Изобретение относится к медицине, более точно к физиологии, космической медицине и может найти применение при устранении негативного последствия гипокинезии или гравитационной разгрузке.
При функциональной разгрузке (или гипокинезии) белки скелетных мышц частично подвергаются распаду, а атрофия m.soleus через 2 недели достигает 40%. Немного известно о триггерах, или молекулярных сигнальных событиях, лежащих в основе этого процесса. Атрофия обусловлена снижением белкового синтеза и резким увеличением белкового распада (S.Kandarian и др., 2004). Деградацию сократительных белков может усиливать разрушение белков теплового шока (Hsp) 90 и 70, концентрация которых обычно очень высока в мышцах. Однако при функциональной разгрузке их уровень падает на 70-75% (параллельно атрофии) (Takashi Sakurai и др., 2005). Некоторые авторы полагают, что невозможность продуцировать мышцами Hsp(s) при разгрузке может иметь существенное значение для их атрофии. Известно, что белки теплового шока выполняют защитную функцию, поддерживая как протеиновый синтез, и перенос, так и защищая клетки от различных повреждающих факторов (Bruce C. Frier and Marius Locke, 2007). Мы предположили, что при разгрузке некоторые функции Hsp90 могут быть снижены (в частности, связанные с синтезом Hsp70). Для увеличения концентрации белков теплового шока в скелетных мышцах ранее были применены следующие подходы. K.Goto и др., 2004, создавали тепловой стресс животным перед восстановлением после функциональной разгрузки крыс и наблюдали увеличение концентрации Hsp72.
Наиболее близким способом (прототипом), направленным на предотвращения атрофии скелетных мышц при их функциональной разгрузке, можно считать способ Sent, S.М. (Senf S.M., Dodd S.L., McClung J.M., Judge A.R. Hsp70 overexpression inhibits NF-kappaB and Foxo3a transcriptional activities and prevents skeletal muscle atrophy // FASEB J., 2008; 22(11): p.3836-3845), согласно которому вызванная введением плазмид экспрессия Hsp70 предотвратила развитие атрофии тех волокон m.soleus, куда она была инкорпорирована.
При этом не наблюдалось увеличение транскрипционной активности Foxo3a и NF-kapaB, а также увеличение концентрации Е3 лигаз atrogin-1 и MuRF1. Авторы пришли к выводу, что введение плазмиды с участком, содержащим Hsp70, эффективно ингибирует ключевые сигнальные пути, инициирующие атрофию. Стоит, правда, отметить, что эффективность трансфекции обычно не превышает 12-15%. Поэтому в практических целях она мало применима. При этом не наблюдалось увеличение транскрипционной активности Foxo3a и NF-kapaB, а также увеличение концентрации Е3 лигаз atrogin-1 и MuRF1. Авторы пришли к выводу, что введение плазмиды с участком, содержащим Hsp70, эффективно ингибирует ключевые сигнальные пути, инициирующие атрофию. Стоит, правда, отметить, что эффективность трансфекции обычно не превышает 12-15%. Поэтому в практических целях она мало применима.
Техническим результатом заявленного изобретения является повышение эффективности степени предотвращения атрофии путем более эффективного ингибирования ключевых сигнальных путей, инициирующих ее.
Этот технический результат достигается тем, что в известном способе предотвращения атрофии скелетных мышц при их функциональной разгрузке путем воздействия на организм фактора, увеличивающего в мышцах количество белков теплового шока в качестве фактора, увеличивающего количество белков теплового шока, используют введение 17-(allylamino)-17-demethoxygeldanamycin (17-AAG), который дополнительно уменьшает уровень µ-кальнаина.
Препарат 17-AAG широко применяется в онкологии, т.к. имеет избирательную антиопухолевую активность. Этот феномен был отмечен во множестве статей, в том числе Francis Burrows et al., 2004. Это происходит из-за того, что в опухолевых клетках существенно увеличивается связывающая способность Hsp90 и 17-AAG. Однако для обычных тканей этого не показано.
