моноолеилглицерола[14С] (55 мКи/ммоль, American Radiochemicals, Сэнт Луис, Монтана, США) или 6,0 мкМ [14C]глицерол-3-фосфата (G-3-P) динатриевой соли (150 мКи/ммоль, American Radiochemicals). Анализы проводили в течение 7,5, 15, или 30 минут.
Анализ липидов
Анализ липидов из анализов листовых лизатов
Липиды из анализов лизатов экстрагировали с использованием хлороформа:метанола:0,1 М KCl (2:1:1) и выделяли. Различные классы липидов в образцах разделяли на пластинах для тонкослойной хроматографии (TCL) Silica gel 60 (MERCK, Дермштадт, Германия), пропитанных 10% борной кислотой. Система растворителей, применяемая для фракционирования TAG из липидного экстракта, состояла из хлороформа/ацетона (90/10 об./об.). Индивидуальные классы липидов визуализировали, подвергая пластины действию паров йода, и идентифицировали путем прогона параллельных аутентичных стандартов на той же пластине TCL. На пластины воздействовали люминесцентными визуализирующими сортировками в течение ночи и анализировали с помощью люминесцентного визуализатора Fujifilm FLA-5000 перед тем, как количественно определять DPM с помощью жидкостного сцинтиляционного подсчета.
Выделение и фракционирование общего липида
Ткани лиофилизировали, взвешивали и экстрагировали общие липиды, как описано Bligh and Dyer (1959). Различные классы липидов в каждом образце разделяли на пластинах для TCL Silica gel 60. Система растворителей, применяемая для фракционирования нейтральных липидов (NL) и полярных липидов (PL), состояла из гексана/диэтилового эфира/уксусной кислоты (70/30/1 об./об./об.). Индивидуальные классы липидов визуализировали, подвергая пластины действию паров йода, и идентифицировали путем прогона параллельных аутентичных стандартов на той же пластине для TCL.
Для определения жирнокислотного состава липидных образцов получали метиловые сложные эфиры жирной кислоты (FAME) из NL, включающие TAG, DAG и/или MAG, и фракции PL путем извлечения соответствующих полос из пластин для TCL и инкубирования их в растворе метанол/HCl/дихлорметан (10/1/1 об./об./об.) в течение 2 часов при 80°C вместе с известным количеством гексадекановой кислоты в качестве внутреннего стандарта. FAME экстрагировали в гексане/дихлорметане, концентрировали до небольшого объема в гексане и впрыскивали в газовый хроматограф.
Количество индивидуальных и общих жирных кислот (TFA), присутствующее в липидных фракциях, определяли количественно с помощью определения площади под каждым пиком и рассчитывали путем сравнения с площадью пика для известного количества внутреннего стандарта.
Капиллярная газо-жидкостная хроматография (GC)
FAME анализировали с помощью газовой хроматографии (GC) с использованием газового хроматографа Agilent Technologies 6890N (Пало Альто, Калифорния, США), оснащенного капиллярной колонкой из кварцевого стекла Equity™-1 (15 м×0,1 мм i.d., 0,1 мкм толщина пленки), FID, инжектором с разделением/без разделения потока и автоматическим пробоотборником и инжектором Agilent Technologies 7683 Series. В качестве газа-носителя применялся гелий. Образцы впрыскивали в режиме без деления потока при температуре термостата 120°C. После впрыскивания температуру термостата повышали до 270°C при 10°C.мин-1 и, наконец, до 30°C при 5°C.мин-1.
Пики определяли количественно с помощью программного обеспечения Agilent Technologies ChemStation (Rev В.03.01 (317), Пало Альто, Калифорния, США).
Анализ DGAT в H1246 Saccharomvces cerevisiae
Штамм H1246 Saccharomyces cerevisiae полностью лишен DGAT активности и в нем отсутствуют TAG и сложные эфиры стерола в результате нокаутных мутаций в четырех генах (DGA1, LROl, ARE1, ARE2). Добавление свободной жирной кислоты (например, 1 мМ 18:1Δ9) к ростовой среде Н1246 является токсичным в отсутствии DGAT активности. Рост на такой среде, следовательно, можно применять в качестве индикатора или отбора на присутствие DGAT активности в этом штамме дрожжей.
HI246 S. cerevisiae трансформировали конструктом pYES2 (отрицательный контроль), конструктом, кодирующим DGAT1 Arabidopsis thaliana в pYES2, или конструктом, кодирующим MGAT2 Mus musculus в pYES2. Трансформанты снабжали [14С] 18:1Δ9 свободными жирными кислотами.
В отдельном эксперименте HI246 S. cerevisiae трансформировали конструктом pYES2 (отрицательный контроль), конструктом, кодирующим DGAT1 Bernadia pulchella в pYES2, или конструктом, кодирующим MGAT1 М. musculus в pYES2, и снабжали 18:1Δ9 свободными жирными кислотами. Штамм дикого типа S288C S. cerevisiae, трансформированный pYES2, служил в качестве положительного контроля.
Дрожжевые трансформанты ресуспендировали в стерильной воде mQ и разбавляли до OD600=1. Образцы дополнительно разбавляли в четырех последовательных разведениях, 1/10 каждое. 2 мкл каждого разведения помещали на каждой из чашек (YNBD, YNBG, YNBG+FA) вместе с 2 мкл воды mQ и 2 мкл нетрансформированной суспензии клеток HI246 (OD600=1). Чашки инкубировали в течение 6 дней при 30°C перед тем, как оценить рост.
Среда в чашках, 40 мл среды на чашку
- YNBD: минимальная отсеивающая среда, где отсутствует урацил и содержащая 2% глюкозы, 0,01% NP40 и 100 мкл этанола.
- YNBG: минимальная отсеивающая среда, где отсутствует урацил и содержащая 2% галактозы, 1% раффинозы, 0,01% NP40 и 100 мкл этанола.
- YNBG+FA: минимальная отсеивающая среда, где отсутствует урацил и содержащая 2% галактозы, 1% раффинозы, 0,01% NP40 и 1 мМ С18:1Л9, растворенной в 100 мкл этанола.
Пример 2. Конститутивная экспрессия моноацилглицерол ацилтрансферазы в растительных клетках
MGAT1
Активность фермента моноацилглицерол ацилтрансферазы 1 (MGAT1), кодируемого геном из М.musculus (Yen et al., 2002), и диацилглицерол ацилтрансферазы (DGAT1) A.thaliana (Bouvier-Nave et al., 2000), использованной здесь в качестве сравнения с MGAT1, демонстрировали в ткани листьев N.benthamiana с применением системы временной экспрессии, как описано в примере 1.
Вектор, обозначенный 35S-pORE04, получали путем вставки фрагмента PstI, содержащего промотор 35S, в сайт SfoI вектора pORE04 после обработки Т4 ДНК-полимеразой для получения тупых концов (Coutu et al., 2007). Химерную ДНК, кодирующую MGAT1 М. musculus, кодон-оптимизированную для Brassica napus, синтезировали с помощью Geneart и обозначили 0954364_MGAT_pMA. Химерную ДНК, обозначенную 35S:MGAT1 и кодирующую MGAT1 М.musculus (№доступа Genbank Q91ZV4), для экспрессии в растительных клетках, получали путем вставки полной кодирующей области из 0954364_MGAT_pMA, содержащейся в пределах фрагмента EcoRI, в 35S-pORE04 в сайт EcoRl. Вектор, содержащий конструкт 35S:MGAT1, обозначили pJP3184. Аналогично, химерную ДНК 35S:DGAT1, кодирующую DGAT1 A. thaliana (№доступа Genbank AAF19262), для экспрессии в растительных клетках, создали путем вставки полной кодирующей области из pXZP513E, содержащейся в пределах фрагмента BatHl-EcoRN, в 35S-pORE04 в сайт ВатШ-EcoKV. Вектор, содержащий конструкт 35S:DGAT1, обозначили pJP2078.
Химерные векторы вводили в штамм AGL1 A.tumefaciens, и клетки из этих культур инфильтровали в ткань листьев растений N. benthamiana в вегетационном домике при 24°C. Чтобы обеспечить возможность прямого сравнения между образцами и для снижения межлистовых вариаций образцы, подлежащие сравнению, инфильтровали с одной из двух сторон одного листа. Эксперименты выполняли в трех повторностях. После инфильтрации растения выращивали в течение дополнительных трех дней перед тем, как листовые диски собирали, лиофилизировали, и липиды, экстрагированные из образцов, фракционировали и определяли количественно, как описано в примере 1. Этот анализ показал, что гены MGAT1 и DGAT1 были функциориющими в отношении повышения уровней масла в листьях у N.benthamiana, как изложено ниже.
Ткань листьев, трансформированная только конструктом 35S:pl9 (отрицательный контроль), содержала в среднем 4 мкг свободной жирной кислоты (FFA), полученной из DAG/мг сухого веса листьев, и 5 мкг FFA, полученной из TAG/мг сухого веса листьев. Ткань листьев, трансформированная конструктами 35S:pl9 и 35S:DGAT1 (контроль на экспрессию DGAT1), содержала в среднем 4 мкг FFA, полученной из DAG/мг сухого веса листьев, и 22 мкг FFA, полученной из TAG/мг сухого веса листьев. Ткань листьев, трансформированная конструктами 35S:pl9 и 35S:MGAT1, содержала в среднем 8 мкг FFA, полученной из DAG/мг сухого веса листьев, и 44 мкг FFA, полученной из TAG/мг сухого веса листьев. Ткань листьев, трансформированная конструктами 35S:pl9, 35S:DGAT1 и 35S:MGAT1, не содержала уровни DAG или TAG выше тех, которые наблюдались при инфильтрации 35S:pl9 и 35S:MGAT1 (фигура 2). Кроме того, снижение уровня насыщенных соединений в семенах отмечалось после экспрессии MGAT при сравнении как с контролем р19, так и с образцами DGAT1 (таблица 2).
Данные, описанные выше, показали, что фермент MGAT1 был намного более активным, чем фермент DGAT1, в стимуляции накопления в ткани листьев как DAG, так и TAG. Экспрессия гена MGAT1 приводила к вдвое большему накоплению TAG и DAG в ткани листьев по сравнению с тем, если экспрессировалась DGAT1. Этот результат был очень удивительным и неожиданным, учитывая то, что MGAT является ферментом, который экспрессируется в мышином кишечнике, совершенно иной биологической системе, нежели растительные листья. Это исследование было первой демонстрацией эктопической экспрессии MGAT в растительных клетках.
Образцы листьев, инфильтрованные MGAT1 М.musculus, накапливали вдвое больше DAG и TAG по сравнению с тканью листьев, которые инфильтровали только DGAT1 A.thaliana. Эффективность образования TAG также была удивительной и неожиданной, принимая во внимание то, что мышиная MGAT обладала очень низкой активностью в качестве DGAT. Ткань листьев, инфильтрованная генами, кодирующими одновременно MGAT1 и DGAT1, не накапливала существенно больше TAG, чем образец листьев только с MGAT1. Фигура 1 является представлением различных путей накопления TAG, большинство из которых сходятся к DAG, центральной молекуле в синтезе липидов. Например, MAG, DAG и TAG могут взаимопревращаться при посредстве различных ферментных активностей, включая трансацилирования, липазную, MGAT, DGAT и PDAT. Снижение уровней насыщенных соединений также было отмечено после экспрессии MGAT.
MGAT2
Химерную ДНК, обозначенную 35S:MGAT2 и кодирующую MGAT2 М.Musculus, для экспрессии в растительных клетках получали путем вставки полной кодирующей области MGAT2, содержавшейся в пределах фрагмента EcoRl, в 35S-pORE04 в сайт EcoRl. Активность фермента моноацилглицерол ацилтрансферазы 2 (MGAT2), кодируемой геном из М.musculus (Yen, 2003) (№доступа Genbank Q80W94), и DGAT1 A.thaliana (Bouvier-Nave et al., 2000), используемой здесь в качестве сравнения с MGAT2, также демонстрировали в ткани листьев N. benthamiana с использованием системы временной экспрессии, как описано в примере 1.
По сравнению с контролями экспрессия DGAT1 повышала TAG в листьях в 5,9 раза, MGAT2 - в 7,3 раза, а комбинация MGAT2+DGAT1 - в 9,8 раза (фигура 3). Способность MGAT2 отдельно давать такие значительные повышения TAG была неожиданной по ряду причин. Во-первых, количество субстрата MAG, присутствующее в ткани листьев, как известно, является низким, и большие повышения в накоплении TAG из этого субстрата не могли ожидаться. Во-вторых, добавление только MGAT активности (т.е., добавление MGAT2, которая не имеет DGAT активности), как ожидалось, даст DAG, а не TAG, особенно в среде листа, где, как правило, присутствует небольшая естественная DGAT активность.
Обсуждение
Изобретателями настоящего изобретения, к удивлению, было продемонстрировано, что трансгенная экспрессия гена MGAT приводит в результате к значительным повышениям выхода липидов в растительных клетках. Изобретатели настоящего изобретения сделали вывод, что Tumaney et al. выделили DGAT с некоторой MGAT активностью, и что они не достигли успеха в попытках клонировать ген, кодирующий MGAT, как определено в данном документе. Tumaney et al. (2001) сообщали о MGAT активности в арахисе и выделили фермент, ответственный за эту активность. Однако, Tumaney et al. не опубликовали результаты тестов на DGAT активность и, следовательно, создается впечатление, что описанный фермент был DGAT с некоторой MGAT активностью. Действительно, предыдущая работа не смогла идентифицировать никакой MGAT активности у других видов (Stobart et al., 1997). Кроме того, удивительно, что фермент, выделенный Tumaney et al., был растворимым цитозольным ферментом, а не связанным с мембраной ферментом. Наконец, хотя Tumaney et al. позднее опубликовали тезисы, утверждающие, что они выделили гены MGAT из арахиса (Rajasekharan et al., 2006) и Arabidopsis (Ghosh et al., 2006), статей, описывающих выделение этих генов, когда-либо опубликовано не было.
