Изобретение относится к новым вирусным частицам, содержащим вектор, происходящий из альфавируса, дефектный в отношении автономного размножения и, следовательно, в отношении репликации. Оно также относится к способу получения указанных частиц.
В следующем ниже описании изобретение более конкретно проиллюстрировано в отношении вируса леса Семлики (SFV), который относится к категории альфавирусов. Конечно, этот конкретный пример никоим образом не ограничивает объем данного изобретения, и могут рассматриваться все альфавирусы, например вирус Синдбис.
Геном альфавирусов существует в форме одноцепочечной РНК с положительной полярностью, содержащей две открытые рамки считывания, соответственно, первая рамка кодирует белки с ферментативной функцией, а вторая рамка кодирует структурные белки. Репликация происходит в цитоплазме клетки. На первом этапе инфекционного цикла на 5'-конце геномной РНК транслируется полипротеин (nsP 1-4) с РНК-полимеразной активностью, который продуцирует отрицательную цепь, комплементарную геномной РНК. На втором этапе отрицательная цепь используется в качестве матрицы для продуцирования двух РНК, соответственно:
- геномной положительной РНК, соответствующей геному вторичных вирусов, продуцирующей при трансляции другие nsp-белки и выполняющей роль генома этого вируса,
- субгеномной РНК, кодирующей структурные белки вируса, образующие инфекционные частицы.
Более конкретно, субгеномная РНК транскрибируется с промотора p26S, находящегося на 3'-конце последовательности, кодирующей белок nsp4. Соотношение геномная положительная РНК/субгеномная РНК регулируется автопротеолизом полипротеина до nsp1, nsp2, nsp3 и nsp4. На практике экспрессия вирусных генов происходит в две стадии. На первой стадии происходит основной синтез геномных положительных цепей и отрицательных цепей. На второй стадии происходит синтез фактически только субгеномной РНК, что, соответственно, приводит к продуцированию очень большого количества структурных белков.
Знание способа репликации альфавирусов и простота их генома привели к появлению систем переноса генов с использованием этих вирусов; последние позволяют получать сильную экспрессию трансгена в клетке-мишени.
Одно из необходимых условий, делающих возможным использование вектора, происходящего из альфавируса, в генной терапии, заключается в том, что он не должен обладать способностью к репликации. Было предложено несколько решений того, как сделать вирус Семлики дефектным по репликации.
Первое решение состоит в делеции структурных генов РНК вируса Семлики в пользу трансгена, который помещен под контроль промотора p26S. Такой вектор может быть перенесен в клетки в форме РНК или в форме ДНК. Однако это решение является не очень предпочтительным для применений in vivo, поскольку наблюдается низкая эффективность переноса этих генетических элементов, используемых в отсутствие частиц.
Другое решение состоит в инфицировании клеток-мишений вектором Семлики, но не в форме одной ДНК или РНК, а в форме рекомбинантных вирусных частиц. Для того чтобы сделать это, клеточную линию трансфицируют по меньшей мере двумя плазмидами, соответственно, плазмидой, несущей РНК вектора Семлики, лишенной структурных генов, и второй плазмидой, несущей структурные гены вируса Семлики под контролем промотора p26S. В такой клетке образуются вирусные частицы, которые инкапсидируют только дефектную РНК, т.е. РНК Семлики, несущую трансген, поскольку только эта РНК также несет последовательность для инкапсидации, содержащуюся в последовательности nsp2. Хотя теоретически считается, что в ходе этого процесса не образуются никакие репликативные частицы, рекомбинационные события происходят часто, в частности, из-за перекрывания комплементирующих последовательностей и последовательностей рекомбинантного вируса и из-за избытка вирусных РНК в цитоплазме продуцирующих клеток.
В документах (1, 2) Rolls описывает вектор SFV, геном которого был модифицирован путем замещения структурных генов геном, кодирующим VSV-G оболочку, возможно в комбинации с трансгеном. Поэтому инфекционные частицы, полученные таким образом, состоят из VSV-G оболочки и содержат вектор, происходящий из альфавируса. Однако из-за своей способности к автономной репликации описанная система является чрезвычайно опасной. Эквивалентная система описана в документе WO 03/072771.
В документе (3) Lebedeva с соавт. описывает совместную электропорацию клеток BKH:
- вектором, обеспечивающим репликативные функции (Srepβgal: ген, кодирующий репликазу вектора Семлики (SFV) и β-галактозидазу в качестве трансгена),
- вектором, кодирующим структурные гены: ScapSenv, кодирующие капсид и оболочку SFV в качестве контроля, или производным вектором, в котором ген env SFV замещен последовательностями env ретровируса MLV (мышиного лейкоза Молони),
и анализ вирусных частиц, продуцируемых таким образом. Однако в этой системе мобилизация вектора SFV, переносящего трансген, в этом примере ген β-галактозидазы, находится под контролем инкапсидации этой рекомбинантной геномной РНК капсидным белком SFV.
Другими словами, задача, решение которой предположено в данном изобретении, состоит в улучшении способа мобилизации векторов, происходящих из альфавируса, в частности вируса леса Семлики (SFV), с тем, чтобы предотвратить любой риск рекомбинации внутри линий-продуцентов, которые могут продуцировать частицы, способные к репликации.
Другая задача, решение которой предложено в данном изобретении, состоит в получении вирусных частиц, содержащих вектор, происходящий из альфавируса, тропизм которых не ограничивается клетками-мишенями вирусов дикого типа.
Автору заявки удалось получить вирусные частицы, которые удовлетворяют одновременно двум вышеуказанным задачам, путем экспрессии in trans, в клеточной линии, генов, кодирующих структурные элементы, не происходящие из альфавируса, и вектора, происходящего из альфавируса, дефектного по репликации.
Согласно первому воплощению гены, кодирующие структурные элементы, не происходящие из альфавируса, соответствуют только гену ENV вируса везикулярного стоматита, кодирующему белок оболочки VSV-G.
