Область техники, к которой относится изобретение
Изобретение относится к области молекулярной биологии, вирусологии и медицине и касается способа идентификации генотипа и подтипа вируса гепатита С (ВГС) на основе анализа последовательностей 5' нетранслируемой области генома ВГС с использованием олигонуклеотидного микрочипа.
Уровень техники
ВГС относится к семейству РНК-содержащих вирусов Flaviviridae и вызывает у людей инфекционный процесс, наиболее часто встречаемым осложнением которого является гепатит, перерастающий в цирроз печени и гепатокарциному (Surveillance. Hepatitis. CDC Report №61; Younossi Z, Kallman J, Kincaid J. The effects of HCV infection and management on health-related quality of life. Hepatology. 2007 Mar; 45(3):806-16). Этим заболеванием поражено более чем 170 миллионов человек на Земле, и число инфицированных продолжается увеличиваться. По всему миру насчитывается порядка 1,5 млн случаев гепатокарциномы, вызванной инфекцией ВГС. Потери, связанные с этим заболеванием только в Соединенных Штатах, оцениваются в 200 миллионов долларов ежегодно.
Распространенной современной тенденцией в лечении ВГС является применение комплексной терапии, включающей совместное введение мегадоз интерферона с коктейлем, содержащим как общие противовирусные препараты, так и один или два ингибитора размножения ВГС (ингибиторы специфической протеазы - геликазы или(и) РНК-полимеразы) (Toniutto P, Fabris С, Bitetto D, Fornasiere E, Rapetti R, Pirisi M. Valopicitabine dihydrochloride:a specific polymerase inhibitor of hepatitis С virus. Curr Opin Investig Drugs. 2007 Feb; 8(2): 150-8; Johnson CL, Owen DM, Gale M Jr. Functional and therapeutic analysis of hepatitis С virus NS3.4A protease control of antiviral immune defense. J Biol Chem. 2007 Apr; 282(14): 10792-803). Подобные коктейли повышают процент излечения, однако, неминуемо ведут к селекции адаптивных мутантов ВГС, устойчивых к данным ингибиторам.
Идентификация генотипа исследуемого образца ВГС имеет важное значение для оценки длительности и эффективности противовирусной терапии, а также для установления пути распространения вируса. Точное установление подтипа ВГС может быть полезно для оценки эффективности действия новейших ингибиторов вирусной РНК-полимеразы, которые могут быть использованы в качестве противовирусных препаратов. (Pauwels F, Mostmans W, Quirynen LM, van der Helm L, Boutton CW, Rueff AS, Cleiren E, Raboisson P, Surleraux D, Nyanguile О, Simmen KA Binding-site identification and genotypic profiling of hepatitis С virus polymerase inhibitors. J Virol. 2007 Jul; 81(13):6909-19).
Для определения генотипов и подтипов вируса гепатита С в настоящее время применяются следующие методы:
I. Прямое определение нуклеотидной последовательности (Секвенирование) 5'-некодирующей области генома вируса гепатита С с последующим анализом полученной последовательности путем сравнения с существующей базой данных, на основании которого делается заключение о принадлежности данного образца к определенному генотипу и подтипу (набор 'TRUGENE HCV5 'НТР' (Bayer HealthCare LLC, США)):
Jeffrey J. Germer, David W. Majewski, Michael Rosser, Amber Thompson, P. Shawn Mitchell, Thomas F. Smith, Slava Elagin, and Joseph D.C. Yao. 2003. Evaluation of the TRUGENE HCV 5'HTP Genotyping Kit with the New GeneLibrarian Module 3.1.2 for Genotyping of Hepatitis С Virus from Clinical Specimens. J Clin Microbiol., Vol.41, No. 10, p.4855-4857;
II. Метод зондов (Line Probe assay (LiPA)):
Verbeeck J, Stanley MJ, Shieh J, Celis L, Huyck E, Wollants E, Morimoto J, Farrior A, Sablon E, Jankowski-Hennig M, Schaper C, Johnson P, Van Ranst M, Van Brussel M. Evaluation of the VERSANT HCV Genotype Assay (LiPA) 2.0. J Clin Microbiol. 2008 Apr 9;
Nadarajah R, Khan GY, Miller SA, Brooks GF. Evaluation of a new-generation line-probe assay that detects 5' untranslated and core regions to genotype and subtype hepatitis С virus. Am J Clin Pathol. 2007 Aug; 128(2):3 00-4;
III. Метод экстенции генотип-специфического праймера (Primer-specific extension analysis):
Antonishyn NA, Ast VM, McDonald RR, Chaudhary RK, Lin L, Andonov AP, Horsman GB. 2005. Rapid genotyping of hepatitis С virus by primer-specific extension analysis. J Clin Microbiol. Oct; 43(10):5158-63.
IV. Масс-спектрометрические методы (Matrix-assisted laser desorption ionization-time of flight (mass spectrometry)):
Ilina EN, Malakhova MV, Generozov EV, Nikolaev EN, Govorun VM. 2005. Matrix-assisted laser desorption ionization-time of flight (mass spectrometry) for hepatitis С virus genotyping. J Clin Microbiol. Jun; 43(6):2810-5.
V. Методы иммуноферментного анализа (Serotyping of hepatitis C virus):
Elsawy ЕМ, Sobh MA, El-Chenawi FA, Hassan IM, Shehab El-Din AB, Ghoneim MA 2005. Serotyping of hepatitis С virus in hemodialysis patients: comparison with a standardized genotyping assay Diagn Microbiol Infect Dis. Feb; 51(2):91-4.
VI. Гетеродуплексный анализ с применением капиллярного электрофореза (heteroduplex mobility analysis using temperature gradient capillary electrophoresis):
Margraf RL, Erali M, Liew M, Wittwer CT. 2004. Genotyping hepatitis С virus by heteroduplex mobility analysis using temperature gradient capillary electrophoresisro J Clin Microbiol. 2004 Oct; 42(10):4545-51.
VII. Метод инвазивных зондов (Invader Assay):
Germer JJ, Majewski DW, Yung B, Mitchell PS, Yao JD. 2006. Evaluation of the invader assay for genotyping hepatitis С virus. J Clin Microbiol. Feb; 44(2):318-23.
VIII. Метод гнездовой ПЦР с последующим структурно-специфичным расщеплением (Nested restriction site-specific PCR):
Krekulova L, Rehak V, Wakil AE, Harris E, Riley LW. 2001. Nested restriction site-specific PCR to detect and type hepatitis С virus (HCV): a rapid method to distinguish HCV subtype 1b from other genotypes. J Clin Microbiol. May; 39(5): 1774-80.
IX. Высокоэффективная жидкостная хроматография (denaturing high-performance liquid chromatography):
Liew M, Erali M, Page S, Hillyard D, Wittwer C. 2004 Hepatitis С genotyping by denaturing high-performance liquid chromatography. J Clin Microbiol. Jan; 42(1):158-63.
X. Транскрипционно-опосредованная амплификация в сочетании с методом зондов (transcription-mediated amplification in conjunction with the line probe assay):
Comanor L, Elkin C, Leung K, Krajden M, Kronquist K, Nicolas K, Horansky E, deMedina M, Kittichai P, Sablon E, Ziermann R, Sherlock C. 2003 Successful HCV genotyping of previously failed and low viral load specimens using an HCV RNA qualitative assay based on transcription-mediated amplification in conjunction with the line probe assay. J Clin Virol. Sep; 28(1): 14-26.
XI. Метод ПЦР с детекцией в режиме реального времени с последующим анализом кривых плавления (Melting curve analysis):
Doris M. Haverstick, Grant С.Bullock, and David E. Bruns. 2004. Genotyping of Hepatitis С Virus by Melting Curve Analysis: Analytical Characteristics and Performance. Clinical Chemistry 50, No. 12, p/2405-2407.
XII. Прямое определение нуклеотидной последовательности (Секвенирование) области NS5B вируса гепатита С с последующим анализом полученной последовательности путем построения филогенетического дерева и определения генотипа и подтипа исследуемого образца на основании локализации анализируемой последовательности в одном из кластеров данного дерева (NS5B sequencing followed by phylogenetic analysis):
К.Sandres-Saune, P.Deny, C.Pasquier, V.Thibaut, G.Duverlie, J.Izopet. 2003. Determining hepatitis С genotype by analyzing the sequence of the NS5b region. Journal of Virological Methods Vol.109, p.187-193;
Laperche S, Lunel F, Izopet J, Alain S, Deny P, Duverlie G, Gaudy C, Pawlotsky JM, Plantier JC, Pozzetto B, Thibault V, Tosetti F, Lefrere JJ. 2005. Comparison of hepatitis C virus NS5b and 5' noncoding gene sequencing methods in a multicenter study. J Clin Microbiol. Feb; 43(2):733-9;
Hnatyszyn, J., Beld M., Gualbertus Hubertus M., Guettouche Т., Gouw R., Van Der Meer, C., Beatrijs Maria. (Bayer Healthcare LLC). Methods and reagents for genotyping HCV. WO/2007/076493. International Application No. PCT/US2006/062582. Publication Date: 05.07.2007.
Методы (I-IV, VI-XI) основаны на анализе генотип- и подтип-специфических последовательностей 5'-нетранслируемой области генома ВГС (5'НТР). Анализ последовательностей 5'НТР области дает возможность однозначно идентифицировать все 6 генотипов ВГС и большинство клинически значимых подтипов. Однако методы II-IV, VI-XI выявляют только ограниченное количество подтипов ВГС. Так, наиболее распространенный коммерческий тест VERSANT HCV Genotype Assay (LiPA) 2.0 определяет 15 подтипов (1a, 1b, 1a/1b, 2a/2c, 2b, 3а, 3b, 3c, 4a, 4c/4d, 4e, 4f, 4h, 5a, 6a).
В настоящее время анализ области NS5B позволяет идентифицировать подтип 1b со специфичностью, близкой к 100%. Более того, исследование последовательностей данной области дает возможность выявить значительно большее число подтипов, чем при анализе последовательностей 5'НТР (Thomas F, Nicot F, Sandres-Saune К, Dubois M, Legrand-Abravanel F, Alric L, Peron JM, Pasquier C, Izopet J. 2007. Genetic diversity of HCV genotype 2 strains in south western France. J Med Viral. Jan; 79(1):26-34; Nicot F, Legrand-Abravanel F, Sandres-Saune K, Boulestin A, Dubois M, Alric L, Vinel JP, Pasquier C, Izopet J. 2005. Heterogeneity of hepatitis С virus genotype 4 strains circulating in south-western France. J Gеn Virol Jan; 86(Pt 1):107-14). В то же время, в силу чрезвычайно высокой вариабельности области NS5B, эффективная амплификация фрагмента области NS5B всех генотипов и подтипов затруднена, что приводит к потери чувствительности анализа в целом, и, как следствие, невозможности создания надежной и эффективной диагностической тест-системы, используемой в качественно рутинного инструмента генотипирования ВГС. (Martró E, González V, Buckton AJ, Saludes V, Femández G, Matas L, Planas R, Ausina V. Evaluation of a new assay in comparison with reverse hybridization and sequencing methods for hepatitis C virus genotyping targeting both 5' noncoding and nonstructural 5b genomic regions. J Clin Microbiol. 2008 Jan; 46(1): 192-7.)
Метод секвенирования последовательности области NS5B ВГС с последующим филогенетическим анализом (XII) требует постановки реакций амплификации и секвенирования, проведения дополнительной очистки продуктов реакций после каждой из вышеперечисленных стадий и последующего анализа на автоматическом секвенаторе. Более того, последующий анализ хроматограмм, конструирование множественного выравнивания и построение филогенетических деревьев предъявляют повышенные требования к квалификации персонала, что препятствует широкому использованию данного подхода для анализа потока клинических образцов в условиях ординарной диагностической лаборатории.