В отличие от предыдущих работ, мы использовали препарат 17-AAG, который регулирует концентрацию Hsp90 и Hsp70 в мышцах при функциональной разгрузке и тем самым предотвращает их атрофию. В связи с этим мы модифицировали функцию Hsp90, которая затрагивает синтез белков теплового шока Hsp(s). Для этого была блокирована работа АТФ-связывающего участка Hsp 90 введением препарата 17-AAG (17-(allylamino)-17-demethoxygeldanamycin) при вывешивании животных. Мы проверяли гипотезу о том, что ингибитор Hsp90 17-AAG может увеличить количество белков теплового шока 90 и 70 (Hsp(s)) в m.soleus и предотвратить ее атрофию при функциональной разгрузке. Препарат 17-AAG модулирует функцию Hsp 90 (схема представлена на Фиг.1). Он специфически прикрепляется к АТФ-связывающему сайту Hsp90 и разрывает АТФ-зависимую функцию этого шаперона. Этот сайт, как известно, инициирует неспецифическое связывание фактора HSF-1. В интактных клетках некоторые шапероны (в частности Hsp90) связывают HSF-1 и держат его в неактивной форме. При стрессе (или увеличении физической активности) эти подавляющие шапероны оказываются связанными неправильно свернутыми белками, которые приводят к диссоциации HSF-1 от комплекса с Hsp90, что позволяет HSF-1 транслоцироваться в ядро. Часть ядерной HSF-1 связывается с гранулами (элементами) теплового шока, которые могут изменять хроматиновую структуру (Jolly et al., 2004). Прикрепление HSF-1 к элементу генов, индуцирующих синтез Hsp(s), разблокирует РНК-полимеразу, которая до этого не могла работать в большинстве этих генов.
Материалы, подтверждающие воспроизводимость способа
21 самец крыс Wistar были разделены на 3 группы по 7 животных в каждой. Одна группа служила интактным контролем (группа К, вес крыс 188±6 г, вес m.soleus 76±2 мг), вторую группу подвергали вывешиванию под углом 45° в течение 3-х дней таким образом, чтобы передние лапы опирались на землю, а задние ее не касались (группа В, вес крыс 200±4 г, вес m.soleus 64±3 мг), третьей группе вывешенных по такой же методике крыс вводили препарат 17-AAG, растворенный в DMSO (диметилсульфоксиде). Препарат вводили в концентрации 60 мг/кг веса животного (группа ВА, вес крыс 181±5 г, вес m.soleus 66±2 мг). Дозы введения препарата и его содержание в различных тканях у крыс были исследованы ранее. Новизна работы заключается в том, что до сих пор не был использован подход увеличения концентрации белков теплового шока с использованием препаратов in vivo при функциональной разгрузке мышц. После окончания эксперимента животных забивали сверхдозой нембутала, m.soleus замораживали в жидком азоте, а затем использовали для выявления отдельных белков методом вестерн-блота и выявления мРНК методом ПЦР в реальном времени.
Электрофорез в ПААГ
С каждой пробы m.soleus были сделаны срезы толщиной 20 мкм (10-15 мг) и немедленно прогомогенизированы 3 раза по 10 сек в течение 25 мин в 120 мкл буфера, содержащего RIPA-буфер («Santa Cruz», США), 5 мМ ЭДТА (pH 8.0),1 мМ PMSF, 1 мМ DTT, протеазный ингибиторный коктейль («Santa Cruz»), протеазный ингибиторный коктейль («Amresco», США), фосфатазный ингибиторный коктейль («Santa Cruz»), пепстатин А («SIGMA», США). Затем образцы центрифугировали 10 мин при 12000 g. Пробы разводили в двукратном буфере для образцов (5,4 мМ Tris-HCl (pH=6,8), 4% SDS, 20% глицерин, 10% бета-mercaptoethanol, 0,02% bromphenol blue). Часть супернатанта отбирали для определения концентрации общего белка с помощью RC DC Protein Assay («Bio-Rad Laboratories», США), оставшийся объем использовали для анализа с помощью электрофореза в ПААГ. Все образцы хранили при -84°C. Электрофорез проводили в 10% разделяющем акриламидном геле (0,2% метилбисакриламид, 0,1% SDS, 375 мМ Tris-HCl (pH=8,8); 0,05% APS; 0,1% TEMED) и 5% концентрирующем акриламидном геле (0,2% метилбисакриламид, 0,1% SDS, 125 мМ Tris-HCl (pH=6,8), 0,05% APS; 0,1% TEMED). Образцы загружались из расчета для pFoxo, Foxo, pAkt, конъюгатов убиквитина - 50 мкг, для p70S6k, µ-кальпаина - 30 мкг общего белка в каждой пробе на дорожку. Образцы каждой группы загружались на один гель с контрольными образцами.