Использование белка оболочки VSV-G имеет несколько преимуществ. Прежде всего белок оболочки вируса везикулярного стоматита допускает способ введения в клетку путем эндоцитоза, который может быть совмещен со способом введения альфавирусов. Кроме того, VSV-G является очень стабильным белком, который может быть сконцентрирован путем ультрацентрифугирования, и это позволяет рассматривать вопрос о парентеральном введении. Более того, этот белок чрезвычайно расширяет тропизм частиц, которые его содержат, таким образом расширяя область применения вирусных частиц по изобретению в таких разных организмах, как дрозофила и млекопитающие.
Согласно этому первому воплощению экспрессию in trans получают путем котрансфекции, предпочтительно осуществляемой в две отдельные стадии, соответственно, трансфекции клеточной линии плазмидой, экспрессирующей ген оболочки VSV-G, а затем второй трансфекции вектором, происходящим из альфавируса. На практике данную котрансфекцию осуществляют на клетках 293Т.
Согласно второму воплощению гены, кодирующие структурные элементы, не происходящие из альфавируса, соответствуют генам, кодирующим структурные элементы ретровируса.
В этом случае экспрессию in trans получают путем трансфекции упаковывающей клеточной линии, которая продуцируют дефектные по репликации ретровирусы вектором, происходящим из альфавируса. Такой тип вектора, например система Phoenix®, хорошо известен специалистам в данной области (http://www.stanford.edu/group/nolan/retroviral systems/phx.html). В частности, могут быть использованы упаковывающие линии, в которых используются структурные гены MLV (вируса мышиного лейкоза).
Эти линии получены известным способом путем стабильной трансфекции первой плазмидой, экспрессирующей гены GAG-POL, и второй плазмидой, экспессирующей ген ENV ретровируса или другого вируса, имеющего оболочку (4).
Однако можно также рассматривать получение вирусных частиц путем тройной трансфекции клеточной линии, например клеток 293Т, путем введения первого вирусного элемента, экспрессирующего ретровирусные гены GAG и POL, второго вирусного элемента, экспрессирующего ретровирусный ген ENV, и вектора, происходящего из альфавируса.
Можно также дополнительно усилить дефектный характер транскомплементирующих ретровирусных последовательностей путем мутации, в частности, делеции нуклеотидных последовательностей гена POL, кодирующего интегразу (IN) и обратную транскриптазу (RT).
В двух вышеописанных воплощениях изобретения вектор, происходящий из альфавируса, делают дефектным по репликации. На практике это свойство получают путем делеции структурных генов или их замещения в пользу интересующего(их) трансгена(ов) в геноме вектора.
Согласно другой характеристике геном вектора, происходящего из альфавируса, содержит сигнал инкапсидации вирусной частицей, называемый psi-последовательностью.
Согласно первому воплощению psi-последовательность соответствует протяженной последовательности для упаковки из векторов MLV, полученной путем амплификации вектора PLNCX (Clontech®) методом ПЦР (полимеразной цепной реакции) с использованием праймеров:
- 5'-праймера: LNCX Psi 2a: 5'-GGGACCACCGACCCACCACC-3'
и
- 3'-праймера: LNCX Psi 2b: 5'-GATCCTCATCCTGTCTCTTG-3'.
Предпочтительно, psi-последовательность является небольшой по размеру и соответствует минимальной последовательности. Эта модификация является предпочтительной до тех пор, пока psi-последовательность может функционировать в качестве сайта внутренней посадки рибосом (IRES). IRES-функция, таким образом, дает возможность элиминировать промотор p26S SVF, так что трансляцию трансгена осуществляют на геномной РНК.
Парадоксально, но автор заявки также показал, что присутствие ретровирусного сигнала инкапсидации не является абсолютно необходимым. Действительно, количество рекомбинантных РНК вектора Семлики, обнаруженное в цитоплазме трансфицированных клеток, является таким, что указанные РНК предпочтительно инкапсидируются в ретровирусные частицы. Этот феномен подчеркивается подавлением клеточных генов, индуцированным в результате экспрессии неструктурных белков вируса Семлики. Важную роль могла бы играть также субклеточная локализация репликативных комплексов вируса SFV (5). Следовательно, в предпочтительном воплощении геном данного вектора лишен psi-последовательности.
Автор изобретения также показал, что описанный выше вектор, содержащий ретровирусную последовательность для инкапсидации, можно мобилизовать при помощи продуцируемых ретровирусных частиц, используя систему транскомплементации на основе векторов, происходящих из вируса леса Семлики (11). Показано, что в этой системе могут быть получены титры порядка 106 частиц на миллилитр. Также показано, что присутствие ретровирусного сигнала для инкапсидации улучшает титр частиц приблизительно на один порядок. Эти частицы эффективно используются для трансдукции клеток, экспрессирующих амфотропный вирусный рецептор (Pit 2), соответствующий оболочке используемых ретровирусов. Это наблюдение имеет прямое отношение к биологической безопасности ретровирусных частиц, продуцируемых способом, предложенным Li и Garoff (11). В данном контексте показано, что частицы, продуцируемые согласно способу, предложенному Li и Garoff, содержат в титре, близком к 106 частиц/мл, геномы рекомбинантных векторов SFV, экспрессирующих ретровирусные последовательности GAG/POL или ENV.
Более того, автор заявки заметил, что способ трансфекции, обычно используемый для рекомбинантных РНК векторов Семлики, а именно электропорация, приводит к значительному повреждению клеток. Таким образом, для того чтобы допустить осуществление трансфекции клеток-продуцентов способами, которые являются более мягкими, чем электропорация, вектор, происходящий из альфавируса, был модифицирован таким образом, чтобы его экспрессия осуществлялась с эукариотического промотора, например CMV-промотора, расположенного на 5'-конце векторной последовательности.
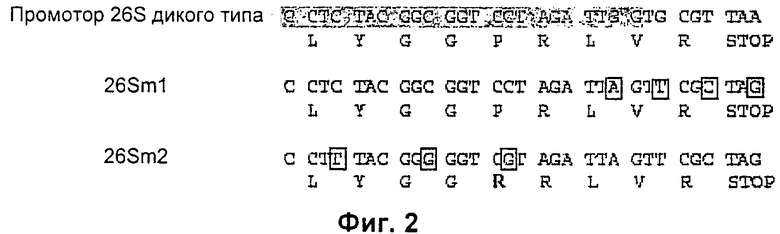
Наконец, предпочтительно, чтобы промотор p26S альфавирусного вектора был мутантным. Вектор SFV 26Sm2, и в меньшей степени вектор SRV 26Sm1, больше не экспрессируют детектируемую субгеномную РНК, которая в результате конкуренции может уменьшать инкапсидацию геномной РНК.
