Настоящее изобретение относится к области клеточной биологии и может быть использовано в клеточной трансплантологии и тканевой инженерии с целью получения клеточного материала для восстановления нервной ткани, поврежденной в результате травм, инсультов или нейродегенеративных заболеваний, а также для тестирования лекарственных препаратов, влияющих на функциональную активность клеток нервной ткани и нейральных предшественников. Предлагается способ, позволяющий с помощью иммуносортинга с антителами против рецептора к нейротрофическому фактору мозга BDNF (TrkB) выделять из суммарной популяции стромальных клеток жировой ткани субпопуляцию прогениторных клеток, предрасположенных к нейральной дифференцировке, для последующей эффективной индукции нейральной дифференцировки этих клеток с помощью соответствующих факторов.
Введение
Потеря трудоспособности в результате повреждений центральной нервной системы и периферических нервных волокон вследствие травм, инсультов или нейродегенеративных заболеваний связана с невозможностью полноценной регенерации нервной ткани. Как и другие соматические ткани человека, нервная ткань содержит небольшие количества так называемых стволовых клеток, которые в случае локального повреждения способны активироваться, подвергаться тканеспецифической дифференцировке и восстанавливать морфофункциональную целостность ткани. Однако при тяжелых повреждениях, а также у пациентов старшего возраста количество таких клеток оказывается недостаточным для полноценной репарации, поэтому наиболее перспективным подходом для лечения таких пациентов является трансплантация стволовых и прогениторных клеток. Эти клетки могут быть получены из собственной ткани пациента, наращены in vitro в необходимом количестве и введены в участок повреждения. В отличие от эмбриональных стволовых клеток, при использовании которых с терапевтической целью возникают этические и религиозные проблемы, опасность передачи инфекционных заболеваний и развития опухолей (тератом), а также отторжение чужеродного материала, применение собственных (аутологичных) клеток не сопряжено с подобными сложностями. Трансплантация аутологичных стволовых клеток, а также тканевых фрагментов, выращенных in vitro на их основе, уже сейчас успешно применяется для восстановления кожного покрова после ожогов, для замещения костных и хрящевых дефектов, а также для восстановления тканей после удаления раковых образований.
В последние годы все большее внимание исследователей привлекает возможность использования в качестве источника аутологичных клеток стромальных клеток из жировой ткани, доступность получения которой в достаточном объеме практически для любого пациента сегодня позволяет рассматривать ее как оптимальный ресурс для клеточной терапии, в том числе и для регенерации нервной ткани.
Уровень техники
Известно, что строма жировой ткани содержит малодифференцированные клетки-предшественники, которые по своему фенотипу и экспрессионному профилю сходны с мезенхимальными стволовыми клетками из костного мозга [6]. Стромальные клетки-предшественники из жировой ткани (СКЖТ) под действием различных индукторов способны дифференцироваться in vitro в различные типы клеток: остеобласты, хондробласты, адипоциты [1], гепатоциты, эндотелиальные клетки [16], миобласты [10], кардиомиоциты [12], эпителиальные клетки [3], а также нейральные предшественники [4].
Нейральная дифференцировка стволовых клеток различного происхождения обычно сопровождается повышением экспрессии специфичных генов и, соответственно, появлением в клетке таких белков цитоскелета, как нестин (маркер ранних нейральных предшественников), нейрональная форма тубулина-β3, глиальный кислый фибриллярный белок GFAP (маркер астроцитарной глии), белок, взаимодействующий с микротрубочками МАР2, специфичная для нейронов енолаза Eno2 и некоторых других маркеров нейральной дифференцировки. Известно, что индуцировать нейральную дифференцировку мезенхимальных стволовых клеток, например клеток, происходящих из таких источников, как костный мозг, можно с помощью ретиноевой кислоты [7] и нейротрофических факторов [8, 15]. Что касается СКЖТ, то несмотря на повышенный интерес к этому источнику прогениторных клеток исследования, касающиеся их способности дифференцироваться в нейральном направлении в ответ на стимуляцию известными индукторами нейральной дифференцировки, носят эпизодический характер и пока не позволяют сделать какие-либо определенные выводы относительно механизмов нейральной дифференцировки и оптимальных условий получения популяции клеток, дифференцирующихся в этом направлении. В патенте США №7,078,230 [18] предлагается многостадийный способ получения предназначенной для трансплантации популяции стромальных клеток из жировой ткани, индуцированных к нейральной дифференцировке под действием ретиноевой кислоты, а в работе [19] в качестве индуктора нейральной дифференцировки СКЖТ, наряду с ретиноевой кислотой, предлагается использовать нейротрофический фактор BDNF. Этот способ предусматривает выделение популяции стромальных клеток из жировой ткани человека, культивирование их в стандартных условиях в течение 3-5 недель и индукцию нейральной дифференцировки, осуществляемую в два этапа: культивированием в течение 3 дней в бессывороточной среде, содержащей bFGF (20нг/мл) и hEGF (20нг/мл), которое сопровождается образованием нейросфер, и последующим 30-дневным культивированием в присутствии 10 нг/мл BDNF и 0,75 мМ ретиноевой кислоты (RA). Очевидными недостатками данного способа являются его длительность и недостаточная эффективность. Как сообщают авторы, после 30-дневной индукции BDNF и RA выраженные морфологические и иммуноцитохимические признаки нейральной дифференцировки проявляются примерно у половины клеток.
Поскольку известные методы получения индуцированной к нейральной дифференцировке культуры СКЖТ, в том числе и рассмотренные выше, наиболее близкие к предлагаемому способу [19], представляются малопригодными для практических целей из-за их низкой эффективности, настоящее изобретение было направлено на решение общей задачи повышения этого показателя.
