Область изобретения
Изобретение относится к получению интересующих пептидов и белков в растениях-хозяевах путем аккумулирования в эндоплазматическом ретикулуме растений, производящем белковые тельца, к нуклеиновой кислоте, кодирующей такие продукты, и к использованию указанных нуклеиновых кислот для производства конструкций и векторов для трансформирования систем растений-хозяев. В частности, раскрыт способ экспрессии и выделения интересующих гетерологичных продуктов, таких как кальцитонин (СТ), в растениях.
Предпосылки создания изобретения
Так как предполагается, что потребность в биофармацевтических препаратах значительно возрастет, благодаря замечательным достижениям в знании генома и в соответствующих биомедицинских исследованиях, то имеется значительный интерес к разработке недорогих рекомбинантных продукционных систем.
Генная инженерия растений для продуцирования биофармацевтических препаратов является относительно новой, в то время как другие трансгенные системы, включая бактерии, грибки и культивируемые клетки млекопитающих, широко и долгое время адаптировали для биопродуцирования. Тем не менее, некоторые рекомбинантные терапевтические белки, использующие растительные экспрессионные системы, уже имеются на рынке или находятся на разных стадиях клинических испытаний на человеке, например гирудин, антикоагулянтный белок для лечения тромбоза (Parmenter et al., 1995), химерная IgG-IgA вакцина против зубного кариеса (Ма et al., 1998), бактериальный вакциноген против энтеротоксигенного штамма Е. coli (Haq et al., 1995), и рекомбинантная желудочная липаза собак для лечения кистозного фиброза (Benicourt et al., 1993).
Растительные экспрессионные системы привлекательны потому, что уровень экспрессии рекомбинантных белков может быть повышен путем использования природных механизмов сортировки и нацеливания, которые используют растения для нацеливания хозяйских белков к органеллам. Кроме того, биофармацевтические препараты, произведенные растениями, могут быть легко увеличены до массового производства и имеют преимущество, обусловленное минимальным риском для здоровья, возникающим из-за контаминации патогенами или токсинами.
Растения оказываются все более и более привлекательными экспрессионными системами из-за их потенциала, предоставляющего неограниченные количества биологически активного материала при низкой стоимости производства и с пониженным риском для здоровья. Способность растений аккумулировать высокие уровни рекомбинантных белков и осуществлять большинство из посттрансляционных модификаций позволяет рассматривать их в качестве биореакторов для молекулярного выращивания рекомбинантных терапевтических препаратов (см. обзор Fischer and Emans, 2000). Однако важные решения, касающиеся выбора видов культур, выбора тканей, стратегий экспрессии и выделения и посттрансляционной обработки, являются определяющими для осуществимости коммерческого производства на основе растений (Cramer et al., 1999).
Внутриклеточное нацеливание рекомбинантных белков является важным условием для высокого уровня аккумулирования и правильной сборки и складывания таких белков в растениях. Компартментализация хозяйских белков во внутриклеточных накопительных органеллах в основном достигается путем использования подходящих сигнальных пептидов или целых слияний белков. Ряд рекомбинантных терапевтических белков был направлен к последующим компартментам растений: апопластическое пространство (McCormick et al., 1999), хлоропласта (Staub et al., 2000), эндоплазматический ретикулум (ЭР) (Stoger et al., 2000). Иммуноглобулины, направленные в ЭР компартмент в трансгенных растениях, оказалось, дают в 10-100 раз большие выходы, чем направленные к другим компартментам, таким как апопласт или цитозоль (Conrad and Fiedler, 1998).
Нацеливание сложных белков, таких как антитела, в ЭР компартмент особенно интересно, потому что большинство посттрансляционных модификаций, требуемых для получения функционального продукта, происходят внутри ЭР (Düring et al., 1990; Ma and Hein, 1995; Conrad and Fiedler, 1998). Несомненно, внутри ЭР сигнальный пептид расщепляется и напряженные белки, такие как связывающий IgG белок (BiP) и энзимы, такие как протеиндисульфидизомераза (PDI), функционируют как компаньоны, связывают несобранный белок и направляют последующее накопление и сборку. Вдобавок к этим особым характеристикам полезно указать, что растительный ЭР является высокоэластичным, что делает его идеальным резервуаром для гетерологичных фармацевтических белков. ЭР, даже если он появляется на входе секреторного пути, также способен сохранять белки в течение короткого или длинного периода времени. Растения сохраняют аминокислоты в течение долгого периода в форме специфических запасных белков. Одним из механизмов защиты этих запасных белков от неконтролируемого преждевременного разрушения является помещение их в ЭР-производные накопительные органеллы, называемые белковыми тельцами (БТ) (см. Müntz, 1998). Сборка таких органелл как простое аккумулирование рекомбинантных белков в ЭР просвете требует, в качестве первого шага, сохранения хозяйского белка. Секреторные белки, верно накопленные и собранные в ЭР, имеют ряд клеточных назначений, главным образом с помощью продвижения через аппарат Гольджи. Однако ЭР сохранение растворимых транспорт-компетентных белков может быть вызвано с помощью карбокси-концевой сохраняющей/восстанавливающей сигнальной KDEL (или HDEL) (Munro et al., 1987; Wandelt et al., 1992; Vitale et al., 1993). Это консервативное С-концевое звено, распознаваемое в аппарате Гольджи через трансмембранные рецепторы, допускает рециклинг выделенных ЭР резидентных белков обратно в ЭР (Vitale and Denecke, 1999; Yamamoto et al., 2001). Многие фрагменты рекомбинантных антител были удлинены KDEL сигналом для того, чтобы стабильно аккумулировались в растительных ЭР (Vitale et al., 1998; Torres et al., 1999). Альтернативным путем генерированию сохранения и аккумулирования рекомбинантных белков в ЭР компартменте является создание подходящего слияния с природным ЭР резидентом, таким как запасной белок семени.
WO 01/75312 раскрывает способ продуцирования цитокина в системе растения-хозяина, где указанная система растения-хозяина трансформирована химерной последовательностью нуклеиновой кислоты, кодирующей указанный цитокин, указанную химерную последовательность нуклеиновой кислоты, содержащую первую последовательность нуклеиновой кислоты, способную к регуляции транскрипции в указанной системе растения-хозяина второй последовательности нуклеиновой кислоты, где указанная вторая последовательность нуклеиновой кислоты кодирует сигнальную последовательность, которая связана в рамке считывания с третьей последовательностью нуклеиновой кислоты, кодирующей цитокин, и четвертую последовательность нуклеиновой кислоты, связанную в рамке считывания с 3′ концом указанной третьей последовательности нуклеиновой кислоты, кодирующую "KDEL" аминокислотную последовательность.
Зеины являются группой белков, которые синтезируются во время развития эндосперма в зерне, и могут быть разделены на четыре группы α, β, γ и δ на основании их растворимости. Зеины могут объединяться в БТ прямо в ЭР. Растения или ткани растений, содержащие руминовые стабильные белковые тельца, экспрессируемые в виде слитых белков, содержащих полный белок зеина и операбельно связанный белковый материал, раскрыты в WO 00/40738.
γ-Зеин, запасный белок маиса, является одним из четырех маисовых проламинов и составляет 10-15% от общего белка в эндосперме маиса. Как и другие зерновые проламины, α и γ-зеины биосинтезируются в мембранно-связанных полисомах на цитоплазматической стороне шероховатого ЭР, собираются внутри просвета и затем секвестируются в ЭР-производных БТ (Herman and Larkins, 1999, Ludevid et al., 1984, Torrent et al., 1986). γ-Зеин состоит из четырех характеристических доменов: i) пептидный сигнал из 19 аминокислот, ii) повторяющийся домен, содержащий восемь единиц гексапептида PPPVHL (53 ак), iii) proX домен, где пролиновые остатки заменены другими аминокислотами (29 ак) и iv) гидрофобный богатый цистеином С-концевой домен (111 ак). Способность γ-зеина собираться в ЭР-производных БТ не ограничивается семенами. Действительно, когда ген γ-зеина коститутивно экспрессировали в трансгенные растения Arabidopsis, запасный белок аккумулировался внутри ЭР-производных БТ в клетках мезофилла листа (Geli et al., 1994). Рассмотрение сигнала, ответственного за помещение γ-зеина в ЭР-производные БТ (проламины не имеют KDEL сигнала), показало, что богатый пролином N-концевой домен, включающий тандем повторяющегося домена, необходим для сохранения ЭР, и что С-концевой домен вовлечен в образование БТ. Однако механизмы, с помощью которых эти домены стимулируют сборку БТ, все еще не известны.
Кальцитонин (СТ), гормональный пептид из 32 аминокислот, является необходимым для правильного метаболизма кальция и нашел широкое клиническое применение при лечении остеопороза, гиперкальцемического шока и болезни Паджета (Reginster et al., 1993; Azria et al., 1995; Silverman et al., 1997). Человеческий СТ синтезировали в виде препропротеина с сигнальным пептидом из 25 аминокислот и двух пропептидов при N- и С-концах (57 ак и 21 ак, соответственно). Результирующий активный пептид имеет в длину 32 аминокислоты с единственной дисульфидной связью (Cysl-Cys7) и амидирован по карбокси-концу. In vitro человеческий СТ собирается в агрегаты, что ограничивает его использование в качестве терапевтических средств. Поэтому лососевый СТ, который менее склонен к агрегации, обычно используется взамен (Cudd et al., 1995). Производство СТ в настоящее время осуществляется путем химического синтеза, но стоимость этого производства подтолкнула некоторые исследовательские группы к разработке альтернативных подходов. Человеческий и лососевый СТ были получены в Е. coli (Ray et al., 1993; Hong et al., 2000), в мышиных клетках (Merli et al., 1996), в нонендокриновых линиях клеток Cos-7 и СНО (Takahashi et al., 1997) и совсем недавно в молоке трансгенных кроликов (McKee et al., 1998). Получение биоактивного кальцитонина с помощью биотехнологических методов включает, по меньшей мере, два этапа: i) генерирование глицин-удлиненного кальцитонина (Bradbury et al., 1988) и ii) образование карбокси-концевого пролинамида посредством действия фермента амидирования, пептидилглицин α-амидирующей монооксигеназы (РАМ) (Eipper et al., 1992). Так как точно не известно, протекает ли карбокси-амидирование в клетках растений, in vitro амидирование растительного глицин-удлиненного кальцитонина с РАМ ферментом дало С-концевой амид (Ray et al., 1993).
Краткое изложение изобретения
Задача, решаемая настоящим изобретением, заключается в разработке альтернативной системы для получения интересующих пептидов и белков в системе растения-хозяина.
Представленное здесь решение основано на способности богатых пролином доменов γ-зеина к самосборке и обеспечению стабильности слитых белков в ЭР системы растения-хозяина. Использование системы, основанной на слитом белке γ-зеина, для аккумулирования интересующего продукта в системе растения-хозяина представляет успешный подход к аккумулированию указанного интересующего продукта внутри ЭР-производных БТ растений.
Изобретение иллюстрируется примерами, где описана основанная на слитом белке система аккумулирования рекомбинантного СТ в ЭР-производных БТ в растениях табака. Ряд богатых пролином доменов были выделены из γ-зеина для использования в качестве партнера слияния через расщепляемый протеазой сайт. Кодирующая область зрелого кальцитонина была слита с С-концом доменов γ-зеина и экспрессирована в трансгенные растения табака. Слитые белки аккумулировались в ЭР-производных БТ листьев табака. После очистки слитые белки подвергали расщеплению энтерокиназой, позволяющему высвобождение кальцитонина.
Соответственно, один из аспектов настоящего изобретения относится к последовательности нуклеиновой кислоты, содержащей (i) нуклеотидную последовательность, кодирующую белок γ-зеин, или ее фрагмент, содержащий нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в эндоплазматическом ретикулуме; (ii) нуклеотидную последовательность, кодирующую аминокислотную последовательность, которая специфически расщепляется с помощью ферментативных или химических способов; и (iii) нуклеотидную последовательность, кодирующую интересующий продукт; где указанные нуклеотидные последовательности являются оперативно связанными между собой.
В другом аспекте изобретение относится к конструкции нуклеиновой кислоты, содержащей указанную последовательность нуклеиновой кислоты.
В следующем аспекте изобретение относится к вектору, содержащему указанные последовательность или конструкцию, и к клетке, трансформированной указанным вектором.
В следующем аспекте изобретение относится к трансформированной системе растения-хозяина, имеющей указанные последовательность нуклеиновой кислоты, конструкцию или вектор.
В следующем аспекте изобретение относится к трансгенной системе растения-хозяина, содержащей интегрированную в ее геном указанную последовательность нуклеиновой кислоты.
В следующем аспекте изобретение относится к способу продуцирования интересующего продукта в системе растения-хозяина.
В следующем аспекте изобретение относится к способу продуцирования кальцитонина в системе растения-хозяина.
В следующем аспекте изобретение относится к слитому белку, причем указанный слитый белок имеет аминокислотную последовательность, соответствующую вышеуказанной последовательности нуклеиновой кислоты.
Краткое описание чертежей
На Фиг.1 показаны нуклеотидные последовательности и трансляции γ-зеина (Фиг.1.А) и производных γ-зеина RX3 (Фиг.1.В, верхняя), R3 (Фиг.1.В, нижняя), Р4 (Фиг.1.С, верхняя) и Х10 (Фиг.1.С, нижняя).