Таким образом, частица по данному изобретению соответствует вирусной частице, состоящей из структурных элементов, не происходящих из альфавируса, и содержащей вектор, происходящий из альфавируса, дефектный по репликации в результате делеции или замещения структурных генов по меньшей мере одним трансгеном, при этом структурные элементы указанной частицы не кодируются геномом вектора, происходящего из альфавируса.
Кроме того, данное изобретение относится к применению вирусных частиц по изобретению для инфицирования клеток in vitro. Автор заявки показал, что частицы, полученные таким образом, могут инфицировать широкий круг эукариотических клеток как человека, так и клеток ,не происходящих из человека.
Данное изобретение также относится к фармацевтической композиции, содержащей вирусные частицы по изобретению.
Аналогично, оно относится к применению данных вирусных частиц для изготовления лекарственного препарата для применения в лечении рака.
Данное изобретение и преимущества, которые из него следуют, станут понятными из следующих примеров.
На Фиг.1 дано схематичное представление структуры вектора, происходящего от вируса леса Семлики (SFV).
На Фиг.2 показаны мутации, оказывающие влияние на промотор p26S. Мутации, введенные в мутанты 26Sm1 и 26Sm2 относительно последовательности дикого типа (Wt), заключены в рамку. Аминокислота, выделенная жирным шрифтом, указывает на изменение в кодирующей последовательности.
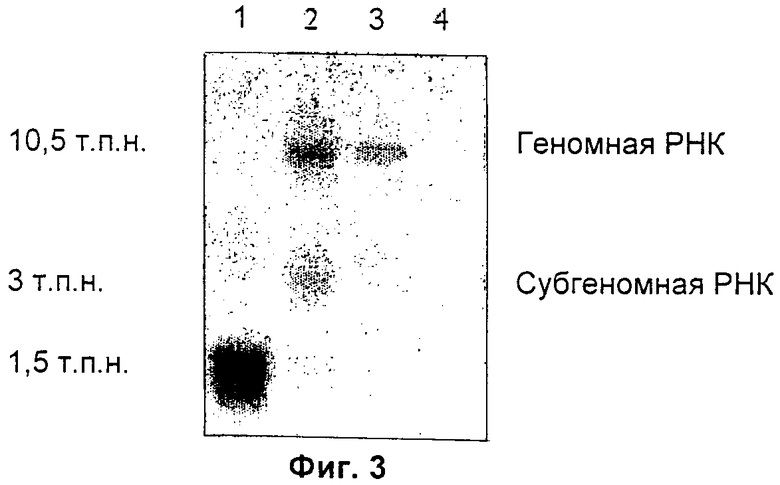
На Фиг.3 приведен результат Нозерн-блоттинга, осуществленного с использованием клеток-продуцентов, экспрессирующих модифицированные векторы SFV (1: pEGFPC1, 2: 26Sm1, 3: 26Sm2, 4: SFV без трансгена), с GFP-зондом из pEGFPC1.
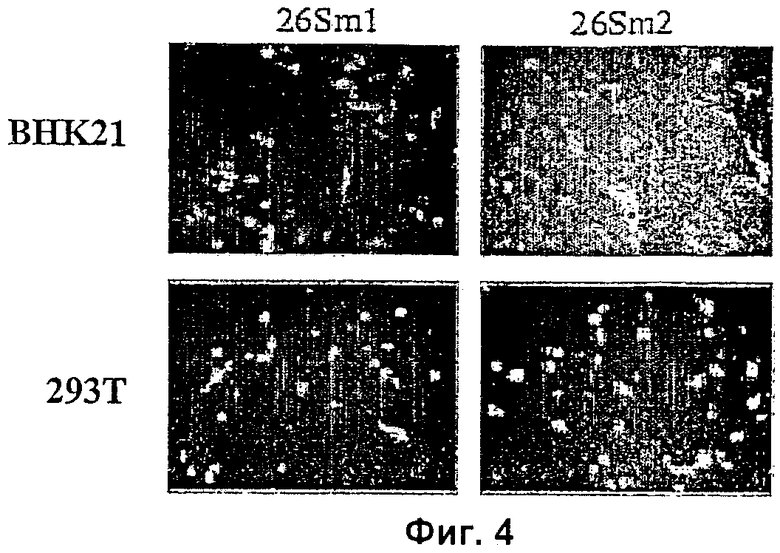
На Фиг.4 показана способность клеток 293Т и BHK 21 экспрессировать производные векторы SFV (26Sm1 и 26Sm2), мобилизованные псевдочастицами VSV-G.
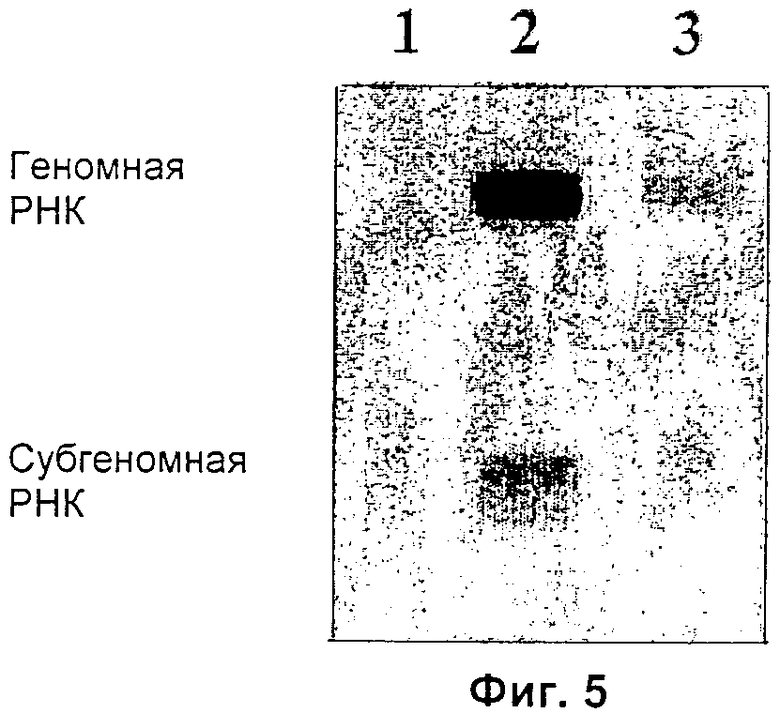
На Фиг.5 приведен результат Нозерн-блоттинга, осуществленного с использованием клеток, инфицированных супернатантом от клеток 293Т, трансфицированных плазмидой pMDG и модифицированными векторами SFV (1: pEGFPC1, 2: 26Sm1, 3: 26Sm2), с GFP-зондом из pEGFPC1.