Метод выявления серотипов с помощью вариантов иммуноферментного анализа (V) позволяет выявлять лишь ограниченный набор генотипов и подтипов (1а, 1b, 2a, 2b, 3а, и 4а) и требует наличия высокоочищенных моноклональных антител для каждого серотипа.
Также, для перечисленных выше методов идентификации генотипов и подтипов отмечены следующие недостатки:
- Коммерческий набор INNO-LiPA (II) и его применение в сочетании с транскрипционно-опосредованной амплификацией (X) отличается высокой себестоимостью и идентифицирует ограниченное количество подтипов. Более того, метод не позволяет однозначно идентифицировать подтип 4d, являющийся наиболее вирулентным и лекарственно устойчивым среди подтипов генотипа 4;
- Метод с использованием генотип-специфических праймеров в пробирке (III) требует постановки независимых реакций по количеству генотипов (т.е. не менее 6) для выявления одного только генотипа;
- ПЦР-гетеродуплексный анализ с применением капиллярного электрофореза (VI) и метод гнездовой ПЦР с последующим структурно-специфичным расщеплением (VIII) трудоемки, занимают большое количество времени и требуют типовых стандартов на каждый определяемый генотип и/или подтип;
- Метод инвазивных зондов (VII) идентифицирует только генотип и не позволяет сделать заключение о подтипе, что является серьезным ограничением для его применения в клинической практике, где необходимо выявление лекарственно-устойчивых разновидностей ВГС (по крайней мере, 1b и 4d) для оценки эффективности и длительности терапии;
- Методы масс-спектрометрии (IV) и ВЭЖХ (IX) требуют наличия дорогостоящего оборудования, дополнительных стадий подготовки образца для анализа и идентифицируют ограниченное число подтипов;
- ПЦР с детекцией в режиме реального времени (XI) выявляет наличие только самых распространенных генотипов и подтипов и при этом весьма дорог для рутинного анализа.
Таким образом, в данной области существует острая потребность в разработке способа идентификации генотипа и подтипа вируса гепатита С, который бы выгодно отличался от известных из уровня техники решений простотой проведения анализа, высокими специфичностью и информативностью в отношении числа идентифицируемых генотипов и подтипов, а также невысокой стоимостью.
Раскрытие изобретения
В результате проведенных обширных научных исследований, анализа баз данных нуклеотидных последовательностей генома ВГС авторы настоящего изобретения обнаружили, что задача разработки способа для идентификации генотипа и подтипа вируса гепатита С может быть успешно решена путем использования олигонуклеотидных микрочипов, содержащих генотип- и подтип-специфичные зонды, последовательности которых комплементарны последовательностям 5'-некодирующей области генома ВГС.
Способ идентификации генотипа и подтипа вируса гепатита С (ВГС) на олигонуклеотидных микрочипах выгодно отличается от известных из уровня техники методов возможностью выявления всех шести генотипов (1-6) и 22 подтипов ВГС (1а, 1b, 1d, 1е, 2а, 2b, 2с, 2i, 2k, 3а, 3b, 4а, 4с, 4d, 4f, 4h, 4k, 4r, 5a, 6a, 6b, 6d) в клинических образцах, а также низкой себестоимостью, малым временем, необходимым для получения результата. Метод не требует дорогостоящего оборудования и высококвалифицированного персонала. Данные, полученные с помощью заявляемого способа, могут быть использованы для оценки и прогнозирования тяжести протекания заболевания (острый/хронический цирроз, вероятность развития рака печени), определения терапевтической дозы лекарственных препаратов и длительности курса терапии, а также для эпидемиологического генотипирования.
В своем первом аспекте данное изобретение обеспечивает способ идентификации генотипа и подтипа ВГС с использованием олигонуклеотидного микрочипа, содержащего генотип- и подтип-специфичные зонды. Процедура, заявляемая в настоящем изобретении, основана на двухстадийной ПЦР с получением флуоресцентно меченого, преимущественно одноцепочечного фрагмента 5'НТР области с последующей гибридизацией этого фрагмента на микрочипе, содержащем набор специфичных дискриминирующих олигонуклеотидов, комплементарных вариантам последовательностей 5'НТР области, специфичных в отношении генотипов и подтипов.
Способ предусматривает следующие стадии:
(а) - обратную транскрипцию, совмещенную с ПЦР (ОТ-ПЦР), с использованием вирусной РНК в качестве матрицы и первой пары праймеров, специфичных к фрагменту 5'НТР области, последовательности которых представлены SEQ ID NO: 53 и 54;
(б) - асимметричную амплификацию фрагмента 5'НТР области с использованием в качестве матрицы продукта ОТ-ПЦР, полученного на стадии (а), второй пары специфичных праймеров, последовательности которых представлены SEQ ID NO: 55 и 56, и смеси четырех дезоксинуклеозидтрифосфатов, в которой один из четырех дезоксинуклеозидтрифосфатов является флуоресцентно меченым, в качестве субстрата, с получением преимущественно одноцепочечного флуоресцентно меченого фрагмента;
(в) - олигонуклеотидный микрочип для идентификации генотипа и подтипа ВГС, представляющий собой подложку, содержащую множество дискретных элементов, в каждом из которых иммобилизован уникальный олигонуклеотидный зонд, имеющий последовательность, комплементарную последовательности одноцепочечного фрагмента, полученного на стадии (б), и выбранную из группы, включающей: а) генотип-специфичные для каждого из генотипов ВГС последовательности фрагмента 5'-нетранслируемой области; и б) подтип-специфичные для каждого из подтипов ВГС последовательности фрагмента 5'-нетранслируемой области, при этом последовательности иммобилизованных олигонуклеотидных зондов представлены SEQ ID NO: 1-52;
(г) - гибридизацию амплифицированного меченого продукта, полученного на стадии (б), на олигонуклеотидном микрочипе с образованием дуплексов с иммобилизованными зондами в условиях, обеспечивающих разрешение в один нуклеотид между образующимися в результате гибридизации совершенными и несовершенными дуплексами;
(д) - регистрацию результатов гибридизации на олигонуклеотидном микрочипе, проведенной на стадии (г), осуществляемую с помощью портативного анализатора флуоресценции и программного обеспечения, что позволяет использовать программную обработку интенсивностей сигналов с последующей интерпретацией результатов;
(е) - интерпретацию результатов гибридизации, полученных на стадии (д), проводимую в два этапа: на первом этапе анализируют сигналы в элементах микрочипа, содержащих олигонуклеотидные зонды, специфичные к генотипам ВГС, тем самым идентифицируя генотип исследуемого образца; в случае идентификации генотипа на втором этапе анализируют только элементы микрочипа, содержащие олигонуклеотидные зонды, специфичные к подтипам идентифицированного генотипа, независимо от наличия сигналов в элементах, содержащих зонды, специфичные к подтипам других генотипов
В одном из своих воплощений способ характеризуется тем, что на стадии (б) один из праймеров второй пары используют, по меньшей мере, в десятикратном молярном избытке по отношению ко второму праймеру.
В своем следующем воплощении способ характеризуется тем, что на стадии (б) в качестве флуоресцентно меченного дезоксинуклеозидтрифосфата используют флуоресцентно меченый дезоксиуридинтрифосфат.
В еще одном из воплощений способ характеризуется тем, что олигонуклеотидный микрочип представляет собой микрочип на основе гидрогелевых ячеек, полученный способом химически или фото-индуцируемой сополимеризации.
Наконец, в еще одном своем воплощении способ дополнительно включает оценку и прогнозирование тяжести протекания заболевания (острый/хронический цирроз, вероятность развития рака печени), определение терапевтической дозы лекарственных препаратов и длительности курса терапии, и/или эпидемиологическое генотипирование на основе проведенной интерпретации результатов гибридизации.
Заявляемый аспект настоящего изобретения будет ясен из прилагаемых фигур, подробного описания и формулы изобретения.
Краткий перечень фигур
Для более ясного понимания сущности заявленного изобретения, а также для демонстрации его характерных черт и преимуществ далее приводится подробное описание изобретения со ссылками на фигуры чертежей, на которых:
Фиг.1 представляет принципиальную схему проведения анализа РНК ВГС с целью идентификации генотипа и подтипа ВГС на олигонуклеотидном микрочипе. Обозначения:
I - стадия обратной транскрипции, совмещенная с ПЦР при использовании вирусной РНК в качестве матрицы для получения двуцепочечной кДНК;
II - асимметричная амплификация фрагмента 5'-нетранслируемой области с включением дезоксинуклеозидтрифосфатов, содержащих флуоресцентную метку, с целью получения одноцепочечного флуоресцентно-меченного продукта для его последующей гибридизации на олигонуклеотидном микрочипе;
III - гибридизация флуоресцентно меченного одноцепочечного фрагмента 5'-нетранслируемой области, полученного на стадии II, с олигонуклеотидами, иммобилизованными на микрочипе;
IV - регистрация и интерпретация результатов гибридизации на олигонуклеотидном микрочипе.
Фиг.2 представляет схему выбора олигонуклеотидов для идентификации генотипов и подтипов ВГС.
Фиг.3 представляет схему размещения дискриминирующих олигонуклеотидов на микрочипе.
Фиг.4 представляет результаты проведения стадии ОТ-ПЦР с праймерами, последовательности которых представлены SEQ ID NO: 53 и 54, для образцов РНК ВГС, имеющих различные генотипы и подтипы. Обозначения (дорожки геля):
1 - подтип 1а; 2 - подтип 1b; 3 - подтип 1е; 4 - подтип 2а; 5 - подтип 2k; 6 - подтип 2i; 7 - подтип 3а; 8 - подтип 4а; 9 - подтип 4d; 10 - подтип 5а; 11 - подтип 6b; 12 - ДНК вируса гепатита В (отр. контр); 13 - РНК ВИЧ (отр. контр.); 14 - Отр. контроль стадии ОТ-ПЦР; М - молекулярный маркер 'FastRuler™ DNA Ladder, Middle Range' (Ферментас, Литва).
Фиг.5 представляет результаты проведения стадии ПЦР с праймерами, последовательности которых представлены SEQ ID NO: 55 и 56, с использованием в качестве матрицы продукта стадии ОТ-ПЦР, для образцов ВГС, имеющих различные генотипы и подтипы. Обозначения (дорожки геля):
1 - подтип 1а; 2 - подтип 1b; 3 - подтип 1е; 4 - подтип 2а; 5 - подтип 2k; 6 - подтип 2i; 7 - подтип 3а; 8 - подтип 4а; 9 - подтип 4d; 10 - подтип 5а; 11 - подтип 6b; 12 - ДНК вируса гепатита В (отр. контр); 13 - РНК ВИЧ (отр. контр.); 14 - Отр. контроль стадии ОТ-ПЦР; М - молекулярный маркер 'FastRuler™ DNA Ladder, Middle Range' (Ферментас, Литва).
Фиг.6 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе в результате анализа образца ВГС, имеющего генотип 1, подтип 1а.
Фиг.7 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца ВГС, имеющего генотип 1, подтип 1b.
Фиг.8 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца ВГС, имеющего генотип 1, подтип 1е.
Фиг.9 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца В ГС, имеющего генотип 2, подтип 2а.
Фиг.10 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца ВГС, имеющего генотип 2, подтип 2i.
Фиг.11 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца ВГС, имеющего генотип 3, подтип 3а.
Фиг.12 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца ВГС, имеющего генотип 4, подтип 4а.