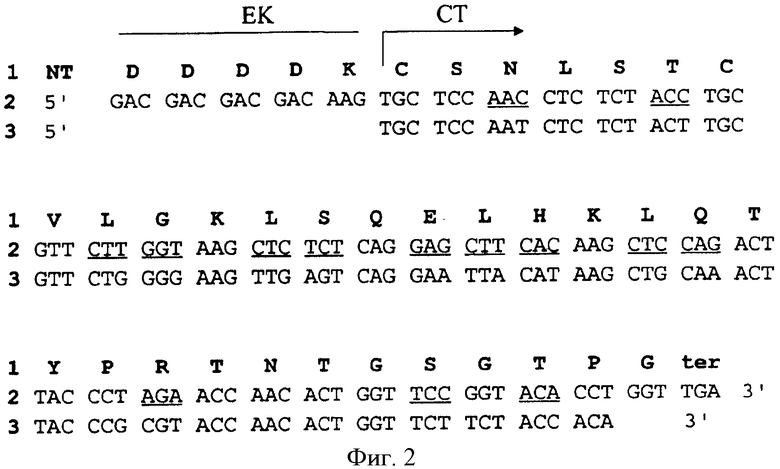
На Фиг.2 показаны нуклеотидная последовательность (полоса 2) и трансляции (полоса 1) синтетического кальцитонина (СТ). Ген синтетического кальцитонина был сконструирован с использованием предпочтительного кодона растения. Модификации кодона подчеркнуты в сравнении с геном лососевого СТ дикого типа (полоса 3). Синтетический ген содержит при 5′ конце линкерную последовательность, соответствующую сайту расщепления энтерокиназой (ЭК), и удлинен при 3′ для получения С-концевого глицина.
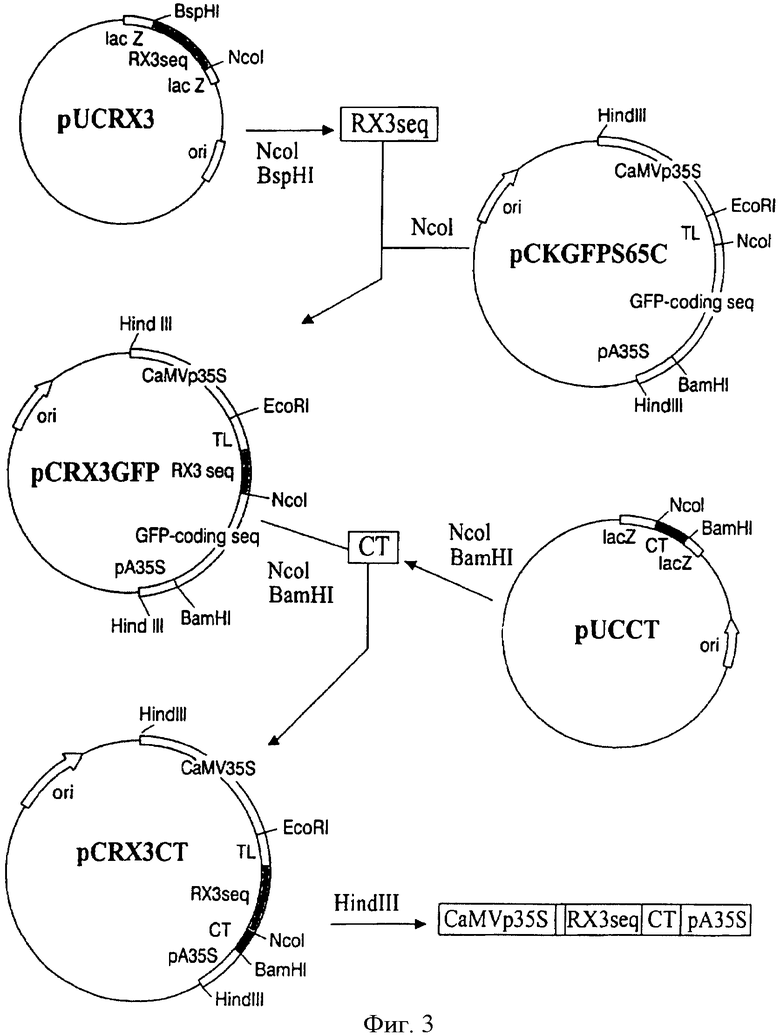
На Фиг.3 показана схема конструирования pCRX3CT плазмиды. Представленный процесс был таким же для получения следующих плазмид pCZeinCT, pCR3CT, рСР4СТ и рСХ10СТ, различие между ними состоит в том, введены соответствующий γ-зеин или производные от γ-зеина последовательности. Разные плазмиды изображены не в пропоции.
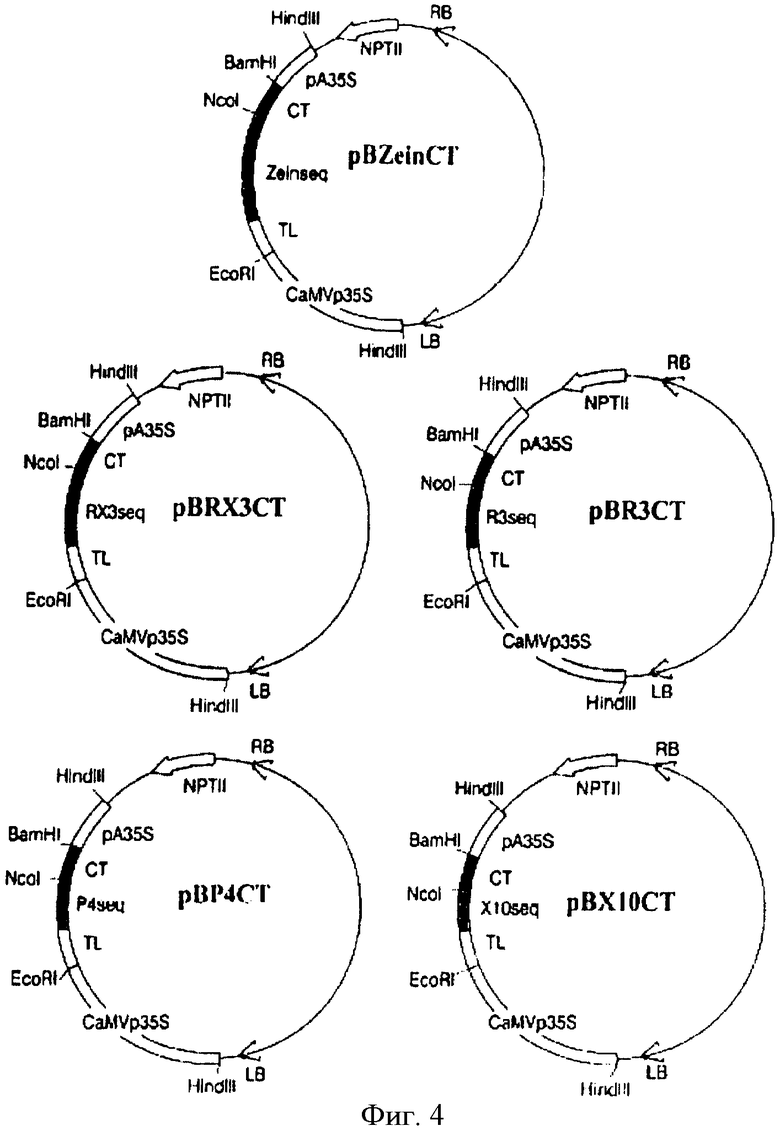
На Фиг.4 показано схематическое изображение плазмид pBZeinCT, рВRХ3СТ, pBR3CT, рВР4СТ и рВХ10СТ. Разные плазмиды изображены не в пропорции.
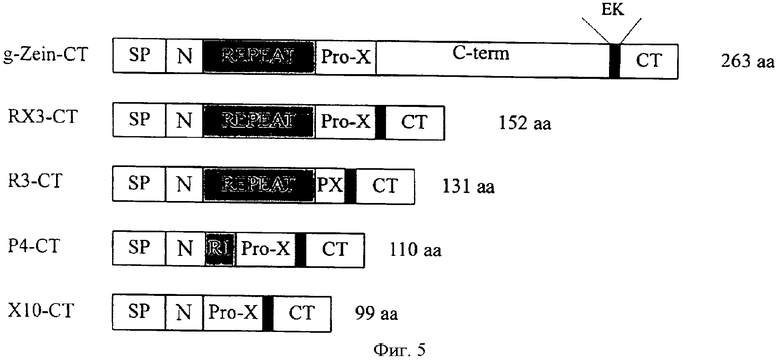
На Фиг.5 показано схематическое изображение различных слитых белков. Домены γ-зеина и производных γ-зеина (RX3, R3, Р4 и Х10) были слиты с кальцитонином (СТ) через сайт расщепления энтерокиназой (ЭК). SP - сигнальный пептид; REPEAT - восемь единиц повторяющегося домена (PPPVHL); RI - одна повторная единица; Рrо-Х - пролин-Хаа; РХ - фрагмент Pro-Х домена; C-term - богатый цистеином С-концевой домен; N - N-концевая последовательность зрелого белка. Число аминокислот для каждого слитого белка указано справа.
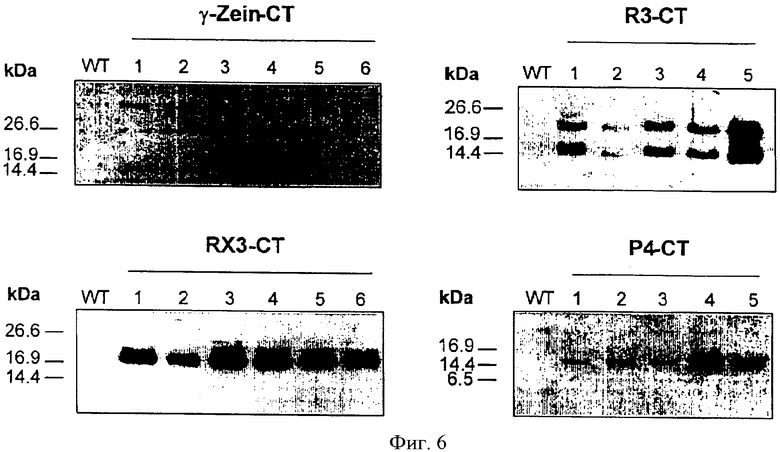
На Фиг.6 показаны результаты иммуноблот-анализа слитых белков в трансгенных растениях табака с использованием антисыворотки γ-зеина. Растворимые белки экстрагировали из листьев дикорастущего (WT) и трансгенного табака (То), разделяли на 15% SDS-полиакриламидном геле (20 мкг на полосу) и переносили в нитроцеллюлозу. Числа указывают независимые трансгенные линии, полученные для различных химерных генов: γ-zein-CT, RX3-CT, R3-CT, Р4-СТ.
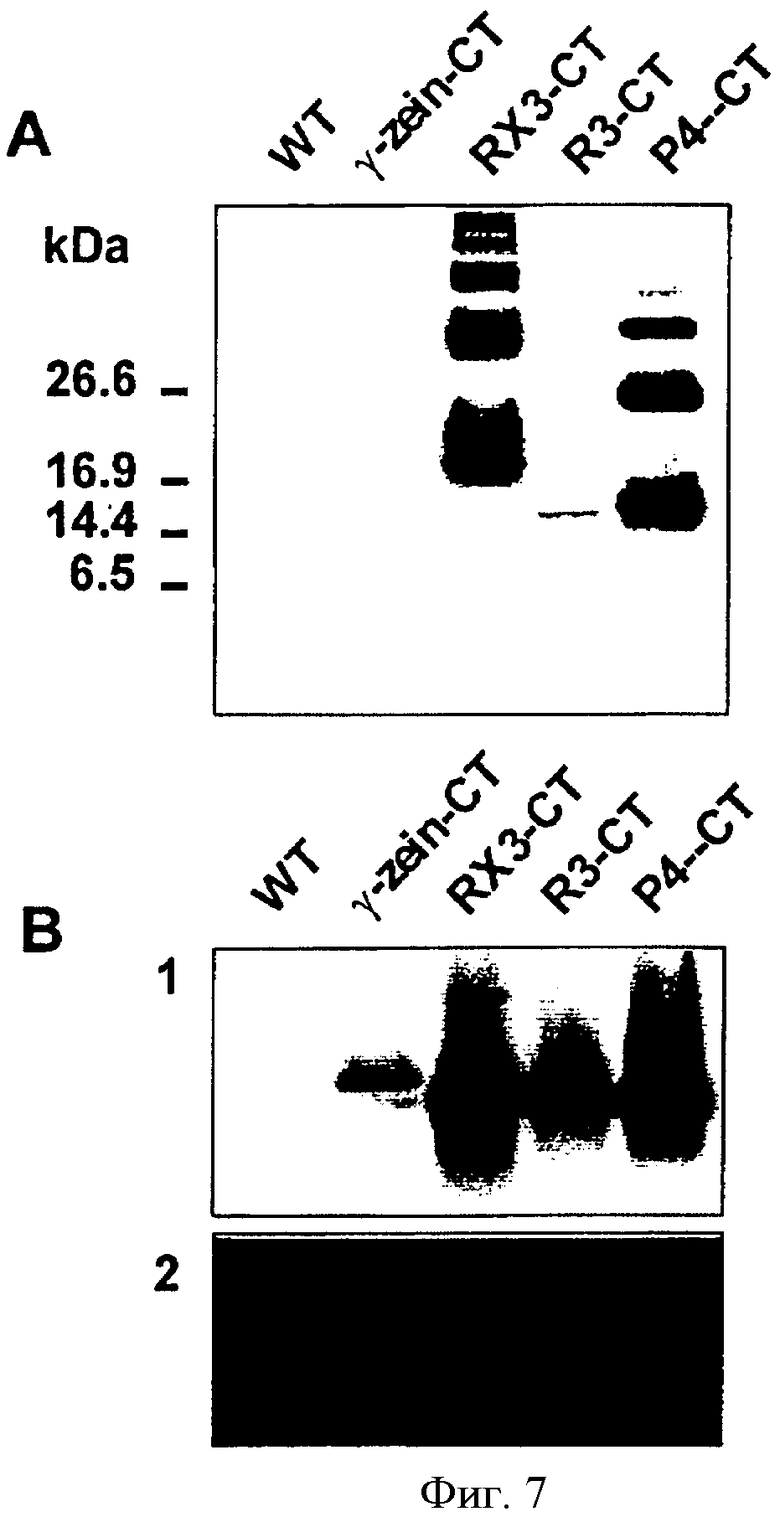
На Фиг.7 показаны: А. Сравнительный вестерн-блот анализ различных рекомбинантных слитых белков с использованием СТ антисыворотки. Экстракты растворимых белков были получены из дикорастущих растений (WT) и линий трансгенного табака (Т1), имеющих максимальную экспрессию слитого белка родственного химерного гена. 8 мкг растворимых белков помещали на 15% SDS-полиакриламидный гель и переносили в нитроцеллюлозу. В. Сравнительный нозерн-блот анализ различных транскриптов химерных генов. Общие РНК были выделены из анализируемых трансгенных линий с помощью иммуноблот-анализа (Фиг.7А), фракционированы с использованием денатурирующего формамидного гель-электрофореза (30 мкг на полосу) и капиллярно блоттированы на нейлоновой мембране. Блоты были гибридизированы с произвольно выбранной зонд-заправкой (129 оснований), полученной из кДНК кальцитонина.