Вестернблоттинг
Электроперенос белков производился в буфере (25 мМ Tris рН 8.3, 192 мМ глицин, 20% метанола, 0,02% SDS) на нитроцеллюлозную мембрану при 100В при температуре 4°C в системе mini Trans-Blot («Bio-Rad Laboratories») в течение 2 часов. Мембраны блокировались в растворе 5%-ого сухого молока («Bio-Rad Laboratories») в TBST ночь при температуре 4°C. Для выявления были использованы первичные моноклональные антитела: анти-pFoxo («Santa Cruz», rabbit, разведение 1:700), анти-Foxo («Santa Cruz», rabbit, разведение 1:700), анти-pAkt («Cell Signalling», США, rabbit, разведение 1:400), анти-убиквитин («Abcam», UK, rabbit, разведение 1:500), aнти-P-p70S6k («Santa Cruz», goat, разведение 1:2000), анти-µ-кальпаина («Santa Cruz», mouse, разведение 1:500), анти-GAPDH («Abm», Canada, mouse, разведение 1:5000). Затем блоты были проинкубированы со вторичными антителами goat-anti-mouse («Bio-Rad Laboratories», разведение 1:200 000) и goat-anti-rabbit («ИМТЕК», Россия, разведение 1:5000), конъюгированными с пероксидазой хрена, в 3% бычьем сывороточном альбумине («SIGMA»). Белковые полосы выявляли на пленке, используя Immun-StarTM Substrate Kit («Bio-Rad Laboratories»). Инкубации с антителами проводили 1 ч при комнатной температуре. Блоты отмывали 3 раза по 10 мин в TBST. Вестерн-блоттинг был повторен не менее 3 раз. Все белковые полосы анализируемых проб нормировались к содержанию в них GAPDH. Анализ белковых полос проводился с помощью денситометра GS800 (QuantityOneTM software, «BioRad Laboratories»). Все измерения плотности изображений проводились в линейном диапазоне проявляющего реагента, сканера и рентгеновской пленки.
Анализ экспрессии генов
Для определения уровня мРНК HSP90, HSP70, µ-кальпаинов была выделена тотальная мРНК из 10 мг мышечной ткани. Все образцы РНК были обработаны протеиназой К и ДНКазой I. Концентрация РНК была определена при 260 нм. Затем водный раствор выделенной РНК был заморожен на -84°C для последующего проведения обратной транскрипции. Для обратной транскрипции использовали водный раствор 1 мкг РНК, олиго(dT)15, случайные гексануклеотиды d(N)6, обратную транкскриптазу MMLV. Обратную транскрипцию проводили при 37°C в течение 60 мин согласно стандартному протоколу, с ферментом MMLV RT, ПЦР в реальном времени с использованием интеркалирующего красителя SYBR Green I в амплификаторе iQ5 Multicolor Real-Time PCR Detection System («BioRad Laboratories»). Полученные образцы кДНК были помещены на хранение при -84°C для проведения ПЦР в реальном времени. Праймеры были сконструированы с помощью программ Primer3 и mfold или взяты из литературы. Для проверки качества работы праймеров и полученных после ПЦР ампликонов был проведен горизонтальный элктрофорез в агарозном геле, выделен нужной сегмент ДНК. Секвенирование ДНК проводилось в центре коллективного пользования «Геном», основанного на базе Института молекулярной биологии им. В.А.Энгельгардта РАН с помощью набора реактивов ABI PRISM® BigDye™ Terminator v.3.1 с последующим анализом продуктов реакции на автоматическом секвенаторе ДНК ABI PRISM 3100-Avant. Для анализа полученных данных использовалось относительное количественное определение целевого гена, нормализованное к референсному, метод 2-ΔΔCt (метод Ливака). GAPDH и β-актин были выбраны в качестве референсных генов, экспрессия которых постоянна в камбаловидной мышце в условиях экспериментов [Stevens et al., 1999; Meissner et al., 2001]. Для оценки относительных изменений в уровне экспрессии специфического гена в пробе мы использовали следующую формулу: ΔCt=Ct(реф)-Ct(тест), где Ct(реф) - точка пересечения базовой линии и графика амплификации референсного гена в пробе, а Ct(тест) - точка пересечения базовой линии и графика амплификации изучаемого гена в той же пробе. Далее определяли в каждой группе среднее значение ΔCt и его стандартную ошибку. Изменение уровня экспрессии анализируемого гена в экспериментальных группах оценивали относительно контрольного уровня по формуле: ΔΔCt=ΔCt(группа)-ΔCt(контроль).