ПРИМЕР 1: Продуцирование вирусных частиц клеточными линиями, экспрессирующими VSV-G оболочку
1/ МЕТОДЫ
1/ Клеточные линии и культуры
- 293Т/17: первичная клеточная линия почки эмбриона человека (ATCC CRL-11268),
- Hela: клеточная линия человека (АТСС CCL-2),
- QM7: клеточная линия мышц перепела (АТСС, CRL-1962),
- LMH: клеточная линия печени курицы (АТСС CRL-2117).
Вышеуказанные четыре клеточные линии культивировали в DMEM (среде Игла, модифицированной по способу Дульбекко) (Invitrogen), содержащей 10% эмбриональной телячьей сыворотки (FCS) (Biowest).
- HepG2: клеточная линия гепатомы человека, культивируемая в среде ЕМ, содержащей 10% FCS (АТСС НВ-8065),
- BHK21: клеточная линия почки сирийского хомячка, культивируемая в среде GMEM, содержащей 5% FCS и 8% жидкого триптозофосфатного раствора (АТСС CCL-10),
- CESC: куриные эмбрионы, полученные и культивируемые согласно ссылке 26,
- High Five клетки, культивируемые при 27°С в среде Грейса для клеток насекомых (№ по каталогу В85502 Invitrogen), содержащей 10% FCS,
- Sp2/0: линия лимфобластоидных клеток мыши, культивируемая в среде RPMI 1640, содержащей 10% FCS (АТСС CRL1581).
2/ Конструирование вектора SFV
Структура вектора SFV представлена на Фиг.1.
а/ Вектор 26Sm1
Внутренний 26S промотор SFV подвергают мутации с помощью ПЦР, используя вектор pSFV1 (Invitrogen), который лишен структурных генов и использован в качестве матрицы, в присутствии двух праймеров, соответственно:
- праймера 26Sm1F, содержащего сайт рестрикции BglI, который выделен жирным шрифтом, имеющего следующую последовательность:
5'-ATCCTCGAAGATCTAGGG-3',
- второго, мутантного, праймера 26Sm1R, содержащего сайт рестрикции
ClaI, который выделен жирным шрифтом, имеющего следующую последовательность:
5'-CAATATCGATTACTAGCGAACTAATCTAGGA-3'.
Затем молчащие мутации вводят в промотор p26S, чтобы получить промотор p26Sm1, представленный на Фиг.2. Амплифицированный таким образам продукт затем клонируют в плазмиду pIRES2-EGFP (Invitrogen) (Фиг.1). Ретровирусную последовательность, обозначенную как RS, происходящую из вируса MLV, затем встраивают между мутантным 26S промотором и IRES-последовательностью. Затем фрагменты, содержащие мутантную 26S последовательность, ретровирусную MLV последовательность и ген EGFP, вырезают с помощью BglII и HpaI и затем клонируют в вектор pSFV1 между рестрикционными сайтами BglII и SmaI. Фрагмент размером 10,5 т.п.н., содержащий этот модифицированный репликон SFV, окончательно клонируют между IE промотором CMV и сигналом полиаденилирования рА из SV40 в векторе pIRES2-EGFP, в котором последовательность IRES GFP была делетирована.
б/ Вектор SFV26Sm2
Внутренний промотор подвергают мутации с помощью ПЦР, используя плазмиду SFV1, которую используют в качестве матрицы, в присутствии двух праймеров, соответственно, первого праймера 26Sm1F и второго праймера 26Sm2R, содержащего рестрикционный сайт, который выделен жирным шрифтом, имеющего следующую последовательность:
5'-ATATCGATTACTAGCGAACTAATCTACGACCCCCGTAAAGGTGT-3'.
Праймер 26Sm2R осуществляет модификации промотора p26S, показанные на Фиг.2. Затем продукт амплификации переваривают с помощью BglII и ClaI и лигируют в вектор 26Sm1, также переваренный BglII и ClaI, чтобы делетировать соответствующий фрагмент.
3/ Трансфекция клеточной линии 293Т векторами SFV 26Sm1 и 26Sm2 и плазмидой pMDG и сбор вирусных частиц
Временную трансфекцию клеток 293Т осуществляют с помощью набора для кальций/фосфатной трансфекции (Invitrogen). Клетки 293Т в количестве 8×105 клеток на лунку засевают в 6-луночные планшеты и инкубируют перед трансфекцией в течение ночи при 37°С. Трансфекцию осуществляют в две стадии. В первый день клетки 293Т трансфицируют 5 мкг плазмиды pMDG, содержащей ген, кодирующий VSV-G оболочку, под контролем IE-промотора CMV (6). На второй стадии, во второй день, эти клетки трансфицируют 5 мкг векторов SFV 26Sm1 или 26Sm2. Вторая среда для трансфекции остается в контакте с клетками в течение 13-17 часов. На третий день эту среду удаляют и заменяют свежей средой, что позволяет инфицирующим частицам высвобождаться. Культуральную среду, содержащую вирусные частицы, собирают через 5-6 часов после этого.