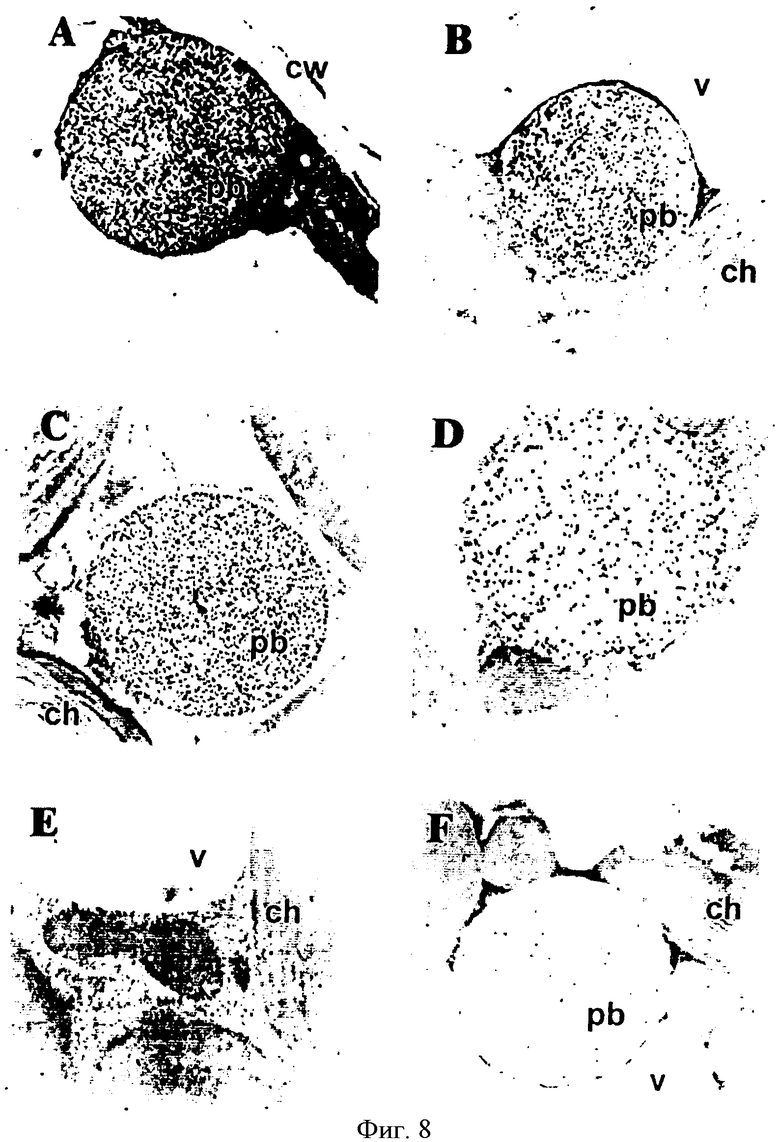
На Фиг.8 показана внутриклеточная локализация RX3-CT и Р4-СТ белков в трансгенных растениях табака: (А) Иммунолокализация RX3-CT белка в RX3-CT трансгенных линиях с использованием антисыворотки СТ (разбавление 1:100). (В) Иммунолокализация Р4-СТ белка в Р4-СТ трансгенных линиях с использованием антисыворотки СТ (разбавление 1:100). (С) Иммунолокализация RX3-CT белка в RX3-CT трансгенных линиях с использованием антисыворотки γ-зеина (разбавление 1:1500). (D) Иммунолокализация BiP белка в RX3-CT трансгенных линиях с использованием антисыворотки BiP (разбавление 1:250). (Е) Иммунолокализация в дикорастущих растениях с использованием антисыворотки γ-зеина (разбавление 1:1500). (F) Иммунолокализация в RX3-CT трансгенных растениях без первичных антител (разбавление 1:1500). Иммуноцитохимия на срезах табачных листьев была выполнена с использованием индикаторных первичных антител и протеинового А-коллоидального золота (15 нм). cw: клеточная стенка; ch: хлоропласт; pb: белковое тельце; v: вакуоль.
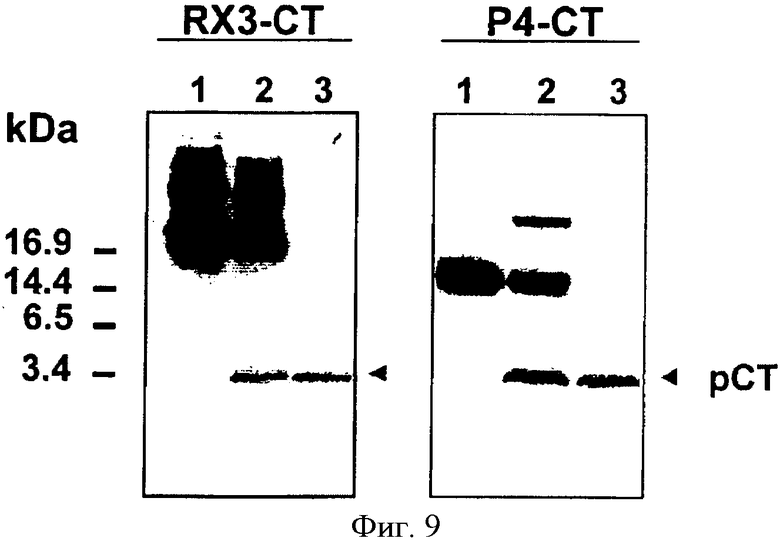
На Фиг.9 показаны результаты иммуноблот-анализа ЭК расщепления RX3-СТ и Р4-СТ слитого белка. 12 мкг каждого частично очищенного слитого белка инкубировали с 0,2 U ЭК в течение 24 часов при 20°С. Гидролизованные слитые белки фракционировали с использованием 18% Трис-трицин-полиакриламидного гель-электрофореза и переносили в нитроцеллюлозу. Полосы 1 - негидролизованные слитые белки (1 мкг); полосы 2 - гидролизованные продукты; полосы 3 - стандарт, синтетический лососевый СТ.
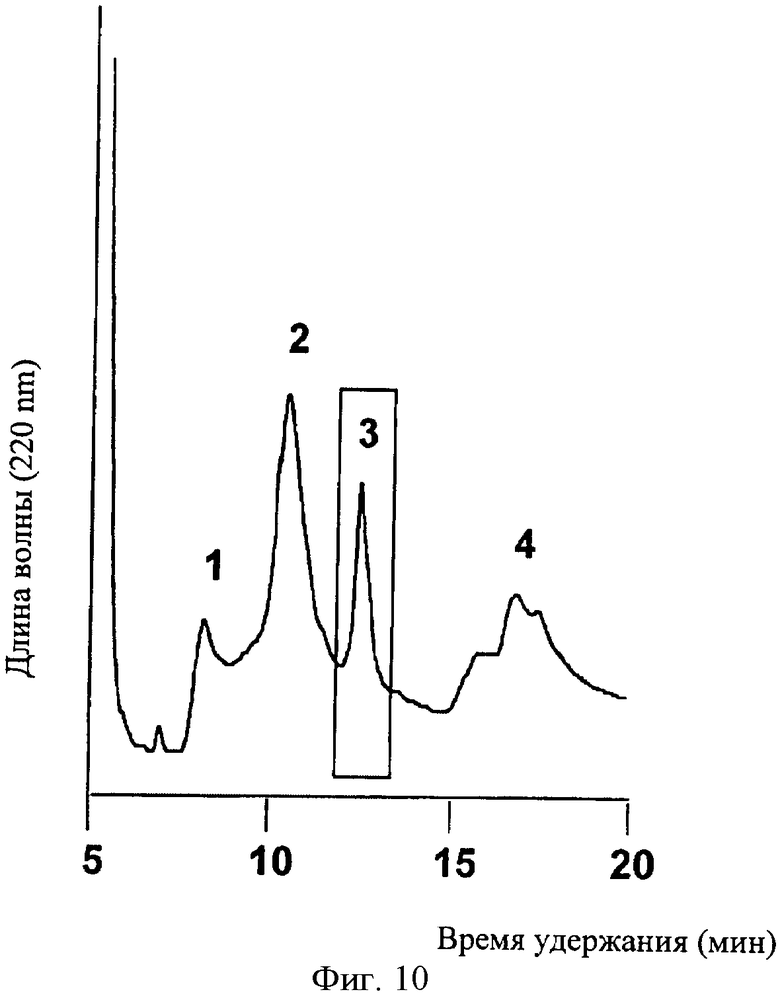
На Фиг.10 показаны результаты RP-HPLC фракционирования RX3-CT слитого белка, гидролизованного с помощью ЭК. рСТ, выделенный из RX3-CT слитого белка, был обнаружен в фракции 3 (Tr=13 мин) с помощью TOF-MALDI с использованием синтетического лососевого СТ в качестве стандарта.
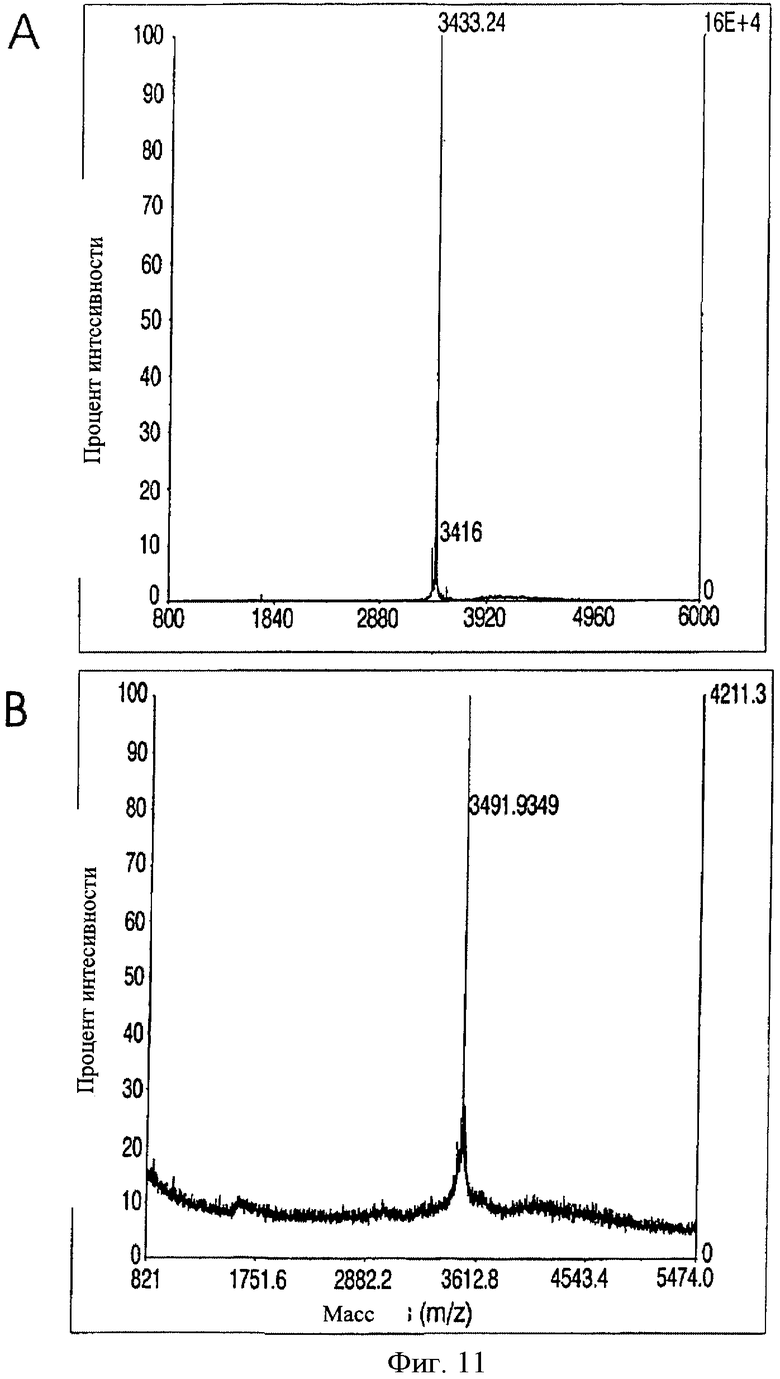
На Фиг.11 показаны результаты TOF-MALDI масс-спектроскопии (А) синтетического лососевого СТ (MW=3433.24) и (В) растительного СТ (MW=3491.93), элюированных при Tr=13 мин при RP-HPLC фракционировании.
Подробное описание изобретения
Первый аспект изобретения представляет последовательность нуклеиновой кислоты, в дальнейшем определяемой как последовательность нуклеиновой кислоты по изобретению, содержащую:
первую последовательность нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует белок γ-зеин, или ее фрагмент, содержащий нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в эндоплазматическом ретикулуме (ЭР);
вторую последовательность нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует аминокислотную последовательность, которая специфически расщепляется с помощью ферментативных или химических способов; и
третью последовательность нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует интересующий продукт;
где 3′ конец указанной первой последовательности нуклеиновой кислоты связан с 5′ концом указанной второй последовательности нуклеиновой кислоты и 3′ конец указанной второй последовательности нуклеиновой кислоты связан с 5′ концом указанной третьей последовательности нуклеиновой кислоты.
Первая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует белок γ-зеин, или ее фрагмент, содержащий нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в ЭР.
Используемый здесь термин "γ-зеин" относится к запасному белку маиса, который состоит из четырех характеристических доменов, упомянутых ранее в разделе "Предпосылки создания изобретения". Указанный термин включает нативные белки γ-зеина, а также его варианты и рекомбинантные белки γ-зеина, которые способны к направлению и сохранению белка в ЭР.
