Изобретение относится к медицине, более точно к физиологии, космической медицине, и может найти применение при устранении негативного последствия гипокинезии или гравитационной нагрузки.
Известно, что при функциональной разгрузке мышц цитоскелетные и сократительные белки подвергаются деструкции (A.Chopard, P.Francoise and J.-F.Marini, 2001). Динамика деструкции неодинакова для различных цитоскелетных, мембранных и сократительных белков. Однако уже через 14 дней разгрузки атрофия m. soleus достигает 35-40%. Клеточные факторы, запускающие процесс атрофии, исследованы недостаточно. Была показана роль кальция в процессе запуска протеолиза цитоскелетных белков (D.L.Enns, Т.Raastad, 2007). Обнаружено, что при «вывешивании» крыс в m. soleus происходит увеличение концентрации кальция (Inglas и соавторы, 1999, Shenkman B.S и соавторы, 2006). Кальций может активировать кальпаины. Через 2 недели разгрузки в мышце увеличивается количество десмин-негативных волокон, снижается количество титина и небуллина, дистрофина, а также сократительных белков. При введении блокатора L-кальциевых каналов нифедипина снижение концентрации кальция во время вывешивания задних конечностей крыс приводит к полному или частичному предотвращению разрушения цитоскелетных белков (Nemirovskaya et al., 2007). Изменения, которые мы наблюдаем при разгрузке мышц, во многом схожи с теми, что происходят в мышце после эксцентрической тренировки или при различных заболеваниях мышц, в том числе дистрофии Дюшена. При гипокинезии или гравитационной разгрузке мышц не определяли концентрацию NO в мышечных волокнах. В то же время при дистрофии Дюшена в поврежденных мышцах отмечали снижение активности нейрональной и эндотелиальной NO-синтаз (nNOS и eNOS) и увеличение протеолитической активности кальпаинов. Barton и соавторы (2005), а также Voisin и соавторы (2005) вводили L-аргинин мышам линии mdx и обнаружили у них увеличение количества утрофина (белка, заменяющего дистрофин при заболевании дистрофией Дюшена) и гамма-саркогликана. Одновременно у этих мышей снизилась проницаемость мембраны для синьки Эванса, снизилось количество коллагена, количество липидов в цитоплазме и увеличилась изометрическая сила. Авторы обращают внимание, что фармакологическая активизация NO путей может использоваться для лечения дистрофии Беккера и Дюшена. Однако механизм этой активации и сигнальные пути, которые могут быть запущены с помощью NO, авторами не исследовались. Функция NO в связи с регуляцией белкового метаболизма при гравитационной разгрузке и гипокинезии остается неизученной. Известно, что уровень концентрации нейрональной NO-синтазы снижается при длительном уменьшении двигательной активности (M.Salanova et al., 2008). А добавление NO в культуру мышечных клеток С2С12 частично предотвращает распад цитоскелетных белков, который провоцируется введением кальциевых ионофоров (T.J.Koh, J.G.Tidball, 2000).
Для предотвращения негативного действия гипокинезии и и/или гравитационной разгрузки использовались различные способы (физическая тренировка во время космического полета, применение нагрузочного костюма «Пингвин», стимуляция опорных зон стопы при гипокинезии, электростимуляция мышц при гипокинезии, применение препарата нифедипина, растяжение мышцы в модели «вывешивания» животных). Наиболее близким является способ, описанный Мухиной A.M. и др. 2006), в Российском физиологическом журнале (A.M.Мухина, Э.Г.Алтаева, Т.Л.Немировская, Б.С.Шенкман. Роль кальциевых каналов L-типа в накоплении Са2+ в волокнах m. soleus крысы и изменении соотношения изоформ миозина и SERCA при гравитационной разгрузке. Росс. Физиол. журнал. 2006, т.92 №11, С.1285-1295) и в обзоре B.S.Shenkman, T.L.Nemirovskaya в 2008 г (B.S.Shenkman, T.L.Nemirovskaya. Calcium-dependent signaling mechanisms and soleus fiber remodeling under gravitational unloading. J. Muscle Res Cell Motil. 2008, Vol.29, No.6-8, P.221-230). Согласно этому способу введение блокатора L-кальциевых каналов нифедипина снижает концентрацию кальция во время вывешивания задних конечностей крыс, что приводит к полному или частичному предотвращению разрушения цитоскелетных белков.
Однако этот известный способ не может применяться для предотвращения атрофии при гипокинезии или в космическом полете человека, т.к. нифедипин является препаратом, который может также оказывать влияние и на сердечную деятельность и требует тщательной индивидуальной подборки дозировки препарата перед применением и может иметь индивидуальные ограничения для применения.
Техническим результатом предлагаемого изобретения является то, что он обеспечивает высокую степень профилактики и устранения атрофии мышц за счет предотвращения снижения синтеза мРНК тяжелых цепей миозина I типа и одновременно исключает негативные побочные эффекты, присущие прототипу.
Этот технический результат достигается тем, что в известном способе профилактики деструкции цитоскелетных белков скелетных мышц при их атрофии, вызванной гипокинезией и/или гравитационной разгрузкой путем введения фармакологического препарата, в качестве фармакологического препарата вводят L-аргинин. При этом L-аргинин вводят в количестве, обеспечивающем снижение синтеза мРНК Е3 лигаз, участвующих в протеасомной деградации белка. L-аргинин вводят в количестве, обеспечивающем предотвращение снижения мРНК тяжелых цепей миозина 1 типа.