Раскрытие изобретения
Одной из наиболее вероятных причин низкой эффективности индукции СКЖТ под действием нейротрофических факторов может быть недостаточный уровень «чувствительности» суммарной популяции клеток к индуктору, поэтому в основу разработки нового метода получения индуцированной к нейральной дифференцировке популяции СКЖТ было положено выявление в суммарной популяции СКЖТ TrkB-позитивных клеток, чувствительных к действию избранных для индукции нейральной дифференцировки нейротрофических факторов, в частности нейротрофического фактора мозга (BDNF), и получение гомогенной культуры клеток, обогащенной TrkB-позитивными клетками, способными эффективно дифференцироваться в нейральном направлении.
Известно, что о готовности клеток воспринимать сигналы нейротрофических факторов можно судить по экспрессии на их поверхности рецепторов к этим факторам, в частности, к действию BDNF чувствительны клетки, экспрессирующие тирозинкиназный рецептор BDNF - TrkB. Этот рецептор экспрессируется на поверхности нейронов, а также на аксонах и дендритах во многих структурах головного мозга [17], а в эмбриогенезе TrkB обнаруживается в клетках моноцитарной-дендритной фракции во время формирования иннервации того или иного органа [5]. Именно посредством взаимодействия с этим рецептором BDNF поддерживает дифференцировку и выживаемость нейронов.
С учетом этого, решаемая задача сводилась к попытке выявить в суммарной популяции СКЖТ клетки, несущие на своей поверхности рецептор нейротрофического фактора мозга (BDNF) TrkB, оценить их содержание в популяции и предложить подходящий метод для максимального обогащения культуры TrkB-содержащими клетками с тем, чтобы повысить «восприимчивость» культуры СКЖТ к последующей индукции с помощью данного фактора, а при установлении такой возможности - и другими известными индукторами нейральной дифференцировки.
При осуществлении изобретения впервые было экспериментально установлено присутствие в суммарной популяции СКЖТ клеток, несущих на своей поверхности рецептор нейротрофического фактора мозга. В частности, с помощью ПЦР-анализа было обнаружено наличие клеток, экспрессирующих мРНК рецептора BDNF (пример 2а, фиг.1), а методом иммунофлуоресцентного окрашивания были выявлены клетки, содержащие антиген TrkB (пример 2б). По результатам цитометрического анализа доля клеток, несущих TrkB рецептор, в суммарной популяции была оценена как равная примерно 6%.
Для обогащения популяции СКЖТ TkrB-экспрессирующими клетками использовали два метода: метод иммуномагнитной селекции и метод проточной цитофлуориметрии; при этом в обоих случаях удалось получить по существу гомогенную популяцию клеток (более 90%) TrkB-содержащих СКЖТ.
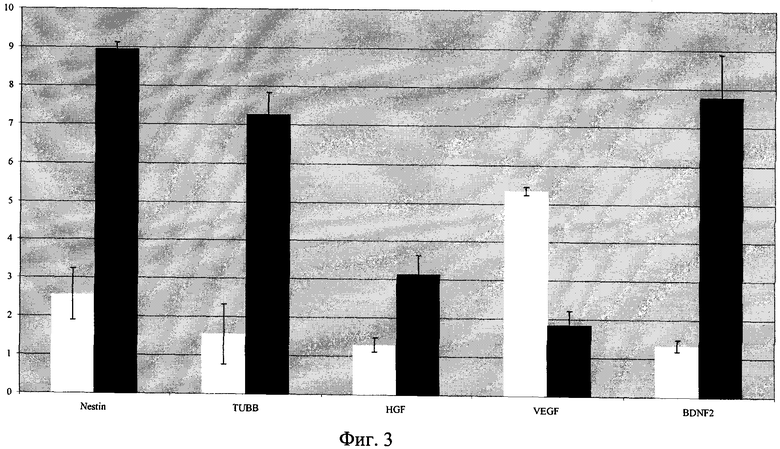
Анализ полученной фракции TrkB-позитивных клеток на экспрессию соответствующих маркеров, свидетельствующих о потенциальной способности этих клеток к нейральной дифференцировке, включавший определение содержания в клетках мРНК специфичных маркерных белков, выявил присутствие мРНК нестина и тубулина-β3, а также нейротрофических факторов роста HGF и BDNF; при этом содержание всех названных мРНК в этих клетках было в несколько раз выше, чем в TrkB-негативных СКЖТ (пример 4, фиг.3).
Полученную субпопуляцию TrkB-содержащих клеток индуцировали к нейральной дифференцировке путем помещения клеток в дифференцировочную среду DMEM/10% FBS, содержащую BDNF (в сравнительном эксперименте - ретиноевую кислоту в сочетании с 5-азацитидином). Оценка эффективности индукции, предусматривавшая определение экспрессии на уровне транскрипции маркерных генов нейральной дифференцировки (нестина, тубулина-β3 и нейронспецифической енолазы Eno2) показала, что в клетках, индуцированных BDNF, уже через 3 дня количество мРНК нестина, тубулина и Eno2 увеличивается в 3-6 раз. Через 7 дней после индукции в этих клетках также возрастало содержание соответствующих белков (Пример 5). При индукции TrkB-экспрессирующих СКЖТ ретиноевой кислотой в целом наблюдались сходные, хотя и менее выраженные, изменения. В результате прямого эксперимента по трансплантации гомогенной популяции Trk-экспрессирующих СКЖТ, полученной и индуцированной согласно предложенному методу, в головной мозг мыши было установлено, что в течение 10 дней наблюдения трансплантированные клетки сохраняли жизнеспособность и демонстрировали миграцию из области введения в паренхиму мозга реципиента, тогда как контрольные клетки суммарной популяции СКЖТ оставались в области трека и не обнаруживали контакта с тканью пациента.
Таким образом, сущность предлагаемого изобретения заключается в получении суммарной популяции СКЖТ, выявлении и выделении из суммарной популяции субпопуляции TrkB-позитивных клеток с помощью иммуномагнитной селекции или иммуносортинга с использованием антител против рецептора TrkB, и индукции нейральной дифференцировки нейротрофическим фактором BDNF или ретиноевой кислотой в сочетании с 5-азацитидином. Преимуществом (техническим результатом) предлагаемого способа перед известными ранее способами является повышение эффективности индукции нейральной дифференцировки и сокращение общих сроков подготовки культуры к трансплантации, о возможности которой однозначно свидетельствует показанная в прямом эксперименте высокая жизнеспособность индуцированных TrkB-позитивных СКЖТ и их способность интегрироваться в ткань реципиента.