Пример 3. Биохимическое обнаружение трансгенной MGAT активности в экстрактах листьев
Клеточные лизаты получали из ткани листьев N. benthamiana, которые были инфильтрованы 35S:MGAT1, 35S:MGAT2 и 35S:DGAT1, как описано в примере 1. Выполняли инфильтрации отдельных листьев, каждую в трех повторностях, штаммами, содержащими только конструкт 35S:p 19 (отрицательный контроль), штаммом 35S:MGAT2 вместе с штаммом 35S:pl9, и смесью 35S:MGAT2 и 35S:DGAT1 штаммов Agrobacterium с штаммом 35S:pl9. Образцы в трех повторностях собирали через три дня, и очищенные клеточные лизаты получали путем механического лизиса тканей и центрифугирования. MGAT активности очищенных клеточных лизатов сравнивали путем снабжения лизатов [14C]MAG, как описано в примере 1. Данные показаны на фигуре 4.
В контрольном образце 35S:pl9 наблюдалась небольшая MGAT активность, так как большая часть радиоактивности оставалась в MAG на протяжении анализа. В отличие от этого большая часть меченого MAG в образце 35S:MGAT2 быстро превратилась в DAG (фигура 4, центральная панель), что указывает на сильную активность MGAT, экспрессированную из конструкта 35S:MGAT2. Кроме того, также образовалось значительное количество TAG. Образование TAG, наблюдавшееся в образце 35S:MGAT2, происходило, вероятно, вследствие нативной DGAT активности N. benthamiana, или он образовался с помощью другого пути синтеза TAG. Количество образования TAG значительно повышалось при дополнительном добавлении 35S:DGAT1 (фигура 4, правая панель), что указывает, что фермент MGAT2 образовывал DAG, который был доступен для превращения в TAG с помощью DGAT1 в вегетативных тканях растений.
Пример 4. Биохимическое обнаружение образования MAG, доступного для MGAT, в экстрактах листьев
В анализах in vitro, описанных в примере 3 с использованием лизатов листьев, субстраты (sn-2 MAG и олеил-СоА) обеспечивались экзогенно в связи с тем, что in vivo MGAT активность в интактных растительных тканях требовала бы нативного присутствия этих субстратов. Присутствие низких уровней MAG в различных растительных тканях уже сообщалось ранее (Hirayama and Hujii, 1965; Panekina et al., 1978; Lakshminarayana et al., 1984; Perry & Harwood, 1993). Чтобы протестировать, может ли MGAT2 иметь доступ к MAG, образованному с помощью нативных путей растения, вышеприведенный эксперимент повторили, но в этот раз со снабжением лизатов [I4C]G-3-P. Полученные в результате данные показаны схематически на фигуре 5.
Образование меченого MAG наблюдалось во всех образцах, что указывает на de novo образование MAG из G-3-P в лизатах листьев растений. Меченые DAG и TAG продукты также наблюдались во всех образцах, хотя они были относительно низки в контрольном образце 35S:pl9, что указывает на то, что образование этих нейтральных липидов с помощью эндогенного пути Кеннеди было относительно низким в этом образце. В отличие от этого, большая часть метки в MGAT2 и MGAT2+DGAT1 образцах оказалось в пулах DAG и TAG, что указывает на то, что экзогенно добавленная MGAT катализирует превращение MAG, который был образован из меченого G-3-P, с помощью нативных путей растения.
Примеры 2-4 показывают несколько ключевых моментов: 1) ткань листьев может синтезировать MAG из G-3-P таким образом, что MAG доступен для экзогенной MGAT, экспрессированной в ткани листьев; 2) даже MGAT, которая происходит из кишечника млекопитающих, может функционировать в тканях растений, которые, как известно, не обладают эндогенной MGAT, при этом требуется успешное взаимодействие с другими растительными факторами, вовлеченными в синтез липидов; 3) DAG, образованный с помощью экзогенной MGAT активности, доступен для растительной DGAT, или экзогенной DGAT, для образования TAG; и 4) экспрессия экзогенной MGAT может давать значительное повышение уровней TAG в тканях растений, уровней, которые по меньшей мере настолько же высокие, как полученные с помощью экзогенной экспрессии DGAT1 A.thaliana.
Пример 5. Экспрессия DGAT1, MGAT1 и MGAT2 в дрожжах
Химерные дрожжевые векторы экспрессии конструировали путем вставки генов, кодирующих DGAT1 A. thaliana, MGAT1 М. musculus и MGAT2 М. musculus в вектор pYES2, что дает pYES2:DGATl, pYES2:MGATl и pYES2:MGAT2. Эти конструкта трансформировали в штамм HI246 Saccharomyces cerevisiae, который полностью лишен DGAT активности и в нем отсутствуют TAG и сложные эфиры стерола в результате нокаутных мутаций в четырех генах (DGA1, LROl, ARE1, ARE2). Штамм дрожжей HI246 способен синтезировать DAG из экзогенно добавленных жирных кислот, но не способен превращать DAG в TAG из-за нокаутных мутаций. Трансформированные дрожжевые культуры снабжали [|4С]18:1Δ9 перед тем, как экстрагировали и фракционировали общие липиды с помощью TCL, как описано в примере 1. Авторадиограмма характерной пластины для TCL показана на фигуре 6.
Образование TAG, указывающее на присутствие DGAT активности, наблюдали для клеток дрожжей, содержащих как DGAT1 (положительный контроль), так и MGAT1 млекопитающих, но не в клетках, содержащих MGAT2. Отсюда сделан вывод, что MGAT1 из мыши также обладала DGAT активностью в клетках дрожжей, и следовательно функционировала как фермент с двойной функцией MGAT/DGAT, в то время как MGAT2 не имела обнаруживаемой DGAT активности и была, следовательно, исключительно MGAT.
Пример 6. Экспрессия моноацилглицерол ацилтрансферазы в растениях, семенах и грибах
Экспрессия MGAT 1 в семенах Arabidopsis thaliana
Ген, кодирующий MGAT1 М. musculus и находящийся под контролем семя-специфического промотора (FP1, усеченный промотор напина из Brassica napus) применяли для создания стабильно трансформированных растений A. thaliana и семян потомства. Вектор, обозначенный pJP3174, получали путем вставки фрагмента Sail, содержащего сайт EcoRl, фланкированный промотором FP1 и лектиновым сигналом полиаденилирования Glycine max, в сайт Sall-Xhol вектора pCW141. Вектор pCW141 также содержал управляемый FP1, прерванный интроном, секретируемый семенами GFP в качестве подлежащего сортировке маркерного гена. Химерный ген, обозначенный FP1:MGAT1-GFP, получали путем вставки полной кодирующей области из конструкта 0954364JVlGAT_pMA, содержавшейся в пределах фрагмента EcoRl, в pJP3174 в сайте EcoRl, что создает pJP3179. Этот химерный вектор вводили в штамм AGL1 A.tumefaciens, и клетки из культуры трансформированной Agrobacterium использовали для обработки растений A. thaliana (экотип Columbia), применяя для трансформации способ погружения цветков (Clough and Bent, 1998). После созревания семена от обработанных растений просматривали под бинокулярным микроскопом Leica MZFLIII и ртутной лампой ebq 100. Выделили пятнадцать трансгенных семян (сильно GFP-положительных) и пятнадцать нетрансгенных (GFP-отрицательных) семян, и каждый набор объединяли. GFP-положительный и GFP-отрицательные пулы анализировали на содержание общей жирной кислоты, как описано в примере 1. Этот анализ представил среднее содержания и состава жирных кислот для семян, трансформированных с помощью конструкта MGAT, но в популяции, которая может содержать как гемизиготные, так и гомозиготные трансформированные семена.
Этот анализ показал, что ген MGAT1 был функционирующим в отношении повышения уровней масла семян в семенах A.Thaliana, при этом пятнадцать нетрансгенных семян (контроль, такой же, как дикий тип) содержат в среднем 69,4 мкг общих жирных кислот, тогда как пятнадцать трансгенных семян, трансформированных геном GFP, и которые, следовательно, вероятно содержат генетический конструкт FP1:MGAT1, содержали в среднем 71,9 мкг общих жирных кислот.Это составило повышение содержания масла на 3,5% по сравнению с контролем (дикий тип). Анализ также показал, что ген MGAT был функционирующим в отношении обогащения семян полиненасыщенными жирными кислотами, как видно из жирнокислотного состава общего экстрагированного липида, полученного из семян. В частности, количество ALA, представленное как процент общей жирной кислоты, экстрагированной из семян, повысилось с 16,0 до 19,6%. Аналогично, процент жирной кислоты 20:2п6 повысился с 1,25% до 1,90% и жирной кислоты 20:ЗпЗ повысился с 0,26% до 0,51% (таблица 2).
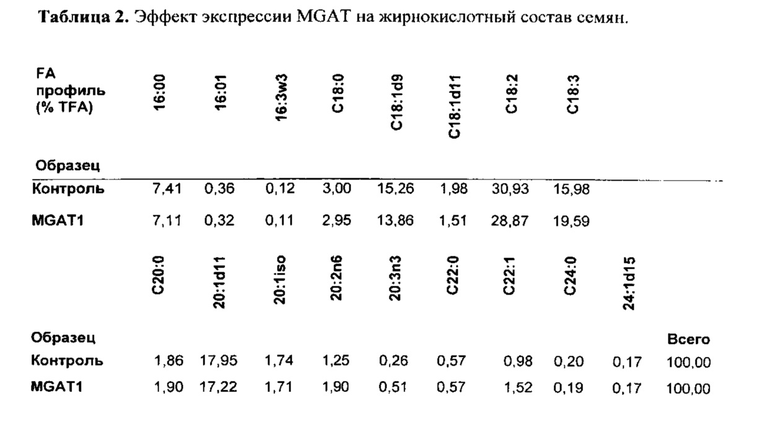
Выполнялся дополнительный эксперимент, где химерную ДНК FPLMGAT1-GFP модифицировали с удалением гена GFP. Эту генетический конструкт, обозначенный FP1:MGAT1, трансформировали в линию A.thaliana, которая была мутантной по FAD2. Общее содержание жирных кислот в семенах Т2 от резистентных к антибиотику Т[растений, а также родительских линий, выращенных рядом с этими растениями, определяли согласно примеру 1. Данные показаны в таблице 3. Среднее общих жирных кислот в семенах из контрольных линий составило 347,9 мкг/100 семян, в то время как среднее трансгенных семян составило 381,0 мкг/100 семян. Когда данные для контрольной линии С6 исключили из определения среднего, среднее для контроля составило 370 мкг/100 семян. Содержание масла в трансгенных семенах показало увеличение на около 3% в относительном выражении по сравнению с содержанием масла в ^трансформированных семенах.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, замещают MGAT1 в конструктах, упомянутых выше, и вводят в Arabidopsis. В семенах от полученных в результате трансгенных растений содержание масла повышено.
