Область техники
Настоящее изобретение относится к микробиологической, генной инженерии и биотехнологии, в частности к способу получения рекомбинантной лакказы лигнинолитического гриба Trametes sp. Рекомбинантная лакказа может применяться в таких индустриальных процессах, как отбеливание бумажной массы, делигнификация целлюлозной пасты и улучшение свойств бумаги и текстильных материалов, очистка окружающей среды от различных загрязнений, окрашивание материалов, получение альдегидов и кетонов из подходящих субстратов и других.
Описание предшествующего уровня техники
Лакказа (ЕС 1.10.3.2 пара-дифенол: кислород оксидредуктаза) является полифенолоксидазой, принадлежащей к семейству голубых оксидаз. Лакказа катализирует окисление редуцирующего субстрата за счет восстановления молекулярного кислорода до воды. Продукты реакции - свободные радикалы редуцирующего субстрата и вода. Обычными субстратами лакказы являются: о-, м-, п-фенолы с различньпйи заместителями, полифенолы, ароматические амины. Недавно были открыты реакции лакказ с участием медиаторов, - низкомолекулярных соединений, повышающих эффективность лакказы в биодеградации лигнина, - устойчивых соединений различной структуры, включая ксенобиотики и загрязнители окружающей среды.
Известны способы получения лакказ из Coprinus cinereus (патенты США 6242232; 6008029), Myceliophthora thermophila (патенты США 5981243; 5925554; 5795760), Scytalidium thermophilum (патенты США 5925554; 5843745; 5750388), Rhizoctonia solani (патент США 5480801), Pycnoporus cinnabarinus (патент Франции FR 2865216) Phlebia (заявка РСТ WO 9201046) с использованием штаммов-продуцентов различных видов и родов.
Описан способ получения рекомбинантной лакказы гриба Polyporus (Trametes) villosa с использованием грибов Aspergillus и Fusarium (патент США 5770418).
Т. versicolor секретирует несколько изоформ лакказы, которые, возможно, призваны выполнять у этого базидиомицета различные функции (Reinhammar В. Copper Proteins and Copper Enzymes, V.3 / Lonti R., ed. CRC Press, Boca Raton, FL. 1-35, (1984)). Наличие у продуцента нескольких изоформ фермента затрудняет очистку индивидуального изофермента.
К настоящему времени описаны факты экспрессии лакказы из Trametes trogii в Pichia pastoris (Colao М.С. et al., Microbial Cell Factories, v.5, №1, p.31, (2006)); лакказы из Trametes versicolor в Pichia pastoris (Jonsson L.J. et al., Current Genetics, 32(6), 425-430 (1997)); лакказы из Trametes sanguinea в дрожжах (Hoshida H. et al., Journal of biosciences and bioenGln,eering, 92(4), 372-80 (2001)) и лакказы из Trametes versicolor в Saccharomyces cerevisiae (Cassland P. et al., Applied Microbiology and Biotechnology, 52(3), 393-400 (1999)).
Основные физико-химические характеристики мажорной изоформы лакказы, секретируемой грибом Trametes hirsuta 072, были недавно описаны, ген этой изоформы был клонирован, и его нуклеотидная последовательность была определена (Ребриков, Д.Н. и др. Прикладная биохимия и микробиология, 42, 6, 645-653 (2006)).
Однако в настоящее время нет сообщений, описывающих получение индивидуальной рекомбинантной лакказы С1 лигнинолитического гриба Trametes hirsuta, обладающей высокой каталитической активностью, с использованием штаммов мицелиальных грибов, например, таких как Penicillium canescens, позволяющих производить высокую гетерологичную экспрессию лигнинолитических ферментов в активной форме.
Краткое описание изобретения
Целями настоящего изобретения являются создание штамма мицелиального гриба - продуцента индивидуальной изоформы фермента лакказы, в частности лакказы С1 лигнинолитического гриба Trametes hirsuta, способного к эффективной секреции целевого белка в культуральную жидкость, и предоставление способа получения индивидуальной рекомбинантной лакказы С1. Поставленная задача была решена путем конструирования рекомбинантной плазмиды pBGlac, содержащей ген, кодирующий лакказу С1 лигнинолитического гриба Trametes hirsuta, и штамма Penicillium canescens PCA-10(niaD-)/BGlac, обеспечивающей синтез и продукцию в секретируемой растворимой форме лакказы С1 лигнинолитического гриба Trametes hirsuta.
Целью настоящего изобретения является предоставление экспрессионой кассеты, содержащей фрагмент ДНК, кодирующий лакказу С1 лигнинолитического гриба Trametes hirsuta (SEQ ID NO:1) или ее вариант.