Краткое описание чертежей
Фиг.1. Содержание мРНК TrkB в суммарной популяции СКЖТ. Продукт амплификации фрагмента транскрипта TrkB (83 пн) указан стрелкой.
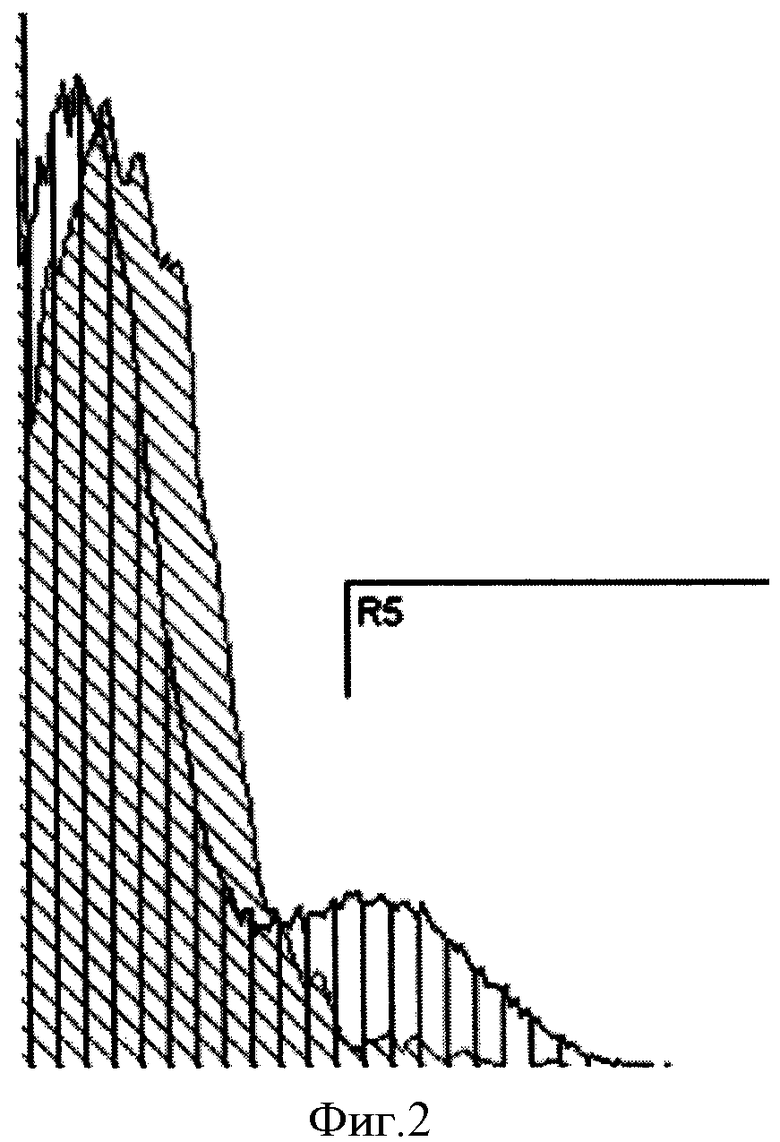
Фиг.2. Анализ содержания клеток, экспрессирующих TrkB, в суммарной популяции СКЖТ с помощью проточной цитометрии. Косая штриховка соответствует клеткам, инкубированным с контрольными иммуноглобулинами, вертикальная штриховка - клеткам, окрашенным антителами против TrkB). R5 - область подсчета позитивно окрашенных клеток.
Фиг.3. Сравнительный анализ содержания мРНК маркерных генов нейральной дифференцировки и нейротрофических факторов в субпопуляциях СКЖТ, экспрессирующих и не экспрессирующих TrkB. По оси абсцисс - аббревиатуры тестируемых генов, по оси ординат - уровень мРНК анализируемых генов, нормированный по содержанию мРНК двух генов домашнего хозяйства (GAPDH, β-actin). Черные столбики - клетки, экспрессирующие TrkB, белые столбики - клетки, не экспрессирующие TrkB.
Фиг.4. Содержание маркерных белков нейральной дифференцировки в субпопуляциях СКЖТ, экспрессирующих и не экспрессирующих TrkB до и после индукции. Левая колонка панелей - детектирование нестина; правая колонка панелей - детектирование β3-тубулина. А, Б - субпопуляции СКЖТ, не экспрессирующих TrkB, до индукции; В, Г - субпопуляции СКЖТ, экспрессирующих TrkB, до индукции; Д, Е - субпопуляции СКЖТ, экспрессирующих TrkB, после воздействия BDNF в сочетании с 5-азацитидином.