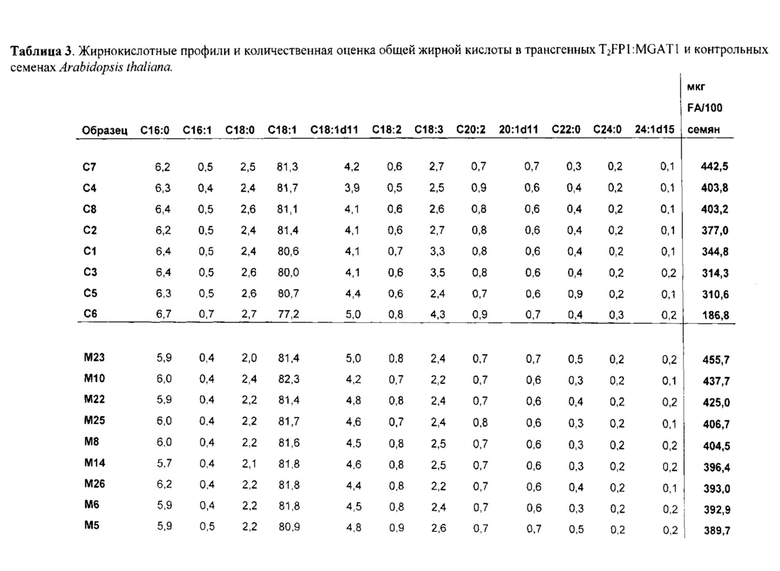
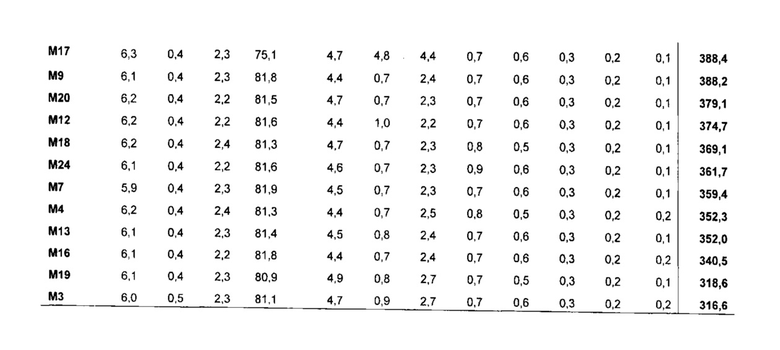
Экспрессия MGAT 1 в семенах Brassica napus
Вектор FP1:MGAT1, используемый для экспрессии MGAT1 М. musculus в семенах Arabidopsis thaliana, использовали для создания трансформированных растений B.napus. Вектор вводили в штамм AGL1 A. tumefaciens путем стандартных процедур электропорации. Культуры выращивали в течение ночи при 28°С в среде LB со встряхиванием при 150 об./мин. Бактериальные клетки собирали с помощью центрифугирования при 4000 об./мин. в течение 5 минут, промывали Winans' АВ (Winans, 1988) и ресуспендировали в 10 мл среды Winans' АВ (рН 5,2), и выращивали с канамицином (50 мг/л) и рифампицином (25 мг/л) в течение ночи с добавлением 100 мкМ ацетосирингона. За два часа перед инфицированием клеток Brassica добавляли спермидин (120 мг/л), и конечную плотность бактерий доводили до OD 600 нм 0,3-0,4 свежей средой АВ. Свежевыделенные семядольные черешки из 8-дневных проростков В. napus, выращенных на 1/2 MS (Murashige-Skoog, 1962), или гипокотильные сегменты, предварительно кондиционированные с помощью 3-4 дней на среде MS с 1 мг/л тидиазурона (TDZ)+0,1 мг/л альфа-нафталинуксусной кислоты (NAA), инфицировали 10 мл культур Agrobacterium в течение 5 минут. Экспланты (семядольный черешок и гипокотиль), инфицированные Agrobacterium, затем промокали стерильной фильтровальной бумагой для удаления избытка. Agrobacterium и переносили в среду для совместного культивирования (среда MS с 1 мг/л TDZ+0,1 мг/л NAA+100 мкМ ацетосирингона) дополненную или без различных антиоксидантов (50 мг/л L-цистеина и 15 мг/л аскорбата). Все чашки герметизировали парафильмом и инкубировали в темноте при 23-24°C в течение 48 часов.
Совместно культивированные экспланты (семядольный черешок и гипокотиль) затем промывали стерильной дистиллированной водой +500 мг/л цефотаксима+50 мг/л тиментина в течение 10 минут, прополаскивали стерильной дистиллированной водой в течение 10 минут, промокнули досуха стерильной фильтровальной бумагой, перенесли в среду предварительного отбора (MS+1 мг/л TDZ+0,1 мг/л NAA+20 мг/л аденин сульфата (ADS)+1,5 мг/л AgN03+250 мг/л цефотаксима и 50 мг/л тиментина) и культивировали в течение пяти дней при 24°C с фотопериодом 16 часов/8 часов. Затем их перенесли в среду для отбора (MS+1 мг/л TDZ+0,1 мг/л NAA+20 мг/л ADS 4 1,5 мг/л Ag N03 4- 250 мг/л цефотаксима и 50 мг/л тиментина) с 1,5 мг/л глюфосинатом аммония и культивировали в течение 4 недель при 24°C с фотопериодом 16 часов/8 часов с субкультивированием через две недели на той же среде. Экспланты с зеленым каллюсом переносили в среду для инициации побегов (MS 4-1 мг/л кинетина 4-20 мг/л ADS 4-1,5 мг/л AgNO3+250 мг/л цефотаксима и 50 мг/л тиментина 4-1,5 мг/л глюфосината аммония) и культивировали в течение следующих 2-3 недель. Побеги, появляющиеся из резистентных эксплантов, переносили в среду для удлинения побегов (среда MS с 0,1 мг/л гибберелиновой кислоты 4-20 мг/л ADS 4-1,5 мг/л AgNQ3+250 мг/л цефотаксима+1,5 мг/л глюфосината аммония и культивировали в течение следующих двух недель. Здоровые побеги длиной 2-3 см отбирали и переносили в среду для укоренения (1/2 MS с 1 мг/л NAA+20 мг/л ADS+1,5 мг/л AgNO3+250 мг/л цефотаксима) и культивировали в течение 2-3 недель. Хорошо укоренившиеся побеги с корнями переносили в вегетационные сосуды (смесь для выращивания проростков) и выращивали в вегетационной камере в течение двух недель и в дальнейшем переносили в теплицу. В отношении шестнадцати индивидуальных трансформантов подтвердили их трансгенность по конструкту FP1:MGAT1, и они нормально росли в тепличных условиях. Рост растений казался нормальным, и растения были фертильными, цвели и нормально завязывали семена. Растения выращивают до зрелости, и семена, полученные из трансформированных растений, собирают и анализируют на содержание масла семян и жирнокислотный состав. Семя-специфическая экспрессия MGAT1 повышает содержание масла и повышает процент полиненасыщенных жирных кислот в масле семян Brassica.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, заменяют MGAT1 в конструктах, упомянутых выше, и вводят в Brassica. В семенах от полученных в результате трансгенных растений содержание масла повышено.
Экспрессия MGAT1 в семенах Gossypium hirsutum
Тот же семя-специфический химерный ген, используемый для экспрессии MGAT1 М. musculus в семенах Arabidopsis thaliana, использовали для создания трансформированных растений Gossypium hirsutum. Вектор, обозначенный FP1:MGAT1, вводили в штамм AGL1 A.tumefaciens путем стандартных процедур электропорации, и клетки из культуры Agrobacterium применяли для введения химерных ДНК в клетки Gossypium hirsutum, сорт Coker315. Семядоли, отделенные от 10-дневных проростков хлопка, использовали в качестве эксплантов, и их инфицировали и совместно культивировали с A. tumefaciens в течение двухдневного периода. За этим последовал шестинедельный отбор на среде MS (Murashige and Skoog, 1962), содержащей 0,1 мг/л 2,4-D, 0,1 мг/л кинетина, 50 мг/л канамицин сульфата и 25 мг/л цефотаксима. Здоровые каллюсы, полученные из эксплантов семядолей, перенесли в среду MS, содержащую 5 мг/л 6-(γ,γ-диметилаллиламино)-пурина (2ip), 0,1 мг/л нафталинуксусной кислоты (NAA), 25 мг/л канамицина и 250 мг/л цефотаксима для второго периода из шести недель при 28°C. Соматические зародыши, образованные после около шести-десяти недель инкубации, проращивали и поддерживали на той же среде, но без добавленного фитогормона или антибиотиков. Сеянцы, развившиеся из соматических эмбрионов, перенесли в почву и содержали в теплице до развития листьев и корней с температурой выращивания 28°C/20°C (день/ночь). Трансгенные растения, содержащие конструкт FP1-MGAT1, вырастили в теплице, они отцвели и образовали коробочки, содержащие семена. Семена собирают после созревания. Семя-специфическая экспрессия MGAT1 повышает содержание масла и повышает процент полиненасыщенных жирных кислот в масле семян хлопка.
Экспрессия генов MGAT1 и MGAT2 в растениях N. benthamiana после стабильной трансформации
N. benthamiana стабильно трансформировали конструктом 35S:MGAT1, описанным в примере 2. 35S:MGAT1 вводили в штамм AGL1 A.tumefaciens путем стандартной процедуры электропорации. Трансформированные клетки выращивали на твердой среде LB, дополненной канамицином (50 мг/л) и рифампицином (25 мг/л), и инкубировали при 28°C в течение двух дней. Одну колонию использовали для инициирования свежей культуры. После 48 часов интенсивного культивирования клетки собирали центрифугированием при 2000 g, и надосадочную жидкость удаляли. Клетки ресуспендировали в свежем растворе, содержащем 50% LB и 50% среды MS при плотности OD600=0,5.
Образцы листьев N. benthamiana, выращенных in vitro, отделяли и разрезали на квадратные участки размером около 0,5-1 см2 острым скальпелем при погружении в раствор A.tumefaciens. Поврежденные кусочки листьев N.benthamiana, покрытые А.tumefaciens, выдерживали при комнатной температуре в течение 10 минут перед тем, как промокнуть насухо стерильной фильтровальной бумагой и перенести на чашки с MS без дополнений. После двухдневного периода совместного культивирования при 24°C экспланты трижды промыли стерильной жидкой средой MS, затем промокнули насухо стерильной фильтровальной бумагой и поместили на селективный агар MS, дополненный 1,0 мг/л бензиламинопурина (ВАР), 0,25 мг/л индолуксусной кислоты (IAA), 50 мг/л канамицина и 250 мг/л цефотаксима. Чашки инкубировали при 24°C в течение двух недель, чтобы дать возможность развиться побегам из трансформированных кусочков листьев N. benthamiana.
Для in vitro создания трансгенных растений с корнями здоровые зеленые побеги отрезали и переносили в 200 мл вегетационные сосуды для тканевых культур, содержащие агаровую среду MS, дополненную 25 мкг/л IAA, 50 мг/л канамицина и 250 мг/л цефотаксима. От трансгенных побегов брали довольно крупные листовые диски, и их лиофилизировали для фракционирования и количественного анализа TAG, как описано в примере 1 (таблица 4). Самое лучшее растение 35S:MGAT1 N. benthamiana имело содержание TAG 204,85 мкг/100 мг сухого веса ткани листьев по сравнению со средним содержанием TAG 85,02 мкг/100 мг сухого веса ткани листьев в контрольных линиях, что представляет собой повышение содержания TAG на 241%).
N. benthamiana также стабильно трансформировалось конструктом 35S:MGAT2, описанным в примере 2, и контрольным бинарным вектором pORE4 (таблица 5). Самое лучшее растение 35S:MGAT2 N.benthamiana имело содержание TAG 79,0 мкг/100 мг сухого веса ткани листьев по сравнению с содержанием TAG 9,5 мкг/100 мг сухого веса ткани листьев в контрольной линии, что представляет собой повышение содержания TAG на 731%. Жирнокислотный профиль фракций TAG также был изменен, при наличии существенно сниженных уровней насыщенных жирных кислот 16:0 и 18:0 и повышенных уровней полиненасыщенных жирных кислот, в частности, 18:3ωЗ (ALA) (таблица 5). Жирнокислотный профиль полярных липидов из тех же образцов листьев не был существенно затронут, что указывает на то, что изменения в составе жирных кислот неполярных липидов были реальными. Контрольные растения в этом эксперименте были меньше и физиологически отличными, нежели в предыдущем эксперименте с конструктом 35S:MGAT1, и это может быть объяснено различными содержаниями масла в контрольных растениях из одного эксперимента по сравнению с другим. Эксперименты для прямого сравнения конструктов 35S:MGAT1 и 35:MGAT2 с контрольными растениями выполняются с использованием растений одного и того же размера и физиологии.
Новый набор конститутивных бинарных векторов экспрессии получали с использованием промотора 35S с дуплицированной энхансерной областью (e35S). 35S:MGAT1#2 (pJP3346), 35S:MGAT2#2 (pJP3347) и 35S:DGAT1#2 (pJP3352) получили с помощью вначале клонирования промотора e35S, содержащегося в пределах фрагмента BamHl-EcoKI, в pORE04 в сайты BamHl-EcoRl для выхода pJP3343. Затем получали pJP3346 и pJP3347 с помощью клонирования генов MGAT1 и MGAT2, соответственно, в сайт ЕсоШ pJP3343. pJP3352 получали с помощью клонирования DGAT1 A. thaliana, содержавшейся в пределах сайта Xhol-AsiSI, в сайты XhoI-AsiSIpJP3343.
Для трансформации N. benthamiana использовали pJP3346, pJP3347 и pJP3352 в штамме AGL1 Agrobacterium, как описано выше. Четырнадцать подтвержденных трансгенных растений получили для pJP3346 и 22 - для pJP3347. Ряд резистентных к канамицину трансформированных побегов были созданы с pJP3352. Анализ экспрессии трансгенов выполняли на растениях, трансформированных MGAT1 или MGAT2. Отбирали растения с высокими уровнями экспрессии. Выполняют анализ экспрессии на растениях, трансформированных DGAT1 A. thaliana. Растения растут нормально и вырастают до зрелости. Семена собирают после созревания. Семена от потомства в высоким уровнем экспрессии высевают прямо в почву для анализа липидов расщепляющейся популяции Т2, которая включает как гомозиготные, так и гетерозиготные растения. Содержание масла в листьях растений, экспрессирующих высокие уровни как MGAT1, так и MGAT2, существенно повышается по сравнению с растениями, трансформированными DGAT1 A. thaliana или контрольными растениями.
pJP3346, pJP3347 и контрольный вектор в AGL1 также использовали для трансформации A.thaliana, как описано выше. Идентифицировали двадцать пять подтвержденных трансгенных растений Т2, содержащих Т-ДНК из pJP3346, и 43 трансгенные растения с pJP3347. Анализ экспрессии выполняют на трансгенных растениях. Семена от потомства с высокими уровнями экспрессии собирают и высевают прямо в почву. Выполняют анализ на липиды, в том числе на содержание масла в листьях от потомства Т2 и ТЗ, в том числе от сегрегантов, в которых отсутствуют трансгены. Самые высокие уровни TAG получат в растениях, которые являются гомозиготными по трансгенам MGAT.