4/ Трансфекция клеточной линии BHK21 векторами SFV 26Sm1 или 26Sm2 и векторами SFV GAGPOL и SFV ENV и сбор вирусных частиц
Клетки BHK21 подвергают электропорации при концентрации 5×106/мл (т.е. 4×106 клеток) при напряжении 350 В и электрической емкости 750 мкФ. Используемые для данной электропорации РНК, соответствующие различным векторам (26Sm1 или m2, SFV GAGPOL и SFV ENV), транскрибируют, используя 1,5 мкг линеаризованной ДНК, с помощью полимеразного набора SP6 (Invitrogen). Для электропорации используют 22 мкл транскрипционного продукта. Рекомбинантные частицы собирают через 14-16 часов. Супернатанты фильтруют и добавляют к клеткам-мишеням в присутствии 2 мкг/мл полибрена.
5/Инфицирование клеточных линий вирусными частицами
Супернатант от трансфицированных линий клеток 293Т собирают и затем фильтруют через 0,45 мкм фильтр (НА Millex®, Millipore), а затем инкубируют с различными клеточными линиями в присутствии свежей среды, содержащей полибрен, используемый в концентрации 5 мкг на мл (Sigma). Экспрессию GFP в инфицированных клетках подтверждают с помощью флуресцентного микроскопа Olimpus 1×50. Трансфекцию количественно характеризуют с помощью проточного цитометра Becton Diskinson FACScalibur®. В качестве контрольных тестов используют супернатанты в различных реагентах:
- 10 мкг на мл РНКазы А (Sigma),
- 1 мкг на мл актиномицина D (Sigma),
- 100 единиц на мл ДНКазы I (invitrogen),
- 1 мг на мл генетицина (Sigma) и
- 3 мкг на мл пуромицина (Cayla).
6/ Концентрирование вирусных частиц
Супернатант от трансфицированных клеток 293 центрифугируют при 150000 g в роторе SW41 в течение 1 часа при 4°С. Концентрированные вирусы ресуспендируют в 300 мкл PBS и 25 мкл данного раствора используют для инфицирования 5×105 клеток (293Т, BHK-21, Hela, HepG2, Sp2/0, LMH, QM7).
7/ Нозерн-блоттинг
РНК экстрагируют из 106 трансфицированных или инфицированных клеток с помощью системы для выделения суммарной РНК (Promega®). РНК из неинфицирорванных клеток 293Т эктрагируют в качестве контроля. 2 мкг каждой РНК подвергают электрофорезу в денатурирующем формальдегидном геле, и эту РНК переносят на положительно заряженную нейлоновую мембрану (Hybond-XL, Amersham). Нозерн-блоттинг-гибридизацию осуществляют согласно стандартным методикам. Зонды соответствуют Agel-BamHI-фрагменту GFP размером 790 п.н. из плазмиды pEGEPC1 (Clontech), перед использованием этот фрагмент метят (система для мечения ДНК Rediprime® II; Amersham) и очищают на колонке (ProbeQuant® G-50 Micro Columns; Amersham).
II/ РЕЗУЛЬТАТЫ
I/ Функциональная активность вектора
Векторы SFV 26Sm1 и SFV 26Sm2 соответствуют векторам SFV, в которых 26S промотор был подвергнут мутации с целью предотвращения любой возможной конкуренции между упаковкой геномной РНК SFV и субгеномной РНК, продуцируемой в результате транскрипции под контролем 26S промотора. Функциональную активность этих двух векторов подтверждали с помощью трансфекции клеток 293Т. Наблюдаемая сильная экспрессия GFP означает, что транскрипция и трансляция модифицированного вектора SFV является правильной. Этот первый результат подтвержден Нозерн-блоттинг-анализом РНК, экстрагированной из клеток 293, трансфицированных вектором SFV 26Sm1.
Как видно из Фиг.3, дорожка 2, GFP зонд обнаруживает существование двух полос, соответствующих геномной РНК и субгеномной РНК, последнее означает, что 26S промотор является по-прежнему функциональным.
Тот же самый тест осуществляют на втором векторе SFV 26Sm2, содержащем дополнительные мутации. Обнаружение GFP и Нозерн-блоттинг-анализ подтверждают, что мутации, введенные в 26Sm2 промотор, ингибируют, на уровне транскрипции, продуцирование субгеномной РНК (смотри Фиг.3, дорожка 3).
2/ Пиодуцирование вирусных частиц
Клетки 293 котрансфицируют плазмидой pMDG, а затем вектором SFV 26Sm1 или SFV 26Sm2, как указано выше. Супернатант от трансфицированных клеток переносят на свежие клетки 293Т или клетки BHK 21. Полученная сильная и быстрая экспрессия GFP указывает на то, что векторы SVF можно мобилизовать с помощью клеток, экспрессирующих VSV-G оболочку (Фиг.4.).
3/ Способность полученных вирусных частиц инфицировать клеточные линии BHK21, 293Т и QM7
Результаты представлены в таблице ниже.
Вирусные титры определяют с помощью FACS-анализа через 24 после инфицирования. Процентное содержание клеток, экспрессирующих GFP, относительно количества клеток в день инфицирования, позволяет рассчитать титр рекомбинантных частиц. (IР(инфекционных частиц)/мл).
Как видно из этой таблицы, самый высокий титр получен на клетках BHK21 при сравнении с клетками 293Т и QM7.
4/ Экспрессия GFP, происходящая в клетках-мишенях, обусловлена реальной трансдукцией вирусными частицами SFV
Для того чтобы убедиться, что экспрессия GFP обусловлена экспрессией векторов SVF, а не мобилизацией плазмид, полученных из первоначальной трансфекции, или псевдотрансдукцией свободного GFP, осуществляют следующие контроли.
