Область техники, к которой относится изобретение
Настоящее изобретение относится в целом к терапии рака. Более конкретно, настоящее изобретение относится к полинуклеотидам, кодирующим слитые белки, причем слитые белки содержат по меньшей мере часть опухолеспецифического полипептидного карциноэмбрионального антигена. Настоящее изобретение также относится к рекомбинантным векторам и хозяевам, содержащим указанные полинуклеотиды, очищенные слитые белки и способы усиления иммунного ответа против СЕА с использованием этих композиций и молекул, описанных здесь.
Уровень техники
Суперсемейство иммуноглобулинов (IgSF) состоит из многочиленных генов, которые кодируют белки с разнообразными функциями, одной из которых является межклеточная адгезия. Белки IgSF содержат по меньшей мере один Ig-связанный домен, который является важным для сохранения правильных взаимодействий межмолекулярного связывания. Поскольку такие взаимодействия являются необходимыми для разнообразных биологических функций членов IgSF, нарушенная или аберрантная экспрессия многих молекул адгезии IgSF коррелировала со многими заболеваниями человека.
Карциноэмбриональный антиген (СЕА) принадлежит к подсемейству суперсемейства Ig, состоящему из гликопротеинов, известных как СЕА-связанные молекулы клеточной адгезии (СЕАСАМ). Было показано, что СЕАСАМ действуют в качестве молекул как гомотипической, так и гетеротипической межклеточной адгезии (Benchimol et al., Cell 57: 327-334 (1989)). Кроме клеточной адгезии, СЕА (также известный как СЕАСАМ5) ингибирует смерть клеток, происходящую при отделении клеток от внеклеточного матрикса, и может способствовать клеточной трансформации (перерождению клеток), связанной с определенными протоонкогенами, такими как Bcl2 и C-Myc (см. Bernstein, J. Clin. Oncol. 20(8): 2197-2207 (2002)). Последовательности, кодирующие СЕА человека, были клонированы и охарактеризованы (патент США № 5274087; патент США № 5571710 и патент США № 6843761. См. также Beauchemin et al., Mol. Cell. Biol. 7:3221-3230 (1987); Zimmerman et al., Proc. Natl. Acad. Sci. USA 84:920-924 (1987); Thompson et al., Proc. Natl. Acad. Sci. USA 84(9):2965-69 (1987)).
Нормальная экспрессия СЕА была обнаружена во время эмбрионального развития и в слизистой оболочке ободочной кишки взрослых. Сверхэкспрессия СЕА была впервые обнаружена в опухолях ободочной кишки человека более тридцати лет тому назад (Gold and Freedman, J. Exp. Med. 121:439-462 (1965)) и с тех пор была обнаружена почти во всех колоректальных опухолях. Кроме того, сверхэкспрессия СЕА обнаруживается в большом проценте аденокарцином поджелудочной железы, печени, молочной железы, яичника, шейки матки и легкого. Вследствие его преобладания в этих типах опухолей и ограниченной экспрессии в нормальных тканях СЕА считают опухолеспецифическим аутоантигеном и мишенью для активной и пассивной иммунотерапии. Недавние клинические данные установили, что различные подходы вакцинации могут образовывать В- и Т-клетки человека, специфические в отношении СЕА, обеспечивая дополнительное доказательство того, что СЕА является мишенью для молекулярного и иммунологического вмешательства для лечения этих типов рака.
Терапевтические подходы нацеливания на СЕА включают в себя использование анти-СЕА-антител (см. Chester et al., Cancer Chemother. Pharmacol. 46 (Suppl): S8-S12 (2000)), а также вакцин на основе СЕА (для обзора см. Berinstein, supra). Развитие и промышленное внедрение многих вакцин тормозились трудностями, связанными с получением высоких уровней экспрессии экзогенных генов. Успех вакцин на основе ДНК также тормозился неспособностью получать иммунный ответ достаточного размера у индивидуумов. Хотя были разработаны ДНК-вакцины, нацеленные на различные белки, полученные иммунные ответы были относительно слабыми по сравнению с общепринятыми вакцинами.
Легкость манипуляции ДНК является благоприятной возможностью разработки вакцин, в том числе стратегий слитой конструкции генов, в которых антигены связаны с различными иммуноусиливающими элементами. Усиление иммунного ответа на антигены-мишени было продемонстрировано на моделях животных с использованием векторов, кодирующих антигены, слитые с белком теплового шока (HSP) 70 (Liu et al., J. Virol. 74: 2888-94 (2000); Cheng et al., J. Immunol. 166: 6218-26 (2001); Chen et al., Cancer Res. 60: 1035-42 (2000)) с Fc-частью IgG1 (You et al., J. Immunol. 165: 4581-92 (2000)), со связанным с лизосомой мембранным белком (LAMP) (Su et al., Cancer Res. 62: 5041-48 (2002)) и универсальным Тh-эпитопом из столбнячного токсина (Renard et al., J. Immunol. 171:1588-95 (2003); King et al., Nature Med. 4: 1281-86 (1998); Lund et al., Cancer Gene Ther. 10: 365-76 (2003); Padua et al., Nature Med. 9(11): 1413-17 (2003); Savelyeva et al., Nature Biotechnol. 19: 760-64 (2001); Wahren et al., WO 2004/092216). Усиление иммунных ответов на антигены-мишени является особенно важным для противораковых вакцин ввиду ограниченной иммуногенности опухолевых антигенов и необходимости преодоления толерантности к проявлению эффективных противоопухолевых эффектов.
Таким образом, несмотря на идентификацию нуклеотидных последовательностей дикого типа, кодирующих белки СЕА, описанные выше, была бы крайне желательна разработка вакцины, которая способна индуцировать усиленный СЕА-специфический иммунный ответ в отношении полноразмерной кДНК СЕА дикого типа при введении млекопитающему. Была бы также желательна разработка способов лечения или профилактики СЕА-обусловленных раков, использующих молекулы нуклеиновых кислот или белки, которые безопасно и эффективно усиливают СЕА-специфический иммунный ответ.
Сущность изобретения
Настоящее изобретение относится к полинуклеотидам, кодирующим слитые белки, причем эти слитые белки содержат по меньшей мере часть опухолеспецифического полипептидного карциноэмбрионального антигена, слитую с существенной частью иммуноусиливающего элемента, такого как бактериальный токсин. В предпочтительных вариантах осуществления СЕА-часть кодируемого слитого белка СЕА делетирована на С-конце якорного домена. В предпочтительных вариантах осуществления иммуноусиливающим элементом является А- или В-субъединица термолабильного энтеротоксина E. coli или его существенная часть. В других предпочтительных вариантах осуществления иммуноусиливающим элементом является минимизированный домен фрагмента С столбнячного токсина (DOM) или его существенная часть. Настоящее изобретение также относится к рекомбинантным векторам, в том числе, но не только, аденовирусам и плазмидным векторам, содержащим указанные полинуклеотиды, и клеткам-хозяевам, содержащим указанные рекомбинантные векторы. Представлены также очищенные слитые белки, кодируемые полинуклеотидами настоящего изобретения.
Дополнительно настоящее изобретение относится к способам ингибирования или профилактики развития рака у млекопитающего путем индукции иммунного ответа на белок СЕА введением вакцины или фармацевтической композиции, содержащей описанные здесь слитые конструкции СЕА или слитые белки. В предпочтительных вариантах описанных здесь способов такой иммунный ответ усилен по сравнению с ответом, индуцированным вакциной СЕА дикого типа.
В контексте всего описания изобретения и прилагаемой формулы изобретения используются следующие определения и аббревиатуры:
Термин «промотор» относится к сайту распознавания на цепи ДНК, с которой связывается РНК-полимераза. Промотор образует инициирующий комплекс с РНК-полимеразой для инициации и запуска транскрипционной активности. Этот комплекс может быть модифицирован активирующими последовательностями, называемыми «энхансерами», или ингибирующими последовательностями, называемыми «сайленсерами».
Термин «кассета» обозначает нуклеотидную или генную последовательность, которая экспрессируется из вектора, например, нуклеотидная или генная последовательность, кодирующая слитую конструкцию hCEA-LTB. Обычно кассета содержит последовательность гена, которая может быть встроена в вектор, который в некоторых вариантах осуществления имеет регуляторные последовательности для экспрессии нуклеотидной или генной последовательности. В других вариантах осуществления эта нуклеотидная или генная последовательность имеет некоторые регуляторные последовательности для их экспрессии. В дополнительных вариантах осуществления, вектор имеет некоторые регуляторные последовательности, а нуклеотидная или генная последовательность имеет другие регуляторные последовательности. Например, вектор может иметь промотор для транскрибирования нуклеотидной или генной последовательности, а нуклеотидная или генная последовательность имеет последовательность терминации транскрипции. Регуляторные последовательности, которые могут находиться в векторе, включают в себя, но ими не ограничиваются, энхансеры, последовательности терминации транскрипции, акцепторные и донорные последовательности сплайсинга, интроны, последовательности связывания рибосом и последовательности поли(А)-добавления.
Термин «вектор» относится к одному из средств, при помощи которого ДНК-фрагменты могут быть введены в организм-хозяин или в ткань-хозяин. Имеются различные типы векторов, в том числе плазмида, вирус (в том числе аденовирус), бактериофаги и космиды.
Термин «первая генерация», в применении к аденовирусным векторам, описывает аденовирусные векторы, которые являются дефектными по репликации. Аденовирусные векторы первой генерации обычно имеют делетированный или инактивированный район гена Е1 и, предпочтительно, имеют делетированный или инактивированный район гена Е3.
Аббревиатура «DOM» обычно обозначает N-концевой домен фрагмента С столбнячного токсоида.
Аббревиатура «LT» обычно обозначает термолабильный энтеротоксин E. coli. «LT» может обозначать полный энтеротоксин, содержащий субъединицы А и В или существенную часть субъединицы А, или существенную часть субъединицы В. Аббревиатура “LTA” обозначает субъединицу А термолабильного энтеротоксина E. coli или ее существенную часть, в том числе субъединицы, которые укорочены на С-конце или N-конце, но сохраняют биологическую активность, а также субъединицы, которые содержат внутренние аминокислотные инсерции, делеции или замены, но сохраняют биологическую активность. Аббревиатура «LTB» обозначает В-субъединицу термолабильного энтеротоксина E. coli или ее существенную часть, в том числе субъединицы, которые укорочены на С-конце или N-конце, но сохраняют биологическую активность, а также субъединицы, которые содержат внутренние аминокислотные инсерции, делеции или замены, но сохраняют биологическую активность.
Под обозначением «pV1J/hCEAopt» понимают плазмидную конструкцию, описанную здесь, содержащую предранний (IE) промотор CMV с интроном А, полноразмерный оптимизированный в отношении кодонов ген СЕА человека, полученные из бычьего гормона роста последовательности полиаденилирования и терминации транскрипции и минимальный каркас pUC (см. пример 2). Под обозначением «pV1J/hCEA» понимают конструкцию, по существу такую же, что и описанная выше, за исключением того, что эта конструкция содержит ген СЕА дикого типа человека вместо оптимизированного в отношении кодонов гена СЕА человека.
Под обозначением «pV1J/hCEA-LTB» понимают плазмидную конструкцию, описанную здесь, содержащую предранний (IE) промотор CMV с интроном А, ген СЕА человека, лишенный его кодирующей якорь GPI последовательности, слитый на его С-конце с субъединицей В термолабильного энтеротоксина E. coli, полученные из бычьего гормона роста последовательности полиаденилирования и терминации транскрипции и минимальный каркас pUC.
Под обозначением «pV1J/hCEAopt-LTB» понимают конструкцию, по существу такую же, как описано непосредственно выше, за исключением того, что эта конструкция содержит оптимизированный в отношении кодонов ген СЕА человека, лишенный его кодирующей якорь GPI последовательности, вместо соответствующей части гена СЕА дикого типа человека.
Под обозначением «pV1J/hCEAopt-LTBopt» понимают конструкцию, по существу такую же, как описано непосредственно выше, за исключением того, что как последовательности СЕА, так и последовательности LTB являются оптимизированными в отношении кодонов для экспрессии высокого уровня в клетках человека.
Под обозначением «pV1J/rhCEAopt-LTBopt» понимают конструкцию, по существу такую же, как описано выше, за исключением того, что оптимизированный в отношении кодонов ген СЕА человека заменен геном СЕА макака-резуса, оптимизированным в отношении кодонов, для экспрессии высокого уровня в клетках человека.
Под обозначением «pV1J/hCEA-LTА» понимают плазмидную конструкцию, описанную здесь, содержащую предранний (IE) промотор CMV с интроном А, ген СЕА человека, лишенный его кодирующей якорь GPI последовательности, слитый на его С-конце с субъединицей А термолабильного энтеротоксина E. coli, полученные из бычьего гормона роста последовательности полиаденилирования и терминации транскрипции и минимальный каркас pUC. Конструирование плазмидных векторов, содержащих различные слитые конструкции СЕА-LT, описано в примере 2.
Под обозначением «pV1J/hCEA-DOM» понимают плазмидную конструкцию, описанную здесь, содержащую предранний (IE) промотор CMV с интроном А, ген СЕА человека, лишенный его кодирующей якорь GPI последовательности, слитый на его С-конце с N-концевым доменом фрагмента С столбнячного токсоида (DOM), полученные из бычьего гормона роста последовательности полиаденилирования и терминации транскрипции, и минимальный каркас pUC (пример 2).
Под обозначением «pV1J/rhCEAopt-DOMopt» понимают конструкцию, по существу такую же, как описано выше, за исключением того, что оптимизированный в отношении кодонов ген СЕА человека заменен геном СЕА макака-резуса, оптимизированным в отношении кодонов, для экспрессии высокого уровня в клетках человека.
Под обозначением «pV1J/hCEA-FcIgG» понимают плазмидную конструкцию, описанную здесь, содержащую предранний (IE) промотор CMV с интроном А, ген СЕА человека, лишенный кодирующей якорь GPI последовательности, слитый на его С-конце c тяжелым фрагментом константной цепи иммуноглобулина G1, полученные из бычьего гормона роста последовательности полиаденилирования и терминации транскрипции, и минимальный каркас pUC (пример 2). pV1J/hCEAopt-FcIgGopt обозначает конструкцию, по существу такую же, как описано, за исключением того, что нуклеотидные последовательности, кодирующие СЕА и FcIgG, были оптимизированы в отношении кодонов для экспрессии высокого уровня в клетках человека.
Под обозначением «Ad5/hCEAopt» и «Ad5/hCEA» понимают две конструкции, описанные здесь, которые содержат аденовирусный геном Ad5, в котором делетированы районы Е1 и Е3. В конструкции «Ad5/hCEAopt» район Е1 заменен оптимизированным в отношении кодонов геном СЕА человека в параллельной Е1 ориентации под контролем промотора CMV человека без интрона А, за которым следует сигнал полиаденилирования бычьего гормона роста. Конструкция «Ad5/hCEA» является по существу такой же, как описано выше, за исключением того, что район Е1 генома Ad5 заменен последовательностью СЕА дикого типа человека. Под обозначением «Ad5/hCEAopt-LTB» понимают конструкцию Ad5, по существу такую же, как описано выше, за исключением того, что оптимизированная в отношении кодонов последовательность СЕА человека лишена кодирующей якорь GPI последовательности и слита на ее С-конце с субъединицей В термолабильного энтеротоксина E. coli. Конструирование аденовирусных векторов, содержащих различные слитые конструкции СЕА-LT, описано в примере 3.
«Иммуноусиливающий элемент» обозначает часть слитых белков СЕА настоящего изобретения, которая способна стимулировать или усиливать иммунный ответ на связанный белок СЕА, родственный полноразмерному СЕА дикого типа. Иммуноусиливающие элементы настоящего изобретения выбраны из группы, состоящей из белка теплового шока (HSP) 70, связанного с лизосомой мембранного белка (LAMP), фрагмента С столбнячного токсоида (FrC), N-концевого домена FrC (DOM), тяжелого фрагмента константной цепи иммуноглобулина G1 (FcIgG), гликопротеина вируса везикулярного стоматита (VSV-G), холерного токсина (СТ) из Vibrio cholerae и термолабильного энтеротоксина E. coli (LT). Термин «иммуноусиливающий элемент» используется взаимозаменяемо с термином «адъювант».
Как используется здесь, «слитый белок» обозначает белок, имеющий по меньшей мере два ковалентно связанных полипептида, где один полипептид происходит из одной последовательности белка или домена, а другой полипептид происходит из второй последовательности белка или домена. Слитые белки по изобретению содержат полипептид СЕА или его фрагмент или вариант и второй полипептид, который содержит существенную часть иммуноусиливающего элемента, который, в некоторых случаях, является бактериальным токсином. Полипептид СЕА, его фрагмент или вариант может быть СЕА человека или СЕА-гомологом другого вида. Полипептиды, которые составляют слитый белок, предпочтительно, связаны по типу N-конец - С-конец. Полипептид СЕА и субъединица токсина могут быть слиты в любом порядке. В некоторых вариантах осуществления настоящего изобретения, С-конец полипептида СЕА слит с N-концом субъединицы токсина, как представлено, например, на фигуре 1А. Однако также рассматриваются слитые белки, в которых иммуноусиливающий элемент слит с N-концом полипептида СЕА. Предполагается, что термин «слитый белок СЕА» является общим термином, который относится к слитой конструкции, описанной выше, которая содержит полипептид СЕА или его фрагмент или вариант, слитый с полипептидом, содержащим иммуноусиливающий элемент.
Термин «слитая конструкция СЕА-LT» обозначает последовательность нуклеиновой кислоты, в которой по меньшей мере часть гена СЕА слита с существенной частью либо субъединицы LTA, либо субъединицы LTB термолабильного энтеротоксина E. coli. Термин «слитый белок СЕА-LT» обозначает полипептид, кодируемый слитой конструкцией СЕА-LT. Предполагается, что термины «слитая конструкция СЕА-LT» и «слитый белок СЕА-LT» также обозначают их фрагменты, их гомологи и их функциональные эквиваленты (вместе называемые «вариантами») такие как фрагменты, гомологи и эквиваленты, в которых одна или несколько аминокислот инсертированы, делетированы или заменены другой аминокислотой (другими аминокислотами). Слитые конструкции СЕА-LT по изобретению при введении млекопитающему, например, человеку, могут стимулировать иммунный ответ посредством хелперных Т-клеток или цитотоксических Т-клеток или могут стимулировать продуцию антител по меньшей мере так же хорошо, как и последовательность СЕА «дикого типа». В предпочтительных вариантах осуществления изобретения слитая конструкция СЕА-LT может усиливать иммунный ответ по сравнению с СЕА дикого типа.
Термин «слитая конструкция СЕА-DOM» обозначает последовательность нуклеиновой кислоты, в которой по меньшей мере часть гена СЕА слита с существенной частью минимизированного домена фрагмента С столбнячного токсина, если контекст ясно указывает на то, что указанный термин относится к этой белковой последовательности. Термин «слитый белок СЕА-DOM» обозначает полипептид, кодируемый описанной слитой конструкцией СЕА-DOM. Предполагается, что термины «слитая конструкция СЕА-DOM» и «слитый белок СЕА-DOM» также обозначают их фрагменты, их гомологи и их функциональные эквиваленты (вместе называемые «вариантами»), такие как фрагменты, гомологи и эквиваленты, в которых одна или несколько аминокислот инсертированы, делетированы или заменены другой аминокислотой (другими аминокислотами). Слитые конструкции СЕА-DOM по изобретению при введении млекопитающему, например, человеку, могут стимулировать иммунный ответ посредством хелперных Т-клеток или цитотоксических Т-клеток или могут стимулировать продукцию антител по меньшей мере так же хорошо, как и последовательность СЕА «дикого типа». В предпочтительных вариантах осуществления изобретения слитая конструкция СЕА-DOM может усиливать иммунный ответ по сравнению с СЕА дикого типа.
Аббревиатура «AD» обозначает якорный домен гена или белка СЕА. Якорный домен СЕА дикого типа человека расположен от приблизительно аминокислоты 679 до приблизительно аминокислоты 702 последовательности SEQ ID NO:20.
Термин «лечение» относится как к терапевтическому лечению, так и профилактическим или превентивным мерам. Индивидуумы, нуждающиеся в лечении, включают в себя как индивидуумов с уже имеющимися нарушениями, так и индивидуумов, которые предрасположены к возникновению этого нарушения, или индивидуумов, у которых должно быть предотвращено это нарушение.
«Нарушение» является любым состоянием, которое бы имело пользу от лечения молекулами по изобретению, в том числе молекулами нуклеиновых кислот, описанных здесь, и слитыми белками, которые кодируются указанными молекулами нуклеиновых кислот. Термин «нарушение» включает в себя хронические и острые нарушения или заболевания, в том числе патологические состояния, которые делают млекопитающее предрасположенным к рассматриваемому нарушению. Предполагается применение молекул изобретения для лечения нарушений или состояний, отличающихся патологической пролиферацией, включающих в себя, но ими не ограничивающихся, рак молочной железы, колоректальный рак и рак легкого.
Термин «эффективное количество» обозначает количество вакцинной композиции, достаточное при его введении для получения адекватных уровней полипептида, так что индуцируется иммунный ответ. Специалисту в данной области будет понятно, что этот уровень может изменяться.
«Консервативная замена аминокислот» обозначает замену одного аминокислотного остатка другим химически сходным аминокислотным остатком. Примерами таких консервативных замен являются замена одного гидрофобного остатка (изолейцина, лейцина, валина или метионина) другим; замена одного полярного остатка другим полярным остатком с таким же зарядом (например, замена лизина на аргинин; аспарагиновой кислоты на глутаминовую кислоту).
«hCEA» и «hCEAopt» обозначают карциноэмбриональный антиген человека и оптимизированный в отношении кодонов карциноэмбриональный антиген человека, соответственно.
«rhCEA» и «rhCEAopt» обозначают карциноэмбриональный антиген макака-резуса и оптимизированный в отношении кодонов карциноэмбриональный антиген макака-резуса, соответственно.
«По существу сходные» обозначает, что конкретная последовательность нуклеиновой кислоты или аминокислотная последовательность идентична по меньшей мере на 75%, предпочтительно, на 85%, более предпочтительно, на 90% и, еще более предпочтительно, на 95% ссылочной последовательности. В изобретении ссылочной последовательностью могут быть релевантные части нуклеотидной или аминокислотной последовательности дикого типа СЕА человека или нуклеотидная или аминокислотая последовательность дикого типа бактериального токсина или его субъединицы, например, субъединиц LTB или LTA термолабильного энтеротоксина E. coli, как указывается в описании. Ссылочной последовательностью может быть, например, последовательность СЕА дикого типа макака-резуса. Таким образом, последовательность белка СЕА, которая является «по существу сходной» с белком СЕА дикого типа человека или его фрагментом, будет идентична по меньшей мере на 75% релевантному фрагменту СЕА дикого типа человека по длине этого фрагмента, идентична, предпочтительно, на 85%, более предпочтительно, на 90% и, еще более предпочтительно, на 95%. Является ли конкретная последовательность белка или нуклеотидная последовательность СЕА, LTB или LTA «по существу сходной» со ссылочной последовательностью, может быть определено, например, сравнением последовательности с использованием программы для анализа последовательности, такой как компьютерная программа GAP, версия 6.0, доступная из University of Wisconsin Genetics Computer Group (UWGCG). Программа GAP использует способ сопоставления Needleman and Wunsch (J. Mol. Biol. 48:443, 1970), проверенный Smith and Waterman (Adv. Appl. Math. 2:482, 1981).
«Существенная часть» гена, его варианта, фрагмента или субъединицы обозначает часть по меньшей мере 50%, предпочтительно, 75%, более предпочтительно, 90% или, еще более предпочтительно, 95%, ссылочной последовательности.
«Ген» обозначает молекулу нуклеиновой кислоты, нуклеотидная последовательность которой кодирует молекулу полипептида. Гены могут быть непрерывными последовательностями нуклеотидов или они могут включать в себя такие встроенные сегменты, как интроны, промоторные районы, сайты сплайсинга и повторяющиеся последовательности. Геном может быть либо РНК, либо ДНК. Предпочтительным геном является ген, который кодирует пептид по изобретению.
Термин «нуклеиновая кислота» или «молекула нуклеиновой кислоты» обозначает рибонуклеиновую кислоту (РНК) или дезоксирибонуклеиновую кислоту (ДНК), зонды, олигонуклеотиды, их фрагмент или их части и праймеры. ДНК может быть либо комплементарной ДНК (кДНК), либо геномной ДНК, например, геном, кодирующим слитый белок СЕА.
«СЕА дикого типа», или «белок дикого типа», или «wt-белок» обозначает белок, содержащий природную последовательность аминокислот, или его вариант. Аминокислотная последовательность СЕА дикого типа человека показана на фигуре 7Е (SEQ ID NO:20). Аминокислотная последовательность СЕА дикого типа макака-резуса была описана ранее (WO 2004/072287, см. фигуры 7А-7В).
«Ген СЕА дикого типа» обозначает ген, содержащий последовательность нуклеотидов, которая кодирует природный белок СЕА, в том числе белки, полученные у человека, или белки, полученные из другого организма, в том числе, но не только, других млекопитающих, таких как крыса, мышь и макак-резус. Нуклеотидная последовательность гена СЕА человека является доступной в этой области (supra). См. также Beauchemin et al., Mol. Cell. Biol. 7:3221-3230 (1987); Zimmerman et al., Proc. Natl. Acad. Sci. USA 84:920-924 (1987); Thompson et al., Proc. Natl. Acad. Sci. USA 84(9):2965-69 (1987). Нуклеотидная последовательность гена дикого типа макака-резуса показана на фигурах 7С-7D.
Термин «млекопитающее» относится к любому млекопитающему, в том числе и человеку.
Аббревиатура «Ag» обозначает антиген.
Аббревиатуры «Ab» и «mAb» обозначают антитело и моноклональное антитело, соответственно.
Аббревиатура «ORF» обозначает открытую рамку считывания гена.
Краткое описание фигур
На фигуре 1 схематически представлены векторы, разработанные в этом исследовании. Показаны основные признаки плазмидных векторов и Ad-векторов, кодирующих слитые конструкции СЕА-LTA и СЕА-LTB. Показаны также инвертированные концевые повторы (ITR) генома Ad5.
На фигуре 2 показаны нуклеотидная (SEQ ID NO:7, панель А) и аминокислотная (SEQ ID NO:8, панель В) последовательности слитой конструкции hCEA-LTA, приведенной в качестве примера. Нуклеотидная последовательность LTA показана жирным шрифтом.
На фигуре 3 показаны нуклеотидная (SEQ ID NO:9, панель А) и аминокислотная (SEQ ID NO:10, панель В) последовательности слитой конструкции hCEA-LTВ, приведенной в качестве примера. Нуклеотидная последовательность LTВ показана жирным шрифтом.