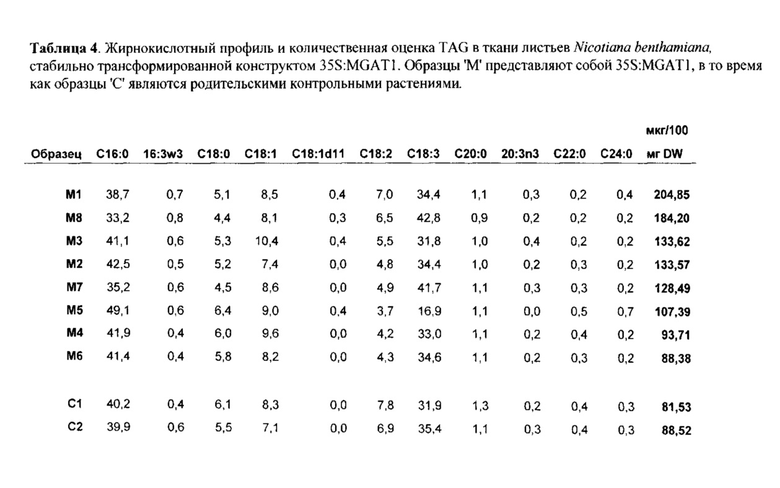
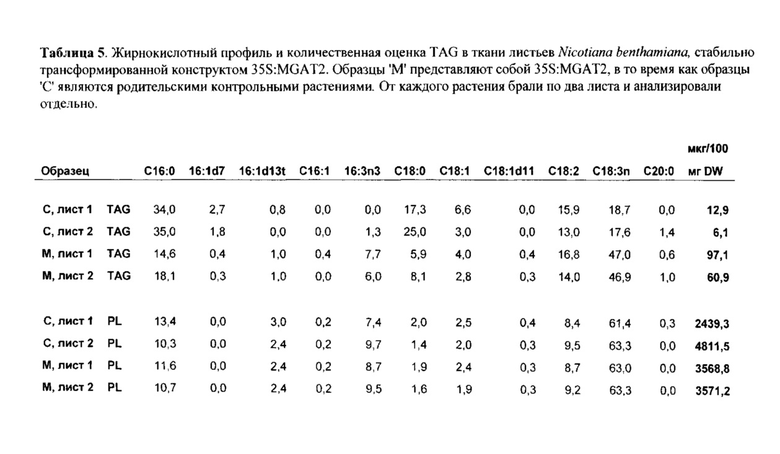
Экспрессия MGAT1 в стабильно трансформированных растениях Trifolium repens
Химерный ген, кодирующий MGAT1 М. musculus, использовали для трансформации Trifolium repens, другого двудольного растения. Векторы, содержащие химерные гены 35S:MGAT1 и 35S:DGAT1, вводили в A. tumefaciens путем стандартной процедуры электропорации. Оба вектора также содержат селектируемый маркерный ген 35S:BAR. Трансформированные клетки Agrobacterium выращивали на твердой среде LB, дополненной канамицином (50 мг/л) и рифампицином (25 мг/л), и инкубировали при 28°C в течение двух дней. Одну колонию использовали для инициации свежей культуры для каждой конструкта. После 48 часов интенсивного культивирования культуры Agrobacterium применяли для обработки семядолей Т.repens (сорт Haifa), которые были отпрепарированы из набухших семян, как описано Larkin et al. (1996). После совместного культивирования в течение трех дней экспланты подвергали действию 5 мг/л РРТ для отбора трансформированных побегов, а затем их перенесли в среду для укоренения для образования корней перед переносом в почву. Получили трансформированное растение, содержащее MGAT1. Промотор 35S конститутивно экспрессируется в клетках трансформированных растений. Содержание масла повышается по меньшей мере в вегетативных тканях, таких как листья.
Экспрессия MGAT в стабильно трансформированном Hordeum vulgare
Химерный вектор, включающий MGAT1 М.musculus, использовали для получения стабильно трансформированного Hordeum vulgare, однодольного растения. Векторы, содержащие химерные гены Ubi:MGATl и Ubi:DGATl, конструировали с помощью клонирования полных кодирующих областей MGAT1 М.musculus и DGAT1 A.thaliana по отдельности в pWVEC8-Ubi. Векторы, содержащие химерные гены Ubi:MGATl и Ubi:DGATl, вводили в штамм AGL1 A.tumefaciens путем стандартной процедуры электропорации. Трансформированные клетки Agrobacterium выращивали на твердой среде LB, дополненной канамицином (50 мг/л) и рифампицином (25 мг/л), и чашки инкубировали при 28°С в течение двух дней. По одной колонии каждого использовали для инициации свежей культуры.
После 48 часов интенсивного культивирования культуры Agrobacterium применяли для трансформации клеток в незрелых зародышах ячменя (сорт Golden Promise) в соответствии с опубликованными способами (Tingay et al., 1997; Bartlett et al., 2008) с некоторыми модификациями. Кратко, зародыши длиной 1,5-2,5 мм выделяли из незрелых зерновок и извлекали зародышевую ось. Полученные экспланты в течение 2-3 дней совместно культивировали с трансгенными Agrobacterium, а затем культивировали в темноте в течение 4-6 недель на среде, содержащей тиментин и гигромицин для получения эмбриогенного каллюса перед тем, как переместить их в переходную среду в условия низкой освещенности на две недели. Затем каллюс переносили в среду для регенерации, чтобы дать возможность регенерации побегов и корней перед переносом в почву. Получили трансформированные растения, и перенесли их в теплицу. Кодирующая область MGAT1 конститутивно экспрессируется под контролем промотора Ubi в клетках трансформированных растений. Содержание масла повышается по меньшей мере в вегетативной ткани.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, замещают MGAT1 в конструктах, упомянутых выше, и вводят в Hordeum, как описано выше. В вегетативных тканях от полученных в результате трансгенных растений содержание масла повышено.
Экспрессия MGAT в клетках дрожжей
Химерный вектор, включающий MGAT1 М. musculus, использовали для трансформации дрожжей, в этом примере Saccharomyces cerevisiae, грибковый микроорганизм, подходящий для получения масла с помощью ферментации. Генетический конструкт Gall:MGATl получили с помощью вставки полной кодирующей области конструкта, обозначенного 0954364_MGAT_pMA, который содержится в пределах фрагмента EcoRI, в pYES2 в сайт EcoRl, при этом создав pJP3301. Аналогично, генетический конструкт Gall:DGAT 1, используемый здесь в качестве сравнения и отдельно кодирующий фермент DGAT1 A. thaliana, получили с помощью вставки полной кодирующей области DGAT1 A.thaliana в pYES2. Эти химерные векторы вводили в штамм S288C S. cerevisiae с помощью теплового шока, и трансформанты отбирали на чашках с минимальной средой для дрожжей (YMM), содержащей 2% раффинозы в качестве единственного источника углерода. Клональные посевные культуры создавали в жидкой YMM с 2% раффинозы в качестве единственного источника углерода. Экспериментальные культуры инокулировали из них в среду YMM, содержащую 1% NP-40, до начальной OD600 около 0,3. Культуры выращивали при 28°С с встряхиванием (около 100 об./мин.) до того, как OD600 составил приблизительно 1,0. На этой стадии добавили галактозу до конечной концентрации 2% (вес./об.). Культуры инкубировали при 25°C с встряхиванием в течение дополнительных 48 часов перед тем, как собрать их с помощью центрифугирования. Осадки клеток промывали водой перед тем, как подвергнуть лиофилизации для фракционирования и количественного анализа классов липидов, как описано в примере 1. Промотор Gal индуцибельно экспрессируется в трансформированных клетках дрожжей, что повышает содержание масла в клетках.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках дрожжей, замещают MGAT1 в конструктах, упомянутых выше, и вводят в дрожжи. В полученных в результате трансгенных клетках содержание масла повышено. Данные гены также вводили в жировые дрожжи, Yarrowia lipolytica, для повышения содержания масла.
Экспрессия MGAT в клетках водорослей Chlamydomonas reinhardtii
Химерный вектор, включающий MGAT1 М.musculus, используют для стабильной трансформации водорослевых клеток. Генетический конструкт, обозначенный 35S:MGAT1, получают с помощью клонирования кодирующей области MGAT1 в клонирующий вектор, содержащий 35S промоторную кассету вируса мозаики цветной капусты и ген резистентности к парамомицину (аминогликозид-О-фосфотрансфераза VIII), который экспрессируется с помощью промотора RBCS2 С.reinhardtii. 35S:MGAT1 вводят отдельно в культуру сс503 при концентрации 5х107, находящуюся в стадии логарифмического роста, штамм Chlamydomonas reinhardtii с недостаточностью клеточной стенки, с помощью модифицированного способа со стеклянными бусами (Kindle, 1990). Оба вектора также содержат ген резистентности BLE в качестве селектируемого маркерного гена. Кратко, колонию нетрансформированных клеток на чашке с ТАР агаром, поддерживаемую при около 24°C, выращивают до около 5×106 клеток/мл в течение четырех дней, полученные в результате клетки осаждают при 3000 g в течение 3 минут при комнатной температуре и ресуспендируют для получения 5×107 клеток в 300 мкл среды ТАР. Добавляют 300 мкл стеклянных бус диаметром 0,6 мм, 0,6 мкг плазмиды в 5 мкл и 100 мкл 20% PEG MW8000, и смесь перемешивают на вортексе при максимальной скорости в течение 30 секунд, затем переносят в 10 мл ТАР и инкубируют в течение 16 часов при встряхивании в темноте. Клетки осаждают, ресуспендируют в 200 мкл ТАР, затем вносят в чашки с ТАР, содержащие 5 мг/л зеоцина, и инкубируют в темноте в течение 3 недель. Трансформированные колонии субкультивируют на чашку Петри со свежей ТАР+зеоцин 5 мг/л, после чего их выращивают в условиях стандартной среды с зеоциновым отбором. После сбора с помощью центрифугирования осадок клеток промывают водой перед тем, как подвергнуть лиофилизированию для фракционирования и количественного анализа классов липидов, как описано в примере 1. Промотор 35S:MGAT1 конститутивно экспрессируется в трансформированных водорослевых клетках. Содержание масла в клетках существенно повышено.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, заменяют MGAT1 в конструктах, упомянутых выше, и вводят в Chlamydomonas. Содержание масла в полученных в результате трансгенных клетках существенно повышено.
Экспрессия MGAT в стабильно трансформированном Lupinus angustifolius
Химерный вектор, включающий MGAT1 М. musculus, применяют для трансформации Lupinus angustifolius, бобового растения. Химерные векторы 35S:MGAT1 и 35S:DGAT1 в Agrobacterium применяют для трансформации L. angustifolius, как описано Pigeaire et al. (1997). Кратко, экспланты из верхушки побега культивируют совместно с трансгенной Agrobacterium, прежде чем тщательно смачивают раствором РРТ (2 мг/мл) и переносят на среду для регенерации без РРТ. Многочисленные аксиллярные побеги, развивающиеся из верхушки побега, отделяют на среду, содержащую 20 мг/л РРТ, и выжившие побеги переносят на свежую среду, содержащую 20 мг/л РРТ. Затем здоровые побеги переносят в почву. Промотор 35S конститутивно экспрессируется в клетках трансформированных растений, что повышает содержание масла в вегетативных тканях и семенах. Семя-специфический промотор используют для дополнительного повышения содержания масла в семенах трансгенного Lupinus.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, заменяют MGAT1 в конструктах, упомянутых выше, и вводят в Lupinus. В семенах и вегетативных тканях от полученных в результате трансгенных растений содержание масла повышено.
Экспрессия MGAT в стабильно трансформированных клетках Sorghum bicolor
Химерный вектор, включающий MGAT1 М.musculus, применяют для стабильной трансформации Sorghum bicolor. Ubi:MGATl и Ubi:DGATl в штамме AGL1 A.tumefaciens применяют для трансформации Sorghum bicolor, как описано Gurel et al. (2009). Agrobacterium вначале центрифугируют при 5000 об./мин. при 4°C в течение 5 минут и разбавляют до OD550=0,4 с помощью жидкой средой для совместного культивирования. Затем предварительно изолированные незрелые зародыши полностью покрывают суспензией Agrobacterium на 15 минут и затем культивируют, щитком зародыша вверх, на среде для совместного культивирования в темноте в течение 2 дней при 24°С.Затем незрелые зародыши переносят в среду для индукции каллюса (CIM) с 100 мг/л карбенициллина, чтобы ингибировать рост Agrobacterium, и оставляют в течение 4 недель. Затем ткани переносят в среду для регенерации для побегов и корней. Промотор Ubi конститутивно экспрессируется в клетках трансформированных растений, что повышает содержание масла по меньшей мере в вегетативных тканях.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, заменяют MGAT 1 в конструктах, упомянутых выше, и вводят в Sorghum. В вегетативных тканях от полученных в результате трансгенных растений содержание масла повышено.