Статистическая обработка данных
Все данные приводились как М±m, где М - среднее арифметическое значение; m - стандартная ошибка среднего значения. Статистическая обработка данных производилась с помощью программы Origin Pro v.8.0 SR5. Все выборки были проверены на принадлежность к нормально распределенной совокупности. Достоверность отличий между группами определялась с помощью двухфакторного дисперсионного анализа (ANOVA) с последующим вычислением значения критерия Ньюмена-Кейлса для множественных сравнений.
В эксперименте с введением крысам 17-AAG (17-(allylamino)-17-demethoxygeldanamycin) в течение 3 дней (60 мг/кг веса животного, n=7) мы получили следующие результаты.
Результаты
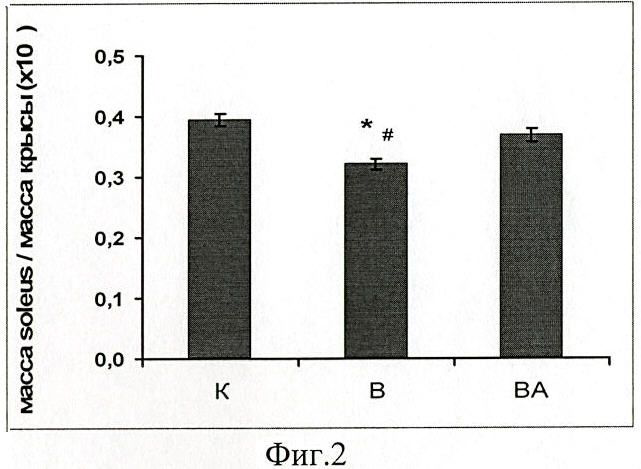
В группе с введением препарата (ВА) атрофии m.soleus не обнаружено, в то время как в группе без введения препарата (В) индекс веса m.soleus/вес крысы был снижен относительно группы контроля (фиг.2).
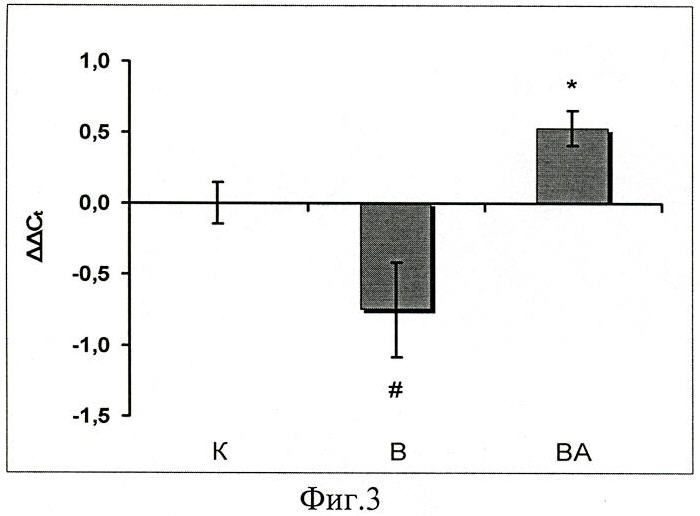
Величина мРНК белков теплового шока 90 (Hsp90) в группе с блокированием работы АТФ-связывающего участка Hsp 90 была даже выше чем в группе контроля (фиг.3), в то время как в группе В она была достоверно снижена.
Количество мРНК Hsp70 в гр. ВА было также выше, чем в группе без введения препарата (фиг.4).
Эти изменения в количестве Hsp(s) свидетельствуют об адекватном воздействии препарата.
Введение 17-AAG (группа ВА) предотвратило увеличение уровня как мРНК, так и белка µ-кальпаина, которое наблюдалось в группе без введения препарата (фиг.5А, Б).