Также целью настоящего изобретения является предоставление рекомбинантной плазмиды, содержащей фрагмент ДНК, кодирующий лакказу С1 лигнинолитического гриба Trametes hirsuta (SEQ ID NO:1) или ее вариант под контролем промотора, функционирующего в эукариотической клетке.
Также целью настоящего изобретения является предоставление рекомбинантной плазмиды, описанной выше, где в качестве плазмиды используют плазмиду pBGlac.
Также целью настоящего изобретения является предоставление эукариотической клетки, трансформированной описанной выше плазмидой, - продуцента лакказы С1 из лигнинолитического гриба Trametes hirsuta.
Также целью настоящего изобретения является предоставление описанной выше эукариотической клетки, при этом в качестве эукариотической клетки используют клетки мицелиального гриба Penicillium canescens.
Также целью настоящего изобретения является предоставление штамма Penicillium canescens PCA-10(niaD-)/BGlac - продуцента лакказы С1 лигнинолитического гриба Trametes hirsuta.
Также целью настоящего изобретения является предоставление способа получения рекомбинантной лакказы С1 лигнинолитического гриба Trametes hirsuta, включающий стадии выращивания описанной выше эукариотической клетки в питательной среде и выделения рекомбинантной лакказы С1 из культуральной жидкости.
Также целью настоящего изобретения является предоставление описанного выше способа, отличающегося тем, что в качестве эукариотической клетки используют клетки мицелиального гриба.
Также целью настоящего изобретения является предоставление описанного выше способа, отличающегося тем, что в качестве клеток мицелиального гриба используют штамм Penicillium canescens PCA-10(niaD-)/BGlac.
Также целью настоящего изобретения является предоставление рекомбинантной лакказы С1 лигнинолитического гриба Trametes hirsuta, полученной описанным выше способом.
Краткое описание чертежей
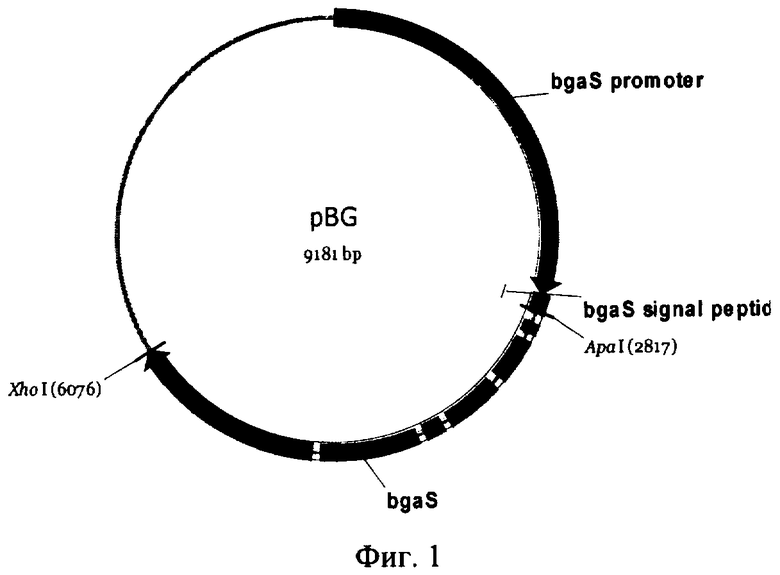
На Фиг.1 изображены схема векторной плазмиды pBG, содержащей регуляторные элементы гена β-галактозидазы Penicillium canescens.
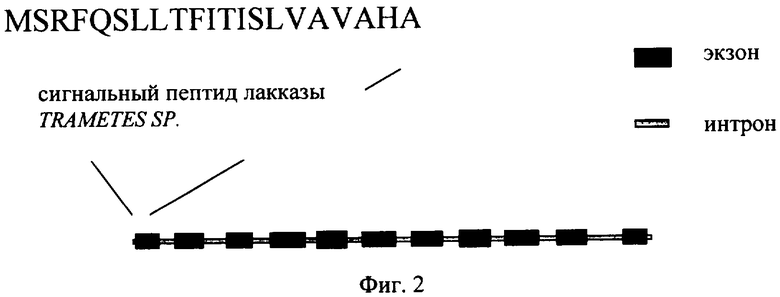
На Фиг.2 изображена структурная часть гена лакказы С1 Trametes sp.,
включающая его собственный стартовый кодон АТГ, сигнальный пептид, внутреннюю часть с нитронами и стоп-кодон на 3′-конце.