Экспрессия MGAT в стабильно трансформированных растениях Glycine max
Химерный ген, кодирующий MGAT1 М. musculus, применяют для стабильной трансформации Glycine max, другого бобового растения, которое может использоваться для получения масла. 35S:MGAT1 в Agrobacterium применяют для трансформации G.max, как описано Zhang et al. (1999). Agrobacterium в течение трех дней культивируют совместно с семядольными эксплантами, полученными от пятидневных проростков. Затем экспланты культивируют на среде Гамборга В5, дополненной 1,67 мг/л BAP и 5,0 мг/л глюфосината, в течение четырех недель, после чего экспланты субкультивируют в среде, содержащей главные и второстепенные соли MS и витамины В5 (MS/B5), дополненной 1,0 мг/л зеатин-рибозида, 0,5 мг/л GA3 и 0,1 мг/л IAA, измененной 1,7 мг/л или 2,0 мг/л глюфосината. Удлиненные побеги укореняют на среде для укоренения MS/B5, дополненной 0,5 мг/л NAA без дополнительного отбора глюфосинатом. Промотор 35S конститутивно экспрессируется в клетках трансформированных растений, что повышает содержание масла в вегетативных тканях и семенах.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, заменяют MGAT1 в конструктах, упомянутых выше, и вводят в Glycine. В вегетативных тканях и семенах от полученных в результате трансгенных растений содержание масла повышено.
Экспрессия MGAT в стабильно трансформированной Zea mays
Химерный ген, кодирующий MGAT1 М.musculus, применяют для стабильной трансформации Zea mays. Векторы, содержащие 35S:MGAT1 и 35S:DGAT1, применяют для трансформации Zea mays, как описано Gould et al. (1991). Кратко, экспланты из верхушки побега культивируют совместно с трансгенной Agrobacterium в течение двух дней перед тем, как перенести их на солевую среду MS, содержащую канамицин и карбенициллин. После нескольких раундов субкультивирования спонтанно образуются трансформированные побеги и корни, и их переносят в почву. Промотор 35S экспрессируется в клетках трансформированных растений, что повышает содержание масла в вегетативных тканях и семенах.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, заменяют MGAT1 в конструктах, упомянутых выше, и вводят в Zea mays. В вегетативных тканях и семенах от полученных в результате трансгенных растений содержание масла повышено. Альтернативно, кодирующие области MGAT экспрессируют под контролем эндосперм-специфического промотора, такого как промотор зеина, или зародыш-специфический промотор, полученный из однодольного растения, для повышенной экспрессии и повышенного содержания масла в семенах. Дополнительный химерный ген, кодирующий GPAT с фосфатазной активностью, такой как GPAT4 или GPAT6 A.thaliana, вводят в Zea mays в комбинации с MGAT, что дополнительно повышает содержание масла в семенах кукурузы.
Экспрессия MGAT в стабильно трансформированном Elaeis guineensis (масличная пальма)
Химерный ген, кодирующий MGAT1 М.musculus, применяют для стабильной трансформации Elaeis guineensis. Применяются химерные векторы, обозначенные Ubi:MGATl и Ubi:DGATl, в Agrobacterium. После 48 часов интенсивного культивирования клетки применяют для трансформации Elaeis guineensis, как описано Izawati et al. (2009). Промотор Ubi конститутивно экспрессируется в клетках трансформированных растений, что повышает содержание масла по меньшей мере в плодах и семенах, и может применяться для получения масла.
Кодирующей областью гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, заменяют MGAT1 в конструктах, упомянутых выше, и вводят в Elaeis. В семенах от полученных в результате трансгенных растений содержание масла повышено.
Экспрессия MGAT в стабильно трансформированном Avena sativa (овес)
Химерный ген, кодирующий MGAT1 М.musculus, применяют для стабильной трансформации Avena sativa, другого однодольного растения. Химерные векторы, обозначенные Ubi:MGATl и Ubi:DGATl, как описано выше, и оба содержащие селектируемый маркер Ubi:BAR, применяли для трансформации Avena sativa, как описано Zhang et al. (1999).
Кодирующей область гена MGAT2 мыши, кодон-оптимизированной для экспрессии в клетках растений, заменяют MGAT1 в конструктах, упомянутых выше, и вводят в Avena. В семенах от полученных в результате трансгенных растений содержание масла повышено.
Пример 7. Конструирование MGAT с DGAT активностью
MGAT со сравнимой DGAT активностью можно сконструировать с помощью выполнения случайного мутагенеза, нацеленного мутагенеза или насыщающего мутагенеза гена(ов) интереса MGAT, и/или подвергая различные гены MGAT и/или DGAT ДНК-шаффлингу. Функцию DGAT можно точно отсортировать с применением, например, штамма дрожжей, который имеет абсолютную потребность в комплементации синтеза TAG при снабжении свободными жирными кислотами, такого как H1246, который содержит мутации в четырех генах (DGA1, LRO1, ARE1, ARE2). Трансформирование вариантов MGAT в такой штамм, а затем снабжение трансформированных дрожжей такой концентрацией свободных жирных кислот, которая предупреждает комплементацию диким типом гена MGAT, будет допускать только рост вариантов с повышенной способностью к синтезу TAG из-за улучшенной DGAT активности. MGAT активность этих мутированных генов можно определять с помощью снабжения их меченым sn-1 или sn-2 MAG и количественным определением образования меченного DAG. Несколько раундов направленной эволюции в комбинации с рациональным конструированием белка приведет к образованию нового гена MGAT с подобными MGAT и DGAT активностями.
Пример 8. Конститутивная экспрессия диацилглицерол ацилтрансферазы 2 А.thaliana в растениях
Экспрессия DGAT2 A. thaliana в дрожжах (Weselake et al., 2009) и клетках насекомых (Lardizabal et al., 2001) не показала DGAT активности. Аналогично, DGAT2 была не способна комплементировать нокаутные мутации DGAT1 A.thaliana (Weselake et at., 2009). Активность фермента DGAT2 A.thaliana в ткани листьев определяли с применением системы временной экспрессии N. benthamiana, как описано в примере 1. DGAT2 A.thaliana (№ доступа Q9ASU1) получали с помощью геномной ПЦР и клонировали в бинарный вектор экспрессии под контроль промотора 35S для создания 35S:DGAT2. Этот химерный вектор вводили в штамм AGL1 A.tumefaciens, и клетки из их культур инфильтровали в ткань листьев растений N. benthamiana в вегетационном домике при 24°C, с использованием 35S:DGAT1 в качестве контроля. Инфильтовали несколько прямых сравнений, при этом образцы, подлежащие сравнению, располагались с обеих стороны одного листа. Эксперименты выполняли в трех повторностях. После инфильтрации растения выращивали в течение дополнительных пяти дней перед тем, как листовые диски собирали и лиофилизировали для фракционирования классов липидов и количественного анализа, как описано в примере 1. Этот анализ показал, что как DGAT1, так и DGAT2 были функционирующими в отношение повышения уровней масла в листьях Nicotiana benthamiana (таблица 6).
Ткань листьев, трансформированная конструктом 35S:pl9 (отрицательный контроль), содержала в среднем 34,9 мкг свободных жирных кислот, происходящих от TAG/100 мг сухого веса листьев. Ткань листьев, трансформированная конструктами 35S:pl9 и 35S:DGAT1 (положительный контроль), содержала в среднем 126,7 мкг свободных жирных кислот, происходящих от TAG/100 мг сухого веса листьев. Ткань листьев, трансформированная конструктами 35S:pl9 и 35S:DGAT2, содержала в среднем 310,0 мкг свободных жирных кислот, происходящих от TAG/100 мг сухого веса листьев.
Данные, описанные выше, демонстрируют, что фермент DGAT2 A.thaliana является более активным, чем фермент DGAT1 A.thaliana, в стимуляции накопления TAG в ткани листьев. Экспрессия гена DGAT2 приводит к не менее чем 245% накоплению TAG в ткани листьев по сравнению с тем, когда экспрессировалась DGAT1.
Пример 9. Совместная экспрессия MGAT и GPAT в трансгенных семенах
Yang et al. (2010) описали две глицерол-3-фосфат ацилтрансферазы (GPAT4 и GPAT6) из A. thaliana, имеющие sn-2 предпочтение (т.е. предпочтительно образующие sn-2 MAG, а не sn-1/3 MAG) и фосфатазную активность, которые были способны образовывать sn-2 MAG из G-3-P (фигура 1). Предполагалось, что эти ферменты составляют часть пути синтеза кутина. GPAT4 и GPAT6 не экспрессировались сильно в ткани семян. Комбинирование такой бифункциональной GPAT/фосфатазы с MGAT дает новый путь синтеза DAG с использованием G-3-P в качестве субстрата, который может заменить или дополнить типичный путь Кеннеди для синтеза DAG в растениях, в частности, в семенах масличных культур или других клетках, что приводит к повышенному содержанию масла, в частности, уровней TAG.
Химерные ДНК, обозначенные pJP3382 и pJP3383, кодирующие GPAT4 и GPAT6 A.thaliana, соответственно, вместе с MGAT2 М. musculus, для экспрессии в семенах растений получали с помощью вначале вставки полной кодирующей области MGAT2, которая содержалась в пределах фрагмента Swat., в pJP3362 в сайт Smal для выхода pJP3378. pJP3362 представлял собой бинарный вектор экспрессии, содержащий пустые кассеты экспрессии FAE1 и FP1 и ген резистентности к канамицину в качестве селектируемого маркера. GPAT4 A.thaliana амплифицировали из кДНК и клонировали в pJP3378 в сайте Notl для выхода pJP3382, в котором GPAT4 экспрессировалась посредством усеченного промотора напина, FP1, и MGAT2 экспрессировались посредством промотора FAE1 A.thaliana. Аналогично, GPAT6 A.thaliana амплифицировали из кДНК и клонировали в pJP3378 в сайт Notl для выхода pJP3384, в котором GPAT6 функционально связан с усеченным промотором напина, FP1, и MGAT2 экспрессировали с помощью промотора FAE1 A.thaliana. pJP3382 и pJP3383 трансформировали в A. thaliana (экотип Columbia) с помощью способа погружения цветков. Семена от обработанных растений высевали на среду, содержащую антибиотик канамицин, для отбора растений-потомков (растения Т1), которые были трансформированы. Трансгенные проростки перенесли в почву и выращивали в теплице. Определяли экспрессию трансгенов в развивающихся зародышах. Трансгенные растения с наивысшим уровнем экспрессии и те, которые показали соотношение 3:1 в отношении трансгенные:нетрансгенные растения на линию, что свидетельствует об одном локусе вставки трансгенов, выбирают и выращивают до зрелости. Из этих растений получают семена (T2), которые включают некоторое количество гомозиготных по трансгенам. Пятьдесят растений-потомков (растения T2) из каждой линии выращивают в почве и определяют содержание липидов, содержание TAG и жирнокислотные состав полученных в результате семян. Содержание нейтральных липидов, в частности TAG, в семенах, содержащих одновременно MGAT и GPAT4 или GPAT6 существенно повышается по сравнению с контролем и по сравнению с семенами, содержащими только MGAT или только DGAT1 A.thaliana.
Содержание MAG и DAG также повышается. Жирнокислотный состав липидов, экстрагированных из семян, модифицирован, в частности, содержание полиненасыщенных жирных кислот, таких как ALA, существенно повышено.
Пример 10. Тестирование эффекта GPAT4 и GPAT6 на опосредованное MGAT повышение TAG с помощью сайленсинга и мутации GPAT
Семейство GPAT является большим, и все известные члены содержат два консервативных домена, домен plsC ацилтрансферазы домен и домен суперсемейства HAD-подобной гидролазы. В дополнение к этому, во всех GPAT4-8 A.thaliana содержится N-концевая область, гомологичная домену фосфосерин фосфатазы. Как GPAT4, так и GPAT6 A.thaliana содержат консервативные остатки, которые известны как критические для фосфатазной активности (Yang et al., 2010).
Вырожденные праймеры на основе консервативной аминокислотной последовательности GDLVICPEGTTCREP (SEQ ID NO:228) конструировали для амплификации фрагментов на GPAT N. benthamiana, экспрессированных в ткани листьев. 3' RACE будет проводиться с использованием этих праймеров и олиго-dT обратных праймеров на РНК, выделенной из ткани листьев N. benthamiana. GPAT с фосфатазной активностью (т.е. СРАТ4/6-подобные) будут идентифицироваться по их гомологии с доменной областью N-концевой фосфосерин фосфатазы, описанной выше. Управляемые 35S РНКи конструкты, нацеленные на эти гены, будут создаваться и трансформироваться в штамм AGL1 A.tumefaciens. Аналогично, конструкт 35S:V2, содержащий вирусный белок-супрессор сайленсинга V2, будут трансформировать в штамм AGL1 A.tumefaciens. V2, как известно, подавляет нативный механизм «сайленсинга» растений, что дает возможность эффективной временной экспрессии, но также позволяет функционирование сайленсинга генов, основанного на РНКи.
Затем будут сравнивать накопление TAG между временно трансформированными образцами листьев, инфильтрованными следующими смесями штаммов: 1) 35S:V2 (отрицательный контроль); 2) 35S:V2+35S:MGAT2 (положительный контроль); 3) 35S:V2+GPAT-РНКи; 4) 35S:V2+GPAT-РНКи t 35S:MGAT2. Ожидается, что смесь 35S:V2+GPAT-РНКи+35S:MGAT2 приведет в результате к меньшему накоплению TAG, чем образец 35S:V2+35S:MGAT2 из-за прерванного синтеза sn-2 MAG в результате сайленсинга GPAT.