Фиг.13 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца ВГС, имеющего генотип 4, подтип 4d.
Фиг.14 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца ВГС, имеющего генотип 5, подтип 5а.
Фиг.15 представляет флуоресцентную картину гибридизации и распределение нормированных сигналов ячеек, полученные на микрочипе, в результате анализа образца ВГС, имеющего генотип 6.
Осуществление изобретения
Задача настоящего изобретения состоит в создании способа идентификации генотипа и подтипа вируса гепатита С на основе анализа 5' нетранслируемой области генома ВГС с помощью олигонуклеотидного микрочипа.
В заявленном способе предложено использование: процедуры обратной транскрипции, совмещенной с полимеразной цепной реакцией (ОТ-ПЦР), для амплификации последовательности фрагмента 5'НТР области генома ВГС; получение с наработанного продукта одноцепочечного флуоресцентно меченого фрагмента, при этом в качестве исходного образца может быть использована вирусная РНК, выделенная из клинического материала, такого как, например, кровь, плазма или биоптат печени. Заявленный способ также предусматривает использование оригинального олигонуклеотидного микрочипа с иммобилизованными специфическими зондами, процедуры гибридизации, регистрации и интерпретации результатов.
Принципиальная схема идентификации генотипа и подтипа исследуемого образца ВГС на микрочипе.
Схема анализа последовательности фрагмента 5'НТР области генома ВГС для идентификации генотипа и подтипа с помощью микрочипа представлена на Фиг.1.
Выделение РНК ВГС из клинического образца осуществляют с помощью известных в данной области способов (например, Hourfar MK, Michelsen U, Schmidt М, Berger A, Seifried E, Roth WK. High-throughput purification of viral RNA based on novel aqueous chemistry for nucleic acid isolation. Clin Chem. 2005 Jul; 51(7): 1217-22), или любого специализированного коммерчески доступного набора реагентов для выделения РНК из крови, плазмы или биоптата печени, например QIAamp DSP Virus Kit (Кат №60704, Qiagen, Германия), MagMAX™ AI/ND Viral RNA Isolation Kits (Кат №AM1939, Ambion, США) или «Комплект реагентов для выделения РНК» (Кат №05-013, ЗАО «НПФ ДНК-технология», Россия).
Амплификацию фрагмента 5'НТР области генома ВГС на первом этапе выполняют с помощью реакции обратной транскрипции, совмещенной с ПЦР (ОТ-ПЦР).
Праймеры для проведения первой стадии амплификации выбирают таким образом, чтобы они фланкировали наиболее полиморфный сегмент 5'-нетранслируемой области генома ВГС, позволяющий дифференцировать существующие генотипы и подтипы ВГС. Предпочтительно, чтобы сегмент области, подлежащий амплификации, включал положения с 40 по 460 генома ВГС согласно Choo, Q.L., К.Н.Richman, J.Н.Наn, К.Berger, С.Lee, С.Dong, С.Gallegos, D.Coit, R.Medina-Selby, P.J.Barr, et al. 1991. Genetic organization and diversity of the hepatitis С virus. Proc. Natl. Acad. Sci. USA 88: 2451-2455.
Последовательности праймеров выбирают таким образом, чтобы проводить эффективную амплификацию РНК исследуемого сегмента 5'-нетранслируемой области любого генотипа и, соответственно, подтипа ВГС. Для этого, например, может быть сконструировано множественное выравнивание всех доступных в базах данных нуклеотидных последовательностей 5'НТР области, находящихся по адресам http://www.ncbi.nlm.nih.gov/Genbank/index.html и http://hcv. lanl. gov/content/hcv-db. Затем находят наиболее консервативные для всех генотипов ВГС участки исследуемой области и выбирают праймеры, специфичные к данным участкам. Используя специализированное программное обеспечение, например Oligo v.6.3 (Molecular Biology Insights Inc., США) или Fast PCR (http://www.biocenter.helsinki.fi/bi/Programs/fastpcr.htm) или другие коммерчески доступные программы, или программы, свободно доступные в сети Internet, рассчитывают температуры плавления праймеров и, варьируя их длину, добиваются того, чтобы разброс температур отжига праймеров внутри пары не превышал 3-4°С. При подборе праймеров избегают таких последовательностей, которые способны формировать вторичные структуры типа шпильки с высокими температурами плавления. Каждый выбранный праймер должен обладать уникальной специфичностью в отношении анализируемого участка. Специфичность праймеров проверяется с помощью программного обеспечения, использующего поиск в базах нуклеотидных последовательностей по алгоритму BLAST (например, www.ncbi.nlm.nih. gov/BLAST). В частности необходимо избегать таких последовательностей, которые способны продуктивно гибридизоваться (отжигаться) с последовательностями генома человека.
На втором этапе получают преимущественно одноцепочечный флуоресцентно меченый продукт методом асимметричной ПЦР с использованием в качестве субстрата для построения цепи de novo смеси дезоксинуклеозидтрифосфатов, в которой один из дезоксинуклеозидтрифосфатов является флуоресцентно меченым. Как известно специалистам в данной области, смесь дезоксинуклеозидтрифосфатов для проведения ПЦР содержит dATP, dGTP, dCTP и dTTP, причем вместо dTTP может использоваться dUTP или смесь dTTP/dUTP в любой молярной пропорции. Любой из указанных дезоксинуклеозидтрифосфатов может нести флуоресцентную метку. Наиболее предпочтительно использование флуоресцентно меченых дезоксиуридинтрифосфата и/или дезоксицитидинтрифосфата, что, с одной стороны, обусловливает эффективное включение данного субстрата во вновь синтезированную цепь ДНК в процессе ПЦР, с другой - обеспечивает возможность предотвращать получение ложных результатов за счет перекрестной кросс-контаминации ампликонами путем использования на первой стадии фермента урацил-ДНК-гликозилазы. Это имеет критическое значение при проведении массовых анализов в условиях специализированной лаборатории.
В качестве флуоресцентного красителя может быть использован любой флуоресцентный краситель, который может быть химически включен в молекулу дезоксинуклеозидтрифосфата таким образом, чтобы не препятствовать в существенной степени прохождению полимеразной цепной реакции и последующей гибридизации полинуклеотидной молекулы, содержащей такие флуоресцентно меченые нуклеотидные остатки, с иммобилизованными олигонуклеотидными зондами. В случае флуоресцентно меченого дезоксиуридинтрифосфата, например, флуоресцентный краситель может быть присоединен к 5'-концу аминоаллильного производного dUTP. Примеры таких красителей хорошо известны специалисту в данной области техники и включают красители флуоресцеинового (TAMRA®, ROX®, JOE®), родаминового (Texas Red®), полиметинового (Су3®, Су5®, Су5.5®, Су7®) рядов (Ranasinghe R. and Brown Т). Fluorescence based strategies for genetic analysis. Chem. Commun., 2005, 5487-5502). Флуоресцентные красители коммерчески доступны, в частности от фирмы Molecular Probes, США. Наиболее предпочтительными являются красители, спектр возбуждения которых лежит в длинноволновой (красной) области спектра, что позволяет использовать для возбуждения флуоресценции недорогие источники возбуждающего излучения типа полупроводниковых лазеров.
Флуоресцентно меченые дезоксинуклеозидтрифосфаты могут быть получены как в лабораторных условиях с использованием таких известных способов, как, например (Kuwahara M, Nagashima J, Hasegawa M, Tamura Т, Kitagata R, Hanawa К, Hososhima S, Kasamatsu T, Ozaki H, Sawai H. Systematic characterization of 2'-deoxynucleoside- 5'-triphosphate analogs as substrates for DNA polymerases by polymerase chain reaction and kinetic studies on enzymatic production of modified DNA. Nucleic Acids Res. 2006 34(19):5383-94), так и являются коммерчески доступными, например CyDye Fluorescent Nucleotides (Кат. №РА55021, РА55032, РА55026, GE Healthcare, США).
Праймеры для проведения второй стадии амплификации выбирают с учетом требований, изложенных выше, с тем отличием, что по меньшей мере один из праймеров выбирается внутри ПЦР-фрагмента, получаемого на первой стадии, что повышает специфичность реакции. Таким образом, получаемый продукт в конечном счете будет являться продуктом полугнездовой или гнездовой реакции амплификации. Размер амплифицируемого на второй стадии фрагмента специально не ограничивается до тех пор, пока это обеспечивает эффективную гибридизацию фрагмента с иммобилизованными на микрочипе зондами. В том случае, когда микрочип представляет собой микрочип на основе гидрогелевых элементов, праймеры для проведения второй стадии амплификации выбирают так, чтобы размер амплифицируемого фрагмента составлял 100-800 п.о. Большая длина ПЦР-продукта, получаемого на второй стадии, затрудняет эффективную диффузию анализируемого фрагмента генома в гелевых элементах микрочипа при гибридизации, что в конечном итоге может привести к уменьшению количества образовавшихся гибридизационных дуплексов и, как следствие, падению флуоресцентного сигнала в ячейках. При подборе праймеров следует учитывать тот факт, что на выходе реакции получают преимущественно одноцепочечные флуоресцентно меченые продукты, которые должны быть комплементарны олигонуклеотидам, иммобилизованным на микрочипе. Поэтому в каждой паре праймер, добавляемый в избытке («ведущий праймер»), выбирается из той цепи, последовательность которой комплементарна последовательностям иммобилизованных на микрочипе олигонуклеотидов. Т.е. в случае, если олигонуклеотиды для иммобилизации выбраны из смысловой цепи, для формирования гибридизационных дуплексов в ячейках микрочипа необходима преимущественная амплификация антисмысловой цепи, тем самьм «ведущий праймер» выбирается из цепи, комплементарной последовательности гена (антисмысловая цепь), и наоборот.
Для получения преимущественно одноцепочечного флуоресцентно меченого продукта ведущий праймер добавляют в молярном избытке по отношению ко второму праймеру. Предпочтительно молярный избыток является по меньшей мере десятикратным.
При выборе дискриминирующих олигонуклеотидов для иммобилизации на микрочипе с учетом размера и сложности анализируемой последовательности и, в частности, наличия повторов и протяженных гомополимерных последовательностей определяют длину дискриминирующих олигонуклеотидов, обеспечивающую их специфичность в отношении анализируемой последовательности.
Выбор дискриминирующих олигонуклеотидов для генотипов и подтипов осуществляют следующим образом. В сконструированном множественном выравнивании последовательностей 5'НТР области для каждого генотипа создается консенсусная последовательность данного региона, на основании которой выбираются уникальные зонды, позволяющие однозначно идентифицировать каждый генотип. При этом в силу высокой вариабельности генома ВГС для повышения надежности процедуры по возможности выбирают два дискриминирующих зонда для каждого из генотипов, комплементарных различным участкам исследуемого фрагмента 5'НТР области. Для каждого из подтипов также создают консенсусную последовательность, а затем выбирают участки фрагмента исследуемой 5'НТР области, позволяющие дифференцировать максимальное количество подтипов внутри одного генотипа. Количество таких участков должно быть достаточно для обеспечения надежной идентификации каждого из подтипов. На основании последовательностей выбранных участков конструируют зонды для идентификации подтипов, при этом последовательность одного зонда может соответствовать сразу двум и более подтипам в отдельном дифференцирующем участке исследуемого фрагмента 5'НТР области. Стратегия выбора зондов для иммобилизации на микрочипе схематично представлена на Фиг.2.
Используя программное обеспечение, например Oligo v. 6.3 (Molecular Biology Insights Inc., США), рассчитывают температуры плавления олигонуклеотидов и, варьируя их длину, добиваются того, чтобы разброс температур плавления олигонуклеотидов составлял не более 2-3°С. Избегают таких олигонуклеотидов, которые способны формировать вторичные структуры типа шпильки с высокими температурами плавления.