Разрабатывая предлагаемый способ, мы предположили, что N0 может являться сигнальной молекулой, имеющей отношение к регуляции белкового метаболизма, а введение донора NO L-аргинина может предотвратить или снизить атрофические процессы в мышце при ее функциональной разгрузке. Действительно, наши эксперименты подтвердили наше предположение и позволили выявить средство, обеспечивающее профилактику и снижение деструкции цитоскелетных белков скелетных мышц при их атрофии, вызванной гипокинезией и/или гравитационной разгрузкой.
Для подтверждения правомерности наших притязаний приводим следующие результаты экспериментов.
Организация и методы исследования
28 самцов 2,5 месячных крыс Wistar массой 220-240 г было разделено на 4 группы (по 7 животных в каждой группе). Одна группа - контроль (гр. К, вес тела 253±9 г). Остальные группы вывешивали в течение 14 дней по методу Morey-Holton таким образом, чтобы задние конечности не касались земли, а передние опирались на пол. Хвост крыс перед процедурой вывешивания был обработан специальным образом для избежания потертости и повреждений и заключен в мягкие специальные шины. Деление на группы: вывешивание крыс без дополнительных воздействий (гр. В, вес крыс 246±10 г); вывешивание с введением L-аргинина с питьевой водой (гр. ВЛ, концентрация L-аргинина 500 мг/кг веса крысы, вес крыс 249±7 г); вывешивание крыс с введением L-NAME (L-NAME - N-nitro L-arginine methyl ester hydrochloride) с питьевой водой (гр. ВН, концентрация L-NAME ежедневно вводили с питьевой водой 90 мг/кг веса крысы, вес крыс 246±7 г).
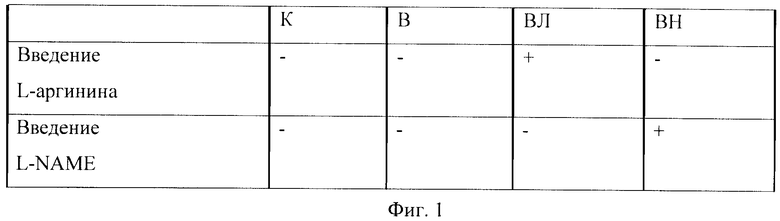
Схема эксперимента и наименования групп отражены на Фиг.1.
Для определения концентрации NO в m. soleus был проведен повторный эксперимент по приведенной выше схеме, где крысам перед забоем вводили специальную ловушку. Для этого в каждой группе было взято по 6 крыс.
Схема вывешивания крыс отражена на Фиг.2.
Крыс забивали сверхдозой нембутала (75 мг/кг веса), m. soleus немедленно замораживали в жидком азоте. Пробы хранили при -85°С. Выявляли концентрацию NO в m. soleus. Определяли содержание дистрофина на поперечных срезах мышцы, десмина и α-актина методом иммуноблота, уровень мРНК Е3 лигаз атрогена-1 и MuRF1, мРНК тяжелых цепей миозина (ТЦМ) I, IIa, IIb, IId/x методом ПЦР в реальном времени.
Определение концентрации NO в m. soleus.
M. soleus была заморожена и хранилась в жидком азоте. Относительное содержание оксида азота в мышце определяли по содержанию в ней парамагнитных центров, количество которых пропорционально концентрации оксида азота в ткани. Электронно-парамагнитный резонанс (ЭПР): крысе вводили спиновую ловушку - диэтилдитиокарбамат (ДЭТК) из расчета 500 мг/кг веса, которая равномерно распределялась в органах и тканях. ДЭТК связывается с оксидом азота, образуя парамагнитные комплексы, которые находятся в равновесии со стационарной концентрацией оксида азота в ткани.
Сразу после введения ДЭТК животному внутримышечно вводили водный раствор смеси 29 мМ FeSO4 и 116 мМ цитрата натрия (2,0 мл/кг веса). Через 30 мин животных декапитировали, m. soleus замораживали в жидком азоте и регистрировали сигнал ЭПР при температуре жидкого азота на ЭПР-спектрометре фирмы "Bruker EMX-8". Сигнал ЭПР представляет собой суперпозицию сигнала комплекса NO - Fe2+(ДЭТК)2, а также других парамагнитных центров, которые обнаруживаются в тканях. Однако в регистрируемом сигнале видны компоненты, которые по форме и положению сигнала совпадают с сигналом ЭПР мононитрозильных комплексов железа с дитиокарбаматом. Это позволяет оценивать относительную концентрацию оксида азота по первой компоненте триплетной сверхтонкой структуры этого комплекса (M.Yu.Obolenskaya et. Al., 1994, A.F.Vanin et al., 2002).
Выявление дистрофина
Поперечные срезы мышечной ткани, сделанные в криостате, толщиной 10 мкм инкубировали с первичными моноклональными антителами против дистрофина (1:20, Novocastra, NCL-DYSI) и вторичными поликлональными антителами, конъюгированными с флуоресцентной меткой (GAM, 1:1000, Alexa546, Molecular probes). Все инкубации проводили в течение часа при комнатной температуре. Подсчитывали процентное содержание волокон с поврежденным дистрофиновым слоем. Анализировали не менее 100 волокон с каждого среза.