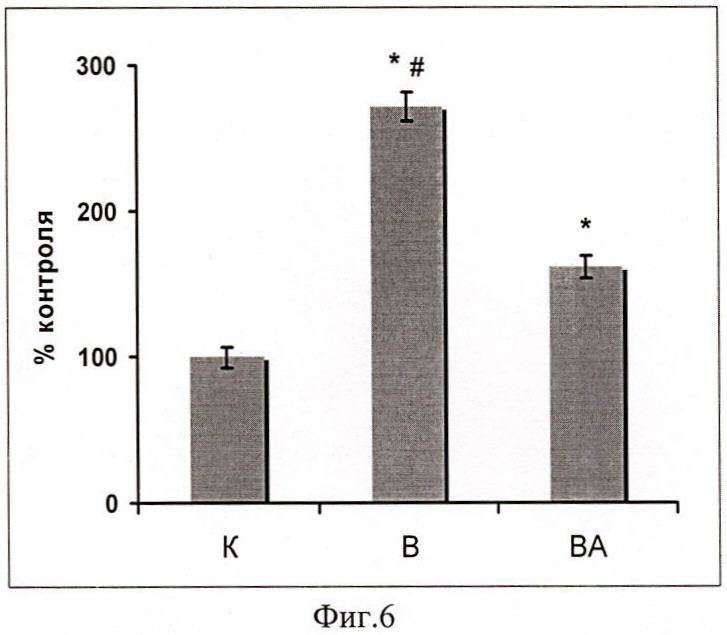
В группе ВА (с введением 17-AAG), уровень убиквитин-конъюгированных белков в m.soleus был существенно ниже по-сравнению с группой В (фиг.6).
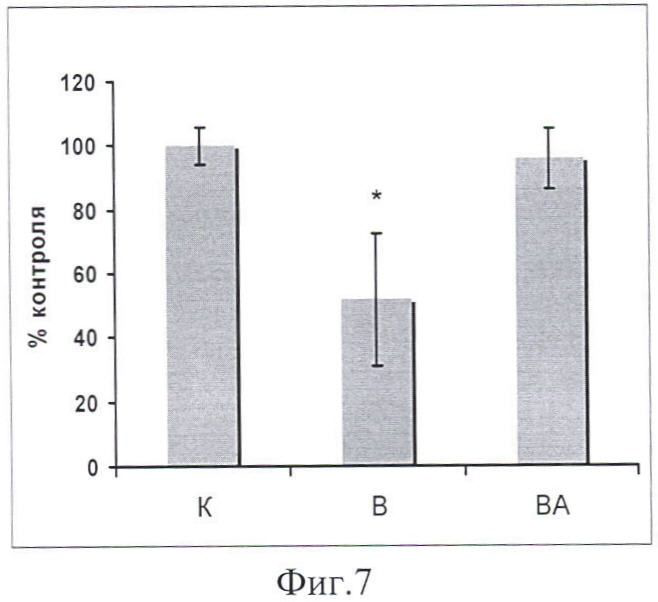
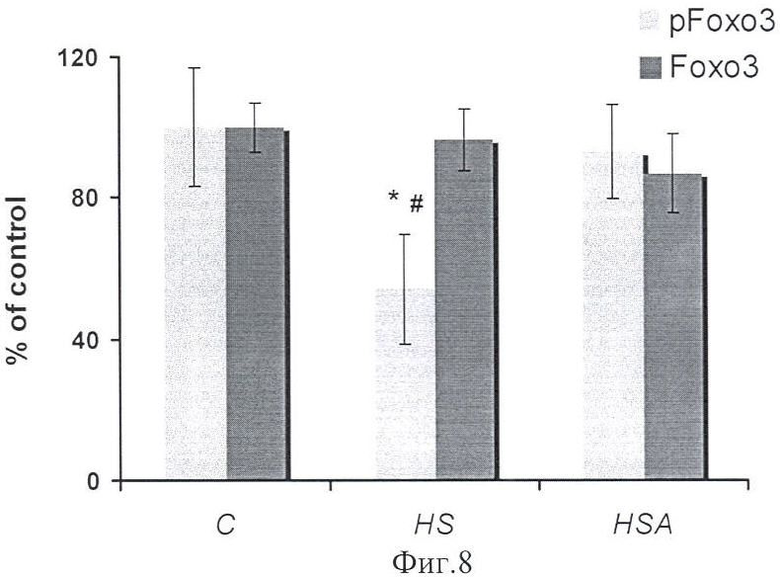
При тестировании звеньев анаболического сигнального пути обнаружено отсутствие снижения белков pAkt и pFOXO3 относительно уровня контроля (в отличие от группы В) (фиг.7, 8).