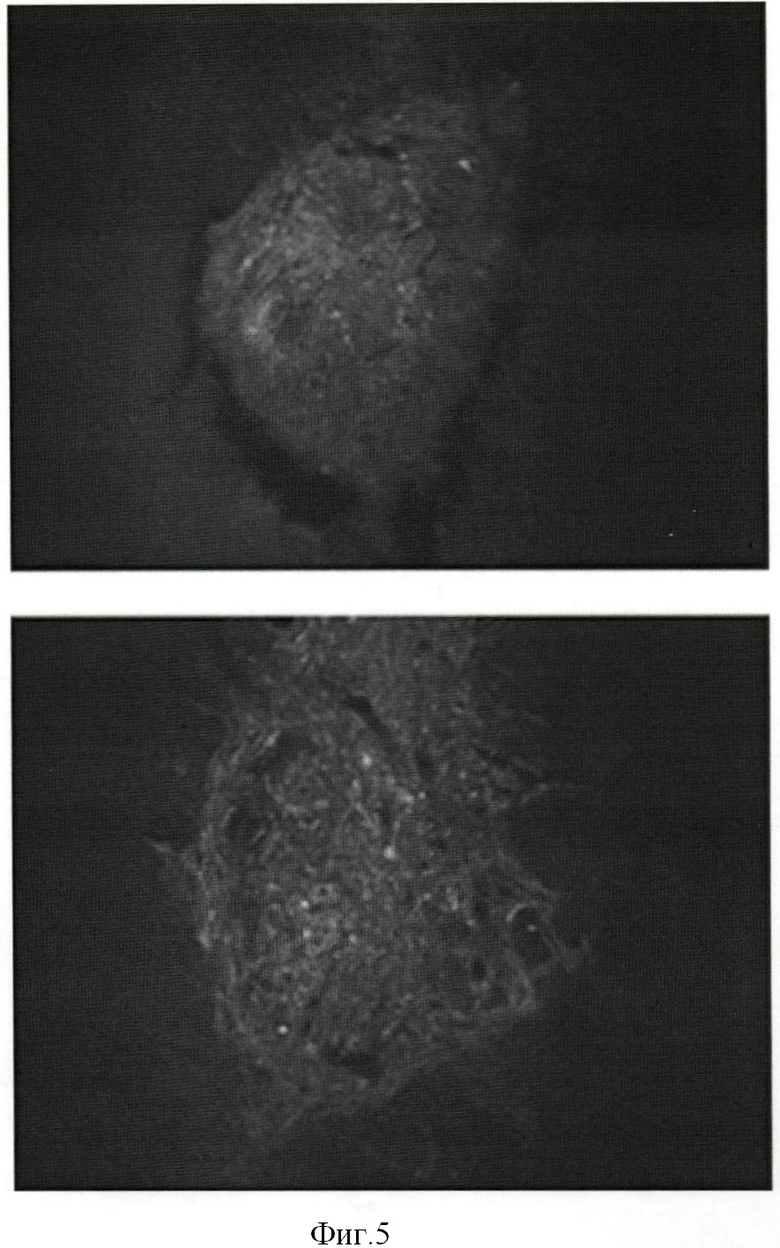
Фиг.5. Распределение GFP-меченных клеток в стриатуме мозга через 10 дней после трансплантации. Верхняя панель - индуцированная (BDNF) субпопуляция СКЖТ, не экспрессирующих TrkB; нижняя панель - индуцированная (BDNF) субпопуляция СКЖТ, экспрессирующих TrkB.
Осуществление изобретения
При осуществлении изобретения, помимо методов, подробно раскрытых в нижеследующих примерах, были использованы хорошо известные специалистам методики по культивированию клеток, описанные также в цитированных источниках.
Пример 1
Выделение стромальных клеток из жировой ткани
Для работы использовали СКЖТ человека и мыши. СКЖТ были выделены по протоколу Zuk et al. [19]. Подкожная жировая клетчатка была получена в ходе полостных операций у неонкологических пациентов, средний возраст которых составлял 40-60 лет. Мышиные СКЖТ были выделены из подкожного жира паховой области самцов линии Black6. Жировую ткань измельчали ножницами, затем обрабатывали ферментами и коллагеназой («Invitrogen», США, 30 ед. на мл ткани) и протеазой («Gibco», США, 200 ед. на мл ткани) в течение 40 мин при 37°С при постоянном помешивании. По прошествии этого времени в суспензию добавляли равный объем среды DMEM («Gibco», США) c 10% фетальной бычьей сывороткой (FBS, «Gibco», США) и центрифугировали 5-10 мин при 900 оборотов в минуту. Осадок, в котором находилась фракция стромально-васкулярных клеток, ресуспендировали в DMEM, с 10% FBS и фильтровали через 100-мм клеточный фильтр (BD Biosciences, Bedford, MA), далее клетки высаживали на культуральные чашки Петри («Costar Corning», США) и помещали в CO2-инкубатор при 37°С и 5%. На следующий день в чашках меняли среду. Таким образом, в культуре оставались только прикрепившиеся клетки. По достижении монослоя клетки снимали трипсином и рассаживали в соотношении 1:2.
Пример 2
Исследование суммарной популяции СКЖТ на наличие клеток, экспрессирующих рецептор TrkB
а) Определение содержания мРНК TrkB
Cуммарную РНК из клеток выделяли с помощью набора для выделения РНК («RNeasy Mini Kit», «QIAGEN», США) согласно протоколу фирмы-производителя. Все процедуры по выделению РНК и синтезу кДНК проводили на льду в ламинарном боксе. Концентрацию тотальной РНК определяли на спектрофотометре Bio Photometer (Eppendorf). Качество выделенной РНК контролировали с помощью электрофореза в 1% агарозном геле. Затем на матрице синтезировали к ДНК согласно стандартному протоколу фирмы «Fermentas», используя олиго-dT-праймеры. Для этого РНК смешивали с Oligo (dT)18 primer (Fermentas, 0,5 мкг/мкл) (на каждые 10 мкл РНК необходимо добавить 1 мкл Oligo (dT)18 primer). Смесь инкубировали 5 минут при 70°С. После инкубации к смеси добавляли 4 мкл 5х reaction buffer (Fermentas) и 2 мкл 10 mM dNTP mix (Fermentas) (на каждые 10 мкл РНК). Смесь инкубировали 5 мин при 37°С. После инкубации в смесь добавляли 2 мкл M-Mul V reverse transcriptase (Fermentas, 20 u/µl) (на каждые 10 мкл PНK). Полученную смесь инкубировали 60 мин при 37°С. Останавливали реакцию нагреванием смеси до 70°С в течение 10 минут. Синтезированную кДНК замораживали и хранили на - 20°С.
Анализ экспрессии гена TrkB проводили методом ПРЦ в реальном времени с применением Sybr Green («Синтол», Россия). При этом использовали праймеры и температуры отжига, представленные в таблице 1.
Результаты электрофоретического анализа ПЦР-продукта (Фиг.1) однозначно свидетельствовали о том, что СКЖТ второго пассажа содержат мРНК TrkB.
б) Цитофлуориметрия
Полученные СКЖТ были проанализированы на наличие экспрессии рецептора TrkB на поверхности анализируемых клеток. Для этого суспензию одиночных клеток (0,5-1 X 106 клеток) нулевого пассажа в 500 µl буфера PBS (phosphate buffered saline) инкубировали с первичными антителами против TrkB (Abeam, # ab51190) в течение 40 минут при комнатной температуре. После трехкратной отмывки в PBS проводили инкубацию клеток со вторичными антителами против антител кролика (Jacson Immunoresearch, # 111-175-144), мечеными Су5, в течение 40 минут. Флуоресценцию клеток после окрашивания оценивали методом проточной цитометрии на приборе MoFlo (Dako Cytomation) и анализировали с помощью программы Summit 4.1. В каждом эксперименте для подтверждения специфичности окрашивания использовался изотипический иммуноглобулиновый контроль: клетки, инкубированные с неспецифическими иммуноглобулинами, а затем со вторичным антителами. В результате анализа более 300000 клеток было подтверждено наличие среди них клеток, экспрессирующих рецептор TrkB. По данным проточной цитометрии доля таких клеток в суммарной популяции СКЖТ составляла 6±2,5%.
в) Оценка способности выделенных СКЖТ к нейральной дифференцировке методом иммунофлуоресцентного окрашивания
Для оценки способности выделенных клеток к нейральной дифференцировке определялось содержание в них основных маркерных белков после индукции in vitro. В эксперименте использовали СКЖТ 2-го пассажа. Клетки в среде, описанной в примере 1, высаживали на 8-луночные стеклянные плашки (Lab-Tek Chamber Slide, Nalge Nunc Int.) и на следующий день добавляли BDNF (20 нг/мл) или ретиноевую кислоту (1mM) в сочетании с 5-азацитидином (1mM). Через 3 дня индуцирующую среду меняли на свежую, также содержащую соответствующие индукторы, культивировали еще 4 дня и фиксировали для анализа методом иммунофлуоресцентного окрашивания. Фиксацию клеток проводили в 4% растворе формалина (Sigma, США) 2 мин при комнатной температуре с последующей 3-кратной отмывкой PBS. После отмывки клетки покрывали 1% раствором БСА (бычий сывороточный альбумин) c 10% козьей сывороткой в течение 30 мин и инкубировали при комнатной температуре с кроличьими антителами против нестина (Chemicon, 1:100), Eno2 (Chemicon, 1:50) и мышиными против тубулина (Abcam, 1:100) в течение 1 часа. В качестве отрицательного контроля использовался 1% раствор БСА c 10% козлиной сывороткой. После отмывки в PBS клетки в течение 30 мин при комнатной температуре инкубировали в растворе PBS с флуоресцентно меченными вторичными козьими антителами (1:100) против иммуноглобулинов мыши (Alexa 488, Molecular Probes, США) или против иммуноглобулина кролика (Alexa 568, Molecular Probes, США). При этом исходно в выделенных СКЖТ наблюдали либо отсутствие экспрессии нестина, либо низкий уровень экспрессии нестина и β3-тубулина, а после инкубации клеток в присутствии BDNF и 5-азацитидина наблюдали увеличение содержания обоих маркерных белков, что свидетельствовало о потенциальной возможности индукции нейральной дифференцировки в полученной популяции клеток (данные не представлены).
Пример 3
Получение субпопуляции TrkB-содержащих клеток из суммарной популяции СКЖТ
Фракцию клеток, несущих TrkB, из суммарной популяции СКЖТ получали с помощью иммуномагнитной селекции или проточной цитофлуориметрии.
Для иммуномагнитного сортинга клеток использовали магнитные частицы размером 40 нм, коньюгированные с антителами против кроличьих IgG, (Dynabeads®Pan Rabbit IgG, prod. No 110.41). Магнитные частицы отмывали от азида натрия и инкубировали с кроличьими антителами против TrkB человека в течение 40 минут на льду при постоянном помешивании. После окончания инкубации пробирку с магнитными частицами помещали в магнит и отмывали от несвязавшихся антител. Все манипуляции проводили в стерильных условиях культурального бокса. Затем подготовленные магнитные частицы инкубировали с суспензией СКЖТ в течение 1 часа при постоянном покачивании на льду. После инкубации клетки помещали в магнит и разделяли на две фракции: связавшиеся с магнитными частицами TrkB-позитивные клетки и оставшиеся в суспензии TrkB-негативные клетки. Чистота полученной фракции TrkB-экспрессирующих клеток составляла более 90%.
Сортинг клеток с использованием метода проточной цитофлуориметрии проводили, как описано в примере 26. Клетки разделяли в режиме сортировки на две популяции, экспрессирующих и не экспрессирующих рецептор TrkB. Чистота полученной фракции TrkB-позитивных клеток составляла 90%-98%.
Пример 4
Сравнительный анализ экспрессии маркерных генов в субпопуляции Trk-позитивных и субпопуляции TrkB- негативных СКЖТ
Суммарную РНК получали, как описано в примере 2а. Анализ экспрессии генов проводили методом ПРЦ в реальном времени с применением Sybr Green («Синтол», Россия).
Для каждого исследуемого гена готовили смесь праймеров forward и reverse в экспериментально подобранных количествах. Для приготовления одной пробы использовали 10 мкл реакционной смеси, содержащей SYBR Green (Синтол), 13 мкл ddH2O (Синтол), 1 мкл смеси праймеров и 1 мкл кДНК ДНК исследуемых клеток. Для приготовления NTC (non-template control) проб вместо кДНК добавляли 1 мкл ddH2O (Синтол). ПЦР с детекцией в режиме реального времени проводили на приборе IQ5 (BioRad) с помощью программного обеспечения IQ5 2.0 Optical system software (BioRad). Температура отжига для смеси праймеров для каждого исследуемого гена также была предварительно подобрана экспериментально.
Последовательности праймеров и температуры отжига для каждого из тестируемых генов представлены в таблице 1.
При этом было установлено, что в субпопуляции TrkB- позитивных СКЖТ представлена мРНК таких маркерных генов нейральной дифференцировки, как нестин и β3 тубулин (Фиг.3). В результате анализа субпопуляции клеток, не содержащих специфического рецептора BDNF (TrkB), также было выявлено присутствие мРНК указанных генов, однако количество соответствующих мРНК в этих клетках было в несколько раз ниже, чем в клетках, экспрессирующих TrkB (Фиг.3). Следует отметить, что в TrkB-позитивных клетках также было обнаружено более высокое содержание мРНК нейротрофических факторов роста, в частности HGF и BDNF.
Пример 5
Индукция нейральной дифференцировки in vitro и исследование показателей нейральной дифференцировки клеток, экспрессирующих TrkB
а) Проверка эффективности различных индукторов дифференцировки
Нейральная дифференцировка клеток сопровождается повышением экспрессии специфичных генов. В частности, клетки, дифференцирующиеся в нейральном направлении, содержат белок цитоскелета нестин (маркер ранних нейральных предшественников), нейрональную форму тубулина-β3, глиальный кислый фибриллярный белок GFAP (маркер астроцитарной глии), белок, взаимодействующий с микротрубочками МАР2, специфичную для нейронов енолазу Eno2 и другие маркеры нейральной дифференцировки.
Нами было проанализировано содержание продуктов перечисленных выше маркерных генов нейральной дифференцировки в СКЖТ, инкубированных в присутствии различных индукционных факторов, включая IBMX, Р-меркаптоэтанол, 5-азацитидин [1, 4, 11], а также нейротрофических факторов роста BDNF и GDNF (таблица 1). Продукт гена GFAP не детектировался ни в контрольных СКЖТ, ни в клетках, инкубированных в присутствии индукторов, перечисленных в таблице 1. Анализ содержания мРНК других маркерных генов показал, что уровень экспрессии Eno2, МАР2, TUBB и Nestin был выше в клетках, инкубированных в присутствии ретиноевой кислоты или BDNF на фоне 5-азацитидина (таблица 2).
Однако в СКЖТ, культивированных в присутствии других индукторов нейральной дифференцировки, увеличения уровня мРТПС маркерных генов не наблюдалось (таблица 3).
На основании полученных результатов для индукции нейральной дифференцировки СКЖТ были выбраны сочетания ретиноевой кислоты или BDNF с 5-азацитидином. При индукции дифференцировки СКЖТ, полученных от 8 доноров, было установлено, что при культивировании клеток в присутствии ретиноевой кислоты в сочетании с 5-азацитидином в них возрастает уровень мРНК нестина и тубулина. Индукция нейральной дифференцировки клеток с помощью сочетания BDNF с 5-азацитидином вызывает увеличение экспрессии всех 4 проанализированных генов (таблица 3).
Эти данные были подтверждены с помощью иммунофлуоресцентного окрашивания индуцированных клеток. Так, было обнаружено, что инкубация клеток с ретиноевой кислотой приводит к повышению содержания в них нестина и β3-тубулина, а в СКЖТ, культивированных в присутствии BDNF, возрастает содержание как нестина и β3-тубулина, так и специфичной для нейронов енолазы Eno2.
б) Определение содержания мРНК до и после индукции
Обе субпопуляции клеток СКЖТ 2-го пассажа (TrkB-позитивые и ТгкВ-негативные), полученные в результате сортинга, как описано в примере 3, высаживали в 6-луночный планшет в плотности 40-50% монослоя в среду DMEM с добавлением 10% FBS. На следующий день среду заменяли на DMEM/3% FBS, к которой добавляли BDNF в конечной концентрации 20 нг/мл в сочетании с 5-азацитидином (1 мM). Через 3 дня индуцирующую среду меняли на свежую, также содержащую индуктор, культивировали еще 4 дня и далее проводили определение содержания мРНК, как описано в примере 4.
Было установлено, что обе субпопуляции клеток демонстрируют после индукции BDNF in vitro возрастание уровня мРНК нестина, тубулина, белка MAP и нейронспецифичной енолазы. Однако индукция нейральной дифференцировки TrkB-экспрессирующих клеток на уровне транскрипции названных генов была выражена в значительно большей степени, чем в клетках, не экспрессирующих этот рецептор. В субпопуляции TrkB-позитивных СКЖТ уже через 3 дня количество мРНК нестина, тубулина, Eno2 и МАР-2 увеличивалось в 3-6 раз. Через 7 дней после индукции в этих клетках также возрастало содержание соответствующих белков.
Следует отметить, что аналогичный, хотя и несколько менее выраженный, эффект наблюдался при индукции клеток ретиноевой кислотой (1 мМ) в сочетании с 5-азацитидином (1 мМ).
в) Определение содержания специфических антигенов методом иммунофлуоресцентного окрашивания
TrkB-экспрессирующие и TrkB-неэкспрессирующие СКЖТ 2-го пассажа клетки высаживали на 8-луночные стеклянные плашки (Lab-Tek Chamber Slide, Nalge Nunc Int.) в среде DMEM с добавлением 3% FBS. На следующий день добавляли BDNF в той же концентрации, что и в примере 5. Через 3 дня индуцирующую среду меняли на свежую, также содержащую индуктор, культивировали еще 4 дня и фиксировали для анализа. Фиксацию проводили в 4% растворе формалина (Sigma, США) в течение 2 мин при комнатной температуре с последующей 3-кратной отмывкой PBS. После отмывки клетки инкубировали в 1% растворе БСА (бычий сывороточный альбумин) с 10% козьей сывороткой в течение 30 мин, далее инкубировали с кроличьими антителами против Nestin (Chemicon, 1:100), Eno2 (Chenicon, 1:50) и мышиными против β3-tubulin (Abeam, 1:100) в течение 1 часа при комнатной температуре. После отмывки в PBS клетки в течение 30 мин инкубировали в растворе PBS с флуоресцентно-меченными вторичными козьими антителами (1:100) против иммуноглобулина мыши (Alexa 488, Molecular Probes, США) или против иммуноглобулина кролика (Alexa 568, Molecular Probes, США). В качестве контроля использовали клетки, окрашенные только вторичными антителами. Полученные результаты показали, что в TrkB-экспрессирующих СКЖТ, культивированных в присутствии BDNF и 5-азацитидина, возрастает содержание нестина и β3-тубулина, а также специфичной для нейронов енолазы Еno2. В TrkB-неэкспрессирующих клетках достоверных отличий в содержании нестина, β3-тубулина и енолазы Eno2 до и после индукции не наблюдалось (данные не приводятся).