Электрофорез в ПААГ nNOS, десмина и α-актина
С каждой пробы m. soleus были сделаны срезы толщиной 20 мкм (10-15 мг) и немедленно прогомогенизированы 3 раза по 10 сек в течение 25 мин в 100 мкл буфера, содержащего для nNOS: 62,5 мМ Tris-HCl (pH 6,8), 150 мМ NaCl, 2% Triton Х-100, 2% SDS, 1 мМ EDTA, 1 мМ PMSF, 1 мМ DTT, Complite mini (коктейль ингибиторов протеаз. Roche, Germany). Состав буфера для десмина и α-актина: 62,5 мМ Tris-HCl (pH 6,8), 2% SDS, 10% Glicerin, 5% β-mercaptoethanol, 0,01% bromphenol blue. Затем образцы отцентрифугировали при 20000 g 25 мин. Пробы для nNOS были разведены в соотношении 1:1 в следующем буфере для образцов (5,4 мМ Tris-HCl (pH 6,8), 4% SDS, 20% Glicerin, 10% β-mercaptoethanol, 0,02% bromphenol blue). Все пробы прокипятили при 95°С в течение 2 мин. Часть супернатанта из всех проб отобрали для определения концентрации общего белка с помощью RC DC Protein Assay (Bio-Rad Laboratories, США), оставшийся объем использовали для электрофореза в ПААГ. Все образцы поместили на хранение при -84°С. Электрофорез nNOS проводили в 8% разделяющем, а десмина и α-актина в 12% разделяющем акриламидных гелях (0,2% метилбисакриламид, 0,1% SDS, 375 мМ Tris-HCl (pH 8,8); 0,05% APS; 0,1% TEMED) и 5% концентрирующем акриламидном геле (0,2% метилбисакриламид, 0,1% SDS, 125 мМ Tris-HCl (pH 6,8), 0,05% APS; 0,1% TEMED). Использовали катодный (192 мМ Tris-Glycine (pH 8,6), 0,1% SDS) и анодный буфер (25 мМ Tris-HCl (pH 8,6)). Образцы загружались из расчета для nNOS 40 мкг, для десмина и α-актина 20 мкг общего белка на одну дорожку. Электрофорез проводили 75 минут при 15 мА на гель в мини-системе Bio-Rad при комнатной температуре.
Вестернблоттинг
Электроперенос белков производился в буфере (25 мМ Tris (pH 8,3), 192 мМ Glycine, 20% метанола, 0,04% SDS) на нитроцеллюлозную мембрану при 350 мА при температуре +4°С в системе mini Trans-Blot (Bio-Rad Laboratories, США) для nNOS в течение 1 часа 20 мин, а для десмина и α-актина 25 мин. Мембраны блокировали в растворе 5% сухого молока (Bio-Rad Laboratories, США) в TBST (4mM Tris-HCl (pH 7,4), 100 тМ NaCl, Tween20 0,1%) ночь при температуре +4°С. Для выявления были использованы поликлональные анти-nNOS (первичные BD Biosciences, разведение 1:250, вторичные GAR, ИМТЕК, 1:1000); против десмина (моноклональные антитела Novocastra, NCL-L-DES-DERII, 1:200), против α-актина (антитела SIGMA, 1:500), конъюгированные с пероксидазой хрена). Ввторичные биотинилированные антитела для выявления десмина и α-актина использованы GAM, ИМТЕК, 1:500. Затем блоты с десмином и α-актином икубировали с авидин-пероксидазным конъюгатом (SIGMA, 1:1000) в течение 30 мин с последующей проявкой с помощью 3% раствора перекиси водорода в TBST с диаминобензидином. nNOS (NO-синтазу) на блоте выявляли, используя Immun-Star™ Substrate Kit (Bio-Rad Laboratories, США). Инкубации с антителами проводили 1 час при комнатной температуре. Блоты отмывали 6Х10 мин в TBST. Вестернблоттинг был повторен не менее 3 раз. Анализ белковых полос производился с помощью денситометра GS-800 (Quantity-One™ software, BioRad). Оптическую плотность полосы белка контрольной группы на анализируемой мембране принимали за 100%, а оптическую плотность полос белков других групп сравнивали с контрольными пробами, расположенными на одной и той же мембране.