Подобный эксперимент будет осуществлен с использованием GPAT4/6-подобных последовательностей A.thaliana и N.benthamiana, которые мутируют для удаления консервативных остатков, известных как критические для фосфатазной активности (Yang et al., 2010). Эти мутированные гены (совместно известные как ОРАТ4/6-дельта) затем будут клонировать в управляемые 35S бинарные векторы экспрессии и трансформировать в A.tumefaciens. Затем будут сравнивать накопление TAG между временно трансформированными образцами листьев, инфильтрованными следующими смесями штаммов: 1) 35S:pl9 (отрицательный контроль); 2) 35S:p 19+35S:MGAT2 (положительный контроль); 3) 35S:pl9+ОРАТ4/6-дельта; 4) 35S:p19+ОРАТ4/6-дельта+35S:MGAT2. Ожидается, что смесь 35S:p19+ОРАТ4/6-дельта+35S: MGAT2 приведет к меньшему накоплению TAG, чем образец 35S:p19+35S.MGAT2 из-за прерванного синтеза sn-2 MAG в результате мутации GPAT. Хотя нативные ОРАТ4/6-подобные гены N. benthamiana будут присутствовать в этом эксперименте, ожидается, что высокий уровень экспрессии конструктов GPAT4/6-дельта будет вытеснять на второй план эндогенные гены при доступе к субстрату G-3-P.
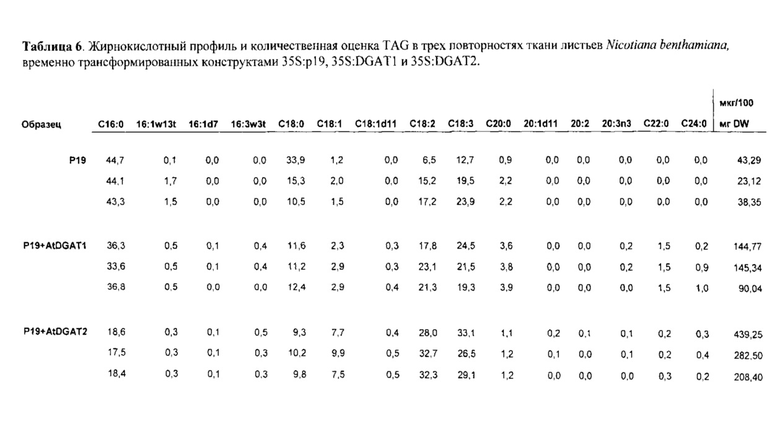
Специалистам в данной области техники будет понятно, что в настоящем изобретении могут производиться многочисленные вариации и/или модификации, как показано в конкретных вариантах осуществления, без отклонения от сущности или объема настоящего изобретения, как в широком смысле описано. Настоящие варианты осуществления, следовательно, должны рассматриваться во всех отношениях как иллюстративные и не ограничительные.
Все публикации, обсуждаемые и/или на которые ссылаются в настоящем документе, включены в настоящий документ во всей своей полноте.
Любое обсуждение документов, действий, материалов, устройств, изделий и т.п., которые были включены в настоящее описание, приводится исключительно для целей обеспечения контекста настоящего изобретения. Это не следует понимать как признание того, что любой или все эти объекты образуют часть основы уровня техники или являлись общеизвестными распространенными знаниями в области, относящейся к настоящему изобретению, как она существовала до даты приоритета в отношении каждого пункта формулы изобретения данной заявки.
REFERENCES
Abdullah et al. (1986) Biotech. 4:1087.
Alvarez et al. (2000) Theor Appl Genet 100:319-327.
Al-Mariri et al. (2002) Infect. Immun. 70:1915-1923.
Bao and Ohlrogge (1999) Plant Physiol. 120:1057-62.
Bartlett et al. (2008) Plant Methods 4:22.
Baumlein et al. (1991) Mol. Gen. Genet. 225:459-467.
Baumlein et al. (1992) Plant J. 2:233-239.
Benghezal et al. (2007) Journal of Biological Chemistry 282:30845-30855.
Bligh and Dyer (1959) Canadian Journal of Biochemistry and Physiology 37:911-7.
Bouvier-Nave et al. (2000) European Journal of Biochemistry / FEBS 267:85-96.
Bradford (1976) Anal. Biochem. 72:248.
Broothaerts et al. (2005) Nature 433:629-633.
Broun etal. (1998) Plant J. 13:201-210.
Cadwell and Joyce (1992) PCR Methods Appl. 2:28-33.
Cao et al. (2003) J. Biol. Chem. 278:13860-13866.
Cao et al. (2007) J. Lipid Res. 48:583.
Cheng et al. (1996) Plant Cell Rep.15:653-657.
Cheng et al. (2003) J. Biol. Chem. 278, 13611-13614.
Chikwamba et al. (2003) Proc. Natl. Acad. Sci. U.S.A. 100:11127-11132.
Chung et al. (2006) BMC Genomics 7:120.
Clough and Bent (1998) Plant J. 16:735-743.
Coco et al. (2001) Nature Biotechnology 19:354-359.
Coco et al. (2002) Nature Biotechnology 20:1246-1250.
Comai et al. (2004) Plant J 37: 778-786.
Courvalin et al. (1995) Life Sci. 318:1209-1212.
Coutu et al. (2007) Transgenic Res. 16:771-781.
Crameri et al. (1998) Nature 391:288-291.
Darji et al. (1997) Cell 91:765-775.
Deshpande and Mukund (1992) Appl. Biochem. Biotechnol, 36:227.
Dietrich et al. (1998) Nature Biotech. 18:181-185.
Eggert et al. (2005) Chembiochem 6:1062-1067.
Ellerstrom et al. (1996) Plant Mol. Biol. 32:1019-1027.
Fennelly et al. (1999) J. Immunol. 162:1603-1610.
Fujimura et al. (1985) Plant Tissue Culture Lett. 2:74.
Ghosal et al. (2007) Biochimica et Biophysica Acta 1771:1457-1463.
Ghosh et al. (2006) International Symposium on Plant Lipids, Abstract.
Glevin et al. (2003) Microbiol. Mol. Biol. Rev. 67:16-37.
Grant et al. (1995) Plant Cell Rep.15:254-258.
Grillot-Courvalin et al. (1998) Nature Biotech. 16:862-866.
Grillot-Courvalin (1999) Curr. Opin. Biotech. 10-477-481.
Gould et al. (1991) Plant Physiol. 95:426-434.
Gurel et al. (2009) Plant Cell Rep.28:429-444.
Harayama (1998) Trends Biotechnol. 16: 76-82.
Hellinga (1997) Proc. Natl. Acad. Sci. 94(19): 10015-10017.
Henikoff et al. (2004) Plant Physiol 135: 630-636.
Hense et al. (2001) Cell Microbiol. 3:599-609.
Hinchee et al. (1988) Biotechnology 6:915-922.
Hirayama and Hujii К (1965) Agricultural and Biological Chemistry 29:1-6.
Horvath et al. (2000) Proc. Natl. Acad. Sci. U.S.A. 97:1914-1919.
Izawati et al. (2009) J. Oil Palm Res. 21:643-652.
Jako et al. (2001) Plant Physiol. 126:861-874.
Jezequel et al. (2008) Biotechniques 45:523-532.
Lacroix et al. (2008) Proc. Natl. Acad. Sci.U.S.A. 105:15429-15434.
Leung et al. (1989) Technique 1:11-15.
Kindle (1990) Proc. Nat. Acad. Sci. USA 87: 1228-1232.
Koziel et al. (1996) Plant Mol. Biol. 32:393-405.
Kunik et al. (2001) Proc. Natl. Acad. Sci. U.S.A. 98:1871-1876.
U.S.A. 105: 15429-15434.
Lakshminarayana et al. (1984) Journal of the American Oil Chemists Society 61:1249-1253.
Lardizabal et al. (2001) J. Biol. Chem. 276:38862-38869.
Larkin et al. (1996) Transgenic Res. 5:325-335.
Murashige and Skoog (1962) Physiologia Plantarum 15:473-497.
Ness et al. (2002) Nature Biotechnology 20:1251-1255.
Niedz et al. (1995) Plant Cell Reports 14:403.
Ostermeier et al. (1999) Nature Biotechnology 17:1205-1209.
Ow et al. (1986) Science 234:856-859.
Panekina (1978) Chemistry of Natural Compounds 14:33-36.
Perez-Vich et al. (1998) JAOCS 75:547-555
Perrin et al. (2000) Mol Breed 6:345-352.
Perry and Harwood (1993) Phytochemistry 33:329-333.
Phillips et al. (2002) Journal of Food Composition and Analysis 12:123-142.
Pigeaire et al. (1997) Mol. Breed. 3:341-349.
Potenza et al. (2004) In Vitro Cell Dev. Biol. Plant 40:1-22.
Powell etal. (1996)Vaccines 183, Abstract.
Prasher et al. (1985) Biochem. Biophys. Res. Commun. 127:31-36.
Rajasekharan et al. (2006) International Symposium on Plant Lipids, Abstract.
Sahaetal. (2006) Plant Physiol. 141:1533-1543.
Schaffher et al. (1980) Proc. Natl. Acad. Sci. U.S.A. 77:2163-2167.
Shiau et al. (2001) Vaccine 19:3947-3956.
Sieber et al. (2001) Nature Biotechnology 19:456-460.
Sizemore et al. (1995) Science 270:299-302.
Sladeand Knauf (2005) Transgenic Res. 14: 109-115.
Stobart et al. (1997) Planta 203:58-66.
Stalker et al. 1988 Science 242: 419-423.
Stemmer (1994a) Proc. Natl. Acad. Sci. USA 91:10747-10751.
Stemmer (1994b) Nature 370(6488):389-391.
Thilletetal. (1988) J. Biol. Chem 263:12500-12508.
Tingay etal. (1997) Plant J. 11:1369-1376.
Toriyama et al. (1986) Theor. Appl. Genet. 205:34.
Tumaney et al. (2001) J. Biol. Chem. 276:10847-10852.
Tzfira and Citovsky (2006) Curr. Opin. Biotech. 17:147-154.
Weiss et al. (2003) Int. J. Med. Microbiol. 293:95:106.
Weselake et al. (2009) Biotechnology Advances 27:866-878.
Winans et al. (1988) Journal of Bacteriology 170:4047-4054.
Wood et al. (2009). Plant Biotech. J. 7: 914-924.
Yang et al. (2003) Planta 216:597-603.
Yang et al. (2010) PNAS 107:12040-12045.
Yen and Farese (2003) J. Biol. Chem. 278:18532-18537.
Yen et al. (2002) Proc. Natl. Acad. Sci. U.S.A. 99:8512-8517.
Yen et al. (2005) J. Lipid Res., 46: 1502-1511.
Voinnet et al. (2003) Plant J. 33:949-956.
Volkov et al. (1999) Nucleic acids research 27(18):el8.
Zhang et al. (1999) Plant Cell Reports 18:959-966.
Zhang et al. (1999) Plant Cell, Tissue and Organ Culture 56:-46.
Zheng etal. (2003) The Plant Cell 15:1872-1887.