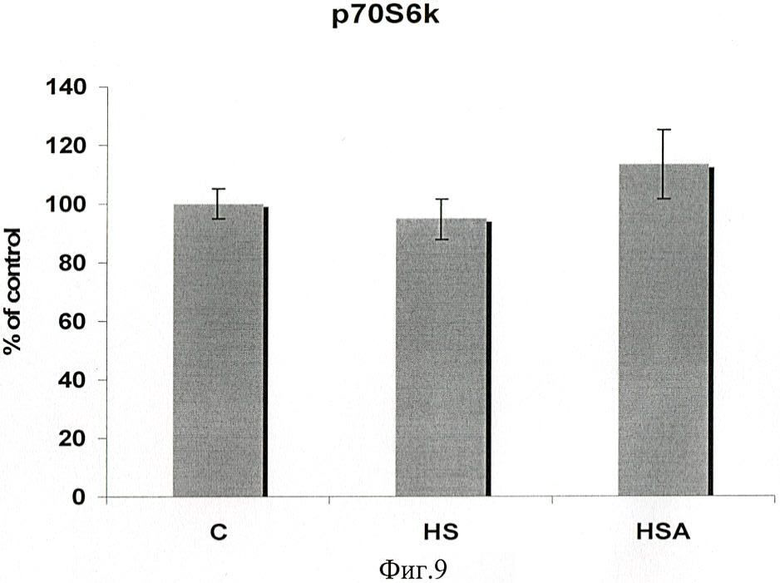
Изменений уровня белков pP70S6k через 3 дня вывешивания мы не заметили ни в одной из групп (фиг.9).
Таким образом, в результате 3-дневного вывешивания крыс мы обнаружили существенное снижение мРНК Hsp90 и 70 относительно группы контроля (фиг 3, 4). Введения 17-AAG во время вывешивания (группа ВА) приводит к гораздо большей экспрессии мРНК Hsp90/70 в soleus крыс, чем в вывешенной группе В. Параллельно увеличению количеству мРНК Hsp(s), атрофия m.soleus была также предотвращена после 3-х дней вывешивания (фиг.2). Раньше было показано только предотвращение атрофии отдельных волокон m.soleus вывешенных крыс, в которые были инкорпорированы плазмиды, содержащие Hsp70 (Senf, S.M. et all., 2008). Нам впервые удалось воздействовать непосредственно на белки теплового шока in vivo при разгрузке для увеличения их экспрессии в мышце. При анализе содержания pAkt в m.soleus мы обнаружили существенное падение его уровня в группе В (фиг.7) и отсутствие его снижения в группе ВА (относительно группы контроля). Т.к. pAkt является звеном анаболического сигнального пути pAkt-mTOR-p70S6k, эти результаты согласуются с данными об отсутствии атрофии в мышце этой группы и повышением уровня Hsp90. В то же время уровень фосфорилированных P70S6k был несущественно выше, чем в группе контроля и В (фиг.9). Следует отметить, что кроме Akt-mTOR-S6K существуют другие факторы, участвующие в регуляции белкового синтеза, например, на уровне трансляции или элонгации.
Мышечная атрофия, вызванная функциональной разгрузкой, является результатом как снижения белкового синтеза, так и увеличения белковой деградации. Но считается, что последний играет более существенную роль (Thomason, D.В., et all, 1989). Белковая деградация первично инициируется активацией убиквитин-протеасомного пути (Jagoe, R.Т., and Goldberg, A.L. (2001)). Полагая, что блокирование АТФ-связывающего участка Hsp90 предотвращает мышечную атрофию, мы проверяли гипотезу о том, что увеличение концентрации Hsp(s) может контролировать главный сигнальный путь, известный регулированием мышечной массы - Foxo. Мы обнаружили падение pFoxo3 только в вывешенной без препарата группе (В) относительно группы контроля, но не в группе ВА. В то же время уровень Рохо3 был одинаков у всех экспериментальных животных (фиг.8). В группе ВА (с введением 17-AAG), уровень убиквитин-конъюгированных белков в m.soleus был существенно ниже по-сравнению с группой В (фиг.6). Стоит еще раз подчеркнуть, что мы определяли только конъюгированный с белком убиквитин. Эти результаты свидетельствуют о снижении разрушения белка в группе ВА. Недавними исследованиями было показано, что Hsp90 очень динамично взаимодействует с различными белками, например NOS (NO-синтазой), которые не являются классическими клиентскими белками Hsp90. Это динамическое соединение клиентского белка (NOS) с Hsp90 защищает его от CHIP-инициированного убиквитинирования (Pratt W.B. et all., 2010). Можно предположить, что развитие событий в разгруженной мышце ведет к открытию специфического лиганда, с которым связывается «клиентский» белок. Это является первым шагом в разворачивании структуры белка. И так долго, как Hsp90 может взаимодействовать с лигандом для стабилизации участка связывания, он будет препятствовать эффекту повреждения белка при разгрузке. Когда связывающий лиганд находится в открытом состоянии, так, что Hsp90 не может больше взаимодействовать с белком, происходит Hsp70-зависимое убиквитинирование белка. Так как