Анализ экспрессии генов. Обратная транскрипция
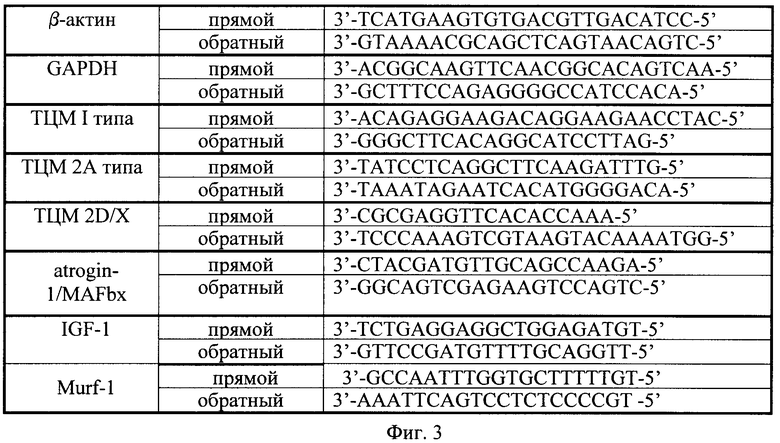
Тотальная РНК была экстрагирована из 10 мг замороженной m. soleus при помощи RNeasy Micro Kit (QIAGEN, Germany). Все образцы РНК были обработаны протеиназой К и ДНКазой I. Концентрация РНК была определена при 260 нм. Затем водный раствор выделенной РНК был заморожен при -84°С для последующего проведения обратной транскрипции. Для обратной транскрипции использовали водный раствор 1 мкг РНК, олиго(dT)15, случайные гексануклеотиды d(N)6, обратную транкскриптазу MMLV. Обратную транскрипцию проводили при 37°С в течение 60 мин согласно стандартному протоколу. Полученные образцы кДНК были помещены на хранение при -84°С для проведения ПЦР в реальном времени. Все образцы были проанализированы не менее 3 раз и все реакции измерены с помощью детектирующего амплификатора iQ5 Multicolor Real-Time PCR Detection System (Bio-Rad Laboratories, США) β-актин и GAPDH использовали в качестве референсных генов. Для проведения ПЦР-РВ применяли следующие праймеры (Фиг.3). Для оценки относительных изменений в уровне экспрессии специфического гена в пробе мы использовали следующую формулу:
ΔCt=Ct(реф)-Ct(тест),
где Ct(реф) - точка пересечения базовой линии и графика амплификации референсного гена в пробе, а Ct(тест) - точка пересечения базовой линии и графика амплификации изучаемого гена в той же пробе. Далее определяли в каждой группе среднее значение ΔCt и его стандартную ошибку. Изменение уровня экспрессии анализируемого гена в экспериментальных группах оценивали относительно контрольного уровня по формуле: ΔΔCt=ΔCt(группа)-ΔCt(контроль).
Статистический анализ
Обработку данных проводили с помощью программ Excel и SigmaPlot. При статистическом анализе результатов использовали одномерный вариант ANOVA и t-критерий Стьюдента.
Результаты
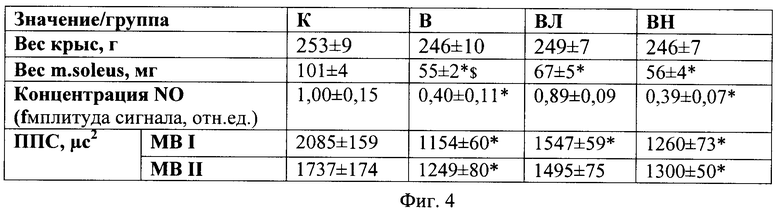
Вес крыс во всех группах не отличался от уровня контроля. Вес m. soleus был снижен в группах В и ВН (на 46 и 45%, p<0,05 соответственно) относительно группы контроля. В то же время у группы ВЛ вес мышцы был на 21% выше, чем в группе В (p<0,05) (Фиг.4), где показан вес животных и вес m. soleus крыс различных экспериментальных групп.
Аналогичные результаты получены для ППС MBI и MBII. В группе В она была снижена на 45% и 28% соответственно (p<0,05), а в группе ВН на 40% и 25% по сравнению с гр. К (p<0,05, табл.1). ППС MBI и MBII волокон в группе ВЛ был существенно выше, чем в группе В (на 25% и 16% (p<0,05) соответственно). Атрофия MBII волокон в гр. ВЛ была предотвращена полностью, т.к. их ППС существенно не отличалась от группы К (см. Фиг.4).
Содержание NO в m. soleus
В группе крыс, вывешенных с введением L-аргинина, концентрация NO в m. soleus не отличалась от группы контроля. У животных, вывешенных без препарата, или с введением L-NAME, концентрация N0 была соответственно на 60 и 61% ниже (p<0,05), чем в группе контроля (Фиг.4).
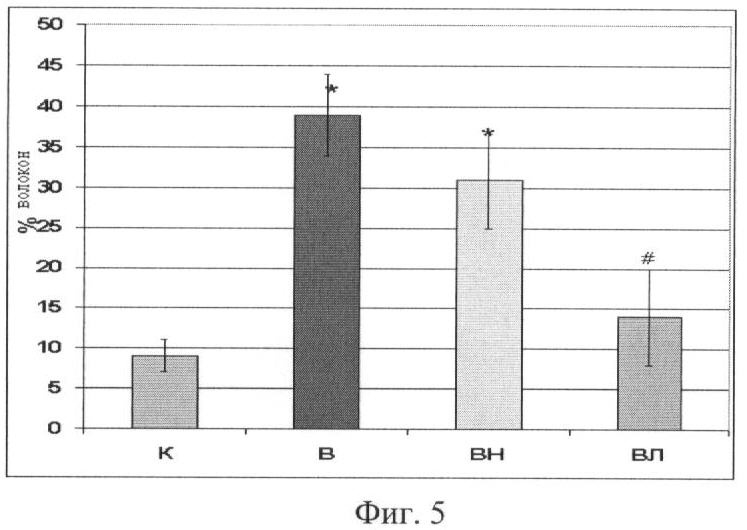
Процент волокон, имеющих разрывы в дистрофиновом слое
Мы обнаружили существенное увеличение числа волокон, имеющих разрывы в дистрофиновом слое, у вывешенных животных, и у вывешенных с введением L-NAME (Фиг.5).
