Настоящее изобретение относится к способам и композициям, полезным для лечения и предупреждения инфекций, вызванных вирусом гепатита С (HCV), и симптомов и заболеваний, ассоциированных с ними. В частности, настоящее изобретение относится к ДНК-вакцинам, содержащим полинуклеотидные последовательности, кодирующие белки HCV, и способам лечения индивидуумов, инфицированных HCV, включающим введение вакцин по настоящему изобретению.
HCV был идентифицирован недавно как главный причинный агент посттрансфузионного и внебольничного заражения гепатитом "ни А, ни В". Приблизительно 170 млн людей хронически инфицированы HCV с частотой заболеваний от 1 до 10%. Расходы на медицинскую помощь в США, где частота заболеваний составляет 1,8%, оцениваются в 2 миллиарда долларов. 40-60% случаев заболеваний печени обусловлены HCV и 30% трансплантаций в Великобритании обусловлены HCV-инфекциями. Хотя изначально HCV-инфекция является бессимптомной, у более чем 90% пациентов развивается хроническое заболевание. Патологический процесс обычно развивается из хронического активного гепатита (70%), фиброза, цирроза (40%) в гепатоцеллюлярную карциному (60%). Средняя продолжительность процесса от инфицирования до развития цирроза или гепатоцеллюлярной карциномы составляет 20 лет (Lauer G. and Walker В. 2001, Hepatitis C virus Infection. N. Engl. J. Med. 345, 41, Cohen J. 2001. The Scientific challenge of Hepatitis C. Science 285 (5424) 26).
Существует большая потребность в улучшенном лечении HCV. В настоящее время нет доступных низкомолекулярных ингибиторов репликации. Широко распространенный золотой стандарт рибовирина и пегилированного интерферона является основой при лечении HCV-инфекции. Однако доступные в настоящее время схемы лечения дают недостаточную защитную реакцию (в целом уровень реакции составляет 50% вплоть до 6 месяцев, однако для генотипа 1b уровень реакции ниже (27%)). Кроме того, это лечение связано с неприятными побочными эффектами. Это приводит к значительному снижению эффективности, особенно после первых 6 месяцев лечения.
В нескольких исследованиях показано, что индивидуальные белки HCV являются иммуногенными в нормальных мышах, включая последующую иммунизацию с использованием ДНК. Несколько вакцин для профилактики или лечения HCV в настоящее время проходят клинические испытания. Наиболее перспективные, в которых используются оболочечные белки Е1 или Е2, в настоящее время находятся в фазе 2 в Chiron и Innogenetics. Эпитопная вакцина, разрабатываемая Transvax, также находится в фазе 2. Несколько вакцин, в которых используют последовательности из коровых (core) и неструктурных антигенов с применением разнообразных систем доставки, включая ДНК, находятся на стадии доклинической разработки.
HCV является вирусом из семейства флавивирусов с нитью РНК, имеющей положительную полярность, чей геном имеет длину 9,4 т.п.н. с одной открытой рамкой считывания. Геном HCV транслируется в виде единственного полипротеина, который затем подвергается процессингу под действием хозяйских и вирусных протеаз с образованием структурных белков (core, оболочечных Е1 и Е2, и р7) и шести неструктурных белков с различными ферментативными активностями. Геном изолята HCV J4L6, который является примером генотипа 1b, имеет регистрационный номер AF054247 (Yanagi M., St Claire M., Shapiro M., Emerson S.U., Purcell R.H. and Bukh J. "Transcripts of a chimeric cDNA clone of hepatitis С virus genotype 1b are infectious in vivo". Virology 244 (1), 161-172 (1998)) и показан на Фиг.1 (а-д).
Оболочечные белки отвечают за распознавание, связывание и проникновение вируса в клетки-мишени. Главные неструктурные белки, вовлеченные в репликацию вируса, включают NS2 (Zn-зависимую металлопротеиназу), NS3 (сериновую протеазу/геликазу), NS4A (кофактор протеазы), NS5A и NS5B (РНК-полимеразу) (Bartenschlager В and Lohmann V. 2000. Replication of hepatitis С virus. J. Gen. Virol. 81, 1631).
Структура полипротеина HCV может быть представлена следующим образом (цифры относятся к положению первой аминокислоты каждого белка; полноразмерный полипротеин изолята J4L6 включает 3010 аминокислот):
Вирус обладает высокой скоростью мутаций, и на основе нуклеотидной последовательности консервативных и неконсервативных областей было определено по меньшей мере шесть основных генотипов. Однако существует дополнительная гетерогенность, поскольку HCV, выделенный из одного пациента, всегда представляет собой смесь близкородственных геномов, или квази-видов.
Геном HCV демонстрирует высокую степень генетической изменчивости и был разделен на 6 основных генотипов (1а, 1b, 2, 3, 4, 5 и 6). Генотипы 1а, 1b, 2 и 3 наиболее распространены в Европе, Северной и Южной Америке, Азии, Китае, Японии и Австралии. Генотипы 4 и 5 преобладают в Африке, а генотип 6 - в Юго-Восточной Азии.
Поэтому существует большая потребность в улучшенных способах лечения HCV-инфекции, а также в обеспечении способов лечения, различающихся по способности лечить некоторые генотипы HCV. В первом аспекте настоящего изобретения предложены новые вакцинные препараты, различающиеся по их защитному действию в отношении различных генотипов.
Описаны вакцины против HCV, содержащие полинуклеотиды, кодирующие один или более чем один белок HCV. Вакцины, содержащие плазмидную ДНК или Semliki Forest Virus векторы, кодирующие NS3, описаны Brinster с соавт. (2002, Journal of General Virology, 83, 369-381). Полинуклеотидные вакцины, кодирующие NS5B, раскрыты в WO 99/51781. Кодон-оптимизированные гены и содержащие их вакцины, кодирующие E1, слитые белки Е1+Е2, белки NS5A и NS5B HCV, описаны в WO 97/47358. WO 01/04149 раскрывает полипептиды или полинуклеотиды, кодирующие мозаичные эпитопы HCV, происходящие из core, NS3, NS4 или NS5A. Слитые белки и ДНК, кодирующая такие слитые белки, содержащие NS3, NS4, NS5A и NS5B, которые полезны для получения вакцин, описаны в WO 01/30812; полагают, что слитые белки могут включать фрагменты core-белка. WO 03/031588 описывает пригодный для использования в качестве вакцины аденовирусный вектор, который кодирует белки NS3-NS4A-NS4B-NS5A-NS5B HCV.
В WO 96/37606 описаны вакцины, содержащие полипептиды, включающие "непроцессированный" core-белок и неструктурный белок.
Настоящее изобретение относится к получению полинуклеотидной вакцины, которая кодирует белки HCV: core, NS3, NS4B и NS5B. Полинуклеотидные вакцины по настоящему изобретению не кодируют белок NS4A HCV и/или белок NS5A. Предпочтительно полинуклеотидные вакцины по настоящему изобретению кодируют белки HCV: core, NS3, NS4B и NS5B, но не другие белки HCV. В настоящем изобретении также предложено применение полинуклеотидной вакцины, кодирующей эти антигены, в медицине и в изготовлении лекарства для лечения или предупреждения HCV-инфекции.
Полинуклеотидные последовательности, используемые в вакцинах по настоящему изобретению, представляют собой предпочтительно последовательности ДНК.
Полинуклеотиды, кодирующие белки HCV, могут быть во многих комбинациях или конфигурациях. Например, белки могут быть экспрессированы в виде индивидуальных белков или слитых белков. Примером слияния, которое может происходить как на уровне ДНК, так и на уровне белка, обычно является двойное слияние, состоящее из одного полипептида или полинуклеотида, содержащего или кодирующего аминокислотные последовательности NS4B и NS5B (NS4B-NS5B); тройное слияние, содержащее или кодирующее аминокислотные последовательности NS3-NS4B-NS5B, или слияние всех четырех антигенов по настоящему изобретению (Core-NS3-NS4B-NS5B).
Предпочтительные слияния по настоящему изобретению представляют собой полинуклеотиды, кодирующие двойное слияние между NS4B и NS5B (NS4B-NS5B или NS5B-NS4B); и между core и NS3 (NS3-Core или Core-NS3). Предпочтительные тройные слияния представляют собой полинуклеотиды, кодирующие аминокислотные последовательности NS3-NS4B-NS5B.
Полинуклеотиды по настоящему изобретению, кодирующие единичные антигены или слитые белки, могут быть представлены в виде единичных или множественных экспрессирующих векторов. Предпочтительно полинуклеотиды, кодирующие каждый антиген, представлены в одном и том же экспрессирующем векторе или плазмиде. В данном контексте полинуклеотиды, кодирующие белки HCV, могут находиться в единичной экспрессионной кассете или во множественных последовательно соединенных экспрессионных кассетах.
Для оптимизации экспрессии других белков HCV полинуклеотид, кодирующий core-белок HCV, предпочтительно находится в экспрессионной кассете, которая расположена в прямом направлении относительно экспрессионной кассеты, содержащей полинуклеотид, кодирующий по меньшей мере один другой белок HCV. Предпочтительно core-белок HCV находится в экспрессионной кассете, которая расположена в прямом направлении относительно экспрессионной кассеты, содержащей полинуклеотид, кодирующий NS5B.
Полипептиды, кодируемые олигонуклеотидными вакцинами по настоящему изобретению, могут содержать полноразмерную аминокислотную последовательность или, альтернативно, данные полипептиды могут быть короче, чем полноразмерные белки, в том смысле, что они содержат достаточную часть полноразмерной полинуклеотидной последовательности, которая позволяет экспрессировать продукт укороченного гена с целью получения иммунного ответа, дающего перекрестную реакцию с полноразмерным белком. Например, полинуклеотид по изобретению может кодировать фрагмент белка HCV, представляющий собой укороченный белок HCV, в котором участки исходной последовательности были делегированы, при этом конечный фрагмент содержит менее 90% исходной полноразмерной аминокислотной последовательности и, возможно, менее 70% или менее 50% исходной последовательности. Иными словами, считают, что полинуклеотид, кодирующий фрагмент длиной по меньшей мере 8, например 8-10 аминокислот или вплоть до 20, 50, 60, 70, 80, 100, 150 или 200 аминокислот, находится в пределах объема данного изобретения до тех пор, пока кодируемый им олиго- или полипептид демонстрирует HCV-антигенность. В частности, но не исключительно, данный аспект изобретения охватывает ситуацию, когда полинуклеотид кодирует фрагмент полной белковой последовательности HCV и может представлять один или более различных эпитопов данного белка.
В предпочтительных вакцинах по настоящему изобретению по меньшей мере один и предпочтительно все полипептиды HCV инактивированы в результате укорочения или мутации. Например, геликазная и протеазная активности NS3 предпочтительно снижены или аннулированы мутацией в соответствующем гене. Предпочтительно NS5В-полимеразная активность экспрессируемого полипептида снижена или аннулирована в результате мутации. Предпочтительно NS4В-активность экспрессируемого полипептида снижена или аннулирована в результате мутации. Предпочтительно активность core-белка экспрессируемого полипептида снижена или аннулирована в результате укорочения или мутации. В этом смысле мутация может быть результатом вставки, делеции, замены или перестройки в полинуклеотиде, кодирующем полипептид. Альтернативно, полноразмерная последовательность может быть экспрессирована в виде двух или более отдельных частей.
Функциональная структура и ферментативная функция полипептидов NS3 и NS5B HCV описаны в данной области техники.
NS5B был описан как РНК-зависимая РНК-полимераза (Qin et al., 2001, Hepatology, 33, р. 728-737; Lohmann et al., 2000, Journal of Viral Hepatitis; Lohmann et al., 1997, Nov., Journal of Virology, 8416-8428; De Francesco et al., 2000, Seminars in Liver Disease, 20(1), 69-83). Было показано, что полипептид NS5B имеет четыре функциональных мотива А, В, С и D.
Предпочтительно последовательность полипептида NS5B, кодируемую полинуклеотидными вакцинами по настоящему изобретению, подвергают мутации для того, что уменьшить или удалить РНК-зависимую РНК-полимеразную активность. Предпочтительно этот полипептид подвергают мутациям с целью нарушения мотива А NS5B, например, путем замены аспарагиновой кислоты (D) в положении 2639 на глицин (G); или замены аспарагиновой кислоты (D) в положении 2644 на глицин (G). Предпочтительно полипептид NS5B, кодируемый вакцинным полинуклеотидом, содержит обе эти мутации по аспарагиновой кислоте.
Предпочтительно кодируемый NS5B содержит нарушение в своем мотиве С. Например, мутация D2737, инвариантного остатка аспарагиновой кислоты, на Н, N или Е приводит к полной инактивации NS5B.
Предпочтительно NS5B, кодируемый ДНК-вакцинами по настоящему изобретению, содержит мутацию в мотиве А, который может возможно содержать мутацию в мотиве С. Предпочтительные мутации в мотиве А включают замену аспарагиновой кислоты (D) 2639 на глицин и аспарагиновой кислоты (D) 2644 на глицин. Предпочтительно присутствуют обе мутации. Кроме того, могут присутствовать дополнительные консенсусные мутации, как изложено ниже в Примере 1.
Было описано, что NS3 имеет и протеазную, и геликазную активность. Предпочтительно полипептиды NS3, кодируемые ДНК-вакцинами по настоящему изобретению, имеют мутации, нарушающие и протеазную, и геликазную активности NS3. Известно, что протеазная активность NS3 связана с "каталитической триадой" Н-1083, D-1107 и S-1165. Предпочтительно NS3, кодируемый вакцинами по настоящему изобретению, содержит мутацию в остатках каталитической триады, и наиболее предпочтительно NS3 содержит единичную точечную мутацию с заменой серина 1165 на валин (De Francesco R., Pessi, A and Steinkuhler С. 1998. The hepatitis С Virus NS3 proteinase: structure and function of zinc containing proteinase. Anti-Viral Therapy 3, 1-18).
Структура и функция NS3 могут быть представлены следующим образом:
Было идентифицировано четыре важных мотива, ответственных за геликазную активность NS3 - I, II, III и IV. Предпочтительно NS3, кодируемый ДНК-вакцинами по настоящему изобретению, содержит нарушающие мутации по меньшей мере в одном из этих мотивов. Более предпочтительно, когда имеется замена аспарагиновой кислоты 1316 на глутамин (Paolini С., Lahm A., De Francesco R. and Gallinari P. 2000, Mutational analysis of hepatitis С virus NS3-associated helicase. J. Gen. Virol. 81, 1649). Ни одна из этих наиболее предпочтительных мутаций NS3, S1165V или D1316Q не находится в пределах известных или предсказанных Т-клеточных эпитопов.