Дискриминирующие олигонуклеотиды иммобилизуют на подложке микрочипа. В качестве подходящих подложек для изготовления микрочипа могут использоваться активированная, например аминированная поверхность предметных стекол (Adessi С., Matton G., Ayala G., Turcatti G., Mermod J., Mayer P., Kawashima E. Solid phase DNA amplification: characterisation of primer attachment and amplification mechanisms Nucleic Acids Research. 2000. V. 51. 28(20).:E87), пластиковые носители (Nikiforov Т., Rendle R., Goelet P., Rogers Y., Kotewicz M., Anderson S., Trainor G., Knapp Michael R. Genetic Bit Analysis: a solid phase method for typing single nucleotide polymorphisms. Nucleic Acids Research. 1994. 22(20):4167-75), подложки с полимерными макропористьми носителями, такими как акриламид (Timofeev E., Kochetkova S., Mirzabekov A., Florentiev V. Regioselective immobilization of short oligonucleotides to acrylic copolymer gels. Nucleic Acids Res. 1996 24(16):3142-8), сефароза (Margulies M, Egholm M, Altman WE с соавт., Genome sequencing in microfabricated high-density picolitre reactors. Nature. 2005 437(7057):376-80) и др. Способы иммобилизации олигонуклеотидов на подложках хорошо известны в данной области и включают:
- хемосорбцию олигонуклеотидов и ДНК, содержащих тиоловую группу, на металлах (Mirkin C.A., Letsinger R.L., Mucic R.C. and Storhoff J.J. A DNA-based method for rationally assembling nanoparticles into macroscopic materials. Nature. 1996 Aug 15; 382(6592):607-9);
- ковалентное связывание модифицированных олигонуклеотидов с функциональными группами поверхности, основанное на реакциях образования амидной (Healey BG, Matson RS, Walt DR. Fiberoptic DNA sensor array capable of detecting point mutations. Anal Biochem. 1997 251(2):270-9), сложноэфирной (Ghosh SS, Musso GF. Covalent attachment of oligonucleotides to solid supports. Nucleic Acids Res. 1987 15(13):5353-72), карбимидоильной (Matson RS, Rampal J, Pentoney SL. Anderson PD, Coassin P. Biopolymer synthesis on polypropylene supports: oligonucleotide arrays. Anal Biochem. 1995 Jan 1; 224(1):110-6) связей и т.д.
- фотохимически, химически и электрохимически индуцируемую сополимеризацию олигонуклеотидов, несущих непредельную группу, с мономерами, составляющими основу формируемой твердой фазы (Vasiliskov A.V., Timofeev E.N., Surzhikov S.A., Drobyshev A.L., Shick V.V. and Mirzabekov A.D., Fabrication of microarray of gel-immobilized compounds on a chip by copolymerization. Biotechniques, 1999, 27, 592-606)
В предпочтительном воплощении настоящего изобретения используют микрочип на основе гидрогелевых элементов. Способы получения таких микрочипов включают полимеризацию аминомодифицированных олигонуклеотидов с образованием ковалентной связи с мономерами гидрогеля при соответствующих условиях (рН, температура, состав полимеризационной смеси и др.) (Rubina AY, Pan'kov SV, Dementieva El et al. Hydrogel drop microchips with immobilized DNA:
properties and methods for large-scale production. Anal Biochem 2004; 325: 92-106). Наиболее предпочтительным является использование микрочипов, содержащих гелевые элементы, которые наносятся на матрицу в виде капель диаметром от 80 до 300 мкм с периодом 150-500 мкм, без использования специальных приспособлений, например, кварцевых масок. В качестве матрицы может использоваться как стеклянная подложка (предметное или покровное стекло), так и более доступные материалы, такие как пластик. Для иммобилизации олигонуклеотидов в гелевых элементах микрочипа используется их совместная полимеризация с основными компонентами геля. В результате этой одностадийной реакции иммобилизуемые молекулы необратимо, ковалентно присоединяются к тем или иным мономерам растущей полимерной цепи и равномерно распределяются во всем объеме геля с высоким выходом (порядка 50% для олигонуклеотидов) (Rubina AY, Pan'kov SV, Dementieva EI et al. Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production. Anal Biochem 2004; 325: 92-106). Концентрация иммобилизованных олигонуклеотидных зондов может быть оценена прокрашиванием гелевых элементов микрочипа красителем с низкой специфичностью к нуклеотидной последовательности ДНК (А.Л.Михейкин, А.В.Чудинов, А.И.Ярощук, А.Ю.Рубина, С.В.Паньков, А.С.Крылов, А.С.Заседателев, А.Д.Мирзабеков. Краситель с низкой специфичностью к нуклеотидной последовательности ДНК: применение для оценки количества олигонуклеотидов, иммобилизованных в ячейках биологических микрочипов. Молекулярная биология 2003; 37(6): 1061-70).
ПЦР-продукты, полученные на второй стадии амплификации, гибридизуют на микрочипе с иммобилизованными дифференцирующими олигонуклеотидами, комплементарными участкам консенсусных последовательностей генотипов и подтипов фрагмента 5'НТР области. Гибридизацию проводят в растворе, содержащем буферный компонент для поддержания рН, соль для создания ионной силы и хаотропный (дестабилизирующий водородные связи) агент, в герметичной гибридизационной камере при температуре, зависящей от температуры плавления иммобилизованных на микрочипе дискриминирующих олигонуклеотидов. В качестве дестабилизирующего водородные связи агента могут быть использованы, например, гуанидин тиоцианат, мочевина или формамид. Выбор оптимальной температуры гибридизации проводят с учетом удобства практического применения системы. Дискриминирующие олигонуклеотиды, заявленные в настоящем изобретении, имеют температуру плавления в интервале от 42 до 44°С, что позволяет проводить гибридизацию при 37°С с использованием хаотропного агента. Температура 37°С удобна тем, что большинство клинических лабораторий оснащены термостатами, поддерживающими эту температуру.
Изучаемый фрагмент ДНК образует совершенные гибридизационные дуплексы только с соответствующими (полностью комплементарными) олигонуклеотидами. Со всеми остальными олигонуклеотидами изучаемый фрагмент ДНК дает несовершенный дуплекс. Дискриминацию совершенных и несовершенных дуплексов выполняют путем сравнения интенсивностей флуоресценции ячеек, в которых образовались дуплексы. Интенсивность сигнала в ячейке, в которой образовался совершенный гибридизационный дуплекс (Iсов), выше, чем в таковой, где образовался несовершенный дуплекс (Iнесов). Проведение гибридизации при оптимальных условиях (температура, подобранная концентрация хаотропного агента и ионная сила гибридизационного буфера) позволяет добиться соотношения Iсов/Iнесов≥1,5 между двумя ячейками, содержащими зонды, принадлежащие одной группе и различающиеся на один нуклеотид.
Регистрация результатов гибридизации на микрочипах может быть осуществлена с помощью коммерческих сканирующих устройств - анализаторов флуоресценции биологических микрочипов, например GenePix 4000B (Ахоn Instruments, США), оснащенных соответствующим программным обеспечением для вычисления интенсивности флуоресцентных сигналов дискретных элементов микрочипа и их последующей нормировки на фоновое значение, например 'GenePix Pro', 'Acuity' (Axon Instruments, США).
Интерпретация результатов гибридизации может быть осуществлена визуально соотнесением фиксированной флуоресцентной картины микрочипа и/или полученного распределения интенсивностей сигналов элементов микрочипа к расположению специфичных дискриминирующих зондов в элементах микрочипа (см. Фиг.3). Обладая распределением интенсивности сигналов в ячейках микрочипа, выделяют максимальный сигнал среди элементов, содержащих генотип-специфичные олигонуклеотиды. Идентификация генотипа может быть осуществлена путем установления элемента микрочипа, обладающего наибольшей интенсивностью флуоресценции, среди ячеек, содержащих зонды для определения генотипа ВГС. Идентификация подтипа исследуемого образца ВГС может быть осуществлена путем определения наибольших сигналов в ячейках, содержащих подтип-специфичные олигонуклеотиды, соответствующие установленному генотипу, в случае, если максимальные сигналы зарегистрированы по меньшей мере в двух различных ячейках, содержащих уникальные олигонуклеотиды, специфичные в отношении отдельного подтипа.
Предпочтительно интерпретацию результатов гибридизации на микрочипах осуществляют следующим образом. На первом этапе проводят выделение достоверных сигналов, т.е. сигналов в ячейках, где возможно образование совершенных дуплексов. Для этого нормированные сигналы интенсивности флуоресценции всех ячеек микрочипа сортируются по возрастанию и сравниваются с усредненным сигналом (Iref) в ячейках, не содержащих олигонуклеотидов. Достоверными сигналами считают такие, которые превосходят Iref, по меньшей мере, в 1,5 раза. Второй этап начинают с анализа отфильтрованных достоверных сигналов Gi в группах ячеек, содержащих олигонуклеотиды, специфичные к различным генотипам (i-номер генотипа). Выделяют максимальный сигнал внутри каждой группы ячеек Gimax и сравнивают между собой. Если сигнал Gimax в одной группе превосходит максимальные сигналы в остальных группах более чем в 1,5 раза (пороговое значение), то делается заключение о принадлежности исследуемого образца к данному генотипу. В случае, если отношение сигналов среди Gimах не превышает порогового значения, то делается заключение о невозможности однозначной идентификации генотипа и о возможном присутствии в анализируемом образце смеси двух и более вариантов ВГС с различными генотипами. В случае, если сигналы в группах, содержащих генотип-специфичные олигонуклеотиды, не проходят первичную фильтрацию относительно Iref, то делается заключение о низкой интенсивности сигналов и возможном отсутствии РНК ВГС в исследуемом образце. При этом идентификация подтипа не проводится.
Таким образом, в случае, если определен генотип на основании значения сигнала Gimax, то далее рассматриваются только группы ячеек, содержащих олигонуклеотиды, специфичные к подтипам, относящимся к идентифицированному генотипу. В соответствии с предложенной стратегий выбора зондов для идентификации подтипа олигонуклеотиды объединены в группы, соответствующие выбранным участкам исследуемого фрагмента, позволяющим дифференцировать максимальное число подтипов. При этом для каждого из подтипов сконструировано 2 олигонуклеотида, объединенных в 2 различные группы последовательностей фрагмента 5'-нетранслируемой области, и диктуется необходимостью надежной дифференциации подтипов внутри генотипа. При идентификации подтипа сначала выбирают сигналы внутри каждой группы ячеек, превосходящие остальные сигналы в этой группе, по меньшей мере, в 1,5 раза, такой сигнал обозначается как Sixj (при этом i - номер генотипа, 'х' - символьное обозначение подтипа, в соответствии с принятой классификации подтипов ВГС, j - номер группы). В случае, если в группе две и более ячейки имеют сигналы, различающиеся между собой менее чем в 1,5 раза, то выбирают все такие сигналы Sixj, Sivj и т.д. В результате получают набор ячеек из разных групп ixl, iy1, ix2, iz2, ix3 и т.д., сигналы в которых превосходят остальные сигналы в своих группах не менее чем в 1,5 раза. При этом, если в полученном наборе присутствуют по меньшей мере 2 ячейки из разных групп, соответствующие одному подтипу, например ix1 и ix3 или ix1 и ixy2, то выдают заключение о принадлежности исследуемого образца к подтипу 'х' генотипа 'i'. В случае, если в полученном наборе ячейки из разных групп соответствуют разным генотипам, например ix1, iy2, iz3 или ix1, iyz3, тогда интенсивности сигналов данных ячеек сравнивают между собой. Если сигнал ячейки, соответствующей подтипу 'х' одной группы, превосходит сигналы ячеек других групп в 3 и более раза, то делают заключение о принадлежности исследуемого образца к подтипу 'х'. В случае, если отношение сигналов Six1/Siy2 не превышает трех, выдают заключение о том, что подтип не установлен и возможна принадлежность к подтипу 'х' или подтипу 'у'. Такое же заключение выдается в случае, если максимальный сигнал в группе имеет ячейка, содержащая олигонуклеотид, специфичный к двум подтипам, например ixy1, при отсутствии достоверных сигналов в других группах ячеек. В случае, если сигналы в группах, содержащих подтип-специфичные олигонуклеотиды, не проходят первичную фильтрацию относительно Iref, то считают, что подтип исследуемого образца не определен.