Введение L-аргинина при вывешивании предотвратило эти изменения.
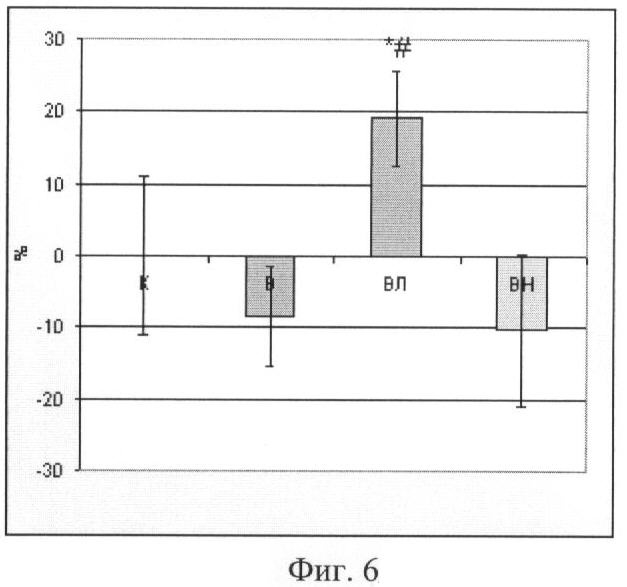
Концентрация α-актина в m. soleus
Концентрация белка α-актина у вывешенных без препарата животных и у крыс, вывешенных с введением блокатора нейрональной NO-синтазы, не отличается от уровня контроля. В то же время у животных, которым вводили донор NO L-аргинин, содержание α-актина существенно выше (p<0,05), чем у других вывешенных животных (Фиг 6.).
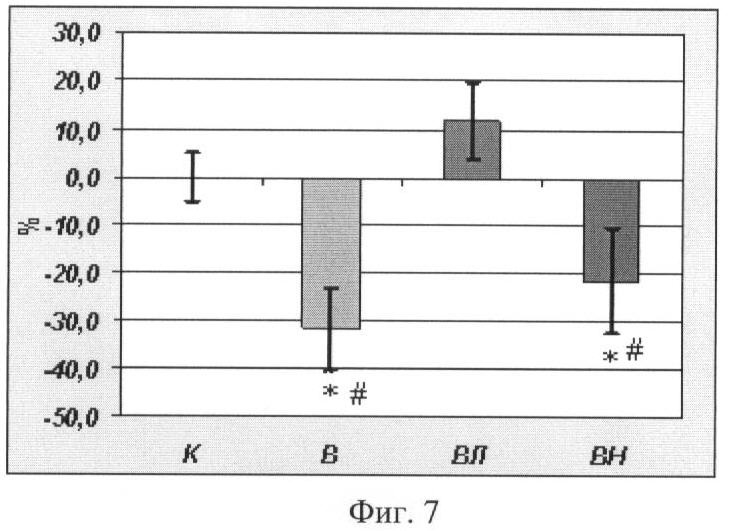
Содержание десмина в m. soleus
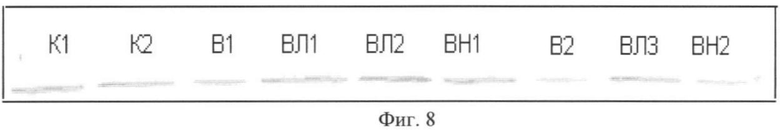
Содержание десмина в группах В и ВН было на 31 и 21% ниже, чем в группе контроля (p<0,05). В группе крыс, которым давали L-аргинин, содержание десмина в m. soleus было существенно выше, чем в группе В и ВН (p<0,05), как и в случае с концентрацией α-актина, см. Фиг.7, где К - контроль, В - вывешивание, ВЛ - вывешивание + L-аргинин, ВН - вывешивание + L-NAME. На рисунке отражено содержание десмина в m. soleus (в процентах от уровня контроля). На Фиг.8 приведено фото мембраны блота десмина.