Может быть использована практически любая нуклеотидная последовательность, кодирующая белок γ-зеин, или ее фрагмент, содержащий нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в ЭР.
Соответственно, в предпочтительном варианте осуществления изобретения первая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую всю длину белка γ-зеина. В частном варианте осуществления изобретения нуклеотидная последовательность, кодирующая всю длину белка γ-зеина, показана на Фиг.1А и идентифицирована в SEQ ID NO:1.
В другом предпочтительном варианте осуществления изобретения первая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую фрагмент белка γ-зеина, указанный фрагмент содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в ЭР. В этом случае первая последовательность нуклеиновой кислоты может содержать:
- одну или более нуклеотидных последовательностей, кодирующих весь или часть повторяющегося домена белка γ-зеина;
- одну или более нуклеотидных последовательностей, кодирующих весь или часть ProX домена белка γ-зеина; или
- одну или более нуклеотидных последовательностей, кодирующих весь или часть повторяющегося домена белка γ-зеина, и одну или более нуклеотидных последовательностей, кодирующих весь или часть ProX домена белка γ-зеина.
В частном предпочтительном варианте осуществления изобретения указанная первая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую фрагмент белка γ-зеина, указанный фрагмент, содержащий нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в ER, выбирали из группы, состоящей из:
- нуклеотидной последовательности, показанной в SEQ ID NO:2 [нуклеотидная последовательность идентифицирована как RX3 (Фиг.1В)],
- нуклеотидной последовательности, показанной в SEQ ID NO:3 [нуклеотидная последовательность идентифицирована как R3 (Фиг.1В)],
- нуклеотидной последовательности, показанной в SEQ ID NO:4 [нуклеотидная последовательность идентифицирована как Р4 (Фиг.1С)], и
- нуклеотидной последовательности, показанной в SEQ ID NO:5 [нуклеотидная последовательность идентифицирована как X10 (Фиг.1С)].
Вторая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность, которая специфически расщепляется ферментативными или химическими способами. В частном варианте осуществления изобретения указанная вторая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует сайт расщепления протеазой, например, аминокислотный сайт, расщепляющийся с помощью протеазы, такой как энтерокиназа, Arg-C эндопротеаза, Glu-C эндопротеаза, Lys-C эндопротеаза, фактор Ха и аналогичных.
Альтернативно, вторая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует аминокислоту, которая специфически расщепляется химическим реагентом, таким как, например, цианбромид, который расщепляет остаток метионина, или любой другой подходящий химический реагент.
Вторая последовательность нуклеиновой кислоты может быть генерирована как результат объединения указанной первой последовательности нуклеиновой кислоты и указанной третьей последовательности нуклеиновой кислоты. В этом случае каждая последовательность содержит ряд нуклеотидов таким образом, что когда указанные первая и третья последовательности нуклеиновой кислоты становятся связанными, образуется функциональная нуклеотидная последовательность, которая кодирует аминокислотную последовательность, которая специфически расщепляется ферментативными или химическими способами, т.е. вторая последовательность нуклеиновой кислоты. В альтернативном варианте осуществления изобретения вторая последовательность нуклеиновой кислоты является чужеродной последовательностью, оперативно вставленной между указанными первой и третьей последовательностями нуклеиновой кислоты.
Третья последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует интересующий продукт. В принципе, любой интересующий продукт может быть экспрессирован с помощью системы, предлагаемой настоящим изобретением. В предпочтительном варианте осуществления изобретения интересующий продукт является белковым (т.е. белок или пептид) лекарственным средством, например, пептидный гормон, такой как кальцитонин, эритропоэтин, тромбопоэтин, гормон роста и подобные, интерферон, т.е., белок, продуцируемый в ответ на вирусную инфекцию, и как цитокин при иммунном ответе, и т.д. Предпочтительно, указанные интересующие терапевтические продукты являются эффективными для лечения человека или животных.
В частном варианте осуществления изобретения третья последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую кальцитонин (СТ), например, человеческий кальцитонин (hCT) или лососевый кальцитонин (sCT). В общем, в этом случае указанная третья последовательность нуклеиновой кислоты, предпочтительно, включает кодон для глицина при 3′ конце указанной последовательности нуклеиновой кислоты, кодирующей кальцитонин, воспроизводя таким образом глицин-удлиненный кальцитонин.
Согласно изобретению, 3′ конец указанной первой последовательности нуклеиновой кислоты связан с 5′ концом указанной второй последовательности нуклеиновой кислоты и 3′ конец указанной второй последовательности нуклеиновой кислоты связан с 5′ концом указанной третьей последовательности нуклеиновой кислоты, т.е. указанные первая, вторая и третья последовательности нуклеиновой кислоты находятся в рамке считывания.
Последовательность нуклеиновой кислоты по изобретению может быть получена с использованием обычных методик, известных специалисту в данной области. В основном, указанные методики включают связывание различных фрагментов последовательности нуклеиновой кислоты по изобретению с подходящим вектором. Обзор указанных традиционных методик можно найти, например, в "Molecular cloning, a Laboratory Manual", 2nd ed., by Sambrook et al., Cold Spring Harbor Laboratory Press, 1989. Конструирование некоторых векторов, содержащих нуклеиновую кислоту по изобретению, приведено в примерах и проиллюстрировано на Фиг.3 и 4. Как там показано, различные богатые пролином домены были выделены из γ-зеина для использования в качестве партнера слияния через расщепляемый протеазой сайт. Кодирующая область зрелого кальцитонина (32 ак) была слита с С-концом доменов γ-зеина и экспрессирована в трансгенные растения табака. Слитые белки аккумулировались в ЭР-производных белковых тельцах в листьях табака. После очистки слитые белки подвергли расщеплению энтерокиназой, обеспечивающему высвобождение кальцитонина, который может быть далее очищен из гидролизной смеси с помощью хроматографии с обращенными фазами.
В другом аспекте изобретение представляет слитый белок, далее называемый слитый белок по изобретению, содержащий (i) аминокислотную последовательность белка γ-зеин, или ее фрагмент, способный к направлению и сохранению белка в ЭР, (ii) аминокислотную последовательность, которая специфически расщепляется с помощью ферментативных или химических способов, и (iii) интересующий продукт; указанный слитый белок является продуктом экспрессии последовательности нуклеиновой кислоты по изобретению в системе растения-хозяина.
Слитый белок по изобретению аккумулируется в стабильных ЭР-производных БТ в системе растения-хозяина. Ферментативно или химически расщепляемый сайт, который находится при С-концах доменов γ-зеина, позволяет получать впоследствии интересующий продукт. Интересующий продукт затем может быть выделен и очищен общеизвестными методами. Следовательно, слитый белок по изобретению представляет новый и успешный подход к аккумулированию интересующего продукта.
В одном варианте осуществления изобретения слитый белок по изобретению содержит полный белок γ-зеина. Специфическая аминокислотная последовательность полного γ-зеина показана на Фиг.1А и идентифицирована как SEQ ID NO:6.
В другом варианте осуществления изобретения слитый белок по изобретению содержит фрагмент белка γ-зеина, указанный фрагмент содержит аминокислотную последовательность, способную к направлению и сохранению белка в ЭР. В частном варианте осуществления изобретения слитый белок по изобретению содержит фрагмент белка γ-зеина, выбранный из группы, состоящей из:
- аминокислотной последовательности, показанной в SEQ ID NO:7 [аминокислотная последовательность, соответствующая RX3 (Фиг.1В)],
- аминокислотной последовательности, показанной в SEQ ID NO:8 [аминокислотная последовательность, соответствующая R3 (Фиг.1В)],
- аминокислотной последовательности, показанной в SEQ ID NO:9 [аминокислотная последовательность, соответствующая Р4 (Фиг.1С)], и
- аминокислотной последовательности, показанной в SEQ ID NO:10 [аминокислотная последовательность, соответствующая X10 (Фиг.1С)].
Слитый белок по изобретению содержит аминокислотную последовательность, которая специфически расщепляется с помощью ферментативных или химических способов. В частном варианте осуществления изобретения указанный расщепляющийся сайт содержит сайт расщепления протеазой, например, аминокислотный сайт, расщепляющийся с помощью протеазы, такой как энтерокиназа, Arg-C эндопротеаза, Glu-C эндопротеаза, Lys-C эндопротеаза, фактор Ха и аналогичных, или аминокислотный сайт, расщепляющийся с помощью химического реагента, такого как, например, цианбромид, который расщепляет остаток метионина, или любой другой подходящий химический реагент.
Слитый белок по изобретению также содержит интересующий продукт, например, белковое (т.е. белок или пептид) лекарственное средство, например, пептидный гормон, интерферон, и аналогичные. Предпочтительно, указанный интересующий продукт является эффективным для лечения человека или животных. В частном варианте осуществления изобретения слитый белок по изобретению содержит кальцитонин (СТ), например, необязательно глицин-удлиненный человеческий кальцитонин (hCT) или лососевый кальцитонин (sCT).
В следующем аспекте изобретение относится к конструкции нуклеиновой кислоты, содержащей (i) последовательность нуклеиновой кислоты по изобретению, и (ii) регуляторную нуклеотидную последовательность, которая регулирует транскрипцию нуклеиновой кислоты по изобретению (i), причем указанная регуляторная последовательность (ii) является функциональной в растениях. Указанные последовательности нуклеиновой кислоты являются оперативно связанными.
Практически может быть использована любая функциональная регуляторная последовательность растения. В одном варианте осуществления изобретения указанная регуляторная последовательность (ii) является, предпочтительно, тканеспецифичной, т.е. она может регулировать транскрипцию нуклеиновой кислоты по изобретению в специфичных тканях, таких как семена, листья, клубеньки и т.д.
Регуляторная последовательность (ii) может содержать промотор, функциональный в растении. Фактически может быть использован любой промотор, функциональный в растении. В частном варианте осуществления изобретения указанная регуляторная последовательность (ii) содержит промотор 35SCaMV. В другом частном варианте осуществления изобретения указанная регуляторная последовательность (ii) содержит промотор "пататина", промотор запасного белка, промотор гена убихитина, регуляторную последовательность гена γ-зеина, или аналогичные.
Регуляторная последовательность (ii) может также содержать последовательность окончания транскрипции. Фактически может быть использована любая последовательность окончания транскрипции, функциональная в растении. В частном варианте осуществления изобретения указанная последовательность окончания транскрипции содержит терминатор 35SCaMV, терминатор гена октопинсинтазы (ocs), терминатор гена нопалинсинтазы (nos), терминатор гена γ-зеина, и аналогичные.
Регуляторная последовательность (ii) может также содержать трансляционный энхансер, функциональный в растении. Фактически может быть использован любой трансляционный энхансер, функциональный в растении, например, промотирующая последовательность для транскрипции вируса гравировки томата, и т.п.
Последовательность нуклеиновой кислоты по изобретению или конструкция, полученная с помощью этого изобретения, может быть вставлена в подходящий вектор. Поэтому в следующем аспекте изобретения представлен вектор, содержащий последовательность нуклеиновой кислоты по изобретению или конструкцию нуклеиновой кислоты, полученной с помощью настоящего изобретения. Подходящие векторы включают плазмиды, космиды и вирусные векторы. В одном варианте осуществления изобретения указанный вектор является подходящим для трансформирования растений. Выбор вектора может зависеть от клетки-хозяина, в которую он впоследствии вводится. В качестве примера, вектор, где вводится последовательность нуклеиновой кислоты по изобретению, может быть плазмидой, космидой или вирусным вектором, который, когда вводится в клетку-хозяин, интегрируется в геном указанной клетки-хозяина и реплицируется вдоль хромосомы (или хромосом), в которую он был интегрирован. Для получения указанного вектора могут использоваться традиционные способы (Sambrook et al., 1989).
В следующем аспекте изобретения представлена система растения-хозяина, указанная система растения-хозяина трансформируется нуклеиновой кислотой по изобретению или конструкцией или вектором, полученными по настоящему изобретению.
Используемый здесь термин "система растения-хозяина" включает растения, включая, но не ограничиваясь ими, односемядольные, двудольные, и, особенно, зерновые (например, кукуруза, рис, овес и т.д.), бобовые (например, соя и т.д.), крестоцветные (например, Arabidopsis thaliana, рапс и т.д.) или пасленовые (например, картофель, томаты, табак и т.д.). Система растения-хозяина также включает клетки растений. Клетки растений включают суспензионные культуры, завязи, меристематические области, ткани каллуса, листья, корни, побеги, гаметофиты, спорофиты, пыльцу, семена и микроспоры. Система растения-хозяина может быть на различных стадиях зрелости и может быть растущей в жидкой или твердой культуре, или в почве или в подходящей среде в горшке, в теплице или в поле. Экспрессия в системе растения-хозяина может быть кратковременной или постоянной. Система растения-хозяина также относится к любому клону такого растения, семени, самого или гибридного продукта, отводку, размножаемому половым путем или бесполым, и потомкам любого из них, таким как отростки или семена.
Трансформация систем растений-хозяев может быть осуществлена с использованием традиционных методов. Обзор генетического переноса в растениях можно посмотреть в руководстве, озаглавленном "Ingenieria genetica and transferencia génica", написанном Marta Izquierdo, Ed. Pyramide (1999), в частности, глава 9,"Transferencia génica a plantas", стр.283-316.
В следующем аспекте изобретения представлена трансгенная система растения-хозяина, сконструированного так, что содержит новый лабораторно определенный трансген, причем указанная трансгенная система растения-хозяина содержит интегрированную в его геном нуклеиновую кислоту по изобретению. Указанная трансгенная система растения-хозяина может быть получена с помощью общеизвестных методик, например, путем использования обычных методик антисмысловой мРНК и/или переэкспрессии (в смысле подавления) или других, например, путем использования бинарных векторов или других векторов, пригодных для ныне используемых методик трансформации различных растений. Примеры трансгенных систем растения-хозяина, представляемых настоящим изобретением, включают односемядольные и двудольные растения и, особенно, зерновые, бобовые, крестоцветные, пасленовые и т.д.
Последовательность нуклеиновой кислоты по изобретению используется для получения интересующего продукта в системе растения-хозяина. Поэтому в следующем аспекте изобретения представлен способ получения интересующего продукта в системе растения-хозяина, который включает выращивание трансформированной или трансгенной системы растения-хозяина, представленной настоящим изобретением, в условиях, которые допускают получение и экспрессию указанного интересующего продукта в форме слитого белка. Как указывалось выше, указанный слитый белок аккумулируется в стабильных ЭР-производных БТ в указанной системе растения-хозяина. Ферментативно или химически расщепляемый сайт, который находится на С-конце домена γ-зеина, позволяет впоследствии получать интересующий продукт. Интересующий продукт затем может быть выделен и очищен с помощью традиционных методов. Следовательно, метод, предлагаемый настоящим изобретением, далее включает, если желательно, выделение и очистку указанного слитого белка, и, необязательно, высвобождение указанного интересующего продукта из указанного слитого белка. Слитый белок расщепляется по сайту расщепления с помощью фермента или химического реагента, по обстановке.
В следующем аспекте изобретение представляет способ продуцирования кальцитонина в системе растения-хозяина, включающий:
а) трансформирование системы растения-хозяина вектором экспрессии или конструкцией нуклеиновой кислоты, содержащей
(i) регуляторную последовательность и
(ii) последовательность нуклеиновой кислоты состоящей из:
первой последовательности нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует белок γ-зеин, или ее фрагмент, содержащий нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в эндоплазматическом ретикулуме (ЭР);
второй последовательности нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует аминокислотную последовательность, которая специфически расщепляется с помощью ферментативных или химических способов; и
третьей последовательности нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует кальцитонин;
где 3′ конец указанной первой последовательности нуклеиновой кислоты связан с 5′ концом указанной второй последовательности нуклеиновой кислоты и 3′ конец указанной второй последовательности нуклеиновой кислоты связан с 5′ концом указанной третьей последовательности нуклеиновой кислоты; и
где регуляторную последовательность регулируют транскрипцией упомянутой последовательности нуклеиновой кислоты
b) генерирование полных растений из указанных систем растений-хозяев, трансформированных указанным вектором экспрессии или конструкцией нуклеиновой кислоты;
c) выращивание таких трансформированных растений при условиях, которые обеспечивают получение и экспрессию кальцитонина в форме слитого белка; и, если желательно,
d) выделение, очистку указанного слитого белка и обработку указанного слитого белка для того, чтобы высвободить кальцитонин.
Изобретение представляет поэтому основанную на слитом белке систему для аккумулирования рекомбинантных интересующих продуктов в ЭР-производных БТ в системе растения-хозяина. Изобретение далее иллюстрируется нелимитирующими примерами.
ПРИМЕР 1
Получение кальцитонина в растениях табака
Успешный пример получения СТ в растениях табака описан ниже. Различные богатые пролином домены были сконструированы из γ-зеина для использования в качестве партнера слияния через расщепляемый протеазой сайт. Кодирующая область (32 ак) зрелого СТ была слита с С-концами доменов γ-зеина и экспрессирована в трансгенные растения табака. Расщепляемый протеазой сайт был вставлен при С-концах доменов γ-зеина для возвращения впоследствии чистого кальцитонина. Этот подход предлагает высокую аккумуляцию слитых белков внутри ЭР и образование ЭР-производных БТ в растениях табака. Слитые белки сильно аккумулируются в ЭР-производных БТ в листьях табака. Уровень экспрессии указанных слитых белков достигал, в некоторых случаях, 12,44% от общих растворимых белков. После только двух стадий очистки слитые белки подвергали расщеплению энтерокиназой, позволяющему высвобождение кальцитонина. Конечный кальцитонин, аккумулированный в растениях табака, был идентифицирован с помощью масс-спектроскопии. Также осуществляли очистку слитых белков, протеазный ферментативный гидролиз и полную характеристику высвобожденного растительного кальцитонина (рСТ).
I. ЭКСПЕРИМЕНТАЛЬНЫЕ МЕТОДИКИ
Конструирование химерных генов и векторов
Ген γ-зеина дикого типа и четырех производных от γ-зеина последовательностей, названных RX3, R3, Р4 и X10, кодирующих различные домены γ-зеина (Фиг.1А, 1В и 1С), были слиты с синтетическим геном СТ, содержащим гидролизуемый энтерокиназой сайт (Фиг.2), и вставлены в векторы трансформации растений, как описано ниже и на Фиг.3.
Последовательности кДНК γ-зеина, RX3 и R3 генерировали с помощью PCR с использованием pKSG2 (Torrent et al., 1994) в качестве шаблона. X10 кДНК амплифицировали из pDR20, плазмиду получали из pKSG2 после делеции последовательности, соответствующей повторяющемуся домену. Праймеры, используемые для различных PCR, представляли собой: для последовательности кДНК γ-зеина:
T1: 5′TCATGAGGGTGTTGCTCGTTGCCCTC3′, и
Т4: 5′CCATGGCGTGGGGGACACCGCCGGC3′,
для последовательностей кДНК RX3 и X10:
Т1 и
Т2: 5′CCATGGTCTGGCACGGGCTTGGATGCGG 3′, и
для последовательности кДНК R3:
Т1 и
Т3: 5′CCATGGTCCGGGGCGGTTGAGTAGGGTA3′.
Продукты PCR были субклонированы в вектор pUC 18 (SureClone Ligation Kit, Pharmacia) и результирующие плазмиды были названы pUCZein, pUCRX3, pUCR3 и pUCX10. Вектор pUCP4, который содержит производную от γ-зеина последовательность Р4, (Фиг.1С), был получен во время скрининга клонов, производных pUCRX3. Фрагменты кДНК γ-зеина, RX3, R3, Р4 и Х10, содержащие "липкие" концы BspHI и NcoI, были вставлены в вектор pCKGFPS65C (Reichel et al., 1996), предварительно гидролизованный с NcoI. Этот вектор был выбран потому, что он содержит регуляторную последовательность для экспрессии в растениях и GFP кодирующую последовательность, которая использовалась бы для изучения параллельного нацеливания белков, производных γ-зеина, в трансгенных растениях. Генерированные векторы pCZeinGFP, pCRX3GFP, pCR3GFP, pCP4GFP и pCXIOGFP содержали следующие регуляторные последовательности для экспрессии в растительных системах: i) усовершенствованный 35S промотор, извлеченный из вируса мозаики цветной капусты (CaMVp35S), ii) трансляционный энхансер из вируса гравировки томатов (TL) и iii) последовательность окончания транскрипции из CaMV35S (pA35S). Химерные конструкции производное γ-зеина/СТ были генерированы путем замещения GFP кодирующей последовательности синтетическим геном СТ, как описано ниже (Фиг.3).
Синтетический ген, кодирующий 32 аминокислоты активного лососевого СТ (Фиг.2), генерировали из двух 122 основных комплементарных олигонуклеотидов. Олигонуклеотиды предназначались для использования предпочтительных кодонов растений, для того чтобы достичь высокой экспрессии в растениях. 5′ фосфорилированные олигонуклеотиды, синтезированные с использованием ДНК синтезатора Applied Biosystems 394, имели следующие последовательности:
CalI:
5′CATGGACGACGACGACAAGTGCTCCAACCTCTCTACCTGCGTTCTTGG
TAAGCTCTCTCAGGAGCTTCACAAGCTCCAGACTTACCCTAGAACCAACACTG
GTTCCGGTACCCCTGGTTGAT 3′,
CalII:
5′CTAGATCAACCAGGGGTACCGGAACCAGTGTTGGTTCTAGGGTAAGTC
TGGAGCTTGTGAAGCTCCTGAGAGAGCTTACCAAGAACGCAGGTAGAGAGGTT
GAGCACTTGTCGTCGTCGTC3′.
После очистки на 12% полиакриламидном геле 60 пмоль каждого нуклеотида использовали для формирования двухниточной молекулы. Смесь гибридизации грели при 95°С в течение 5 мин, выдерживали при 70°С в течение 1 часа и охлаждали до комнатной температуры. Фрагмент синтетической кДНК содержал NcoI и XbaI "липкие" концы при 5′ и 3′ концах, соответственно. Синтетическая кДНК СТ включала 5′ линкерную последовательность, соответствующую сайту специфического расщепления энтерокиназой ((Asp)4-Lys), и была удлинена при 3′ конце, чтобы получить единичный глицин для дальнейшего амидирования СТ пептида. NcoI/XbaI СТ кДНК субклонировали в pUC 18 вектор и затем вставили в NcoI и BamHI сайты рестрикции векторов pCZeinGFP, pCRX3GFP, pCR3GFP, pCP4GFP и pCX10GFP, содержащие кодирующие производные от γ-зеина последовательности, и делетировали из GFP кодирующей последовательности. Результирующие последовательности были названы pCZeinCT, pCRX3CT, pCR3CT, рСР4СТ и рСХ10СТ (Фиг.3). Эффективные векторы трансформации растений pBZeinCT, pBRX3CT, pBR3CT, pBP4CT и рВХ10СТ (Фиг.4) были, в конце концов, получены путем вставки различных HindIII/HindIII кассет экспрессии в бинарный вектор pBin19 (Bevan, 1984).
Трансформация устойчивых растений табака
Бинарные векторы переносили в LBA 4404 штаммы Agrobacterium tumefaciens. Диски листьев табака (Nicotiana tobaccum, W38) трансформировали согласно методу Draper et al. (1988). Регенерированные растения отбирали на среде, содержащей 200 мг/л канамицина, и переносили в теплицу. Трансгенные растения табака, имеющие наивысшие уровни трансгенного продукта, культивировали для получения Т1 генерации. Развивающиеся листья (примерно 12 см длиной) собирали, немедленно замораживали в жидком азоте и хранили при -80°С для дальнейших экспериментов.
Экстракция и вестерн-блот анализ рекомбинантных белков
Листья табака измельчали в жидком азоте и гомогенизировали с использованием 4 мл экстракционного буфера (50 мМ Tris-HCl рН 8, 200 мМ дитиотреитола (DTT) и ингибиторов протеазы (10 мкМ апротинина, 1 мкМ пепстатина, 100 мкМ лейпептина, 100 мкМ фенилметилсульфонилфторида и 100 мкМ Е64 [(N-(N-(L-3-транс-карбоксиоксиран-2-карбонил)-Lлейцил)агмантина] на грамм свежего листового материала. Гомогенаты перемешивали в течение 30 мин при 4°С и затем дважды центрифугировали (15000 об/мин, 30 мин, 4°С) для удаления нерастворимого материала. Общее количество белков измеряли с использованием анализа белков по Брэдфорду (Bio-Rad). Белки разделяли на 15% SDS полиакриламидном геле и переносили на нитроцеллюлозные мембраны (0,22 мкМ) с использованием полусухого аппарата. Мембраны инкубировали с антисывороткой γ-зеина (разбавление 1/7000) (Ludevid et al., 1985) или антисывороткой против KLH-кальцитонина (СТ-антисыворотка) (разбавление 1/1000) и затем инкубировали с конъюгированными антителами пероксидазы хрена (разбавление 1/10000). Иммунореактивные связи определяли с помощью усовершенствованной химиолюминесценции (ECL вестерн-блоттинг система, Amersham). Антитела кальцитонина выращивали в кроликах путем инокуляции синтетического лососевого кальцитонина, связанного с KLH. После четырех инокуляций антигена (200 г каждая) сыворотку собирали, делили на аликвоты и хранили при -80°С. Титрование сыворотки осуществляли с помощью точечного иммуноблот-анализа с применением синтетического кальцитонина и ELISA анализа с использованием BSA-кальцитонина в качестве антигена.
Нозерн-блот анализ
Общая РНК была выделена из листьев дикорастущего и трансгенного табака согласно Logemann et al., 1987. РНК фракционировали с использованием денатурированного формамид-агарозного гель-электрофореза (30 мкг на полосу) и капиллярно блоттировали на нейлоновые мембраны (Hybond N, Amersham Pharmacia Biotech). Блоты РНК гибридизировали с 129 основной ДНК пробой, полученной из СТ кДНК, и метили (α-32Р) dCTP, используя набор для мечения со случайно заправленной ДНК (Roche). Гибридизацию осуществляли в течение ночи при 42°С и фильтры мыли три раза в течение 15 мин в 3Х SSC и 0,5% SDS(WN) при 65°С. Блоты определяли с помощью фосфор-визуализирующего сканера (Fluor-S™ Multilmager, BIO-RAD).
ELISA анализ (метод твердофазного иммуноферментного анализа)
ELISA анализы были проведены для количественного анализа растительного кальцитонина (рСТ) в экстрактах растворимых листовых белков и частично очищенных γ-зеин-СТ слитых белках. Титрационные микропланшеты (MaxiSorp, Nalgene Nunc International) заряжали растворимыми белками (100 мкл), разбавленными фосфатно-буферным солевым раствором рН 7,5 (PBS) и инкубировали в течение ночи при 4°С. После трехкратной промывки лунок неспецифические связывающие сайты блокировали 3% бычьим сывороточным альбумином (BSA) в PBS-T (PBS, содержащий 0,1% Tween 20) в течение одного часа при комнатной температуре. Планшеты инкубировали с антисывороткой СТ (разбавление 1/1000) в течение двух часов и после четырех промывок PBS-T инкубировали с пероксидаза-конъюгированными вторичными антителами (разбавление 1/8000) (Sigma) в течение двух часов. Первичные и вторичные антитела разбавляли PBS-T, содержащим 1% BSA. После интенсивной промывки PBS-T осуществляли ферментативную реакцию при 37°С со 100 мкл основного буфера (100 мМ ацетата натрия рН 6, 0,01 мг/мл ТМВ (3,3′,5,5′-тетраметилбензилиден) и 0,01% перекиси водорода). Реакцию останавливали через 10 мин 2Н серной кислотой и измеряли оптическую плотность при 450 нм, используя спектрофотометр Multiskan EX (Labsystems). Концентрацию антигена в растительных экстрактах экстраполировали из стандартных кривых, полученных с помощью использования кальцитонин-BSA и СТ антисыворотки (разбавление 1/1000).