Наиболее предпочтительно, когда полипептид NS3, кодируемый ДНК-вакцинами по настоящему изобретению, содержит мутацию: серин (S) 1165 на валин (V) и аспарагиновая кислота (D) 1316 на глутамин (Q). Дополнительно может присутствовать одна или более чем одна консенсусная мутация, как изложено в Примере 1.
Биологические функции core-белка HCV являются сложными и не коррелируют с отдельными точечными мутациями (McLauchlan J. 2000. Properties of the hepatitis С virus core protein: a structural protein that modulate cellular processes. J. of Viral Hepatitis 7, 2-4). Имеются доказательства того, что core непосредственно взаимодействует с рецептором лимфотоксина β, а также может вмешиваться в NFkB- и PKR-пути и может оказывать влияние на клеточное выживание и апоптоз. Обнаружили, что рекомбинантная конструкция на основе вируса коровьей оспы, экспрессирующая core, ингибирует клеточные ответы на вирус коровьей оспы, делая его более вирулентным in vivo.
Во время инфицирования core-белок из вирусного полипротеина расщепляется по двум сайтам под действием протеаз клетки-хозяина. Первое расщепление, которое происходит в положении 191, дает N-терминальный конец Е1. Остаток, по которому происходит второе расщепление, точно не установлен и находится в интервале между аминокислотами 174 и 191, в результате чего высвобождается короткая последовательность core-пептида длиной приблизительно 17 аминокислот (McLauchlan J. (2000) J. Viral Hepatitis. 7, 2-14; Yasui К, Lau JYN, Mizokami M., et al., J. Virol. 1998. 726048-6055).
Core-полипептиды, используемые в вакцинах по настоящему изобретению, находятся как в полноразмерной, так и в укороченной форме. Core-полипептид может быть полноразмерным, но его последовательность перегруппирована таким образом, чтобы аннулировать какую-либо активность core-белка. Core-полипептид может быть расщеплен по меньшей мере на два фрагмента и наиболее предпочтительно образует полипептид, содержащий core-аминокислоты 66-191, следующие за аминокислотами 1-65, и, альтернативно, core-аминокислоты 105-191, следующие за core-аминокислотами 1-104.
Для минимизации отрицательного эффекта core на продукцию других белков HCV в той же клетке наиболее предпочтительно, чтобы используемый core-белок представлял собой укороченный белок. В предпочтительном аспекте настоящего изобретения кодируемый core-белок укорачивают с карбокситерминального конца на величину, достаточную для уменьшения ингибиторного эффекта core на экспрессию других белков HCV. Наиболее предпочтительно core-белок укорачивают с карбокситерминального конца таким образом, что в последовательности продуцируемого белка отсутствует высвобождающаяся в природных условиях С-концевая пептидная последовательность, появляющаяся в результате второго расщепления core; более предпочтительно, когда в белке отсутствуют по меньшей мере последние 10 аминокислот, предпочтительно отсутствуют по меньшей мере последние 15 аминокислот, более предпочтительно отсутствуют последние 20 аминокислот, более предпочтительно отсутствуют последние 26 аминокислот и наиболее предпочтительно отсутствуют последние 40 аминокислот. Наиболее предпочтительными полинуклеотидами, кодирующими core, которые подходят для использования в настоящем изобретении, являются те, которые кодируют укороченный core, содержащий аминокислоты 1-171, 1-165, 1-151. Наиболее предпочтительным полинуклеотидом, кодирующим core, который подходит для использования в настоящем изобретении, является такой, который кодирует укороченный core-белок между аминокислотами 1-151. Может присутствовать одна или более чем одна консенсусная мутация, как изложено в Примере 1.
Предпочтительный полипептид NS4B, кодируемый полинуклеотидами по настоящему изобретению, содержит N-терминальное укорочение для удаления участка, который является гипервариабельным среди изолятов и генотипов HCV. Предпочтительно полипептид NS4B содержит делецию 30-100 аминокислот с N-конца, более предпочтительно 40-80 аминокислот и наиболее предпочтительно делецию первых 48 N-концевых аминокислот (в контексте изолята J4L6 это соответствует укорочению по аминокислоте 1760, что означает потерю первых 48 аминокислот NS4B; эквивалентные укорочения в других изолятах HCV также составляют часть настоящего изобретения). Кроме того, последовательность NS4B может быть разделена на два или более фрагментов и экспрессирована с образованием полипептида, имеющего последовательность NS4B, организованную в другом порядке по сравнению с последовательностью, обнаруженной в молекуле дикого типа.
Полинуклеотиды, которые присутствуют в вакцинах по настоящему изобретению, могут содержать природную нуклеотидную последовательность, которая обнаружена у вируса HCV, однако предпочтительно, чтобы нуклеотидная последовательность была кодон-оптимизированной для экспрессии в клетках млекопитающих.
В дополнение к кодоновой оптимизации предпочтительно, чтобы использование кодонов в полинуклеотидах по настоящему изобретению, кодирующих core, NS3, NS4B и NS5B HCV, было изменено таким образом, чтобы редкие кодоны не появлялись в концентрированных кластерах и были, наоборот, либо относительно случайно распределены по всей полинуклеотидной последовательности, либо исключены из кодон-оптимизированного гена.
Код ДНК состоит из 4 букв (А, Т, С и G) и использует их для составления трехбуквенных "кодонов", которые отображают аминокислоты белков, закодированных в генах организма. Линейная последовательность кодонов вдоль молекулы ДНК транслируется в линейную последовательность аминокислот в белке(ах), кодируемом этими генами. Код является сильно вырожденным с 61 кодоном, кодирующим 20 природных аминокислот, и 3 кодонами, представляющими "стоп"-сигналы. Таким образом, большинство аминокислот кодируется более чем одним кодоном - фактически некоторые кодируются четырьмя или более различными кодонами.
Отмечено, что когда в распоряжении имеется более чем один кодон для кодирования данной аминокислоты, то картины использования кодонов в организмах являются далеко не случайными. Различные виды демонстрируют различное отклонение в выборе ими кодонов, и, более того, использование кодонов может заметно различаться в пределах одного вида между генами, которые экспрессируются с высокими и низкими уровнями. Такое отклонение различно в вирусах, растениях, бактериях и клетках млекопитающих, и некоторые виды демонстрируют более строгое, по сравнению со случайным, отклонение в выборе кодонов, чем другие виды. Например, у людей и других млекопитающих это отклонение является менее строгим, чем у некоторых видов бактерий или вирусов. По этим причинам существует большая вероятность того, что ген млекопитающих, экспрессируемый в E. coli, или вирусный ген, экспрессируемый в клетках млекопитающих, будет иметь неподходящее для эффективной экспрессии распределение кодонов. Однако ген с картиной использования кодонов, подходящей для экспрессии в Е. coli, также может эффективно экспрессироваться в клетках человека. Считают, что присутствие в гетерологической последовательности ДНК кластеров кодонов, которые редко наблюдаются у хозяина, в клетках которого происходит экспрессия, является прогностическим признаком низких уровней гетерологичной экспрессии в клетках этого хозяина.
Существует несколько примеров, в которых замена кодонов, которые редко встречаются в клетках хозяина, на кодоны, которые предпочитаются хозяином ("оптимизация кодонов"), усиливала уровни гетерологичной экспрессии, например поздние гены L1 и L2 BPV (вирус папилломы быка) были подвергнуты оптимизации кодонов относительно картин использования кодонов у млекопитающих, и было показано, что это приводит к повышенным уровням экспрессии в культуре клеток млекопитающих (Cos-1) по сравнению с последовательностями BPV дикого типа (Zhou et. al. J. Virol. 1999. 73, 4972-4982). В этой работе каждый кодон BPV, который присутствовал более чем в два раза чаще в BPV, чем в клетках млекопитающих (коэффициент использования более 2), и большинство кодонов с коэффициентом использования более 1,5 были консервативно заменены кодоном, предпочтительно используемым у млекопитающих. В WO 97/31115, WO 97/48370 и WO 98/34640 (Merk&Co., Inc.) было показано, что кодоновая оптимизация генов или сегментов генов ВИЧ приводит к усилению экспрессии белка и улучшенной иммуногенности в том случае, когда кодон-оптимизированные последовательности используются в качестве ДНК-вакцин в млекопитающем-хозяине, для которого разрабатывалась оптимизация. Согласно этим документам последовательности полностью состояли из оптимизированных кодонов (за исключением тех случаев, когда это привело бы к образованию нежелательного сайта рестрикции, сайта сплайсинга интрона и т.д.), так как каждый вирусный кодон консервативно заменяли оптимальным кодоном для предполагаемого хозяина.
Термин "картина использования кодонов" относится к средним частотам для всех кодонов в рассматриваемых нуклеотидной последовательности, гене или классе генов (например высокоэкспрессируемых генах млекопитающих). Картину использования кодонов для млекопитающих, включая людей, можно найти в литературе (см., например, Nakamura et al. Nucleic Acids Research 1996, 24: 214-215).
В полинуклеотидах по настоящему изобретению картина использования кодонов предпочтительно отличается от таковой, типичной для HCV, тем, что чаще представлено отклонение в использовании кодонов для организма-мишени, например E. coli или млекопитающего, в частности человека. "Коэффициент использования кодонов" или индекс адаптации кодонов (Sharp P.M., Li W.H. Nucleic Acids Research. 15 (3): 1281-95, 1987) является мерой того, насколько близко картина использования кодонов данной полинуклеотидной последовательности напоминает таковую в видах-мишенях. Частоты встречаемости кодонов для каждого из 61 кодона (выраженные как количество случаев наблюдения на 1000 кодонов для выбранного класса генов) нормируют для каждой из двадцати природных аминокислот таким образом, что величину для наиболее часто используемого кодона для каждой аминокислоты принимают за 1, а частоты для менее распространенных кодонов пропорционально наносят на шкалу в интервале от нуля до 1. Таким образом, каждому из 61 кодона присваивают величину, равную 1 или ниже для высокоэкспрессируемых генов видов-мишеней. Эту величину обозначают как показатель предпочтения (W). Для того чтобы подсчитать коэффициент использования кодонов для конкретного полинуклеотида относительно высокоэкспрессируемых генов этого вида, отмечают нормированную величину для каждого кодона конкретного полинуклеотида и берут среднее геометрическое всех этих величин (путем деления суммы натуральных логарифмов этих величин на общее количество кодонов и взятия антилогарифма). Этот коэффициент будет иметь величину в интервале от нуля до 1, и чем выше коэффициент, тем больше кодонов в полинуклеотиде являются часто используемыми кодонами. Если полинуклеотидная последовательность имеет коэффициент использования кодонов, равный 1, то все кодоны являются "наиболее частыми" кодонами для высокоэкспрессируемых генов видов-мишеней.
Согласно настоящему изобретению предложены полинуклеотидные последовательности, кодирующие аминокислотные последовательности core, NS3, NS4B или NS5B HCV, где картина использования кодонов полинуклеотидной последовательности похожа на таковую для высокоэкспрессируемых генов млекопитающих. Предпочтительно полинуклеотидная последовательность является последовательностью ДНК. Желательно, чтобы картина использования кодонов полинуклеотидной последовательности была похожа на таковую для высокоэкспрессируемых генов человека.
Кодон-оптимизированная полинуклеотидная последовательность, кодирующая core HCV (1-191), показана на Фиг.2. Кодон-оптимизированная полинуклеотидная последовательность, кодирующая NS3 HCV, содержащая полипептидные мутации S1165V и D1316Q, показана на Фиг.3. Кодон-оптимизированная полинуклеотидная последовательность, кодирующая NS4B HCV, содержащая N-концевое укорочение 1-48 полипептида, показана на Фиг.4. Кодон-оптимизированная полинуклеотидная последовательность, кодирующая NS5B HCV, содержащая полипептидные мутации D2639G и D2644G, показана на Фиг.5.
Таким образом, предложен синтетический ген, содержащий большое количество кодонов, совместно кодирующих аминокислотные последовательности core, NS3, NS4B или NS5B HCV, где выбор возможных кодонов, используемых для кодирования аминокислотной последовательности, изменен таким образом, чтобы иметь сходство с оптимальным вариантом использования кодонов у млекопитающих, так что частота использования кодонов в синтетическом гене более близка к таковой у высокоэкспрессируемых генов млекопитающих, чем у генов вируса гепатита С. Предпочтительно картина использования кодонов по существу такая же, как у высокоэкспрессируемых генов человека. "Природные" последовательности core, NS3, NS4B и NS5B HCV были проанализированы на предмет использования кодонов. Коэффициент использования кодонов для белков HCV составляет: core (0,487), NS3 (0,482), NS4B (0,481) и NS5B (0,459). Полинуклеотид по настоящему изобретению в общем случае будет иметь коэффициент использования кодонов (как определено выше) для высокоэкспрессируемых генов человека более чем 0,5, предпочтительно более чем 0,6, наиболее предпочтительно более чем 0,7, но менее 1. Желательно, когда полинуклеотид будет также иметь коэффициент использования кодонов для высокоэкспрессируемых генов E. coli более чем 0,5, предпочтительно более чем 0,6, наиболее предпочтительно более чем 0,7.
В дополнение к кодоновой оптимизации синтетические гены также подвергают мутациям таким образом, чтобы исключить появление кластеров редких кодонов. Это может быть достигнуто одним из двух путей. Предпочтительный путь достижения этого заключается в исключении редких кодонов из генной последовательности. Согласно одному способу определения редкими кодонами будут кодоны, представляющие менее 20% кодонов, используемых для конкретной аминокислоты, и предпочтительно менее 10% кодонов, используемых для конкретной аминокислоты в высокоэкспрессируемых генах организма-мишени. Альтернативно, редкие кодоны могут быть определены как кодоны с относительной величиной использования синонимичных кодонов (RSCU) менее 0,3 или предпочтительно менее 0,2 в высокоэкспрессируемых генах организма-мишени. Величина RSCU представляет собой наблюдаемое количество кодонов, деленное на ожидаемое количество, если все кодоны для данной аминокислоты были использованы с одинаковой частотой. Соответствующее определение редкого кодона будет очевидно для специалиста в данной области техники.