Прежде всего, с помощью Нозерн-блоттинга выявляют РНК SFV, используя РНК, экстрагированную из инфицированных клеток (смотри Фиг.5). Что касается клеток-продуцентов, то в клетках, инфицированных вектором SVF 26Sm1, обнаружена как геномная РНК, так и субгеномная РНК. С другой стороны, в клетках, инфицированных вектором SVF 26Sm2, обнаружена только геномная РНК. Интенсивность сигнала предполагает интенсивную репликацию векторов SVF. Однако для того чтобы убедиться в том, что сильная экспрессия GFP в клетках-мишенях действительно соответствует мобилизации РНК SFV, и что данные плазмиды действительно были перенесены в клетки-мишени, в данном примере в клетки 293Т, к трансдуцирующему супернатанту добавляют ДНКазу I в высокой концентрации (1000 МЕ/мл). Титры вирусных частиц SFV в этом случае схожи с титрами, полученными в отсутствие ДНКазы I, что предполагает трансдукцию, а не вторичную трансфекцию. Однако такой результат можно было бы получить, если плазмида после своего проникновения инкапсидируется в трансфицированных клетках и затем доставляется в трансдуцированную клетку. Для того чтобы проверить этот возможный феномен, клетки-мишени предварительно обрабатывают актиномицином D в концентрации один микрограмм на миллилитр и затем инкубируют с инфицирующим супернатантом. Актиномицин D ингибирует экспрессию генов, контролируемую РНК POL II (РНК-полимеразой II), такую как экспрессия генома вектора SFV в плазмиде pSFV26Sm1 или m2, но не оказывает действия на репликазу SFV. В присутствии и в отсутствие актиномицина D наблюдается сходная экспрессия GFP, что ясно подтверждает тот факт, что РНК переносится (смотри таблицу 2).
Затем решали вопрос о том, действительно ли экспрессия GFP обусловлена экспрессией векторов SFV или псевдотрансдукцией в клетки-миишени. Этот вопрос возникает потому, что в некоторых публикациях (7) было показано, что GFP может пассивно переноситься посредством ретровирусных частиц независимо от любой экспрессии. Для того чтобы убедиться, что это не так, клетки-мишени предварительно обрабатывают двумя ингибиторами трансляции, соответственно, генетицином и пуромицином. После обработки клетки-мишени показывают едва обнаруживаемую экспрессию GFP, что показывает, что наблюдаемый GFP является результатом трансляции, а не пассивного переноса (таблица 2). Кроме того, котрансфекция плазмиды pEGFPC1, интенсивно экспрессирующей GFP, с плазмидой, кодирующей VSV-G, не приводит к какой-либо псевдотрансдукции GFP. Аналогично, супернатанты, происходящие из клеток, трансфицированных только векторами SVF, не вызывают экспрессию GFP, что доказывает, что должен присутствовать VSV-G для того, чтобы стимулировать образование псевдочастиц. Для подтверждения того, что РНК SFV защищена в VSV-G везикулах, супернатанты обрабатывают РНКазой А перед трансдукцией. Показано, что обработка РНКазой А не оказывает никакого влияния на инфекционные титры, что подтверждает, что РНК SFV действительно защищена (таблица 2). Принимая во внимание все эти результаты, сделали вывод, что экспрессия GFP в клетках-мишенях обусловлена реальной трансдукцией вирусными частицами SFV.
ПРИМЕР 2
I/ МЕТОДЫ
1/ Конструкции
Были использованы конструкции, описанные в примере 1.
Были также использованы другие производные конструкции, представляющие собой замещение промотора CMV прокариотическим промотором SP6:
- первая конструкция, spSFV26Sm1, является прямым производным SFV26Sm1;
- вторая конструкция, spSFV26Sm1ψ, получена путем βglII-SmaI-переваривания плазмиды pSFV1 (Invitrogen®), в которую клонирован βglII-HpaI-фрагмент плазмиды pIRES2 GFP (Clontech®), модифицированный путем введения ПЦР-фрагмента, содержащего 3'-конец гена nsp4 и образованного с использованием праймеров 26Sm1F n26Sm1R (сравни пример 1, раздел 2а).
Эти конструкции транскрибируют in vitro, и затем РНК вводят путем электропорации в клетки-продуценты. Транскрипцию in vitro осуществляют после линеаризации плазмид путем βstBI-переваривания. Транскрипцию осуществляют в присутствии кэп-аналога (Invitrogen®), SP6-полимеразы (Invitrogen®) и рибонуклеотидов (Promega®).
2/ Клетки:
Происходящие из клеток 293 клетки Phoenix®, продуцирующие рекомбинантный ретровирус (http://www.stanford.edu/group/nolan/retroviral systems/phx.html), культивируют в среде DMEM (GIBCO) в присутствии не содержащей комплемент эмбриональной телячьей сыворотки (Abcys).
Данные клетки-продуценты трансфицируют плазмидами SFV26Sm1 и 26Sm2 в количестве 4 мкг ДНК на 5×105 клеток, в лунке шестилуночного планшета. Трансфекцию осуществляют, используя фосфат кальция (набор для кальций-фосфатной трансфекции, Invitrogen®).
Для этих двух конструкций, экспрессирующих векторы SFV в форме РНК, трансфекцию осуществляют путем электропорации: 40 мкл репликонов, продуцируемых in vitro, помещают вместе с 40×105 клеток и подвергают электропорации, используя систему EasyjecT Plus (Equibio®).
Через 20 часов после трансфекции, независимо от используемого метода трансфекции, меняют среду. Через 16 часов после этой смены среду собирают для того, чтобы осуществить инфицирование. Во время сбора среду фильтруют, используя 0,45 мкм фильтры (Millipore®).
3/ Инициирование
Отфильтрованные супернатанты используют для инфицирования клеток 293Т, помещенных в культуру в 12-луночные планшеты. Инфицирование осуществляют в присутствии поликатиона, полибрена (Sigma®), необходимого для вирусно-клеточных взаимодействий, в концентрации 5 мкг/мл. В день инфицирования одну лунку с клетками-мишенями 293Т обрабатывают трипсином для подсчета.
Через 24 часа после инфицирования клетки обрабатывают трипсином для анализа методом проточной цитометрии (FACScalibur, Becton-Dickinson®). Процент клеток, экспрессирующих GFP, относительно количества клеток в день инфекции, позволяет рассчитать титр рекомбинантных частиц (IP/мл), (таблица 3).
4/ Контроли
Осуществляли контроли, идентичные контролям, осуществленным в примере 1:
- 10 мкг на мл РНКазы А (Sigma®),
- 1 мкг на мл актиномицина D (Sigma®),
- 100 единиц на мл ДНКазы I (invitrogen®),
- 1 мг на мл генетицина (Sigma®).