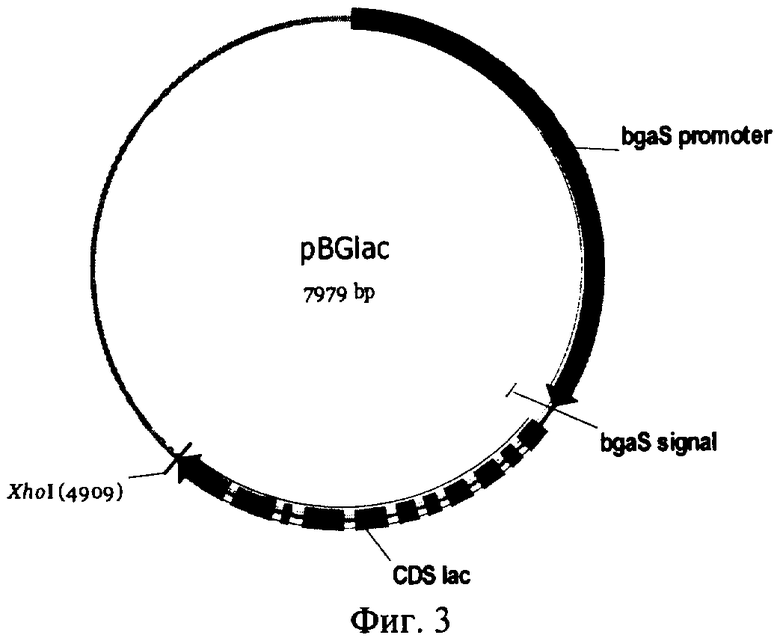
На Фиг.3 изображена схема рекомбинантной плазмиды pBGlac, использованной для создания штамма-продуцента лакказы С1 Trametes sp.
Подробное описание настоящего изобретения
Для реализации настоящего изобретения главной технической задачей явилось создание штамма мицелиального гриба - продуцента рекомбинантной лакказы С1 лигнинолитического гриба Trametes hirsuta, способного к эффективной продукции и секреции активного целевого белка в культуральную жидкость. Поставленная задача была решена путем конструирования рекомбинантной плазмиды pBGlac, содержащей ген, кодирующий синтез лакказы С1 лигнинолитического гриба Trametes hirsuta, и штамма Penicillium canescens PCA-10(niaD-)/BGlac, обеспечивающего синтез и продукцию в секретируемой растворимой форме лакказы С1 лигнинолитического гриба Trametes hirsuta.
Термин «экспрессионная кассета» означает фрагмент ДНК, содержащий все необходимые генетические элементы для экспрессии внедренного в нее гена, например, такие как промотор, терминатор. Конкретными примерами генетических элементов, необходимых для экспрессии рекомбинантной лакказы в составе экспрессионной кассеты согласно настоящему изобретению, являются, но не ограничиваются ими, промотор и терминатор β-галактозидазы Penicillium canescens.
Фрагментом ДНК, кодирующим лакказу С1 лигнинолитического гриба Trametes hirsuta согласно настоящему изобретению, является, например, ген мажорной лакказы, описанный в статье Ребрикова Д.Н. и др. (Прикладная биохимия и микробиология, 42, 6, 645-653 (2006)). Структура гена, кодирующего лакказу С1 лигнинолитического гриба Trametes hirsuta, представлена на Фиг.2. Последовательность гена лакказы С1 лигнинолитического гриба Trametes hirsuta представлена в Перечне последовательностей под номером SEQ ID NO:1. Аминокислотная последовательность лакказы С1 представлена в Перечне последовательностей под номером SEQ ID NO:2.
Термин «вариант белка» в значении, в котором он используется в настоящем изобретении, означает белок, который имеет изменения в аминокислотной последовательности, а именно делеции, вставки, добавления или замены аминокислот, при условии, что при этом сохраняется необходимый уровень активности белка, например как минимум 10% от активности нативной лакказы. Ряд изменений в варианте белка зависит от положения или от типа аминокислотного остатка в трехмерной структуре белка. Количество изменений может составлять от 1-го до 30-ти, предпочтительно от 1-го до 15-ти и наиболее предпочтительно от 1-го до 5-ти изменений в последовательности белка под номером 2 (SEQ ID NO:2). Эти изменения могут иметь место в областях белка, которые не являются критичными для его функции. Это становится возможным благодаря тому, что аминокислоты обладают высокой гомологией друг с другом, и поэтому третичная структура или активность белка не нарушаются при таком изменении. Поэтому в качестве варианта белка может выступать белок, который имеет гомологию не менее чем 70%, предпочтительно не менее чем 80%, более предпочтительно не менее чем 90% и наиболее предпочтительно не менее чем 95% по отношению к полной аминокислотной последовательности, представленной на SEQ ID NO:2, при условии, что активность белка сохраняется. Гомология между аминокислотными последовательностями может быть установлена с использованием хорошо известных методов, например с помощью выравнивания последовательностей в компьютерной программе BLAST 2.0, которая вычисляет три параметра: счет, идентичность и сходство.
Замена, делеция, вставка, добавление или замена одного или нескольких аминокислотных остатков будут представлять собой консервативную мутацию или консервативные мутации при условии, что активность белка при этом сохраняется. Примером консервативной мутации(ий) является консервативная замена(ы). Примеры консервативных замен включают замену Ala на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или Ala, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Tyr, замену Ile на Leu, Met, Val или Phe, замену Leu на Ile, Met, Val или Phe, замену Lys на Asn, Glu, Gln, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Trp, Tyr, Met, Ile или Leu, замену Ser на Thr или Ala, замену Thr на Ser или Ala, замену Trp на Phe или Tyr, замену Tyr на His, Phe или Trp и замену Val на Met, Ile или Leu.
Фрагменты ДНК, которые кодируют по существу тот же функциональный белок, могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК (SEQ ID NO:1), кодирующего лакказу, например посредством метода сайт-направленного мутагенеза, так что один или несколько аминокислотных остатков в определенном сайте будут делетированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации.
Фрагменты ДНК, которые кодируют по существу тот же функциональный белок лакказы, могут быть получены путем экспрессии фрагментов ДНК, имеющих мутацию, описанную выше, в соответствующей клетке, и установления активности экспрессируемого продукта. Фрагменты ДНК, которые кодируют по существу тот же функциональный белок лакказы, также могут быть получены путем выделения фрагментов ДНК, которые гибридизуются с зондами, имеющими нуклеотидную последовательность, которая содержит, например, нуклеотидную последовательность, приведенную в Перечне последовательностей под номером SEQ ID NO:1, в жестких условиях, и кодирует белок, обладающие активностью лакказы. "Жесткие условия", в том значении, которое приписывается этому выражению в рамках настоящего изобретения, означает такие условия, при которых так называемые специфические гибриды образуются, а неспецифические гибриды не образуются. Например, демонстрацией жестких условий могут быть такие условия, при которых фрагменты ДНК, имеющие высокую гомологию, например фрагменты ДНК, имеющие гомологию не менее чем 50%, предпочтительней не менее чем 60%, более предпочтительней не менее чем 70%, еще более предпочтительней не менее чем 80%, еще более предпочтительней не менее чем 90% и наиболее предпочтительней не менее чем 95% способны гибридизоваться друг с другом, а фрагменты ДНК, имеющие гомологию более низкую, чем описано выше, не способны гибридизоваться друг с другом. Кроме того, демонстрацией жестких условий могут служить такие условия, при которых фрагменты ДНК гибридизуются при концентрации соли, эквивалентной условиям однократной отмывки при гибридизации по Саузерну, которые составляют 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS, при 60°С. Продолжительность отмывки зависит от типа мембраны, используемой для блотинга и, как правило, рекомендуется производителем набора. Например, рекомендуемая продолжительность отмывки для мембраны Hybond™ N+ nylon (Amersham, США) в жестких условиях составляет приблизительно 15 минут. Желательно отмывку повторять 2-3 раза. В качестве зондов может быть также использована неполная нуклеотидная последовательность, приведенная в Перечне последовательностей под номерами SEQ ID NO:1. Зонды могут быть приготовлены с помощью ПЦР с использованием праймеров, синтезированных на основе нуклеотидной последовательности, приведенной в Перечне последовательностей под номером SEQ ID NO:1, и на основе фрагментов ДНК, содержащих нуклеотидную последовательность, приведенную в Перечне последовательностей под номером SEQ ID NO:1 в качестве матрицы. В тех случаях, когда фрагмент ДНК, имеющий длину около 300 п.о., используется в качестве зонда, условия отмывки в процессе гибридизации включают, например, 50°С, 2×SSC и 0.1% SDS.
Замена, делеция, вставка или добавление нуклеотидов, так, как они описаны выше, также включает мутации, которые имеют место в природе (мутант или вариант), например обусловленные видовой изменчивостью.
Показатели функциональной активности, при которой считается, что полученный белок обладает свойствами лакказы, определяются по его способности окислять различные субстраты фенольной природы, такие, например, как о-, м-, п-фенолы с различными заместителями, полифенолы, ароматические амины. Также лакказы способны повышать эффективность биодеградации лигнина и устойчивых ароматических ксенобиотиков в присутствии медиаторов. Так, например, активность лакказы можно детектировать спектрофотометрически при 410 нм, используя в качестве хромогенного субстрата 10 мМ раствор пирокатехина в 0,1 М Na-ацетатном буфере, рН 4,9, при этом за "условную единицу активности" принимают увеличение оптической плотности в 1 мл реакционной смеси за 1 мин, пересчитанной на 1 мг вносимого белка (Ребриков Д.Н. и др. Прикладная биохимия и микробиология, 42, 6, 645-653 (2006)).
Активность лакказы также можно определить по измерению начальных скоростей ферментативной реакции с помощью кислородного электрода Кларка в ячейке объемом 0,7 мл при 25°С и постоянном перемешивании. В качестве органического субстрата используют 10 мМ раствор пирокатехина в 0,015 М фосфатном буфере, рН 4,9. Концентрацию кислорода в растворе принимают равной 260 мкМ (согласно значению коэффициента Генри 773 атм/моль × кг воды). Активность лакказы выражают в мкмолях поглощенного кислорода на 1 мг белка за 1 мин.
Считается, что вариант белка обладает свойствами лакказы при условии, что активность указанного варианта составляет не ниже 10% активности нативной лакказы.
Рекомбинантная плазмида согласно настоящему изобретению содержит фрагмент ДНК, кодирующий лакказу С1 лигнинолитического гриба Trametes hirsuta (SEQ ID NO:1) или ее вариант, под контролем промотора, функционирующего в эукариотической клетке.
В качестве рекомбинантной плазмиды согласно настоящему изобретению может быть использована любая бактертриальная плазмида, такая, например, как pBGlac, но список таких плазмид не ограничивается ею.
Рекомбинантная плазмида pBGlac была сконструирована путем клонирования гена, кодирующего лакказу С1, в плазмиду pBG (), содержащую последовательность промотора, сигнального пептида, и терминатора гена β-галактозидазы Penicillium canescens, вместо структурной части гена β-галактозидазы.
Рекомбинантная плазмида pBGlac содержит ген, кодирующий лакказу С1, в составе экспрессионной кассеты под контролем регулируемого промотора гена β-галактозидазы Penicillium canescens, обеспечивающего высокий уровень транскрипции целевого гена, и терминатора гена β-галактозидазы Penicillium canescens. В структуру экспрессионной кассеты входят нуклеотидная последовательность, содержащая промотор гена β-галактозидазы, последовательность сигнального пептида гена β-галактозидазы Penicillium canescens; последовательность ДНК, кодирующая белок, обладающий активностью лакказы, или его вариант; и область терминации транскрипции гена β-галактозидазы Penicillium canescens. При введении данной плазмиды в клетку достигается высокий уровень транскрипции гена, кодирующего лакказу, благодаря использованию высокоэффективного промотора гена β-галактозидазы.
При помощи созданной плазмиды можно трансформировать любую эукариотическую клетку, восприимчивую к подобной трансформации указанной плазмидой. Выбор клетки не является критическим, поскольку методология и приемы трансформации хорошо известны специалисту в данной области техники. И хотя в зависимости от вида клетки и условий культивирования полученного трансформанта уровень экспрессии лакказы может варьироваться, факт экспрессии целевого белка будет иметь место при условии успешной трансформации клетки-реципиента.
«Трансформация клетки плазмидой» означает введение плазмиды в клетку с помощью методов, хорошо известных специалисту в данной области техники. Трансформация этой плазмидой приводит к усилению экспрессии гена, кодирующего белок согласно настоящему изобретению, и к увеличению активности белка в бактериальной клетке. Методы трансформации включают любые стандартные методы, известные специалисту в данной области техники, например метод, описанный в Примере 3.
Трансформация клетки также может быть осуществлена линейным фрагментом ДНК, содержащим ген, кодирующий лакказу С1 лигнинолитического гриба Trametes hirsuta (SEQ ID NO:1) или ее вариант, под контролем промотора, функционирующего в такой эукариотической клетке.
Согласно настоящему изобретению «эукариотическая клетка - продуцент лакказы» означает эукариотическую клетку, обладающую способностью к продукции и выделению лакказы согласно настоящему изобретению в питательную среду, когда эукариотическая клетка согласно настоящему изобретению выращивается в указанной питательной среде. Используемый здесь термин «эукариотическая клетка - продуцент лакказы» также означает клетку, которая способна к продукции лакказы и вызывает накопление лакказы в питательной среде в больших количествах, по сравнению с природным или родительским штаммом, и предпочтительно означает, что указанная клетка способна накапливать в среде лакказу в количестве не менее чем 0,1 условной единицы активности в 1 мл реакционной смеси (определение "условной единицы активности" с использованием хромогенного субстрата пирокатехина дано выше).
Предпочтительно использование клеток мицелиальных грибов в качестве реципиентов для трансформации рекомбинантной плазмидой, содержащей фрагмент ДНК, кодирующий лакказу.
Примером штамма-реципиента для получения продуцента рекомбинантной лакказы согласно настоящему изобретению является, но не ограничивается им, штамм мицелиальных грибов Penicillium canescens PCA-10(niaD-) (Aleksenko A.Y. et al., Curr. Genet., 28, 474-478 (1995)).
Способ получения рекомбинантной лакказы C1 лигнинолитического гриба Trametes hirsuta включает стадии выращивания эукариотических клеток, трансформированных плазмидой согласно настоящему изобретению в питательной среде, подходящей для выращивания указанньк эукариотических клеток и выделения рекомбинантной лакказы С1 из культуральной жидкости.
Использование указанного выше способа позволяет проводить синтез и продукцию в секретируемой растворимой форме лакказы С1 лигнинолитического гриба Trametes hirsuta с активностью не менее 0,1 условной единицы активности в 1 мл реакционной смеси.
Рекомбинантная лакказа С1 лигнинолитического гриба Trametes hirsuta 072, полученная способом согласно настоящему изобретению, обладает следующими характеристиками: молекулярная масса - 70±2 кДа, изоэлектрическая точка - 4,0, содержание углеводов - 12%, состав углеводной цепи - манноза и N-ацетилглюкозамид, количество ионов меди - 4, рН-оптимум - 4,5-5,0, температурный оптимум 55±3°С.
Примеры
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение примеры.
Пример 1. Клонирование гена лакказы С1 лигнинолитического гриба Trametes hirsuta.
Препараты тотальной ДНК Trametes sp.получали, как описано ранее (Ребриков Д.Н. и др. Прикладная биохимия и микробиология, 42, 6, 645-653 (2006)). Фрагмент ДНК с геном лакказы амплифицировали с использованием ДНК Thrametes sp. 072 в качестве матрицы и праймеров Ecolac (SEQ ID NO:3) и Xholac (SEQ ID NO:4), комплементарных последовательностям 5′- и 3′-концов гена лакказы и содержащих сайты узнавания рестриктазы Eco52I и XhoI соответственно. Амплификацию проводили методом ПЦР в амплификаторе Gradient Palm-Cycler в специально подобранных условиях. Реакционная смесь содержала 5 мкл образца хромосомной ДНК, 1-кратный реакционный буфер Pwo DNA Polymerase (Roche), 200 мкМ дНТФ (дезоксинуклеотидтрифосфатов), 0,3 мкМ праймеров, общий объем смеси - 50 мкл. Температурный режим: денатурация 95°С - 5 мин; 95°С - 20 сек; 56°С - 20 сек; 72°С - 2 мин, в течение 30 циклов; достройка 72°С - 5 мин. Продукты реакции разделяли с помощью гель-электрофореза в агарозе. Фрагмент ДНК вьщеляли из агарозы и очищали с использованием набора Wizard SV Gel and PGR Clean-Up System.
Пример 2. Конструирование рекомбинантной плазмиды pBGlac.
Клонирование фрагмента в векторную плазмиду pBG производили по сайтам ApaI и XhoI. Правильность вставки подтверждали секвенированием полученных плазмидных конструкций. Нуклеотидную последовательность определяли с помощью автоматического секвенатора Beckman - Coulter CEQ-2000 в соответствии с протоколом, рекомендованным производителем. Вьщеление и очистку плазмидной ДНК проводили с использованием набора Wizard Plus Minipreps DNA Purification System. Процедуры клонирования выполняли по стандартным методикам (Sambrook J., Fritsch Е., Maniatis Т. Molecular cloning: a laboratory manual. New York, Cold Spring Harbor Lab Press, 1989).
Полученная в результате плазмида была названа pBGlac.
Пример 3. Получение штаммов-продуцентов лакказы С1 лигнинолитического гриба Trametes hirsuta.
Для трансформации мицелиальных грибов использовали штамм Р. canescens РСА-10(niaD-) (Aleksenko A.Y., et al., Curr. Genet., 28, 474-478 (1995)).
P. canescens PC A-10 (niaD-) выращивали 16 часов при 30°C на полной питательной среде: пептон ферментативный - 3 г/л, дрожжевой экстракт - 2 г/л, глюкоза - 10 г/л, NH4Cl - 10 mM, 50х раствор минеральных солей - 20 мл/л. Раствор минеральных солей (50х) имел состав: KCl - 26 г/л, MgSO4×7Н2О - 26 г/л, KH2PO4 - 76 г/л, микроэлементы - 50 мл/л. 50х раствор минеральных солей включал в себя CuSO4×5Н2О - 400 мг/л, FeSO4×5Н2О - 800 мг/л, MgSO4×2Н2О - 800 мг/л, Na2MoO4×2Н2О - 800 мг/л, ZnSO4×7Н2О - 800 мг/ л, B4Na2O7 - 40 мг/л. Переносили мицелий в раствор 1.2 М MgSO4 и 10 мМ NaH2PO4, рН 5.8 и добавляли лизирующий фермент Т. harzianum (5 мг/мл). Протопластирование проводили в течение 2 часов при 30°С в условиях перемешивания. Суспензию переносили в центрифужную пробирку и наслаивали 1-2 см раствора: 0.6 М сорбитол, 10 mM CaCl, 10 mM Трис-HCl. После центрифугирования при 3000 об/мин и 4°С в течение 10 мин отбирали интерфазу, содержащую протопласты. Протопласты промывали 2 раза в стабилизирующем растворе SCT (1.2 М сорбитола, 10 мМ Трис, рН 7.5, 10 мМ CaCl2) и ресуспендировали в нем же до концентрации 108 протопластов/мл. Трансформацию проводили следующим образом: к 200 мкл суспензии протопластов добавляли 10 мкг трансформируемой ДНК pBGlac, инкубировали в ледяной бане 20 мин, после чего проводили осмотический шок в течение 5 мин в РСТ (50% полиэтиленгликоль, 10 мМ CaCl2, 10 мМ Трис-HCl) и высевали протопласты в верхнем слое на агаризованную селективную минимальную среду, содержащую 1.2 М сорбитол. Для селекции рекомбинантных клонов применялась система котрансформации по гену niaD, контролирующему синтез нитратредуктазы (Aleksenko A.Y., et al., Curr. Genet., 28, 474-478 (1995)). В качестве селективного маркера выступала способность утилизировать нитраты. В качестве реципиентов были использованы штаммы, дефектные по нитратредуктазе, неспособные расти на средах с добавлением нитратов. Для трансформации niaD-признака использовался ген нитратредуктазы А. niger, клонированный в плазмиде pSTA10 (23) (Unkles et al., Gene, 78, 157-166 (1989)). Протопласты трансформировали смесью ДНК плазмиды pSTA10 и ДНК плазмиды pBGlac с геном лакказы Thrametes sp. при их соотношении 1:10. Так как маркерный ген и ген лакказы не объединены в общей плазмиде, а добавлялись по-отдельности, то для образования трансформанта по гену лакказы требовалась котрансформация, при которой протопласт должен получить обе плазмиды. Котрансформация является событием более редким, чем трансфомация единичной плазмидой. Около 40% котрансформантов, способных расти на среде, содержащей нитраты, также содержали плазмиду pBGlac с геном лакказы, что обнаруживалось по наличию активности лакказы в культуральной жидкости.
Таким образом были отобраны трансформанты: 21 клон Р. canescens, названные Р. canescens PCA-10(niaD-)/BGlac.
Для отбора клонов, продуцирующих лакказу, проводилось их ферментационное культивирование и тестирование культуральных жидкостей на наличие соответствующей ферментативной активности.
Пример 4. Определение продуктивности штаммов-продуцентов лакказы.
Ферментационное культивирование отобранных клонов проводили согласно следующим протоколам. Штамм Р. canescens PCA-10(niaD-)/BGlac для получения посевного материала выращивали на агаризованной среде при 30°С в течение 5-7 суток. Водной суспензией конидий инокулировали 100 мл среды (конечная концентрация - 4×10-5 конидий/мл). Культивирование проводили в ферментационной среде на орбитальной качалке при 240-250 об/мин в качалочных колбах на 750 мл в течение 140 часов. По окончании ферментации культуральную жидкость отделяли от мицелия и твердых остатков среды центрифугированием (10000 g, 10 мин). В качестве ферментационной среды для Р. canescens использована среда, имеющая следующий состав: свекловичный жом - 30 г/л, пептон - 50 г/л, KH2PO4 - 25 г/л. Агаризованая среда имела следующий состав: 50х раствор минеральных солей - 20 мл/л, глюкоза - 10 г/л, NaNO3 1 М - 10 мл/л, агар-бакто - 20 г/л. Раствор минеральных солей (50х) имел состав: KCl - 26 г/л, MgSO4×7Н2О - 26 г/л, KH2PO4 - 76 г/л, микроэлементы - 50 мл/л. 50х раствор минеральных солей включал в себя CuSO4×5Н2О - 400 мг/л, FeSO4×5Н2О - 800 мг/л, MgSO4×2Н2О - 800 мг/л, Na2MoO4×2Н2О - 800 мг/л, ZnSO4×7Н2О - 800 мг/ л, B4Na2O7 - 40 мг/л.
Наиболее продуктивный клон Р. canescens PCA-10(niaD-)/BGlac продуцировал лакказу с общей активностью 3,4 условных единиц активности в мл. Эффективная экспрессия лакказы наблюдалась при культивировании штамма-продуцента на среде, содержащей в качестве углеводного компонента свекловичный жом.
Пример 5. Определение биологической активности рекомбинантной лакказы.
Активность лакказы в культуральной жидкости штаммов детектировали спектрофотометрически (длина волны 410 нм), используя в качестве хромогенного субстрата 10 мМ раствор пирокатехина (ПКХ) в 0,1 М Na-ацетатном буфере рН 4,9. Пирокатехин был предварительно очищен возгонкой в вакууме. Культуральную жидкость штамма Р. canescens PCA-10(niaD-) использовали в качестве отрицательного контроля. Результаты измерения активности лакказы в культуральных жидкостях, полученных при культивировании штамма-продуцента Р. canescens PCA-10(niaD-)/BGlac при разных температурах, приведены в Таблице 1.
Известно, что при экспрессии генов в гетерологичных системах часто не происходит корректный сплайсинг интронов. Однако в данном случае наличие активной лакказы в культуральных жидкостях отобранных рекомбинантных штаммов свидетельствует о правильном процессинге мРНК, транскрибируемой с гена лакказы в штаммах Р. canescens.







Изобретение относится к биотехнологии и представляет собой клетку мицелиального гриба, принадлежащего к роду Penicillium, трансформированную плазмидой. Плазмида содержит фрагмент ДНК, кодирующий лакказу С1 лигнинолитического гриба Trametes hirsuta или фрагмент ДНК, который гибридизуется с SEQ ID NO:1 в жестких условиях, под контролем промотора, функционирующего в указанной клетке. Полученная клетка является продуцентом лакказы С1. Изобретение относится также к способу получения лакказы с использованием указанной клетки. Изобретение позволяет эффективно получать лакказу с высокой каталитической активностью. 3 н. и 3 з.п. ф-лы, 1 табл., 3 ил.
1. Клетка мицелиального гриба, принадлежащего к роду Penicillium, трансформированная плазмидой, содержащей фрагмент ДНК, кодирующий лакказу С1 лигнинолитического гриба Trametes hirsuta (SEQ ID NO:1), или фрагмент ДНК, который гибридизуется с SEQ ID NO:1 в жестких условиях, под контролем промотора, функционирующего в указанной клетке, - продуцент лакказы С1 из лигнинолитического гриба Trametes hirsuta.
2. Клетка по п.1, отличающаяся тем, что в качестве указанной клетки используют клетки мицелиального гриба Penicillium canescens.
3. Штамм Penicillium canescens PCA-10(niaD-)/BGlac - продуцент лакказы С1 лигнинолитического гриба Trametes hirsuta.
4. Способ получения рекомбинантной лакказы С1 лигнинолитического гриба Trametes hirsuta, включающий стадии выращивания клеток мицелиального гриба, принадлежащего к роду Penicillium, по п.1 в питательной среде и выделения рекомбинантной лакказы С1 из культуральной жидкости.
5. Способ по п.4, отличающийся тем, что в качестве указанных клеток используют клетки мицелиального гриба Penicillium canescens.
6. Способ по п.5, отличающийся тем, что в качестве клеток мицелиального гриба используют клетки штамма Penicillium canescens РСА-10 (niaD-)/BGlac.
| РЕБРИКОВ Д.Н | |||
| и др | |||
| Лакказа лигнолитического гриба Trametes hirsute: очистка и характеристика фермента, клонирование и первичная структура гена, Прикладная биохимия и микробиология, 2006, т.42, №6, с.645-653 | |||
| ФРАГМЕНТ ДНК МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM, КОДИРУЮЩИЙ СИНТЕЗ СЕКРЕТИРУЕМОЙ ЭНДОГЛЮКАНАЗЫ III И ГРУППА ШТАММОВ PENICILLIUM CANESCENS, СИНТЕЗИРУЮЩИХ ЭНДОГЛЮКАНАЗУ III PENICILLIUM VERRUCULOSUM, СКОНСТРУИРОВАННЫХ МЕТОДАМИ ТРАНСФОРМАЦИИ И ГЕНЕТИЧЕСКОЙ ИНЖЕНЕРИИ НА ОСНОВЕ ЭТОГО ФРАГМЕНТА ДНК | 2001 |
|
RU2238974C2 |
| US 5925345, 20.07.1999. | |||