Оценка длительности противовирусной терапии и прогноз течения заболевания могут быть сделаны на основании данных идентификации генотипа и подтипа. Так, в случае установления генотипа 1, приводящего к циррозу, хроническому гепатиту и гепатоцеллюлярной карциноме, длительность терапии пегилированным интерфероном и рибавирином должна составлять не менее 24 недель для подтипов 1a, 1c, 1d, 1е, а в случае обнаружения подтипа 1b, устойчивого к интерферону, - не менее 48 недель (Week К. Molecular methods of hepatitis C genotyping. Expert Rev Mol Diagn. 2005 Jul; 5(4):507-20).
Инфекция, вызванная ВГС с генотипом 4, дает клиническую картину, схожую с заражением вирусом, имеющим генотип 1 (Legrand-Abravanel F, Nicot F, Boulestin A, Sandres-Sauné K, Vinel JP, Alric L, Izopet J. Pegylated interferon and ribavirin therapy for chronic hepatitis С virus genotype 4 infection. J Med Virol. 2005 Sep; 77(1):66-9). При этом, в отличие от генотипа 1, для генотипа 4 характерно расщепление на значительно большее число подтипов (Nicot F, Legrand-Abravanel F, Sandres-Saune K, Boulestin A, Dubois M, Alric L, Vinel JP, Pasquier C, Izopet J. 2005. Heterogeneity of hepatitis С virus genotype 4 strains circulating in south-western France. J Gen Virol Jan; 86(Pt 1):107-14). Клиническая значимость ряда из них, например, 4а и 4d, уже установлена - 4d является устойчивым к интерферону и требует пролонгированного курса лечения (не менее 48 недель) (Roulot D, Bourcier V, Grando V с соавт. Epidemiological characteristics and response to peginterferon plus ribavirin treatment of hepatitis С virus genotype 4 infection. J Viral Hepat. 2007 Jul; 14(7):460-7).
Подтипы генотипов 2 и 3 ВГС являются чувствительными к терапии лекарственными препаратами и значительно реже приводят к хронизации процесса. Длительность курса терапии рибавирином и интерфероном для больных, инфицированных ВГС с данными генотипами, составляет от 6 до 12 недель.
В случае установления у пациента вируса гепатита С, имеющего генотип 5 или 6, рекомендован тот же курс терапии, что и для ВГС, имеющего генотип 1 (Pawlotsky JM. Therapy of hepatitis C: from empiricism to eradication. Hepatology. 2006 Feb; 43(2 Suppl 1):S207-20).
Помимо прогностических целей и оценки длительности терапии определение генотипа и подтипа дает информацию в плане этиологии инфекции. Субтипы 1а, 3а, 4а, 4d чаще ассоциируются с «шприцевым» гепатитом у лиц, употребляющих парэнтеральные наркотики, в то время как генотип 2 и субтип 1b связан с гемотрансфузионным путем передачи (Simmonds P, Bukh J, Combet С с соавт. Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology. 2005 Oct; 42(4):962-73).
Наконец, результаты, полученные с помощью способа, заявленного в настоящем изобретении, могут быть использованы для эпидемиологического генотипирования. Распределение генотипов и подтипов ВГС варьирует в различных географических регионах (Zein NN. Clinical significance of hepatitis C virus genotypes. Clin Microbiol Rev. 2000 13(2):223-35). Некоторые из подтипов являются универсальными, другие циркулируют лишь в пределах ограниченных географических зон. Подтип 1b превалирует на юге Европы, в Китае, Японии, России (50-80%). В США и странах Южной Америки, на севере Западной Европе чаще встречаются генотипы 1a и 1b, за ним следуют генотипы 2 и 3. Генотип 4 превалирует в Северной и Центральной Африке, в то время как на юге континента основным является генотип 5. Генотип 3 встречается почти всюду и является основным в Австралии и Юго-Восточной Азии. Генотип 6 также распространен в Юго-Восточной Азии и является основным типом во Вьетнаме, одним из основных в Таиланде, Индонезии.
Далее изобретение будет проиллюстрировано примерами, которые предназначены для обеспечения лучшего понимания сущности заявленного изобретения, но не должны рассматриваться как ограничивающие данное изобретение.
Примеры
Пример 1. Микрочип для идентификации генотипа и подтипа ВГС на основе анализа 5'-нетранслируемой области генома ВГС.
1. Олигонуклеотиды для иммобилизации на микрочипе и праймеры для амплификации были синтезированы на автоматическом синтезаторе 394 DNA/RNA synthesizer (Applied Biosystems, США) и содержали спейсер со свободной аминогруппой 3'-Ammo-Modifier C7 CPG 500 (Glen Research, США) для последующей иммобилизации в гель. Изготовление микрочипов производили по процедуре, описанной ранее (Rubina AY, Pan'kov SV, Dementieva EI et al. Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production. Anal Biochem 2004; 325: 92-106). Микрочипы содержали полусферические ячейки диаметром 100 мкм и отстоящие друг от друга на расстояние 300 мкм. Равномерность нанесения ячеек и их диаметр оценивали с помощью программного обеспечения 'Тест-чип' (Микрочип-ИМБ, Россия). Контроль качества микрочипов выполняли измерением концентраций иммобилизованных олигонуклеотидов. Микрочипы окрашивали флуоресцентным красителем ImD-310 (Микрочип-ИМБ), концентрацию иммобилизованных зондов оценивали, как было описано ранее (А.Л.Михейкин, А.В.Чудинов, А.И.Ярощук, А.Ю.Рубина, С.В.Паньков, А.С.Крылов, А.С.Заседателев, А.Д.Мирзабеков. Краситель с низкой специфичностью к нуклеотидной последовательности ДНК: применение для оценки количества олигонуклеотидов, иммобилизованных в ячейках биологических микрочипов. Молекулярная биология 2003; 37(6): 1061-70).
Структура микрочипа
Микрочип содержит 52 иммобилизованных олигонуклеотида, список которых представлен в таблице 1, четыре маркерные точки (М) для правильного позиционирования (захвата изображения), выполняемого компьютерной программой, и 3 ячейки пустого геля (0), необходимые для вычисления референсного (фонового) значения интенсивности флуоресценции Iref. Расположение иммобилизованных на микрочипе олигонуклеотидов показано на Фиг.3.
В двух верхних строках иммобилизованы олигонуклеотиды с индексом 'G', позволяющие идентифицировать генотип ВГС. Микрочип идентифицирует все шесть генотипов ВГС. Для надежной идентификации каждого генотипа сконструировано 2 уникальных генотип-специфичных олигонуклеотида.
Ниже иммобилизованы олигонуклеотиды, позволяющие идентифицировать подтипы:
- внутри генотипа 1:1a, 1b, 1d, 1e. Для надежной идентификации каждого из подтипов генотипа 1 сконструировано 2 группы олигонуклеотидов. Каждая группа соответствует отдельному участку внутри исследуемого фрагмента 5'НТР области;
- внутри генотипа 2: 2а, 2b, 2c, 2i, 2k. Для надежной идентификации каждого из подтипов генотипа 2 сконструировано 2 группы олигонуклеотидов. Каждая группа соответствует отдельному участку внутри исследуемого фрагмента 5'НТР области;
- внутри генотипа 3: 3а, 3b. Для надежной идентификации каждого из подтипов генотипа 3 сконструировано 2 группы олигонуклеотидов. Каждая группа соответствует отдельному участку внутри исследуемого фрагмента 5'НТР области;
- внутри генотипа 4: 4а, 4 с, 4d, 4f, 4h, 4k, 4r. Для надежной идентификации каждого из подтипов генотипа 4 сконструировано 2 группы олигонуклеотидов. Каждая группа соответствует отдельному участку внутри исследуемого фрагмента 5'НТР области;
- внутри генотипа 5: 5а. Генотип 5 представляет собой единственный подтип 5а, тем самым зонды, идентифицирующие генотип, выявляют и подтип 5а;
- внутри генотипа 6: 6а, 6b, 6d. Для надежной идентификации каждого из подтипов генотипа 6 сконструировано 2 группы олигонуклеотидов. Каждая группа соответствует отдельному участку внутри исследуемого фрагмента 5'НТР области.
Перечень последовательностей олигонуклеотидов, иммобилизованных на микрочипе.
Пример 2. Обратная транскрипция, совмещенная с амплификацией (ОТ-ПЦР) фрагмента 5'НТР области; получение одноцепочечных флуоресцентно меченых фрагментов методом асимметричной ПЦР.
На первой стадии проводили обратную транскрипцию, совмещенную с амплификацией (ОТ-ПЦР) фрагмента 5'НТР области длиной 418 п.о.
В 40 мкл смеси для ОТ-ПЦР вносили 10 мкл выделенного препарата РНК (конечный объем 50 мкл).
Состав ПЦР-смеси:
- 1X ОТ-ПЦР-буфер: 70 мМ Трис-HCl, рН 8,3, 16,6 мМ (NH4)2SO4, 7,5 мМ MgCl2;
- 200 мкМ каждого dATP, dCTP, dGTP, dUTP (Силекс, Россия);
- Смесь праймеров (последовательности представлены в Таблице 2) Pr3_f/ Pr2_r в концентрации 200 нМ каждый;
- 10 ед. термостабильной ST-полимеразы (Силекс, Россия);
- 1 ед. урацил-ДНК-гликозилазы (Силекс, Россия);
- 10 ед. ингибитора РНКазы (Fermentas, Литва).
Амплификацию проводили на термоциклере РТС-200 Dyad (MJ Research, США) со следующим режимом: обратная транскрипция при 50°С - 30 минут, затем 50 циклов ПЦР: 95°С - 30 с, 63°С - 30 с, 72°С - 30 с; финальная элонгация при 72°С - 10 минут.
Результат проведения стадии ОТ-ПЦР при использовании РНК ВГС, выделенной из клинических образцов, содержащих ВГС различных генотипов и подтипов, представлен на Фиг.4.
1 мкл реакционной смеси, полученной после первой стадии ПЦР, использовали в качестве матрицы при проведении второй стадии.
Вторую стадию ПЦР проводили в полугнездовом варианте, с праймерами P3_f/Pr5_r, фланкирующими район 5'НТР области длиной 382 п.о. Последовательности праймеров приведены в таблице 2.
Состав ПЦР-смеси (25 мкл):
- 1X ПЦР-буфер: 10 мМ КСl, 10 мМ Трис-НСl (рН 8,3) (Силекс, Россия);
- 1,5 мМ MgCl2;
- 200 мкМ каждого dNTP (Силекс, Россия);
- 10 мкМ флуоресцентно меченого dUTP («Микрочип-ИМБ», Россия);
- Смесь праймеров P3_f/Pr5_r в концентрации 20 нм/100 нМ, соответственно;
- 10 ед. термостабильной Taq ДНК-полимеразы (Силекс, Россия).
Амплификацию проводили на термоциклере РТС-200 Dyad (MJ Research, США) со следующим режимом: 95°С - 2 мин, затем 36 циклов: 95°С - 20 с, 60°С - 20 с, 72°С - 30 с, заключительная элонгация 72°С - 5 минут.
Результаты проведения стадии ПЦР при использовании в качестве матрицы продукта стадии ОТ-ПЦР, для образцов ВГС, имеющих различные генотипы и подтипы, представлены на Фиг.5.
12 мкл полученного продукта использовались в гибридизации на микрочипе.

Пример 3. Гибридизация амплифицированного меченого продукта на микрочипе.
К 12 мкл реакционной смеси, полученной после второй стадии ПЦР и содержавшей преимущественно одноцепочечные флуоресцентно меченые фрагменты ДНК, соответствующие исследуемому фрагменту 5'НТР области, добавляли концентрированный раствор гибридизационного буфера так, чтобы конечная концентрация гуанидина тиоцианата составляла 1 М, HEPES - 50 мМ, рН 7,5, ЭДТА - 5 мМ. Полученную смесь (32 мкл) помещали в гибридизационную камеру микрочипа. Гибридизацию проводили при 37°С в течение 12-18 часов. По окончании гибридизации микрочип трижды промывали дистиллированной водой при 37°С в течение 30 секунд и высушивали.
Пример 4. Регистрация и интерпретация результатов гибридизации
Регистрация флуоресцентного изображения микрочипа выполнялась на универсальном аппаратно-программном комплексе (УАПК) (ИМБ РАН, Россия) для анализа биологических микрочипов с использованием специализированного программного обеспечения 'ImageWare®' (Микрочип-ИМБ, Россия).
Интерпретацию результатов проводили согласно приведенному выше алгоритму, реализованному в модуле программного обеспечения для анализа флуоресцентных изображений микрочипов 'Imageware®'.
Пример 5. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 1а, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.6 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,22. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип и подтип-специфических ячейках. В результате было получено, что сигналы в ячейках группы G1, содержащих зонды, специфичные в отношении генотипа 1, превосходят Iref в 1,5 и более раза. Сигналы в других группах ячеек, содержащих генотип-специфичные зонды, были близки к фоновым. Далее, выбирается максимальный сигнал из группы Gl, G1-1 (5,7). Следовательно, последовательность РНК анализируемого образца относится к генотипу 1.
Анализ в группах ячеек, содержащих зонды, специфичные для подтипов генотипа 1, выявляет следующее: В группе 1 максимальные (т.е. превосходящие пороговое значение 1,5 по отношению к другим ячейкам) сигналы имеют 1а1 (16,7). В группе 2 - 1а2 (16,4). Таким образом, во всех группах максимальный сигнал имеют ячейки, содержащие зонды, специфичные в отношении подтипа 1а. Следовательно, анализируемый образец относится к подтипу 1а.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 1а.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 1 и подтип 1а, что полностью совпадает с результатами секвенирования.
Пример 6. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 1b, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.7 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,31. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип- и подтип-специфических ячейках. В результате было получено, что сигналы в ячейках группы G1, содержащих зонды, специфичные в отношении генотипа 1, превосходят Iref в 1,5 и более раза (максимальный сигнал имеет ячейка G1-2 (0,802)). Сигналы в других группах ячеек, содержащих генотип-специфичные зонды, были близки к фоновым. Следовательно, последовательность РНК анализируемого образца относится к генотипу 1.
Анализ в группах ячеек, содержащих зонды, специфичные для подтипов генотипа 1, выявляет следующее: В группе 1 максимальный (т.е. превосходящий пороговое значение 1,5 по отношению к другим ячейкам) сигнал имеет 1b1 (8,79). В группе 2 - 1b2 (2,58). Таким образом, в обеих группах совершенные гибридизационные дуплексы образовались в ячейках, содержащих олигонуклеотиды, специфичные к подтипу 1b. Следовательно, анализируемый образец относится к подтипу 1b.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа Ib.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 1 и подтип 1b, что полностью совпадает с результатами секвенирования.
Пример 7. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 1е, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.8 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,04. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип- и подтип-специфических ячейках. В результате было получено, что сигнал в ячейке G1-2 (0,632), содержащей зонд, специфичный в отношении генотипа 1, превосходит Iref в 1,5 и более раза. Также сигнал в ячейке G2-1 (0,121) проходит порог относительно Iref. Сигналы в других группах ячеек, содержащих генотип-специфичные зонды, близки к фоновым. Сигнал в ячейке G1-2 более чем в 3 раза превышает сигнал в ячейке G2-1. Следовательно, последовательность РНК анализируемого образца относится к генотипу 1.
Анализ в группах ячеек, содержащих зонды, специфичные для подтипов генотипа 1, выявляет следующее: В группе 1 максимальный (т.е. превосходящий пороговое значение 1,5 по отношению к другим ячейкам) сигнал имеет 1е1(4,18). В группе 2 сигналы ни в одной из ячеек не прошли порог относительно Iref. Таким образом, в группе 1 совершенный дуплекс с исследуемой ДНК формирует олигонуклеотид, последовательность которого является уникальной для подтипа 1е. Следовательно, анализируемый образец относится к подтипу 1е.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 1е.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 1 и подтип 1е, что полностью совпадает с результатами секвенирования.
Пример 8. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 2а, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.9 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,11. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип и подтип-специфических ячейках. В результате было получено, что сигналы в ячейках группы G2, содержащих зонды, специфичные в отношении генотипа 2, превосходят Iref в 1,5 и более раза. Максимальный сигнал в группе G2 имеет ячейка G2-2 (2,71). Сигналы в других группах ячеек, содержащих генотип-специфичные зонды, близки к фоновым. Следовательно, последовательность РНК анализируемого образца относится к генотипу 2.
Анализ в группах ячеек, содержащих зонды, специфичные для подтипов генотипа 2, выявляет следующее: В группе 1 максимальный (т.е. превосходящий пороговое значение 1,5 по отношению к другим ячейкам) сигнал имеет 2а1 (3,02). В группе 2 - 2а2 (2,01). В обеих группах максимальный сигнал имеют ячейки, содержащие зонды, специфичные в отношении подтипа 2а. Следовательно, анализируемый образец относится к подтипу 2а.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 2а.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 2 и подтип 2а, что полностью совпадает с результатами секвенирования.
Пример 9. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 2i, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.10 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,06. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип- и подтип-специфических ячейках. В результате было получено, что только сигналы в ячейках группы G2, содержащих зонды, специфичные в отношении генотипа 2, превосходят Iref в 1,5 и более раза.
Максимальный сигнал в группе G2 имеет ячейка G2-2 (1,559). Следовательно, последовательность РНК анализируемого образца относится к генотипу 2.
Анализ в группах ячеек, содержащих зонды, специфичные для подтипов генотипа 2, выявляет следующее: В группе 1 максимальный (т.е. превосходящий пороговое значение 1,5 по отношению к другим ячейкам) сигнал имеет 2il (0,187). В группе 2 - 2i2 (0,627). Таким образом, в двух группах максимальный сигнал принадлежит ячейкам, содержащим уникальные олигонуклеотиды, специфичные в отношении подтипа 2i. В соответствии с заявленным алгоритмом интерпретации анализируемый образец относится к подтипу 2i.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 2i.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 2 и подтип 2i, что полностью совпадает с результатами секвенирования.
Пример 10. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 3а, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.11 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,03. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип- и подтип-специфических ячейках. В результате было получено, что только сигнал в ячейке G3-1 (0,37), содержащей зонд, специфичный в отношении генотипа 3, превосходит Iref в 1,5 и более раза. Сигналы в других группах ячеек, содержащих генотип-специфичные зонды, близки к фоновым. Следовательно, последовательность РНК анализируемого образца относится к генотипу 3.
Анализ в группах ячеек, содержащих зонды, специфичные для подтипов генотипа 3, выявляет следующее: В группе 1 максимальный (т.е. превосходящий пороговое значение 1,5 по отношению к другим ячейкам) сигнал имеет ячейка 3а1 (3,51). В группе 2 - 3а2 (0,141). Таким образом, во всех группах максимальный сигнал имеют ячейки, содержащие зонды, специфичные в отношении подтипа 3а. Следовательно, анализируемый образец относится к подтипу 3а.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 3а.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 3 и подтип 3а, что полностью совпадает с результатами секвенирования.
Пример 11. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 4а, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.12 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,08. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип- и подтип-специфических ячейках. В результате было получено, что только сигналы в ячейках группы G4, содержащих зонды, специфичные в отношении генотипа 4, превосходят Iref в 1,5 и более раза. Сигналы в остальных ячейках, содержащих генотип-специфичные олигонуклеотиды, были близки к фоновым. Максимальный сигнал в группе G4 зарегистрирован в ячейке G4-1 (1,25). Следовательно, последовательность РНК анализируемого образца относится к генотипу 4.
Анализ в группах ячеек, содержащих зонды, специфичные для подтипов генотипа 4, выявляет следующее: в обеих группах зондов, специфичных к генотипу 4, максимальные сигналы имеют ячейки, содержащие зонды для выявления подтипа 4а: 4ас1 (1,71), 4а2 (0,2). Следовательно, в соответствии с описанным алгоритмом, анализируемый образец относится к подтипу 4а.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 4а.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 4 и подтип 4а, что полностью совпадает с результатами секвенирования.
Пример 12. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 4d, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.13 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,16. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип- и подтип-специфических ячейках. В результате было получено, что сигналы в ячейках группы G4, содержащих зонды, специфичные в отношении генотипа 4, превосходят Iref в 1,5 и более раза. Сигналы в остальных ячейках, содержащих генотип-специфичные олигонуклеотиды, были близки к фоновым. Максимальный сигнал в группе G4 имеет ячейка G4-2 (1,69). Следовательно, последовательность РНК анализируемого образца относится к генотипу 4.
Анализ в группах ячеек, содержащих зонды, специфичные для подтипов генотипа 4, выявляет следующее: в группе 1 максимальный сигнал имеет ячейка, содержащая олигонуклеотид, специфичный к подтипам 4d и 4f- 4df1 (1,69), в группе 2 - ячейка 4d2 (0,54). Во всех группах максимальный сигнал имеют ячейки, содержащие зонды, специфичные к подтипу 4d. Следовательно, анализируемый образец относится к подтипу 4d.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 4d.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 4 и подтип 4d, что полностью совпадает с результатами секвенирования.
Пример 13. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 5а, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.14 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,03. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип- и подтип-специфических ячейках. В результате получаем, что только сигналы в ячейках группы G5, содержащих зонды, специфичные в отношении генотипа 5, превосходят Iref в 1,5 и более раза - G5-1/G5-2 (0,35). Следовательно, последовательность РНК анализируемого образца относится к генотипу 5. Поскольку генотип 5 имеет всего один подтип, 5а, то делается заключение, что исследуемый образец имеет подтип 5а.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 5а.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 5 и подтип 5а, что полностью совпадает с результатами секвенирования.
Пример 14. Анализ 5'НТР области генома вируса гепатита С, имеющего подтип 6b, с использованием заявляемого способа.
Образец крови, полученный от пациента, был подвергнут обработке набором для выделения вирусной РНК Qiamp viral RNA (Qiagen, Германия) согласно протоколу фирмы-производителя. Выделенный препарат РНК был использован в ОТ-ПЦР, как описано в примере 2. По окончании ОТ-ПЦР наличие амплифицированного фрагмента заданной длины (418 п.о.) было протестировано электрофорезом в агарозном геле, после чего продукт ОТ-ПЦР был разделен на 2 части, одна из которых была обработана и проанализирована в соответствии с методикой, описанной в Примерах 2-4 (стадия ПЦР с последующей гибридизацией на микрочипе, отмывкой, регистрацией и интерпретацией флуоресцентной картины микрочипа). Вторую часть продукта первой стадии после дополнительной очистки использовали в реакции секвенирования с последующим разделением на автоматическом секвенаторе, получением и коррекцией хроматограммы, анализом обработанной последовательности 5'НТР фрагмента, конструированием множественного выравнивания и филогенетического дерева, на основании которых осуществляли установление генотипа и подтипа.
На фиг.15 представлены гибридизационная картина микрочипа и распределение нормированных гибридизационных сигналов ячеек микрочипа.
В соответствии с предложенным алгоритмом интерпретации результатов анализ начинается с вычисления усредненного сигнала (Iref) в ячейках, не содержащих олигонуклеотидов. В данном случае значение Iref составило 0,13. В соответствии с этой величиной и пороговым значением, равным 1,5, осуществляют фильтрацию сигналов в генотип и подтип-специфических ячейках. В результате получаем, что сигнал в ячейке группы G6-1 (10,54), содержащей зонд, специфичный в отношении генотипа 6, превосходят Iref в 1,5 и более раза. Сигнал в ячейке G3-1 (1,923) также следует считать достоверным. Ячейка G6 превосходит сигнал в ячейке G3-1 более чем в 1,5 раза, что означает принадлежность РНК анализируемого образца к генотипу 6.
Из двух групп, содержащих зонды, специфичные к подтипам генотипа 6, в ячейке 6b1 сигнал (1,147) является достоверным относительно Iref, превзошел пороговое значение. Следовательно, подтип данного образца ВГС классифицируется как 6b.
Методом секвенирования с последующим филогенетическим анализом было показано, что исследуемая последовательность попадает в кластер последовательностей подтипа 6b.
Таким образом, установлено, что анализируемый образец РНК ВГС имеет генотип 6 и подтип 6b, что полностью совпадает с результатами секвенирования.
Таким образом, представленное изобретение позволяет идентифицировать генотип и подтип вируса гепатита С на основе анализа 5'-нетраслируемой области генома ВГС с использованием олигонуклеотидного микрочипа. Способ позволяет идентифицировать все 6 генотипов и 22 подтипа вируса гепатита С, в том числе наиболее вирулентные и лекарственно-устойчивые формы. Способ выгодно отличается от существующих в настоящее время аналогов высокой специфичностью в отношении идентификации подтипов генотипа 1 и 4, в частности подтипов 1b и 4d, а также простотой выполнения и низкой себестоимостью. Данные, полученные с помощью процедуры гибридизации на микрочипах настоящего изобретения, могут быть использованы для прогнозирования и оценки тяжести протекания заболевания (острый/хронический цирроз, вероятность развития рака печени), определения терапевтической дозы лекарственных препаратов и длительности курса терапии, а также для эпидемиологического генотипирования.
Все патенты, публикации, научные статьи и другие документы и материалы, цитируемые или упоминаемые здесь, включены в настоящее описание путем отсылки в такой степени, как если бы каждый из этих документов был включен путем отсылки индивидуально или приведен здесь в его полном виде.
Хотя предпочтительные воплощения настоящего изобретения и их преимущества были подробно описаны выше, специалист в данной области сможет внести различные изменения, дополнения, не выходя при этом за рамки данного изобретения, которые определяются прилагаемой ниже формулой изобретения.














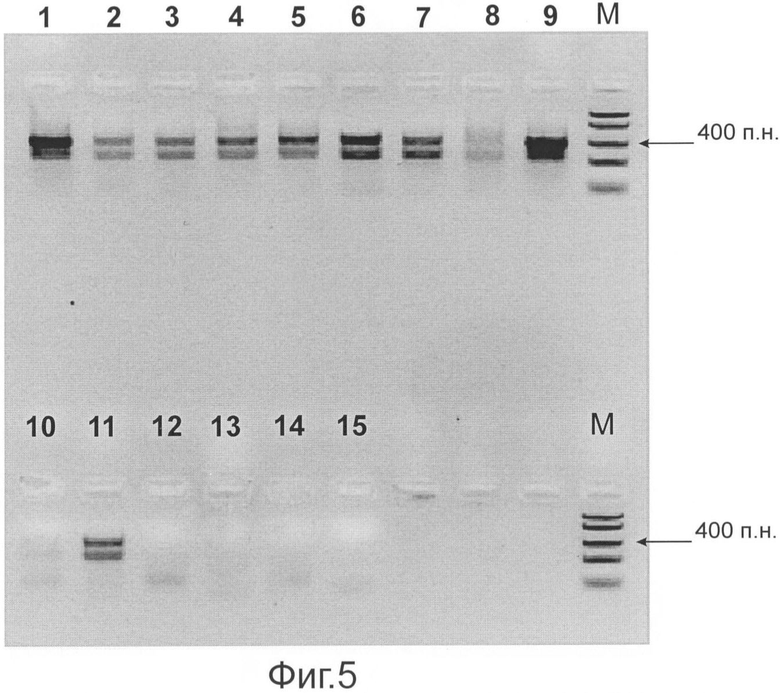



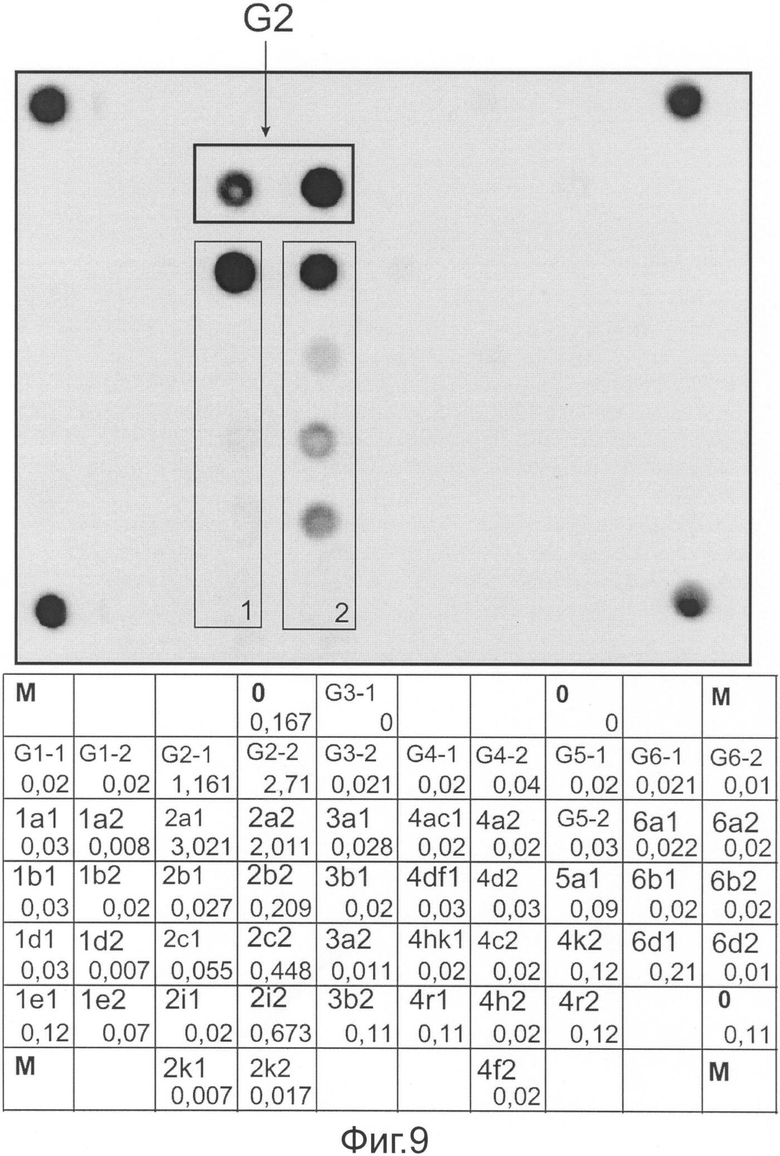

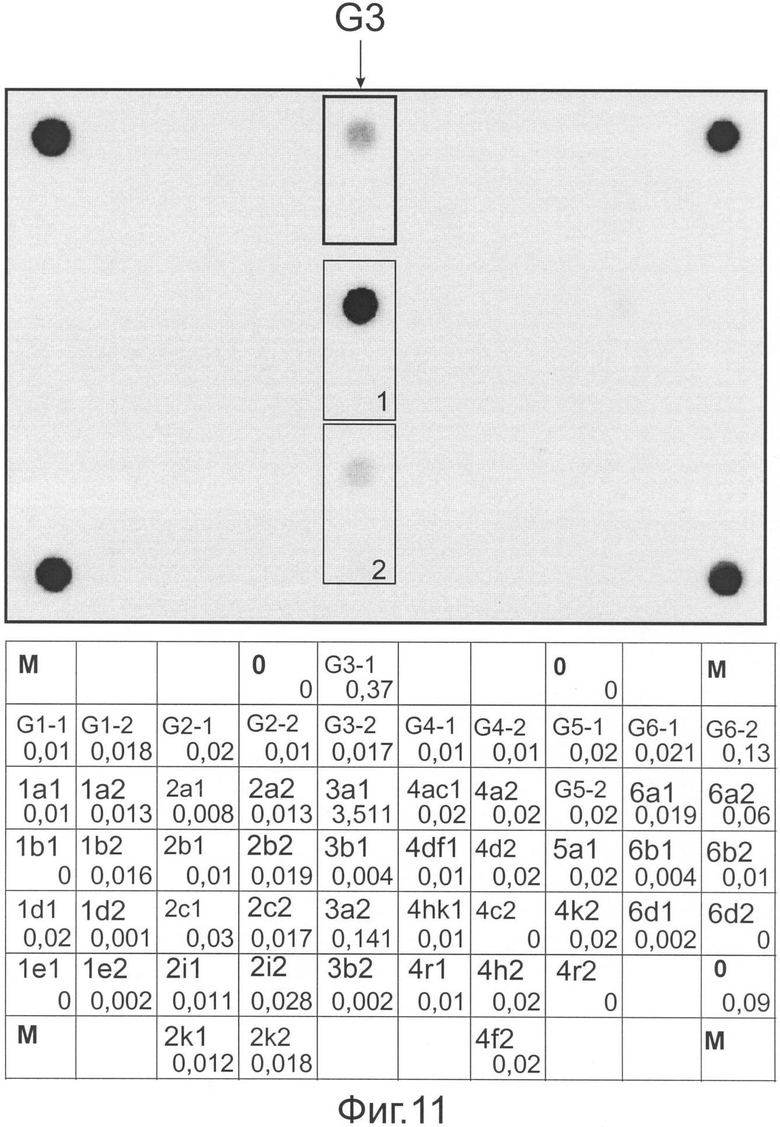

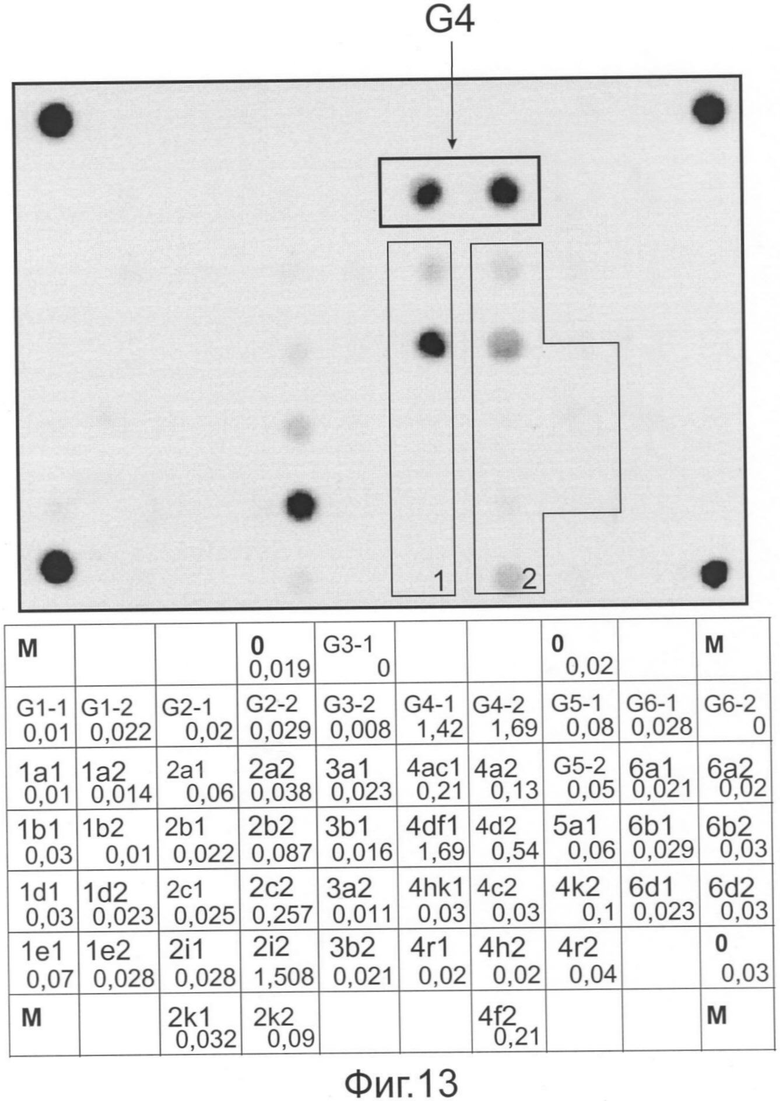

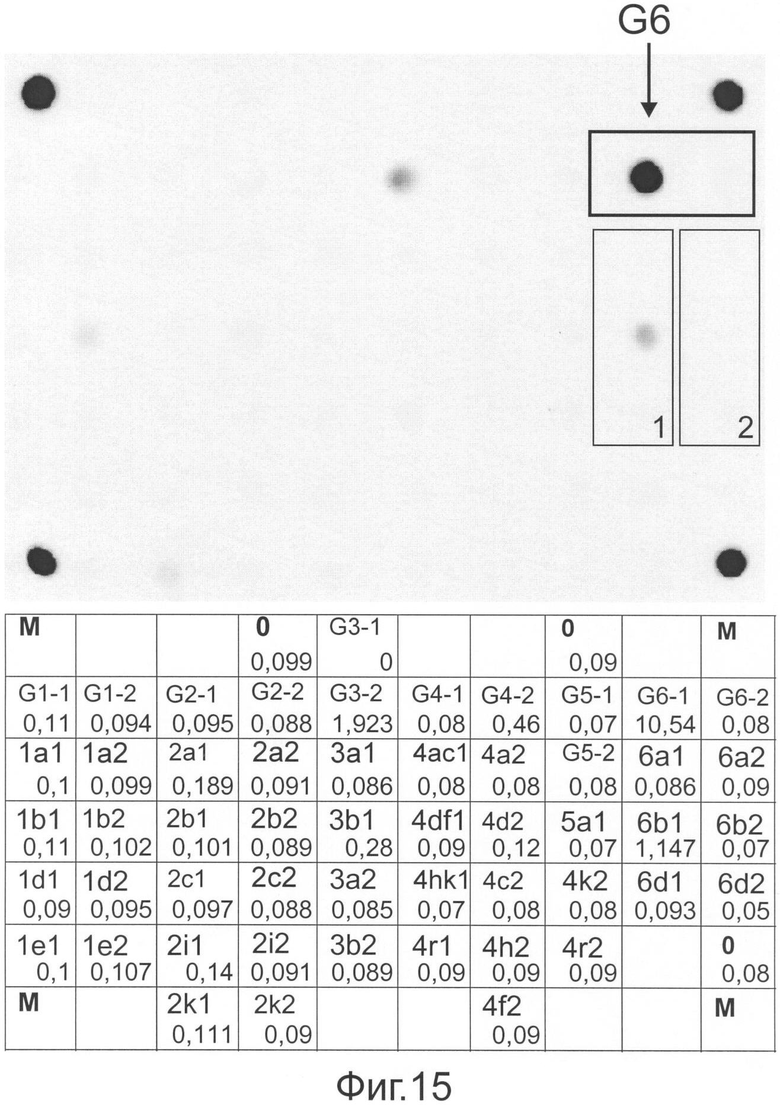
Изобретение относится к молекулярной биологии, вирусологии и медицине. Способ основан на анализе 5'-нетранслируемой области генома ВГС с использованием дифференцирующего олигонуклеотидного микрочипа. Способ предусматривает двухстадийную ПЦР с получением флуоресцентно меченого одноцепочечного фрагмента 5'-нетранслируемой области генома ВГС и последующую гибридизацию этого фрагмента на биологическом микрочипе, содержащем набор специфичных дискриминирующих олигонуклеотидов. Идентификацию генотипа и подтипа ВГС проводят путем определения специфичных последовательностей участков фрагмента 5'-нетранслируемой области. Изобретение позволяет проводить анализ непосредственно из клинического образца, определять 6 генотипов и 22 подтипа ВГС, в том числе наиболее вирулентные и лекарственно-устойчивые формы, снизить себестоимость анализа. Изобретение может быть использовано в медицине. 4 з.п. ф-лы, 15 ил., 2 табл.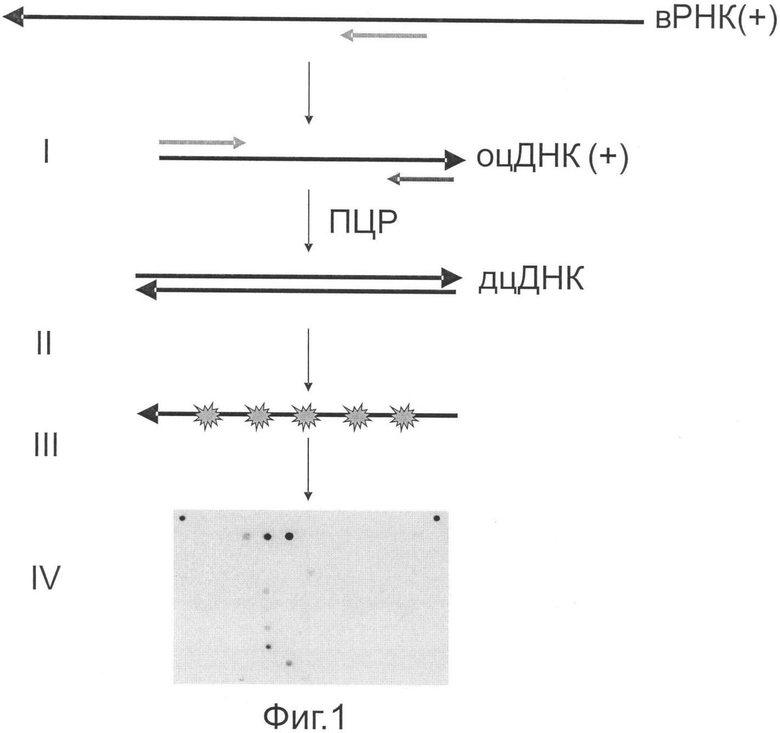
1. Способ идентификации генотипа и подтипа вируса гепатита С (ВГС) на основе анализа последовательностей 5'-нетранслируемой области генома ВГС, включающий:
(а) - обратную транскрипцию, совмещенную с ПЦР (ОТ-ПЦР), с использованием вирусной РНК в качестве матрицы и первой пары праймеров, специфичных к фрагменту 5'-нетранслируемой области генома ВГС, последовательности которых представлены SEQ ID NO:53 и 54;
(б) - асимметричную амплификацию фрагмента 5'-нетранслируемой области генома ВГС с использованием в качестве матрицы продукта ОТ-ПЦР, полученного на стадии (а), второй пары специфичных праймеров, последовательности которых представлены SEQ ID NO:55 и 56, и смеси четырех дезоксинуклеозидтрифосфатов, в которой один из четырех дезоксинуклеозидтрифосфатов является флуоресцентно меченым, в качестве субстрата, с получением преимущественно одноцепочечного флуоресцентно меченого фрагмента;
(в) - олигонуклеотидный микрочип для идентификации генотипа и подтипа ВГС, представляющий собой подложку, содержащую множество дискретных элементов, в каждом из которых иммобилизован уникальный олигонуклеотидный зонд, имеющий последовательность, комплементарную последовательности одноцепочечного фрагмента, полученного на стадии (б), и выбранную из группы, включающей: а) генотип-специфичные для каждого из генотипов ВГС последовательности фрагмента 5'-нетранслируемой области; и б) подтип-специфичные для каждого из подтипов ВГС последовательности фрагмента 5'-нетранслируемой области, при этом последовательности иммобилизованных олигонуклеотидных зондов представлены SEQ ID NO:1-52;
(г) гибридизацию амплифицированного меченого продукта, полученного на стадии (б), на олигонуклеотидном микрочипе с образованием дуплексов с иммобилизованными зондами в условиях, обеспечивающих разрешение в один нуклеотид между образующимися в результате гибридизации совершенными и несовершенными дуплексами;
(д) регистрацию результатов гибридизации на олигонуклеотидном микрочипе, проведенной на стадии (г), осуществляемую с помощью портативного анализатора флуоресценции и программного обеспечения, что позволяет использовать программную обработку интенсивностей сигналов с последующей интерпретацией результатов;
(е) интерпретацию результатов гибридизации, полученных на стадии (д), проводимую в два этапа: на первом этапе анализируют сигналы в элементах микрочипа, содержащих олигонуклеотидные зонды, специфичные к генотипам ВГС, тем самым идентифицируя генотип исследуемого образца; в случае идентификации генотипа на втором этапе анализируют только элементы микрочипа, содержащие олигонуклеотидные зонды, специфичные к подтипам идентифицированного генотипа, независимо от наличия сигналов в элементах, содержащих зонды, специфичные к подтипам других генотипов.
2. Способ по п.1, характеризующийся тем, что на стадии (б) один из праймеров второй пары используют, по меньшей мере, в десятикратном молярном избытке по отношению ко второму праймеру.
3. Способ по п.1, характеризующийся тем, что на стадии (б) в качестве флуоресцентно меченого дезоксинуклеозидтрифосфата используют флуоресцентно меченый дезоксиуридинтрифосфат.
4. Способ по п.1, характеризующийся тем, что олигонуклеотидный микрочип представляет собой микрочип на основе гидрогелевых ячеек, полученный способом химически или фото-индуцируемой сополимеризации.
5. Способ по п.1, характеризующийся тем, что он дополнительно предназначен для оценки и прогнозирования тяжести протекания заболевания (острый/хронический цирроз, вероятность развития рака печени), определения терапевтической дозы лекарственных препаратов и длительности курса терапии, и/или эпидемиологического генотипирования на основе проведенной интерпретации результатов гибридизации.
| GERMER J.J | |||
| et al | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| KREKULOVA L | |||
| et al., "Nested restriction site-specific PCR to detect and type hepatitis С virus (HCV): a rapid method to distinguish | |||

