Область техники, к которой относится изобретение
Изобретение относится к способам получения ферментных препаратов, применяемых в биотехнологии, в данном случае к получению новой термостабильной лакказы со щелочным оптимумом активности. Клонирование фрагмента ДНК бактерии Streptomyces griseoflavus Ac-993, кодирующего двухдоменную лакказу, в составе экспрессионных векторов и введение таких рекомбинантных ДНК в клетки, например бактерий, дрожжей, грибов, насекомых или млекопитающих, позволяет проводить наработку, выделение, очистку рекомбинантной двухдоменной лакказы, которая может быть использована в текстильной промышленности, для нужд химического синтеза, трансформации ксенобиотиков, в целлюлозно-бумажной промышленности, пищевой промышленности, а также в научно-исследовательских целях.
Уровень техники.
Лакказа (ЕС 1.10.3.2, n-дифенол - кислород оксидоредуктаза) является ферментом, катализирующим окисление различных соединений кислородом, который в ходе реакции восстанавливается до воды. В качестве субстратов лакказы могут выступать разные вещества: ароматические амины, замещенные и полиароматические фенолы. Способность лакказы катализировать окисление указанных соединений и обуславливает применение этого фермента в биотехнологии. В результате окисления фенольных соединений в присутствии лакказы образуются продукты их конъюгации, хиноны, полимерные соединения, что позволяет использовать лакказу в органическом синтезе (Kunamneni A., Camarero S., García-Burgos С., Plou F.J, Ballesteros A., Alcalde M. «Engineering and Applications of fungal laccases for organic synthesis.» Microbial Cell Factories, 2008, V.7, p.32). Лакказа способна взаимодействовать с различными ксенобиотиками - загрязнителями окружающей среды, в результате чего может снижаться их токсичность или происходит полное удаление из загрязненной среды. Поэтому лакказу применяют для ремедиации загрязненных сред (Strong P.J., Claus H. «Laccase: a review of its past and its future in bioremediation» Critical Reviews in Environmental Science and Technology, 2011, V.41, p.373). В текстильной промышленности лакказа применяется для замены химических окислителей при окраске или обесцвечивании волокон (WO 2001044563 А1; WO 2003016615 А1). В целлюлозно-бумажной промышленности лакказа используется вместо химических окислителей либо совместно с ними для отбеливания бумажной массы (Gamelas J.A.F., Tavares A.P.M., Evtuguin D.V., Xavier A.M.В. «Oxygen bleaching of kraft pulp with polyoxometalates and laccase applying a novel multi-stage process». J Mol Catal B-Enzym, 2005, V.33, p.57; WO 2006126983 А1). В пищевой промышленности лакказу применяют для улучшения свойств теста, осветления соков и вина, удаления неприятных запахов продуктов (Minussi R.C., Pastore G.M., Duran N. «Potential applications of laccase in the food industry». Trends Food Sci Technol, 2002, V.13, p.205; WO 9428728 А1; US 6074631 A).
Лакказа широко распространена среди живых организмов. Фермент обнаружен у растений, грибов, бактерий, насекомых. Наиболее активными продуцентами лакказы являются дереворазрушающие грибы, поэтому наиболее изучена лакказа этих продуцентов. В состав молекулы лакказы входят четыре атома меди, образующие активный центр фермента. Ионы меди координируются аминокислотами полипептидной цепи - гистидинами и цистеином. Последовательности аминокислот, содержащие гистидины и цистеин, координирующие ионы меди, являются консервативными для различных организмов (Solomon E.I., Sundaram U.M., Machonkin Т.Е. «Multicopper oxidases and oxygenases». Chem Rev, 1996, V.96, p.2563). Молекула лакказы организована тремя доменами, между которыми расположен активный центр, молекулярная масса молекулы белка около 60-70 кДа. Такое строение молекулы характерно для грибов, растений и бактерий (Thurston C.F. The structure and function of fungal laccases. - Microbiol, 1994, vol.140, p.19; Mayer A., Staples R.C. «Laccase: new function for an old enzyme». Phytochem, 2002, V.60, p.551; Claus H. «Laccases and their occurrence in prokaryotes». Arch Microbiol, 2003, V.179, p.145). Лакказа указанных продуцентов активна в кислых значениях рН, оптимум окисления фенольных субстратов обычно составляет 4-5 единиц рН (Baldrian P. «Fungal laccases - occurrence and properties». FEMS Microbiol Rev, 2006, V.30, p.215). Термостабильность различных лакказ значительно варьирует, большинство лакказ стабильны в диапазоне температур 30-50°С и быстро инактивируются при температурах выше 60°С (Wood D.A. «Inactivation of extracellular laccase during fruiting of Agaricus bisporus». J Gen Microbiol, 1980, V.117, p.339; Xu F., Shin W., Brown S., Wahleithner J.A., Sundaram U.M., Solomon E.I. «A study of a series of recombinant fungal laccases and bilirubin oxidase that exhibit significant differences in redox potential, substrate specificity, and stability». Biochim Biophys Acta, 1996, V.1292, p.303). Однако существует пример термостабильного фермента у грибного продуцента. Лакказа гриба Steccherinum ochraceum 1833 сохраняла активность десятки минут при 80°С (Chernykh A., Myasoedova N., Kolomytseva M., Ferraroni M., Briganti F., Scozzafava A., Golovleva L. «Laccase isoforms with unusual properties from the basidiomycete Steccherinum ochraceum strain 1833». J Appl Microbiol, 2008, V.205, p.2065). Наряду с лакказой, молекула которой состоит из трех доменов, была обнаружена лакказа, состоящая из двух доменов, чья молекулярная масса составляла ~2/3 молекулярной массы трехдоменных лакказ (Machczynski M.C., Vijgenboom E., Samyn В., Canters G.W. Characterization of SLAC: «A small laccase from Streptomyces coelicolor with unprecedented activity» Protein Science, 2004, V.13, p.2388). Двухдоменная лакказа содержит активный центр, состоящий из четырех атомов меди и организованный аналогично активному центру трехдоменной лакказы (Skálová Т, Dohnáiek J, Østergaard LH, Østergaard PR, Kolenko P, Dušková J, Štĕpánková A, Hašek J «The structure of the small laccase from Streptomyces coelicolor reveals a link between laccases and nitrite reductases». J Mol Biol, 2009, V.385, p.1165). Каталитические свойства двухдоменных лакказ сильно отличаются от свойств трехдоменных. Двухдоменные лакказы обладают щелочным оптимумом рН окисления фенольных соединений и высокой термостабильностью, фермент сохранял активность при 80 и 90°С (Molina-Guijarro JM, Pérez J, Muñoz-Dorado J, Guillén F, Moya R, Hernández M, Arias ME «Detoxification of azo dyes by a novel pH-versatile, salt-resistant laccase from Streptomyces ipomoea». Int Microbiol, 2009, V.12, p.13; Dubé E, Shareck F, Hurtubise Y, Daneault C, Beauregard M «Homologous cloning, expression, and characterization of a laccase from Streptomyces coelicolor and enzymatic ecolourisation of an indigo dye». Appl Microbiol Biotechnol, 2008, V.79, p.597).
Существуют патенты, в которых описываются рекомбинантные лакказы грибных продуцентов: Coriolus hirsutus (EP 0388166), Melanocarpus albomyces (US 7183090), гриба рода Scytalidium (US 5843745). Однако эти патенты описывают трехдоменную лакказу, обладающую кислым рН оптимумом и относительно низкой термостабильностью. Существуют патенты, в которых описываются рекомбинантные термостабильные трехдоменные лакказы бактериальных продуцентов: Streptomyces lavendulae (JP 2003230392 А2), Thermus thermophilus (JP 2006158252 A2). Тем не менее, эти ферменты также имеют кислый рН оптимум окисления фенольных соединений.
Изобретение было сделано с помощью последовательности гена двухдоменной лакказы бактерии Streptomyces griseoflavus Tu4000, последовательность которой размещена в GenBank (NCBI Reference Sequence: ZP_07309495.1). Однако последовательность гена двухдоменной лакказы SEQ ID №1 бактерии Streptomyces griseoflavus Ac-993, полученная нами, отличается от последовательности, взятой за основу. Идентичность последовательностей составляет 86%. Данных о патентовании, экспрессии, получении и изучении свойств двухдоменной лакказы бактерии Streptomyces griseoflavus Tu4000 не обнаружено.
Сущность изобретения.
Технической задачей изобретения является: 1) клонирование и определение последовательности гена двухдоменной лакказы; 2) создание экспрессионной системы, экспрессия и очистка белка двухдоменной лакказы; 3) определение рН оптимума двухдоменной лакказы в отношении фенольного соединения и определение ее термостабильности.
Одним объектом изобретения является двухдоменная лакказа Streptomyces griseoflavus Ac-993, которая катализирует окисление фенольных соединений кислородом при щелочных значениях рН и устойчива к термоинактивации при 90 и 100°С. Двухдоменная лакказа имеет длину 322 а.о., в составе гена есть ТАТ-сигнальная последовательность с 1 по 30 а.о. Другим объектом изобретения является участок ДНК генома бактерии Streptomyces griseoflavus Ac-993, обозначенный как SEQ ID №1, кодирующий двухдоменную лакказу. Данный фрагмент ДНК может быть использован для продукции двухдоменной лакказы как in vivo, так и in vitro. Еще одним объектом изобретения является способ получения двухдоменной лакказы в активном виде. Способ получения двухдоменной лакказы включает клонирование гена, последовательность которого представлена в SEQ ID №1, двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993 в бактерии Escherichia coli, продукцию этого фермента в клетках бактерии Escherichia coli, внесение в среду культивирования ионов меди с целью наработки активного белка, очистку фермента с помощью металлхелатной хроматографии и гельфильтрации.
Способ получения фермента подразумевает клонирование в экспрессионном векторе в клетках бактерии Escherichia coli гена двухдоменной лакказы. Клонирование возможно, как полного белка (включая сигнальный пептид), так и белка без сигнального пептида. В полученной в результате клонирования рекомбинантной плазмиде гена двухдоменной лакказы помещается под контроль эффективных сигналов транскрипции и трансляции. Клонирование может быть проведено таким образом, что при экспрессии гена в клетках бактерии Escherichia coli будет синтезироваться фермент, слитый на N- и/или С-концах с дополнительными аминокислотными последовательностями, предназначенными для выделения и очистки продуцированного фермента с помощью металлхелатной хроматографии. Затем полученную рекомбинантную плазмиду вводят в штамм бактерии Escherichia coli, в котором проводится экспрессия клонированного гена. Фермент, не содержащий атомов меди в активном центре, накапливается в штамме-продуценте бактерии Escherichia coli в неактивной форме. Однако если в среду культивирования рекомбинантного штамма вносить ионы меди, то происходит накопление активного фермента. После экспрессии фермент клетки лизируют, фермент экстрагируют из лизата, затем проводят очистку фермента путем металлхелатной хроматографии и гельфильтрации.
Сущность изобретения состоит в следующем.
1) Ген, кодирующий двухдоменную лакказу с последовательностью SEQ ID №1, находится в геноме бактерии Streptomyces griseoflavus Ac-993. Для получения гена были сконструированы праймеры на основе последовательности гена двухдоменной лакказы Streptomyces griseoflavus Tu4000, находящейся в свободном доступе в GenBank. С помощью олигонуклеотидных праймеров к соответствующему гену при помощи полимеразной цепной реакции (ПЦР) был амплифицирован соответствующий фрагмент гена двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993. Последовательность гена была определена из амплифицированного фрагмента ДНК и представлена как SEQ ID №1. Затем установленная нуклеотидная последовательность была транслирована в аминокислотную последовательность (последовательность SEQ ID №2). Анализ базы данных последовательностей и белков GenBank (http://www.ncbi.nlm.nih.gov/Genbank/index.html) показал, что идентифицированная нами двухдоменная лакказа является уникальным ферментом, т.к. идентичных белков обнаружено не было. Идентичность полученной аминокислотной последовательности двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993 с последовательностью двухдоменной лакказы Streptomyces griseoflavus Tu4000 составляла 86%. Это указывает на уникальность полученной последовательности гена двухдоменной лакказы.
2) Клонированный ген был вставлен в экспрессионный вектор, который позволяет вводить в состав последовательности белка 6 гистидинов, что облегчает очистку рекомбинатного белка с помощью металлхелатной хроматографии на носителе Ni-сефароза. Для получения активного белка в среду культивирования рекомбинантного штамма был внесен CuSO4×5H2O в концентрации 0,25 мМ. Без внесения сульфата меди активный белок не образовывался. Затем клетки рекомбинантной Escherichia coli разрушали и из полученного клеточного гомогената выделяли фермент. Элюция белка с колонки с Ni-сефарозой осуществляется изократически 0,3 М имидазолом. После металлхелатной хроматографии белок окончательно очищается до электрофоретически гомогенного состояния с помощью гельфильтрации на носителе Superdex 200.
3) Полученная двухдоменная лакказа обладает уникальными свойствами - катализирует окисление фенольных соединений при щелочных значениях рН, стабильность при 90 и 100°С. Для определения рН-зависимости окисления фенольного соединения в качестве субстрата использовали 2,6-диметоксифенол. Определение активности проводилось в универсальной буферной системе при различных значениях рН. Темостабильность двухдоменной лакказы определяли путем изучения скорости инактивации фермента при 90 и 100°С.
Перечень фигур
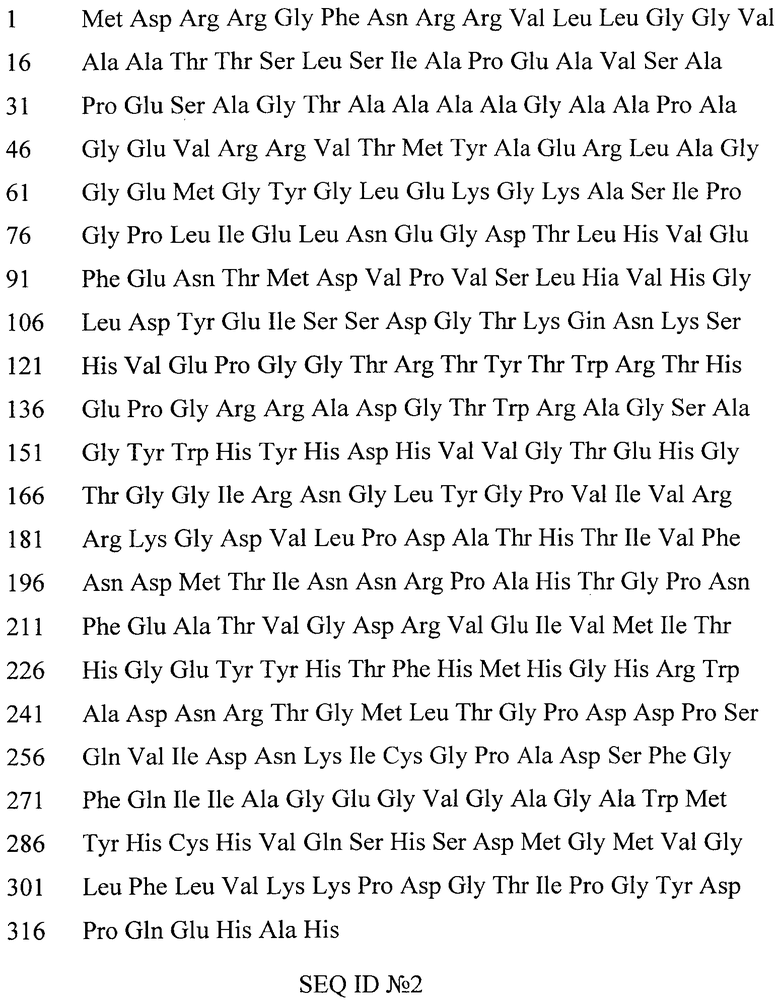
На Фиг.1 представлен электрофорезочищенного препарата лакказы бактерии Streptomyces griseoflavus Ac-993, полоса А - маркеры молекулярной массы, полоса Б - двухдоменная лакказа.
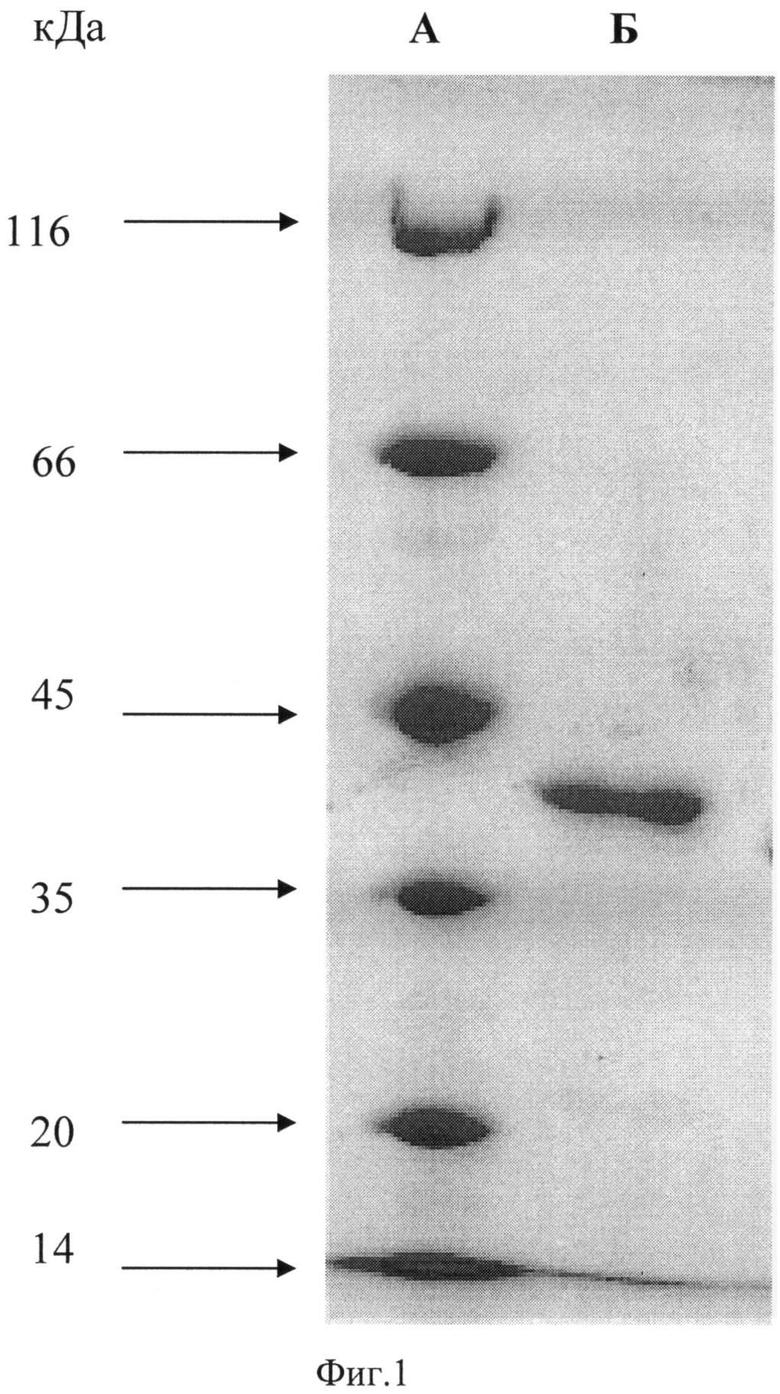
На Фиг.2 - зависимость активности двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993 относительно фенольного соединения 2,6-диметоксифенола от величины рН. Оптимум окисления фенольного соединения составил 9,2 единицы рН.
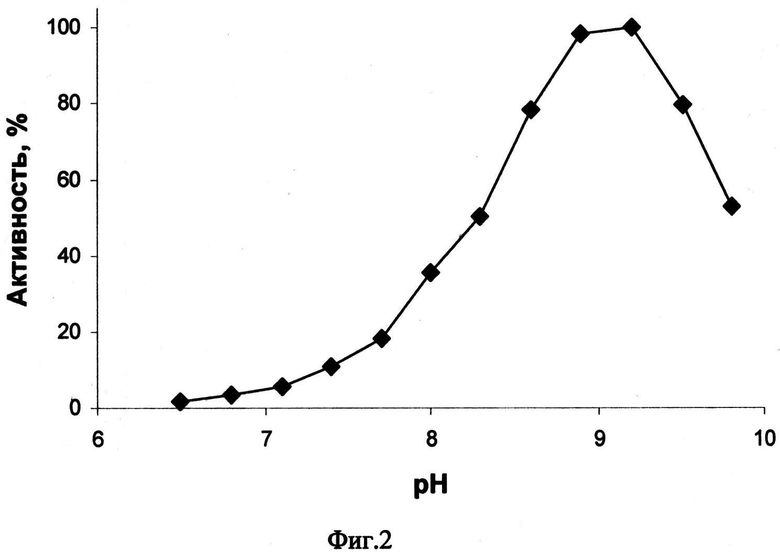
На Фиг.3 - зависимость активности двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993 от времени инкубации при 90 и 100°С. Фермент сохранял 35% активности при 90°С и 6% активности при 100°С в течение часа.
Сведения, подтверждающие возможность осуществления изобретения
Пример 1. Клонирование гена двухдоменной лакказы в экспрессионный вектор pQE и экспрессия его в клетках бактерии Escherichia coli M15(pREP4).
Ген двухдоменной лакказы амплифицируют из тотальной ДНК бактерии Streptomyces griseoflavus Ac-993 при помощи ПЦР с использованием олигонуклеотидных праймеров:
ZP_07309495.1SNA - 5'-GAGGCTGAAGCTTACatggacagacgcggtttcaac-3'
ZP_07309495.1ECO - 5'-GTGCTGATGGAATTCgtgcgcgtgctcctgg-3'
Определена последовательность ампликона, наработанного с геномной ДНК штамма Streptomyces griseoflavus Ac-993, предположительно включающего в себя ген двухдоменной лакказы. Ампликон содержит последовательность гена двухдоменной лакказы (SEQ ID №1). Нуклеотидную последовательность гена транслировали в аминокислотную последовательность белка (SEQ ID №2).
Для проведения последующей процедуры клонирования были разработаны следующие праймеры:
S.gfFe - 5'-AGTGGATCC-atggacagacgcggtttcaac-3'
S.gfRe - 5'-TCAAAGCTT-tcagtgcgcgtgctcctgg-3'
В последовательность праймера S.gfFe содержится сайт эндонуклеазы рестрикции BamHI, а в последовательность праймера - S.gfRe сайт эндонуклеазы рестрикции HindIII.
ПЦР-смесь объемом 20 мкл содержит 1×SE-буфер для Т4-ДНК полимеразы, 125 мкМ каждого дезоксирибонуклеозидтрифосфата, 1,5 мкМ каждого праймера, 20 нг геномной ДНК бактерии Streptomyces griseoflavus Ac-993 и 1 единицу активности ДНК-полимеразы Т4. ПЦР проводят на приборе MJ Mini (Bio-Rad, США) при следующем режиме: 98°С - 10 сек, 50°С - 15 сек, 72°С - 3 мин 30 сек - 1 цикл. Затем в последующих циклах температура отжига праймеров составляет: 2-й цикл - 54°С, 3-й цикл - 58°С и далее 60°С, 62°С, 64°С, 65°С по 15 сек, денатурация и элонгация при этом составляют 98°С - 10 сек и 72°С - 3 мин 30 сек соответственно. Затем 29 циклов: 98°С - 30 сек, 72°С - 3 мин 30 сек.
Полученный ПЦР-фрагмент и плазмиду pQE-30 (Qiagen, Германия) гидролизуют эндонуклеазами рестрикции BamHI и HindIII и лигируют между собой при помощи ДНК-лигазы фага Т4 (Литва).
Лигазной смесью трансформируют клетки штамма бактерии Escherichia coli M15(pREP4) (Qiagen, Германия) и проводят скрининг устойчивых к ампициллину и канамицину клонов на наличие рекомбинантной плазмиды.
Проводят выделение рекомбинантных плазмид, их рестрикционный анализ и секвенируют в составе этих плазмид фрагмент ДНК, соответствующий последовательности гена медьсодержащей оксидазы штамма Streptomyces griseoflavus Ac-993. В результате получают рекомбинантную плазмиду, обозначенную pQE-993, которая имеет размер 4394 п.н. и содержит ген двухдоменной медьсодержащей оксидазы бактерии Streptomyces griseoflavus Ac-993 под контролем фагового промотора Т5.
Клетки штамма бактерии Escherichia coli M15(pREP4) трансформируют плазмидой pQE-993 и подращивают на агаризованной среде LB (бакто-триптон - 10 г/л, бакто-дрожжевой экстракт - 5 г/л, NaCl - 10 г/л, агар - 17,5 г/л), содержащей ампициллин в концентрации 100 мкг/мл и канамицин в концентрации 25 мкг/мл.
Клетки бактерии Escherichia coli M15(pREP4), содержащие плазмиду pQE-993 (M15(pREP4)/pQE-993) инокулируют в 5 мл жидкой среды LB, содержащей ампициллин в концентрации 100 мкг/мл и канамицин в концентрации 25 мкг/мл. Пробирки инкубируют при 37°С ночь, затем отбирают 50 мкл подросшей за ночь культуры и инокулируют в 5 мл среды LB с ампициллином и канамицином в концентрации 100 мкг/мл и 25 мкг/мл соответственно. Культуру подращивают при 37°С и постоянном перемешивании (250 об/мин) 3 часа, затем вносят в среду с культурой изопропил-β-D-1-тиогалактопиранозид (ИПТГ) до конечной концентрации 1 мМ. Культуру клеток подращивают при 37°С и постоянном перемешивании (250 об/мин) 3 часа. Об экспрессии гена медьсодержащей оксидазы судят по проявлению индукции синтеза рекомбинантного белка исходя их сравнения белковых профилей штамма M15(pREP4) до внесения и после внесения индуктора - ИПТГ. Продукцию белка проверяют с помощью SDS-электрофореза в полиакриламидном геле.
Пример 2. Экспрессия белка в клетках бактерии Escherichia coli M15(Rep4) и очистка давухдоменной лакказы.
Клетки бактерии Escherichia coli штамма M15(pREP4) трансформируют плазмидой pQE-993 (аналогично примеру 1) и выращивают трансформантов на агаризованной среде LB, содержащей ампициллин в концентрации 100 мкг/мл и канамицин в концентрации 25 мкг/мл.
Подросших трансформантов M15(pREP4)/pQE-993 инокулируют в 5 мл среды LB, содержащей ампициллин в концентрации 100 мкг/мл и канамицин в концентрации 25 мкг/мл, подращивают в течение 16 часов при 37°С и постоянном перемешивании (250 об/мин).
Среду LB, содержащей ампициллин в концентрации 100 мкг/мл и канамицин в концентрации 25 мкг/мл, объем 200 мл, засевают инокулятом клеток бактерии Escherichia coli M15(pREP4)/pQE-993, выращенным в течение 16 часов при 37°С, и инкубируют на роторной качалке при 37°С и 250 об/мин до достижения культурой оптической плотности 0,03 при длине волны 600 нм. Начальная оптическая плотность среды составляет: D600=0,003 о.е. Экспрессию индуцируют добавлением изопропил-β-D-тиогалактопиранозида до концентрации 0,1 мМ, также одновременно с индуктором в среду вносят 0,25 мМ раствор CuSO4×5H2O. Индуцированную культуру инкубируют на роторной качалке при 18°С и 50 об/мин в течение 17 часов.
Осаждают выращенную биомассу центрифугированием при 4°С и 6000×g в течение 15 минут.
Осажденную биомассу ресуспендируют в 10 мл 20 мМ натрий-фосфатного буфера рН 7,4, содержащего 500 мМ NaCl и 1 мМ имидазол, после чего разрушают клетки при помощи ультразвуковой дезинтеграции на ледяной бане. Полученный лизат клеток центрифугируют 30 минут при 10000×g и температуре 4°С для удаления неразрушенных клеточных фрагментов. Таким образом, получают клеточный экстракт.
Клеточный экстракт анализируют SDS-электрофорезом в 12% полиакриламидном геле в денатурирующих условиях на наличие индуцированного белка.
В данных условиях медьсодержащая оксидаза накапливается внутриклеточно, однако не включается в тельца включений, поэтому обнаруживается в растворимой фракции клеточного лизата.
Супернатант наносят на 5 мл сорбента Ni-NTA (Qiagen, Германия), собирают проскок, затем колонку промывают 25 мл натриево-фосфатного буфера рН 7,4, содержащего 500 мМ NaCl и 1 мМ имидазол.
Элюируют белок с сорбента Ni-NTA 15 мл 20 мМ натриево-фосфатного буфера рН 7,4, содержащего 500 мМ NaCl и 20 мМ имидазол.
Фракции, содержащие индуцированный белок, наносят на колонку Superdex200 (гель-фильтрация), уравновешенную 20 мМ натрий-ацетатным буфером рН 5,0, содержащим 0,1М NaCl. Таким образом, проводят доочистку целевого белка от небольшого количества примесных белков. На выходе получают электрофоретически чистый белковый препарат. Чистоту препарата определяют с помощью SDS-электрофорезом в 12% полиакриламидном геле в денатурирующих условиях. Результат очистки изображен на Фиг.1 (Фиг.1 представляет электрофорезочищенный препарат лакказы бактерии Streptomyces griseoflavus Ac-993, полоса А - маркеры молекулярной массы, полоса Б - двухдоменная лакказа).
Диализуют препарат при 4°С в течение 15 часов против 50 объемов 20 мМ натрий-ацетатного буфера рН 5,0 для удаления NaCl из препарата.
Оценивают концентрацию полученного белкового препарата спектрофотометрически при длине волны 280 нм. Выход белка составляет ~10 мг при очистке из 1000 мл культуры.
Пример 3. Определение рН оптимума окисления двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993.
Изучение окисления фенольного соединений, 2,6-диметоксифенола, проводили в универсальной буферной системе, которая была получена путем смешивания равных объемов 0,05 М уксусной кислоты, 0,05 М H3PO4, 0,05 М борной кислоты. Необходимое значение рН буферного раствора доводили внесением 1 М раствора NaOH, величину рН в ходе внесения NaOH контролировали с помощью рН-метра. Буферные растворы приготовляли с шагом рН 0,3 единицы, начиная с 6,5. На основе полученных буферных растворов с различным значением рН приготовляли реакционные смеси с концентрацией 2,6-диметоксифенола равной 1 мМ. Активность двухдоменной лакказы определяли спектрофотометрически по увеличению поглощения при 469 нм за счет образования в ходе реакции продукта окисления 2,6-диметоксифенола. Реакцию проводили в кварцевой кювете объемом 1 мл в течение 180 секунд при 30°С. Реакцию начинали внесением фермента, который вносили в количестве 2,57 мкг/мл. Активность выражали в условных единицах активности, которая равнялась количеству фермента, осуществляющему изменение поглощения при 469 нм за минуту. Самую высокую активность при оптимальном значении рН принимали за 100%, активность при остальных значениях рН выражали в процентах в отношении самой высокой активности. Результаты определения рН оптимума показаны на Фиг.2 (Фиг.2 - зависимость активности двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993 относительно фенольного соединения 2,6-диметоксифенола от величины рН. Оптимум окисления фенольного соединения составил 9,2 единицы рН).
Пример 4. Определение термостабильности двухдоменной лакказы.
Исследование термостабильности двухдоменной лакказы проводили в 0,05 М универсальной буферной системе рН 4,5. Универсальная буферная система была получена путем смешивания равных объемов 0,05 М уксусной кислоты, 0,05 М H3PO4, 0,05 М борной кислоты. Необходимое значение рН буферного раствора доводили внесением 1 М раствора NaOH. Термоинактивацию определяли по скорости потери активности фермента в ходе его инкубирования при различных температурах. Двухдоменную лакказу разделяли на порции объемом 200 мкл, концентрация белка составляла 0,66 мг/мл. Проводили измерение исходной активности двухдоменной лакказы. Активность измеряли с помощью реакции окисления субстрата АБТС (2,2-азино-бис-(3-этилбензтиазолин-6-сульфонат). Реакцию отслеживали спектрофотометрически по образованию продукта окисления АБТС при 420 нм. Реакцию проводили в кювете объемом 1 мл в течение 1 минуты при 30°С. Затем порции фермента помещали на водяную баню с температурой 90 или 100°С. Через определенные промежутки времени отдельные порции фермента отбирали, центрифугировали на скорости 5000 g в течение 5 минут для охлаждения и осаждения конденсата, который образовывался на стенках сосуда с ферментом. После этого проводили измерение активности в отобранной порции. Вывод о степени инактивации двухдоменной лакказы делали по остаточной активности фермента, которую вычисляли путем определения отношения активности в ферменте после температурного воздействия на момент времени t к исходной активности фермента, выраженной в процентах по формуле:
А(ост)=[A(t)/A(исх)]*100,
где А(ост) - остаточная активность;
A(t) - активность на момент времени t;
А(исх) - исходная активность.
Результаты исследования термоинактивации фермента показаны на Фиг.3 (Фиг.3 - зависимость активности двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993 от времени инкубации при 90 и 100°С. Фермент сохранял 35% активности при 90°С и 6% активности при 100°С в течение часа).

Изобретение относится к области биотехнологии, в частности к способам получения ферментных препаратов. Изобретение касается термостабильной двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993 со щелочным оптимумом активности, последовательности ДНК, кодирующей данный фермент, и способа получения фермента, включающий клонирование в клетках бактерии Escherichia coli гена фермента, продукцию фермента в клетках Escherichia coli, внесение ионов меди в среду для культивирования рекомбинантного штамма для образования активного фермента, получение ферментного препарата методами металлхелатной хроматографии и гельфильтрации. Изобретение позволяет расширить ассортимент ферментов, таких как лакказа. 3 н.п. ф-лы,3 ил.
1. Двухдоменная лакказа бактерии Streptomyces griseoflavus Ac-993, обладающая высокой термостабильностью и оптимумом рН окисления фенольных соединений в щелочной области с аминокислотной последовательностью, представленной в SEQ ID №:2.
2. Фрагмент ДНК, обозначенный в SEQ ID №1 и кодирующий аминокислотную последовательность двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993, представленную в SEQ ID №:2.
3. Способ получения двухдоменной лакказы бактерии Streptomyces griseoflavus Ac-993, включающий клонирование в клетках бактерии Escherichia coli гена двухдоменной лакказы, имеющего нуклеотидную последовательность SEQ ID №1, продукцию двухдоменной лакказы в клетках Escherichia coli, внесение в среду культивирования рекомбинантного штамма CuSO4 x 5 H2O в концентрации 0,25 мМ с целью синтеза бактерией активной двухдоменной лакказы, получение препарата двухдоменной лакказы методами металлхелатной хроматографии и гельфильтрации.
| Лигатура для чугуна | 1984 |
|
SU1294859A1 |
| Буферное запоминающее устройство | 1978 |
|
SU767836A1 |
| Прибор для измерения площадей | 1928 |
|
SU15438A1 |
| Груз для погружения в воду гидрометрических приборов | 1933 |
|
SU37864A1 |
| DUBE E et.al | |||
| Homologous cloning, expression, and characterisation of a | |||