Zhao et al. (1998) Nature Biotechnology 16:258-261.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ЛИПИДОВ | 2012 |
|
RU2656996C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИПИДОВ | 2012 |
|
RU2804627C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| ФЕРМЕНТАТИВНАЯ ОБРАБОТКА МАСЕЛ | 2004 |
|
RU2377307C2 |
| АНАЛИЗ ДИАЦИЛГЛИЦЕРИН-АЦИЛТРАНСФЕРАЗЫ | 2005 |
|
RU2415946C2 |
| ДЕЛЬТА-5-ДЕСАТУРАЗА И ЕЕ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ПОЛИНЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ | 2007 |
|
RU2469092C2 |
| ГЕНЫ ДИАЦИЛГЛИЦЕРОЛ-АЦИЛТРАНСФЕРАЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 2010 |
|
RU2514655C2 |
| ПЕСТИЦИДНЫЕ КОМПОЗИЦИИ И ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ | 2013 |
|
RU2662672C2 |
| СПОСОБЫ ПОВЫШЕНИЯ ТОЛЕРАНТНОСТИ К АБИОТИЧЕСКОМУ СТРЕССУ И/ИЛИ БИОМАССЫ У РАСТЕНИЙ И ПОЛУЧАЕМЫЕ ТАКИМ ОБРАЗОМ РАСТЕНИЯ | 2004 |
|
RU2350653C2 |

























































































































































































































































































































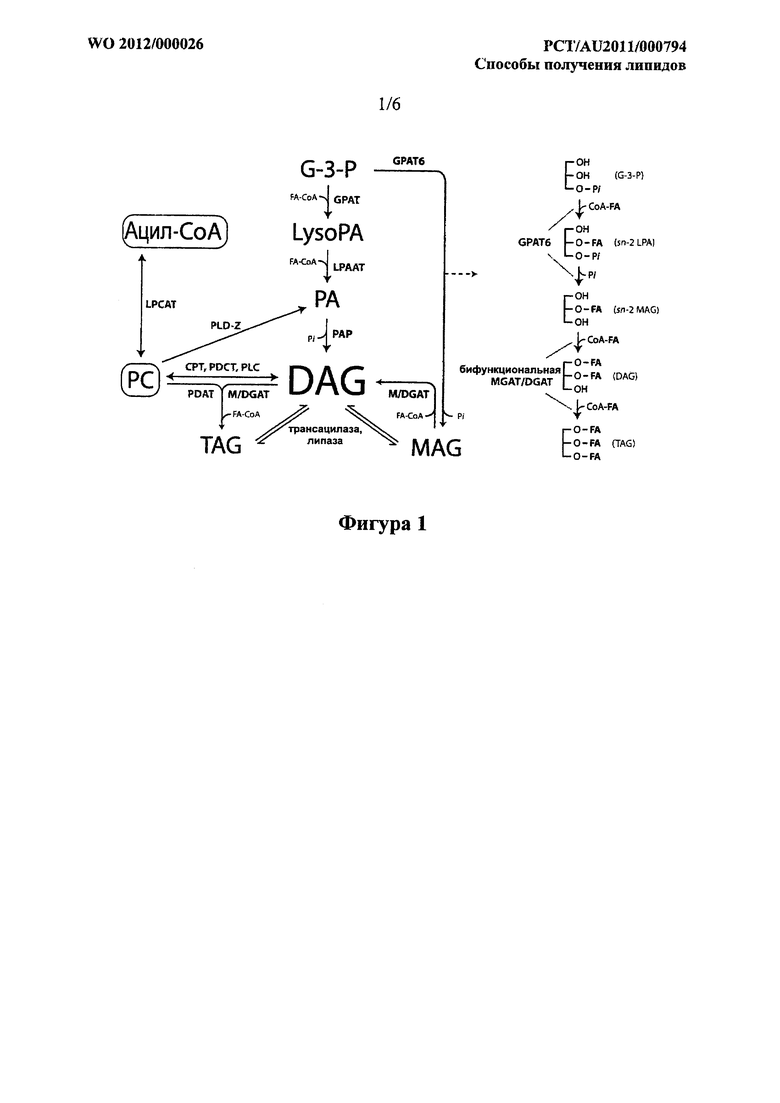

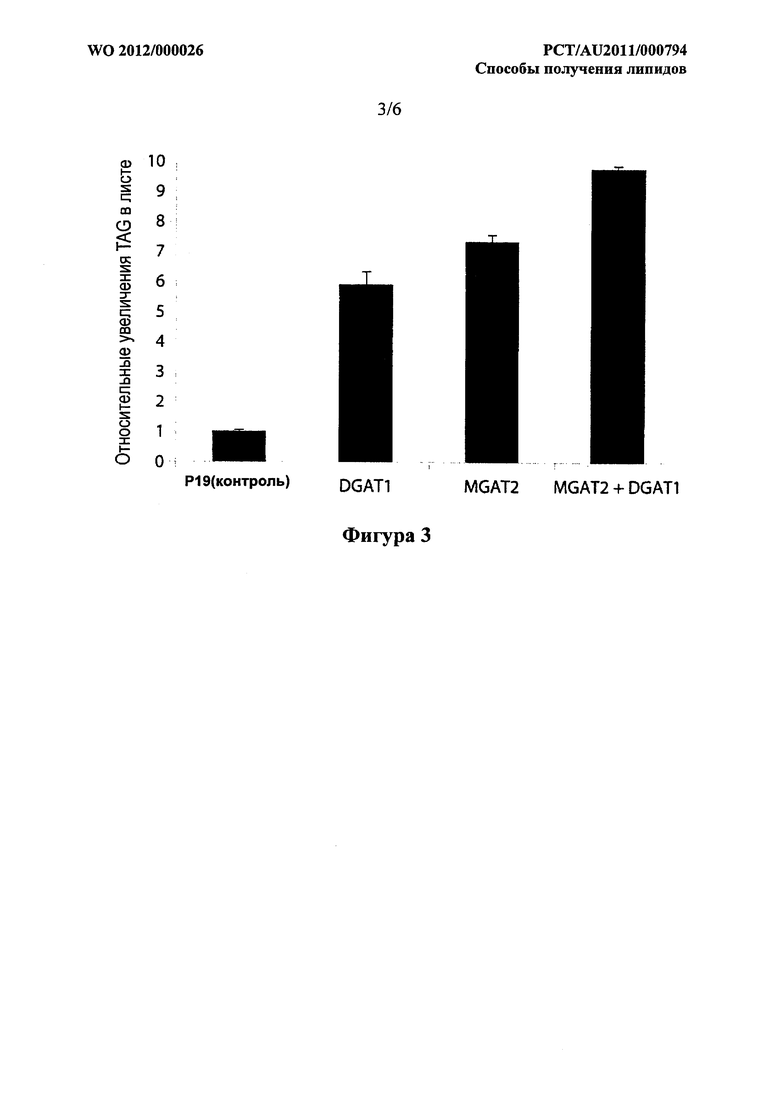
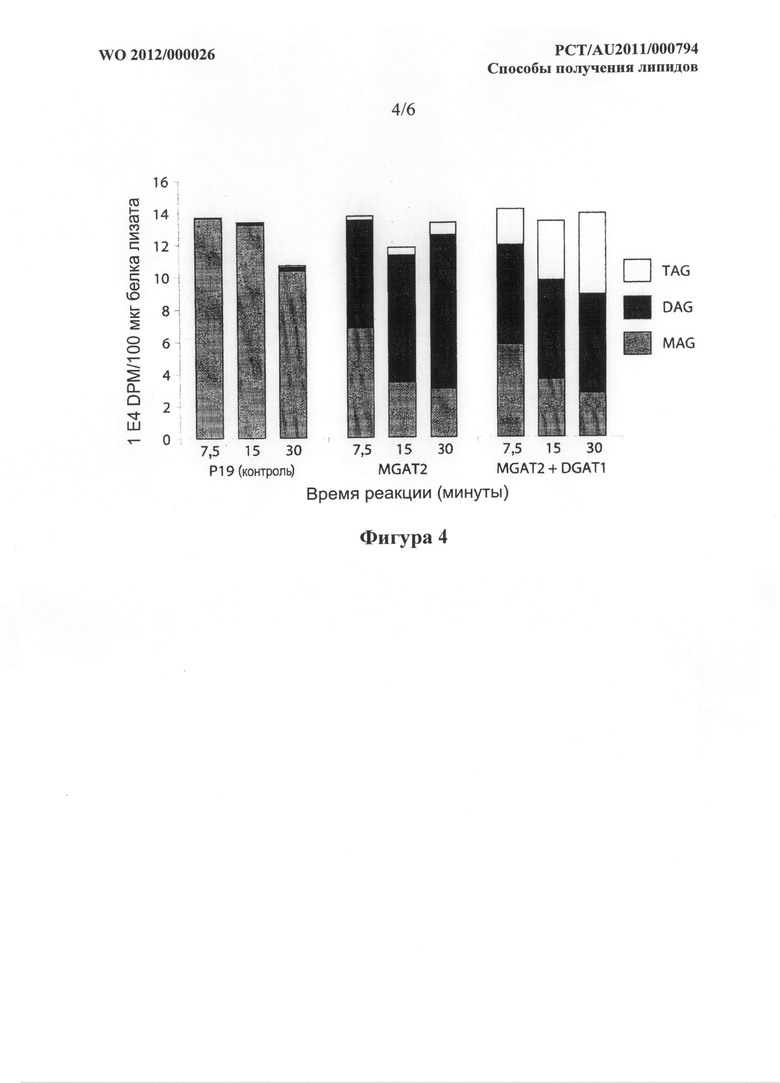
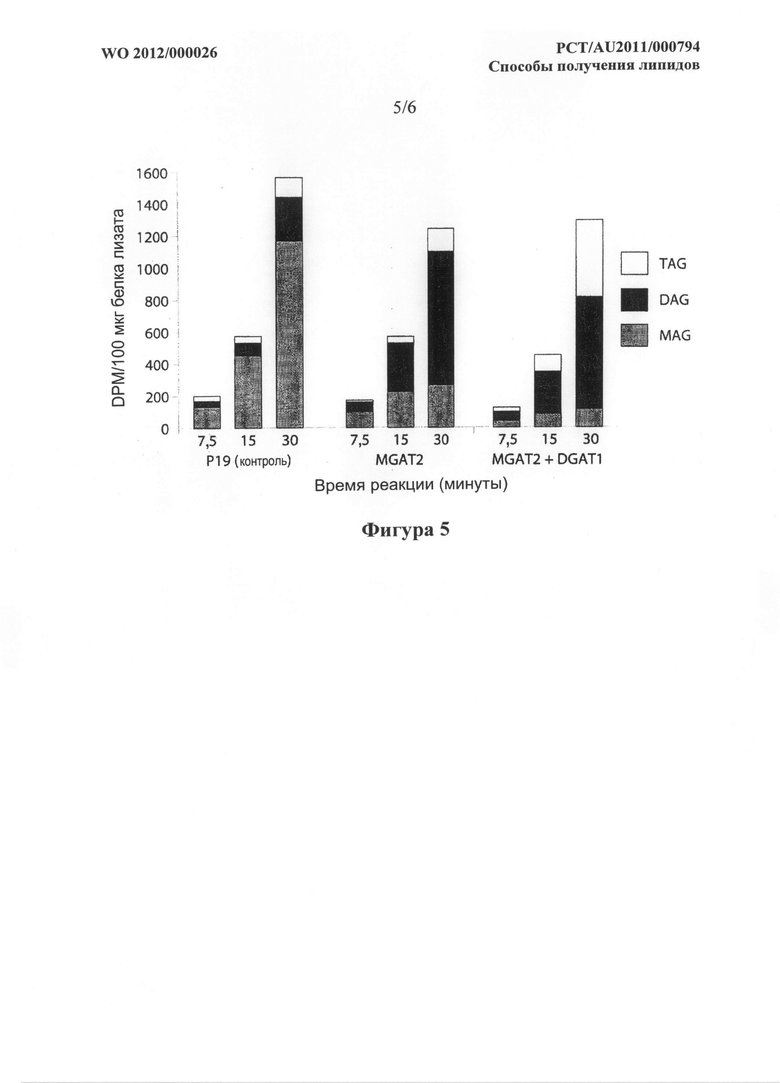
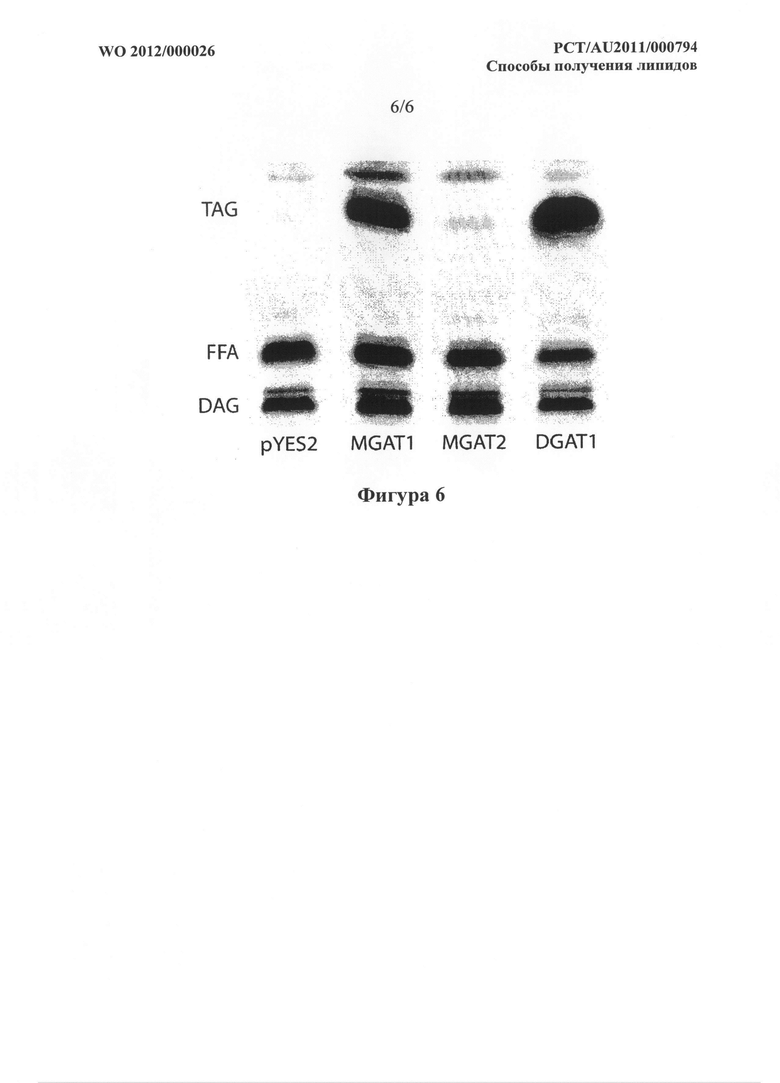
Изобретение относится к области биохимии, в частности к трансгенной растительной клетке, трансгенному растению, части трансгенного растения и к семени трансгенного растения, содержащим экзогенный полинуклеотид, кодирующий моноацилглицерол-ацилтрансферазу (MGAT), и имеющим повышенный уровень триацилглицерола (TAG), а также к способу получения из них экстрагированного липида и их применению для получения промышленного продукта. Также раскрыт способ получения растительной клетки или семени, которые имеют повышенный уровень TAG, предусматривающий введение в клетку или в семя экзогенного полинуклеотида, кодирующего MGAT. Изобретение также относится к способу получения алкиловых эфиров жирных кислот, способу получения пищевого продукта, каждый из которых предусматривает применение вышеуказанной клетки, растения, части растения или его семени, а также к самому полученному пищевому продукту. Также раскрыт способ ферментации, предусматривающий использование вышеуказанной растительной клетки. Изобретение позволяет эффективно повышать уровень TAG в клетке растения. 14 н. и 15 з.п. ф-лы, 6 ил., 6 табл., 9 пр.
1. Трансгенная растительная клетка, содержащая экзогенный полинуклеотид, кодирующий моноацилглицерол-ацилтрансферазу (MGAT), где трансгенная растительная клетка имеет повышенный уровень триацилглицерола (TAG) по сравнению с соответствующей растительной клеткой, в которой отсутствует указанный экзогенный полинуклеотид.
2. Трансгенная растительная клетка по п. 1, где экзогенный полинуклеотид кодирует моноацилглицерол-ацилтрансферазу 2 (MGAT2) или моноацилглицерол-ацилтрансферазу 1 (MGAT1).
3. Трансгенная растительная клетка по п. 1, где уровень триацилглицерола (TAG) в трансгенной растительной клетке на 0,5% (вес./вес.) - 25% (вес./вес.) выше на основе веса, чем у соответствующей трансгенной растительной клетки, в которой отсутствует указанный экзогенный полинуклеотид.
4. Трансгенная растительная клетка по п. 1, которая характеризуется одним или несколькими из:
(i) где клетка дополнительно содержит один или несколько экзогенных полинуклеотидов, кодирующих глицерол-3-фосфат-ацилтрансферазу (GPAT), и/или диацилглицерол-ацилтрансферазу (DGAT), предпочтительно где GPAT также имеет фосфатазную активность для образования моноацилглицерола (MAG) и предпочтительно где DGAT представляет собой диацилглицерол-ацилтрансферазу 2 (DGAT2);
(ii) где общее содержание неполярных липидов трансгенной растительной клетки увеличено по сравнению с соответствующей растительной клеткой, где отсутствует указанный экзогенный полинуклеотид;
(iii) где общее содержание неполярных липидов трансгенной растительной клетки на 0,5% (вес./вес.) - 25% (вес./вес.) больше на основе веса, чем в соответствующей растительной клетке, где отсутствует указанный экзогенный полинуклеотид;
(iv) где содержание полиненасыщенной жирной кислоты (PUFA) в липиде трансгенной растительной клетки увеличено по сравнению с липидом соответствующей растительной клетки, где отсутствует указанный экзогенный полинуклеотид;
(v) где содержание полиненасыщенной жирной кислоты (PUFA) в липиде трансгенной растительной клетки увеличено по сравнению с липидом соответствующей растительной клетки, где отсутствует указанный экзогенный полинуклеотид, где PUFA включает эйкозадиеновую кислоту (EDA), арахидоновую кислоту (ARA), альфа-линоленовую кислоту (ALA), стеаридоновую кислоту (SDA), эйкозатриеновую кислоту (ЕТЕ), эйкозатетраеновую кислоту (ЕТА), эйкозапентаеновую кислоту (ЕРА), докозапентаеновую кислоту (DPA), докозагексаеновую кислоту (DHA) или комбинацию двух или более из них;
(vi) где трансгенная растительная клетка происходит от Brassica sp., Gossypium hirsutum, Linum usitatissimum, Helianthus sp., Carthamus tinctorius, Glycine max, Zea mays, Arabidopsis thaliana, Sorghum bicolor, Sorghum vulgare, Avena sativa, Trifolium sp., Camelina sativa, Miscanthus x giganteus или Miscanthus sinensis или от люпина, арахиса или подсолнечника;
(vii) где общее содержание масла или общее содержание жирной кислоты в растительной клетке на 0,5% (вес./вес.) - 25% (вес./вес.) больше на основе веса, чем в соответствующей растительной клетке, где отсутствует указанный экзогенный полинуклеотид;
(viii) где содержание триацилглицерола (TAG), диацилглицерола (DAG), TAG и DAG или моноацилглицерола (MAG) в трансгенной растительной клетке по меньшей мере на 10% (вес./вес.) больше на относительной основе, чем содержание TAG, DAG, TAG и DAG или MAG соответствующей растительной клетки, где отсутствует указанный экзогенный полинуклеотид;
(ix) где по меньшей мере 60% (моль %) содержания жирной кислоты липида трансгенной растительной клетки представляет собой олеиновую кислоту; и
(x) где уровень одного или нескольких неполярных липидов и/или общее содержание неполярных липидов трансгенной растительной клетки на 0,5% (вес./вес.) - 25% (вес./вес.) больше на основе веса, чем у соответствующей растительной клетки, где отсутствует указанный экзогенный полинуклеотид, кодирующий MGAT, но которая содержит экзогенный полинуклеотид, кодирующий DGAT1 Arabidopsis thaliana.
5. Трансгенная растительная клетка по любому из пп. 1-4, где триацилглицерол содержит специфическую полиненасыщенную жирную кислоту (PUFA), которая представляет собой эйкозадиеновую кислоту (EDA), арахидоновую кислоту (ARA), альфа-линоленовую кислоту (ALA), стеаридоновую кислоту (SDA), эйкозатриеновую кислоту (ЕТЕ), эйкозатетраеновую кислоту (ЕТА), эйкозапентаеновую кислоту (ЕРА), докозапентаеновую кислоту (DPA), докозагексаеновую кислоту (DHA) или комбинацию двух или более из них.
6. Трансгенное растение, содержащее экзогенный полинуклеотид, кодирующий моноацилглицерол-ацилтрансферазу (MGAT), где трансгенное растение имеет повышенный уровень триацилглицерола (TAG) по сравнению с соответствующим растением, в котором отсутствует указанный экзогенный полинуклеотид.
7. Растение по п. 6, представляющее собой водоросль.
8. Часть трансгенного растения, содержащая экзогенный полинуклеотид, кодирующий моноацилглицерол-ацилтрансферазу (MGAT), где часть трансгенного растения имеет повышенный уровень триацилглицерола (TAG) по сравнению с соответствующей частью растения, в которой отсутствует указанный экзогенный полинуклеотид.
9. Часть растения по п. 8, представляющая собой вегетативную часть растения, надземную часть растения, зеленую часть растения, лист растения или стебель растения.
10. Семя трансгенного растения, содержащее экзогенный полинуклеотид, кодирующий моноацилглицерол-ацилтрансферазу (MGAT), где семя трансгенного растения имеет повышенный уровень триацилглицерола (TAG) по сравнению с соответствующим семенем растения, в котором отсутствует указанный экзогенный полинуклеотид.
11. Способ получения экстрагированного липида, причем способ включает этапы:
i) получения трансгенной растительной клетки по любому из пп. 1-5, растения по п. 6 или 7, части растения по п. 8 или 9 или семени растения по п. 10, и
ii) экстрагирования липида из указанных трансгенной растительной клетки, растения, части растения или семени растения, таким образом получая экстрагированный липид.
12. Способ по п. 11, где семя растения представляет собой семя канолы, которое включает по меньшей мере 45% масла семян на основе веса.
13. Способ по п. 11, где семя растения представляет собой семя кукурузы, которое включает по меньшей мере 5% масла семян на основе веса.
14. Способ по п. 11, где семя растения представляет собой семя сои, которое включает по меньшей мере 20% масла семян на основе веса.
15. Способ по любому из пп. 11-14, где способ дополнительно включает одно или несколько из сбора семян из трансгенного растения, отжима масла семян из семени, очищения масла семян и получения промышленного продукта из экстрагированного липида, где промышленный продукт представляет собой предпочтительно алкиловые сложные эфиры, такие как метиловые сложные эфиры жирных кислот.
16. Способ получения растительной клетки, имеющей повышенный уровень триацилглицерола (TAG), где способ включает следующее:
i) введение в растительную клетку экзогенного полинуклеотида, кодирующего моноацилглицерол-ацилтрансферазу MGAT, где экзогенный полинуклеотид функционально связан с одним или несколькими промоторами, которые способны управлять экспрессией одного или нескольких экзогенных полинуклеотидов в клетке,
ii) экспрессирование одного или нескольких экзогенных полинуклеотидов в клетке или клетке-потомке,
iii) анализ содержания липидов клетки или клетки-потомка, и
iv) выбор клетки, имеющей повышенный уровень триацилглицерола (TAG) по сравнению с соответствующей клеткой, где отсутствует указанный экзогенный полинуклеотид.
17. Способ по п. 16, где выбранная клетка представляет собой клетку, определенную в любом из пп. 1-5.
18. Способ по п. 16, где экзогенный полинуклеотид является стабильно интегрированным в геном клетки.
19. Способ по любому из пп. 16-18, дополнительно включающий этап регенерации трансгенного растения из клетки этапа i).
20. Применение экзогенного полинуклеотида, кодирующего моноацилглицерол-ацилтрансферазу (MGAT), для получения трансгенной растительной клетки, имеющей повышенный уровень триацилглицерола (TAG) по сравнению с соответствующей растительной клеткой, где отсутствует указанный экзогенный полинуклеотид.
21. Применение по п. 20, где трансгенная растительная клетка представляет собой клетку по любому из пп. 1-5.
22. Способ получения семени, причем способ включает:
i) выращивание трансгенного растения, содержащего экзогенный полинуклеотид, кодирующий моноацилглицерол-ацилтрансферазу (MGAT), и
ii) сбор семени от этого растения, где семя имеет повышенный уровень триацилглицерола (TAG) по сравнению с соответствующим семенем, в котором отсутствует указанный экзогенный полинуклеотид.
23. Применение трансгенной растительной клетки по любому из пп. 1-5, растения по п. 6 или 7, части растения по п. 8 или 9 или семени растения по п. 10 для производства промышленного продукта, предпочтительно алкиловых сложных эфиров, таких как метиловые сложные эфиры жирных кислот.
24. Применение по п. 23, где промышленный продукт представляет собой топливо.
25. Способ получения алкиловых эфиров жирных кислот, причем способ включает:
i) экстрагирование липида из трансгенной растительной клетки по любому из пп. 1-5, из растения по п. 6 или 7, из части растения по п. 8 или 9 или из семени по п. 10; и
ii) реакцию липида со спиртом, необязательно, в присутствии катализатора для образования указанных алкиловых сложных эфиров жирных кислот.
26. Способ получения пищевого продукта, причем способ включает примешивание трансгенной растительной клетки по любому из пп. 1-5, растения по п. 6 или 7, части растения по п. 8 или 9 или семени по п. 10 к по меньшей мере одному другому пищевому ингредиенту.
27. Пищевой продукт, содержащий трансгенную растительную клетку по любому из пп. 1-5, растение по п. 6 или 7, часть растения по п. 8 или 9 или семя по п. 10.
28. Способ ферментации, включающий этапы:
i) обеспечения сосуда, содержащего жидкую композицию, содержащую трансгенную растительную клетку по любому из пп. 1-5, который является подходящим для ферментации, и составляющих, необходимых для ферментации и биосинтеза жирной кислоты, и
ii) обеспечения условий, способствующих ферментации жидкой композиции, содержащейся в указанном сосуде.
29. Применение экзогенного полинуклеотида, кодирующего моноацилглицерол-ацилтрансферазу (MGAT) в растительной клетке для повышения образования триацилглицерола (TAG), где растительная клетка, содержащая указанный экзогенный полинуклеотид, имеет повышенный уровень триацилглицерола (TAG) по сравнению с соответствующей растительной клеткой, в которой отсутствует указанный экзогенный полинуклеотид.
| WO 2008147935 A2, 04.12.2008 | |||
| COLETTE JAKO ET AL., Seed-Specific Over-Expression of an Arabidopsis cDNA Encoding a Diacylglycerol Acyltransferase Enhances Seed Oil Content and Seed Weight, Plant Physiology, June 2001, Vol | |||
| Ударно-вращательная врубовая машина | 1922 |
|
SU126A1 |
| Наконечник для брандспойта | 1911 |
|
SU861A1 |
| WO 2004011671 A2, 05.02.2004 | |||
| KATHRYN D | |||
| LARDIZABAL et al., DGAT2 Is a New Diacylglycerol Acyltransferase Gene Family, THE JOURNAL OF BIOLOGICAL CHEMISTRY, 2001, Vol | |||
| СПОСОБ СОСТАВЛЕНИЯ ЗВУКОВОЙ ЗАПИСИ | 1921 |
|
SU276A1 |
| Устройство для усиления микрофонного тока с применением самоиндукции | 1920 |
|
SU42A1 |
| Катушечная зеркальная камера | 1931 |
|
SU38862A1 |
| US 2005106697 A1, 19.05.2005 | |||
| МОДИФИКАЦИЯ ЛИПИДОВ РАСТЕНИЙ И МАСЕЛ СЕМЯН ПУТЕМ ИСПОЛЬЗОВАНИЯ ДРОЖЖЕВЫХ ГЕНОВ SLC | 1996 |
|
RU2190665C2 |