уровень мРНК Hsp90 в группе с введением 17-AAG (ВА) превышал не только уровень группы вывешивания (В), но и контрольный (К), вполне может быть, что этот способ защиты белков от разрушения мог иметь место. Мы обнаружили существенно более высокий уровень µ-кальпаина (как белка, так и мРНК) только в группе крыс, вывешенных без введения препарата (В) (параллельно с развитием у них атрофии soleus и увеличением убиквитин-конъюгированных белков) по-сравнению с группами ВА и К (фиг 5). Убиквитирующие кальпаины (µ-кальпаин и m-кальпаин), а также мышечно-специфический кальпаин-3 - кальцийзависимые протеазы, реагирующие на изменение концентрации кальция. Ранее было показано, что увеличение в soleus концентрации кальция при вывешивании крыс может быть стимулом для повышения уровня µ-кальпаина в вывешенной группе В (фиг.5). Однако механизм, который бы мог вести к предотвращению повышения уровня µ-кальпаина в группе ВА, неизвестен. Ранее выдвинули предположение, что активация кальпаинов может ингибировать анаболический сигнальный путь Akt. Это может происходить потому, что молекулярные шапероны могут стимулировать увеличение активности Akt, a Hsp90 в свою очередь являются субстратами кальпаинов. Интересно отметить, что обратный процесс - чтобы ингибирование АТФ-связывающего участка Hsp90 вело к снижению экспрессии µ-кальпаина - ранее показан не был.
Краткое описание рисунков
Фиг.1. Схема модулирования функции Hsp 90 препаратом 17-AAG.
Фиг.2. Отношение массы m.soleus к массе тела животного у вывешенных (В) и вывешенных с введением 17-AAG (ВА) крыс оценивали относительно контрольного (К) уровня. Результаты приведены в виде М±m (n=7 для каждой группы). * - достоверные отличия от К, р<0,05, # - достоверные отличия от ВА, р<0,05.
Фиг.3. Содержание мРНК HSP90 в m.soleus у вывешенных (В) и вывешенных с введением 17-AAG (ВА) крыс оценивали относительно контрольного (К) уровня. Результаты приведены в виде М±m (n=7 для каждой группы). * - достоверные отличия от К, p<0,05, # - достоверные отличия от ВА, p<0,05.
Фиг 4. Содержание мРНК HSP70 в m.soleus у вывешенных (В) и вывешенных с введением 17-AAG (ВА) крыс оценивали относительно контрольного (К) уровня. Результаты приведены в виде М±m (n=7 для каждой группы). * - достоверные отличия от К, р<0,05, # - достоверные отличия от ВА, р<0,05.
Фиг.5. (А,Б) А - содержание мРНК µ-кальпаина; Б - большой и малой субъединиц белка кальпаина.
Фиг.6. Содержание убиквитинированных белков в m.soleus у вывешенных (В) и вывешенных с введением 17-AAG (ВА) крыс оценивали относительно уровня соответствующих белков в контроле (К). Результаты приведены в виде М±m (n=7 для каждой группы). * - достоверные отличия от К, p<0,05; # - достоверные отличия от ВА, p<0,05.
Фиг.7. Содержание pAkt в m. soleus у вывешенных (В) и вывешенных с введением 17-AAG (ВА) крыс оценивали относительно уровня соответствующих белков в контроле (К). Результаты приведены в виде М±m (n=7 для каждой группы). * - достоверные отличия от К, р<0,05.
Фиг.8. Содержание рFОХО3 в m.soleus у вывешенных (В) и вывешенных с введением 17-AAG (ВА) крыс оценивали относительно уровня соответствующих белков в контроле (К). Результаты приведены в виде М±m (n=7 для каждой группы). * - достоверные отличия от К, р<0,05; # - достоверные отличия от ВА, р<0,05.
Фиг.9. Содержание p70S6k в m.soleus у вывешенных (В) и вывешенных с введением 17-AAG (ВА) крыс оценивали относительно уровня соответствующих белков в контроле (К). Результаты приведены в виде М±m (n=7 для каждой группы).
Литература
1. Goto K, Honda M, Kobayashi T, Uehara K, Kojima A, Akema T, Sugiura T, Yamada S, Ohira Y, Yoshioka T. Heat stress facilitates the recovery of atrophied soleus muscle in rat. Jpn J Physiol. 2004 Jun; 54(3):285-293.
2. Sent S.M., Dodd S.L., McClung J.M., Judge A.R. Hsp70 overexpression inhibits NF-kappaB and Foxo3a transcriptional activities and prevents skeletal muscle atrophy // FASEB J., 2008; 22(11): p.3836-3845.
3. Kayani AC, Close GL, Broome CS, Jackson MJ, McArdle A. Enhanced recovery from contraction-induced damage in skeletal muscles of old mice following treatment with the heat shock protein inducer 17-(allylamino)-17-demethoxygeldanamycin. Rejuvenation Res. 2008 Dec; 11(6):1021-1030.
4. Waza M, Adachi H, Katsuno M, Minamiyama M, Tanaka F, Sobue G. Alleviating neurodegeneration by an anticancer agent: an Hsp90 inhibitor (17-AAG). Ann NY Acad Sci. 2006 Nov; 1086: 21-34.
5. Kamal A, Burrows FJ. Hsp90 inhibitors as selective anticancer drugs. Discov Med. 2004, Oct; 4 (23): 277-280.
6. Le Brazidec JY, Kamal A, Busch D, Thao L, Zhang L, Timony G, Grecko R, Trent K, Lough R, Salazar T, Khan S, Burrows F, Boehm MF. Synthesis and biological evaluation of a new class of geldanamycin derivatives as potent inhibitors of Hsp90. J Med Chem. 2004 Jul 15; 47(15): 3865-73.
| название | год | авторы | номер документа |
|---|---|---|---|
| СРЕДСТВО ДЛЯ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517576C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517259C1 |
| ПРИМЕНЕНИЕ VX 745 ДЛЯ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2019 |
|
RU2714923C1 |
| Способ предотвращения атрофии скелетных мышц при их функциональной разгрузке | 2022 |
|
RU2797216C1 |
| Способ предотвращения развития утомления скелетной мышцы | 2023 |
|
RU2814077C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2010 |
|
RU2444354C2 |
| Способ предотвращения атрофии, повышенной утомляемости медленных мышечных волокон и их трансформации в быстрые мышечные волокна в камбаловидной мышце | 2024 |
|
RU2834246C1 |
| ИНГИБИТОР ПРОТЕОЛИЗА ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ФИЗИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2389486C1 |
| СПОСОБ ПРОФИЛАКТИКИ РАЗРУШЕНИЯ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ЭКСЦЕНТРИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2391968C1 |
| ИНДУЦИРОВАНИЕ ГИБЕЛИ КЛЕТОК ПУТЕМ ИНГИБИРОВАНИЯ АДАПТИВНОГО ТЕПЛОВОГО ШОКОВОГО ОТВЕТА | 2009 |
|
RU2474612C2 |



Изобретение относится к медицине, а именно к физиологии и космической медицине. Способ для предотвращения атрофии скелетных мышц при их функциональной разгрузке включает введение 17-(allylamino)-17-demethoxygeldanamycin (17-AAG), который увеличивает в мышцах количество белков теплового шока и дополнительно уменьшает уровень µ-кальпаина. Изобретение позволяет устранить негативные последствия гипокинезии или гравитационной разгрузки с помощью более эффективного ингибирования ключевых сигнальных путей, инициирующих атрофию. 9 ил.
Способ предотвращения атрофии скелетных мышц при их функциональной разгрузке путем воздействия на организм фактора, увеличивающего в мышцах количество белков теплового шока, отличающийся тем, что в качестве фактора, увеличивающего количество белков теплового шока, используют введение 17-(allylamino)-17-demethoxygeldanamycin (17-AAG), который дополнительно уменьшает уровень µ-кальпаина.
| SENF S.M | |||
| et al | |||
| Деревянный торцевой шкив | 1922 |
|
SU70A1 |
| СПОСОБ ПРОФИЛАКТИКИ РАЗРУШЕНИЯ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ЭКСЦЕНТРИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2391968C1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| НОВОСЕЛОВА Т.В | |||
| и др | |||
| Влияние гелданамицина на экспрессию сигнальных белков и белков теплового шока в нормальных лимфоцитов мышей // Цитология | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |