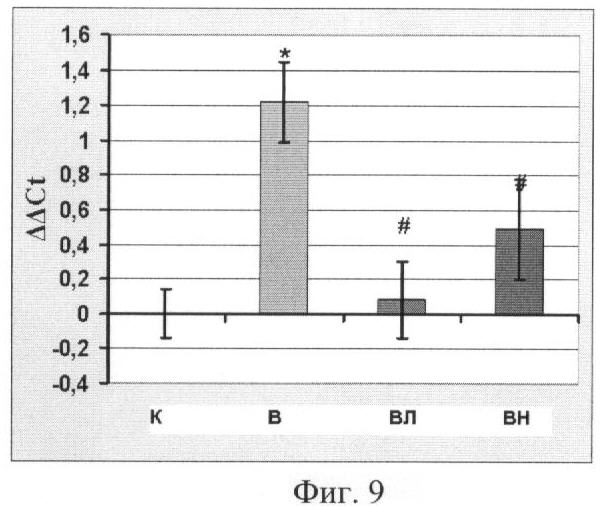
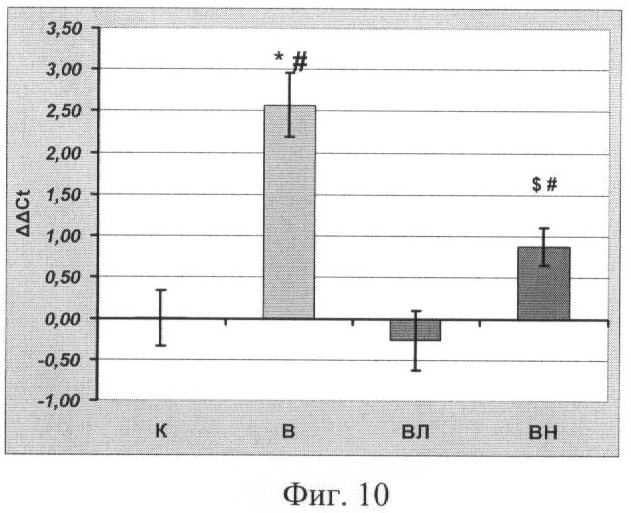
Концентрация мРНК Е3 лигаз атрогина-1 и MuRF-1 в m. soleus
Концентрация мРНК Е3 лигаз атрогина-1 и MuRF-1, отвечающих за протеолитический распад белков, была существенно повышена в группах В и ВН (p<0,05). Фиг.9 и Фиг.10. В противоположность этому у крыс, вывешенных с введением L-аргинина, концентрация мРНК этих лигаз не отличается от группы контроля. Это согласуется с данными по распаду цитоскелетных белков, приведенными выше. Распада белка в группе ВЛ также не наблюдалось, см. Фиг 5, 6, 7.
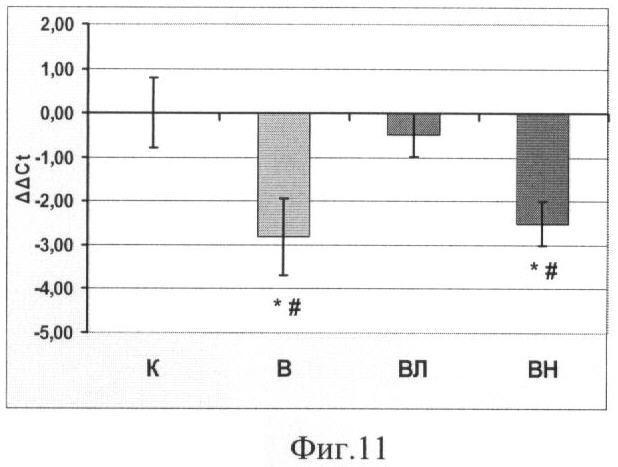
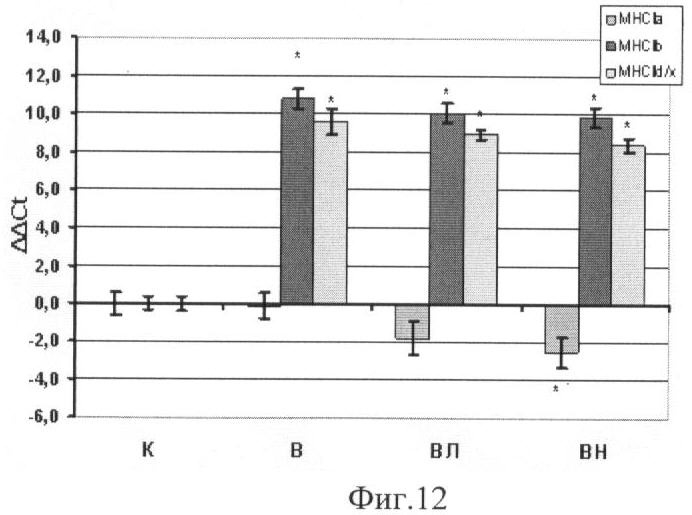
Концентрация мРНК тяжелых цепей миозина (МНС) I и IIa,b,d/x типов.
Введение L-аргинина при вывешивании предотвратило снижение мРНК МСН I (Фиг.11), которая наблюдалась как в группе В, так и в группе ВН. По концентрации мРНК МНС II в группе, получавшей L-аргинин, никаких отличий от других вывешенных животных не наблюдалось как в гр.В, так и ВН (Фиг.12.).
Т.о. нами впервые протестирована функция NO в разгруженной мышце и механизмы, с помощью которых эта молекула может быть включена в процессы регуляции белкового метаболизма. Мы впервые показали, что NO может выполнять роль сигнальной молекулы, регулирующей белковый метаболизм in vivo при гипокинезии и гравитационной разгрузке.
Для проверки эффективности введения дозы L-аргинина при вывешивании для изменения продукции NO в m. soleus мы измерили концентрацию NO, используя технику парамагнитного резонанса. Мы обнаружили, что 2-недельное вывешивание приводит к существенному падению концентрации NO в m. soleus, в то время как введение L-аргинина полностью предотвращает эти изменения (Фиг.4). Неожиданным было то, что введение блокатора NO-синтазы L-NAME не привело к более выраженному падению концентрации NO при вывешивании, чем в группе В.