Электронная микроскопия
Листья дикорастущих и трансгенных растений закрепляли путем вакуумной инфильтрации с 1% глутарового альдегида и 2,5% параформа в 20 мМ фосфатного буфера с рН 7,4 в течение одного часа при комнатной температуре. После последовательной промывки 20 мМ фосфатного буфера и 200 мМ хлорида аммония образцы обезвоживали в этаноле и вводили в Lowicryl К4М полимер. Иммунохимию выполняли по существу как описано Moore et al., 1991. Ультратонкие срезы инкубировали с антисывороткой против KLH-кальцитонин (1/500), αBiP (1/500) и γ-зеина (1/1500). Для определения антител использовали протеиновое А-коллоидальное золото (частицы золота 15 нм). В качестве контроля осуществляли параллельное инкубирование нетрансгенных растительных образцов, используя идентичные разбавления первичных антител, и трансгенных образцов без первичных антител. Срезы окрашивали уранилацетатом и цитратом свинца и исследовали с помощью 301 модели электронного микроскопа (Phillips, Eindhoven, The Netherlans).
Очистка и расщепление энтерокиназой RX3-CT и Р4-СТ слитых белков
Растворимые экстракты RX3-CT и Р4-СТ получали из листьев трансгенного табака (Т1) в экстракционном буфере, как описано выше. Твердый (NH4)2SO4 постепенно добавляли при 0°С к растворимым экстрактам RX3-CT и Р4-СТ до 45% и 60% насыщения, соответственно. Образцы перемешивали в течение 30 мин при 0°С и затем центрифугировали при 15000 об/мин в течение 45 мин при 4°С. Осажденные белки ресуспендировали в 20 мМ Tris-HCl рН 8,6 и обессоливали на PD 10 колонке (Sephadex G-25 М, Amersham Pharmacia). Обессоленные белковые экстракты фракционировали с помощью быстродействующей жидкостной хроматографии (FPLC), с использованием анионообменной колонки (HiTrap Q sepharose, Amersham Pharmacia), уравновешенной с 20 мМ Tris-HCl рН 8,6, 100 мМ DTT. Элюирование белка осуществляли с линейным солевым градиентом от 0 до 200 мМ NaCl в 20 мМ Tris-HCl рН 8,6, 100 мМ DTT. Наличие RX3-CT и Р4-СТ в элюированных фракциях оценивали с помощью электрофореза в 15% SDS полиакриламидном геле и иммуноблот-анализа с использованием антисыворотки СТ. Положительные фракции обессоливали и концентрировали на 5 K NMWL центробежных фильтрах (BIOMAX, Millipore). Количественное определение RX3-CT и Р4-СТ слитых белков выполняли методом ELISA.
Для ЭК ферментативного гидролиза 15 мкг частично очищенных слитых белков инкубировали с 0,2 U EK (ЕК Max, Invitrogen) в 30 мкл гидролизного буфера (50 мМ Tris-HCl рН 8,1, 1 мМ NaCl, 0,1% Tween-20) в течение 24 часов при 20°С. Буфер ЭК гидролиза дополняли 100 мМ DTT. Наличие восстанавливающего средства позволяет оптимизировать расщепление энтерокиназой. Продукты гидролиза анализировали с помощью 18% Трис-трицин-полиакриамидного гель-электрофореза, а высвобожденный рСТ определяли с помощью иммуноблот-анализа. Синтетический лососевый СТ использовали в качестве положительного контроля.
Очистка и анализ высвобожденного рСТ
Растительный кальцитонин (рСТ), высвобожденный из слитых белков с помощью ЭК гидролиза, был очищен с помощью RP-HPLC. Гидролизную смесь помещали на аналитическую колонку RP-C18 (250×4 мм, размер частиц 10 мкМ, размер пор 120 Å) и колонку элюировали с использованием градиента, изменяющегося от 25 до 60% ацетонитрила с 0,036% TFA за 20 мин при расходе 1 мл/мл. Собранные фракции концентрировали путем лиофилизации и хранили при -20°С для определения характеристик рСТ. В отдельном эксперименте стандартный лососевый СТ элюировали при тех же хроматографических условиях. Для определения характеристик рСТ использовали TOF-MALDI масс-спектрометрию. Аликвоты фракций RP-HPLC смешивали с равным объемом матричного раствора (10 мг/мл α-циано-4-гидроксикоричной кислоты и 0,1% TFA) и 1 мкл помещали на держатель и анализировали с помощью масс-спектрометра Voyager-DE-RP (Applied Biosystems). Стандартный лососевый СТ использовали в экспериментах TOF-MALDI масс-спектрометрии в качестве контроля. С-концевой анализ рСТ выполняли путем инкубирования очищенного пептида (20 пмоль/мкл) в течение 60 мин при 37°С с карбоксипептидазой Y (0,1 U/мкл) и анализировали продукты гидролиза с помощью TOF-MALDI масс-спектрометрии.
II. РЕЗУЛЬТАТЫ
Конструирование химерных генов различных производных γ-зеин-СТ
Экспрессия и успешная сборка богатых пролином доменов γ-зеина в ЭР-производных белковых тельцах в листьях растений (Geli et al., 1994) представляют полноценное средство для аккумулирования терапевтических белков в ЭР растительных тканей. Ген γ-зеина делетировали для создания различных обогащенных пролином укороченных белков, используемых в качестве связывающего партнера для продуцирования СТ в растениях табака. Химерные гены, содержащие домены γ-зеина и синтетический ген СТ, связывали с помощью линкера, соответствующего протеазой расщепляемому сайту. Синтетический ген, кодирующий 32 аминокислотный активный лососевый кальцитонин, генерировали из двух комплементарных олигонуклеотидов (122 основания), предназначенных для использования предпочтительных кодонов растения, для того чтобы достигнуть высокой экспрессии рекомбинантных белков в растениях. Синтетическая кДНК СТ (Фиг.2) включала при 5′ конце линкерную последовательность, соответствующую сайту расщепления энтерокиназой ((Asp)4-Lys), и при 3′ конце дополнительный кодон для продуцирования глицина. Этот глицин является необходимым субстратом для амидирующего фермента (РАМ), чтобы генерировать С-концевой пролинамид, необходимый для биологической активности СТ. кДНК кальцитонина была слита с последовательностями, кодирующими домены γ-зеина, С-концевым слиянием. Для оптимальной экспрессии химерных генов производных γ-зеин-СТ в системах растений векторы трансформации растений содержали следующие регуляторные последовательности: i) конститутивный усовершенствованный 35S промотор и 35S терминатор из вируса мозаики цветной капусты и ii) трансляционный энхансер из вируса гравировки томатов (TL). Различные генерированные слитые белки представлены на Фиг.5. Слитый белок γ-зеин-СТ содержит целый γ-зеин, слитый с СТ. Слитые белки RX3-CT, R3-CT, Р4-СТ и Х10-СТ содержат домены производных γ-зеина, связанные с СТ тем же путем, как и целый γ-зеин. Эти слитые белки существенно различаются по наличию или отсутствию повторяющегося и proX доменов.
Получение слитых белков в растениях табака
Все гены слияния использовались для трансформации устойчивых табачных растений с помощью Agrobacterium tumefaciens. По меньшей мере, двадцать независимых канамицин-устойчивых растений (То) генерировали для каждого гена слияния. Скрининг трансгенных растений был выполнен с помощью вестерн-блот анализа экстрактов растворимых белков с использованием поликлональной антисыворотки γ-зеина. Конфигурации иммуноблотов трансгенных линий, отображающие каждый ген слияния, показаны на Фиг.6. Как показали наблюдения, рекомбинантные слитые белки присутствовали во всех трансгенных линиях, за исключением гена слияния Х10-СТ, где не определялись следы слитых белков. Этот маленький слитый белок (80 аминокислот) вероятно нестабилен в растениях табака. Две иммуно-меченых полосы были обнаружены в трансгенных линиях R3-CT, одна с нетипично высокой кажущейся молекулярной массой. Этот слитый белок, вероятно, был подвергнут посттрансдукционным модификациям, таким как гликозилирование. Кроме того, было показано, что богатый пролином повторяющийся домен γ-зеина способен гликозилироваться, когда экспрессирован в растениях Arabidopsis (Alvarez et al., 1998). Уровень экспрессии белка совершенно изменялся для различных линий одного и того же гена слияния, за исключением гена слияния RX3-СТ, который показал высокий уровень экспрессии рекомбинантного белка во всех трансгенных линиях. Дополнительный скрининг иммуноблотов был выполнен с использованием антисыворотки, специально выращенной против пептида sCT (Фиг.7А). Как показали наблюдения, RX3-CT и Р4-СТ хорошо распознавались с помощью антисыворотки sCT, указывая, что эти слияния обеспечивают лучшее аккумулирование СТ пептида в растениях табака. Следует отметить, что изображения иммуноблотов RX3-CT и Р4-СТ показали различные меченые полосы, с главной полосой, соответствующей правильной кажущейся молекулярной массе родственного рекомбинантного белка. По одной из гипотез меченые полосы высокой молекулярной массы были результатом процесса олигомеризации доменов γ-зеина, которые образовывались во время аккумулирования слитых белков в тканях растений. Для того чтобы контролировать уровни экспрессии генов слияния относительно уровней белка, был выполнен сравнительный нозерн-блот анализ (Фиг.7В) с использованием трансгенных линий, анализированных с помощью иммуноблот-анализа (Фиг.7А). Как было показано, транскрипты RX3-CT и Р4-СТ были более обильными, демонстрируя устойчивое аккумулирование этих транскриптов. Неожиданно транскрипты R3-CT были более обильны по сравнению с низким уровнем слитого белка R3-CT, определяемым с помощью иммуноблот-анализа. Вероятно, посттрансляционная модификация отменяет правильную самосборку слитого белка и, следовательно, его стабильность в ЭР.
Максимальные уровни экспрессии RX3-CT и Р4-СТ белков, измеренные методом ELISA в экстрактах листовых белков из Т1 растений, были соответственно 12,44% и 10,65% от общего количества растворимых белков. Что касается этих результатов, трансгенные линии RX3-CT и Р4-СТ были выбраны для дальнейших экспериментов, способствующих получению растительного кальцитонина (рСТ).
Внутриклеточная локализация слитых белков RX3-CT и Р4-СТ
Экспрессия γ-зеина и двух мутантов делеции γ-зеина в растениях Arabidopsis показали, что эти белки локализовались внутри ЭР мезофильных клеток, образующих ЭР-производные БТ (Geli et al., 1994). Однако было неочевидно, что кальцитонин, слитый с производными γ-зеина, распределяется в похожих органеллах, ЭР-БТ. Чтобы исследовать внутриклеточную локализацию в листьях табака слитых белков γ-зеина, содержащих кальцитонин, изобретатели использовали иммуноэлектронную микроскопию (Фиг.8). Ультратонкие срезы листьев трансгенного табака, экспрессированные RX3-CT и Р4-СТ белками, инкубировали с антителом СТ и протеиновым А-золотом. Сильно меченые БТ-подобные органеллы наблюдали в мезофильных клетках табака, экспрессированных RX3-CT и Р4-СТ (Фиг.8А и В, соответственно). Были определены несколько везикул на клетку и их размер был совсем неоднородным. Поскольку слитые белки содержали белок кальцитонин и фрагменты γ-зеина, ультратонкие срезы также инкубировали с антителом γ-зеина. Как ожидалось, БТ были помечены антителом γ-зеина, подтверждая, что слитые белки аккумулировались внутри этих органелл (Фиг.8С). Чтобы продемонстрировать, что БТ образовывались из ЭР, срезы инкубировали с антителами против ЭР резидентного белка BiP (Фиг.8 D). Сопутствующее распространение СТ-слитых белков и BiP в этих органеллах показало, что RX3-CT и Р4-СТ аккумулировали внутри ЭР просвета с последующим образованием независимых ЭР-производных везикул. Поскольку изобретатели не смогли определить БТ-подобные органеллы в ультратонких срезах нетрансгенных растений (Фиг.8Е), контрольные эксперименты были выполнены без первичного антитела в трансгенных растениях (Фиг.8F). Как ожидалось, неспецифическая метка была определена в контрольных экспериментах.
Очистка слитых белков и высвобождение рСТ
RX3-CT и Р4-СТ слитые белки эффективно экстрагировали из листьев трансгенного табака (Т1), используя экстракционный буфер, включающий восстанавливающий агент, такой как DDT (200 мМ). Около 85 мкг RX3-CT и 75 мкг Р4-СТ получали из грамма свежего материала. RX3-CT и Р4-СТ белки концентрировали осаждением с 45% и 60% сульфатом аммония, соответственно. Обессоленные экстракты белков фракционировали методом FPLC, используя анионообменную хроматографию, и выделенные слитые белки анализировали методом ELISA. RX3-CT белок составлял около 80% от общих очищенных белков, в то время как Р4-СТ составлял только около 50% от общих очищенных белков. Это различие можно объяснить с помощью того факта, что больше белков осаждается с 60% сульфата аммония, чем с 45%, и что, следовательно, осажденные Р4-СТ белки содержали много больше сопутствующих белков. Частично очищенные слитые белки RX3-CT и Р4-СТ гидролизовали с помощью ЭК и высвобождение рСТ контролировали с помощью Трис-трицин-полиакриламидного гель-электрофореза и иммунологического анализа. Как показано на Фиг.9, единственная меченая полоса, соответствующая кальцитонину, была генерирована при расщеплении обоих RX3-СТ и Р4-СТ белков. Малые количества слитых белков RX3-CT и Р4-СТ оставались негидролизованными, вероятно, вследствие недоступности для фермента некоторых сайтов расщепления.
Очистка и определение характеристик рСТ
Растительный кальцитонин (рСТ) выделяли фракционированием смесей ферментативного ЭК гидролиза на аналитической колонке С18 RP-HPLC (Фиг.10) и анализировали элюируемые фракции с помощью TOF-MALDI масс-спектроскопии, используя синтетический sCT в качестве стандарта (М.м. 3433.24, Фиг.11.А). рСТ кальцитонин элюировали 13 мин (синтетический sCT Tr=14 min) и получили один спектр с массой 3491,93 Да с помощью TOF-MALDI масс-спектроскопии, что согласуется с теоретической молекулярной массой полученного кальцитонина, удлиненного глицином при С-конце (Фиг.11В). Масс-спектрометрический анализ рСТ, подвергнутого ферментативному гидролизу с карбоксипептидазой Y, подтвердил целостность С-концевого глицина, который является необходимым для получения С-концевого пролинамида.
III. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Предложена удачная система на основе слитых белков для аккумулирования лососевого кальцитонина в листьях табака. Были найдены два слитых белка RX3-Cal и Р4-Cal для сильного аккумулирования в ЭР-производных БТ листьев табака. Эти слитые белки содержат СТ пептид и богатые пролином домены γ-зеина, которые включают i) повторяющийся домен, содержащий восемь единиц гексапептида PPPVHL (только одна единица в Р4-Cal слитом белке) и ii) proX домен, где пролиновые остатки заменены другими аминокислотами. Богатые пролином домены γ-зеина необходимы для правильного сохранения и сборки γ-зеина внутри ЭР растений Arabidopsis (Geli et al., 1994). Складывание и стабилизация полипептидных цепей γ-зеина в ЭР объясняется способностью повторяющегося и proX доменов осуществлять самосборку и промотировать образование олигомеров. Особая конформация, принимаемая с помощью этих высокогидрофобных доменов, вероятно, обусловлена богатыми пролином последовательностями, которые способны образовывать амфипатическую вторичную структуру. В результате их характерной конформации богатые пролином домены будут индуцировать механизмы агрегации, включающие протеин-протеиновые взаимодействия и дисульфидные поперечные связи, способствующие сохранению ЭР и образованию ЭР-производных БТ. Этот пример показывает, что экспрессированные методом N-концевого слияния богатые пролином домены γ-зеина сохраняют полностью способность к самосборке и промотированию множества явлений, приводящих к сохранению и аккумулированию ЭР-производных БТ. Было также найдено, что лососевый СТ, включенный в слитый белок, сильно аккумулируется в БТ. Высокий уровень экспрессии СТ в трансгенных растениях табака можно объяснить способностью богатых пролином доменов складывать и стабилизировать слитый белок. Депонирование слитого белка в БТ безусловно вносит вклад в обогащение тканей растений кальцитонином, путем перемещения его из гидролитического внутриклеточного окружения. Так как малые белки нестабильны в биологических системах, теперь решили использовать слитые белки для получения кальцитонина в гетерологичных системах, например в Е. coli (Ray et al., 1993; Yabuta et al., 1995; Hong et al., 2000), в Staphylococcus carnosus (Dilsen et al., 2000) и в молоке трансгенных кроликов (Mckee et al., 1998). В этом последнем случае целью слияния СТ с человеческим альфа-лактальбумином была маскировка активности кальцитонина, чтобы избежать возможной интерференции с нормальным развитием животного.
Изобретатели преуспели в быстром получении удлиненного глицином sCT из растений табака:
i) RX3-Cal и P4-Cal слитые белки эффективно выделяли из тканей табака благодаря их высокой растворимости в присутствии восстанавливающих агентов,
ii) энтерокиназное высвобождение кальцитонина из слитых белков выполняли после одной стадии очистки слитого белка с помощью анионообменной хроматографии, и
iii) обращенно-фазовая хроматография привела к очищенному СТ путем выделения его из смеси ЭК ферментативного гидролиза.
Масс-спектрометрический анализ высвобожденного СТ подтвердил, что точно удлиненный глицином СТ был получен с помощью растений табака.
Источники информации
Álvarez, I., Gelim M.I., Pimentel, Е., Ludevid, D., Torrent, M. 1998. Lysine-rich γ-zeins are secreted in transgenic Arabidopsis plants. Planta 205: 420-427.
Azria, M., Copp, D.H., Zanelli, J.M. 1995. 25 years of salmon calcitonin: from synthesis to therapeutic use. Calcif. Tissue Int. 57 (6): 405-408.
Bénicourt, C, Blanchard, C, Carriére, F., Verger, R. and Junien, J.L. 1993. Potential use of a recombinant dog gastric lipase as an enzymatic supplement to pancreatic extracts in cystic fibrosis, pp.291-295 in Clinical Ecology of Cystic Fibrosis. Escobar H, Baquero CF, Suarez L (eds). Elsevier Sciences Publishers BV, Amsterdam.
Bevan, M. 1984. Binary Agrobacterium vectors for plant transformation. Nucleic Acids Research 12: 8711-8721.
Bradbury, A.F. and Smyth, D.G. 1988. Biosynthesis of peptide neurotransmitters: studies on the formation of peptide amides. Physiol. Bohemoslov. 37: 267-274.
Conrad, U. and Fiedler, U. 1998. Compartment-specific accumulation of recombinant immunoglobulins in plant cells: an essential tool for antibody production and immunomodulation of physiological functions and pathogen activity. Plant Mol. Biol. 38: 101-109.
Cramer, C.L., Boothe, J.G. and Oishi, K.K. 1999. Transgenic plants for therapeutic proteins linking upstream and downstream strategies. Curr. Top. Microbiol. Immunol. 240: 95-118.
Cudd, A., Arvinte, Т., Das, R.E., Chinni, C. and MacIntyre, I. 1995. Enhanced potency of human calcitonin when fibrillation is avoided. J. Pharm. Sci. 84: 717-719.
Dilsen, S., Paul, W., Sandgathe, D., Tippe, R., Freudl, J., Kula, M.R., Takors, C, Wandrey, D. and Weuster-Botz, D. 2000. Fed-batch production of recombinant human calcitonin precursor fusion protein using Staphylococcus carnosus as an expression-secretion system. Appl. Microbiol. Biotechnol. 54: 361-369.
Draper, J., Scott, R. and Hamil, J. 1988. In: Plant Genetic Transformation and Gene Expression. A Laboratory Manual (Eds. Draper, J., Scott, R., Armitage, P. and Walden, R.), Blackwell Scientific Publications.
Düring, K., Hippe, S., Kreuzaler, F. and Schell, J. 1990. Synthesis and self-assembly of a functional monoclonal antibody in transgenic Nicotiana tabacum. Plant Mol. Biol. 15: 281-293.
Eipper, B.A., Stoffers D.A. and Mains R.E. 1992. The biosynthesis of neuropeptides: peptide α-amidation. Annu. Rev. Neurosci. 15: 57-85.
Fischer, R. and Emans, N. 2000. Molecular farming of pharmaceutical proteins. Transgenic Research 9: 279-299.
Geli, M.I., Torrent, M. and Ludevid, M.D. 1994. Two structural domains mediate two sequential events in γ-zein targeting: protein endoplasmic reticulum retention and protein body formation. Plant Cell 6: 1911-1922.
Haq, T.A., Mason, H.S., Clements, J.D. and Arntzen, C.J. 1995. Oral immunization with a recombinant bacterial antigen produced in transgenic plants. Science 268: 714-715.
Herman, E.M. and Larkins, B.A. 1999. Protein storage bodies. Plant Cell 11: 601-613.
Hong, D., Mingqiang, Z., Min, L., Changqing, C. and Jifang, M. 2000. Production of recombinant salmon calcitonin by amidation of precursor peptide using enzymatic transacylation and photolysis in vitro. Biochem. Biophys. Res. Com. 267: 362-367.
Kogan, M.J., Dalcol, I., Gorostiza, P., López-Iglesias, C., Pons, M., Sanz, F., Ludevid, M.D., and Giralt, E. 2001. Self-assembly of the amphipathic helix (VHLPPP)8. A mechanism for zein protein body formation. J. Mol. Biol. 00: 1-7.
Logemann, J., Schell, J., Willmitzer, L. 1987. Improved method for the isolation of RNA from plant tissues. Anal. Biochem. 163: 16-20.
Ludevid, M.D., Martinez-Izquierdo, J.A., Armengol, M., Torrent, M., Puigdomènech, P. and Palau, P., J. 1985. Immunological relations between glutelin-2 and low molecular weight zein-2 proteins from maize (Zea mays) endosperm. Plant sci. 41: 41-48.
Ludevid, M.D., Torrent, M., Martinez-Izquierdo, J.A., Puigdomènech, P. and Palau, P. 1984. Subcellular localization of glutelin-2 in maize. Plant Mol. Biol. 3: 277-234.
Ma, J.K., Hikmat, B.Y., Wycoff, K., Vine, N.D., Chargelegue. D., Yum L., Hein, M.B. and Lehner, T. 1998. Characterization of a recombinant plant monoclonal secretory antibody and preventive immunotherapy in humans. Nat. Med. 4: 601-606.
Ma, J.K. and Hein, M.B. 1995. Plant antibodies for immunotherapy. Plant Physiol. 109: 341-346.
McCormick, A.A., Kumagai, M.H., Hanley, K., Turpen, Т.Н., Hakim, I., Grill, L.K., Tusé, D., Levy, S. and Levy, R. 1999. Rapid production of specific vaccines for lymphoma by expression of the tumor-derived single-chain Fv epitopes in tobacco plants. Proc. Natl. Аса. Sci. USA 96 (2) 703-708.
McKee, C, Gibson, A., Dalrymple, M., Emslie, L., Garner, I. and Cottingham, I. 1998. Production of biologically active salmon calcitonin in the milk of transgenic rabbits. Nature Biotechnology 16: 647-651.
Merli, S., De Falco, S., Verdoliva, A., Tortora, M., Villain, M., Silvi, P., Cassani, G. and Fassina, G. 1996. An expression system for the single-step production of recombinant human amidated calcitonin. Protein Express. Purif. 7: 347-354.
Moore, P.J., Swords, K.M.M., Lynch, M.A. and Staehelin, L.A. 1991. Spatial organization of the assembly pathways of glycoproteins and complex polysaccharides in the Golgi apparatus of plants. J. Cell Biol. 112: 589-602.
Munro, S. and Pelham, H.R.B. 1987. A C-terminal signal prevents secretion of luminal ER proteins. Cell 48: 899-907.
Müntz, K. 1998. Deposition of storage proteins. Plant Mol. Biol. 38: 77-99.
Parmenter, D.L., Boothe, J.G., Van Rooijen, G.J., Yeung, E.C. and Moloney, M.M. 1995. Production of biologically active hirudin in plant seeds using oleosin partitioning. Plant Mol. Biol. 29: 1167-1180.
Ray, M.V., Van Duyne, P., Bertelsen, A.H., Jackson, Matthews, D.E., Sturmer, A.M., Merkler, D.J., Consalvo, A.P., Young, S.D., Gilligan, J.P. and Shields, P.P. 1993. Production of recombinant salmon calcitonin by in vitro amidation of an Escherichia coli produced precursor peptide. Bio/Technology 11: 64-70.
Reginster, J.Y. 1993. Calcitonin for prevention and treatment of osteoporosis. Am. J. Med. 95 (5A): 44S-47S.
Reichel, C, Mathur, J., Eckes, P., Langenkemper, K., Koncz, C, Schell, J., Reiss, В., and Maas, C. 1996. Enhanced green fluorescence by the expression of an Aequorea victoria green fluorescent protein mutant in mono- and dicotyledonous plant cells. Proc. Natl. Acad. Sci. USA 93: 5888-5893.
Sambrook, J., Fritsch, E., Maniatis, Т., 1989. Molecular Cloning: A laboratory manual. Cols Spring Harbor Laboratory, Cold Spring Harbor, New York.
Silverman, S.L. 1997. Calcitonin. Am. J. Med. Sci. 313 (1): 13-16.
Staub, J.M., Garcia, В., Graves, J., Hajdukiewicz, P.T., Hunter, P., Nehra, N., Paradkar, V., Schlittler, M., Caroll, J.A., Spatola, L., Ward, D., Ye, G. and Russell, D.A. 2000. High-yield production of a human therapeutic protein in tobacco chloroplasts. Nature Biotechnology 18: 333-338.
Stöger, E., Vaquero, C, Torres, E., Sack, M., Nicholson, L., Drossard, J., Williams, S., Keen, D., Perrin, Y., Christou, P. and Fischer, R. 2000. Cereal crops as viable production and storage systems for pharmaceutical scfv antibodies. Plant Mol. Biol. 42: 583-590.
Takahashi, K.I., Liu, Y.C., Hayashi, N., Goto, F., Kato, M., Kawashima, H. and Takeuchi, T. 1997. Production of bioactive salmon calcitonin from the nonendocrine cell lines COS-7 and CHO. Peptides 18: 439-444.
Torrent, M., Роса, E., Campos, N., Ludevid, M.D. and Palau, J. 1986. In maize, glutelin-2 and low molecular weight zeins are synthesized by membrane-bound polyribosomes and translocated into microsomal membranes. Plant Mol. Biol. 7: 393-403.
Torrent, M., Geli, M.I., Ruir-Avila, L., Canals, JM., Puigdomènech, P. and Ludevid, D. 1994. Role of structural domains for maize g-zein retention in Xenopus oocytes. Planta 192: 512-518.
Torres, E., Vaquero, C, Nicholson, L., Sack, M., Stöger, E., Drossard, J., Christou, P., Fischer, R. and Perrin, Y. 1999. Rice cell culture as an alternative production system for functional diagnostic and therapeutic antibodies. Transgenic Research 8: 441-449.
Verch, Т., Yusibov, V. and Koprowski, H. 1998. Expression and assembly of a full-length monoclonal antibody in plants using a plant virus vector. J. Immunol. Methods 220: 69-75.
Vitale, A., Ceriotti, A. and Denecke, J. 1993. The role of the endoplasmic reticulum in protein synthesis, modification and intracellular transport. J. Exp. Bot. 44: 1417-1444.
Vitale, A. and Denecke, J. 1999. The endoplasmic reticulum-gateway of the secretory pathway. Plant Cell 11: 615-628.
Wandelt, С.I., Khan, M.R.I., Craig, S., Schroeder, H.E., Spencer, D. and Higgins, T.J.V. 1992. Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulum and accumulates to high levels in the leaves of transgenic plants. Plant J. 2: 181-192.
Yabuta, M., Suzuki, Y. and Ohsuye, K. 1995. High expression of a recombinant human calcitonin precursor peptide in Escherichia coli. Appl. Microbiol. Biotechnol. 42 (5): 703-708.
Yamamoto, K., Fujii, R., Toyofuku, Y., Saito, Т., Koseki, H., Hsu V.W. and Aoe, T. 2001. The KDEL receptor mediates a retrieval machanism that contributes to quality control at the endoplasmic reticulum. EMBO J. 20 (12): 3082-3091.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОИЗВОДСТВО БЕЛКОВ | 2005 |
|
RU2429243C2 |
| ВЫДЕЛЕНИЕ И ОЧИСТКА БЕЛКА | 2005 |
|
RU2420585C2 |
| ПОЛУЧЕНИЕ БИОЛОГИЧЕСКИ АКТИВНЫХ БЕЛКОВ | 2007 |
|
RU2441911C2 |
| ПЕСТИЦИДНЫЕ КОМПОЗИЦИИ И ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ | 2013 |
|
RU2662672C2 |
| РАСТИТЕЛЬНЫЙ ПЕПТИД ГАММА-ЗЕИН ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2011 |
|
RU2563805C2 |
| РАСТЕНИЯ ПШЕНИЦЫ С ПОВЫШЕННОЙ УСТОЙЧИВОСТЬЮ К ИМИДАЗОЛИНОНОВЫМ ГЕРБИЦИДАМ | 2002 |
|
RU2337532C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТА ИЗ BRASSICA | 2013 |
|
RU2636035C2 |
| УСТОЙЧИВЫЕ К БАКТЕРИЯМ ТРАНСГЕННЫЕ РАСТЕНИЯ | 2012 |
|
RU2640246C2 |
| ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ОПЕРАТИВНО СЛИТЫЙ ИНТРАМОЛЕКУЛЯРНЫЙ ДИМЕР БЕЛКА KillerRed, КАССЕТА ЭКСПРЕССИИ, КЛЕТКА, ПРОДУЦИРУЮЩАЯ ХИМЕРНЫЙ БЕЛОК И СОДЕРЖАЩАЯ КАССЕТУ ЭКСПРЕССИИ, ВЫДЕЛЕННЫЙ ХИМЕРНЫЙ БЕЛОК | 2011 |
|
RU2491342C1 |
| ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ БЕЛОК KillerRed (ВАРИАНТЫ), ВЫДЕЛЕННЫЙ БЕЛОК (ВАРИАНТЫ), КАССЕТА ЭКСПРЕССИИ, КЛЕТКА, СОДЕРЖАЩАЯ КАССЕТУ ЭКСПРЕССИИ | 2011 |
|
RU2458129C1 |







Растения трансформируют с использованием нуклеиново-кислотных конструктов, содержащих первую нуклеотидную последовательность, которая кодирует γ-зеин, или ее фрагмент, способный направлять и удерживать белок в эндоплазматическом ретикулуме клетки растения, вторую последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность, которая специфическим образом расщепляется ферментными или химическими средствами, и третью последовательность нуклеиновой кислоты, которая кодирует целевой пептид или белок. Трансформация растения такими конструктами позволяет получать слитые белки, накапливающиеся в эндоплазматическом ретикулуме клеток в форме белковых телец, из которых могут быть выделены целевые белки, в частности кальцитонин. 10 н.и 37 з.п. ф-лы, 19 ил.
1. Последовательность нуклеиновой кислоты для экспрессии слитого белка в системе хозяина, включающая:
первую последовательность нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует белок γ-зеин, или ее фрагмент, содержащий нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в эндоплазматическом ретикулуме (ЭР) растительной клетки;
вторую последовательность нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует аминокислотную последовательность, которая специфически расщепляется с помощью ферментативных или химических способов; и
третью последовательность нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует интересующий продукт;
где 3′-конец указанной первой последовательности нуклеиновой кислоты связан с 5′-концом указанной второй последовательности нуклеиновой кислоты и 3′-конец указанной второй последовательности нуклеиновой кислоты связан с 5′-концом указанной третьей последовательности нуклеиновой кислоты.
2. Последовательность нуклеиновой кислоты по п.1, где указанная первая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует полный белок γ-зеина.
3. Последовательность нуклеиновой кислоты по п.1, где указанная первая последовательность нуклеиновой кислоты содержит:
одну или более нуклеотидных последовательностей, кодирующих весь или часть повторяющегося домена белка γ-зеина;
одну или более нуклеотидных последовательностей, кодирующих весь или часть ProX домена белка γ-зеина; или
одну или более нуклеотидных последовательностей, кодирующих весь или часть повторяющегося домена белка γ-зеина, и одну или более нуклеотидных последовательностей, кодирующих весь или часть РrоХ домена белка γ-зеина.
4. Последовательность нуклеиновой кислоты по п.1, где указанная первая последовательность нуклеиновой кислоты выбрана из следующей группы:
нуклеотидная последовательность, показанная в SEQ ID NO:l [нуклеотидная последовательность, которая кодирует γ-зеин (фиг.1А)],
нуклеотидная последовательность, показанная в SEQ ID NO:2 [нуклеотидная последовательность, идентифицированная как RX3 (фиг.1В)],
нуклеотидная последовательность, показанная в SEQ ID NO:3 [нуклеотидная последовательность, идентифицированная как R3 (фиг.1В)],
нуклеотидная последовательность, показанная в SEQ ID NO:4 [нуклеотидная последовательность, идентифицированная как Р4 (фиг.1С)], и
нуклеотидная последовательность, показанная в SEQ ID NO:5 [нуклеотидная последовательность, идентифицированная как Х10 (фиг.1С)].
5. Последовательность нуклеиновой кислоты по п.1, где указанная вторая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность, которая определяет сайт расщепления протеазой.
6. Последовательность нуклеиновой кислоты по п.5, где указанная протеаза является энтерокиназой, Arg-C эндопротеазой, Glu-C эндопротеазой, Lys-C эндопротеазой или фактором Ха.
7. Последовательность нуклеиновой кислоты по п.1, где указанная вторая последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует аминокислоту, которая специфически расщепляется с помощью химических реагентов.
8. Последовательность нуклеиновой кислоты по п.7, где указанный химический реагент является цианбромидом.
9. Последовательность нуклеиновой кислоты по п.1, где указанный интересующий продукт является белковым лекарственным средством.
10. Последовательность нуклеиновой кислоты по п.9, где указанное белковое лекарственное средство представляет собой пептидный гормон или интерферон, причем указанное лекарственное средство эффективно для лечения человека и животного.
11. Последовательность нуклеиновой кислоты по п.10, где указанный пептидный гормон выбирают из кальцитонина, эритропоэтина, тромбопоэтина и гормона роста.
12. Последовательность нуклеиновой кислоты по п.1, где указанная третья последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую кальцитонин, и кодон для глицина при 3′-конце указанной последовательности нуклеиновой кислоты, кодирующей кальцитонин.
13. Слитый белок, содержащий:
(i) аминокислотную последовательность белка γ-зеина, или ее фрагмент, способный к направлению и сохранению белка в ЭР растительной клетки,
(ii) аминокислотную последовательность, которая специфически расщепляется с помощью ферментативных или химических способов, и
(iii) интересующий продукт;
причем указанный слитый белок является продуктом экспрессии последовательности нуклеиновой кислоты по любому из пп.1-12 в системе растения-хозяина.
14. Слитый белок по п.13, содержащий полный белок γ-зеина.
15. Слитый белок по п.14, содержащий аминокислотную последовательность, показанную на фиг.1А и идентифицированную как SEQ ID NO:6.
16. Слитый белок по п.15, содержащий фрагмент белка γ-зеина, при этом указанный фрагмент содержит аминокислотную последовательность, способную к направлению и сохранению белка в ЭР растительной клетки.
17. Слитый белок по п.16, содержащий фрагмент белка γ-зеина, выбранный из группы, состоящей из:
аминокислотной последовательности, показанной в SEQ ID NO:7 [аминокислотная последовательность, соответствующая RX3 (фиг.1В)],
аминокислотной последовательности, показанной в SEQ ID NO:8 [аминокислотная последовательность, соответствующая R3 (фиг.1В)],
аминокислотной последовательности, показанной в SEQ ID NO:9 [аминокислотная последовательность, соответствующая Р4 (фиг.1С)], и
аминокислотной последовательности, показанной в SEQ ID NO:10 [аминокислотная последовательность, соответствующая X10 (фиг.1С)].
18. Слитый белок по п.13, где указанная аминокислотная последовательность, которая специфически расщепляется с помощью ферментативных способов, содержит сайт расщепления протеазой.
19. Слитый белок по п.13, где указанная аминокислотная последовательность, которая специфически расщепляется с помощью химических способов, содержит сайт, расщепляемый с помощью химического реагента.
20. Слитый белок по п.13, где указанный интересующий продукт является белковым лекарственным средством.
21. Конструкция нуклеиновой кислоты, содержащая (i) последовательность нуклеиновой кислоты по п.1, и (ii) регуляторную нуклеотидную последовательность, которая регулирует транскрипцию нуклеиновой кислоты по изобретению (i), причем указанная регуляторная последовательность (ii) является функциональной в растениях.
22. Конструкция по п.21, где указанная регуляторная последовательность (ii) является тканеспецифичной.
23. Конструкция по п.21, где указанная регуляторная последовательность (ii) содержит промотор, функциональный в растениях.
24. Конструкция по п.22, где указанная регуляторная последовательность (ii) содержит промотор 35SCaMV, промотор пататина, промотор запасного белка, промотор гена убихитина или регуляторную последовательность гена γ-зеина.
25. Конструкция по п.21, где указанная регуляторная последовательность (ii) содержит последовательность окончания транскрипции, функциональную в растении.
26. Конструкция по п.25, где указанная регуляторная последовательность (ii) содержит терминатор 35SCaMV, терминатор гена октопинсинтазы (ocs), терминатор гена нопалинсинтазы (nos) или терминатор гена γ-зеина.
27. Конструкция по п.21, где указанная регуляторная последовательность (ii) далее содержит трансляционный энхансер, функциональный в растении.
28. Конструкция по п.27, где указанный трансляционный энхансер, функциональный в растении, содержит промотирующую последовательность для транскрипции вируса гравировки томата.
29. Вектор, содержащий экспрессибельную последовательность нуклеиновой кислоты по п.1 для трансформации системы-хозяина.
30. Вектор, содержащий экспрессибельную конструкцию нуклеиновой кислоты по п.21 для трансформации системы растения-хозяина.
31. Трансформированная система растения-хозяина, причем указанная система растения-хозяина трансформируется нуклеиновой кислотой по п.1, или конструкцией по п.21, или вектором по п.29, или вектором по п.30.
32. Трансгенная система растения-хозяина, причем указанная трансгенная система растения-хозяина содержит интегрированную в его геном последовательность нуклеиновой кислотой по п.1.
33. Система растения-хозяина по п.31, где указанное растение является однодольным или двудольным растением.
34. Система растения-хозяина по п.33, где указанное растение представляет собой зерновые, бобовые, крестоцветные или пасленовые.
35. Система растения-хозяина по п.31, где растение содержит семена.
36. Система растения-хозяина по п.32, где указанное растение является однодольным или двудольным растением.
37. Система растения-хозяина по п.36, где указанное растение представляет собой зерновые, бобовые, крестоцветные или пасленовые.
38. Система растения-хозяина по п.32, где растение содержит семена.
39. Способ получения интересующего продукта в системе растения-хозяина, который включает выращивание трансформированной системы растения-хозяина по п.31 в условиях, которые допускают получение и экспрессию указанного интересующего продукта в форме слитого белка.
40. Способ получения интересующего продукта в системе растения-хозяина, который включает выращивание трансгенной системы растения-хозяина по п.32 в условиях, которые допускают получение и экспрессию указанного интересующего продукта в форме слитого белка.
41. Способ по п.39, который далее включает выделение и очистку указанного слитого белка.
42. Способ по п.39, который далее включает высвобождение указанного интересующего продукта из указанного слитого белка.
43. Способ по п.40, который далее включает выделение и очистку указанного слитого белка.
44. Способ по п.40, который далее включает высвобождение указанного интересующего продукта из указанного слитого белка.
45. Способ получения кальцитонина в системе растения-хозяина, включающий:
а) трансформирование системы растения-хозяина вектором экспрессии или конструкцией нуклеиновой кислоты, содержащей (i) регуляторную последовательность и (ii) последовательность нуклеиновой кислоты состоящей из:
первой последовательности нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует белок γ-зеин, или ее фрагмент, содержащий нуклеотидную последовательность, которая кодирует аминокислотную последовательность, способную к направлению и сохранению белка в эндоплазматическом ретикулуме (ЭР) растительных клеток;
второй последовательности нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует аминокислотную последовательность, которая специфически расщепляется с помощью ферментативных или химических способов; и
третьей последовательности нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует кальцитонин;
где 3′-конец указанной первой последовательности нуклеиновой кислоты связан с 5′-концом указанной второй последовательности нуклеиновой кислоты и 3′-конец указанной второй последовательности нуклеиновой кислоты связан с 5′-концом указанной третьей последовательности нуклеиновой кислоты; и
где регуляторная последовательность регулирует транскрипцию упомянутой последовательности нуклеиновой кислоты
b) генерирование полных растений из указанных систем растений-хозяев, трансформированных указанным вектором экспрессии или конструкцией нуклеиновой кислоты;
c) выращивание таких трансформированных растений при условиях, которые обеспечивают получение и экспрессию кальцитонина в форме слитого белка; и, если желательно,
d) выделение, очистку указанного слитого белка и обработку указанного слитого белка для того, чтобы высвободить кальцитонин.
46. Способ по п.41, где указанная первая последовательность нуклеиновой кислоты кодирует полный белок γ-зеина.
47. Способ по п.41, где указанная первая последовательность нуклеиновой кислоты кодирует фрагмент белка γ-зеина.
| WO 00/40738 А, 13.07.2000 | |||
| US 5948682 А, 07.09.1999. |