Альтернативно, полинуклеотиды core, NS3, NS4B и NS5B HCV оптимизируют таким образом, чтобы предотвратить кластеризацию редких, неоптимальных кодонов, присутствующих в концентрированных областях. Поэтому полинуклеотиды оптимизируют таким образом, чтобы отдельные редкие кодоны, такие как кодоны с RSCU менее 0,4 (и более предпочтительно с RSCU менее 0,3) равномерно располагались на всем протяжении полинуклеотидов.
Было показано, что уровни экспрессии кодон-оптимизированных мутантных core, NS3 и NS5B увеличены по сравнению с диким типом, как определено вестерн-блоттингом. Укороченный кодон-оптимизированный NS4B был экспрессирован в виде слияния с NS5B, и это слияние экспрессируется хорошо.
Вакцины по настоящему изобретению могут включать вектор, который направляет индивидуальную экспрессию полипептидов HCV, альтернативно полипептиды HCV могут быть экспрессированы в виде одного или более слитых белков.
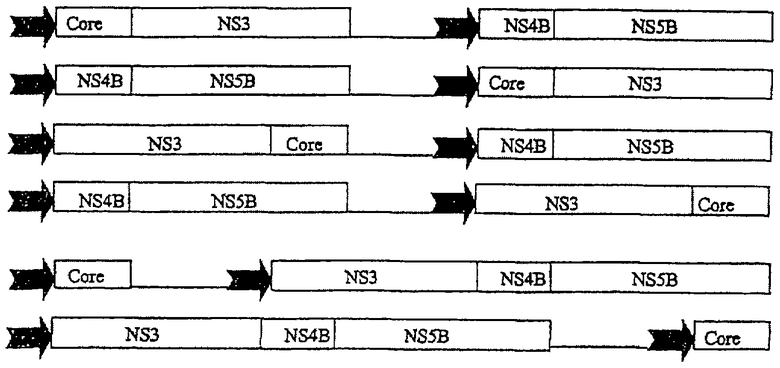
Предпочтительные вакцины по настоящему изобретению включают тетра-слияния либо на уровне белков, либо на уровне полинуклеотидов, в том числе:

Другие предпочтительные слияния аналогичны HCV-комбинациям 1, 2 и 3, но в них core-белок представляет собой укороченный core-белок, типично Core1-151. Другие предпочтительные вакцины по настоящему изобретению представлены ниже и включают полинуклеотидные двойные и тройные слияния, представленные в различных экспрессионных кассетах в пределах одной плазмиды, при этом каждая кассета находится под контролем независимой промоторной единицы (например, IE HCMV) (показано стрелкой). Такие двойные промоторные конструкции управляют экспрессией четырех белковых антигенов в виде двух отдельных белков (как показано ниже) в одной и той же клетке.
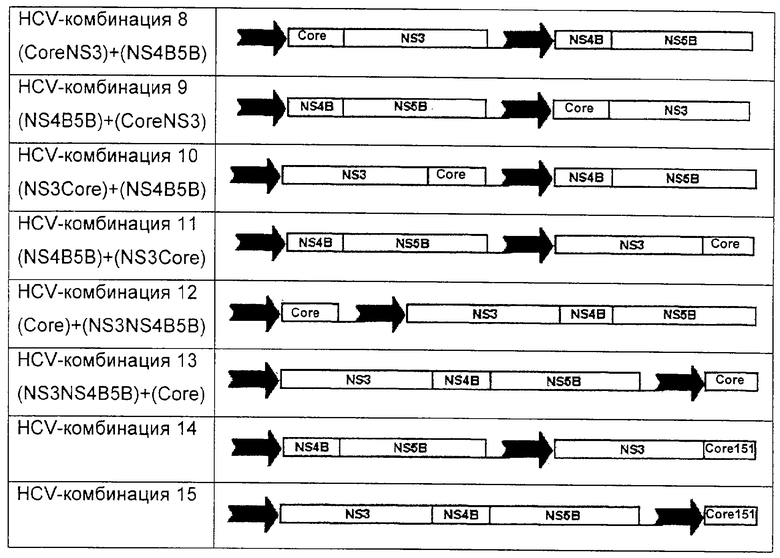
Предпочтительными конструкциями являются HCV-комбинации 7, 9, 11 или 12. Особенно предпочтительны 7 и 11.
В альтернативном аспекте настоящего изобретения полинуклеотидные вакцины возможно не содержат полинуклеотид, кодирующий core-белок. Например, предпочтительные полинуклеотиды по этому аспекту настоящего изобретения включают:
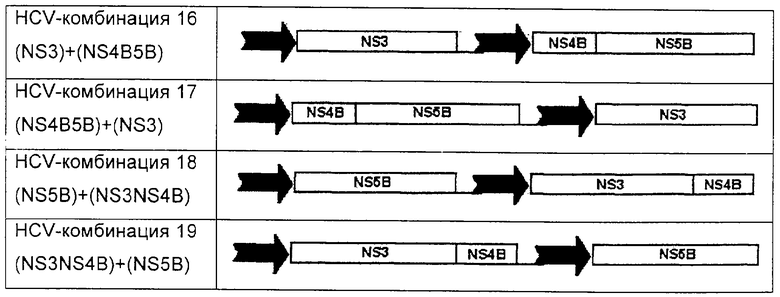
Для представленных выше HCV-комбинаций 8-19 подразумевается, что используемая терминология, например (CoreNS3)+(NS4B5B), раскрывает полинуклеотидный вектор, содержащий две экспрессионные кассеты, каждая из которых управляется независимо индивидуальным промотором, и в случае этого примера содержащий одну экспрессионную кассету, кодирующую двойной слитый белок CoreNS3, и другую, кодирующую двойной слитый белок NS4B-NS5B. Каждую HCV-комбинацию 8-19 следует интерпретировать соответственно.
В представленных выше HCV-комбинациях 1-19 раскрыты относительные ориентации белков, полипротеиновых слияний или полинуклеотидов HCV. В частности, в данном описании также раскрыто, что все представленные выше HCV-комбинации 1-19 также раскрыты с каждой из предпочтительных мутаций или укорочений для удаления активности входящих в состав белков. Например, предпочтительные варианты комбинаций 1-19 (если не указано иначе в обратном смысле) включают нуклеотидные последовательности для core (1-191 (целую последовательность, но разделенную при этом на два или более фрагментов с ограниченной биологической активностью), или предпочтительно core, представленный в своих укороченных формах 1-151, или 1-165, или 1-171); NS3 1027-1657 (мутации с целью инактивации геликазной (аспарагиновая кислота 1316 на глутамин) и протеазной (серин 1165 на валин) активности; NS5B 2420-3010 (мутация с заменой аспарагиновой кислоты 2639 на глицин и аспарагиновой кислоты 2644 на глицин, мотив А, для инактивации полимеразной активности); и NS4B 1712-1972 (возможно укороченный до 1760-1972 с удалением N-концевого высоковариабельного фрагмента).
Согласно настоящему изобретению предложены новые ДНК-вакцины и полипептиды, которые описаны выше. Кроме того, в настоящем изобретении предложены аналоги описанных полипептидов и содержащие их ДНК-вакцины.
Термин "аналог" относится к полинуклеотиду, который кодирует ту же аминокислотную последовательность, что и другой полинуклеотид по настоящему изобретению, но который, вследствие избыточности генетического кода, имеет отличающуюся нуклеотидную последовательность, сохраняющую при этом ту же самую картину использования кодонов, например, имеющую тот же самый коэффициент использования кодонов или коэффициент использования кодонов в пределах 0,1, предпочтительно в пределах 0,05 от такового для другого полинуклеотида.
Полинуклеотидные последовательности HCV могут быть получены из любых различных генотипов, штаммов или изолятов HCV. Изоляты HCV можно классифицировать на следующие шесть основных генотипов, содержащих один или более подтипов: HCV 1 (1а, 1b или 1с), HCV 2 (2а, 2b или 2с), HCV 3 (3а, 3b, 10а), HCV 4 (4а), HCV 5 (5а) и HCV 6 (6а, 6b, 7b, 8b, 9а и 11а); Simmonds, J. Gen. Virol., 2001, 693-712. В контексте настоящего изобретения каждый белок HCV может быть получен из полинуклеотидной последовательности одного и того же генотипа или подтипа HCV либо, альтернативно, из любой комбинации генотипа или подтипа HCV, и белок HCV может быть использован. Предпочтительно данные гены происходят из генотипа типа 1b, например, инфекционного клона J4L6 (регистрационный № AF 0542478 - см. Фиг.1).
Конкретные штаммы, которые были секвенированы, включают HCV-J (Kato et al., 1990, PNAS, USA, 87: 9724-9528) и ВК (Takamizawa et al., 1991, J. Virol. 65: 1105-1113).
Полинуклеотиды согласно изобретению находят применение в получении посредством экспрессии кодируемых белков, при этом экспрессия может иметь место in vitro, in vivo или ex vivo. Таким образом, эти нуклеотиды могут быть вовлечены в рекомбинантный белковый синтез, например с целью увеличения выходов, или реально находить применение в качестве подходящих терапевтических агентов, используемых в способах ДНК-вакцинации. Когда полинуклеотиды по настоящему изобретению используют в получении кодируемых белков in vitro или ex vivo, клетки, например в клеточной культуре, будут модифицированы с включением полинуклеотида, который должен быть экспрессирован. Такие клетки включают временные или предпочтительно стабильные клеточные линии млекопитающих. Конкретные примеры клеток, которые могут быть модифицированы посредством вставки векторов, кодирующих полипротеины по изобретению, включают клетки млекопитающих НЕК293Т, СНО, HeLa, 293 и COS. Предпочтительно, когда выбранная клеточная линия будет представлять собой линию, являющуюся не только стабильной, но также разрешающей "зрелое" гликозилирование и экспрессию полипротеина на клеточной поверхности. Экспрессия может осуществляться в трансформированных ооцитах. Полипептид может быть экспрессирован из полинуклеотида по настоящему изобретению в клетках трансгенного, отличного от человека животного, предпочтительно мыши. Трансгенное, отличное от человека животное, экспрессирующее полипептид из полинуклеотида по изобретению, включено в объем данного изобретения.
Настоящее изобретение включает экспрессирующие векторы, которые содержат нуклеотидные последовательности по изобретению. Такие экспрессирующие векторы конструируют в области молекулярной биологии стандартными способами, и они могут, например вовлекать использование плазмидной ДНК и соответствующих инициаторов, промоторов, энхансеров и других элементов, таких как, например, сигналы полиаденилирования, которые могут быть необходимы, и которые располагаются в правильной ориентации для того, чтобы сделать возможной экспрессию белка. Другие подходящие векторы очевидны специалистам в данной области техники. В качестве дополнительного примера в этом отношении авторы ссылаются на Sambrook et al. Molecular Cloning: a Laboratory Manual. 2nd Edition. CSH Laboratory Press (1989).
Предпочтительно полинуклеотид по изобретению или для использования в данном изобретении в векторе функциональным образом связан с контролирующей последовательностью, способной обеспечивать экспрессию кодирующей последовательности клеткой-хозяином, то есть данный вектор является экспрессирующим вектором. Термин "функциональным образом связанный" относится к смежному положению, при котором описанные элементы взаимосвязаны, что позволяет им функционировать надлежащим образом. Регуляторная последовательность, например промотор, "функциональным образом связанный" с кодирующей последовательностью, располагается таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с регуляторной последовательностью.
Экспрессионная кассета представляет собой конструкцию, которая способна направлять экспрессию интересующей последовательности или гена. Экспрессионная кассета включает регуляторные элементы, такие как промотор, который функциональным образом связан с интересующим геном.
Векторы могут представлять собой, например плазмиды, искусственные хромосомы (например ВАС, РАС, YAC), векторы на основе вирусов или фагов, снабженные ориджином репликации, возможно промотором для экспрессии полинуклеотида и возможно регулятором промотора. Векторы могут содержать один или более селективных маркерных генов, например ген устойчивости к ампициллину или канамицину в случае бактериальной плазмиды или ген устойчивости для вектора на основе грибов. Векторы могут быть использованы in vitro, например для продукции ДНК или РНК либо для трансфекции или трансформации клетки-хозяина, например клетки-хозяина млекопитающих, в частности для продукции белка, кодируемого этим вектором. Кроме того, векторы могут быть адаптированы для использования in vivo, например в способе ДНК-вакцинации или генной терапии.
Промоторы и другие сигналы, регулирующие экспрессию, могут быть выбраны по их совместимости с клеткой-хозяином, для которой предназначена экспрессия. Например, промоторы млекопитающих включают металлотионеиновый промотор, который может индуцироваться в ответ на тяжелые металлы, такие как кадмий, и β-актиновый промотор. Также могут быть использованы вирусные промоторы, такие как промотор большого Т-антигена SV40, немедленно ранний (immediate early, IE) промотор цитомегаловируса человека, промотор LTR вируса саркомы Рауса, аденовирусный промотор или промотор HPV, в частности регуляторная область (upstream regulatory region, URR) HPV. Все эти промоторы хорошо описаны и легко доступны в данной области техники.
Примеры подходящих вирусных векторов включают векторы на основе вируса простого герпеса, векторы на основе вируса коровьей оспы (vaccinia virus) или альфа-вируса и ретровирусы, включая лентивирусы, аденовирусы и адено-ассоциированные вирусы. Методы генного переноса с использованием этих вирусов известны специалистам в данной области техники. Ретровирусные векторы, например, могут быть использованы для стабильной интеграции полинуклеотида по изобретению в геном хозяина, хотя такая рекомбинация не является предпочтительной. В противоположность этому, дефектные по репликации аденовирусные векторы остаются эписомными и, следовательно, разрешают временную экспрессию. Векторы, способные осуществлять экспрессию в клетках насекомых (например, бакуловирусные векторы), в клетках человека или в бактериях, могут быть использованы для продукции количеств белка HCV, кодируемого полинуклеотидами по настоящему изобретению, например для использования в качестве субъединичных вакцин или в иммуноанализах.