Одновременно со снижением степени атрофии и площади поперечного сечения мышечных волокон (Фиг.4) в группе крыс, которым при вывешивании вводили L-аргинин (ВЛ), наблюдается предотвращение распада дистрофина (Фиг.5), а также существенно более высокое содержание α-актина и десмина (Фиг.6 и Фиг.7), чем в группах с вывешиванием без введения донора NO, или блокированием активности nNOS (гр. В и ВН). Разрушение дистрофина в soleus при вывешивании ранее было показано рядом авторов (CHOCKALINGAM et al. 2002, CHOPARD et al, 2001). Некоторые авторы ранее показывали, что при некоторых заболеваниях (например, дистрофии Дюшена) падение дистрофина может быть частично компенсировано при введении L-аргинина экспрессией утрофина у мышей mdx (E.R.Barton, 2005, V.Voisin at. Al., 2005). Однако не было сообщений о том, что повреждения дистрофинового слоя могут быть предотвращены с помощью повышения концентрации NO в мышце при гипокинезии.
Мы обнаружили падение концентрации десмина после 14 суток вывешивания. Däpp и соавторы показали снижение экспрессии десмина после 7 дней вывешивания крыс (2004). Enns и соавторы обнаружили снижение десмина после 3 дней вывешивания (2007). В то же время Chopard с соавторами не нашли каких-либо изменений в содержании десмина после 3 и 6 недель вывешивания, или после продолжительной гипокинезии у (2001, 2005). Можно предположить, что концентрация десмина снижается на первых фазах функциональной разгрузки мышц, а затем на более поздних фазах способна восстанавливаться. Введение L-аргинина предотвращает снижение концентрации десмина после 14 суток вывешивания животных. Т.о. можно заключить, что поддержание концентрации NO при функциональной разгрузке предотвращает снижение в ней концентрации таких цитоскелетных белков, как дистрофина, десмина, а также сократительного белка α-актина.
Известно, что в скелетных мышцах при функциональной разгрузке (и некоторых дистрофических заболеваниях) особенно интенсивно начинают активироваться такие Е3 лигазы, как атрогин-1 и MuRFl, которые задействованы в процессах протеасомной деградации (R.W.Jackman and S.C.Kandarian, 2004, E.E.Dupont-Versteegden et. Al., 2006, M.B.Reid, 2005). Концентрация их мРНК повышена у вывешенных животных (гр. В), (Фиг.9 и Фиг.10) и не отличается от группы контроля у животных, которым вводили L-аргинин. Таким образом, повышение концентрации NO при вывешивании крыс ведет к снижению концентрации мРНК Е3 лигаз, участвующих в протеасомной деградации белков.
Очевидно, что повышение концентрации NO при вывешивании ведет к снижению синтеза мРНК основных Е3 лигаз, участвующих в протеасомной деградации белка, таких как атрогин-1 и MuRF1.
Мы обнаружили снижение синтеза мРНК тяжелых цепей миозина (ТЦМ) I типа и повышение концентрации ТЦМ IIb и II d/x типа у вывешенных животных (группы В). Аналогичные изменения были описаны ранее (D.B.Thomason, R.B.Biggs and F.W.Booth, 1989). В то же время введение L-аргинина (группа ВЛ) полностью предотвратило эти изменения (Фиг. 11, 12). Ранее было показано, что введение L-NAME крысам в длительном эксперименте, где мышца подвергалась перегрузке, привело к предотвращению трансформации мышечных волокон в медленную сторону (Smith, Lori et al., 2002). Таким образом, был сделан вывод, что NO может участвовать в трансформации тяжелых цепей миозина. Мы показали, что увеличение концентрации NO при разгрузке мышц предотвращает снижение мРНК ТЦМ I типа.
Выводы:
1. Повышение концентрации NO в мышце при гипокинезии или гравитационной разгрузке снижает степень атрофии мышцы.
2. Поддержание концентрации NO при функциональной разгрузке предотвращает снижение в ней концентрации некоторых цитоскелетных белков.
3. Повышение концентрации NO при вывешивании ведет к снижению синтеза мРНК основных Е3 лигаз (таких как атрогин-1 и MuRF1), участвующих в протеасомной деградации белка.
4. Мы показали, что увеличение концентрации NO при разгрузке мышц предотвращает снижение в них концентрации мРНК тяжелых цепей миозина I типа.
Краткое описание чертежей
Фиг.1. Таблица - схема эксперимента и наименования групп.
Где К - контроль, В - вывешивание, ВЛ - вывешивание + введение L-аргинина, ВН - вывешивание + введение L-NAME.
Фиг.2. Схема вывешивания крыс.
Фиг.3. Праймеры, которые применяли для проведения ПЦР-РВ.
Фиг.4. Вес животных, вес m. soleus, ППС MB и концентрация NO у крыс различных экспериментальных групп.
Где * - отличия от группы контроля существенны при p<0.05,
$ - отличия от группы вывешивания существенны при p<0.05,
ППС - площадь поперечного сечения,
MB I, II - мышечные волокна I и II типа.
К - контроль, В - вывешивание, ВЛ - вывешивание + введение L-аргинина, ВН - вывешивание + введение L-NAME.
Фиг.5. Процент волокон, имеющих разрывы в дистрофиновом слое.
Где * - существенные отличия от К, p<0,05,
# - существенные отличия от В, p<0,05,
К - контроль, В - вывешивание, ВЛ - вывешивание + введение L-аргинина,
ВН - вывешивание + введение L-NAME
Фиг.6. Содержание α-актина в m. soleus. Где К - контроль, В - вывешивание, ВЛ - вывешивание + введение L-аргинина, ВН - вывешивание + введение L-NAME
Фиг.7. Содержание десмина в m. soleus (в процентах от уровня контроля). Где
К - контроль, В - вывешивание, ВЛ - вывешивание + L-аргинин, ВН - вывешивание + L-NAME
* - достоверные отличия от К, p<0,05,
# - достоверные отличия от гр. ВЛ.
Фиг.8. Фото мембраны блота десмина. Где
К - контроль, В - вывешивание, ВЛ - вывешивание + L-аргинин, ВН - вывешивание + L-NAME.
Фиг.9. Уровень мРНК атрогина-1. Где
К - контроль, В - вывешивание, ВЛ - вывешивание + L-аргинин, ВН - вывешивание + L-NAME.
* - достоверные отличия от К, p<0,05,
# - достоверные отличия от В, p<0,05.
Фиг 10. Концентрация Е3 лигазы MuRF-1 (референсный ген - β-актин). Где
* - достоверные отличия от К, p<0,05,
# - достоверные отличия от ВЛ, p<0,05,
$ - достоверные отличия от В, p<0,05,
К - контроль, В - вывешивание, ВЛ - вывешивание + L-аргинин, ВН - вывешивание + L-NAME.
Фиг.11. Концентрация мРНК тяжелых цепей миозина I типа в m. soleus. Где
* - существенные отличия от группы К, p<0,05,
# - существенные отличия от группы ВЛ, p<0,05,
К - контроль, В - вывешивание, ВЛ - вывешивание + L-аргинин, ВН - вывешивание + L-NAME.
Фиг.12. Концентрация мРНК тяжелых цепей миозина IIa, IIb, IId/x типов в m. soleus. Где
* - существенные отличия от группы К, p<0,05,
К - контроль, В - вывешивание, ВЛ - вывешивание + L-аргинин, ВН - вывешивание + L-NAME.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНГИБИТОР ПРОТЕОЛИЗА ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ФИЗИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2389486C1 |
| СПОСОБ ПРОФИЛАКТИКИ РАЗРУШЕНИЯ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ЭКСЦЕНТРИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2391968C1 |
| СРЕДСТВО ДЛЯ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517576C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517259C1 |
| ПРИМЕНЕНИЕ VX 745 ДЛЯ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2019 |
|
RU2714923C1 |
| СПОСОБ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2012 |
|
RU2481105C1 |
| Способ предотвращения атрофии, повышенной утомляемости медленных мышечных волокон и их трансформации в быстрые мышечные волокна в камбаловидной мышце | 2024 |
|
RU2834246C1 |
| Способ предотвращения атрофии скелетных мышц при их функциональной разгрузке | 2022 |
|
RU2797216C1 |
| Способ поддержания силы и жесткости постуральных мышц млекопитающих при их функциональной разгрузке | 2022 |
|
RU2800317C1 |
| Способ предотвращения развития утомления скелетной мышцы | 2023 |
|
RU2814077C1 |
Изобретение относится к медицине, более точно к физиологии, космической медицине, и может найти применение для профилактики деструкции цитоскелетных белков скелетных мышц при их атрофии, вызванной гипокинезией и/или гравитационной разгрузкой с целью устранения их негативного последствия. Для этого вводят L-аргинин. При этом L-аргинин можно вводить в количестве, обеспечивающем снижение синтеза мРНК Е3 лигаз, участвующих в протеасомной деградации белка. L-аргинин также можно вводить в количестве, обеспечивающем предотвращение снижения мРНК тяжелых цепей миозина 1 типа. Способ обеспечивает высокую степень профилактики и устранения атрофии мышц и одновременно исключает негативные побочные эффекты. 2 з.п. ф-лы, 12 ил.
1. Способ профилактики деструкции цитоскелетных белков скелетных мышц при их атрофии, вызванной гипокинезией и/или гравитационной разгрузкой, путем введения фармакологического препарата, отличающийся тем, что в качестве фармакологического препарата вводят L-аргинин.
2. Способ по п.1, отличающийся тем, что L-аргинин вводят в количестве, обеспечивающем снижению синтеза мРНК Е3 лигаз, участвующих в протеасомной деградации белка.
3. Способ по п.1, отличающийся тем, что L-аргинин вводят в количестве, обеспечивающем предотвращение снижения мРНК тяжелых цепей миозина 1-го типа.
| МУХИНА A.M | |||
| и др | |||
| Роль кальциевых каналов L-типа в накоплении Ca в волокнах m.soleus крысы и изменении соотношения изоформ миозина и SERCA при гравитационной разгрузке //Росс | |||
| Физиол | |||
| Журнал | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Спринклерная головка для водяных и химических огнетушителей с сигнализацией | 1924 |
|
SU1285A1 |
| УСТРОЙСТВО ДЛЯ РЕАБИЛИТАЦИИ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА В УСЛОВИЯХ НЕВЕСОМОСТИ | 1999 |
|
RU2148981C1 |
| ЛИТВИНОВА К.С | |||
| Сократительные характеристики отдельных скинированных волокон | |||