В аналогичном эксперименте с индукцией ретиноевой кислотой наблюдалось повышение содержания двух белков: нестина и β3-тубулина.
Пример 6
Трансплантация индуцированных мышиных СКЖТ в стриатум мыши
Проведено сравнение способности клеток СКЖТ TrkB-позитивных и TrkB-негативных интегрироваться в ткани мозга после трансплантации. Для этого свежевыделенные СКЖТ мышей линии В16, трансгенных по гену GFP, сортировали по экспрессии TrkB, получая TrkB-позитивные и TrkВ-негативные субпопуляции клеток, индуцировали их нейральную дифференцировку, как описано в примере 5, и далее инъецировали в стриатум мышей линии В16 при помощи шприца.
Через 10 дней мышей перфузировали 4% формалином, извлекали головной мозг и фиксировали 3-5 часов в 4% формалине, затем инкубировали 10-16 часов в 30% сахарозе и замораживали. Далее делали срезы толщиной 20 мкм на криостате (НМ 505 Е, Microm, Германия). Готовые препараты хранили при +4°С в темноте. Полученные препараты (Фиг.5) анализировали при помощи флуоресцентного микроскопа Axiovert 200М (Zeiss). Документирование изображений производили с помощью цифровой видеокамеры Axiocam HRC (Zeiss, Германия) и обработки изображения в программе Axiovision 3.1. Индуцированные in vitro СКЖТ клетки, не экспрессирующие TrkB, за указанное время не обнаруживали вступления в контакт с тканью реципиента и оставались в области трека. За тот же период времени клетки, экспрессирующие TrkB, будучи индуцированы в направлении нейральной дифференцировки, демонстрировали выраженную миграцию в паренхиму мозга (Фиг.5).
Статистический анализ всех данных проводился с использованием программы SigmaStat 9.0. Для сравнения маленьких групп и ненормальных распределений использовался U-критерий Манна-Уитни. Различия считались статистически значимыми при уровне значимости р<0,05.
Литература
1. Ashjian, Р.Н., et al., In vitro differentiation of human processed lipoaspirate cells into early neural progenitors. Plast Reconstr Surg, 2003. 111(6): p.1922-31.
2. Balmer, J.E. and R. Blomhoff, Gene expression regulation by retinoic acid. J Lipid Res, 2002. 43(11): p.1773-808.
3. Brzoska, M., et al, Epithelial differentiation of human adipose tissue-derived adult stem cells. Biochem Biophys Res Commun, 2005. 330(1): p.142-50.
4. Fujimura, J., et al., Neural differentiation of adipose-derived stem cells isolated from GFP transgenic mice. Biochem Biophys Res Commun, 2005. 333(1): p.116-21.
5. Garcia-Suarez, O., et al., Expression of the neurotrophin receptor TrkB in the mouse liver. Anat Embryol (Berl), 2006. 211(5): p.465-73.
6. Gronthos, S., et al., Surface protein characterization of human adipose tissue-derived stromal cells. J Cell Physiol, 2001. 189(1): p.54-63.
7. Kim, В J., et al., Differentiation of adult bone marrow stem cells into neuroprogenitor cells in vitro. Neuroreport, 2002. 13(9): p.1185-8.
8. Lim, J.Y., et al., Brain-derived neurotrophic factor stimulates the neural differentiation of human umbilical cord blood-derived mesenchymal stem cells and survival of differentiated cells through MAPK/ERK and PI3K/Akt-dependent signaling pathways. J Neurosci Res, 2008.
9. Miranville, A., et al, Improvement of postnatal neovascularization by human adipose tissue-derived stem cells. Circulation, 2004. 110(3): p.349-55.
10. Mizuno, H., et al., Myogenic differentiation by human processed lipoaspirate cells. Plast Reconstr Surg, 2002. 109(1): p.199-209; discussion 210-1.
11. Ning, H., et al., Neuron-like differentiation of adipose tissue-derived stromal cells and vascular smooth muscle cells. Differentiation, 2006. 74(9-10): p.510-8.
12. Planat-Benard, V., et al., Spontaneous cardiomyocyte differentiation from adipose tissue stroma cells. Circ Res, 2004. 94(2): p.223-9.
13. Planat-Benard, V., et al., Plasticity of human adipose lineage cells toward endothelial cells: physiological and therapeutic perspectives. Circulation, 2004. 109(5): p.656-63.
14. Reichardt, L.F., Neurotrophin-regulated signalling pathways. Philos Trans R Soc bond В Biol Sci, 2006. 361(1473): p.1545-64.
15. Rivera, F.J., et al., Adult hippocampus derived soluble factors induce a neuronal-like phenotype in mesenchymal stem cells. Neurosci Lett, 2006. 406(1-2): p.49-54.
16. Seo, M.J., et al., Differentiation of human adipose stromal cells into hepatic lineage in vitro and in vivo. Biochem Biophys Res Commun, 2005. 328(1): p.258-64.
17. Yan, Q., et al, Immunocytochemical localization of TrkB in the central nervous system of the adult rat. J Comp Neurol, 1997. 378(1): p.135-57.
18. US 7,078,230; опубликован 18.07.2006.
19. Anghileri, E., et al., Neuronal differentiation potential of human adipose-derived mesenchymal stem cells. Stem Cells Dev, 2008. 17: p.909-916.
| название | год | авторы | номер документа |
|---|---|---|---|
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях теплового шока, и способ ее получения | 2019 |
|
RU2752906C2 |
| ПОЛУЧЕНИЕ ТЕРМИНАЛЬНО ДИФФЕРЕНЦИРОВАННЫХ ДОФАМИНЕРГИЧЕСКИХ НЕЙРОНОВ ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2004 |
|
RU2345133C2 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях гипоксии, и способ ее получения | 2019 |
|
RU2732600C1 |
| КОНВЕРСИЯ СОМАТИЧЕСКИХ КЛЕТОК В ИНДУЦИРОВАННЫЕ РЕПРОГРАМИРОВАННЫЕ НЕЙРОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ (ИРНСК) | 2011 |
|
RU2562111C2 |
| ПРИМЕНЕНИЕ СТВОЛОВЫХ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ МЕЗЕНХИМАЛЬНЫЕ И НЕЙРОНАЛЬНЫЕ МАРКЕРЫ, И ИХ КОМПОЗИЦИЙ ДЛЯ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ (ВАРИАНТЫ) | 2017 |
|
RU2742828C2 |
| СТВОЛОВЫЕ КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ МЕЗЕНХИМАЛЬНЫЕ И НЕЙРОНАЛЬНЫЕ МАРКЕРЫ, ИХ КОМПОЗИЦИИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2017 |
|
RU2741839C2 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА В ДОФАМИНЭРГИЧЕСКИЕ НЕЙРОНЫ | 2023 |
|
RU2823290C1 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из глиальных прогениторных клеток в условиях теплового шока, и способ её получения | 2019 |
|
RU2732599C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК МЛЕКОПИТАЮЩИХ, МОДИФИЦИРОВАННЫХ ГЕНОМ ФАКТОРА РОСТА НЕРВОВ ЧЕЛОВЕКА | 2011 |
|
RU2458984C1 |
| ПОЛУЧЕНИЕ ГАМКергических НЕЙРОНОВ in vitro ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ НЕВРОЛОГИЧЕСКИХ РАССТРОЙСТВ | 2004 |
|
RU2380410C2 |

Изобретение относится к области биотехнологии, конкретно к получению клеточных популяций, и может быть использовано в клеточной трансплантологии и тканевой инженерии с целью получения клеточного материала для восстановления нервной ткани, поврежденной в результате травм, инсультов или нейродегенеративных заболеваний. Способ заключается в выделении популяции стромальных клеток из жировой ткани (СКЖТ), иммуносортинге, основанном на использовании антител против рецептора (TrkB) к нейротрофическому фактору мозга (BDNF), с получением по существу гомогенной субпопуляции TrkB-экспрессирующих клеток, культивировании их в стандартной среде для клеток млекопитающих и индукции дифференцировки путем переноса в среду, содержащую индуктор нейральной дифференцировки - BDNF в сочетании с 5-азацитидином или ретиноевую кислоту в сочетании с 5-азацитидином. Изобретение позволяет получить популяцию стромальных клеток жировой ткани, индуцированных к нейральной дифференцировке. 6 з.п. ф-лы, 5 ил., 4 табл.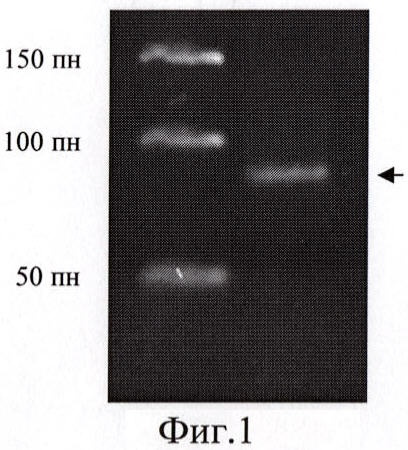
1. Способ получения популяции стромальных клеток жировой ткани, индуцированных к нейральной дифференцировке, включающий выделение популяции стромальных клеток из жировой ткани (СКЖТ), культивирование их в стандартной среде для клеток млекопитающих и индукцию дифференцировки путем переноса в среду, содержащую индуктор нейральной дифференцировки, и отличающийся тем, что после выделения суммарной популяции СКЖТ ее подвергают иммуносортингу, основанному на использовании антител против рецептора (TrkB) к нейротрофическому фактору мозга (BDNF), с получением, по существу, гомогенной субпопуляции TrkB-экспрессирующих клеток, которые далее культивируют и индуцируют в направлении нейральной дифференцировки, используя в качестве индуктора BDNF в сочетании с 5-азацитидином или ретиноевую кислоту в сочетании с 5-азацитидином.
2. Способ по п.1, отличающийся тем, что в качестве индуктора используют BDNF в сочетании с 5-азацитидином.
3. Способ по п.2, отличающийся тем, что концентрация BDNF и 5-азацитидина в дифференцировочной среде составляет 20нг/мл и 1 мМ соответственно.
4. Способ по п.1, отличающийся тем, что в качестве индуктора используют ретиноевую кислоту в сочетании с 5-азацитидином.
5. Способ по п.4, отличающийся тем, что концентрация ретиноевой кислоты и 5-азацитидина в дифференцировочной среде составляет 1 мМ и 1 мМ соответственно.
6. Способ по п.1, отличающийся тем, что иммуносортинг осуществляют с помощью иммуномагнитной селекции или проточной цитофлуориметрии.
7. Способ по любому из пп.1-6, отличающийся тем, что культивирование клеток проводят в среде DMEM с 10%-ной фетальной бычьей сывороткой, а индукцию - в той же среде с добавлением индуктора.
| US 7078230, 18.07.2006 | |||
| ANGHILERI E | |||
| ET AL., Neuronal differentiation potential of human adipose-derived mesenchymal stem cells, Stem Cells Dev., 2008, v.l7, n.5, p.909-916 | |||
| ЛОПАТИНА Т.В | |||
| и др | |||
| Индукция нейральной дифференцировки стромальных клеток жировой ткани нейротрофическими факторами и ретиноевой кислотой | |||
| - Тезисы V Конференции молодых |