На фигуре 4 показана нуклеотидная последовательность (SEQ ID NO:11) слитой конструкции hCEAopt-LTB, приведенной в качестве примера. Нуклеотидная последовательность LTB показана жирным шрифтом.
На фигуре 5 показаны нуклеотидная (SEQ ID NO:12, панель А) и аминокислотная (SEQ ID NO:13, панель В) последовательности полностью оптимизированной слитой конструкции hCEA-LTB, обозначенной здесь hCEAopt-LTBopt, приведенной в качестве примера. Нуклеотидная и аминокислотная последовательности LTB показаны жирным шрифтом. Последовательности мест соединений, созданные стратегией клонирования, используемой для слитой конструкции последовательностей СЕА и LTB, подчеркнуты.
На фигуре 6 показаны нуклеотидная (SEQ ID NO:14, панель А) и аминокислотная (SEQ ID NO:15, панель В) последовательности полностью оптимизированной слитой конструкции CEA-LTB макака-резуса, обозначенной здесь rhCEAoptLTBopt, приведенной в качестве примера. Нуклеотидная и аминокислотная последовательности LTB показаны жирным шрифтом. Последовательности мест соединений, созданные стратегией клонирования, используемой для слитой конструкции последовательностей СЕА и LTB, подчеркнуты.
На фигуре 7 показаны нуклеотидные последовательности генов дикого типа, кодирующих СЕА макака-резуса (панели А и В, SEQ ID NO:16 и 17) и аминокислотные последовательности соответствующих белков (панели С и D, SEQ ID NO:18 и 19), описанных ранее (U.S.S.N. 60/447203). Панель Е показывает аминокислотную последовательность СЕА дикого типа человека (SEQ ID NO:20), которая была описана ранее (см., например, патент США № 5274087).
На фигуре 8 показано сравнение эффективности экспрессии в клетках, трансфицированных различными конструкциями СЕА. Панель А изображает эффективности экспрессии клеток HeLa, трансфицированных 3 мкг плазмид, несущих последовательности дикого типа hCEA, hCEA-LTA и hCEA-LTB, вместе с 0,2 мкг плазмиды pV1J/mEPO в качестве индикатора. Панель В показывает результаты из сходного эксперимента по трансфекции с использованием pV1J/hCEAopt и pV1J/hCEAopt-LTB. Эффективность экспрессии определяли через 3 дня после трансфекции измерением количества белка СЕА, присутствующего в клеточных экстрактах и нормализацией этой величины в отношении экспрессии ЕРО. Показанные результаты относятся к средним величинам экспрессии СЕА двух независимых трансфекций.
На фигуре 9 показано сравнение эффективности экспрессии различных аденовирусных рекомбинантных векторов, экспрессирующих СЕА. Клетки HeLa инфицировали при множественности заражения (moi) 100 и 1000 Ad/hCEAopt и Ad/hCEAopt-LTB. Эффективность экспрессии определяли измерением через 3 дня после инфицирования эффективного количества белка СЕА, высвобождаемого в клеточные экстракты. Показанные результаты отражают средние величины экспрессии СЕА двух независимых инфицирований.
На фигуре 10 показан анализ клеточно-опосредованного иммунного ответа, индуцированного различными плазмидными векторами, кодирующими СЕА человека. Трем группам мышей С57BL/6 внутримышечно электроинъецировали 50 мкг указанной плазмиды (СЕА, слитая конструкция СЕА-LTA или слитая конструкция СЕА-LTB) при 0-3 неделях. Четвертую группу мышей иммунизировали смесью 25 мкг pV1J/hCEA-LTA и 25 мкг pV1J/hCEA-LTB. Панель А. Через две недели после стимуляции количество IFNγ-секретирующих Т-клеток, специфических в отношении СЕА, определяли анализом ELISPOT на спленоцитах индивидуальных мышей (белые кружки) с использованием пулов пептидов, которые включают в себя весь этот белок. Указаны также геометрические средние величины (черные ромбы). Панель В изображает внутриклеточное окрашивание IFNγ объединенных спленоцитов из иммунизированных мышей с использованием пула пептидов D. Для каждой группы показано неспецифическое продуцирование IFNγ (ДМСО).
На фигуре 11 показаны титры антител мышей, иммунизированных плазмидными ДНК-векторами, кодирующими СЕА. Индивидуальные титры против очищенного белка СЕА человека измеряли при помощи ELISA на сыворотке из индивидуальных мышей, иммунизированных плазмидами pV1J/hCEA, pV1J/hCEA-LTА и pV1J/hCEA-LTВ. Показаны также средние величины (черные ромбы).
На фигуре 12 показан анализ клеточно-опосредованного иммунного ответа, индуцированного различными плазмидными векторами, кодирующими СЕА. Группы из 4 мышей BALB/c электроинъецировали указанной плазмидой, как описано выше (фигура 4). Через две недели после последней инъекции количество IFNγ-секретирующих Т-клеток, специфических в отношении СЕА, определяли анализом ELISPOT на спленоцитах из индивидуальных мышей (белые кружки) с использованием пулов пептидов, которые включают в себя весь этот белок. Указаны также средние величины (черные ромбы).
На фигуре 13 показан анализ реакции СЕА-специфических CD8+ Т-клеток, индуцированной различными плазмидными векторами, кодирующими СЕА. Мышей С57/DR4 электроинъецировали указанной плазмидой, как описано выше (см. фигуру 4). Через две недели после последней инъекции IFNγ-секретирующие Т-клетки окрашивали с использованием пула пептидов D. Для каждой группы показано неспецифическое продуцирование IFNγ (ДМСО).
На фигуре 14 показан анализ реакции СЕА-специфических CD8+ Т-клеток, индуцированной различными плазмидными векторами, кодирующими СЕА. Мышей HHD электроинъецировали указанной плазмидой, как описано выше (см. фигуру 4). Через две недели после последней инъекции выполняли внутриклеточное окрашивание IFNγ с использованием пулов пептидов В и D. Для каждой группы показана неспецифическая продуция IFNγ (ДМСО).
На фигуре 15 показан клеточно-опосредованный и гуморальный иммунный ответ СЕА-трансгенных мышей (N=9), иммунизированных 5 электроинъекциями один раз в неделю указанных плазмид. Общее количество 50 мкг плазмидной ДНК инъецировали внутримышечно при каждой вакцинации. Панель А. Через две недели после последней инъекции количество IFNγ-секретирующих Т-клеток, специфических в отношении СЕА, определяли внутриклеточным окрашиванием на спленоцитах из индивидуальных мышей (кружки) с использованием пула пептидов D. Показаны также величины геометрического среднего (треугольники). Панель В. Индивидуальные титры против очищенного белка СЕА человека измеряли при помощи ELISA на каждой сыворотке из мышей, иммунизированных плазмидами pV1J/hCEAopt и pV1J/hCEA-LTВ. Показаны также величины геометрического среднего (черные ромбы). Эти результаты указывают на то, что слитая конструкция СЕА-LTB разрушает толерантность к СЕА в трансгенных мышах.
На фигуре 16 показан анализ реакции СЕА-специфических CD8+ Т-клеток, индуцированной различными аденовирусными векторами, кодирующими СЕА. СЕА-трансгенных мышей иммунизировали различными дозами Ad/hCEAopt и Ad/hCEAopt-LTB при 0 и 2 неделях. Через две недели после последней инъекции выполняли внутриклеточное окрашивание IFNγ PBMC из каждой индивидуальной мыши с использованием пула пептидов D (черные кружки). Показаны также величины геометрического среднего (черные ромбы). Неспецифическое продуцирование IFNγ (ДМСО) каждой инъецированной группы было меньше или равно 0,01%.
На фигуре 17 показаны результаты исследований по защите от опухолей иммунизированных СЕА-трансгенных мышей, провоцированных клетками МС38-СЕА. Группы из 10 СЕА-трансгенных мышей иммунизировали 5 еженедельными электроинъекциями указанной плазмидной ДНК (50 мкг/инъекция). Через две недели после последней инъекции мышей повторно иммунизировали единственной инъекцией 1·1010 вч (вирусных частиц) соответствующего Ad-вектора. Через 14 дней после повторной иммунизации аденовирусом мышей иммунизировали подкожной инъекцией 5·105 клеток МС38-СЕА. Панель А показывает процент не имеющих опухолей мышей в указанной временной точке. Панель В показывает средние объемы опухолей каждой иммунизированной группы. Эти результаты показывают, что иммунизация СЕА-трансгенных мышей СЕА-LTB защищает мышей от развития опухолей.
Фигура 18. На панели А схематически представлены репрезентативные слитые белки СЕА, использованные в данном исследовании. Векторы, экспрессирующие слитые белки СЕА, были получены из плазмиды pV1Jns, как описано в примере 2. Эти конструкции содержат нуклеотидную последовательность СЕА нуклеотид (nt) 1 - нуклеотид (nt) 2037 с общей делецией 64 аминокислот (аа), соответствующих GPI-якорной последовательности, и экспрессируют СЕА аминокислота (аа) 1 - аминокислота (аа) 679. Указаны также координаты последовательности каждого белка, слитого с СЕА. Панель В показывает экспрессию pV1J-производных конструкций в трансфицированных клетках. Клетки HeLa трансфицировали плазмидами pV1J/CEA-VSV-G, pV1J/CEA-FcIgG, pV1J/CEA-DOM, pV1J/CEA-HSP70, pV1J/CEA-LAMP или pV1J/СЕА и обрабатывали для Вестерн-блот-анализа, как описано в примере 5. Указана специфичность антитела, используемого для Вестерн-блота. Показан белок СЕА (черная стрелка). Показаны также положения стандартов молекулярных масс (в килодальтонах).
На фигуре 19 показано сравнение эффективности экспрессии слитых конструкций СЕА. Клетки HeLa трансфицировали указанными плазмидами и произведенный из СЕА белок, присутствующий в клеточных лизатах (А) и супернатантах (В), измеряли при помощи ELISA, как описано в примере 8. Полученные результаты являются репрезентативными результатами двух независимых экспериментов.
На фигуре 20 показано сравнение иммуногенности различных конструкций, кодирующих слитые белки СЕА. Мышей С57BL/6 электропорировали внутримышечно с использованием 5 или 50 мкг на дозу указанных плазмид. Инъекции проводили в дни 0 и 14. Количество IFNγ-секретирующих Т-клеток в PBMC в каждой индивидуальной мыши определяли с использованием пула пептидов, охватывающего аминокислоты 497-703 (пул D), как описано в примерах 6 и 15. Показано также среднее количество IFNγ-секретирующих Т-клеток (черные кружки). Величины SFC (образующих пятна клеток) pV1J/CEA-DOM и pV1J/CEA-FcIgG значимо отличаются от величин SFC pV1J/CEA. Панель В. Титр антител измеряли при помощи ELISA с использованием очищенного СЕА в качестве субстрата. Показаны средние величины каждой группы, иммунизированной дозой 50 мкг указанной плазмиды. Титры, которые являются значимо отличающимися от титров мышей, инъецированных pV1J/CEA, указаны звездочкой.
На фигуре 21 показана индукция СЕА-специфических иммунных ответов в СЕА-трансгенных мышах. Группы из 12 СЕА-трансгенных мышей иммунизировали плазмидной ДНК (50 мкг/доза, электроинъецированные в четырехглавую мышцу) или аденовирусными векторами (109 вч/доза), несущими оптимизированную в отношении использования кодонов кДНК СЕА, СЕА-DOM или СЕА-FcIgG. СЕА-специфические CD8+ Т-клетки, индуцированные схемой иммунизации ДНК/ДНК (А) и Ad/Ad (С), измеряли внутриклеточным окрашиванием IFNγ на PBMC каждой иммунизированной мыши. Показаны также средние величины для каждой группы (черные кружки). Группы СЕА-DOM и СЕА-FcIgG, иммунизированные со схемами иммунизации ДНК/ДНК и Ad/Ad, значимо отличались от СЕА-вакцинированной группы. Титры СЕА-специфических антител каждой индивидуальной мыши, вакцинированной по схеме иммунизации ДНК/ДНК (В) или Ad/Ad (D), измеряли при помощи ELISA. Титры, индуцированные векторами CEA-DOM и CEA-FcIgG, значимо отличались от титров, индуцированных СЕА.
На фигуре 22 показана иммуногенность схемы ДНК/Ad. Группы из 12 СЕА-трансгенных мышей иммунизировали плазмидной ДНК (50 мкг/доза, электроинъецированные в четырехглавую мышцу) или аденовирусными векторами (109 вч/доза), несущими оптимизированную в отношении использования кодонов кДНК СЕА, СЕА-DOM или СЕА-FcIgG. СЕА-специфические CD8+ Т-клетки измеряли внутриклеточным окрашиванием IFNγ на PBMC каждой иммунизированной мыши (А). Показаны также средние величины для каждой группы (черные кружки). Группы СЕА-DOM и СЕА-FcIgG значимо отличались от СЕА-вакцинированной группы. Титры СЕА-специфических антител каждой индивидуальной мыши измеряли при помощи ELISA (В). Титры, индуцированные векторами CEA-DOM и CEA-FcIgG, значимо отличались от титров, индуцированных СЕА. Показаны средние величины (черные кружки).
На фигуре 23 показано обнаружение CD4+ Т-клеточной реакции на белок столбнячного токсоида. СЕА-трансгенных мышей иммунизировали pV1J/CTF-DOMopt, как описано в примере 16. Внутриклеточное окрашивание IFNγ на объединенных PBMC из индивидуальных мышей выполняли с пептидом р30. Показаны дискриминация целых лимфоцитов и дискриминация для CD8+ (R3) и CD4+ Т-клеток (R4).
На фигуре 24 показано противоопухолевое действие вакцинации векторами, несущими оптимизированные в отношении кодонов кДНК СЕА, СЕА-DOM или СЕА-FcIgG. Группы из 10 СЕА-трансгенгых мышей иммунизировали по схемам вакцинации ДНК/ДНК (А), Ad/Ad (В) и ДНК/Ad (С) с использованием плазмидных ДНК-векторов и Ad-векторов, несущих оптимизированные в отношении использования кодонов кДНК СЕА, СЕА-DOM или СЕА-FcIgG, как описано в примере 18. Через две недели после последней инъекции мышей иммунизировали sc-инокуляцией 5·105 опухолевых клеток МС38-СЕА. Процент не имеющих опухолей мышей в вакцинированных группах определяли с недельными интервалами и сравнивали с процентом необработанных контролей. Мыши, вакцинированные векторами СЕА-DOM (схема ДНК/Ad), значимо отличались от контрольных мышей (логарифмический ранговый критерий р<0,05).
На фигуре 25 показано действие элиминации CD4, CD8 или НК на индукцию противоопухолевого действия, индуцированного иммунизацией СЕА-DOM по схеме ДНК/Ad. СЕА-трансгенных мышей иммунизировали повторяющимися еженедельными инъекциями 50 мкг pV1J/CEA-DOMopt с последующей стимуляцией 1·109 вч Ad/СЕА-DOMopt (пример 19). Через неделю после последней инъекции мышей либо истощали, либо не истощали в отношении CD4+ Т-клеток, CD8+ Т-клеток или НК-клеток. Через две недели после последней инъекции мышей иммунизировали sc-инокуляцией 5·105 опухолевых клеток МС38-СЕА. Процент не имеющих опухолей мышей в вакцинированных группах определяли с недельными интервалами и сравнивали с процентом необработанных контролей. Эти результаты показывают, что процент не имеющих опухолей мышей в вакцинированной группе значимо отличался от необработанных контролей и истощенных групп.
На фигуре 26 показана нуклеотидная последовательность (SEQ ID NO:21) примерного, полностью оптимизированной слитой конструкции hCEA-DOM, называемой здесь hCEAoptDOMopt. Аминокислотная последовательность кодируемого белка также показана (SEQ ID NO:45). СЕА-часть нуклеотидной последовательности этой конкретной слитой конструкции СЕА состоит из нуклеотидов 1-2037, которые являются оптимизированными в отношении кодонов для экспрессии высокого уровня в клетке-хозяине человека. DOM-часть этой нуклеотидной последовательности показана жирным шрифтом и является также оптимизированной в отношении кодонов для экспрессии высокого уровня в клетках человека. Последовательности мест соединения, созданные стратегией клонирования, используемой для слитой конструкции последовательностей СЕА и LTB, подчеркнуты.
На фигуре 27 показана примерная нуклеотидная (SEQ ID NO:25) последовательность слитой конструкции hCEA-FcIgGopt, называемой здесь hCEAoptFcIgGopt. Показана также последовательность кодируемого белка (SEQ ID NO:46). СЕА-часть нуклеотидной последовательности этой конкретной слитой конструкции СЕА состоит из нуклеотидов 1-2037, которые являются оптимизированными в отношении кодонов для экспрессии высокого уровня в клетке-хозяине человека. FcIgG-часть этой нуклеотидной последовательности показана жирным шрифтом и является также оптимизированной в отношении кодонов для экспрессии высокого уровня в клетках человека. Последовательности мест соединения, созданные стратегией клонирования, используемой для слитой конструкции последовательностей СЕА и LTB, подчеркнуты.
На фигуре 28 показана нуклеотидная последовательность части кДНК СЕА дикого типа человека от нуклеотида 1 по нуклеотид 2037 (SEQ ID NO:22, панель А), кодирующая часть белка hCEA от аминокислоты 1 по аминокислоту 679 (SEQ ID NO:23, панель В).
На фигуре 29 показана неоптимизированная нуклеотидная последовательность минимизированного домена фрагмента С столбнячного токсина (DOM) кДНК от нуклеотида 1 по нуклеотид 825 (SEQ ID NO:47), кодирующая белок DOM, также показанный (SEQ ID NO:48).
На фигуре 30 показана неоптимизированная нуклеотидная последовательность примерной слитой конструкции СЕА-DOM человека (SEQ ID NO:49). СЕА-часть нуклеотидной последовательности этой конкретной слитой конструкции СЕА состоит из нуклеотидов 1-2037. DOM-часть этой нуклеотидной последовательности показана жирным шрифтом.
На фигуре 31 показана примерная нуклеотидная последовательность (SEQ ID NO:50) слитой конструкции CEA-DOM макака-резуса, называемой здесь rhCEA-DOMopt. Последовательность кодируемого белка также показана (SEQ ID NO:51). СЕА-часть нуклеотидной последовательности этой конкретной слитой конструкции СЕА состоит из нуклеотидов 1-2037, которые являются оптимизированными в отношении кодонов для экспрессии высокого уровня в клетке-хозяине человека. DOM-часть этой нуклеотидной последовательности, которая является также оптимизированной в отношении кодонов для экспрессии высокого уровня в клетках человека, показана жирным шрифтом.
На фигуре 32 показана нуклеотидная последовательность (SEQ ID NO:52) слитой конструкции CEA-СТВ макака-резуса, называемой здесь rhCEA-СВТopt. Последовательность кодируемого белка также показана (SEQ ID NO:53). СЕА-часть нуклеотидной последовательности этой конкретной слитой конструкции СЕА состоит из нуклеотидов 1-2037, которые являются оптимизированными в отношении кодонов для экспрессии высокого уровня в клетке-хозяине человека. СТВ-часть этой нуклеотидной последовательности, которая является также оптимизированной в отношении кодонов для экспрессии высокого уровня в клетках человека, показана жирным шрифтом.
Подробное описание изобретения
Карциноэмбриональный антиген (СЕА) обычно связан с развитием аденокарцином. Настоящее изобретение относится к композициям и способам индукции или усиления иммунитета к белковому продукту, экспрессируемому опухолеспецифическим антигеном СЕА, причем аберрантная экспрессия СЕА связана с карциномой или ее развитием. Ассоциация аберрантной экспрессии СЕА с карциномой не требует, чтобы белок СЕА экспрессировался в ткани опухоли во всех временных точках ее развития, так как аберрантная экспрессия СЕА может присутствовать при возникновении опухоли и не определяться позднее при прогрессировании опухоли, или наоборот.
Для этой цели настоящее изобретение относится к полинуклеотидам, векторам, клеткам-хозяевам и кодируемым белкам, содержащим последовательность СЕА или ее вариант, для использования в вакцинах и фармацевтических композициях для лечения и/или профилактики рака. Полинуклеотиды по изобретению содержат нуклеотидную последовательность, кодирующую белок СЕА или его вариант, слитую с нуклеотидной последовательностью, кодирующей по меньшей мере субъединицу иммуноусиливающего элемента, такого как бактериальный энтеротоксин или его существенная часть, которая может эффективно способствовать иммунному ответу на связанный СЕА.
Нуклеотидные последовательности СЕА по изобретению могут быть получены у человека или могут быть гомологом СЕА другого вида. Нуклеотидная последовательность СЕА дикого типа человека описана (см., например, патент США № 5274087; патент США № 5571710 и патент США № 6843761). Последовательность СЕА макака-резуса была недавно описана (WO 2004/072287). СЕА-часть слитой конструкции СЕА может быть полноразмерной или может быть любым вариантом, достаточным для индукции СЕА-специфического иммунного ответа у млекопитающего. Варианты СЕА по изобретению включают в себя, но ими не ограничиваются, последовательности, которые являются укороченными на С- или N-конце, последовательности с консервативными заменами и последовательности с внутренними делециями или инсерциями.
В предпочтительных вариантах осуществления настоящего изобретения СЕА-часть слитой конструкции СЕА является СЕА человека или его функциональным эквивалентом. В других предпочтительных вариантах осуществления, СЕА-часть является СЕА макака-резуса или его функциональным вариантом.
Таким образом, настоящее изобретение относится к синтетическому полинуклеотиду, содержащему последовательность нуклеотидов, кодирующую слитый белок СЕА, причем указанный слитый белок содержит белок СЕА или его биологически активный фрагмент или мутантную форму белка СЕА, слитые с иммуноусиливающим элементом или его субъединицей, который может эффективно усиливать иммунный ответ на белок СЕА. Указанные мутантные формы белка СЕА включают в себя, но ими не ограничиваются, консервативные замены аминокислот, укороченные на аминоконце формы, укороченные на карбоксиконце формы, делеции или добавления. Любой подобный биологически активный фрагмент и/или мутант будет кодировать либо белок, либо фрагмент белка, который по меньшей мере по существу повторяет иммунологические свойства белка СЕА SEQ ID NO:20. Синтетические полинуклеотиды по изобретению кодируют молекулы мРНК, которые экспрессируют функциональный слитый белок СЕА таким образом, что он может использоваться для разработки терапевтической или профилактической противораковой вакцины.
В предпочтительных вариантах осуществления изобретения СЕА-часть кодируемого слитого белка СЕА представляет собой СЕА человека (SEQ ID NO:20) или его функциональный эквивалент, например, СЕА человека с делецией его С-концевого якорного домена (AD) (SEQ ID NO:23), который расположен от приблизительно аминокислоты 679 до приблизительно аминокислоты 702 полноразмерного СЕА человека. Не связываясь с теорией, авторы настоящего изобретения полагают, что делеция якорного домена увеличивает секрецию слитого белка СЕА, усиливая посредством этого перекрестный прайминг иммунного ответа СЕА-LTB. В других предпочтительных вариантах осуществления СЕА-часть является СЕА макака-резуса (SEQ ID NO:18 и 19) или его функциональным эквивалентом.
Часть иммуноусиливающего элемента слитых белков СЕА изобретения способна стимулировать или усиливать иммунный ответ на связанный белок СЕА и выбрана из группы, состоящей из белка теплового шока (HSP) 70, связанного с лизосомой мембранного белка (LAMP), фрагмента С столбнячного токсоида (FrC), N-концевого домена FrC (DOM), тяжелого фрагмента константной цепи иммуноглобулина G1 (FcIgG), гликопротеина вируса везикулярного стоматита (VSV-G), холерного токсина (СТ) из Vibrio cholerae и термолабильного энтеротоксина E. coli (LT). В предпочтительных вариантах осуществления изобретения эта адъювантная часть слитой конструкции СЕА содержит N-концевой домен FrC (DOM), который, как было показано, эффективно усиливает иммуногенность одновременно доставляемых антигенов. В следующих предпочтительных вариантах осуществления, адъювантная часть слитой конструкции СЕА содержит субъединицу LT или ее существенную часть. В других предпочтительных вариантах осуществления адъювантной частью слитой конструкции СЕА является существенная часть FcIgG.
Была описана слитая конструкция СЕА, содержащая укороченный СЕА человека, слитая с единственным эпитопом столбнячного токсина (Q830-L844) (Lund et al., Cancer Gene Therapy 10: 365-376 (2003)). В отличие от этой слитой конструкции с единственным эпитопом, слитые конструкции СЕА по настоящему изобретению содержат существенную часть иммуноусиливающего элемента или его субъединицы, как описано выше, которая способна усиливать иммуногенность белка СЕА или его варианта. Существенная часть иммуноусиливающего элемента, которая используется для описанных здесь композиций и способов, не включает в себя части, которые составляют менее чем 50% полноразмерной субъединицы токсина. Используемая здесь стратегия, которая использует полноразмерные субъединицы адъюванта или их существенные части, была применена для обеспечения большей иммунной реакции на последовательность слитого СЕА. Не основываясь на какой-либо теории, авторы настоящего изобретения полагают, что, если бактериальный токсин, выбранный в качестве адъюванта, содержит более одного хелперного эпитопа, то ограничение последовательности токсина слитого белка единственным эпитопом приводило бы доказуемо к уменьшенному действию на иммуногенность рассматриваемого белка. Кроме того, авторы изобретения считают, что, если опосредованное адъювантом усиление иммунного ответа зависит от взаимодействия этого адъюванта со специфическими рецепторами клетки и не основано на универсальном эпитопе, то взаимодействие с рецептором могло бы зависеть от специфической структурной конфигурации, которая требовала бы существенной части иммуноусиливающего элемента для проявления действия адъюванта. В таком случае, короткая последовательность адъюванта, содержащая единственный эпитоп, была бы недостаточной для опосредования увеличения иммунного ответа.
В рамках настоящего изобретения рассматриваются также нуклеотидные последовательности, кодирующие варианты или мутанты описанных здесь иммуноусиливающих элементов, в том числе, но не только, замены, делеции, добавления нуклеотидов, аминоконцевые укорочения и карбоксиконцевые укорочения. В некоторых случаях может быть выгодным добавление специфических точковых мутаций к нуклеотидной последовательности, кодирующей адъювант или его субъединицу, для уменьшения или элиминации токсичности кодируемого белка. В примерах вариантов этого аспекта изобретения субъединица LT слита с СЕА-последовательностью слитой конструкции СЕА, причем субъединица LT укорочена удалением ее сигнальной последовательности. Не основываясь на какой-либо теории, авторы изобретения считают, что делеция сигнальной последовательности этого токсина, например, сигнальной последовательности LTB, обеспечивает запуск посттрансляционного процессинга этой слитой конструкции СЕА сигнальной последовательностью СЕА.
Иммуноусиливающий элемент, его субъединица или его существенная часть могут быть слиты с аминоконцом или карбоксиконцом последовательности СЕА. Дополнительно, последовательность иммуноусиливающего элемента и последовательность СЕА могут быть слиты по типу N-конец - N-конец, С-конец - С-конец, С-конец - N-конец или N-конец - С-конец. В предпочтительных вариантах осуществления изобретения С-конец полипептида СЕА слит с N-концом иммуноусиливающего элемента.
Настоящее изобретение относится к синтетической молекуле нуклеиновой кислоты (полинуклеотиду), содержащей последовательность нуклеотидов, которая кодирует мРНК, которая экспрессирует новый слитый белок СЕА; например, нуклеотидные последовательности, кодирующие слитые белки, представленные в SEQ ID NO:8, 10, 13, 15, 45, 51 и 53. Молекулы нуклеиновых кислот настоящего изобретения по существу не содержат других нуклеиновых кислот.
Настоящее изобретение относится также к рекомбинантным векторам и рекомбинантным клеткам-хозяевам как прокариотическим, так и эукариотическим, которые содержат молекулы нуклеиновых кислот, описанные в описании настоящего изобретения. Эти синтетические молекулы ДНК, связанные векторы и хозяева по изобретению могут использоваться для развития противораковой вакцины.
Примеры молекул нуклеиновых кислот настоящего изобретения содержат нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:7, 9, 11, 12, 14, 21, 25, 49, 50 и 52, показанных на фигурах 2-6, 26-27, 30-32, которые кодируют примеры слитых белков настоящего изобретения СЕА-LTА, СЕА-LTB, СЕА-DOM, CEA-FcIgG и СЕА-СТВ.
Настоящее изобретение включает в себя также биологически активные фрагменты или мутанты SEQ ID NO:7, 9, 11, 12, 14, 21, 25, 49, 50 и 52, которые кодируют мРНК, экспрессирующую примеры слитых белков СЕА. Любой такой биологически активный фрагмент и/или мутант будет кодировать либо белок, либо фрагмент белка, который по меньшей мере по существу имитирует фармакологические свойства белка hCEA, в том числе, но не только, белка hCEA, представленного в SEQ ID NO:20. Любой такой полинуклеотид включает в себя, но ими не ограничивается, замены, делеции, добавления нуклеотидов, аминоконцевые укорочения и карбоксиконцевые укорочения. Мутации изобретения кодируют молекулы мРНК, которые экспрессируют функциональный слитый белок СЕА в эукариотической клетке таким образом, чтобы он был применим в развитии противораковой вакцины.
В объем настоящего изобретения включены также мутации в ДНК-последовательности, которые по существу не изменяют конечные физические свойства экспрессируемого белка. Например, замена лейцина на валин, лизина на аргинин или глутамина на аспарагин может не вызывать изменения функциональности полипептида.
Как утверждалось выше, настоящее изобретение дополнительно относится к рекомбинантным векторам, которые содержат молекулы нуклеиновых кислот, описанные в этом описании изобретения. Эти векторы могут состоять из ДНК или РНК. В большинстве случаев, при клонировании предпочтительными являются векторы на основе ДНК. Характерные векторы включают в себя плазмиды, модифицированные вирусы, бакуловирус, бактериофаг, космиды, искусственные хромосомы дрожжей и другие формы эписомной или интегрированной ДНК, которые могут кодировать слитый белок СЕА. Специалист в данной области может определить подходящий вектор для переноса конкретного гена или другого использования.
Настоящее изобретение относится также к очищенным слитым белкам СЕА, кодируемым нуклеиновыми кислотами, описанными в настоящем описании. В примерных вариантах этого аспекта изобретения слитый белок СЕА содержит последовательность аминокислот, выбранную из группы, состоящей из SEQ ID NO:8, 10, 13, 15, 45, 46, 51 и 53.
Настоящее изобретение относится также к ДНК-последовательностям, которые гибридизуются с SEQ ID NO:7, 9, 11, 12, 14, 21, 25, 49, 50 или 52 в жестких условиях. В качестве примера, но им не ограничивается, процедура, использующая условия высокой жесткости, является следующей. Предгибридизацию фильтров, содержащих ДНК, проводят в течение приблизительно 2 часов в течение ночи при приблизительно 65°С в буфере, состоящем из 6×SSC, 5× раствора Денхардта и 100 мкг/мл денатурированной ДНК спермы лосося. Фильтры гибридизуют в течение приблизительно 12-48 часов при 65°С в смеси для предгибридизации, содержащей 100 мкг/мл денатурированной ДНК спермы лосося и 5-20×106 имп/мин 32Р-меченного зонда. Промывание фильтров выполняют при 37°С в течение приблизительно 1 часа в растворе, содержащем 2×SSC, 0,1% SDS. Затем следует промывание в 0,1×SSC, 0,1% SDS при 50°С в течение 45 минут перед радиоавтографией. Другие процедуры, использующие условия высокой жесткости, включают в себя либо стадию гибридизации, проводимую в 5×SSC, 5х растворе Денхардта, 50% формамиде при приблизительно 42°С в течение приблизительно 12-48 часов, либо стадию промывания проводят в 0,2×SSPE, 0,2% SDS при приблизительно 65°С в течение приблизительно 30-60 минут. Реагенты, упомянутые в вышеуказанных процедурах для проведения гибридизации высокой жесткости, хорошо известны в данной области. Подробный состав этих реагентов можно найти в Sambrook et al., Molecular Cloning: A Laboratory Manual 2nd Edition; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989) или Sambrook and Russel, Molecular Cloning: A Laboratory Manual 3rd Edition; Cold Spring Harbor Laboratory Press, Plainview, NY (2001). Кроме вышеуказанных условий, другие условия высокой жесткости, которые могут быть использованы, также хорошо известны в данной области.
Экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты, кодирующую слитый белок СЕА, может быть использован для экспрессии высокого уровня слитого белка СЕА в рекомбинантной клетке-хозяине. Экспрессирующие векторы могут включать в себя, но ими не ограничиваются, клонирующие векторы, модифицированные клонирующие векторы, специально сконструированные плазмиды или вирусы. Могут быть также использованы различные бактериальные экспрессирующие векторы для экспрессии рекомбинантных слитых последовательностей СЕА в бактериальных клетках, если это желательно. Кроме того, различные экспрессирующие векторы клеток грибов могут быть использованы для экспрессии слитых последовательностей СЕА в клетках грибов. Далее, различные экспрессирующие векторы клеток насекомых могут быть использованы для экспрессии рекомбинантного белка в клетках насекомых.
Изобретение относится также к клеткам-хозяевам, трансформированным или трансфицированным векторами, содержащими молекулы нуклеиновых кислот настоящего изобретения. Рекомбинантные клетки-хозяева могут быть прокариотическими или эукариотическими, включающими в себя, но ими не ограничивающимися, бактерии, такие как E. coli, клетки грибов, такие как дрожжи, клетки млекопитающих, в том числе, но не только, клеточные линии, полученные из быков, свиней, обезьян и грызунов; и клетки насекомых, в том числе, но не только, клеточные линии, полученные из Drosophila и тутового шелкопряда. Такие рекомбинантные клетки-хозяева могут культивироваться в подходящих условиях для получения слитого белка СЕА или биологически эквивалентной формы. В предпочтительном варианте осуществления клетка-хозяин является клеткой человека. Как определено здесь, термин «клетка-хозяин» не предназначен для включения в него клетки-хозяина в теле трансгенного человека, плода человека или эмбриона человека.
Как отмечалось выше, экспрессирующий вектор, содержащий ДНК, кодирующую слитый белок СЕА, может быть использован для экспрессии слитого белка СЕА в рекомбинантной клетке-хозяине. Таким образом, другим аспектом настоящего изобретения является способ экспрессии слитого белка СЕА в рекомбинантной клетке-хозяине, предусматривающий (а) введение вектора, содержащего нуклеиновую кислоту, содержащую последовательность нуклеотидов, которая кодирует слитый белок СЕА, в подходящую клетку-хозяина человека, где этот слитый белок содержит белок СЕА или его вариант, слитый с существенной частью иммуноусиливающего элемента или его субъединицы, где этот иммуноусиливающий элемент или его субъединица выбраны из группы, состоящей из белка теплового шока (HSP) 70, связанного с лизосомой мембранного белка (LAMP), фрагмента С столбнячного токсоида (FrC), N-концевого домена FrC (DOM), тяжелого фрагмента константной цепи иммуноглобулина G1 (FcIgG), гликопротеина вируса везикулярного стоматита (VSV-G), холерного токсина (СТ) из Vibrio cholerae и термолабильного энтеротоксина E. coli (LT); и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) культивирование этой клетки-хозяина при условиях, которые делают возможной экспрессию указанного слитого белка СЕА.
Предпочтительные иммуноусиливающие элементы для применения в настоящем аспекте изобретения выбраны из группы, состоящей из LTB, LTA, DOM и FcIgG.
В следующем предпочтительном варианте этого аспекта настоящего изобретения нуклеотидная последовательность СЕА-части этой слитой конструкции и/или части иммуноусиливающего элемента этой слитой конструкции являются оптимизированными в отношении кодонов для экспрессии высокого уровня в клетках человека.
Настоящее изобретение относится также к способу экспрессии слитого белка СЕА-LT в рекомбинантной клетке-хозяине, предусматривающему (а) введение вектора, содержащего нуклеиновую кислоту, содержащую последовательность нуклеотидов, которая кодирует слитый белок СЕА-LT, в подходящую клетку-хозяина человека, где этот слитый белок содержит белок СЕА или его вариант, слитый с существенной частью субъединицы LT, и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) культивирование этой клетки-хозяина при условиях, которые делают возможной экспрессию указанного слитого белка СЕА-LT.
В предпочтительном варианте способа экспрессии слитого белка СЕА-LT, описанного выше, субъединица LT является существенной частью LTB, причем в последовательности LTB делетирована ее сигнальная последовательность. В других вариантах осуществления субъединица LT является LTA или ее существенной частью.
Настоящее изобретение относится также к способу экспрессии слитого белка СЕА-DOM в рекомбинантной клетке-хозяине, предусматривающему (а) введение вектора, содержащего нуклеиновую кислоту, содержащую последовательность нуклеотидов, которая кодирует слитый белок СЕА-DOM, в подходящую клетку-хозяина человека, где этот слитый белок содержит белок СЕА или его вариант, слитый с существенной частью N-концевого домена фрагмента С столбнячного токсина (DOM) и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) культивирование этой клетки-хозяина при условиях, которые делают возможной экспрессию указанного слитого белка СЕА-DOM.
В предпочтительных вариантах способа экспрессии слитого белка СЕА-DOM, описанного выше, DOM-часть является оптимизированной в отношении кодонов для экспрессии высокого уровня в клетках человека. В других предпочтительных вариантах осуществления СЕА-часть слитой конструкции СЕА является оптимизированной в отношении кодонов для экспрессии высокого уровня в клетках человека. В других предпочтительных вариантах осуществления как СЕА-часть, так и DOM-часть являются оптимизированными в отношении кодонов для экспрессии высокого уровня в клетках человека.
После экспрессии слитой конструкции СЕА в клетке-хозяине слитый белок СЕА может быть выделен с получением слитого белка СЕА в активной форме. Несколько процедур очистки белков являются доступными и подходящими для использования. Рекомбинантный белок может быть очищен из клеточных лизатов и экстрактов различными комбинациями или индивидуальным применением высаливания, ионообменной хроматографии, гель-фильтрации, адсорбционной хроматографии на гидроксиапатите и гидрофобной хроматографии. Кроме того, слитый белок СЕА может быть отделен от других клеточных белков с использованием иммуноаффинной колонки, приготовленной с моноклональными или поликлональными антителами, специфическими в отношении белка СЕА или полипептидных фрагментов белка СЕА.
Были сконструированы молекулы нуклеиновых кислот, содержащие слитые конструкции СЕА, и кодируемые слитые белки по изобретению для усиления СЕА-специфического иммунного ответа, родственные полноразмерной кДНК, кодирующей СЕА, для использования в развитии вакцин. Для дополнительного усиления иммуногенных свойств слитых последовательностей СЕА по изобретению в некоторых описанных здесь вариантах осуществления полинуклеотиды, кодирующие слитые белки СЕА, содержат оптимизированные кодоны для последующей экспрессии высокого уровня в клетке-хозяине, как описано выше. В этих вариантах осуществления, по меньшей мере часть кодонов слитых конструкций СЕА сконструированы таким образом, чтобы использовать кодоны, предпочтительные для предполагаемой клетки-хозяина, которая в предпочтительных вариантах осуществления является клеткой человека. Эти оптимизированные слитые конструкции СЕА могут быть использованы для разработки рекомбинантных ДНК-вакцин на основе аденовируса или на основе плазмид, которые обеспечивают эффективную профилактику против СЕА-обусловленного рака посредством иммунитета, обеспечиваемого нейтрализующими антителами, и клеточно-опосредованного иммунитета. Эти синтетические молекулы могут быть использованы в качестве иммуногенной композиции. Настоящее изобретение относится к оптимизированным в отношении кодонов слитым полинуклеотидам СЕА, которые, при непосредственном введении позвоночному животному in vivo, в том числе млекопитающим, таким как приматы и человек, индуцируют экспрессию кодируемых белков у этого животного.
Как указано выше, в некоторых вариантах настоящего изобретения, эти синтетические молекулы содержат последовательность нуклеотидов, в которой некоторые из нуклеотидов были изменены таким образом, чтобы использовались кодоны, предпочтительные для клетки человека, что делает возможным экспрессию белка высокого уровня в клетке-хозяине человека. Эти синтетические молекулы могут быть использованы в качестве источника слитого белка СЕА, например, слитого белка СЕА-LTВ, который может быть использован в противораковой вакцине для обеспечения эффективной иммунопрофилактики против СЕА-обусловленных карцином посредством иммунитета, обеспечиваемого нейтрализующими антителами, и клеточно-опосредуемого иммунитета. Описанные здесь молекулы нуклеиновых кислот могут также служить основой для противораковой вакцины на основе ДНК.
«Триплетный» кодон из четырех возможных нуклеотидных оснований может существовать в более чем 60 вариантных формах. Поскольку эти кодоны обеспечивают транскрипт (информацию) только для 20 различных аминокислот (а также инициацию и терминацию транскрипции), некоторые аминокислоты могут кодироваться более чем одним кодоном, что известно как феномен вырожденности кодонов. По не вполне понятным причинам альтернативные кодоны неоднородно присутствуют в эндогенной ДНК различных типов клеток. Действительно, по-видимому, существует вариабельная природная иерархия или «предпочтение» в отношении определенных кодонов в некоторых типах клеток. Например, аминокислота лейцин кодируется любым из шести ДНК-кодонов, включающих в себя СТА, СТС, CTG, CTT, TTA и TTG. Исчерпывающий анализ частоты кодонов в геноме для микроорганизмов выявил, что эндогенная ДНК E. coli обычно содержит кодон CTG, определяющий лейцин, тогда как ДНК дрожжей и миксомицетов наиболее часто включает в себя кодон ТТА, определяющий лейцин. Ввиду этой иерархии, обычно считают, что вероятность получения высоких уровней экспрессии богатого лейцином полипептида хозяином E. coli будет зависеть до некоторой степени от частоты использования кодона. Например, существует вероятность, что ген, богатый кодонами ТТА, будет слабо экспрессироваться в E. coli, тогда как ген, богатый CTG, будет, возможно, высоко экспрессироваться в этом хозяине. Подобным образом, предпочтительным кодоном для экспрессии богатого лейцином полипептида в дрожжевых клетках-хозяевах будет ТТА.
Причастность феномена предпочтения кодонов проявляется в способах рекомбинантных ДНК, и этот феномен может служить объяснением многих прежних неудач в достижении высоких уровней экспрессии экзогенных генов в успешно трансформированных организмах-хозяевах: менее «предпочтительный» кодон может повторяемым образом присутствовать во встраиваемом гене, но аппарат экспрессии этой клетки-хозяина не может работать эффективно. Этот феномен предполагает, что синтетические гены, которые были сконструированы таким образом, что они включают в себя предпочтительные кодоны предполагаемой клетки-хозяина, обеспечивают оптимальную форму чужеродного генетического материала для использования на практике способов рекомбинантных ДНК. Таким образом, одним из аспектов настоящего изобретения является ген слитой конструкции СЕА, который оптимизирован в отношении кодонов для экспрессии в клетке человека. В предпочтительном варианте осуществления настоящего изобретения было обнаружено, что использование альтернативных кодонов, кодирующих одну и ту же последовательность белка, может устранять ограничения экспрессии экзогенного слитого белка СЕА в клетках человека.
В соответствии с некоторыми вариантами осуществления настоящего изобретения, молекулы нуклеиновых кислот, которые кодируют слитые белки СЕА, превращают в полинуклеотидную последовательность, имеющую идентичную транслируемую последовательность, но с использованием альтернативных кодонов, как описано в работе Lathe, “Synthetic Oligonucleotide Probes Deduced from Amino Acid Data: Theoretical and Practical Considerations” J. Molec. Biol. 183:1-12 (1985), приведенной здесь в качестве ссылки. Этот метод обычно предусматривает идентификацию кодонов в последовательности дикого типа, которые обычно не связаны с высоко экспрессируемыми генами человека, и замену их оптимальными кодонами для экспрессии высокого уровня в клетках человека. Затем эту новую последовательность гена исследуют на нежелательные последовательности, генерируемые этими заменами кодонов (например, последовательности «ATTTA», непреднамеренное создание сайтов узнавания сплайсинга интронов, нежелательных сайтов рестриктаз и т.д.). Нежелательные последовательности элиминируют заменой существующих кодонов различными кодонами, кодирующими ту же самую аминокислоту. Затем эти синтетические сегменты гена испытывают на улучшенную экспрессию.
Описанные здесь выше способы используют для создания синтетических последовательностей генов, которые кодируют слитые белки СЕА, с получением гена, содержащего кодоны, оптимизированные для экспрессии высокого уровня. Хотя описанная выше процедура обеспечивает суммирование методологии авторов для конструирования оптимизированных в отношении кодонов генов для использования в противораковых вакцинах, специалисту в данной области понятно, что сходная эффективность вакцин или увеличенная экспрессия генов может быть также достигнута минорными вариациями в этой процедуре или минорными вариациями в данной последовательности.
Специалисту в данной области будет также понятно, что могут быть сконструированы дополнительные молекулы нуклеиновых кислот, которые могут обеспечивать высокие уровни экспрессии слитой конструкции СЕА в клетках человека, причем только часть кодонов в этих ДНК-молекулах являются оптимизированными в отношении кодонов. Например, в некоторых вариантах осуществления настоящего изобретения кодоны, составляющие СЕА-часть слитой конструкции СЕА, являются оптимизированными для экспрессии высокого уровня в клетках человека, а кодоны, составляющие адъювантную часть этой слитой конструкции СЕА, являются по существу сходными с нуклеотидной последовательностью адъюванта дикого типа. В других вариантах осуществления настоящего изобретения кодоны, составляющие адъювантную часть слитой конструкции СЕА, являются оптимизированными для экспрессии высокого уровня в клетках человека, а кодоны, составляющие СЕА-часть этой слитой конструкции СЕА, являются по существу сходными с геном СЕА дикого типа. В других вариантах осуществления настоящего изобретения как СЕА-часть, так и часть адъюванта слитой конструкции СЕА являются оптимизированными в отношении кодонов для экспрессии высокого уровня в клетках человека. Слитые конструкции СЕА, в которых только субпопуляция кодонов является оптимизированной в СЕА-части и/или в части адъюванта слитой конструкции СЕА, также рассматриваются изобретением.
Нуклеиновые кислоты по изобретению могут быть собраны в экспрессирующую кассету, которая содержит последовательности, сконструированные для обеспечения эффективной экспрессии белка в клетке человека. Эта кассета, предпочтительно содержит ген, кодирующий слитый белок СЕА, с соответствующими регуляторными последовательностями транскрипции и трансляции, функционально связанными с ним, такими как промотор и последовательности терминации. В предпочтительном варианте осуществления этим промотором может быть промотор цитомегаловируса без последовательности интрона А (CMV), хотя специалистам в данной области будет понятно, что любой из ряда других известных промоторов, таких как сильный промотор иммуноглобулина или промоторы других эукариотических генов, могут быть использованы. Предпочтительным терминатором транскрипции является терминатор бычьего гормона роста, хотя могут быть также использованы другие известные терминаторы транскрипции. Особенно предпочтительной является комбинация CMV-BGH-терминатора.
В соответствии с изобретением, экспрессионная кассета слитой конструкции СЕА встроена в вектор. Этим вектором предпочтительно является аденовирусный или плазмидный вектор, хотя могут быть также использованы линейная ДНК, связанная с промотором, или другие векторы, такие как аденосвязанный вирус или модифицированный вирус коровьей оспы, ретровирусный или лентивирусный вектор.
Если выбранным вектором является аденовирус, предпочтительно, чтобы им был так называемый аденовирусный вектор первой генерации. Эти аденовирусные векторы отличаются наличием нефункционального района гена Е1 и предпочтительно делетированным районом гена Е1 аденовируса. В некоторых вариантах осуществления экспрессионную кассету встраивают в положении, в котором обычно расположен аденовирусный ген Е1. Кроме того, эти векторы, необязательно, имеют нефункциональный или делетированный район Е3. Предпочтительно, чтобы используемый геном аденовируса имел делетированные районы Е1 и Е3 (ΔЕ1ΔЕ3). Эти аденовирусы могут быть размножены в известных клеточных линиях, которые экспрессируют вирусный ген Е1, таких как клетки 293 или клетки PERC.6, или в клеточных линиях, произведенных из клетки 293 или PERC.6, которая транзиторно или стабильно трансформирована для экспрессии дополнительного количества белка. Например, при использовании конструкций, которые имеют регулируемую экспрессию генов, например, регулируемую промотором тетрациклина систему, эта клеточная линия может экспрессировать компоненты, участвующие в этой регуляторной системе. Одним примером такой клеточной линии является T-Rex-293; в данной области известны и другие примеры.
Для удобства манипулирования аденовирусным вектором этот аденовирус может находиться в форме челночной плазмиды. Настоящее изобретение относится также к челночному плазмидному вектору, который содержит плазмидную часть и аденовирусную часть, причем аденовирусная часть содержит аденовирусный геном, имеющий делецию Е1 и, необязательно, делецию Е3, и имеет встроенную экспрессионную кассету, содержащую кодирующую слитый белок СЕА нуклеотидную последовательность. В предпочтительных вариантах осуществления имеется сайт рестрикции, фланкирующий аденовирусную часть этой плазмиды, так что аденовирусный вектор может быть легко удален. Эта челночная плазмида может реплицироваться в прокариотических клетках или эукариотических клетках.
В предпочтительном варианте осуществления изобретения эта экспрессионная кассета встроена в аденовирусную плазмиду pMRKAd5-HVO (см. Emini et al., WO 02/22080, который включен здесь в качестве ссылки). Эта плазмида содержит геном аденовируса Ad5 с делецией районов Е1 и Е3. Конструирование плазмиды pMRKAd5-HV0 было улучшено относительно прежних аденовекторов удлинением 5'-цис-действующего упаковочного района дополнительно в ген Е1 для включения элементов, которые, как было обнаружено, являются важными в оптимизации упаковки вируса, приводящей к усиленной амплификации вируса. Предпочтительно, этот усиленный аденовирусный вектор способен сохранять генетическую стабильность после размножения с высоким числом пассажей.
Стандартные способы молекулярной биологии для получения и очистки ДНК-конструкций позволяют получать аденовирусы, челночные плазмиды и ДНК-иммуногены настоящего изобретения.
В соответствии с настоящим изобретением было определено, что молекулы, кодирующие слитый белок СЕА-LT, описанные здесь (например, SEQ ID NO:12), которые содержат существенную часть субъединиц LTA или LTB термолабильного энтеротоксина E. coli, экспрессируются с эквивалентной эффективностью в сравнении с соответствующей последовательностью СЕА дикого типа (см. пример 4). Было показано также, что плазмиды pV1J/hCEA-LTA и pV1J/hCEA-LTВ индуцируют более высокий ответ в виде антител, чем pV1J/hCEA, подтверждая адъювантное действие, проявляемое субъединицами LT на СЕА-специфический иммунный ответ (см. пример 11). Таким образом, описанные здесь результаты демонстрируют, что слитая конструкция кодирующей СЕА последовательности с кДНК LTA или LTВ приводит к увеличению СЕА-специфического иммунного ответа. По-видимому, LTВ проявляет большее усиливающее действие на этот иммунный ответ с преимущественной индукцией CD8+ Т-клеток, в то время как LTA индуцирует преимущественную CD4+ реакцию.
В соответствиии с настоящим изобретением было также показано, что толерантность к аутоантигену СЕА может быть разрушена более эффективно относительно полноразмерной кДНК СЕА дикого типа вследствие увеличенных иммуногенных свойств слитой конструкции СЕА-LTВ. Усиливающий эффект LTB на иммуногенные свойства СЕА также наблюдался при инъекции плазмиды, несущей полностью оптимизированные кодоны кДНК слитой конструкции СЕА-LTВ. Наконец, описанные здесь результаты с использованием аденовирусных векторов, несущих слитые конструкции СЕА-LT, свидетельствуют о том, что усиленная иммуногенность слитых конструкций СЕА-LT не ограничивается иммунизацией плазмидной ДНК (см. пример 13).
Дополнительно, в соответствии с настоящим изобретением было показано, что плазмиды pV1J/hCEA-DOM и pV1J/hCEA-FcIgG индуцируют большую СЕА-специфическую клеточно-опосредованную и гуморальную реакцию, чем СЕА (см. пример 15). В соответствии с настоящим изобретением было также показано, что толерантность к аутоантигену СЕА может быть разрушена более эффективно описанными здесь слитыми конструкциями СЕА с DOM и FcIgG относительно полноразмерной кДНК СЕА дикого типа вследствие увеличенных иммуногенных свойств этих слитых конструкций СЕА. Усиленные иммуногенные свойства этих слитых белков наблюдали после иммунизации ДНК- или Ad-векторами, что свидетельствует о том, что усиленная иммуногенность слитых конструкций СЕА-LT не ограничивается иммунизацией плазмидной ДНК (см. пример 16).
Таким образом, описанные выше векторы могут быть использованы в иммуногенных композициях и вакцинах для профилактики развития аденокарцином, связанных с аберрантной экспрессией СЕА, и/или для лечения существующих раков. Векторы по изобретению делают возможными развитие и организацию производства вакцин устранением трудностей с получением высоких уровней экспрессии экзогенного СЕА в успешно трансформированных организмах-хозяевах и обеспечением слитого белка СЕА, который может индуцировать усиленный иммунный ответ при введении млекопитающему, такому как человек.
Для этой цели одним из аспектов настоящего изобретения является способ профилактики или лечения СЕА-обусловленного рака, предусматривающий введение млекопитающему вакцинного вектора, содержащего полинуклеотид, содержащий последовательность нуклеотидов, которая кодирует слитый белок СЕА, причем этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с существенной частью иммуноусиливающего элемента, выбранного из группы, состоящей из белка теплового шока (HSP) 70, связанного с лизосомой мембранного белка (LAMP), фрагмента С столбнячного токсоида (FrC), N-концевого домена FrC (DOM), тяжелого фрагмента константной цепи иммуноглобулина G1 (FcIgG), гликопротеина вируса везикулярного стоматита (VSV-G), холерного токсина (СТ) из Vibrio cholerae и термолабильного энтеротоксина E. coli (LT); причем этот слитый белок способен вызывать иммунный ответ у млекопитающего.
В предпочтительных вариантах описанных здесь способов иммуноусиливающий элемент выбран из группы, состоящей из LTA, LTВ, DOM и FcIgG.
В соответствии с описанным выше способом вакцинный вектор может вводиться для лечения или профилактики рака у любого млекопитающего, в том числе, но не только, рака легкого, рака молочной железы и колоректального рака. В предпочтительном варианте осуществления настоящего изобретения этим млекопитающим является человек.
Дополнительно, специалист в данной области может выбрать любой тип вектора для использования в описанном способе лечения и профилактики. Предпочтительно, этим вектором является аденовирусный вектор или плазмидный вектор. В предпочтительном варианте осуществления настоящего изобретения этим вектором является аденовирусный вектор, содержащий аденовирусный геном с делецией в районе Е1 аденовируса и инсерцией в этом районе Е1 аденовируса, причем инсерция содержит экспрессионную кассету, содержащую (а) последовательность нуклеотидов, которая кодирует слитый белок СЕА, где этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с иммуноусиливающим элементом или его существенной частью; где этот иммуноусиливающий элемент выбран из группы, состоящей из белка теплового шока (HSP) 70, связанного с лизосомой мембранного белка (LAMP), фрагмента С столбнячного токсоида (FrC), N-концевого домена FrC (DOM), тяжелого фрагмента константной цепи иммуноглобулина G1 (FcIgG), гликопротеина вируса везикулярного стоматита (VSV-G), холерного токсина (СТ) из Vibrio cholerae и термолабильного энтеротоксина E. coli (LT); и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) промотор, функционально связанный с этим полинуклеотидом.
Дополнительно настоящее изобретение относится к аденовирусному вакцинному вектору, содержащему аденовирусный геном с делецией в районе Е1 аденовируса и инсерцией в этом районе Е1 аденовируса, причем инсерция содержит экспрессионную кассету, содержащую (а) последовательность нуклеотидов, которая кодирует слитый белок СЕА, где этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с существенной частью иммуноусиливающего элемента; где этот иммуноусиливающий элемент выбран из группы, состоящей из HSP70, LAMP, FrC, DOM, FcIgG, VSV-G, CT, LTA и LTB; и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) промотор, функционально связанный с этим полинуклеотидом.
В предпочтительном варианте этого аспекта изобретения аденовирусным вектором является вектор Ad5.
В другом предпочтительном варианте осуществления изобретения аденовирусным вектором является вектор Ad6.
Еще в одном предпочтительном варианте осуществления изобретения аденовирусным вектором является вектор Ad24.
Для использования в настоящем изобретении рассматривается также аденовирусный вакцинный вектор, содержащий аденовирусный геном, который природно инфицирует виды, другие, чем человек, в том числе, но не только, аденовирусные векторы шимпанзе. Предпочтительным вариантом осуществления настоящего изобретения является вакцинный вектор chimp Ad3.
В другом аспекте, настоящее изобретение относится к вакцинной плазмиде, содержащей плазмидную часть и часть экспрессионной кассеты, где эта часть экспрессионной кассеты содержит (а) последовательность нуклеотидов, которая кодирует слитый белок СЕА, где этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с иммуноусиливающим элементом или его существенной частью, выбранными из группы, состоящей из HSP70, LAMP, FrC, DOM, FcIgG, VSV-G, CT, LTA и LTB; и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) промотор, функционально связанный с этим полинуклеотидом.
В некоторых вариантах осуществления настоящего изобретения рекомбинантные аденовирусные или полинуклеотидные вакцины на основе плазмид, описанные здесь, используют в различных комбинациях прайминга/стимуляции для индукции усиленного иммунного ответа. В этом случае эти два вектора вводят в виде схемы «прайминг и стимуляция». Например, первый тип вектора вводят один раз или несколько раз, затем после заданного периода времени, например, 2 недель, 1 месяца, шести месяцев или другого подходящего интервала вводят второй тип вектора один раз или несколько раз. Предпочтительно, эти векторы несут экспрессионные кассеты, кодирующие один и тот же полинуклеотид или комбинацию полинуклеотидов. В варианте осуществления, в котором используют также плазмидную ДНК, предпочтительно, чтобы этот вектор содержал один или несколько промоторов, узнаваемых клетками млекопитающих или насекомых. В предпочтительном варианте осуществления эта плазмида содержит сильный промотор, такой как, но не только, промотор CMV. К такому промотору могут быть присоединены синтетический слитый ген СЕА или другой экспрессирующийся ген. Примером такой плазмиды может быть экспрессионная плазмида V1Jns млекопитающих, описанная в работе J. Shiver et al., in DNA Vaccines, M. Liu et al., eds., N.Y. Acad. Sci., N.Y., 772: 198-208 (1996), которая включена здесь в качестве ссылки.
Как утверждалось выше, вакцина в виде аденовирусного вектора и плазмидная вакцина могут вводиться позвоночному животному в виде части единой терапевтической схемы введения для индукции иммунного ответа. Для этой цели настоящее изобретение относится к способу защиты млекопитающего от СЕА-обусловленного рака, предусматривающему (а) введение млекопитающему первого вектора, содержащего i) последовательность нуклеотидов, которая кодирует слитый белок СЕА, где слитый белок СЕА содержит белок СЕА или его вариант, слитый с существенной частью иммуноусиливающего элемента, выбранного из группы, состоящей из HSP70, LAMP, FrC, DOM, FcIgG, VSV-G, CT, LTA и LTB; и где слитый белок способен вызывать иммунный ответ у млекопитающего; и ii) промотор, функционально связанный с этим полинуклеотидом; (b) выдерживание в течение заданного периода времени; и (с) введение млекопитающему второго вектора, содержащего i) последовательность нуклеотидов, которая кодирует слитый белок СЕА, где этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с существенной частью иммуноусиливающего элемента, выбранного из группы, состоящей из HSP70, LAMP, FrC, DOM, FcIgG, VSV-G, CT, LTA и LTB; и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и ii) промотор, функционально связанный с этим полинуклеотидом.
В одном из вариантов способа защиты, описанного выше, первый вектор является плазмидой, а второй вектор является аденовирусным вектором. В альтернативном варианте осуществления первый вектор является аденовирусным вектором, а второй вектор является плазмидой.
В описанном выше способе первый тип вектора может быть введен более одного раза, причем каждое введение этого вектора отделено от предыдущего заданным периодом времени. За таким рядом введений первого типа вектора может следовать введение второго типа вектора один раз или несколько раз после прохождения заданного периода времени. Подобно обработке первым типом вектора, второй тип вектора может также предоставляться один раз или более чем один раз с заданными интервалами времени.
Кроме того, настоящее изобретение относится к способу лечения млекопитающего, страдающего СЕА-обусловленной аденокарциномой, предусматривающему (а) введение млекопитающему первого вектора, содержащего i) последовательность нуклеотидов, которая кодирует слитый белок СЕА, где этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с существенной частью иммуноусиливающего элемента, выбранного из группы, состоящей из HSP70, LAMP, FrC, DOM, FcIgG, VSV-G, CT и LTB; и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и ii) промотор, функционально связанный с этим полинуклеотидом; (b) выдерживание в течение заданного периода времени; и (с) введение млекопитающему второго вектора, содержащего i) последовательность нуклеотидов, которая кодирует слитый белок СЕА, где этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с существенной частью иммуноусиливающего элемента, выбранного из группы, состоящей из HSP70, LAMP, FrC, DOM, FcIgG, VSV-G, CT и LT; и где этот слитый белок способен вызывать иммунный ответ у млекопитающего; и ii) промотор, функционально связанный с этим полинуклеотидом.
В одном из вариантов описанного выше способа лечения первый вектор является плазмидой, а второй вектор является аденовирусным вектором. В альтернативном варианте осуществления первый вектор является аденовирусным вектором, а второй вектор является плазмидой.
В предпочтительных вариантах описанных выше способов эти векторы содержат нуклеотиды, которые кодируют слитый белок СЕА-LT, где этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с существенной частью субъединицы LT. В дополнительных предпочтительных вариантах этот вектор содержит последовательность нуклеотидов, которая кодирует слитый белок СЕА-LTB. В дополнительных предпочтительных вариантах описанных здесь способов векторы содержат последовательность нуклеотидов, которая кодирует слитый белок СЕА-DOM, где этот слитый белок СЕА содержит белок СЕА или его вариант, слитый с существенной частью субъединицы DOM. В дополнительных предпочтительных вариантах этот вектор содержит последовательность нуклеотидов, которая кодирует слитый белок СЕА-FcIgG.
Количество экспрессируемой ДНК или транскрибируемой РНК, которое должно вводиться в вакцине реципиенту, будет зависеть от силы используемых промоторов и от иммуногенности экспрессируемого продукта гена. Обычно, иммунологически или профилактически эффективную дозу приблизительно 1 нг - 100 мг и предпочтительно приблизительно 10-300 мкг плазмидного вакцинного вектора вводят непосредственно в мышечную ткань. Эффективная доза для рекомбинантного аденовируса равна приблизительно 106-1012 частиц и предпочтительно приблизительно 107-1011 частиц. Рассматриваются также подкожная инъекция, внутрикожное введение, вдавливание через кожу и другие способы введения, такие как внутрибрюшинная, внутривенная, внутримышечная или ингаляционная доставка.
В предпочтительных вариантах осуществления настоящего изобретения вакцинные векторы вводят реципиенту посредством внутримышечной инъекции.
Вакцинные векторы по настоящему изобретению могут быть «голыми», т.е. не связанными с какими-либо белками или другими агентами, которые влияют на иммунную систему реципиента. В этом случае, желательно, чтобы эти вакцинные векторы находились в физиологически приемлемом растворе, таком как, но не только, стерильный физиологический раствор или стерильный забуференный физиологический раствор. Альтернативно, может быть предпочтительным введение агента, который способствует клеточному поглощению ДНК, такого как, но не только, ион кальция. Эти агенты обычно называют усиливающими трансфекцию реагентами и фармацевтически приемлемыми носителями. Специалисты в данной области смогут определить конкретный реагент или фармацевтически приемлемый носитель, а также подходящее время и схему введения.
Все упоминаемые публикации приведены здесь в качестве ссылки для описания и раскрытия способов и материалов, которые могут быть использованы вместе с настоящим изобретением. Ничто не должно рассматриваться как признание того, что настоящее изобретение может быть противопоставлено источнику информации с более ранним приоритетом.
После описания предпочтительных вариантов осуществления настоящего изобретения со ссылкой на сопутствующие фигуры должно быть понятно, что это изобретение не ограничивается приведенными определенными вариантами осуществления и что в них могут быть произведены различные изменения и модификации специалистом в данной области без отклонения от объема или идеи настоящего изобретения, определенных в прилагаемой формуле изобретения.
Приведенные ниже примеры иллюстрируют, но не ограничивают настоящее изобретение.
ПРИМЕР 1
Конструирование слитых белков СЕА
Для определения иммуногенности слитых белков СЕА конструировали ряд векторов, кодирующих аминокислоты (далее называемые аа) 1-679 белка СЕА человека, слитые с панелью выбранных полипептидов (см. пример 2). Эти последовательности были выбраны в связи с их сообщенными иммуноусиливающими свойствами, которые были продемонстрированы в различных экспериментальных системах. Слитые конструкции СЕА конструировали соединением кДНК белка СЕА с делецией якорной последовательности GPI с чужеродными полипептидами (примеры конструкций изображены на фигуре 18А). Опухолевый антиген связывали с последовательностями HSP70, FcIgG или LAMP для определения, будет ли усиление поглощения антигена или повторное нацеливание на эндосомный компартмент приводить к увеличенному иммунному ответу. Подобным образом, слитая конструкция с фрагментом С столбнячного токсина (FrC) или с минимальным доменом, лишенным потенциально конкурентных связывающих эпитопов МНС класса I (DOM, см. фигуру 29) (Rice et al., J. Immunol. 168: 3908-13 (2002)), конструировали для усиления гуморальной и CD4+ Т-клеточной реакций. СЕА связывали также с кодирующей VSV-G последовательностью для определения, влияет ли слитая конструкция с вирусным гликопротеином на иммуногенные свойства СЕА.
Кодирующие последовательности этих слитых конструкций СЕА клонировали в векторы pV1Jns под контролем промотора CMV/интрона А человека плюс сигнал полиаденилирования бычьего гормона роста (BGH) (пример 2). Плазмиды pV1J/hCEA-FrC, pV1J/hCEA-DOM, pV1J/hCEA-FcIgG, pV1J/hCEA-LAMP, pV1J/hCEA-VSV-G и pV1J/hCEA-HSP70 несут кДНК СЕА дикого типа, слитую с кодирующими последовательностями указанных чужеродных полипептидов. Примеры нуклеотидных и аминокислотных последовательностей слитых конструкций hCEA-DOM и hCEA-FcIgG показаны на фигурах 26, 27 и 30.
Для оценки действия субъединиц LTA и LTВ термолабильного энтеротоксина E. coli на иммуногенность СЕА генерировали ряд дополнительных слитых конструкций, кодирующих аминокислоты 1-679 белка СЕА, слитых с кодирующими либо LTA (аминокислоты 18-259), либо LTВ (аминокислоты 21-125) последовательностью. Схематическое представление структуры примеров слитых конструкций СЕА-LTA и СЕА-LTВ, полученных для этого исследования, показаны на фигуре 1. Примеры нуклеотидных и аминокислотных последовательностей слитых конструкций СЕА-LT показаны на фигурах 2-6.
Слитые конструкции СЕА-LT конструировали соединением кДНК белка СЕА с делецией якорной последовательности с субъединицами LT, у которых была удалена кодирующая последовательность сигнального пептида. Кодирующие последовательности этих слитых конструкций СЕА клонировали в векторы pV1Jns под контролем промотора цитомегаловируса (CMV)/интрона А человека плюс сигнал полиаденилирования бычьего гормона роста (BGH). Плазмиды pV1J/hCEA-LTA и pV1J/hCEA-LTВ несут кДНК СЕА дикого типа, слитую с кодирующими последовательностями LTА и LTВ, соответственно (см. пример 2).
Все конструкции, несущие слитую конструкцию СЕА-LTВ, генерировали слитой конструкцией кДНК СЕА (нуклеотиды 1-2037) с кДНК-фрагментом LTВ, включающим в себя нуклеотиды 64-375. Кодирующую LTВ последовательность получали ПЦР-амплификацией геномной ДНК E. coli с использованием последовательность-специфических праймеров LTB-S1 5'- T A T T C T A G A T G C T C C C C A G A C T A T T A C A G A A -3' (SEQ ID NO:1) и LTB-А1 5'- T A T G C G G C C G C C T A G T T T T C C A T A C T G A T T G C C G C-3' (SEQ ID NO:2). Амплифицированную ДНК вводили на 3'-конце кодирующей последовательности СЕА с получением плазмид.
ПРИМЕР 2
Плазмидные конструкции
pV1J/CEA opt и pV1J/CEA: Эти две конструкции несут оптимизированную в отношении использования кодонов кДНК и кДНК СЕА дикого типа, соответственно. Кодирующая последовательность СЕА расположена между предранним промотором CMV/интроном А цитомегаловируса и сигналом полиаденилирования бычьего гормона роста. Для генерирования pV1J/hCEAopt плазмиду pCR-hCEAopt расщепляли EcoRI в течение 1 часа при 37єС. Полученную инсерцию 2156 п.н. очищали и клонировали в сайт EcoRI плазмиды pV1JnsB ((Montgomery et al., DNA Cell Biol. 12(9): 777-83 (1993)).
Для генерирования pV1J/hCEA, плазмиду pCR-hCEA (Song et al., Regulation of T-helper-1 versus T-helper-2 activity and enhancement of tumour immunity by combined DNA-based vaccination and nonviral cytokine gene transfer. Gene Therapy 7: 481-492 (2000)) расщепляли EcoRI. Полученную инсерцию 2109 п.н. клонировали в сайт EcoRI плазмиды pV1JnsA (Montgomery et al., supra).
pV1J/hCEA-LTВ и pV1J/hCEA opt -LTВ: Оптимизированную в отношении кодонов кДНК LTВ синтезировали сборкой олигонуклеотидов (Geneart GmbH, Regensburg, Germany) и клонировали в вектор pCR-script (Stratagene, La Jolla, CA). Для генерирования pV1J/hCEAopt-LTВopt, LTВopt амплифицировали при помощи ПЦР с использованием следующих ПЦР-праймеров: LTВopt-5'XbaI (5'-конец) 5'- G C T C T A G A G C C C C C C A G A G C A T C A C C G A G C T G T G C - 3' (SEQ ID NO:3) и LTВopt-3'BglII (3'-конец) 5'- G C T C T A G A A C C C C T C A G A A C A T C A C C G A T C T G T G C G C C - 3' (SEQ ID NO:4). Затем этот амплифицированный продукт встраивали в сайты XbaI/BglII плазмиды pV1J/hCEAopt.
pV1J/hCEA-LTA: Кодирующую LTA последовательность, соответствующую нуклеотидам 54-774, которые кодируют аминокислоты 18-259, амплифицировали при помощи ПЦР из геномной ДНК E. coli с использованием последовательность-специфических праймеров LTА-S1 5'- T A T T C T A G A T A A T G G C G A C A A A T T A T A C C G - 3' (SEQ ID NO:5) и LTА-А1 5'- T A T G C G G C C G C T C A T A A T T C A T C C C G A A T T C T G T T - 3' (SEQ ID NO:6). Амплифицированную ДНК расщепляли подходящими рестриктазами и встраивали в плазмиду pV1J/hCEA.
pV1J/rhCEAopt-LTВ: 3'-фрагмент кДНК СЕА макака-резуса (нуклеотиды 1641-2026), который был оптимизирован в отношении кодонов для экспрессии высокого уровня в клетках человека, амплифицировали при помощи ПЦР из pV1J-rhCEAopt. Амплифицированная кДНК была лишена кодирующей GPI-якорь последовательности и несла сайты рестрикции XbaI/BglII. Этот фрагмент встраивали в сайт PstI pCR-blunt-rhCEAopt с получением таким образом промежуточной плазмиды pCR-blunt-rhCEAopt XbaI/BglII. rhCEAopt экстрагировали в виде BglII/SalI-фрагмента и клонировали в те же самые сайты в pV1J-nsB с получением таким образом pV1J-rhCEAopt XbaI/BglII. LTBopt амплифицировали при помощи ПЦР из pCR-script-LTBopt с добавлением сайтов XbaI и BglII на 5'- и 3'-концах, соответственно, и клонировали в pV1J-rhCEAopt XbaI/BglII с получением таким образом pV1J-rhCEA-LTВopt.
pV1J/CEA-FrC, pV1J/CEA-DOM, pV1J/CEA-FcIgG, pV1J/CEA-LAMP, pV1J/CEA-HSP70 и pV1J/CEA-VSV-G: Все эти конструкции, кодирующие указанные слитые белки СЕА, генерировали слитой конструкцией кДНК СЕА с нуклеотидами 1-2037 (SEQ ID NO:22, фигура 28А), соответствующими аминокислотам 1-679 (SEQ ID NO:23, фигура 28В), с кДНК-фрагментом, соответствующим следующим фрагментам: фрагменту С столбнячного токсоида (СЕА-FrC, SEQ ID NO:24), N-концевому домену FrC (СЕА-DOM, SEQ ID NO:21 и 49), тяжелому фрагменту константной цепи иммуноглобулина G1 (СЕА-FcIgG, SEQ ID NO:25), связанному с лизосомой мембранному белку (СЕА-LAMP, SEQ ID NO:26), белку теплового шока 70 (СЕА-HSP70, SEQ ID NO:27) или гликопротеину вируса везикулярного стоматита (СЕА-VSV-G, SEQ ID NO:28).
Кодирующие последовательности FrC и DOM получали при помощи ПЦР-амплификации из плазмиды pRep-TeT.C, как описано в Rice et al. (J. Immunol. 169: 3908-13 (2002)). FcIgG получали из тотальной РНК PBMC человека. VSV-G и HSP70 получали из p-FAST-VSV-G и из плазмиды pY3111, соответственно. LAMP1 получали сборкой генов. Амплификации проводили с использованием следующих праймеров: FrC смысловой (5'-T A T T C T A G A T T C A A C A C C A A T T C C A T T T T C T T A T T C -3' (SEQ ID NO:29), FrC антисмысловой (5'-G C G G C C G C T A G A A T C A T T T G T C C A T C C T T C A T C -3' (SEQ ID NO:30), DOM смысловой (5'-T A T T C T A G A T T C A A C A C C A A T T C C A T T T T C T T A T T C -3' (SEQ ID NO:31), DOM антисмысловой (5'- T T A G C G G C C G C T A G T T C T G T A T C A T A T C G T A A A G G G -3' (SEQ ID NO:32), FcIgG смысловой (5'- T C T A G A T A A A A C T C A C A C A T G C C C A -3' (SEQ ID NO:33), FcIgG антисмысловой (5'- G C C G A C T C A T T T A C C C G G A G A C A G G G A G -3' (SEQ ID NO:34), LAMP смысловой (5'- T C T A G A T T T G A T C C C C A T T G C T G T G G G C G G T G C C C T G -3'(SEQ ID NO:35), LAMP антисмысловой (5'- G G C G T G A C T C C T C T T C C T G C C A A T G A G G T A G G C A A T G A G -3' (SEQ ID NO:36), VSV-G смысловой (5'-A T A T C T A G A T T T C A C C A T A G T T T T T C C A C A C A A C C -3' (SEQ ID NO:37), VSV-G антисмысловой (5'-G C G G C C G C C T T C C T T C C A A G T C G G T T C A T C T C T A T G -3' (SEQ ID NO:38), HSP70 смысловой (5'-G C T C T A G A T A T G G C T C G T G C G G T C G G G A T C G A C C -3' (SEQ ID NO:39)) и HSP70 антисмысловой (5'-G C C G C G G C C G C T C A C T T G G C C T C C C G G C C G T C G T C G -3' (SEQ ID NO:40). Амплифицированную ДНК вводили на 3'-конце кодирующей СЕА последовательности с получением плазмид pV1J/CEA-FrC, pV1J/CEA-DOM, pV1J/CEA-FcIgG, pV1J/CEA-LAMP, pV1J/CEA-HSP70 и pV1J/CEA-VSV-G.
pV1J/CEA-DOMopt и pV1J/CEA-FcIgGopt: Оптимизированные в отношении использования кодонов кДНК DOM и FcIgG синтезировали сборкой олигонуклеотидов (Geneart GmbH, Regensburg, Germany) и клонировали в вектор pCR-script (Stratagene, La Jolla, CA). Для генерирования pV1J/CEA-DOMopt, DOMopt амплифицировали при помощи ПЦР с использованием следующих праймеров: DOMopt смысловой DOM (5'- G T T A T C T A G A A G C A C C C C C A T C C C -3' (SEQ ID NO:41)) и DOMopt обратный (5'-T T A A G A T C T C T A A G A T C T G G T G T C G T A T C T C A G G G G -3' (SEQ ID NO:42). Затем амплифицированный продукт встраивали в сайты XbaI/BglII плазмиды pV1J/CEAopt. Для генерирования pV1J/CEA-FcIgGopt, FcIgGopt амплифицировали при помощи ПЦР с использованием следующих праймеров: FcIgGopt смысловой (5'- T T A T C T A G A A A G A C C C A C A C C T G C C C C C C T T G C -3' (SEQ ID NO:43)) и FcIgGopt обратный (5'- T A T A G A T C T T A G G G T A C C T T A C T T G C C G G G G -3' (SEQ ID NO:44)), амплифицированный продукт встраивали в сайты XbaI/BglII плазмиды pV1J/CEAopt.
ПРИМЕР 3
Аденовирусные векторы
Ad5/hCEA opt: Плазмиду pCR-hCEAopt расщепляли EcoRI. Полученную инсерцию 2156 п.н. очищали и клонировали в EcoRI челночной плазмиды polyMRK-Ad5.
Ad5/CEA: Челночную плазмиду pMRK-hCEA для генерирования вектора Ad5 получали расщеплением плазмиды pDelta1sp1B/hCEA SspI и EcoRV. Затем фрагмент 9,52 т.п.н. лигировали с 1272 п.н. BglII/BamHI-рестрицированным, обработанным фрагментом Кленова продуктом из плазмиды polyMRK. PacI/StuI-фрагмент из pMRK-hCEA и pMRK-hCEAopt, содержащий экспрессионную кассету для hCEA и Е1 фланкирующих районов Ad5, рекомбинировали с линеаризованной ClaI плазмидой pAd5 в клетках E. coli BJ5183. Полученными плазмидами были pAd5-hCEA и pAd5-hCEAopt, соответственно. Обе плазмиды разрезали PacI для высвобождения ITR Ad и трансфицировали в клетки PerC-6. Амплификацию векторов Ad5 проводили при помощи серийного пассирования. MRKAd5/hCEA и MRKAd5/hCEAopt очищали при помощи стандартной очистки в градиенте CsCl и интенсивно диализовали против буфера А105 (5 мМ Трис-Cl рН 8,0, 1 мМ MgCl2, 75 мМ NaCl, 5% сахароза, 0,005 Твин 20).
Ad5/hCEAopt-LTB: Плазмиду pMRK-hCEAopt-LTB конструировали разрезанием челночной плазмиды polyMRK-Ad5 SwaI и лигированием этого линеаризованного вектора с ДНК-фрагментом 2300 п.н., полученным из pV1J/hCEAopt-LTB, которая была рестрицирована EcoRI, BglII и обработана фрагментом Кленова. Эту pMRK-hCEAopt-LTB линеаризовали и рекомбинировали в геном Ad, как указано выше.
Ad5/CEA-DOMopt и Ad5/CEA-FcIgGopt: Плазмиды pMRK-CEA-DOMopt и pMRK-CEA-FcIgGopt конструировали разрезанием челночной плазмиды polyMRK-Ad5 SwaI и лигированием этого линеаризованного вектора с ДНК-фрагментом 2,9 т.п.н., полученным из pV1J/CEA-DOMopt, или лигированием этого линеаризованного вектора с ДНК-фрагментом 2700 п.н., полученным из pV1J/CEA-FcIgG1opt, которые были рестрицированы EcoRI, BglII и обработаны фрагментом Кленова. pMRK-CЕA-FcIgGopt и pMRK-CEA-DOMopt линеаризовали и рекомбинировали в геном Ad, как указано выше.
ПРИМЕР 4
Сравнительная эффективность экспрессии различных слитых конструкций СЕА-LT
Было показано, что применение оптимизированных в отношении кодонов кДНК для генетической вакцинации против вирусных заболеваний индуцирует более высокий иммунный ответ, частично вследствие увеличенной экспрессией белка-мишени. Для проверки, будет ли кодирующая LTВ последовательность также усиливать иммуногенные свойства кДНК СЕА, сконструированной для включения предпочтительных для человека (гуманизированных) кодонов для каждого аминокислотного остатка, конструировали также плазмиду pV1J/hCEAopt-LTB. Наконец, конструировали также полный оптимизированный в отношении кодонов вариант слитой конструкции СЕА-LTВ с использованием синтетической оптимизированной в отношении кодонов кДНК LTВ для генерирования плазмиды pV1J/hCEA-LTBopt.
Для определения, не ограничивается ли действие LTВ на иммуногенность СЕА иммунизацией плазмидной ДНК, конструировали также вектор аденовируса типа 5, кодирующий слитую конструкцию СЕАopt-LTB, фланкированную промотором CMV/интроном А и сигналом полиаденилирования BGH. Молекулярная масса этих слитых белков СЕА, экспрессируемых как плазмидным, так и аденовирусным векторами, не отличалась от молекулярной массы, полученной из соответствующих векторов, кодирующих полноразмерную форму кДНК СЕА (данные не показаны).
Для сравнения эффективности экспрессии векторов, кодирующих слитые конструкции СЕА-LTA и СЕА-LTВ, и эффективности экспрессии кДНК полноразмерного СЕА, клетки HeLa трансфицировали плазмидами pV1J/hCEA-LTA и pV1J/hCEA-LTB. Экспрессию СЕА этих конструкций сравнивали с экспрессией соответствующей плазмиды, несущей кДНК СЕА дикого типа, pV1J/hCEA. Подобным образом, эффективность экспрессии плазмиды pV1J/hCEAopt-LTВ сравнивали с эффективностью экспрессии pV1J/hCEAopt. Эффективность экспрессии этих конструкций определяли через два дня после трансфекции мониторингом количества белка СЕА в клеточных экстрактах.
Трансфекция плазмид pV1J/hCEA-LTA и pV1J/hCEA-LTВ давала приблизительно в два раза более высокие количества белка СЕА (183 и 139 мкг/л, соответственно, фигура 8А), детектируемые в культуральном супернатанте, в сравнении с плазмидой pV1J/hCEA (91 мкг/л). Подобным образом, эффективность экспрессии конструкций pV1J/hCEAopt и pV1J/hCEAopt-LTВ была также сравнимой (113 и 136 мкг/л, соответственно; фигура 8В). Наконец, эффективность экспрессии Ad/hCEAopt и Ad/hCEAopt-LTB также сравнивали посредством инфицирования клеток HeLa при различной moi (множественности заражения). Эффективность экспрессии СЕА этих двух векторов была сравнимой при moi 1000 (1790 и 1400 мкг/л, соответственно, фигура 9), тогда как при moi 100 вектор Ad/hCEAopt-LTB давал приблизительно в четыре раза более низкие количества белка СЕА, детектируемого в культуральном супернатанте, чем Ad/hCEAopt (390 и 1500 мкг/л, соответственно).
Таким образом, эти результаты показывают, что кДНК, кодирующая слитые белки СЕА-LTA и СЕА-LTВ, экспрессируется с эквивалентной эффективностью относительно эффективности экспрессии соответствующей кДНК, кодирующей полноразмерный белок СЕА. Кроме того, сравнимая экспрессия СЕА этих кДНК не зависела от типа вектора-переносчика генов, используемых для их доставки.
ПРИМЕР 5
Детектирование экспрессии СЕА
Экспрессию СЕА плазмидными и Ad-векторами подвергали мониторингу с использованием Вестерн-блот-анализа и ELISA. Плазмиды трансфицировали в клетки HeLa с использованием Липофектамина 2000 (Life Technologies). Аденовирусные инфицирования клеток HeLa выполняли в бессывороточной среде в течение 30 минут при 37оС и затем добавляли свежую среду. После 48 часов инкубирования собирали лизаты целых клеток. Белок СЕА, присутствующий в этих клеточных лизатах, детектировали с использованием Вестерн-блот-анализа с применением кроличьей поликлональной антисыворотки. Этот белок детектировался в виде полосы 180-200 кДа. Количество экспрессированного СЕА детектировали в клеточных лизатах с использованием набора Direct Elisa CEA (DBC-Diagnostics Biochem Canada Inc.).
Экспрессию слитых белков в трансфицированных клетках исследовали при помощи Вестерн-блот-анализа с использованием антител, специфических в отношении СЕА, VSV-G, FcIgG, столбнячного токсина или HSP70. Клетки HeLa либо трансфицировали указанной плазмидой, либо инфицировали выбранным Ad-вектором. После 48 часов инкубирования собирали лизаты целых клеток и культуральные супернатанты.
Экспрессию СЕА в клеточном лизате или супернатанте также подвергали мониторингу с использованием набора Direct Elisa CEA (DBC-Diagnostics Biochem Canada Inc.). Белок СЕА детектировался антителом, специфическим в отношении этого слитого полипептида, в трансфицированных клеточных лизатах, в то время как не наблюдали экспрессии антигена-мишени в ложнотрансфицированных контрольных пробах (фигура 18В). Молекулярная масса этих слитых белков не отличалась значимо от молекулярной массы СЕА. Это кажущееся отсутствие различия в молекулярной массе между различными полипептидами СЕА обусловлено, возможно, высокой степенью гликозилирования этого опухолевого антигена.
Для сравнения эффективности экспрессии векторов, кодирующих эти cлитые конструкции СЕА, с эффективностью экспрессии pV1J/CEA, клетки HeLa трансфицировали различными плазмидами и экспрессию СЕА этими конструкциями определяли через два дня после трансфекции при помощи ELISA. Плазмиды pV1J/CEA-FrC, pV1J/CEA-DOM, pV1J/CEA-FcIgG, pV1J/CEA-LAMP, pV1J/CEA-VSV-G и pV1J/CEA-HSP70 экспрессировали СЕА со сравнимой эффективностью относительно pV1J/CEA (фигура 19А). Большинство слитых белков секретировались и детектировались в клеточном супернатанте; однако, СЕА-LAMP не высвобождался из трансфицированных клеток, возможно, вследствие его перехода в лизосомный компартмент (фигура 19В). Таким образом, эти результаты показывает, что кДНК, кодирующия слитые белки СЕА-FrC, СЕА-DOM, СЕА-VSV-G, СЕА-FcIgG, СЕА HSP70 и СЕА-LAMP, экспрессируются с эффективностью, эквивалентной эффективности кДНК, кодирующей полноразмерный белок СЕА.
ПРИМЕР 6
Пептиды
Лиофилизированные пептиды hCEA покупали из Bio-Synthesis и ресуспендировали в ДМСО при 40 мг/мл. Пулы пептидов с длиной 15 аминокислот, перекрывающиеся 11 остатками, собирали, как описано (Facciabene et al., J. Virol. 78: 8663-72 (2004). Конечные концентрации были следующими: пул А=1,2 мг/мл, пул В 0,89 мг/мл, пул С 0,89 мг/мл, пул D 0,8 мг/мл. Пептиды хранили при -80оС. Иммунную реакцию на DOM подвергали мониторингу с использованием пептида столбнячного токсоида р30 (F947NNFTVSFWLRVPKVSASHLE967 (SEQ ID NO:54) (Rice et al., J. Immunol. 167: 1558-65 (2001)).
ПРИМЕР 7
Иммунизация мышей и стимуляция опухолевым антигеном
Все исследования на животных были одобрены IRBM institutional animal care and use committeе. Самок мышей С57BL/6 (Н-2b) покупали у Charles River (Lecco, Italy). Мыши HLA-А2.1 (HHD) были любезно предоставлены F. Lemmonier (Institute Pasteur, Paris, France). Мышей С57BL/DR4 покупали у Taconic (Germantown, NY). Мыши СЕА.tg (трансгенные) (H-2b) были любезно предоставлены J. Primus (Vanderbilt University) и их содержали в стандартных условиях (Clarke et al., Cancer Res. 58:1469-77 (1998)). Пятьдесят микрограммов плазмидной ДНК электроинъецировали в объеме 50 мкл в четырехглавые мышцы мышей, как описано ранее (Rizzuto et al., Proc. Natl. Acad. Sci. USA 96(11): 6417-22 (1999)). Инъекции Ad проводили в четырехглавые мышцы мышей в объеме 50 мкл. Гуморальную и клеточно-опосредованную иммунные реакции анализировали в указанное время.
Мышей C57BL/6 подвергали двум инъекциям ДНК в четырехглавую мышцу с последующей электрической стимуляцией, как описано выше (Rizzuto et al., supra). Инъекции проводили при трехнедельных интервалах. СЕА-трансгенных мышей подвергали либо 5 еженедельным инъекциям плазмидной ДНК (50 мкг/инъекция), 2 инъекциям Ad-векторов (1×109 вирусных частиц/инъекция), либо 5 еженедельным инъекциям с последующей стимуляцией Ad. Через две недели после последней инъекции анализировали гуморальную и клеточно-опосредованную реакцию. Мышам вводили также подкожную (s.c) инъекцию 5×105 клеток МС38-СЕА (Clarke et al., supra). При интервалах, равных одной неделе, мышей исследовали на рост опухоли.
ПРИМЕР 8
Обнаружение и титрование антител
Сыворотки для титрования антител получали ретроглазничным кровоизвлечением. Планшеты ELISA (Nunc maxiprep) покрывали 100 нг на лунку высокоочищенного белка СЕА (Fitzgerald), разведенного в буфере для сенсибилизации поверхностей (50 мМ NaHCO3, рН 9,4) и инкубированного в течение ночи при 4оС, как описано ранее (Facciabene et al., supra). Затем планшеты блокировали PBS, содержащим 5% БСА, в течение 1 часа при 37°С. Сыворотки мышей разбавляли в PBS с 5% БСА (разведение 1/50 для оценки степени сероконверсии; разведение 1:10 - 1:31,2150 для оценки титра). Преиммунизированные сыворотки использовали в качестве фона. Разведенные сыворотки инкубировали в течение ночи при 4°С. Промывки проводили при помощи PBS с 1% БСА, 0,05% Твином 20. Вторичные антитела (козьи антитела против мышиного IgG, конъюгированные с пероксидазой, Sigma) разводили в PBS, 5% БСА и инкубировали 2-3 часа при комнатной температуре на шейкере. После промывания планшеты проявляли с использованием 100 мкл на лунку субстрата ТМВ (Pierce Biotechnology, Inc., Rockford, IL). Реакцию останавливали 25 мкл на лунку 1 М раствора H2SO4 и планшеты считывали при 450 нм/620 нм. Титры анти-СЕА-сывороток рассчитывали в виде реципрокного лимитирующего разведения сыворотки, дающего оптическую плотность, по меньшей мере, в 3 раза более высокую, чем оптическая плотность аутологичной преиммунизированной сыворотки при том же самом разведении.
ПРИМЕР 9
IFN-γ-ELISPOT-анализ
Анализы проводили с использованием спленоцитов мыши и СЕА-специфических пептидов, как описано ранее (Facciabene et al., supra). 96-луночные планшеты MAIP (Millipore Corp., Billerica, MA) сенсибилизировали 100 мкл на лунку очищенного крысиного антитела против мышиного IFN-γ (IgG1, клон R4-6A2, Pharmingen), разведенного до 2,5 мкг/мл в стерильном PBS. После промывания PBS проводили блокирование планшетов 200 мкл на лунку среды R10 в течение 2 часов при 37°С.
Спленоциты получали стерильным удалением селезенки из эвтанизированных мышей и разрушением селезенки протиранием на металлической решетке. Эритроциты удаляли осмотическим лизисом добавлением 1 мл 0,1Х PBS к осадку клеток и встряхиванием на вортексе в течение приблизительно 15 секунд. Затем добавляли один мл 2× PBS и объем доводили до 4 мл 1× PBS. Клетки осаждали центрифугированием при 1200 об/мин в течение 10 минут при комнатной температуре и осадок суспендировали в 1 мл среды R10. Жизнеспособные клетки считали с использованием окрашивания Türks.
Спленоциты высевали при 5×105 и 2,5×105 клеток на лунку в двух повторностях и инкубировали в течение 20 часов при 37°С с 1 мкг/мл суспензии каждого пептида. Конканавалин А (ConA) использовали в качестве положительного внутреннего контроля для каждой мыши в концентрации 5 мкг/мл. После промывания PBS, 0,05% Твином 20 планшеты инкубировали в течение ночи при 4°С с 50 мкл на лунку конъюгированных с биотином крысиных антител против мышиного IFNγ (крысиных RatIgG1, клон XMG 1.2, PharMingen), разведенных 1:2500 в буфере для анализа. После интенсивного промывания планшеты проявляли добавлением 50 мкл на лунку NBT/B-CIP (Pierce Biotechnology, Inc., Rockford, IL), пока проявление пятен не становилось ясно видимым. Реакцию останавливали тщательным промыванием планшетов дистиллированной водой. Планшеты сушили на воздухе и затем считали с использованием автоматического ELISPOT-ридера.
ПРИМЕР 10
Внутриклеточное окрашивание цитокинов
Один-два миллиона спленоцитов мыши или PBMC в 1 мл RPMI с 10% FCS инкубировали с пулом пептидов (конечная концентрация каждого пептида 5-6 мкг/мл) и брефелдином А (1 мкг/мл; BD Pharmingen cat. #555028/2300kk) при 37°С и 5% СО2 в течение 12-16 часов, как описано ранее (Facciabene et al., supra). Затем клетки промывали FACS-буфером (PBS-1% FCS, 0,01% NaN3) и инкубировали с очищенным анти-CD16/CD32 Fc-антителом (блокирующим реагентом) (BD Pharmingen cat # 553142) в течение 15 мин при 4°С. Затем клетки промывали и окрашивали поверхностными антителами: CD4-РЕ-конъюгированное антимышиное антитело (BD Pharmingen cat. #553049), PercP CD8-конъюгированное антимышиное антитело (BD Pharmingen cat. #553036) и АРС-конъюгированное антимышиное антитело против CD3е (BD Pharmingen cat. #553066) в течение 30 мин при комнатной температуре в темноте. После промывания клетки фиксировали и увеличивали их проницаемость раствором Cytofix-Cytoperm (BD Pharmingen cat. #555028/2300kk) в течение 20 мин при 4˚С в темноте. После промывания раствором PermWash (BD Pharmingen cat. #555028/2300kk) клетки инкубировали с IFNγ-FITC-антителами (BD Pharmingen). Затем клетки промывали, фиксировали 1% формальдегидом в PBS и анализировали на проточном цитометре FACS-Calibur с использованием программы CellQuest (Becton Dickinson, San Jose, CA).
ПРИМЕР 11
Иммуногенность слитых конструкций СЕА-LT
Для испытания иммунных реакций, индуцируемых плазмидами, кодирующими слитые конструкции СЕА-LTA и СЕА-LTВ, группы из 9 мышей С57BL/6 иммунизировали двумя инъекциями i.m. 50 мкг каждой из плазмид pV1J/hCEA, pV1J/hCEA-LTA и pV1J/hCEA-LTВ. Дополнительно, для проверки, может ли коэкспрессия слитых белков СЕА-LTA и СЕА-LTВ иметь дополнительное действие на иммуногенность белка СЕА, группу мышей иммунизировали совместной инъекцией 25 мкг каждой из плазмид pV1J/hCEA-LTA и pV1J/hCEA-LTВ. Иммунизации проводили с трехнедельными интервалами. Плазмидную ДНК рутинным образом электроинъецировали в скелетную мышцу мыши в связи с усиленной трансдукцией и иммуногенностью, связанными с этой конкретной процедурой (Zucchelli et al. J. Virol. 74: 11598-11607 (2000); Widera et al. J. Immunol. 164: 4635-4640 (2000)).
Клеточный иммунитет, индуцируемый различными плазмидами, измеряли при помощи ELISPOT-анализа через 2 недели после последней инъекции. Антигенспецифическую секрецию IFNγ из стимулированных спленоцитов измеряли с использованием четырех пулов 15-мерных пептидов, перекрывающихся 11 аминокислотами и включающими в себя полный гликопротеин СЕА. Пул А включает в себя аминокислоты 1-147, пул В включает в себя аминокислоты 137-237, пул С включает в себя аминокислоты 317-507 и пул D включает в себя аминокислоты 497-703. В качестве отрицательного контроля продукцию цитокинов измеряли также после стимуляции спленоцитов ДМСО в той же самой концентрации, которую использовали для солюбилизации пептидов СЕА.
Иммунный ответ, индуцируемый ДНК-вакцинацией у мышей С57BL/6, был прежде всего смещен к С-концу этого белка, так как величины SFC (образующих пятна клеток), детектированные с пулом А пептидов, были слегка выше фона со всеми конструкциями (фигура 10). Схема вакцинации pV1J/hCEA-LTB превосходила схему вакцинации pV1J/hCEA, как показано более высокими величинами геометрического среднего SFC, детектированными с пулами В, С и D (pV1J/hCEA-LTB: 482, 1436 и 2054 SFC/106 спленоцитов, соответственно; pV1J/hCEA: 45, 350 и 264 SFC/106 спленоцитов, соответственно). Подобным образом, плазмида pV1J/hCEA-LTA имела также усиливающее действие на СЕА-специфический иммунный ответ в сравнении с pV1J/hCEA. Однако, увеличение иммунного ответа наблюдали только с пулами С и D пептидов (925 и 528 SFC/106 спленоцитов, соответственно), тогда как иммунный ответ, измеренный с пулом В пептидов, был низким (15 SFC/106 спленоцитов). Кроме того, совместная инъекция плазмид pV1J/hCEA-LTA и pV1J/hCEA-LTВ не обнаруживала синергического действия на иммунный ответ СЕА в сравнении с иммунным ответом, измеренным в обработанной pV1J/hCEA-LTB группе, а, скорее, приводила к уменьшению величин SFC, детектируемых с пептидами пула В и D (210 и 528 SFC/106 спленоцитов, соответственно).
Для определения Т-клеточной специфичности, индуцируемой после вакцинации различными конструкциями СЕА, внутриклеточное окрашивание IFNγ проводили на объединенных спленоцитах из инъецированных мышей с использованием пула D пептидов. CD8+-специфический ответ детектировали у мышей, инъецированных pV1J/hCEA-LTB, превосходящий (4,5%) CD8+-специфический ответ, детектируемый с pV1J/hCEA-LTA и pV1J/hCEA (0,14% и 0,8%, соответственно, фигура 10В). В отличие от этого, pV1J/CEA-LTA индуцировал сильную CD4+-специфическую реакцию, более высокую (1,21%), чем CD4+-специфическая реакция, наблюдаемая с pV1J/hCEA-LTВ и pV1J/hCEA (0,55% и 0,58%, соответственно).
Индукцию гуморального иммунного ответа на СЕА испытывали измерением антигенспецифических антител (фигура 11). Обе плазмиды pV1J/hCEA-LTA и pV1J/hCEA-LTВ индуцировали более высокую реакцию антител, чем pV1J/hCEA, подтверждая адъювантное действие, проявляемое субъединицами LT, на СЕА-специфический иммунный ответ. Таким образом, эти результаты демонстрируют, что слитая конструкция кодирующей СЕА последовательности с кДНК LTA и LTВ приводит к увеличению СЕА-специфического иммунного ответа. Однако LTВ, по-видимому, имеет большее усиливающее действие на этот иммунный ответ с преобладающей индукцией CD8+ Т-клеток, тогда как LTA индуцирует преобладающую CD4+ реакцию.
ПРИМЕР 12
Иммуногенность слитых конструкций СЕА-LTВ в различных штаммах мышей
Для определения, не ограничивается ли усиливающее действие субъединиц LT на СЕА-специфический иммунный ответ только единственным генетическим фоном мышей, иммунизации на основе ДНК проводили на мышах BALB/c, C57/DR4 и HLA-А2.1 (ННD). Мыши BALB/c были выбраны в связи с их иммунокомпетентностью, так как этот штамм мышей является чрезвычайно реактивным на схемы иммунизации различных сортов. HHD-трансгенные мыши экспрессируют гены МНС класса I человека. Подобным образом, трансгенные мыши С57/DR4 несут гены МНС класса II человека. Таким образом, эти два штамма трансгенных мышей могут обеспечить информацию в отношении иммунореактивности слитых конструкций СЕА-LT в контексте с гаплотипами как МНС класса I, так и МНС класса II человека.
СЕА-специфический иммунный ответ у мышей BALB/c сначала оценивали с использованием ELISPOT-анализа. Усиление антигенспецифического иммунного ответа после иммунизации плазмидой pV1J/hCEA-LTВ детектировали с пулами А, В, С, D пептидов (pV1J/hCEA-LTВ: 166, 1353, 796, 899 SFC/106 спленоцитов, соответственно; pV1J/hCEA: 57, 312, 327, 318 SFC/106 спленоцитов, соответственно; фигура 12). Как наблюдали и у мышей С57BL/6, N-конец белка СЕА, по-видимому, является наименее иммуногенным в сравнении с другими участками этого опухолевого антигена. Иммунизация pV1J/hCEA-LTA также давала увеличение антигенспецифического иммунного ответа в сравнении с pV1J/hCEA. Это увеличение иммунного ответа детектировали с пулами пептидов В, С и D (936, 727 и 650 SFC/106 спленоцитов, соответственно). Кроме того, совместная инъекция этих двух плазмид pV1J/hCEA-LTA и pV1J/hCEA-LTВ давала значимый аддитивный эффект, который детектировали в основном с пулами пептидов С и D (1783 и 2141 SFC/106 спленоцитов, соответственно).
СЕА-специфический иммунный ответ у мышей С57/DR4 значительно усиливался иммунизацией pV1J/hCEA-LTB и детектировался только с одним пулом пептидов D (фигура 13). Окрашивание внутриклеточного IFNγ, выполняемое на объединенных PBMC из инъецированных мышей, показало, что CD8+ реакция на СЕА была наивысшей у мышей, иммунизированных pV1J/hCEA-LTB (15,32%), тогда как была очень слабой в обработанной pV1J/hCEA группе (0,5%). Иммунизация pV1J/hCEA-LTA увеличивала антигенспецифический иммунный ответ лишь умеренно (0,43%) и не усиливала дополнительно иммуногенность СЕА при совместной инъекции с конструкцией, кодирующей слитую конструкцию СЕА-LTB (13,44%). Интересно, что в этих иммунизированных мышах не детектировалась значимая CD4+ Т-клеточная реакция (данные не показаны).
Иммунный ответ, индуцируемый различными СЕА-кодирующими плазмидами, оценивали у мышей HHD выполнением внутриклеточного окрашивания IFNγ на объединенных PBMC. Иммунный ответ детектировали только с пулами пептидов В и D, и, как показано на фигуре 14, иммунизация pV1J/hCEA-LTВ приводила к более чем 10-кратному увеличению CD8+ реакции на антигены-мишени. В отличие от этого, увеличение иммунного ответа не детектировали с использованием pV1J/hCEA-LTA ни отдельно, ни после совместной инъекции с pV1J/hCEA-LTB. CD4+ Т-клеточную реакцию не детектировали в этих иммунизированных мышах (данные не показаны).
Взятые вместе, эти результаты подтверждают, что слитая конструкция кодирующей LTB последовательности с СЕА приводит к значительному увеличению антигенспецифического иммунного ответа. Интересно, что этот ответ является преимущественно CD8+-специфическим и может наблюдаться в разных штаммах мышей, что указывает, следовательно, на то, что усиливающее действие, проявляемое субъединицей LT, не является генотип-рестриктированным.
ПРИМЕР 13
Толерантность к СЕА человека в трансгенных мышах
Для определения, могут ли усиленные иммуногенные свойства слитой конструкции hCEA-LTB разрушать толерантность к СЕА человека более эффективно, hCЕA-трансгенных мышей иммунизировали векторами, несущими либо полностью оптимизированную в отношении кодонов кДНК hCEA, либо CЕА-LTВ. Эти трансгенные мыши несут полный ген СЕА человека и фланкирующие последовательности этого гена и экспрессируют белок hCEA в кишечнике, в основном в слепой кишке и ободочной кишке. Таким образом, эта мышиная линия является полезной моделью для исследования безопасности и эффективности стратегий иммунотерапии, нацеленных против этого опухолевого аутоантигена (Clarke et al., Cancer Research 58: 1469-1477 (1998)).
Иммунизация pV1J/hCEA-LTBopt приводила к значимому увеличению СЕА-специфического иммунного ответа, измеренного посредством внутриклеточного окрашивания IFNγ на PBMC инъецированных мышей (фигура 15А). Усиление Т-клеточной реакции детектировали с пулом пептидов D, и оно было преимущественно усилением CD8+-реакции. Кроме того, СЕА-специфический гуморальный ответ увеличивался в СЕА-LTВ-обработанных мышах, как показано 47-кратным увеличением величин геометрического среднего титра Ab в сравнении с pV1J/hCEAopt-обработанной группой (фигура 15В).
Для определения, наблюдается ли усиливающее действие, проявляемое LTВ на СЕА-специфическом иммунном ответе, также и после иммунизации векторами, другими, чем плазмидная ДНК, группы из 12 СЕА-трансгенных мышей иммунизировали Ad5/hCEAopt-LTB и Ad/hCEAopt в дозе 1×107, 1×108 и 1×109 вч. Мышей подвергали двум инъекциям с двухнедельным интервалом и иммунный ответ измеряли внутриклеточным окрашиванием IFNγ на PBMC через две недели после последней инъекции. Иммунный ответ оценивали с использованием пула пептидов D. Ad/hCEAopt-LTB был более иммуногенным, чем Ad/hCEAopt, так как значимые иммунные ответы на СЕА могли быть детектированы с дозой 1×108 вч, тогда как дозы 1×109 вч Ad/hCEAopt были необходимы для разрушения толерантности к этому антигену-мишени (фигура 16). CD4+-реакция не могла быть обнаружена ни в одной из иммунизированных мышей (данные не показаны).
Эти данные подтверждают, что толерантность к этому аутоантигену может быть разрушена более эффективно вследствие увеличенных иммуногенных свойств слитой конструкции СЕА-LTВ. Кроме того, усиливающее действие LTB на иммуногенные свойства СЕА могут наблюдаться также после инъекции плазмиды, несущей полностью оптимизированную в отношении кодонов кДНК слитую конструкцию СЕА-LTB. Наконец, эти результаты показывают, что усиленная иммуногенность СЕА-LTB не ограничивается иммунизацией плазмидной ДНК.
ПРИМЕР 14
Кинетика роста опухоли у СЕА-трансгенных мышей, иммунизированных слитыми конструкциями СЕА-LTВ
Авторы считали полезным определение, приводит ли увеличенная иммуногенность слитой конструкции СЕА-LTВ также и к усиленному терапевтическому действию, способному препятствовать прогрессированию опухоли. Для этой цели группы из 10 СЕА-трансгенных (tg) мышей подвергали 5 инъекциям один раз в неделю плазмид pV1J/hCEAopt или pV1J/hCEAopt-LTB с последующей конечной стимуляцией 1×1010 вч соответствующего Ad-вектора. В связи с недавними сообщениями, которые указывают на то, что высокие уровни клеточного иммунитета могут быть индуцированы против вирусных и бактериальных антигенов с использованием схемы прайминга плазмидой ДНК - стимуляции Ad, в этом исследовании использовали тот же самый протокол иммунизации. Через две недели после последней иммунизации СЕА-трансгенных мышей провоцировали подкожной инъекцией 5х105 опухолевых клеток МС38-СЕА. Эту сингенную клеточную линию получали из химически индуцированного рака ободочной кишки, и она экспрессирует СЕА. Развитие опухоли в ложнообработанных мышах детектировали на 22 день после стимуляции (введения опухолевого антигена), когда все обработанные мыши уже не были не имеющими опухоли (фигура 17А). Кроме того, было сопутствующее увеличение среднего размера массы опухоли, которая достигала значительного объема на 34 день после стимуляции. Мыши, вакцинированные векторами, кодирующими pV1J/hCEAopt, показали частичную устойчивость к развитию опухоли, так как 3 из 10 обработанных мышей оставались не имеющими опухоли в день 34 после стимуляции. Средний размер опухолей этой группы был меньше, чем объем опухолей, наблюдаемых в ложнообработанных мышах. Иммунизация векторами, кодирующими слитую конструкцию СЕАopt-LTB, приводила к значимому протективному действию против развития опухоли. Пять из 10 обработанных мышей оставались не содержащими опухоли в день 34 после стимуляции, и средний размер массы опухоли в этой группе был значимо меньшим, чем средний размер массы опухоли в ложнообработанных или обработанных pV1J/hCEAopt мышах. Таким образом, эти результаты указывают на то, что усиленный СЕА-специфический иммунный ответ, связанный с векторами, кодирующими слитую конструкцию СЕА-LTB, коррелирует со значимым противоопухолевым действием, приводящим к частичной защите от опухолевого роста и пониженной кинетике роста массы опухоли.
ПРИМЕР 15
Слитые конструкции СЕА-DOM и СЕА-FcIgG, усиливающие иммуногенность белка СЕА
Для исследования иммунных ответов, индуцированных плазмидами, кодирующими слитые конструкции СЕА-FrC, CЕА-DOM, СЕА-VSV-G, СЕА-FcIgG, СЕА-HSP70 и СЕА-LAMP, группы из 9 мышей С57BL/6 иммунизировали двумя i.m. инъекциями 50 или 5 мкг каждой плазмиды. Иммунизации проводили с трехнедельными интервалами. Ввиду усиленной трансдукции и иммуногенности, сообщенной для электропорации (Zucchelli et al. J. Virology 74: 11598 (2000), Widera et al., J. Immunol. 164: 4635 (2000)), плазмидные ДНК рутинным образом электропорировали (DNA-EP) в скелетную мышцу мыши.
Иммунный ответ, индуцированный разными плазмидами, измеряли IFNγELISPOT-анализом через 2 недели после последней инъекции. Антигенспецифическую секрецию IFNγ из стимулированных спленоцитов измеряли с использованием пула 15-мерных пептидов, перекрывающихся 11 аминокислотами и включающих в себя С-конец СЕА (пул D, аминокислоты 497-703) (Zucchelli et al., supra). Анализ иммунного ответа на СЕА проводили с пулом пептидов D, так как клеточный иммунный ответ на СЕА у мышей С57BL/6 первично смещен в сторону С-конца этого белка (Zucchelli et al., supra). В качестве отрицательного контроля продукцию цитокина измеряли также после стимуляции спленоцитов ДМСО в той же концентрации, которую использовали для солюбилизации пептидов СЕА.
Инъекция pV1J/CEA-DOM или pV1J/CEA-Fc индуцировала более высокий иммунный ответ на СЕА в сравнении с pV1J/CEA. Более высокая иммуногенность этих двух слитых белков приводила к более высоким величинам геометрических средних значений образующих пятна клеток (SFC) на 106 спленоцитов (фигура 20А). Плазмиды pV1J/CEA-DOM и pV1J/CEA-FcIgG имели сходные иммуногенные свойства и проявляли 3-4-кратное увеличение СЕА-специфических иммунных ответов после инъекции 5 или 50 мкг плазмидной ДНК (pV1J/CEA-DOM: 590 и 1098 SFC/106 спленоцитов, pV1J/CEA-FcIgG: 510 и 1160, pV1J/hCEA: 146 и 264 SFC/106 спленоцитов, соответственно). Не отмечали значимых различий между величинами SFC, индуцированными pV1J/CEA-FrC, pV1J/CEA-LAMP, pV1J/CEA-HSP70 и pV1J/CEA. Не детектировали СЕА-специфических иммунных ответов в отрицательных контрольных пробах.
Для определения действия слитых конструкций СЕА на гуморальную реакцию на СЕА, сыворотки из иммунизированных мышей тестировали при помощи ELISA с использованием очищенного белка СЕА в качестве субстрата (фигура 20В). Увеличение титра СЕА-специфических антител наблюдали после инъекции 50 мкг плазмид pV1J/CEA-DOM, pV1J/CEA-FcIgG, pV1J/CEA-FrC и pV1J/CEA-HSP70. Напротив, инъекция pV1J/CEA-LAMP и pV1J/CEA-VSV-G приводила к СЕА-специфической реакции антител, сходной с реакцией, наблюдаемой после иммунизации pV1J/CEA. Взятые вместе, эти данные демонстрируют, что слитая конструкция кодирующей СЕА последовательности с кДНК DOM или FcIgG приводит к увеличению СЕА-специфического клеточно-опосредованного и гуморального иммунного ответа.
ПРИМЕР 16
Слитые конструкции СЕА-DOM и СЕА-FcIgG, разрушающие толерантность к антигену-мишени у СЕА-трансгенных мышей
Толерантность к антигену-мишени является одним из барьеров, которые противораковая вакцина должна преодолеть для индукции иммунного ответа и для проявления эффективного противоракового действия. Таким образом, авторы считали важным определение, будут ли усиленные иммуногенные свойства слитых конструкций СЕА-DOM и СЕА-FcIgG разрушать толерантность к СЕА более эффективно, чем белок СЕА. Для этой цели СЕА-трансгенных мышей использовали для проведения сравнительных исследований иммунизации. Эти трансгенные мыши несут полный ген СЕА человека и фланкирующие последовательности гена СЕА человека и экспрессируют белок СЕА в кишечнике, в основном в слепой кишке и ободочной кишке. Таким образом, эта мышиная линия является полезной моделью для исследования безопасности и эффективности стратегий иммунотерапии, нацеленных против этого опухолевого аутоантигена (Clarke et al., supra).
Ввиду усиленных иммуногенных свойств векторов, несущих оптимизированную в отношении кодонов кДНК (cDNAopt) СЕА, конструировали как плазмидный, так и аденовирусный векторы, которые несут оптимизированную в отношении кодонов кДНК (cDNAopt) СЕА-DOM (СЕА-DOMopt) или СЕА-FcIgG (СЕА-FcIgGopt) слитых конструкций. Как наблюдали для СЕА, кДНК СЕА-DOMopt и СЕА-FcIgGopt, как было показано, экспрессируются с большей эффективностью, чем соответствующая кДНК дикого типа, приводя к усиленному иммунному ответу на СЕА (данные не показаны).
Иммуногенность этих двух слитых белков сравнивали с иммуногенностью СЕА посредством серии исследований иммунизации, основанных на использовании плазмидной ДНК и Ad-векторов, вводимых по отдельности или в комбинации. Группы СЕА-трансгенных мышей иммунизировали с использованием следующих варьирующихся схем введения: i) 5 инъекций с однонедельными интервалами 50 мкг плазмидной ДНК (ДНК/ДНК), ii) 2 инъекции один раз в две недели плазмидной ДНК с последующей конечной инъекцией аденовируса в дозах в диапазоне 1×107-1×109 вирусных частиц (вч) аденовируса (Ad/Ad) или iii) 5 инъекций один раз в неделю плазмидной ДНК с последующей конечной инъекцией 1×109 вч аденовируса (ДНК/Ad). Иммунные ответы анализировали внутриклеточным окрашиванием IFNγ на PBMC или спленоцитах каждой иммунизированной мыши с использованием пептидов пула D. Кроме того, индукцию СЕА-специфических антител подвергали мониторингу при помощи ELISA.
ДНК/ДНК-иммунизация СЕА-трансгенных мышей выявила, что векторы СЕА-DOMopt и СЕА-FcIgGopt проявляли измеримую CD8+ Т-клеточную реакцию на антиген-мишень (фигура 21А). Таким образом, обе конструкции были способны разрушать толерантность к СЕА у этих мышей. Антигенспецифическая реакция, индуцируемая слитыми белками СЕА-DOM и СЕА-FcIgG, была сравнимой, как показано средними величинами внутриклеточного окрашивания IFNγ (0,22 и 0,34%, соответственно). Тем не менее, иммунный ответ, индуцируемый этими двумя конструкциями, был большим, чем иммунный ответ, который наблюдали после вакцинации pV1J/hCEAopt (0,07%). Подобным образом, гуморальная реакция против СЕА была также большей после вакцинации этими слитыми белками. Титр СЕА-специфических антител детектировали во всех мышах, иммунизированных pV1J/CEA-DOMopt и pV1J/CEA-FcIgGopt, и средний титр антител был равен 56136 и 24725, соответственно. В противоположность этому, иммунизированная pV1J/CEAopt группа показала по меньшей мере в 77 раз более низкий титр СЕА-специфического антитела (318) (фигура 21В).
СЕА-трансгенные мыши, обработанные по схеме вакцинации Ad/Ad также показали лучшую эффективность в разрушении толерантности в СЕА после вакцинации Ad-векторами СЕА-DOMopt и СЕА-FcIgGopt, чем Ad-CЕАopt. СЕА-специфическая CD8+ Т-клеточная реакция могла наблюдаться у вакцинированных мышей после инъекции всего лишь 107 вч Ad-СЕА-DOM или Ad-СЕА-FcIgG, эта СЕА-специфическая реакция была сравнима между этими двумя антигенами, и увеличивалась после инъекции 109 вч (1,55% и 1,15%, соответственно). В отличие от этого, 109 вч Ad-СЕАopt требовались для индукции значимых частот предшественника CD8+ Т-клеток (2,1%) (фигура 21С). СЕА-специфические антитела детектировались у всех мышей, иммунизированных Ad-СЕА-DOMopt и Ad-CEA-FcIgGopt. Средний титр антител был 19600 и 33000, соответственно. Инъекция Ad-СЕАopt приводила к измеримой СЕА-специфической реакции только у 2 из обработанных мышей, и титр антител был значимо более низким (Zucchelli et al., supra) (фигура 21D). Интересно, что иммунизация ДНК/Ad показала уменьшенные различия в частотах предшественника CD8+ Т-клеток, индуцируемых векторами СЕА, СЕА-DOM и СЕА-FcIgG (фигура 22А). Однако, средние титры СЕА-специфических антител были более высокими после вакцинации векторами, экспрессирующими СЕА-DOM и СЕА-FcIgG, чем СЕА (31200, 26120 и 412, соответственно) (фигура 22В).
Интересно, что, независимо от антигена, не детектировали видимой CD4+ клеточной Тh1-реакции на СЕА ни в одной из трех схем вакцинации (данные не показаны). Однако значимая CD4+ клеточная Тh1-реакция против хелперного эпитопа, р30, присутствующая в последовательности DOM (Rice et al., J. Immunol. 167: 1558-65 (2001)), была обнаружена после ДНК/ДНК-вакцинации (0,4%) (фигура 23).
Таким образом, эти результаты демонстрируют, что слитые белки СЕА-DOM и СЕА-FcIgG могут разрушать толерантность к СЕА у трансгенных мышей с большей эффективностью, чем белок СЕА. Усиленные иммуногенные свойства этих слитых белков можно наблюдать после иммунизации ДНК-векторами или Ad-векторами. Однако более высокая способность этих двух слитых белков в индуцировании CD8+ Т-клеток на СЕА может быть преодолена, по меньшей мере частично, схемой вакцинации ДНК/Ad.
ПРИМЕР 17
Исследования с истощением Т-клеток
Иммунизированных животных истощали в отношении CD4+ Т-клеток, CD8+ Т-клеток, НК-клеток внутрибрюшинной инъекцией (i.p.) анти-CD4 (гибридома GK1.5), анти-CD8 (гибридома Lyt2.2) или анти-Asialo GM1 (Wako Chemicals, Richmobd, VA), как описано (Perricone et al., J. Immunother. 27(4): 273-81 (2004); Yoon et al., J. Ethnopharmacol. 93 (2-3): 247-53 (2004)). Антитела (100 мкл разведенной асцитной жидкости на дозу) инъецировали в день -7 относительно стимуляции опухолевым антигеном и затем инъецировали каждую неделю в течение 3 недель после инъекции 5×105 клеток МС38-СЕА. Условия истощения валидизировали анализом проточной цитометрией периферической крови с использованием конъюгированных с фикоэритрином mAb анти-CD4, анти-CD8 и анти-НК (PharMimgen, San Diego, CA); 99% релевантной субпопуляции клеток истощалось, тогда как все другие субпопуляции оставались в пределах нормальных уровней.
ПРИМЕР 18
СЕА-DOM-иммунизация, проявляющая противоопухолевое действие у СЕА-трансгенных мышей
Затем авторы настоящего изобретения определяли, приводит ли увеличенная иммуногенность слитых конструкций СЕА-DOM или СЕА-FcIgG также к усиленному терапевтическому действию, способному препятствовать прогрессированию опухоли. Для этой цели группы из 10 СЕА-трансгенных мышей подвергали схемам вакцинации ДНК/ДНК, Ad/Ad или ДНК/Ad с использованием векторов СЕА-DOM, СЕА-FcIgG или СЕА. Через две недели после последней иммунизации СЕА-трансгенных мышей стимулировали подкожной инъекцией 5×105 МС38-СЕА-клеток, сингенной линии опухолевых клеток, которая экспрессирует СЕА (Сlarke et al., supra). Иммунизация по схемам ДНК/ДНК или Ad/Ad не приводила к какому-либо значимому противоопухолевому действию, независимо от белка, экспрессируемого инъецированными векторами (фигура 24). В противоположность этому, иммунизация ДНК-EP/Ad векторами, кодирующими слитый белок СЕА/DOM, приводила к значимому противоопухолевому действию, причем 7 из 10 обработанных мышей оставались не имеющими опухоли в день 34 после стимуляции опухолевым антигеном. Таким образом, эти результаты показывают, что усиленная СЕА-специфическая иммунная реакция, связанная с кДНК СЕА-DOMopt, и схема вакцинации ДНК/Ad коррелируют со значимым противоопухолевым действием в СЕА-трансгенных мышах.
ПРИМЕР 19
Противоопухолевое действие СЕА-DOM, зависящее от CD4 + Т-клеток, CD8 + Т-клеток и НК-клеток
Были охарактеризованы эффекторные клетки, участвующие в противоопухолевом действии после электропорации ДНК (ДНК-EP) и иммунизации Ad векторами, кодирующими слитую конструкцию СЕА-DOM. После ДНК/Ad-иммунизации, но перед опухолевой стимуляцией, мышей истощали в отношении CD4+, CD8+ Т-клеток или НК-клеток с использованием MAb. Антитела вводили во время проведения опухолевой стимуляции для гарантии непрерывного истощения релевантных НК- и Т-клеточных субпопуляций. Истощение всех трех типов клеток подвергали мониторингу с использованием анализа проточной цитометрии с применением антител, специфических в отношении маркеров клеточной поверхности (данные не показаны). Истощение CD4+, CD8+ Т-клеток или НК-клеток оказывало отрицательное действие на выживание иммунизированных мышей, приводя к разительному уменьшению не имеющих опухолей мышей в сравнении с вакцинированной группой (фигура 25). Таким образом, эти результаты показывают, что НК-, CD4+ и CD8+ Т-клетки играют важную роль в противопухолевом действии, проявляемом вакцинацией СЕА-DOM.
ПРИМЕР 20
Статистический анализ
Как указано, результаты анализировали с использованием логарифмического рангового или двухстороннего t-критерия Стьюдента. Величину р<0,05 считали значимой.









































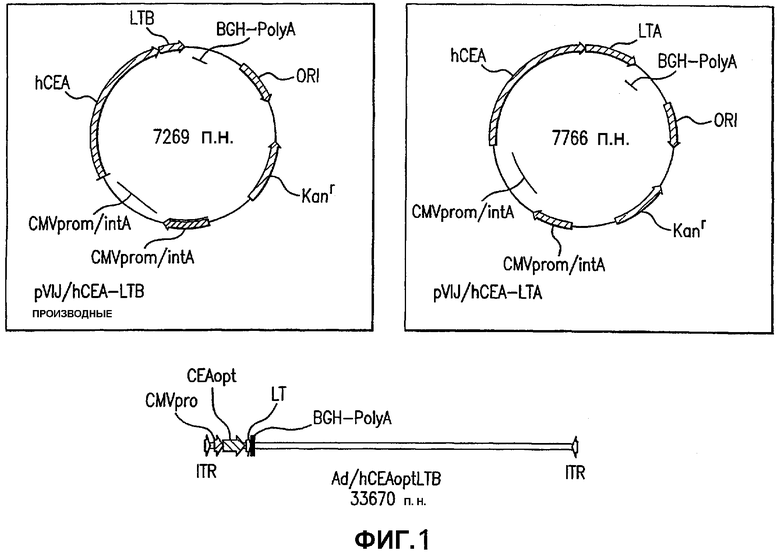










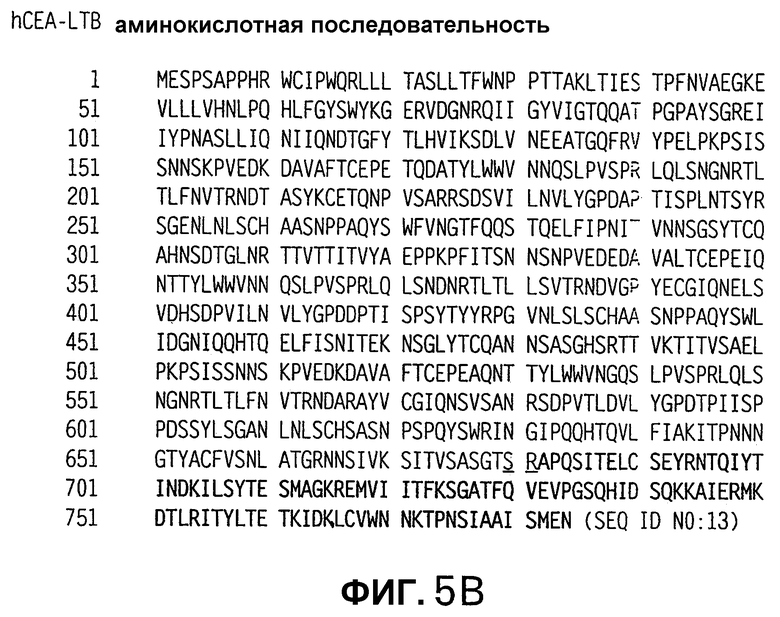











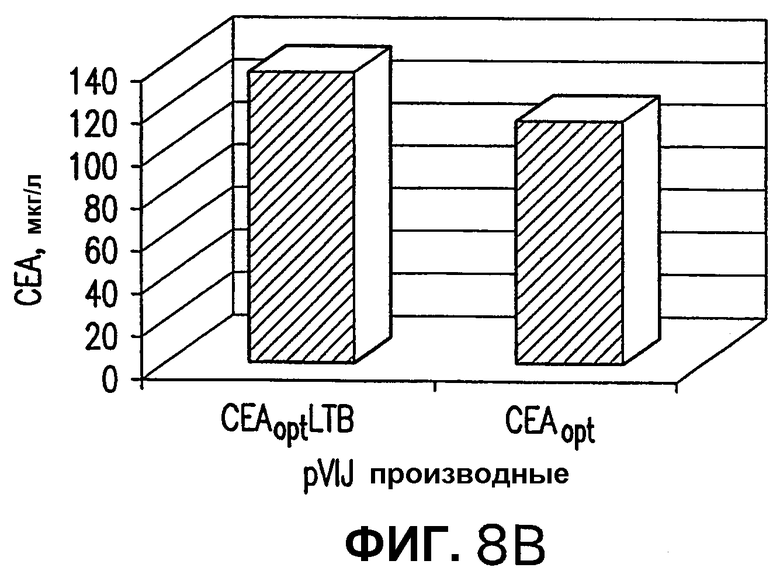
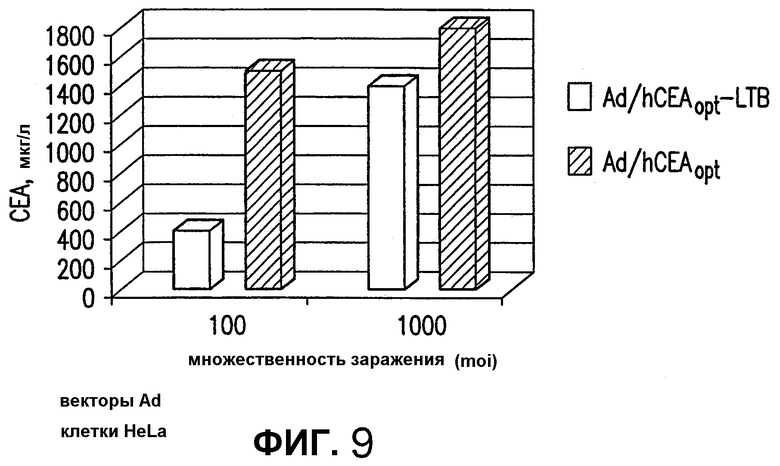

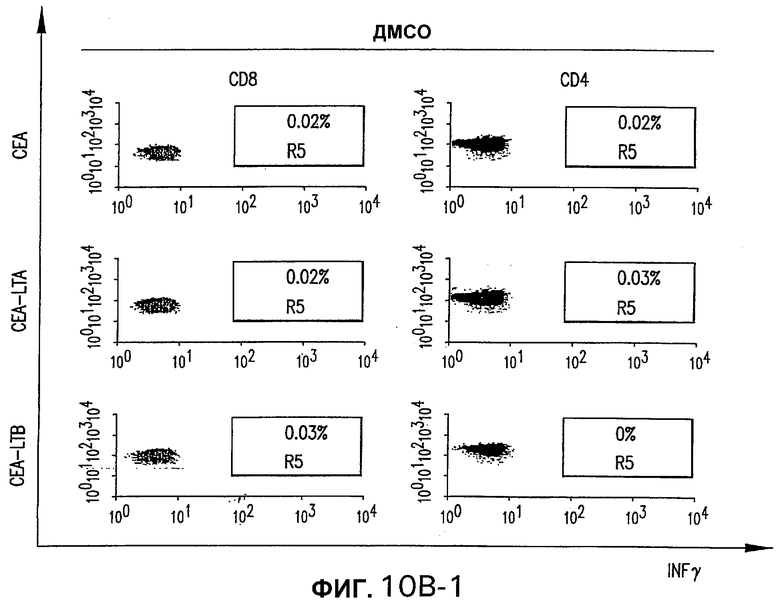
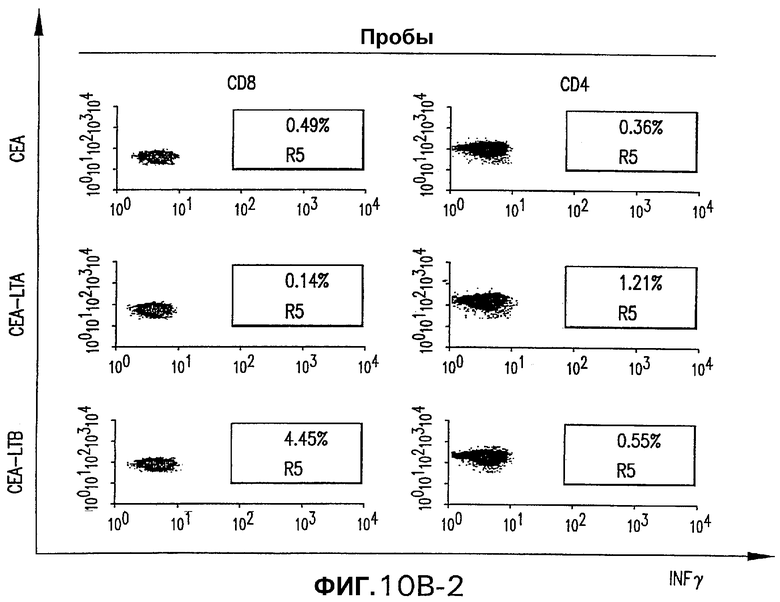
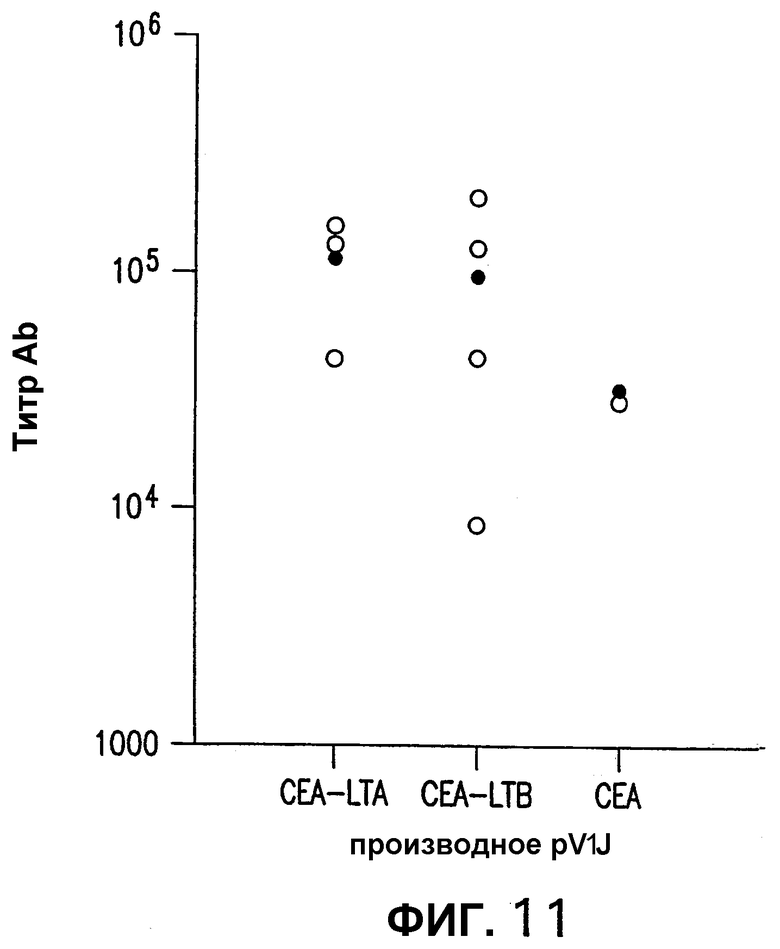
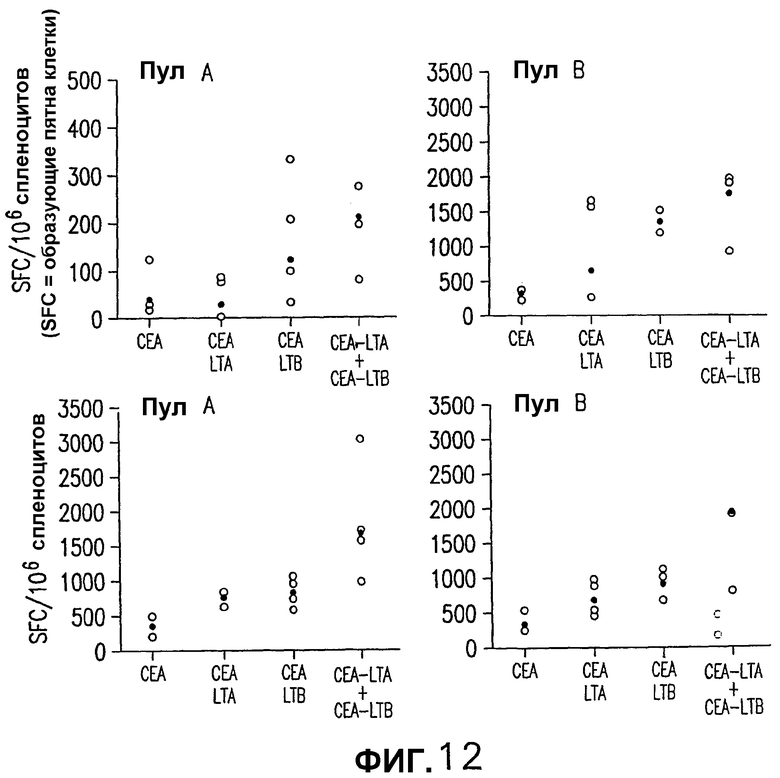
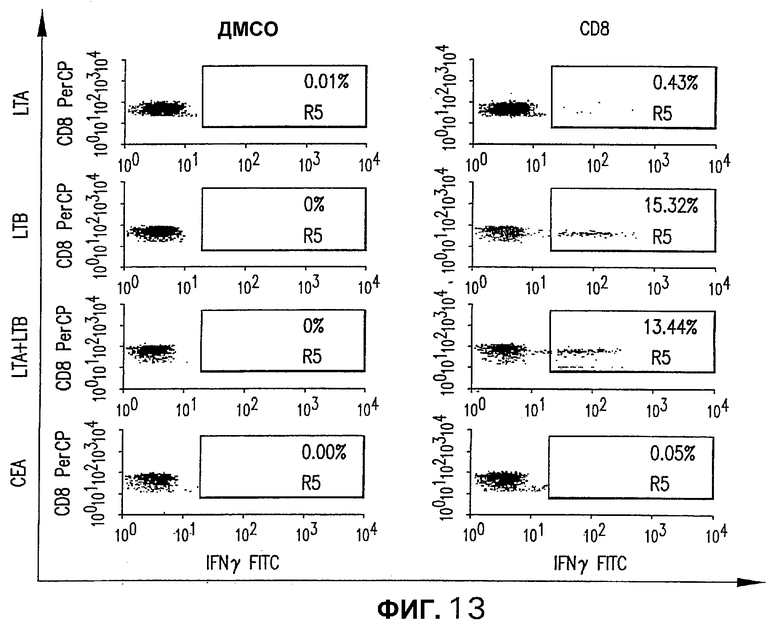
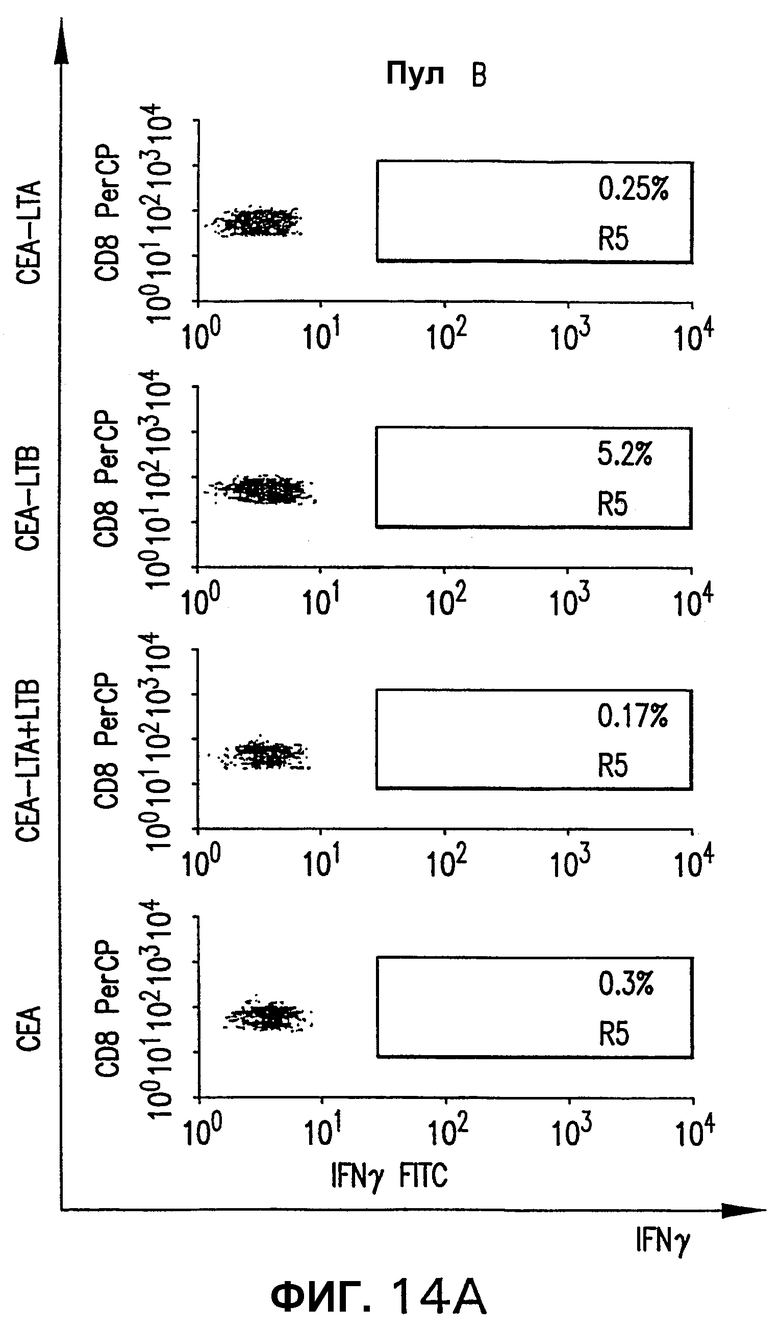
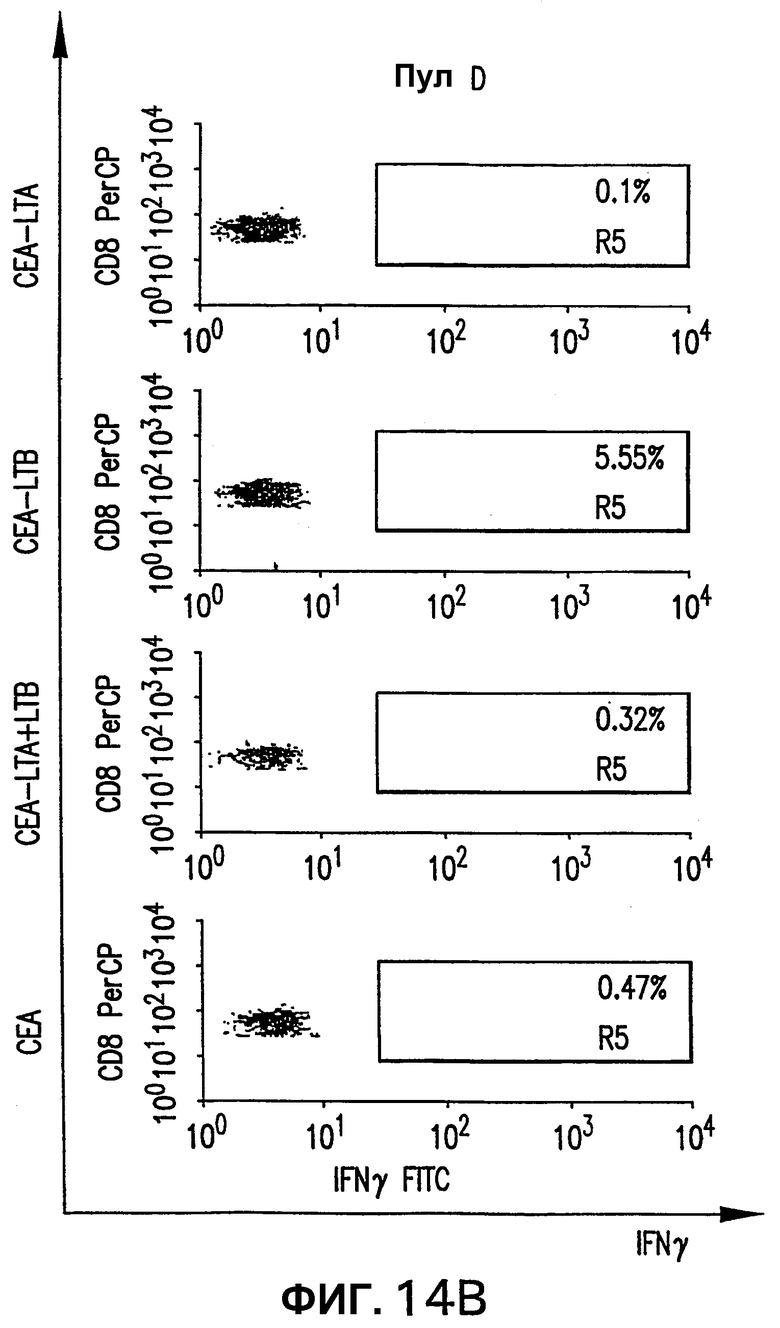
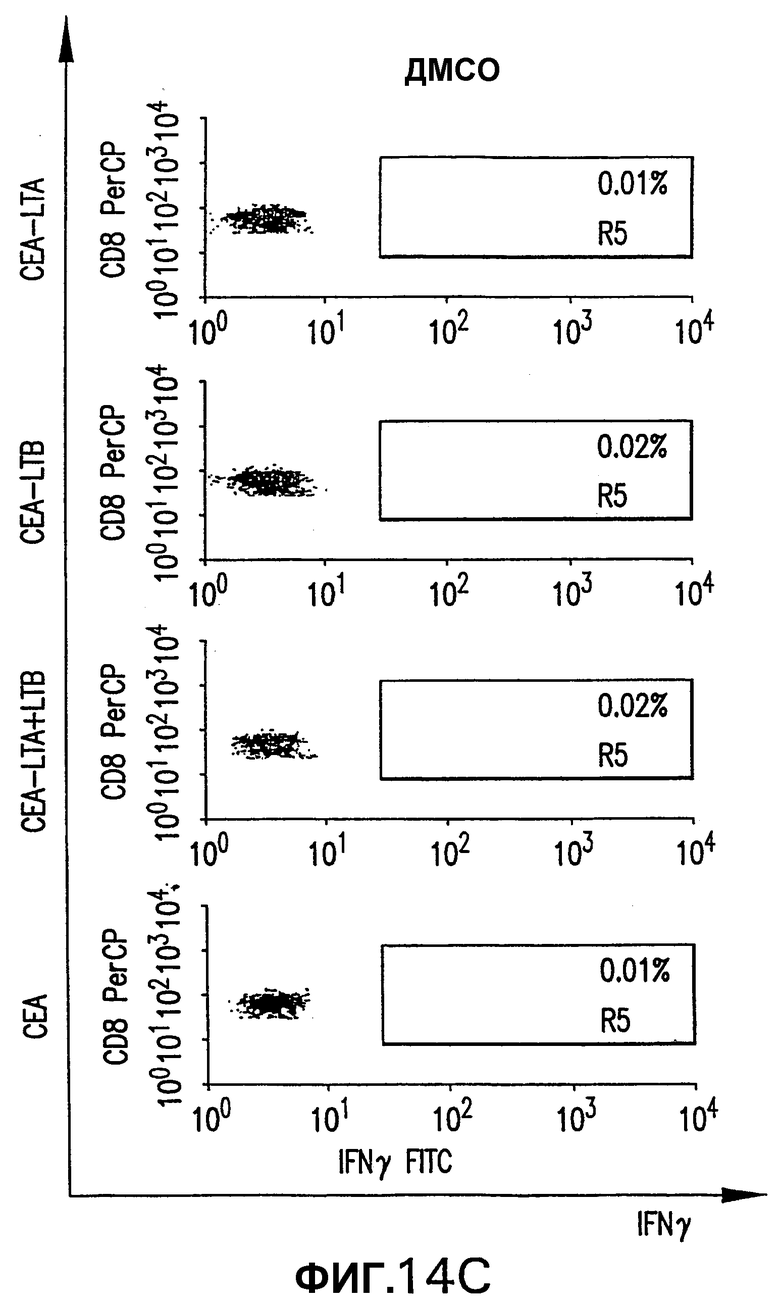
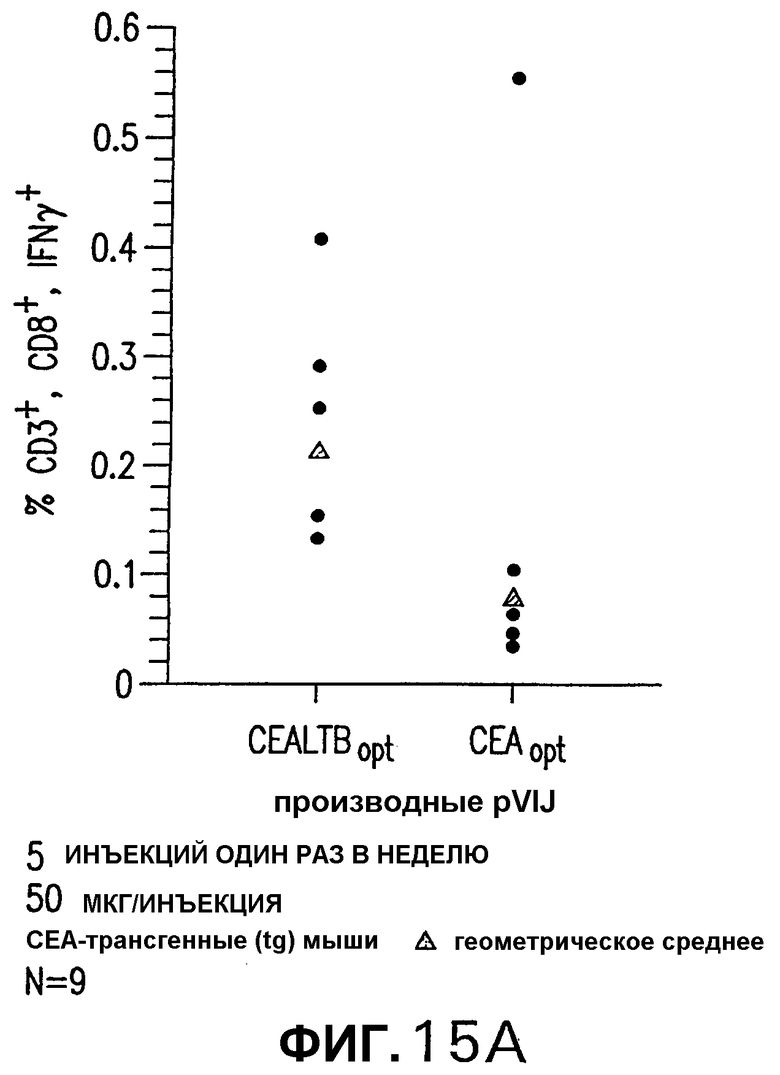

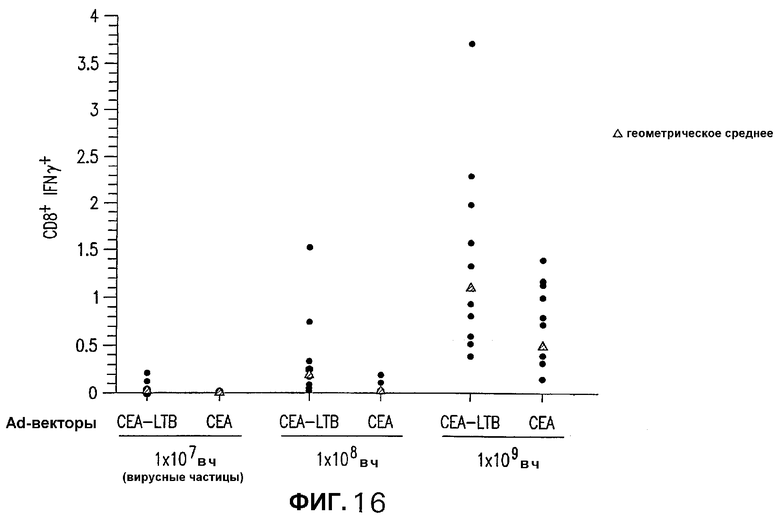
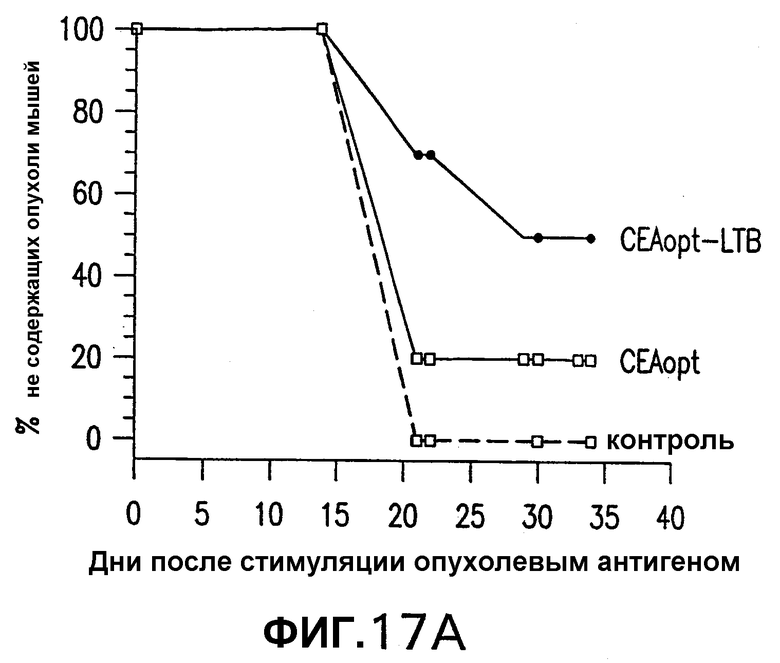
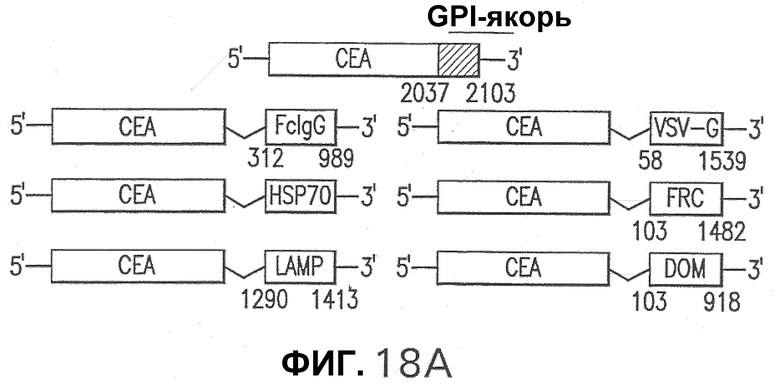
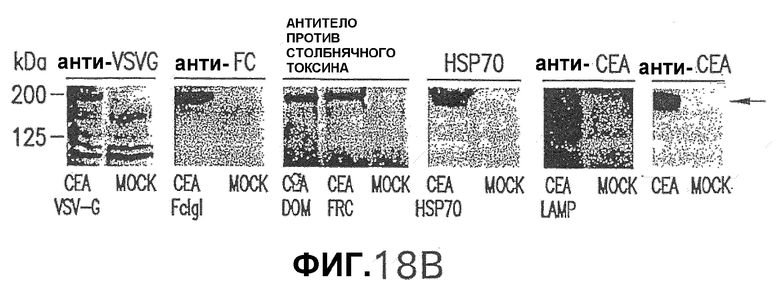
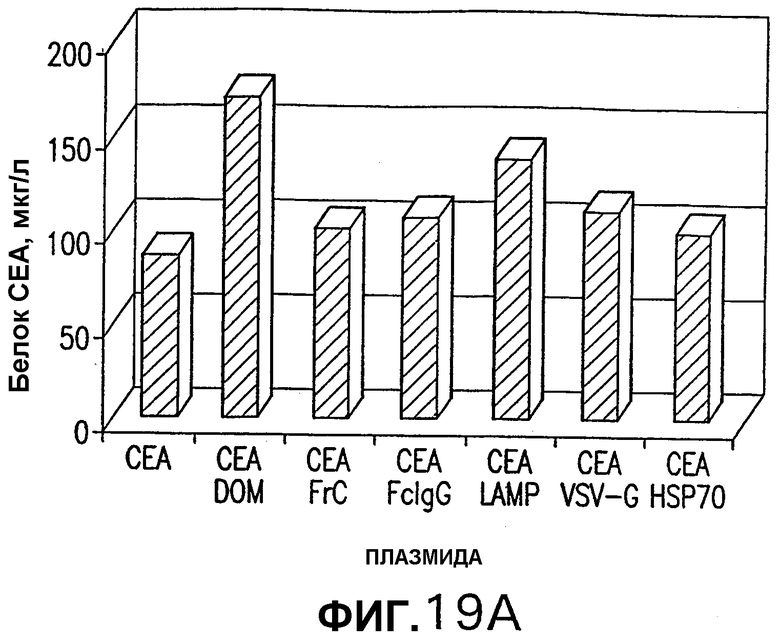

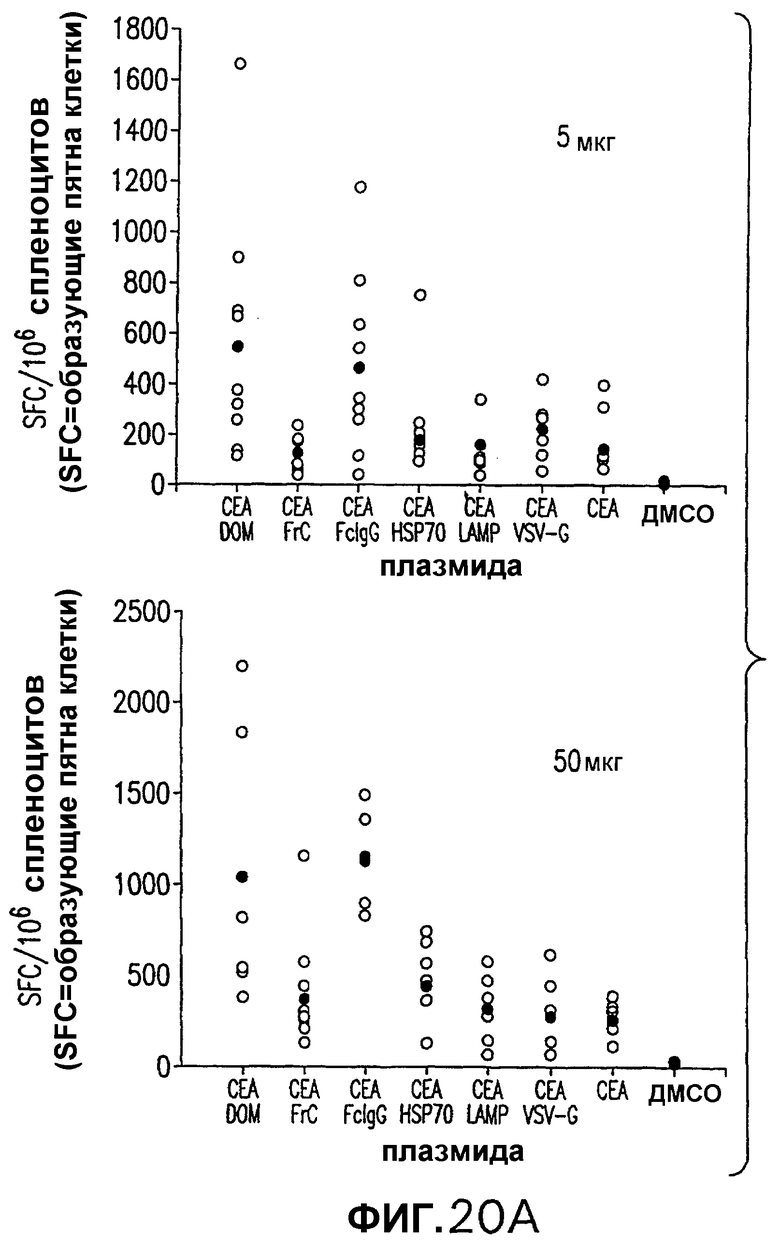

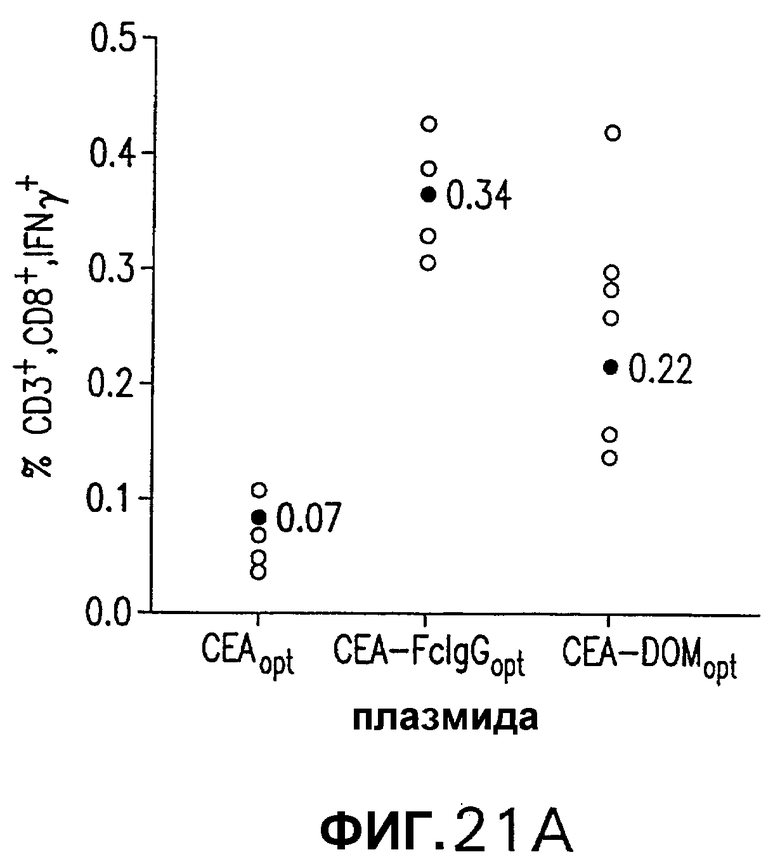
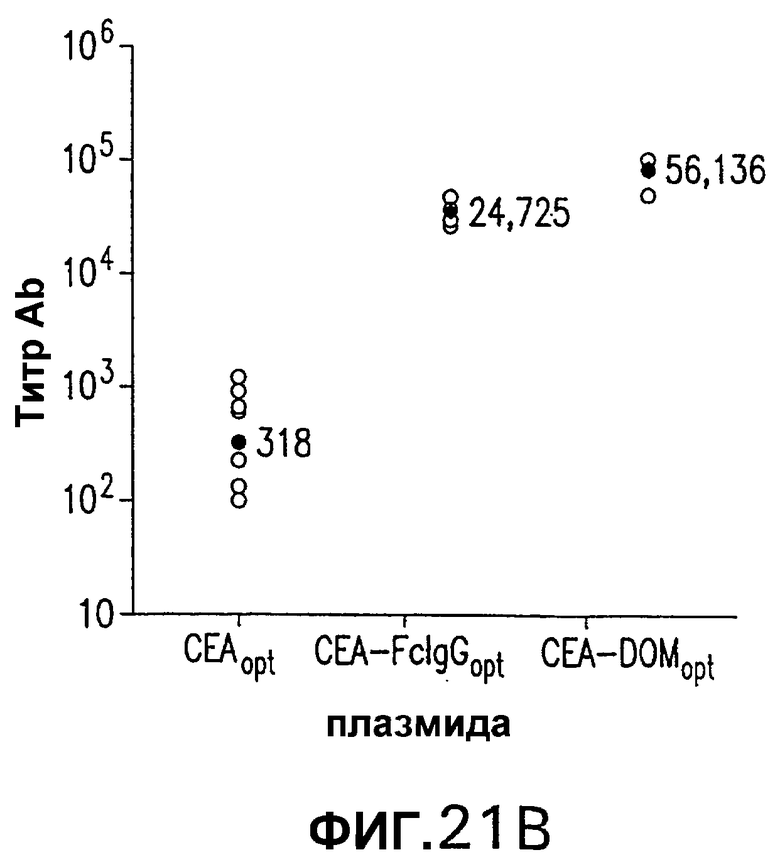

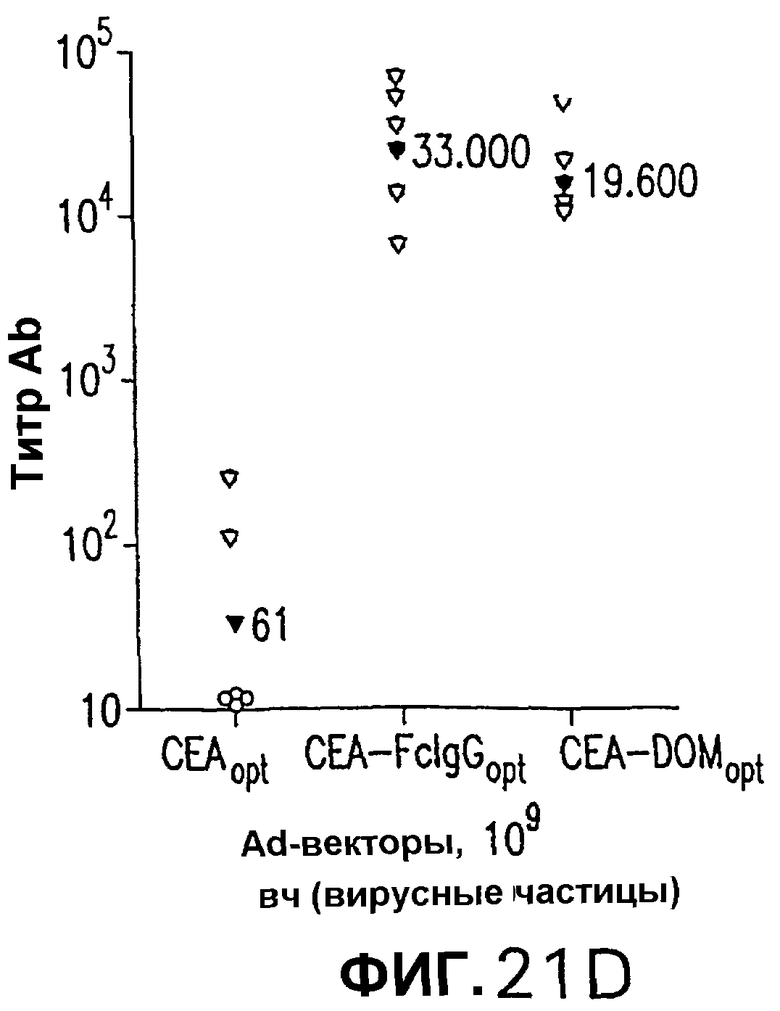
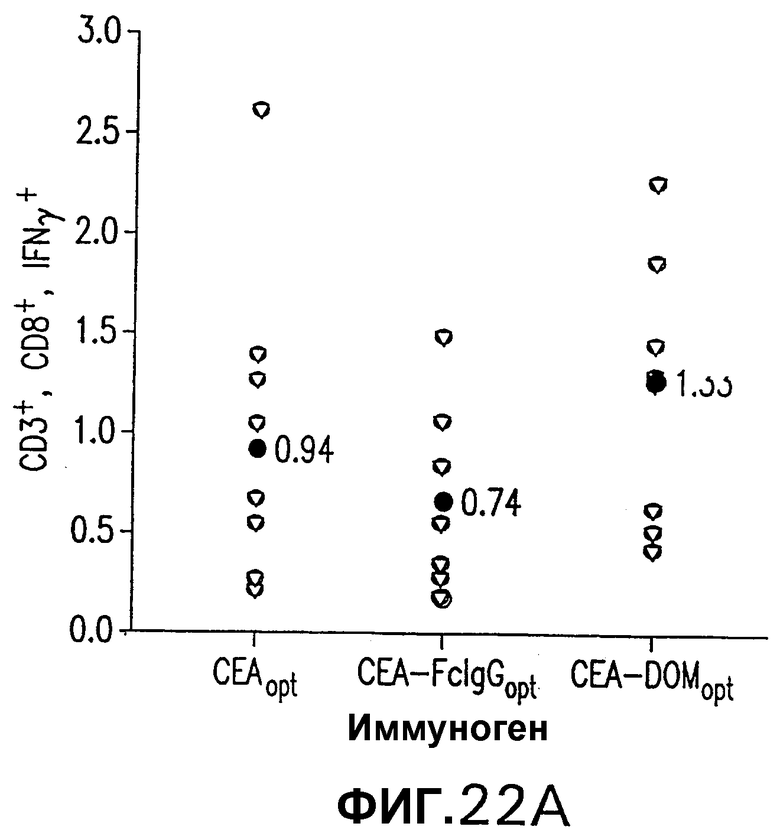
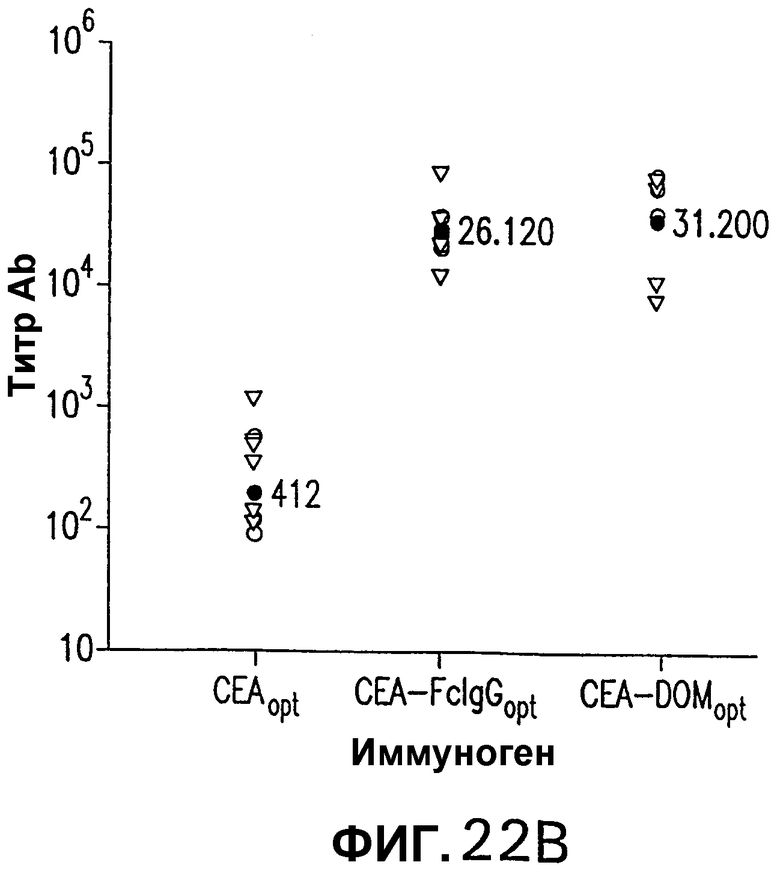
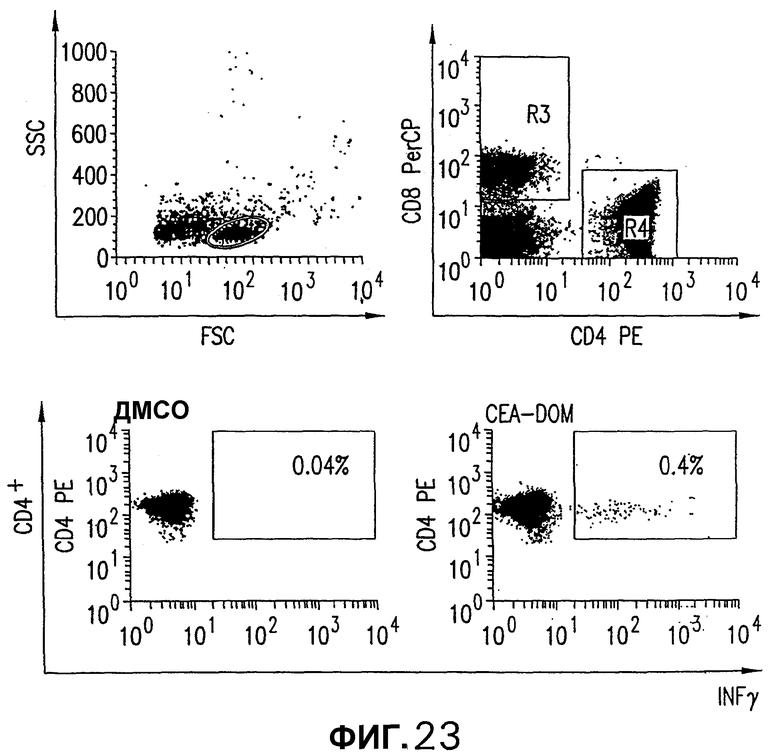
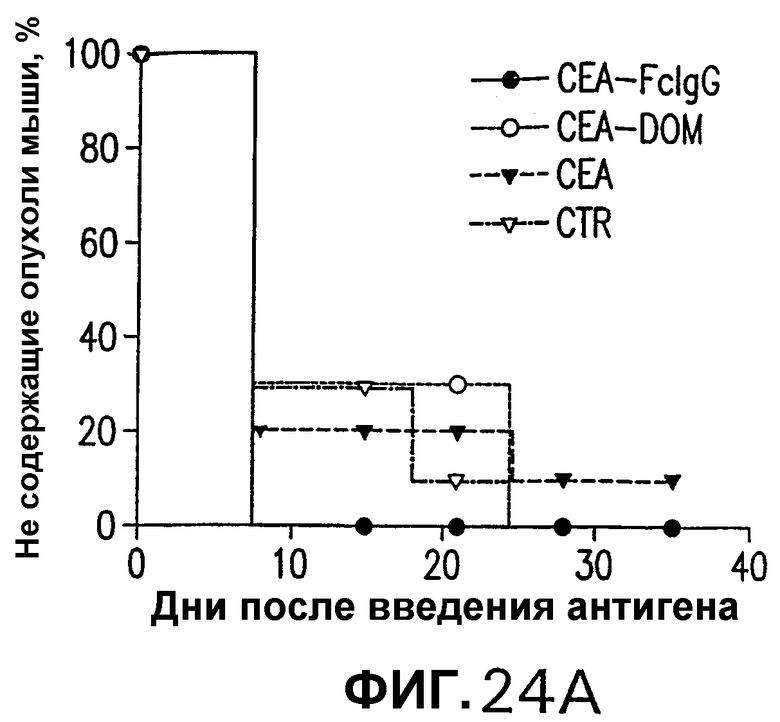
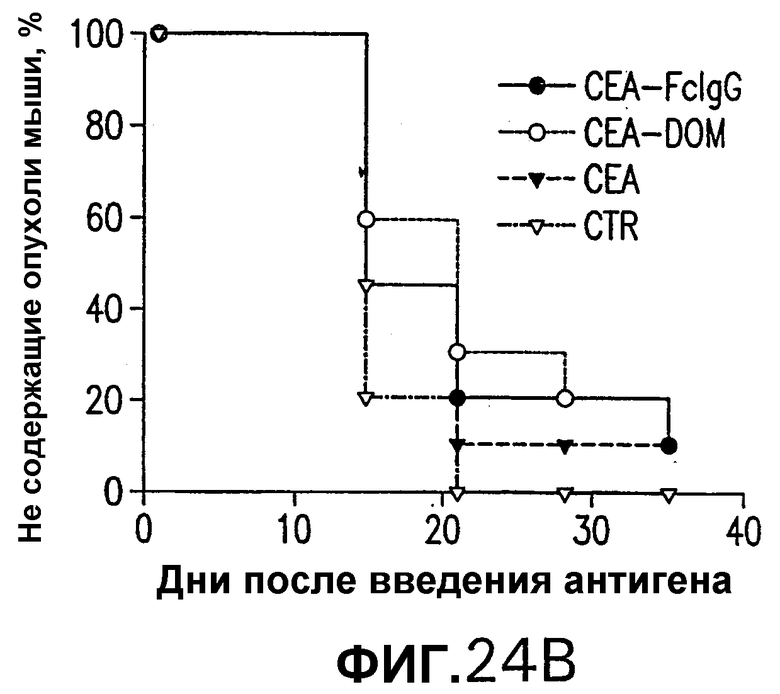
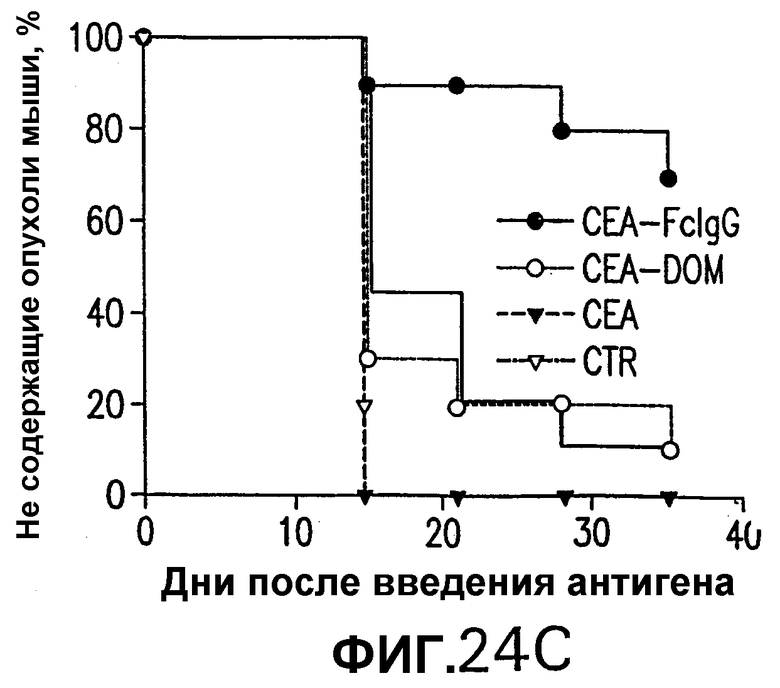
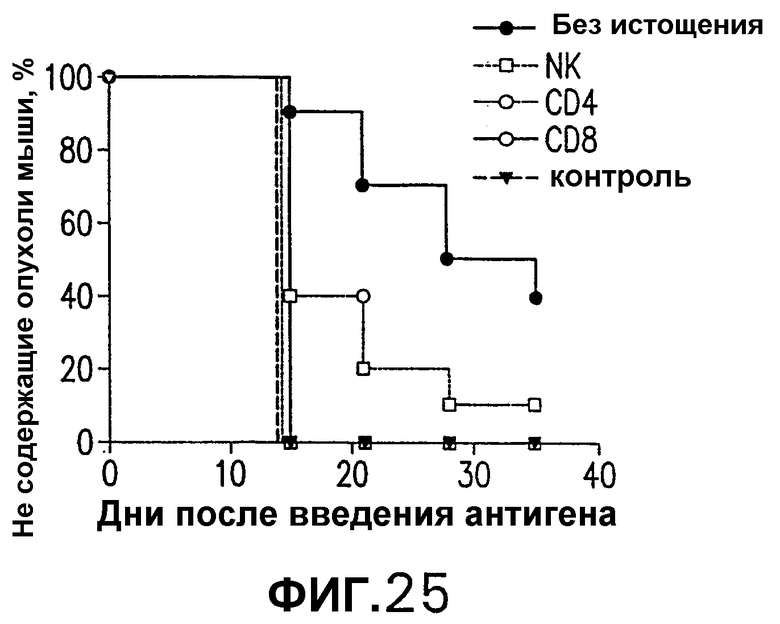

















Настоящее изобретение относится к области иммунологии и биотехнологии. Предложена молекула нуклеиновой кислоты, индуцирующая иммунный ответ к СЕА, содержащая нуклеотидную последовательность, которая кодирует слитый белок на основе карциноэмбрионального антигена (СЕА) или его функционального варианта, слитого с субъединицей В термолабильного энтеротоксина E.coli. Описаны ее варианты, а также соответствующий очищенный белок. Раскрыт вектор экспрессии, содержащий молекулу нуклеиновой кислоты, и клетка-хозяин, содержащая указанный вектор. Описаны аденовирусный вакцинный вектор для индукции иммунного ответа и вакцинная плазмида на основе указанной молекулы. Использование изобретения позволяет индуцировать иммунный ответ у млекопитающего, который является более сильным, чем ответ, индуцируемый СБА дикого типа, что может найти применение в медицине для лечения рака. 8 н. и 12 з.п. ф-лы, 32 ил.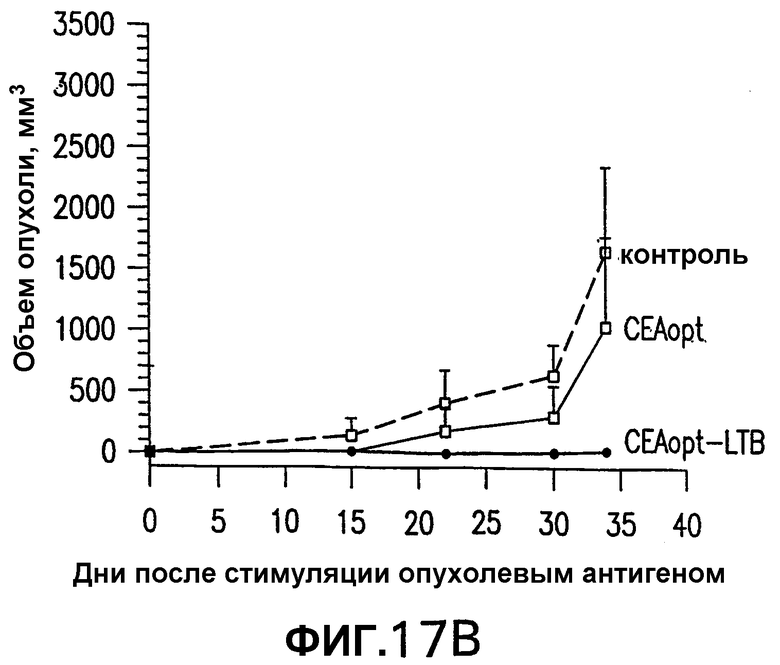
1. Молекула нуклеиновой кислоты, индуцирующая иммунный ответ против карциномоэмбрионального антигена (СEА), содержащая нуклеотидную последовательность, которая кодирует слитый белок СEА, где слитый белок СEА содержит белок СEА или его функциональный вариант, слитый с субъединицей В термолабильного энтеротоксина Е.coli (LTB) или функционального варианта; где указанный белок СEА содержит последовательность аминокислот, по существу, соответствующую SEQ ID NO: 20, и где слитый белок способен вызывать иммунный ответ у млекопитающего.
2. Молекула нуклеиновой кислоты по п.1, где белок СEА является укороченным на С-конце и С-концевое укорочение состоит из аминокислот 679-702 последовательности SEQ ID NO: 20.
3. Молекула нуклеиновой кислоты по п.1, где LTB укорочен на сигнальную последовательность.
4. Молекула нуклеиновой кислоты по п.1, где нуклеотидная последовательность содержит нуклеотидную последовательность SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 12.
5. Молекула нуклеиновой кислоты, индуцирующая иммунный ответ против СEА, содержащая последовательность нуклеотидов, которая кодирует слитый белок СEА, где нуклеотидная последовательность представлена в SEQ ID NO: 12.
6. Молекула нуклеиновой кислоты по п.3, где С-конец белка СEА слит с N-концом субъединицы B LT.
7. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п.1, которая функционально связана с промотором, индуцирующий иммунный ответ против СEА у млекопитающих.
8. Вектор экспрессии по п.7, где вектор является аденовирусным вектором или плазмидным вектором.
9. Вектор экспрессии по п.8, где вектором является вектор Ad5.
10. Вектор экспрессии по п.8, где вектором является вектор Ad6 или вектор Ad24.
11. Вектор экспрессии по п.8, где вектором является вектор Ad шимпанзе.
12. Вектор экспрессии по п.8, где вектором является pV1JnsB.
13. Клетка-хозяин, содержащая экспрессионный вектор по п.8, экспрессирующая слитый белок CEA-LTB.
14. Способ экспрессии слитого белка СEА в рекомбинантной клетке-хозяине, предусматривающий
(a) введение вектора, содержащего молекулу нуклеиновой кислоты по п.1, в подходящую клетку-хозяина; и
(b) культивирование клетки-хозяина при условиях, которые делают возможной экспрессию указанного слитого белка СEА человека.
15. Очищенный слитый белок СEА, кодируемый молекулой нуклеиновой кислоты по п.1, для индукции иммунного ответа против СEА.
16. Очищенный слитый белок СEА по п.15, где слитый белок содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 10, 13 и 15.
17. Аденовирусный вакцинный вектор для индукции иммунного ответа против СEА у млекопитающих, содержащий аденовирусный геном с делецией в районе Е1 и инсерцией в районе Е1, где инсерция содержит экспрессионную кассету, содержащую
(a) полинуклеотид, содержащий последовательность нуклеотидов, которая кодирует слитый белок СЕА, где слитый белок СEА содержит белок СEА или его вариант, слитый с белком LTB; и где слитый белок способен вызывать иммунный ответ у млекопитающего; и
(b) промотор, функционально связанный с полинуклеотидом.
18. Аденовирусный вектор по п.17, который является вектором Ad5.
19. Аденовирусный вектор по п.17, который является вектором Ad6 или вектором Ad24.
20. Вакцинная плазмида для индукции иммунного ответа против СЕА, содержащая плазмидную часть и часть экспрессионной кассеты, причем часть экспрессионной кассеты содержит
(a) полинуклеотид, содержащий последовательность нуклеотидов, которая кодирует слитый белок СЕА, где слитый белок СЕА содержит белок СЕА или его вариант, слитый с белком LTB; и где слитый белок способен вызывать иммунный ответ у млекопитающего; и
(b) промотор, функционально связанный с полинуклеотидом.
| Rice J et al | |||
| «DNA fusion vaccine designed to induce cytotoxic T cell responses against defined peptide motifs: implications for cancer vaccines», J Immunol | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| ПОЛИНУКЛЕОТИДНАЯ ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИНА | 1995 |
|
RU2186109C2 |