II/ РЕЗУЛЬТАТЫ
1/ Инфицирование
Результаты инфицирования представлены в таблице 3.
IP/мл: количество инфекционных частиц на мл; н/о: не определяли.
Присутствие клеток, экспрессирующих GFP, подтверждает возможность мобилизации рекомбинантных РНК SFV посредством интервенции ретровирусной частицы. Однако наблюдаемые низкие титры указывают на то, что для получения более высоких титров необходимо контролировать цитотоксичность вектора SFV. Причиной низких титров является антагонизм, который существует между продукцией РНК SFV и продукцией ретровирусных белков. Продукция последних уменьшается, когда увеличивается продукция белков SFV. В настоящее время описано и может быть успешно использовано несколько мутантов SFV (8).
Присутствие или отсутствие ретровирусных последовательностей для инкапсидации, по-видимому, не оказывает существенного влияния на эффективность инкапсидации. В соответствии с наблюдениями Muriaux с соавт.(9), определяющую роль в стимуляции инкапсидации здесь, по-видимому, играет высокая внутриклеточная концентрация РНК. В контексте низкотоксичных векторов необходимо будет переоценить влияние ретровирусной psi-последовательности.
Кроме того, эти результаты, по-видимому, указывают на то, что при использовании "хелперной" системы на основе векторов SFV (10, 11), возможно, существует контаминация продуцируемыми рекомбинантными ретровирусами. Эти контаминации состоят из ретровирусных частиц, содержащих либо векторы SFV, используемые для экспрессии ретровирусных последовательностей, предназначенных для транскомплементации, либо векторы SFV, содержащие последовательность рекомбинантного ретровируса. Это наблюдение ставит под сомнение применение этих способов продуцирования ретровирусных векторов для клинических целей, в отличие от вирусных частиц по данному изобретению.
Источники информации
1. Rolls, M.M., Webster, P., Balba, N.H. & Rose, J.K. Novel infectious particles generated by expression of the vesicular stomatitis virus glycoprotein from a self-replicating RNA. Cell 79, 497-506. (1994).
2. Rolls, M.M., Haglund, K. & Rose, J.К. Expression of additional genes in a vector derived from a minimal RNA virus. Virology 218, 406-411. (1996).
3. Lebedeva, I., Fujita, K., Nihrane, A. & Silver, J. Infectious particles derived from Semliki Forest Virus Vectors encoding Murine Leukemia virus envelopes. Journal of Virology 71 (9), 7061-7067. (1997).
4. Russell SJ, Cosset FL. Modifying the host range properties of retroviral vectors. J Gene Med 1, 300-11. (1999).
5. Salonen A, Vasiljeva L, Merits A, Magden J, Jokitalo E, Kaariainen L. Properly folded nonstructural polyprotein directs the semliki forest virus replication complex to the endosomal compartment. J Virol. 77,1691-702. (2003).
6. Naldini, L. et al. In vivo gene delivery and stable transduction of non-dividing cells by a lentiviral vector. Science 272,263-267 (1996).
7. Liu, M.L, Winther, B.L. & Kay, M.A. Pseudotransduction of hepatocytes by using concentrated pseudotyped vesicular stomatitis virus G glycoprotein (VSV-G)-Moloney murine leukemia virus-derived retrovirus vectors: comparison of VSV-G and amphotropic vectors for hepatic gene transfer. J Virol 70, 2497-2502. (1996).
8. Lundstrom К, Abenavoli A, Malgaroli A, Ehrengruber ML). Novel semliki forest virus vectors with reduced cytotoxicity and temperature sensitivity for long-term enhancement of transgene expression. Mol Ther. 2, 7202-9. (2003).
9. Muriaux, D., J. Mirro, et al.. RNA is a structural element in retrovirus particles. Proc Natl Acad Sci USA 98, 5246-51.(2001).
10. Wahlfors JJ, Xanthopoulos KG, Morgan RA. Semliki Forest virus-mediated production of retroviral vector RNA in retroviral packaging cells. Hum Gene Ther 8, 2031-41. (1997).
11. Li KJ, Garoff H. Packaging of intron-containing genes into retrovirus vectors by alphavirus vectors. Proc NatI Acad Sci USA 95, 3650-4. (1998).
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОДУЦИРОВАНИЯ РЕПЛИКАТИВНОЙ ЧАСТИЦЫ ВИРУСА ГРИППА, КОМПОЗИЦИЯ КЛЕТОК (ВАРИАНТЫ), КОМПОЗИЦИЯ КЛЕТОЧНОЙ КУЛЬТУРЫ И ЕЕ ПРИМЕНЕНИЕ | 2005 |
|
RU2435855C2 |
| СПОСОБ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ ВИРУСНОЙ ТРАНСДУКЦИИ | 2011 |
|
RU2478711C1 |
| СИСТЕМА ОЧИСТКИ ВИРУСНЫХ ВЕКТОРОВ | 2012 |
|
RU2607044C2 |
| АДЕНОВИРУСНЫЙ/АЛЬФА-ВИРУСНЫЙ ГИБРИДНЫЙ ВЕКТОР ДЛЯ ЭФФЕКТИВНОГО ВВЕДЕНИЯ И ЭКСПРЕССИИ ТЕРАПЕВТИЧЕСКИХ ГЕНОВ В ОПУХОЛЕВЫЕ КЛЕТКИ | 2005 |
|
RU2394104C2 |
| Способ получения рекомбинантного вируса везикулярного стоматита | 2022 |
|
RU2807751C1 |
| ВИРУСНЫЕ ВЕКТОРЫ С ЗАВИСИМОЙ ОТ УСЛОВИЙ РЕПЛИКАЦИЕЙ И ИХ ПРИМЕНЕНИЕ | 2001 |
|
RU2301260C2 |
| ЧАСТИЦА РЕПЛИКОНА АЛЬФАВИРУСА | 2018 |
|
RU2795596C2 |
| СПОСОБ ВРЕМЕННОЙ ТРАНСФЕКЦИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСА | 2016 |
|
RU2749717C2 |
| БИОПРОИЗВОДСТВО ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2016 |
|
RU2729385C2 |
| СТАБИЛЬНЫЕ КЛЕТОЧНЫЕ ЛИНИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСОВ | 2016 |
|
RU2752498C2 |
Изобретение относится к области биотехнологии, генной инженерии, вирусологии и медицины. Вирусная частица состоит из структурных элементов, не происходящих из альфавируса, и содержит вектор, происходящий из альфавируса, дефектный по репликации в результате делеции или замещения по меньшей мере одним трансгеном. Указанная частица содержит также структурные элементы, которые не кодируются геномом вектора, происходящего из альфавируса. Способ получения указанной частицы согласно изобретению состоит в экспрессии in trans генов, кодирующих структурные элементы, не происходящие из альфавируса, и вектора, происходящего из альфавируса, в клеточной линии и затем в выделении вирусных частиц, содержащихся в супернатанте клеточной культуры. Изобретение позволяет получить вирусные частицы с пониженным риском рекомбинации. Изобретение может быть использовано в медицине. 5 н. и 19 з.п. ф-лы, 5 ил., 3 табл.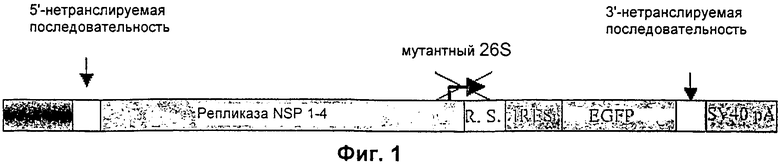
1. Вирусная частица для экспрессии трансгена в клетках-мишени, состоящая из структурных элементов, не происходящих из альфавируса, и содержащая вектор, происходящий из альфавируса, дефектный по репликации в результате делеции или замещения структурных генов по меньшей мере одним трансгеном, отличающаяся тем, что структурные элементы указанной частицы не кодируются геномом вектора, происходящего из альфавируса.