В следующем аспекте настоящего изобретения предложена фармацевтическая композиция, содержащая полинуклеотидную последовательность, как описано здесь. Предпочтительно данная композиция содержит ДНК-вектор согласно второму аспекту настоящего изобретения. В предпочтительных воплощениях композиция содержит большое количество частиц, предпочтительно частиц золота, покрытых ДНК, содержащей вектор, включающий полинуклеотидную последовательность, которая кодирует аминокислотную последовательность HCV, причем картина использования кодонов данной полинуклеотидной последовательности имеет сходство с таковой для высокоэкспрессируемых генов млекопитающих, в частности генов человека. В альтернативных воплощениях композиция содержит фармацевтически приемлемый эксципиент и ДНК-вектор согласно второму аспекту настоящего изобретения. Композиция также может включать адъювант.
ДНК-вакцины могут быть доставлены посредством интерстициального введения жидких вакцин в мышцу (WO 90/11092) либо посредством механизмов, отличных от внутримышечного введения. Например, преимущество доставки в кожу заключается в том, что иммунные механизмы очень активны в тканях, являющихся барьерами для инфекции, таких как кожа и слизистые оболочки. Доставка в кожу может быть осуществлена посредством инъекции, посредством безыгольного шприца (при этом жидкость нагнетается в кожу или расположенные под ней ткани, включая мышцы, под давлением) либо посредством бомбардировки частицами, причем ДНК может быть нанесена в виде покрытия на частицы достаточной плотности для того, чтобы проходить сквозь эпителий (патент US 5371015). Например, нуклеотидные последовательности могут быть включены в плазмиду, которую наносят в виде покрытия на частицы золота, которые затем вводят под большим давлением в эпидермис, как описано, например, в Haynes et al. J. Biotechnology 44: 37-42 (1996). Бомбардировка кожи этими частицами приводит к прямой трансфекции как эпидермальных клеток, так и клеток Лангерганса. Клетки Лангерганса являются антиген-представляющими клетками (АРС), которые поглощают ДНК, экспрессируют кодируемые пептиды и процессируют их для презентации белков МНС на клеточной поверхности. Трансфицированные клетки Лангерганса мигрируют в лимфатические узлы, где они представляют презентированные антигенные фрагменты лимфоцитам, вызывая иммунный ответ. Очень небольшие количества ДНК (менее 1 мкг, часто менее 0,5 мкг) требуются для индукции иммунного ответа путем опосредованной частицами доставки в кожу, и это контрастирует с миллиграммовыми количествами ДНК, которые, как известно, требуются для получения иммунных ответов после непосредственной внутримышечной инъекции.
В тех случаях, когда полинуклеотиды по настоящему изобретению находят применение в качестве терапевтических агентов, например в ДНК-вакцинации, нуклеиновую кислоту будут вводить млекопитающему, например человеку, которого необходимо подвергнуть вакцинации. Нуклеиновая кислота, такая как РНК или ДНК, предпочтительно ДНК, предложена в форме вектора, такого, как описано выше, который может экспрессироваться в клетках млекопитающего. Полинуклеотиды могут быть введены любым доступным способом. Например, нуклеиновая кислота может быть введена посредством игольной инъекции, предпочтительно внутрикожно, подкожно или внутримышечно. Альтернативно, нуклеиновая кислота может быть доставлена непосредственно в кожу с использованием способа доставки нуклеиновой кислоты, такого как опосредованная частицами доставка ДНК (particle-mediated DNA delivery, PMDD). Согласно этому способу инертные частицы (например, золотые гранулы) покрывают нуклеиновой кислотой и ускоряют их при скоростях, достаточных для того, чтобы способствовать их прохождению через поверхность реципиента (например, кожу), например, посредством доставки в условиях повышенного давления из проецируемого устройства. (Частицы, покрытые молекулой нуклеиновой кислоты по настоящему изобретению, находятся в пределах объема настоящего изобретения, как представляющие собой устройства для доставки, нагруженные такими частицами). Желательно, чтобы композиция содержала частицы золота, имеющие средний диаметр 0,5-5 мкм, предпочтительно около 2 мкм. В предпочтительных воплощениях золотые гранулы, имеющие покрытие, загружают в пробирки, которые служат в качестве картриджей, так, чтобы каждый картридж содержал 0,1-1 мг, предпочтительно 0,5 мг золота, покрытого 0,1-5 мкг, предпочтительно около 0,5 мкг ДНК/картридж.
Согласно другому аспекту изобретения предложена клетка-хозяин, содержащая полинуклеотидную последовательность, как описано здесь. Клетка-хозяин может быть бактериальной, например Е. coli, клеткой млекопитающего, например человека, или может быть клеткой насекомого. Клетки млекопитающих, содержащие вектор по настоящему изобретению, могут быть культивируемыми клетками, трансфицированными in vitro, или могут быть трансфицированы in vivo посредством введения вектора млекопитающему.
В другом аспекте настоящего изобретения предложен способ приготовления фармацевтической композиции, как описано выше, включающий стадию изменения картины использования кодонов нуклеотидной последовательности HCV дикого типа или создания синтетической полинуклеотидной последовательности с целью получения последовательности, имеющей картину использования кодонов, сходную с таковой для высокоэкспрессируемых генов млекопитающих, и кодирующей аминокислотную последовательность HCV дикого типа, или мутантной аминокислотной последовательности HCV, содержащей последовательность дикого типа с аминокислотными заменами, достаточными для инактивации одной или более природных функций полипептида.
Также предложено применение полинуклеотида или вакцины, как описано здесь, в лечении или профилактике HCV-инфекции.
Подходящие методы введения "голого" полинуклеотида или вектора пациенту включают местное применение с использованием соответствующего носителя. Нуклеиновая кислота может быть нанесена местно на кожу или слизистые оболочки, например путем интраназального, перорального, интравагинального или интраректального введения. "Голый" полинуклеотид или вектор могут быть представлены вместе с фармацевтически приемлемым эксципиентом, таким как фосфатно-солевой буфер (PBS). "Захват" ДНК может быть дополнительно облегчен путем использования вспомогательных агентов, таких как бупивакаин, либо по отдельности, либо включенными в препарат ДНК. Другие способы введения нуклеиновой кислоты непосредственно реципиенту включают ультразвуковую, электрическую стимуляцию, электропорацию и microseeding (прямой перенос генетического материала в клетку-мишень с помощью микроигл), который описан в патенте US 5697901.
Захват конструкций нуклеиновой кислоты может быть усилен с помощью некоторых известных методов трансфекции, например таких, которые включают применение агентов трансфекции. Примеры этих агентов включают катионные агенты, например фосфат кальция и ДЭАЭ-декстран, и липофектанты, например липофектам и трансфертам. Дозировка вводимой нуклеиновой кислоты может изменяться. Обычно нуклеиновую кислоту вводят в количестве в интервале от 1 пг до 1 мг, предпочтительно от 1 пг до 10 мкг нуклеиновой кислоты на частицу, опосредующую доставку гена, и от 10 мкг до 1 мг для других путей.
Последовательность нуклеиновой кислоты по настоящему изобретению также может быть введена посредством специализированных векторов доставки, полезных в генной терапии. Генотерапевтические подходы обсуждаются, например, в Verme et al., Nature 1997, 389: 239-242. Могут быть использованы как вирусные, так и невирусные векторные системы. Системы на основе вирусов включают системы на основе ретровирусов, лентивирусов, аденовирусов, адено-ассоциированных вирусов, вирусов герпеса, вируса оспы канареек и вируса коровьей оспы. Предпочтительными являются аденовирусные векторы, происходящие из приматов, не являющихся людьми. В частности, Pan 9 (С68), как описано в патенте US 6083716, Pan 5, 6 или 7, как описано в WO 03/046124.
Невирусные системы включают непосредственное введение нуклеиновых кислот, технологию инкапсулирования микросфер из (поли(лактидко-гликолида)) и системы на основе липосом. Вирусные и невирусные системы доставки могут быть объединены, если это желательно, с целью обеспечения бустерных инъекций после первичной вакцинации, например "первичной" ДНК-вакцинации с использованием невирусного вектора, например плазмиды, с последующей одной или более "бустерными" вакцинациями с использованием вирусных векторов или невирусных систем. Кроме того, протоколы первичных и бустерных вакцинаций могут иметь преимущество, состоящее в сенсибилизации (priming) белком в адъюванте и ревакцинации ДНК или вирусным вектором, кодирующим полинуклеотид по изобретению. Альтернативно, в качестве бустера может быть использована вакцина на основе белка. Предпочтительно, чтобы такая белковая вакцина содержала все антигены, которые будет содержать ДНК-вакцина на основе вирусного вектора. Однако эти белки могут быть представлены индивидуально или в виде полипротеина.
Последовательность нуклеиновой кислоты по настоящему изобретению также может быть введена посредством трансформированных клеток. Такие клетки включают клетки, полученные от субъекта. "Голый" полинуклеотид или вектор по настоящему изобретению может быть введен в такие клетки in vitro, и трансформированные клетки затем могут быть возвращены данному субъекту. Полинуклеотид по изобретению может встраиваться в нуклеиновую кислоту, уже присутствующую в клетке, посредством гомологичной рекомбинации. При желании трансформированная клетка может выращиваться in vitro, и одна или более полученных клеток может быть использовано в настоящем изобретении. Клетки могут быть доставлены в соответствующий участок пациента с помощью известных хирургических или микрохирургических методов (например, трансплантации, микроинъекции и т.д.).
Подходящие клетки включают антигенпредставляющие клетки (АРС), такие как дендритные клетки, макрофаги, В-клетки, моноциты и другие клетки, которые могут быть сконструированы как эффективные АРС. Такие клетки могут быть, но не обязательно, генетически модифицированы с целью увеличения способности к представлению антигена для улучшения активации и/или поддержания Т-клеточного ответа, с целью приобретения противоопухолевых эффектов, например против цервикальной карциномы per se, и/или иммунологической совместимости с реципиентом (то есть подходящего HLA-гаплотипа). В общем случае АРС могут быть выделены из любой биологической жидкости или органа, включая опухолевые и околоопухолевые ткани, и могут представлять собой аутологичные, аллогенные, сингенные или ксеногенные клетки.
В некоторых предпочтительных воплощениях по настоящему изобретению используют дендритные клетки или их предшественники в качестве антигенпредставляющих клеток либо для трансформации in vitro и возвращения пациенту, либо в качестве in vivo мишени для нуклеотидов, доставляемых в вакцине, например, посредством частицы, опосредующей доставку ДНК. Дендритные клетки являются высокоэффективными АРС (Banchereau and Steinman, Nature 392: 245-251, 1998), и было показано, что они эффективны в качестве физиологического адъюванта для обеспечения профилактического или терапевтического противоопухолевого иммунитета (см. Timmerman and Levy, Ann. Rev. Med. 50: 507-529, 1999). В общем случае дендритные клетки могут быть идентифицированы на основе их типичной формы (звездообразная in situ, с заметными цитоплазматическими отростками (дендритами), видимыми in vitro), их способности поглощать, процессировать и представлять антигены с высокой эффективностью и их способности активировать первичные Т-клеточные ответы. Дендритные клетки могут, несомненно, быть сконструированы для экспрессии специфических рецепторов клеточной поверхности или лигандов, которые обычно не обнаруживаются на дендритных клетках in vivo или ех vivo, например антигена(ов), кодируемого в конструкциях по изобретению, и такие модифицированные дендритные клетки рассматриваются в настоящем изобретении. В качестве альтернативы дендритным клеткам в вакцине могут быть использованы антигеннагруженные дендритные клетки, секретирующие везикулы (так называемые экзосомы) (см. Zitvogel et al., Nature Med. 4: 594-600, 1998).
Дендритные клетки и предшественники могут быть получены из периферической крови, костного мозга, опухольинфильтрирующих клеток, клеток, инфильтрирующих околоопухолевые ткани, лимфатических узлов, селезенки, кожи, пуповинной крови или любой другой подходящей ткани или жидкости. Например, дендритные клетки могут быть дифференцированы ех vivo путем добавления комбинации цитокинов, таких как GM-CSF, IL-4, IL-13 и/или TNF, к культурам моноцитов, полученных из периферической крови. Альтернативно, СD34-положительные клетки, полученные из периферической крови, пуповинной крови или костного мозга, могут быть дифференцированы в дендритные клетки путем добавления в культуральную среду комбинаций GM-CSF, IL-3, TNF, СD40-лиганда, липополисахарида LPS, fltS-лиганда (цитокин, важный в генерации профессиональных антигенпредставляющих клеток, в частности дендритных клеток) и/или другого соединения(й), которые индуцируют дифференцировку, созревание и пролиферацию дендритных клеток.
В общем случае АРС могут быть трансфицированы полинуклеотидом, кодирующим антигенную аминокислотную последовательность HCV, таким как кодон-оптимизированный полинуклеотид, как предусмотрено в настоящем изобретении. Такая трансфекция может иметь место ex vivo, и композиция или вакцина, содержащая такие трансфицированные клетки, может быть затем использована для терапевтических целей, как описано здесь. Альтернативно, пациенту может быть введен носитель для доставки гена, который нацелен на дендритную или другую антиген-представляющую клетку, что приводит к трансфекции, происходящей in vivo. Например, in vivo и ех vivo трансфекция дендритных клеток в общем случае может быть проведена с использованием любых способов, известных в данной области техники, например способов, описанных в WO 97/24447, или подхода, опосредованного частицами, описанного Mahvi et al., Immunology and Cell Biology 75: 456-460, 1997.
Вакцины и фармацевтические композиции по изобретению могут быть использованы в сочетании с противовирусными агентами, такими как α-интерферон, предпочтительно пегилированный α-интерферон, и рибоварин. Вакцины и фармацевтические композиции могут быть представлены в однодозовых или многодозовых контейнерах, таких как герметично закрытые ампулы или флаконы. Такие контейнеры предпочтительно герметично запаяны для сохранения стерильности препарата до использования. Обычно препараты могут храниться в виде суспензий, растворов или эмульсий в масляных или водных растворителях. Альтернативно, вакцина или фармацевтическая композиция может храниться в лиофилизированном состоянии, что требует лишь добавления стерильного жидкого носителя непосредственно перед использованием. Вакцины, содержащие нуклеотидные последовательности, предназначенные для введения через опосредованную частицами доставку, могут быть представлены в виде картриджей, подходящих для использования с устройством для доставки посредством сжатого газа, в этом случае картриджи могут состоять из полых трубок, внутренняя поверхность которых покрыта частицами, несущими вакцинную нуклеотидную последовательность, возможно в присутствии других фармацевтически приемлемых ингредиентов.
Фармацевтические композиции по настоящему изобретению могут включать адъювантные соединения или другие вещества, которые могут быть полезными для модуляции или усиления иммунного ответа, индуцированного белком, кодируемым данной ДНК. Они могут кодироваться ДНК, либо отдельно от антигена, либо в виде слияния с антигеном, или могут быть включены в виде не-ДНК элементов препарата. Примеры адъювантных веществ, которые могут быть включены в препараты по настоящему изобретению, включают убиквитин, лизосомально-ассоциированный мембранный белок (LAMP), core-антиген вируса гепатита В, flt3-лиганд и другие цитокины, такие как IFN-γ и GMCSF.
Другие подходящие адъюванты имеются в продаже, такие как, например, неполный адъювант и полный адъювант Фрейнда (Difco Laboratories, Detroit, MI); имиквимод (Imiquimod) (3M, St. Paul, MN); резимиквимод (Resimiquimod) (3M, St. Paul, MN); адъювант 65 фирмы Merck (Merck and Company, Inc., Rahway, NJ); соли алюминия, например, гель гидроксида алюминия (квасцы) или фосфат алюминия; соли кальция, железа или цинка; нерастворимая суспензия ацилированного тирозина; ацилированные сахара; катионные или анионные производные полисахаридов; полифосфазены; биодеградируемые микросферы; монофосфориллипид А и квил А. Цитокины, такие как GM-CSF или интерлейкин-2, -7 или -12, также могут быть использованы в качестве адъювантов.
В препаратах по изобретению предпочтительно, чтобы адъювантная композиция индуцировала иммунный ответ преимущественно Тh1-типа. Таким образом, адъювант может быть полезен для модуляции иммунного ответа, генерируемого в ответ на кодируемые ДНК антигены, от ответа преимущественно Th2- до преимущественно Тh1-типа. Высокие уровни цитокинов Тh1-типа (например IFN-, TNF, IL-2 и IL-12) имеют тенденцию способствовать индукции клеточно-опосредованных иммунных ответов на вводимый антиген. Согласно предпочтительному воплощению, в котором ответ является преимущественно ответом Тh1-типа, уровень цитокинов Тh1-типа будет увеличиваться в большей степени, чем уровень цитокинов Тh2-типа. Уровни этих цитокинов могут быть легко оценены с использованием стандартных анализов. Для обзора семейств цитокинов см. Mosmann and Coffman, Ann. Rev. Immunol. 7: 145-173, 1989.
Так, подходящие адъюванты, использующиеся для того, чтобы вызывать преимущественно ответ Тh1-типа, включают, например комбинацию монофосфориллипида А, предпочтительно 3-де-O-ацилированного монофосфориллипида A (3D-MPL), вместе с солью алюминия. Другие известные адъюванты, которые предпочтительно индуцируют иммунный ответ TH1-типа, включают CpG-содержащие олигонуклеотиды. Эти олигонуклеотиды характеризуются тем, что динуклеотид CpG не метилирован. Такие олигонуклеотиды хорошо известны и описаны, например в WO 96/02555. Также описаны иммуностимуляторные последовательности ДНК, например Sato et al., Science 273: 352, 1996. CpG-содержащие олигонуклеотиды могут кодироваться отдельно от антигена(ов) папилломавируса в той же или другой полинуклеотидной конструкции либо могут находиться в непосредственной близости к ним, например, в виде слияния с ними. Альтернативно, CpG-содержащие олигонуклеотиды могут быть введены отдельно, то есть не как часть композиции, которая включает кодируемый антиген. CpG-олигонуклеотиды могут быть использованы как таковые или в комбинации с другими адъювантами. Например, улучшенная система включает комбинацию CpG-содержащего олигонуклеотида и производного сапонина, в частности комбинацию CpG и QS21, как описано в WO 00/09159 и WO 00/62800. Предпочтительно, когда препарат дополнительно содержит эмульсию масло-в-воде и/или токоферол.
Другим предпочтительным адъювантом является сапонин, предпочтительно QS21 (Aquila Biopharmaceuticals Inc., Framingham, MA), который может быть использован как таковой или в комбинации с другими адъювантами. Например, улучшенная система включает комбинацию монофосфориллипида А и производного сапонина, такую как комбинация QS21 и 3D-MPL, как описано в WO 94/00153, или менее реактогенную композицию, где QS21 гасят холестерином, как описано в WO 96/33739. Другие предпочтительные препараты содержат эмульсию масло-в-воде и токоферол. Особенно эффективный адъювантный препарат, включающий QS21, 3D-MPL и токоферол в эмульсии масло-в-воде, описан в WO 95/17210.
Другие предпочтительные адъюванты включают Монтанид ISA 720 (Seppic, France), SAF (Chiron, California, США), ISCOMS (CSL), MF-59 (Chiron), Детокс (Ribi, Hamilton, MT), RC-529 (Corixa, Hamilton, MT) и другие аминоалкил-глюкозаминид-4-фосфаты (AGP).
Если вакцина включает адъювант, то вакцинный препарат может быть введен в два приема. Например, часть препарата, содержащая нуклеотидную конструкцию, кодирующую антиген, может быть введена первой, например, посредством подкожной или внутримышечной инъекции либо посредством интрадермальной опосредованной частицами доставки, а затем может быть введена часть препарата, содержащая адъювант, либо сразу, либо через подходящий период времени, который будет очевиден врачу-специалисту в области вакцин. В этих случаях адъювант может быть введен тем же способом, что и антигенный препарат, или альтернативным способом. В других воплощениях адъювантная часть препарата будет вводиться до введения антигенной части. В одном воплощении адъювант вводят в виде местного препарата, наносимого на кожу в месте опосредованной частицами доставки нуклеотидных последовательностей, кодирующих антиген(ы), либо до, либо после его опосредованной частицами доставки.
Предпочтительно ДНК-вакцины по настоящему изобретению стимулируют эффективный иммунный ответ, обычно CD4+ и CD8+ иммунитет против антигенов HCV. Предпочтительно против широкого диапазона эпитопов. С терапевтической точки зрения предпочтительно, чтобы фиброз печени и/или воспаление были уменьшены после вакцинации.
Используемый здесь термин "содержащий" предназначен для использования в его не ограничивающем смысле, то есть присутствие других элементов не исключается. Однако предполагается также, что слово "содержащий" может быть также понято в своем исключающем смысле, соизмеримым с "состоящим" или "состоящим из". Настоящее изобретение проиллюстрировано следующими не ограничивающими примерами.
Пример 1. Мутации, введенные в панель антигенов
1) Консенсусные мутации
Было проведено сравнение полных геномных последовательностей всех известных изолятов HCV. Некоторые положения в полипротеине J4L6 были идентифицированы как необычные/отклоняющиеся от большинства других изолятов HCV. Особое значение представляли те положения, для которых было обнаружено отклонение от более консенсусных остатков среди родственных изолятов группы 1b, распространяющееся на изоляты групп 1а, 2, 3 и других групп, где в противном случае один или два альтернативных аминокислотных остатка доминировали в эквивалентном положении. Ни одна из выбранных консенсусных мутаций не вносила изменений в известный CD4- или CD8-эпитоп. Два изменения в NS3 фактически восстанавливали иммунодоминантный HLA-В35-ограниченный CD8-эпитоп [изолейцин (I) 1365 на валин (V) и глицин (G) 1366 на аланин (А)].
Первая 51 аминокислота NS4B была удалена ввиду бесполезной изменчивости.
Core
Аланин (А) 52 на треонин (Т)
NS3
Валин (V) 1040 на лейцин (L)
Лейцин (L) 1106 на глутамин (Q)
Серин (S) 1124 на треонин (Т)
Валин (V) 1179 на изолейцин (I)
Треонин (Т) 1215 на серин (S)
Глицин (G) 1289 на аланин (А)
Серин (S) 1290 на пролин (Р)
Изолейцин (I) 1365 на валин (V)
Глицин (G) 1366 на аланин (А)
Треонин (Т) 1408 на серин (S)
Пролин (Р) 1428 на треонин (Т)
Изолейцин (I) 1429 на серин (S)
Изолейцин (I) 1636 на треонин (Т)
NS4B
Стартовая ОРС на фенилаланине (F) 1760
NS5B
Изолейцин (I) 2824 на валин (V)
Треонин (Т) 2892 на серин (S)
Треонин (Т) 2918 на валин (V)
Примечание: Нумерация соответствует положению в полипротеине для изолята J4L6.
Пример 2. Конструирование плазмидных ДНК-вакцин
Полинуклеотидные последовательности, кодирующие core, NS3, укороченный NS4B и NS5B HCV, были кодон-оптимизированы для использования кодонов у млекопитающих с применением программного обеспечения SynGene 2e. Коэффициент использования кодонов был улучшен до значения, превышающего 0,7 для каждого полинуклеотида. Смысловые и антисмысловые нити каждой новой полинуклеотидной последовательности, включающие кодоновую оптимизацию, фермент-нокаутирующие мутации и консенсусные мутации, были разбиты на участки по 40-60 нуклеотидов с перекрыванием в 20 нуклеотидов. Эти участки были синтезированы на заказ, и полинуклеотид получали методом ПЦР с помощью набора олигонуклеотидов.
Ниже приведены внешние прямые и обратные ПЦР-праймеры для каждого полинуклеотида, иллюстрирующие уникальные рестрикционные эндонуклеазные сайты, используемые для клонирования:

Все полинуклеотиды, кодирующие единичные антигены, были клонированы в экспрессирующий вектор p7313ie для млекопитающих через уникальные сайты клонирования NotI и BamHI (см. Фиг.7).
Кодируемые полипротеины представляют собой следующие последовательности (включая мутации и кодоновую оптимизацию):

Пример 3. Анализы иммунного ответа
Мышей C57BL или BALB/c иммунизировали либо WT (дикий тип), либо кодон-оптимизированными + мутантными вариантами четырех антигенов HCV, экспрессированными индивидуально в векторе р7313. Мышей иммунизировали PMID (particle-mediated immunotherapeutic delivery) в стандартной дозе 1,0 мкг/картридж и повторно иммунизировали на 21-й день (бустер 1) и затем на 49-й день (бустер 2). Клетки селезенки собирали от отдельных мышей и повторно стимулировали в клеточном иммуноферментном анализе (ELISPOT) различными препаратами антигенов HCV. Измеряли оба ответа как на IL2, так и на IFNγ. Реагенты, используемые для измерения иммунных ответов, представляли собой очищенные белки core, NS3, NS4 и NS5B HCV (генотип 1b) от фирмы Mikrogen, core и NS3-5 вируса коровьей оспы (генотип 1b).
Core HCV
Мышей C57BL, иммунизированных полноразмерным (FL-1-191) WT или укороченным core (TR 1-115), повторно стимулировали core-белком HCV и наблюдали хорошие ответы с очищенным core-белком (Фиг.8).
NS3 HCV
Мышей иммунизировали р7313 WT и кодон-оптимизированным NS3, используя PMID. Хорошие ответы на NS3 после иммунизации и однократной повторной иммунизации были продемонстрированы на мышах C57BL с использованием как белка NS3, так и коровьей оспы 3-5 для прочтения ответа посредством ELISPOT. Были выявлены оба ответа и на IL2, и на IFNγ. Никаких существенных различий между вариантом конструкции дикого типа и кодон-оптимизированным (со+m) вариантом конструкции в этом эксперименте не обнаружено (Фиг.9). Однако при экспрессии in vitro после временной трансфекции наблюдали различия между конструкцией дикого типа и кодон-оптимизированной конструкцией. Эксперименты, где проводят сравнение конструкций при низкой дозе ДНК или при первичном ответе, могут выявлять различия в эффективности плазмид.
NS4B HCV
Ответы на полноразмерный NS4B р7313 WT наблюдали после PMID-иммунизации мышей BALB/c. Оба ELISPOT-ответа, как на IL2, так и на IFNγ, наблюдали после повторной стимуляции in vitro как белком NS4B, так и коровьей оспы 3-5 (Фиг.10).
Белок NS4B был укорочен по N-концу с целью удаления высоковариабельного участка, однако экспрессию этого белка невозможно было выявить после исследований трансфекции in vitro, поскольку доступная антисыворотка была получена против N-терминального участка. Для подтверждения экспрессии этого участка его сливали с белком NS5B. Недавние эксперименты подтвердили, что иммунные ответы могут быть выявлены против укороченного белка NS4B как такового, так и в виде слияния с белком NS5B, с использованием белка NS4B и NS3-5 коровьей оспы. Хорошие ответы наблюдали на NS4B WT и кодон-оптимизированный NS4B.
NS5B HCV
Иммунный ответ на NS5B после PMID исследовали после иммунизации WT и кодон-оптимизированной (со+m) последовательностями. Хорошие ответы на NS5B после иммунизации и однократной повторной иммунизации были продемонстрированы на мышах C57BL с использованием как белка NS3, так и 3-5 коровьей оспы для прочтения ответа посредством ELISPOT. Как и с NS3, никаких различий в иммунном ответе между вариантами конструкций WT и со+m в этом эксперименте не наблюдали (Фиг.11).
Пример 4. Экспрессия полипротеинов HCV
Четыре выбранных антигена HCV - core, NS3, NS4B и NS5B - были сконструированы в p7313ie для экспрессии в виде единого слитого полипротеина. Антигены были экспрессированы в различном порядке в различных конструкциях, как показано ниже (панель конструкций, кодирующих экспрессию единых полипротеинов, была разработана так, чтобы аминоконцевое положение было занято каждым из четырех антигенов по очереди для того, чтобы контролировать, в каком случае уровень экспрессии был значительно выше или ниже в зависимости от присутствия того или другого антигена в этом важном положении). В дополнение к этому были созданы две конструкции, в которых core-белок был преобразован в 2 фрагмента, то есть Core 66-191>1-65 и 105-191>1-104.

Стандартизированное количество ДНК трасфицировали в клетки НЕК 293Т, используя реагент для трансфекции липофектамин 2000 (Invitrogen/Life Technologies), следуя стандартному протоколу производителя. Клетки собирали через 24 часа после трансфекции и проводили электрофорез в полиакриламидном геле, используя предварительно приготовленные 4-12%-ные NuPAGE бис-трис-гели с готовыми буферами MOPS или MES (Invitrogen/Life Technologies). Разделенные белки переносили на мембрану PVDF и экспрессию белков контролировали, используя кроличью антисыворотку, полученную против целого белка NS5B. Вторичным зондом являлась антисыворотка против иммуноглобулинов кролика, конъюгированная с пероксидазой хрена (hrp), с последующей хемилюминесцентной детекцией с использованием реагентов ECL (Amersham Biosciences).
Результаты исследования этой экспрессии показаны на Фиг.12. Эти результаты показывают, что все полипротеины экспрессируются в одинаковой степени, хотя и с более низкими уровнями, чем уровень, наблюдаемый в случае единичного антигена, экспрессирующего NS5B. Несколько более низкая молекулярная масса HCV500 обусловлена отщеплением core HCV от N-терминального положения. HCV502 не определялся в этом эксперименте вследствие ошибки клонирования. В повторном эксперименте с другим клоном уровень экспрессии HCV502 был аналогичен уровню экспрессии других полипротеинов.
Пример 5. Детекция иммунного ответа на полипротеины HCV
Мышей C57BL иммунизировали PMID с ДНК (1 мкг), кодирующей каждый из полипротеинов, с последующей повторной иммунизацией через 3 недели, как описано в Примере 4. Иммунные ответы контролировали через 7 дней после повторной иммунизации, используя ELISPOT или продукцию внутриклеточных цитокинов в ответ на антигены HCV.
ELISPOT-анализы Т-клеточных ответов на продукты генов HCV
Получение спленоцитов
На 7 сутки после повторной иммунизации из иммунизированных животных выделяли селезенку. Для получения клеточной суспензии селезенку обрабатывали путем измельчения между предметными стеклами. Эритроциты лизировали обработкой хлоридом аммония и удаляли дебрис, оставляя очищенную суспензию спленоцитов. Клетки ресуспендировали в концентрации 4×106/мл в полной среде RPMI для использования в ELISPOT-анализах, когда мыши получали только первичную иммунизацию, и в концентрации 2×106/мл, когда мышей подвергали повторной иммунизации.
ELISPOT-анализ
Планшеты покрывали 15 мкг/мл (в PBS) крысиного антимышиного IFNγ или крысиного антимышиного IL-2 (Pharmingen). Планшеты покрывали в течение ночи при +4°С. Перед использованием планшеты три раза промывали PBS. В планшеты добавляли спленоциты 4×105 клеток/лунку. Рекомбинантные антигены HCV получали от Mikrogen и использовали в концентрации 1 мкг/мл. Для измерения CD4- или СD8-ответов в анализах использовали пептиды в конечной концентрации 1-10 мкМ. Эти пептиды получали от Genemed Synthesis. Общий объем в каждой лунке составлял 200 мкл. Планшеты, содержащие антигенстимулированные клетки, инкубировали в течение 16 часов во влажном инкубаторе при 37°С. В некоторых экспериментах в качестве антигенов в ELISPOT-анализе использовали клетки, инфицированные рекомбинантной коровьей оспой, экспрессирующей NS3-5, или коровьей оспой дикого типа.
Обработка планшетов в ELISPOT-анализе
Клетки удаляли из планшетов однократной промывкой водой (с выдерживанием в течение 1 минуты для обеспечения лизиса клеток) и три раза PBS. Добавляли конъюгированный с биотином крысиный антимышиный IFN-γ или IL-2 (Pharmingen) в концентрации 1 мкг/мл в PBS. Планшеты инкубировали со встряхиванием в течение 2 часов при комнатной температуре. Затем планшеты три раза промывали PBS перед добавлением стрептавидин-щелочной фосфатазы (Caltag) в разведении 1/1000. После трех промывок в PBS пятна выявляли путем инкубации с субстратом BCICP (Biorad) в течение 15-45 мин. Субстрат удаляли, промывая водой, и планшеты оставляли сушиться. Пятна подсчитывали, используя систему анализа изображений.
Проточная цитометрия для определения продукции IFNγ и IL2 Т-клетками в ответ на стимуляцию пептидами
Приблизительно 3×106 спленоцитов отбирали в тест-пробирку, центрифугировали с образованием осадка. Супернатант удаляли и образцы интенсивно перемешивали для разрушения осадка. В каждую пробирку добавляли 0,5 мкг анти-СD28 + 0,5 мкг анти-CD49d (Pharmingen) и оставляли инкубироваться при комнатной температуре в течение 10 минут. В соответствующие пробирки добавляли 1 мл среды, которая содержала либо одну среду, либо среду с антигенами HCV. Затем образцы инкубировали в течение часа при 37°С в нагретой водяной бане. В каждую пробирку добавляли брефелдин А (10 мкг/мл), и инкубацию при 37°С продолжали в течение еще 5 часов. Затем температуру в программируемой водяной бане возвращали к 6°С и поддерживали при этой температуре в течение ночи.
Образцы затем окрашивали с помощью анти-мышиного CD-4-CyChrome (Pharmingen) и антимышиного CD8-биотин (Immunotech). Образцы промывали и окрашивали с использованием стрептавидин-ECD. Образцы промывали и добавляли 100 мкл Fixative из набора "Intraprep Permeabilization Reagent" (Immunotech) на 15 минут при комнатной температуре. После промывки к каждому образцу добавляли 100 мкл реагента для пермеабилизации из набора Intraprep с анти-IFN-γ-PE + анти-IL-2-FITC. Образцы инкубировали при комнатной температуре в течение 15 минут и промывали. Образцы ресуспендировали в 0,5 мл буфера и анализировали на проточном цитометре.
В целом образец включал 500000 клеток, и затем CD4- и CD8-клетки пропускали для определения популяций клеток, секретирующих IFNγ и/или IL-2 в ответ на стимул.
Результаты показывают, что все полипротеины, кодирующие core, NS3, NS4B и NS5B в различном порядке, способны стимулировать иммунные ответы на NS3 (то есть HCV 500, 510, 520, 530). Результаты представлены на Фиг.13. Ответы на белок NS3 похожи между каждым из полипротеинов HCV (HCV 500, 510, 520 и 530), когда контролируются с помощью IL2 (Фиг.13А) и IFNγ (Фиг.13B) ELISPOT.
Фенотип иммунореактивных клеток анализировали более детально с использованием ICS. Был выявлен хороший CD4+ Т-клеточный ответ на иммунодоминантный NS3 СD4-специфический пептид, который был похожим между HCV 500, 510, 520, 530.
Частота NS3 CD4- и СD8-специфичных Т-клеток, продуцирующих IFNγ после иммунизации полипротеинами HCV
IFNγ-специфические Т-клеточные ответы определяли после стимуляции спленоцитов в присутствии или в отсутствие антигена в течение 6 часов, в присутствии брефелдина А в течение последних 4 часов. IFNg определяли путем пропускания CD4 или СD8 Т-клеток и окрашивания с использованием IFNγFITC.
Сильный CDS-ответ на иммунодоминантный МS3-специфический пептид, достигающий частот до 2,5-6% CD8+ клеток, был также получен после иммунизации с использованием HCV 500, 510, 520 и 530.
Иммунизация с использованием HCV 500, 510, 520 и 530 также приводила к детекции CD4- и CD8-ответов как на NS4B, так и на NS5B антигены, хотя CD8-ответы на эти полипротеины были слабее, чем после иммунизации единичным антигеном.
Частота NS5B CD4- или CD8-специфичных Т-клеток, продуцирующих IFNγ после иммунизации полипротеинами HCV
IFNγ-специфические Т-клеточные ответы определяли после стимуляции спленоцитов в присутствии или в отсутствие антигена в течение 6 часов, в присутствии брефелдина А в течение последних 4 часов. IFNg определяли путем пропускания CD4 или CD8 Т-клеток и окрашивания с использованием IFNγFITC.
Частота NS4B CD4- или CD8-специфичных T-клеток, продуцирующих IFNγ после иммунизации полипротеинами HCV
IFNy-специфические Т-клеточные ответы определяли после стимуляции спленоцитов в присутствии или в отсутствие антигена в течение 6 часов, в присутствии брефелдина А в течение последних 4 часов. IFNg определяли путем пропускания CD4 или CD8 Т-клеток и окрашивания с использованием IFNyFITC.
Используемые пептиды имели следующую последовательность.
Распознавание эндогенно процессированного антигена
Для того чтобы определить, индуцировала ли PMID-иммунизация полипротеинами HCV ответ, который может распознавать эндогенно процессированный антиген, в качестве стимуляторов в ELISPOT-анализе использовали клетки-мишени, инфицированные рекомбинантным вирусом коровьей оспы, экспрессирующим NS3-5. Результаты показывают, что хорошие ELISPOT-ответы на IL2 и IFNγ были определены после иммунизации с использованием 500, 510, 520 и 530 (Фиг.14).
Иммунизация полипротеинами HCV индуцирует функциональную CTL-активность
Мышей C57BL иммунизировали 0,01 мкг ДНК, кодирующей только NS3, HCV 500, 510 и 520. После первичной и однократной повторной иммунизации клетки селезенки из каждой группы повторно стимулировали in vitro с использованием NS3 СD8-пептида и IL2 в течение 5 дней. CTL-активность (CTL, цитотоксические Т лимфоциты) измеряли против EL-4-клеток, импульсно обработанных тем же пептидом. Мыши, иммунизированные всеми конструкциями, показали похожие уровни цитолиза в этом виде анализа.
Эти результаты показывают, что PMID-иммунизация полипротеинами HCV может индуцировать функциональные СD8-ответы. Эти результаты показаны на Фиг.15.
Пример 6. Доставка антигенов HCV посредством конструкции с двойным промотором
Конструкции с двойным промотором создавали, используя следующий способ. Фрагмент, несущий экспрессионную кассету 1 (включая Iowa-length промотор CMV, Экзон 1, ген, который кодирует представляющий интерес белок/слитый белок, плюс поли-А-сигнал глобина кролика), вырезали из его вектора-хозяина, а именно p7313ie, по уникальным эндонуклеазным рестрикционным сайтам Clal и Xmnl. Xmnl дает тупой конец на 3′-конце вырезанного фрагмента.
Реципиентный плазмидный вектор представлял собой p7313ie, содержащий экспрессионную кассету 2. Она была получена путем переваривания с использованием уникальной рестриктазы Sse83871 с последующей инкубацией с ДНК-полимеразой Т4 для удаления созданных 3′-выступов, что давало в результате тупые концы как 5′-, так и 3′- в линейной молекуле. Ее вырезали с использованием уникальной рестриктазы Clal, которая удаляет фрагмент длиной 259 п.н.
Экспрессионную кассету 1 клонировали в р7313iе/экспрессионную кассету 2 через Clal/тупые совместимые концы, образуя р7313iе/экспрессионную кассету 1 + экспрессионную кассету 2, где кассета 1 находится в обратном направлении относительно кассеты 2.
Были созданы следующие плазмиды p7313ie:
Примечание:
стрелка = промотор IE гена цитомегаловируса человека (IE HCMV).
NS4B = укороченный NS4B, содержащий аминокислоты 49-260, как изложено выше.
Core = core-белок, содержащий аминокислоты 1-191.
Показанная выше панель конструкций является полной, и она была подвергнута мониторингу на предмет экспрессии в результате временной трансфекции в клетках 293Т посредством вестерн-блоттинга. Результаты вестерн-блоттинг-анализа показаны на Фиг.16. Обозначение дорожек:

Каждая пара конструкций несет две независимые экспрессионные кассеты. Не ожидали, что порядок, в котором кассеты встроены в вектор, будет оказывать какое-либо влияние на экспрессию с каждой кассеты. Однако эти результаты указывают на значительное препятствие в экспрессии слитых белков NS4B5B или NS34B5B, когда их соответствующие экспрессионные кассеты расположены в прямом направлении относительно кассеты Core, NS3Core или CoreNS3.
Уровень экспрессии не является настолько позитивным, как для конструкций с единичным антигеном, однако некоторое снижение следует ожидать в результате значительного увеличения в размере (175-228%), трансляции на плазмиде с уменьшенным количеством копий, доставленной в клетку в количестве примерно 50% от той же массы ДНК.
In vitro иммуногенность, индуцированная конструкциями с двойным промотором
Для исследований иммуногенности были отобраны три конструкции с двойным промотором, которые показали наибольшую экспрессию всех четырех антигенов. Это p7313ieNS4B/NS5B + Core/NS3, p7313ieNS4B/NS5B + NS3Core и p7313ieNS3/NS4B/NS5B + Core. Мышей C57BL иммунизировали 1 мкг ДНК с использованием PMID, и ответы на доминантный NS3 CD8 Т-клеточный эпитоп определяли через 7 суток, используя ELISPOT для IL2. Результаты (представлены на Фиг.17) показывают, что ответы наблюдали для всех трех конструкций с двойным промотором после однократной иммунизации (спленоциты, стимулированные CD4 и CD8 NS3 Т-клеточными специфическими пептидами).
Пример 7. Делеционные мутации core
Конструировали и полностью секвенировали ряд генов, кодирующих ОРС core, последовательно делегированных по участку, охватывающему 20 аминокислот, за один раз с 3′-конца.
На Фиг.18 представлен ДНК агарозный гель, демонстрирующий диапазон генов, кодирующих фрагменты core. Эти конструкции были протестированы на экспрессию совместно с их влиянием на уровень экспрессии слитой конструкции NS4B5B (p7313ie/NS4B5B) посредством котрансфекции клеток 293Т. Результаты показаны на Фиг.19. На дорожки были нанесены следующие пробы.
Экспрессия Core191, CoreΔ15, Core171, Core151 и Core131 четко определяется, когда вестерн-блоттинг обрабатывают зондом анти-core после анти-NS5B детекции экспрессии NS4B5B. Более укороченные формы core не определялись, возможно из-за размера удерживаемых рестриктов в используемой гель-системе.
Результаты показывают значительное снижение уровня экспрессии NS4B5B в присутствии Core191 и Δ15, который восстанавливается с Core171 и, кроме того, с Core151, несмотря на сильную экспрессию обоих видов core. Это наблюдение повторяли дважды с NS4B5B и один раз с NS3 и NS5B.
Пример 8. Влияние Core и Core151 на экспрессию NS3, NS5B, слитой конструкции NS4B-NS5B и тройной слитой конструкции NS3-NS4B-NS5B
Эксперимент 1. Экспрессия в Транс-формате
Был проведен эксперимент для контроля влияния экспрессии Core191 в сравнении с Core151 на экспрессию неструктурных антигенов, когда core экспрессируется в транс или кодируется отдельной плазмидой. Протокол эксперимента такой же, как описано в Примере 7. Вкратце, 0,5 мкг каждого из двух ДНК-плазмидных векторов, приведенных ниже в таблице, котрансфицировали в клетки НЕК 293Т, используя реагент для трансфекции липофектамин 2000 в соответствии со стандартным протоколом (Invitrogen/Life Technologies). (Трансфекция и вестерн-блоттинг как в Примере 4).
Результаты показаны на Фиг.20, где на дорожки наносили, как описано в предыдущей таблице, и вестерн-блоттинг-анализы проводили с целью определения экспрессии сначала неструктурных белков, используя анти-NS3 и анти-NS5В антисыворотки, и экспрессии core с использованием второго образца того же геля с анти-core.
Во всех случаях было продемонстрировано, что количество неструктурного белка или слитого белка (NS3, NS5B, NS4B-5B) при продуцировании in trans с Core151 было значительно увеличено по сравнению с уровнем, продуцируемым при экспрессии in trans с Core191.
Эксперимент 2. Экспрессия в Цис-формате
Был проведен эксперимент для контроля влияния экспрессии Core191 в сравнении с Core151 на экспрессию неструктурных антигенов, когда core экспрессируется в цис или кодируется той же плазмидой в слиянии с неструктурными элементами. В каждом случае Core151 был замещен на Core191 в С-концевом слиянии с заданной неструктурной областью.
1 мкг ДНК плазмидного вектора, описанного в таблице ниже, трансфицировали в клетки НЕК 293Т, используя реагент для трансфекции липофектамин 2000 в соответствии со стандартным протоколом (Invitrogen/Life Technologies). (Трансфекция и вестерн-блоттинг как в Примере 4).
Результаты показаны на Фиг.21. Вестерн-блоттинг-анализ проводили в Геле А для определения экспрессии сначала неструктурных белков, используя анти-NS3 и анти-NS5В антисыворотки, и экспрессии core с использованием второго образца того же геля с анти-core. На дорожки наносили, как описано в следующей таблице.
Эти результаты указывают на то, что в Цис-формате, где антигены находятся в слитом полипротеине, укорочение core усиливает экспрессию слитого белка.
Сравнение влияния Core191 и Core151 на иммунные ответы на NS3
Мышей C57BL иммунизировали двумя "выстрелами" по 1,5 мкг тотальной ДНК с использованием PMID. Группы для иммунизации включали пустой вектор p7313ie как таковой, золотые гранулы, покрытые совместно p7313ieNS3, p7313ieNS5B и p7313ieCore191 или p7313ieNS3, p7313ieNS5B и p7313ieCore151. Использовали совместное покрытие, поскольку оно осуществляло доставку всех плазмид в одну и ту же клетку, что должно было имитировать описанные выше исследования котрансфекции in vitro. Иммунные ответы на доминантные CD8 и CD4 Т-клеточные эпитопы NS3 определяли через 14 суток после первичной иммунизации, используя внутриклеточное окрашивание цитокинов для измерения IFNγ- и IL2-антигенспецифических ответов. Результаты (приведены на Фиг.22) показывают, что и CD4, и CD8 NS3-ответы были приблизительно в два раза выше в присутствии Core151 по сравнению с Core191.
В другом эксперименте мышей C57BL иммунизировали золотыми гранулами, покрытыми совместно плазмидами, экспрессирующими тройную слитую конструкцию p7313ieNS3/NS4B/NS5B вместе либо с Core191, либо с Core151. Животных затем повторно иммунизировали теми же конструкциями, и ответы на NS3 контролировали через 7 дней после повторной иммунизации, используя внутриклеточное окрашивание цитокинов для измерения ответов. Результаты, представленные на Фиг.23, показывают, что оба NS3-антигенспецифических CD4- и CD8-ответа были приблизительно в 2 раза выше в присутствии Core151 по сравнению с Core191.
Общие исследования in vivo, сравнивающие ответ на NS3 в присутствии core, подтверждают данные по экспрессии in vitro о том, что совместная доставка FL-Core и неструктурных белков может снижать экспрессию неструктурных антигенов, и это снижает иммуногенность конструкций. Этот эффект может быть по меньшей мере частично преодолен путем совместного покрытия укороченным core, из которого удалены 40 С-концевых аминокислот.














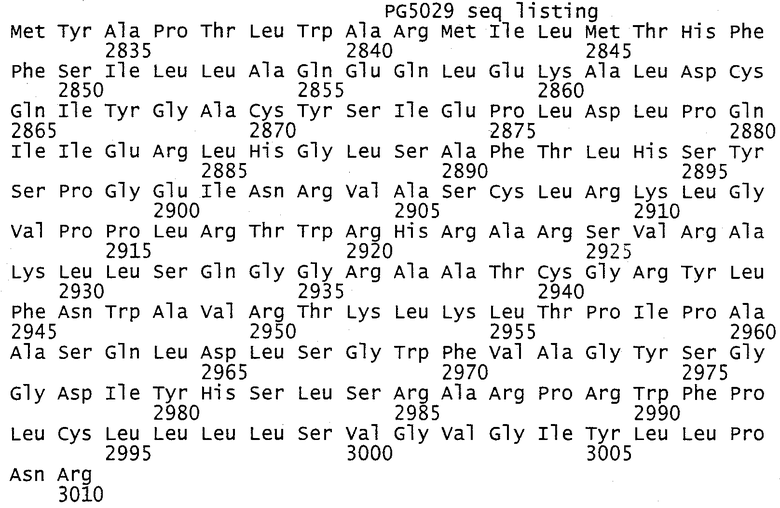
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИНА | 2003 |
|
RU2363492C2 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ КАТИОННЫЕ МИКРОЧАСТИЦЫ И ДНК HCV Е1Е2, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2364419C2 |
| ОПРЕДЕЛЕНИЕ КОМПЛЕКСА HCV-АНТИГЕН/АНТИТЕЛО | 2001 |
|
RU2274863C2 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛЯЦИИ ИММУННОГО ОТВЕТА (ВАРИАНТЫ), СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ, ПРИМЕНЕНИЯ И СПОСОБЫ СТИМУЛЯЦИИ ИММУННОГО ОТВЕТА С ИСПОЛЬЗОВАНИЕМ ЭТИХ КОМПОЗИЦИЙ | 2002 |
|
RU2316347C2 |
| БИОМАРКЕР ДЛЯ МОНИТОРИНГА ПАЦИЕНТОВ | 2010 |
|
RU2542435C2 |
| БИОМАРКЕР ДЛЯ ОТБОРА ПАЦИЕНТОВ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ | 2009 |
|
RU2517719C2 |
| Иммунобиологическое средство для повышения клеточного ответа против вируса гепатита С | 2023 |
|
RU2811136C1 |
| БИОМАРКЕР ДЛЯ МОНИТОРИНГА ПАЦИЕНТОВ | 2010 |
|
RU2555340C2 |
| ОЛИГОМЕРНАЯ ЧАСТИЦА, ИНДУЦИРУЮЩАЯ ИММУНИТЕТ ПРОТИВ ВИРУСА ГЕПАТИТА С, СПОСОБ ПОЛУЧЕНИЯ ОЛИГОМЕРНОЙ ЧАСТИЦЫ, КОМПОЗИЦИЯ, СПЕЦИФИЧЕСКОЕ АНТИТЕЛО, НАБОР (ВАРИАНТЫ), ИММУНОЛОГИЧЕСКИЙ АНАЛИЗ И ВАКЦИНА ПРОТИВ ВИРУСА ГЕПАТИТА С | 1999 |
|
RU2247729C2 |
| ПОЛИПЕПТИД И СПОСОБ ЕГО ПОЛУЧЕНИЯ, РЕАГЕНТ ДЛЯ ИММУНОАНАЛИЗА, СПОСОБ ОПРЕДЕЛЕНИЯ ПРИСУТСТВИЯ АНТИТЕЛ И СПОСОБ ИНДУКЦИИ ИММУННОГО ОТВЕТА | 1992 |
|
RU2148587C1 |





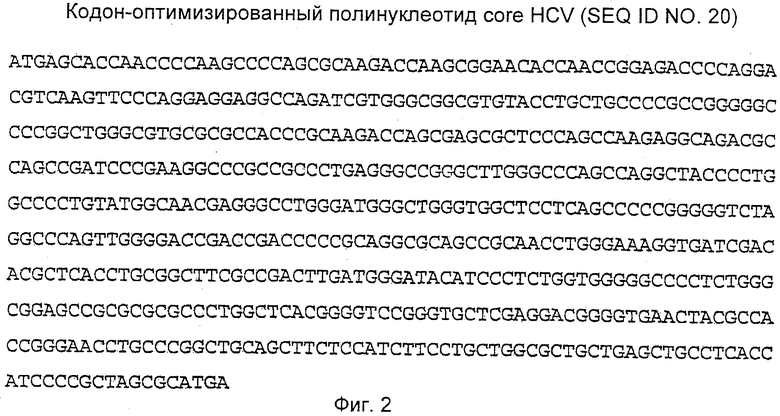

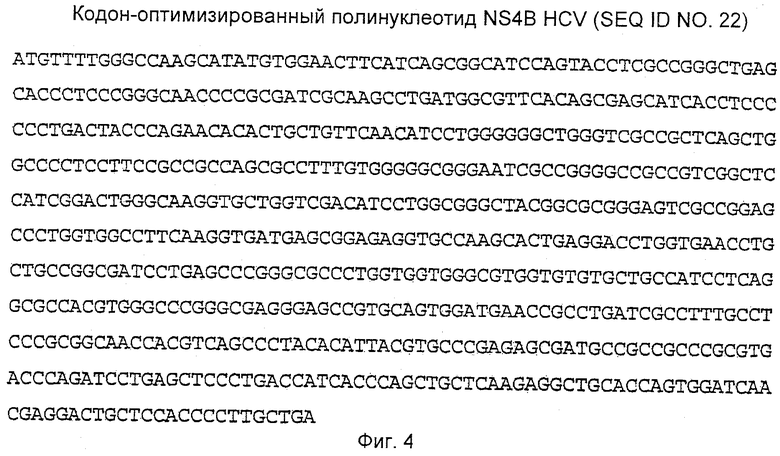





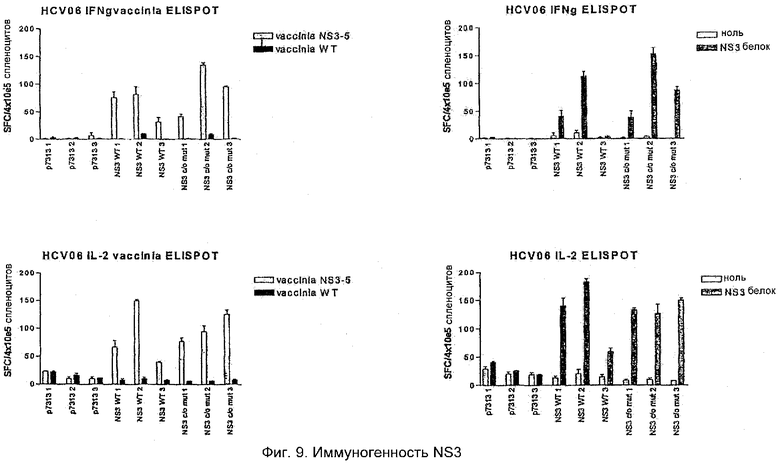
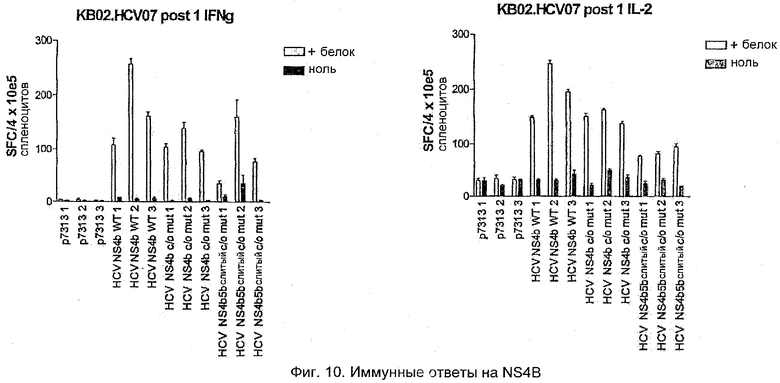
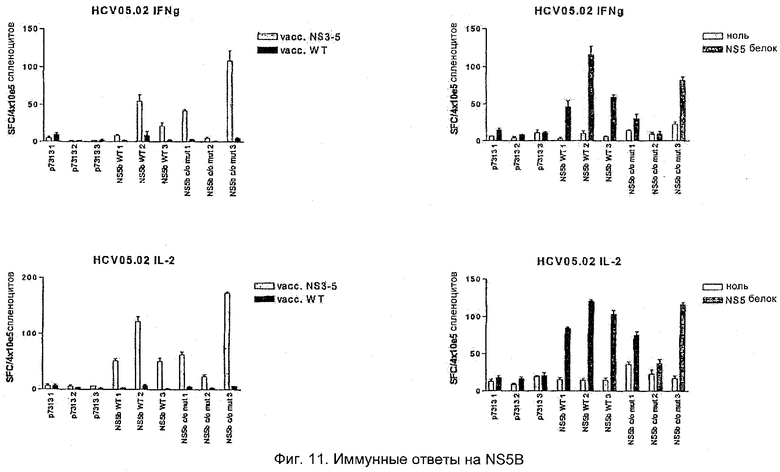
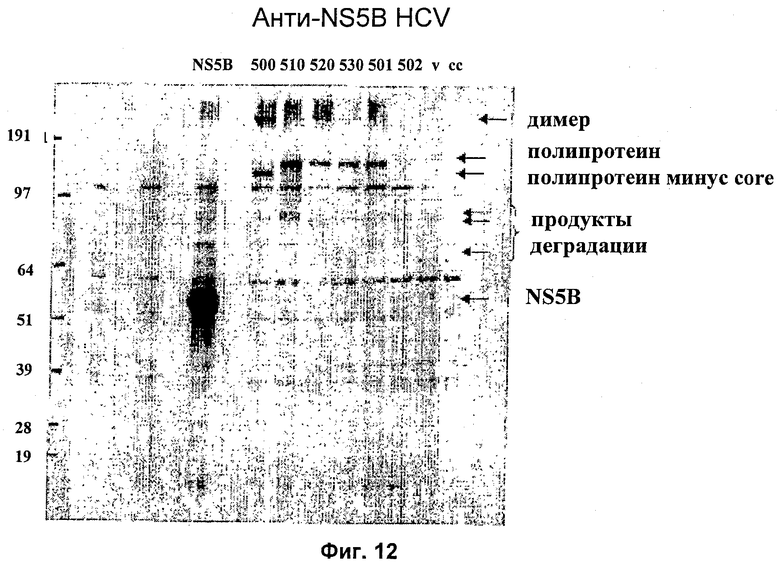
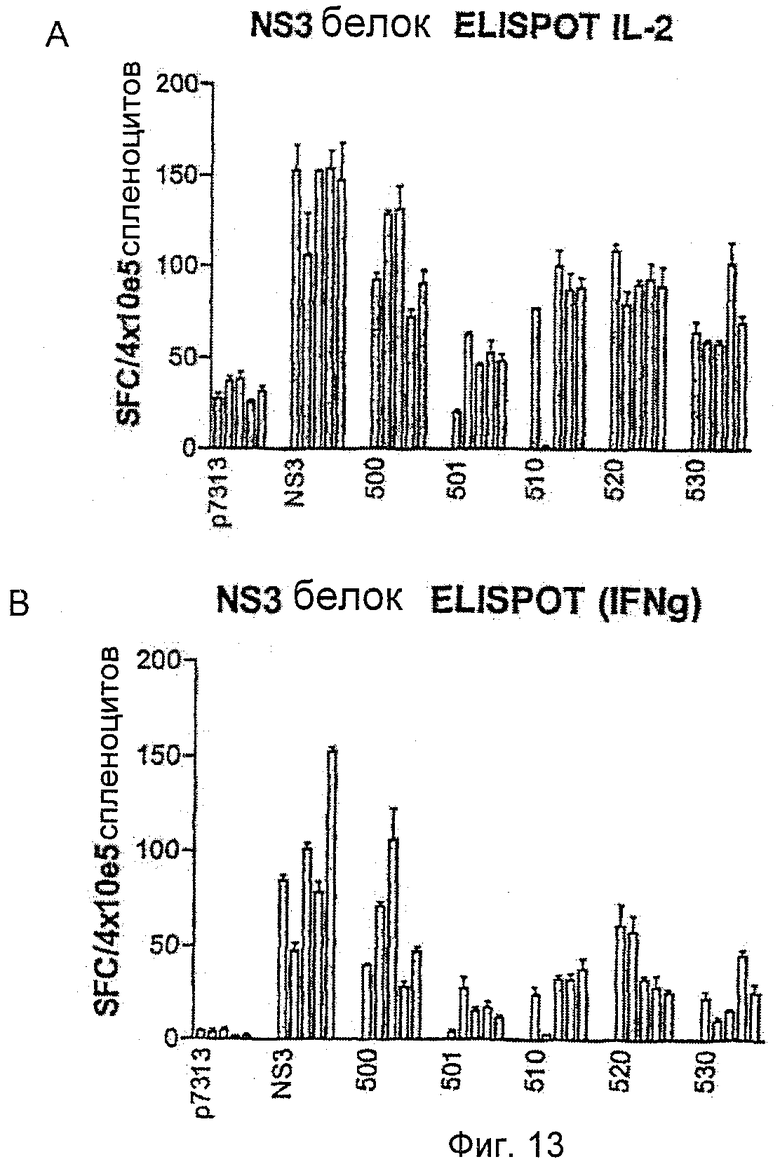
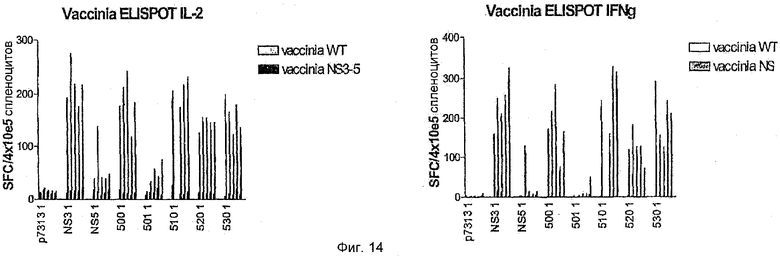
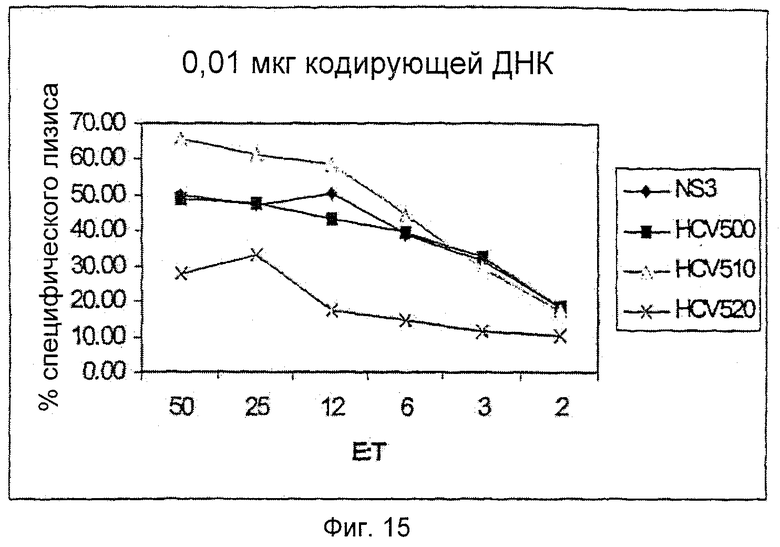
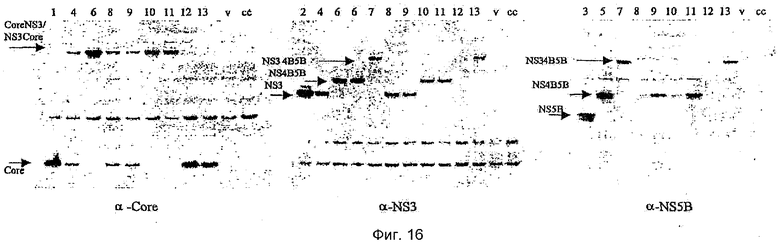
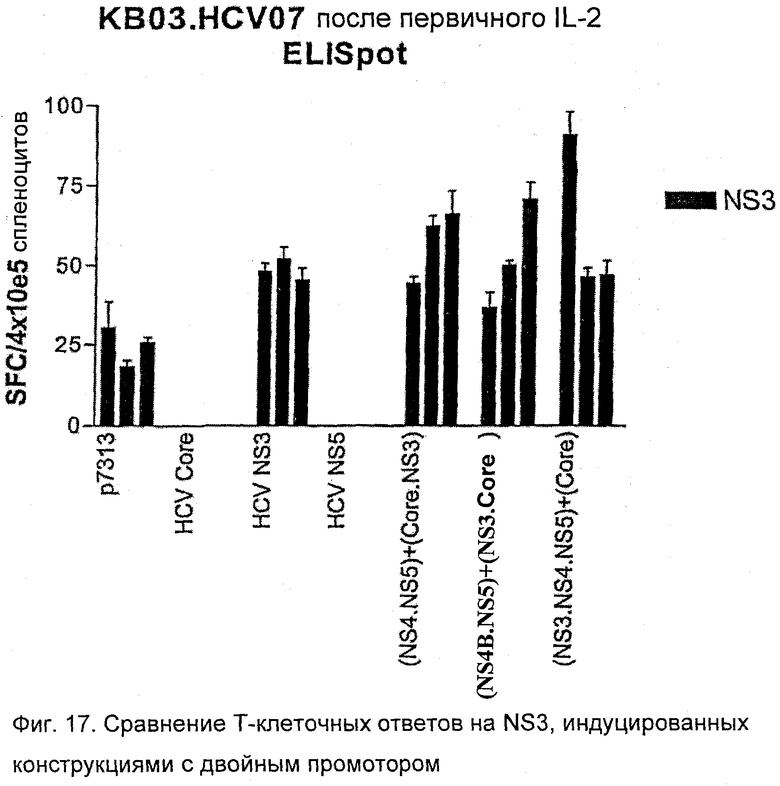

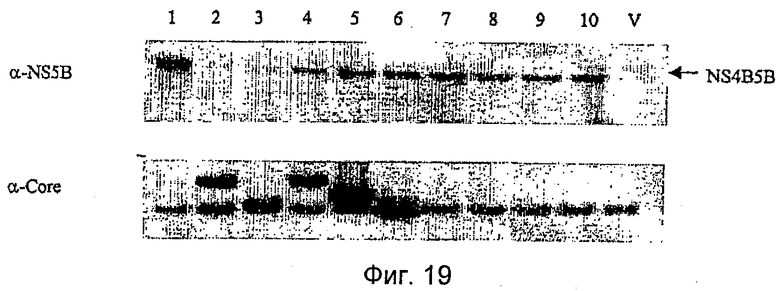
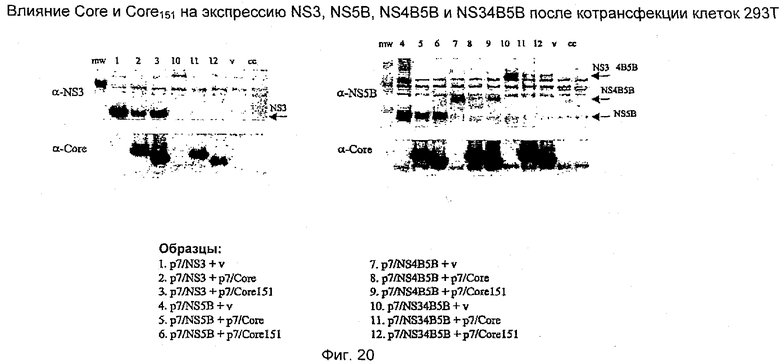
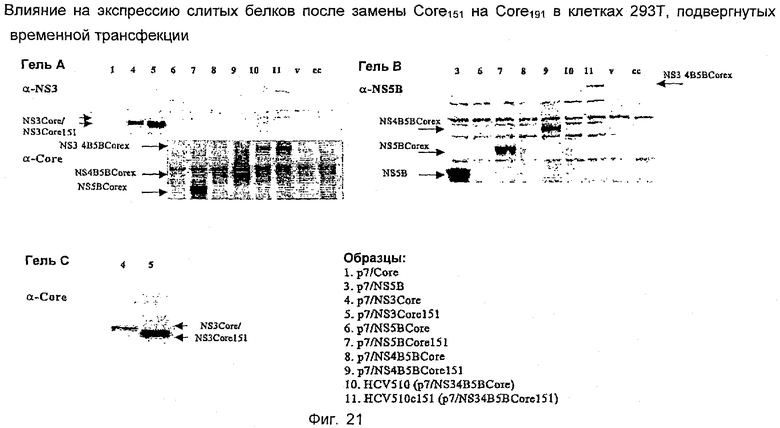
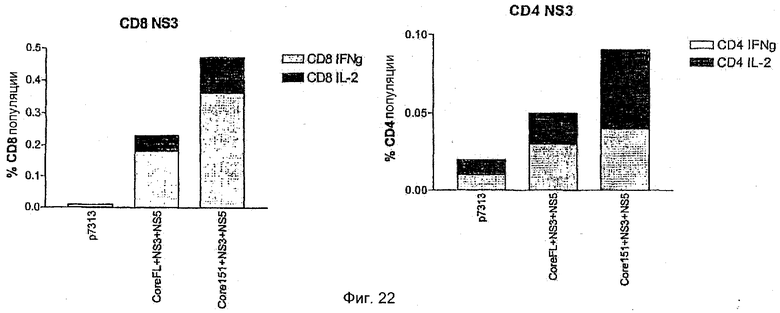
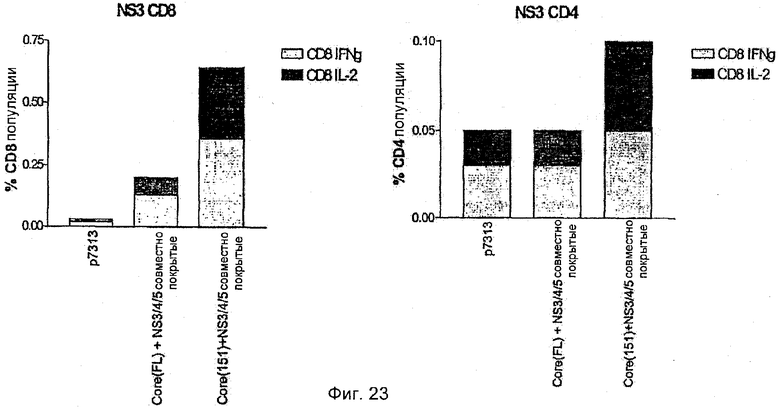
Изобретение относится к способам и композициям, полезным для предупреждения и лечения инфекций, вызванных вирусом гепатита С (HCV). Предложены ДНК-вакцина против HCV, содержащая полинуклеотидные последовательности, кодирующие белки HCV, способ предупреждения и лечения HCV-инфекции, способ вакцинации индивида, а также применение вышеуказанной вакцины для изготовления лекарства. Изобретение может быть использовано для лечения или предупреждения HCV. 5 н. и 19 з.п. ф-лы, 23 ил.
| РЕКОМБИНАНТНЫЙ МОДИФИЦИРОВАННЫЙ ВИРУС ВАКЦИНЫ АНКАРА, СПОСОБНЫЙ ЭКСПРЕССИРОВАТЬ СТРУКТУРНЫЕ АНТИГЕНЫ ВИРУСА ГЕПАТИТА С | 2002 |
|
RU2270860C2 |
| JP 2006117681, 11.05.2006 | |||
| WO 9951781, 14.10.1999 | |||
| Электрод-инструмент для электрохимической обработки | 1974 |
|
SU518313A2 |
| WO 03031588, 17.04.2003 | |||
| US 5371015, 06.12.1994. | |||