2. Вирусная частица по п.1, отличающаяся тем, что структурные элементы соответствуют одному белку оболочки VSV-G.
3. Вирусная частица по п.1, отличающаяся тем, что структурные элементы соответствуют структурным белкам ретровируса.
4. Частица по п.1, отличающаяся тем, что альфавирус представляет собой вирус леса Семлики.
5. Частица по п.1, отличающаяся тем, что геном вектора, происходящего из альфавируса, содержит протяженную последовательность для упаковки из MLV-векторов.
6. Частица по п.1, отличающаяся тем, что геном вектора, происходящего из альфавируса, лишен psi-последовательности.
7. Частица по п.1, отличающаяся тем, что геном вектора, происходящего из альфавируса, содержит расположенный на 5'-конце эукариотический промотор.
8. Частица по п.1, отличающаяся тем, что геном вектора, происходящего из альфавируса, содержит мутантный промотор p26S.
9. Применение вирусной частицы по любому из пп.1-8 для инфицирования эукариотической клетки in vitro.
10. Фармацевтическая композиция для экспрессии трансгена в клетке-мишени, содержащая вирусную частицу по любому из пп.1-8 и среду, содержащую эту частицу.
11. Применение вирусной частицы по любому из пп.1-8 для изготовления лекарственного препарата для применения в лечении рака.
12. Способ получения вирусных частиц, состоящих из структурных элементов, не происходящих из альфавируса, и содержащих вектор, происходящий из альфавируса, дефектный по репликации в результате делеции или замещения структурных генов по меньшей мере одним трансгеном, заключающийся:
в экспрессии in trans, в клеточной линии, генов, кодирующих структурные элементы, не происходящие из альфавируса и вектора, происходящего из альфавируса,
в выделении вирусных частиц, присутствующих в супернатанте клеточной культуры.
13. Способ по п.12, отличающийся тем, что структурные элементы соответствуют белку оболочки VSV-G.
14. Способ по п.13, отличающийся тем, что экспрессия in trans получена путем котрансфекции клеточной линии вектором для экспрессии VSV-G-оболочки и вектором, происходящим из альфавируса, причем котрансфекцию осуществляют в две отдельные стадии, соответственно, трансфекции клеточной линии вектором, экспрессирующим ген оболочки VSV-G, и затем второй трансфекции вектором, происходящим из альфавируса.
15. Способ по п.14, отличающийся тем, что трансфицированная клеточная линия представляет собой клеточную линию 293Т.
16. Способ по п.12, отличающийся тем, что структурные элементы соответствуют структурным белкам ретровируса.
17. Способ по п.16, отличающийся тем, что экспрессия in trans получена путем трансфекции упаковывающей клеточной линии, которая продуцирует дефектные по репликации ретровирусы, вектором, происходящим из альфавируса.
18. Способ по п.17, отличающийся тем, что упаковывающая клеточная линия получена путем стабильной трансфекции клеточной линии первым вирусным элементом, экспрессирующим ретровирусные гены GAG и POL, и вторым вирусным элементом, экспрессирующим ретровирусный ген ENV.
19. Способ по п.16, отличающийся тем, что экспрессия in trans получена тройной трансфекцией клеточной линии 293 Т путем введения первого вирусного элемента, экспрессирующего ретровирусные гены GAG и POL, второго вирусного элемента, экспрессирующего ретровирусный ген ENV, и вектора, происходящего из альфавируса.
20. Способ по п.12, отличающийся тем, что альфавирус представляет собой вирус леса Семлики.
21. Способ по п.12, отличающийся тем, что геном вектора, происходящего из альфавируса, содержит протяженную последовательность для упаковки из MLV-векторов.
22. Способ по п.12, отличающийся тем, что геном вектора, происходящего из альфавируса, лишен psi-последовательности.
23. Способ по п.12, отличающийся тем, что геном вектора, происходящего из альфавируса, содержит расположенный на 5'-конце эукариотический промотор.
24. Способ по п.12, отличающийся тем, что вектор, происходящий из альфавируса, содержит мутантный промотор p26S.
| LEBEDEVA I | |||
| et al., "Infectious particles derived from Semliki Forest virus vectors encoding murine leukemia virus envelopes", J Virol | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| WO 03072771, 04.09.2003 | |||
| LUNDSTROM K., "Alphavirus vectors as tools in cancer gene therapy", Technol Cancer Res Treat | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |

