По данной заявке испрашивается приоритет по патентным заявкам США с серийными номерами 60/671657 (поданная 15 апреля 2005 года; истекшая); 11/409339 (поданная 17 апреля 2006 года; на рассмотрении); PCT/US06/014481 (поданная 17 апреля 2006 года; истекшая); 60/945523 (поданная 21 июля 2007 года; истекшая), 61/019051 (поданная 4 января 2008 года; истекшая); 12/138867 (поданная 13 июня 2008 года, на рассмотрении), PCT/US08/066957 (поданная 13 июня 2008 года, на рассмотрении), 61/139352 (поданная 19 декабря 2008 года, на рассмотрении), 61/156035 (поданная 27 февраля 2009 года, на рассмотрении) и 61/256779 (поданная 30 октября 2009 года; на рассмотрении), которые все включены в настоящий документ в качестве ссылок в полном объеме.
1. ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к молекулам диантител, иначе называемым "перенацеливающими реагентами с двойной аффинностью" ("DARTS"), и к их применению для лечения различных заболеваний и нарушений, включающих иммунологические нарушения и злокачественные опухоли. Молекулы диантител по изобретению содержат по меньшей мере две полипептидных цепи, которые ассоциируют с образованием по меньшей мере двух участков связывания эпитопа, которые могут распознавать один и тот же или различные эпитопы. Кроме того, эпитопы могут быть из одной или из различных молекул или они могут быть расположены на одной или различных клетках. Отдельные полипептидные цепи молекулы диантитела могут быть ковалентно связаны через непептидные ковалентные связи, такие как, но не ограничиваясь ими, дисульфидная связь между остатками цистеина, расположенными в каждой полипептидной цепи. В конкретных вариантах осуществления, молекулы диантител по настоящему изобретению дополнительно содержат Fc-область, которая позволяет внесение в молекулу функциональности, подобной антителу.
2. УРОВЕНЬ ТЕХНИКИ
Конструкция ковалентных антител основана на конструкции одноцепочечных Fv (scFv) (Holliger et al. (1993) "'Diabodies': Small Bivalent And Bispecific Antibody Fragments", Proc. Natl. Acad. Sci. USA 90:6444-6448; включенная в настоящий документ в качестве ссылки в полном объеме). В целом немодифицированном IgG домены VL и VH расположены на отдельных полипептидных цепях, т.е. на легкой цепи и тяжелой цепи, соответственно. Взаимодействие легкой цепи антитела и тяжелой цепи антитела и, в частности, взаимодействие доменов VL и VH, приводит к образованию одного из участков связывания эпитопа антитела. Напротив, конструкция scFv содержит домены VL и VH антитела, содержащиеся в одной полипептидной цепи, где эти домены разделены подвижным линкером достаточной длины для обеспечения самосборки двух доменов в функциональный участок связывания эпитопа. Когда самосборка является невозможной вследствие недостаточной длины линкера (менее чем приблизительно 12 аминокислотных остатков), две конструкции scFv взаимодействуют друг с другом с образованием двухвалентной молекулы, причем VL одной цепи ассоциирует с VH другой (рассмотрено в Marvin et al. (2005) "Recombinant Approaches To IgG-Like Bispecific Antibodies", Acta Pharmacol. Sin. 26:649-658). Более того, показано, что добавление остатка цистеина на C-конец конструкции позволяет образование дисульфидных связей между полипептидными цепями, стабилизируя полученный димер без препятствования характеристикам связывания двухвалентной молекулы (см. например, Olafsen et al. (2004) "Covalent Disulfide-Linked Anti-CEA Diabldy Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications", Prot. Engr. Des. Sel. 17:21-27). Кроме того, когда выбирают домены VL и VH с отличающейся специфичностью, можно конструировать не только двухвалентную, но также и биспецифическую молекулу.
Двухвалентные диантитела имеют широкий диапазон применений, включая терапию и иммунодиагностику. Двухвалентность позволяет значительную гибкость при разработке и конструировании диантитела для различных применений, обеспечивая увеличенную авидность к мультимерным антигенам, связывание с различными антигенами и направленное нацеливание в конкретные типы клеток на основании наличия обоих антигенов-мишеней. Также было показано, что вследствие их увеличенной валентности, низких констант диссоциации и быстрого выведения из кровотока (для диантител небольшого размера с массой ~50 кДа или менее), молекулы диантител, известные в данной области, особенно применимы в области визуализации опухоли (Fitzgerald et al. (1997) "Improved Tumour Targeting By Disulphide Stabilized Diantibodies Expressed In Pichia pastoris", Protein Eng. 10:1221). Особенно важным является связывание различных клеток, например, связывание цитотоксических T-клеток с клетками опухоли (Staerz et al. (1985) "Hybrid Antibodies Can Target Sites For Attack By T Cells", Nature 314:628-631, и Holliger et al. (1996) "Specific Killing Of Limphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diantibody", Protein Eng. 9:299-305). Связывающие эпитоп домены диантитела также могут быть нацелены на поверхностную детерминанту любой иммунной эффекторной клетки, такую как CD3, CD16, CD32 или CD64, которые экспрессируются на T-лимфоцитах, естественных киллерных (NK) клетках или других мононуклеарных клетках. Во многих исследованиях также выявлено, что связывание диантитела с детерминантами эффекторных клеток, например, рецепторами Fcγ (FcγR), активирует эффекторные клетки (Holliger et al. (1996) "Specific Killing Of Limphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody", Protein Eng. 9:299-305; Holliger et al. (1999) "Carcinoembryonic Antigen (CEA)-Specific T-cell Activation In Colon Carcinoma Induced By Anti-CD 3×Anti-CEA Bispecific Diabodies And B7×Anti-CEA Bispecific Fusion Proteins", Cancer Res. 59:2909-2916). Обычно активация эффекторных клеток запускается связыванием связанного с антигеном антитела с эффекторной клеткой путем взаимодействия Fc-FcγR; таким образом, в этом отношении, молекулы диантитела по изобретению могут проявлять Ig-подобную функциональность, независимо от того, содержат ли они Fc-домен (например, как анализируют в любом анализе эффекторных функций, известном в данной области или проиллюстрированном в настоящем документе (например, в анализе ADCC)). Путем связывания опухолевых и эффекторных клеток, диантитело не только сближает эффекторную клетку с опухолевой клеткой, но также приводит к эффективному уничтожению опухоли (см. например, Cao et al. (2003) "Bispecific Antibody Conjugates In Therapeutics", Adv. Drug. Deliv. Rev. 55:171-197, включенную в настоящий документ в качестве ссылки в полном объеме).
2.1 РЕЦЕПТОРЫ ЭФФЕКТОРНЫХ КЛЕТОК И ИХ РОЛИ В ИММУННОЙ СИСТЕМЕ
При обычном функционировании иммунной системы взаимодействие комплексов антитело-антиген иммунной системы приводит к широкому набору ответов, варьирующему от эффекторных функций, таких как антителозависимая цитотоксичность, дегрануляция тучных клеток и фагоцитоз, до иммуномодулирующих сигналов, таких как регуляция пролиферации лимфоцитов и секреции антител. Все эти взаимодействия инициируются путем связывания Fc-домена антител или иммунных комплексов со специализированными рецепторами клеточной поверхности на кроветворных клетках. Разнообразие клеточных ответов, запускаемых антителами и иммунными комплексами, является следствием структурной гетерогенности Fc-рецепторов. Fc-рецепторы обладают общими структурно родственными антигенсвязывающими доменами, которые предположительно опосредуют внутриклеточную передачу сигнала.
Fcγ-рецепторы, представители суперсемейства белков генов иммуноглобулинов, представляют собой поверхностные гликопротеины, которые могут связывать Fcγ-часть молекул иммуноглобулинов. Каждый представитель этого семейства распознает иммуноглобулины одного или нескольких изотипов через домен распознавания на альфа-цепи рецептора Fcγ. Fcγ-рецепторы определяются их специфичностью к подтипам иммуноглобулинов. Fcγ-рецепторы для IgG обозначают как FcγR, для IgE как FcεR, и для IgA как FcαR. Различные вспомогательные клетки содержат Fcγ-рецепторы для антител различного изотипа, и изотип антитела определяет, какие вспомогательные клетки будут вовлечены в данный ответ (рассмотрено Ravetch J.V. et al. (1991) "Fc Receptors", Annu. Rev. Immunol. 9:457-92; Gerber J.S. et al. (2001) "Stimulatory And Inhibitory Signals Originating From The Macrophage Fcgamma Receptors", Microbes and Infection, 3:131-139; Billadeau D.D. et al. (2002), "ITAMs Versus ITIMs: Striking A Balance During Cell Regulation", The Journal of Clinical Investigation, 2(109):161-1681; Ravetch J.V. et al. (2000) "Immune Inhibitory Receptors", Science, 290:84-89; Ravetch J.V. et al., (2001) "IgG Fc Receptors", Annu. Rev. Immunol. 19:275-90; Ravetch J.V. (1994) "Fc Receptors: Rubor Redux", Cell, 78(4):553-60). Различные Fcγ-рецепторы, клетки, которые их экспрессируют, и их изотипическая специфичность обобщенно представлены в таблице 1 (взятой из Immunobiology: The Immune System in Health and Disease, 4th ed. 1999, Elsevier Science Ltd/Garland Publishing, New York).
Fcγ-Рецепторы
Каждый представитель этого семейства представляет собой интегральный мембранный гликопротеин, обладающий внеклеточными доменами, родственными C2-группе доменов, родственных иммуноглобулинам, однократно проходящим через мембрану доменом и внутрицитоплазматическим доменом переменной длины. Существует три известных FcγR, обозначаемых как FcγRI(CD64), FcγRII(CD32) и FcγRIII(CD16). Эти три рецептора кодируются различными генами; однако высокая гомология между этими тремя представителями указывает на то, что они возникли из общего предшественника, возможно, дупликацией гена.
FcγRII(CD32)
FcγRII-белки представляют собой интегральные мембранные гликопротеины размером 40 кДа, которые связываются только с IgG в комплексе вследствие низкой аффинности к мономерному Ig (106 M-1). Этот рецептор представляет собой наиболее широко экспрессируемый FcγR, присутствующий на всех кроветворных клетках, включая моноциты, макрофаги, B-клетки, NK-клетки, нейтрофилы, тучные клетки и тромбоциты. FcγRII имеет только две иммуноглобулин-подобных области в его иммуноглобулин-связывающей цепи и, таким образом, обладает значительно более низкой аффинностью в отношении IgG, чем FcγRI. Существует три гена FcγRII человека (FcγRII-A, FcγRII-B, FcγRII-C), все из которых связывают IgG в агрегатах или иммунных комплексах.
Четкие различия цитоплазматических доменов FcγRII-A и FcγRII-B создают два функционально гетерогенных ответа на лигирование рецептора. Фундаментальное отличие состоит в том, что изоформа A инициирует внутриклеточную передачу сигнала, ведущую к активации клеток, такой как фагоцитоз и дыхательный взрыв, в то время как изоформа B инициирует ингибиторные сигналы, например, ингибирование активации B-клеток.
FcγRIII (CD16)
Вследствие гетерогенности в этом классе размер FcγRIII варьирует от 40 до 80 кДа у мыши и человека. Два гена кодируют два транскрипта, FcγRIIIA - интегральный мембранный гликопротеин, и FcγRIIIB - гликозилфосфатидилинозитол (GPI)-связанный вариант. Один ген мыши кодирует FcγRIII, гомологичный проходящему через мембрану FcγRIIIA человека. FcγRIII имеет общие структурные характеристики с каждым из двух других FcγR. Подобно FcγRII, FcγRIII связывает IgG с низкой аффинностью и содержит соответствующие два внеклеточных Ig-подобных домена. FcγRIIIA экспрессируется в макрофагах, тучных клетках и является единственным FcγR в NK-клетках. В настоящее время известно, что связанный с GPI FcγRIIIB экспрессируется только в нейтрофилах человека.
Передача сигнала через FcγR
После лигирования через FcγR передаются как активирующие, так и ингибиторные сигналы. Эти диаметрально противоположные функции являются следствием структурных отличий среди различных изоформ рецептора. За различные ответы отвечают два различных домена в цитоплазматических доменах передачи сигнала рецептора, называемых тирозинсодержащими активационными мотивами иммунорецепторов (ITAM) или тирозинсодержащими ингибиторными мотивами иммунорецепторов (ITIM). Привлечение различных цитоплазматических ферментов в эти структуры определяет исход опосредуемых FcγR клеточных ответов. ITAM-содержащие комплексы FcγR включают FcγRI, FcγRIIA, FcγRIIIA, в то время как ITIM-содержащие комплексы включают только FcγRIIB.
Нейтрофилы человека экспрессируют ген FcγRIIA. Кластеризация FcγRIIA посредством иммунных комплексов или специфического связывания антителом служит для агрегации ITAM вместе с рецептор-ассоциированными киназами, которые способствуют фосфорилированию ITAM. Область фосфорилирования ITAM служит в качестве участка для присоединения Syk-киназы, активация которой приводит к активации нижерасположенных субстратов (например, PI3K). Активация клеток приводит к высвобождению провоспалительных медиаторов.
Ген FcγRIIB экспрессируется на B-лимфоцитах; его внеклеточный домен на 96% идентичен FcγRIIA и они связывают комплексы IgG неотличимо. Этот ингибиторный подкласс FcγR определяет присутствие ITIM в цитоплазматическом домене FcγRIIB. Недавно была установлена молекулярная основа для этого ингибирования. При совместном лигировании с активирующим FcγR, ITIM в FcγRIIB становится фосфорилированным и привлекает SH2-домен инозитолполифосфат-5'-фосфатазы (SHIP), которая гидролизует фосфоинозитоловые посредники в результате опосредуемой ITAM-содержащим FcγR активации тирозинкиназы, таким образом, препятствуя входящему потоку внутриклеточного Ca++. Таким образом, связывание FcγRIIB сдерживает активационный ответ на лигирование FcγR и ингибирует способность клеток к ответу. Таким образом, предотвращаются активация B-клеток, пролиферация B-клеток и секреция антител.
Рецепторы для Fc-областей изотипов иммуноглобулинов
108 M-1
Нейтрофилы
Эозинофилы
Дендритные клетки
Активация дыхательного взрыва,
Индукция уничтожения
2×106 M-1
Нейтрофилы
Эозинофилы
Дендритные клетки
Тромбоциты
Клетки Лангерганса
2×106 M-1
Нейтрофилы
Эозинофилы
2×106 M-1
Тучные клетки
5×105 M-1
Эозинофилы
Макрофаги
Нейтрофилы
Тучные клетки
1010 M-1
Эозинофилы
Базофилы
107 M-1
Нейтрофилы
Эозинофилы
3. СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к ковалентным диантителам и/или молекулам ковалентных диантител и к их применению для лечения различных заболеваний и нарушений, включая злокачественную опухоль, аутоиммунные нарушения, аллергические нарушения и инфекционные заболевания, вызываемые бактериями, грибами или вирусами. Предпочтительно, диантитело по настоящему изобретению может связываться с двумя различными эпитопами на двух различных клетках, где первый эпитоп экспрессируется на типе клеток, отличающемся от типа клеток, на котором экспрессируется второй эпитоп, так что диантитело может сближать эти две клетки.
В одном из вариантов осуществления настоящее изобретение относится к ковалентному биспецифическому диантителу, которое содержит первую и вторую полипептидную цепи, причем первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичную к первому эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичную ко второму эпитопу, и, необязательно, (iii) третий домен, содержащий по меньшей мере один остаток цистеина, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа; причем вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), и, необязательно, (iii) шестой домен, содержащий по меньшей мере один остаток цистеина, где четвертый и пятый домены ковалентно связаны, так что четвертый и пятый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен ассоциируют с образованием первого участка связывания (VL1)(VH1), который связывает первый эпитоп; где второй домен и четвертый домен ассоциируют с образованием второго участка связывания (VL2)(VH2), который связывает второй эпитоп.
В другом варианте осуществления настоящее изобретение относится к ковалентному биспецифическому диантителу, которое содержит первую и вторую полипептидные цепи, причем первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичную к первому эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичную ко второму эпитопу, и (iii) третий домен, содержащий Fc-домен или его часть, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа; причем вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), где четвертый и пятый домены ковалентно связаны, так что третий и четвертый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен ассоциируют с образованием первого участка связывания (VL1)(VH1), который связывает первый эпитоп; где второй домен и четвертый домен ассоциируют с образованием второго участка связывания (VL2)(VH2), который связывает второй эпитоп.
В некоторых аспектах настоящее изобретение относится к молекуле диантитела, которая содержит первую и вторую полипептидные цепи, причем первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичную к первому эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичную ко второму эпитопу, и (iii) третий домен, содержащий Fc-домен или его часть, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа; причем вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), и (iii) шестой домен, содержащий по меньшей мере один остаток цистеина, где четвертый и пятый домены ковалентно связаны, так что четвертый и пятый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен ассоциируют с образованием первого участка связывания (VL1)(VH1), который связывает первый эпитоп; где второй домен и четвертый домен ассоциируют с образованием второго участка связывания (VL2)(VH2), который связывает второй эпитоп.
В определенных вариантах осуществления настоящее изобретение относится к ковалентному биспецифическому диантителу, которое представляет собой димер молекулы диантитела, причем каждая молекула диантитела содержит первую и вторую полипептидную цепи, причем первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичную к первому эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичную ко второму эпитопу, и (iii) третий домен, содержащий Fc-домен или его часть, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа; и где вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), и (iii) шестой домен, содержащий по меньшей мере один остаток цистеина, где четвертый и пятый домены ковалентно связаны, так что четвертый и пятый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь каждой молекулы диантитела ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен каждой молекулы диантитела ассоциируют с образованием первого участка связывания (VL1)(VH1), который связывает первый эпитоп; где второй домен и четвертый домен каждой молекулы диантитела ассоциируют с образованием второго участка связывания (VL2)(VH2), который связывает второй эпитоп.
В других вариантах осуществления настоящее изобретение относится к ковалентному тетраспецифическому диантителу, которое представляет собой димер молекул диантител, причем первая молекула диантитела содержит первую и вторую полипептидные цепи, причем первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичную к первому эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичную ко второму эпитопу, и (iii) третий домен, содержащий Fc-домен или его часть, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа; и где вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), и (iii) шестой домен, содержащий по меньшей мере один остаток цистеина, где четвертый и пятый домены ковалентно связаны, так что четвертый и пятый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен ассоциируют с образованием первого участка связывания (VL1)(VH1), который связывает первый эпитоп; где второй домен и четвертый домен ассоциируют с образованием второго участка связывания (VL2)(VH2), который связывает второй эпитоп; и вторая молекула диантитела содержит первую и вторую полипептидные цепи, причем первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи третьего иммуноглобулина (VL3), специфичную к третьему эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи четвертого иммуноглобулина (VH4), специфичную к четвертому эпитопу, и (iii) третий домен, содержащий Fc-домен или его часть, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа; и где вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи четвертого иммуноглобулина (VL4), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи третьего иммуноглобулина (VH3), и (iii) шестой домен, содержащий по меньшей мере один остаток цистеина, где четвертый и пятый домены ковалентно связаны, так что четвертый и пятый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен ассоциируют с образованием первого участка связывания (VL3)(VH3), который связывает третий эпитоп; где второй домен и четвертый домен ассоциируют с образованием второго участка связывания (VL4)(VH4), который связывает четвертый эпитоп.
В определенных аспектах изобретения первый эпитоп, второй эпитоп и, где это применимо, третий эпитоп и четвертый эпитоп могут быть одинаковыми. В других аспектах каждый из первого эпитопа, второго эпитопа и, где это применимо, третьего эпитопа и четвертого эпитопа могут отличаться друг от друга. В определенных аспектах изобретения, включающих связывающий третий эпитоп домен, первый эпитоп и третий эпитоп могут быть одинаковыми. В определенных аспектах изобретения, включающих связывающий четвертый эпитоп домен, первый эпитоп и четвертый эпитоп могут быть одинаковыми. В определенных аспектах изобретения, включающих связывающий третий эпитоп домен, второй эпитоп и третий эпитоп могут быть одинаковыми. В определенных аспектах изобретения, включающих связывающий четвертый эпитоп домен, второй эпитоп и четвертый эпитоп могут быть одинаковыми. В предпочтительных аспектах изобретения первый эпитоп и второй эпитоп отличаются. В других аспектах изобретения, включающих связывающий третий эпитоп домен и связывающий четвертый эпитоп домен, третий эпитоп и четвертый эпитоп могут отличаться. Следует понимать, что настоящее изобретение охватывает любую комбинацию из указанных выше.
В конкретных аспектах изобретения первый домен и пятый домен диантитела или молекулы диантитела могут происходить из одного иммуноглобулина. В другом аспекте второй домен и четвертый домен диантитела или молекулы диантитела могут происходить из одного иммуноглобулина. В другом аспекте первый домен и пятый домен диантитела или молекулы диантитела могут происходить из различных иммуноглобулинов. В другом аспекте второй домен и четвертый домен диантитела или молекулы диантитела могут происходить из различных иммуноглобулинов. Следует понимать, что настоящее изобретение охватывает любую комбинацию из указанных выше.
В определенных аспектах изобретения ковалентная связь между первой полипептидной цепью и второй полипептидной цепью диантитела или молекулы диантитела может быть через дисульфидную связь между по меньшей мере одним остатком цистеина первой полипептидной цепи и по меньшей мере одним остатком цистеина второй полипептидной цепи. Остатки цистеина первой или второй полипептидных цепей, которые ответственны за образование дисульфидных связей, могут находиться в любом месте на полипептидной цепи, включая первый, второй, третий, четвертый, пятый и шестой домены. В конкретном варианте осуществления остаток цистеина на первой полипептидной цепи находится в первом домене и остаток цистеина на второй полипептидной цепи находится в пятом домене. Первый, второй, четвертый и пятый домены соответствуют вариабельным областям, ответственным за связывание. В предпочтительных вариантах осуществления остатки цистеина, ответственные за образование дисульфидных связей между первой и второй полипептидными цепями, расположены в третьем и шестом доменах, соответственно. В конкретном аспекте этого варианта осуществления третий домен первой полипептидной цепи содержит 6 C-концевых аминокислот легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23), которые могут кодироваться аминокислотной последовательностью (SEQ ID NO: 17). В другом аспекте этого варианта осуществления шестой домен второй полипептидной цепи содержит 6 C-концевых аминокислот легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23), которые могут кодироваться аминокислотной последовательностью (SEQ ID NO: 17). В другом аспекте этого варианта осуществления третий домен первой полипептидной цепи содержит аминокислотную последовательность VEPKSC (SEQ ID NO: 77), происходящую из шарнирного домена IgG человека, которая может кодироваться нуклеотидной последовательностью (SEQ ID NO: 78). В другом аспекте этого варианта осуществления шестой домен второй полипептидной цепи содержит аминокислотную последовательность VEPKSC (SEQ ID NO: 77), происходящую из шарнирного домена IgG человека, которая может кодироваться нуклеотидной последовательностью (SEQ ID NO: 78). В некоторых аспектах этого варианта осуществления третий домен первой полипептидной цепи содержит 6 C-концевых аминокислот легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23); и шестой домен второй полипептидной цепи содержит аминокислотную последовательность VEPKSC (SEQ ID NO: 77). В других аспектах этого варианта осуществления шестой домен второй полипептидной цепи содержит 6 C-концевых аминокислот легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23); и третий домен первой полипептидной цепи содержит аминокислотную последовательность VEPKSC (SEQ ID NO: 77). В других аспектах этого варианта осуществления третий домен первой полипептидной цепи содержит 6 C-концевых аминокислот легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23); и шестой домен второй полипептидной цепи содержит шарнирный домен. В других аспектах этого варианта осуществления шестой домен второй полипептидной цепи содержит 6 C-концевых аминокислот легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23); и третий домен первой полипептидной цепи содержит шарнирный домен. В других аспектах этого варианта осуществления третий домен первой полипептидной цепи содержит 6 C-концевых аминокислот легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23); и шестой домен первой полипептидной цепи содержит Fc-домен или его часть. В других аспектах этого варианта осуществления шестой домен второй полипептидной цепи содержит 6 C-концевых аминокислот легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23); и третий домен первой полипептидной цепи содержит Fc-домен или его часть.
В других вариантах осуществления остатки цистеина на первом или втором полипептиде, которые ответственны за образование дисульфидной связи, могут быть расположены вне первого, второго или третьего доменов на первой полипептидной цепи и вне четвертого, пятого и шестого домена на второй полипептидной цепи. В частности, остаток цистеина на первой полипептидной цепи может быть N-концевым относительно первого домена или он может быть C-концевым относительно первого домена. Остаток цистеина на первой полипептидной цепи может быть N-концевым для второго домена или он может быть C-концевым для второго домена. Остаток цистеина на первой полипептидной цепи может быть N-концевым для третьего домена или он может быть C-концевым для третьего домена. Кроме того, остаток цистеина на второй полипептидной цепи может быть N-концевым для четвертого домена или он может быть C-концевым для четвертого домена. Остаток цистеина на второй полипептидной цепи может быть N-концевым для пятого домена или он может быть C-концевым для пятого домена. Таким образом, остаток цистеина на второй полипептидной цепи может быть C-концевым для шестого домена или он может быть N-концевым для шестого домена. В конкретном аспекте дисульфидная связь может быть между по меньшей мере двумя остатками цистеина на первой полипептидной цепи и по меньшей мере двумя остатками цистеина на второй полипептидной цепи. В конкретном аспекте, где третий домен и шестой домен не содержат Fc-домен или его часть, остаток цистеина может быть на C-конце первой полипептидной цепи и на C-конце второй полипептидной цепи. Следует понимать, что настоящее изобретение охватывает любую комбинацию из указанных выше.
В конкретных вариантах осуществления изобретения, описанных выше, ковалентное диантитело по изобретению охватывает димеры молекул диантитела, где каждая молекула диантитела содержит первую и вторую полипептидные цепи. В некоторых аспектах этого варианта осуществления молекулы диантител могут быть ковалентно связаны с образованием димера, при условии, что ковалентная связь не является пептидной связью. В предпочтительных аспектах этого варианта осуществления ковалентная связь представляет собой дисульфидную связь между по меньшей мере одним остатком цистеина на первой полипептидной цепи каждой из молекул диантител димера. В более предпочтительных аспектах этого изобретения ковалентная связь представляет собой дисульфидную связь между по меньшей мере одним остатком цистеина на первой полипептидной цепи каждой из молекул диантител, образующих димер, где указанный по меньшей мере один остаток цистеина расположен в третьем домене каждой первой полипептидной цепи.
В определенных аспектах изобретения первый домен на первой полипептидной цепи может быть N-концевым для второго домена или он может быть C-концевым для второго домена. Первый домен на первой полипептидной цепи может быть N-концевым для третьего домена или он может быть C-концевым для третьего домена. Второй домен на первой полипептидной цепи может быть N-концевым для первого домена или он может быть C-концевым для первого домена. Более того, второй домен на первой полипептидной цепи может быть N-концевым для третьего домена или он может быть C-концевым для третьего домена. Таким образом, третий домен на первой полипептидной цепи может быть N-концевым для первого домена или он может быть C-концевым для первого домена. Третий домен на первой полипептидной цепи может быть N-концевым для второго домена или он может быть C-концевым для второго домена. Что касается второй полипептидной цепи, четвертый домен может быть N-концевым для пятого домена или он может быть C-концевым для пятого домена. Четвертый домен может быть N-концевым для шестого домен или он может быть C-концевым для шестого домена. Пятый домен на второй полипептидной цепи может быть N-концевым для четвертого домена или он может быть C-концевым для четвертого домена. Пятый домен на второй полипептидной цепи может быть N-концевым для шестого домена или он может быть C-концевым для шестого домена. Таким образом, шестой домен на второй полипептидной цепи может быть N-концевым для четвертого домена или он может быть C-концевым для четвертого домена. Шестой домен на второй полипептидной цепи может быть N-концевым для пятого домена или может быть C-концевым для пятого домена. Следует понимать, что настоящее изобретение охватывает любую комбинацию из указанных выше.
В определенных вариантах осуществления первый домен и второй домен могут быть расположены у C-конца третьего домена на первой полипептидной цепи; или первый домен и второй домен могут быть расположены у N-конца третьего домена на первой полипептидной цепи. Что касается второй полипептидной цепи, четвертый домен и пятый домен могут быть расположены у C-конца шестого домена, или четвертый домен и пятый домен могут быть расположены у N-конца шестого домена. В некоторых аспектах этого варианта осуществления настоящее изобретение относится к ковалентному биспецифическому диантителу, которое является димером молекул диантител, причем каждая молекула диантитела содержит первую и вторую полипептидные цепи, причем первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичную к первому эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичную ко второму эпитопу, и (iii) третий домен, содержащий Fc-домен или его часть, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа, и где третий домен расположен у N-конца как первого домена, так и второго домена; и где вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), и (iii) шестой домен, содержащий по меньшей мере один остаток цистеина, где четвертый и пятый домены ковалентно связаны, так что четвертый и пятый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь каждой молекулы диантитела ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен каждой молекулы диантитела ассоциируют с образованием первого связывающего участка (VL1)(VH1), который связывает первый эпитоп; где второй домен и четвертый домен каждой молекулы диантитела ассоциируют с образованием второго связывающего участка (VL2)(VH2), который связывает второй эпитоп.
В другом варианте осуществления настоящее изобретение относится к ковалентному тетраспецифическому диантителу, которое представляет собой димер молекул диантител, причем первая молекула диантитела содержит первую и вторую полипептидные цепи, и первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичную к первому эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичную ко второму эпитопу, и (iii) третий домен, содержащий Fc-домен или его часть, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа, и где третий домен расположен с N-конца как первого домена, так и второго домена; и где вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VHl), и (iii) шестой домен, содержащий по меньшей мере один остаток цистеина, где четвертый и пятый домены ковалентно связаны, так что четвертый и пятый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен ассоциируют с образованием первого участка связывания (VL1)(VH1), который связывает первый эпитоп; где второй домен и четвертый домен ассоциируют с образованием второго участка связывания (VL2)(VH2), который связывает второй эпитоп; и вторая молекула диантитела содержит первую и вторую полипептидные цепи, причем первая полипептидная цепь содержит (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи третьего иммуноглобулина (VL3), специфичную к третьему эпитопу, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи четвертого иммуноглобулина (VH4), специфичную к четвертому эпитопу, и (iii) третий домен, содержащий Fc-домен или его часть, где первый и второй домены ковалентно связаны, так что первый и второй домены не ассоциируют с образованием участка связывания эпитопа, и где третий домен расположен у N-конца как первого домена, так и второго домена; и где вторая полипептидная цепь содержит (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи четвертого иммуноглобулина (VL4), (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи третьего иммуноглобулина (VH3), и (iii) шестой домен, содержащий по меньшей мере один остаток цистеина, где четвертый и пятый домены ковалентно связаны, так что четвертый и пятый домены не ассоциируют с образованием участка связывания эпитопа; и где первая полипептидная цепь и вторая полипептидная цепь ковалентно связаны, при условии, что ковалентная связь не является пептидной связью; где первый домен и пятый домен ассоциируют с образованием первого участка связывания (VL3)(VH3), который связывает третий эпитоп; где второй домен и четвертый домен ассоциируют с образованием второго участка связывания (VL4)(VH4), который связывает четвертый эпитоп.
Как рассмотрено выше, домены отдельных полипептидных цепей связаны ковалентно. В конкретных аспектах ковалентная связь между первым и вторым доменом, первым и третьим доменом, вторым и третьим доменом, четвертым и пятым доменом, четвертым и шестым доменом и/или пятым и шестым доменом может представлять собой пептидную связь. В частности, первый и второй домены и четвертый и пятый домены могут быть разделены третьим доменом и шестым доменом, соответственно, или дополнительными аминокислотными остатками, при условии, что первый и второй и четвертый и пятый домены не ассоциируют с образованием связывающего участка. Количество аминокислотных остатков может составлять 0, 1, 2, 3, 4, 5, 6, 7, 8 или 9 аминокислотных остатков. В одном предпочтительном аспекте количество аминокислотных остатков между доменами равно 8.
В определенных аспектах изобретения домены первой и второй полипептидных цепей, содержащие Fc-домен, т.е. необязательно третий и шестой домены, соответственно, могут далее содержать шарнирный домен, так чтобы этот домен содержал шарнирную-Fc область. В альтернативных вариантах осуществления первая полипептидная цепь или вторая полипептидная цепь может содержать шарнирный домен и при этом не содержать Fc-домен. Тяжелые цепи, легкие цепи, шарнирные области, Fc-домены и/или шарнирные-Fc домены для применения в изобретении могут происходить из любого типа иммуноглобулина, включая IgA, IgD, IgE, IgG или IgM. В предпочтительном аспекте тип иммуноглобулина представляет собой IgG или любой его подтип, т.е. IgG1, IgG2, IgG3 или IgG4. В других аспектах иммуноглобулин, из которого происходят легкая и тяжелая цепи, является гуманизированным или химеризованным.
Кроме того, первый эпитоп и второй эпитоп и, где это применимо, третий эпитоп и четвертый эпитоп, с которыми связывается диантитело или молекула диантитела, могут представлять собой различные эпитопы из одного антигена или они могут представлять собой различные эпитопы из различных антигенов. Антигены могут представлять собой любую молекулу, против которой можно получать антитело, например, белки, нуклеиновые кислоты, бактериальные токсины, маркеры клеточной поверхности, аутоиммунные маркеры, вирусные белки, лекарственные средства и т.д. В конкретных аспектах по меньшей мере один участок связывания эпитопа в диантителе является специфичным к антигену на конкретной клетке, такой как B-клетка, T-клетка, фагоцитарная клетка, естественная киллерная (NK) клетка или дендритная клетка.
В некоторых аспектах настоящего варианта осуществления по меньшей мере один участок связывания эпитопа в диантителе или молекуле диантитела является специфичным к Fc-рецептору, который может представлять собой активирующий Fc-рецептор или ингибиторный Fc-рецептор. В конкретных аспектах Fc-рецептор представляет собой Fcγ-рецептор, и Fcγ-рецептор представляет собой FcγRI-, FcγRII- или FcγRIII-рецептор. В более предпочтительных аспектах FcγRIII-рецептор представляет собой FcγRIIIA(CD16A)-рецептор или FcγRIIIB(CD16B)-рецептор, и более предпочтительно FcγRIII-рецептор представляет собой FcγRIIIA(CD16A)-рецептор. В другом предпочтительном аспекте FcγRII-рецептор представляет собой FcγRIIA(CD32A)-рецептор или FcγRIIB(CD32B)-рецептор, и более предпочтительно FcγRIIB(CD32B)-рецептор. В особенно предпочтительном аспекте один участок связывания диантитела является специфичным к CD32B, а другой участок связывания является специфичным к CD16A. В конкретном варианте осуществления изобретения по меньшей мере один участок связывания эпитопа в диантителе или молекуле диантитела является специфичным к активирующему Fc-рецептору, и по меньшей мере один другой участок является специфичным к ингибиторному Fc-рецептору. В некоторых аспектах этого варианта осуществления активирующий Fc-рецептор представляет собой CD32A, и ингибиторный Fc-рецептор представляет собой CD32B. В других аспектах этого варианта осуществления активирующий Fc-рецептор представляет собой BCR, и ингибиторный Fc-рецептор представляет собой CD32B. В других аспектах этого варианта осуществления активирующий Fc-рецептор представляет собой IgERI, и ингибиторный Fc-рецептор представляет собой CD32B.
В случаях, когда один участок связывания эпитопа является специфичным к CD16A, домены VL и VH могут быть одинаковыми или сходными с доменами VL и VH антитела мыши 3G8, последовательность которого клонирована и представлена в настоящем документе. В других случаях, когда один участок связывания эпитопа является специфичным к CD32A, домены VL и VH могут быть одинаковыми или сходными с доменами VL и VH антитела мыши IV.3. В других случаях, когда один участок связывания эпитопа является специфичным к CD32B, домены VL и VH могут быть одинаковыми или сходными с доменами VL и VH антитела мыши 2B6, последовательность которого была клонирована и указана в настоящем документе. Следует понимать, что любой из доменов VL или VH антител 3G8, 2B6 и IV.3 можно использовать в любой комбинации. Настоящее изобретение также относится к биспецифическому диантителу или молекуле диантитела, где первый эпитоп является специфичным к CD32B, и второй эпитоп является специфичным к CD16A.
В других аспектах участок связывания эпитопа может быть специфичным к патогенному антигену. Как используют в настоящем документе, патогенный антиген представляет собой антиген, вовлеченный в определенное патогенное заболевание, включая злокачественную опухоль, инфекцию и аутоиммунное заболевание. Таким образом, патогенный антиген может представлять собой опухолевый антиген, бактериальный антиген, вирусный антиген или аутоиммунный антиген. Иллюстративные патогенные антигены включают, но не ограничиваются ими, липополисахарид, вирусные антигены, выбранные из группы, состоящей из вирусных антигенов вируса иммунодефицита человека, аденовируса, респираторного синцитиального вируса, вируса лихорадки западного Нила (например, антигены E16 и/или E53) и вируса гепатита, нуклеиновые кислоты (ДНК и РНК) и коллаген. Предпочтительно патогенный антиген представляет собой нейтрализующий антиген. В предпочтительном аспекте, где один участок связывания эпитопа является специфичным к CD16A или CD32A, другой участок связывания эпитопа является специфичным к патогенному антигену, исключая аутоиммунные антигены. В другом предпочтительном аспекте, где один участок связывания эпитопа является специфичным к CD32B, другой участок связывания эпитопа является специфичным к любому патогенному антигену. В конкретных вариантах осуществления молекула диантитела по изобретению связывает два различных антигена на одной клетке, например, один участок связывания антигена является специфичным к активирующему Fc-рецептору, а другой является специфичным к ингибиторному Fc-рецептору. В других вариантах осуществления молекула диантитела связывает два различных нейтрализующих вирус эпитопа, например, но не ограничиваясь ими, E16 и E53 вируса лихорадки западного Нила.
В другом варианте осуществления настоящего изобретения диантитела по изобретению можно применять для лечения различных заболеваний и нарушений. Таким образом, настоящее изобретение относится к способу лечения заболевания или нарушения, включающему введение пациенту, нуждающемуся в этом, эффективного количества ковалентного диантитела или молекулы диантитела по изобретению, в котором по меньшей мере один участок связывания является специфичным к патогенному антигену, такому как антиген, экспрессируемый на поверхности злокачественной клетки или на поверхности бактерии или вириона, и по меньшей мере один другой участок связывания является специфичным к Fc-рецептору, например, CD16A.
В другом варианте осуществления изобретение относится к способу лечения заболевания или нарушения, включающему введение пациенту, нуждающемуся в этом, эффективного количества диантитела или молекулы диантитела по изобретению, в котором по меньшей мере один участок связывания является специфичным к CD32B и по меньшей мере один другой участок связывания является специфичным к CD16A.
В другом варианте осуществления изобретение относится к способу индукции иммунной толерантности к патогенному антигену, включающему введение пациенту, нуждающемуся в этом, эффективного количества ковалентного диантитела или молекулы ковалентного диантитела по изобретению, в котором по меньшей мере один участок связывания является специфичным к CD32B и по меньшей мере один другой участок связывания является специфичным к указанному патогенному антигену. В аспектах этого варианта осуществления патогенный антиген может представлять собой аллерген или другую молекулу, к которой является желательной иммунная толерантность, такую как белок, экспрессируемый на трансплантированной ткани.
В другом варианте осуществления настоящее изобретение относится к способу детоксикации, включающему введение пациенту, нуждающемуся в этом, эффективного количества ковалентного диантитела или молекулы диантитела по изобретению, в котором по меньшей мере один участок связывания является специфичным к маркеру клеточной поверхности и по меньшей мере один другой участок связывания является специфичным к токсину. В конкретных аспектах вводимое диантитело по изобретению представляет собой диантитело, где один участок связывания является специфичным к маркеру клеточной поверхности, такому как Fc, а другой участок связывания является специфичным к бактериальному токсину или лекарственному средству. В одном аспекте маркер клеточной поверхности не встречается на эритроцитах.
3.1 ОПРЕДЕЛЕНИЯ
Если не определено иначе, подразумевают, что все термины данной области, обозначения и другие научные термины или терминология, используемые в настоящем документе, имеют значения, обычно понимаемые специалистами в области, к которой относится это изобретение. В некоторых случаях термины с обычно понятными значениями определены в настоящем документе для ясности и/или для простоты отсылки, и включение таких определений в настоящий документ не обязательно следует истолковывать как представляющее существенное отличие от того, как их обычно понимают в данной области. Для практики этого изобретения можно использовать, если нет иных указаний, общепринятые способы молекулярной биологии (включая рекомбинантные технологии), микробиологии, клеточной биологии, биохимии, химии нуклеиновых кислот и иммунологии, которые входят в пределы квалификации в данной области. Такие способы подробно объяснены в литературе, такой как Current Protocols in Immunology (J.E. Coligan et al., eds., 1999, включая дополнения по 2001 год); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987, включая дополнения по 2001 год); Molecular Cloning: A Laboratory Manual, третье издание (Sambrook and Russel, 2001); ПЦР: The Polimarase Chain Reaction, (Mullis et al., eds., 1994); The Immunoassay Handbook (D. Wild, ed., Stockton Press NY, 1994); Bioconjugate Techniques (Greg T. Hermanson, ed., Academic Press, 1996); Methods of Immunological Analysis (R. Masseyeff, W.H. Albert, and N.A. Staines, eds., Weinheim: VCH Verlags gesellschaft mbH, 1993), Harlow and Lane Using Antibodies: A Laboratory Manual Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y., 1999; и Beaucage et al. eds., Current Protocols in Nucleic Acid Chemistry John Wiley & Sons, Inc., New York, 2000).
Как используют в настоящем документе, термины "антитело" и "антитела" относятся к моноклональным антителам, полиспецифическим антителам, антителам человека, гуманизированным антителам, синтетическим антителам, химерным антителам, поликлональным антителам, камелизованным антителам, одноцепочечным Fv (scFv), одноцепочечным антителам, Fab-фрагментам, F(ab')-фрагментам, связанным дисульфидной связью Fv (sdFv), интраантителам и антиидиотипическим (анти-Id) антителам (включая, например, анти-Id- и анти-анти-Id-антитела к антителам по изобретению) и связывающим эпитоп фрагментам любого из указанных выше. В частности, антитела включают молекулы иммуноглобулинов и иммунологически активные фрагменты молекул иммуноглобулинов, т.е. молекулы, которые содержат антигенсвязывающий центр. Молекулы иммуноглобулинов могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса.
Как используют в настоящем документе, термины "иммуноспецифично связывает", "иммуноспецифично распознает", "специфично связывает", "специфично распознает" и аналогичные термины относятся к молекулам, которые специфично связываются с антигеном (например, эпитоп или иммунный комплекс) и не связываются специфично с другой молекулой. Молекула, которая специфично связывается с антигеном, может связываться с другими пептидами или полипептидами с более низкой аффинностью при определении, например, с помощью иммуноанализов, BIAcore или других анализов, известных в данной области. Предпочтительно, молекулы, которые специфично связываются с антигеном, не реагируют перекрестно с другими белками. Молекулы, которые специфично связывают антиген, можно идентифицировать, например, с помощью иммуноанализов, BIAcore или других способов, известных специалистам в данной области.
Как используют в настоящем документе, "иммунный комплекс" относится к структуре, которая образуется, когда по меньшей мере одна молекула-мишень и по меньшей мере один содержащий гетерологичную Fcγ-область полипептид связываются друг с другом с образованием более высокомолекулярного комплекса. Примерами иммунных комплексов являются комплексы антиген-антитело, которые могут быть либо растворимыми, либо в форме частиц (например, комплекс антиген/антитело на клеточной поверхности.).
Как используют в настоящем документе, термины "тяжелая цепь", "легкая цепь", "вариабельная область", "каркасная область", "константный домен" и т.д. имеют их обычное значение в области иммунологии и относятся к доменам во встречающихся в природе иммуноглобулинах и соответствующим доменам синтетических (например, рекомбинантных) связывающих белков (например, гуманизированных антител, одноцепочечных антител, химерных антител и т.д.). Основным структурным элементом встречающихся в природе иммуноглобулинов (например IgG) является тетрамер, имеющий две легкие цепи и две тяжелые цепи, обычно экспрессирующиеся в качестве гликопротеина массой приблизительно 150000 Да. N-концевая ("N") часть каждой цепи включает вариабельную область размером приблизительно от 100 до 110 или более аминокислот, ответственную, главным образом, за распознавание антигена. С-концевая ("C") часть каждой цепи определяет константную область, причем легкие цепи имеют один константный домен и тяжелые цепи обычно имеют три константных домена и шарнирную область. Таким образом, структура легких цепей молекулы IgG представляет собой n-VL-CL-с, и структура тяжелых цепей IgG представляет собой n-VH-CH1-H-CH2-CH3-c (где H представляет собой шарнирную область). Вариабельные области молекулы IgG состоят из определяющих комплементарность областей (CDR), которые содержат остатки, контактирующие с антигеном, и не-CDR-сегменты, называемые каркасными сегментами, которые, как правило, поддерживают структуру и определяют расположение петель CDR (хотя определенные каркасные остатки также могут контактировать с антигеном). Таким образом, домены VL и VH имеют структуру n-FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4-с.
При указании на связывающие белки или антитела (как широко определено в настоящем документе), отнесение аминокислот каждому из доменов осуществлено согласно определениям Kabat, Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md., 1987 и 1991). Аминокислоты из вариабельных областей зрелых тяжелой и легкой цепей иммуноглобулинов обозначены по положению аминокислоты в цепи. Kabat описал множество аминокислотных последовательностей для антител, идентифицировал консенсусные аминокислотные последовательности для каждой подгруппы и присвоил номер остатка каждой аминокислоте. Схема нумерации Kabat распространяется на антитела, не включенные в его справочник, путем выравнивания рассматриваемого антитела с одной из консенсусных последовательностей по Kabat, исходя из сохранившихся аминокислот. Этот способ присвоения номеров остатков стал стандартным в данной области, и он легко идентифицирует аминокислоты в эквивалентных положениях различных антител, включая химерные или гуманизированные варианты. Например, аминокислота в положении 50 легкой цепи антитела человека занимает эквивалентное положение аминокислоте в положении 50 легкой цепи антитела мыши.
Как используют в настоящем документе, термин "тяжелая цепь" используют для определения тяжелой цепи IgG-антитела. В целом нативном IgG тяжелая цепь содержит домены VH, CH1, шарнирной области, CH2 и CH3 иммуноглобулина. На протяжении настоящего описания нумерация остатков в тяжелой цепи IgG представляет собой нумерацию согласно индексу EU по Kabat et al., Sequences of Protein of Immunological Interest, 5th Ed. Public Health Service, NH1, MD (1991), включенной в настоящий документ в качестве ссылки в полном объеме. "Индекс EU по Kabat" относится к нумерации IgG1-антитела EU человека. Примеры аминокислотных последовательностей, содержащих шарнирную область, CH2- и CH3-домены IgG1 человека, представлены на ФИГ. 1A и 1B, как описано ниже. На ФИГ. 1A и 1B также указаны аминокислотные последовательности шарнирной области, доменов CH2 и CH3 тяжелых цепей IgG2, IgG3 и IgG4. Аминокислотные последовательности изотипов IgG2, IgG3 и IgG4 выравниваются с последовательностью IgG1 путем помещения первого и последнего остатков цистеина соответствующих шарнирных областей, которые образуют S-S-связи между тяжелыми цепями, в одинаковые положения. Для шарнирной области IgG2 и IgG3 не все остатки пронумерованы согласно индексу EU.
"Шарнирная область" или "шарнирный домен" обычно определяют как участок с Glu216 по Pro230 IgG1 человека. Пример аминокислотной последовательности шарнирной области IgG1 человека представлен на ФИГ. 1A (аминокислотные остатки на ФИГ. 1A пронумерованы согласно системе Kabat). Шарнирные области других изотипов IgG можно выравнивать с последовательностью IgG1, помещая первый и последний остатки цистеина, образующие связи S-S между тяжелыми цепями, в одинаковые положения, как показано на ФИГ. 1A.
Как используют в настоящем документе, термин "Fc-область", "Fc-домен" или аналогичные термины используют для определения C-концевой области тяжелой цепи IgG. Пример аминокислотной последовательности, содержащей IgG1 человека, представлен на ФИГ. 1B. Хотя границы могут немного варьировать при нумерации согласно системе Kabat, Fc-домен располагается от аминокислоты 231 до аминокислоты 447 (аминокислотные остатки на ФИГ. 1B пронумерованы согласно системе Kabat). На ФИГ. 1B также представлены примеры аминокислотных последовательностей Fc-областей изотипов IgG IgG2, IgG3 и IgG4.
Fc-область IgG содержит два константных домена, CH2 и CH3. CH2-домен Fc-области IgG человека обычно располагается с аминокислоты 231 по аминокислоту 341 согласно системе нумерации по Kabat (ФИГ. 1B). CH3-домен Fc-области IgG человека обычно располагается с аминокислоты 342 по аминокислоту 447 согласно системе нумерации Kabat (ФИГ. 1B). CH2-домен Fc-области IgG человека (также называемый "Cγ2"-доменом) является уникальным в том, что он не является близкорасположенным к другому домену. Вместо этого, между двумя CH2-доменами целого нативного IgG расположены две N-связанных разветвленных углеводных цепи.
Как используют в настоящем документе, термины "FcγR-связывающий белок", "антитело к FcγR" и "антитело против FcγR" используют взаимозаменяемо и они относятся к различным иммуноглобулин-подобным или происходящим из иммуноглобулинов белкам. "FcγR-связывающие белки" связывают FcγR путем взаимодействия с доменами VL и/или VH (что отличается от опосредуемого Fcγ связывания). Примеры связывающих FcγR белков включают полностью человеческие, поликлональные, химерные и гуманизированные антитела (например, содержащие 2 тяжелых и 2 легких цепи), их фрагменты (например, Fab-, Fab'-, F(ab')2 и Fv-фрагменты), бифункциональные или полифункциональные антитела (см., например, Lanzavecchia et al. (1987) "The Use Of Hybrid Hybridomas To Target Human Cytotoxic T Lymphocytes", Eur. J. Immunol. 17:105-111), одноцепочечные антитела (см., например, Bird et al. (1988) "Single-Chain Antigen-Binding Proteins", Science 242:423-26), слитые белки (например, слитые белки фагового дисплея), "миниантитела" (см., например, патент США № 5837821) и другие связывающие антиген белки, содержащие домен VL и/или VH или его фрагмент. В одном аспекте связывающий FcγRIIIA белок представляет собой "тетрамерное антитело", т.е. имеющее, главным образом, структуру встречающегося в природе IgG и содержащее вариабельный и константный домены, т.е. две легких цепи, содержащих VL-домен и константный домен легкой цепи, и две тяжелых цепи, содержащих VH-домен и шарнирную область тяжелой цепи и константные домены.
Как используют в настоящем документе, термин "антагонисты FcγR" и аналогичные термины относятся к белковым и небелковым веществам, включающим небольшие молекулы, которые осуществляют антагонизм по меньшей мере одному виду биологической активности FcγR, например, блокируют передачу сигнала. Например, молекулы по изобретению блокируют передачу сигнала путем блокирования связывания IgG с FcγR.
Как используют в настоящем документе, термин "производное" в контексте полипептидов или белков относится к полипептиду или белку, который содержит аминокислотную последовательность, которая изменена путем внесения замен, делеций или вставок аминокислотных остатков. Термин "производное", как используют в настоящем документе, также относится к полипептиду или белку, который является модифицированным, т.е. путем ковалентного присоединения любого типа молекулы к полипептиду или белку. В качестве неограничивающего примера, антитело может быть модифицированным, например, путем гликозилирования, ацетилирования, пегилирования, фосфорилирования, амидации, преобразования в производное с помощью известных защищающих/блокирующих групп, протеолитического расщепления, связывания с клеточным антигеном или другим белком и т.д. Производное полипептида или белка можно получать химическими модификациями с использованием способов, известных специалистам в данной области, включая, но не ограничиваясь ими, специфическое химическое расщепление, ацетилирование, формилирование, метаболический синтез туникамицина и т.д. Кроме того, производное полипептида или производное белка обладают сходной или идентичной функцией с полипептидом или белком, из которых они получены.
Как используют в настоящем документе, термин "производное" в контексте небелкового производного относится ко второй органической или неорганической молекуле, которая образована на основе структуры первой органической или неорганической молекулы. Производное органической молекулы включает, но не ограничивается ими, молекулу, модифицированную, например, путем добавления или удаления гидроксильной, метильной, этильной, карбоксильной группы или аминогруппы. Органический молекула также может быть этерифицированной, алкилированной и/или фосфорилированной.
Как используют в настоящем документе, термин "молекула диантитела" относится к комплексу из двух или более полипептидных цепей или белков, каждый из которых содержит по меньшей мере один VL-домен и один VH-домен или его фрагмент, где оба домена находятся в одной полипептидной цепи. В определенных вариантах осуществления "молекула диантитела" включает молекулы, содержащие Fc-домен или шарнирный-Fc домен. Указанные полипептидные цепи в комплексе могут быть одинаковыми или могут отличаться, т.е. молекула диантитела может быть гомомультимерной или гетеромультимерной. В конкретных аспектах "молекула диантитела" включает димеры или тетрамеры указанных полипептидных цепей, содержащих как VL-домен, так и VH-домен. Отдельные полипептидные цепи, содержащие мультимерные белки, могут быть ковалентно связаны по меньшей мере с одним другим пептидом мультимера межцепочечными дисульфидными связями.
Как используют в настоящем документе, термины "нарушение" и "заболевание" используют взаимозаменяемо для обозначения состояния у индивидуума. В частности, термин "аутоиммунное заболевание" используют взаимозаменяемо с термином "аутоиммунное нарушение" для обозначения состояния у индивидуума, характеризующегося повреждением клеток, ткани и/или органа, вызываемым иммунологической реакцией индивидуума на его собственные клетки, ткани и/или органы. Термин "воспалительное заболевание" используют взаимозаменяемо с термином "воспалительное нарушение" для обозначения состояния у индивидуума, характеризующегося воспалением, предпочтительно хроническим воспалением. Аутоиммунные нарушения могут быть ассоциированы или могут не быть ассоциированы с воспалением. Более того, воспаление может быть вызвано аутоиммунным нарушением или может не быть вызвано им. Таким образом, определенные нарушения могут быть охарактеризованы в качестве как аутоиммунных, так и воспалительных нарушений.
"Идентичные полипептидные цепи", как используют в настоящем документе, также относятся к полипептидные цепям, имеющим практически идентичную аминокислотную последовательность, например, включая цепи, имеющие отличия в одной или нескольких аминокислотах, предпочтительно консервативные аминокислотные замены, так что активность двух полипептидных цепей значительно не отличается.
Как используют в настоящем документе, термин "злокачественная опухоль" относится к новообразованию или опухоли, являющимся следствием аномального неконтролируемого роста клеток. Как используют в настоящем документе, злокачественная опухоль конкретно включает лейкозы и лимфомы. В некоторых вариантах осуществления злокачественная опухоль относится к низкозлокачественной опухоли, которая остается локализованной. В других вариантах осуществления злокачественная опухоль относится к высокозлокачественной опухоли, которая вторгается и разрушает соседние структуры организма и распространяется в отдаленные области. В некоторых вариантах осуществления злокачественная опухоль ассоциирована с конкретным антигеном злокачественной опухоли.
Как используют в настоящем документе, термин "иммуномодулирующее средство" и его варианты относится к средству, которое модулирует иммунную систему хозяина. В определенных вариантах осуществления иммуномодулирующее средство представляет собой иммунодепрессивное средство. В некоторых других вариантах осуществления иммуномодулирующее средство представляет собой иммуностимулирующее средство. Иммуномодулирующие средства включают, но не ограничиваются ими, небольшие молекулы, пептиды, полипептиды, слитые белки, антитела, неорганические молекулы, миметики и органические молекулы.
Как используют в настоящем документе, термин "эпитоп" относится к фрагменту полипептида или белка или небелковой молекулы, обладающему антигенной или иммуногенной активностью у животного, предпочтительно у млекопитающего, и наиболее предпочтительно у человека. Эпитоп, обладающий иммуногенной активностью, представляет собой фрагмент полипептида или белка, который вызывает антительный ответ у животного. Эпитоп, обладающий антигенной активностью, представляет собой фрагмент полипептида или белка, с которым антитело иммуноспецифически связывается, при определении любым способом, хорошо известным специалисту в данной области, например, способами иммуноанализа. Антигенные эпитопы не должны быть обязательно иммуногенными.
Как используют в настоящем документе, термин "фрагмент" относится к пептиду или полипептиду, содержащему аминокислотную последовательность по меньшей мере из 5 соседних аминокислотных остатков, по меньшей мере из 10 соседних аминокислотных остаток, по меньшей мере из 15 соседних аминокислотных остатков, по меньшей мере из 20 соседних аминокислотных остатков, по меньшей мере из 25 соседних аминокислотных остатков, по меньшей мере из 40 соседних аминокислотных остатков, по меньшей мере из 50 соседних аминокислотных остатков, по меньшей мере из 60 соседних аминокислотных остатков, по меньшей мере из 70 соседних аминокислотных остатков, по меньшей мере из 80 соседних аминокислотных остатков, по меньшей мере из 90 соседних аминокислотных остатков, по меньшей мере из 100 соседних аминокислотных остатков, по меньшей мере из 125 соседних аминокислотных остатков, по меньшей мере из 150 соседних аминокислотных остатков, по меньшей мере из 175 соседних аминокислотных остатков, по меньшей мере из 200 соседних аминокислотных остатков или по меньшей мере из 250 соседних аминокислотных остатков аминокислотной последовательности другого полипептида. В конкретном варианте осуществления фрагмент полипептида сохраняет по меньшей мере одну функцию полипептида.
Как используют в настоящем документе, термины "нуклеиновые кислоты" и "нуклеотидные последовательности" включают молекулы ДНК (например, кДНК или геномной ДНК), молекулы РНК (например, мРНК), комбинации молекул ДНК и РНК или гибридные молекулы ДНК/РНК, и аналоги молекул ДНК или РНК. Такие аналоги можно получать с использованием, например, нуклеотидных аналогов, которые включают, но не ограничиваются ими, инозин или тритилированные основания. Такие аналоги также могут содержать молекулы ДНК или РНК, содержащие модифицированные основные цепи, которые придают молекулам благоприятные свойства, например, такие как устойчивость к нуклеазе или повышенная способность проходить через клеточные мембраны. Нуклеиновые кислоты или нуклеотидные последовательности могут быть одноцепочечными, двухцепочечными или могут содержать как одноцепочечные, так и двухцепочечные части, и они могут содержать трехцепочечные части, однако предпочтительно они представляют собой двухцепочечную ДНК.
Как используют в настоящем документе, "терапевтически эффективное количество" относится к количеству лекарственного средства, достаточному для лечения или управления течением заболевания или нарушения. Терапевтически эффективное количество может относиться к количеству лекарственного средства, достаточному для замедления или сведения к минимуму возникновения заболевания, например, замедления или сведения к минимуму распространения злокачественной опухоли. Терапевтически эффективное количество также может относиться к количеству лекарственного средства, которое обеспечивает терапевтическую пользу при лечении или управлении течением заболевания. Кроме того, терапевтически эффективное количество в отношении лекарственного средства по изобретению означает количество лекарственного средства отдельно или в комбинации с другими способами лечения, которое обеспечивает терапевтическую пользу при лечении или управлении течением заболевания.
Как используют в настоящем документе, термины "профилактическое средство" и "профилактические средства" относятся к любому средству(ам), которое можно использовать для предупреждения нарушения или предупреждения рецидива или распространения нарушения. Профилактически эффективное количество может относиться к количеству профилактического средства, достаточному для предупреждения рецидива или распространения гиперпролиферативного заболевания, в частности злокачественной опухоли, или его возникновения у пациента, включая, но не ограничиваясь ими, пациентов, предрасположенных к гиперпролиферативному заболеванию, например, пациентов, генетически предрасположенных к злокачественной опухоли или ранее подвергнутых воздействию канцерогенов. Профилактически эффективное количество также может относиться к количеству профилактического средства, которое обеспечивает профилактическую пользу в отношении предупреждения заболевания. Кроме того, профилактически эффективное количество в отношении профилактического средства по изобретению означает количество профилактического средства отдельно или в комбинации с другими средствами, которое обеспечивает профилактическую пользу для предупреждения заболевания.
Как используют в настоящем документе, термины "предупреждать", "предупреждающий" и "предупреждение" относятся к предупреждению рецидива или возникновения одного или нескольких симптомов нарушения у индивидуума в результате введения профилактического или терапевтического средства.
Как используют в настоящем документе, термин "в комбинации" относится к применению более одного профилактического и/или терапевтического средства. Применение термина "в комбинации" не ограничивает порядок, в котором профилактические и/или терапевтические средства вводят индивидууму с нарушением. Первое профилактическое или терапевтическое средство можно водить до (например, за 5 минут, 15 минут, 30 минут, 45 минут, 1 час, 2 часа, 4 часа, 6 часов, 12 часов, 24 часа, 48 часов, 72 часа, 96 часов, 1 неделю, 2 недели, 3 недели, 4 недели, 5 недель, 6 недель, 8 недель или 12 недель до), одновременно или после (например, через 5 минут, 15 минут, 30 минут, 45 минут, 1 час, 2 часа, 4 часа, 6 часа, 12 часов, 24 часа, 48 часов, 72 часа, 96 часов, 1 неделю, 2 недели, 3 недели, 4 недели, 5 недель, 6 недель, 8 недель или 12 недель после) введения второго профилактического или терапевтического средства индивидууму с нарушением.
"Эффекторная функция", как используют в настоящем документе, означает биохимическое явление, которое является результатом взаимодействия Fc-области антитела с Fc-рецептором или антигеном. Эффекторные функции включают, но не ограничиваются ими, антителозависимую клеточноопосредуемую цитотоксичность (ADCC), антителозависимый клеточноопосредуемый фагоцитоз (ADCP) и комплементзависимую цитотоксичность (CDC). Эффекторные функции включают как функции, которые действуют после связывания антигена, так и функции, которые действую независимо от связывания антигена.
"Эффекторная клетка", как используют в настоящем документе, означает клетку иммунной системы, которая экспрессирует один или несколько Fc-рецепторов и опосредует одну или несколько эффекторных функций. Эффекторные клетки включают, но не ограничиваются ими, моноциты, макрофаги, нейтрофилы, дендритные клетки, эозинофилы, тучные клетки, тромбоциты, B-клетки, большие гранулярные лимфоциты, клетки Лангерганса, естественные киллерные (NK) клетки, и они могут быть из любого организма, включая, но не ограничиваясь ими, людей, мышей, крыс, кроликов и обезьян.
Как используют в настоящем документе, термин "специфично связывает иммунный комплекс" и аналогичные термины относятся к молекулам, которые специфично связываются с иммунным комплексом и не связываются специфично с другой молекулой. Молекула, которая специфично связывается с иммунным комплексом, может связываться с другими пептидами или полипептидами с более низкой аффинностью при определении, например, способами иммуноанализов, BIAcore или другими анализами, известными в данной области. Предпочтительно молекулы, которые специфично связывают иммунный комплекс, не реагируют перекрестно с другими белками. Молекулы, которые специфично связывают иммунный комплекс, можно идентифицировать, например, способами иммуноанализов, BIAcore или другими способами, известными специалистам в данной области.
"Стабильный слитый белок", как используют в настоящем документе, относится к слитому белку, который подвергается деградации на уровне от минимального до не поддающегося детекции в ходе продукции и/или хранении, как оценивают с использованием распространенных биохимических и функциональных анализов, известных специалисту в данной области, и который можно хранить в течение длительного времени без потери биологической активности, например, связывания с FcγR.
4. КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
ФИГ. 1A-B АМИНОКИСЛОТНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ CH1-ОБЛАСТИ, ШАРНИРНОЙ ОБЛАСТИ И Fc-ОБЛАСТИ IgG ЧЕЛОВЕКА
На фигуре 1 представлены аминокислотные последовательности домена шарнирной области (A) и Fc-домена (B) IgG1, IgG2, IgG3 и IgG4 человека. (Шарнирный домен IgG1 (SEQ ID NO: 1); шарнирный домен IgG2 (SEQ ID NO: 2); шарнирный домен IgG3 (SEQ ID NO: 3); шарнирный домен IgG4 (SEQ ID NO: 4); Fc-домен IgG1 (SEQ ID NO: 5); Fc-домен IgG2 (SEQ ID NO: 6); Fc-домен IgG3 (SEQ ID NO: 7); Fc-домен IgG1 (SEQ ID NO: 8)). Аминокислотные остатки, представленные на ФИГ. 1A и 1B, пронумерованы согласно системе нумерации EU по Kabat. Изотипические последовательности выровнены с последовательностью IgG1 путем помещения первого и последнего остатков цистеина соответствующих шарнирных областей, которые образуют S-S-связи между тяжелыми цепями, в одинаковые положения. Для фигуры 1B остатки в CH2-домене обозначены с помощью +, а остатки в CH3-домене указаны с помощью ~.
ФИГ. 2 СХЕМАТИЧНОЕ ПРЕДСТАВЛЕНИЕ ПОЛИПЕПТИДНЫХ ЦЕПЕЙ КОВАЛЕНТНЫХ БИФУНКЦИОНАЛЬНЫХ ДИАНТИТЕЛ
Полипептиды ковалентного бифункционального диантитела состоят из VL-домена антитела и VH-домена антитела, разделенных коротким пептидным линкером. Линкер из 8 аминокислотных остатков препятствует самосборке единой полипептидной цепи в конструкции scFv, и, вместо этого, преобладают взаимодействия между VL- и VH-доменами различных полипептидных цепей. Было создано 4 конструкции (каждая конструкция представлена от N-конца ("n"), с левой стороны конструкции, к C-концу ("c"), с правой стороны фигуры): конструкция (1) (SEQ ID NO: 9) содержала n-VL домен Hu2B6-линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - и C-концевую последовательность (LGGC)-c; конструкция (2) (SEQ ID NO: 11) содержала n-VL-домен Hu3G8 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - и C-концевую последовательность (LGGC)-c; конструкция (3) (SEQ ID NO: 12) содержала n-VL-домен Hu3G8 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - и C-концевую последовательность (LGGC)-c; конструкция (4) (SEQ ID NO: 13) содержала n-VL-домен Hu2B6 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - и C-концевую последовательность (LGGC)-c.
ФИГ. 3 АНАЛИЗ SDS-PAGE ДИАНТИТЕЛ, ПОДВЕРГНУТЫХ АФФИННОЙ ОЧИСТКЕ
Диантитела, подвергнутые аффинной очистке, подвергали анализу SDS-PAGE в восстанавливающих (дорожки 1-3) или невосстанавливающих (дорожки 4-6) условиях. Указана приблизительная молекулярная масса стандарта (между дорожками 3 и 4). Дорожки 1 и 4, h3G8 CMD; дорожки 2 и 5, h2B6 CMD; и дорожки 3 и 6, h2B6-h3G8 CBD.
ФИГ. 4A-B SEC-АНАЛИЗ ДИАНТИТЕЛ, ПОДВЕРГНУТЫХ АФФИННОЙ ОЧИСТКЕ
Подвергнутые аффинной очистке диантитела подвергали SEC-анализу. (A) Профиль элюирования известных стандартов: полноразмерный (~150 кДа), Fab-фрагмент IgG (~50 кДа) и scFv (~30 кДа); (B) Профиль элюирования h2b6 CMD, h3G8 CMD и h2B6-h3G8 CBD.
ФИГ. 5 СВЯЗЫВАНИЕ h2B6-h3G8 CBD С sCD32B И sCD16A
Связывание h2B6-h3G8 CBD с sCD32B и sCD16A анализировали в сэндвич-ELISA. В качестве белка-мишени использовали sCD32B. Вторичный зонд представлял собой sCD16A, конъюгированный с HRP. h3G8 CMD, который связывает CD16A, использовали в качестве контроля.
ФИГ. 6A-C АНАЛИЗ BIACORE СВЯЗЫВАНИЯ ДИАНТИТЕЛА С sCD16A, sCD32B И sCD32B
Связывание h2B6-h3G8 CBD, h2B6 CMD и h3G8 CMD с sCD16A, sCD32B и sCD32A (отрицательный контроль) анализировали с помощью анализа SPR. Также тестировали h3G8 scFv в качестве контроля. (A) Связывание с sCD16; (B) связывание с sCD32B и (C) связывание с sCD32A. Диантитела инъецировали в концентрации 100 нМ, и scFv в концентрации 200 нМ, над рецепторными поверхностями со скоростью потока 50 мл/мин в течение 60 секунд.
ФИГ. 7A-C АНАЛИЗ BIACORE СВЯЗЫВАНИЯ ДИАНТИТЕЛА С sCD16A И SCD32B
Связывание h2B6-h3G8 CBD, h2B6 CMD и h3G8 CMD с sCD16A и sCD32B анализировали с помощью анализа SPR. Также тестировали h3G8 scFv в качестве контроля. (A) Связывание h3G8 CMD с sCD16A; (B) связывание h2B6-h3G8 CBD с sCD16A; (C) связывание h3G8 scFv с sCD16A; (D) связывание h2B6 CMD с sCD32B; и (E) связывание h2B6-h3G8 CBD с sCD32B. Диантитела инъецировали в концентрациях 6,25-200 нМ над рецепторными поверхностями со скоростью потока 70 мл/мин в течение 180 секунд.
ФИГ. 8 СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ ВЗАИМОДЕЙСТВИЯ ПОЛИПЕПТИДНЫХ ЦЕПЕЙ, СОДЕРЖАЩИХ VL- И VH-ДОМЕНЫ, С ОБРАЗОВАНИЕМ МОЛЕКУЛЫ КОВАЛЕНТНОГО БИСПЕЦИФИЧЕСКОГО ДИАНТИТЕЛА
NH2 и COOH соответствуют N-концу и С-концу, соответственно, каждой полипептидной цепи. S представляет собой C-концевой остаток цистеина на каждой полипептидной цепи. VL и VH указывают на вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, соответственно. Точечная и пунктирная линии представлены для различения двух полипептидных цепей и, в частности, соответствуют линкерным частям указанных цепей. h2B6 Fv и h3G8 Fv указывают на участок связывания эпитопа, специфичный к CD32B и CD16, соответственно.
ФИГ. 9 СХЕМАТИЧНОЕ ПРЕДСТАВЛЕНИЕ ПОЛИПЕПТИДНЫХ ЦЕПЕЙ, СОДЕРЖАЩИХ Fc-ДОМЕНЫ КОВАЛЕНТНЫХ БИСПЕЦИФИЧЕСКИХ ДИАНТИТЕЛ
Представление полипептидных конструкций молекул диантител по изобретению (каждая конструкция описана от N-конца ("n"), с левой стороны конструкции, к С-концу ("c"), с правой стороны фигуры). Конструкция (5) (SEQ ID NO: 14) содержала n-VL домен Hu2B6 - первый линкер (GGGSGGGG (SEQ ID NO: 10)) - VH домен Hu3G8 - второй линкер (LGGC) - и C-концевой Fc-домен IgG1 человека-c; конструкция (6) (SEQ ID NO: 15) содержала n-VL домен Hu3G8 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH домен Hu2B6 - и второй линкер (LGGC) - и C-концевой Fc домен IgG1 человека-c; конструкция (7) (SEQ ID NO: 16) содержала n-VL домен Hu2B6 - первый линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - и C-концевую последовательность (LGGCFNRGEC) (SEQ ID NO: 17)-c; конструкция (8) (SEQ ID NO: 18) содержала n-VL домен Hu3G8 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH домен Hu2B6 - и второй линкер (LGGC) - и C-концевой шарнирный/Fc-домен IgG1 человека (с аминокислотной заменой A215V)-c.
ФИГ. 10 СВЯЗЫВАНИЕ МОЛЕКУЛ ДИАНТИТЕЛ, СОДЕРЖАЩИХ Fc-ДОМЕНЫ, С sCD32B И sCD16A
Связывание молекул диантител, содержащих Fc-домены, с sCD32B и sCD16A анализировали в сэндвич-ELISA. Анализированные диантитела получали с помощью 3 рекомбинантных экспрессирующих систем: котрансфекция pMGX669 и pMGX674, экспрессирующих конструкции 1 и 6, соответственно; котрансфекция pMGX667 и pMGX676, экспрессирующих конструкции 2 и 5, соответственно; и котрансфекция pMGX674 и pMGX676, экспрессирующих конструкции 5 и 6, соответственно. В качестве белка-мишени использовали sCD32B. Вторичный зонд представлял собой sCD16A, конъюгированный с HRP.
ФИГ. 11 СХЕМАТИЧЕСКОЕ ПРЕДСТАВЛЕНИЕ ВЗАИМОДЕЙСТВИЯ ДВУХ ПОЛИПЕПТИДНЫХ ЦЕПЕЙ, КАЖДАЯ ИЗ КОТОРЫХ СОДЕРЖАЛА Fc-ДОМЕН, С ОБРАЗОВАНИЕМ ДВУХВАЛЕНТНОГО КОВАЛЕНТНОГО ДИАНТИТЕЛА
NH2 и COOH представляют собой N-конец и С-конец, соответственно, каждой полипептидной цепи. S представляет собой по меньшей мере одну дисульфидную связь между остатком цистеина во второй линкерной последовательности каждой полипептидной цепи. VL и VH указывают на вариабельный домен легкой цепи и вариабельный домен тяжелой цепи. Точечная и пунктирная линии представлены для различения двух полипептидных цепей и, в частности, соответствуют первым линкерным частям указанных цепей. CH2 и CH3 представляют собой константные домены CH2 и CH3 Fc-домена. h2B6 Fv и h3G8 Fv указывают на участок связывания эпитопа, специфичный к CD32B и CD16, соответственно.
ФИГ. 12 СВЯЗЫВАНИЕ МОЛЕКУЛ ДИАНТИТЕЛ, СОДЕРЖАЩИХ ШАРНИРНЫЕ/Fc-ДОМЕНЫ C sCD32B И sCD16A
Связывание молекул диантител, содержащих Fc-домены, с sCD32B и sCD16A анализировали в сэндвич-ELISA. Анализированные диантитела получали с помощью 4 рекомбинантных экспрессирующих систем: котрансфекция pMGX669 + pMGX674, экспрессирующих конструкции 1 и 6, соответственно; котрансфекция pMGX669 + pMGX678, экспрессирующих конструкции 2 и 8, соответственно; котрансфекция pMGX677 + pMGX674, экспрессирующих конструкции 7 и 6, соответственно; и котрансфекция pMGX677 + pMGX678, экспрессирующих конструкции 7 и 8, соответственно. В качестве белка-мишени использовали sCD32B. Вторичный зонд представлял собой sCD16A, конъюгированный с HRP.
ФИГ. 13 СХЕМАТИЧНОЕ ПРЕДСТАВЛЕНИЕ ВЗАИМОДЕЙСТВИЯ ПОЛИПЕПТИДНЫХ ЦЕПЕЙ С ОБРАЗОВАНИЕМ ТЕТРАМЕРНОЙ МОЛЕКУЛЫ ДИАНТИТЕЛА
NH2 и COOH представляют собой N-конец и С-конец, соответственно, каждой полипептидной цепи. S представляет собой по меньшей мере одну дисульфидную связь между остатком цистеина во второй линкерной последовательности - содержащей Fc "более тяжелой" полипептидной цепи, и остатком цистеина на C-концевой последовательности - не содержащей Fc "более легкой" полипептидной цепи. VL и VH указывают на вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, соответственно. Точечная и пунктирная линии представлены для различения полипептидных цепей и, в частности, соответствуют первым линкерным частям указанных более тяжелых цепей или линкеру указанных более легких цепей. CH2 и CH3 представляют собой константные домены CH2 и CH3 Fc-домена. h2B6 Fv и h3G8 Fv указывают на участок связывания эпитопа, специфичный к CD32B и CD16, соответственно.
ФИГ. 14 СХЕМАТИЧНОЕ ПРЕДСТАВЛЕНИЕ ПОЛИПЕПТИДНЫХ ЦЕПЕЙ, СОДЕРЖАЩИХ Fc-ДОМЕНЫ, КОТОРЫЕ ОБРАЗУЮТ КОВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ ДИАНТИТЕЛА
Представление полипептидных конструкций, которые образуют молекулы диантител по изобретению (каждая конструкция представлена от N-конца ("n"), с левой стороны конструкции, к С-концу ("c"), с правой стороны фигуры). Конструкция (9) (SEQ ID NO: 19) содержала n-шарнирный/Fc-домен IgG1 человека - VL-домен Hu3G8 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - линкер (GGGSGGGG (SEQ ID NO: 10))- и C-концевую последовательность LGGC-c; конструкция (10) (SEQ ID NO: 20) содержала n-Fc-домен IgG1 человека - VL-домен Hu3G8 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - линкер (GGGSGGGG (SEQ ID NO: 10))- и C-концевую последовательность LGGC-c; конструкция (11) (SEQ ID NO: 21) содержала n-VL-домен Hu2B6 (G105C) - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - и C-концевой шарнирный/Fc-домен IgG1 человека с аминокислотной заменой A215V-c; конструкция (12) (SEQ ID NO: 22) содержала n-VL-домен Hu3G8 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 (G44C) - и C-концевую последовательность FNRGEC (SEQ ID NO: 23)-c.
ФИГ. 15A-B SDS-PAGE И ВЕСТЕРН-БЛОТ АНАЛИЗ АФФИННОСТИ ТЕТРАМЕРНЫХ АНТИТЕЛ
Диантитела, продуцированные рекомбинантными экспрессирующими системами, котрансфицированными векторами, экспрессирующими конструкции 10 и 1, конструкции 9 и 1 и конструкции 11 и 12, подвергали SDS-PAGE-анализу в невосстанавливающих условиях (A) и вестерн-блот анализу с использованием антител козы против H+L IgG1 человека в качестве зонда (B). Белки на геле SDS-PAGE визуализировали с помощью Simply Blue Safestain (Invitrogen). На обеих панелях A и B молекулы диантител, содержащие конструкции 10 и 1, конструкции 9 и 1 и конструкции 11 и 12A, приведены на дорожках 1, 2 и 3, соответственно.
ФИГ. 16 СВЯЗЫВАНИЕ МОЛЕКУЛ ДИАНТИТЕЛ, СОДЕРЖАЩИХ Fc-ДОМЕНЫ И ВСТРОЕННЫЕ СПОСОБАМИ ИНЖЕНЕРИИ МЕЖЦЕПОЧЕЧНЫЕ ДИСУЛЬФИДНЫЕ СВЯЗИ, С sCD32B И sCD16A
Связывание молекул диантител, содержащих Fc-домены и встроенные способами инженерии дисульфидные связи между "более легкой" и "более тяжелой" полипептидными цепями, с sCD32B и sCD16A анализировали в сэндвич-ELISA. Анализированные диантитела продуцировались 3 рекомбинантными экспрессирующими системами: экспрессирующие конструкции 1 и 10, экспрессирующие конструкции 1 и 9 и экспрессирующие конструкции 11 и 12, соответственно. В качестве белка-мишени использовали sCD32B. Вторичный зонд представлял собой sCD16A, конъюгированный с HRP. Связывание h3G8 использовали в качестве контроля.
ФИГ. 17 СХЕМАТИЧНОЕ ПРЕДСТАВЛЕНИЕ ПОЛИБЕЛКОВОГО ПРЕДШЕСТВЕННИКА МОЛЕКУЛЫ ДИАНТИТЕЛА И СХЕМАТИЧНОЕ ПРЕДСТАВЛЕНИЕ ПОЛИПЕПТИДНЫХ ЦЕПЕЙ, СОДЕРЖАЩИХ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА И/ИЛИ ШАРНИРНЫЕ ДОМЕНЫ
Представление полипептидных конструкций, которые содержат молекулы диантител по изобретению (каждая конструкция представлена от N-конца ("n"), с левой стороны конструкции, к С-концу ("c"), с правой стороны фигуры). Конструкция (13) (SEQ ID NO: 95) содержала n-VL домен 3G8 - первый линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен 2.4G2VH - второй линкер (LGGC) - участок распознавания фурином (RAKR (SEQ ID NO: 93)) - VL-домен 2.4G2 - третий линкер (GGGSGGG (SEQ ID NO: 10) - VH-домен 3G8 - и C-концевой домен LGGC; (нуклеотидная последовательность, кодирующая SEQ ID NO: 95, представлена в SEQ ID NO: 96). Конструкция (14) (SEQ ID NO: 97) содержала n-VL домен 3G8 - первый линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен 2.4G2VH - второй линкер (LGGC) - участок распознавания фурином (RAKR (SEQ ID NO: 93))- участок FMD (протеаза C3 вируса ящура) - VL домен 2.4G2 - третий линкер (GGGSGGG (SEQ ID NO: 10) - VH домен 3G8 - и C-концевой домен LGGC; (нуклеотидная последовательность, кодирующая SEQ ID NO: 97, представлена в SEQ ID NO: 98). Конструкция (15) (SEQ ID NO: 99) содержала n-VL домен Hu2B6 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - и C-концевой домен FNRGEC (SEQ ID NO: 23); (нуклеотидная последовательность, кодирующая SEQ ID NO: 99, представлена в SEQ ID NO: 100). Конструкция (16) (SEQ ID NO: 101) содержала n-VL домен Hu3G8 - линкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6- и C-концевой домен VEPKSC (SEQ ID NO: 77); (нуклеотидная последовательность, кодирующая SEQ ID NO: 101, представлена в SEQ ID NO: 102).
ФИГ. 18 СВЯЗЫВАНИЕ МОЛЕКУЛ ДИАНТИТЕЛ, ПРОИСХОДЯЩИХ ИЗ МОЛЕКУЛЫ ПОЛИБЕЛКОВОГО ПРЕДШЕСТВЕННИКА, С mCD32B И sCD16A
Связывание молекул диантител, происходящих из конструкции 13 молекулы полибелкового предшественника (SEQ ID NO: 95), с CD32B мыши (mCD32B) и растворимым CD16A (sCD16A) анализировали в сэндвич-ELISA. В качестве белка-мишени использовали mCD32B. Вторичный зонд представлял собой sCD16A, конъюгированный с биотином.
ФИГ. 19 СВЯЗЫВАНИЕ МОЛЕКУЛ ДИАНТИТЕЛ, СОДЕРЖАЩИХ ЦЕПЬ ЛЯМБДА И/ИЛИ ШАРНИРНЫЕ ДОМЕНЫ С sCD32B И sCD16A
Связывание молекул диантител, содержащих домены, происходящие из C-конца легкой цепи лямбда человека и/или шарнирного домена IgG, с sCD32B и sCD16A анализировали и сравнивали со связыванием диантитела, содержащего конструкции 1 и 2 (ФИГ. 5), в сэндвич-ELISA. Анализированные диантитела продуцировались рекомбинантной экспрессирующей системой, экспрессирующей конструкции 15 и 16 (SEQ ID NO: 99 и SEQ ID NO: 101, соответственно). В качестве белка-мишени использовали sCD32B. Вторичный зонд представлял собой sCD16A, конъюгированный с HRP. Столбики с мелкими прямоугольниками соответствуют комбинации конструкций 15/16, в то время как столбики с крупными прямоугольниками соответствуют комбинации конструкций 1/2.
ФИГ. 20 СХЕМАТИЧНОЕ ПРЕДСТАВЛЕНИЕ DART 2B6/4420, СВЯЗАННОГО С CD32B, РАСПОЛОЖЕННЫМ НА ПОВЕРХНОСТИ КЛЕТКИ, И КОНЪЮГИРОВАННОЙ С ФЛУОРЕСЦЕИНОМ МОЛЕКУЛОЙ
На диаграмме представлена гибкость "универсального адаптора", плеча DART против флуоресцеина, а также возможность замены на другие специфичности для плеча 2B6. V-области заключены в рамки, линкеры GGGSGGGG (SEQ ID NO: 10) представлены в качестве линий, указана дисульфидная связь, соединяющая две цепи. Компоненты одной цепи представлены синим цветом, а компоненты другой цепи окрашены розовым. N, амино-конец; C, карбокси-конец; FL, флуоресцеин, VL, вариабельная область легкой цепи; VH, вариабельная область тяжелой цепи.
ФИГ. 21 (ПАНЕЛИ A И B) DART 2B6/4420 СПЕЦИФИЧНО СВЯЗЫВАЕТСЯ С КОНЪЮГИРОВАННЫМИ С ФЛУОРЕСЦЕИНОМ МОЛЕКУЛАМИ И МОЖЕТ ОДНОВРЕМЕННО СВЯЗЫВАТЬ CD32B.
(A) 2B6/4420 или 2B6/3G8 связывали с планшетами для ELISA, покрытыми FITC-S-белком. Связывание и функцию плеча 2B6 выявляли путем контактирования с растворимым CD32B, а затем с антителом, специфичным к CD32B, и вторичным антителом для детекции, конъюгированным с HRP. (B) 2B6/4420 или 2B6/3G8 связывали с планшетами ELISA, покрытыми HuIgG или FITC-HuIgG (конъюгированный с флуоресцеином). Связывание выявляли контактированием с поликлональной сывороткой, специфичной к 2B6 Fv, а затем со вторичным антителом, конъюгированным с HRP.
ФИГ. 22 (ПАНЕЛИ A И B) АКТИВАЦИЯ ОЧИЩЕННЫХ B-КЛЕТОК С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ ПРОТИВ CD79B ЧЕЛОВЕКА.
Очищенные B-клетки активировали с использованием возрастающих концентраций антител против CD79b человека, конъюгированных с FITC, CB3.1-FITC (A) или CB3.2-FITC (B) и 50 мкг/мл F(ab')2-фрагмента Fc-специфичного IgG GAM (ось x). B-клетки активировали в присутствии PBS (белые столбцы) или 5 мкг/мл либо αFITCαCD32BDART (черные столбцы), либо αCD16αCD32BDART (серые столбцы). Реакции проводили в трех экземплярах и вычисляли стандартные отклонения.
ФИГ. 23 (ПАНЕЛИ A И B) АКТИВАЦИЯ ОЧИЩЕННЫХ B-КЛЕТОК
Очищенные B-клетки от второго здорового донора активировали, как описано на ФИГ. 22, панель B. Индекс пролиферации измеряли в клетках, активированных в присутствии антитела против CD79b, конъюгированного с FITC, CB3.2-FITC (A), и сравнивали с индексом пролиферации клеток, активированных в присутствии немеченого антитела CB3.2 (B).
ФИГ. 24 (Верхняя и нижняя панели) ИСТОЩЕНИЕ B-КЛЕТОК МЫШИ IN VIVO У ТРАНСГЕННЫХ МЫШЕЙ С ИСПОЛЬЗОВАНИЕМ MGD261
Мышам mCD32-/- hCD16A+ C57B1/6, mCD32-/- hCD32B+ C57B1/6 и mCD32-/- hCD16A+ hCD32B+ C57B1/6 из колонии для выведения MacroGenics инъецировали внутривенно на 0, 3, 7, 10, 14 и 17 сутки MGD261 (10, 3, 1 или 0,3 мг/кг) или постороннее антитело (hE16 10 мг/кг). На сутки -19 (до отбора крови), 4, 11, 18, 25 и 32 проводили взятие крови для анализа FACS. Здоровье и активность животных регистрировали три раза в неделю. Верхняя панель: h2B6-3G8 и WNV mAb; нижняя панель: мыши h2B6-3G8-hCD16A или -hCD32B и мыши WNV mAb-hCD16A или - hCD32B.
ФИГ. 25 ИСТОЩЕНИЕ B-КЛЕТОК МЫШИ IN VIVO У ТРАНСГЕННЫХ МЫШЕЙ HCD16A/B ПРИ ИСПОЛЬЗОВАНИИ 2.4G2-3G8 DB
Мышам mCD16-/-, mCD16-/- hCD16A+ C57B1/6, mCD16-/- hCD16B+ и mCD16-/- hCD16A+ hCD16B+ из колонии для выведения MacroGenics инъецировали внутрибрюшинно на 0, 2, 4, 7, 9, 11, 14, 16 и 18 сутки 2.4G2-3G8 DB (75 мкг/мышь) или PBS. На -10 (до отбора крови), 4, 11 и 18 проводили взятие крови для анализа FACS. Здоровье и активность животных регистрировали три раза в неделю.
ФИГ. 26 ДЕМОНСТРАЦИЯ ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТИ MGD261 С ИСПОЛЬЗОВАНИЕМ ВНУТРИВЕННОЙ (IV) МОДЕЛИ С ОПУХОЛЕВОЙ КЛЕТОЧНОЙ ЛИНИЕЙ ЧЕЛОВЕКА RAJI.
Мышам в возрасте двенадцати-двадцати недель mCD16-/-, hCD16A+, RAGl-/- C57B1/6 из колонии для выведения MacroGenics инъецировали внутривенно на 0 сутки 5×106 клеток Raji. На 6, 9, 13, 16, 20, 23, 27 и 30 мышам также внутрибрюшинно (IP) вводили 250, 25 или 2,5 мкг MGD261 или PBS (отрицательный контроль). Затем мышей наблюдали каждые сутки и массу тела регистрировали два раза в неделю. Мышей, у которых развивался паралич задних конечностей, умерщвляли.
ФИГ. 27 ЭКСПРЕССИЯ DART У НЕ ЯВЛЯЮЩЕГОСЯ МЛЕКОПИТАЮЩИМ ХОЗЯИНА
Клетки BL21DE3 (Novagen) трансформировали плазмидой pET25b(+) T7-lac+ 3G8/3G8, и устойчивую к amp колонию использовали для посева в бульонную культуру. Когда культура достигала 0,5 OD600 единиц, добавляли 0,5 мМ IPTG для индукции экспрессии. Культуру выращивали при 30°С в течение 2 часов и бесклеточную среду собирали.
ФИГ. 28 ELISA ДЛЯ DART
ELISA связывания h3G8-h3G8 DART проводили с использованием 96-луночных планшетов Maxisorp. После реакции планшет промывали три раза PBS-T и проявляли 80 мкл/лунка субстрата TMB. После инкубации в течение 5 минут реакцию останавливали 40 мкл/лунка 1% H2SO4. Проводили считывание при OD450 нм с использованием устройства для считывания 96-луночных планшетов и программного обеспечения SOFTmax. Данные считывания наносили на график с использованием программного обеспечения GraphPadPrism 3.03.
ФИГ. 29 ИНДУЦИРУЕМАЯ DART ГИБЕЛЬ B-КЛЕТОК ЧЕЛОВЕКА
PBMC человека инкубировали в течение ночи с указанными молекулами. Апоптоз анализировали с помощью анализа FACS в качестве процента популяции B-клеток PI+аннексин-V+ (клетки CD20+) от всей популяции вне области FSC/SSC.
ФИГ. 30 КОНСТРУКЦИИ 8B5-CB3.1 DART
Для иллюстрации настоящего изобретения получали множество конструкций 8B5-CB3.1 DART. Экспрессирующие плазмиды, кодирующие конструкции 5 и 6, или 6 и 7, или 8 и 9, или 9 и 10, котрансфицировали в клетки HEK-293 для экспрессии 8B5-CB3.1 DART с антителом против flag-метки или без него с использованием Lipofectamine 2000 (Invitrogen). Кондиционированную среду собирали каждые трое суток три раза. Затем кондиционированную среду очищали с использованием аффинной колонки для CD32B.
ФИГ. 31 ELISA для 8B5-CB3.1 DART
Конкурентный ELISA для 8B5-CB3.1 DART/ch8B5 проводили с использованием 96-луночных планшетов Maxisorp. После реакции планшет три раза промывали PBS-T и проявляли 80 мкл/лунка субстрата TMB. После инкубации в течение 5 минут реакцию останавливали 40 мкл/лунка 1% H2SO4. Считывание при OD450 нм проводили с использованием устройства для считывания 96-луночных планшетов и программного обеспечения SOFTmax. Данные считывания наносили на график с использованием программного обеспечения GraphPadPrism 3.03.
ФИГ. 32 СХЕМАТИЧНАЯ ИЛЛЮСТРАЦИЯ ЧЕТЫРЕХВАЛЕНТНОЙ СТРУКТУРЫ DART
Проиллюстрирована общая структура типов DART, продуцируемых путем сборки четырех полипептидных цепей. Четыре антигенсвязывающих домена Ig-подобного DART представлены в качестве полосатых и темно-серых эллипсов.
ФИГ. 33 Ig-ПОДОБНЫЙ ЧЕТЫРЕХВАЛЕНТНЫЙ DART
Представлена схема участков связывания эпитопа Ig-подобного четырехвалентного DART.
ФИГ. 34 ELISA СВЯЗЫВАНИЯ mCD32-hCD16A
Представлены результаты ELISA, которые демонстрируют, что Ig-подобные четырехвалентные типы DART примера 6.10 связывают антиген с более высокой аффинностью, чем контрольное антитело (ch-mCD32 mAb) или другие типы DART.
ФИГ. 35 СХЕМАТИЧНАЯ ИЛЛЮСТРАЦИЯ МОЛЕКУЛ Ig DART
Представлена схема молекул Ig DART. Специфичность указана затемненными, мозаичными или белыми областями, константные области указаны черным цветом, и дисульфидные связи указаны точечными черными линиями. N-концы всех белковых цепей ориентированы в направлении верхней части фигуры, а C-концы всех белковых цепей ориентированы в направлении нижней части фигуры. Иллюстрации A-E являются биспецифическими, и иллюстрации F-J являются триспецифическими. Иллюстрации A и E являются четырехвалентными. Иллюстрации B, C, F, I и J являются шестивалентными. Иллюстрации D, G и H являются восьмивалентными. См. фигуры 1, 2, 9, 14 и 17 и раздел 3.1 для подробных описаний отдельных доменов.
ФИГ. 36 СВЯЗЫВАЮЩАЯ СПОСОБНОСТЬ БИСПЕЦИФИЧЕСКОГО ДИАНТИТЕЛА HU2B6 4.5-HU3G8 5.1
На фигуре 36 представлена способность биспецифического диантитела Hu2B6 4.5-Hu3G8 5.1 (прямоугольники) связывать CD32b и CD16a относительно диантител Hu2B6 4.5 или Hu3G8 5.1 (треугольники).
ФИГ. 37 СХЕМА ПРОИЗВОДНЫХ DART С E-СПИРАЛЬЮ И K-СПИРАЛЬЮ
На фигуре 37 проиллюстрирована общая конформация производных DART с E-спиралью и K-спиралью.
ФИГ. 38 ОРГАНИЗАЦИЯ СПИРАЛИ ПРЕДПОЧТИТЕЛЬНЫХ РАЗДЕЛИТЕЛЕЙ В ВИДЕ E-СПИРАЛИ И K-СПИРАЛИ
На фигуре 38 представлена организация спирали предпочтительной последовательности "E-спирали" (EVAALEK)4 (SEQ ID NO: 299) и предпочтительной последовательности "K-спирали" (KVAALKE)4 (SEQ ID NO: 300).
ФИГ. 39 Fc-СОДЕРЖАЩИЕ ПРОИЗВОДНЫЕ DART C E-СПИРАЛЬЮ И K-СПИРАЛЬЮ
На фигуре 39 проиллюстрированы различные типы Fc-содержащих производных DART с E-спиралью и K-спиралью, которые могут быть получены путем обмена цепями.
ФИГ. 40 ЭКСКЛЮЗИОННАЯ ХРОМАТОГРАФИЯ ПРОИЗВОДНЫХ С E-СПИРАЛЬЮ И/ИЛИ K-СПИРАЛЬЮ И FC-СОДЕРЖАЩИХ ПРОИЗВОДНЫХ С E-СПИРАЛЬЮ И/ИЛИ K-СПИРАЛЬЮ ДЛЯ DART h2B6YAhCB3.
На фигуре 40 представлены результаты эксклюзионной хроматографии производных с E-спиралью и/или K-спиралью и Fc-содержащих производных с E-спиралью и/или K-спиралью для DART h2B6YAhCB3. Анализировали четыре типа таких молекул; все имели E-спираль и K-спираль: EK (без Fc-области), 2,1 мг; EFc/K (Fc, связанный с E-спиралью), 2,7 мг; E/KFc (Fc, связанный с K-спиралью), 1,8 мг; EFc/KFc (Fc, связанный с K-спиралью и E-спиралью того же DART), 2,0 мг.
ФИГ. 41. СТРУКТУРА ПРОДУЦИРОВАННЫХ ДИМЕРНЫХ МОЛЕКУЛ
На фигуре 41 представлена возможная структура продуцированной димерной молекулы, идентифицированной эксклюзионной хроматографией, представленной на фигуре 40.
ФИГ. 42 АНАЛИЗ СПОСОБОМ SDS-ПОЛИАКРИЛАМИДНОГО ГЕЛЬ-ЭЛЕКТРОФОРЕЗА ПРОИЗВОДНЫХ С E-СПИРАЛЬЮ И/ИЛИ K-СПИРАЛЬЮ ИЛИ FC-СОДЕРЖАЩИХ ПРОИЗВОДНЫХ С E-СПИРАЛЬЮ И/ИЛИ K-СПИРАЛЬЮ ДЛЯ DART h2B6YAhCB3
На фигуре 42 представлены результаты анализы путем SDS-полиакриламидного гель-электрофореза фракций, полученных эксклюзионной хроматографией (фигура 40), производных с E-спиралью и/или K-спиралью и Fc-содержащих производных с E-спиралью и/или K-спиралью для DART h2B6YAhCB3. Крайние дорожки: контроли в виде молекулярного маркера; дорожка 1: EK (без Fc-области); дорожка 2: EFc/K, фракция агрегата; дорожка 3: EFc/K, фракция мономера; дорожка 4: E/KFc, фракция агрегата; дорожка 5: E/KFc, фракция мономера; дорожка 6: EFc/KFc, фракция агрегата; дорожка 7: EFc/KFc, фракция димера; дорожка 8: EFc/KFc, фракция мономера.
ФИГ. 43 ELISA-АНАЛИЗ БИСПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ
На фигуре 43 представлен ELISA-анализ биспецифического связывания, сравнивающего Fc-содержащие производные с E-спиралью/K-спиралью DART h2B6YAhCB3 (EFc/K или E/KFc), DART h2B6YAhCB3, контроль и производное DART EFc/KFc h2B6YAhCB3.
ФИГ. 44 СПОСОБНОСТЬ ПРОИЗВОДНЫХ С E-СПИРАЛЬЮ И/ИЛИ K-СПИРАЛЬЮ И FC-СОДЕРЖАЩИХ ПРОИЗВОДНЫХ С E-СПИРАЛЬЮ И/ИЛИ K-СПИРАЛЬЮ ДЛЯ DART h2B6YAhCB3 ИНГИБИРОВАТЬ ПРОЛИФЕРАЦИЮ T-КЛЕТОК
На фигуре 44 представлена способность производных с E-спиралью и/или K-спиралью и Fc-содержащих производных с E-спиралью и/или K-спиралью для DART h2B6YAhCB3 ингибировать пролиферацию T-клеток.
ФИГ. 45 ABD-DART hCD16-hCD32B
На фигуре 45 представлена схема молекулы рекомбинантного антитела ABD-DART hCD16-hCD32B, состоящего из ABD3-домена белка стрептококков G, слитого с рекомбинантным биспецифическим DART, который является иммунореактивным в отношении антигенов hCD16 и hCD32B.
ФИГ. 46A/46B АФФИННОСТЬ СВЯЗЫВАНИЯ ABD-DART hCD16-hCD32B С ИСПОЛЬЗОВАНИЕМ ELISA С ДВОЙНОЙ СПЕЦИФИЧНОСТЬЮ
Планшеты для ELISA покрывали либо антигеном CD16 (фигура 46A), либо сывороточным альбумином человека (фигура 46B) в концентрации 2 мкг/мл. Связывали различные концентрации ABD-DART hCD16-hCD32B (■) и контрольного DART hCD16-hCD32B (○), начиная с 2 мкг/мл. В планшеты добавляли биотинилированный антиген sCD32B, а затем конъюгированный с HRP стрептавидин для детекции.
ФИГ. 47 ОПОСРЕДУЕМАЯ PBMC ЦИТОТОКСИЧНОСТЬ БЕЛКОВ DART
Опосредуемая PBMC цитотоксичность белков DART. Анализы ADCC проводили с использованием B-клеточных линий человека Daudi в качестве клеток-мишеней, инкубированных с PBMC в качестве эффекторных клеток. Индивидуальные анализы проводили в трех экземплярах в соотношении эффектора к мишени 20:1 и при титровании антител: DART hCD16A-hCD32B (●) и ABD-DART hCD16A-hCD32B (■). Клеточноопосредуемую цитотоксичность измеряли с помощью анализа высвобождения LDH. Нижняя кривая на уровне 100 соответствует ABD-DART hCD16A-hCD32B (■).
ФИГ. 48 УСОВЕРШЕНСТВОВАННЫЕ ФАРМАКОКИНЕТИЧЕСКИЕ СВОЙСТВА hCD 16-hCD32B ABD-DART У МЫШЕЙ C57B1/6
Мышам (n=3) проводили однократную внутривенную инъекцию (A) ABD-DART hCD16-hCD32B (●) и (B) DART hCD16-hCD32B (▲) в концентрации 5 мг/кг. Сыворотку мышей собирали в различные моменты времени и количественно определяли концентрации белка в сыворотке с помощью ELISA. Фармакокинетические вычисления проводили с использованием WinNonlin Professional 5.1.
ФИГ. 49A-E АКТИВНОСТЬ DART HER2×TCRb НА ПАНЕЛИ КЛЕТОЧНЫХ ЛИНИЙ С НИЗКОЙ ЭКСПРЕССИЕЙ HER2
Молекулы DART, имеющие домены, связывающие Her2 и T-клеточный рецептор (TCR), тестировали в отношении их способности опосредовать цитотоксичность во множестве клеточных линий рака молочной железы, рака толстого кишечника и рака мочевого пузыря, которые были ранее охарактеризованы, как проявляющие низкие уровни экспрессии HER2 (и, таким образом, являющиеся устойчивыми к лечению антителом против Her2/neu, Herceptin®). Протестированные клеточные линии рака молочной железы представляют собой ZR75-1 (HER2 2+) (ФИГ. 49A), MCF-7 (HER2 1+) (ФИГ. 49B) и MDA-MB468 (HER2-ve) (ФИГ. 49C). Протестированные клеточные линии рака не молочной железы представляют собой HT-29 (клеточная линия рака толстого кишечника) (ФИГ. 49D) и SW780 (клеточная линия рака мочевого пузыря) (ФИГ. 49E).
5. ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Каждая полипептидная цепь молекулы диантитела содержит VL-домен и VH-домен, которые ковалентно связаны, так чтобы эти домены удерживались от самосборки. Взаимодействие двух полипептидных цепей приводит к образованию пар VL-VH, образующих участки связывания эпитопов, т.е. бивалентную молекулу. Ни VH-, ни VL-домен не ограничены каким-либо положением в полипептидной цепи, т.е. не ограничены ни амино (N)-, ни (C)-концом, а также эти домены не ограничены по их относительным положениям друг относительно друга, т.е. VL-домен может быть N-концевым для VH-домена и наоборот. Единственное ограничение состоит в том, чтобы была доступна комплементарная полипептидная цепь для образования функционального диантитела. Когда VL- и VH-домены происходят из одного и того же антитела, две комплементарные полипептидные цепи могут быть идентичными. Например, когда связывающие домены происходят из антитела, специфичного к эпитопу A (т.е. связывающий домен образуется при взаимодействии VLA-VHA), каждый полипептид содержит VHA и VLA. Гомодимеризация двух полипептидных цепей антитела приводит к образованию двух участков связывания VLA-VHA, что приводит к двухвалентному моноспецифическому антителу. Когда VL- и VH-домены происходят из антител, специфичных к различным антигенам, образование функционального биспецифического диантитела требует взаимодействия двух различных полипептидных цепей, т.е. образования гетеродимера. Например, для биспецифического диантитела одна полипептидная цепь содержит VLA и VLB; гомодимеризация указанной цепи приводит к образованию двух участков связывания VLA-VHB, либо без связывания, либо с непрогнозируемым связыванием. Напротив, когда две различных полипептидных цепи свободны для взаимодействия, например, в рекомбинантной экспрессирующей системе, причем одна из них содержит VLA и VHB, а другая содержит VLB и VHA, два различных участка связывания будут образовывать: VLA-VHA и VLB-VHB. Для всех пар полипептидных цепей диантитела возможно ошибочное расположение или ошибочное связывание двух цепей, т.е. взаимодействие доменов VL-VL или VH-VH; однако очистку функциональных диантител легко проводить на основе иммуноспецифичности правильно димеризованного участка связывания с использованием любого способа на основе аффинности, известного в данной области или проиллюстрированного в настоящем документе, например, аффинной хроматографии.
В других вариантах осуществления одна или несколько полипептидных цепей диантитела содержит Fc-домен. Fc-домены в полипептидных цепях молекул диантител предпочтительно димеризуются, что приводит к образованию молекулы диантитела, которая проявляет иммуноглобулин-подобные свойства, например, взаимодействия Fc-FcγR. Fc-содержащие диантитела могут представлять собой димеры, например, состоящие из двух полипептидных цепей, каждая из которых содержит VH-домен, VL-домен и Fc-домен. Димеризация указанных полипептидных цепей приводит к двухвалентному диантителу, содержащему Fc-домен, хотя и со структурой, отличающейся от структуры немодифицированного двухвалентного антитела (ФИГ. 11). Такие молекулы диантител обладают измененными фенотипами относительно иммуноглобулина дикого типа, например, измененным временем полужизни в сыворотке, свойствами связывания и т.д. В других вариантах осуществления молекулы диантител, содержащие Fc-домены, могут быть тетрамерными. Такие тетрамеры содержат две "более тяжелые" полипептидные цепи, т.е. полипептидные цепи, содержащие VL, VH и Fc-домен, и две "более легкие" полипептидные цепи, т.е. полипептидные цепи, содержащие VL и VH. Указанные более легкие и более тяжелые цепи взаимодействуют с образованием мономера, и указанные мономеры взаимодействуют через их неспаренные Fc-домены с образованием Ig-подобной молекулы. Такое Ig-подобное диантитело является четырехвалентным и может быть моноспецифическим, биспецифическим или тетраспецифическим.
По меньшей мере два участка связывания молекулы диантитела могут распознавать одинаковые или различные эпитопы. Различные эпитопы могут быть из одного антигена или из разных антигенов. В одном из вариантов осуществления эпитопы происходят из различных клеток. В другом варианте осуществления эпитопы представляют собой антигены клеточной поверхности на одной клетке или вирусе. Участки связывания эпитопов могут распознавать любой антиген, к которому может быть получено антитело. Например, белки, нуклеиновые кислоты, бактериальные токсины, маркеры клеточной поверхности, аутоиммунные маркеры, вирусные белки, лекарственные средства и т.д. В конкретных аспектах по меньшей мере один участок связывания эпитопа в диантителе является специфичным к антигену на конкретной клетке, такой как B-клетка или T-клетка, фагоцитарная клетка, естественная киллерная (NK) клетка или дендритная клетка.
Любые домены полипептидной цепи диантитела, т.е. VL-, VH- и FC-домен, могут быть разделены пептидным линкером. Пептидный линкер может иметь 0, 1, 2, 3, 4, 5, 6, 7, 8 или 9 аминокислот. В определенных вариантах осуществления последовательность аминокислотного линкера представляет собой GGGSGGGG (SEQ ID NO: 10) и кодируется последовательностью нуклеиновой кислоты (SEQ ID NO: 74).
В определенных вариантах осуществления каждую полипептидную цепь молекулы диантитела конструируют так, чтобы она содержала по меньшей мере один остаток цистеина, который будет взаимодействовать с аналогичным по меньшей мере одним остатком цистеина на второй полипептидной цепи по изобретению с образованием межцепочечной дисульфидной связи. Указанные межцепочечные дисульфидные связи служат для стабилизации молекулы диантитела, улучшая экспрессию и выход в рекомбинантных системах, что приводит к стабильному и единообразному составу, а также к повышению стабильности выделенного и/или очищенного продукта in vivo. Указанный по меньшей мере один остаток цистеина можно вносить в качестве одной аминокислоты или в качестве более крупной аминокислотной последовательности, например шарнирного домена, в любой части полипептидной цепи. В конкретном варианте осуществления указанный по меньшей мере один остаток цистеина встроен так, чтобы он находился на C-конце полипептидной цепи. В некоторых вариантах осуществления указанный по меньшей мере один остаток цистеина встраивают в аминокислотную последовательность LGGC полипептидной цепи. В конкретном варианте осуществления C-конец полипептидной цепи, содержащий молекулу диантитела по изобретению, содержит аминокислотную последовательность LGGC. В другом варианте осуществления указанный по меньшей мере один остаток цистеина встраивают в аминокислотную последовательность полипептида, содержащую шарнирный домен, например SEQ ID NO: 1 или SEQ ID NO: 4. В конкретном варианте осуществления C-конец полипептидной цепи молекулы диантитела по изобретению содержит аминокислотную последовательность шарнирного домена IgG, например SEQ ID NO: 1. В другом варианте осуществления C-конец полипептидной цепи молекулы диантитела по изобретению содержит аминокислотную последовательность VEPKSC (SEQ ID NO: 77), которая может кодироваться нуклеотидной последовательностью (SEQ ID NO: 78). В других вариантах осуществления указанный по меньшей мере один остаток цистеина вносят в аминокислотную последовательность LGGCFNRGEC (SEQ ID NO: 17) полипептидной цепи, которая может кодироваться нуклеотидной последовательностью (SEQ ID NO: 76). В конкретном варианте осуществления C-конец полипептидной цепи, содержащейся в диантителе по изобретению, содержит аминокислотную последовательность LGGCFNRGEC (SEQ ID NO: 17), которая может кодироваться нуклеотидной последовательностью (SEQ ID NO: 76). В других вариантах осуществления указанный по меньшей мере один остаток цистеина вносят в полипептидную цепь в аминокислотной последовательности FNRGEC (SEQ ID NO: 23), которая может кодироваться нуклеотидной последовательностью (SEQ ID NO: 75). В конкретном варианте осуществления C-конец полипептидной цепи, содержащейся в диантителе по изобретению, содержит аминокислотную последовательность FNRGEC (SEQ ID NO: 23), которая может кодироваться нуклеотидной последовательностью (SEQ ID NO: 75).
В определенных вариантах осуществления молекула диантитела содержит по меньшей мере две полипептидных цепи, каждая из которых содержит аминокислотную последовательность LGGC и ковалентно связана дисульфидной связью между остатками цистеина в указанных последовательностях LGGC. В другом конкретном варианте осуществления молекула диантитела содержит по меньшей мере две полипептидных цепи, одна из которых содержит последовательность FNRGEC (SEQ ID NO: 23), а другая содержит шарнирный домен (содержащий по меньшей мере один остаток цистеина), где указанные по меньшей мере две полипептидных цепи ковалентно связаны дисульфидной связью между остатком цистеина в FNRGEC (SEQ ID NO: 23) и остатком цистеина в шарнирном домене. В конкретных аспектах остаток цистеина, ответственный за дисульфидную связь, расположенную в шарнирном домене, представляет собой Cys-128 (при нумерации согласно EU по Kabat; расположен в шарнирном домене немодифицированной целой тяжелой цепи IgG), и аналогичный остаток цистеина в SEQ ID NO: 23 представляет собой Cys-214 (при нумерации согласно EU по Kabat; расположен на C-конце немодифицированной целой легкой цепи IgG) (Elkabetz et al. (2005) "Cysteins In CH1 Underlie Retention Of Unassembled Ig Heavy Chains", J. Biol. Chem. 280:14402-14412; включенная в настоящий документ в качестве ссылки в полном объеме). В других вариантах осуществления по меньшей мере один остаток цистеина встроен так, чтобы он располагался на N-конце аминокислотной цепи. В других вариантах осуществления по меньшей мере один остаток цистеина встроен так, чтобы он располагался в линкерной части полипептидной цепи молекулы диантитела. В следующих вариантах осуществления VH- или VL-домен встроены так, чтобы они содержали по меньшей мере одну аминокислотную модификацию относительно исходных VH- или VL-домена, так чтобы указанная аминокислотная модификация содержала замену исходной аминокислоты на цистеин.
Изобретение охватывает молекулы диантител, содержащие Fc-домен или его часть (например, CH2-домен или CH3-домен). Fc-домен или его часть могут происходить из любого изотипа или аллотипа иммуноглобулина, включая, но не ограничиваясь ими, IgA, IgD, IgG, IgE и IgM. В предпочтительных вариантах осуществления Fc-домен (или его часть) происходит из IgG. В конкретных вариантах осуществления изотип IgG представляет собой IgG1, IgG2, IgG3 или IgG4 или их аллотипы. В одном из вариантов осуществления молекула диантитела содержит Fc-домен, где Fc-домен содержит CH2-домен и CH3-домен, независимо выбранный из любого изотипа иммуноглобулина (т.е. Fc-домен, содержащий CH2-домен, происходящий из IgG, и CH3-домен, происходящий из IgE, или CH2-домен, происходящий из IgG1, и CH3-домен, происходящий из IgG2, и т.д.). Указанный Fc-домен может быть встроен в полипептидную цепь, содержащую молекулу диантитела по изобретению в любом положении относительно других доменов или частей указанной полипептидной цепи (например, Fc-домен или его часть может быть c-концевым как для VL-домена, так и для VH-домена полипептидной цепи; он может быть n-концевым как для VL-домена, так и для VH-домена; или он может быть N-концевым для одного домена и c-концевым для другого домена (т.е. между доменами полипептидной цепи)).
Также настоящее изобретение охватывает молекулы, содержащие шарнирный домен. Шарнирный домен может происходить из любого изотипа или аллотипа иммуноглобулина, включая IgA, IgD, IgG, IgE и IgM. В предпочтительных вариантах осуществления шарнирный домен происходит из IgG, где изотип IgG представляет собой IgG1, IgG2, IgG3 или IgG4 или его аллотип. Указанный шарнирный домен может быть встроен в полипептидную цепь, содержащуюся в молекуле диантитела, вместе с Fc-доменом, так чтобы молекула диантитела содержала шарнирный-Fc-домен. В определенных вариантах осуществления шарнирная область и Fc-домен независимо выбраны из любого изотипа иммуноглобулина, известного в данной области или проиллюстрированного в настоящем документе. В других вариантах осуществления шарнирная область и Fc-домен разделены по меньшей мере одним другим доменом полипептидной цепи, например, VL-доменом. Шарнирный домен, или необязательно шарнирный-Fc-домен, можно встраивать в полипептид по изобретению в любом положении относительно других доменов или частей указанной полипептидной цепи. В определенных вариантах осуществления полипептидная цепь по изобретению содержит шарнирный домен, который находится на C-конце полипептидной цепи, где указанная полипептидная цепь не содержит Fc-домен. В других вариантах осуществления полипептидная цепь по изобретению содержит шарнирный-Fc-домен, который находится на C-конце полипептидной цепи. В следующих вариантах осуществления полипептидная цепь по изобретению содержит шарнирный-Fc-домен, который находится на N-конце полипептидной цепи.
Как рассмотрено выше, изобретение охватывает мультимеры полипептидных цепей, каждая из которых содержит VH- и VL-домен. В некоторых аспектах полипептидные цепи в указанных мультимерах дополнительно содержат Fc-домен. Димеризация Fc-доменов приводит к образованию молекулы диантитела, которая обладает иммуноглобулин-подобной функциональностью, т.е. Fc-опосредуемой функцией (например, взаимодействие Fc-FcγR, связывание комплемента и т.д.). В определенных вариантах осуществления VL- и VH-домены, составляющие каждую полипептидную цепь, обладают одной специфичностью, и указанная молекула диантитела является двухвалентной и моноспецифической. В других вариантах осуществления VL- и VH-домены, составляющие каждую полипептидную цепь, обладают отличающейся специфичностью, и диантитело является двухвалентным и биспецифическим.
В других вариантах осуществления молекулы диантител по изобретению охватывают тетрамеры полипептидных цепей, каждая из которых содержит VH- и VL-домен. В определенных вариантах осуществления две полипептидных цепи тетрамера, кроме того, содержат Fc-домен. Тетрамер, таким образом, содержит две "более тяжелых" полипептидных цепи, каждая из которых содержит VL-, VH- и Fc-домен, и две "более легких" полипептидных цепи, содержащих VL- и VH-домен. Взаимодействие более тяжелой и более легкой цепи в двухвалентном мономере, связанном путем димеризации указанных мономеров через Fc-домены более тяжелых цепей, приводит к образованию четырехвалентной иммуноглобулин-подобной молекулы (проиллюстрированной в примере 6.2 и примере 6.3). В некоторых аспектах мономеры являются одинаковыми, и четырехвалентная молекула диантитела является моноспецифической или биспецифической. В других аспектах мономеры отличаются, и четырехвалентная молекула является биспецифической или тетраспецифической.
Образование молекулы тетраспецифического диантитела, как описано выше, требует взаимодействия четырех различных полипептидных цепей. Таких взаимодействий трудно эффективно достигнуть в пределах одной клеточной рекомбинантной системы продукции, вследствие многих вариантов потенциальных ошибочных спариваний цепей. Одним из решений для повышения вероятности ошибочных спариваний является встраивание мутаций типа "выступы-в-полостях" в желаемые пары полипептидных цепей. Такие мутации способствуют гетеродимеризации, а не гомодимеризации. Например, что касается взаимодействий Fc-Fc, аминокислотную замену (предпочтительно замена аминокислотой, содержащей объемную боковую группу, образующую "выступ", например, триптофан) можно встраивать в CH2- или CH3-домен, так чтобы пространственное препятствие предотвращало взаимодействие с доменом с аналогичной мутацией и вынуждало мутантный домен образовывать пару с доменом, в который внесена комплементарная или обеспечивающая соответствие мутация, т.е. "полость" (например, замена глицином). Такие наборы мутаций можно вносить в любую пару полипептидов, составляющих молекулу диантитела, и, кроме того, их можно вносить в любую часть полипептидных цепей указанной пары. Способы инженерии белков для способствования гетеродимеризации, а не гомодимеризации, хорошо известны в данной области, в частности, в отношении инженерии иммуноглобулин-подобных молекул, и они охватываются настоящим изобретением (см. например, Ridgway et al. (1996) "'Knobs-Into-Holes' Engineering Of Antibody CH3 Domains For Heavy Chain Heterodimerization", Protein Engr. 9:617-621, Atwell et al. (1997) "Stable Heterodimers From Remodeling The Domain Interface Of A Homodimer Using A Phage Display Library", J. Mol. Biol. 270: 26-35, и Xie et al. (2005) "A New Format Of Bispecific Antibody: Highly Efficient Heterodimerization, Expression And Tumor Cell Lysis", J. Immunol. Methods 296:95-101; каждая из которых включена в настоящий документ в качестве ссылок в полном объеме).
Также изобретение относится к молекулам диантител, содержащим вариант Fc-домен или вариант домена шарнирная область-Fc (или их часть), причем вариант Fc-домена содержит по меньшей мере одну аминокислотную модификацию (например, замену, инсерцию, делецию) относительно сравнимого Fc-домена или домена шарнирная область-Fc (или их части) дикого типа. Молекулы, содержащие варианты Fc-доменов или доменов шарнирная область-Fc (или из частей) (например, антитела), обычно обладают измененными фенотипами относительно молекул, содержащих Fc-домены или домены шарнирная область-Fc дикого типа или их части. Вариант фенотипа может проявляться в качестве измененного времени полужизни, измененной стабильности, измененной чувствительности к клеточным ферментам или измененной эффекторной функции при анализе в анализе на основе NK или на основе макрофагов. Варианты Fc-доменов, идентифицированные как изменяющие эффекторную функцию, описаны в международной заявке WO04/063351, публикациях патентных заявок США 2005/0037000 и 2005/0064514, предварительных заявках США 60/626510, поданной 10 ноября 2004 года, 60/636663, поданной 15 декабря 2004 года, и 60/781564, поданной 10 марта 2006 года, и в патентных заявках США 11/271140, поданной 10 ноября 2005 года, и 11/305787, поданной 15 декабря 2005, совместных заявках авторов настоящего изобретении, которые все включены в настоящий документ в качестве ссылок в полном объеме.
Биспецифические диантитела по изобретению могут одновременно связывать два различных и обособленных эпитопа. В определенных вариантах осуществления эпитопы находятся в одном антигене. В других вариантах осуществления эпитопы находятся на различных антигенах. В предпочтительных вариантах осуществления по меньшей мере один участок связывания эпитопа является специфичным к детерминанте, экспрессированной на иммунной эффекторной клетке (например CD3, CD16, CD32, CD64 и т.д.), которая экспрессируется на T-лимфоцитах, естественных киллерных (NK) клетках или других мононуклеарных клетках. В одном из вариантов осуществления молекула диантитела связывается с детерминантой эффекторной клетки и также активирует указанную эффекторную клетку. В этом отношении молекулы диантител по изобретению могут проявлять Ig-подобную функциональность, независимо от того, содержат ли они, кроме того, Fc-домен (например, в каком-либо анализе эффекторных функций, известном в данной области или проиллюстрированном в настоящем документе (например, в анализе ADCC). В определенных вариантах осуществления биспецифическое диантитело по изобретению связывает как антиген злокачественной опухоли на опухолевой клетке, так и детерминанту эффекторной клетки, одновременно активируя указанную клетку. В альтернативных вариантах осуществления биспецифическое диантитело или молекула диантитела по изобретению может ингибировать активацию мишени, например, эффекторной клетки, путем одновременного связывания, и, таким образом, сшивания, активирующего и ингибиторного рецептора на той же клетке (например, связывает CD32A и CD32B, BCR и CD32B или IgERI и CD32B), как описано выше (см., раздел "Уровень техники"). В следующем аспекте этого варианта осуществления биспецифическое диантитело может проявлять противовирусные свойства путем одновременного связывания двух нейтрализующих эпитопов на вирусе (например, эпитопов RSV; эпитопов WNV, таких как E16 и E53).
В определенных вариантах осуществления биспецифические молекулы диантител по изобретению обеспечивают уникальную возможность нацеливания на конкретные типы клеток. Например, биспецифическое диантитело или молекулу диантитела можно конструировать так, чтобы она содержала комбинацию участков связывания эпитопов, которые распознают набор антигенов, уникальный для клетки-мишени или типа ткани-мишени. Кроме того, когда какой-либо или оба из отдельных антигенов достаточно часто встречаются по отдельности в другой ткани и/или типах клеток, для конструирования диантитела или молекулы диантитела можно использовать низкоаффинные связывающие домены. Такие низкоаффинные связывающие домены будут неспособны связываться с отдельным эпитопом или антигеном с достаточной авидностью для терапевтических целей. Однако когда оба эпитопа или антигена присутствуют на одной клетке-мишени или ткани-мишени, авидность диантитела или молекулы диантитела к клетке или ткани, относительно клетки или ткани, экспрессирующих только один из антигенов, будет возрастать, так что изобретение может эффективно нацеливаться на указанную клетку-мишень или ткань-мишень. Такая биспецифическая молекула может проявлять усиленное связывание с одним или с обоими из ее антигенов-мишеней на клетках, экспрессирующих оба указанных антигена, относительно моноспецифического диантитела или антитела со специфичностью только к одному из антигенов.
Предпочтительно связывающие свойства диантител по изобретению охарактеризовывают в функциональных анализах in vitro для определения активности связывания и/или одной или нескольких опосредуемых FcγR функций эффекторных клеток (опосредуемых взаимодействиями Fc-FcγR или иммуноспецифическим связыванием молекулы диантитела с FcγR) (cм. раздел 5.4.2 и 5.4.3). Аффинность и связывающие свойства молекул, например диантител, по изобретению к FcγR можно определять с использованием анализов in vitro (биохимических или иммунологических анализов), известных в данной области, для определения связывания домен-антиген или взаимодействий Fc-FcγR, т.е. специфического связывания антигена со связывающим доменом или специфического связывания Fc-области с FcγR, соответственно, включая, но не ограничиваясь ими, анализ ELISA, анализ поверхностного плазмонного резонанса, анализы иммунопреципитации (см. раздел 5.4.2). В наиболее предпочтительных вариантах осуществления молекулы по изобретению обладают сходными связывающими свойствами в моделях in vivo (таких как модели, описанные и раскрытые в настоящем документе) со свойствами в анализах in vitro. Однако настоящее изобретение не исключает молекулы по изобретению, которые не проявляют желаемый фенотип в анализах in vitro, но проявляют желаемый фенотип in vivo.
В некоторых вариантах осуществления молекулы по изобретению конструируют так, чтобы они обладали измененным паттерном гликозилирования или содержали измененную гликоформу относительно сравнимой части молекулы-матрицы. Сконструированные гликоформы могут быть пригодны для различных целей, включая, но не ограничиваясь ими, усиление эффекторной функции. Сконструированные гликоформы можно получать способом, известный специалисту в данной области, например, с использованием сконструированных или вариантных экспрессирующих штаммов, путем коэкспрессии одного или нескольких ферментов, например, DI N-ацетилглюкозамилтрансферазы III (GnTIII), путем экспрессии диантитела по изобретению в различных организмах и клеточных линиях из различных организмов, или путем модификации углевода(ов) после экспрессии и очистки диантитела. Способы получения сконструированных гликоформ известны в данной области и включают, но не ограничиваются ими, способы, описанные в Umana et al. (1999) "Engineered Glycoforms Of An Antineuroblastoma IgG1 With Optimized Antibody-Dependent Cellular Cytotoxic Activity", Nat. Biotechnol 17:176-180; Davies et al. (2001) "Expression Of GnTIII In A Recombinant Anti-CD20 CHO Production Cell Line: Expression Of Antibodies With Altered Glycoforms Leads To An Increase In Adcc Through Higher Affinity For Fc Gamma RIII", Biotechnol Bioeng 74:288-294; Shields et al. (2002) "Lack Of Fucose On Human IgGl N-Linked Oligosaccharide Improves Binding To Human Fcgamma RIII And Antibody-Dependent Cellular Toxicity", J. Biol Chem 277:26733-26740; Shinkawa et al. (2003) "The Absence Of Fucose But Not The Presence Of Galactose Or Bisecting N-Acetylglucosamine Of Human IgGl Complex-Type Oligosaccharides Shows The Critical Role Of Enhancing Antibody-Dependant Cellular Cytotoxicity", J. Biol Chem 278:3466-3473) US 6602684; USSN 10/277370; USSN 10/113929; PCT WO 00/61739A1; PCT WO 01/292246A1; PCT WO 02/311140Al; PCT WO 02/30954A1; технологию PotillegentTM (Biowa, Inc. Princeton, NJ); инженерную технологию гликозилирования GlycoMAbTM (GLYCART biotechnology AG, Zurich, Switzerland); каждый из которых включен в настоящий документ в качестве ссылки в полном объеме. См., например, WO 00061739; EA01229125; US 20030115614; Okazaki et al. (2004) "Fucose Depletion From Human IgGl Oligosaccaride Enhances Binding Enthalpy And Association Rate Between IgG1 And FcGammaRIIIA", JMB, 336: 1239-49, каждый из которых включен в настоящий документ в качестве ссылки в полном объеме.
Кроме того, изобретение охватывает включение неприродных аминокислот для получения диантител по изобретению. Такие способы известны специалистам в данной области, как, например, способы с использованием природного биосинтетического аппарата для обеспечения включения неприродных аминокислот в белки, см., например, Wang et al. (2002) "Expanding The Genetic Code", Chem. Comm. 1:1-11; Wang et al. (2001) "Expanding The Genetic Code Of Escherichia coli", Science, 292:498-500; van Hest et al. (2001) "Protein-Based Materials, Toward A New Level Of Structural Control", Chem. Comm. 19:1897-1904, каждый из которых включен в настоящий документ в качестве ссылки в полном объеме. Альтернативные стратегии сосредоточены на ферментах, ответственных за биосинтез аминоацил-тРНК, см., например, Tang et al. (2001) "Biosynthesis Of A Highly Stable Coiled-Coil Protein Containing Hexafluoroleucine In An Engineered Bacterial Host", J. Am. Chem. Soc. 123(44): 11089-11090; Kiick et al. (2001) "Identification Of An Expanded Set Of Translationally Active Methionine Analogues In Escherichia coli", FEBS Lett. 502(1-2):25-30; каждый из которых включен в настоящий документ в качестве ссылки в полном объеме.
В некоторых вариантах осуществления изобретение охватывает способы модификации VL-, VH- или Fc-домена молекулы по изобретению путем добавления или делеции участка гликозилирования. Способы модификации углевода белков хорошо известны в данной области и охватываются изобретением, см., например, патент США № 6218149; EP 0 359 096 B1; публикацию США № US 2002/0028486; WO 03/035835; публикацию США № 2003/0115614; патент США № 6218149; патент США № 6472511; которые все включены в настоящий документ в качестве ссылок в полном объеме.
Молекулы диантител по настоящему изобретению можно конструировать так, чтобы они содержали домен, который представляет собой связывающий лиганд для рецептора естественных киллеров группы 2D (NKG2D). Такие связывающие лиганды, и, в частности, лиганды, которые не экспрессируются на нормальных клетках, включают молекулу гистосовместимости 60 (H60), продукт раннего индуцируемого ретиноевой кислотой гена-1 (RAE-1) и UL16-связывающий подобный белку транскрипт 1 мыши (MULT1) (Raulet D.H. (2003) "Roles Of The NKG2D Immunoreceptor And Its Ligands", Nature Rev. Immunol. 3:781-790; Coudert, J.D. et al. (2005) "Altered NKG2D Function In NK Cells Induced By Chronic Exposure To Altered NKG2D Ligand-Expressing Tumor Cells", Blood 106:1711-1717). Дополнительные лиганды, реактивные к NKG2D человека, включают полиморфные родственные цепи MHC класса I молекулы MICA и MICB (Diefenbach, A. et al. (1999) "Natural Killer Cells: Stress Out, Turn On, Tune In", Curr. Biol. 9(22):R851-R8533; Bauer, S. et al. (1999) "Activation Of NK Cells And T Cells By NKG2D, A Receptor For Stress-Inducible MICA", Science 285(5428):727-729; Stephens, H.A. (2001) "MICA and MICB genes: can the enigma of their polymorphism be resolved?", Trends Immunol. 22:378-385.
Последовательность MICA представляет собой SEQ ID NO: 311:
MGLGPVFLLL AGIFPFAPPG AAAEPHSLRY NLTVLSWDGS VQSGFLTEVH
LDGQPFLRCD RQKCRAKPQG QWAEDVLGNK TWDRETRDLT GNGKDLRMTL
AHIKDQKEGL HSLQEIRVCE IHEDNSTRSS QHFYYDGELF LSQNLETKEW
TMPQSSRAQT LAMNVRNFLK EDAMKTKTHY HAMHADCLQE LRRYLKSGVV
LRRTVPPMVN VTRSEASEGN ITVTCRASGF YPWNITLSWR QDGVSLSHDT
QQWGDVLPDG NGTYQTWVAT RICQGEEQRF TCYMEHSGNH STHPVPSGKV
LVLQSHWQTF HVSAVAAAAI FVIIIFYVRC CKKKTSAAEG PELVSLQVLD
QHPVGTSDHR DATQLGFQPL MSDLGSTGST EGA
Последовательность MICB представляет собой SEQ ID NO: 312:
PHSLRYNLMV LSQDGSVQSG FLAEGHLDGQ PFLRYDRQKR RAKPQGQWAE
DVLGAKTWDT ETEDLTENGQ DLRRTLTHIK DQKGGLHSLQ EIRVCEIHED
SSTRGSRHFY YDGELFLSQN LETQESTVPQ SSRAQTLAMN VTNFWKEDAM
KTKTHYRAMQ ADCLQKLQLP PMVNVICSEV SEGNITVTCR ASSFYPRNIT
LTWRQDGVSL SHNTQQWGDV LPDGNGTYQT WVATRIRQGE EQRFTCYMEH
SGNHGTHPVP SGKALVLQSQ RTDFPYVSAA MPCFVIIIIL CVPCCKKKTS
AAEGP
Антитела, которые специфично связываются с T-клеточным рецептором, включают антитело против TCR BMA 031 (Kurrle, R. et al. (1989) "BMA 031 - A TCR-Specific Monoclonal Antibody For Clinical Application", Transplant Proc. 21 (1 Pt 1):1017-1019; Nashan, B. et al. (1987) "Fine Specificity Of A Panel Of Antibodies Against The TCR/CD3 Complex", Transplant Proc. 19(5):4270-4272; Shearman, C.W. et al. (1991) "Construction, Expression, And Biologic Activity Of Murine/Human Chimeric Antibodies With Specificity For The Human α/β T Cell", J. Immunol. 146(3):928-935; Shearman, C.W. et al. (1991) "Construction, Expression And Characterization of Humanized Antibodies Directed Against The Human α/β T Cell Receptor", J. Immunol. 147(12):4366-4373). Антитела, которые специфично связываются с рецептором NKG2D, включают KYK-2.0 (Kwong, K.Y. et al. (2008) "Generation, Affinity Maturation, And Characterization Of A Human Anti-Human NKG2D Monoclonal Antibody With Dual Antagonistic And Agonistic Activity", J. Mol. Biol. 384:1143-1156; и PCT/US09/54911).
При использовании такого диантитела клетка-мишень становится клеткой, которая может связываться клетками, на которых расположен рецептор (NKG2D). Рецептор NKG2D экспрессируется на всех естественных киллерных клетках человека (и других млекопитающих) (Bauer, S. et al. (1999) "Activation Of NK Cells And T Cells By NKG2D, A Receptor For Stress-Inducible MICA", Science 285(5428):727-729; Jamieson, A.M. et al. (2002) "The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing", Immunity 17(1):19-29), а также на всех CD8+ T-клетках (Groh, V. et al. (2001) "Costimulation Of CD8αβ T Cells By NKG2D Via Engagement By MIC Induced On Virus-Infected Cells", Nat. Immunol. 2(3):255-260; Jamieson, A.M. et al. (2002) "The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing", Immunity 17(1):19-29).
Альтернативно молекулы диантител по настоящему изобретению можно конструировать так, чтобы они содержали домен, который представляет собой связывающий лиганд для T-клеточного рецептора ("TCR"). TCR экспрессируется в природе CD4+ или CD8+ T-клетками и позволяет таким клеткам распознавать антигенные пептиды, которые связываются и представляются белками MHC класса I или класса II антигенпредставляющих клеток. Распознавание комплекса pMHC (пептид-MHC) посредством TCR запускает распространение клеточного иммунного ответа, который приводит к продукции цитокинов и лизису антигенпредставляющей клетки (см., например, Armstrong, K.M. et al. (2008) "Conformational Changes And Flexibility In T-Cell Receptor Recognition Of Peptide-MHC Complexes", Biochem. J. 415(Pt 2):183-196; Willemsen, R. (2008) "Selection Of Human Antibody Fragments Directed Against Tumor T-Cell Epitopes For Adoptive T-Cell Therapy", Cytometry A. 73(11):1093-1099; Beier, K.C. et al. (2007) "Master Switches Of T-Cell Activation And Differentiation", Eur. Respir. J. 29:804-812; Mallone, R. et al. (2005) "Targeting T-Lymphocytes For Immune Monitoring And Intervention In Autoimmune Diabetes", Am. J. Ther. 12(6):534-550).
При конструировании таких молекул диантител, чтобы они дополнительно содержали по меньшей мере один связывающий эпитоп домен, способный связываться, например, с рецептором, присутствующим на поверхности клетки-мишени, такие молекулы диантител будут представлять собой молекулы DART и, таким образом, они будут способны связываться с клетками-мишенями и, тем самым, приводить к тому, чтобы клетки-мишени экспонировали связывающий лиганд для рецептора естественных киллеров группы 2D (NKG2D) или с TCR (в зависимости от того, который из них присутствует на связанном с клеткой-мишенью антителе) (см., например, Germain, C. et al. (2008) "Redirecting NK Cells Mediated Tumor Cell Lysis By A New Recombinant Bifunctional Protein", Prot. Engineer. Design Selection 21(11):665-672).
Такие диантитела можно использовать для преобразования желаемой клетки-мишени в клетку, которая является мишенью опосредуемого NK-клетками лизиса или опосредуемой T-клетками цитотоксичности. В одном из вариантов осуществления связывающий эпитоп домен диантитела, способного связываться с рецептором, присутствующим на поверхности клетки-мишени, представляет собой эпитоп, который связывается с ассоциированным с опухолью антигеном, так чтобы преобразовывать злокачественные клетки в субстраты для опосредуемого NK-клетками лизиса клеток или опосредуемой T-клетками цитотоксичности. Особый интерес представляют ассоциированные с опухолью антигены, которые представляют собой антиген рака молочной железы, антиген рака яичника, антиген рака предстательной железы, антиген рака шейки матки, антиген карциномы поджелудочной железы, антиген рака легкого, антиген рака мочевого пузыря, антиген рака толстого кишечника, антиген рака яичка, антиген глиобластомы, антиген, ассоциированный с B-клеточной злокачественной опухолью, антиген, ассоциированный с множественной миеломой, антиген, ассоциированный с неходжкинской лимфомой, или антиген, ассоциированный с хроническим лимфоцитарным лейкозом.
Пригодные ассоциированные с опухолью антигены для такого применения включают A33 (антиген карциномы ободочной и прямой кишки; Almqvist, Y. 2006, Nucl Med Biol. Nov; 33(8):991-998); B1 (Egloff, A.M. et al. 2006, Cancer Res. 66(1):6-9); BAGE (Bodey, B. 2002 Expert Opin Biol Ther. 2(6):577-84); бета-катенин (Prange W. et al. 2003 J Pathol. 201(2):250-9); CA125 (Bast, R.C. Jr. et al. 2005 Int J. Gynecol Cancer 15 Suppl 3:274-81); CD5 (Calin, G.A. et al. 2006 Semin Oncol. 33(2):167-73; CD19 (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48); CD20 (Thomas, D.A. et al. 2006 Hematol Oncol Clin North Am. 20(5):1125-36); CD22 (Kreitman, R.J. 2006 AAPS J. 18; 8(3):E532-51); CD23 (Rosati, S. et al. 2005 Curr Top Microbiol Immunol. 5; 294:91-107); CD25 (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48); CD27 (Bataille, R. 2006 Haematologica 91(9): 1234-40); CD28 (Bataille, R. 2006 Haematologica 91(9):1234-40); CD36 (Ge, Y. 2005 Lab Hematol. 11(1):31-7); CD40/CD154 (Messmer, D. et al. 2005 Ann N Y Acad Sci. 1062:51-60); CD45 (Jurcic, J.G. 2005 Curr Oncol Rep. 7(5):339-46); CD56 (Bataille, R. 2006 Haematologica 91(9):1234-40); CD79a/CD79b (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48; Chu, P.G. et al. 2001 Appl Immunohistochem Mol. Morphol. 9(2):97-106); CD103 (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48); CDK4 (Lee, Y.M. et al. 2006 Cell Cycle 5(18):2110-4); CEA (карциноэмбриональный антиген; Mathelin, C. 2006 Gynecol Obstet Fertil. 34(7-8):638-46; Tellez-Avila, F.I. et al. 2005 Rev Invest Clin. 57(6):814-9); CTLA4 (Peggs, K.S. et al. 2006 Curr Opin Immunol. 18(2):206-13); EGF-R (рецептор эпидермального фактора роста; Adenis, A. et al. 2003 Bull Cancer. 90 Spec No:S228-32); Erb (ErbBl; ErbB3; ErbB4; Zhou, H. et al. 2002 Oncogene 21(57):8732-40; Rimon, E. et al. 2004 Int J. Oncol. 24(5):1325-38); GAGE (GAGE-1; GAGE-2; Akcakanat, A. et al. 2006 Int J. Cancer. 118(1): 123-8); GD2/GD3/GM2 (Livingston, P.O. et al. 2005 Cancer Immunol Immunother. 54(10):1018-25); gplOO (Lotem, M. et al. 2006 J. Immunother. 29(6):616-27); HER-2/neu (Kumar, Pal S et al. 2006 Semin Oncol. 33(4):386-91); E6 вируса папилломы человека/E7 вируса папилломы человека (DiMaio, D. et al. 2006 Adv Virus Res. 66:125-59; KSA (17-1A) (Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); MAGE (MAGE-1; MAGE-3; (Bodey, B. 2002 Expert Opin Biol Ther. 2(6):577-84); MART (Kounalakis, N. et al. 2005 Curr Oncol Rep. 7(5):377-82); MUC-1 (Mathelin, C. 2006 Gynecol Obstet Fertil. 34(7-8):638-46); MUM-1 (Castelli, C. et al. 2000 J. Cell Physiol. 182(3):323-31); N-ацетилглюкозаминилтрансферазу (Dennis, J.W. 1999 Biochim Biophys Acta. 6;1473(l):21-34); pl5 (Gil, J. et al. 2006 Nat Rev Mol. Cell Biol. 7(9):667-77); PSA (простатспецифический антиген; Cracco, C.M. et al. 2005 Minerva Urol Nefrol. 57(4):301-11); PSMA (Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); sTn (Holmberg, L.A. 2001 Expert Opin Biol Ther. 1(5):881-91); рецептор TNF (рецептор TNF-α, рецептор TNF-β или рецептор TNF-γ рецептор; van Horssen, R. et al. 2006 Oncologist. 11(4):397-408; Gardnerova, M. et al. 2000 Curr Drug Targets. l(4):327-64); или рецептор VEGF (O'Dwyer. PJ. 2006 Oncologist. 11(9):992-8).
Дополнительные ассоциированные с опухолью антигены для такого применения (и публикации, в которых описаны специфически реактивные антитела к таким антигенам) включают ADAM-9 (публикация патента США № 2006/0172350; публикация PCT WO 06/084075); ALCAM (публикация PCT WO 03/093443); карбоксипептидазу M (публикация патента США № 2006/0166291); CD46 (патент США № 7148038; публикация PCT WO 03/032814); цитокератин 8 (публикация PCT WO 03/024191); рецепторы эфрина (и, в частности, EphA2 (патент США № 7569672; публикация PCT WO 06/084226)); интегрин альфа-V-Beta-6 (публикация PCT WO 03/087340); JAM-3 (публикация PCT WO 06/084078); KID3 (публикация PCT WO 05/028498); KID31 (публикация PCT WO 06/076584); LUCA-2 (публикация патента США № 2006/0172349; публикация PCT WO 06/083852); онкостатин M (рецептор-бета онкостатина) (патент США № 7572896; публикация PCT WO 06/084092); PIPA (патент США № 7405061; публикация PCT WO 04/043239); RAAG10 (патент США № 7527969; публикация PCT WO 04/001381); ROR1 (патент США № 5843749); TES7 (публикация PCT WO 08/066691); и рецептор трансферрина (патент США № 7572895; публикация PCT WO 05/121179).
Также интерес представляют антигены, специфичные к конкретным инфекционным агентам, например, вирусным агентам, включая, но не ограничиваясь ими, вирус иммунодефицита человека (HIV), вирус гепатита B (HBV), вирус гриппа, вирус папилломы человека (HPV), вирус ящура (вирус Коксаки), вирус бешенства, вирус простого герпеса (HSV) и этиологические факторы гастроэнтерита, включая ротавирусы, аденовирусы, калицивирусы, астровирусы и вирус Норволк; бактериальные агенты, включая, но не ограничиваясь ими, E. coli, Salmonella thyphimurium, Pseudomonas aeruginosa, Vibrio cholerae, Neisseria gonorrhoeae, Helicobacter pylori, Hemophilus influenzae, Shigella dysenteriae, Staphylococcus aureus, Mycobacterium tuberculosis и Streptococcus pneumoniae, грибные агенты и паразиты, такие как Giardi.
Альтернативно такой эпитоп может связываться с Fc-рецептором (например, FcγRI или FcγRII), так чтобы, например, превращать клетки острого моноцитарного лейкоза в субстраты для опосредуемого NK-клетками лизиса клеток.
5.1 СВЯЗЫВАЮЩИЕ ДОМЕНЫ ДИАНТИТЕЛА
Диантитела по настоящему изобретению содержат антигенсвязывающие домены, главным образом, происходящие из иммуноглобулинов или антител. Антитела, из которых происходят связывающие домены, используемые в способах по изобретению, могут быть из любого животного, включая птиц и млекопитающих (например, человека, не являющегося человеком примата, мыши, осла, овцы, кролика, козы, морской свинки, верблюда, лошади или курицы). Предпочтительно антитела представляют собой моноклональные антитела человека или гуманизированные моноклональные антитела. Как используют в настоящем документе, антитела "человека" включают антитела, имеющие аминокислотную последовательность иммуноглобулина человека, и включают антитела, выделенные из библиотек иммуноглобулинов человека или библиотек кодирующих последовательностей для синтетических иммуноглобулинов человека, или из мышей, которые экспрессируют антитела с генов человека.
Изобретение относится к применению любых антител, известных в данной области, для лечения и/или профилактики злокачественной опухоли, аутоиммунного заболевания, воспалительного заболевания или инфекционного заболевания, в качестве источника связывающих доменов для диантител по изобретению. Неограничивающие примеры известных антител против злокачественной опухоли представлены в разделе 5.7.1, а также ими являются другие антитела, специфичные к приведенным антигенам-мишеням, и антитела против антигенов злокачественной опухоли, приведенных в разделе 5.6.1; неограничивающие примеры известных антител для лечения и/или профилактики аутоиммунного заболевания и воспалительного заболевания представлены в разделе 5.7.2., а также ими являются антитела против приведенных антигенов-мишеней и антитела против антигенов, приведенных в разделе 5.6.2; в других вариантах осуществления можно использовать антитела против эпитопов, ассоциированных с инфекционными заболеваниями, приведенными в разделе 5.6.3. В определенных вариантах осуществления антитела содержат вариант Fc-области, содержащий одну или несколько модификаций аминокислот, которые идентифицированы способами по изобретению в качестве имеющих приданную им эффекторную функцию и/или усиленную аффинность к FcγRIIB и сниженную аффинность к FcγRIIIA относительно сравнимой молекулы, содержащей Fc-область дикого типа. Неограничивающий пример антител, которые используют для лечения или профилактики воспалительных нарушений, которые могут быть сконструированы согласно изобретению, представлен в таблице 9, и неограничивающий пример антител, которые используют для лечения или профилактики аутоиммунных нарушений, представлен в таблице 10.
Для некоторых применений, включая применение антител in vivo у человека и в анализах детекции in vitro, может быть предпочтительным использовать диантитела с вариабельными доменами, происходящими из антител человека, химерных или гуманизированных антител. Вариабельные домены из полностью человеческих антител являются особенно желательными для терапевтического лечения людей. Антитела человека можно получать различными способами, известными в данной области, включая способы фагового дисплея, описанные выше, с использованием библиотек антител, полученных из последовательностей иммуноглобулинов человека. Также см. патенты США №№ 4444887 и 4716111; и международные публикации WO 98/46645, WO 98/50433, WO 98/24893, WO 98/16654, WO 96/34096, WO 96/33735 и WO 91/10741; которые все включены в настоящий документ в качестве ссылок в полном объеме.
Гуманизированное антитело представляет собой антитело, его вариант или фрагмент, которые способны связываться с заданным антигеном и которые содержат каркасную область, по существу имеющую аминокислотную последовательность иммуноглобулина человека, и CDR, по существу имеющую аминокислотную последовательность иммуноглобулина, не являющегося человеческим. Гуманизированное антитело может содержать по существу все из по меньшей мере одного, и, как правило, двух, вариабельных доменов, в которых все или по существу все из CDR-областей соответствуют CDR-областям не являющегося человеческим иммуноглобулина (т.е. донорного антитела), и все или по существу все из каркасных областей представляют собой каркасные области консенсусной последовательности иммуноглобулина человека.
Каркасная область и CDR-области гуманизированного антитела не должны точно соответствовать исходным последовательностям, например, донорная CDR или консенсусная каркасная область могут быть подвергнуты мутагенезу путем замены, вставки или делеции по меньшей мере одного остатка, так чтобы остаток CDR или каркасной области в этом участке не соответствовал ни консенсусному, ни и донорному антителу. Однако такие мутации предпочтительно не являются протяженными. Обычно, по меньшей мере 75% остатков гуманизированного антитела соответствуют остаткам исходной последовательности каркасной области (FR) и CDR, более часто 90%, и наиболее предпочтительно более 95%. Гуманизированные антитела можно получать с использованием различных способов, известных в данной области, включая, но не ограничиваясь ими, пересадку CDR (патент Европы EP 239400; международная публикация WO 91/09967; и патенты США №№ 5225539, 5530101 и 5585089), гиперхимеризацию или изменение поверхности (патенты Европы EP 592106 и EP 519596; Padlan (1991) "A Possible Procedure For Reducing The Immunogenecity Of Antibody Variable Domains While Preserving Their Ligand-Binding Properties", Molecular Immunology 28(4/5):489-498; Studnicka et al. (1994) "Human-Engineered Monoclonal Antibodies Retain Full Specific Binding Activity By Preserving Non-CDR Complementarity-Modulating Residues", Protein Engineering 7(6):805-814; и Roguska et al. (1994) "Humanization Of Murine Monoclonal Antibodies Through Variable Domain Resurfacing", Proc Natl Acad Sci USA 91:969-973), перетасовку цепей (патент США № 5565332), и способы, описанные, например, в патентах США №№ 6407213, 5766886, 5585089, международной публикации WO 9317105, Tan et al. (2002) "'Super humanized' Antibodies: Reduction Of Immunogenic Potential By Complementarity-Determining Region Grafting With Human Germline Sequences: Application To An Anti-CD28", J. Immunol. 169:1119-25, Caldas et al. (2000) "Design And Synthesis Of Germline-Based Hemi-Humanized Single-Chain Fv Against The CD 18 Surface Antigen", Protein Eng. 13:353-60, Morea et al. (2000) "Antibody Modeling: Implications For Engineering And Design", Methods 20:267-79, Baca et al. (1997) "Antibody Humanization Using Monovalent Phage Display", J. Biol. Chem. 272:10678-84, Roguska et al. (1996) "A Comparison Of Two Murine Monoclonal Antibodies Humanized By CDR-Grafting And Variable Domain Resurfacing", Protein Eng. 9:895-904, Couto et al. (1995) "Designing Human Consensus Antibodies With Minimal Positional Templates", Cancer Res. 55 (23 Supp):5973s-5977s, Couto et al. (1995) "Anti-BA46 Monoclonal Antibody Mc3: Humanization Using A Novel Positional Consensus And In Vivo And In Vitro Characterization", Cancer Res. 55:1717-22, Sandhu (1994) "A Rapid Procedure For The Humanization Of Monoclonal Antibodies", Gene 150:409-10, Pedersen et al. (1994) "Comparison Of Surface Accessible Residues In Human And Murine Immunoglobulin Fv Domains. Implication For Humanization Of Murine Antibodies", J. Mol. Biol. 235:959-973, Jones et al. (1986) "Replacing The Complementarity-Determining Regions In A Human Antibody With Those From A Mouse", Nature 321:522-525, Riechmann et al. (1988) "Reshaping Human Antibodies For Therapy", Nature 332:323-327, и Presta (1992) "Antibody Engineering", Curr. Op. Biotech. 3(4):394-398. Часто каркасные остатки в каркасных областях заменяют соответствующим остатком из антитела-донора CDR для изменения, предпочтительно повышения, связывания антигена. Эти замены в каркасной области идентифицируют способами, хорошо известными в данной области, например, путем моделирования взаимодействий остатков CDR и каркасной области для идентификации остатков каркасной области, важных для связывания антигена, и сравнения последовательностей для идентификации необычных остатков каркасной области в конкретных положениях. (См., например, Queen et al., патент США № 5585089; публикации США №№ 2004/0049014 и 2003/0229208; патенты США №№ 6350861; 6180370; 5693762; 5693761; 5585089 и 5530101, и Riechmann et al. (1988) "Reshaping Human Antibodies For Therapy", Nature 332:323-327, которые все включены в настоящий документ в качестве ссылок в полном объеме).
В наиболее предпочтительном варианте осуществления гуманизированный связывающий домен специфично связывается с тем же эпитопом, что и донорное антитело мыши. Специалисту в данной области будет понятно, что изобретение, в целом, охватывает пересадку CDR антител. Таким образом, донорное и акцепторное антитела могут происходить из животных одного вида и даже из одного класса или подкласса антител. Однако более часто донорное и акцепторное антитела происходят из животных различных видов. Как правило, донорное антитело представляет собой не являющееся человеческим антитело, такое как mAb грызуна, и акцепторное антитело представляет собой антитело человека.
В некоторых вариантах осуществления пересаживают по меньшей мере одну CDR из донорного антитела в антитело человека. В других вариантах осуществления в антитело человека пересаживают по меньшей мере две и предпочтительно все три CDR каждой из вариабельных областей тяжелой и/или легкой цепей. CDR могут содержать CDR по Kabat, CDR структурных петель или их комбинации. В некоторых вариантах осуществления изобретение охватывает гуманизированное антитело против FcγRIIB, содержащее по меньшей мере одну тяжелую цепь с пересаженным CDR и по меньшей мере одну легкую цепь с пересаженным CDR.
Диантитела, используемые в способах по изобретению, включают производные, которые являются модифицированными, т.е. путем ковалентного присоединения любого типа молекулы к диантителу. Например, но не ограничиваясь этим, производные диантитела включают диантитела, модифицированные, например, гликозилированием, ацетилированием, пегилированием, фосфорилированием, амидацией, преобразованием в производное с помощью известных защитных/блокирующих групп, протеолитическим расщеплением, связыванием с клеточным лигандом или другим белком и т.д. Любые из множества химических модификаций можно проводить известными способами, включая, но не ограничиваясь ими, специфическое химическое расщепление, ацетилирование, формилирование, метаболический синтез туникамицина и т.д. Кроме того, производные могут содержать одну или несколько неклассических аминокислот.
Химерное антитело представляет собой молекулу, в которой различные части антитела происходят из различных молекул иммуноглобулинов, таких как антитела, имеющие вариабельную область, происходящую из не являющегося человеческим антитела, и константную область иммуноглобулина человека. Способы получения химерных антител известны в данной области. См. например, Morrison (1985) "Transfectomas Provide Novel Chimeric Antibodies", Science 229:1202-1207; Oi et al. (1986) "Chimeric Antibodies", BioTechniques 4:214-221; Gillies et al. (1989) "High-Level Expression Of Chimeric Antibodies Using Adapted cDNA Variable Region Cassettes", J. Immunol. Methods 125:191-202; и патенты США №№ 6311415, 5807715, 4816567 и 4816397, которые включены в настоящий документ в качестве ссылок в полном объеме.
Часто каркасные остатки в каркасных областях заменяют соответствующим остатком из антитела-донора CDR для изменения, предпочтительно повышения, связывания антигена. Эти замены в каркасной области идентифицируют способами, хорошо известными в данной области, например, путем моделирования взаимодействий CDR и каркасных остатков для идентификации каркасных остатков, важных для связывания антигена, и сравнения последовательностей для идентификации необычных каркасных остатков в конкретных положениях (см., например, патент США № 5585089; и Riechmann et al. (1988) "Reshaping Human Antibodies For Therapy", Nature 332:323-327, которые включены в настоящий документ в качестве ссылок в полном объеме).
Моноклональные антитела, из которых можно получать связывающие домены диантител по изобретению, можно получать с использованием широкого множества способов, известных в данной области, включая применение гибридомной, рекомбинантной технологий и технологии фагового дисплея или их комбинации. Например, моноклональные антитела можно получать с использованием способов гибридом, включая способы, известные в данной области и описанные, например, в Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling, et al.: Monoclonal Antibodies and T-Cell Hybridomas, pp. 563-681 (Elsevier, N.Y., 1981) (обе из которых включены в качестве ссылок в полном объеме). Термин "моноклональные антитело", как используют в настоящем документе, не ограничивается антителами, полученными с помощью технологии гибридом. Термин "моноклональное антитело" относится к антителу, полученному из одного клона, включая любой эукариотический, прокариотический или фаговый клон, а не к способу, которым оно получено.
Способы получения и скрининга специфических антител с использованием технологии гибридом являются общепринятыми и хорошо известны в данной области. В неограничивающем примере мышей можно иммунизировать представляющим интерес антигеном или клеткой, экспрессирующей такой антиген. После выявления иммунного ответа, например, детекции антител, специфичных к антигену, в сыворотке мыши, селезенку мышей извлекают и выделяют спленоциты. Затем спленоциты подвергают слиянию хорошо известными способами с любыми пригодными миеломными клетками. Гибридомы отбирают и клонируют способом лимитирующих разведений. Затем клоны гибридом анализируют способами, известными в данной области, в отношении клеток, которые секретируют антитела, способные связываться с антигеном. Асцитную жидкость, которая обычно содержит высокие уровни антител, можно получать, инокулируя внутрибрюшинно мышам положительные клоны гибридом. Представляющие интерес антигены включают, но не ограничиваются ими, антигены, ассоциированные со злокачественными опухолями, представленные в разделе 5.8.1, антигены, ассоциированные с аутоиммунными заболеваниями и воспалительными заболеваниями, представленные в разделе 5.8.2, антигены, ассоциированные с инфекционными заболеваниями, представленные в разделе 5.8.3, и токсины, представленные в разделе 5.8.4.
Также антитела можно получать с использованием различных способов фагового дисплея, известных в данной области. В способах фагового дисплея функциональные домены антитела экспонируют на поверхности фаговых частиц, которые обладают кодирующими их полинуклеотидными последовательностями. В конкретном варианте осуществления такой фаг можно использовать для экспонирования антигенсвязывающих доменов, таких как Fab или Fv или стабилизированный дисульфидной связью Fv, экспрессированных из библиотеки набора антител или комбинаторной библиотеки антител (например, человека или мыши). Фаг, экспрессирующий антигенсвязывающий домен, который связывает представляющий интерес антиген, можно подвергать селекции и идентифицировать с помощью антигена, например, с использованием меченого антигена или антигена, связанного или фиксированного на твердой поверхности или гранулах. Фаг, используемый в этих способах, как правило, представляет собой нитевидный фаг, включающий fd и M13. Антигенсвязывающие домены экспрессируются в качестве белка, слитого рекомбинантными способами с фаговым белком гена III или гена VIII. Примеры способов фагового дисплея, которые можно использовать для получения антител по настоящему изобретению, включают способы, описанные в Brinkmann et al. (1995) "Phage Display Of Disulfide-Stabilized Fv Fragments", J. Immunol. Methods, 182:41-50; Ames et al. (1995) "Conversion Of Murine Fabs Isolated From A Combinatorial Phage Display Library To Full Length Immunoglobulins", J. Immunol. Method, 184:177-186; Kettleborough et al. (1994) "Isolation Of Tumor Cell-Specific Single-Chain Fv From Immunized Mice Using Phage-Antibody Libraries And The Re-Construction Of Whole Antibodies From These Antibody Fragments", Eur. J. Immunol., 24:952-958; Persic et al. (1997) "An Integrated Vector System For The Eucariotic Expression Of Antibodies Or Their Fragments After Selection From Phage Display Libraries", Gene, 187:9-18; Burton et al. (1994) "Human Antibodies From Combinatorial Libraries", Advances in Immunology, 57:191-280; заявке PCT № PCT/GB91/01134; заявках PCT WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401; и патентах США №№ 5698426; 5223409; 5403484; 5580717; 5427908; 5750753; 5821047; 5571698; 5427908; 5516637; 5780225; 5658727; 5733743 и 5969108; которые все включены в настоящий документ в качестве ссылок в полном объеме.
Технологию фагового дисплея можно использовать для повышения аффинности антитела к его антигену. Этот способ может быть пригоден для получения высокоаффинных антител. В технологии, называемой созреванием аффинности, используется мутагенез или "прогулка" по CDR и реселекция с использованием собственно антигена для идентификации антител, которые связываются с антигеном с более высокой аффинностью к антигену по сравнению с исходным или родительским антителом (см., например, Glaser et al. (1992) "Dissection Of The Combining Site In A Humanized Anti-Tac Antibody", J. Immunology 149:2607-2614). Мутагенез полных кодонов, а не отдельных нуклеотидов, приводит к полурандомизированному набору аминокислотных мутаций. Можно конструировать библиотеки, состоящие из совокупности вариантов клонов, каждый из которых отличается изменением одной аминокислоты в одной CDR, которые содержат варианты, соответствующие каждой возможной аминокислотной замене для каждого остатка CDR. Скрининг мутантов с увеличенной аффинностью связывания с антигеном можно проводить путем контактирования иммобилизованных мутантов с меченым антигеном. Для идентификации мутантных антител с увеличенной авидностью к антигену можно использовать любой способ скрининга, известный в данной области (например, ELISA) (см. Wu et al. (1998) "Stepwise In Vitro Affinity Maturation Of Vitaxin, An Alphav Beta3-Specific Humanized mAb", Proc Natl. Acad Sci. USA 95:6037-6042; Yelton et al. (1995) "Affinity Maturation Of The Br96 Anti-Carcinoma Antibody By Codon-Based Mutagenesis", J. Immunology 155:1994-2004). Также возможна "прогулка" по CDR, которая рандомизирует легкую цепь (см. Schier et al. (1996) "Isolation Of Picomolar Affinity Anti-C-ErbB-2 Single-Chain Fv By Molecular Evolution Of The Complementarity Determining Regions In The Center Of The Antibody Binding Site", J. Mol. Bio. 263:551-567).
Также настоящее изобретение охватывает применение связывающих доменов, содержащих аминокислотную последовательность любого из связывающих доменов, описанных в настоящем документе или известных в данной области, с мутациями (например, одной или несколькими аминокислотными заменами) в каркасных областях или CDR. Предпочтительно мутации в этих связывающих доменах сохраняют или повышают авидность и/или аффинность связывающих доменов к FcγRIIB, с которым они иммуноспецифически связываются. Для анализа аффинности антитела к конкретному антигену можно использовать стандартные способы, известные специалистам в данной области (например, иммуноанализы).
Для внесения мутаций в нуклеотидную последовательность, кодирующую антитело или его фрагмент, можно использовать стандартные способы, известные специалистам в данной области, включая, например, сайт-направленный мутагенез и опосредуемый ПЦР мутагенез, которые приводят к аминокислотным заменам. Предпочтительно производные включают менее 15 аминокислотных замен, менее 10 аминокислотных замен, менее 5 аминокислотных замен, менее 4 аминокислотных замен, менее 3 аминокислотных замен или менее 2 аминокислотных замен относительно исходного антитела или его фрагмента. В предпочтительном варианте осуществления производные имеют консервативные аминокислотные замены, внесенные в один или несколько предсказанных не необходимых аминокислотных остатков.
5.1.1 ДИАНТИТЕЛА, СОДЕРЖАЩИЕ УЧАСТКИ СВЯЗЫВАНИЯ ЭПИТОПОВ, КОТОРЫЕ ИММУНОСПЕЦИФИЧЕСКИ СВЯЗЫВАЮТ FcγRIIB
В конкретном варианте осуществления по меньшей мере один из связывающих доменов диантител по изобретению осуществляет агонизм по меньшей мере одному виду активности FcγRIIB. В одном из вариантов осуществления изобретения указанная активность представляет собой ингибирование опосредуемой B-клеточным рецептором передачи сигнала. В другом варианте осуществления связывающий домен ингибирует активацию B-клеток, пролиферацию B-клеток, продукцию антител, вхождение кальция внутрь B-клеток, прогрессирование клеточного цикла или активность одной или нескольких нижерасположенных молекул передачи сигнала в каскаде передачи сигнала FcγRIIB. В другом варианте осуществления связывающий домен усиливает фосфорилирование FcγRIIB или привлечение SHIP. В следующем варианте осуществления изобретения связывающий домен ингибирует активность MAP-киназы или привлечение Akt в опосредуемом B-клеточным рецептором каскаде передачи сигнала. В другом варианте осуществления связывающий домен осуществляет агонизм опосредуемого FcγRIIB ингибирования передачи сигнала FcεRI. В конкретном варианте осуществления указанный связывающий домен ингибирует индуцируемую FcεRI активацию тучных клеток, мобилизацию кальция, дегрануляцию, продукцию цитокинов или высвобождение серотонина. В другом варианте осуществления связывающие домены по изобретению стимулируют фосфорилирование FcγRIIB, стимулируют привлечение SHIP, стимулируют фосфорилирование SHIP и его связывание с Shc или ингибирует активацию членов семейства MAP-киназ (например, Erk1, Erk2, JNK, p38 и т.д.). В другом варианте осуществления связывающие домены по изобретению усиливают фосфорилирование тирозина p62dok и его связывание с SHIP и rasGAP. В другом варианте осуществления связывающие домены по изобретению ингибируют опосредуемый FcγR фагоцитоз в моноцитах или макрофагах.
В другом варианте осуществления связывающие домены осуществляют антагонизм по меньшей мере одному виду активности FcγRIIB. В одном из вариантов осуществления указанная активность представляет собой активацию опосредуемой B-клеточным рецептором передачу сигнала. В конкретном варианте осуществления связывающий домен усиливает активацию B-клеток, пролиферацию B-клеток, продукцию антител, вхождение кальция внутрь B-клеток, прогрессирование клеточного цикла или активность одной или нескольких нижерасположенных молекул передачи сигнала в каскаде передачи сигнала FcγRIIB. В другом варианте осуществления связывающий домен снижает фосфорилирование FcγRIIB или привлечение SHIP. В следующем варианте осуществления изобретения связывающий домен повышает активность MAP-киназы или привлечение Akt в опосредуемом B-клеточным рецептором каскаде передачи сигнала. В другом варианте осуществления связывающий домен осуществляет антагонизм опосредуемого FcγRIIB ингибирования передачи сигнала FcεRI. В конкретном варианте осуществления указанный связывающий домен усиливает индуцируемую FcεRI активацию тучных клеток, мобилизацию кальция, дегрануляцию, продукцию цитокинов или высвобождение серотонина. В другом варианте осуществления связывающие домены по изобретению ингибируют фосфорилирование FcγRIIB, ингибируют привлечение SHIP, ингибируют фосфорилирование SHIP и его связывание с Shc, или усиливают активацию членов семейства MAP-киназ (например, Erk1, Erk2, JNK, p38 и т.д.). В другом варианте осуществления связывающие домены по изобретению ингибируют фосфорилирование тирозина p62dok и его связывание с SHIP и rasGAP. В другом варианте осуществления связывающие домены по изобретению усиливают опосредуемый FcγR фагоцитоз в моноцитах или макрофагах. В другом варианте осуществления связывающие домены препятствуют фагоцитозу, выведению опсонизированных частиц макрофагами селезенки.
В других вариантах осуществления по меньшей мере один из связывающих доменов можно использовать для нацеливания диантител по изобретению на клетки, которые экспрессируют FcγRIIB.
В одном конкретном варианте осуществления один из связывающих доменов происходит из моноклонального антитела мыши, продуцированного клонами 2B6 или 3H7, имеющими регистрационные номера ATCC PTA-4591 и PTA-4592, соответственно. Гибридомы, продуцирующие антитела 2B6 и 3H7, были депонированы в American Type Culture Collection (10801 University Blvd., Manassas, VA. 20110-2209) 13 августа 2002 года согласно условиям Будапештского договора о международном признании депонирования микроорганизмов для целей патентных процедур, и им были присвоены регистрационные номера PTA-4591 (для гибридомы, продуцирующей 2B6) и PTA-4592 (для гибридомы, продуцирующей 3H7), соответственно, и они включены в настоящий документ в качестве ссылок. В предпочтительном варианте осуществления связывающие домены являются человеческими или гуманизированными, предпочтительно они происходят из гуманизированного варианта антитела, продуцированного клоном 3H7 или 2B6.
Также изобретение охватывает диантитела со связывающими доменами из других антител, которые специфично связывают FcγRIIB, предпочтительно FcγRIIB человека, более предпочтительно нативный FcγRIIB человека, которые получены из клонов, включающих, но не ограничивающихся ими, 1D5, 2E1, 2H9, 2D11 и 1F2, имеющие регистрационные номера ATCC PTA-5958, PTA-5961, PTA-5962, PTA-5960 и PTA-5959, соответственно. Гибридомы, продуцирующие указанные выше клоны, были депонированы согласно условиям Будапештского договора в American Type Culture Collection (10801 University Blvd., Manassas, VA. 20110-2209) 7 мая 2004 года и включены в настоящий документ в качестве ссылок. В предпочтительных вариантах осуществления связывающие домены из антител, описанных выше, являются гуманизированными.
В конкретном варианте осуществления связывающие домены, используемые в диантителах по настоящему изобретению, происходят из антитела или его антигенсвязывающего фрагмента (например, содержащего одну или несколько определяющих комплементарность областей (CDR), предпочтительно все 6 CDR) антитела, продуцированного клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2. В другом варианте осуществления связывающий домен связывается с тем же эпитопом, что и моноклональное антитело мыши, продуцированное клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2, соответственно, и/или конкурирует с моноклональным антителом мыши, продуцированным клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2, при определении, например, с помощью анализа ELISA или другого подходящего конкурентного иммуноанализа, и также связывает FcγRIIB с более высокой аффинностью, чем аффинность, с которой связывающий домен связывает FcγRIIA.
Также настоящее изобретение охватывает диантитела со связывающими доменами, содержащими аминокислотную последовательность вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, которая по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99% идентична аминокислотной последовательности вариабельной области тяжелой цепи и/или легкой цепи моноклонального антитела мыши, продуцированного клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2. Кроме того, настоящее изобретение охватывает диантитела со связывающими доменами, которые специфично связывают FcγRIIB с более высокой аффинностью, чем аффинность, с которой указанные антитело или его фрагмент связывают FcγRIIA, и которые содержат аминокислотную последовательность одной или нескольких CDR, которая по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99% идентична аминокислотной последовательности одного или нескольких CDR моноклонального антитела мыши, продуцированного клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2. Определение процентной идентичности двух аминокислотных последовательностей можно проводить любым способом, известным специалисту в данной области, включая поиск белков в BLAST.
Также настоящее изобретение охватывает применение диантител, содержащих связывающие домены, которые специфично связывают FcγRIIB с более высокой аффинностью, чем аффинность, с которой связывающий домен связывает FcγRIIA, которые кодируются нуклеотидной последовательностью, гибридизующейся с нуклеотидной последовательностью моноклонального антитела мыши, продуцированного клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2 в строгих условиях. В предпочтительном варианте осуществления связывающий домен специфично связывает FcγRIIB с более высокой аффинностью, чем FcγRIIA, и содержит вариабельную область легкой цепи и/или вариабельную область тяжелой цепи, кодируемые нуклеотидной последовательностью, которая гибридизуется в строгих условиях с нуклеотидной последовательностью вариабельной области легкой цепи и/или вариабельной области тяжелой цепи моноклонального антитела мыши, продуцированного клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2 в строгих условиях. В другом предпочтительном варианте осуществления связывающие домены специфично связывают FcγRIIB с более высокой аффинностью, чем FcγRIIA, и содержат одну или несколько CDR, кодируемых нуклеотидной последовательностью, которая гибридизуется в строгих условиях с нуклеотидной последовательностью одной или нескольких CDR моноклонального антитела мыши, продуцированного клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2. Строгие условия гибридизации включают, но не ограничиваются ими, гибридизацию со связанной с фильтром ДНК в 6X растворе хлорида натрия/цитрата натрия (SSC) при приблизительно 45°C, а затем одно или несколько промываний в 0,2X SSC/0,1% SDS при приблизительно 50-65°С, условия высокой строгости, такие как гибридизация со связанной с фильтром ДНК в 6X SSC при приблизительно 45°C, а затем одно или несколько промываний в 0,1X SSC/0,2% SDS при приблизительно 60°С, или любые другие строгие условия гибридизации, известные специалистам в данной области (см., например, Ausubel, F.M. et al., eds. 1989 Current Protocols in Molecular Biology, vol. 1, Green Publishing Associates, Inc. and John Wiley and Sons, Inc., NY на стр. с 6.3.1 по 6.3.6 и 2.10.3, включенную в настоящий документ в качестве ссылки).
Также настоящее изобретение охватывает применение связывающих доменов, содержащих аминокислотную последовательность любого из связывающих доменов, описанных выше, с мутациями (например, одной или несколькими аминокислотными заменами) в каркасных областях или CDR. Предпочтительно мутации в этих связывающих доменах сохраняют или повышают авидность и/или аффинность связывающих доменов к FcγRIIB, с которым они иммуноспецифически связываются. Для анализа аффинности антитела к конкретному антигену можно использовать стандартные способы, известные специалистам в данной области (например, иммуноанализы).
Для внесения мутаций в нуклеотидную последовательность, кодирующую антитело или его фрагмент, можно использовать стандартные способы, известные специалистам в данной области, включая, например, сайт-направленный мутагенез и ПЦР-опосредуемый мутагенез, которые приводят к аминокислотным заменам. Предпочтительно производные включают менее 15 аминокислотных замен, менее 10 аминокислотных замен, менее 5 аминокислотных замен, менее 4 аминокислотных замен, менее 3 аминокислотных замен или менее 2 аминокислотных замен относительно исходного антитела или его фрагмента. В предпочтительном варианте осуществления производные имеют консервативные аминокислотные замены, внесенные вместо одного или нескольких предсказанных не необходимых аминокислотных остатков.
В предпочтительных вариантах осуществления связывающие домены происходят из гуманизированных антител. Гуманизированное специфичное к FcγRIIB антитело может содержать по существу все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или по существу все из областей CDR соответствуют областям не являющегося человеческим иммуноглобулина (т.е. донорного антитела), и все или по существу все из каркасных областей представляют собой каркасные области консенсусной последовательности иммуноглобулина человека.
Диантитела по настоящему изобретению содержат гуманизированные вариабельные домены, специфичные к FcγRIIB, в которых один или несколько участков одного или нескольких CDR вариабельных областей тяжелой и/или легкой цепей антитела человека (реципиентного антитела) заменены аналогичными частями одного или нескольких CDR донорного моноклонального антитела, которое специфично связывает FcγRIIB с более высокой аффинностью, чем FcγRIIA, например, моноклонального антитела, продуцированного клоном 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2. В других вариантах осуществления гуманизированные антитела связываются с тем же эпитопом, что и 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2, соответственно.
В предпочтительном варианте осуществления области CDR гуманизированного связывающего FcγRIIB домена происходят из антитела мыши, специфичного к FcγRIIB. В некоторых вариантах осуществления гуманизированные антитела, описанные в настоящем документе, содержат изменения, включая, но не ограничиваясь ими, делеции, инсерции, модификации аминокислот, в акцепторном антителе, т.е. каркасных областях вариабельного домена тяжелой и/или легкой цепей человека, которые необходимы для сохранения специфичности связывания донорного моноклонального антитела. В некоторых вариантах осуществления каркасные области гуманизированных антител, описанных в настоящем документе, не обязательно состоят из точной аминокислотной последовательности каркасной области природной вариабельной области антитела человека, а содержит различные изменения, включая, но не ограничиваясь ими, делеции, инсерции, модификации аминокислот, которые изменяют свойство гуманизированного антитела, например, повышают связывающие свойства гуманизированной области антитела, которое специфично к той же мишени, что и антитело мыши, специфичное к FcγRIIB. В наиболее предпочтительных вариантах осуществления в каркасную область вносят минимальное количество изменений, чтобы избежать масштабного внесения не являющихся человеческими остатков каркасной области и для обеспечения минимальной иммуногенности гуманизированного антитела у человека. Донорное моноклональные антитело предпочтительно представляет собой моноклональное антитело, продуцированное клонами 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2.
В конкретном варианте осуществления связывающий домен охватывает вариабельные домены антитела с пересаженными CDR, которые специфично связывают FcγRIIB с более высокой аффинностью, чем аффинность, с которой указанное антитело связывает FcγRIIA, где антитело с пересаженной CDR содержит домен вариабельной области тяжелой цепи, содержащий каркасные остатки реципиентного антитела и остатки из донорного моноклонального антитела, которое специфично связывает FcγRIIB с аффинностью, превышающей аффинность, с которой указанное антитело связывает FcγRIIA, например, моноклонального антитела, продуцированного из клонов 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2. В другом конкретном варианте осуществления диантитела по изобретению содержат вариабельные домены из антитела с пересаженными CDR, которые специфично связывают FcγRIIB с более высокой аффинностью, чем аффинность, с которой указанное антитело связывает FcγRIIA, где антитело с пересаженными CDR содержит домен вариабельной области легкой цепи, содержащий каркасные остатки реципиентного антитела и остатки из донорного моноклонального антитела, которое специфично связывает FcγRIIB с более высокой аффинностью, чем аффинность, с которой указанное антитело связывает FcγRIIA, например, моноклонального антитела, продуцированного клонами 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2.
Вариабельные домены гуманизированного антитела против FcγRIIB, используемые в изобретении, могут иметь вариабельную область тяжелой цепи, содержащую аминокислотную последовательность CDR1 (SEQ ID NO: 24 или SEQ ID NO: 25) и/или CDR2 (SEQ ID NO: 26 или SEQ ID NO: 27) и/или CDR3 (SEQ ID NO: 28 или SEQ ID NO: 29), и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность CDR1 (SEQ ID NO: 30 или SEQ ID NO: 31) и/или CDR2 (SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35) и/или CDR3 (SEQ ID NO: 36 или SEQ ID NO: 37).
В одном конкретном варианте осуществления диантитело содержит вариабельные домены из гуманизированного антитела 2B6, где VH-область состоит из FR-сегментов, взятых из VH-сегментов VH1-18 эмбрионального типа человека (Matsuda et al. (1998) "The Complete Nucleotide sequence Of The Human Immunoglobulin Heavy Chain Variable Region Locus", J. Exp. Med. 188:2151-2162) и JH6 (Ravetch et al. (1981) "Structure Of The Human Immunoglobulin Mu Locus: Characterization Of Embryonic And Rearranged J And D Genes", Cell 27(3 Pt. 2):583-91), и одну или несколько CDR-областей VH 2B6, имеющего аминокислотную последовательность SEQ ID NO: 24, SEQ ID NO: 26 или SEQ ID NO: 28. В одном варианте осуществления VH 2B6 имеет аминокислотную последовательность SEQ ID NO: 38. В другом варианте осуществления VH-домен 2B6 имеет аминокислотную последовательность Hu2B6VH, SEQ ID NO: 85, и может кодироваться нуклеотидной последовательность SEQ ID NO: 86. В другом конкретном варианте осуществления диантитело дополнительно содержит VL-область, которая состоит из FR-сегментов, взятых из VL-сегмента VK-A26 эмбрионального типа человека (Lautner-Rieske et al. (1992) "The Human Immunoglobulin Kappa Locus. Characterization Of The Duplicated A Regions", Eur. J. Immunol. 22:1023-1029) и JK4 (Hieter et al. (1982) "Evolution Of Human Immunoglobulin Kappa J. Region Genes", J. Biol. Chem. 257:1516-22), и одну или несколько областей CDR VL 2B6, имеющего аминокислотную последовательность SEQ ID NO: 30, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 и SEQ ID NO: 36. В одном из вариантов осуществления VL 2B6 имеет аминокислотную последовательность SEQ ID NO: 39, SEQ ID NO: 40 или SEQ ID NO: 41. В конкретном варианте осуществления VL 2B6 имеет аминокислотную последовательность VL Hu2B6, SEQ ID NO: 87, и может кодироваться нуклеотидной последовательностью, представленной в SEQ ID NO: 88.
В другом конкретном варианте осуществления диантитело имеет вариабельные домены из гуманизированного антитела 3H7, где VH-область состоит из FR-сегментов, взятых из VH-сегмента человека эмбрионального типа, и CDR-областей VH 3H7, имеющей аминокислотную последовательность SEQ ID NO: 35. В другом конкретном варианте осуществления гуманизированное антитело 3H7, кроме того, содержит VL-области, которые состоят из FR-сегментов, взятых из VL-сегмента эмбрионального типа человека и CDR-областей VL 3H7, имеющей аминокислотную последовательность SEQ ID NO: 42.
В частности, связывающие домены иммуноспецифично связываются с внеклеточными доменами нативного FcγRIIB человека и содержат (или альтернативно состоят из) последовательности CDR 2B6, 3H7, 1D5, 2E1, 2H9, 2D11 или 1F2, любую из следующих комбинаций: CDR1 VH и CDR1 VL; CDR1 VH и CDR2 VL; CDR1 VH и CDR3 VL; CDR2 VH и CDR1 VL; CDR2 VH и CDR2 VL; CDR2 VH и CDR3 VL; CDR3 VH и CDR1 VH; CDR3 VH и CDR2 VL; CDR3 VH и CDR3 VL; CDR1 VH1, CDR2 VH и CDR1 VL; CDR1 VH, CDR2 VH и CDR2 VL; CDR1 VH, CDR2 VH и CDR3 VL; CDR2 VH, CDR3 VH и CDR1 VL, CDR2 VH, CDR3 VH и CDR2 VL; CDR2 VH, CDR2 VH и CDR3 VL; CDR1 VH, CDR1 VL и CDR2 VL; CDR1 VH, CDR1 VL и CDR3 VL; CDR2 VH, CDR1 VL и CDR2 VL; CDR2 VH, CDR1 VL и CDR3 VL; CDR3 VH, CDR1 VL и CDR2 VL; CDR3 VH, CDR1 VL и CDR3 VL; CDR1 VH, CDR2 VH, CDR3 VH и CDR1 VL; CDR1 VH, CDR2 VH, CDR3 VH и CDR2 VL; CDR1 VH, CDR2 VH, CDR3 VH и CDR3 VL; CDR1 VH, CDR2 VH, CDR1 VL и CDR2 VL; CDR1 VH, CDR2 VH, CDR1 VL и CDR3 VL; CDR1 VH, CDR3 VH, CDR1 VL и CDR2 VL; CDR1 VH, CDR3 VH, CDR1 VL и CDR3 VL; CDR2 VH, CDR3 VH, CDR1 VL и CDR2 VL; CDR2 VH, CDR3 VH, CDR1 VL и CDR3 VL; CDR2 VH, CDR3 VH, CDR2 VL и CDR3 VL; CDR1 VH, CDR2 VH, CDR3 VH, CDR1 VL и CDR2 VL; CDR1 VH, CDR2 VH, CDR3 VH, CDR1 VL и CDR3 VL; CDR1 VH, CDR2 VH, CDR1 VL, CDR2 VL и CDR3 VL; CDR1 VH, CDR3 VH, CDR1 VL, CDR2 VL и CDR3 VL; CDR2 VH, CDR3 VH, CDR1 VL, CDR2 VL и CDR3 VL; или любую комбинацию из CDR VH и CDR VL, описанных в настоящем документе.
Антитела, в качестве источников связывающих доменов для включения в диантитела по изобретению, можно далее охарактеризовывать картированием эпитопов, так чтобы можно было выбирать антитела, которые имеют наиболее высокую специфичность к FcγRIIB по сравнению с FcγRIIA. Способы картирования эпитопов антител хорошо известны в данной области и охватываются способами по изобретению. В определенных вариантах осуществления для картирования эпитопа антитела по изобретению можно использовать слитые белки, содержащие один или несколько участков FcγRIIB. В конкретном варианте осуществления слитый белок содержит аминокислотную последовательность участка FcγRIIB, слитую с Fc-областью IgG2 человека. Кроме того, каждый слитый белок может содержать аминокислотные замены и/или изменения определенных областей рецептора соответствующей областью из гомологичного рецептора, например, FcγRIIA, как показано в таблице 2 ниже. pMGX125 и pMGX132 содержат связывающийся с IgG участок рецептора FcγRIIB, причем первый из них имеет на C-конце FcγRIIB, а второй имеет на C-конце FcγRIIA, и их можно использовать для различения C-концевого связывания. Другие белки имеют замены в связывающемся с IgG участке рецептора FcγRIIA и FcγIIA или FcγIIB на N-конце. Эти молекулы могут помочь определить часть молекулы рецептора, с которой связываются антитела.
Таблица 2. Перечень слитых белков, которые можно использовать для исследования эпитопов моноклональных антител против FcγRIIB. Остатки с 172 по 180 относятся к связывающему IgG участку FcγRIIA и B. Специфичные аминокислоты из последовательности FcγRIIA указаны полужирным шрифтом.
Слитые белки можно использовать в любом биохимическом анализе для определения связывания с антителом против FcγRIIB по изобретению, например, ELISA. В других вариантах осуществления можно проводить дальнейшее подтверждение специфичности к эпитопу с использованием пептидов, в котором определенные остатки заменены остатками из последовательности FcγRIIA.
Антитела можно охарактеризовывать с использованием анализов дли идентификации функции антител по изобретению, в частности, активности модулирования передачи сигнала FcγRIIB. Например, в анализах для охарактеризации изобретения можно измерять фосфорилирование остатков тирозина в ITIM-мотиве FcγRIIB или измерять ингибирование сгенерированной B-клеточным рецептором мобилизации кальция. Анализы для охарактеризации изобретения могут представлять собой анализы на клеточной основе или бесклеточные анализы.
В данной области установлено, что в тучных клетках коагрегация FcγRIIB с высокоаффинным рецептором IgE, FcεRI, приводит к ингибированию индуцируемой антигеном дегрануляции, мобилизации кальция и продукции цитокинов (Metcalfe D.D. et al. (1997) "Mast Calls", Physiol. Rev. 77:1033-1079; Long E.O. (1999) "Regulation Of Immune Responses Through Inhibitory Receptors", Annu. Rev. Immunol. 17:875-904). Молекулярные детали этого каскада передачи сигнала недавно были установлены (Ott V. L. (2002) "Downstream Of Kinase, p62(dok), Is A Mediator Of FcgammaIIB Inhibition Of Fc Epsilon RI Signaling", J. Immunol. 62(9):4430-4439). После коагрегации с FcεRI, FcγRIIB быстро фосфорилируется по тирозину в его ITIM-мотиве, а затем привлекает Src Homology-2, содержащую инозитол-5-фосфатазу (SHIP), и содержащую SH2-домен инозитолполифосфат-5-фосфатазу, которая, в свою очередь, фосфорилируется и связывается с Shc и p62dok (p62dok представляет собой прототип семейства адаптерных молекул, которые включают домены передачи сигнала, такие как N-концевой домен гомологии плекстрина (PH-домен), PTB-домен, и С-концевую область, содержащую мотивы PXXP и многочисленные участки фосфорилирования (Carpino et al. (1997) "p62(dok): A Constitutively Tyrosine-Phosphorilated, GAP-Associated Protein In Chronic Myelogenous Leukemia Progenitor Cells", Cell, 88:197-204; Yamanshi et al. (1997) "Identification Of The Abl- And rasGAP-Associated 62 kDa Protein As A Docking Protein, Dok", Cell, 88:205-211).
Антитела против FcγRIIB для применения в изобретении аналогично могут быть охарактеризованы по способности модулировать один или несколько опосредуемых IgE ответов. Предпочтительно клеточные линии, коэкспрессирующие высокоаффинный рецептор для IgE и низкоаффинный рецептор для FcγRIIB, можно использовать для охарактеризации антител против FcγRIIB в отношении модулирования опосредуемых IgE ответов. В конкретном варианте осуществления используют клетки из линии лейкоза базофильных клеток крысы (RBL-H23; Barsumian E.L. et al. (1981) "IgE-Induced Histamine Release From Rat Basophilic Leukemia Cell Lines: Isolation Of Releasing And Nonreleasing Clones", Eur. J. Immunol. 11:317-323, которая включена в настоящий документ в качестве ссылки в полном объеме), трансфицированные полноразмерным FcγRIIB человека. RBL-2H3 представляет собой хорошо охарактеризованную клеточную линию, которую широко использовали для исследования механизмов передачи сигнала после опосредуемой IgE активации клеток. При экспрессии в клетках RBL-2H3 и коагрегации с FcεRI, FcγRIIB ингибирует индуцируемую FcεRI мобилизацию кальция, дегрануляцию и продукцию цитокинов (Malbec et al. (1998) "Fc Epsilon Receptor I-Associated Lyn-Dependent Phosphorylation Of Fc Gamma Receptor IIB During Negative Regulation Of Mast Cell Activation", J. Immunol. 160:1647-1658; Daeron et al. (1995) "Regulation Of High-Affinity IgE Receptor-Mediated Mast Cell Activation By Murine Low-Affinity IgG Receptors", J. Clin. Invest. 95:577; Ott V.L. (2002) "Downstream Of Kinase, p62(dok), Is A Mediator Of FcgammaIIB Inhibition Of Fc Epsilon RI Signaling", J. Immunol. 162(9):4430-4439).
Антитела для применения в изобретении также могут быть охарактеризованы по ингибированию индуцируемой FcεRI активации тучных клеток. Например, клетки из линии лейкоза базофильных клеток крысы (RBL-H23; Barsumian E.L. et al. (1981) "IgE-Induced Histamine Release From Rat Basophilic Leukemia Cell Lines: Isolation Of Releasing And Nonreleasing Clones", Eur. J. Immunol. 11:317-323), трансфицированных FcγRIIB, сенсибилизируют IgE и стимулируют либо F(ab')2-фрагментами антитела кролика против IgG мыши для агрегации FcεRI отдельно, или полным антителом кролика против IgG мыши для коагрегации FcγRIIB и FcεRI. В этой системе непрямое модулирование нижерасположенных молекул передачи сигнала можно анализировать путем добавления антител по изобретению к сенсибилизированным и стимулированным клеткам. Например, можно анализировать фосфорилирование тирозина FcγRIIB и привлечение и фосфорилирование SHIP, активацию представителей семейства MAP-киназ, включая, но не ограничиваясь ими, Erk1, Erk2, JNK или p38; и фосфорилирование тирозина p62dok и его связывание с SHIP и RasGAP.
Один из иллюстративных анализов для определения ингибирования индуцируемой FcεRI активации тучных клеток антителами по изобретению может включать следующее: трансфекция клеток RBL-H23 FcγRIIB человека; сенсибилизация клеток RBL-H23 посредством IgE; либо стимуляция клеток RBL-H23 F(ab')2 антитела кролика против IgG мыши (для агрегации FcεRI отдельно и возникновения опосредуемой FcεRI передачи сигнала, в качестве контроля), либо стимуляция клеток RBL-H23 целым антителом кролика против IgG мыши (для коагрегации FcγRIIB и FcεRI, что приводит к ингибированию опосредуемой FcεRI передачи сигнала). Клетки, стимулированные целыми антителами кролика против IgG мыши, можно далее преинкубировать с антителами по изобретению. Измерение FcεRI-зависимой активности клеток, преинкубированных с антителами по изобретению, и клеток, не преинкубированных с антителами по изобретению, и сравнение уровней FcεRI-зависимой активности в этих клетках может указать на модулирование FcεRI-зависимой активности антителами по изобретению.
Иллюстративный анализ, описанный выше, можно использовать, например, для идентификации антител, которые блокируют связывание лиганда (IgG) с FcγRIIB-рецептором и осуществляют антагонизм FcγRIIB-опосредуемого ингибирования передачи сигнала FcεRI путем предотвращения коагрегации FcγRIIB и FcεRI. В этом анализе аналогично идентифицируют антитела, которые усиливают коагрегацию FcγRIIB и FcεRI и осуществляют антагонизм опосредуемого FcγRIIB ингибирования передачи сигнала FcεRI путем обеспечения коагрегации FcγRIIB и FcεRI.
В некоторых вариантах осуществления диантитела против FcγRIIB по изобретению, содержащие связывающие эпитоп домены антител против FcγRIIB, описанных в настоящем документе или известных в данной области, характеризуются их способностью модулировать опосредуемый IgE ответ путем мониторинга и/или измерения дегрануляции тучных клеток или базофилов, предпочтительно в анализе на клеточной основе. Предпочтительно тучные клетки или базофилы для применения в таких анализах конструируют, чтобы они содержали FcγRIIB человека, с использованием стандартных рекомбинантных способов, известных специалисту в данной области. В конкретном варианте осуществления антитела против FcγRIIB по изобретению характеризуются их способностью модулировать IgE-опосредуемый ответ в клеточном анализе высвобождения β-гексозаминидазы (фермента, содержащегося в гранулах); высвобождение β-гексозаминидазы из тучных клеток и базофилов является основным событием при остром аллергическом и воспалительном состоянии (Aketani et al. (2001) "Correlation Between Cytosolic Calcium Concentration And Degranulation In RBL-2H3 Cells In Presence Of Various Concentrations Of Antigen-Specific IgEs", Immunol. Lett. 75:185-189; Aketani et al. (2000) "A Screening Method For Antigen-Specific IgE Using Mast Cells Based On Intracellular Calcium Signaling", Anal. Chem. 72:2653-2658). Высвобождение других медиаторов воспаления, включая, но не ограничиваясь ими, серотонин и гистамин, можно анализировать для измерения IgE-опосредуемого ответа согласно способам по изобретению. Без связи с каким-либо конкретным механизмом действия, высвобождение гранул, таких как гранулы, содержащие β-гексозаминидазу, из тучных клеток и базофилов является процессом, зависимым от внутриклеточной концентрации кальция, который инициируется связыванием FcγRI с поливалентным антигеном.
Возможность исследования тучных клеток человека была ограничена отсутствием пригодных длительных культур тучных клеток. Недавно были получены две новых зависимых от фактора стволовых клеток линии тучных клеток человека, обозначенных LAD1 и LAD2, из аспиратов костного мозга пациента с саркомой тучных клеток/лейкозом (Kirshenbaum et al. (2003) "Characterization Of Novel Stem Cell Factor Responsive Human Mast Cell Lines LAD 1 And 2 Established From A Patient With Mast Cell Sarcoma/Leukemia; Activation Following Aggregation Of FcRI Or FcγRI", Leukemia research, 27:677-82, которая включена в настоящий документ в качестве ссылки в полном объеме). Было описано, что обе клеточных линий экспрессируют FcεRI и несколько маркеров тучных клеток человека. Клетки LAD1 и 2 можно использовать для оценки эффекта антител по изобретению на опосредуемые IgE ответы. В конкретном варианте осуществления для определения какого-либо модулирования IgE-опосредуемого ответа антителами против FcγRIIB по изобретению в клетках LAD можно использовать клеточные анализы высвобождения β-гексозаминидазы, такие как анализы, описанные выше. В иллюстративном анализе тучные клетки человека, например LAD1, стимулируют химерным антителом человека против IgE-нитрофенола (NP) и нагружают BSA-NP, поливалентным антигеном, и проводят мониторинг дегрануляции клеток путем измерения высвобожденной β-гексозаминидазы в супернатанте (Kirshenbaum et al. (2003) "Characterization Of Novel Stem Cell Factor Responsive Human Mast Cell Lines LAD 1 And 2 Established From A Patient With Mast Cell Sarcoma/Leukemia; Activation Following Aggregation Of FcRI Or FcyRI", Leukemia research, 27:677-82, которая включена в настоящий документ в качестве ссылки в полном объеме).
В некоторых вариантах осуществления, если тучные клетки человека имеют низкую экспрессию эндогенного FcγRIIB, при определении с использованием стандартных способов, известных в данной области, например, окрашивания FACS, может быть трудным осуществлять мониторинг и/или детекцию различий в активации ингибиторного каскада, опосредуемого диантителами против FcγRIIB по изобретению. Таким образом, изобретение охватывает альтернативные способы, в которых экспрессия FcγRIIB может повышаться при использовании цитокинов и конкретных условий роста. Описано, что FcγRIIB в высокой степени активируется в моноцитарных клеточных линиях человека, например, THP1 и U937, (Tridandapani et al. (2002) "Regulated Expression And Inhibitory Function Of Fcgamma RIIB In Human Monocytic Cells", J. Biol. Chem., 277(7):5082-5089) и в первичных моноцитах человека (Pricop et al. (2001) "Differential Modulation Of Stimulatory And Inhibitory Fc Gamma Receptors On Human Monocytes By ThI And Th2 Cytokines", J. of Immunol, 166:531-537) посредством IL4. Описано, что дифференцировка клеток U937 с помощью дибутирилциклического AMP увеличивает экспрессию FcγRII (Cameron et al. (2002) "Differentiation Of The Human Monocyte Cell Line, U937, With Dibutyryl Cyclic AMP Induces The Expression Of The Inhibitory Fc Receptor, FcgammaRIIB", Immunology Letters 83, 171-179). Таким образом, для применения в способах по изобретению для повышения чувствительности детекции эндогенную экспрессию FcγRIIB в тучных клетках человека можно повышать с использованием цитокинов, например, IL-4, IL-13.
Диантитела против FcγRIIB также можно анализировать в отношении ингибирования опосредуемой B-клеточным рецептором (BCR) передачи сигнала. Опосредуемая BCR передача сигнала может включать по меньшей мере один или несколько последующих биологических ответов, таких как активация и пролиферация B-клеток, продукция антител и т.д. Коагрегация FcγRIIB и BCR приводит к ингибированию прогрессирования клеточного цикла и выживания клеток. Кроме того, коагрегация FcγRIIB и BCR приводит к ингибированию опосредуемой BCR передачи сигнала.
В частности, BCR-опосредуемая передача сигнала включает по меньшей мере одно или несколько из следующих: модулирование нижерасположенных молекул передачи сигнала, например, фосфорилированное состояние FcγRIIB, привлечение SHIP, локализация Btk и/или PLCγ, активность MAP-киназы, привлечение Akt (антиапоптотический сигнал), мобилизация кальция, прогрессирование клеточного цикла и пролиферация клеток.
Хотя многочисленные эффекторные функции опосредуемого FcγRIIB ингибирования передачи сигнала BCR опосредуются SHIP, недавно было продемонстрировано, что активированные липополисахаридом (LPS) B-клетки из дефицитных по SHIP мышей проявляют значительное опосредуемое FcγRIIB ингибирование мобилизации кальция, продукции Ins(1,4,5)P3, и фосфорилирования Erk и Akt (Brauweiler et al. (2001) "Partially Distinct Molecular Mechanisms Mediate Inhibitory FcgammaRIIB Signaling In Resting And Activated B Cells", Journal of Immunology, 167(1): 204-211). Таким образом, для охарактеризации антител по изобретению можно использовать B-клетки ex vivo из дефицитных по SHIP мышей. Один из иллюстративных анализов для определения опосредуемого FcγRIIB ингибирования передачи сигнала BCR антителами по изобретению может включать следующее: выделение B-клеток селезенки из дефицитных по SHIP мышей, активация указанных клеток липополисахаридом и стимуляция указанных клеток либо F(ab')2 против IgM для агрегации BCR, либо антителом против IgM для коагрегации BCR с FcγRIIB. Клетки, стимулированные целым антителом против IgM для коагрегации BCR с FcγRIIB, можно далее преинкубировать с антителами по изобретению. FcγRIIB-зависимую активность клеток можно измерять стандартными способами, известными в данной области. Сравнение уровня FcγRIIB-зависимой активности в клетках, преинкубированных с антителами, и в клетках, которые не преинкубировали, указывает на модулирование FcγRIIB-зависимой активности антител.
Измерение FcγRIIB-зависимой активности может включать, например, измерение внутриклеточной мобилизации кальция проточной цитометрией, измерение фосфорилирования Akt и/или Erk, измерение опосредуемого BCR накопления PI(3,4,5)P3 или количественное определение опосредуемой FcγRIIB пролиферации B-клеток.
Анализы можно использовать, например, для идентификации диантител или антител против FcγRIIB для применения в изобретении, которые модулируют опосредуемое FcγRIIB ингибирование передачи сигнала BCR путем блокирования участка связывания лиганда (IgG) с FcγRIIB-рецептором и антагонизма опосредуемого FcγRIIB ингибирования передачи сигнала BCR путем предотвращения коагрегации FcγRIIB и BCR. Также анализы можно использовать для идентификации антител, которые усиливают коагрегацию FcγRIIB и BCR и осуществляют антагонизм опосредуемого FcγRIIB ингибирования передачи сигнала BCR.
Антитела против FcγRIIB также можно анализировать в отношении опосредуемой FcγRII передачи сигнала в моноцитах/макрофагах человека. Коагрегация FcγRIIB с рецептором, несущим тирозинсодержащий активационный мотив иммунорецепторов (ITAM), действует, подавляя опосредуемый FcγR фагоцитоз с использованием SHIP в качестве эффектора (Tridandapani et al. (2002) "Regulated Expression And Inhibitory Function Of Fcgamma RIIB In Human Monocytic Cells", J. Biol. Chem., 277(7):5082-5089). Коагрегация FcγRIIA с FcγRIIB приводит к быстрому фосфорилированию остатка тирозина на мотиве ITIM в FcγRIIB, вызывая усиление фосфорилирования SHIP, связывания SHIP Shc и фосфорилирования белков, имеющих молекулярную массу 120 и 60-65 кДа. Кроме того, коагрегация FcγRIIA с FcγRIIB приводит к подавлению фосфорилирования Akt, которая представляет собой серин-треониновую киназу, вовлеченную в клеточную регуляцию, и служит для подавления апоптоза.
Диантитела против FcγRIIB также можно анализировать в отношении ингибирования опосредуемого FcγR фагоцитоза в моноцитах/макрофагах человека. Например, клетки из моноцитарной клеточной линии человека THP-1 можно стимулировать либо Fab-фрагментами моноклонального антитела мыши IV.3 против FcγRII и антителом козы против антитела мыши (для агрегации FcγRIIA отдельно), либо целым моноклональным антителом мыши IV.3 и антителом козы против антител мыши (для коагрегации FcγRIIA и FcγRIIB). В этой системе модулирование нижерасположенных молекул передачи сигнала, такое как фосфорилирование тирозина в FcγRIIB, фосфорилирование SHIP, связывание SHIP с Shc, фосфорилирование Akt и фосфорилирование белков, имеющих молекулярную массу 120 и 60-65 кДа, можно анализировать путем добавления молекул по изобретению к стимулированным клеткам. Кроме того, FcγRIIB-зависимую эффективность фагоцитоза моноцитарной клеточной линии можно прямо измерять в присутствии и отсутствие антител по изобретению.
Другой иллюстративный анализ для определения ингибирования опосредуемого FcγR фагоцитоза в моноцитах/макрофагах человека антителами по изобретению может включать следующее: стимуляция клеток THP-1 либо Fab антитела мыши IV.3 против FcγRII и антителом козы против антител мыши (для агрегации FcγRIIA отдельно и возникновения опосредуемой FcγRIIA передачи сигнала); либо антителом мыши против FcγRII и антителом козы против антител мыши (для коагрегации FcγRIIA и FcγRIIB и ингибирования опосредуемой FcγRIIA передачи сигнала). Клетки, стимулированные антителом мыши против FcγRII и антителом козы против антител мыши, можно далее преинкубировать с молекулами по изобретению. Измерение FcγRIIA-зависимой активности стимулированных клеток, преинкубированных с молекулами по изобретению, и клеток, которые не преинкубировали с антителами по изобретению, и сравнение уровней FcγRIIA-зависимой активности в этих клетках указывает на модулирование FcγRIIA-зависимой активности антителами по изобретению.
Иллюстративный описанный анализ можно использовать, например, для идентификации связывающих доменов, которые блокируют связывание лиганда рецептора FcγRIIB и осуществляют антагонизм опосредуемого FcγRIIB ингибирования передачи сигнала FcγRIIA путем препятствования коагрегации FcγRIIB и FcγRIIA. Аналогично, этот анализ идентифицирует связывающие домены, которые усиливают коагрегацию FcγRIIB и FcγRIIA и осуществляют антагонизм опосредуемому FcγRIIB ингибированию передачи сигнала FcγRIIA.
Представляющие интерес связывающие домены FcγRIIB, когда они содержатся в антителах, можно анализировать путем определения способности клеток THP-1 фагоцитировать эритроциты овцы (SRBC), опсонизированные связанным с флуоресцеином IgG, описанными выше способами (Tridandapani et al. (2000) "The Adapter Protein LAT Enhances Fcgamma Receptor-Mediated Signal Transduction In Myeloid Cells", J. Biol. Chem. 275:20480-7). Например, иллюстративный анализ для определения фагоцитоза включает: обработку клеток THP-1 антителами по изобретению или контрольным антителом, которое не связывается с FcγRII, сравнение уровней активности указанных клеток, где различия в активности клеток (например, активности розеткообразования (количество клеток THP-1, связывающихся с покрытыми IgG SRBC), активности прикрепления (общее количество SRBC, связанных с клетками THP-1), и уровня фагоцитоза указывает на модулирование FcγRIIA-зависимой активности антителами по изобретению. Этот анализ можно использовать для идентификации, например, антител, которые блокируют связывание лиганда рецептора FcγRIIB и осуществляют антагонизм опосредуемого FcγRIIB ингибирования фагоцитоза. Также этот анализ может идентифицировать антитела, которые усиливают опосредуемое FcγRIIB ингибирование передачи сигнала FcγRIIA.
В предпочтительном варианте осуществления связывающие домены модулируют FcγRIIB-зависимую активность в моноцитах/макрофагах человека по меньшей мере одним или несколькими из следующих способов: модулирование нижерасположенных молекул передачи сигнала (например, модулирование состояния фосфорилирования FcγRIIB, модулирование фосфорилирования SHIP, модулирование связывания SHIP и Shc, модулирование фосфорилирования Akt, модулирование фосфорилирования дополнительных белков массой приблизительно 120 и 60-65 кДа) и модулирование фагоцитоза.
5.1.2 СВЯЗЫВАЮЩИЕ CD16A ДОМЕНЫ
В данном разделе обсуждаются связывающие CD16A белки, которые можно использовать в качестве источников вариабельных областей легкой и тяжелой цепей для продукции ковалентного диантитела. В настоящем изобретении связывающие CD16A белки включают молекулы, содержащие VL- и VH-домены антител против CD16A, которые использованы для продукции диантител по настоящему изобретению.
Применительно к настоящему изобретению можно использовать множество связывающих CD16A белков. Пригодные связывающие CD16A белки включают моноклональные антитела человека или гуманизированные моноклональные антитела, а также связывающие CD16A фрагменты антител (например, scFv или одноцепочечные антитела, Fab-фрагменты, миниантитела) и другие антителоподобные белки, которые связываются с CD16A путем взаимодействия с доменом вариабельной области легкой цепи, доменом вариабельной области тяжелой цепи или с обоими из них.
В некоторых вариантах осуществления связывающий CD16A белок для применения согласно изобретению содержит VL- и/или VH-домен, который имеет одну или несколько CDR с последовательностями, происходящими из антитела против CD16A, не являющегося человеческим, такого как антитело против CD16A, и одну или несколько каркасных областей с каркасными последовательностями, происходящими из одного или нескольких иммуноглобулинов человека. Известно множество не являющихся человеческими моноклональных антител против CD16A, из которых можно получать CDR и другие последовательности (см., например, Tamm et al. (1996) "The Binding Epitops Of Human CD 16 (Fc gamma RIII) Monoclonal Antibodies. Implications For Ligand Binding", J. Imm. 157:1576-81; Fleit et al. (1989) p.159; LEUKOCYTE TYPING II: HUMAN MYELOID AND HEMATOPOIETIC CELLS, Reinherz et al, eds. New York: Springer-Verlag; (1986); LEUCOCYTE TYPING III: WHITE CELL DIFFERENTIATION ANTIGENS, McMichael A.J., ed., Oxford: Oxford University Press, 1986); LEUKOCYTE TYPING IV: WHITE CELL DIFFERENTIATION ANTIGENS, Kapp et al., eds. Oxford Univ. Press, Oxford; LEUKOCYTE TYPING V: WHITE CELL DIFFERENTIATION ANTIGENS, Schlossman et al., eds. Oxford Univ. Press, Oxford; LEUKOCYTE TYPING VI: WHITE CELL DIFFERENTIATION ANTIGENS, Kishimoto, ed. Taylor & Francis. Кроме того, как показано в разделе "Примеры", новые связывающие CD16A белки, которые распознают CD16A человека, экспрессированный на клетках, можно получать с использованием хорошо известных способов получения и селекции моноклональных антител или сходных связывающих белков (например, технология гибридом, фиговый дисплей и т.п.). См., например, O'Connell et al. (2002) "Phage Versus Phagemid Libraries For Generation Of Human Monoclonal Antibodies", J. Mol. Biol. 321:49-56; Hoogenboom et al. (2000) "Natural And Designer Binding Sites Made By Phage Display Technology", Imm. Today 21:371078; Krebs et al. (2001) "High-Throughput Generation And Engineering Of Recombinant Human Antibodies", J. Imm. Methods 254:67-84; и другие ссылки, цитированные в настоящем документе. Моноклональные антитела из не являющихся человеком видов можно подвергать химеризации или гуманизации с использованием способов гуманизации антител, известных в данной области.
Альтернативно можно получать полностью человеческие антитела против CD16A с использованием трансгенных животных, имеющих элементы иммунной системы человека (см., например, патенты США №№ 5569825 и 5545806), с использованием клеток периферической крови человека (Casali et al. (1986) "Human Monoclonals From Antigen-Specific Selection Of B Lymhocytes And Transformation By EBV", Science 234:476-479), путем скрининга библиотеки ДНК из B-клеток человека согласно общему протоколу, описанному Huse et al. (1989) "Generation Of A Large Combinatorial Library Of The Immunoglobulin Repertoire In Phage Lambda", Science 246:1275-1281, и другими способами.
В предпочтительном варианте осуществления связывающий донор происходит из антитела 3G8 или его гуманизированной версии, например, такой как версии, описанные в публикации патентной заявки США 2004/0010124, которая включена в настоящий документ в качестве ссылки в полном объеме. Предусматривается, что для некоторых целей может быть преимущественным использование связывающих CD16A белков, которые связывают тот же эпитоп рецептора CD16A, который связывает 3G8, или эпитоп, расположенный по меньшей мере достаточно близко к этому эпитопу, для блокирования связывания 3G8. Способы картирования эпитопов и эксперименты по конкурентному связыванию для идентификации связывающих белков с желаемыми связывающими свойствами связывания хорошо известны специалистам в области экспериментальной иммунологии. См., например, Harlow and Lane, цитированную выше; Stähli et al. (1983) "Distinction Of Epitops By Monoclonal Antibodies", Methods in Enzymology 92:242-253; Kirkland et al. (1986) "Analysis Of The Fine Specificity And Cross-Reactivity Of Monoclonal Anti-Lipid A Antibodies", J. Immunol. 137:3614-3619; Morel et al. (1988) "Monoclonal Antibodies To Bovine Surum Albumin: Affinity And Specificity Determinations", Molec. Immunol. 25:7-15; Cheung et al. (1990) "Epitop-Specific Antibody Response To The Surface Antigen Of Duck Hepatitis B Virus In Infected Ducks", Virology 176:546-552; и Moldenhauer et al. (1990) "Identity Of HML-1 Antigen On Intestinal Intraepithelial T Cells And Of B-ly7 Antigen On Hairy Cell Leukemia", Scand. J. Immunol. 32:77-82. Например, то, связывают ли два антитела один участок, можно определить с использованием одного из антител для иммобилизации антигена на планшете для ELISA, а затем определения способности второго антитела связываться с уловленным антигеном. Сравнение эпитопов также можно проводить путем мечения первого антитела, прямо или непрямо, ферментом, радионуклидом или флуорофором, и измерения способности немеченого второго антитела ингибировать связывание первого антитела с антигеном на клетках, в растворе или на твердой фазе.
Также можно определять способность антител блокировать связывание рецептора CD16A с иммунными комплексами, образованными на планшетах для ELISA. Такие иммунные комплексы образуются путем первоначального покрытия планшета антигеном, таким как флуоресцеин, а затем нанесения на планшет специфического антитела против флуоресцеина. Затем этот иммунный комплекс служит в качестве лиганда для растворимых Fc-рецепторов, таких как sFcRIIIa. Альтернативно можно получать растворимый иммунный комплекс и метить его, прямо или непрямо, ферментом, радионуклидом или флуорофором. Затем можно измерять способность антител ингибировать связывание этих меченых иммунных комплексов с Fc-рецепторами на клетках, в растворе или на твердой фазе.
Связывающие CD16A белки по изобретению могут содержать Fc-область иммуноглобулина человека, или могут не содержать ее. Fc-области не присутствуют, например, в связывающих белках scFv. Fc-области присутствуют, например, в тетрамерных моноклональных IgG-антителах человека или гуманизированных тетрамерных моноклональных IgG-антителах. Как описано выше, в некоторых вариантах осуществления настоящего изобретения связывающий CD16A белок включает Fc-область, которая имеет измененную эффекторную функцию, например, сниженную аффинность к эффекторному лиганду, такому как Fc-рецептор или C1-компонент комплемента, по сравнению с неизмененной Fc-областью (например, Fc встречающихся в природе белков IgG1). В одном из вариантов осуществления Fc-область не является гликозилированной по аминокислоте в Fc-области, соответствующей положению 297. Такие антитела лишены эффекторной функции Fc.
Таким образом, связывающий CD16A белок может не проявлять опосредуемое Fc связывание с эффекторным лигандом, таким как Fc-рецептор или C1-компонент комплемента, вследствие отсутствия Fc-домена в связывающем белке, в то время как в других случаях отсутствие связывающей или эффекторной функции является следствием изменения константной области антитела.
5.1.2.1 Связывающие CD16A белки, содержащие последовательности CDR, сходные с последовательностями CDR mAb 3G8.
Связывающие CD16A белки, которые можно использовать для практики изобретения, включают белки, содержащие последовательность CDR, происходящую из (т.е. имеющую одинаковую или сходную последовательность c) CD моноклонального антитела мыши 3G8. Комплементарные кДНК, кодирующие вариабельные области тяжелой и легкой цепей моноклонального антитела мыши 3G8, включая кодирующие CDR последовательности, клонировали и секвенировали, как описано. Последовательности нуклеиновых кислот и белков 3G8 представлены ниже. С использованием последовательностей вариабельной области и CDR мыши было получено большое количество химерных и гуманизированных моноклональных антител, содержащих определяющие комплементарность области, происходящие из CDR 3G8, и их свойства были проанализированы. Для идентификации гуманизированных антител, которые связывают CD16A с высокой аффинностью и имеют другие желательные свойства, получили тяжелые цепи антител, содержащие VH-область с CDR, происходящими из 3G8, и комбинировали их (путем коэкспрессии) с легкими цепями антител, содержащими VL-область с CDR, происходящими из 3G8, с получением тетрамерного антитела для анализа. Свойства полученных тетрамерных антител определяли, как описано ниже. Как описано ниже, согласно изобретению можно использовать связывающие CD16A белки, содержащие CDR 3G8, такие как белки гуманизированных антител, описанные в настоящем документе.
5.1.2.1.1 VH-область
В одном аспекте связывающий CD16A белок по изобретению может содержать вариабельный домен тяжелой цепи, в котором по меньшей мере одна CDR (и обычно три CDR) имеет последовательность CDR (и более конкретно всех трех CDR) тяжелой цепи моноклонального антитела мыши 3G8 и в котором остальные части являются по существу человеческими (происходящими из вариабельной области тяжелой цепи антитела или антител человека и по существу сходными с ней).
В одном аспекте изобретение относится к гуманизированному антителу 3G8 или фрагменту антитела, содержащим CDR, происходящие из антитела 3G8, по существу в человеческом каркасе, но в которых по меньшей мере одна из CDR вариабельного домена тяжелой цепи отличается по последовательности от соответствующей CDR тяжелой цепи антитела мыши 3G8. Например, в одном из вариантов осуществления CDR отличается от последовательности CDR 3G8 по меньшей мере наличием одной или нескольких замен CDR, для которых показано, что они влияют на связывание 3G8 с CD16A, как известно в данной области или как представлено в таблицах 3 и 4A-H. Пригодные связывающие CD16 белки могут содержать 0, 1, 2, 3 или 4, или более из этих замен (и часто они имеют от 1 до 4 из этих замен) и необязательно они также могут иметь дополнительные замены.
Замены в домене VH
Tyr или
Asp
Lys или
Ala или His
Последовательности VH, происходящие из VH 3G8*
FR1
CDR1
FR2
CDR2
FR3
CDR3
FR4
В одном из вариантов осуществления связывающий CD16A белок может содержать последовательность вариабельного домена тяжелой цепи, которая является одинаковой или сходной с VH-доменом конструкции Hu3G8VH-1, последовательность которой представлена в SEQ ID NO: 68. Например, изобретение относится к связывающему CD16A белку, содержащему VH-домен с последовательностью, которая (1) отличается от VH-домена Hu3G8VH-1 (SEQ ID NO: 68) нулем, одной или несколькими заменами, указанными в таблице 1; (2) отличается от VH-домена Hu3G8VH-1 нулем, одной или несколькими заменами в каркасной области, представленными в таблице 1; и (3) является по меньшей мере приблизительно на 80% идентичной, часто по меньшей мере приблизительно на 90%, и иногда по меньшей мере приблизительно на 95% идентичной или даже по меньшей мере приблизительно на 98% идентичной последовательности VH Hu3G8VH-1 в остальных положениях.
Иллюстративные VH-домены связывающих CD16 белков по изобретению имеют последовательность 3G8VH, Hu3G8VH-5 и Hu3G8VH-22 (SEQ ID NO: 79, SEQ ID NO: 69 и SEQ ID NO: 70, соответственно). Иллюстративные нуклеотидные последовательности, кодирующие последовательности 3G8VH и Hu3G8VH-5 (SEQ ID NO: 79 и SEQ ID NO: 69, соответственно), представлены в SEQ ID NO: 80 и SEQ ID NO: 81, соответственно.
VH-домен может иметь последовательность, которая отличается от Hu3G8VH-1 (SEQ ID NO: 68) по меньшей мере одной, по меньшей мере двумя, по меньшей мере тремя, по меньшей мере четырьмя, по меньшей мере пятью или по меньшей мере шестью заменами, представленными в таблице 3. Полагают, что эти замены приводят к повышенной аффинности к CD16A и/или снижают иммуногенность связывающего CD16A белка при введении человеку. В определенных вариантах осуществления степень идентичности последовательности с VH-доменом Hu3G8VH-1 в остальных положениях составляет по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 98%.
Для иллюстрации и не для ограничения, последовательности ряда VH-доменов связывающего CD16 белка представлены в таблице 4. Тяжелые цепи, содержащие эти последовательности, слитые с константной областью Cγ1 человека, коэкспрессировали с легкой цепью hu3G8VL-1 (описанной ниже) с образованием тетрамерных антител, и связывание антител с CD16A измеряли для оценки эффекта аминокислотных замен по сравнению с VH-доменом hu3G8VH-1. Конструкции, в которых VH-домен имеет последовательность hu3G8VH-1, 2, 3, 4, 5, 8, 12, 14, 16, 17, 18, 19, 20, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 42, 43, 44 и 45, продемонстрировали высокоаффинное связывание, а конструкции с VH-доменами hu3G8VH-6 и 40 продемонстрировали среднее связывание. Связывающие CD16A белки, содержащие VH-домены hu3G8VH-5 и hu3G8VH-22 (SEQ ID NO: 69 и SEQ ID NO: 70, соответственно), считаются имеющими особенно благоприятные свойства связывания.
5.1.2.2 VL-область
Сходные исследования проводили для идентификации последовательностей вариабельных доменов легкой цепи с благоприятными свойствами связывания. В одном аспекте изобретение относится к связывающему CD16A белку, содержащему вариабельный домен легкой цепи, в котором по меньшей мере одна CDR (и обычно три CDR) имеет последовательность CDR (и более конкретно, всех трех CDR) легкой цепи моноклонального антитела мыши 3G8 и в котором остальные части являются по существу человеческими (происходящими из вариабельной области тяжелой цепи антитела или антител человека и по существу сходными с ней).
В одном аспекте изобретение относится к фрагменту гуманизированного антитела 3G8, содержащему CDR, происходящие из антитела 3G8, по существу в человеческом каркасе, но в котором по меньшей мере одна из CDR вариабельного домена легкой цепи отличается по последовательности от соответствующей CDR легкой цепи антитела мыши 3G8. В одном из вариантов осуществления CDR отличается от последовательности CDR 3G8 наличием по меньшей мере одной или нескольких замен CDR, таких как одна или несколько замен, представленных в таблице 2 (например, аргинин в положении 24 в CDR1; серин в положении 25 в CDR1; тирозин в положении 32 в CDR1; лейцин в положении 33 в CDR1; аспарагиновая кислота, триптофан или серин в положении 50 в CDR2; серин в положении 53 в CDR2; аланин или глутамин в положении 55 в CDR2; треонин в положении 56 в CDR2; серин в положении 93 в CDR3; и/или треонин в положении 94 в CDR3). В различных вариантах осуществления вариабельный домен может иметь 0, 1, 2, 3, 4, 5 или более из этих замен (и часто он имеет от 1 до 4 из этих замен), и необязательно он также может иметь дополнительные замены.
В одном из вариантов осуществления пригодный связывающий CD16A белок может содержать последовательность вариабельного домена легкой цепи, которая является одинаковой или сходной с VL-доменом конструкции Hu3G8VL-1 (SEQ ID NO: 71), последовательность которого представлена в таблице 6. Например, изобретение относится к связывающему CD16A белку, содержащему VL-домен с последовательностью, которая (1) отличается от VL-домена Hu3G8VL-1 (SEQ ID NO: 71) нулем, одной или несколькими заменами, указанными в таблице 5; (2) отличается от VL-домена Hu3G8VL-1 нулем, одной или несколькими заменами в каркасной области, представленными в таблице 5; и (3) является по меньшей мере приблизительно на 80% идентичной, часто по меньшей мере приблизительно на 90%, и иногда по меньшей мере приблизительно на 95% идентичной или даже по меньшей мере приблизительно на 98% идентичной последовательности VL Hu3G8VL-1 (SEQ ID NO: 71) в остальных положениях.
Замены в VL-домене 3G8
Последовательности VL, происходящие из VL 3G8*
FR1
CDR1
FR2
CDR2
FR3
CDR3
FR4
Иллюстративные VL-домены связывающих CD16 белков по изобретению имеют последовательность 3G8VL, Hu3G8VL-1 или Hu3G8VL-43, (SEQ ID NO: 82, SEQ ID NO: 71 и SEQ ID NO: 72, соответственно), как показано в таблицах 5 и 6. Иллюстративные нуклеотидные последовательности, кодирующие 3G8VL (SEQ ID NO: 82) и Hu3G8VL-1 (SEQ ID NO: 71), представлены в SEQ ID NO: 83 и SEQ ID NO: 84, соответственно.
VL-домен может иметь последовательность, которая отличается от Hu3G8VL-1 (SEQ ID NO: 71) нулем, одной, по меньшей мере двумя, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8 или по меньшей мере 9 заменами, представленными в таблице 2. Полагают, что эти замены приводят к увеличенной аффинности к CD16A и/или к сниженной иммуногенности связывающего белка CD16A при введении человеку. В определенных вариантах осуществления степень идентичности последовательностей в остальных положениях составляет по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 98%.
Для иллюстрации и не для ограничения, последовательности ряда VL-доменов связывающих CD16A белков представлены в таблице 6. Легкие цепи, содержащие эти последовательности, слитые с константным доменом Cκ человека, коэкспрессировали с тяжелой цепью Hu3G8VH (описанной выше) с получением тетрамерных антител, и измеряли связывание антител с CD16A для оценки эффекта аминокислотных замен по сравнению с VL-доменом Hu3G8VL-1 (SEQ ID NO: 71). Конструкции, в которых VL-домен имеет последовательность hu3G8VL-1, 2, 3, 4, 5, 10, 16, 18, 19, 21, 22, 24, 27, 28, 32, 33, 34, 35, 36, 37 и 42, продемонстрировали высокую аффинность связывания, и hu3G8VL-15, 17, 20, 23, 25, 26, 29, 30, 31, 38, 39, 40 и 41 продемонстрировали среднее связывание. Связывающие CD16A белки, содержащие VL-домены hu3G8VL-1, hu3G8VL-22 и hu3G8VL-43, считаются имеющими особенно благоприятные свойства связывания (SEQ ID NO: 71, SEQ ID NO: 73 и SEQ ID NO: 72, соответственно).
5.1.2.2.1 Комбинации VL- и/или VH-доменов
Как известно в данной области и описано в настоящем документе, легкие и тяжелые цепи иммуноглобулинов могут быть рекомбинантно экспрессированы в условиях, в которых они ассоциируют с образованием диантитела, или их можно комбинировать таким путем in vitro. Таким образом, понятно, что происходящий из 3G8 VL-домен, описанный в настоящем документе, можно комбинировать с происходящим из 3G8 VH-доменом, описанным в настоящем документе, с получением связывающего CD16A диантитела, и предусматриваются все такие комбинации.
Для иллюстрации и не для ограничения, примерами пригодных диантител CD16A являются диантитела, содержащие по меньшей мере один VH-домен и по меньшей мере один VL-домен, где VH-домен происходит из hu3G8VH-1, hu3G8VH-22 или hu3G8VH-5 (SEQ ID NO: 68, SEQ ID NO: 70 и SEQ ID NO: 69, соответственно) и VL-домен происходит из hu3G8VL-1, hu3G8VL-22 или hu3G8VL-43 (SEQ ID NO: 71, SEQ ID NO: 73 и SEQ ID NO: 41, соответственно). В частности, гуманизированные антитела, которые содержат hu3G8VH-22 (SEQ ID NO: 22) и любой из hu3G8VL-1, hu3G8VL-22 или hu3G8VL-43 (SEQ ID NO: 71, SEQ ID NO: 70 и SEQ ID NO: 72, соответственно) или hu3G8VH-5 (SEQ ID NO: 69) и hu3G8VL-1 (SEQ ID NO: 71), имеют благоприятные свойства.
Специалистам в данной области будет понятно, что последовательности VL- и VH-доменов, описанные в настоящем документе, можно далее модифицировать известными в данной области способами, такими как созревание аффинности (см. Schier et al. (1996) "Isolation Of Picomolar Affinity Anti-C-ErbB-2 Single-Chain Fv By Molecular Evolution Of The Complementarity-Determining Regions In The Center Of The Antibody Binding Site", J. Mol. Biol. 263:551-567; Daugherty et al. (1998) "Antibody Affinity Maturation Using Bacterial Surface Display", Protein Eng. 11:825-832; Boder et al. (1997) "Yeast Surface Display For Screening Combinatorial Polypeptide Libraries", Nat. Biotechnol. 15:553-557; Boder et al. (2000) "Directed Evolution Of Antibody Fragments With Monovalent Femtomolar Antigen-Binding Affinity", Proc. Natl. Acad. Sci. U.S.A 97:10701-10705; Hudson et al. (2003) "Engineered Antibodies", Nature Medicine 9:129-39). Например, связывающие CD16A белки можно модифицировать с использованием способов созревания аффинности для идентификации белков с увеличенной аффинностью к CD16A и/или сниженной аффинностью к CD16B.
Одним иллюстративным связывающим CD16 белком является антитело мыши 3G8. Аминокислотная последовательность, содержащая VH- и VL-домены гуманизированного 3G8, описана на ФИГ. 2, 9, 14, и представлена в SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 и SEQ ID NO: 72.
5.2 ДИАНТИТЕЛА, СОДЕРЖАЩИЕ Fc-ОБЛАСТИ ИЛИ ИХ ЧАСТИ
Изобретение охватывает молекулы диантител, содержащие Fc-домены или их части (например, CH2- или CH3-домен). В определенных вариантах осуществления Fc-домен, или его часть(и), содержит один или несколько константный домен(ов) Fc-области IgG2, IgG3 или IgG4 (например, CH2 или CH3). В других вариантах осуществления изобретение охватывает молекулы, содержащие и Fc-домен или его часть, где указанный Fc-домен или его часть содержит по меньшей мере одну модификацию аминокислот (например, замену) относительно сравнимого Fc-домена дикого типа или его части. Варианты Fc-доменов хорошо известны в данной области, и их используют, главным образом, для изменения фенотипа антитела, содержащего указанный вариант Fc-домена при исследовании в любом из анализов активности связывания или эффекторной функции, хорошо известных в данной области, например в ELISA, анализе SPR или ADCC. Такие варианты Fc-доменов или их частей применимы в настоящем изобретении вследствие придания или модификации эффекторной функции, проявляемой молекулой диантитела по изобретению, содержащей Fc-домен (или его часть), в функциональном анализе, например, в анализе с NK-клетками или макрофагами. Варианты Fc-доменов, идентифицированные как изменяющие эффекторную функцию, описаны в международной заявке WO04/063351, публикациях патентных заявок США 2005/0037000 и 2005/0064514, условных заявках США 60/626510, поданной 10 ноября 2004 года, 60/636663, поданной 15 декабря 2004 года, и 60/781564, поданной 10 марта 2006 года, и в патентных заявках США 11/271140, поданной 10 ноября 2005 года, и 11/305787, поданной 15 декабря 2005 года, аналогичных заявках авторов изобретения, все из которых включены в настоящий документ в качестве ссылок в полном объеме.
В других вариантах осуществления изобретение охватывает применение любого варианта Fc, известного в данной области, такого как варианты, описанные в Duncan et al. (1988) "Localization Of The Binding Site For The Human High-Affinity Fc Receptor On IgG", Nature 332:563-564; Lund et al. (1991) "Human Fc Gamma RI And Fc Gamma RII Interact With Distinct But Overlapping Sites On Human IgG", J. Immunol. 147:2657-2662; Lund et al. (1992) "Multiple Binding Sites On The CH2 Domain OfIgG For Mouse Fc Gamma RII", Mol. Immunol. 29:53-59; Alegre et al. (1994) "A Non-Activating "Humanized" Anti-CD3 Monoclonal Antibody Retains Immunosuppressive Properties In Vivo", Transplantation 57:1537-1543; Hutchins et al. (1995) "Improved Biodistribution, Tumor Targeting, And Reduced Immunogenicity In Mice With A Gamma 4 Variant Of Campath-1H", Proc. Natl. Acad. Sci. U S A 92:11980-11984; Jefferis et al. (1995) "Recognition Sites On Human IgG For Fc Gamma Receptors: The Role Of Glycosylation", Immunol. Lett. 44:111-117; Lund et al. (1995) "Oligosaccharide-Protein Interactions In IgG Can Modulate Recognition By Fc Gamma Receptors", FASEB J. 9:115-119; Jefferis et al. (1996) "Modulation Of Fc(Gamma)R And Human Complement Activation By IgGS-Core Oligosaccharide Interactions", Immunol. Lett. 54:101-104; Lund et al. (1996) "Multiple Interactions Of Igg With Its Core Oligosaccharide Can Modulate Recognition By Complement And Human Fc Gamma Receptor I And Influence The Synthesis Of Its Oligosaccharide Chains", J. Immunol. 157:4963-4969; Armour et al. (1999) "Recombinant Human IgG Molecules Lacking Fcgamma Receptor I Binding And Monocyte Triggering Activities", Eur. J. Immunol. 29:2613-2624; Idusogie et al. (2000) "Mapping Of The ClQ Binding Site On Rituxan, A Chimeric Antibody With A Human IgGl Fc", J. Immunol. 164:4178-4184; Reddy et al. (2000) "Elimination OfFc Receptor-Dependent Effector Functions Of A Modified IgG4 Monoclonal Antibody To Human CD4", J. Immunol. 164:1925-1933; Xu et al. (2000) "In Vitro Characterization Of Five Humanized OKT3 Effector Function Variant Antibodies", Cell. Immunol. 200:16-26; Idusogie et al. (2001) "Engineered Antibodies With Increased Activity To Recruit Complement", J. Immunol. 166:2571-2575; Shields et al. (2001) "High Resolution Mapping Of The Binding Site On Human IgGl For Fc gamma RI, Fc gamma RII, Fc gamma RIII, And FcRn And Design Of IgG1 Variants With Improved Binding To The Fc gamma R", J. Biol. Chem. 276:6591-6604; Jefferis et al. (2002) "Interaction Sites On Human IgG-Fc For FcgammaR: Current Models", Immunol. Lett. 82:57-65; Presta et al. (2002) "Engineering Therapeutic Antibodies For Improved Function", Biochem. Soc. Trans. 30:487-490); US 5624821; US 5885573; US 6194551; PCT WO 00/42072; PCT WO 99/58572; все из которых включены в настоящий документ в качестве ссылок в полном объеме.
В определенных вариантах осуществления указанная одна или несколько модификаций аминокислот Fc-области снижают аффинность и авидность Fc-области, и, таким образом, молекулы диантитела по изобретению, в отношении одного или нескольких FcγR-рецепторов. В конкретном варианте осуществления изобретение охватывает диантитела, содержащие вариант Fc-области или ее части, где указанный вариант Fc-области содержит по меньшей мере одну модификацию аминокислот относительно Fc-области дикого типа, и этот вариант Fc-области связывает только один FcγR, где указанный FcγR представляет собой FcγRIIIA. В другом конкретном варианте осуществления изобретение охватывает диантитела, содержащие вариант Fc-области или ее части, где указанный вариант Fc-области содержит по меньшей мере одну модификацию аминокислот относительно Fc-области дикого типа, и этот вариант Fc-области связывает только один FcγR, где указанный FcγR представляет собой FcγRIIA. В другом конкретном варианте осуществления изобретение охватывает диантитела, содержащие вариант Fc-области или ее части, где указанный вариант Fc-области содержит по меньшей мере одну модификацию аминокислот относительно Fc-области дикого типа, и этот вариант Fc-области связывает только один FcγR, где указанный FcγR представляет собой FcγRIIB. В определенных вариантах осуществления изобретение охватывает молекулы, содержащие вариант Fc-домена, где указанный вариант обеспечивает или опосредует увеличенную активность ADCC и/или повышенное связывание с FcγRIIA (CD32A) относительно молекулы, не содержащей Fc-домен или содержащей Fc-домен дикого типа, при определении с использованием способов, известных специалисту в данной области и описанных в настоящем документе. В альтернативных вариантах осуществления изобретение охватывает молекулы, содержащие вариант Fc-домена, где указанный вариант обеспечивает или опосредует сниженную активность ADCC (или другую эффекторную функцию) и/или увеличенное связывание с FcγRIIB (CD32B), относительно молекулы, не содержащей Fc-домен или содержащей Fc-домен дикого типа, при измерении с использованием способов, известных специалисту в данной области и описанных в настоящем документе.
Также изобретение охватывает применение Fc-домена, содержащего домены или области из двух или более изотипов IgG. Как известно в данной области, модификация аминокислот Fc-области может глубоко влиять на опосредуемую Fc эффекторную функцию и/или активность связывания. Однако эти изменения функциональных характеристик можно далее усовершенствовать и/или на них можно далее воздействовать, при осуществлении в контексте выбранных изотипов IgG. Аналогично, на отрицательные характеристики Fc изотипов можно воздействовать путем одной или нескольких модификаций аминокислот. Различные изотипы IgG (т.е. IgG1, IgG2, IgG3 и IgG4) проявляют различные физические и функциональные свойства, включая время полужизни в сыворотке, фиксацию комплемента, аффинность связывания FcγR и активность эффекторных функций (например, ADCC, CDC) вследствие отличий в аминокислотных последовательностях их шарнирных и/или Fc-доменов. В определенных вариантах осуществления модификацию аминокислот и Fc-области IgG независимо выбирают, исходя из их соответствующей отдельной активности связывания и/или эффекторной функции, для конструирования диантитела с желаемыми характеристиками. В большинстве вариантов осуществления указанные модификации аминокислот и шарнирной области/Fc-области IgG анализируют по отдельности в отношении активности связывания и/или эффекторной функции, как описано в настоящем документе или как известно в данной области в контексте IgG1. В определенных вариантах осуществления указанная модификация аминокислот и шарнирной области/Fc-области IgG проявляет сходную функциональность, например, повышенную аффинность к FcγRIIA, при анализе отдельно связывания FcγR или эффекторной функции в контексте молекулы диантитела или другой содержащей Fc молекулы (например, иммуноглобулина). Комбинация указанной модификации аминокислот и выбранной Fc-области IgG затем действует аддитивно или, более предпочтительно, синергично, для модификации указанной функциональности в молекуле диантитела по изобретению, относительно молекулы диантитела по изобретению, содержащей Fc-область дикого типа. В других вариантах осуществления указанная модификация аминокислот и Fc-области IgG проявляет противоположную функциональность, например, повышенную и сниженную, соответственно, аффинность к FcγRIIA, при анализе по отдельности в отношении связывания FcγR и/или эффекторной функции в контексте молекулы диантитела или другой содержащей Fc молекулы (например иммуноглобулина), содержащей Fc-область дикого типа, как описано в настоящем документе или известно в данной области; затем комбинация указанной "противоположной" модификации аминокислот и выбранной области IgG действует, селективно сдерживая или снижая специфическую функциональность в диантителе по изобретению относительно диантитела по изобретению, не содержащего Fc-область или содержащего Fc-область дикого типа того же изотипа. Альтернативно изобретение охватывает варианты Fc-областей, содержащие комбинации модификаций аминокислот, известных в данной области, и выбранных областей IgG, которые проявляют новые свойства, которые не поддаются детекции, когда указанные модификации и/или области анализируют независимо, как описано в настоящем документе.
Функциональные характеристики множества изотипов IgG и их доменов хорошо известны в данной области. Аминокислотные последовательности IgG1, IgG2, IgG3 и IgG4 представлены на ФИГ. 1A-1B. Выбор и/или комбинации двух или более доменов из конкретных изотипов IgG для применения в способах по изобретению может быть основан на любом известном параметре исходных изотипов, включая аффинность к FcγR (таблица 7; Flesch et al. (2000) "Functions Of The Fc Receptors For Immunoglobulin G", J. Clin. Lab. Anal. 14:141-156; Chappel et al. (1993) "Identification Of A Secondary Fc Gamma RI Binding Site Within A Genetically Engineered Human IgG Antibody", J. Biol. Chem. 33:25124-25131; Chappel et al. (1991) "Identification Of The Fc Gamma Receptor Class I Binding Site In Human IgG Through The Use Of Recombinant IgGI/IgG2 Hybrid And Point-Mutated Antibodies", Proc. Natl. Acad. Sci. USA 88:9036-9040, все из которых включены в настоящий документ в качестве ссылок в полном объеме). Например, использование областей или доменов из изотипов IgG, которые проявляют ограниченное связывание или отсутствие связывания с FcγRIIB, например, IgG2 или IgG4, может быть особенно применимым, когда желательно сконструировать диантитело для максимального увеличения связывания с активирующим рецептором и минимизации связывания с ингибиторным рецептором. Аналогично применение Fc-областей или доменов из изотипов IgG, о которых известно, что они предпочтительно связывают C1q или FcγRIIIA, например, IgG3 (Bruggemann et al. (1987) "Comparison Of The Effector Functions Of Human Immunoglobulin Using A Matched Set Of Chimeric Antibodies", J. Exp. Med. 166:1351-1361), можно комбинировать с модификациями аминокислот Fc, известными в данной области, для усиления ADCC, в целях конструирования молекулы диантитела, так чтобы активность эффекторной функции, например, активации комплемента или ADCC, максимально увеличивалась.
Основные характеристики связывания IgG с FcγR, взятые из Flesch and Neppert, 1999, J. Clin. Lab. Anal. 14:141-156
отсутствие связывания: IgG2
отсутствие связывания: IgG2, IgG4
отсутствие связывания: IgG4
отсутствие связывания: IgG2
отсутствие связывания: IgG2, IgG4
5.3 МОЛЕКУЛЯРНЫЕ КОНЪЮГАТЫ
Молекулы диантител по изобретению можно подвергать рекомбинантному слиянию или химической конъюгации (включая как ковалентные, так и нековалентные конъюгации) с гетерологичными полипептидами, т.е. неродственным полипептидом или его частью, предпочтительно по меньшей мере из 10, по меньшей мере из 20, по меньшей мере из 30, по меньшей мере из 40, по меньшей мере из 50, по меньшей мере из 60, по меньшей мере из 70, по меньшей мере из 80, по меньшей мере из 90 или по меньшей мере из 100 аминокислот полипептида, с получением слитых белков. Слияние не обязательно должно быть прямым, а может происходить через линкерные последовательности.
Кроме того, молекулы диантител по изобретению (т.е. полипептиды) можно конъюгировать с лекарственным средством или группой лекарственного средства, которая модифицирует данный биологический ответ. В качестве альтернативы прямой конъюгации, благодаря множеству участков связывания эпитопа на поливалентных, например тетравалентных, молекулах диантител по изобретению, по меньшей мере один участок связывания на диантителе может быть предназначен для связывания лекарственного средства или желаемой группы лекарственного средства без влияния на связывание диантитела.
Лекарственные средства или группы лекарственных средств не следует понимать как ограниченные классическими химическими лекарственными средствами. Например, группа лекарственного средства может представлять собой белок или полипептид, обладающий желаемой биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин A, экзотоксин pseudomonas (т.е. PE-40), или дифтерийный токсин, рицин, гелонин и противовирусный белок лаконоса, такой белок, как фактор некроза опухоли, интерфероны, включая, но не ограничиваясь ими, α-интерферон (IFN-α), β-интерферон (IFN-β), фактор роста нервов (NGF), тромбоцитарный фактор роста (PDGF), тканевой активатор плазминогена (TPA), апоптотическое средство (например, TNF-α, TNF-β, AIM I, как описано в публикации PCT WO 97/33899), AIM II (см., публикация PCT WO 97/34911), Fas-лиганд, и VEGI (публикация PCT No. WO 99/23105), тромботическое средство или антитромботическое средство (например, ангиостатин или эндостатин) или модификатор биологического ответа, например, такой как лимфокин, например, интерлейкин-1 ("IL-1"), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6"), гранулоцитарно-макрофагальный колониестимулирующий фактор ("GM-CSF") и гранулоцитарный колониестимулирующий фактор ("G-CSF"), макрофагальный колониестимулирующий фактор, ("M-CSF") или фактор роста, например, гормон роста ("GH"); протеазы или рибонуклеазы.
Молекулы диантител по изобретению (т.е. полипептиды) можно подвергать слиянию с маркерными последовательностями, такими как пептид, для упрощения очистки. В предпочтительных вариантах осуществления маркерная аминокислотная последовательность представляет собой гексагистидиновый пептид, такой как, среди прочих, метка, находящаяся в векторе pQE (QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, CA, 91311), многие из которых коммерчески доступны. Как описано в Gentz et al. (1989) "Bioassay For Trans-Activation Using Purified Human Immunodeficiency Virus TAT-Encoded Protein: Trans-Activation Requires mRNA Synthesis", Proc. Natl. Acad. Sci. USA, 86:821-824, например, гексагистидин обеспечивает удобную очистку слитого белка. Другие пептидные метки, пригодные для очистки, включают, но не ограничиваются ими, гемагглютининовую метку "HA", которая соответствует эпитопу, происходящему из белка гемагглютинина вируса гриппа (Wilson et al. (1984) "The Structure Of An Antigenic Determinant In A Protein", Cell, 37:767-778) и "flag"-метку (Knappik et al. (1994) "An Improved Affinity Tag Based On The FLAG Peptide For The Detection And Purification Of Recombinant Antibody Fragments", Biotechniques, 17(4):754-761).
Дополнительные слитые белки можно получать способами "перетасовки" генов, "перетасовки" мотивов, "перетасовки" экзонов и/или "перетасовки" кодонов (в совокупности, называемыми "перетасовкой ДНК"). "Перетасовку" ДНК можно использовать для изменения активности молекул по изобретению (например, участки связывания эпитопа с более высокой аффинностью и более низкими скоростями диссоциации). См., главным образом, патенты США №№ 5605793; 5811238; 5830721; 5834252 и 5837458, и Patten et al. (1997) "Applications Of DNA Shuffling To Pharmaceuticals And Vaccines", Curr. Opinion Biotechnol. 8:724-733; Harayama (1998) "Artificial Evolution By DNA Shuffling", Trends Biotechnol. 16:76-82; Hansson et al. (1999) "Evolution Of Differential Substrate Specificities In Mu Class Glutathione Transferases Probed By DNA Shuffling", J. Mol. Biol. 287:265-276; и Lorenzo et al. (1998) "PCR-Based Method For The Introduction Of Mutations In Genes Cloned And Expressed In Vaccinia Virus", BioTechniques 24:308-313 (все из этих патентов и публикаций включены в настоящий документ в качестве ссылок в полном объеме). Молекулы диантител по изобретению или нуклеиновых кислот, кодирующих молекулы по изобретению, можно далее изменять, подвергая их случайному мутагенезу посредством ПЦР с пониженной точностью, случайному встраиванию нуклеотидов или другим способам, перед рекомбинацией. Одну или несколько частей полинуклеотида, кодирующего молекулу по изобретению, можно рекомбинировать с одним или несколькими компонентами, мотивами, сегментами, частями, доменами, фрагментами и т.д. одной или нескольких гетерологичных молекул.
Также настоящее изобретение охватывает молекулы диантител по изобретению, конъюгированные с диагностическим или лекарственным средством или любой другой молекулой, для которых желательно повышение/снижение времени полужизни в сыворотке, или иммуноспецифично распознающие его, и/или нацеленные на конкретную группу клеток. Молекулы по изобретению можно использовать в диагностике, например, для мониторинга развития или прогрессирования заболевания, нарушения или инфекции, в качестве части процедуры клинического тестирования, например, для определения эффективности данного режима лечения. Детекцию можно упрощать путем связывания молекул по изобретению с поддающимся детекции веществом или с помощью молекул, иммуноспецифично распознающих поддающееся детекции вещество. Примеры поддающихся детекции веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы, радиоактивные материалы, позитронно-эмиссионные металлы и нерадиоактивные ионы парамагнитных металлов. Поддающееся детекции вещество можно связывать или конъюгировать либо прямо с молекулами по изобретению, либо непрямо, через промежуточное соединение (например, такое как линкеры, известные в данной области) с использованием способов, известных в данной области, или молекула может иммуноспецифично распознавать поддающееся детекции вещество, иммунологически связывая указанное вещество. См., например, патент США № 4741900 для ионов металлов, которые можно конъюгировать с антителами для применения в качестве диагностических средств в соответствии с настоящим изобретением. Такую диагностику и детекцию можно проводить, конструируя молекулы для иммуноспецифического распознавания поддающегося детекции вещества или путем связывания молекул по изобретению с поддающимися детекцию веществами, включая, но не ограничиваясь ими, различные ферменты, включающие, но не ограничивающиеся ими, пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; комплексы простетических групп, такие как, но не ограничиваясь ими, стрептавидин/биотин и авидин/биотин; флуоресцентные материалы, такие как, но не ограничиваясь ими, умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин; люминесцентный материал, такой как, но не ограничиваясь этим, люминол; биолюминесцентные материалы, такие как, но не ограничиваясь ими, люцифераза, люциферин и экворин; радиоактивный материал, такой как, но не ограничиваясь ими, висмут (213Bi), углерод (14C), хром (51Cr), кобальт (57Co), фтор (18F), гадолиний (153Gd, 159Gd), галлий (68Ga, 67Ga), германий (68Ge), гольмий (166Ho), индий (115In, 113In, 112In, 111In), йод (131I, 125I, 123I, 121I), лантан (140La), лютеций (177Lu), марганец (54Mn), молибден (99Mo), палладий (103Pd), фосфор (32P), празеодимий (142Pr), прометий (149Pm), рений (186Re, 188Re), родий (105Rh), рутений (97Ru), самарий (153Sm), скандий (47Sc), селен (75Se), стронций (85Sr), сера (35S), технеций (99Tc), таллий (201Ti), олово (113Sn, 117Sn), тритий (3H), ксенон (133Xe), иттербий (169Yb, 175Yb), иттрий (90Y), цинк (65Zn); позитронно-эмисионные металлы с использованием различных позитронно-эмиссионных томографий, и нерадиоактивные ионы парамагнитных металлов.
Молекулы диантител по изобретению могут иммуноспецифично распознавать терапевтическую группу, такую как цитотоксин (например, цитостатическое или цитоцидное средство), лекарственное средство или радиоактивный элемент (например, альфа-эмиттеры, гамма-эмиттеры и т.д.). Цитотоксины или цитотоксические средства включают любое средство, вредное для клеток. Их примеры включают паклитаксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи, или они могут быть конъюгированы с ними. Лекарственные средства включают, но не ограничиваются ими, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацилдекарбазин), алкилирующие средства (например, мехлорэтамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин C и цисдихлордиаминплатину (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)), и антимитотические средства (например, винкристин и винбластин).
Более того, молекулу диантитела по изобретению можно конъюгировать с терапевтическими группами, такими как радиоактивные материалы или макроциклические хелаторы, пригодные для конъюгации ионов радиоактивных металлов (см. выше для примеров радиоактивных материалов), или она может быть сконструирована для иммуноспецифичного распознавания их. В определенных вариантах осуществления макроциклический хелатор представляет собой 1,4,7,10-тетраазациклододекан-N,N',N",N'"-тетрауксусную кислоту (DOTA), которая может быть связана с полипептидом через линкерную молекулу. Такие линкеры широко известны в данной области и описаны в Denardo et al. (1998) "Comparison Of 1,4, 7,10-Tetraazacyclododecane-N,N',N",N'"-Tetraacetic acid (DOTA)-Peptide-ChL6, A Novel Immunoconjugate With Catabolizable Linker, To 2-Iminothiolane-2-[p-(bromoacetamido)benzyl]-DOTA-ChL6 In Breast Cancer Xenografts", Clin. Cancer Res. 4:2483-2490; Peterson et al. (1999) "Enzymatic Cleavage Of Peptide-LinkedRadiolabels From Immunoconjugates", Bioconjug. Chem. 10:553-; и Zimmerman et al., (1999) "A Triglycerine Linker Improves Tumor Uptake And Biodistributions Of 67-Cu-Labeled Anti-Neuroblastoma mAb chCE7 F(ab)'2 Fragments", Nucl. Med. Biol. 26:943-950, все из которых включены в настоящий документ в качестве ссылок в полном объеме.
Способы конъюгации таких терапевтических групп с полипептидами, включая например, Fc-домены, хорошо известны; см., например, Arnon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), 1985, pp. 243-56, Alan R. Liss, Inc.); Hellstrom et al, "Antibodies For Drug Delivery", Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), 1987, pp. 623-53, Marcel Dekker, Inc.); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera et al. (eds.), 1985, pp. 475-506); "Analysis, Results And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), 1985, pp. 303-16, Academic Press; и Thorpe et al. (1982) "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-158.
Молекулу диантитела по изобретению можно вводить с терапевтической группой, конъюгированной с ним, или без нее, отдельно или в комбинации с цитотоксическим фактором(ами) и/или цитокином(ами) для применения в качестве терапевтического лечения. При введении отдельно по меньшей мере один эпитоп поливалентной, например тетравалентной, молекулы диантитела может быть сконструирован для иммуноспецифического распознавания лекарственного средства, например, цитотоксичесокого фактора(ов) и/или цитокина(ов), которые можно вводить одновременно с молекулой по изобретению или после нее. Таким путем молекула диантитела может нацеливать лекарственное средство аналогично прямой конъюгации. Альтернативно молекулу по изобретению можно конъюгировать с антителом с образованием гетероконъюгата, как описано Segal в патенте США № 4676980, который включен в настоящий документ в качестве ссылки в полном объеме. Молекулы диантител по изобретению также можно связывать с твердыми подложками, которые особенно пригодны для иммуноанализов или очистки антигена-мишени. Такие твердые подложки включают, но не ограничиваются ими, стекло, целлюлозу, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен.
5.4 ОХАРАКТЕРИЗАЦИЯ СВЯЗЫВАНИЯ МОЛЕКУЛАМИ ДИАНТИТЕЛ
Молекулы диантител по настоящему изобретению могут быть охарактеризованы различными путями. В частности, молекулы по изобретению можно анализировать в отношении способности иммуноспецифично связываться с антигеном, например, FcRIIIA или FcRIIB, или, где молекула содержит Fc-домен (или его часть), в отношении способности проявлять взаимодействия Fc-FcγR, т.е. специфическое связывание Fc-домена (или его части) с FcγR. Такой анализ можно проводить в растворе (например, Houghten (1992) "The Use Of Synthetic Peptide Combinatorial Libraries For The Identification Of Bioactive Peptides", BioTechniques, 13:412-421), на гранулах (Lam (1991) "A New Type Of Synthetic Peptide Library For Identifying Ligand-Binding Activity", Nature, 354:82-84), на чипах (Fodor (1993) "Multiplexed Biochemical Assays With Biological Chips", Nature, 364:555-556), на бактериях (патент США № 5223409), на спорах (патенты США №№ 5571698; 5403484 и 5223409), на плазмидах (Cull et al. (1992) "Screening For Receptor Ligands Using Large Libraries Of Peptides Linked To The C Terminus Of The Lac Repressor", Proc. Natl. Acad. Sci. USA, 89:1865-1869) или на фаге (Scott et al. (1990) "Searching For Peptide Ligands With An Epitope Library", Science, 249:386-390; Devlin (1990) "Random Peptide Libraries: A Source Of Specific Protein Binding Molecules", Science, 249:404-406; Cwirla et al. (1990) "Peptides On Phage: A Vast Library Of Peptides For Identifying Ligands, "Proc. Natl. Acad. Sci. USA, 87:6378-6382; и Felici (1991) "Selection Of Antibody Ligands From A Large Library Of Oligopeptides Expressed On A Multivalent Exposition Vector", J. Mol. Biol., 222:301-310) (каждая из этих ссылок включена в настоящий документ в качестве ссылки в полном объеме). Затем молекулы, идентифицированные как связывающиеся с антигеном, например, FcγRIIIA, можно анализировать в отношении их специфичности и аффинности к антигену.
Молекулы по изобретению, сконструированные так, чтобы они содержали множество связывающих эпитоп доменов, можно анализировать в отношении иммуноспецифического связывания с одним или несколькими антигенами (например, антиген злокачественной опухоли и перекрестная реактивность с другими антигенами (например, FcγR)) или, где молекулы содержат Fc-домен (или его часть) в отношении взаимодействий Fc-FcγR любым способом, известным в данной области. Иммуноанализы, которые можно использовать для анализа иммуноспецифического связывания, перекрестной реактивности и взаимодействий Fc-FcγR, включают, но не ограничиваются ими, конкурентные и неконкурентные системы анализа с использованием способов, таких как вестерн-блоттинг, радиоиммунные анализы, ELISA (твердофазный иммуноферментный анализ), "сэндвич"-иммуноанализы, анализы иммунопреципитации, реакции преципитации, реакции преципитации с диффузией в геле, анализы иммунодиффузии, анализы агглютинации, анализы фиксации комплемента, иммунорадиометрические анализы, флуоресцентные иммуноанализы, иммуноанализы с белком, среди прочих. Такие анализы являются общепринятыми и хорошо известны в данной области (см., например, Ausubel et al., eds, 1994, Current Protocols in Molecular Biology, Vol. 1, John Wiley & Sons, Inc., New York, которая включена в настоящий документ в качестве ссылки в полном объеме).
Аффинность связывания и константу диссоциации при взаимодействии антиген-связывающий домен или Fc-FcγR можно определять конкурентными анализами связывания. Одним из примеров конкурентного анализа связывания является радиоиммунный анализ, включающий инкубацию меченого антигена, такого как тетрамерный FcγR (например, 3H или 125I, см. раздел 5.4.1), с представляющей интерес молекулой (например, молекулами по настоящему изобретению, содержащими множество связывающих эпитоп доменов, в присутствии возрастающих количеств немеченого эпитопа, такого как тетрамерный FcγR (см. раздел 5.4.1)), и детекцию молекулы, связавшейся с меченым антигеном. Аффинность молекулы по настоящему изобретению к антигену и константы диссоциации при связывании можно определять из данных по насыщению в анализе Скэтчарда.
Аффинность и свойства связывания молекул по изобретению в отношении антигена или FcγR можно первоначально определять с использованием анализов in vitro (биохимические или иммунологические анализы), известных в данной области, для взаимодействий антиген-связывающий домен или Fc-FcγR, включая, но не ограничиваясь ими, анализ ELISA, анализ поверхностного плазмонного резонанса, анализы иммунопреципитации. Предпочтительно свойства связывания молекул по изобретению также охарактеризовывают функциональными анализами in vitro для определения одной или нескольких опосредуемых FcγR функций эффекторных клеток, как описано в разделе 5.4.2. В наиболее предпочтительных вариантах осуществления молекулы по изобретению обладают свойствами связывания в моделях in vivo (таких как описаны и раскрыты в настоящем документе), сходными со свойствами связывания в анализах in vitro. Однако настоящее изобретение не исключает молекулы по изобретению, которые не проявляют желаемый фенотип в анализах in vitro, но проявляют желаемый фенотип in vivo.
В некоторых вариантах осуществления скрининг и идентификацию молекул, содержащих множество связывающих эпитоп доменов и, необязательно, Fc-доменов (или их частей), проводят в функциональных анализах, предпочтительно, высокопроизводительным способом. Функциональные анализы могут представлять собой любой анализ, известный в данной области, для охарактеризации одной или нескольких опосредуемых FcγR функций эффекторных клеток, таких как функции, описанные в настоящем документе в разделах 5.4.2 и 5.4.3. Неограничивающие примеры функций эффекторных клеток, которые можно использовать в соответствии со способами по изобретению, включают, но не ограничиваются ими, антителозависимую клеточно-опосредуемую цитотоксичность (ADCC), антителозависимый фагоцитоз, фагоцитоз, опсонизацию, опсонофагоцитоз, связывание клеток, розеткообразование, связывание C1q и комплементзависимую клеточноопосредуемую цитотоксичность.
В предпочтительном варианте осуществления используют кинетический анализ BIAcore для определения констант ассоциации и диссоциации при связывании молекул по настоящему изобретению с антигеном или/и FcγR. Кинетический анализ BIAcore включает анализ связывания антигена или FcγR с чипами с иммобилизованными молекулами (например, молекулы, содержащие связывающие эпитоп домены или Fc-домены (или их части), соответственно) на их поверхности и диссоциации антигена или FcγR с этих чипов. Анализ BIAcore описан в разделе 5.4.3.
Предпочтительно, для иммунологического или функционального анализа в целях охарактеризации молекул по изобретению используют активированную флуоресценцией сортировку клеток (FACS), с использованием любого из способов, известных специалистам в данной области. Проточные сортеры способны быстро исследовать большое количество отдельных клеток, связанных, например, опсонизированных, молекулами по изобретению (например, 10-100 миллион клеток в час) (Shapiro et al. (1995) Practical Flow Cytometry). Кроме того, конкретные параметры, используемые для оптимизации воздействия диантитела, включают но не ограничиваются ими, концентрацию антигена (т.е. тетрамерного комплекса FcγR, см. раздел 5.4.1), время кинетического конкурирования или строгость FACS, каждый из которых можно варьировать для отбора молекул диантител, содержащих молекулы по изобретению, которые проявляют определенные свойства связывания, например, одновременное связывание со множеством эпитопов. Проточные цитометры для сортировки и биологического исследования клеток хорошо известны в данной области. Известные проточные цитометры описаны, например, в патентах США №№ 4347935; 5464581; 5483469; 5602039; 5643796 и 6211477; полное содержание которых включено в настоящий документ в качестве ссылок. Другие проточные цитометры представляют собой систему FACS VantageTM, изготовленную Becton Dickinson and Company, и систему COPASTM, изготовленную Union Biometrica.
Охарактеризацию аффинности связывания антигена-мишени или аффинности связывания Fc-FcγR, и оценку плотности антигена мишени или FcγR на поверхности клетки можно проводить способами, хорошо известными в данной области, такими как анализ Скэтчарда, или с использованием наборов в соответствии с инструкциями изготовителя, таких как QuantumTM Simply Cellular® (Bangs Laboratories, Inc., Fishers, IN). Один или несколько функциональных анализов может представлять собой любой анализ, известный в данной области, для охарактеризации одной или нескольких опосредуемых FcγR функций эффекторных клеток, как известно специалисту в данной области или описано в настоящем документе. В конкретных вариантах осуществления молекулы по изобретению, содержащие множество связывающих эпитоп доменов и, необязательно, и Fc-домен (или его часть), анализируют в анализе ELISA в отношении связывания с одним или несколькими антигенами-мишенями или одним или несколькими FcγR, например, FcγRIIIA, FcγRIIA, FcγRIIA; а затем в одном или нескольких анализах ADCC. В некоторых вариантах осуществления молекулы по изобретению далее анализируют с использованием анализа на основе поверхностного плазмонного резонанса, например, BIAcore. Анализы на основе поверхностного плазмонного резонанса хорошо известны в данной области, и далее рассмотрены в разделе 5.4.3, и проиллюстрированы в настоящем документе, например, в примере 6.1.
В наиболее предпочтительных вариантах осуществления молекулы по изобретению, содержащие множество связывающих эпитоп доменов и, необязательно, и Fc-домен (или его часть), далее охарактеризовывают в модели на животных для взаимодействия с антигеном-мишенью (например, FcγR) или для взаимодействия Fc-FcγR. Когда оценке подлежат взаимодействия Fc-FcγR, предпочтительными моделями на животных для применения в способах по изобретению являются, например, трансгенные мыши, экспрессирующие FcγR человека, например, любая модель на мышах, описанная в патентах США №№ 5877397 и 6676927, которые включены в настоящий документ в качестве ссылок в полном объеме. Кроме того, трансгенные мыши для применения в таких способах включают, но не ограничиваются ими, мышей nude c нокаутом FcγRIIIA, несущих FcγRIIIA человека; мышей nude с нокаутом FcγRIIIA, несущих FcγRIIA человека; мышей nude c нокаутом FcγRIIIA, несущих FcγRIIB человека и FcγRIIIA человека; мышей nude c нокаутом FcγRIIIA, несущих FcγRIIB человека и FcγRIIA человека; мышей nude c нокаутом FcγRIIIA и FcγRIIA, несущих FcγRIIIA и FcγRIIA человека, и мышей nude с нокаутом FcγRIIIA, FcγRIIA и FcγRIIB, несущих FcγRIIIA, FcγRIIA и FcγRIIB человека.
5.4.1 АНАЛИЗЫ СВЯЗЫВАНИЯ, ВКЛЮЧАЮЩИЕ FcγR
Охарактеризацию связывания FcγR молекулами, содержащими Fc-домен (или его часть) и/или содержащими связывающий эпитоп домен, специфичный к FcγR, можно проводить с использованием любого FcγR, включая, но не ограничиваясь ими полиморфные варианты FcγR. В некоторых вариантах осуществления используют полиморфный вариант FcγRIIIA, который содержит фенилаланин в положении 158. В других вариантах осуществления охарактеризацию проводят с использованием полиморфного варианта FcγRIIIA, который содержит валин в положении 158. FcγRIIIA 158V проявляет более высокую аффинность к IgG1, чем 158F, и увеличенную активность ADCC (см., например, Koene et al. (1997) "Fc gammaRIIIa-158V/F Polymorphism Influences The Binding Of IgG By Natural Killer Cell Fc gammaRIIIa, Independant Of The Fc gammaRIIIa-48L/R/H Phenotype", Blood, 90:1109-14; Wu et al. (1997) "A Novel Polymorphism Of FcgammaRIIIa (CDI 6) Alters Receptor Function And Predisposes To Autoimmune Disease", J. Clin. Invest. 100: 1059-70, которые обе включены в настоящий документ в качестве ссылок в полном объеме); этот остаток в действительности прямо взаимодействует с нижней шарнирной областью IgG1, как недавно показано в исследованиях кокристаллизации IgG1-FcγRIIIA, см., например, Sondermann et al. (2000) "The 3.2-A Crystal Structure Of The Human IgG1 Fc Fragment-FcgammaRIII complex", Nature, 406(6793):267-273, которая включена в настоящий документ в качестве ссылки в полном объеме. В исследованиях было показано, что в некоторых случаях терапевтические антитела обладают улучшенной эффективностью у гомозиготных по FcγRIIIA-158V пациентов. Например, гуманизированное моноклональное антитело против CD20 ритуксимаб было терапевтически более эффективным у гомозиготных по FcγRIIIA158V пациентов по сравнению с гомозиготными по FcγRIIIA 158F пациентами (см., например, Cartron et al. (2002) "Therapeutic Activity Of Humanized Anti-CD20 Monoclonal Antibody And Polymorphysm In IgG Fc Receptor FcgammaRIIIA Gene", Blood, 99(3):754-758). В других вариантах осуществления терапевтические молекулы, содержащие эту область, также могут быть более эффективными у пациентов, гетерозиготных по FcγRIIIA-158V и FcγRIIIA-158F, и у пациентов с FcγRIIA-131H. Без связи с конкретным механизмом действия, выбор молекул по изобретению с альтернативными аллотипами может обеспечить варианты, которые после встраивания в терапевтические диантитела будут клинически более эффективными у пациентов, гомозиготных по указанному аллотипу.
Анализ связывания FcγR был разработан для определения связывания молекул по изобретению с FcγR и, в частности, для определения связывания Fc-доменов с FcγR. Этот анализ позволил выявление и количественное определение взаимодействий Fc-FcγR, несмотря на слабую по своей природе аффинность рецептора к его лиганду, например, в микромолярном диапазоне для FcγRIIB и FcγRIIIA. Этот способ подробно описан в международной заявке WO04/063351 и публикациях патентных заявок США 2005/0037000 и 2005/0064514, которые все включены в настоящий документ в качестве ссылок в полном объеме. В кратком изложении, способ вовлекает образование комплекса FcγR, который можно использовать в любом стандартном иммуноанализе, известном в данной области, например, FACS, ELISA, поверхностный плазмонный резонанс и т.д. Кроме того, комплекс FcγR обладает повышенной авидностью к Fc-области, относительно FcγR не в комплексе. Согласно изобретению, предпочтительным молекулярным комплексом является тетрамерный иммунный комплекс, содержащий: (a) растворимую область FcγR (например, растворимую область FcγRIIIA, FcγRIIA или FcγRIIB); (b) биотинилированную последовательность из 15 аминокислот AVITAG (AVITAG), функционально связанную с C-концом растворимой области FcγR (например, растворимой области FcγRIIIA, FcγRIIA или FcγRIIB); и (c) стрептавидин-фикоэритрин (SA-PE); в таком молярном соотношении, чтобы образовывался тетрамерный комплекс FcγR (предпочтительно в молярном соотношении 5:1). Слитый белок биотинилируют ферментативно с использованием, например, фермента Bir A E. coli, биотинлигазы, которая специфично биотинилирует остаток лизина в последовательности из 15 аминокислот AVITAG. Затем биотинилированные растворимые белки FcγR смешивают с SA-PE в молярном соотношении 1X SA-PE:5X биотинилированный растворимый FcγR с получением тетрамерного комплекса FcγR.
Было показано, что полипептиды, содержащие Fc-области, связывают тетрамерные комплексы FcγR с аффинностью, по меньшей мере в 8 раз превышающей аффинность мономерного FcγR не в комплексе. Связывание полипептидов, содержащих Fc-области, с тетрамерными комплексами FcγR, можно определять с использованием стандартных способов, известных специалистам в данной области, например, таких как активированная флуоресценцией сортировка клеток (FACS), радиоиммунные анализы, анализы ELISA и т.д.
Изобретение охватывает применение иммунных комплексов, содержащих молекулы по изобретению, и образованных согласно способам, описанным выше, для определения функциональности молекул, содержащих Fc-область, в клеточных или бесклеточных анализах.
Для удобства реагенты могут быть предоставлены в наборе для анализа, т.е. в упакованной комбинации реагентов для анализа способности молекул, содержащих Fc-области, связывать тетрамерные комплексы FcγR. Также предусматриваются другие формы молекулярных комплексов для применения в определении взаимодействий Fc-FcγR в целях применения в способах по изобретению, например, формы слитых белков, как описано в предварительной заявке США 60/439709, поданной 13 января 2003 года, которая включена в настоящий документ в качестве ссылки в полном объеме.
5.4.2 ФУНКЦИОНАЛЬНЫЕ АНАЛИЗЫ МОЛЕКУЛ С ВАРИАНТАМИ ТЯЖЕЛЫХ ЦЕПЕЙ
Изобретение охватывает охарактеризацию молекул по изобретению, содержащих множество связывающих эпитоп доменов и, необязательно, Fc-доменов (или их частей) с использованием анализов, известных специалистам в данной области, для идентификации функции молекул в отношении эффекторных клеток. В частности, изобретение охватывает охарактеризацию молекул по изобретению в отношении опосредуемой FcγR функции эффекторных клеток. Кроме того, когда по меньшей мере один из антигенов-мишеней молекулы диантитела по изобретению представляет собой FcγR, связывание FcγR молекулой диантитела может служить для активации опосредуемых FcγR каскадов, сходных с каскадами, активируемыми связыванием FcγR-Fc. Таким образом, когда по меньшей мере один связывающий эпитоп домен молекулы диантитела распознает FcγR, молекула диантитела может вызывать опосредуемую FcγR эффекторную функцию без наличия в ней Fc-домена (или его части), или без сопутствующего связывания Fc-FcγR. Примеры функций эффекторных клеток, которые можно анализировать согласно изобретению, включают, но не ограничиваются ими, антителозависимую клеточноопосредуемую цитотоксичность, фагоцитоз, опсонизацию, опсонофагоцитоз, связывание C1q и комплементзависимую клеточноопосредуемую цитотоксичность. Для определения активности в отношении функции эффекторных клеток можно использовать любой клеточный или бесклеточный анализ, известный специалистам в данной области (для анализа эффекторных клеток см. Perussia et al. (2000) "Assays For Antibody-Dependant Cell-Mediated Cytotoxicity (ADCC) And Reverse ADCC (Redirected Cytotoxicity) In Human Natural Killer Cells", Methods Mol. Biol. 121:179-92; Baggiolini et al. (1988) "Cellular Models For The Detection And Evaluation Of Drugs That Modulate Human Phagocyte Activity", Experientia, 44(10):841-848; Lehmann et al. (2000) "Phagocytosis: Measurement By Flow Cytometry", J. Immunol. Methods, 243(1-2):229-42; Brown (1994) "In Vitro Assays Of Phagocytic Function Of Human Peripheral Blood Leukocytes: Receptor Modulation And Signal Transduction", Methods Cell Biol, 45: 147-64; Munn et al. (1990) "Phagocytosis Of Tumor Cells By Human Monocytes Cultured In Recombinant Macrophage Colony-Stimulating Factorx", J. Exp. Med., 172:231-237, Abdul-Majid et al. (2002) "Fc Receptors Are Critical For Autoimmune Inflammatory Damage To The Central Nervous System In Experimental Autoimmune Encephalomyelitis", Scand. J. Immunol. 55:70-81; Ding et al. (1998) "Two Human T Cell Receptors Bind In A Similar Diagonal Mode To The HLA-A2/Tax Peptide Complex Using Different TCR Aminoacids", Immunity 8:403-411, которые все включены в настоящий документ в качестве ссылки в полном объеме).
В одном из вариантов осуществления молекулы по изобретению можно анализировать в отношении опосредуемого FcγR фагоцитоза в моноцитах человека. Альтернативно опосредуемый FcγR фагоцитоз молекул по изобретению можно анализировать в других фагоцитах, например, нейтрофилах (полиморфноядерные лейкоциты; PMN); моноцитах периферической крови человека, происходящих из моноцитов макрофагов, которые можно получать с использованием стандартных способов, известных специалистам в данной области (например, см. Brown (1994) "In Vitro Assays Of Phagocytic Function Of Human Peripheral Blood Leukocytes: Receptor Modulation And Signal Transduction", Methods Cell Biol., 45:147-164). В одном из вариантов осуществления функцию молекул по изобретению охарактеризовывают измерением способности клеток THP-1 фагоцитировать эритроциты, опсонизированные связанным с флуоресцеином IgG (SRBC) способами, описанными ранее (Tridandapani et al. (2000) "The Adapter Protein LAT Enhances Fcgamma Receptor-Mediated Signal Transduction In Myeloid Cells", J. Biol. Chem. 275:20480-20487).
Другим иллюстративным анализом для определения фагоцитоза молекул по изобретению является антителозависимый анализ опсонофагоцитоза (ADCP), который может включать следующее: покрытие заданной биочастицы, такой как Escherichia coli, меченные FITC (Molecular Probes) или Staphylococcus aureus-FITC (i) антителом 4-4-20 дикого типа, антителом к флуоресцеину (см. Bedzyk et al. (1989) "Comparison Of Variable Region Primary Structures Within An Anti-Fluorescein Idiotype Family", J. Biol. Chem, 264(3):1565-1569, которая включена в настоящий документ в качестве ссылки в полном объеме), в качестве контрольного антитела для FcγR-зависимого ADCP; или (ii) антителом 4-4-20, содержащим мутацию D265A, которая подавляет связывание с FcγRIII, в качестве фонового контроля для FcγR-зависимого ADCP (iii) диантителом, содержащим связывающий эпитоп домен 4-4-20 и Fc-домен и/или связывающий эпитоп домен, специфичный к FcγRIII; и образование опсонизированной частицы; добавление любой из описанных опсонизированных частиц (i-iii) к эффекторным клеткам THP-1 (моноцитарная клеточная линия, доступная от ATCC) в соотношении 1:1, 10:1, 30:1, 60:1, 75:1 или 100:1 для обеспечения опосредуемого FcγR фагоцитоза; предпочтительно инкубацию клеток и E. coli-FITC/антитела при 37°С в течение 1,5 часов; добавление трипанового синего после инкубации (предпочтительно при комнатной температуре в течение 2-3 минут) к клеткам без тушения флуоресценции бактерий, прикрепившихся снаружи клеточной поверхности без интернализации; перенос клеток в буфер для FACS (например, 0,1%, BSA в PBS, 0,1% азид натрия), анализ флуоресценции клеток THP1 с использованием FACS (например, BD FACS Calibur). Предпочтительно, клетки THP-1, используемые в этом анализе, анализируют с помощью FACS в отношении FcγR на клеточной поверхности. Клетки THP-1 экспрессируют как CD32A, так и CD64. CD64 представляет собой высокоаффинный FcγR, который блокируют при проведении анализа ADCP в соответствии со способами по изобретению. Клетки THP-1 предпочтительно блокируют 100 мкг/мл растворимого IgG1 или 10% сывороткой человека. Для анализа степени ADCP область предпочтительно устанавливают на клетки THP-1 и измеряют срединную интенсивность флуоресценции. Активность ADCP в отношении отдельных мутаций вычисляют и выражают в качестве величины, нормализованной к величине, полученной для chMab 4-4-20 дикого типа. Опсонизированные частицы добавляют к клеткам THP-1, так чтобы соотношение опсонизированных частиц и клеток THP-1 составляло 30:1 или 60:1. В наиболее предпочтительных вариантах осуществления анализ ADCP проводят с контролями, такими как E. coli-FITC в среде, E. coli-FITC и клетки THP-1 (в качестве FcγR-независимой активности ADCP), E. coli-FITC, клетки THP-1 и антитело 4-4-20 дикого типа (в качестве FcγR-зависимой активности ADCP), E. coli-FITC, клетки THP-1, 4-4-20 D265A (в качестве фонового контроля для FcγR-зависимой активности ADCP).
В другом варианте осуществления молекулы по изобретению можно анализировать в отношении опосредуемой FcγR активности ADCC в эффекторных клетках, например, естественных киллерных клетках, с использованием любого из стандартных способов, известных специалистам в данной области (см. например, Perussia et al. (2000) "Assays For Antibody-Dependant Cell-Mediated Cytotoxicity (ADCC) And Reverse ADCC (Redirected Cytotoxicity) In Human Natural Killer Cells", Methods Mol. Biol. 121:179-92; Weng et al. (2003) "Two Immunoglobulin G Fragment C Receptor Polymorphisms Independent Predict Response To Rituximab In Patients With Follicular Lymphoma", J. Clin. Oncol. 21:3940-3947; Ding et al. (1998) "Two Human T Cell Receptors Bind In A Similar Diagonal Mode To The HLA-A2/Tax Peptide Complex Using Different TCR Amino Acids", Immunity 8:403-411). Иллюстративный анализ для определения активности молекул по изобретению в отношении ADCC основан на анализе высвобождения 51Cr, включающем: мечение клеток-мишеней [51Cr]Na2CrO4 (эту проникающую через клеточную мембрану молекулу часто используют для мечения, поскольку она связывает цитоплазматические белки, и хотя она самопроизвольно выходит из клеток с медленной кинетикой, она массированно высвобождается после некроза клетки-мишени); опсонизацию клеток-мишеней молекулами по изобретению, содержащими варианты тяжелых цепей; комбинирование опсонизированных меченных радиоактивной меткой клеток-мишеней с эффекторными клетками в микропланшете для титрования в подходящем соотношении клеток-мишеней и эффекторных клеток; инкубацию смеси клеток в течение 16-18 часов при 37°C; сбор супернатантов; и анализ радиоактивности. Затем можно определять цитотоксичность молекул по изобретению, например, с использованием следующей формулы: %лизис = (экспериментальное значение cpm - значение cpm для выхода из мишени)/(значение cpm при лизисе детергентом - значение cpm для выхода из мишени)×100%. Альтернативно %лизис = (ADCC-AICC)/(максимальное высвобождение - самопроизвольное высвобождение). Специфический лизис можно вычислять с использованием формулы: специфический лизис = %лизис с молекулами по изобретению - %лизис в отсутствие молекул по изобретению. График можно строить, варьируя либо соотношение клетка-мишень:эффекторная клетка, либо концентрацию антитела.
Предпочтительно эффекторные клетки, используемые в анализах ADCC по изобретению, представляют собой мононуклеарные клетки периферической крови (PBMC), которые предпочтительно являются очищенными из нормальной крови человека с использованием стандартных способов, известных специалисту в данной области, например, с использованием центрифугирования в градиенте плотности Ficoll-Paque. Предпочтительные эффекторные клетки для применения в способах по изобретению экспрессируют различные активирующие рецепторы FcγR. Изобретение охватывает эффекторные клетки THP-1, экспрессирующие FcγRI, FcγRIIA и FcγRIIB, и происходящие из моноцитов первичные макрофаги, полученные из цельной крови человека, экспрессирующие как FcγRIIIA, так и FcγRIIB, для определения того, демонстрируют ли мутанты антител по тяжелой цепи увеличенную активность ADCC и фагоцитоз относительно антител IgG1 дикого типа.
В клеточной линии моноцитов человека THP-1 фагоцитоз активируется путем экспрессии высокоаффинного рецептора FcγRI и низкоаффинного рецептора FcγRIIA (Fleit et al. (1991) "The Human Monocyte-Like Cell Line THP-1 Expresses Fc Gamma RI And Fc Gamma RII", J. Leuk. Biol. 49:556-565). Клетки THP-1 не экспрессируют конститутивно FcγRIIA или FcγRIIB. Стимуляция этих клеток цитокинами влияет на паттерн экспрессии FcR (Pricop et al. (2001) "Differential Modulation Of Stimulatory And Inhibitory Fc Gamma Receptors On Human Monocytes By Th1 And Th2 Cytokines", J. of Immunol., 166:531-537). Рост клеток THP-1 в присутствии цитокина IL4 индуцирует экспрессию FcγRIIB и вызывает снижение экспрессии FcγRIIA и FcγRI. Экспрессия FcγRIIB также может усиливаться при увеличении плотности клеток (Tridandapani et al. (2002) "Regulated Expression And Inhibitory Function Of Fcgamma RIIB In Human Monocytic Cells", J. Biol. Chem., 277(7):5082-5089). Напротив, было описано, что IFNγ может приводить к экспрессии FcγRIIIA (Pearse et al. (1993) "Interferon Gamma-Induced Transcription Of The High-Affinity Fc Receptor For IgG Requires Assembly Of A Complex That Includes The 91-kDa Subunit Of Transcription Factor ISGF 3", Proc. Nat. Acad. Sci. USA 90:4314-4318). Присутствие или отсутствие рецепторов на поверхности клеток можно определять с помощью FACS с использованием обычных способов, известных специалисту в данной области. Индуцируемая цитокинами экспрессия FcγR на поверхности клеток обеспечивает систему для тестирования как активации, так и ингибирования, в присутствии FcγRIIB. Если клетки THP-1 неспособны экспрессировать FcγRIIB, изобретение также охватывает другую моноцитарную клеточную линию человека U937. Было показано, что эти клетки терминально дифференцируются в макрофаги в присутствии IFNγ и TNF (Koren et al. (1979) "In Vitro Activation Of A Human Macrophage-Like Cell Line", Nature 279:28-331).
FcγR-зависимое уничтожение опухоли опосредуется макрофагами и NK-клетками в моделях опухолей на мышах (Clynes et al. (1998) "Fc Receptors Are Required In Passive And Active Immunity To Melanoma", Proc. Nat. Acad. Sci. USA 95:652-656). Изобретение охватывает применение подвергнутых элютриации моноцитов от доноров в качестве эффекторных клеток для анализа эффективности мутантов по Fc в отношении запуска цитотоксичности клеток-мишеней в анализе как фагоцитоза, так и ADCC. На паттерны экспрессии FcγRI, FcγRIIIA и FcγRIIB влияют различные условия роста. Экспрессию FcγR из замороженных подвергнутых элютриации моноцитов, свежих подвергнутых элютриации моноцитов, моноцитов, содержащихся в 10% FBS, и моноцитов, культивированных в FBS + GM-CSF и/или в сыворотке человека, можно определять с использованием общепринятых способов, известных специалистам в данной области. Например, клетки можно окрашивать специфичными к FcγR антителами и анализировать посредством FACS для определения профилей FcR. Затем для способов по изобретению используют условия, которые наилучшим образом имитируют экспрессию FcγR макрофагами in vivo.
В некоторых вариантах осуществления изобретение охватывает применение клеток мышей, особенно, когда невозможно получить клетки человека с правильными профилями FcγR. В некоторых вариантах осуществления изобретение охватывает клеточную линию макрофагов мыши RA W264.7(ATCC), которую можно трансфицировать FcγRIIIA человека, и стабильные трансфектанты можно выделять с использованием способов, известных в данной области, см., например, Ralph et al. (1977) "Antibody-Dependent Killing Of Erythrocyte And Tumor Targets By Macrophage-Related Cell Lines: Enhancement By PPD And LPS", J. Immunol. 119:950-4). Экспрессию FcγRIIIA в трансфектантах можно количественно определять FACS-анализом с использованием общепринятого экспериментирования, и трансфектанты с высокой экспрессией можно использовать в анализах ADCC по изобретению. В других вариантах осуществления изобретение охватывает выделение перитонеальных макрофагов селезенки, экспрессирующих FcγR человека, из трансгенных мышей с нокаутом, таких как мыши, описанные в настоящем документе.
Лимфоциты можно собирать от доноров периферической крови (PBM) с использованием градиента Ficoll-Paque (Pharmacia). В популяции выделенных мононуклеарных клеток большая часть активности ADCC происходит через естественные киллерные клетки (NK), содержащие FcγRIIIA, но не FcγRIIB, на их поверхности. Результаты для этих клеток указывают на эффективность мутантов в отношении запуска ADCC NK-клеток и выявляют реагенты для тестирования с подвергнутыми элютриации моноцитами.
Клетки-мишени, используемые в анализах ADCC по изобретению, включают, но не ограничиваются ими, клеточные линии рака молочной железы, например SK-BR-3 с регистрационным номером ATCC HTB-30 (см., например, Tremp et al. (1976) "Human Brest Cancer In Culture", Recent Results Cancer Res. 33-41); B-лимфоциты; клетки, происходящие из лимфомы Беркитта, например, клетки Raji с регистрационным номером ATCC CCL-86 (см., например, Epstein et al. (1965) "Characteristics And Mode Of Growth Of Tissue Culture Strain (EBl) Of Human Lymphoblasts From Burkitt's Lymphoma", J. Natl. Cancer Inst. 34:231-240), и клетки Daudi с регистрационным номером ATCC CCL-213 (см., например, Klein et al. (1968) "Surface IgM-Kappa Specificity On A Burkitt Lymphoma Cell In Vivo And In Derived Culture Lines", Cancer Res. 28:1300-1310). Клетки-мишени необходимо выбирать по участку связывания антигена в молекуле диантитела, участвующей в анализе.
Анализ ADCC основан на способности NK-клеток опосредовать гибель клеток через апоптотический путь. NK-клетки опосредуют гибель клеток частично путем распознавания посредством FcγRIIIA Fc-домена IgG, связанного с антигеном на клеточной поверхности. Анализы ADCC, используемые в соответствии со способами по изобретению, могут представлять собой анализы на основе радиоактивности или флуоресценции. Анализ ADCC, использованный для охарактеризации молекул по изобретению, содержащих варианты Fc-областей, включает мечение клеток-мишеней, например, клеток SK-BR-3, MCF-7, OVC AR3, Raji, Daudi, опсонизацию клеток-мишеней антителом, которое распознает рецептор клеточной поверхности на клетке-мишени через ее антигенсвязывающий центр; комбинирование меченых опсонизированных клеток-мишеней и эффекторных клеток в соответствующем соотношении, которое можно определять общепринятым экспериментированием; сбор клеток; детекцию метки в супернатанте лизированных клеток-мишеней с использованием подходящей схемы детекции, исходя из используемой метки. Клетки-мишени можно метить либо радиоактивной меткой, либо флуоресцентной меткой, с использованием стандартных способов, известных в данной области. Например, метки включают, но не ограничиваются ими, [51Cr]Na2CrO4; и ацетоксиметиловый эфир усиливающего флуоресценцию лиганда, 2,2':6',2"-терпиридин-6-6"-дикарбоксилата (TDA).
В конкретном предпочтительном варианте осуществления для измерения активности ADCC против клеток-мишеней, меченных ацетоксиметиловым эфиром усиливающего флуоресценцию лиганда, 2,2':6',2"-терпиридин-6-6"-дикарбоксилата (TDA), используют флуориметрический анализ с временным разрешением. Такие флуориметрические анализы известны в данной области, см., например, Blomberg et al. (1996) "Time-Resolved Fluorometric Assay For Natural Killer Activity Using Target Cells Labelled With A Fluorescence Enhancing Ligand", Journal of Immunological Methods, 193:199-206; которая включена в настоящий документ в качестве ссылки в полном объеме. В кратком изложении, клетки-мишени метят проникающим через мембрану ацетоксиметиловым диэфиром TDA (бис(ацетоксиметил)-2,2':6',2"-терпиридин-6-6"-дикарбоксилат, (BATDA)), который быстро диффундирует через клеточную мембрану жизнеспособных клеток. Внутриклеточные эстеразы отщепляют сложноэфирные группы, и регенерированная не проходящая через мембрану молекула TDA улавливается внутрь клетки. После инкубации эффекторных клеток и клеток-мишеней, например, в течение по меньшей мере двух часов, вплоть до 3,5 часов, при 37°C, в атмосфере 5% CO2, TDA, высвобожденный из лизированных клеток-мишеней, хелатируют посредством Eu3+, и флуоресценцию образовавшихся хелатов европий-TDA количественно определяют на флуориметре с временным разрешением (например, Victor 1420, Perkin Elmer/Wallace).
В другом конкретном варианте осуществления анализ ADCC, используемый для охарактеризации молекул по изобретению, содержащих множество участков связывания эпитопа и, необязательно, Fc-домен (или его часть), включает следующие стадии: предпочтительно 4-5×106 клеток-мишеней (например, клеток SK-BR-3, MCF-7, OVCAR3, Raji) метят бис(ацетоксиметил)-2,2':6',2"-терпиридин-6-6"-дикарбоксилатом (реагент DELFIA BATDA, Perkin Elmer/Wallac). Для оптимальной эффективности мечения количество клеток-мишеней, используемых в анализе ADCC, предпочтительно не должно превышать 5×106. Реагент BATDA добавляют к клеткам и смесь инкубируют при 37°С предпочтительно в 5% CO2 в течение по меньшей мере 30 минут. Затем клетки промывают физиологическим буфером, например, PBS с 0,125 мМ сульфинпиразолом, и средой, содержащей 0,125 мМ сульфинпиразол. Затем меченые клетки-мишени опсонизируют (покрывают) молекулой по изобретению, содержащей связывающий эпитоп домен, специфичный к FcγRIIA и, необязательно, Fc-домен (или его часть). В предпочтительных вариантах осуществления молекула, используемая в анализе ADCC, также является специфичной к рецептору клеточной поверхности, опухолевому антигену или антигену злокачественной опухоли. Молекула диантитела по изобретению может специфично связывать любой антиген злокачественной опухоли или опухолевый антиген, такой как антигены, приведенные в разделе 5.6.1. Клетки-мишени в анализе ADCC выбирают согласно участкам связывания эпитопа, встроенным в диантитело по изобретению, так что диантитело специфично связывает рецептор клеточной поверхности клетки-мишени.
Клетки-мишени добавляют к эффекторным клеткам, например PBMC, для обеспечения соотношений эффектор:мишень приблизительно 1:1, 10:1, 30:1, 50:1, 75:1 или 100:1. Эффекторные клетки и клетки-мишени инкубируют в течение по меньшей мере двух часов, вплоть до 3,5 часов, при 37°C, в 5% CO2. Клеточные супернатанты собирают и добавляют к раствору европия в кислоте (например, раствор европия в DELFIA, Perkin Elmer/Wallac). Флуоресценцию образовавшихся хелатов европий-TDA количественно определяют на флуориметре с временным разрешением (например, Victor 1420, Perkin Elmer/Wallac). Максимальное высвобождение (MR) и спонтанное высвобождение (SR) определяют путем инкубации клеток-мишеней с 1% TX-100 и средой отдельно, соответственно. Антителозависимую клеточную цитотоксичность (AICC) измеряют путем инкубации клеток-мишеней и эффекторных клеток в отсутствии тестируемой молекулы, например, диантитела по изобретению. Каждый анализ предпочтительно проводят в трех экземплярах. Средний процентный специфичный лизис вычисляют, как: экспериментальное высвобождение (ADCC) - AICC)/(MR-SR)×100.
Изобретение охватывает анализы, известные в данной области и проиллюстрированные в настоящем документе, для охарактеризации связывания C1q и опосредования молекулами по изобретению, содержащими Fc-домены (или их части), комплементзависимой цитотоксичности (CDC). Для определения связывания C1q можно проводить ELISA связывания C1q. Иллюстративный анализ может включать следующее: планшеты для анализа можно покрывать в течение ночи при 4°С полипептидом, содержащим молекулу по изобретению, или исходным полипептидом (контролем) в буфере для покрытия. Затем планшеты можно промывать и блокировать. После промывания в каждую лунку можно добавлять аликвоту C1q человека и инкубировать в течение 2 ч при комнатной температуре. После последующего промывания в каждую лунку можно добавлять 100 мкл конъюгированного с пероксидазой антитела овцы против C1q-компонента комплемента и инкубировать в течение 1 часа при комнатной температуре. Планшет можно вновь промывать буфером для промывания и в каждую лунку можно добавлять 100 мкл буфера для субстрата, содержащего OPD (O-фенилендиаминдигидрохлорид (Sigma)). Реакции окисления, наблюдаемой по появлению желтого цвета, можно позволять протекать в течение 30 минут, и ее можно останавливать добавлением 100 мкл 4,5 NH2SO4. Затем можно считывать поглощение при (492-405) нм.
Для оценки активации комплемента можно проводить анализ комплементзависимой цитотоксичности (CDC), например, как описано в Gazzano-Santoro et al. (1997) "A Non-Radioactive Complement-Dependant Cytotoxicity Assay For Anti-CD20 Monoclonal Antibody", J. Immunol. Methods 202:163-171, которая включена в настоящий документ в качестве ссылки в полном объеме. В кратком изложении, различные концентрации молекулы, содержащей (вариантный) Fc-домен (или его часть) и комплемент человека, можно разбавлять в буфере. Клетки, которые экспрессируют антиген, с которым связывается молекула диантитела, можно разбавлять до плотности приблизительно 1×106 клеток/мл. Смеси молекул диантител, содержащих (вариантный) Fc-домен (или его часть), разбавленный комплемент человека и клетки, экспрессирующие антиген, можно добавлять в 96-луночный планшет для культивированная тканей с плоским дном и им можно позволять инкубироваться в течение 2 ч при 37°C и 5% CO2 для способствования опосредуемому комплементом лизису клеток. Затем в каждую лунку можно добавлять 50 мкл Alamar Blue (Accumed International) и инкубировать в течение ночи при 37°C. Поглощение измеряют с использованием флуориметра для 96-луночных планшетов с возбуждением при 530 нм и испусканием при 590 нм. Результаты можно выражать в относительных единицах флуоресценции (RFU). Концентрации образца можно вычислять из стандартной кривой, и для представляющего интерес варианта описывают процентную активность по сравнению с невариантной молекулой, т.е. молекулой, не содержащей Fc-домен или содержащей невариантный Fc-домен.
5.4.3 ДРУГИЕ АНАЛИЗЫ
Молекулы по изобретению, содержащие связывающий домен с множеством эпитопов и, необязательно, Fc-домен, можно анализировать с использованием любых анализов на основе поверхностного плазмонного резонанса, известных в данной области, для охарактеризации кинетических параметров связывания антиген-связывающий домен или Fc-FcγR. Для настоящего изобретения можно использовать любое коммерчески доступное устройство SPR, включая, но не ограничиваясь ими, устройства BIAcore, доступные от Biacore AB (Uppsala, Швеция); устройства IAsys, доступные от Affinity Sensors (Franklin, MA.); систему IBIS, доступную от Windsor Scientific Limited (Berks, Великобритания), системы SPR-CELLIA, доступные от Nippon Laser and Electronics Lab (Hokkaido, Япония), и SPR Detector Spreeta, доступное от Texas Instruments (Dallas, TX). Для обзора технологии на основе SPR см. Mullet et al. (2000) "Surface Plasmon Resonance-Based Immunoassays", Methods 22:77-91; Dong et al. (2002) "Some new aspects in biosensors", Reviews in Mol. Biotech. 82:303-23; Fivash et al. (1998) "BIAcore For Macromolecular Interaction", Current Opinion in Biotechnology 9:97-101; Rich et al. (2000) "Advances In Surface Plasmon Resonance Biosensor Analysis", Current Opinion in Biotechnology 11:54-61; которые все включены в настоящий документ в качестве ссылок в полном объеме. Кроме того, в способах по изобретению предусматриваются любые из устройств SPR и способов на основе SPR для определения белок-белковых взаимодействий, описанных в патентах США №№ 6373577; 6289286; 5322798; 5341215; 6268125, которые все включены в настоящий документ в качестве ссылок в полном объеме.
В кратком изложении, анализы на основе SPR вовлекают иммобилизацию члена связывающейся пары на поверхности и мониторинг его взаимодействия с другим членом связывающей пары в растворе в реальном времени. SPR основан на измерении изменения индекса рефракции растворителя вблизи поверхности, которое происходит при образовании или диссоциации комплекса. Поверхность, на которой происходит иммобилизация, представляет собой сенсорный чип, который находится в основе технологии SPR; он состоит из стеклянной поверхности, покрытой тонким слоем золота, и образует основу для ряда специализированных поверхностей, предназначенных для оптимизации связывания молекулы с поверхностью. Множество сенсорных чипов доступны коммерчески, особенно от компаний, приведенных выше, и все из них можно использовать в способах по изобретению. Примеры сенсорных чипов включают чипы, доступные от BIAcore AB, Inc., например, Sensor Chip CM5, SA, NTA и HPA. Молекулу по изобретению можно иммобилизовывать на поверхности сенсорного чипа с использованием любых способов иммобилизации и химических групп, известных в данной области, включая, но не ограничиваясь ими, прямое ковалентное связывание через группы аминов, прямое ковалентное связывание через сульфгидрильные группы, связывание биотина с покрытой авидином поверхностью, связывание альдегида с углеводными группами и связывание через гистидиновую метку с чипами NTA.
В некоторых вариантах осуществления кинетические параметры связывания молекул по изобретению, содержащих множество участков связывания эпитопа и, необязательно, и Fc-домен, с антигеном или FcγR можно определять с использованием устройства BIAcore (например, устройство BIAcore 1000, BIAcore Inc., Piscataway, NJ). Как рассмотрено выше, см. раздел 5.4.1, любой FcγR можно использовать для оценки связывания молекул по изобретению, либо где по меньшей мере один участок связывания эпитопа в молекуле диантитела иммуноспецифично распознает FcγR, и/либо где молекула диантитела содержит Fc-домен (или его часть). В конкретном варианте осуществления FcγR представляет собой FcγRIIIA, предпочтительно растворимый мономерный FcγRIIIA. Например, в одном из вариантов осуществления растворимый мономерный FcγRIIIA представляет собой внеклеточную область FcγRIIIA, связанную с последовательностью линкер-AVITAG (см., предварительную заявку США № 60/439498, поданную 9 января 2003 года, и предварительную заявку США № 60/456041, поданную 19 марта 2003 года, которые включены в настоящий документ в качестве ссылок в полном объеме). В другом конкретном варианте осуществления FcγR представляет собой FcγRIIB, предпочтительно растворимый димерный FcγRIIB. Например, в одном из вариантов осуществления растворимый димерный белок FcγRIIB получают в соответствии со способами, описанными в предварительной заявке США № 60/439709, поданной 13 января 2003 года, которая включена в настоящий документ в качестве ссылки в полном объеме.
Для всех иммунологических анализов распознавание/связывание FcγR молекулой по изобретению может осуществляться множеством доменов: в определенных вариантах осуществления молекулы по изобретению иммуноспецифично распознают FcγR через один из множества связывающих эпитоп доменов; в других вариантах осуществления, где молекула по изобретению содержит Fc-домен (или его часть), молекула диантитела может иммуноспецифично распознавать FcγR через взаимодействия Fc-FcγR; в следующих вариантах осуществления, где молекула по изобретению содержит как Fc-домен (или его часть), так и участок связывания эпитопа, который иммуноспецифично распознает FcγR, молекула диантитела может распознавать FcγR через один или оба из связывающего эпитоп домена и Fc-домена (или его части). Иллюстративный анализ для определения кинетических параметров молекулы, содержащей множество связывающих эпитоп доменов и, необязательно, и Fc-домен (или его часть) с антигеном и/или FcγR, с использованием устройства BIAcore включает следующее: первый антиген иммобилизуют на одной из четырех проточных ячеек поверхности сенсорного чипа, предпочтительно химической реакцией связывания аминов, так чтобы приблизительно 5000 единиц ответа (RU) указанного антигена было иммобилизовано на поверхности. После подготовки поверхности молекулы по изобретению, которые иммуноспецифично распознают указанный первый антиген, пропускают над поверхностью, предпочтительно путем инжекций в течение одной минуты раствора 20 мкг/мл при скорости потока 5 мкл/мин. Уровни молекул по изобретению, связанных с поверхностью на этой стадии, как правило, находятся в диапазоне от 400 до 700 RU. Далее серии разведений второго антигена (например, FcγR) или рецептора FcγR в буфере HBS-P (20 мМ HEPES, 150 мМ NaCl, 3 мМ ЭДТА, pH 7,5) инжектируют на поверхность при 100 мкл/мин. Регенерацию молекул между разведениями отличающегося второго антигена или рецептора предпочтительно проводят путем однократных инъекций в течение 5 секунд 100 мМ NaHCO3, pH 9,4; 3 M NaCl. В способе по изобретению предусмотрен любой способ регенерации, известный в данной области.
После сбора всех данных полученные кривые связывания подвергают глобальной аппроксимации с использованием компьютерных алгоритмов, предоставляемых производителем устройства SPR, например, BIAcore, Inc. (Piscataway, NJ). Эти алгоритмы вычисляют как Kon, так и Koff, из которых устанавливают кажущуюся константу равновесного связывания Kd в качестве соотношения двух констант скоростей (т.е. Koff/Kon). Более подробное описание того, как могут быть установлены отдельные константы, может быть найдено в BIAevaluaion Software Handbook (BIAcore, Inc., Piscataway, NJ). Анализ полученных данных можно проводить с использованием любого способа, известного в данной области. Для обзора различных способов интерпретации полученных кинетических данных см. Myszka (1997) "Kinetic Analysis Of Macromolecular Interactions Using Surface Plasmon Resonance Biosensors", Current Opinion in Biotechnology 8:50-7; Fisher et al. (1994) "Surface Plasmon Resonance Based Methods For Measuring The Kinetics And Binding Affinities Of Biomolecular Interactions", Current Opinion in Biotechnology 5:389-95; O'Shannessy (1994) "Determination Of Kinetic Rate And Equilibrium Binding Constants For Macromolecular Interactions: A Critique Of The Surface Plasmon Resonance Literature", Current Opinion in Biotechnology, 5:65-71; Chaiken et al. (1992) "Analysis Of Macromolecular Interactions Using Immobilized Ligands", Analytical Biochemistry, 201:197-210; Morton et al. (1995) "Interpreting Complex Binding Kinetics From Optical Biosensors: A Comparison Of Analysis By Linearization, The Integrated Rate Equation, And Numerical Integration", Analytical Biochemistry 227:176-85; O'Shannessy et al., 1996, Analytical Biochemistry 236:275-83; которые все включены в настоящий документ в качестве ссылок в полном объеме.
В предпочтительных вариантах осуществления кинетические параметры, определенные с использованием анализа SPR, например, BIAcore, можно использовать в качестве прогностического показателя того, как молекула по изобретению будет функционировать в функциональном анализе, например ADCC. Иллюстративный способ предсказания эффективности молекулы по изобретению на основе кинетических параметров, полученных из анализа SPR, может включать следующее: определение величин Koff для связывания молекулы по изобретению с FcγRIIIA и FcγRIIB (через домен связывания эпитопа и/или Fc-домен (или его часть)); построение графика (1) Koff (wt)/Koff (mut) для FcγRIIIA; (2) Koff (mut)/Koff (wt) для FcγRIIB против данных ADCC. Числа, превышающие единицу, демонстрируют сниженную скорость диссоциации для FcγRIIIA и увеличенную скорость диссоциации для FcγRIIB относительно дикого типа; и наличие усиленной функции ADCC.
5.5 СПОСОБЫ ПОЛУЧЕНИЯ МОЛЕКУЛ ДИАНТИТЕЛ ПО ИЗОБРЕТЕНИЮ
Молекулы диантител по настоящему изобретению можно получать с использованием различных способов, хорошо известных в данной области, включая синтез белка de novo и рекомбинантную экспрессию нуклеиновых кислот, кодирующих связывающие белки. Желаемые последовательности нуклеиновых кислот можно получать рекомбинантными способами (например, мутагенезом посредством ПЦР ранее полученного варианта желаемого полинуклеотида) или твердофазным синтезом ДНК. Как правило, используют рекомбинантные способы экспрессии. В одном аспекте изобретение относится к полинуклеотиду, который содержит последовательность, кодирующую VH и/или VL CD16A; в другом аспекте изобретение относится к полинуклеотиду, который содержит последовательность, кодирующую VH и/или VL CD32B. Вследствие вырожденности генетического кода каждую аминокислотную последовательность иммуноглобулина кодируют различные последовательности нуклеиновых кислот, и настоящее изобретение включает все нуклеиновые кислоты, кодирующие связывающие белки, описанные в настоящем документе.
5.5.1 ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ МОЛЕКУЛЫ ПО ИЗОБРЕТЕНИЮ
Также настоящее изобретение включает полинуклеотиды, которые кодируют молекулы по изобретению, включая полипептиды и антитела. Полинуклеотиды, кодирующие молекулы по изобретению, можно получать, и нуклеотидную последовательность полинуклеотидов можно определять, любым способом, известным в данной области.
После определения нуклеотидной последовательности молекул, которые идентифицированы способами по изобретению, нуклеотидную последовательность можно изменять с использованием способов, хорошо известных в данной области, например, способов рекомбинантных ДНК, сайт-направленного мутагенеза, ПЦР и т.д. (см., например, способы, описанные в Sambrook et al., 2001, Molecular Cloning, A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY; и Ausubel et al., eds., 1998, Current Protocols in Molecular Biology, John Wiley & Sons, NY, которые обе включены в настоящий документ в качестве ссылок в полном объеме), для получения, например, антител, имеющих отличающуюся аминокислотную последовательность, например путем создания замен, делеций и/или вставок аминокислот.
В одном из вариантов осуществления для клонирования нуклеиновых кислот, кодирующих молекулы по изобретению можно проводить скрининг библиотек нуклеиновых кислот человека или любых других библиотек, доступных в данной области, стандартными способами, известными в данной области.
5.5.2 РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ МОЛЕКУЛ ПО ИЗОБРЕТЕНИЮ
После получения последовательности нуклеиновой кислоты, кодирующей молекулы по изобретению (т.е. антитела), можно получать вектор для продукции молекул с помощью технологии рекомбинантных ДНК с использованием способов, хорошо известных в данной области. Для конструирования экспрессирующих векторов, содержащих кодирующие последовательности для молекул по изобретению и подходящие сигналы контроля транскрипции и трансляции, можно использовать способы, которые хорошо известны специалистам в данной области. Эти способы включают, например, технологии рекомбинантных ДНК in vitro, способы синтеза и генетическую рекомбинацию in vivo. (См., например, способы, описанные в Sambrook et al., 1990, Molecular Cloning, A Laboratory Manual, 2d Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY и Ausubel et al. eds., 1998, Current Protocols in Molecular Biology, John Wiley & Sons, NY).
Экспрессирующий вектор, содержащий нуклеотидную последовательность молекулы, идентифицированной способами по изобретению, можно переносить в клетку-хозяина общепринятыми способами (например, электропорация, липосомальная трансфекция и преципитация с фосфатом кальция), и трансфицированные клетки затем культивируют общепринятыми способами для получения молекул по изобретению. В конкретных вариантах осуществления экспрессия молекул по изобретению регулируется конститутивным, индуцибельным или тканеспецифическим промотором.
Клетки-хозяева, используемые для экспрессии молекул, идентифицированных способами по изобретению, могут представлять собой либо бактериальные клетки, такие как Escherichia coli, либо, предпочтительно, эукариотические клетки, особенно для экспрессии целой рекомбинантной молекулы иммуноглобулина. В частности, эффективной экспрессирующей системой для иммуноглобулинов являются клетки млекопитающих, такие как клетки яичника китайского хомячка (CHO), вместе с вектором, таким как основной предранний промоторный элемент гена из цитомегаловируса человека (Foecking et al. (1986) "Powerful And Versatile Enhancer-Promoter Unit For Mammalian Expression vectors", Gene 45:101-106; Cockett et al. (1990) "High Level Expression Of Tissue Inhibitor Of Metalloproteinases In Chinese Hamster Ovary Cells Using Glutamine Synthetase Gene Amplification", Biotechnology 8:662-667).
Для экспрессии молекул, идентифицированных способами по изобретению, можно использовать различные системы хозяин-экспрессирующий вектор. Такие системы хозяин-экспрессирующий вектор представляют собой носители, с помощью которых можно продуцировать и впоследствии очищать кодирующие последовательности для молекул по изобретению, а также они представляют собой клетки, которые могут при трансформации или трансфекции соответствующими нуклеотидными кодирующими последовательностями экспрессировать молекулы по изобретению in situ. Они включают, но не ограничиваются ими, микроорганизмы, такие как бактерии (например, E. coli и B. subtilis), трансформированные экспрессирующими векторами на основе рекомбинантной ДНК бактериофага, плазмидной ДНК или космидной ДНК, содержащими кодирующие последовательности для молекул, идентифицированных способами по изобретению; дрожжи (например, Saccharomyces pichia), трансформированные рекомбинантными дрожжевыми экспрессирующими векторами, содержащими последовательности, кодирующие молекулы, идентифицированные способами по изобретению; системы клеток насекомых, инфицированных рекомбинантными вирусными экспрессирующими векторами (например, бакуловирусом), содержащими последовательности, кодирующие молекулы, идентифицированные способами по изобретению; системы растительных клеток, инфицированных рекомбинантными вирусными экспрессирующими векторами (например, вирусом мозаики цветной капусты (CaMV) и вирусом табачной мозаики (TMV)) или трансформированных рекомбинантными плазмидными экспрессирующими векторами (например, плазмидой Ti), содержащими последовательности, кодирующие молекулы, идентифицированные способами по изобретению; или системы клеток млекопитающих (например, клетки COS, CHO, BHK, 293, 293T, 3T3, лимфатические клетки (см. U.S. 5807715), клетки Per C.6 (клетки сетчатки человека, разработанные Crucell), содержащие рекомбинантные экспрессирующие конструкции, содержащие промоторы, происходящие из генома клеток млекопитающих (например, промотор металлотионеина) или из вирусов млекопитающих (например, позднего промотора аденовируса; промотора 7,5-K вируса осповакцины).
В бактериальных системах можно преимущественно выбрать ряд экспрессирующих векторов в зависимости от предполагаемого применения для экспрессируемой молекулы. Например, когда предстоит получить большое количество такого белка для получения фармацевтических композиций антитела, могут быть желательными векторы, которые направляют экспрессию высоких уровней слитых белковых продуктов, которые легко очищать. Такие векторы включают, но не ограничиваются ими, экспрессирующий вектор E. coli pUR278 (Ruther et al. (1983) "Easy Identification Of cDNA Clones", EMBO J. 2:1791-1794), в котором последовательность, кодирующая антитело, может быть индивидуально лигирована в вектор в рамке считывания с кодирующей областью для lac Z, так чтобы продуцировался слитый белок; векторы pIN (Inouye et al. (1985) "Up-Promoter Mutations In The lpp Gene Of Escherichia Coli", Nucleic Acids Res. 13:3101-3110; Van Heeke et al. (1989) "Expression Of Human Asparagine Synthetase In Escherichia Coli", J. Biol. Chem. 24:5503-5509) и т.п. Векторы pGEX также можно использовать для экспрессии чужеродных полипептидов в качестве слитых белков с глутатион-S-трансферазой (GST). Как правило, такие слитые белки являются растворимыми и их можно легко очищать из лизированных клеток путем адсорбции и связывания с гранулами матрицы из глутатион-агарозы с последующим элюированием в присутствии свободного глутатиона. Векторы pGEX конструируют так, чтобы они включали участки расщепления протеазой, тромбином или фактором Xa, чтобы клонированный намеченный продукт гена можно было освобождать от группы GST.
В системе насекомых в качестве вектора для экспрессии чужеродных генов используют вирус ядерного полиэдроза Autographa californica (AcNPV). Этот вирус растет в клетках Spodoptera frugiperda. Последовательность, кодирующую антитело, можно индивидуально клонировать в не являющиеся необходимыми области (например, ген полиэдрина) вируса и помещать под контроль промотора AcNPV (например, промотора полиэдрина).
В клетках-хозяевах млекопитающих можно использовать множество экспрессирующих систем на основе вирусов. В случаях, когда в качестве экспрессирующего вектора используют аденовирус, представляющую интерес кодирующую антитело последовательность можно лигировать с аденовирусным комплексом контроля транскрипции/трансляции, например, поздней промоторной и тройной лидерной последовательностью. Затем этот химерный ген можно встраивать в геном аденовируса посредством рекомбинации in vitro или in vivo. Встраивание в не являющуюся необходимой область генома вируса (например, области E1 или E3) приводит к тому, что рекомбинантный вирус является жизнеспособным и способен экспрессировать молекулу иммуноглобулина в инфицированных хозяевах (например, см. Logan et al. (1984) "Adenovirus Tripartite Leader Sequence Enhances Translation Of mRNAs Late After Infection", Proc. Natl. Acad. Sci. USA 81:3655-3659). Также для эффективной трансляции встроенных кодирующих антитело последовательностей могут требоваться определенные сигналы инициации. Эти сигналы включают кодон инициации ATG и соседние последовательности. Более того, кодон инициации должен находиться в фазе с рамкой считывания желаемой кодирующей последовательности для обеспечения трансляции всей вставки. Эти экзогенные сигналы контроля трансляции и кодоны инициации могут быть различного происхождения, как природного, так и синтетического. Эффективность экспрессии можно усиливать путем включения подходящих энхансерных транскрипционных элементов, терминаторов транскрипции и т.д. (см. Bitter et al. (1987) "Expression And Secretion Vectors For Yeast", Methods in Enzymol. 153:516-544).
Кроме того, можно выбрать клетку-хозяина, которая модулирует экспрессию встроенных последовательностей или модифицирует и процессирует продукт гена конкретным желаемым образом. Такие модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут быть важны для функционирования белка. Например, в определенных вариантах осуществления полипептиды, содержащие молекулу диантитела по изобретению, можно экспрессировать в качестве единого продукта гена (например, в качестве одной полипептидной цепи, т.е. в качестве полибелкового предшественника), требующего протеолитического расщепления с помощью природных или рекомбинантных клеточных механизмов с образованием отдельных полипептидов молекул диантител по изобретению. Таким образом, изобретение охватывает конструирование последовательности нуклеиновой кислоты, чтобы она кодировала полибелковую молекулу-предшественника, содержащую полипептиды по изобретению, которая включает кодирующие последовательности, способные обеспечивать посттрансляционное расщепление указанного полибелкового предшественника. Посттрансляционное расщепление полибелкового предшественника дает полипептиды по изобретению. Посттрансляционное расщепление молекулы-предшественника, содержащей полипептиды по изобретению, может происходить in vivo (т.е. в клетке-хозяине посредством природных или рекомбинантных клеточных систем/механизмов, например, расщепления фурином в соответствующем участке), или оно может происходить in vitro (например, инкубация указанной полипептидной цепи в композиции, содержащей протеазы или пептидазы с известной активностью, и/или в композиции, содержащей условия или реагенты, известные тем, что они содействуют желаемому протеолитическому действию). Очистка и модификация рекомбинантных белков хорошо известны в данной области, так что конструкция полибелкового предшественника может включать ряд вариантов осуществления, хорошо понятных квалифицированному специалисту. Для описанной модификации молекулы предшественника можно использовать любые протеазы или пептидазы, известные в данной области, например, тромбин (который распознает аминокислотную последовательность LVPRˆGS (SEQ ID NO: 89)), или фактор Xa (который распознает аминокислотную последовательность I(E/D)GRˆ (SEQ ID NO: 90) (Nagai et al. (1985) "Oxigen Binding Properties Of Human Mutant Hemoglobins Synthesized In Escherichia Coli", Proc. Nat. Acad. Sci. USA 82:7252-7255, и обзор в Jenny et al. (2003) "A Critical Review Of Methods For Cleavage Of Fused Proteins With Thrombin And Factor Xa", Protein Expr. Purif. 31:1-11, которые все включены в настоящий документ в качестве ссылки в полном объеме)), энтерокиназу (которая распознает аминокислотную последовательность DDDDK˄ (SEQ ID NO: 91) (Collins-Racie et al. (1995) "Production Of Recombinant Bovine Enterokinase Catalytic Subunit In Escherichia Coli Using The Novel Secretory Fusion Partner DsbA", Biotechnology 13:982-987, включенная в настоящий документ в качестве ссылки в полном объеме)), фурин (который распознает аминокислотную последовательность RXXRˆ, предпочтительно RX(K/R)Rˆ (SEQ ID NO: 92 и SEQ ID NO: 93, соответственно) (дополнительный R в положении P6, по-видимому, усиливает расщепление)), и AcTEV (который распознает аминокислотную последовательность ENLYFQˆG (SEQ ID NO: 94) (Parks et al. (1994) "Release Of Proteins And Peptides From Fused Proteins Using A Recombinant Plant Virus Protease", Anal. Biochem. 216:413-417, включенная в настоящий документ в качестве ссылки в полном объеме)) и протеазу C3 вируса ящура. См. например, раздел 6.4, выше.
Различные клетки-хозяева обладают характерными и специфичными механизмами посттрансляционного процессинга и модификации белков и продуктов генов. Для обеспечения правильной модификации и процессинга чужеродного экспрессированного белка можно выбирать подходящие клеточные линии или системы хозяев. Для этого можно использовать эукариотические клетки-хозяева, которые обладают клеточным аппаратом для надлежащего процессинга первичного транскрипта, гликозилирования и фосфорилирования продукта гена. Такие клетки-хозяева млекопитающих включают, но не ограничиваются ими, CHO, VERY, BHK, HeLa, COS, MDCK, 293, 293T, 3T3, WI38, BT483, Hs578T, HTB2, BT20 и T47D, CRL7030 и Hs578Bst.
Для длительной продукции рекомбинантных белков с высоким выходом предпочтительной является стабильная экспрессия. Например, можно конструировать клеточные линии, которые стабильно экспрессируют антитело по изобретению. Вместо использования экспрессирующих векторов, которые содержат вирусные ориджины репликации, клетки-хозяева можно трансформировать ДНК, контролируемой соответствующими элементами контроля экспрессии (например, промоторными, энхансерными последовательностями, терминаторами транскрипции, участками полиаденилирования и т.д.) и селективным маркером. После встраивания чужеродной ДНК сконструированным клеткам можно позволять расти в течение 1-2 суток в обогащенной среде, а затем их можно переключать на селективную среду. Селективный маркер в рекомбинантной плазмиде придает устойчивость при селекции и позволяет клеткам стабильно встраивать плазмиду в их хромосомы и расти с образованием очагов, которые, в свою очередь, можно клонировать и размножать в клеточные линии. Этот способ можно преимущественно использовать для конструирования клеточных линий, которые экспрессируют антитела по изобретению. Такие сконструированные клеточные линии могут быть особенно пригодны при скрининге и оценке соединений, которые прямо или непрямо взаимодействуют с молекулами по изобретению.
Можно использовать множество систем селекции, включая, но не ограничиваясь ими, гены тимидинкиназы вируса простого герпеса (Wigler et al. (1977) "Transfer Of Purified Herpes Virus Thymidine Kinase Gene To Cultured Mouse Cells", Cell 11:223-232), гипоксантингуанинфосфорибозилтрансферазы (Szybalska et al. (1992) "Use Of The HPRT Gene And The HAT Selection Technique In DNA-Mediated Transformation Of Mammalian Cells: First Steps Toward Developing Hybridoma Techniques And Gene Therapy", Bioessays 14:495-500), и аденинфосфорибозилтрансферазы (Lowy et al. (1980) "Isolation Of Transforming DNA: Cloning The Hamster aprt Gene", Cell 22:817-823), которые можно использовать в клетках tk-, hgprt- или aprt-, соответственно. Также в качестве основы для селекции можно использовать устойчивость к антиметаболиту, обеспечиваемую следующими генами: dhfr, который придает устойчивость к метотрексату (Wigler et al. (1980) "Transformation Of Mammalian Cells With An Amplifiable Dominant-Acting Gene", Proc. Natl. Acad. Sci. USA 77:3567-3570; O'Hare et al. (1981) "Transformation Of Mouse Fibroblasts To Methotrexate Resistance By A Recombinant Plasmid Expressing A Procaryotic Dihydrofolate Reductase", Proc. Natl. Acad. Sci. USA 78:1527-1531); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan et al. (1981) "Selection For Animal Cells That Express The Escherichia coli Gene Coding For Xanthine-Guanine Phosphoribosyltransferase", Proc. Natl. Acad. Sci. USA 78:2072-2076); neo, который придает устойчивость к аминогликозиду G-418 (Tolstoshev (1993) "Gene Therapy, Concepts, Current Trials And Future Directions", Ann. Rev. Pharmacol. Toxicol. 32:573-596; Mulligan (1993) "The Basic Science Of Gene Therapy", Science 260:926-932; и Morgan et al. (1993) "Human Gene Therapy", Ann. Rev. Biochem. 62:191-217), и hygro, который придает устойчивость к гигромицину (Santerre et al. (1984) "Expression Of Procaryotic Genes For Hygromycin B And G418 Resistance As Dominant-Selection Markers In Mouse L Cells", Gene 30:147-156). Способы, широко известные в области технологии рекомбинантных ДНК, которые можно использовать, описаны в Ausubel et al. (eds.), 1993, Current Protocols in Molecular Biology, John Wiley & Sons, NY; Kriegler, 1990, Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NY; и в главах 12 и 13, Dracopoli et al. (eds), 1994, Current Protocols in Human Genetics, John Wiley & Sons, NY.; Colberre-Garapin et al. (1981) "A New Dominant Hybrid Selective Marker For Higher Eucaryotic Cells", J. Mol. Biol. 150:1-14.
Уровни экспрессии молекулы по изобретению можно увеличивать путем амплификации вектора (для обзора см. Bebbington and Hentschel, The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells in DNA cloning. Vol. 3 (Academic Press, New York, 1987)). Когда маркер в векторной системе, экспрессирующей антитело, является амплифицируемым, увеличение уровня ингибитора, присутствующего в культуре клеток-хозяев, приводит к увеличению количества копий маркерного гена. Поскольку амплифицированная область ассоциирована с нуклеотидной последовательностью полипептида молекулы диантитела, продукция полипептида также будет возрастать (Crouse et al. (1983) "Expression And Amplification Of Engineered Mouse Dihydrofolate Reductase Minigenes", Mol. Cell. Biol. 3:257-266).
Клетку-хозяина можно котрансфицировать двумя экспрессирующими векторами по изобретению, причем первый из векторов кодирует первый полипептид молекулы диантитела, а второй вектор кодирует второй полипептид молекулы диантитела. Эти два вектора могут содержать идентичные селективные маркеры, которые обеспечивают равную экспрессию обоих полипептидов. Альтернативно можно использовать один вектор, который кодирует оба полипептида. Кодирующие последовательности для полипептидов молекул по изобретению могут содержать кДНК или геномную ДНК.
После рекомбинантной экспрессии молекулы по изобретению (т.е. диантитела), ее можно очищать любым способом очистки полипептидов, полибелков или диантител, известным в данной области (например, аналогичным схемам очистки антител на основе антигенной селективности), например, посредством хроматографии (например, ионообменной, аффинной, в частности по аффинности к специфическому антигену (необязательно после селекции с белком A, где молекула диантитела содержит Fc-домен (или его часть)), и сортирующей по размеру колоночной хроматографии), центрифугирования, дифференциальной растворимости или любым другим стандартным способом очистки полипептидов, полибелков или диантител.
5.6 ПРОФИЛАКТИЧЕСКИЕ И ТЕРАПЕВТИЧЕСКИЕ СПОСОБЫ
Молекулы по изобретению особенно пригодны для лечения и/или профилактики заболевания, нарушения или инфекции, где является желаемой функция эффекторных клеток (например, ADCC), опосредуемая FcγR (например, злокачественная опухоль, инфекционное заболевание). Как рассмотрено выше, диантитела по изобретению могут проявлять антитело-подобную функциональность в отношении возникновения эффекторной функции, хотя молекула диантитела не содержит Fc-домен. Вследствие содержания по меньшей мере одного связывающего эпитоп домена, который иммуноспецифично распознает FcγR, молекула диантитела может проявлять связывание и активность FcγR, аналогичные взаимодействиям Fc-FcγR. Например, молекулы по изобретению могут связывать антиген клеточной поверхности и FcγR (например, FcγRIIIA) на иммунной эффекторной клетке (например, NK-клетке), стимулируя эффекторную функцию (например, ADCC, CDC, фагоцитоз, опсонизацию и т.д.) против указанной клетки.
В других вариантах осуществления молекула диантитела по изобретению содержит Fc-домен (или его часть). В таких вариантах осуществления Fc-домен может дополнительно содержать по меньшей мере одну модификацию аминокислот относительно Fc-домена дикого типа (или его части) и/или он может содержать домены из одного или нескольких изотипов IgG (например, IgG1, IgG2, IgG3 или IgG4). Молекулы по изобретению, содержащие варианты Fc-доменов, могут проявлять сообщенные им или измененные фенотипы относительно молекул, содержащих Fc-домен дикого типа, такие как измененная или сообщенная им активность эффекторной функции (например, при анализе в анализе на основе NK-клеток или макрофагов). В указанных вариантах осуществления молекулы по изобретению с сообщенной им или измененной активностью эффекторной функции пригодны для лечения и/или профилактики заболевания, нарушения или инфекции, где является желательной усиленная эффективность активности эффекторной функции. В определенных вариантах осуществления молекулы диантител по изобретению, содержащие Fc-домен (или его часть), опосредуют комплементзависимый каскад. Варианты Fc-доменов, идентифицированные как изменяющие эффекторную функцию, описаны в международной заявке WO04/063351, публикациях патентных заявок США 2005/0037000 и 2005/0064514, условных заявках США 60/626510, поданных 10 ноября 2004 года, 60/636663, поданной 15 декабря 2004 года, и 60/781564, поданной 10 марта 2006 года, и в патентных заявках США 11/271140, поданной 10 ноября 2005 года, и 11/305787, поданной 15 декабря 2005 года, совместных заявках авторов изобретения, которые все включены в настоящий документ в качестве ссылок в полном объеме.
Изобретение охватывает способы и композиции для лечения, профилактики или управления течением злокачественной опухоли у индивидуума, включающие введение индивидууму терапевтически эффективного количества одной или нескольких молекул, содержащих один или несколько участков связывания эпитопа, и необязательно, Fc-домен (или его часть), сконструированный в соответствии с изобретением, и эта молекула, кроме того, связывает антиген злокачественной опухоли. Молекулы по изобретению являются особенно пригодными для профилактики, ингибирования, снижения роста первичных опухолей или для их регрессии, метастазирования злокачественных клеток и инфекционных заболеваний. Без связи с конкретным механизмом действия, молекулы по изобретению опосредуют эффекторную функцию, приводящую к устранению опухоли, уменьшению опухоли или их комбинации. В альтернативных вариантах осуществления диантитела по изобретению опосредуют терапевтическую активность путем связывания антигенов клеточной поверхности и/или рецепторов и усиленного апоптоза или отрицательной регулирующей рост передачи сигнала.
Без связи с конкретным механизмом действия, молекулы диантител по изобретению проявляют усиленную терапевтическую эффективность относительно терапевтических антител, известных в данной области, частично, вследствие способности диантитела иммуноспецифично связывать клетку-мишень, которая экспрессирует конкретный антиген (например, FcγR) на сниженных уровнях, например, благодаря способности диантитела оставаться на клетке-мишени дольше вследствие увеличенной авидности взаимодействия диантитело-эпитоп.
Диантитела по изобретению с повышенной аффинностью и авидностью к антигенам (например, FcγR) особенно пригодны для лечения, профилактики или управления течением злокачественной опухоли или другого заболевания или нарушения у индивидуума, где FcγR экспрессируются на низких уровнях в популяциях клеток-мишеней. Как используют в настоящем документе, экспрессию FcγR в клетках определяют в единицах плотности таких молекул на клетку при измерении с использованием обычных способов, известных специалистам в данной области. Молекулы по изобретению, содержащие множество участков связывания эпитопа и, необязательно, и FcγR (или его часть), предпочтительно также обладают сообщенной им или усиленной авидностью и аффинностью и/или эффекторной функцией в клетках, которые экспрессируют антиген-мишень, например, антиген злокачественной опухоли, при плотности от 30000 до 20000 молекул/клетка, при плотности от 20000 до 10000 молекул/клетка, при плотности 10000 молекул/клетка или менее, при плотности 5000 молекул/клетка или менее, или при плотности 1000 молекул/клетка или менее. Молекулы по изобретению особенно применимы при лечении, профилактике или управлении течением заболевания или нарушения, такого как злокачественная опухоль, в субпопуляции, где антиген-мишень экспрессируется в популяции клеток-мишеней на низких уровнях.
Молекулы по изобретению также можно преимущественно использовать в комбинации с другими лекарственными средствами, известными в данной области, для лечения или профилактики заболеваний, таких как злокачественная опухоль, аутоиммунное заболевание, воспалительные нарушения и инфекционные заболевания. В конкретном варианте осуществления молекулы по изобретению можно использовать в комбинации с моноклональными или химерными антителами, лимфокинами или гемопоэтическими факторами роста (например, такими как IL-2, IL-3 и IL-7), которые, например, служат для увеличения количества или активности эффекторных клеток, которые взаимодействуют с молекулами и повышают иммунный ответ. Молекулы по изобретению также можно преимущественно использовать в комбинации с одним или несколькими лекарственными средствами, используемыми для лечения заболевания, нарушения или инфекции, например, такими как средства против злокачественной опухоли, противовоспалительные средства, противовирусные средства, например, как подробно описано в разделе 5.7.
5.6.1 ЗЛОКАЧЕСТВЕННЫЕ ОПУХОЛИ
Изобретение охватывает способы и композиции для лечения или профилактики злокачественной опухоли у индивидуума, включающие введение индивидууму терапевтически эффективного количества одной или нескольких молекул, содержащих множество связывающих эпитоп доменов. В некоторых вариантах осуществления изобретение охватывает способы и композиции для лечения или профилактики злокачественной опухоли у индивидуума с полиморфизмами FcγR, такого как индивидуумы, гомозиготные по аллелям FγRIIIA-158V или FcγRIIIA-158F. В некоторых вариантах осуществления изобретение охватывает конструирование по меньшей мере одного связывающего эпитоп домена молекулы диантитела для иммуноспецифического связывания FcγRIIIA (158F). В других вариантах осуществления изобретение охватывает конструирование по меньшей мере одного связывающего эпитоп домена молекулы диантитела для иммуноспецифического связывания FcγRIIIA (158V).
Эффективность стандартной терапии моноклональным антителом зависит от полиморфизма FcγR индивидуума (Cartron et al. (2002) "Therapeutic Activity Of Humanized Anti-CD20 Monoclonal Antibody And Polymorphism In IgG Fc Receptor FcRIIIa Gene", Blood 99:754-758; Weng et al. (2003) "Two Immunoglobulin G Fragment C Receptor Polymorphisms Independently Predict Response To Rituximab In Patients With Follicular Lymphoma", J Clin Oncol. 21(21):3940-3947, которые обе включены в настоящий документ в качестве ссылок в полном объеме). Эти рецепторы экспрессируются на поверхности эффекторных клеток и опосредуют ADCC. Высокоаффинные аллели низкоаффинных активирующих рецепторов увеличивают способность эффекторных клеток опосредовать ADCC. В противоположность принципу взаимодействий Fc-FcγR для достижения эффекторной функции, способы по изобретению охватывают конструирование молекул так, чтобы они иммуноспецифично распознавали низкоаффинные активирующие рецепторы, позволяя разработку молекул для конкретного полиморфизма. Альтернативно или дополнительно, молекулу по изобретению можно конструировать так, чтобы она содержала вариант Fc-домена, который проявляет усиленную аффинность к FcγR (относительно Fc-домена дикого типа) на эффекторных клетках. Сконструированные молекулы по изобретению обеспечивают лучшие реагенты для иммунотерапии пациентов, независимо от их полиморфизма FcγR.
Молекулы диантител, сконструированные в соответствии с изобретением, тестируют в отношении ADCC с использованием либо культивируемой клеточной линии, либо полученных от пациента клеток PMBC, для определения способности мутаций в Fc усиливать ADCC. Стандартный анализ ADCC проводят с использованием способов, описанных в настоящем документе. Лимфоциты собирают из периферической крови с использованием градиента Ficoll-Paque (Pharmacia). Клетки-мишени, т.е. культивируемые клеточные линии или полученные от пациента клетки, нагружают европием (PerkinElmer) и инкубируют с эффекторами в течение 4 ч при 37°C. Детекцию высвобожденного европия проводят с использованием флуоресцентного устройства для считывания планшетов (Wallac). Полученные данные ADCC указывают на эффективность молекул по изобретению в отношении запуска опосредуемой NK-клетками цитотоксичности и показывают, какие молекулы можно тестировать как с образцами пациентов, так и с подвергнутыми элютриации моноцитами. Затем молекулы диантител, демонстрирующие наибольший потенциал в отношении обеспечения активности ADCC, тестируют в анализе ADCC с использованием PBMC от пациентов. В качестве эффекторных клеток используют PBMC от здоровых доноров.
Таким образом, изобретение относится к способам профилактики или лечения злокачественной опухоли, характеризующейся антигеном злокачественной опухоли, путем конструирования молекулы диантитела для иммуноспецифического распознавания указанного антигена злокачественной опухоли, так чтобы молекула диантитела была сама цитотоксической (например, путем связывания рецепторов поверхности, ведущего к повышенному апоптозу или отрицательной регуляции пролиферативных сигналов) и/или содержала Fc-домен согласно изобретению и/или опосредовала одну или несколько эффекторных функций (например, ADCC, фагоцитоз). Диантитела, сконструированные согласно изобретению, пригодны для профилактики или лечения злокачественной опухоли, поскольку они обладают цитотоксической активностью (например, усиленным уничтожением опухолевых клеток и/или, например, усиленной активностью ADCC или активностью CDC).
Злокачественные опухоли, ассоциированные с антигеном злокачественной опухоли, можно лечить или предупреждать путем введения диантитела, которое связывает антиген злокачественной опухоли и является цитотоксическим и/или которое сконструировано способами по изобретению так, чтобы оно проявляло эффекторную функцию. Например, но не ограничиваясь этим, с помощью способов и композиций по изобретению можно лечить или предупреждать злокачественные опухоли, ассоциированные со следующими антигенами злокачественной опухоли: общий антиген карцином KS 1/4 (Perez et al. (1989) "Isolation And Characterization Of A Cdna Encoding The Ks1/4 Epithelial Carcinoma Marker", J. Immunol. 142:3662-3667; Moller et al. (1991) "Bispecific-Monoclonal-Antibody-Directed Lysis Of Ovarian Carcinoma Cells By Activated Human T Lymphocytes", Cancer Immunol. Immunother. 33(4):210-216), антиген карциномы яичника (CA125) (Yu et al. (1991) "Coexpression Of Different Antigenic Markers On Moieties That Bear CA 125 Determinants", Cancer Res. 51(2):468-475), простатический кислый фосфат (Tailor et al. (1990) "Nucleotide Sequence Of Human Prostatic Acid Phosphatase Determined From A Full-Length cDNA Clone", Nucl. Acids Res. 18(16):4928), простатспецифический антиген (Henttu et al. (1989) "cDNA Coding For The Entire Human Prostate Specific Antigen Shows High Homologies To The Human Tissue Kallikrein Genes", Biochem. Biophys. Res. Comm. 10(2):903-910; Israeli et al. (1993) "Molecular Cloning Of A Complementary DNA Encoding A Prostate-Specific Membrane Antigen", Cancer Res. 53:227-230), ассоциированный с меланомой антиген p97 (Estin et al. (1989) "Transfected Mouse Melanoma Lines That Express Various Levels Of Human Melanoma-Associated Antigen p97", J. Natl. Cancer Instit. 81(6):445-454), антиген меланомы gp75 (Vijayasardahl et al. (1990) "The Melanoma Antigen Gp75 Is The Human Homologue Of The Mouse B (Brown) Locus Gene Product", J. Exp. Med. 171(4):1375-1380), высокомолекулярный антиген меланомы (HMW-MAA) (Natali et al. (1987) "Immunohistochemical Detection Of Antigen In Human Primary And Metastatic Melanomas By The Monoclonal Antibody 140.240 And Its Possible Prognostic Significance", Cancer 59:55-63; Mittelman et al. (1990) "Active Specific Immunotherapy In Patients With Melanoma. A Clinical Trial With Mouse Antiidiotypic Monoclonal Antibodies Elicited With Syngeneic Anti-High-Molecular-Weight-Melanoma-Associated Antigen Monoclonal Antibodies", J. Clin. Invest. 86:2136-2144), простатспецифический мембранный антиген, карциноэмбриональный антиген (CEA) (Foon et al. (1995) "Immune Response To The Carcinoembryonic Antigen In Patients Treated With An Anti-Idiotype Antibody Vaccine", J. Clin. Invest. 96(1):334-42), полиморфный эпителиальный муциновый антиген, антиген жировых глобул молока человека, ассоциированные с опухолью толстой и прямой кишки антигены, такие как: CEA, TAG-72 (Yokota et al. (1992) "Rapid Tumor Penetration Of A Single-Chain Fv And Comparison With Other Immunoglobulin Forms", Cancer Res. 52:3402-3408), CO17-1A (Ragnhammar et al. (1993) "Effect Of Monoclonal Antibody 17-1A And GM-CSF In Patients With Advanced Colorectal Carcinoma-Long-Lasting, Complete Remissions Can Be Induced", Int. J. Cancer 53:751-758); GICA 19-9 (Herlyn et al. (1982) "Monoclonal Antibody Detection Of A Circulating Tumor-Associated Antigen. I. Presence Of Antigen In Sera Of Patients With Colorectal, Gastric, And Pancreatic Carcinoma", J. Clin. Immunol. 2:135-140), CTA-1 и LEA, антиген лимфомы Беркитта 38.13, CD19 (Ghetie et al. (1994) "Anti-CD 19 Inhibits The Growth Of Human B-Cell Tumor Lines In Vitro And Of Daudi Cells In SCID Mice By Inducing Cell Cycle Arrest", Blood 83:1329- 1336), антиген B-лимфомы человека CD20 (Reff et al. (1994) "Depletion Of B Cells In Vivo By A Chimeric Mouse Human Monoclonal Antibody To CD20", Blood 83:435-445), CD33 (Sgouros et al. (1993) "Modeling And Dosimetry Of Monoclonal Antibody M195 (Anti-CD33) In Acute Myelogenous Leukemia", J. Nucl. Med. 34:422-430), меланома-специфические антигены, такие как ганглиозид GD2 (Saleh et al. (1993) "Generation Of A Human Anti-Idiotypic Antibody That Mimics The GD2 Antigen", J.Immunol., 151, 3390-3398), ганглиозид GD3 (Shitara et al. (1993) "A Mouse/Human Chimeric Anti-(Ganglioside GD3) Antibody With Enhanced Antitumor Activities", Cancer Immunol. Immunother. 36:373-380), ганглиозид GM2 (Livingston et al. (1994) "Improved Survival In Stage III Melanoma Patients With GM2 Antibodies: A Randomized Trial Of Adjuvant Vaccination With GM2 Ganglioside", J. Clin. Oncol. 12:1036-1044), ганглиозид GM3 (Hoon et al. (1993) "Molecular Cloning Of A Human Monoclonal Antibody Reactive To Ganglioside GM3 Antigen On Human Cancers", Cancer Res. 53:5244-5250), опухолеспецифический трансплантационный тип антигена клеточной поверхности (TSTA), такой как индуцируемые вирусом антигены опухоли, включая T-антиген ДНК-вирусов опухолей и антигены оболочки РНК-вирусов опухолей, онкофетальный антиген альфа-фетопротеин, такой как онкофетальный CEA толстого кишечника, онкофетальный антиген мочевого пузыря (Hellstrom et al. (1985) "Monoclonal Antibodies To Cell Surface Antigens Shared By Chemically Induced Mouse Bladder Carcinomas", Cancer. Res. 45:2210-2188), антиген дифференцировки, такой как антиген карциномы легких человека L6, L20 (Hellstrom et al. (1986) "Monoclonal Mouse Antibodies Raised Against Human Lung Carcinoma", Cancer Res. 46:3917-3923), антигены фибросаркомы, антиген T-клеточного лейкоза человека Gp37 (Bhattacharya-Chatterjee et al. (1988) "Idiotype Vaccines Against Human T CeIl Leukemia. II. Generation And Characterization Of A Monoclonal Idiotype Cascade (AbI, Ab2, and Ab3)", J. Immunol. 141:1398-1403), неогликопротеин, сфинголипиды, антиген рака молочной железы, такой как EGFR (рецептор эпидермального фактора роста), антиген HER2 (p185HER2), полиморфный эпителиальный муцин (PEM) (Hilkens et al. (1992) "Cell Membrane-Associated Mucins And Their Adhesion-Modulating Property", Trends in Biochem. Sci. 17:359-363), злокачественный лимфоцитарный антиген человека APO-1 (Trauth et al. (1989) "Monoclonal Antibody-Mediated Tumor Regression By Induction Of Apoptosis", Science 245:301-304), антиген дифференцировки (Feizi (1985) "Demonstration By Monoclonal Antibodies That Carbohydrate Structures Of Glycoproteins And Glycolipids Are Onco-Developmental Antigens", Nature 314:53-57), такой как антиген I, встречающийся в эмбриональных эритроцитах и первичной эндодерме, I(Ma), встречающийся в аденокарциномах желудка, M18 и M39, встречающиеся в эпителии молочной железы, SSEA-I, встречающийся в миелоидных клетках, VEP8, VEP9, Myl, VIM-D5 и D156-22, встречающийся в раке толстой и прямой кишки, TRA-1-85 (группа крови H), C14, встречающийся в аденокарциноме толстой кишки, F3, встречающийся в аденокарциноме легких, AH6, встречающийся в раке желудка, гаптен Y, Ley, встречающийся в клетках эмбринальной карциномы, TL5 (группа крови A), рецептор EGF, встречающийся в клетках A431, серия E1 (группа крови B), встречающаяся в раке поджелудочной железы, FC10.2, встречающийся в клетках эмбриональной карциномы, аденокарциномы желудка, CO-514 (группа крови Lea), встречающийся в аденокарциноме, NS-10, встречающийся в аденокарциномах, CO-43 (группа крови Leb), G49, рецептор EGF, (группа крови ALeb/Ley), встречающийся в аденокарциноме толстой кишки, 19.9, встречающийся в раке толстой кишки, муцины рака желудка, T5A7, встречающийся в миелоидных клетках, R24, встречающийся в меланоме, 4.2, GD3, D1.1, OFA-1, GM2, OFA-2, GD2, M1:22:25:8, встречающийся в клетках эмбриональной карциномы SSEA-3, SSEA-4, встречающиеся в эмбрионах стадии 4-8 клеток. В другом варианте осуществления антиген представляет собой пептид, происходящий из T-клеточного рецептора при T-клеточной лимфоме кожи (см. Edelson (1998) "Cutaneous T-Cell Lymphoma: A Model For Selective Immunotherapy", Cancer J Sci Am. 4:62-71).
Злокачественные опухоли и сходные нарушения, которые можно лечить или предупреждать способами и композициями по настоящему изобретению, включают, но не ограничиваются ими, следующие: лейкозы, включающие, но не ограничивающиеся ими, острый лейкоз, острый лимфоцитарный лейкоз, острые миелоцитарные лейкозы, такие как миелобластный, промиелоцитарный, миеломоноцитарный, моноцитарный, эритроцитарный лейкозы и миелодиспластический синдром, хронические лейкозы, такие как, но не ограничиваясь ими, хронический миелоцитарный (гранулоцитарный) лейкоз, хронический лимфоцитарный лейкоз, волосатоклеточный лейкоз; истинную полицитемию; лимфомы, такие как, но не ограничиваясь ими, болезнь Ходжкина, неходжкинскую лимфому; множественные миеломы, такие как, но не ограничиваясь ими, вялотекущая множественная миелома, несекреторная миелома, остеосклеротическая миелома, плазмоклеточный лейкоз, изолированная плазмацитома и экстрамедуллярная плазмацитома; макроглобулинемию Вальденстрема; моноклональную гаммапатию неясного значения; доброкачестванную моноклональную гаммапатию; болезнь тяжелых цепей; саркомы костей и соединительной ткани, такие как, но не ограничиваясь ими, саркома кости, остеосаркома, хондросаркома, саркома Юинга, злокачественная гигантоклеточная опухоль, фибросаркома кости, хордома, периостальная саркома, саркомы мягких тканей, ангиосаркома (гемангиосаркома), фибросаркома, саркома Капоши, лейомиосаркома, липосаркома, лимфангиосаркома, неврилеммома, рабдомиосаркома, синовиальная саркома; опухоли головного мозга включая, но не ограничиваясь ими, глиому, астроцитому, глиому ствола головного мозга, эпендимому, олигоденроглиому, неглиальную опухоль, неврилеммому слухового нерва, краниофарингиому, медуллобластому, менингиому, пинеоцитому, пинеобластому, первичную лимфому головного мозга; рак молочной железы, включая, но не ограничиваясь ими, аденокарциному, лобулярную (мелкоклеточную) карциному, внутрипротоковую карциному, медуллярный рак молочной железы, муцинозный рак молочной железы, тубулярный рак молочной железы, папиллярный рак молочной железы, болезнь Педжета и воспалительный рак молочной железы; рак надпочечника, включая, но не ограничиваясь ими, феохромацитому и карциному надпочечников; рак щитовидной железы, такой как, но не ограничиваясь ими, папиллярный или фолликулярный рак щитовидной железы, медуллярный рак щитовидной железы и анапластический рак щитовидной железы; рак поджелудочной железы, включая, но не ограничиваясь ими, инсулиному, гастриному, глюкагоному, випому, соматостатин-секретирующую опухоль и карциноид или опухоль островковых клеток; злокачественные опухоли гипофиза, включая, но не ограничиваясь ими, болезнь Кушинга, пролактин-секретирующую опухоль, акромегалию и несахарный диабет; злокачественные опухоли глаза, включая, но не ограничиваясь ими, меланому глаза, такую как меланома радужной оболочки, хороидальная меланома и меланома ресничного тела, и ретинобластома; злокачественные опухоли влагалища, включая, но не ограничиваясь ими, плоскоклеточную карциному, аденокарциному и меланому; рак наружных женских половых органов, включая, но не ограничиваясь ими, плоскоклеточную карциному, меланому, аденокарциному, базально-клеточную карциному, саркому и болезнь Педжета; злокачественные опухоли шейки матки, включая, но не ограничиваясь ими, плоскоклеточную карциному и аденокарциному; злокачественные опухоли матки, включая, но не ограничиваясь ими, карциному эндометрия и саркому матки; злокачественные опухоли яичника, включая, но не ограничиваясь ими, эпителиальную карциному яичника, пограничную опухоль, опухоль половых клеток и стромальную опухоль; злокачественные опухоли пищевода, включая, но не ограничиваясь ими, плоскоклеточный рак, аденокарциному, кистозную карциному аденоидов, микоэпидермоидную карциному, плоскоклеточную аденокарциному, саркому, меланому, плазмацитому, бородавчатый рак и овсяно-клеточную (мелкоклеточную) карциному; злокачественные опухоли желудка, включая, но не ограничиваясь ими, аденокарциному, грибовидную (полипоидную), изъязвляющуюся, поверхностно распространяющуюся, диффузно распространяющуюся злокачественную лимфому, липосаркому, фибросаркому и карциносаркому; злокачественные опухоли толстого кишечника; злокачественные опухоли прямой кишки; злокачественные опухоли печени, включая, но не ограничиваясь ими, печеночно-клеточную карциному и гепатобластому, злокачественные опухоли желчного пузыря, включая, но не ограничиваясь ими, аденокарциному; холангиокарциномы, включая, но не ограничиваясь ими, папиллярную, узелковую и диффузную; злокачественные опухоли легкого, включая, но не ограничиваясь ими, немелкоклеточный рак легкого, плоскоклеточную карциному (эпидермоидную карциному), аденокарциному, крупноклеточную карциному и мелкоклеточный рак легкого; злокачественные опухоли яичка, включая, но не ограничиваясь ими, гоноцитому, семиному, анапластическую, классическую (типичную), сперматоцитарную, несеминому, эмбриональную карциному, тератомную карциному, хориокарциному (опухоль желточного мешка), злокачестенные опухоли предстательной железы, включая, но не ограничиваясь ими, аденокарциному, лейомиосаркому и рабдомиосаркому; злокачественные опухоли полового члена; злокачественные опухоли полости рта, включая, но не ограничиваясь ими, плоскоклеточную карциному; базально-клеточную карциному; злокачественные опухоли слюнных желез, включая, но не ограничиваясь ими, аденокарциному, мукоэпидермоидную карциному и кистозную карциному аденоидов; злокачественные опухоли глотки, включая, но не ограничиваясь ими, плоскоклеточный рак и бородавчатый рак; злокачественные опухоли кожи, включая, но не ограничиваясь ими, базально-клеточную карциному, плоскоклеточную карциному и меланому, поверхностно распространяющуюся меланому, узелковую меланому, злокачественную меланому лентиго, лентигиноз конечностей; злокачественные опухоли почки, включая, но не ограничиваясь ими, рак почечных клеток, аденокарциному, гипернефрому, фибросаркому, рак переходных клеток (почечная лоханка и/или мочеточник); опухоль Вильмса; злокачественные опухоли мочевого пузыря, включая, но не ограничиваясь ими, карциному переходных клеток, плоскоклеточный рак, аденокарциному, карциносаркому. Кроме того, злокачественные опухоли включают миксосаркому, остеогенную саркому, эндотелиосаркому, лимфангиоэндотелиосаркому, мезотелиому, синовиому, геманиобластому, карциному эпителия, цистаденокарциному, бронхогенную карциному, карциному потовых желез, карциному сальных желез, папиллярную карциному и папиллярные аденокарциномы (для обзора таких нарушений см. Fishman et al. (1985) Medicine, 2d Ed., J.B. Lippincott Co., Philadelphia; и Murphy et al. (1997) Informed Decisions: The Complete Book of Cancer Diagnosis, Treatment, and Recovery, Viking Penguin, Penguin Books U.S.A., Inc., United States of America).
Таким образом, способы и композиции по изобретению также пригодны для лечения или профилактики различных злокачественных опухолей или других заболеваний с нарушенной пролиферацией, включая (но не ограничиваясь ими) следующие: карцинома, включая карциному мочевого пузыря, молочной железы, толстого кишечника, почки, печени, легкого, яичника, поджелудочной железы, желудка, предстательной железы, шейки матки, щитовидной железы и кожи; включая плоскоклеточную карциному; гемопоэтические опухоли лимфоидного ростка, включая лейкоз, острый лимфоцитарный лейкоз, острый лимфобластный лейкоз, B-клеточную лимфому, T-клеточную лимфому, лимфом Беркитта; гемопоэтические опухоли миелоидного ростка, включая острые и хронические миелогенные лейкозы и промиелоцитарный лейкоз; опухоли мезенхимного происхождения, включая фибросаркому и рабдомиосаркому; другие опухоли, включая меланому, семиному, тератокарциному, нейробластому и глиому; опухоли центральной и периферической нервной системы, включая астроцитому, нейробластому, глиому и шванномы; опухоли мезенхимного происхождения, включая фибросаркому, рабдомиосаркому и остеосаркому; и другие опухоли, включая меланому, пигментную ксеродерму, кератоакантому, семиному, фолликулярный рак щитовидной железы и тератокарциному. Также предусматривается, что способами и композициями по изобретению можно лечить злокачественные опухоли, вызванные нарушениями в апоптозе. Такие злокачественные опухоли могут включать, но не ограничиваться ими, фолликулярные лимфомы, карциномы с мутациями p53, гормонзависимые опухоли молочной железы, предстательной железы и яичника и предлозкачественные нарушения, такие как семейный аденоматозный полипоз и миелодиспластические синдромы. В конкретных вариантах осуществления способами и композициями по изобретению лечат злокачественные или диспролиферативные изменения (такие как метаплазии и дисплазии), или гиперпролиферативные нарушения в яичнике, мочевом пузыре, молочной железе, толстом кишечнике, легком, коже, поджелудочной железе или матке. В других конкретных вариантах осуществления способами и композициями по изобретению лечат или предупреждают саркому, меланому или лейкоз.
В конкретном варианте осуществления молекула по изобретению (например, диантитело, содержащее множество связывающих эпитоп доменов и, необязательно, и Fc-домен (или его часть)) ингибирует или снижает рост злокачественных клеток по меньшей мере на 99%, по меньшей мере на 95%, по меньшей мере на 90%, по меньшей мере на 85%, по меньшей мере на 80%, по меньшей мере на 75%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 45%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 35%, по меньшей мере на 30%, по меньшей мере на 25%, по меньшей мере на 20% или по меньшей мере на 10% относительно роста злокачественных клеток в отсутствие указанной молекулы по изобретению.
В конкретном варианте осуществления молекула по изобретению (например, диантитело, содержащее множество связывающих эпитоп доменов и, необязательно, и Fc-домен (или его часть)) уничтожает клетки или ингибирует или снижает рост злокачественных клеток по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 100% лучше, чем исходная молекула.
5.6.2 АУТОИММУННОЕ ЗАБОЛЕВАНИЕ И ВОСПАЛИТЕЛЬНЫЕ ЗАБОЛЕВАНИЯ
В некоторых вариантах осуществления молекулы по изобретению содержат связывающий эпитоп домен, специфичный к FcγRIIB, и/или вариант Fc-домена (или его часть), сконструированный способами по изобретению, причем этот Fc-домен проявляет более высокую аффинность к FcγRIIB и сниженную аффинность к FcγRIIIA и/или FcγRIIA относительно Fc-домена дикого типа. Молекулы по изобретению с такими характеристиками связывания пригодны для регуляции иммунного ответа, например, для ингибирования иммунного ответа применительно к аутоиммунным заболеваниям или воспалительным заболеваниям. Без связи с каким-либо механизмом действия, молекулы по изобретению с аффинностью к FcγRIIB и/или содержащие Fc-домен с увеличенной аффинностью к FcγRIIB и сниженной аффинностью к FcγRIIIA и/или FcγRIIA, могут привести к ослаблению активирующего ответа на FcγR и ингибированию способности клеток к ответам и, таким образом, обладают терапевтической эффективностью для лечения и/или профилактики аутоиммунного нарушения.
Также изобретение относится к способам профилактики, лечения или управления течением одного или нескольких симптомов, ассоциированных с воспалительным нарушением у индивидуума, кроме того, включающим введение указанному индивидууму терапевтически или профилактически эффективного количества одного или нескольких противовоспалительных средств. Также изобретение относится к способам профилактики, лечения или управления течением одного или нескольких симптомов, ассоциированных с аутоиммунным заболеванием, кроме того, включающим введение указанному индивидууму терапевтически или профилактически эффективного количества одного или нескольких иммуномодулирующих средств. В разделе 5.7 представлены неограничивающие примеры противовоспалительных средств и иммуномодулирующих средств.
Примеры аутоиммунных нарушений, которые можно лечить путем введения молекул по настоящему изобретению, включают, но не ограничиваются ими, очаговую алопецию, анкилозирующий спондилит, антифосфолипидный синдром, аутоиммунную болезнь Аддисона, аутоиммунные заболевания надпочечника, аутоиммунную гемолитическую анемию, аутоиммунный гепатит, аутоиммунный оофорит и орхит, аутоиммунную тромбоцитопению, болезнь Бехчета, буллезный пемфигоид, кардиомиопатию, брюшные афты-дерматит, синдром хронической усталости вследствие иммунной дисфункции (CFIDS), хроническую воспалительную демиелинизирующую полиневропатию, синдром Черджа-Стросс, рубцовый пемфигоид, синдром CREST, синдром холодовой агглютинации, болезнь Крона, дискоидную волчанку, эссенциальную смешанную криоглобулинемию, фибромиалгию-фибромиозит, гломерулонефрит, болезнь Грэйвса, синдром Гийена-Барре, тиреоидит Хашимото, идиопатический фиброз легких, идиопатическую тромбоцитопеническую пурпуру (ITP), IgA-невропатию, ювенильный артрит, плоский лишай, красную волчанку, болезнь Меньера, смешанное заболевание соединительной ткани, рассеянный склероз, сахарный диабет 1 типа или иммуноопосредуемый сахарный диабет, миастению, пемфигус обыкновенный, пернициозную анемию, узелковый полиартериит, полихондрит, полигландулярные синдромы, ревматическую полимиалгию, полимиозит и дерматомиозит, первичную агаммаглобулинемию, первичный билиарный цирроз, псориаз, псориатический артрит, явление Рейно, синдром Рейтера, ревматоидный артрит, саркоидоз, склеродермию, синдром Шегрена, синдром ригидности мышц, системную красную волчанку, красную волчанку, артериит Такаясу, темпоральный артериит/гигантоклеточный артериит, язвенный колит, увеит, васкулиты, такие как герпетиформный дерматит васкулит, витилиго и гранулематоз Вегенера. Примеры воспалительных нарушений включают, но не ограничиваются ими, астму, энцефалит, воспалительное заболевание кишечника, хроническое обструктивное заболевание легких (COPD), аллергические нарушения, септический шок, фиброз легких, недифференцированную спондилоартропатию, недифференцированную артропатию, артрит, воспалительный остеолизис и хроническое воспаление вследствие хронических вирусных или бактериальных инфекций. Как описано в настоящем документе в разделе 2.2.2, некоторые аутоиммунные нарушения ассоциированы с воспалительным состоянием. Таким образом, существует перекрывание между тем, что считают аутоиммунным нарушением и воспалительным нарушением. Таким образом, некоторые аутоиммунные нарушения также можно охарактеризовать как воспалительные нарушения. Примеры воспалительных нарушений, которые можно предупреждать, лечить или контролировать в соответствии со способами по изобретению, включают, но не ограничиваются ими, астму, энцефалит, воспалительное заболевание кишечника, хроническое обструктивное заболевание легких (COPD), аллергические нарушения, септический шок, фиброз легких, недифференцированную спондилоартропатию, недифференцированную артропатию, артрит, воспалительный остеолизис и хроническое воспаление вследствие хронических или бактериальных инфекций.
Молекулы по изобретению, содержащие по меньшей мере один связывающий эпитоп домен, специфичный к FcγRIIB, и/или вариант Fc-домена с повышенной аффинностью к FcγRIIB и сниженной аффинностью к FcγRIIIA, также можно использовать для снижения воспаления у животных, в частности млекопитающих, с воспалительными нарушениями. В конкретном варианте осуществления молекула по изобретению снижает воспаление у животного по меньшей мере на 99%, по меньшей мере на 95%, по меньшей мере на 90%, по меньшей мере на 85%, по меньшей мере на 80%, по меньшей мере на 75%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 45%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 35%, по меньшей мере на 30%, по меньшей мере на 25%, по меньшей мере на 20% или по меньшей мере на 10% относительно воспаления у животного, которому не вводят указанную молекулу.
Молекулы по изобретению, содержащие по меньшей мере один связывающий эпитоп домен, специфичный к FcγRIIB, и/или вариант Fc-домена с повышенной аффинностью к FcγRIIB и сниженной аффинностью к FcγRIIIA, также можно использовать для предупреждения отторжения трансплантатов.
5.6.3 ИНФЕКЦИОННОЕ ЗАБОЛЕВАНИЕ
Также изобретение охватывает способы лечения или профилактики инфекционного заболевания у индивидуума, включающие введение терапевтически или профилактически эффективного количества одной или нескольких молекул по изобретению, содержащих по меньшей мере один связывающий эпитоп домен, специфичный к инфекционному агенту, ассоциированному с указанным инфекционным заболеванием. В определенных вариантах осуществления молекулы по изобретению являются токсичными для инфекционного агента, усиливают иммунный ответ против указанного агента или усиливают эффекторную функцию против указанного агента, относительно иммунного ответа в отсутствие указанной молекулы. Инфекционные заболевания, которые можно лечить или предупреждать с помощью молекул по изобретению, вызываются инфекционными агентами, включая, но не ограничиваясь ими, вирусы, бактерии, грибы, простейших и вирусы.
Вирусные заболевания, которые можно лечить или предупреждать с использованием молекул по изобретению совместно со способами по настоящему изобретению, включают, но не ограничиваются ими, заболевания, вызываемыми вирусом гепатита типа A, вирусом гепатита типа B, вирусом гепатита типа C, вирусом гриппа, вирусом ветрянки, аденовирусом, вирусом простого герпеса типа I (HSV-I), вирусом простого герпеса типа II (HSV-II), вирусом чумы рогатого скота, риновирусом, echo-вирусом, ротавирусом, респираторно-синцитиальным вирусом, вирусом папилломы, паповавирусом, цитомегаловирусом, эхиновирусом, арбовирусом, гунтавирусом, вирусом коксаки, вирусом свинки, вирусом кори, вирусом краснухи, вирусом полиомиелита, вирусом оспы, вирусом Эпштейна-Барр, вирусом иммунодефицита человека типа I (ВИЧ-I), вирусом иммунодефицита человека типа II (ВИЧ-II), и агентами вирусных заболеваний, таких как вирусный менингит, энцефалит, денге или оспа.
Бактериальные заболевания, которые можно лечить или предупреждать с использованием молекул по изобретению совместно со способами по настоящему изобретению, которые вызываются бактериями, включают, но не ограничиваются ими, заболевания, вызываемые микобактериями, риккетсиями, микоплазмой, нейсериями, S. pneumoniae, Borrelia burgdorferi (болезнь Лайма), Bacillus antracis (сибирская язва), столбнячными бактериями, стрептококками, стафилококками, микобактериями, столбнячными бактериями, коклюшными бактериями, холерными бактериями, чумными бактериями, дифтерийными бактериями, хламидиями, S. aureus и легионеллами.
Вызываемые простейшими заболевания, которые можно лечить или предупреждать с использованием молекул по изобретению совместно со способами по настоящему изобретению, которые вызываются простейшими, включают, но не ограничиваются ими, заболевания, вызываемые лейшманиями, возбудителями кокцидоза, трипаносомами, или малярию.
Паразитарные заболевания, которые можно лечить или предупреждать с использованием молекул по изобретению совместно со способами по настоящему изобретению, которые вызываются паразитами, включают, но не ограничиваются ими, болезни, вызываемые хламидиями и риккетсиями.
Согласно одному аспекту изобретения молекулы по изобретению, содержащие по меньшей мере один связывающий эпитоп домен, специфичный к инфекционному агенту, проявляют эффекторную функцию антитела в отношении указанного агента, например, патогенного белка. Примеры инфекционных агентов включают, но не ограничиваются ими, бактерии (например, Escherichia coli, Klebsiella pneumoniae, Staphylococcus aureus, Enterococcus faecials, Candida albicans, Proteus vulgaris, Staphylococcus viridans и Pseudomonas aeruginosa), патоген (например, B-лимфотропный паповавирус (LPV); Bordatella pertussis; вирус болезни Борна (BDV); коронавирус крупного рогатого скота; вирус хориоменингита; вирус Денге; E. coli; вирус Эбола; Echo-вирус 1; Echo-вирус 11 (EV); эндотоксин (LPS); кишечные бактерии; кишечный "сиротский" вирус; энтеровирусы; вирус лейкоза кошек; вирус ящура; вирус лейкоза гиббонов (GALV); грамотрицательные бактерии; Heliobacter pylori; вирус гепатита B (HBV); вирус простого герпеса; ВИЧ-I; цитомегаловирус человека; коронавирус человека; вирус гриппа A, B и C; легионеллы; Leishmania mexicana; Listeria monocytogenes; вирус кори; менингококки; морбилливирусы; вирус гепатита мышей; вирус лейкоза мышей; вирус герпеса гамма мышей; ретровирус мышей; коронавирус мышей, вирус гепатита мышей; Mycobacterium avium-M; Neisseria gonorrhoeae; вирус болезни Ньюкасла; парвовирус B19; Plasmodium falciparum; поксвирус; Pseudomonas; ротавирус; Samonella typhiurium; шигеллы; стрептококки; T-клеточный лимфотропный вирус 1; вирус осповакцины).
5.6.4 ДЕТОКСИКАЦИЯ
Также изобретение охватывает способы детоксикации у индивидуума, подвергаемого воздействию токсина (например, молекулы токсичного лекарственного средства), включающие введение терапевтически или профилактически эффективного количества одной или нескольких молекул по изобретению, содержащих по меньшей мере один связывающий эпитоп домен, специфичный к молекуле токсичного лекарственного средства. В определенных вариантах осуществления связывание молекулы по изобретению с токсином снижает или устраняет неблагоприятный физиологический эффект указанного токсина. В других вариантах осуществления связывание диантитела по изобретению с токсином увеличивает или усиливает выведение, деградацию или нейтрализацию токсина относительно выведения, деградации или нейтрализации в отсутствие указанного диантитела. Иммунотоксикотерапию в соответствии со способами по изобретению можно использовать для лечения передозировок или воздействия лекарственных средств, включая, но не ограничиваясь ими, дигоксин, PCP, кокаин, колхицин и трициклические антидепрессанты.
5.7 КОМБИНИРОВАННАЯ ТЕРАПИЯ
Кроме того, изобретение охватывает введение молекул по изобретению в комбинации с другими способами лечения, известными специалистам в данной области, для лечения или профилактики злокачественной опухоли, аутоиммунного заболевания, инфекционного заболевания или интоксикации, включая, но не ограничиваясь ими, современные стандартные и экспериментальные способы химиотерапии, способы гормональной терапии, способы биологической терапии, способы иммунотерапии, способы лучевой терапии или хирургическую операцию. В некоторых вариантах осуществления молекулы по изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или нескольких средств, терапевтических антител или других средств, известных специалистам в данной области, для лечения и/или профилактики злокачественной опухоли, аутоиммунного заболевания, инфекционного заболевания или интоксикации.
В определенных вариантах осуществления одну или несколько молекул по изобретению вводят млекопитающему, предпочтительно человеку, одновременно с одним или несколькими другими лекарственными средствами, пригодными для лечения злокачественной опухоли. Термин "одновременно" не ограничивается введением профилактических или терапевтических средств точно в одно время, а вместо этого он подразумевает, что молекулу по изобретению и другое средство вводят млекопитающему последовательно и в пределах такого временного интервала, чтобы молекула по изобретению могла действовать совместно с другим средством, обеспечивая увеличенную пользу по сравнению с их введением иным образом. Например, каждое профилактическое или терапевтическое средство (например, химиотерапию, лучевую терапию, гормональную терапию или биологическую терапию) можно проводить одновременно или последовательно в любом порядке в различные моменты времени; однако если их не проводят одновременно, их следует проводить достаточно близко по времени, чтобы обеспечить желаемый терапевтический или профилактический эффект. Каждое из лекарственных средств можно вводить отдельно, в любой подходящей форме или любым пригодным путем. В различных вариантах осуществления профилактические или терапевтические средства вводят с интервалом менее 1 часа, приблизительно 1 час, от приблизительно 1 часа до приблизительно 2 часов, от приблизительно 2 часов до приблизительно 3 часов, от приблизительно 3 часов до приблизительно 4 часов, от приблизительно 4 часов до приблизительно 5 часов, от приблизительно 5 часов до приблизительно 6 часов, от приблизительно 6 часов до приблизительно 7 часов, от приблизительно 7 часов до приблизительно 8 часов, от приблизительно 8 часов до приблизительно 9 часов, от приблизительно 9 часов до приблизительно 10 часов, от приблизительно 10 часов до приблизительно 11 часов, от приблизительно 11 часов до приблизительно 12 часов, не более чем 24 часа или не более чем 48 часов. В предпочтительных вариантах осуществления два или более компонентов вводят за один визит пациента.
В других вариантах осуществления профилактические или терапевтические средства вводят с интервалом приблизительно от 2 до 4 суток, от 4 до 6 суток, приблизительно 1 неделю, приблизительно от 1 до 2 недель или более 2 недель. В предпочтительных вариантах осуществления профилактические или терапевтические средства вводят во временном интервале, когда оба средства все еще являются активными. Специалист в данной области способен определить такой временной интервал путем определения времени полужизни вводимых средств.
В определенных вариантах осуществления профилактические или терапевтические средства по изобретению вводят индивидууму курсами. Курсовая терапия вовлекает введение первого средства в течение периода времени с последующим введением второго средства и/или третьего средства в течение периода времени и повторение этого последовательного введения. Курсовая терапия может снизить развитие устойчивости к одному или нескольким способам терапии, предотвратить или снизить побочные эффекты одного из способов лечения и/или повысить эффективность лечения.
В определенных вариантах осуществления профилактические или терапевтические средства вводят курсом менее чем приблизительно из 3 недель, приблизительно один раз в две недели, приблизительно один раз каждые 10 суток или приблизительно один раз в неделю. Один курс может включать введение терапевтического или профилактического средства путем инфузии в течение приблизительно 90 минут в каждом курсе, приблизительно 1 час в каждом курсе, приблизительно 45 минут в каждом курсе. Каждый курс может включать по меньшей мере 1 неделю покоя, по меньшей мере 2 недели покоя, по меньшей мере 3 недели покоя. Количество проводимых курсов составляет от приблизительно 1 до приблизительно 12 курсов, более конкретно от приблизительно 2 до приблизительно 10 курсов, и более конкретно от приблизительно 2 до приблизительно 8 курсов.
В других вариантах осуществления терапевтические и профилактические средства по изобретению вводят в метрономических режимах дозирования либо путем непрерывной инфузии, либо путем частого введения без длительных периодов покоя. Такое метрономическое введение может вовлекать дозирование с постоянными интервалами без периодов покоя. Как правило, лекарственные средства, в частности цитотоксические средства, используют в более низких дозах. Такие режимы дозирования охватывают длительное введение каждые сутки относительно низких доз в течение длительных периодов времени. В предпочтительных вариантах осуществления применение более низких доз может минимизировать токсические побочные эффекты и устранить периоды покоя. В определенных вариантах осуществления терапевтические и профилактические средства доставляют с помощью длительной инфузии в низкой дозе или непрерывной инфузии в диапазоне от приблизительно 24 часов до приблизительно 2 суток, до приблизительно 1 недели, до приблизительно 2 недель, до приблизительно 3 недель, до приблизительно 1 месяца, до приблизительно 2 месяцев, до приблизительно 3 месяцев, до приблизительно 4 месяцев, до приблизительно 5 месяцев, до приблизительно 6 месяцев. Расписание таких режимов дозирования может оптимизировать квалифицированный онколог.
В других вариантах осуществления курсы лечения проводят млекопитающему одновременно, т.е. отдельные дозы лекарственных средств вводят по отдельности, но в пределах такого временного интервала, чтобы молекулы по изобретению могли действовать вместе с другим средством или средствами. Например, один компонент можно вводить один раз в неделю в комбинации с другими компонентами, которые можно вводить один раз в две недели или один раз в три недели. Иными словами, режимы дозирования для лекарственных средств проводят одновременно, даже если лекарственные средства не вводят одновременно или за один и тот же визит пациента.
При использовании в комбинации с другими профилактическими и/или терапевтическими средствами, молекулы по изобретению и профилактическое и/или терапевтическое средство могут действовать аддитивно или, более предпочтительно, синергично. В одном из вариантов осуществления молекулу по изобретению вводят одновременно с одним или несколькими лекарственными средствами в одной фармацевтической композиции. В другом варианте осуществления молекулу по изобретению вводят одновременно с одним или несколькими другими лекарственными средствами в отдельных фармацевтических композициях. В другом варианте осуществления молекулу по изобретению вводят до или после введения другого профилактического или терапевтического средства. Изобретение предусматривает введение молекулы по изобретению в комбинации с другими профилактическими или терапевтическими средствами одним или различными путями введения, например, пероральным и парентеральным. В определенных вариантах осуществления, когда молекулу по изобретению вводят одновременно с другим профилактическим или терапевтическим средством, которое потенциально вызывает неблагоприятные побочные эффекты, включая, но не ограничиваясь ими, токсичность, профилактическое или терапевтическое средство можно преимущественно вводить в дозе, которая находится ниже порога, который вызывает неблагоприятный побочный эффект.
Вводимые количества и частота введения, представленные в настоящем документе, охватываются терминами "терапевтически эффективный" и "профилактически эффективный". Дозировка и частота, кроме того, обычно варьируют в соответствии с факторами, определенными для каждого пациента в зависимости от конкретных вводимых терапевтических или профилактических средств, тяжести и типа злокачественной опухоли, пути введения, а также возраста, массы тела, ответа и медицинского анамнеза пациента. Пригодные режимы может выбирать специалист в данной области с учетом таких факторов и в соответствии, например, с дозировками, описанными в литературе и рекомендованными в Physician's Desk Reference (56th ed., 2002).
5.7.1 СРЕДСТВА ПРОТИВ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ
В конкретном варианте осуществления способы по изобретению охватывают введение одной или нескольких молекул по изобретению с одним или несколькими лекарственными средствами, используемыми для лечения и/или профилактики злокачественной опухоли. В одном из вариантов осуществления в комбинации с молекулами по изобретению можно вводить ингибиторы ангиогенеза. Ингибиторы ангиогенеза, которые можно использовать в способах и композициях по изобретению, включают, но не ограничиваются ими: ангиостатин (фрагмент плазминогена); антиангиогенный антитромбин III; ангиозим; ABT-627; Bay 12-9566; бенефин; бевацизумаб; BMS-275291; происходящий из хряща ингибитор (CDI); CAI; фрагмент комплемента CD59; CEP-7055; Col 3; комбретастатин A-4; эндостатин (фрагмент коллагена XVIII); фрагмент фибронектина; Gro-beta; галофунгинон; гепариназы; гексасахаридный фрагмент гепарина; HMV833; хорионический гонадотропин человека (hCG); IM-862; интерферон альфа/бета/гамма; индуцируемый интерфероном белок (IP-10); интерлейкин-12; Kringle 5 (фрагмент плазминогена); маримастат; ингибиторы металлопротеиназ (TIMP); 2-метоксиэстрадиол; MMI 270 (CGS 27023 A); MoAb IMC-IC11; неовастат; NM-3; панзем; PI-88; плацентарный ингибитор рибонуклеазы; ингибитор активатора плазминогена; фактор тромбоцитов-4 (PF4); приномастат; фрагмент пролактина массой 16 кДа; родственный пролиферину белок (PRP); PTK 787/ZK 222594; ретиноиды; солимастат; скваламин; SS 3304; SU 5416; SU6668; SU11248; тетрагидрокортизол-S; тетратиомолибдат; талидомид; тромбоспондин-1 (TSP-1); TNP-470; трансформирующий фактор роста-бета (TGF-b); васкулостатин; вазостатин (фрагмент кальретикулина); ZD6126; ZD 6474; ингибиторы фарнезилтрансферазы (FTI) и бисфосфонаты.
Средства против злокачественной опухоли, которые можно использовать в комбинации с молекулами по изобретению в различных вариантах осуществления изобретения, включая фармацевтические композиции и дозированные формы и наборы по изобретению, включают, но не ограничиваются ими: активицин; акларубицин; акозадола гидрохлорид; акронин; адозелезин; альдеслейкин; алтретамин; амбомицин; аметантрона ацетат; аминоглутетимид; амсакрин; анастрозол; антрамицин; аспарагиназу; асперлин; азацитидин; азетепу; азотомицин; батимастат; бензодепа; бикалутамид; бисантрена гидрохлорид; биснафида димезилат; бизелезин; блеомицина сульфат; бреквинар натрий; бропиримин; бусульфан; cактиномицин; калустерон; карацемид; карбетимер; карбоплатин; кармустин; карубицина гидрохлорид; карзелезин; цедефингол; хлорамбуцил; циролемицин; цисплатин; кладрибин; криснатола мезилат; циклофосфамид; цитарабин; дакарбазин; дактиномицин; даунорубицина гидрохлорид; децитабин; дексормаплатин; дезагуанин; дезагуанина мезилат; диазиквон; доцетаксел; доксорубицин; доксорубицина гидрохлорид; дролоксифен; дролоксифена цитрат; дромостанолона пропионат; дуазомицин; эдатрексат; эфлорнитина гидрохлорид; элсамитруцин; энлоплатин; энпромат; эпипропидин; эпирубицина гидрохлорид; эрбулозол; эзорубицина гидрохлорид; эстрамустин; эстрамустина фосфат натрий; этанидазол; этопозид; этопозида фосфат; этоприн; фадрозола гидрохлорид; фазарабин; фенретинид; флоксуридин; флударабина фосфат; фторурацил; флуроцитабин; фосквидон; фостриецина натрий; гемцитабин; гемцитабина гидрохлорид; гидроксимочевину; идарубицина гидрохлорид; ифосфамид; илмофозин; интерлейкин II (включая рекомбинантный интерлейкин II, или rIL2), интерферон альфа-2a; интерферон альфа-2b; интерферон альфа-n1; интерферон альфа-n3; интерферон бета-Ia; интерферон гамма-Ib; ипроплатин; иринотекана гидрохлорид; ланреотида ацетат; лектрозол; леупролида ацетат; лиарозола гидрохлорид; лометрексол натрий; ломустин; лозоксантрона гидрохлорид; мазопрокол; майтанзин; мехлорэтамина гидрохлорид; магестрола ацетат; меленгестрола ацетат; мелфалан; меногарил; меркаптопурин; метотрексат; метотрексат натрий; метоприн; метуредепу; митиндомид; митокарцин; митокромин; митогиллин; митомалцин; митомицин; митоспер; митотан; митоксантрона гидрохлорид; микофеноловую кислоту; нокодазол; ногаламицин; ормаплатин; оксисуран; паклитаксел; пегаспаргазу; пелиомицин; пентамустин; пепломицина сульфат; перфосфамид; пипоброман; пипосульфан; пироксантрона гидрохлорид; пликамицин; пломестан; порфимер натрий; порфиромицин; преднемустин; прокарбазина гидрохлорид; пуромицин; пуромицина гидрохлорид; пиразофурин; рибоприн; роглетимид; сафингол; сафингола гидрохлорид; семустин; симтразен; спарфосат натрий; спарсомицин; спирогермания гидрохлорид; спиромустин; спироплатин; стрептонигрин; стрептозоцин; сулофенур; тализомицин; текогалан натрий; тегафур; телоксантрона гидрохлорид; темопорфин; тенипозид; тероксирон; тестолактон; тиамиприн; тиогуанин; тиотепу; тиазофурин; тирапазамин; торемифена цитрат; трестолона ацетат; трицирибина фосфат; триметрексат; триметрексата глюкуронат; трипторелин; тубулозола гидрохлорид; урацил иприт; уредепу; вапреотид; вертепорфин; винбластина сульфат; винкристина сульфат; виндезин; виндезина сульфат; винепидина сульфат; винглицината сульфат; винлеурозина сульфат; винорелбина тартрат; винрозидина сульфат; винзолидина сульфат; ворозол; зениплатин; зиностатин; зорубицина гидрохлорид. Другие средства против злокачественной опухоли включают, но не ограничиваются ими: 20-эпи-1,25-дигидроксивитамин D3; 5-этинилурацил; абиратерон; акларубицин; ацилфулвен; адеципенол; адозелезин; альдеслейкин; ALL-TK антагонисты; алтретамин; амбамустин; амидокс; амифостин; аминолевулиновую кислоту; амрубицин; амсакрин; анагрелид; анастрозол; андрографолид; ингибиторы ангиогенеза; антагонист D; антагонист G; антареликс; антидорсализирующий морфогенный белок-1; антиандроген против карциномы предстательной железы; антиэстроген; антинеопластон; антисмысловые олигонуклеотиды; афидиколина глицинат; модуляторы генов апоптоза; регуляторы апоптоза; апуриновую кислоту; ara-CDP-DL-PTBA; аргининдезаминазу; асулакрин; атаместан; атримустин; аксинастатин 1; аксинастатин 2; аксинастатин 3; азастерон; азатоксин; азатирозин; производные баккатина III; баланол; батимастат; антагонисты BCR/ABL; бензохлорины; бензоилстауроспорин; производные бета-лактамов; бета-алетин; бетакламицин B; бетулиновую кислоту; ингибитор bFGF; бикалутамид; бисантрен; бисазиридинилспермин; биснафид; бистратен A; бизелезин; брефлат; бропиримин; будотитан; бутионина сулофоксимин; кальципотриол; кальфостин C; производные камптотецина; канапирокс IL-2; капецитабин; карбоксамид-аминотриазол; карбоксиамидотриазол; CaRest M3; CARN 700; происходящий из хряща ингибитор; карзелезин; казеин киназа ингибиторы (ICOS); кастаноспермин; цекропин B; цетрореликс; хлорины; хлорхиноксалина сульфонамид; цикапрост; цис-порфирин; кладрибин; аналоги кломифена; клотримазол; коллисмицин A; коллисмицин B; комбретастатин A4; аналог комбретастатина; конагенин; крамбесцидин 816; криснатол; криптофицин 8; производные криптофицина A; курацин A; циклопентантрахиноны; циклоплатам; ципемицин; цитарабина окфосфат; цитолитический фактор; цитостатин; дакликсимаб; децитабин; дегидродидемнин B; деслорелин; дексаметазон; дексифосфамид; дексразоксан; дексверапамил; диазиквон; дидемнин B; дидокс; диэтилнормспермин; дигидро-5-азацитидин; дигидротаксол, 9-диоксамицин; дифенилспиромустин; доцетаксел; доказанол; долазетрон; доксифлуридин; дролоксифен; дронабинол; дуокармицин SA; эбселен; экомустин; эдельфозин; эдреколомаб; эфлорнитин; элемен; эмитефур; эпирубицин; эпристерид; аналог эстрамустина; агонисты эстрогенов; антагонисты эстрогенов; этанидазол; этопозида фосфат; экземестан; фадрозол; фазарабин; фенретинид; филграстим; финастерид; флавопиридол; флезеластин; флуастерон; флударабин; фтордауноруницина гидрохлорид; форфенимекс; форместан; фостриецин; фотемустин; гадолиния тексафирин; нитрат галлия; галоцитабин; ганиреликс; ингибиторы желатиназы; гемцитабин; ингибиторы глутатиона; гепсульфам; херегулин; гексаметилена бисацетамид; гиперицин; ибандроновую кислоту; идарубицин; идоксифен; идрамантон; илмофозин; иломастат; имидазоакридоны; имиквимод; иммуностимулирующие пептиды; ингибитор рецептора инсулиноподобного фактора роста-1; агонисты интерферона; интерфероны; интерлейкины; иобенгуан; йододоксорубицин; ипомеанол, 4-ироплакт; ирсогладин; изобенгазол; изогомохаликондрин B; итасетрон; ясплакинолид; кагалалид F; ламелларин-N триацетат; ланреотид; леинамицин; ленограстим; лентинана сульфат; лептолстатин; лектрозол; ингибирующий фактор лейкоза; альфа-интерферон лейкоцитов; леупролид+эстроген+прогестерон; леупрорелин; левамизол; лиарозол; линейный аналог полиамина; липофильный дисахаридный пептид; липофильные соединения платины; лиссоклинамид 7; лобаплатин; ломбрицин; лометрексол; лонидамин; лозоксантрон; ловастатин; локсорибин; луртотекан; лютеций тексафирин; лизофиллин; литические пептиды; майтанзин; манностатин A; маримастат; мазопрокол; маспин; ингибиторы матрилизина; ингибиторы матриксных металлопротеиназ; меногарил; мербарон; метерелин; метиониназу; метоклопрамид; ингибитор MIF; мифепристон; милтефозин; миримостим; двухцепочечную РНК с несовпадающими основаниями; митогуазон; митолактол; аналоги митомицина; митонафид; митотоксиновый фибробластный фактор роста-сапорин; митоксантрон; мофаротен; молграмостим; моноклональное антитело, хорионический гонадотропин человека; монофосфориллипид A + sk клеточной стенки микобактерий; мопидамол; ингибитор множественной устойчивости к лекарственным средствам; терапию на основе множественного супрессора опухолей 1; средство против злокачественной опухоли на основе иприта; микапероксид B; экстракт клеточной стенки микобактерий; мириапорон; N-ацетилдиналин; N-замещенные бензамиды; нафарелин; нагрестип; налоксон + пентазоцин; напавин; нафтерпин; нартограстим; недаплатин; неморубицин; неридроновую кислоту; нейтральную эндопептидазу; нилутамид; низамицин; модуляторы оксида азота; нитроксидный антиоксидант; нитруллин; O6-бензилгуанин; октреотид; окиценон; олигонуклеотиды; онапристон; ондансетрон; ондансетрон; орацин; пероральный индуктор цитокинов; ормаплатин; осатерон; оксалиплатин; оксауномицин; паклитаксел; аналоги паклитаксела; производные паклитаксела; палауамин; пальмитоилризоксин; памидроновую кислоту; панакситриол; паномифен; парабактин; пазеллиптин; пегаспаргазу; пелдезин; пентосан полисульфат натрия; пентостатин; пентрозол; перфлуброн; перфосфамид; периллиловый спирт; феназиномицин; фенилацетат; ингибиторы фосфатазы; пицибанил; пилокарпина гидрохлорид; пирарубицин; пиритрексим; плацетин A; плацетин B; ингибитор активатора плазминогена; комплекс платины; соединения платины; комплекс платины-триамина; порфимер натрий; порфиромицин; преднизон; пропил-бис-акридон; простагландин J2; ингибиторы протеасом; иммунный модулятор на основе белка A; ингибитор протеинкиназы C; ингибиторы протеинкиназы C, микроалгал; ингибиторы тирозинфосфатазы белков; ингибиторы фосфорилазы пуриновых нуклеозидов; пурпурины; пиразолoакридин; конъюгат пиридоксилированного гемоглобина и полиоксиэтилена; антагонисты raf; ралтитрексед; рамосетрон; ингибиторы фарнезилпротеинтрансферазы ras; ингибиторы ras; ингибитор ras-GAP; деметилированный ретеллиптин; этидронат рения Re 186; ризоксин; рибозимы; RII ретинамид; роглетимид; рокситукин; ромуртид; роквинимекс; рубигинон B1; рубоксил; сафингол; саинтопин; SarCNU; саркофитол A; сарграмостим; миметики Sdi 1; семустин; образующийся при старении ингибитор 1; смысловые олигонуклеотиды; ингибиторы передачи сигнала; модуляторы передачи сигнала; одноцепочечный антигенсвязывающий белок; сизофиран; собузоксан; натрий борокаптат; натрий фенилацетат; солверол; связывающий соматомедин белок; сонермин; спарфозовую кислоту; спикамицин D; спиромустин; спленопентин; спонгистатин 1; скваламин; ингибитор стволовых клеток; ингибиторы деления стволовых клеток; стрипиамид; ингибиторы стромелизина; сульфмосин; антагонист суперактивного вазоактивного интестинального пептида; сурадисту; сурамин; сваинсонин; синтетические гликозаминогликаны; таллимустин; метйодид тамоксифена; тауромустин; тазаротен; текогалан натрий; тегафур; теллурапирилий; ингибиторы теломеразы; темопорфин; темозоломид; тенипозид; тетрахлордекаоксид; тетразомин; талибластин; тиокоралин; тромбопоэтин; миметик тромбопоэтина; тималфазин; агонист рецептора тимопоэтина; тимотринан; тиреостимулирующий гормон; этилэтиопурпурин олова; тирапазамин; титаноцен бихлорид; топсентин; торемифен; фактор тотипотентных стволовых клеток; ингибиторы трансляции; третиноин; триацетилуридин; трицирибин; триметрексат; трипторелин; трописетрон; туростерид; ингибиторы тирозинкиназы; тирфостины; ингибиторы UBC; убенимекс; происходящий из мочеполовой пазухи ингибирующий рост фактор; антагонисты рецептора урокиназы; вапреотид; вариолин B; эритроцитарную генную терапию векторной системой; веларесол; верамин; вердины; вертепорфин; винорелбин; винксалтин; витаксин; ворозол; занотерон; зениплатин; зиласкорб и зиностатин стималамер. Предпочтительными дополнительными лекарственными средствами против злокачественной опухоли являются 5-фторурацил и лейковорин.
Примеры терапевтических антител, которые можно использовать в способах по изобретению, включают, но не ограничиваются ими, ZENAP AXВ® (даклизумаб) (Roche Pharmaceuticals, Switzerland), который представляет собой иммунодепрессивное гуманизированное моноклональное антитело против CD25 для предупреждения острого отторжения аллотрансплантата почки; PANOREXTM, который представляет собой IgG2a-антитело мыши против антигена клеточной поверхности 17-IA антитело (Glaxo Wellcome/Centocor); BEC2, которое представляет собой антиидиотипическое IgG-антитело мыши (эпитоп GD3) (ImClone System); IMC-C225, которое представляет собой химерное IgG-антитело против EGFR (ImClone System); VITAXINTM, которое представляет собой гуманизированное антитело против αVβ3 интегрина (Applied Molecular Evolution/MedImmune); Smart M 195, которое представляет собой гуманизированное IgG-антитело против CD33 (Protein Design Lab/Kanebo); LYMPHOCIDETM, которое представляет собой гуманизированное IgG-антитело против CD22 (Immunomedics); ICM3, которое представляет собой гуманизированное антитело против ICAM3 (ICOS Pharm); IDEC-114, которое представляет собой приматизированное антитело против CD80 (IDEC Pharm/Mitsubishi); IDEC-131, которое представляет собой гуманизированное антитело против CD40L (IDEC/Eisai); IDEC-151, которое представляет собой приматизированное антитело против CD4 (IDEC); IDEC-152, которое представляет собой приматизированное антитело против CD23 (IDEC/Seikagaku); SMART anti-CD3, которое представляет собой гуманизированное антитело против CD3 IgG (Protein Design Lab); 5G1.1, которое представляет собой гуманизированное антитело против фактора 5 комплемента (C5) антитело (Alexion Pharm); D2E7, которое представляет собой гуманизированное антитело против TNF-α (CAT/BASF); CDP870, которое представляет собой гуманизированный Fab-фрагмент против TNF-α (Celltech); IDEC-151, которое представляет собой приматизированное IgG1-антитело против CD4 (IDEC Pharm/SmithKline Beecham); MDX-CD4, которое представляет собой IgG-антитело человека против CD4 (Medarex/Eisai/Genmab); CDP571, которое представляет собой гуманизированное IgG4-антитело против TNF-α (Celltech); LDP-02, которое представляет собой гуманизированное антитело против α4β7 (LeukoSite/Genentech); OrthoClone OKT4A, которое представляет собой гуманизированное IgG-антитело против CD4 (Ortho Biotech); ANTOVATM, которое представляет собой гуманизированное IgG-антитело против CD40L (Biogen); ANTEGRENTM, которое представляет собой гуманизированное IgG-антитело против VLA-4 (Elan); и CAT-152, которое представляет собой антитело человека против TGF-β2 (Cambridge Ab Tech). Другие примеры терапевтических антител, которые можно использовать в соответствии с изобретением, представлены в таблице 8.
Терапевтические антитела против злокачественной опухоли
(алемтузумаб)
5.7.2 ИММУНОМОДУЛИРУЮЩИЕ СРЕДСТВА И ПРОТИВОВОСПАЛИТЕЛЬНЫЕ СРЕДСТВА
Настоящее изобретение относится к способам лечения аутоиммунных заболеваний и воспалительных заболеваний, включающим введение молекул по изобретению вместе с другими средствами для лечения. Примеры иммуномодулирующих средств включают, но не ограничиваются ими, метотрексат, ENBREL, REMACADETM, лефлуномид, циклофосфамид, циклоспорин A и макролидные антибиотики (например, FK506 (такролимус)), метилпреднизолон (MP), кортикостероиды, стероиды, микофенолат мофетил, рапамицин (сиролимус), мизорибин, дезоксиспергуалин, бреквинар, малононитрилоамиды (например, лефлунамид), модуляторы T-клеточных рецепторов и модуляторы рецепторов цитокинов.
Противовоспалительные средства демонстрируют успех при лечении воспалительных и аутоиммунных нарушений и в настоящее время являются распространенным и стандартным средством для лечения таких нарушений. В способах по изобретению можно использовать противовоспалительное средство, хорошо известное специалисту в данной области. Неограничивающие примеры противовоспалительных средств включают нестероидные противовоспалительные лекарственные средства (NSAID), стероидные противовоспалительные лекарственные средства, бета-агонисты, антихолинергические средства и метилксантины. Примеры NSAID включают, но не ограничиваются ими, аспирин, ибупрофен, целекоксиб (CELEBREXTM), диклофенак (VOLTARENTM), этодолак (LODINETM), фенопрофен (NALFONTM), индометацин (INDOCINTM), кеторолак (TORADOLTM), оксапрозин (DAYPROTM), набументон (RELAFENTM), сулиндак (CLINORILTM), толментин (TOLECTINTM), рофекоксиб (VIOXXTM), напроксен (ALEVETM, NAPROSYNTM), кетопрофен (ACTRONTM) и набуметон (RELAFENTM). Такие NSAID действуют путем ингибирования фермента циклооксигеназы (например, COX-1 и/или COX-2). Примеры стероидных противовоспалительных лекарственных средств включают, но не ограничиваются ими, глюкокортикоиды, дексаметазон (DECADRONTM), кортизон, гидрокортизон, преднизон (DELTASONETM), преднизолон, триамцинолон, азулфидин и эйкозаноиды, такие как простагландины, тромбоксаны и лейкотриены.
Неограничивающий пример антител, которые можно использовать для лечения или профилактики воспалительных нарушений применительно к молекулам по изобретению, представлен в таблице 9, и неограничивающий пример антител, которые можно использовать для лечения или профилактики аутомиммунных нарушений, представлен в таблице 10.
Терапевтические антитела для лечения воспалительных заболеваний
Терапевтические антитела для лечения аутоиммунных нарушений
5.7.3 СРЕДСТВА ДЛЯ ПРИМЕНЕНИЯ В ЛЕЧЕНИИ ИНФЕКЦИОННОГО ЗАБОЛЕВАНИЯ
В некоторых вариантах осуществления молекулы по изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или дополнительных лекарственных средств, известных специалистам в данной области, для лечения и/или профилактики инфекционного заболевания. Изобретение предусматривает применение молекул по изобретению в комбинации с антибиотиками, известными специалистам в данной области, для лечения и/или профилактики инфекционного заболевания. Антибиотики, которые можно использовать в комбинации с молекулами по изобретению, включают, но не ограничиваются ими, макролид (например, тобрамицин (Tobi®)), цефалоспорин (например, цефалексин (Keflex®), цефрадин (Velosef®), цефуроксим (Ceftin®), цефпрозил (Cefzil®), цефаклор (Ceclor®), цефиксим (Suprax®) или цефадроксил (Duricef®)), кларитромицин (например, кларитромицин (Biaxin®)), эритромицин (например, эритромицин (EMycin®)), пенициллин (например, пенициллин V (V-Cillin K® или Pen Vee K®)) или хинолон (например, офлоксацин (Floxin®), ципрофлоксацин (Cipro®) или норфлоксацин (Noroxin®)), аминогликозидные антибиотики (например, апрамицин, арбекацин, бамбермицины, бутиросин, дибекацин, неомицин, неомицин, ундециленат, нетилмицин, паромомицин, рибостамицин, сизомицин и спектиномицин), амфеникольные антибиотики (например, азидамфеникол, хлорамфеникол, флорфеникол и тиамфеникол), ансамициновые антибиотики (например, рифамид и рифампин), карбацефемы (например, лоракабеф), карбапенемы (например, биапенем и имипенем), цефалоспорины (например, цефаклор, цефадроксил, цефандол, цефатризин, цефазедон, цефозопран, цефпимизол, цефпирамид и цефпиром), цефамицины (например, цефбуперазон, цефметазол и цефминокс), монобактамы (например, азтреонам, карумонам и тигемонам), оксацефемы (например, фломоксеф и моксалактам), пенициллины (например, амдиноциллин, амдиноциллин пивоксил, амоксициллин, бакампициллин, бензилпенициллиновую кислоту, бензилпенициллин натрий, эпициллин, фенбенциллин, флоксициллин, пенамциллин, пенетамат гидройодид, пенициллин о-бенэтамин, пенициллин O, пенициллин V, пенициллин V бензатин, пенициллин V гидрабамин, пенимепицилин и фенцигициллин калий), линкозамиды (например, клиндамицин и линкомицин), амфомицин, бацитрацин, капреомицин, колистин, эндурацидин, энвиомицин, тетрациклины (например, апициклин, хлортетрациклин, хломоциклин и демеклоциклин), 2,4-диаминопиримидины (например, бродимоприм), нитрофлираны (например, флиралтадон и фуразолий хлорид), хинолоны и их аналоги (например, циноксацин, клинафлоксацин, флумеквин и грепаглоксацин), сульфонамиды (например, ацетилсульфаметоксипиразин, бензилсульфамид, ноприлсульфамид, фталилсульфацетамид, сульфахризоидин и сульфацитин), сульфоны (например, диатимосульфон, глюкосульфон натрий и соласульфон), циклосерин, мупироцин и туберин.
В определенных вариантах осуществления молекулы по изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или нескольких противогрибковых средств. Противогрибковые средства, которые можно использовать в комбинации с молекулами по изобретению, включают, но не ограничиваются ими, амфотерицин B, итраконазол, кетоконазол, флуконазол, интратекал, флуцитозин, миконазол, бутоконазол, клотримазол, нистатин, терконазол, тиоконазол, циклопирокс, эконазол, галопрогрин, нафтифин, тербинафин, ундециленат и гризеофулдин.
В некоторых вариантах осуществления молекулы по изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или нескольких противовирусных средств. Пригодные противовирусные средства, которые можно использовать в комбинации с молекулами по изобретению, включают, но не ограничиваются ими, ингибиторы протеаз, нуклеозидные ингибиторы обратной транскриптазы, ненуклеозидныые ингибиторы обратной транскриптазы и аналоги нуклеозидов. Примеры противовирусных средств включают, но не ограничиваются ими, зидовудин, ацикловир, ганцикловир, видарабин, идоксуридин, трифлуридин и рибавирин, а также фоскарнет, амантадин, римантадин, саквинавир, индинавир, ампренавир, лопинавир, ритонавир, альфа-интерфероны; адефовир, клевадин, энтекавир, плеконарил.
5.8 ВАКЦИННАЯ ТЕРАПИЯ
Кроме того, изобретение охватывает использование композиции по изобретению для индукции иммунного ответа против антигенного или иммуногенного агента, включая, но не ограничиваясь ими, антигены злокачественной опухоли и антигены инфекционных заболеваний (примеры, которые описаны ниже). Вакцинные композиции по изобретению содержат одно или несколько антигенных или иммуногенных средств, на которые желателен иммунный ответ, где одно или несколько антигенных или иммуногенных средств покрыты вариантом антитела по изобретению, которое обладает увеличенной аффинностью к FcγRIIIA. Вакцинные композиции по изобретению особенно эффективны в отношении индукции иммунного ответа, предпочтительно защитного иммунного ответа, против антигенного или иммуногенного агента.
В некоторых вариантах осуществления антигенный или иммуногенный агент в вакцинных композициях по изобретению содержит вирусный агент, против которого желателен иммунный ответ. Вирусы могут быть рекомбинантными или химерными и предпочтительно являются аттенуированными. Рекомбинантные, химерные и аттенуированные вирусы можно получать с использованием стандартных способов, известных специалисту в данной области. Изобретение охватывает живую рекомбинантную вирусную вакцину или инактивированную рекомбинантную вирусную вакцину для изготовления состава в соответствии с изобретением. Живая вакцина может быть предпочтительной вследствие того, что размножение в хозяине приводит к длительному стимулу сходного типа и величины с тем стимулом, который встречается при природных инфекциях, и, таким образом, обеспечивает устойчивый длительный иммунитет. Получение таких рекомбинантных вирусных вакцинных составов можно проводить с использованием общепринятых способов, вовлекающих размножение вируса в клеточной культуре или в аллантоисе куриного эмбриона с последующей очисткой.
В конкретном варианте осуществления рекомбинантный вирус является непатогенным для индивидуума, которому его вводят. В этом отношении, применение полученных способами генетической инженерии вирусов для вакцинных целей может требовать наличия свойства аттенуации в этих штаммах. Внесение соответствующих мутаций (например, делеций) в матрицы, используемые для трансфекции, может обеспечить новые вирусы со свойствами аттенуации. Например, определенные миссенс-мутации, которые ассоциированы с температурной чувствительностью и адаптацией к холоду, можно преобразовывать в делеционные мутации. Эти мутации должны быть более стабильными, чем точковые мутации, ассоциированные с чувствительными к холоду или температурой мутантами, и частоты реверсии являются крайне низкими. Технологии рекомбинантных ДНК для конструирования рекомбинантных вирусов известны в данной области и охватываются изобретением. Например, способы модификации вирусов с отрицательной РНК-цепью известны в данной области, см., например, патент США № 5166057, который включен в настоящий документ в качестве ссылки в полном объеме.
Альтернативно можно конструировать химерные вирусы со свойством "самоубийства" для применения во внутрикожных вакцинных составах по изобретению. Такие вирусы проходят только один или несколько раундов репликации в хозяине. При использовании в качестве вакцины рекомбинантный вирус проходит через ограниченный цикл(ы) репликации и индуцирует достаточный уровень иммунного ответа, но не пойдет далее в хозяине-человеке и не вызовет заболевание. Альтернативно в соответствии с изобретением можно изготавливать состав с инактивированным (убитым) вирусом. Инактивированные вакцинные составы можно получать с использованием общепринятых способов, чтобы "убить" химерные вирусы. Инактивированные вакцины являются "погибшими" в том смысле, что их инфекционность является нарушенной. В идеальном случае инфекционность вируса нарушается без влияния на его иммуногенность. Для получения инактивированных вакцин химерный вирус можно выращивать в клеточной культуре или в аллантоисе куриного эмбриона, очищать зональным ультрацентрифугированием, инактивировать формальдегидом или β-пропиолактоном и объединять.
В определенных вариантах осуществления в вирус можно встраивать полностью чужеродные эпитопы, включая антигены, происходящие из других вирусных или невирусных патогенов, для применения во внутрикожных вакцинных составах по изобретению. Например, в аттенуированный штамм можно встраивать антигены неродственных вирусов, таких как ВИЧ (gp160, gp120, gp41), антигены паразитов (например, малярии), антигены бактерий или грибов или опухолевые антигены.
Для применения во внутрикожных вакцинных составах в химерные вирусы по изобретению можно встраивать практически любую последовательность гетерологичного гена. Предпочтительно последовательности гетерологичных генов представляют собой структуры и пептиды, которые действуют в качестве модификаторов биологического ответа. Предпочтительно эпитопы, которые индуцируют защитный иммунный ответ на любой из множества патогенов или антигенов, которые связывают нейтрализующие антитела, можно экспрессировать посредством химерных вирусов или в качестве части химерных вирусов. Например, последовательности гетерологичных генов, которые можно встраивать в химерные вирусы по изобретению, включают, но не ограничиваются ими, гемагглютинин и нейраминидазу вируса гриппа или парагриппа и слитые гликопротеины, такие как гены HN и F PIV3 человека. В другом варианте осуществления последовательности гетерологичных генов, которые можно встраивать в химерные вирусы, включают последовательности, которые кодируют белки с иммуномодулирующей активностью. Примеры иммуномодулирующих белков включают, но не ограничиваются ими, цитокины, интерферон типа 1, гамма-интерферон, колониестимулирующие факторы, интерлейкин-1, -2, -4, -5, -6, -12 и антагонисты этих агентов.
В других вариантах осуществления изобретение охватывает патогенные клетки или вирусы, предпочтительно аттенуированные вирусы, которые экспрессируют вариант антитела на их поверхности.
В альтернативных вариантах осуществления вакцинные композиции по изобретению содержат слитый полипептид, где антигенный или иммуногенный агент функционально связан с вариантом антитела по изобретению, который обладает увеличенной аффинностью к FcγRIIIA. Конструирование слитых полипептидов для применения в вакцинных композициях по изобретению проводят с использованием общепринятых способов технологии рекомбинантных ДНК и относится к пределам обычной квалификации.
Кроме того, изобретение охватывает способы индукции толерантности у индивидуума путем введения композиции по изобретению. Предпочтительно композиция, пригодная для индукции толерантности у индивидуума, содержит антигенный или иммуногенный агент, покрытый вариантом антитела по изобретению, где вариант антитела обладает более высокой аффинностью к FcγRIIB. Без связи с конкретным механизмом действия, такие композиции эффективны в отношении индукции толерантности путем активации опосредуемого FcγRIIB ингибиторного каскада.
5.9 КОМПОЗИЦИИ И СПОСОБЫ ВВЕДЕНИЯ
Изобретение относится к способам и фармацевтическим композициям, включающим молекулы по изобретению (т.е. диантитела), содержащие множество связывающих эпитоп доменов и необязательно Fc-домен (или его часть). Также изобретение относится к способам лечения, профилактики и смягчения одного или нескольких симптомов, ассоциированных с заболеванием, нарушением или инфекцией, путем введения индивидууму эффективного количества слитого белка или конъюгированной молекулы по изобретению, или фармацевтической композиции, содержащей слитый белок или конъюгированную молекулу по изобретению. В предпочтительном аспекте антитело, слитый белок или конъюгированная молекула являются по существу очищенными (т.е. по существу свободными от других субстратов, которые ограничивают их эффект или вызывают нежелательные побочные эффекты). В конкретном варианте осуществления индивидуумом является животное, предпочтительно млекопитающее, такое как не примат (например, коровы, свиньи, лошади, кошки, собаки, крысы и т.д.) и примат (например, обезьяна, такая как яванский макак, и человек). В предпочтительном варианте осуществления индивидуумом является человек. В другом предпочтительном варианте осуществления антитело по изобретению происходит из того же вида, что и индивидуум.
Известны различные системы для доставки и их можно использовать для доставки композиции, содержащей молекулы по изобретению, например, инкапсулирование в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать антитело или слитый белок, опосредуемый рецептором эндоцитоз (см., например, Wu et al. (1987) "Receptor-Mediated In Vitro Gene Transformation By A Soluble DNA Carrier System", J. Biol. Chem. 262:4429-4432), конструирование нуклеиновой кислоты в качестве части ретровирусного или другого вектора и т.д. Способы введения молекулы по изобретению включают, но не ограничиваются ими, парентеральное введение (например, внутрикожное, внутримышечное, внутрибрюшинное, внутривенное и подкожное), эпидуральное и через слизистую оболочку (например, интраназальный и пероральный пути). В конкретном варианте осуществления молекулы по изобретению вводят внутримышечно, внутривенно или подкожно. Композиции можно вводить любым общепринятым путем, например, путем инфузии или болюсной инъекции, путем всасывания через эпителиальные или кожно-слизистые выстилки (например, слизистую оболочку полости рта, слизистую оболочку прямой кишки и слизистую оболочку кишечника, и т.д.), и их можно вводить совместно с другими биологически активными средствами. Введение может быть системным или локальным. Кроме того, также можно использовать легочное введение, например с использованием ингалятора или распылителя, и состава с аэрозолизирующего средства. См., например, патенты США №№ 6019968; 5985320; 5985309; 5934272; 5874064; 5855913; 5290540 и 4880078; и публикации PCT WO 92/19244; WO 97/32572; WO 97/44013; WO 98/31346 и WO 99/66903, каждая из которых включена в настоящий документ в качестве ссылки в полном объеме.
Также изобретение относится к тому, что молекулы по изобретению упакованы в герметично закрытый контейнер, такой как ампула или саше, на которых указано количество антитела. В одном из вариантов осуществления молекулы по изобретению предоставляют в качестве сухого стерилизованного лиофилизированного порошка или не содержащего воды концентрата в герметично закрытом контейнере, и его можно восстанавливать, например, водой или физиологическим раствором до соответствующей концентрации для введения индивидууму. Предпочтительно молекулы по изобретению предоставляют в качестве сухого стерильного лиофилизированного порошка в герметично закрытом контейнере в единичной дозировке по меньшей мере 5 мг, более предпочтительно по меньшей мере 10 мг, по меньшей мере 15 мг, по меньшей мере 25 мг, по меньшей мере 35 мг, по меньшей мере 45 мг, по меньшей мере 50 мг или по меньшей мере 75 мг. Лиофилизированные молекулы по изобретению должны храниться при от 2 до 8°C в их исходном контейнере, и молекулы должны быть введены в пределах 12 часов, предпочтительно в пределах 6 часов, в пределах 5 часов, в пределах 3 часов или в пределах 1 часа после восстановления. В альтернативном варианте осуществления молекулы по изобретению предоставляют в жидкой форме в герметично закрытом контейнере, на котором указано количество и концентрация молекулы, слитого белка или конъюгированной молекулы. Предпочтительно жидкие формы молекул по изобретению предоставляют в герметично закрытом контейнере с концентрацией молекул по меньшей мере 1 мг/мл, более предпочтительно по меньшей мере 2,5 мг/мл, по меньшей мере 5 мг/мл, по меньшей мере 8 мг/мл, по меньшей мере 10 мг/мл, по меньшей мере 15 мг/кг, по меньшей мере 25 мг/мл, по меньшей мере 50 мг/мл, по меньшей мере 100 мг/мл, по меньшей мере 150 мг/мл, по меньшей мере 200 мг/мл.
Количество композиции по изобретению, которое является эффективным для лечения, профилактики или смягчения одного или нескольких симптомов, ассоциированных с нарушением, можно определять стандартными клиническими способами. Точная доза, подлежащая применению в составе, также зависит от пути введения и тяжести состояния, и она должна быть установлена в соответствии с решением специалиста и всех обстоятельств пациента. Эффективные дозы можно экстраполировать из кривых доза-эффект из тест-систем in vitro или в моделях на животных.
Для диантител, охватываемых изобретением, дозировка, вводимая пациенту, как правило, составляет от 0,0001 мг/кг до 100 мг/кг массы тела пациента. Предпочтительно дозировка, вводимая пациенту, составляет от 0,0001 мг/кг до 20 мг/кг, от 0,0001 мг/кг до 10 мг/кг, от 0,0001 мг/кг до 5 мг/кг, от 0,0001 до 2 мг/кг, от 0,0001 до 1 мг/кг, от 0,0001 мг/кг до 0,75 мг/кг, от 0,0001 мг/кг до 0,5 мг/кг, от 0,0001 мг/кг до 0,25 мг/кг, от 0,0001 до 0,15 мг/кг, от 0,0001 до 0,10 мг/кг, от 0,001 до 0,5 мг/кг, от 0,01 до 0,25 мг/кг или от 0,01 до 0,10 мг/кг массы тела пациента. Дозировку и частоту введения диантител по изобретению можно снижать или изменять путем усиления захвата и проникновения в ткани диантител с помощью модификаций, например, таких как липидация.
В одном из вариантов осуществления дозировка молекул по изобретению, вводимых пациенту, может составлять от 0,01 мг до 1000 мг/сутки при применении в качестве терапии одним средством. В другом варианте осуществления молекулы по изобретению используют в комбинации с другими терапевтическими композициями, и дозировка, вводимая пациенту, является более низкой, чем когда указанные молекулы используют в качестве терапии одним средством.
В конкретном варианте осуществления может быть желательным вводить фармацевтические композиции по изобретению местно в область, нуждающуюся в лечении; этого можно достигать, например, посредством, и не ограничиваясь этим, местной инфузии, инъекции или с помощью инмплантата, причем указанный имплантат представляет собой пористый, непористый или гелеобразный материал, включая мембраны, такие как силиконовые мембраны или волокна. Предпочтительно, при введении молекулы по изобретению, необходимо уделять внимание тому, чтобы использовать материалы, на которые молекула не адсорбируется.
В другом варианте осуществления композиции можно доставлять в носителе, в частности, в липосоме (см. Langer (1990) "New Methods Of Drug Delivery", Science 249:1527-1533); Treat et al., Liposomes in the Therapy of Infectious Disease and Cancer, Lopez-Berestein and Fidler (eds.), Liss, New York, pp. 353-365 (1989); Lopez-Berestein, там же, pp. 317-327; см., главным образом, там же).
В другом варианте осуществления композиции можно доставлять в системе с контролируемым высвобождением или замедленным высвобождением. Для получения составов с замедленным высвобождением, содержащих одну или несколько молекул по изобретению, можно использовать любой способ, известный специалисту в данной области. См., например, патент США № 4526938; публикацию PCT WO 91/05548; публикацию PCT WO 96/20698; Ning et al. (1996) "Intratumoral Radioimmunotheraphy Of A Human Colon Cancer Xenograft Using A Sustained-Release Gel", Radiotherapy & Oncology 39:179-189, Song et al. (1995) "Antibody Mediated Lung Targeting Of Long-Circulating Emulsions", PDA Journal of Pharmaceutical Science & Technology 50:372-397; Cleek et al. (1997) "Biodegradable Polymeric Carriers For A bFGF Antibody For Cardiovascular Application", Pro. Int'l. Symp. Control. Rel. Bioact. Mater. 24:853-854; и Lam et al. (1997) "Microencapsulation Of Recombinant Humanized Monoclonal Antibody For Local Delivery", Proc. Int'l. Symp. Control Rel. Bioact. Mater. 24:759-760, которые все включены в настоящий документ в качестве ссылок в полном объеме. В одном варианте осуществления в системе с контролируемым высвобождением можно использовать насос (см. Langer, выше; Sefton, (1987) "Implantable Pumps", CRC Crit. Rev. Biomed. Eng. 14:201-240; Buchwald et al. (1980) "Long-Term, Continuous Intravenous Heparin Administration By An Implantable Infusion Pump In Ambulatory Patients With Recurrent Venous Thrombosis", Surgery 88:507-516; and Saudek et al. (1989) "A Preliminary Trial Of The Programmable Implantable Medication System For Insulin Delivery", N. Engl. J. Med. 321:574-579). В другом варианте осуществления для достижения контролируемого высвобождения антител можно использовать полимерные материалы (см., например, Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Florida (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Levy et al. (1985) "Inhibition Of Calcification Of Bioprosthetic Heart Valves By Local Controlled-Release Diphosphonate", Science 228:190-192; During et al. (1989) "Controlled Release Of Dopamine From A Polymeric Brain Implant: In Vivo Characterization", Ann. Neurol. 25:351-356; Howard et al. (1989) "Intracerebral Drug Delivery In Rats With Lesion-Induced Memory Deficits", J. Neurosurg. 7(1): 105-112); патент США № 5679377; патент США № 5916597; патент США № 5912015; патент США № 5989463; патент США № 5128326; публикацию PCT WO 99/15154; и публикацию PCT WO 99/20253). Примеры полимеров, используемых в составах с замедленным высвобождением, включают, но не ограничиваются ими, поли(2-гидроксиэтилметакрилат), поли(метилметакрилат), полиакриловую кислоту, сополимер этилена и винилацетата, полиметакриловую кислоту, полигликолиды (PLG), полиангидриды, поли(N-винилпирролидон), поливиниловый спирт, полиакриламид, полиэтиленгликоль, полилактиды (PLA), сополимеры лактидов и гликолидов (PLGA) и сложные полиортоэфиры. В другом варианте осуществления систему с контролируемым высвобождением можно помещать вблизи мишени лечения (например, легких), что, таким образом, требует только части системной дозы (см., например, Goodson, Medical Applications of Controlled Release, supra, vol. 2, pp. 115-138 (1984)). В другом варианте осуществления полимерные композиции, пригодные в качестве имплантатов с замедленным высвобождением, используют согласно Dunn et al. (см. U.S. 5945155). Этот конкретный способ основан на терапевтическом эффекте контролируемого высвобождения биоактивного материала из полимерной системы in situ. Имплантация, как правило, может происходить в любой области организма пациента, нуждающегося в терапевтическом лечении. В другом варианте осуществления используют неполимерную систему для замедленной доставки, в которой в качестве системы для доставки используют неполимерный имплантат в организм индивидуума. При имплантации в организм органический растворитель имплантата рассеивается, диспергируется или вымывается из композиции в окружающую тканевую жидкость, и неполимерный материал постепенно коагулирует или преципитирует с образованием твердой микропористой матрицы (см. U.S. 5888533).
Системы с контролируемым высвобождением рассмотрены в обзоре Langer (1990, "New "New Methods Of Drug Delivery", Science 249:1527-1533). Для получения составов с замедленным высвобождением, содержащих одно или несколько терапевтических средств по изобретению, можно использовать любой способ, известный специалисту в данной области. См, например, патент США № 4526938; международные публикации WO 91/05548 и WO 96/20698; Ning et al. (1996) "Intratumoral Radioimmunotheraphy Of A Human Colon Cancer Xenograft Using A Sustained-Release Gel", Radiotherapy & Oncology 39:179-189, Song et al. (1995) "Antibody Mediated Lung Targeting Of Long-Circulating Emulsions", PDA Journal of Pharmaceutical Science & Technology 50:372-397; Cleek et al. (1997) "Biodegradable Polymeric Carriers For A bFGF Antibody For Cardiovascular Application", Pro. Int'l. Symp. Control. Rel. Bioact. Mater. 24:853-854; и Lam et al. (1997) "Microencapsulation Of Recombinant Humanized Monoclonal Antibody For Local Delivery", Proc. Int'l. Symp. Control Rel. Bioact. Mater. 24:759-760, которые все включены в качестве ссылки в полном объеме.
В конкретном варианте осуществления, где композиция по изобретению представляет собой нуклеиновую кислоту, кодирующую диантитело по изобретению, нуклеиновую кислоту можно вводить in vivo для стимуляции экспрессии кодируемого ей диантитела, путем конструирования ее в качестве соответствующего экспрессирующего нуклеиновую кислоту вектора и введения ее так, чтобы она становилась внутриклеточной, например, с использованием ретровирусного вектора (см. патент США № 4980286), или путем прямой инъекции или с использованием бомбардировки микрочастицами (например, генная пушка; Biolistic, Dupont), или путем покрытия липидами или рецепторами клеточной поверхности или агентами для трансфекции, или путем введения ее связанной с гомеобокс-подобным пептидом, известным тем, что он проникает в ядро (см. например, Jo Hot et al. (1991) "Antennapedia Homeobox Peptide Regulates Neural Morphogenesis", Proc. Natl. Acad. Sci. USA 88:1864-1868) и т.д. Альтернативно для экспрессии нуклеиновую кислоту можно вводить внутриклеточно и встраивать в ДНК клетки-хозяина путем гомологичной рекомбинации.
Лечение индивидуума терапевтически или профилактически эффективным количеством молекул по изобретению может включать однократное лечение или, предпочтительно, оно может включать серию лечений. В предпочтительном примере индивидуума лечат молекулами по изобретению в диапазоне приблизительно от 0,1 до 30 мг/кг массы тела, один раз в неделю в течение приблизительно от 1 до 10 недель, предпочтительно от 2 до 8 недель, более предпочтительно приблизительно от 3 до 7 недель и еще более предпочтительно в течение приблизительно 4, 5 или 6 недель. В других вариантах осуществления фармацевтические композиции по изобретению вводят один раз в сутки, два раза в сутки или три раза в сутки. В других вариантах осуществления фармацевтические композиции вводят один раз в неделю, два раза в неделю, один раз в две недели, один раз в месяц, один раз каждые шесть недель, один раз каждые два месяца, два раза в год или один раз в год. Также понятно, что эффективную дозировку молекул, используемых для лечения, можно повышать или снижать в ходе конкретного лечения.
5.9.1 ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ
Композиции по изобретению включают бестарные лекарственные композиции, пригодные для изготовления фармацевтических композиций (например, нечистые или нестерильные композиции), и фармацевтические композиции (т.е. композиции, которые пригодные для введения индивидууму или пациенту), которые можно использовать для получения единичных дозированных форм. Такие композиции содержат профилактически или терапевтически эффективное количество профилактического и/или терапевтического средства, описанного в настоящем документе, или комбинацию этих средств и фармацевтически приемлемый носитель. Предпочтительно, композиции по изобретению содержат профилактически или терапевтически эффективное количество одной или нескольких молекул по изобретению и фармацевтически приемлемый носитель.
Также изобретение охватывает фармацевтические композиции, содержащие молекулу диантитела по изобретению и терапевтическое антитело (например, специфичное к опухоли моноклональные антитело), которое специфично к конкретному антигену злокачественной опухоли, и фармацевтически приемлемый носитель.
В конкретном варианте осуществления термин "фармацевтически приемлемый" означает "одобренный контролирующим органом федерального или штатного правительства" или "приведенный в фармакопее США или другой общепринятой фармакопее для применения у животных, и, более конкретно, у человека". Термин "носитель" относится к разбавителю, адъюванту (например, адъюванту Фрейнда (полному и неполному)), эксципиенту или носителю, с которым вводят лекарственное средство. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода и масла, включая масла нефтяного, животного или растительного происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п. Вода является предпочтительным носителем, когда фармацевтическую композицию вводят внутривенно. Также в качестве жидких носителей можно использовать солевые растворы и водные растворы декстрозы и глицерина, в частности, для инъецируемых растворов. Пригодные фармацевтические эксципиенты включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое снятое молоко, глицерин, пропиленгликоль, воду, этанол и т.п. Композиция, если желательно, также может содержать небольшие количества смачивающих веществ или эмульгаторов, или pH-буферных веществ. Эти композиции могут иметь форму растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков, составов с замедленным высвобождением и т.п.
Как правило, ингредиенты композиций по изобретению предоставляют либо по отдельности, либо смешанными вместе в единичной дозированной форме, например, в качестве сухого лиофилизированного порошка и не содержащего воды концентрата в герметично закрытом контейнере, таком как ампула или саше, на которых указано количество активного вещества. Когда композиция предназначена для введения путем инфузии, она может распространяться в бутылях для инфузий, содержащих стерильную фармацевтически пригодную воду или физиологический раствор. Когда композицию вводят путем инъекции, может быть предоставлена ампула стерильной воды для инъекции или физиологического раствора, так чтобы ингредиенты можно было смешивать перед введением.
Композиции по изобретению можно изготавливать в виде нейтральных или солевых форм. Фармацевтически приемлемые соли включают, но не ограничиваются ими, соли, образованные с анионами, такими как анионы, полученные из хлористоводородной, фосфорной, уксусной, щавелевой, виннокаменной кислот и т.д., и соли, образованные с катионами, такими как катионы, образованные натрием, калием, аммонием, кальцием, гидроксидами железа, изопропиламином, триэтиламином, 2-этиламиноэтанолом, гистидином, прокаином и т.д.
5.9.2 ГЕННАЯ ТЕРАПИЯ
В конкретном варианте осуществления нуклеиновые кислоты, содержащие последовательности, кодирующие молекулы по изобретению, вводят для лечения, профилактики или смягчения одного или нескольких симптомов, ассоциированных с заболеванием, нарушением или инфекцией, посредством генной терапии. Генная терапия относится к терапии, проводимой путем введения индивидууму экспрессированной или экспрессируемой нуклеиновой кислоты. В этом варианте осуществления изобретения нуклеиновые кислоты обеспечивают продукцию кодируемого ими антитела или слитого белка, которые опосредует терапевтический или профилактический эффект.
В соответствии с настоящим изобретением можно использовать любые способы генной терапии, доступные в данной области. Иллюстративные способы описаны ниже.
Для общего обзора способов генной терапии см. Goldspiel et al. (1993) "Human Gene Therapy", Clinical Pharmacy 12:488-505; Wu et al. (1991) "Delivery Systems For Gene Therapy", Biotherapy 3:87-95; Tolstoshev (1993) "Gene Therapy, Concepts, Current Trials And Future Directions", Ann. Rev. Pharmacol. Toxicol. 32:573-596; Mulligan (1993) "The Basic Science Of Gene Therapy", Science 260:926-932; и Morgan et al. (1993) "Human Gene Therapy", Ann. Rev. Biochem. 62:191-217. Способы, широко известные в области технологии рекомбинантных ДНК, которые можно использовать, описаны в Ausubel et al. (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, NY (1993); и Kriegler, Gene Transfer and Expression, A Laboratory Manual. Stockton Press, NY (1990).
В предпочтительном аспекте композиция по изобретению содержит нуклеиновые кислоты, кодирующие диантитело по изобретению, причем указанные нуклеиновые кислоты являются частью экспрессирующего вектора, который экспрессирует антитело в пригодном хозяине. В частности, такие нуклеиновые кислоты имеют промоторы, предпочтительно гетерологичные промоторы, функционально связанные с кодирующей областью антитела, причем указанный промотор является индуцибельным или конститутивным и, необязательно, тканеспецифичным. В другом конкретном варианте осуществления используют молекулы нуклеиновой кислоты, в которых кодирующие антитело последовательности и любые другие желаемые последовательности фланкированы областями, которые обеспечивают гомологичную рекомбинацию в желаемой области генома, таким образом, обеспечивая внутрихромосомную экспрессию кодирующих антитело нуклеиновых кислот (Koller et al. (1989) "Inactivating The Beta 2-Microglobulin Locus In Mouse Embryonic Stem Cells By Homologous Recombination", Proc. Natl. Acad. Sci. USA 86:8932-8935; и Zijlstra et al. (1989) "Germ-Line Transmission Of A Disrupted Beta 2-Microglobulin Gene Produced By Homologous Recombination In Embryonic Stem Cells", Nature 342:435-438).
В другом предпочтительном аспекте композиция по изобретению содержит нуклеиновые кислоты, кодирующие слитый белок, причем указанные нуклеиновые кислоты являются частью экспрессирующего вектора, который экспрессирует слитый белок в пригодном хозяине. В частности, такие нуклеиновые кислоты имеют промоторы, предпочтительно гетерологичные промоторы, функционально связанные с кодирующей областью слитого белка, причем указанный промотор является индуцибельным или конститутивным и, необязательно, тканеспецифическим. В другом конкретном варианте осуществления используют молекулы нуклеиновых кислот, в которых кодирующая последовательность слитого белка и любых других желаемых последовательностей фланкирована областями, которые обеспечивают гомологичную рекомбинацию в желаемой области в геноме, таким образом, обеспечивая внутрихромосомную экспрессию слитого белка.
Доставка нуклеиновых кислот индивидууму может быть либо прямой, в случае которой на индивидуума прямо воздействуют нуклеиновой кислотой или несущими нуклеиновую кислоту векторами, либо непрямой, в случае которой клетки сначала трансформируют нуклеиновыми кислотами in vitro, затем трансплантируют индивидууму. Эти подходы известны, соответственно, как генная терапия in vivo или ex vivo.
В конкретном варианте осуществления последовательности нуклеиновых кислот прямо вводят in vivo, где они экспрессируются с образованием кодируемого продукта. Это можно осуществлять любым из множества способов, известных в данной области, например, путем конструирования их в качестве части соответствующего экспрессирующего нуклеиновую кислоту вектора и введения их так, чтобы они становились внутриклеточными, например, путем инфицирования с использованием дефектных или аттенуированных ретровирусных или других вирусных векторов (см. патент США № 4980286), или путем прямой инъекции простой ДНК, или с использованием бомбардировки микрочастицами (например, генная пушка; Biolistic, Dupont), или путем покрытия липидами или рецепторами клеточной поверхности или агентами для трансфекции, инкапсулирования в липосомы, микрочастицы или микрокапсулы, или путем введения их связанными с пептидом, известным тем, что он проникает в ядро, путем введения их связанными с антигеном, подвергаемым рецептор-опосредуемому эндоцитозу (см., например, Wu et al. (1987) "Receptor-Mediated In Vitro Gene Transformation By A Soluble DNA CArrier System", J. Biol. Chem. 262:4429-4432) (который можно использовать для нацеливания на типы клеток, специфично экспрессирующие рецепторы), и т.д. В другом варианте осуществления можно формировать комплексы нуклеиновая кислота-антиген, в которых антиген содержит осуществляющий слияние вирусный пептид для разрушения эндосом, позволяя нуклеиновой кислоте избежать лизосомальной деградации. В другом варианте осуществления нуклеиновая кислота может быть направлена in vivo на клеточно-специфичный захват и экспрессию путем нацеливания на определенный рецептор (см., например, публикации PCT WO 92/06180; WO 92/22635; W092/20316; W093/14188; WO 93/20221). Альтернативно нуклеиновая кислота может быть введена внутриклеточно и может включаться в ДНК клетки-хозяина для экспрессии, посредством гомологичной рекомбинации (Koller et al. (1989) "Inactivating The Beta 2-Microglobulin Locus In Mouse Embryonic stem cells By Homologous Recombination", Proc. Natl. Acad. Sci. USA 86:8932-8935; и Zijlstra et al. (1989) "Germ-Line Transmission Of A Disrupted Beta 2-Microglobulin Gene Produced By Homologous Recombination In Embryonic stem cells", Nature 342:435-438).
В конкретном варианте осуществления используют вирусные векторы, которые содержат последовательности нуклеиновых кислот, кодирующие молекулу по изобретению (например, диантитело или слитый белок). Например, можно использовать ретровирусный вектор (см. Miller et al. (1993) "Use Of Retroviral Vectors For Gene Transfer And Expression", Meth. Enzymol. 217:581-599). Эти ретровирусные векторы содержат компоненты, необходимые для правильного упаковывания вирусного генома и встраивания в ДНК клетки-хозяина. Последовательности нуклеиновых кислот, кодирующие антитело или слитый белок, подлежащие применению в генной терапии, клонируют в один или несколько векторов, которые облегчают доставку нуклеотидной последовательности индивидууму. Более подробное описание ретровирусных векторов может быть найдено в Boesen et al. (1993) "Circumvention Of Chemotherapy-Induced Myelosupression By Transfer Of The Mdr1 Gene", Biotherapy 6:291-302), в которой описано применение ретровирусного вектора для доставки гена mdr1 в гемопоэтические стволовые клетки для того, чтобы сделать стволовые клетки более устойчивыми к химиотерапии. Другие ссылки, иллюстрирующие применение ретровирусных векторов в генной терапии, представляют собой: Clowes et al. (1994) "Long-Term Biological Response Of Injured Rat Carotid Artery Seeded With Smooth Muscle Cells Expressing Retrovirally Introduced Human Genes", J. Clin. Invest. 93:644-651; Keim et al. (1994) "Retrovirus-Mediated Gene Transduction Into Canine Peripheral Blood Repopulating Cells", Blood 83:1467-1473; Salmons et al. (1993) "Targeting Of Retroviral Vectors For Gene Therapy", Human Gene Therapy 4:129-141; и Grossman et al. (1993) "Retroviruses: Delivery Vehicle To The Liver", Curr. Opin. Genetics и Devel. 3:110-114.
Аденовирусы представляют собой другие вирусные векторы, которые можно использовать в генной терапии. Аденовирусы являются особенно привлекательными носителями для доставки генов в респираторный эпителий. Аденовирусы в природе инфицируют респираторный эпителий, где они могут вызывать мягкое заболевание. Другие мишени для систем доставки на основе аденовирусов представляют собой печень, центральную нервную систему, эндотелиальные клетки и мышцы. Аденовирусы имеют преимущество, состоящее в том, что они способны инфицировать неделящиеся клетки. В Kozarsky et al. (1993, "Gene Therapy: Adenovirus Vectors", Current Opinion in Genetics and Development 3:499-503) представлен обзор генной терапии на основе аденовирусов. Bout et al. (1994, "Lung Gene Therapy: In Vivo Adenovirus-Mediated Gene Transfer To Rhesus Monkey Airway Epithelium", Human Gene Therapy, 5:3-10) продемонстрировали применение аденовирусных векторов для переноса генов в респираторный эпителий макак-резус. Другие примеры применения аденовирусов в генной терапии могут быть найдены в Rosenfeld et al. (1991) "Adenovirus-Mediated Transfer Of A Recombinant Alpha 1-Antitrypsin Gene To The Lung Epithelium In Vivo", Science 252:431-434; Rosenfeld et al. (1992) "In Vivo Transfer Of The Human Cystic Fibrosis Transmembrane Conductance Regulator Gene To The Airway Epithelium", Cell 68:143-155; Mastrangeli et al. (1993) "Diversity Of Airway Epithelial Cell Targets For In Vivo Recombinant Adenovirus-Mediated Gene Transfer", J. Clin. Invest. 91:225-234; публикации PCT W094/12649 и Wang et al. (1995) "A Packaging Cell Line For Propagation Of Recombinant Adenovirus Vectors Containing Two Lethal Gene-Region Deletions", Gene Therapy 2:775-783. В предпочтительном варианте осуществления используют аденовирусные векторы.
Также для применения в генной терапии предложен аденоассоциированный вирус (AAV) (см., например, Walsh et al. (1993) "Gene Therapy For Human Hemoglobinopathies", Proc. Soc. Exp. Biol. Med. 204:289-300 и патент США № 5436146).
Другой подход для генной терапии вовлекает перенос гена в клетки в тканевой культуре такими способами, как электропорация, липофекция, опосредуемая фосфатом кальция трансфекция или вирусная инфекция. Обычно способ переноса включает перенос селективного маркера в клетки. Затем клетки подвергают селекции для выделения тех клеток, которые захватываются и экспрессируют перенесенный ген. Затем эти клетки доставляют индивидууму.
В этом варианте осуществления нуклеиновую кислоту встраивают в клетку до введения in vivo полученной рекомбинантной клетки. Такое введение можно проводить любым способом, известным в данной области, включая, но не ограничиваясь ими, трансфекцию, электропорацию, микроинъекцию, инфицирование вирусным или бактериофаговым вектором, содержащим последовательности нуклеиновых кислот, слияние клеток, опосредуемый хромосомами перенос генов, опосредуемый микроклетками перенос генов, слияние сферопластов и т.д. В данной области известно множество способов введения чужеродных генов в клетки (см., например, Loeffler et al. (1993) "Gene Transfer Into Primary And Established Mammalian Cell Lines With Lipopolyamine-Coated DNA", Meth. Enzymol. 217:599-618, Cotten et al. (1993) "Receptor-Mediated Transport Of DNA Into Eucaryotic cells", Meth. Enzymol. 217:618-644) и их можно использовать в соответствии с настоящим изобретением, при условии, что необходимые связанные с развитием и физиологические функции клеток-реципиентов не нарушаются. Способ должен обеспечить стабильный перенос нуклеиновой кислоты в клетку, так чтобы нуклеиновая кислота экспрессировалась клеткой и предпочтительно была наследуемой и экспрессировалась ее потомками.
Полученные рекомбинантные клетки можно доставлять индивидууму различными способами, известными в данной области. Рекомбинантные клетки крови (например, гемопоэтические стволовые клетки или клетки-предшественники) предпочтительно вводят внутривенно. Количество клеток, предусмотренных для применения, зависит от желаемого эффекта, состояния пациента и т.д., и его может определить специалист в данной области.
Клетки, в которые можно вводить нуклеиновую кислоту для целей генной терапии, охватывают любой желаемый доступный тип клеток и включают, но не ограничиваются ими, эпителиальные клетки, эндотелиальные клетки, кератиноциты, фибробласты, мышечные клетки, гепатоциты; клетки крови, такие как T-лимфоциты, B-лимфоциты, моноциты, макрофаги, нейтрофилы, эозинофилы, мегакариоциты, гранулоциты; различные стволовые клетки или клетки-предшественники, в частности, гемопоэтические стволовые клетки или клетки-предшественники, например, получаемые из костного мозга, пуповинной крови, периферической крови, печени эмбриона и т.д.
В предпочтительном варианте осуществления клетка, используемая для генной терапии, является аутологичной для индивидуума.
В варианте осуществления, в котором в генной терапии используют рекомбинантные клетки, последовательности нуклеиновых кислот, кодирующие антитело или слитый белок, вводят в клетки, так чтобы они экспрессировались клетками или их потомством, а затем рекомбинантные клетки вводят in vivo для терапевтического эффекта. В конкретном варианте осуществления используют стволовые клетки или клетки-предшественники. В соответствии с этим вариантом осуществления настоящего изобретения потенциально можно использовать любые стволовые клетки и/или клетки-предшественники, которые можно выделять и поддерживать in vitro (см. например, публикацию PCT WO 94/08598; Stemple et al. (1992) "Isolation Of A Stem Cell For Neurons And Glia From The Mammalian Neural Crest", Cell 71:973-985; Rheinwald (1980) "Serial Cultivation Of Normal Human Epidermal Keratinocytes", Meth. Cell Bio. 21A:229-254; и Pittelkow et al. (1986) "New Techniques For The In Vitro Culture Of Human Skin Keratinocytes And Perspectives On Their Use For Grafting Patients With Extensive Burns", Mayo Clinic Proc. 61:771-777).
В конкретном варианте осуществления нуклеиновая кислота, подлежащая введению для целей генной терапии, содержит индуцибельный промотор, функционально связанный с кодирующей областью, так чтобы экспрессия нуклеиновой кислоты контролировалась присутствием или отсутствием соответствующего индуктора транскрипции.
5.9.3 НАБОРЫ
Изобретение относится к фармацевтической упаковке или набору, содержащим один или несколько контейнеров, заполненных молекулами по изобретению. Кроме того, также в фармацевтическую упаковку или набор может быть включено одно или несколько других профилактических или терапевтических средств, пригодных для лечения заболевания. Также изобретение относится к фармацевтической упаковке или набору, содержащим один или несколько контейнеров, заполненных одним или несколькими ингредиентами фармацевтических композиций по изобретению. Необязательно к такому контейнеру(ам) может прилагаться уведомление в форме, предписанной государственным органом, контролирующим производство, применение или продажу фармацевтических препаратов или биологических продуктов, в котором отражено одобрение государственным органом по изготовлению, применению или продаже для введения человеку.
Настоящее изобретение относится к наборам, которые можно использовать в описанных выше способах. В одном из вариантов осуществления набор содержит одну или несколько молекул по изобретению. В другом варианте осуществления набор, кроме того, содержит одно или несколько других профилактических или терапевтических средств, пригодных для лечения злокачественной опухоли, в одном или нескольких контейнерах. В другом варианте осуществления набор, кроме того, содержит одно или несколько цитотоксических антител, которые связывают один или несколько антигенов злокачественной опухоли, ассоциированных со злокачественной опухолью. В определенных вариантах осуществления другое профилактическое или терапевтическое средство представляет собой химиотерапевтическое средство. В других вариантах осуществления профилактическое или терапевтическое средство представляет собой биологическое или гормональное терапевтическое средство.
5.10 ОХАРАКТЕРИЗАЦИЯ И ДЕМОНСТРАЦИЯ ТЕРАПЕВТИЧЕСКОЙ ПРИМЕНИМОСТИ
Некоторые аспекты фармацевтических композиций, профилактических или лекарственных средств по изобретению предпочтительно тестируют in vitro в отношении желаемой терапевтической активности, в клеточной культуральной системе и модельном организме животного, таком как модельная система на животных-грызунах, перед применением у человека. Например, анализы, которые можно использовать для определения того, является ли желательным введение определенной фармацевтической композиции, включают анализы клеточной культуры, в которых образец ткани пациента выращивают в культуре, и экспонируют или иным образом контактируют с фармацевтической композицией по изобретению, и наблюдают эффект такой композиции на образец ткани. Образец ткани можно получать от пациента путем биопсии. Этот тест позволяет идентификацию терапевтически наиболее эффективной профилактической или терапевтической молекулы (молекул) для каждого отдельного пациента. В различных конкретных вариантах осуществления анализы in vitro можно проводить с иллюстративными типами клеток, вовлеченными в аутоиммунное или воспалительное нарушение (например, T-клетки), для определения того, обладает ли фармацевтическая композиция по изобретению желаемым эффектом на такие типы клеток.
Комбинации профилактических и/или терапевтических средств перед применением у человека можно тестировать в пригодных модельных системах на животных. Такие модельные системы на животных включают, но не ограничиваются ими, крыс, мышей, кур, коров, обезьян, свиней, собак, кроликов и т.д. Можно использовать любую животную систему, хорошо известную в данной области. В конкретном варианте осуществления изобретения комбинации профилактических и/или терапевтических средств тестируют в модельной системе на мышах. Такие модельные системы широко используют и они хорошо известны квалифицированному специалисту. Профилактические и/или терапевтические средства можно вводить многократно. Некоторые аспекты этой процедуры могут варьировать. Указанные аспекты включают временный режим введения профилактических и/или терапевтических средств, и то, вводят ли такие средства по отдельности или в смеси.
Предпочтительные модели на животных для применения в способах по изобретению представляют собой, например, трансгенных мышей, экспрессирующих FcγR человека на эффекторных клетках мыши, например, в настоящем изобретении можно использовать любую модель, описанную в U.S. 5877396 (который включен в настоящий документ в качестве ссылки в полном объеме). Трансгенные мыши для применения в способах по изобретению включают, но не ограничиваются ими, мышей, имеющих FcγRIIIA человека; мышей, имеющих FcγRIIA человека; мышей, имеющих FcγRIIB человека и FcγRIIIA человека; мышей, имеющих FcγRIIB человека и FcγRIIA человека. Предпочтительно мутации, демонстрирующие наиболее высокие уровни активности в функциональных анализах, описанных выше, тестируют в отношении применения в модельных исследованиях на животных перед применением у человека. Достаточные количества антител можно получать для применения в моделях на животных с использованием способов, описанных выше, например, с использованием экспрессирующих систем млекопитающих и способов очистки, описанных и проиллюстрированных в настоящем документе.
Для исследования эффективности антител мыши, полученных против опухолеспецифической мишени, можно использовать модели на мышах с ксенотрансплантатами, исходя из аффинности и специфичности связывающих эпитоп доменов молекулы диантитела по изобретению и способности диантитела индуцировать иммунный ответ (Wu et al. (2001) "Mouse Models For Multistep Tumorigenesis", Trends Cell Biol. 11:S2-9). Трансгенные мыши, экспрессирующие FcγR человека на эффекторных клетках мыши, являются уникальными и представляют собой специализированные модели на животных для тестирования эффективности взаимодействий Fc-FcγR человека. Можно использовать пары трансгенных по FcγRIIIA, FcγRIIIB и FcγRIIA линий мышей, полученных в лаборатории Dr. Jeffrey Ravetch (путем лицензионного соглашения с Rockefeller U. и Sloan Kettering Cancer center), такие как линии, приведенные в таблице 11 ниже.
Линии мышей
Противовоспалительную активность комбинированных способов лечения по изобретению можно определять с использованием различных экспериментальных моделей воспалительного артрита на животных, известных в данной области и описанных в Crofford L.J. and Wilder R.L., "Arthritis and Autoimmunity in Animals", Arthritis and Allied Conditions: A Textbook of Rheumatology, McCarty et al.(eds.), Chapter 30 (Lee and Febiger, 1993). Также для оценки противовоспалительной активности комбинированных способов лечения по изобретению можно использовать экспериментальные и спонтанные модели воспалительного артрита и аутоиммунных ревматических заболеваний на животных. Ниже в качестве неограничивающих примеров представлены некоторые анализы.
Основные модели на животных для артрита или воспалительного заболевания известны в данной области, и широко используемые модели включают: модели индуцируемого адъювантом артрита на крысах, модели индуцируемого коллагеном артрита на крысах и мышах и модели индуцируемого антигеном артрита на крысах, кроликах и хомяках, все из которых описаны в Crofford L.J. and Wilder R.L., "Arthritis and Autoimmunity in Animals", Артрит and Allied Conditions: A Textbook of Rheumatology, McCarty et al.(eds.), Chapter 30 (Lee and Febiger, 1993), включенной в настоящий документ в качестве ссылки в полном объеме.
Противовоспалительную активность комбинированных способов лечения по изобретению можно оценивать с использованием модели индуцируемого каррагенаном артрита на крысах. Индуцируемый каррагенаном артрит также используют у кроликов, собак и свиней при исследовании хронического артрита или воспаления. Для оценки терапевтической эффективности используют количественную гистоморфометрическую оценку. Способы с использованием такой модели индуцируемого каррагенаном артрита описаны в Hansra P. et al. (2000) "Carrageenan-induced Arthritis In The Rat", Inflammation, 24(2):141-155. Также широко используют модели индуцируемого зимосаном воспаления на животных, как известно и описано в данной области.
Противовоспалительную активность комбинированных способов лечения по изобретению также можно оценивать путем измерения ингибирования индуцируемого каррагенаном отека лапы у крыс с использованием модификации способа, описанного в Winter C.A. et al. (1962) "Carrageenan-induced Edema In Hind Paw Of The Rat As An Assay For Anti-Inflammatory Drugs", Proc. Soc. Exp. Biol Med. 111, 544-547. Этот анализ использовали в качестве первичного скрининга in vivo противовоспалительной активности большинства NSAID, и он считается прогнозирующим эффективность у человека. Противовоспалительную активность тестируемых профилактических или терапевтических средств выражают в качестве процентного ингибирования увеличения массы задней лапы в тестируемой группе относительно контрольной группы, в которую вводили носитель.
Кроме того, также для оценки эффективности комбинированных способов лечения по изобретению можно использовать модели воспалительного заболевания кишечника на животных (Kim et al. (1992) "Experimental Colitis In Animal Models", Scand. J. Gastroentrol. 27:529-537; Strober (1985) "Animal Models Of Inflammatory Bowel Disease - An Overview", Dig. Dis. Sci. 30 (12 Suppl):3S-10S). Язвенный колит и болезнь Крона представляют собой воспалительные заболевания кишечника человека, которые можно индуцировать у животных. Для индукции воспалительных заболеваний кишечника животным перорально можно вводить сульфатированные полисахариды, включая, но не ограничиваясь ими, амилопектин, карраген, сульфат амилопектина и декстрансульфат или химические раздражители, включая, но не ограничиваясь ими, тринитробензолсульфоновую кислоту (TNBS) и уксусную кислоту.
Модели аутоиммунных нарушений на животных также можно использовать для оценки эффективности комбинированных способов лечения по изобретению. Разработаны модели на животных для аутоиммунных нарушений, таких как диабет 1 типа, аутоиммунное заболевание щитовидной железы, системная красная волчанка и гломерулонефрит (Flanders et al. (1999) "Prevention Of Type 1 Diabetes From Laboratory To Public Health", Autoimmunity 29:235-246; Rasmussen et al. (1999) "Models To Study The Pathogenesis Of Thyroid Autoimmunity", Biochimie 81:511-515; Foster (1999) "Relevance Of Systemic Lupus Erythematosus Nephritis Animal Models To Human Disease", Semin. Nephrol. 19:12-24).
Кроме того, для оценки профилактической и/или терапевтической применимости комбинированных способов лечения, описанных в настоящем документе для аутоиммунных и/или воспалительных заболеваний, можно использовать любые анализы, известные специалистам в данной области.
Токсичность и эффективность профилактических и/или терапевтических протоколов по настоящему изобретению можно определять с помощью стандартных фармацевтических способов в клеточных культурах или у экспериментальных животных, например, для определения LD50 (доза, летальная для 50% популяции) и ED50 (доза, терапевтически эффективная для 50% популяции). Соотношение доз между токсическими и терапевтическими эффектами представляет собой терапевтический индекс и его можно выражать в качестве соотношения LD50/ED50. Предпочтительными являются профилактические и/или терапевтические средства, которые проявляют высокие терапевтические индексы. Хотя можно использовать профилактические и/или терапевтические средства, которые проявляют токсические побочные эффекты, следует уделить внимание тому, чтобы создать систему для доставки, которая нацеливает такие средства в область пораженной ткани для минимизации потенциального повреждения неинфицированных клеток и, тем самым, снижения побочных эффектов.
При изготовлении ряда дозировок профилактических и/или терапевтических средств для применения у человека можно использовать данные, полученные из анализов клеточных культур и исследований на животных. Дозировка таких средств предпочтительно находится в диапазоне концентраций в кровотоке, которые включают ED50 с небольшой токсичностью или без токсичности. Дозировка может варьировать в этом диапазоне, в зависимости от используемой дозированной формы и используемого пути введения. Для любого средства, используемого в способе по изобретению, терапевтически эффективная доза может быть первоначально оценена из анализов в клеточной культуре. Дозу можно изготавливать в моделях для животных так, чтобы достигнуть диапазона циркулирующих концентраций в плазме, который включает IC50 (т.е. концентрацию тестируемого соединения, которая достигает полумаксимального ингибирования симптомов) при определении в клеточной культуре. Такую информацию можно использовать для более точного определения пригодных доз у человека. Уровни в плазме можно измерять, например, высокоэффективной жидкостной хроматографией.
Активность средств, используемых в соответствии с настоящим изобретением, в отношении злокачественной опухоли также можно определять с использованием различных экспериментальных моделей на животных для исследования злокачественной опухоли, таких как модель на мышах SCID или трансгенных мышах или мышах nude с ксенотрансплантатами человека, в моделях на животных, таких как хомяки, кролики и т.д., известных в данной области и описанных в Relevance of Tumor Models for Anticancer Drug Development (1999, eds. Fiebig and Burger); Contributions to Oncology (1999, Karger); The Nude Mouse in Oncology Research (1991, eds. Boven and Winograd); и Anticancer Drug Development Guide (1997 ed. Teicher), включенных в настоящий документ в качестве ссылок в полном объеме.
Предпочтительные модели на животных для определения терапевтической эффективности молекул по изобретению представляют собой модели на мышах с ксенотрансплантатами. Опухолевые клеточные линии, которые можно использовать в качестве источника ксенотрансплантированных опухолей, включают, но не ограничиваются ими, клетки SKBR3 и MCF7, которые могут быть получены от пациентов с аденокарциномой молочной железы. Эти клетки имеют как рецепторы erbB2, так и рецепторы пролактина. Клетки SKBR3 широко используют в данной области в качестве моделей ADCC и моделей ксенотрансплантированных опухолей. Альтернативно в качестве источника ксенотрансплантированных опухолей можно использовать клетки OVCAR3, происходящие из аденокарциномы яичника человека.
Протоколы и композиции по изобретению предпочтительно тестируют in vitro, а затем in vivo, в отношении желаемой терапевтической или профилактической активности, перед применением у человека. Лекарственные средства и способы можно подвергать скринингу с использованием клеток опухоли или злокачественной клеточной линии. Для оценки такого выживания и/или роста можно использовать многие анализы, стандартные в данной области; например, пролиферацию клеток можно анализировать путем измерения включения 3H-тимидина, путем прямого подсчета клеток, путем детекции изменений транскрипционной активности известных генов, таких как протоонкогены (например, fos, myc) или маркеров клеточного цикла; жизнеспособность клеток можно оценивать путем окрашивания трипановым синим, дифференцировку можно визуально оценивать на основании изменений морфологии, снижения роста и/или образования колоний в мягком агаре или структуре трубчатой сети в трехмерном препарате базальной мембраны или внеклеточного матрикса и т.д.
Соединения для применения в терапии можно тестировать в пригодных модельных системах на животных перед тестированием у человека, включая, но не ограничиваясь ими, крыс, мышей, кур, коров, обезьян, кроликов, хомяков и т.д., например, в моделях на животных, описанных выше. Затем соединения можно использовать в соответствующих клинических испытаниях.
Кроме того, для оценки профилактической и/или терапевтической применимости комбинированных способов лечения, описанных в настоящем документе, для лечения или профилактики злокачественной опухоли, воспалительного нарушения или аутоиммунного заболевания можно использовать любые анализы, известные специалистам в данной области.
6. ПРИМЕРЫ
6.1 КОНСТРУИРОВАНИЕ И ОХАРАКТЕРИЗАЦИЯ КОВАЛЕНТНЫХ БИСПЕЦИФИЧЕСКИХ ДИАНТИТЕЛ
Моноспецифическое ковалентное диантитело и биспецифическое ковалентное диантитело конструировали для оценки рекомбинантной продукции, очистки и характеристик связывания каждого из них. В анализах SDS-PAGE и SEC было выявлено, что аффинно очищенные молекулы диантител, которые продуцировали с помощью рекомбинантных экспрессирующих систем, описанных в настоящем документе, состоят из одного типа димера. Кроме того, анализ ELISA и SPR выявил, что ковалентное биспецифическое диантитело проявляло аффинность к обоим антигенам-мишеням и могло связывать оба антигена одновременно.
Материалы и способы:
Конструирование и разработка полипептидных молекул: Экспрессирующие нуклеиновую кислоту векторы конструировали так, чтобы продуцировались четыре полипептидных конструкции, схематично представленные на ФИГ. 2. Конструкция 1 (SEQ ID NO: 9) содержала VL-домен гуманизированного антитела 2B6, который распознает FcγRIIB, и VH-домен гуманизированного антитела 3G8, который распознает FcγRIIIA. Конструкция 2 (SEQ ID NO: 11) содержала VL-домен Hu3G8 и VH-домен Hu2B6. Конструкция 3 (SEQ ID NO: 12) содержала VL-домен Hu3G8 и VH-домен Hu3G8. Конструкция 4 (SEQ ID NO: 13) содержала VL-домен Hu2B6 и VH-домен Hu2B6.
ПЦР и конструирование экспрессирующих векторов: Кодирующие последовательности VL- или VH-доменов амплифицировали с матричной ДНК с использованием прямого и обратного праймеров, сконструированных так, чтобы исходные продукты ПЦР содержали перекрывающиеся последовательности, обеспечивая получение в ПЦР с перекрыванием кодирующих последовательностей для желаемых полипептидных конструкций.
Первичная ПЦР-амплификация ДНК-матрицы: Приблизительно 35 нг ДНК-матрицы, например, легкой цепи и тяжелой цепи представляющего интерес антитела; 1 мкл 10 мкМ прямого и обратного праймеров; 2,5 мкл 10x буфера pfuUltra (Stratagene, Inc.); 1 мкл 10 мМ dNTP; 1 мкл 2,5 единиц/мкл ДНК-полимеразы pfuUltra (Stratagene, Inc.); и дистиллированную воду до общего объема 25 мкл осторожно смешивали в микроцентрифужной пробирке и кратковременно центрифугировали в микроцентрифуге для скапливания реакционной смеси на дне пробирки. Реакции ПЦР проводили с использованием системы для ПЦР GeneAmp PCR System 9700 (PE Applied Biosystem) и следующих условий: 94°С, 2 минуты; 25 циклов по 94°С, каждый по 15 секунд; 58°С, 30 секунд; и 72°С, 1 минута.
VL Hu2B6 амплифицировали с легкой цепи Hu2B6 с использованием прямого и обратного праймеров SEQ ID NO: 55 и SEQ ID NO: 56, соответственно. VH Hu2B6 амплифицировали с тяжелой цепи Hu2B6 с использованием прямого и обратного праймеров SEQ ID NO: 57 и SEQ ID NO: 58, соответственно. VL Hu3G8 амплифицировали с легкой цепи Hu3G8 с использованием прямого и обратного праймеров SEQ ID NO: 55 и SEQ ID NO: 59, соответственно. VH Hu3G8 амплифицировали с тяжелой цепи Hu3G8 с использованием прямого и обратного праймеров SEQ ID NO: 60 и SEQ ID NO: 61, соответственно.
Продукты ПЦР подвергали электрофорезу в 1% агарозном геле в течение 30 минут при 120 вольтах. Продукты ПЦР вырезали из геля и очищали с использованием набора для экстракции из геля MinElute GEl Extraction Kit (Qiagen, Inc.).
ПЦР с перекрыванием: Продукты первичной ПЦР объединяли, как описано ниже, и амплифицировали с использованием тех же условий ПЦР, которые описаны для первичной амплификации ДНК-матрицы. Продукты ПЦР с перекрыванием также очищали, как описано выше.
Последовательность нуклеиновой кислоты, кодирующую конструкцию, SEQ ID NO: 9 (схематично представленную на ФИГ. 2), амплифицировали путем объединения продуктов амплификаций посредством ПЦР VL Hu2B6 и VH Hu3G8, и прямого и обратного праймеров SEQ ID NO: 55 и SEQ ID NO: 61, соответственно. Последовательность нуклеиновой кислоты, кодирующую конструкцию 2, SEQ ID NO: 11 (схематично представленную на ФИГ. 2), амплифицировали, объединяя продукты ПЦР после амплификаций VL Hu3G8 и VH Hu2B6, и прямой и обратный праймеры SEQ ID NO: 55 и SEQ ID NO: 58, соответственно. Последовательности нуклеиновых кислот, кодирующие конструкцию 3, SEQ ID NO: 12 (схематично представленную на ФИГ. 2), амплифицировали, объединяя продукты ПЦР после амплификаций VL Hu3G8 и VH Hu3G8, и прямой и обратный праймеры SEQ ID NO: 55 и SEQ ID NO: 61, соответственно. Последовательность нуклеиновой кислоты, кодирующую конструкцию 4, SEQ ID NO: 13 (схематично представленную на ФИГ. 2), амплифицировали, объединяя продукты ПЦР после амплификаций VL Hu2B6 и VH Hu2B6, и прямой и обратный праймеры SEQ ID NO: 55 и SEQ ID NO: 58, соответственно.
Прямые праймеры для VL-доменов (т.е. SEQ ID NO: 55) и обратные праймеры для VH-доменов (т.е. SEQ ID NO: 58 и SEQ ID NO: 61) содержали уникальные участки рестрикции для обеспечения клонирования конечного продукта в экспрессирующий вектор. Очищенные продукты ПЦР с перекрыванием расщепляли эндонуклеазами рестрикции Nhe I и EcoR I и клонировали в экспрессирующий вектор млекопитающих pCIneo (Promega, Inc.). Плазмиды, кодирующие конструкции, были обозначены так, как указано в таблице 12:
ПЛАЗМИДНЫЕ КОНСТРУКЦИИ
Экспрессия полипептида/диантитела: pMGX0669, кодирующий конструкцию 1, котрансфицировали с pMGX0667, кодирующим конструкцию 2, в клетки HEK-293 с использованием Lipofectamine 2000 согласно инструкциям изготовителя (Invitrogen). Котрансфекция этих двух плазмид была предназначена для экспрессии ковалентного биспецифического диантитела (CBD), иммуноспецифичного как к FcγRIIB, так и к FcγRIIIA (диантитело h2B6-h3G8). pMGX0666 и pMGX0668, кодирующие конструкции 3 и 4, соответственно, по отдельности трансфицировали в клетки HEK-293 для экспрессии ковалентного моноспецифического диантитела (CMD), иммуноспецифичного к FcγRIIIA (диантитело h3G8) и FcγRIIB (диантитело h2B6), соответственно. После трех дней в культуре секретируемые продукты очищали из кондиционированной среды.
Очистка: Диантитела улавливали из кондиционированной среды с использованием соответствующих антигенов, связанных с Sepharose 4B, активированной CNBr. Аффинную смолу Sepharose уравновешивали в 20 мМ Tris/HCl, pH 8,0, перед загрузкой. После загрузки перед элюированием смолу промывали буфером для уравновешивания. Диантитела элюировали с промытой смолы с использованием 50 мМ глицина, pH 3,0. Элюированные диантитела сразу нейтрализовывали 1 M Tris/HCl, pH 8,0, и концентрировали с использованием концентратора центрифужного типа. Концентрированные диантитела далее очищали эксклюзионной хроматографией с использованием колонки Superdex 200, уравновешенной в PBS.
SEC: Эксклюзионную хроматографию использовали для анализа приблизительного размера и гетерогенности диантител, элюированных с колонки. Анализ SEC проводили на колонке GE healthcare Superdex 200HR 10/30, уравновешенной PBS. В качестве контролей использовали сравнение с профилями элюирования полноразмерного IgG (~150 кДа), Fab-фрагмента (~50 кДа) и одноцепочечного Fv (~30 кДа).
ELISA: Связывание элюированных и очищенных диантител охарактеризовывали с помощью анализа ELISA, как описано в 5.4.2. 50 мкл/лунка раствора sCD32B-Ig в концентрации 2 мкг/мл наносили на 96-луночный планшет Maxisorp в карбонатном буфере при 4°С в течение ночи. Планшет промывали три раза посредством PBS-T (PBS, 0,1% Tween 20) и блокировали 0,5% BSA в PBS-T в течение 30 минут при комнатной температуре. После этого CBD h2B6-h3G8, CMD h2B6 или CMD h3G8 разбавляли в блокирующем буфере в серийных двукратных разведениях с получением диапазона концентраций диантител от 0,5 мкг/мл до 0,001 мкг/мл. Затем планшет инкубировали при комнатной температуре в течение 1 часа. После промывания PBS-T три раза в каждую лунку добавляли 50 мкл/лунка sCD16A-биотина в концентрации 0,2 мкг/мл. Планшет вновь инкубировали при комнатной температуре в течение 1 часа. После промывания три раза PBS-T, для детекции использовали 50 мкл/лунка конъюгированного с HRP стрептавидина в разведении 1:5000 (Amersham Pharmacia Biotech). HRP-стрептавидину позволяли инкубироваться в течение 45 минут при комнатной температуре. Планшет три раза промывали PBS-T и проявляли с использованием 80 мкл/лунка субстрата TMB. После инкубации в течение 10 минут реакцию HRP-TMB останавливали добавлением 40 мкл/лунка 1% H2SO4. Считывание OD450 нм проводили с использованием 96-луночного устройства для считывания планшетов и программного обеспечения SOFTmax, и результаты наносили на график с использованием программного обеспечения GraphPadPrism 3.03.
Анализ BIAcore: Кинетические параметры связывания элюированных и очищенных диантител анализировали с использованием анализа BIAcore (устройство BIAcore 1000, BIAcore Inc., Piscataway, N.J.) и прилагаемого к нему программного обеспечения, как описано в разделе 5.4.3.
sCD16A, sCD32B или sCD32A (отрицательный контроль) иммобилизовывали на одной из четырех проточных ячеек (проточная ячейка 2) поверхности сенсорного чипа путем реакции связывания аминов (посредством модификации карбоксиметильных групп смесью NHS/EDC), так чтобы приблизительно 1000 единиц ответа (RU) каждого рецептора было иммобилизовано на поверхности. После этого не вступившие в реакцию активные сложные эфиры "кэпировали" путем инъекции 1 M Et-NH2. После приготовления пригодной поверхности ковалентные биспецифические диантитела (h2B6-h3G8 CBD) или ковалентные моноспецифические диантитела (h2B6 CMD или h3G8 CMB) пропускали над поверхностью посредством инъекций в течение 180 секунд 6,25-200 нМ раствора при скорости потока 70 мл/мин. Также для сравнения тестировали h3G8 scFV.
После сбора полного набора данных полученные кривые связывания подвергали глобальной аппроксимации с использованием компьютерных алгоритмов, предоставляемых изготовителем, BIAcore, Inc. (Piscataway, NJ). Эти алгоритмы вычисляют как Kon, так и Koff, из которых устанавливают кажущуюся константу равновесия KD в качестве соотношения двух констант скорости (т.е. Koff/Kon). Более подробное описание того, как получают отдельные константы скоростей, может быть найдено в BIAevaluaion Software Handbook (BIAcore, Inc., Piscataway, NJ).
Фазы ассоциации и диссоциации аппроксимировали по отдельности. Константу скорости диссоциации получали для интервала 32-34 с фазой диссоциации 180 с; аппроксимацию фазы ассоциации проводили с помощью модели Ленгмюра 1:1, и основную аппроксиацию выбирали, исходя из Rmax и критериев chi2 для биспецифических диантител и scFv; для связывания CMD использовали двухвалентную аппроксимацию анализируемого образца.
Результаты
Анализ SDS-PAGE в невосстанавливающих условиях показал, что каждый очищенный продут экспрессирующих систем CMD h3G8, CMD h2B6 и CBD h2B6-h3G8 был отдельным типом с оцененной молекулярной массой приблизительно 50 кДа (ФИГ. 3, дорожки 4, 5 и 6, соответственно). В восстанавливающих условиях продукт, очищенный из любой из экспрессирующих систем CMD, мигрирует в качестве отдельной полосы (дорожки 1 и 2), в то время как было выявлено, что продукт, очищенный из системы CBD h2B6-h3G8, представлял собой 2 отдельных белка (ФИГ. 3, дорожка 3). Все полипептиды, очищенные из экспрессирующей системы и визуализированные посредством SDS-PAGE, в восстанавливающих условиях мигрировали до уровня приблизительно 28 кДа.
Анализ SEC каждого из продуктов экспрессирующих систем также продемонстрировал отдельные молекулярные типы (ФИГ. 4B), каждый из которых элюировался в то же время, что и Fab-фрагмент IgG (~50 кДа) (ФИГ. 4A). Результаты указывают на то, что подвергнутый аффинной очистке продукт представлял собой гомогенный ковалентный гомодимер в случае экспрессирующей системы CMD и гомогенный ковалентный гетеродимер в случае CBD h2B6-h3G8.
Сэндвич-анализ ELISA использовали для тестирования специфичности связывания CBD h2B6-h3G8 с любым из или с обоими из CD32B и/или CD16A (ФИГ. 5). CD32B служил в качестве антигена-мишени, и CD16A использовали в качестве вторичного зонда. Положительный сигнал в ELIZA показал, что гетеродимерное CBD h2B6-h3G8 обладал специфичностью к обоим антигенам. Сходное тестирование CMD h3G8 (который не должен связывать CD32B) продемонстрировало отсутствие сигнала.
Анализ SPR показал, что CMD h3G8 иммуноспецифично распознает sCD16, но не sCD32B, что CMD h2B6 иммуноспецифично распознает sCD32B, но не sCD16, и что CBD h2B6-h3G8 иммуноспецифично распознает как sCD16, так и sCD32B (ФИГ. 6A-B). Ни одно из протестированных диантител не связывало контрольный рецептор sCD32A (ФИГ. 6C).
Анализ SPR также использовали для оценки кинетических и равновесных констант CMD и CBD h2B6-h3G8 в отношении sCD16 и/или sCD32B. Результаты сравнивали с теми же константами, которые были вычислены для scFV h3G8. На ФИГ. 7A-E представлены графические результаты анализа SPR. Кинетические константы ассоциации и диссоциации, а также равновесная константа, вычисленная из результатов, представленных на ФИГ. 7, представлены в таблице 13.
Кинетические и равновесные константы, вычисленные из данных BIAcore
В совокупности с результатами анализа ELISA исследования подтверждают, что ковалентный гетеродимер h2B6-h3G8 сохраняет специфичность в отношении как CD32B, так и CD16, и способен связывать оба антигена одновременно. Эта молекула схематично представлена на ФИГ. 8.
6.2 КОНСТРУИРОВАНИЕ И ОХАРАКТЕРИЗАЦИЯ КОВАЛЕНТНЫХ БИСПЕЦИФИЧЕСКИХ ДИАНТИТЕЛ, СОДЕРЖАЩИХ FC-ДОМЕНЫ
В попытках создать IgG-подобную молекулу, т.е. содержащую Fc-домен, один из полипептидов, содержащих гетеродимерную молекулу CBD, представленную в примере 6.1, модифицировали, чтобы он дополнительно содержал Fc-домен (создавая "более тяжелую" и "более легкую цепь", аналогичную тяжелой и легкой цепям антитела). После этого гетеродимерная биспецифическая молекула содержала Fc-домен, который димеризуется с гомологичной молекулой, образуя четырехвалентную тетрамерную IgG-подобную молекулу (т.е. образованную димеризацией через Fc-домены гетеродимерных биспецифических молекул). Интересно, что такие тетрамерные молекулы не выявлялись в кондиционированной среде рекомбинантных экспрессирующих систем с использованием функциональных анализов, например, тестирования кондиционированной среды в отношении иммуноспецифического связывания с антигенами-мишенями. Вместо этого, в таких функциональных анализах выявлялась только димерная молекула, содержащая мономеры, состоящие из VL, VH и Fc-домена. Для тестирования того, нарушается ли стабильность теоретической тетрамерной молекулы, полипептиды, содержащие Fc-домен, конструировали, чтобы они дополнительно содержали шарнирную область, в то время как полипептиды, содержащие "более легкую" цепь, конструировали так, чтобы они дополнительно содержали 6 C-концевых аминокислот константного домена легкой цепи каппа человека. Когда такие перестроенные "более тяжелая" и "более легкая" цепи коэкспрессировали в рекомбинантных экспрессирующих системах, в функциональных анализах детектировались молекулы диантител, которые были способны иммуноспецифично связывать как антигены-мишени, так и антитела против Fc.
Материалы и способы
Конструирование и разработка полипептидных молекул: Экспрессирующие нуклеиновую кислоту векторы конструировали, чтобы они продуцировали модифицированные варианты конструкций 1 и 2, представленных в примере 6.1. Конструкции 5 (SEQ ID NO: 14) и 6 (SEQ ID NO: 15) создавали модификацией способами инженерии конструкций 1 и 2, соответственно, чтобы они дополнительно содержали Fc-домен. Конструкцию 7 (SEQ ID NO: 16) создавали модификацией способами инженерии конструкции 1, чтобы она дополнительно содержала последовательность FNRGEC (SEQ ID NO: 23) на ее C-конце. Конструкцию 8 (SEQ ID NO: 18) создавали модификацией способами инженерии конструкции 2, чтобы она дополнительно содержала шарнирную область и Fc-домен (содержащий мутацию V215A). Схематическое представление конструкций 5-8 представлено на ФИГ. 9.
ПЦР и конструирование экспрессирующего вектора: Все протоколы ПЦР и очистки продуктов ПЦР были такими, как описано в примере 6.1. Плазмиды pMGX0669 и pMGX0667 служили в качестве матриц для кодирующих последовательностей для конструкций 1 и 2, соответственно. Кодирующие последовательности для Fc-домена и/или шарнирного домена HuIgG представляли собой SEQ ID NO: 5 или SEQ ID NO: 1 и SEQ ID NO: 5, соответственно. Кодирующие последовательности для матричных ДНК амплифицировали с использованием прямого и обратного праймеров, так чтобы продукты ПЦР содержали перекрывающиеся последовательности, позволяя ПЦР с перекрыванием образовывать кодирующие последовательности для желаемых продуктов.
Кодирующую последовательность конструкции 1 амплифицировали с pMGX0669 с использованием прямого и обратного праймеров SEQ ID NO: 55 и SEQ ID NO: 62, соответственно. Кодирующую последовательность конструкции 2 амплифицировали с pMGX0667 с использованием прямого и обратного праймеров SEQ ID NO: 55 и SEQ ID NO: 63, соответственно. Шарнинрую область HuIgG-Fc амплифицировали с использованием прямого и обратного праймеров SEQ ID NO: 65 и SEQ ID NO: 66, соответственно. Конструкцию 7 (SEQ ID NO: 16) амплифицировали с pMGX0669 с использованием прямого и обратного праймеров SEQ ID NO: 55 и SEQ ID NO: 67.
ПЦР с перекрыванием. Продукты первичной ПЦР объединяли, как описано ниже, амплифицировали и очищали, как описано в примере 6.1.
Последовательность нуклеиновой кислоты, кодирующую конструкцию 5, SEQ ID NO: 14 (схематично представленную на ФИГ. 9), амплифицировали, объединяя продукты ПЦР после амплификаций конструкции 1 и HuIgG Fc, и прямого и обратного праймеров SEQ ID NO: 55 и SEQ ID NO: 64, соответственно. Последовательность нуклеиновой кислоты, кодирующую конструкцию 6, SEQ ID NO: 15 (схематично представленную на ФИГ. 9), амплифицировали, объединяя продукты ПЦР после амплификаций конструкции 2 и HuIgG Fc, и прямого и обратных праймеров SEQ ID NO: 55 и SEQ ID NO: 66, соответственно. Последовательность нуклеиновой кислоты, кодирующую конструкцию 8, SEQ ID NO: 18 (схематично представленную на ФИГ. 9), амплифицировали, объединяя продукты ПЦР после амплификации конструкции 2 и шарнирной области HuIgG-Fc, и прямой и обратный праймеры SEQ ID NO: 55 и SEQ ID NO: 66, соответственно.
Конечные продукты клонировали в экспрессирующий вектор млекопитающих pCIneo (Promega, Inc.), как описано ранее. Плазмиду, кодирующую конструкции, обозначали, как указано в таблице 14:
ПЛАЗМИДНЫЕ КОНСТРУКЦИИ
Экспрессия полипептида/диантитела: Проводили четыре отдельных котрансфекции в клетки HEK-293 с использованием Lipofectamine 2000, как описано в разделе 6.1: pMGX0669 и pMGX0674, кодирующие конструкции 1 и 6, соответственно; pMGX0667 и pMGX0676, кодирующие конструкции 2 и 5, соответственно; и pMGX0677 и pMGX0678, кодирующие конструкции 7 и 8, соответственно.
Котрансфекция этих плазмид была предназначена для обеспечения экспрессии четырехвалентного биспецифического диантитела (CBD) с IgG-подобной структурой, иммуноспецифичного как к FcγRIIB, так и к FcγRIIIA. Также проводили дополнительную котрансфекцию: pMGX0674 и pMGX0676, кодирующие конструкции 6 и 5, соответственно. После трех суток в культуре кондиционированные среды собирали. Количество секретируемого продукта в кондиционированной среде количественно определяли с помощью ELISA против Fc IgG с использованием очищенного Fc в качестве стандарта. Затем концентрации продукта в образцах нормализовали, исходя из количественного определения, и нормализованные образцы использовали для остальных анализов.
ELISA: Связывание молекул диантител, секретированных в среду, анализировали посредством сэндвич-ELISA, как описано выше. Если не указано, для покрытия планшета использовали CD32B, т.е. в качестве белка-мишени, и в качестве зонда использовали конъюгированный с HRP CD16.
Результаты
Для тестирования нормализованных образцов из рекомбинантных экспрессирующих систем, содержащих конструкции 1 и 6 (pMGX669-pMGX674), конструкции 2 и 5 (pMGX667-pNGX676) и конструкции 5 и 6 (pMGX674-pMGX676), в отношении экспрессии молекул диантител, способных одновременно связывать CD32B и CD16A (ФИГ. 10), использовали анализ ELISA. Данные ELISA показали, что котрансфекция конструкциями 1 и 6 или котрансфекция конструкциями 2 и 5 не продуцировала продукт, который мог связывать любой или оба антигена (ФИГ. 10, □ и ▲, соответственно). Однако котрансфекция конструкций 5 и 6 привела к секреции продукта, способного связывать оба антигена CD32B и CD16. Последний продукт представлял собой димер конструкций 5 и 6, содержащий один участок связывания для каждого антигена со структурой, схематично представленной на ФИГ. 11.
Для запуска образования IgG-подобной гетеротетрамерной структуры к C-концу конструкции 1 присоединяли кодирующую последовательность для шести дополнительных аминокислот, создавая конструкцию 7 (SEQ ID NO: 16 и схематично представленная на ФИГ. 9). Шесть дополнительных аминокислот FNRGEC (SEQ ID NO: 23) были взяты с C-конца легкой цепи каппа, и в норме они взаимодействуют с верхним шарнирным доменом тяжелой цепи в молекуле IgG. Затем в конструкцию 6 встраивали шарнирный домен, создавая конструкцию 8 (SEQ ID NO: 18 и ФИГ. 9). Конструкция 8 дополнительно содержала мутацию аминокислоты в верхней шарнирной области, A215V. Затем экспрессирующие плазмиды, кодирующие конструкцию 7 и конструкцию 8, pMGX677 и pMGX678, соответственно, котрансфицировали в клетки HEK-293 и экспрессировали, как описано.
Молекулы диантител, продуцированные с рекомбинантной экспрессирующей системы, содержащей конструкции 7 и 8 (pMGX0677 + pMGX0678), сравнивали в анализе ELISA в отношении связывания с CD32B и CD16A молекул диантител, продуцированных с экспрессирующих систем, содержащих конструкции 1 и 6 (pMGX669 + pMGX674), конструкции 2 и 8 (pMGX669 + pMGX678) и конструкции 6 и 7 (pMGX677 + pMGX674) (ФИГ. 12).
Как и ранее, молекула, продуцируемая экспрессирующей системой, содержащей конструкции 1 и 6 (pMGX669 + pMGX674), оказалась неспособна связывать как CD32A, так и CD16A (ФИГ. 10 и ФИГ. 12). Напротив, продукты коэкспрессии любой из конструкций 7 и 6 (pMGX0677 + pMGX0674) или коэкспрессии конструкций 7 и 8 (pMGX0677-pMGX0678) были способны связывать как CD32B, так и CD16 (ФИГ. 12). Следует отметить, что конструкция 7 аналогична конструкции 1, за исключением того, что конструкция 7 содержит C-концевую последовательность FNRGEC (SEC ID NO:23); и что конструкция 8 аналогична конструкции 6, за исключением того, что конструкция 8 содержит шарнирный домен и мутацию A215V. Данные указывают на то, что добавление 6 дополнительных аминокислот с C-конца легкой цепи C-каппа (FNRGEC; SEQ ID NO: 23) к не несущей Fc "более легкой цепи" помогло стабилизировать образование тетрамерных IgG-подобных молекул диантител, независимо от того, содержит ли соответствующая "более тяжелая" цепь шарнирный домен (т.е. pMGX0677 + pMGX0674 и pMGX0677-pMGX0678, ФИГ. 12). Добавление шарнирного домена несущему Fc "более тяжелому" полипептиду без добавления C-концевой последовательности FNRGEC (SEQ ID NO: 23) к соответствующей "более легкой" цепи, по-видимому, оказалось неспособным обеспечить сходную стабилизацию (т.е. отсутствие связывания продуктом котрансфекции конструкций 2 и 8 (pMGX669 + pMGX678)). Структура тетрамерной молекулы диантитела схематично представлена на ФИГ. 13.
6.3 ЭФФЕКТ ПОРЯДКА ДОМЕНОВ И ДОПОЛНИТЕЛЬНЫХ ДИСУЛЬФИДНЫХ СВЯЗЕЙ НА ОБРАЗОВАНИЕ ТЕТРАМЕРНОГО IgG-ПОДОБНОГО ДИАНТИТЕЛА
Эффект дополнительной стабилизации между "более легкой" и "более тяжелой" полипептидными цепями тетрамерной IgG-подобной молекулы диантитела исследовали путем замены выбранных остатков на полипептидных цепях остатками цистеина. Дополнительные остатки цистеина обеспечивают дополнительные дисульфидные связи между "более тяжелыми" и "более легкими цепями". Кроме того, порядок доменов в отношении активности связывания исследовали смещением Fc-домена или домена шарнирная область-Fc с C-конца полипептидной цепи на N-конец. Хотя в случае таких связей активность связывания молекулы, содержащей дополнительные дисульфидные связи, не изменялась относительно ранее сконструированных молекул диантител, перенос Fc или шарнирного-Fc домена на N-конец "более тяжелой" полипептидной цепи, содержащей диантитело, неожиданно улучшил аффинность связывания и/или авидность биспецифической молекулы к одному или обоим из ее антигенов-мишеней.
Материалы и способы
Конструирование и разработка полипептидных молекул: Экспрессирующие векторы для нуклеиновых кислот конструировали так, чтобы они продуцировали модифицированные варианты конструкций 5, 6 и 8, представленных в примере 6.2. Конструкция 9 (SEQ ID NO: 19) и конструкция 10 (SEQ ID NO: 20) (обе схематично представлены на ФИГ. 13) были аналогичны конструкциям 8 и 6, за исключением того, что Fc-домен или шарнирный-Fc домен, соответственно, были сдвинуты с C-конца полипептида на N-конец. Кроме того, все использованные Fc-домены представляли собой Fc-домены IgG1 дикого типа. Конструкция 11, SEQ ID NO: 21, (схематично представленная на ФИГ. 14) была аналогична конструкции 2 из примера 6.1, за исключением того, что C-конец был предназначен, чтобы он дополнительно содержал последовательность FNRGEC (SEQ ID NO: 23). Конструкция 12, SEQ ID NO: 22, (схематично представленная на ФИГ. 14) была аналогична конструкции 5 из примера 6.2 за исключением того, что Fc-домен дополнительно содержал шарнирную область. Также для конструкций 11 и 12, VL-домен 2B6 и VH-домен 2B6 содержали одну модификацию аминокислот (G105C и G44C, соответственно), так чтобы глицин в каждом домене был заменен цистеином.
ПЦР и конструирование экспрессирующих векторов: Все протоколы ПЦР и очистки продуктов ПЦР были такими, как описано в примере 6.1 и 6.2.
ПЦР c перекрыванием: Конечные продукты конструировали, амплифицировали и очищали с использованием способов, описанных в примере 6.1 и примере 6.2.
Конечные продукты клонировали в экспрессирующий вектор млекопитающих pCIneo (Promega, Inc.), как описано ранее. Плазмиды, кодирующие конструкции, были обозначены, как указано в таблице 15:
ПЛАЗМИДНЫЕ КОНСТРУКЦИИ
Экспрессия полипептида/диантитела: Проводили три отдельных котрансфекции в клетки HEK-293 с использованием Lipofectamine 2000, как описано в разделе 6.1: pMGX0669 и pMGX0719, кодирующие конструкции 1 и 9, соответственно; pMGX0669 и pMGX0718, кодирующие конструкции 1 и 10, соответственно; и pMGX0617 и pMGX0717, кодирующие конструкции 11 и 12, соответственно. Котрансфекция этих плазмид была предназначена для обеспечения экспрессии четырехвалентного биспецифического диантитела (CBD) с IgG-подобной структурой, иммуноспецифичного как к FcγRIIB, так и к FcγRIIIA. После трех суток в культуре кондиционированную среду собирали. Количество секретируемого продукта в кондиционированной среде количественно определяли посредством ELISA против Fc IgG с использованием очищенного Fc в качестве стандарта. Затем концентрации продукта в образцах нормализовали, исходя из количественного определения, и нормализованные образцы использовали в остальных анализах.
ELISA: Связывание молекул диантител, секретированных в среду, анализировали посредством сэндвич-ELISA, как описано выше. Если не указано, для покрытия планшета использовали CD32B, т.е. в качестве белка-мишени, и в качестве зонда использовали конъюгированный с HRP CD16.
Вестерн-блоттинг: Приблизительно 15 мл кондиционированной среды после трех описанных выше котрансфекций анализировали посредством SDS-PAGE в невосстанавливающих условиях. Один гель окрашивали Simply Blue Safestain (Invitrogen) и идентичный гель переносили на мембрану PVDF (Invitrogen) с использованием стандартных способов переноса. После переноса мембрану блокировали 5% сухим обезжиренным молоком в 1X PBS. Затем мембрану инкубировали в 10 мл конъюгированного с HRP антитела козы против H+L IgG1 человека, разбавленного 1:8000, в 2% сухом обезжиренном молоке 1X PBS/0,1% Tween 20 при комнатной температуре в течение 1 ч при осторожном встряхивании. После промывания 1X PBS/0,3% Tween 20, 2X 5 мин каждый раз, затем 20 мин при комнатной температуре, мембрану проявляли системой для детекции вестерн-блоттинга ECL (Amersham Biosciences) согласно инструкциям изготовителя. Пленку проявляли устройством для обработки рентгеновских данных.
Результаты
Кондиционированную среду от рекомбинантных экспрессирующих систем, содержащих конструкции 1 и 9, конструкции 1 и 10 и конструкции 11 и 12, анализировали посредством анализа SDS-PAGE (в невосстанавливающих условиях) и вестерн-блоттинга (с использованием антитела против IgG в качестве зонда). Вестерн-блоттинг выявил, что продукт систем, содержащих конструкции 11 и 12 или содержащих конструкции 9 и 1, главным образом, образовывал отдельные типы молекул размером приблизительно 150 кДа (ФИГ. 14, дорожки 3 и 2, соответственно). Оба из этих продукта имеют встроенные способами инженерии внутренние дисульфидные связи между "более легкими" и "более тяжелыми" цепями, содержащими диантитело. Напротив, молекула со встроенными в нее способами инженерии внутренними дисульфидными связями между "более легкими" и "более тяжелыми" цепями, образованными из конструкций 10 и 1, образовывала по меньшей мере два типа молекул с молекулярной массой ~75 и ~100 кДа (ФИГ. 14, дорожка 1).
Несмотря на результаты вестерн-блоттинга, было выявлено, что каждый из трех продуктов был способен связывать как CD32A, так и CD16 (ФИГ. 15). Неожиданно, относительно продукта, содержащего C-концевой шарнирный-Fc домен (образованный из конструкций 11 и 12), продукт обеих систем, где Fc-домен (или домен Fc-шарнирный домен) был на N-конце содержащей Fc полипептидной цепи (т.е. "более тяжелой" цепи) (конструкции 9+1 и конструкции 10+1), продемонстрировал увеличенную аффинность и/или авидность в отношении одного или обоих из его пептидов-мишеней (т.е. CD32B и/или CD16).
6.4 ЭФФЕКТ ВНУТРЕННЕГО/ВНЕШНЕГО УЧАСТКА РАСЩЕПЛЕНИЯ НА ПРОЦЕССИНГ ПОЛИБЕЛКОВОГО ПРЕДШЕСТВЕННИКА И ЭКСПРЕССИЮ КОВАЛЕНТНОГО БИСПЕЦИФИЧЕСКОГО ДИАНТИТЕЛА; КОНСТРУИРОВАНИЕ И ОХАРАКТЕРИЗАЦИЯ БИСПЕЦИФИЧЕСКОГО ДИАНТИТЕЛА, СОДЕРЖАЩЕГО ЧАСТИ ЛЯМБДА-ЦЕПИ IgG ЧЕЛОВЕКА И ШАРНИРНЫЙ ДОМЕН
Как описано в настоящем документе, отдельные полипептидные цепи диантитела или молекулы диантитела по изобретению можно экспрессировать в качестве единой полибелковой молекулы-предшественника. Способность рекомбинантных систем, описанных в примерах 6.1-6.3, надлежащим образом осуществлять процессинг и экспрессировать функциональный CBD из такого полибелкового предшественника, тестировали путем конструирования нуклеиновой кислоты, кодирующей как первую, так и вторую полипептидные цепи CBD, разделенные внутренним участком расщепления, в частности, участком расщепления фурином. Функциональный CBD выделяли из рекомбинантной системы, содержащей полибелковую молекулу-предшественника.
Как описано в примере 6.3, было выявлено, что добавление 6 C-концевых аминокислот из легкой цепи каппа человека, FNRGEC (SEQ ID NO: 23), стабилизирует образование диантитела - предположительно посредством усиленного межцепочечного взаимодействия между двумя доменами, содержащими SEQ ID NO: 23, и доменами, содержащими Fc-домен или шарнирный-Fc домен. Стабилизирующий эффект этого взаимодействия, подобного взаимодействию лямбда-цепь/Fc, тестировали в CBD, где ни одна полипептидная цепь не содержала Fc-домен. Одну полипептидную цепь диантитела конструировали так, чтобы она содержала SEQ ID NO: 23 на ее C-конце; полипептидную цепь-партнер конструировали так, чтобы она содержала аминокислотную последовательность VEPKSC (SEQ ID NO: 77), которая происходила из шарнирого домена IgG. Сравнение этого CBD с CBD, состоящим из конструкций 1 и 2 (из примера 6.1) показало, что CBD, содержащее домены, происходящие из шарнирного домена и лямбда-цепи, проявляли немного более высокую аффинность к одному или обоим из их эпитопов-мишеней.
Материалы и способы
Конструирование и разработка полипептидных молекул: Полибелковый предшественник: Экспрессирующие нуклеиновые кислоты векторы конструировали, чтобы они продуцировали 2 полибелковых молекулы-предшественника, обе из которых схематично представлены на ФИГ. 17. Конструкция 13 (SEQ ID NO: 95) состояла из N-конца полипептидной цепи, VL-домена 3G8, VH-домена 2.4G2 (который связывает mCD32B), участка расщепления фуринов, VL-домена 2.4G2 и VH-домена 3G8. Нуклеотидная последовательность, кодирующая конструкцию 13, представлена в SEQ ID NO: 96. Конструкция 14 (SEQ ID NO: 97) (ФИГ. 17) состояла из N-конца полипептидной цепи, VL-домена 3G8, VH-домена 2.4G2 (который связывает mCD32B), участка расщепления фурином, участка FMD (протеаза C3 вируса ящура), VL-домена 2.4G2 и VH-домена 3G8. Нуклеотидная последовательность, кодирующая конструкцию 14, представлена в SEQ ID NO: 98.
Векторы, экспрессирующие нуклеиновые кислоты, конструировали, чтобы они продуцировали модифицированные варианты конструкций 1 и 2, представленных в примере 6.1. Конструкция 15 (SEQ ID NO: 99) (ФИГ. 17) была аналогична конструкции 1 (SEQ ID NO: 9), представленной в примере 6.1, за исключением того, что C-конец конструкции 15 содержал аминокислотную последовательность FNRGEC (SEQ ID NO: 23). Последовательность нуклеиновой кислоты, кодирующая конструкцию 15, представлена в SEQ ID NO: 100. Конструкция 16 (SEQ ID NO: 101) (ФИГ. 17) была аналогична конструкции 2, представленной в примере 6.1, за исключением того, что C-конец конструкции 16 содержал аминокислотную последовательность VEPKSC (SEQ ID NO: 77). Последовательность нуклеиновой кислоты, кодирующая конструкцию 16, представлена в SEQ ID NO: 102.
ПЦР и конструирование экспрессирующих векторов: Все протоколы ПЦР и очистки продуктов ПЦР были такими, как описано в примере 6.1 и 6.2.
ПЦР c перекрыванием: Конечные продукты конструировали, амплифицировали и очищали с использованием способов, описанных в примере 6.1 и примере 6.2, с соответствующими праймерами.
Конечные продукты клонировали в экспрессирующий вектор млекопитающих pCIneo (Promega, Inc.), как описано ранее. Плазмиды, кодирующие конструкции, были обозначены, как указано в таблице 16:
ПЛАЗМИДНЫЕ КОНСТРУКЦИИ
Экспрессия полипептида/диантитела: Проводили одну трансфекцию и одну котрансфекцию в клетки HEK-293 с использованием Lipofectamine 2000, как описано в разделе 6.1: единичная: pMGX0750, кодирующий конструкцию 13; и котрансфекция: pMGX0752 и pMGX0753, кодирующие конструкции 15 и 16, соответственно. После трех суток в культуре кондиционированные среды собирали, и секретированный продукт подвергали аффинной очистке, как описано.
ELISA: Связывание молекул диантител, секретируемых в среду, анализировали посредством сэндвич-ELISA, как описано выше. Для покрытия планшета использовали CD32B мыши, т.е. в качестве белка-мишени, и конъюгированный с HRP CD16A использовали в качестве зонда для продукта котрансфекции конструкций 15 и 16. mCD32B использовали в качестве белка-мишени и конъюгированный с биотином CD16A использовали в качестве зонда для рекомбинантной системы, содержащей конструкцию 13.
Результаты
Кондиционированные среды из рекомбинантных экспрессирующих систем, содержащих конструкции 13, анализировали посредством сэндвич-ELISA. Анализ ELISA тестировал специфичность связывания CBD с любым или с обоими из mCD32B и/или CD16 (ФИГ. 18). CD32B служил в качестве антигена-мишени и CD16A использовали в качестве вторичного зонда. Положительный сигнал в ELISA показал, что гетеродимерное CBD h2.4FD2-h3G8, продуцированное из полибелкового предшественника, обладало специфичностью к обоим антигенам.
Аналогично, очищенный продукт получали котрансфекцией векторов, кодирующих конструкции 15 и 16, тестировали в анализе ELISA и сравнивали с продуктом, состоящим из конструкций 1 и 2 (пример 6.1). CD32B служил в качестве антигена-мишени и CD16A использовали в качестве вторичного зонда. Как и в случае продукта, состоящего из конструкций 1 и 2, было выявлено, что продукт из конструкций 15 и 16 способен одновременно связывать CD32B и CD16A. В действительности, продукт из конструкций 15 и 16 продемонстрировал немного увеличенную аффинность в отношении одного или обоих из антигенов-мишеней, т.е. CD32B или CD16A. Возможно, это является следствием увеличенной стабильности и/или точности (относительно взаимодействия доменов VH-VL дикого типа) ассоциации между цепями, обеспечиваемой взаимодействием области лямбда-цепи FNRGEC (SEQ ID NO: 23) и шарнирной области VEPKSC (SEQ ID NO: 77), которая отсутствует в продукте, состоящем из конструкций 1 и 2.
6.5 ПРИМЕНЕНИЕ ПЕРЕНАЦЕЛИВАЮЩИХ РЕАГЕНТОВ С ДВОЙНОЙ АФФИННОСТЬЮ ("DART") ДЛЯ ОБЪЕДИНЕНИЯ МНОЖЕСТВА АФФИННОСТЕЙ ВМЕСТЕ
Один из аспектов настоящего изобретения относится к новым перенацеливающим реагентам с двойной аффинностью ("DART"), а также к новым способам объединения нескольких аффинностей вместе. "DART" могут быть моноспецифическими, биспецифичскими, триспецифическими и т.д., таким образом, являясь способными одновременно связывать один, два, три или более различных эпитопов (которые могут быть из одного или различных антигенов). Кроме того, "DART" могут быть моновалентными, двухвалентными, трехвалентными, четырехвалентными, пятивалентными, шестивалентными и т.д., таким образом, являясь способными одновременно связывать одну, две, три, четыре, пять, шесть или более молекул. Как показано на фигуре 35, эти два свойства DART можно комбинировать, например, для получения биспецифических антител, которые являются четырехвалентными и т.д.
Одним из продвижений является разработка DART, который обладает аффинностью в отношении прототипного иммунного рецептора, huCD32B, а также аффинностью к гаптену, флуоресцеину. Этот DART, названный "2B6/4420", служит в качестве универсального адаптера, способного колигировать huCD32B с молекулами, которые взаимодействуют с конъюгированными с флуоресцеином партнерами по связыванию. CD32B представляет собой Fc-рецептор, который способен гасить активирующие сигналы путем кластеризации с активирующими передачу сигнала иммунными комплексами. В этом первоначальном воплощении, эта технология позволяет быстрый скрининг нескольких биологических мишеней в отношении кластеризации с huCD32B без необходимости в получении новых конструкций DART. 2B6/4420 можно просто смешивать с конъюгированным с флуоресцеином антителом против рецептора клеточной поверхности, и, тем самым, имитировать действие DART с аффинностью в отношении этого рецептора (ФИГ. 20). Кроме того, этот реагент позволяет эффективное связывание аффинных реагентов, которые нелегко экспрессировать или продуцировать, что позволяет преодолеть технические ограничения. Содержащие 2B6/4420 DART, очевидно, пригодны в качестве инструментов для исследования, а также в качестве кандидатов для клиники. 2B6/4420, продуцированный из клеток HEK293, может одновременно связывать CD32B и флуоресцеин в анализе ELISA. Кроме того, он может ингибировать пролиферацию клеток путем привлечения CD32B в комплекс BCR путем колигирования с CD79. Плечо 2B6 в DART можно заменять отличающейся последовательностью антитела или связывающей последовательностью, имеющей другую подходящую специфичность.
Материалы и способы:
Плазмидные конструкции: 2B6/4420 происходит из последовательностей гуманизированного MAb 2B6 (hu2B6, MGA321) и химерного варианта Fv мыши/Fc человека MAb против флуоресцеина 4420. Полностью собранный DART состоит из двух полипептидов, что приводит к ковалентному связыванию двух Fv-областей. Первый полипептид состоит из секреторной сигнальной последовательности, за ним следует hu2B6VL, продуцируемый в качестве слитого белка с 4420VH, разделенный линкером, состоящим из аминокислотных остатков GGGSGGGG. Последовательность FNRGEC, происходящая из C-конца легкой цепи каппа, присоединена к C-концу этого полипептида. Другой полипептид состоит из сигнальная последовательность-4420VL-GGGSGGGG-hu2B6VH, с последовательностью VEPKSC, происходящей из C-конца Fd-фрагмента IgG1 человека, присоединенной к C-концу. Остатки цистеина в двух цепях образуют дисульфидную связь, ковалентно связывающую два полипептида вместе (ФИГ. 20). Последовательности ДНК, кодирующие описанные полипептиды, подвергали амплификации способом ПЦР из существующих плазмид, комбинировали с помощью ПЦР с перекрыванием и клонировали в pCIneo (Promega) между участками NheI и EcoRI. Наконец, DART с аффинностью в отношении huCD32B и huCD16 (2B6/3G8), который ранее конструировали с использованием способов, сходных со способами, описанными выше, использовали в качестве контроля.
Антитела: Моноклональные антитела мыши против CD79b, CB3.1 и CB3.2 человека (гибридомы) получали от Dr. Cooper MD, University of Alabama at Birmingham, Birmingham AL. CB3.1 и CB3.2 метили флуоресцеинизотиоцианатом (FITC) в соответствии с инструкциями изготовителя (Pierce, Rockford IL). F(ab')2-фрагмент специфического к Fc-фрагменту антитела козы против (GAM) IgG мыши получали от Jackson Laboratories (West Grove, PA). MAb мыши против huCD32B, 3H7, продуцировали и очищали самостоятельно. Антитело козы против 2B6Fv продуцировали путем иммунизации коз целым антителом hu2B6 и аффинной очистки против Fv-области hu2B6. HuIgG, FITC-huIgG и конъюгированное с HRP антитело против IgG мыши получали от Jackson Immunoresearch. Конъюгированное с HRP антитело против антител козы получали от Southern Biotech.
Экспрессия DART: Плазмиды, кодирующие каждую цепь, котрансфицировали в клетки 293H (Invitrogen) с использованием Lipofectamine 2000 (Invitrogen) согласно инструкциям изготовителя. Секретированный белок собирали 3-4 раза с интервалами трое суток и очищали жидкостной хроматографией против иммобилизованной растворимой формы CD32B.
ELISA: DART 2B6/4420 или 2B6/3G8 улавливали на планшетах MaxiSorp (Nalge Nunc), покрытых меченным FITC белком S (Novagen), IgG человека или FITC-huIgG. Детекцию проводили по связыванию растворимого эктодомена CD32B, а затем 3H7 (моноклональное антитело мыши, специфичное к CD32B), и наконец антитела против антител мыши, конъюгированного с HRP. Альтернативно детекцию проводили путем связывания поликлональной подвергнутой аффинной очистке антисыворотки козы против Fv 2B6, а затем антителом против антител козы, конъюгированным с HRP. Детекцию активности HRP проводили с использованием колориметрического субстрата TMB (Bio FX) и считывания на устройстве для считывания планшетов VersaMax ELISA.
Очистка B-клеток и анализ пролиферации: Мононуклеарные клетки периферической крови разделяли способом с градиентом Ficoll/Paque Plus (Amersham Pharmacia Biotech, UK) с использованием крови от здоровых доноров. B-лимфоциты выделяли с использованием набора Dynal B Cell Negative Isolation Kit (Dynal Biotechnology Inc., NY) в соответствии с инструкциями изготовителя. Чистота выделенных B-клеток (CD20+) была выше 90% при оценке посредством анализа FACS. Для анализа пролиферации очищенные B-клетки высевали в полную среду RPMI 1640 в 96-луночных микропланшетах для титрования с плоским дном с плотностью клеток 1×105 клеток на лунку в конечном объеме 200 мкл и инкубировали в течение 48 ч в присутствии или отсутствие антител и диантител при 37°C в 5% CO2. Затем добавляли 1 мкКи/лунка [3H]тимидина (Perkin Elmer, Wellesley, MA) и инкубацию продолжали в течение дополнительных 16-18 ч перед сбором. Включение [3H]тимидина измеряли путем жидкостного сцинтилляционного подсчета.
Результаты
Чтобы продемонстрировать, что DART 2B6/4420 является активным и специфичным, проводили два эксперимента ELISA. Сначала, 2B6/4420 или 2B6/3G8 (в качестве отрицательного контроля) связывали с конъюгированным с флуоресцеином белком (S-белок), который наносили на планшеты для ELISA. Далее плечо 2B6 в DART приводили в контакт с растворимым CD32B. Детекцию связывания проводили с помощью другого антитела к CD32B с эпитопом, который не перекрывает эпитоп 2B6, а затем конъюгированным с HRP вторичным антителом. Хотя DART 2B6/4420 способно одновременно связывать флуоресцеин и CD32B, 2B6/3G8 не способно к этому (ФИГ. 21, панель A). При улавливании DART на планшетах, покрытых растворимым CD32B, и детекции связывания с помощью антитела, специфичного к Fv hu2B6, оба DART демонстрируют хорошее связывание. Чтобы показать, что DART 2B6/4420 способен связывать флуоресцеин, конъюгированный с IgG человека (учитывая, что это является контекстом первоначального воплощения этого реагента), HuIgG, немеченное или меченное флуоресцеином, связывали с планшетами для ELISA и использовали для улавливания 2B6/4420. Вновь в качестве отрицательного контроля использовали 2B6/3G8. Детекцию связывания проводили с использованием антитела, специфичного к Hu2B6 Fv. DART 2B6/4420 очевидно связывается с FITC-HuIgG, но не связывается с немеченым HuIgG, демонстрируя, что этот DART способен связывать флуоресцеин, конъюгированный с антителом, и что отсутствует существенное связывание с антителом отдельно. Как и ожидалось, не было выявлено связывания посредством DART 2B6/3G8 ни в одном из этих контекстов.
Проводили эксперименты, чтобы продемонстрировать, что DART 2B6/4420 способен функционировать в качестве реагента с двойной аффинностью, который мог иметь эффект при передаче сигнала в контексте клеточного анализа. Было показано, что коагрегация CD32B с BCR ингибирует активацию B-клеток. Исследовали способность DART 2B6/4420 совместно связывать CD32B с BCR, покрытым антителами αCD79b, меченными флуоресцеином, и запускать ингибирование пролиферации клеток. B-клетки подвергали отрицательной селекции из крови человека и активировали путем обработки возрастающими концентрациями клонами CB3.1 и CB3.2 меченных FITC антител мыши против CD79b человека, и путем добавления F(ab')2-фрагмента Fc-специфического GAM в качестве вторичного реагента для связывания BCR, вместе с фиксированной концентрацией (5 мкг/мл) DART 2B6/4420 или эквивалентным количеством DART 2B6/3G8, молекулы, которая не нацелена на флуоресцеин, таким образом, использованной в качестве контроля. Пролиферация клеток, измеренная как включение [3H]-тимидина, возрастала при увеличении концентраций моноклонального антитела-активатора против CD79b-FITC в отсутствие DARTS или в присутствии контрольного DART 2B6/3G8. Присутствие DART 2B6/4420 привело к выраженной пролиферации B-клеток при всех концентрациях антитела против CD79b человека-FITC (ФИГ. 22, панели A и B, и ФИГ. 23, панель A).
Ингибирования пролиферации не наблюдали, когда B-клетки, покрытые немеченым CB3.2 и активированные с использованием тех же экспериментальных условий, обрабатывали DART 2B6/4420, обеспечивающим его специфичность к мишени (ФИГ. 23, панель B). Эти данные демонстрируют, что DART 2B6/4420 способен связывать CD32B и BCR и доставлять ингибиторный сигнал, способный блокировать индуцируемую антигеном-рецептором активацию клеток.
6.6 ИММУНОТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО DART ПРОТИВ CD32B, ЭКСПРЕССИРУЕМОГО B-КЛЕТОЧНЫМИ ЗЛОКАЧЕСТВЕННЫМИ ОПУХОЛЯМИ
В настоящее время B-клеточные злокачественные опухоли лечат с использованием антитела против CD20 Rituxane®. Однако некоторые B-клеточные злокачественные опухоли не экспрессируют CD20 или становятся устойчивыми к Rituxane. DART по настоящему изобретению обеспечивают альтернативное иммунотерапевтическое средство, способное преодолевать проблемы, ассоциированные с антителом против CD20 Rituxane®.
MGD261 представляет собой перенацеливающую молекулу с двойной аффинностью (DART), связывающуюся с hCD32B (через антитело h2B6) и hCD16A и hCD16B (через антитело h3G8).
Эффективность (истощение B-клеток) и безопасность MGD261 тестировали на mCD32-/- hCD16A+ C57B1/6, mCD32-/- hCD32B+ C57B1/6 и mCD32-/- hCD16A+ hCD32B+ C57B1/6. В этом эксперименте с повторяющейся дозой мышам проводили 6 внутривенных инъекций (два раза в неделю в течение 3 недель). Мониторинг истощения B-клеток проводили посредством FACS. Мониторинг безопасности проводили путем наблюдения за клетками.
Данные указывают на то, что MGD261 способно истощать B-клетки у двойных трансгенных мышей без индукции каких-либо существенных побочных эффектов.
Данные: Мышам mCD32-/- hCD16A+ C57B1/6, mCD32-/- hCD32B+ C57B1/6 и mCD32-/- hCD16A+ hCD32B+ C57B1/6 из колонии для выведения MacroGenics внутривенно инъецировали на 0, 3, 7, 10, 14 и 17 сутки MGD261 (10, 3, 1 или 0,3 мг/кг), или постороннее антитело (hE16 10 мг/кг). Взятие крови проводили на сутки -19 (до отбора крови), 4, 11, 18, 25 и 32 для анализа FACS. Здоровье и активность животных регистрировали три раза в неделю.
Способ FACS-анализа: Образцы цельной крови собирали за 18 суток до введения h2B6-h3G8 и через 4, 11, 18, 25 и 32 суток после введения. Образцы крови анализировали для определения эффекта h2B6-h3G8 на количество B-клеток в анализе на основе FACS. Для определения количества B-клеток, T-клеток и PMN использовали протокол без промывания с использованием гранул FlowCount, полученных от Beckman Coulter. Панель антител, использованная в анализе, представляла собой 1A8-FITC для PMN, CD3-PE для T-клеток, CD19-APC для B-клеток и CD45-PerCP для общих лейкоцитов.
Результаты
Мыши, которым вводили hE16 или MGD261 (в любой концентрации), не продемонстрировали каких-либо признаков дискомфорта в какое-либо время на протяжении эксперимента.
Истощение B-клеток наблюдали у двойных трансгенных мышей hCD16A и hCD32B. Диантитело h2B6-3G8 связывает экспрессирующие hCD16A эффекторные клетки и экспрессирующие hCD32B B-клетки; связывание требовалось для уничтожения B-клеток. Истощение B-клеток не наблюдали у однократно трансгенных мышей (ФИГ. 24). В процессе исследования не было значительных изменений уровня T-клеток и PMN.
В качестве дальнейшей демонстрации альтернативных иммунотерапевтических средств по настоящему изобретению, конструировали замену MGD261, названную "2.4G2-3G8 DB". 2.4G2-3G8 DB представляет собой перенацеливающую молекулу с двойной аффинностью (DART), связывающуюся с mCD32B (через антитело 2.4G2) и hCD16A и hCD16B (через антитело h3G8).
Эффективность (истощение B-клеток) и безопасность 2.4G2-3G8 DB тестировали у мышей mCD16-/-, mCD16-/- hCD16A+ C57B1/6, mCD16-/- hCD16B+ и mCD16-/- hCD16A+ hCD16B+. В этом эксперименте с повторяющейся дозой мышам проводили 9 внутрибрюшинных инъекций (три раза в неделю в течение 3 недель). Мониторинг истощения B-клеток проводили посредством FACS. Мониторинг безопасности проводили посредством наблюдения за клеткой.
Данные указывают на то, что 2.4G2-3G8 DB способно истощать B-клетки у трансгенных мышей hCD16 без индукции каких-либо значительных побочных эффектов.
Данные: мышам mCD16-/-, mCD16-/- hCD16A+ C57B1/6, mCD16-/- hCD16B+ и mCD16-/- hCD16A+ hCD16B+ из колонии для выведения MacroGenics на 0, 2, 4, 7, 9, 11, 14, 16 и 18 сутки внутрибрюшинно инъецировали 2.4G2-3G8 DB (75 мкг/мышь) или PBS. Для анализа FACS взятие крови проводили на сутки -10 (до кровопускания), 4, 11 и 18. Здоровье и активность животных регистрировали три раза в неделю.
Способ FACS-анализа: Образцы цельной крови собирали за 10 суток до введения 2.4G2-3G8 и через 4, 11 и 18 после начала введения. Образцы крови анализировали для определения эффекта 2.4G2-3G8 на количество B-клеток с помощью анализа на основе FACS. Для определения количества B-клеток, T-клеток и PMN применяли протокол без промывания с использованием гранул пробирок TruCOUNT, полученных от BD Immunocytometry System. Панель антител, использованных для анализа, представляла собой 1A8-FITC для PMN, CD3-PE для T-клеток, CD19-APC для B-клеток и CD45-PerCP для общего количества лейкоцитов.
Результаты
Мыши, которым вводили hE16 или 2.4G2-3G8 DB, не продемонстрировали каких-либо признаков дискомфорта в какое-либо время в течение эксперимента.
Истощение B-клеток наблюдали у мышей mCD16-/- hCD16A+ или mCD16-/- hCD16A+ hCD16B+, но не у мышей mCD16-/-. Эти данные указывают на то, что для уничтожения B-клеток требовались эффекторные клетки, несущие hCD16A (ФИГ. 25). Не было значимых изменений для уровня T-клеток и PMN в процессе исследования.
Внутривенная (IV) модель: Противоопухолевую активность MGD261 тестировали с использованием внутривенной (IV) модели на опухолевой клеточной линии человека Raji. Raji представляет собой клеточную линию лимфомы Беркитта, экспрессирующую hCD32B. При внутривенной инъекции мышам mCD16-/-, hCD16A+, RAG1-/-, опухолевые клетки располагаются в позвоночнике и приводят к параличу задних лап.
Данные указывают на то, что MGD261 способно блокировать рост опухолевых клеток Raji in vivo у мышей mCD16-/-, hCD16A+, RAG1-/-. Данные указывают на то, что MGD261 можно использовать для лечения экспрессирующих CD32B B-клеточных злокачественных опухолей у человека.
Данные: Мышам mCD16-/-, hCD16A+, RAG1-/- C57B1/6 из колонии для выведения MacroGenics внутривенно инъецировали на 0 сутки 5×106 клеток Raji. На сутки 6, 9, 13, 16, 20, 23, 27 и 30 мышам также внутрибрюшинно (IP) вводили 250, 25 или 2,5 мкг MGD261 или PBS (отрицательный контроль). Затем мышей наблюдали каждые сутки и массу тела регистрировали два раза в неделю. Мышей, у которых развивался паралич задних лап, умерщвляли.
Результаты: Мыши, которым вводили PBS, погибали между 25 сутками и 50 сутками. Мыши, которым вводили MGD261, выживали по меньшей мере до 90 суток (ФИГ. 26). Увеличение выживания является статистически значимым. Сравнение кривых выживания с использованием логарифмического рангового критерия дало χ2, равный 96,46 (df 9; значение P <0,0001).
6.7 ЭКСПРЕССИЯ DART У ПРОКАРИОТ
Эксперименты проводили для того, чтобы продемонстрировать возможность продуцировать DART в хозяевах, не являющихся млекопитающими. Таким образом, Escherichia coli трансформировали экспрессирующей DART плазмидой, и проводили мониторинг экспрессии DART.
Материалы и способы:
Конструирование плазмиды: 3G8 представляет собой гуманизированное моноклональное антитело против HuCD16. DART, описанный здесь, состоит из двух ковалентно связанных цепей, каждая из которых имеет VL, за которым следует спейсер, затем VH, за которым следует Cys в подходящем контексте для образования дисульфидной связи на противоположной цепи. Последовательность DART, кодирующую 3G8VL-GlyGlyGlySerGlyGlyGlyGly (SEQ ID NO: 10)-3G8VH-LeuGlyGlyCys амплифицировали посредством ПЦР с существующей эукариотической экспрессирующей конструкцией и расщепляли посредством Nco I и EcoR I. Вектор-мишень представлял собой pET25b (+) (Novagen), который содержит лидерную последовательность apelB для секреции в E. coli. Перед встраиванием последовательностей DART 3G8/3G8, вектор модифицировали следующим образом: сначала промотор T7 заменяли промотором lac с более низкой активностью для способствования экспрессии растворимых, хотя и на более низком уровне, белков под его контролем. Кроме того, вносили две точковые мутации для устранения двух внутренних кодонов Met, присутствующих в начале участка множественного клонирования (MCS) для обеспечения инициации в Met, находящемся в начале лидерной последовательности pelB. DART, который продуцировался посредством этой конструкции, состоит из двух плечей V-области, которые обладают одной специфичностью, а именно HuCD16.
Экспрессия: Клетки BL21DE3 (Novagen) трансформировали плазмидой pET25b(+) T7-lac+ 3G8/3G8 и для посева в культуральный бульон использовали устойчивую к amp колонию. Когда культура достигала OD600 0,5 единиц, добавляли 0,5 мМ IPTG для индукции экспрессии. Культуру выращивали при 30°С в течение 2 часов и бесклеточную среду собирали.
Очистка: DART 3G8/3G8 очищали в двухстадийном процессе с использованием аффинной и эксклюзионной хроматографии. DART улавливали из кондиционированной среды с использованием аффинной хроматографии. Конкретно, CD16A связывали с активированной посредством CNBr Sepharose 4B (GE Healthcare). Смолу CD16A-Sepharose уравновешивали в 20 мМ Tris/HCl, pH 8,0, перед загрузкой. После завершения загрузки смолу промывали буфером для уравновешивания перед элюированием связанного DART посредством 50 мМ глицина pH 3,0. Элюированный DART сразу нейтрализовывали посредством 1 M Tris/HCl, pH 8,0, и концентрировали с использованием концентратора центрифужного типа (Vivaspin 20, 10k MWCO PES, VivaScience Inc.). Концентрированный DART далее очищали эксклюзионной хроматографией с использованием колонки Superdex 200 (GE Healthcare), уравновешенной в PBS.
Результаты
1,7 литров культуральной кондиционированной среды E. coli пропускали через колонку с CD16A Sepharose. Выход DART составил 0,12 мг. Анализ очищенного DART посредством SDS-PAGE и SEC продемонстрировал сопоставимость с клеткой млекопитающих (CHO), экспрессировавшей контрольный DART (ФИГ. 27).
ELISA связывания DART h3G8-h3G8, экспрессированного из E. coli: Экспрессию DART h3G8-h3G8 в E. coli измеряли с использованием ELISA. 50 мкл/лунка антитела, специфичного к Fv h3G8 2C11, в концентрации 2 мкг/мл наносили на 96-луночный планшет Maxisorp в карбонатном буфере при 4°C в течение ночи. Планшет промывали три раза посредством PBS-T (PBS, 0,1% Tween 20), а затем блокировали посредством 0,5% BSA в PBS-T в течение 30 минут при комнатной температуре перед добавлением тестируемого DART. В процессе блокирования E. coli, экспрессировавшие DART h3G8-h3G8, DART h2B6-h3G8 и DART h2B6-h2B6 (отрицательный контроль), разбавляли до 1 мкг/мл и 0,3 мкг/мл в PBST/BSA. В каждую лунку добавляли 50 мкл/лунка разбавленных DART. Планшет инкубировали при комнатной температуре в течение 1 часа. После промывания посредством PBS-T три раза, в планшет добавляли 50 мкл/лунка биотинилированной слитой молекулы sCD16-Fc в концентрации 0,1 мкг/мл. Планшет инкубировали при комнатной температуре в течение 1 часа. После промывания посредством PBS-T три раза для детекции использовали 50 мкл/лунка конъюгированного с HRP стрептавидина в разведении 1:5000 (Amersham Pharmacia Biotech) и инкубировали при комнатной температуре в течение 1 часа. Планшет промывали посредством PBS-T три раза и проявляли с использованием 80 мкл/лунка субстрата TMB. После инкубации в течение 5 минут реакцию останавливали посредством 40 мкл/лунка 1% H2SO4. Считывание при OD450 нм проводили с использованием 96-луночного устройства для считывания планшетов и программного обеспечения SOFTmax. Данные считывания наносили на график с использованием программного обеспечения GraphPadPrism 3.03 (ФИГ. 28).
6.8 ИНДУЦИРОВАННАЯ DART ГИБЕЛЬ B-КЛЕТОК ЧЕЛОВЕКА
PBMC человека инкубировали в течение ночи с: CD16-CD32B-hu3G8-hu2b6 (описанным выше); агликозилированным химерным антителом 2B6 ch2B6-aglyc (описанным в совместно рассматриваемой патентной заявке США с серийным № 11/108135, опубликованной в качестве US2005/0260213, включенной в настоящий документ в качестве ссылки) и CD16-CD79. ДНК и кодируемые белковые последовательности CD16-CD79 являются следующими:
H3G8VL-CB3.1VH
Нуклеотидная последовательность (SEQ ID NO: 226):

Аминокислотная последовательность (SEQ ID NO: 227):

CB3.1VL-h3G8VH
Нуклеотидная последовательность (SEQ ID NO: 228):

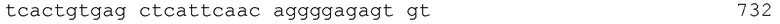
Аминокислотная последовательность (SEQ ID NO: 229):

Апоптоз анализировали посредством FACS-анализа в качестве процента популяции B-клеток (CD20+ клеток) PI+Аннексин-V+ от общей популяции FSC/SSC, не находящейся в области (ФИГ. 29).
6.9 DART 8B5-CB3.1
8B5VL-CB3.1VH-VEPKSC
8B5VL амплифицировали с использованием H9 и lgh630R в качестве праймеров, ch8B5Lc в качестве матрицы. CB3.1VH амплифицировали с использованием lgh628F и lgh629R в качестве праймеров, ch8B5Hc в качестве матрицы. В праймеры lgh630R и lgh628F была включена линкерная последовательность. В праймер lgh629R были включены c-концевой линкер и стоп-кодон. Продукты ПЦР очищали из геля и смешивали в равном молярном соотношении, затем амплифицировали с использованием H9 и lgh629R в качестве праймеров. Затем продукт ПЦР с перекрыванием расщепляли эндонуклеазами рестрикции NheI/EcoRI и клонировали в вектор pCIneo.
CB3.1VL-8B5VH-FNRGEC
VL CB3.1 амплифицировали с использованием H9 и lgh630R, которые обладали той же последовательностью, что и 8B5VL в FR4, в качестве праймеров, и chCB3.1Lc в качестве матрицы. 8B5VH амплифицировали с использованием lgh631F и lgh640R в качестве праймеров и ch8B5Hc в качестве матрицы. Линкерную последовательность включали в праймеры lgh630R и lgh631F. В праймер lgh640R были включены c-концевой линкер и стоп-кодон. Продукты ПЦР очищали из геля и смешивали вместе в равном молярном соотношении, затем амплифицировали с использованием H9 и lgh640R в качестве праймеров. Затем продукт ПЦР с перекрыванием расщепляли эндонуклеазами рестрикции NheI/EcoRI и клонировали в вектор pCIneo.
Антитело против Flag-метки-8B5VL-CB3.1VH-VEPKSC
Антитело против Flag-метки встраивали между сигнальной последовательностью и 8B5VL посредством ПЦР с перекрыванием. Сигнальную последовательность и Flag-метку амплифицировали с использованием H9 и lgh647R в качестве праймеров и ch8B5Lc в качестве матрицы. 8B5VL-CB3.1VH-VEPKSC повторно амплифицировали с использованием lgh647F и lgh629R в качестве праймеров и 8B5VL-CB3.1VH-VEPKSC в качестве матрицы. Продукты ПЦР очищали из геля и смешивали вместе в равном молярном соотношении, затем амплифицировали с использованием H9 и lgh629R в качестве праймеров. Затем продукт ПЦР с перекрыванием расщепляли эндонуклеазами рестрикции NheI/EcoRI и клонировали в вектор pCIneo.
8B5VL-CB3.1VH-LGGC
Для получения отличающегося C-концевого линкера в конструкции 8B5VL-CB3.IVH-VEPKSC конструкцию повторно амплифицировали с использованием H9 и lgh646R в качестве праймеров. В праймер lgh646R был встроен C-концевой линкер LGGC. Затем продукт ПЦР расщепляли эндонуклеазами рестрикции NheI/EcoRI и клонировали в вектор pCIneo.
CB3.1VL-8B5VH-LGGC
Ту же стратегию использовали для создания CB3.1VL-8B5VH-LGGC. В праймер lgh648R был встроен C-концевой линкер LGGC и в качестве матрицы использовали CB3.1VL-8B5VH-FNRGEC. Затем продукт ПЦР расщепляли эндонуклеазами рестрикции NheI/EcoRI и клонировали в вектор pCIneo.
Антитело против Flag-метки-8B5VL-CB3.1VH-LGGC
Ту же стратегию также использовали для создания антитела против Flag-метки-8B5VL-CB3.1VH-LGGC. В праймер lgh648R был встроен C-концевой линкер LGGC и в качестве матрицы использовали антитело против Flag-метки-8B5VL-CB3.1VH-VEPKSC. Затем продукт ПЦР расщепляли эндонуклеазами рестрикции NheI/EcoRI и клонировали в вектор.
Нуклеотидная последовательность 8B5-CB3.1-VEPKSC (SEQ ID NO: 230):

Аминокислотная последовательность 8B5-CB3.1-VEPKSC (SEQ ID NO: 231):

Нуклеотидная последовательность CB3.1-8B5-FNRGEC (SEQ ID NO: 232):

Аминокислотная последовательность CB3.1-8B5-FNRGEC (SEQ ID NO: 233):

8B5VL-CB3.1VH-LGGC
8B5VL амплифицировали с использованием H9 и lgh694R в качестве праймеров, ch8B5Lc в качестве матрицы. 8B5VH амплифицировали с использованием lgh695F и lgh696R в качестве праймеров, ch8B5Hc в качестве матрицы. Линкерную последовательность включали в праймеры lgh694R и lgh695F. HuIgG1Fc амплифицировали с использованием lgh355F и lgh366R в качестве праймеров, ch8B5Hc в качестве матрицы. Продукты ПЦР очищали из геля и смешивали в равном молярном соотношении, затем амплифицировали с использованием H9 и lgh366R в качестве праймеров. Затем продукт ПЦР с перекрыванием расщепляли эндонуклеазами рестрикции NheI/EcoRI и клонировали в вектор pCIneo.
Нуклеотидная последовательность 8B5VL-CB3.1VH-LGGC (SEQ ID NO: 234):

Аминокислотная последовательность 8B5VL-CB3.1VH-LGGC (SEQ ID NO: 235):

Нуклеотидная последовательность CB3.1-8B5-LGGC (SEQ ID NO: 236):

Аминокислотная последовательность CB3.1-8B5-LGGC (SEQ ID NO: 237):

Праймеры:

Экспрессия: Экспрессирующие плазмиды, кодирующие конструкции 5 и 6, или 6 и 7, или 8 и 9, или 9 и 10 (ФИГ. 30), котрансфицировали в клетки HEK-293 для экспрессии DART 8B5-CB3.1 с антителом против flag-метки или без него с использованием Lipofectamine 2000 (Invitrogen). Кондиционированную среду собирали три раза каждые трое суток. Затем кондиционированную среду очищали с использованием аффинной колонки CD32B.
ELISA: ELISA проводили следующим образом: 50 мкл/лунка CD32B-Fc в концентрации 2 мкг/мл наносили на 96-луночный планшет Maxisorp в карбонатном буфере при 4°C в течение ночи. Планшет промывали три раза посредством PBS-T (PBS, 0,1% Tween 20), а затем блокировали посредством 0,5% BSA в PBS-T в течение 30 минут при комнатной температуре, а затем добавляли тестируемый одноцепочечный слитый белок Fc. В процессе блокирования DART 8B5-CB3.1 серийно разбавляли двукратными разведениями, начиная с 2 мкг/мл. 25 мкл/лунка разбавленного DART, смешанного с 25 мкл/лунка ch8B5 в концентрации 50 нг/мл переносили из планшета для разведения в планшет для ELISA. Планшет инкубировали при комнатной температуре в течение 1 часа. После промывания три раза посредством PBS-T в планшет добавляли 50 мкл/лунка разбавленного 1:10000 конъюгированного с HRP F(ab')2 козы против F(ab')2 IgG человека (Jackson ImmunoResearch). Планшет инкубировали при комнатной температуре в течение 1 часа. Планшет промывали три раза посредством PBS-T и проявляли посредством 80 мкл/лунка субстрата TMB. После инкубации в течение 5 минут реакцию останавливали посредством 40 мкл/лунка 1% H2SO4. Считывание при OD450 нм проводили с использованием 96-луночного устройства для считывания планшетов и программного обеспечения SOFTmax. Данные считывания наносили на график с использованием программного обеспечения GraphPadPrism 3.03 (ФИГ. 31).
6.10 КОНСТРУИРОВАНИЕ И ОХАРАКТЕРИЗАЦИЯ Ig-ПОДОБНОГО ЧЕТЫРЕХВАЛЕНТНОГО DART
Для получения Ig-подобных видов DART использовали четыре полипептидных цепи, имеющих четырехвалентные антигенсвязывающие центры (фигура 32; фигура 33). Ig-подобные типы DART имеют уникальные свойства, поскольку их домены можно конструировать так, чтобы они связывались с тем же эпитопом (так чтобы образовывать четырехвалентный специфичный к одному эпитопу Ig-подобный DART, способный связывать четыре идентичных молекулы антигена), или с различными эпитопами или антигенами. Например, его домены можно конструировать, чтобы они связывались с двумя эпитопами одного антигена (так чтобы образовывался четырехвалентный специфичный к одному антигену, специфичный к двум эпитопам Ig-подобный DART), или с эпитопами на различных молекулах антигенов, чтобы образовывался четырехвалентный Ig-подобный DART, имеющий пару участков связывания, специфичных к первому антигену, и вторую пару участков связывания, специфичную ко второму антигену). Можно легко получать гибридные молекулы, обладающие комбинациями таких свойств.
Для иллюстрации характеристик таких Ig-подобных типов DART продуцировали иллюстративый четырехвалентный Ig-подобный тип DART, имеющий пару участков связывания для CD32 и вторую пару участков связывания, специфичных к CD16A. Этот Ig-подобный тип DART продуцировали с использованием следующих четырех полипептидных цепей:
Нуклеотидная последовательность 2.4G2-3G8-hKappa (SEQ ID NO: 247):

Аминокислотная последовательность, кодируемая 2.4G2-3G8-hKappa (SEQ ID NO: 248):

Нуклеотидная последовательность 3G8-2.4G2-hG1 (SEQ ID NO: 249):


Аминокислотная последовательность, кодируемая 3G8-2.4G2-hG1 (SEQ ID NO: 250):

Препараты Ig-подобных молекул DART, имеющие указанные выше последовательности, получали из различных плазмидных изолятов и были обозначены как "Ig DART 1" и "Ig DART 2". Способность этих Ig-подобных типов DART связывать mCD32-hCD16A в ELISA сравнивали с этой способностью среды отдельно, DART, имеющим единичный участок связывания CD32 и единичный участок связывания CD16A ("DART"), и контрольным mAb против ch-mCD32 (фигура 34). Было выявлено, что Ig-подобный DART по настоящему изобретению имеет значительно более высокую аффинность связывания антигена, чем DART или контрольное антитело.
6.11 КОНСТРУИРОВАНИЕ И ОХАРАКТЕРИЗАЦИЯ БИСПЕЦИФИЧЕСКИХ ДИАНТИТЕЛ CD32B-CD79-1 и CD32B-CD79-2
Гены, кодирующие CD79VL-CD32BVH (последовательность 1), CD32BVL-CD79VH-1 (последовательность 2) и CD32BVL-CD79VH-2 (последовательность 3) клонировали в экспрессирующий вектор pEE13 с получением экспрессирующих конструкций 1, 2 и 3, соответственно. Экспрессирующую конструкцию 1 плазмиду котрансфицировали либо с экспрессирующей плазмидой 2, либо с экспрессирующей плазмидой 3 в клетки HEK-293 с получением биспецифических диантител CD32B-CD79-1 и CD32B-CD79-2, соответственно. Кондиционированную среду собирали три раза каждые трое суток. Затем кондиционированную среду очищали с использованием аффинной колонки к CD32B.
ELISA проводили следующим образом: 50 мкл/лунка CD32B-Fc в концентрации 2 мкг/мл наносили на 96-луночный планшет Maxisorp в карбонатном буфере при 4°C в течение ночи. Планшет промывали три раза посредством PBS-T (PBS, 0,1% Tween 20), а затем блокировали посредством 0,5% BSA в PBS-T в течение 30 минут при комнатной температуре, а затем добавляли тестируемый одноцепочечный слитый белок Fc. В процессе блокирования биспецифическое диантитело CD32B-CD79-1 или CD32B-CD79-2 серийно разбавляли двукратными разведениями, начиная с 2 мкг/мл. 25 мкл/лунка разбавленного биспецифического диантитела смешивали с 25 мкл/лунка антитела против CD32B в концентрации 50 нг/мл и добавляли в планшет для ELISA. Планшет инкубировали при комнатной температуре в течение 1 часа. После промывания три раза посредством PBS-T, в планшет добавляли 50 мкл/лунка разбавленного 1:10000 конъюгированного с HRP F(ab')2 козы против F(ab')2 IgG человека (Jackson ImmunoResearch). Планшет инкубировали при комнатной температуре в течение 1 часа. Планшет промывали три раза посредством PBS-T и проявляли посредством 80 мкл/лунка субстрата TMB. После инкубации в течение 5 минут реакцию останавливали посредством 40 мкл/лунка 1% H2SO4. Считывание при OD450 нм проводили с использованием 96-луночного устройства для считывания планшетов и программного обеспечения SOFTmax. Данные считывания наносили на график с использованием программного обеспечения GraphPadPrism 3.03. Этот эксперимент показал, что биспецифические диантитела CD32B-CD79-1 и CD32B-CD79-2 были способны иммуноспецифично связываться с CD32-Fc с аффинностью, эквивалентной аффинности контрольного антитела против CD32B. Нуклеотидные и кодируемые ими аминокислотные последовательности описанных выше конструкций представлены ниже:
Последовательность 1 - нуклеотидная последовательность CD79VL-CD32BVH (SEQ ID NO: 251):


Последовательность 2 - аминокислотная последовательность CD79VL-CD32BVH (SEQ ID NO: 252):

Последовательность 3 - нуклеотидная последовательность CD32BVL-CD79VH-1 (SEQ ID NO: 253):

Последовательность 4 - аминокислотная последовательность CD32BVL-CD79VH-1 (SEQ ID NO: 254):

Последовательность 5 - нуклеотидная последовательность CD32BVL-CD79VH-2 (SEQ ID NO: 255):


Последовательность 6 - аминокислотная последовательность CD32BVL-CD79VH-2 (SEQ ID NO: 256):

6.12 КОНСТРУИРОВАНИЕ И ОПТИМИЗАЦИЯ БИФУНКЦИОНАЛЬНЫХ ДИАНТИТЕЛ H8B5-HBCRC
Конструировали диантитело, которое содержит вариабельные области, способные связываться с CD32 и B-клеточным рецепторным комплексом ("BCRC").
Клонирование. Конструкции конструировали с использованием стандартной ПЦР/ПЦР с перекрыванием: h8B5VL-G3SG4-hBCRCVH M48I-LGGC:
Полностью гуманизированный VL 8B5 (распознающий CD32) амплифицировали с использованием lgh321F и lgh788R в качестве праймеров. hBCRCVH M48I амплифицировали с использованием lgh784F и lgh386R в качестве праймеров. Продукты ПЦР очищали из геля и смешивали и амплифицировали с использованием lgh321F и lgh386R. Затем фрагмент ПЦР с перекрыванием встраивали в pEE6 в участке XbaI-EcoRI. "G3SG4" представляет собой линкер, имеющий последовательность: GGGSGGGG (SEQ ID NO: 10).
hBCRCVL R45N-G3SG4-h8B5VH-LGGC
hBCRCVL R45N амплифицировали с использованием lgh321F и lgh785R в качестве праймеров. h8B5VH амплифицировали с использованием lgh787F и lgh786R в качестве праймеров. Продукты ПЦР очищали из геля и смешивали и амплифицировали с использованием lgh321F и lgh786R. Затем фрагмент ПЦР с перекрыванием клонировали в pEE13 в участке XbaI-EcoRI.
Конструирование единого вектора. pEE6hHBCRCVL R45N-h8B5VH расщепляли в участках Bgl II-Sal I и фрагмент размером 3,3 т.п.н. очищали и встраивали в pEE13 hHBCRCVL 45N-h8B5VH в участках BamHI-SalI. Bgl II и BamH I обладают общими совместимыми липкими концами. Последовательность DART и праймеров, использованных для конструирования DART, представлены ниже:
Нуклеотидная последовательность hHBCRCVL. R45N-h8B5VH-LGGC (SEQ ID NO: 257):

Аминокислотная последовательность hHBCRCVL. R45N-h8B5VH-LGGC (SEQ ID NO: 258):

Нуклеотидная последовательность H8B5VL-hHBCRCVH M48I-LGGC (SEQ ID NO: 259):

Аминокислотная последовательность H8B5VL-HBCRCVH M48I-LGGC (SEQ ID NO: 260):

Праймер Lgh321F (SEQ ID NO: 261):
cgagctagct ctagatgaga tcacagttct ctctac 36
Праймер Lgh386R (SEQ ID NO: 262):
tttgaattct agcagcctcc cagtgaggag acggtgaccg tggtc 45
Праймер Lgh784F (SEQ ID NO: 263):
ggcggatccg gaggcggagg ccaggttcag ctggtgcag 39
Праймер Lgh785R (SEQ ID NO: 264):
cctccggatc cgcctccttt gatctcaagc ttggtccc 38
Праймер Lgh786R (SEQ ID NO: 265):
tttgaattct agcagcctcc caggctggag acggtcacca gg 42
Праймер Lgh787F (SEQ ID NO: 266):
ggaggcggat ccggaggcgg aggcgaagtg cagcttgtgg agtc 44
Экспрессирующую плазмиду Hu3G8VL 1-G3SG4-Hu2B6VH 4-LGGC котрансфицировали вместе с Hu2B6VL 5-G3SG4-Hu3G8VH 5-LGGC в клетки HEK-293 с получением биспецифического диантитела Hu2B6 4.5-Hu3G8 5.1, распознающего CD32 и CD79. В то же время Hu2B6VL 5-G3SG4-Hu2B6VH 4-LGGC и Hu3G8VL 1-G3SG4-Hu3G8VH 5-LGGC трансфицировали по отдельности в клетки HEK-293 с получением диантитела Hu2B6 4.5 и Hu3G8 5.1. После трех суток в культуре кондиционированную среду собирали и охарактеризовывали посредством связывающего ELISA. Результат этого эксперимента представлен на фигуре 36.
Схема эксперимента: 100 нг/лунка растворимого FcRIIb-G2-Agly наносили на 96-луночный планшет Maxisorp в карбонатном буфере при 40°C в течение ночи. Планшет промывали три раза посредством PBS/0,1% Tween20, а затем блокировали 0,5% BSA в PBS/0,1% Tween 20 в течение 30 мин при комнатной температуре перед добавлением диантител. В каждую лунку добавляли серийное двукратное разведение среды, кондиционированной биспецифическим диантитеом Hu2B6 4.5-Hu3G8 5.1, диантителом Hu2B6 4.5 и диантителом hu3G8 5.1, начиная с 25 нг/лунка. Планшет инкубировали при комнатной температуре в течение 1 часа. После промывания три раза посредством PBS/0,1% Tween20 в планшет добавляли 10 нг/лунка FcRIIIa-G2-биотин. Планшет инкубировали при комнатной температуре в течение 1 часа. После промывания три раза посредством PBS/0,1% Tween20 50 мкл разведения 1:5000 конъюгированного с HRP стрептавидина (Amersham Pharmacia Biotech) использовали для детекции. После инкубации в течение 45 минут при комнатной температуре планшет промывали три раза посредством PBS/0,1% Tween20 и проявляли с использованием субстрата TMB. После инкубации в течение 10 минут реакцию останавливали посредством 1% H2SO4. Считывание при OD450 нм проводили с помощью программы SOFTmax. Данные считывания наносили на график с использованием программного обеспечения GraphPadPrism 3.03.
6.13 КОНСТРУИРОВАНИЕ ДИАНТИТЕЛ IgDART
Конструировали диантитела IgDART, которые содержат вариабельные области, способные связываться с CD32 и B-клеточным рецепторным комплексом ("BCRC"). В первом диантителе использовали линкер LGGCGGGS (SEQ ID NO: 267) между последовательностями VH и последовательностями Fc молекулы. Во втором диантителе использовали либо линкер LEIK, имеющий последовательность: LEIK (SEQ ID NO: 268), либо линкер TVSS, имеющий последовательность TVSS (SEQ ID NO: 269). Последовательности цепей этих диантител и кодирующих полинуклеотидов представлены ниже:
H8B5VL-hBCRCVH M48I, M62K_LGGCG3S_hKappa (SEQ ID NO: 270):

Последовательности H8B5VL слиты с последовательностями hBCRCVH посредством линкера GGGSGGGG (SEQ ID NO: 10), расположенного в положении 108-115. Последовательности hBCRCVH слиты с последовательностями Fc посредством линкера LGGCGGGS (SEQ ID NO: 267), расположенного в положении 229-236 (оба показаны посредством подчеркивания, выше). Полинуклеотид, кодирующий последовательность H8B5VL-hBCRCVH M48I, M62K_LGGCG3S_hKappa, представляет собой: (SEQ ID NO: 271):


где последовательности, кодирующие линкеры: GGGSGGGG (SEQ ID NO: 10) и LGGCGGGS (SEQ ID NO: 267), расположены в положении 322-345 и 685-708, соответственно (оба показаны подчеркиванием, выше).
HBCRCVL R45N-h8B5VH_LGGCGGGS-hG1 (SEQ ID NO: 272):

Последовательности hBCRCVL слиты с последовательностями H8B5VH посредством линкера GGGSGGGG (SEQ ID NO: 10), расположенного в положении 113-120. Последовательности H8B5VH слиты с Fc-последовательностями посредством линкера LGGCGGGS (SEQ ID NO: 267), расположенного в положении 237-244 (оба показаны подчеркиванием, выше). Полинуклеотид, кодирующий последовательность HBCRCVL R45N-h8B5VH_LGGCGGGS-hG1, представляет собой: (SEQ ID NO: 273):


где последовательности, кодирующие линкеры: GGGSGGGG (SEQ ID NO: 10) и LGGCGGGS (SEQ ID NO: 267), расположены в положении 337-360 и 709-732, соответственно (оба показаны подчеркиванием, выше).
H8B5VL-HBCRCVH M48I, M62K_(-4)LEIK_hKappa (SEQ ID NO: 274):

Последовательности H8B5VL слиты с последовательностями HBCRCVH посредством линкера GGGSGGGG (SEQ ID NO: 10), расположенного в положении 108-115. Последовательности HBCRCVH слиты с последовательностями Fc посредством линкера LEIK (SEQ ID NO: 268), расположенного в положении 225-228 (оба показаны подчеркиванием, выше). Полинуклеотид, кодирующий последовательность H8B5VL-HBCRCVH M48I, M62K_(-4)LEIK_hKappa, представляет собой: (SEQ ID NO: 275):


где последовательности, кодирующие линкеры: GGGSGGGG (SEQ ID NO: 10) и LEIK (SEQ ID NO: 268), расположены в положении 322-345 и 673-684, соответственно (обе показаны подчеркиванием, выше).
HBCRCVL R45N-h8B5VH_(-4)TVSS-hG1 = HBCRCVL R45N-h8B5VH_-hG1 (SEQ ID NO: 276):

Последовательности HBCRCVL слиты с последовательностями h8B5VH посредством линкера GGGSGGGG (SEQ ID NO: 10), расположенного в положении 113-120. Последовательности h8B5VH слиты с последовательностями Fc посредством линкера TVSS (SEQ ID NO: 269), расположенного в положении 233-236 (оба показаны подчеркиванием, выше). Полинуклеотид, кодирующий HBCRCVL R45N-h8B5VH_(-4)TVSS-hG1, представляет собой: (SEQ ID NO: 277):


где последовательности, кодирующие линкеры: GGGSGGGG (SEQ ID NO: 10) и TVSS (SEQ ID NO: 269), расположены в положении 337-360 и 697-708, соответственно (обе показаны подчеркиванием, выше).
6.14 ОПТИМИЗАЦИЯ ЛИНКЕРОВ
Как рассмотрено выше, диантитела IgDART по настоящему изобретению предпочтительно содержат линкеры между последовательностями VH и последовательностями Fc молекулы. Эксперименты проводили для оптимизации линкеров в целях максимизации выхода и активности. Использовали следующие линкеры.
Описанные выше линкеры встраивали в плазмиды для получения набора диантител IgDART, имеющих различные комбинации линкеров:
(мкг/мл)
Определяли агрегационные свойства продуцированных IgDART.
Данные неожиданно показали, что конструкции, имеющие линкеры, такие как линкеры, использованные в 901A/901B, 903A/903B и 908A/908B, привели к значительно лучшим результатам (меньшая олигомеризация и/или меньшее образование фрагментов), чем конструкции, имеющие линкеры, такие как 910A/911B.
6.15 DART С E-СПИРАЛЬЮ/K-СПИРАЛЬЮ
Как будет понятно ввиду вышесказанного, отдельные полипептиды биспецифического DART могут образовывать два типа гомодимеров и один тип гетеродимера. В одном из вариантов осуществления настоящего изобретения, на C-конец одного или, более предпочтительно, обоих полипептидов DART можно добавлять заряженный полипептид. Путем выбора заряженных полипептидов с противоположным зарядом для отдельных полипептидов биспецифического DART, включение таких заряженных полипептидов способствует образованию гетеродимеров и уменьшает образование гомодимеров. Предпочтительно, положительно заряженный полипептид имеет значительное содержание аргинина, глутамина, гистидина и/или лизина (или смеси таких аминокислот), и отрицательно заряженный полипептид имеет значительное содержание аспартата или глутамата (или смеси таких аминокислоты). Положительно заряженные полипептиды, имеющие значительное содержание лизина, и отрицательно заряженные полипептиды, имеющие значительное содержание глутамата, являются особенно предпочтительными. Для максимизации электростатического притяжения между такими противоположно заряженными полипептидами, предпочтительно использовать полипептиды, способные самопроизвольно принимать конформацию спирали.
Таким образом, в предпочтительном варианте осуществления положительно заряженная "E-спираль" присоединена к одному из полипептидов, используемых для образования биспецифического DART, и отрицательно заряженная "K-спираль" присоединена ко второму из полипептидов DART (фигура 37).
Особенно предпочтительная E-спираль имеет последовательность: (EVAALEK)4:
SEQ ID NO: 299 EVAALEKEVAALEKEVAALEKEVAALEK
Особенно предпочтительная K-спираль имеет последовательность: (KVAALKE)4:
SEQ ID NO: 300 KVAALKEKVAALKEKVAALKEKVAALKE
Предпочтительный полипептид DART, обладающий такой E-спиралью, имеет общую последовательность: [VL домен]- [GGGSGGGG]-[VH-домен]-[(EVAALEK)4]-GGGNS, где VL представляет собой вариабельный домен легкой цепи Ig DART, GGGSGGGG представляет собой SEQ ID NO: 10, VH представляет собой вариабельный домен тяжелой цепи Ig DART, (EVAALEK)4 представляет собой SEQ ID NO: 299, и GGGNS представляет собой SEQ ID NO: 301. Предпочтительный полипептид DART, обладающий такой K-спиралью, имеет общую последовательность: [VL-домен]-[GGGSGGGG]-[VH-домен] -[(KVAALKE)4]-GGGNS, где VL представляет собой вариабельный домен легкой цепи Ig DART, GGGSGGGG представляет собой SEQ ID NO: 10, VH представляет собой вариабельный домен тяжелой цепи Ig DART, (KVAALKE)4 представляет собой SEQ ID NO: 300, и GGGNS представляет собой SEQ ID NO: 301.
6.16 FC-СОДЕРЖАЩИЕ DART С E-СПИРАЛЬЮ/K-СПИРАЛЬЮ
В следующем варианте осуществления Fc-области могут быть связаны с E- и/или K-спиралями в DART с E-спиралью или K-спиралью.
Далее, разделение между Fc-областями и VH-доменом DART в Fc-содержащем DART является желательным в случаях, в которых менее разделенное расположение таких доменов приводит к сниженному взаимодействию между такими доменами и их связывающими лигандами или иным образом препятствует сборке DART. Хотя можно использовать разделители с любой аминокислотной последовательностью, предпочтительно использовать разделители, которые образуют витки α-спирали, так чтобы максимально вытянуть и расположить Fc-домен в отдалении от вариабельных доменов (фигура 37). Поскольку описанные выше спиралевидные полипептиды с противоположным зарядом, кроме того, функционируют, обеспечивая образование гетеродимера, такие молекулы являются особенно предпочтительными разделителями. Такие содержащие спираль молекулы Fc-DART обеспечивают преимущества, сходные с преимуществами Fc-DARTS, включая улучшенное время полужизни в сыворотке и приращение эффекторной функции. Описанные выше полипептиды с E-спиралью и K-спиралью особенно предпочтительны для этой цели.
Таким образом, в предпочтительном варианте осуществления Fc-содержащий DART с E-спиралью имеет общую последовательность: [VL-домен]-[GGGSGGGG]-[VH-домен]-[(EVAALEK)4]-GGG-Fc-домен, начиная с D234 (нумерация по Kabat), где VL представляет собой вариабельный домен легкой цепи Ig DART, GGGSGGGG представляет собой SEQ ID NO: 10, VH представляет собой вариабельный домен тяжелой цепи Ig DART, и (EVAALEK)4 представляет собой SEQ ID NO: 299.
Аналогично, в предпочтительном варианте осуществления Fc-содержащий DART с K-спиралью имеет общую последовательность: [VL-домен]-[GGGSGGGG]-[VH-домен]-[(KVAALKE)4]-GGG-Fc-домен, начиная с D234 (нумерация по Kabat), где VL представляет собой вариабельный домен легкой цепи Ig DART, GGGSGGGG представляет собой SEQ ID NO: 10, VH представляет собой вариабельный домен тяжелой цепи Ig DART, и (KVAALKE)4 представляет собой SEQ ID NO: 300.
Как указано выше, содержащая спираль молекула DART или содержащая спираль Fc-содержащая молекула DART может содержать только один такой разделитель спиралей, или она может содержать более одного такого разделителя (например, два разделителя, предпочтительно противоположного заряда, один из которых связан с каждым VH-доменом полипептидов DART). Путем связывания Fc-области такой молекулы (молекул) разделителя способность создавать двухвалентные, четырехвалентные и т.д. варианты молекул Fc-DART посредством обмена цепями увеличивается (фигура 39). Как показано на фигуре 39, можно продуцировать молекулы Fc-DART, которые образуют мономеры или димеры, в зависимости от того, связан Fc-домен с одним или с обоими из VH-доменов DART.
6.17 ФУНКЦИОНАЛЬНАЯ АКТИВНОСТЬ FC-СОДЕРЖАЩИХ DART С E-СПИРАЛЬЮ/K-СПИРАЛЬЮ
Типы Fc-DART с E-спиралью и/или K-спиралью продуцировали из биспецифических молекул DART, имеющих: (1) вариабельные области легкой и тяжелой цепей антитела CB3, реактивного к CD79b (комплексу BCR), и (2) вариабельные области легкой и тяжелой цепей низкоаффинного варианта (названного вариантом "YA") реактивного к CD32B антитела 2B6. Эта вариабельная область легкой цепи этого антитела отличается от вариабельной области антитела 2B6 наличием мутаций: N50Y и V51A. Таким образом, антитело YA2B6 имеет последовательность вариабельной области легкой цепи:

Последовательность вариабельной области тяжелой цепи этого антитела представляет собой:

или
Низкоаффинное антитело было выбрано, поскольку оно предпочтительно связывает CD32B в цис-положении на клетках, экспрессирующих CD79b (B-клетки). По существу эта конфигурация уменьшает взаимодействие с другими экспрессирующими CD32B клетками (моноциты, эндотелиальные клетки, печень), а также нежелательное транс-взаимодействие.
Получали производные с E-спиралью и/или K-спиралью и Fc-содержащие производные с E-спиралью и/или K-спиралью таких DART h2B6YAhCB3. Для анализа приблизительного размера и гетерогенности продуцированных молекул использовали эксклюзионную хроматографию. Как показано на фигуре 40, димеры образовывались из DART с E-спиралью/K-спиралью, имеющих одну присоединенную Fc-область, связанную с доменом K-спирали, а также из DART с E-спиралью/K-спиралью, имеющих одну присоединенную Fc-область, связанную с доменом E-спирали. Желаемые мономерные, а также димерные молекулы выделяли из препаратов, в которых Fc-области были связаны как с E-спиралями, так и с K-спиралями одной молекулы DART, причем мономер представляет собой основной образованный продукт. На фигуре 41 представлена возможная структура продуцированных молекул димеров.
Фракции, полученные эксклюзионной хроматографией, анализировали с использованием SDS-полиакриламидного гель-электрофореза для дальнейшего анализа структур продуцированных молекул (фигура 42). Производные DART с E-спиралью/K-спиралью (без Fc-области) мигрировали в качестве двух преобладающих полос, каждая из которых имеет массу приблизительно 28 кДа (соответствующую KFc-содержащему полипептиду и немного меньшему EFc-содержащему полипептиду), и менее заметную полосу на уровне приблизительно 49 кДа (соответствующую DART с E-спиралью/K-спиралью). Фракции мономеров Fc-содержащих производных DART с E-спиралью/K-спиралью (EFc/K или E/KFc) из эксклюзионной хроматографии продемонстрировали только полосу либо с большей, либо с меньшей молекулярной массой, на уровне приблизительно 28 кДа (соответствующую либо DART, представляющему собой KFc-содержащий DART (полоса, соответствующая большей молекулярной массой), либо EFc-содержащему DART (полоса, соответствующая меньшей молекулярной массе)). Материал преимущественно мигрировал на уровне приблизительно 49 кДа (что соответствовало DART с E-спиралью/K-спиралью). Также наблюдали значительно более высокую молекулярную массу.
Для охарактеризации продуцированных молекул проводили ELISA биспецифического связывания. Планшет для ELISA покрывали CD79. Затем с планшетом связывали DART. Детекцию связывания DART проводили с использованием sCD32B-биотина, с последующей инкубацией со стрептавидином-HRP. Как показано на фигуре 43, Fc-содержащие производные DART h2B6YAhCB3 с E-спиралью/K-спиралью (EFc/K или E/KFc) продемонстрировали значительное усиление связывания относительно DART h2B6YAhCB3, или производного DART h2B6YAhCB3 с EFc/KFc.
Связывание антител, которые связались с CD79b, приводит к активации B-клеток (Van Kooten, C. et al. (1997) "Cross-Linking Of Antigen Receptor Via Ig-B (B29, CD79b) Can Induce Both Positive And Negative Signals In CD40-Activated Human B Cells", Clin. Exp. Immunol. 110:509-515). Поскольку молекулы DART h2B6YAhCB3 способны связываться как с CD79b, так и с ингибиторным рецептором CD32B, они способны "привлекать" CD32B в участки связывания CD79b и, тем самым, блокировать пролиферацию B-клеток. Чтобы продемонстрировать эту способность, DART инкубировали с B-клетками, подвергнутыми воздействию антител, способных связывать связанные антитела против CD79b. Результаты этого эксперимента представлены на фигуре 44. Результаты демонстрируют, что антитела, направленные только против CD79b или CD32B (Ch2B6N297Q и ChCB3.1N297Q, соответственно), не ингибируют пролиферацию B-клеток. Производные DART с EFc/KFc h2B6YA×hCB3 были по существу более эффективными в отношении пролиферации B-клеток, также как и сам DART h2B6YA×hCB3 и контроль h2B6YA×hCB3VF. Было выявлено, что DART c E-спиралью/K-спиралью, имеющие только одну связанную Fc-область (производные DART E/KFc h2B6YA×hCB3 и производные DART EFc/K h2B6YA×hCB3), проявляют наиболее высокое ингибирование пролиферации B-клеток.
6.18 МОДИФИКАЦИИ DART ДЛЯ ИЗМЕНЕНИЯ ВРЕМЕНИ ПОЛУЖИЗНИ В СЫВОРОТКЕ IN VIVO
Как рассмотрено выше, небольшие рекомбинантные молекулы антител, такие как биспецифические одноцепочечные молекулы (например, имеющие молекулярную массу приблизительно 55 кДа), быстро выводятся из кровотока. Исследования фармакокинетики молекул DART in vivo у мышей продемонстрировало ожидаемое короткое терминальное время полужизни, составляющее приблизительно 2 часа.
В некоторых вариантах осуществления, например, при лечении острого воспалительного состояния, такое короткое время полужизни является желательным, однако в других вариантах осуществления, например, при лечении злокачественной опухоли и хронических заболеваний и состояний, предпочтительно, чтобы молекулы DART по настоящему изобретению проявляли более длительное время полужизни.
Для улучшения фармакокинетических свойств молекул DART in vivo для таких применений, молекулы DART можно модифицировать так, чтобы они содержали полипептидную часть связываемого сывороткой белка на одном или нескольких из концов молекулы DART. Наиболее предпочтительно, такая полипептидная часть связываемого сывороткой белка находится на C-конце молекулы DART. Особенно предпочтительная полипептидная часть связываемого сывороткой белка для этой цели представляет собой альбумин-связывающий домен (ABD) из стрептококкового белка G. Альбумин-связывающий домен 3 (ABD3) белка G штамма Streptococcus G148 является особенно предпочтительным.
Альбумин-связывающий домен 3 (ABD3) белка G штамма Streptococcus G148 состоит из 46 аминокислотных остатков, образующих стабильный трехспиральный пучок, и имеет широкую альбумин-связывающую специфичность (Johansson, M.U. et al. (2002) "Structure, Specificity, And Mode Of Interaction For Bacterial Albumin-Binding Modules", J. Biol. Chem. 277(10):8114-8120). Альбумин является наиболее распространенным белком в плазме и имеет время полужизни у человека 19 суток. Альбумин обладает несколькими участками связывания небольших молекул, которые позволяют ему нековалентно связываться с другими белками и, тем самым, продлить их время полужизни в сыворотке.
Чтобы продемонстрировать способность полипептидной части сывороточного белка продлевать время полужизни DART, ABD3-домен стрептококкового белка G подвергали слиянию с рекомбинантным биспецифическим DART (иммунореактивным к антигенам hCD16 и hCD32B) с получением рекомбинантной молекулы антитела hCD16-hCD32B ABD-DART (фигура 45). Этот ABD-DART продемонстрировал специфическое связывание с обоими антигенами, а также с сывороточным альбумином человека (HSA), и был способен перенацеливать эффекторные клетки in vitro. По сравнению с контрольным DART этот ABD-DART продемонстрировал значительное увеличение времени полужизни в сыворотке у мышей. Этот подход можно использовать в качестве целесообразного пути для увеличения времени полужизни потенциально важных фармацевтических средств, таких как DART, до более чем 90 минут, более 2 часов, более 5 часов, более 10 часов, более 20 часов и, наиболее предпочтительно, более 30 часов.
МАТЕРИАЛЫ И СПОСОБЫ:
Разработка и конструирование ABD DART: ABD DART hCD16-hCD32B получали с использованием в качестве цепи 1:
hCD16VL-G3SG4-hCD32BVH-K-спираль[(KVAALKE)4],
где CD16VL обозначает CD16VL 3G8, G3SG4 обозначает SEQ ID NO: 10, hCD32BVH обозначает CD32BVH 2B6, и (KVAALKE)4 обозначает SEQ ID NO: 300;
и в качестве цепи 2:
hCD32BVL-G3SG4-hCD16VH-GGCGGG-E-спираль[(EVAALEK)4]-GGGNS-ABD, где CD32BVL обозначает CD32BVL, G3SG4 обозначает SEQ ID NO: 10, hCD16VH обозначает CD16VH, GGCGGG представляет собой остатки 2-7 SEQ ID NO: 267, E-спираль [(EVAALEK)4] представляет собой SEQ ID NO: 299, GGGNS представляет собой SEQ ID NO: 301, и ABD представляет собой:
LAEAKVLANR ELDKYGVSDY YKNLINNAKT VEGVKALID EILAALP (SEQ ID NO: 304).
Таким образом, последовательность цепи 1 (h3G8VL1-G3SG4-h2B6VH4-K-спираль-GGGNS) представляет собой:

Предпочтительный полинуклеотид, кодирующий цепь 1 (h3G8VL1-G3SG4-h2B6VH4-K-спираль-GGGNS), представляет собой:

Таким образом, последовательность цепи 2 (h2B6VL5-G3SG4-h3G8VH5-E-спираль-GGGNS-ABD) представляет собой:

Предпочтительный полинуклеотид, кодирующий цепь 2 (h2B6VL5-G3SG4-h3G8VH5-E-спираль-GGGNS-ABD), представляет собой:

Каждый сегмент VL и VH амплифицировали посредством ПЦР с использованием DART hCD16-hCD32B в качестве матрицы. Для цепи 2 нуклеотидные последовательности, содержащие E-спираль и ABD, получали с помощью димера праймеров, а затем субклонировали на C-конце VH-области hCD16 с использованием рестрикционного расщепления и лигирования. Обе цепи клонировали в вектор pCIneo (Promega, inc.) в участках NheI-NotII. Затем отдельные плазмиды, несущие соответствующую цепь 1, расщепленные по NgoMIV-NheI, и экспрессирующие цепь 2 кассеты, расщепленные по BstBI-PmeI, клонировали в единую плазмиду для трансфекции в клетки CHO для получения стабильных клеточных линий.
Экспрессия и очистка белка. Для стабильной трансфекции клетки CHO-S трансфицировали плазмидной ДНК hCD16-hCD32B EK ABD-DART. Белок ABD-DART очищали аффинной хроматографией с использованием растворимого варианта антигена FcRIIB, связанного с активированной посредством CNBr Sepharose 4B. Концентрированный белок далее очищали эксклюзионной хроматографией с использованием Superdex 200HR 10/30.
Анализ связывания посредством ELISA: Для улавливания на основе CD-16 планшеты покрывали антигеном FcRIIB в концентрации 2 мкг/мл при 4°C в течение ночи. Затем планшеты блокировали 0,5% пептоном в PBS-T. Очищенные белки, разбавленные в двукратном серийном разведении, связывали с планшетом в течение 1 ч при комнатной температуре. Наконец, проводили детекцию с использованием биотинилированного CD32B (50 нг/мл), а затем конъюгированного с HRP стрептавидина (1/1000, BD-Pharm). Активность HRP измеряли путем добавления TMB и считывание планшета проводили в устройстве для считывания планшетов при OD450 нм.
Для улавливания сывороточного альбумина человека (HSA) планшеты покрывали посредством HSA в концентрации 2 мкг/мл при 4°C в течение ночи. После этого следовали той же процедуре для проведения ELISA с двойной аффинностью.
Анализ опосредуемой мононуклеарными клетками периферической крови ADCC: Цитотоксичность измеряли посредством анализа LDH. Мононуклеарные клетки периферической крови (PBMC) очищали из цельной крови человека (Lonza Walkersville, Inc, Gaithersburg, MD) центрифугированием в градиенте плотности Ficoll-Hypaque (Amersham Biosciences, Piscataway, NJ) в соответствии с инструкциями изготовителя. 2×104 клеток-мишеней высевают в каждую лунку круглодонного 96-луночного планшета для культивирования тканей. К клеткам в планшете добавляют от одного до четырех серийных разведений различных молекул DART или антител. После этого в те же лунки добавляют 6×105 PBMC. Затем планшет инкубируют в течение ночи в инкубаторе при 37°С и 5% CO2. Затем планшет центрифугируют при 1200 об./мин в течение 5 минут, 50 мкл супернатанта переносят в планшет для ELISA с плоским дном. В каждую лунку добавляют 50 мкл раствора субстрата LDH (Promega) и планшет инкубируют в течение 30 мин в темноте при комнатной температуре. Затем в каждую лунку добавляют 50 мкл стоп-раствора и проводят считывание планшета при 490 нм в течение одного часа. Процентную цитотоксичность каждой лунки вычисляют с помощью исходных данных O.D. в качестве
(образец- AICC)/(максимум для мишени - самопроизвольно для мишени)×100,
где AICC представляет собой антителонезависимую клеточную цитотоксичность. Кривую доза-эффект получают с использованием программного обеспечения Prism.
Фармакокинетические исследования: мышам C57B1/6 проводили однократную внутривенную инъекцию DART hCD16-hCD32B в дозе 5 мг/кг. Сыворотку мышей собирали до введения дозы, через 2, 30 мин; 1, 3, 6, 24 и 72 ч. Количественно определяли концентрацию DART hCD16-hCD32B в сыворотке. Фармакокинетические вычисления для DART hCD16-hCD32B проводили с помощью фармакокинетического пакета программ WinNonlin Professional 5.1 (Pharsight Corporation, США). Параметры определяли посредством некомпартментного анализа (NCA). Некомпартментный анализ был основан на модели (Model 201), требующей внутривенной инъекции лекарственного средства. Для вычисления параметров использовали линейный трапецеидальный способ.
РЕЗУЛЬТАТЫ
Исследование экспрессии и связывания посредством ELISA: ABD-DART hCD16-hCD32B экспрессировался эффективно в концентрации 6,5 мг на литр в клетках млекопитающих CHO-S. Активность связывания очищенного белка ABD-DART в отношении соответствующих антигенов оценивали посредством ELISA. Результаты показали, что ABD-DART hCD16-hCD32B связывается одновременно с обоими антигенами, CD16, а также CD32B (фигура 46A). Профиль связывания совпадает с контрольным связыванием белка DART hCD16-hCD32B. Аффинность очищенного ABD-DART hCD16-hCD32B в отношении сывороточного альбумина человека (HSA) также продемонстрировали посредством ELISA (фигура 46B). Результат показал сильное связывание с HSA слитой с ABD молекулы DART, в то время как не наблюдали связывания с контрольным DART hCD16-hCD32B.
Цитотоксичность ABD-DART in vitro: Чтобы продемонстрировать одновременное связывание этого биспецифического ABD-DART с двумя антигенами, одним на эффекторной клетке и другим на клетке-мишени, проводили анализ перенаправленного уничтожения клеток. С использованием PBMC человека в качестве эффекторных клеток ABD-DART hCD16-hCD32B индуцировало сильную дозозависимую цитотоксичность против CD32B-положительных B-клеточных линий Daudi (фигура 47). Результат продемонстрировал, что эффективность ABD-DART была эквивалентна эффективности исходного DART.
Фармакокинетические свойства ABD-DART: Фармакокинетические свойства ABD-DART hCD16-hCD32B анализировали посредством ELISA образцов сыворотки после внутривенной инъекции однократной дозы мышам C57B1/6 (фигура 48). Оба белка, DART и ABD-DART, продемонстрировали двухфазное выведение из кровотока. PK-исследование ABD-DART продемонстрировало более длительное время в кровотоке с увеличенным терминальным временем полужизни, составляющим 35,1 ч, по сравнению с 1,2 ч для обычного DART (фигура 48, таблица 17). Улучшение фармакокинетических свойств также было продемонстрировано посредством сравнения площади под кривой (AUC). Для конструкции ABD-DART AUC возрастала практически на коэффициент 30 после слияния с ABD (таблица 17).
В общем, был успешно разработан и продуцирован слитый с альбумин-связывающим доменом белок DART (обозначаемый как ABD-DART). Было выявлено, что ABD-DART hCD16-hCD32B сохраняет специфичность к двум его распознаваемым антигенным детерминантам: CD16 и CD32B. Было выявлено, что ABD-DART демонстрирует высокую аффинность в отношении сывороточного альбумина человека. Слияние с ABD не снижало биологическую активность (т.е. эффективность DART в отношении перенаправленного уничтожения опухолевых клеток). Слияние молекулы DART с ABD привело к значительному улучшению (увеличению) его времени полужизни in vivo и достигало этой цели без существенного увеличения размера. Способность сохранять небольшой размер является значительной и преимущественной, поскольку она способствует способности DART диффундировать в опухолевые ткани.
6.19 DART ДЛЯ Her2/B-КЛЕТОЧНОГО РЕЦЕПТОРА
Конструировали диантитело IgDART, которое содержало вариабельные области, способные связываться с Her2/neu и с T-клеточным рецептором ("TCR").
Как рассмотрено выше, TCR в природе экспрессируется CD4+ или CD8+ T-клетками и позволяет таким клеткам распознавать антигенные пептиды, которые связываются и презентируются белками MHC класса I или класса II антигенпредставляющих клеток. Распознавание комплекса pMHC (пептид-MHC) посредством TCR инициирует развитие клеточного иммунного ответа, который приводит к продукции цитокинов и лизису антигенпредставляющих клеток. HER2/neu, важный представитель семейства ErbB, широко исследован вследствие его роли в нескольких карциномах человека и в развитии млекопитающих. (Hynes and Stern (1994) Biochim. et Biophys. Acta 1198:165-184; и Dougall et al. (1994) Oncogene 9:2109-2123; Lee et al. (1995) Nature 378:394-398). Ген HER2/neu и белок HER2/neu человека описаны в Semba et al. (1985) Proc. Natl. Acad. Sci. (U.S.A.) 82:6497-6501 и Yamamoto et al. (1986) Nature 319:230-234, и последовательность доступна под регистрационным номером GenBank X03363. HER2/neu содержит четыре домена: внеклеточный домен, с которым связывается лиганд; липофильный трансмембранный домен; консервативный внутриклеточный тирозинкиназный домен; и С-концевой домен передачи сигнала, содержащий несколько остатков тирозина, которые могут быть фосфорилированными. (Plowman et al. (1993) Proc. Natl. Acad. Sci. (U.S.A.) 90:1746-1750). Последовательность внеклеточного домена (ECD) HER2/neu описана Franklin et al. (2004) Cancer Cell. 5(4):317-328, и доступна в реестре белков DataBank 1S78 (2004).
HER2/neu функционирует как рецептор фактора роста и часто экспрессируется опухолями, такими как рак молочной железы, рак толстого кишечника, рак мочевого пузыря, рак яичника и рак легкого. HER2/neu сверхэкспрессируется в 25-30% случаев рака молочной железы и рака яичника человека и ассоциирован с агрессивным клиническим прогрессированием и плохим прогнозом у этих пациентов. (Slamon et al. (1987) Science 235:177-182; Slamon et al. (1989) Science 244:707-712). Сверхэкспрессию HER2/neu также наблюдали при других карциномах, включая карциномы желудка, эндометрия, слюнной железы, легкого, почки, толстого кишечника, щитовидной железы, поджелудочной железы и молочного пузыря. (См., например, King et al. (1985) Science 229:974; McCann et al. (1990) Cancer 65:88-92; Yonemura et al. (1991) Cancer Research 51:1034).
Разработан ряд моноклональных антител и низкомолекулярных ингибиторов тирозинкиназ, нацеленных на HER-1 или HER2/neu, включая, в частности, гуманизированный вариант моноклонального антитела мыши, известный как 4D5 (HERCEPTIN®, Genentech, Inc.), который распознает внеклеточный эпитоп (аминокислоты с 529 по 627) в цистеин-богатом домене II HER2/neu, который расположен очень близко к трансмембранной области белка. В исследованиях было показано, что в клетках рака молочной железы, сверхэкспрессирующих HER2/neu, введение антител, специфичных к HER2/neu, в комбинации с химиотерапевтическими средствами (например, цисплатином, доксорубицином, таксолом) вызывает более высокий цитотоксический ответ, чем лечение химиотерапией отдельно. (Hancock et al. (1991) Cancer Res. 51:4575-4580; Arteaga et al. (1994) Cancer 54:3758-3765; Pietras et al. (1994) Oncogene 9:1829-1838). Полагают, что одним из возможных механизмов, посредством которого антитела против HER2/neu могут усиливать ответ на химиотерапевтические средства, является модулирование экспрессии белка HER2/neu или препятствование репарации ДНК. (Stancovski et al. (1991) Proc. Natl. Acad. Sci. (U.S.A.) 88:8691-8695; Bacus et al. (1992) Cell Growth & Diff. 3:401-411; Bacus et al. (1993) Cancer Res. 53:5251-5261; Klapper et al. (1997) Oncogene 14:2099-2109; Klapper et al. (2000) Cancer Res. 60:3384-3388; Arteaga et al. (2001) J Clinical Oncology 19(18s):32s-40s. Хотя в некоторых случаях антитела против HER2/neu, такие как HERCEPTIN®, обеспечивают терапевтическую пользу у пациентов, большинство пациентов с раком молочной железы и других пациентов обладают рефрактерными ответами на такие антитела. Эти ответы отражают, частично, различия в степени сверхэкспрессии HER2/neu злокачественными клетками пациента.
В качестве следствия наличия вариабельных областей, способных связываться с Her2/neu и с T-клеточным рецептором ("TCR"), DART способен связываться с экспрессирующими HER2 клетками и, тем самым, присоединять к таким клеткам домен, способный связываться с T-клеточным рецептором. Когда такие T-клетки связываются с этим доменом, они активируются, инициируя иммунный ответ, который приводит к уничтожению клеток, экспрессирующих HER2.
Аминокислотные последовательности и последовательности нуклеиновых кислот для такого DART представлены ниже, причем последовательности VL и VH показаны обычным шрифтом, линкер VL-VH показан подчеркнутым шрифтом, и последовательность, кодирующая C-концевой мотив гетеродимеризации (SEQ ID NO: 313: GFNRGEC или SEQ ID NO: 314: GVEPKSC), показана полужирным шрифтом и курсивом.
Аминокислотная последовательность TCRVL-HER2VH (SEQ ID NO: 315)

Кодирующая TCRVL-HER2VH последовательность нуклеиновой кислоты (SEQ ID NO: 316)

Аминокислотная последовательность HER2VL-TCRVH (SEQ ID NO: 317)

Кодирующая HER2VL-TCRVH последовательность нуклеиновой кислоты (SEQ ID NO: 318)

В предпочтительном варианте осуществления такие конструкции модифицируют так, чтобы они содержали домен с E-спиралью или K-спиралью, который способствует образованию гетеродимеров (т.е. димеров TCRVL-HER2VH×HER2VL-TCRVH). Последовательности аминокислот и последовательности нуклеиновых кислот для такого DART представлены ниже, причем последовательности VL и VH показаны обычным шрифтом, линкер VL-VH показан подчеркнутым шрифтом, и последовательность, кодирующая Cys-содержащий линкер для димеризации (GGCGGG; остатки 2-7 SEQ ID NO: 267), показана курсивом. Домен гетеродимеризации с E-спиралью или K-спиралью подчеркнут двойным подчеркиванием (предпочтительная последовательность "E-спирали" представляет собой 4 гептамерных повтора EVAALEK; SEQ ID NO: 299; предпочтительная последовательность "K-спирали" представляет собой 4 гептамерных повтора r KVAALKE (SEQ ID NO: 300). Последовательность после E-спирали или K-спирали не имеет приписываемой ей функции.
Аминокислотная последовательность TCRVL-HER2VH-E-спирали (SEQ ID NO: 319)

Кодирующая TCRVL-HER2VH-E-спираль последовательность нуклеиновой кислоты (SEQ ID NO: 320)

Аминокислотная последовательность HER2VL-TCRVH-K-спираль (SEQ ID NO: 321)

Кодирующая HER2VL-TCRVH-K-спираль последовательность нуклеиновой кислоты (SEQ ID NO: 322)


Молекулы DART, имеющие домены, связывающие Her2 и T-клеточный рецептор (TCR), тестировали в отношении их способности опосредовать цитотоксичность во множестве клеточных линий рака молочной железы, рака толстого кишечника и рака мочевого пузыря, которые ранее были охарактеризованы как проявляющие низкие уровни экспрессии HER2 (и таким образом, являющиеся рефрактерными к лечению антителом против Her2/neu, Herceptin®). Протестированные клеточные линии рака молочной железы представляют собой ZR75-1 (HER2 2+) (ФИГ. 49A), MCF-7 (HER2 1+) (ФИГ. 49B) и MDA-MB468 (HER2-ve) (ФИГ. 49C). Протестированные клеточные линии рака не молочной железы представляют собой HT-29 (клеточная линия рака толстого кишечника) (ФИГ. 49D) и SW780 (клеточная линия рака мочевого пузыря) (ФИГ. 49E). Как показано на ФИГ.49A-E, такие молекулы DART были по существу более эффективными, чем HERCEPTIN®, в опосредовании цитотоксичности происходящих из опухоли клеточных линий, как с точки зрения концентраций, требуемых для достижения эквивалентной цитотоксичности, так и с точки зрения наблюдаемых максимальных уровней цитотоксичности.
Можно проводить множество модификаций и изменений этого изобретения без отклонения от его сущности и объема, как будет очевидно специалистам в данной области. Конкретные варианты осуществления, описанные в настоящем документе, предоставлены только в качестве примера, и изобретение ограничивается только прилагаемой формулой изобретения, наряду с полным объемом эквивалентов, на которые эта формулы изобретения предоставляет право. Подразумевают, что такие модификации находятся в объеме прилагаемой формулы изобретения. Все ссылки, патентные и непатентные, цитированные в настоящем документе, включены в настоящий документ в качестве ссылок в полном объеме и для всех целей в той же степени, как если бы каждая отдельная публикация или патент или патентная заявка были конкретно и индивидуально указаны, как включенные в качестве ссылок в полном объеме для любых целей.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MacroGenics, Inc.
Johnson, Leslie S.
Huang, Ling S.
<120> КОВАЛЕНТНЫЕ ДИАНТИТЕЛА И ИХ ПРИМЕНЕНИЕ
<130> 1301.0007I4
<150> US 60/671,657
<151> 2005-04-15
<150> US 11/409,339
<150> PCT/US06/014481
<151> 2006-04-17
<150> US 60/945,523
<151> 2007-06-21
<150> US 61/019,051
<151> 2008-01-04
<150> US 12/138,867
<151> 2008-06-13
<150> PCT/US08/66957
<151> 2008-06-13
<150> US 61/139,352
<151> 2008-12-19
<150> US 61/156,035
<151> 2009-02-27
<150> US 61/256,779
<151> 2009-10-30
<160> 322
<170> PatentIn version 3.5
<210> 1
<211> 13
<212> Белок
<213> homo sapiens
<400> 1
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
1 5 10
<210> 2
<211> 10
<212> Белок
<213> homo sapiens
<400> 2
Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
1 5 10
<210> 3
<211> 60
<212> Белок
<213> homo sapiens
<400> 3
Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro Arg Cys
1 5 10 15
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
20 25 30
Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro Glu
35 40 45
Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
50 55 60
<210> 4
<211> 10
<212> Белок
<213> homo sapiens
<400> 4
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
1 5 10
<210> 5
<211> 217
<212> Белок
<213> homo sapiens
<400> 5
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 6
<211> 216
<212> Белок
<213> homo sapiens
<400> 6
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
1 5 10 15
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
20 25 30
Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
35 40 45
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
50 55 60
Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln
65 70 75 80
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
85 90 95
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro
100 105 110
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
115 120 125
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
130 135 140
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
145 150 155 160
Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
165 170 175
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
180 185 190
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
195 200 205
Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 7
<211> 217
<212> Белок
<213> homo sapiens
<400> 7
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
35 40 45
Val Asp Gly Val Gln Val His Asn Ala Lys Thr Lys Pro Arg Glu Gln
50 55 60
Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asn Trp Leu Asp Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 8
<211> 217
<212> Белок
<213> homo sapiens
<400> 8
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 9
<211> 237
<212> Белок
<213> homo sapiens
<400> 9
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Phe Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Glu Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys
115 120 125
Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu
130 135 140
Ser Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys
145 150 155 160
Ala Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr
165 170 175
Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys
180 185 190
Asn Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala
195 200 205
Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly
210 215 220
Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly Cys
225 230 235
<210> 10
<211> 8
<212> Белок
<213> homo sapiens
<400> 10
Gly Gly Gly Ser Gly Gly Gly Gly
1 5
<210> 11
<211> 244
<212> Белок
<213> homo sapiens
<400> 11
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala
115 120 125
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
130 135 140
Gly Tyr Thr Phe Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Gln Gly Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr Tyr
165 170 175
Pro Asn Tyr Asn Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val Val
180 185 190
Val Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp
195 200 205
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr Tyr
210 215 220
Ser Gly Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Leu Gly Gly Cys
<210> 12
<211> 240
<212> Белок
<213> homo sapiens
<400> 12
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Phe Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Glu Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr Tyr Pro Asn Tyr Asn
165 170 175
Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val Val Val Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr Tyr Ser Gly Met Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Leu Gly Gly Cys
225 230 235 240
<210> 13
<211> 241
<212> Белок
<213> homo sapiens
<400> 13
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Gln Val Thr Leu Arg Glu Ser Gly Pro
115 120 125
Ala Leu Val Lys Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Phe Ser
130 135 140
Gly Phe Ser Leu Ser Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln
145 150 155 160
Pro Pro Gly Lys Ala Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp
165 170 175
Asp Lys Arg Tyr Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys
180 185 190
Asp Thr Ser Lys Asn Gln Val Val Leu Thr Met Thr Asn Met Asp Pro
195 200 205
Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe
210 215 220
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly
225 230 235 240
Cys
<210> 14
<211> 470
<212> Белок
<213> homo sapiens
<400> 14
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Phe Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Glu Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys
115 120 125
Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu
130 135 140
Ser Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys
145 150 155 160
Ala Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr
165 170 175
Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys
180 185 190
Asn Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala
195 200 205
Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly
210 215 220
Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly Cys Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 15
<211> 477
<212> Белок
<213> homo sapiens
<400> 15
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala
115 120 125
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
130 135 140
Gly Tyr Thr Phe Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Gln Gly Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr Tyr
165 170 175
Pro Asn Tyr Asn Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val Val
180 185 190
Val Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp
195 200 205
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr Tyr
210 215 220
Ser Gly Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Leu Gly Gly Cys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
245 250 255
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
260 265 270
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
275 280 285
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
290 295 300
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
305 310 315 320
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
325 330 335
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
340 345 350
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
355 360 365
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
370 375 380
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
385 390 395 400
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
405 410 415
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
420 425 430
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
435 440 445
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
450 455 460
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470 475
<210> 16
<211> 243
<212> Белок
<213> homo sapiens
<400> 16
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Phe Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Glu Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys
115 120 125
Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu
130 135 140
Ser Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys
145 150 155 160
Ala Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr
165 170 175
Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys
180 185 190
Asn Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala
195 200 205
Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly
210 215 220
Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly Cys Phe Asn Arg
225 230 235 240
Gly Glu Cys
<210> 17
<211> 10
<212> Белок
<213> искусственная
<220>
<223> линкер плюс C-конец каппа цепи IgG человека
<400> 17
Leu Gly Gly Cys Phe Asn Arg Gly Glu Cys
1 5 10
<210> 18
<211> 477
<212> Белок
<213> homo sapiens
<400> 18
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala
115 120 125
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
130 135 140
Gly Tyr Thr Phe Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Gln Gly Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr Tyr
165 170 175
Pro Asn Tyr Asn Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val Val
180 185 190
Val Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp
195 200 205
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr Tyr
210 215 220
Ser Gly Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Leu Gly Gly Cys Ala Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
245 250 255
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
260 265 270
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
275 280 285
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
290 295 300
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
305 310 315 320
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
325 330 335
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
340 345 350
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
355 360 365
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
370 375 380
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
385 390 395 400
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
405 410 415
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
420 425 430
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
435 440 445
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
450 455 460
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470 475
<210> 19
<211> 485
<212> Белок
<213> homo sapiens
<400> 19
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Ser Gly Gly Gly
225 230 235 240
Gly Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu
245 250 255
Gly Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe
260 265 270
Asp Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro
275 280 285
Pro Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro
290 295 300
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
305 310 315 320
Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser
325 330 335
Asn Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
340 345 350
Gly Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly
355 360 365
Ala Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala
370 375 380
Ser Gly Tyr Thr Phe Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala
385 390 395 400
Pro Gly Gln Gly Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr
405 410 415
Tyr Pro Asn Tyr Asn Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val
420 425 430
Val Val Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser
435 440 445
Asp Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr
450 455 460
Tyr Ser Gly Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
465 470 475 480
Ser Leu Gly Gly Cys
485
<210> 20
<211> 469
<212> Белок
<213> homo sapiens
<400> 20
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Ser Gly Gly Gly
210 215 220
Gly Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu
225 230 235 240
Gly Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe
245 250 255
Asp Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro
260 265 270
Pro Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro
275 280 285
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
290 295 300
Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser
305 310 315 320
Asn Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
325 330 335
Gly Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly
340 345 350
Ala Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala
355 360 365
Ser Gly Tyr Thr Phe Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala
370 375 380
Pro Gly Gln Gly Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr
385 390 395 400
Tyr Pro Asn Tyr Asn Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val
405 410 415
Val Val Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser
420 425 430
Asp Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr
435 440 445
Tyr Ser Gly Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
450 455 460
Ser Leu Gly Gly Cys
465
<210> 21
<211> 466
<212> Белок
<213> homo sapiens
<400> 21
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Phe Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Glu Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys
115 120 125
Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu
130 135 140
Ser Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys
145 150 155 160
Ala Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr
165 170 175
Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys
180 185 190
Asn Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala
195 200 205
Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly
210 215 220
Gln Gly Thr Leu Val Thr Val Ser Ser Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 22
<211> 246
<212> Белок
<213> homo sapiens
<400> 22
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala
115 120 125
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
130 135 140
Gly Tyr Thr Phe Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Gln Cys Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr Tyr
165 170 175
Pro Asn Tyr Asn Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val Val
180 185 190
Val Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp
195 200 205
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr Tyr
210 215 220
Ser Gly Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Phe Asn Arg Gly Glu Cys
245
<210> 23
<211> 6
<212> Белок
<213> homo sapiens
<400> 23
Phe Asn Arg Gly Glu Cys
1 5
<210> 24
<211> 5
<212> Белок
<213> искусственная
<220>
<223> CDR1 вариабельной области тяжелой цепи 2B6
<400> 24
Asn Tyr Trp Ile His
1 5
<210> 25
<211> 5
<212> Белок
<213> искусственная
<220>
<223> CDR1 вариабельной области тяжелой цепи 3H7
<400> 25
Asp Ala Trp Met Asp
1 5
<210> 26
<211> 17
<212> Белок
<213> искусственная
<220>
<223> CDR2 вариабельной области тяжелой цепи 2B6
<400> 26
Val Ile Asp Pro Ser Asp Thr Tyr Pro Asn Tyr Asn Lys Lys Phe Lys
1 5 10 15
Gly
<210> 27
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR2 вариабельной области тяжелой цепи 3H7
<400> 27
Glu Ile Arg Asn Lys Ala Asn Asn Leu Ala Thr Tyr Tyr Ala Glu Ser
1 5 10 15
Val Lys Gly
<210> 28
<211> 12
<212> Белок
<213> искусственная
<220>
<223> CDR3 вариабельной области тяжелой цепи 2B6
<400> 28
Asn Gly Asp Ser Asp Tyr Tyr Ser Gly Met Asp Tyr
1 5 10
<210> 29
<211> 6
<212> Белок
<213> искусственная
<220>
<223> CDR3 вариабельной области тяжелой цепи 3H7
<400> 29
Tyr Ser Pro Phe Ala Tyr
1 5
<210> 30
<211> 11
<212> Белок
<213> искусственная
<220>
<223> CDR1 вариабельной области легкой цепи 2B6
<400> 30
Arg Thr Ser Gln Ser Ile Gly Thr Asn Ile His
1 5 10
<210> 31
<211> 11
<212> Белок
<213> искусственная
<220>
<223> CDR1 вариабельной области легкой цепи 3H7
<400> 31
Arg Ala Ser Gln Glu Ile Ser Gly Tyr Leu Ser
1 5 10
<210> 32
<211> 7
<212> Белок
<213> искусственная
<220>
<223> CDR2 вариабельной области легкой цепи 2B6
<400> 32
Asn Val Ser Glu Ser Ile Ser
1 5
<210> 33
<211> 7
<212> Белок
<213> искусственная
<220>
<223> CDR2 вариабельной области легкой цепи 2B6
<400> 33
Tyr Val Ser Glu Ser Ile Ser
1 5
<210> 34
<211> 7
<212> Белок
<213> искусственная
<220>
<223> CDR2 вариабельной области легкой цепи 2B6
<400> 34
Tyr Ala Ser Glu Ser Ile Ser
1 5
<210> 35
<211> 7
<212> Белок
<213> искусственная
<220>
<223> CDR2 вариабельной области легкой цепи 3H7
<400> 35
Ala Ala Ser Thr Leu Asp Ser
1 5
<210> 36
<211> 9
<212> Белок
<213> искусственная
<220>
<223> CDR3 вариабельной области легкой цепи 2B6
<400> 36
Gln Gln Ser Asn Thr Trp Pro Phe Thr
1 5
<210> 37
<211> 9
<212> Белок
<213> искусственная
<220>
<223> CDR3 вариабельной области легкой цепи 3H7
<400> 37
Leu Gln Tyr Val Ser Tyr Pro Tyr Thr
1 5
<210> 38
<211> 121
<212> Белок
<213> искусственная
<220>
<223> вариабельная область тяжелой цепи 2B6
<400> 38
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Asp Pro Ser Asp Thr Tyr Pro Asn Tyr Asn Lys Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Gly Asp Ser Asp Tyr Tyr Ser Gly Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 39
<211> 107
<212> Белок
<213> искусственная
<220>
<223> вариабельная область легкой цепи 2B6 Hu2B6VL-1
<400> 39
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Asn Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 40
<211> 107
<212> Белок
<213> искусственная
<220>
<223> гуманизированная вариабельная область тяжелой цепи 2B6 Hu2B6VL-2
<400> 40
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 41
<211> 107
<212> Белок
<213> искусственная
<220>
<223> гуманизированная вариабельная область легкой цепи 2B6 Hu2B6VL-3
<400> 41
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 42
<211> 107
<212> Белок
<213> Mus sp.
<400> 42
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Asp Gly Thr Ile Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Trp Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Ala Asp Tyr Tyr Cys Leu Gln Tyr Val Ser Tyr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 43
<211> 9
<212> Белок
<213> искусственная
<220>
<223> участок связывания IgG в рецепторе FcgammaRIIB
<400> 43
Lys Lys Phe Ser Arg Ser Asp Pro Asn
1 5
<210> 44
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 44
Gln Lys Phe Ser Arg Leu Asp Pro Asn
1 5
<210> 45
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 45
Gln Lys Phe Ser Arg Leu Asp Pro Thr
1 5
<210> 46
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 46
Lys Lys Phe Ser Arg Leu Asp Pro Thr
1 5
<210> 47
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 47
Gln Lys Phe Ser His Leu Asp Pro Thr
1 5
<210> 48
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 48
Lys Lys Phe Ser His Leu Asp Pro Thr
1 5
<210> 49
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 49
Gln Lys Phe Ser Arg Leu Asp Pro Asn
1 5
<210> 50
<211> 9
<212> Белок
<213> искусственная
<220>
<223> IgG binding site for FcgammaRIIB receptor
<400> 50
Lys Lys Phe Ser Arg Ser Asp Pro Asn
1 5
<210> 51
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 51
Gln Lys Phe Ser Arg Leu Asp Pro Thr
1 5
<210> 52
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 52
Gln Lys Phe Ser His Leu Asp Pro Thr
1 5
<210> 53
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 53
Lys Lys Phe Ser Arg Leu Asp Pro Thr
1 5
<210> 54
<211> 9
<212> Белок
<213> искусственная
<220>
<223> Участок связывания IgG в рецепторе FcgammaRIIB
<400> 54
Lys Lys Phe Ser His Leu Asp Pro Thr
1 5
<210> 55
<211> 36
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh321F
<400> 55
cgagctagct ctagatgaga tcacagttct ctctac 36
<210> 56
<211> 47
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh318R
<400> 56
gcctccgcct ccggatccgc ctcctttgat ctccaccttg gtccctc 47
<210> 57
<211> 47
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh319F
<400> 57
ggaggcggat ccggaggcgg aggccaggtt cagctggtgc agtctgg 47
<210> 58
<211> 43
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh320R
<400> 58
tttgaattct agcagcctcc cagtgaggag acggtgaccg tgg 43
<210> 59
<211> 46
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh315R
<400> 59
gcctccgcct ccggatccgc ctcctttgat ctcaagcttg gtcccc 46
<210> 60
<211> 48
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh316F
<400> 60
ggaggcggat ccggaggcgg aggccaggtt accctgagag agtctggc 48
<210> 61
<211> 47
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh317R
<400> 61
tttgaattcc tagcagcctc ccagtgagct cacagtgacc agagtcc 47
<210> 62
<211> 27
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh339R
<400> 62
ctcaacgcag cctcccagtg agctcac 27
<210> 63
<211> 31
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh340R
<400> 63
aacgcagcct cccagtgagg agacggtgac c 31
<210> 64
<211> 27
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh366R
<400> 64
tttgaattct atttacccgg agacagg 27
<210> 65
<211> 33
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lhg367F
<400> 65
ctgggaggct gcgcagagcc caaatcttgt gac 33
<210> 66
<211> 33
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh368R
<400> 66
gtcacaagat ttgggctctg cgcagcctcc cag 33
<210> 67
<211> 50
<212> ДНК
<213> искусственная
<220>
<223> Праймер Lgh369R
<400> 67
tttgaattct aacactctcc cctgttgaag cagcctccca gtgaggagac 50
<210> 68
<211> 118
<212> Белок
<213> искусственная
<220>
<223> вариабельная область тяжелой цепи гуманизированного антитела против
CD16A Hu3G8VH-1
<400> 68
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 69
<211> 118
<212> Белок
<213> искусственная
<220>
<223> вариабельная область тяжелой цепи гуманизированного антитела против
CD16A Hu3G8VH-5
<400> 69
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 70
<211> 117
<212> Белок
<213> искусственная
<220>
<223> вариабельная область тяжелой цепи гуманизированного антитела против
CD16A Hu3G8VH-22
<400> 70
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Val Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Ser Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Ile Asn Pro Ala Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser
115
<210> 71
<211> 111
<212> Белок
<213> искусственная
<220>
<223> вариабельная область легкой цепи гуманизированного антитела против
CD16A Hu3G8VL-1
<400> 71
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 72
<211> 111
<212> Белок
<213> искусственная
<220>
<223> вариабельная область легкой цепи гуманизированного антитела против
CD16A Hu3G8VL-43
<400> 72
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Ser Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Ser Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 73
<211> 111
<212> Белок
<213> искусственная
<220>
<223> вариабельная область легкой цепи гуманизированного антитела против
CD16A Hu3G8VL-22
<400> 73
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Thr Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Ser Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 74
<211> 24
<212> ДНК
<213> Последовательность ДНК SEQ ID NO:10
<400> 74
ggaggcggat ccggaggcgg aggc 24
<210> 75
<211> 18
<212> ДНК
<213> homo sapiens
<400> 75
ttcaacaggg gagagtgt 18
<210> 76
<211> 30
<212> ДНК
<213> искусственная
<220>
<223> Линкер плюс C-конец каппа цепи IgG человека
<400> 76
ctgggaggct gcttcaacag gggagagtgt 30
<210> 77
<211> 6
<212> Белок
<213> homo sapiens
<400> 77
Val Glu Pro Lys Ser Cys
1 5
<210> 78
<211> 18
<212> ДНК
<213> homo sapiens
<400> 78
gttgagccca aatcttgt 18
<210> 79
<211> 118
<212> Белок
<213> homo sapiens
<400> 79
Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Arg Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser Asn Gln Val
65 70 75 80
Phe Leu Lys Ile Ala Ser Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 80
<211> 354
<212> ДНК
<213> homo sapiens
<400> 80
caggttaccc tgaaagagtc tggccctggg atattgcagc cctcccagac cctcagtctg 60
acttgttctt tctctgggtt ttcactgagg acttctggta tgggtgtagg ctggattcgt 120
cagccttcag ggaagggtct agagtggctg gcacacattt ggtgggatga tgacaagcgc 180
tataatccag ccctgaagag ccgactgaca atctccaagg atacctccag caaccaggta 240
ttcctcaaaa tcgccagtgt ggacactgca gatactgcca catactactg tgctcaaata 300
aaccccgcct ggtttgctta ctggggccaa gggactctgg tcactgtgag ctca 354
<210> 81
<211> 354
<212> ДНК
<213> homo sapiens
<400> 81
caggttaccc tgagagagtc tggccctgcg ctggtgaagc ccacacagac cctcacactg 60
acttgtacct tctctgggtt ttcactgagc acttctggta tgggtgtagg ctggattcgt 120
cagcctcccg ggaaggctct agagtggctg gcacacattt ggtgggatga tgacaagcgc 180
tataatccag ccctgaagag ccgactgaca atctccaagg atacctccaa aaaccaggta 240
gtcctcacaa tgaccaacat ggaccctgtg gatactgcca catactactg tgctcaaata 300
aaccccgcct ggtttgctta ctggggccaa gggactctgg tcactgtgag ctca 354
<210> 82
<211> 111
<212> Белок
<213> homo sapiens
<400> 82
Asp Thr Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Ala Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Thr Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 83
<211> 333
<212> ДНК
<213> homo sapiens
<400> 83
gacactgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atctcctgca aggccagcca aagtgttgat tttgatggtg atagttttat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctata ctacatccaa tctagaatct 180
gggatcccag ccaggtttag tgccagtggg tctgggacag acttcaccct caacatccat 240
cctgtggagg aggaggatac tgcaacctat tactgtcagc aaagtaatga ggatccgtac 300
acgttcggag gggggaccaa gctggaaata aaa 333
<210> 84
<211> 332
<212> ДНК
<213> искусственная
<220>
<223> Вариабельная область легкой цепи гуманизированного антитела против
CD16A Hu3G8VL-1
<400> 84
gacatcgtga tgacccaatc tccagactct ttggctgtgt ctctagggga gagggccacc 60
atcaactgca aggccagcca aagtgttgat tttgatggtg atagttttat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctata ctacatccaa tctagaatct 180
ggggtcccag acaggtttag tggcagtggg tctgggacag acttcaccct caccatcagc 240
agcctgcagg ctgaggatgt ggcagtttat tactgtcagc aaagtaatga ggatccgtac 300
acgttcggac aggggaccaa gcttgagatc aa 332
<210> 85
<211> 121
<212> Белок
<213> искусственная
<220>
<223> Гуманизированная вариабельная область тяжелой цепи 2B6
<400> 85
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Asp Pro Ser Asp Thr Tyr Pro Asn Tyr Asn Lys Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Val Val Val Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Gly Asp Ser Asp Tyr Tyr Ser Gly Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 86
<211> 363
<212> ДНК
<213> искусственная
<220>
<223> Гуманизированная вариабельная область тяжелой цепи 2B6
<400> 86
caggttcagc tggtgcagtc tggagctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg cttctggtta cacctttacc aactactgga tacactgggt gcgacaggcc 120
cctggacaag ggcttgagtg gattggagtg attgatcctt ctgatactta tccaaattac 180
aataaaaagt tcaagggcag agtcaccatg accgtagtcg tatccacgag cacagcctac 240
atggagctga ggagcctgag atctgacgac acggccgtgt attactgtgc gagaaacggt 300
gattccgatt attactctgg tatggactac tgggggcaag ggaccacggt caccgtctcc 360
tca 363
<210> 87
<211> 111
<212> Белок
<213> искусственная
<220>
<223> Гуманизированная вариабельная область легкой цепи 2B6
<400> 87
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 88
<211> 321
<212> ДНК
<213> искусственная
<220>
<223> Гуманизированная вариабельная область легкой цепи 2B6
<400> 88
gaaattgtgc tgactcagtc tccagacttt cagtctgtga ctccaaagga gaaagtcacc 60
ttcacctgca ggaccagtca gagcattggc acaaacatac actggtacca gcagaaacca 120
gatcagtctc caaagctcct catcaaggag gtttctgagt ctatctctgg agtcccatcg 180
aggttcagtg gcagtggatc tgggacagat ttcaccctca ccatcaatag cctggaagct 240
gaagatgctg caacgtatta ctgtcaacaa agtaatacct ggccgttcac gttcggcgga 300
gggaccaagg tggagatcaa a 321
<210> 89
<211> 6
<212> Белок
<213> искусственная
<220>
<223> участок распознавания для расщепления тромбином
<400> 89
Leu Val Pro Arg Gly Ser
1 5
<210> 90
<211> 4
<212> Белок
<213> искусственная
<220>
<223> участок распознавания для расщепления фактором Xa
<220>
<221> VARIANT
<222> (2)..(2)
<223> Xaa=Glu или Asp
<400> 90
Ile Xaa Gly Arg
1
<210> 91
<211> 5
<212> Белок
<213> искусственная
<220>
<223> участок распознавания для расщепления энтерокиназой
<400> 91
Asp Asp Asp Asp Lys
1 5
<210> 92
<211> 4
<212> Белок
<213> искусственная
<220>
<223> участок распознавания для расщепления фурином
<220>
<221> VARIANT
<222> (2)..(3)
<223> Xaa=любая аминокислота
<400> 92
Arg Xaa Xaa Arg
1
<210> 93
<211> 4
<212> Белок
<213> искусственная
<220>
<223> предпочтительный участок распознавания для расщепления фурином
<220>
<221> VARIANT
<222> (2)..(2)
<223> Xaa=любая аминокислота
<220>
<221> VARIANT
<222> (3)..(3)
<223> Xaa=Lys или Arg
<400> 93
Arg Xaa Xaa Arg
1
<210> 94
<211> 7
<212> Белок
<213> искусственная
<220>
<223> участок распознавания для расщепления AcTEV
<400> 94
Gln Asn Leu Tyr Phe Gln Gly
1 5
<210> 95
<211> 487
<212> Белок
<213> искусственная
<220>
<223> полибелковый предшественник ковалентного диантитела
<400> 95
Asp Thr Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Ala Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Thr Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Glu Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro
145 150 155 160
Thr Thr Gly Leu Glu Trp Val Ala Ser Ile Ser Tyr Asp Gly Gly Asp
165 170 175
Thr His Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
180 185 190
Asn Ala Lys Ser Ser Leu Tyr Leu Gln Met Asp Ser Leu Arg Ser Glu
195 200 205
Asp Thr Ala Thr Tyr Tyr Cys Ala Thr Glu Thr Thr Gly Ile Pro Thr
210 215 220
Gly Val Met Asp Ala Trp Gly Gln Gly Val Ser Val Thr Val Ser Ser
225 230 235 240
Leu Gly Gly Cys Gly Gly Arg Ala Lys Arg Asp Val Gln Met Thr Gln
245 250 255
Ser Pro Ser Asn Leu Ala Ala Ser Pro Gly Glu Ser Val Ser Ile Asn
260 265 270
Cys Lys Ala Ser Glu Ser Ile Ser Lys Tyr Leu Ala Trp Tyr Leu Gln
275 280 285
Lys Pro Gly Lys Ala Asn Lys Leu Leu Met Tyr Asp Gly Ser Thr Leu
290 295 300
Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
305 310 315 320
Phe Thr Leu Thr Ile Arg Ser Leu Glu Pro Glu Asp Phe Gly Leu Tyr
325 330 335
Tyr Cys Gln Gln His Tyr Glu Tyr Pro Ala Thr Phe Gly Ser Gly Thr
340 345 350
Lys Leu Glu Ile Lys Gly Gly Gly Ser Gly Gly Gly Gly Gln Val Thr
355 360 365
Leu Lys Glu Ser Gly Pro Gly Ile Leu Gln Pro Ser Gln Thr Leu Ser
370 375 380
Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Arg Thr Ser Gly Met Gly
385 390 395 400
Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu Trp Leu Ala
405 410 415
His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala Leu Lys Ser
420 425 430
Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser Asn Gln Val Phe Leu Lys
435 440 445
Ile Ala Ser Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
450 455 460
Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
465 470 475 480
Val Ser Ser Leu Gly Gly Cys
485
<210> 96
<211> 1464
<212> ДНК
<213> искусственная
<220>
<223> Последовательность ДНК SEQ ID NO:97 (полибелковый предшественник
ковалентного диантитела)
<400> 96
gacactgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atctcctgca aggccagcca aagtgttgat tttgatggtg atagttttat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctata ctacatccaa tctagaatct 180
gggatcccag ccaggtttag tgccagtggg tctgggacag acttcaccct caacatccat 240
cctgtggagg aggaggatac tgcaacctat tactgtcagc aaagtaatga ggatccgtac 300
acgttcggag gggggaccaa gctggaaata aaaggaggcg gatccggagg cggaggcgag 360
gtggagctag tggagtctgg gggaggctta gtgcagcctg gaaggtccct gaaactctcg 420
tgtgcagcct caggattcac tttcagtgac tattacatgg cctgggtccg gcaggctcca 480
acgacgggtc tggagtgggt cgcatccatt agttatgatg gtggtgacac tcactatcga 540
gactccgtga agggccgatt tactatttcc agagataatg caaaaagcag cctatacctg 600
caaatggaca gtctgaggtc tgaggacacg gccacttatt actgtgcaac agagactacg 660
ggaataccta caggtgttat ggatgcctgg ggtcaaggag tttcagtcac tgtctcctca 720
ctgggaggct gcggcgggag agctaagagg gatgtccaga tgacccagtc tccatctaat 780
cttgctgcct ctcctggaga aagtgtttcc atcaattgca aggcaagtga gagcattagc 840
aagtatttag cctggtatct acagaaacct gggaaagcaa ataagcttct tatgtacgat 900
gggtcaactt tgcaatctgg aattccatcg aggttcagtg gcagtggatc tggtacagat 960
ttcactctca ccatcagaag cctggagcct gaagattttg gactctatta ctgtcaacag 1020
cattatgaat atccagccac gttcggttct gggaccaagc tggagatcaa aggaggcgga 1080
tccggaggcg gaggccaggt taccctgaaa gagtctggcc ctgggatatt gcagccctcc 1140
cagaccctca gtctgacttg ttctttctct gggttttcac tgaggacttc tggtatgggt 1200
gtaggctgga ttcgtcagcc ttcagggaag ggtctagagt ggctggcaca catttggtgg 1260
gatgatgaca agcgctataa tccagccctg aagagccgac tgacaatctc caaggatacc 1320
tccagcaacc aggtattcct caaaatcgcc agtgtggaca ctgcagatac tgccacatac 1380
tactgtgctc aaataaaccc cgcctggttt gcttactggg gccaagggac tctggtcact 1440
gtgagctcac tgggaggctg ctag 1464
<210> 97
<211> 511
<212> Белок
<213> искусственная
<220>
<223> полибелковый предшественник ковалентного диантитела
<400> 97
Asp Thr Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Ala Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Thr Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Glu Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro
145 150 155 160
Thr Thr Gly Leu Glu Trp Val Ala Ser Ile Ser Tyr Asp Gly Gly Asp
165 170 175
Thr His Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
180 185 190
Asn Ala Lys Ser Ser Leu Tyr Leu Gln Met Asp Ser Leu Arg Ser Glu
195 200 205
Asp Thr Ala Thr Tyr Tyr Cys Ala Thr Glu Thr Thr Gly Ile Pro Thr
210 215 220
Gly Val Met Asp Ala Trp Gly Gln Gly Val Ser Val Thr Val Ser Ser
225 230 235 240
Leu Gly Gly Cys Gly Gly Arg Ala Lys Arg Ala Pro Val Lys Gln Thr
245 250 255
Leu Asn Phe Asp Leu Leu Lys Leu Ala Gly Asp Val Glu Ser Asn Pro
260 265 270
Gly Pro Asp Val Gln Met Thr Gln Ser Pro Ser Asn Leu Ala Ala Ser
275 280 285
Pro Gly Glu Ser Val Ser Ile Asn Cys Lys Ala Ser Glu Ser Ile Ser
290 295 300
Lys Tyr Leu Ala Trp Tyr Leu Gln Lys Pro Gly Lys Ala Asn Lys Leu
305 310 315 320
Leu Met Tyr Asp Gly Ser Thr Leu Gln Ser Gly Ile Pro Ser Arg Phe
325 330 335
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg Ser Leu
340 345 350
Glu Pro Glu Asp Phe Gly Leu Tyr Tyr Cys Gln Gln His Tyr Glu Tyr
355 360 365
Pro Ala Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly
370 375 380
Ser Gly Gly Gly Gly Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile
385 390 395 400
Leu Gln Pro Ser Gln Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe
405 410 415
Ser Leu Arg Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser
420 425 430
Gly Lys Gly Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys
435 440 445
Arg Tyr Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr
450 455 460
Ser Ser Asn Gln Val Phe Leu Lys Ile Ala Ser Val Asp Thr Ala Asp
465 470 475 480
Thr Ala Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr
485 490 495
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly Cys
500 505 510
<210> 98
<211> 1536
<212> ДНК
<213> искусственная
<220>
<223> Последовательность ДНК SEQ ID NO:99 (полибелковый предшественник
ковалентного диантитела)
<400> 98
gacactgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atctcctgca aggccagcca aagtgttgat tttgatggtg atagttttat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctata ctacatccaa tctagaatct 180
gggatcccag ccaggtttag tgccagtggg tctgggacag acttcaccct caacatccat 240
cctgtggagg aggaggatac tgcaacctat tactgtcagc aaagtaatga ggatccgtac 300
acgttcggag gggggaccaa gctggaaata aaaggaggcg gatccggagg cggaggcgag 360
gtggagctag tggagtctgg gggaggctta gtgcagcctg gaaggtccct gaaactctcg 420
tgtgcagcct caggattcac tttcagtgac tattacatgg cctgggtccg gcaggctcca 480
acgacgggtc tggagtgggt cgcatccatt agttatgatg gtggtgacac tcactatcga 540
gactccgtga agggccgatt tactatttcc agagataatg caaaaagcag cctatacctg 600
caaatggaca gtctgaggtc tgaggacacg gccacttatt actgtgcaac agagactacg 660
ggaataccta caggtgttat ggatgcctgg ggtcaaggag tttcagtcac tgtctcctca 720
ctgggaggct gcggcgggag agctaagagg gcccctgtga agcagaccct gaacttcgac 780
ctgctgaagc tggccggaga cgtggagagc aaccccggcc ccgatgtcca gatgacccag 840
tctccatcta atcttgctgc ctctcctgga gaaagtgttt ccatcaattg caaggcaagt 900
gagagcatta gcaagtattt agcctggtat ctacagaaac ctgggaaagc aaataagctt 960
cttatgtacg atgggtcaac tttgcaatct ggaattccat cgaggttcag tggcagtgga 1020
tctggtacag atttcactct caccatcaga agcctggagc ctgaagattt tggactctat 1080
tactgtcaac agcattatga atatccagcc acgttcggtt ctgggaccaa gctggagatc 1140
aaaggaggcg gatccggagg cggaggccag gttaccctga aagagtctgg ccctgggata 1200
ttgcagccct cccagaccct cagtctgact tgttctttct ctgggttttc actgaggact 1260
tctggtatgg gtgtaggctg gattcgtcag ccttcaggga agggtctaga gtggctggca 1320
cacatttggt gggatgatga caagcgctat aatccagccc tgaagagccg actgacaatc 1380
tccaaggata cctccagcaa ccaggtattc ctcaaaatcg ccagtgtgga cactgcagat 1440
actgccacat actactgtgc tcaaataaac cccgcctggt ttgcttactg gggccaaggg 1500
actctggtca ctgtgagctc actgggaggc tgctag 1536
<210> 99
<211> 239
<212> Белок
<213> homo sapiens
<400> 99
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Phe Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Glu Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys
115 120 125
Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu
130 135 140
Ser Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys
145 150 155 160
Ala Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr
165 170 175
Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys
180 185 190
Asn Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala
195 200 205
Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly
210 215 220
Gln Gly Thr Leu Val Thr Val Ser Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 100
<211> 720
<212> ДНК
<213> homo sapiens
<400> 100
gaaattgtgc tgactcagtc tccagacttt cagtctgtga ctccaaagga gaaagtcacc 60
ttcacctgca ggaccagtca gagcattggc acaaacatac actggtacca gcagaaacca 120
gatcagtctc caaagctcct catcaaggag gtttctgagt ctatctctgg agtcccatcg 180
aggttcagtg gcagtggatc tgggacagat ttcaccctca ccatcaatag cctggaagct 240
gaagatgctg caacgtatta ctgtcaacaa agtaatacct ggccgttcac gttcggcgga 300
gggaccaagg tggagatcaa aggaggcgga tccggaggcg gaggccaggt taccctgaga 360
gagtctggcc ctgcgctggt gaagcccaca cagaccctca cactgacttg taccttctct 420
gggttttcac tgagcacttc tggtatgggt gtaggctgga ttcgtcagcc tcccgggaag 480
gctctagagt ggctggcaca catttggtgg gatgatgaca agcgctataa tccagccctg 540
aagagccgac tgacaatctc caaggatacc tccaaaaacc aggtagtcct cacaatgacc 600
aacatggacc ctgtggatac tgccacatac tactgtgctc aaataaaccc cgcctggttt 660
gcttactggg gccaagggac tctggtcact gtgagctcat tcaacagggg agagtgttag 720
<210> 101
<211> 246
<212> Белок
<213> Homo sapiens
<400> 101
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala
115 120 125
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
130 135 140
Gly Tyr Thr Phe Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Gln Gly Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr Tyr
165 170 175
Pro Asn Tyr Asn Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val Val
180 185 190
Val Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp
195 200 205
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr Tyr
210 215 220
Ser Gly Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Val Glu Pro Lys Ser Cys
245
<210> 102
<211> 741
<212> ДНК
<213> Homo sapiens
<400> 102
gacatcgtga tgacccaatc tccagactct ttggctgtgt ctctagggga gagggccacc 60
atcaactgca aggccagcca aagtgttgat tttgatggtg atagttttat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctata ctacatccaa tctagaatct 180
ggggtcccag acaggtttag tggcagtggg tctgggacag acttcaccct caccatcagc 240
agcctgcagg ctgaggatgt ggcagtttat tactgtcagc aaagtaatga ggatccgtac 300
acgttcggac aggggaccaa gcttgagatc aaaggaggcg gatccggagg cggaggccag 360
gttcagctgg tgcagtctgg agctgaggtg aagaagcctg gggcctcagt gaaggtctcc 420
tgcaaggctt ctggttacac ctttaccaac tactggatac actgggtgcg acaggcccct 480
ggacaagggc ttgagtggat tggagtgatt gatccttctg atacttatcc aaattacaat 540
aaaaagttca agggcagagt caccatgacc gtagtcgtat ccacgagcac agcctacatg 600
gagctgagga gcctgagatc tgacgacacg gccgtgtatt actgtgcgag aaacggtgat 660
tccgattatt actctggtat ggactactgg gggcaaggga ccacggtcac cgtctcctca 720
gttgagccca aatcttgtta g 741
<210> 103
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH FR1
<400> 103
Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Arg
20 25 30
<210> 104
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH FR1
<400> 104
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser
20 25 30
<210> 105
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH FR1
<400> 105
Gln Val Thr Leu Lys Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser
20 25 30
<210> 106
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH FR1
<400> 106
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Arg
20 25 30
<210> 107
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH FR1
<400> 107
Gln Ile Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser
20 25 30
<210> 108
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR1
<400> 108
Thr Ser Gly Met Gly Val Gly
1 5
<210> 109
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR1
<400> 109
Thr Ser Gly Val Gly Val Gly
1 5
<210> 110
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR2
<400> 110
Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu Trp Leu Ala
1 5 10
<210> 111
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR2
<400> 111
Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Ala
1 5 10
<210> 112
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 112
His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala Leu Lys Ser
1 5 10 15
<210> 113
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 113
His Ile Tyr Trp Asn Asp Asp Lys Arg Tyr Asn Pro Ala Leu Lys Ser
1 5 10 15
<210> 114
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 114
His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Ser Pro Ser Leu Lys Ser
1 5 10 15
<210> 115
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 115
His Ile Tyr Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala Leu Lys Ser
1 5 10 15
<210> 116
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 116
His Ile Trp Trp Asn Asp Asp Lys Arg Tyr Asn Pro Ala Leu Lys Ser
1 5 10 15
<210> 117
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 117
Leu Ile Asp Trp Asp Asp Asp Lys Arg Tyr Ser Pro Ser Leu Lys Ser
1 5 10 15
<210> 118
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 118
His Ile Phe Trp Asp Asp Asp Lys Arg Tyr Ser Pro Ser Leu Lys Ser
1 5 10 15
<210> 119
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 119
Leu Ile Trp Trp Asp Asp Asp Lys Arg Tyr Ser Pro Ser Leu Lys Ser
1 5 10 15
<210> 120
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 120
His Ile Asp Trp Asp Asp Asp Lys Arg Tyr Ser Pro Ser Leu Lys Ser
1 5 10 15
<210> 121
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR2
<400> 121
Leu Ile Trp Trp Asn Asp Asp Lys Arg Tyr Ser Pro Ser Leu Lys Ser
1 5 10 15
<210> 122
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 122
Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser Asn Gln Val Phe Leu Lys
1 5 10 15
Ile Asp Thr Ala Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 123
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 123
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25
<210> 124
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 124
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 125
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 125
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Thr
20 25
<210> 126
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 126
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Lys
20 25
<210> 127
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 127
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Ala
20 25
<210> 128
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 128
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala His
20 25
<210> 129
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 129
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25
<210> 130
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 130
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala His
20 25
<210> 131
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 131
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 132
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 132
Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser Asn Gln Val Phe Leu Lys
1 5 10 15
Ala Asp Thr Ala Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 133
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 133
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25
<210> 134
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 134
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 135
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 135
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Thr
20 25
<210> 136
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 136
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Lys
20 25
<210> 137
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 137
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Ala
20 25
<210> 138
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 138
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala His
20 25
<210> 139
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 139
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25
<210> 140
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 140
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala His
20 25
<210> 141
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 141
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Thr Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 142
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 142
Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser Asn Gln Val Phe Leu Lys
1 5 10 15
Ser Asp Thr Ala Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 143
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 143
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25
<210> 144
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 144
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 145
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 145
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Thr
20 25
<210> 146
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 146
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Lys
20 25
<210> 147
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 147
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Ala
20 25
<210> 148
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 148
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala His
20 25
<210> 149
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 149
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25
<210> 150
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 150
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala His
20 25
<210> 151
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 151
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Asn Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 152
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 152
Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser Asn Gln Val Phe Leu Lys
1 5 10 15
Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 153
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 153
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25
<210> 154
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 154
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 155
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 155
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Thr
20 25
<210> 156
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 156
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Lys
20 25
<210> 157
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 157
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Ala
20 25
<210> 158
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 158
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala His
20 25
<210> 159
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 159
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25
<210> 160
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 160
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala His
20 25
<210> 161
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR3
<400> 161
Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln
20 25
<210> 162
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR3
<400> 162
Ile Asn Pro Ala Trp Phe Ala Tyr
1 5
<210> 163
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR3
<400> 163
Ile Asn Pro Ala Trp Phe Asp Tyr
1 5
<210> 164
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR3
<400> 164
Ile Asn Pro Ala Tyr Phe Ala Tyr
1 5
<210> 165
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * CDR3
<400> 165
Ile Asn Pro Ala Tyr Phe Asp Tyr
1 5
<210> 166
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR4
<400> 166
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
1 5 10
<210> 167
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VH, происходящие из 3G8 VH * FR4
<400> 167
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 168
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* FR1
<400> 168
Asp Thr Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu
1 5 10 15
<210> 169
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* FR1
<400> 169
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu
1 5 10 15
<210> 170
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 170
Lys Ala Ser Gln Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 171
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 171
Arg Ala Ser Gln Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 172
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 172
Lys Ser Ser Gln Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 173
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 173
Lys Ala Ser Gln Asp Gly Asp Ser Tyr Met Asn
1 5 10
<210> 174
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 174
Lys Ala Ser Gln Asp Gly Asp Ser Phe Leu Asn
1 5 10
<210> 175
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 175
Lys Ala Ser Gln Asp Gly Asp Ser Phe Met Ala
1 5 10
<210> 176
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 176
Lys Ala Ser Gln Asp Gly Asp Ser Tyr Leu Ala
1 5 10
<210> 177
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 177
Lys Ala Ser Ser Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 178
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 178
Arg Ala Ser Ser Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 179
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 179
Lys Ser Ser Ser Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 180
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 180
Lys Ala Ser Ser Asp Gly Asp Ser Tyr Met Asn
1 5 10
<210> 181
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 181
Lys Ala Ser Ser Asp Gly Asp Ser Phe Leu Asn
1 5 10
<210> 182
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 182
Lys Ala Ser Ser Asp Gly Asp Ser Phe Met Ala
1 5 10
<210> 183
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 183
Lys Ala Ser Ser Asp Gly Asp Ser Tyr Leu Ala
1 5 10
<210> 184
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 184
Lys Ala Ser Val Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 185
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 185
Arg Ala Ser Val Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 186
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 186
Lys Ser Ser Val Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 187
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 187
Lys Ala Ser Val Asp Gly Asp Ser Tyr Met Asn
1 5 10
<210> 188
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 188
Lys Ala Ser Val Asp Gly Asp Ser Phe Leu Asn
1 5 10
<210> 189
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 189
Lys Ala Ser Val Asp Gly Asp Ser Phe Met Ala
1 5 10
<210> 190
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 190
Lys Ala Ser Val Asp Gly Asp Ser Tyr Leu Ala
1 5 10
<210> 191
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 191
Lys Ala Ser Asp Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 192
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 192
Arg Ala Ser Asp Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 193
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 193
Lys Ser Ser Asp Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 194
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 194
Lys Ala Ser Asp Asp Gly Asp Ser Tyr Met Asn
1 5 10
<210> 195
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 195
Lys Ala Ser Asp Asp Gly Asp Ser Phe Leu Asn
1 5 10
<210> 196
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 196
Lys Ala Ser Asp Asp Gly Asp Ser Phe Met Ala
1 5 10
<210> 197
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 197
Lys Ala Ser Asp Asp Gly Asp Ser Tyr Leu Ala
1 5 10
<210> 198
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 198
Lys Ala Ser Phe Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 199
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 199
Arg Ala Ser Phe Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 200
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 200
Lys Ser Ser Phe Asp Gly Asp Ser Phe Met Asn
1 5 10
<210> 201
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 201
Lys Ala Ser Phe Asp Gly Asp Ser Tyr Met Asn
1 5 10
<210> 202
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 202
Lys Ala Ser Phe Asp Gly Asp Ser Phe Leu Asn
1 5 10
<210> 203
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 203
Lys Ala Ser Phe Asp Gly Asp Ser Phe Met Ala
1 5 10
<210> 204
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR1
<400> 204
Lys Ala Ser Phe Asp Gly Asp Ser Tyr Leu Ala
1 5 10
<210> 205
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* FR2
<400> 205
Trp Tyr Gln Gln Lys Ala Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 206
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 206
Thr Thr Ser Asn Leu Glu Ser
1 5
<210> 207
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 207
Asp Ala Ser Asn Leu Glu Ser
1 5
<210> 208
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 208
Trp Ala Ser Asn Leu Glu Ser
1 5
<210> 209
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 209
Thr Thr Ser Asn Leu Glu Thr
1 5
<210> 210
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 210
Asp Ala Ser Asn Leu Glu Thr
1 5
<210> 211
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 211
Asp Ala Ser Asn Leu Ala Thr
1 5
<210> 212
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 212
Ser Ala Ser Asn Leu Gln Ser
1 5
<210> 213
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 213
Ser Thr Ser Asn Leu Glu Ser
1 5
<210> 214
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 214
Ser Thr Ser Asn Leu Gln Ser
1 5
<210> 215
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 215
Thr Thr Ser Asn Leu Gln Ser
1 5
<210> 216
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR2
<400> 216
Thr Thr Ser Ser Leu Gln Ser
1 5
<210> 217
<211> 32
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* FR3
<400> 217
Gly Ile Pro Ala Arg Phe Ser Ala Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Asn Ile His Pro Val Glu Glu Glu Asp Thr Ala Thr Tyr Tyr Cys
20 25 30
<210> 218
<211> 32
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* FR3
<400> 218
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys
20 25 30
<210> 219
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR3
<400> 219
Gln Gln Ser Asn Glu Asp Pro Tyr Thr
1 5
<210> 220
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR3
<400> 220
Gln Gln Ser Tyr Ser Thr Pro Tyr Thr
1 5
<210> 221
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR3
<400> 221
Gln Gln Ser Tyr Glu Asp Pro Tyr Thr
1 5
<210> 222
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR3
<400> 222
Gln Gln Ser Asn Ser Asp Pro Tyr Thr
1 5
<210> 223
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* CDR3
<400> 223
Gln Gln Ser Asn Glu Thr Pro Tyr Thr
1 5
<210> 224
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* FR4
<400> 224
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 225
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательности VL, происходящие из 3G8 VL* FR4
<400> 225
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 226
<211> 714
<212> ДНК
<213> Искусственная последовательность
<220>
<223> H3G8VL-CB3.1VH
<400> 226
gacatcgtga tgacccaatc tccagactct ttggctgtgt ctctagggga gagggccacc 60
atcaactgca aggccagcca aagtgttgat tttgatggtg atagttttat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctata ctacatccaa tctagaatct 180
ggggtcccag acaggtttag tggcagtggg tctgggacag acttcaccct caccatcagc 240
agcctgcagg ctgaggatgt ggcagtttat tactgtcagc aaagtaatga ggatccgtac 300
acgttcggac aggggaccaa gcttgagatc aaaggaggcg gatccggagg cggaggccag 360
gtccaactgc agcagcctgg ggctgagctg gtgaggcctg gggcttcagt gaagctgtcc 420
tgcaaggctt ctggctacac cttcaccagc tactggatga actgggtgaa gcagaggcct 480
ggacaaggcc ttgaatggat tggtatggtt gatccttcag acagtgaaac tcactacaat 540
caaatgttca aggacaaggc cacattgact gttgacaaat cctccagcac agcctacatg 600
cagctcagca gcctgacatc tgaggactct gcggtctatt actgtgcaag agctatgggc 660
tactggggtc aaggaacctc agtcaccgtc tcctcagttg agcccaaatc ttgt 714
<210> 227
<211> 238
<212> Белок
<213> Искусственная последовательность
<220>
<223> H3G8VL-CB3.1VH
<400> 227
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Gln Gln Pro Gly Ala
115 120 125
Glu Leu Val Arg Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser
130 135 140
Gly Tyr Thr Phe Thr Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro
145 150 155 160
Gly Gln Gly Leu Glu Trp Ile Gly Met Val Asp Pro Ser Asp Ser Glu
165 170 175
Thr His Tyr Asn Gln Met Phe Lys Asp Lys Ala Thr Leu Thr Val Asp
180 185 190
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
195 200 205
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln
210 215 220
Gly Thr Ser Val Thr Val Ser Ser Val Glu Pro Lys Ser Cys
225 230 235
<210> 228
<211> 732
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CB3.1VL-h3G8VH
<400> 228
gatgttgtga tgacccagac tccactcact ttgtcggtta acattggaca accagcctcc 60
atctcttgta agtcaagtca gagcctctta gatactgatg gaaagacata tttgaattgg 120
ttgttacaga ggccaggcca gtctccaaac cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggaatt tattattgct ggcaaggtac acattttccg 300
ctcacgttcg gtgctgggac caagctggag ctgaaaggag gcggatccgg aggcggaggc 360
caggttaccc tgagagagtc tggccctgcg ctggtgaagc ccacacagac cctcacactg 420
acttgtacct tctctgggtt ttcactgagc acttctggta tgggtgtagg ctggattcgt 480
cagcctcccg ggaaggctct agagtggctg gcacacattt ggtgggatga tgacaagcgc 540
tataatccag ccctgaagag ccgactgaca atctccaagg atacctccaa aaaccaggta 600
gtcctcacaa tgaccaacat ggaccctgtg gatactgcca catactactg tgctcaaata 660
aaccccgcct ggtttgctta ctggggccaa gggactctgg tcactgtgag ctcattcaac 720
aggggagagt gt 732
<210> 229
<211> 244
<212> Белок
<213> Искусственная последовательность
<220>
<223> CB3.1VL-h3G8VH
<400> 229
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Asn Ile Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Thr
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Gln Val Thr Leu Arg Glu Ser Gly
115 120 125
Pro Ala Leu Val Lys Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Phe
130 135 140
Ser Gly Phe Ser Leu Ser Thr Ser Gly Met Gly Val Gly Trp Ile Arg
145 150 155 160
Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Ala His Ile Trp Trp Asp
165 170 175
Asp Asp Lys Arg Tyr Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser
180 185 190
Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr Met Thr Asn Met Asp
195 200 205
Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp
210 215 220
Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Phe Asn
225 230 235 240
Arg Gly Glu Cys
<210> 230
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 8B5-CB3.1-VEPKSC
<400> 230
gacattcaga tgacacagtc tccatcctcc ctacttgcgg cgctgggaga aagagtcagt 60
ctcacttgtc gggcaagtca ggaaattagt ggttacttaa gctggcttca gcagaaacca 120
gatggaacta ttaaacgcct gatctacgcc gcatccactt tagattctgg tgtcccaaaa 180
aggttcagtg gcagtgagtc tgggtcagat tattctctca ccatcagcag tcttgagtct 240
gaagattttg cagactatta ctgtctacaa tattttagtt atccgctcac gttcggtgct 300
gggaccaagc tggagctgaa aggaggcgga tccggaggcg gaggccaggt ccaactgcag 360
cagcctgggg ctgagctggt gaggcctggg gcttcagtga agctgtcctg caaggcttct 420
ggctacacct tcaccagcta ctggatgaac tgggtgaagc agaggcctgg acaaggcctt 480
gaatggattg gtatggttga tccttcagac agtgaaactc actacaatca aatgttcaag 540
gacaaggcca cattgactgt tgacaaatcc tccagcacag cctacatgca gctcagcagc 600
ctgacatctg aggactctgc ggtctattac tgtgcaagag ctatgggcta ctggggtcaa 660
ggaacctcag tcaccgtctc ctcagttgag cccaaatctt gt 702
<210> 231
<211> 234
<212> Белок
<213> Искусственная последовательность
<220>
<223> 8B5-CB3.1-VEPKSC
<400> 231
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Leu Ala Ala Leu Gly
1 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Ala Asp Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
115 120 125
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Val Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Met Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
180 185 190
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Ser Val
210 215 220
Thr Val Ser Ser Val Glu Pro Lys Ser Cys
225 230
<210> 232
<211> 726
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CB3.1-8B5-FNRGEC
<400> 232
gatgttgtga tgacccagac tccactcact ttgtcggtta acattggaca accagcctcc 60
atctcttgta agtcaagtca gagcctctta gatactgatg gaaagacata tttgaattgg 120
ttgttacaga ggccaggcca gtctccaaac cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggaatt tattattgct ggcaaggtac acattttccg 300
ctcacgttcg gtgctgggac caagctggag ctgaaaggag gcggatccgg aggcggaggc 360
gaagtgaagc ttgaggagtc tggaggaggc ttggtgcaac ctggaggatc catgaaactc 420
tcttgtgaag cctctggatt cacttttagt gacgcctgga tggactgggt ccgtcagtct 480
ccagagaagg ggcttgagtg ggttgctgaa attagaaaca aagctaaaaa tcatgcaaca 540
tactatgctg agtctgtgat agggaggttc accatctcaa gagatgattc caaaagtagt 600
gtctacctgc aaatgaacag cttaagagct gaagacactg gcatttatta ctgtggggct 660
ctgggccttg actactgggg ccaaggcacc actctcacag tctcctcgtt caacagggga 720
gagtgt 726
<210> 233
<211> 242
<212> Белок
<213> Искусственная последовательность
<220>
<223> CB3.1-8B5-FNRGEC
<400> 233
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Asn Ile Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Thr
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Lys Leu Glu Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Met Lys Leu Ser Cys Glu Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ser
145 150 155 160
Pro Glu Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ser Lys Ser Ser Val Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Gly Ile Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Phe Asn Arg Gly
225 230 235 240
Glu Cys
<210> 234
<211> 696
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 8B5VL-CB3.1VH-LGGC
<400> 234
gacattcaga tgacacagtc tccatcctcc ctacttgcgg cgctgggaga aagagtcagt 60
ctcacttgtc gggcaagtca ggaaattagt ggttacttaa gctggcttca gcagaaacca 120
gatggaacta ttaaacgcct gatctacgcc gcatccactt tagattctgg tgtcccaaaa 180
aggttcagtg gcagtgagtc tgggtcagat tattctctca ccatcagcag tcttgagtct 240
gaagattttg cagactatta ctgtctacaa tattttagtt atccgctcac gttcggtgct 300
gggaccaagc tggagctgaa aggaggcgga tccggaggcg gaggccaggt ccaactgcag 360
cagcctgggg ctgagctggt gaggcctggg gcttcagtga agctgtcctg caaggcttct 420
ggctacacct tcaccagcta ctggatgaac tgggtgaagc agaggcctgg acaaggcctt 480
gaatggattg gtatggttga tccttcagac agtgaaactc actacaatca aatgttcaag 540
gacaaggcca cattgactgt tgacaaatcc tccagcacag cctacatgca gctcagcagc 600
ctgacatctg aggactctgc ggtctattac tgtgcaagag ctatgggcta ctggggtcaa 660
ggaacctcag tcaccgtctc ctcactggga ggctgc 696
<210> 235
<211> 232
<212> Белок
<213> Искусственная последовательность
<220>
<223> 8B5VL-CB3.1VH-LGGC
<400> 235
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Leu Ala Ala Leu Gly
1 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Ala Asp Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
115 120 125
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Val Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Met Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
180 185 190
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Ser Val
210 215 220
Thr Val Ser Ser Leu Gly Gly Cys
225 230
<210> 236
<211> 720
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CB3.1-8B5-LGGC
<400> 236
gatgttgtga tgacccagac tccactcact ttgtcggtta acattggaca accagcctcc 60
atctcttgta agtcaagtca gagcctctta gatactgatg gaaagacata tttgaattgg 120
ttgttacaga ggccaggcca gtctccaaac cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggaatt tattattgct ggcaaggtac acattttccg 300
ctcacgttcg gtgctgggac caagctggag ctgaaaggag gcggatccgg aggcggaggc 360
gaagtgaagc ttgaggagtc tggaggaggc ttggtgcaac ctggaggatc catgaaactc 420
tcttgtgaag cctctggatt cacttttagt gacgcctgga tggactgggt ccgtcagtct 480
ccagagaagg ggcttgagtg ggttgctgaa attagaaaca aagctaaaaa tcatgcaaca 540
tactatgctg agtctgtgat agggaggttc accatctcaa gagatgattc caaaagtagt 600
gtctacctgc aaatgaacag cttaagagct gaagacactg gcatttatta ctgtggggct 660
ctgggccttg actactgggg ccaaggcacc actctcacag tctcctcgct gggaggctgc 720
<210> 237
<211> 240
<212> Белок
<213> Искусственная последовательность
<220>
<223> CB3.1-8B5-LGGC
<400> 237
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Asn Ile Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Thr
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Lys Leu Glu Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Met Lys Leu Ser Cys Glu Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ser
145 150 155 160
Pro Glu Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ser Lys Ser Ser Val Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Gly Ile Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Leu Gly Gly Cys
225 230 235 240
<210> 238
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh628F
<400> 238
ggaggcggat ccggaggcgg aggccaggtc caactgcagc agcctgg 47
<210> 239
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh629R
<400> 239
tttgaattct aacaagattt gggctcaact gaggagacgg tgactgagg 49
<210> 240
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh630R
<400> 240
gcctccgcct ccggatccgc ctcctttcag ctccagcttg gtccc 45
<210> 241
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh631F
<400> 241
ggaggcggat ccggaggcgg aggcgaagtg aagcttgagg agtctgg 47
<210> 242
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh640R
<400> 242
tttgaattct aacactctcc cctgttgaac gaggagactg tgagagtgg 49
<210> 243
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh644R
<400> 243
tttgtcgtca tcatcgtctt tgtagtcgga gtggacacct gtggagag 48
<210> 244
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh646R
<400> 244
tttgaattct agcagcctcc cagtgaggag acggtgactg ag 42
<210> 245
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh647F
<400> 245
caaagacgat gatgacgaca aagacattca gatgacacag tctcc 45
<210> 246
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh648R
<400> 246
tttgaattct agcagcctcc cagcgaggag actgtgagag tgg 43
<210> 247
<211> 1044
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 2.4G2-3G8-hKappa
<400> 247
gatgtccaga tgacccagtc tccatctaat cttgctgcct ctcctggaga aagtgtttcc 60
atcaattgca aggcaagtga gagcattagc aagtatttag cctggtatct acagaaacct 120
gggaaagcaa ataagcttct tatgtacgat gggtcaactt tgcaatctgg aattccatcg 180
aggttcagtg gcagtggatc tggtacagat ttcactctca ccatcagaag cctggagcct 240
gaagattttg gactctatta ctgtcaacag cattatgaat atccagccac gttcggttct 300
gggaccaagc tggagatcaa aggaggcgga tccggaggcg gaggccaggt taccctgaaa 360
gagtctggcc ctgggatatt gcagccctcc cagaccctca gtctgacttg ttctttctct 420
gggttttcac tgaggacttc tggtatgggt gtaggctgga ttcgtcagcc ttcagggaag 480
ggtctagagt ggctggcaca catttggtgg gatgatgaca agcgctataa tccagccctg 540
aagagccgac tgacaatctc caaggatacc tccagcaacc aggtattcct caaaatcgcc 600
agtgtggaca ctgcagatac tgccacatac tactgtgctc aaataaaccc cgcctggttt 660
gcttactggg gccaagggac tctggtcact gtgagctcac tgggaggctg cggcggaggg 720
agccgtacgg tggctgcacc atcggtcttc atcttcccgc catctgatga gcagttgaaa 780
tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta 840
cagtggaagg tggataacgc cctccaatcg ggtaactccc aggagagtgt cacagagcag 900
gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac 960
gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca 1020
aagagcttca acaggggaga gtgt 1044
<210> 248
<211> 348
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2.4G2-3G8-hKappa
<400> 248
Asp Val Gln Met Thr Gln Ser Pro Ser Asn Leu Ala Ala Ser Pro Gly
1 5 10 15
Glu Ser Val Ser Ile Asn Cys Lys Ala Ser Glu Ser Ile Ser Lys Tyr
20 25 30
Leu Ala Trp Tyr Leu Gln Lys Pro Gly Lys Ala Asn Lys Leu Leu Met
35 40 45
Tyr Asp Gly Ser Thr Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Gly Leu Tyr Tyr Cys Gln Gln His Tyr Glu Tyr Pro Ala
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Gln
115 120 125
Pro Ser Gln Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu
130 135 140
Arg Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys
145 150 155 160
Gly Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr
165 170 175
Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser
180 185 190
Asn Gln Val Phe Leu Lys Ile Ala Ser Val Asp Thr Ala Asp Thr Ala
195 200 205
Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly
210 215 220
Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly Cys Gly Gly Gly
225 230 235 240
Ser Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
245 250 255
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
260 265 270
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
275 280 285
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
290 295 300
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
305 310 315 320
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
325 330 335
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
340 345
<210> 249
<211> 1734
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3G8-2.4G2-hG1
<400> 249
gacactgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atctcctgca aggccagcca aagtgttgat tttgatggtg atagttttat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctata ctacatccaa tctagaatct 180
gggatcccag ccaggtttag tgccagtggg tctgggacag acttcaccct caacatccat 240
cctgtggagg aggaggatac tgcaacctat tactgtcagc aaagtaatga ggatccgtac 300
acgttcggag gggggaccaa gctggaaata aaaggaggcg gatccggagg cggaggcgag 360
gtggagctag tggagtctgg gggaggctta gtgcagcctg gaaggtccct gaaactctcg 420
tgtgcagcct caggattcac tttcagtgac tattacatgg cctgggtccg gcaggctcca 480
acgacgggtc tggagtgggt cgcatccatt agttatgatg gtggtgacac tcactatcga 540
gactccgtga agggccgatt tactatttcc agagataatg caaaaagcag cctatacctg 600
caaatggaca gtctgaggtc tgaggacacg gccacttatt actgtgcaac agagactacg 660
ggaataccta caggtgttat ggatgcctgg ggtcaaggag tttcagtcac tgtctcctca 720
ctgggaggct gcggcggagg gagcgcctcc accaagggcc catcggtctt ccccctggca 780
ccctcctcca agagcacctc tgggggcaca gcggccctgg gctgcctggt caaggactac 840
ttccccgaac cggtgacggt gtcgtggaac tcaggcgccc tgaccagcgg cgtgcacacc 900
ttcccggctg tcctacagtc ctcaggactc tactccctca gcagcgtggt gaccgtgccc 960
tccagcagct tgggcaccca gacctacatc tgcaacgtga atcacaagcc cagcaacacc 1020
aaggtggaca agagagttga gcccaaatct tgtgacaaaa ctcacacatg cccaccgtgc 1080
ccagcacctg aactcctggg gggaccgtca gtcttcctct tccccccaaa acccaaggac 1140
accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa 1200
gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca 1260
aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg 1320
caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca 1380
gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac 1440
accctgcccc catcccggga tgagctgacc aagaaccagg tcagcctgac ctgcctggtc 1500
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 1560
aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct ctacagcaag 1620
ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat 1680
gaggctctgc acaaccacta cacgcagaag agcctctccc tgtctccggg taaa 1734
<210> 250
<211> 578
<212> Белок
<213> Искусственная последовательность
<220>
<223> 3G8-2.4G2-hG1
<400> 250
Asp Thr Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Ala Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Thr Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Glu Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro
145 150 155 160
Thr Thr Gly Leu Glu Trp Val Ala Ser Ile Ser Tyr Asp Gly Gly Asp
165 170 175
Thr His Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
180 185 190
Asn Ala Lys Ser Ser Leu Tyr Leu Gln Met Asp Ser Leu Arg Ser Glu
195 200 205
Asp Thr Ala Thr Tyr Tyr Cys Ala Thr Glu Thr Thr Gly Ile Pro Thr
210 215 220
Gly Val Met Asp Ala Trp Gly Gln Gly Val Ser Val Thr Val Ser Ser
225 230 235 240
Leu Gly Gly Cys Gly Gly Gly Ser Ala Ser Thr Lys Gly Pro Ser Val
245 250 255
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
260 265 270
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
275 280 285
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
290 295 300
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
305 310 315 320
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
325 330 335
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
340 345 350
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
385 390 395 400
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
465 470 475 480
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
485 490 495
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
565 570 575
Gly Lys
<210> 251
<211> 720
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CD79VL-CD32BVH
<400> 251
gatgttgtga tgactcagtc tccactctcc ctgcccgtca cccttggaca gccggcctcc 60
atctcctgca agtcaagtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttcagcaga ggccaggcca atctccaaac cgcctaattt atctggtgtc taaactggac 180
tctggggtcc cagacagatt cagcggcagt gggtcaggca ctgatttcac actgaaaatc 240
agcagggtgg aggctgagga tgttggggtt tattactgct ggcaaggtac acattttccg 300
ctcacgttcg gcggagggac caagcttgag atcaaaggag gcggatccgg aggcggaggc 360
gaagtgaagc ttgaggagtc tggaggaggc ttggtgcaac ctggaggatc catgaaactc 420
tcttgtgaag cctctggatt cacttttagt gacgcctgga tggactgggt ccgtcagtct 480
ccagagaagg ggcttgagtg ggttgctgaa attagaaaca aagctaaaaa tcatgcaaca 540
tactatgctg agtctgtgat agggaggttc accatctcaa gagatgattc caaaagtagt 600
gtctacctgc aaatgaacag cttaagagct gaagacactg gcatttatta ctgtggggct 660
ctgggccttg actactgggg ccaaggcacc actctcacag tctcctcgct gggaggctgc 720
<210> 252
<211> 240
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD79VL-CD32BVH
<400> 252
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly Cys
225 230 235 240
<210> 253
<211> 696
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CD32BVL-CD79VH-1
<400> 253
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctgtgggaga tagagtcacc 60
atcacttgtc gggcaagtca ggaaattagt ggttacttaa gctggctgca gcagaaacca 120
ggcaaggccc ctagacgcct gatctacgcc gcatccactt tagattctgg tgtcccatcc 180
aggttcagtg gcagtgagtc tgggaccgag ttcaccctca ccatcagcag ccttcagcct 240
gaagattttg caacctatta ctgtctacaa tattttagtt atccgctcac gttcggaggg 300
gggaccaagg tggaaataaa aggaggcgga tccggaggcg gaggccaggt tcagctggtg 360
cagtctggag ctgaggtgaa gaagcctggc gcctcagtga aggtctcctg caaggcttct 420
ggttacacct ttaccagcta ctggatgaac tgggtgcgac aggcccctgg acaagggctt 480
gagtggatcg gaatgattga tccttcagac agtgaaactc actacaatca aatgttcaag 540
gacagagtca ccatgaccac agacacatcc acgagcacag cctacatgga gctgaggagc 600
ctgagatctg acgacacggc cgtgtattac tgtgcgagag ctatgggcta ctgggggcaa 660
gggaccacgg tcaccgtctc ctcactggga ggctgc 696
<210> 254
<211> 232
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD32BVL-CD79VH-1
<400> 254
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Met Phe Lys Asp Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val
210 215 220
Thr Val Ser Ser Leu Gly Gly Cys
225 230
<210> 255
<211> 696
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CD32BVL-CD79VH-2
<400> 255
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctgtgggaga tagagtcacc 60
atcacttgtc gggcaagtca ggaaattagt ggttacttaa gctggctgca gcagaaacca 120
ggcaaggccc ctagacgcct gatctacgcc gcatccactt tagattctgg tgtcccatcc 180
aggttcagtg gcagtgagtc tgggaccgag ttcaccctca ccatcagcag ccttcagcct 240
gaagattttg caacctatta ctgtctacaa tattttagtt atccgctcac gttcggaggg 300
gggaccaagg tggaaataaa aggaggcgga tccggaggcg gaggccaggt tcagctggtg 360
cagtctggag ctgaggtgaa gaagcctggc gcctcagtga aggtctcctg caaggcttct 420
ggttacacct ttaccagcta ctggatgaac tgggtgcgac aggcccctgg acaagggctt 480
gagtggatcg gaatgattga tccttcagac agtgaaactc actacaatca aaagttcaag 540
gacagagtca ccatgaccac agacacatcc acgagcacag cctacatgga gctgaggagc 600
ctgagatctg acgacacggc cgtgtattac tgtgcgagag ctatgggcta ctgggggcaa 660
gggaccacgg tcaccgtctc ctcactggga ggctgc 696
<210> 256
<211> 232
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD32BVL-CD79VH-2
<400> 256
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Lys Phe Lys Asp Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val
210 215 220
Thr Val Ser Ser Leu Gly Gly Cys
225 230
<210> 257
<211> 720
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hHBCRCVL. R45N-h8B5VH-LGGC
<400> 257
gatgttgtga tgactcagtc tccactctcc ctgcccgtca cccttggaca gccggcctcc 60
atctcctgca agtcaagtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttcagcaga ggccaggcca atctccaaac cgcctaattt atctggtgtc taaactggac 180
tctggggtcc cagacagatt cagcggcagt gggtcaggca ctgatttcac actgaaaatc 240
agcagggtgg aggctgagga tgttggggtt tattactgct ggcaaggtac acattttccg 300
ctcacgttcg gcggagggac caagcttgag atcaaaggag gcggatccgg aggcggaggc 360
gaagtgaagc ttgaggagtc tggaggaggc ttggtgcaac ctggaggatc catgaaactc 420
tcttgtgaag cctctggatt cacttttagt gacgcctgga tggactgggt ccgtcagtct 480
ccagagaagg ggcttgagtg ggttgctgaa attagaaaca aagctaaaaa tcatgcaaca 540
tactatgctg agtctgtgat agggaggttc accatctcaa gagatgattc caaaagtagt 600
gtctacctgc aaatgaacag cttaagagct gaagacactg gcatttatta ctgtggggct 660
ctgggccttg actactgggg ccaaggcacc actctcacag tctcctcgct gggaggctgc 720
<210> 258
<211> 240
<212> Белок
<213> Искусственная последовательность
<220>
<223> hHBCRCVL. R45N-h8B5VH-LGGC
<400> 258
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly Cys
225 230 235 240
<210> 259
<211> 696
<212> ДНК
<213> Искусственная последовательность
<220>
<223> H8B5VL-hHBCRCVH M48I-LGGC
<400> 259
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctgtgggaga tagagtcacc 60
atcacttgtc gggcaagtca ggaaattagt ggttacttaa gctggctgca gcagaaacca 120
ggcaaggccc ctagacgcct gatctacgcc gcatccactt tagattctgg tgtcccatcc 180
aggttcagtg gcagtgagtc tgggaccgag ttcaccctca ccatcagcag ccttcagcct 240
gaagattttg caacctatta ctgtctacaa tattttagtt atccgctcac gttcggaggg 300
gggaccaagg tggaaataaa aggaggcgga tccggaggcg gaggccaggt tcagctggtg 360
cagtctggag ctgaggtgaa gaagcctggc gcctcagtga aggtctcctg caaggcttct 420
ggttacacct ttaccagcta ctggatgaac tgggtgcgac aggcccctgg acaagggctt 480
gagtggatcg gaatgattga tccttcagac agtgaaactc actacaatca aatgttcaag 540
gacagagtca ccatgaccac agacacatcc acgagcacag cctacatgga gctgaggagc 600
ctgagatctg acgacacggc cgtgtattac tgtgcgagag ctatgggcta ctgggggcaa 660
gggaccacgg tcaccgtctc ctcactggga ggctgc 696
<210> 260
<211> 232
<212> Белок
<213> Искусственная последовательность
<220>
<223> H8B5VL-HBCRCVH M48I-LGGC
<400> 260
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Met Phe Lys Asp Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val
210 215 220
Thr Val Ser Ser Leu Gly Gly Cys
225 230
<210> 261
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh321F
<400> 261
cgagctagct ctagatgaga tcacagttct ctctac 36
<210> 262
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh386R
<400> 262
tttgaattct agcagcctcc cagtgaggag acggtgaccg tggtc 45
<210> 263
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh784F
<400> 263
ggcggatccg gaggcggagg ccaggttcag ctggtgcag 39
<210> 264
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh785R
<400> 264
cctccggatc cgcctccttt gatctcaagc ttggtccc 38
<210> 265
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh786R
<400> 265
tttgaattct agcagcctcc caggctggag acggtcacca gg 42
<210> 266
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Lgh787F
<400> 266
ggaggcggat ccggaggcgg aggcgaagtg cagcttgtgg agtc 44
<210> 267
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 267
Leu Gly Gly Cys Gly Gly Gly Ser
1 5
<210> 268
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 268
Leu Glu Ile Lys
1
<210> 269
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 269
Thr Val Ser Ser
1
<210> 270
<211> 343
<212> Белок
<213> Искусственная последовательность
<220>
<223> H8B5VL-hBCRCVH M48I, M62K_LGGCG3S_hKappa
<400> 270
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Lys Phe Lys Asp Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val
210 215 220
Thr Val Ser Ser Leu Gly Gly Cys Gly Gly Gly Ser Arg Thr Val Ala
225 230 235 240
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
245 250 255
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
260 265 270
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
275 280 285
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
290 295 300
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
305 310 315 320
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
325 330 335
Ser Phe Asn Arg Gly Glu Cys
340
<210> 271
<211> 1032
<212> ДНК
<213> Искусственная последовательность
<220>
<223> H8B5VL-hBCRCVH M48I, M62K_LGGCG3S_hKappa
<400> 271
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctgtgggaga tagagtcacc 60
atcacttgtc gggcaagtca ggaaattagt ggttacttaa gctggctgca gcagaaacca 120
ggcaaggccc ctagacgcct gatctacgcc gcatccactt tagattctgg tgtcccatcc 180
aggttcagtg gcagtgagtc tgggaccgag ttcaccctca ccatcagcag ccttcagcct 240
gaagattttg caacctatta ctgtctacaa tattttagtt atccgctcac gttcggaggg 300
gggaccaagg tggaaataaa aggaggcgga tccggaggcg gaggccaggt tcagctggtg 360
cagtctggag ctgaggtgaa gaagcctggc gcctcagtga aggtctcctg caaggcttct 420
ggttacacct ttaccagcta ctggatgaac tgggtgcgac aggcccctgg acaagggctt 480
gagtggatcg gaatgattga tccttcagac agtgaaactc actacaatca aaagttcaag 540
gacagagtca ccatgaccac agacacatcc acgagcacag cctacatgga gctgaggagc 600
ctgagatctg acgacacggc cgtgtattac tgtgcgagag ctatgggcta ctgggggcaa 660
gggaccacgg tcaccgtctc ctcactggga ggctgcggcg gagggagccg aactgtggct 720
gcaccatcgg tcttcatctt cccgccatct gatgagcagt tgaaatctgg aactgcctct 780
gttgtgtgcc tgctgaataa cttctatccc agagaggcca aagtacagtg gaaggtggat 840
aacgccctcc aatcgggtaa ctcccaggag agtgtcacag agcaggacag caaggacagc 900
acctacagcc tcagcagcac cctgacgctg agcaaagcag actacgagaa acacaaagtc 960
tacgcctgcg aagtcaccca tcagggcctg agctcgcccg tcacaaagag cttcaacagg 1020
ggagagtgtt ag 1032
<210> 272
<211> 574
<212> Белок
<213> Искусственная последовательность
<220>
<223> HBCRCVL R45N-h8B5VH_LGGCGGGS-hG1
<400> 272
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Leu Gly Gly Cys
225 230 235 240
Gly Gly Gly Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
340 345 350
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
355 360 365
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
370 375 380
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
385 390 395 400
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
405 410 415
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
420 425 430
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
435 440 445
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
450 455 460
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
465 470 475 480
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
485 490 495
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
500 505 510
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
515 520 525
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
530 535 540
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
545 550 555 560
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
565 570
<210> 273
<211> 1722
<212> ДНК
<213> Искусственная последовательность
<220>
<223> HBCRCVL R45N-h8B5VH_LGGCGGGS-hG1
<400> 273
gatgttgtga tgactcagtc tccactctcc ctgcccgtca cccttggaca gccggcctcc 60
atctcctgca agtcaagtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttcagcaga ggccaggcca atctccaaac cgcctaattt atctggtgtc taaactggac 180
tctggggtcc cagacagatt cagcggcagt gggtcaggca ctgatttcac actgaaaatc 240
agcagggtgg aggctgagga tgttggggtt tattactgct ggcaaggtac acattttccg 300
ctcacgttcg gcggagggac caagcttgag atcaaaggag gcggatccgg aggcggaggc 360
gaagtgcagc ttgtggagtc tggaggaggc ttggtgcaac ctggaggatc cctgagactc 420
tcttgtgccg cctctggatt cacttttagt gacgcctgga tggactgggt ccgtcaggcc 480
ccaggcaagg ggcttgagtg ggttgctgaa attagaaaca aagctaaaaa tcatgcaaca 540
tactatgctg agtctgtgat agggaggttc accatctcaa gagatgacgc caaaaacagt 600
ctgtacctgc aaatgaacag cttaagagct gaagacactg ccgtgtatta ctgtggggct 660
ctgggccttg actactgggg ccaaggcacc ctggtgaccg tctccagcct gggaggctgc 720
ggcggaggga gcgcctccac caagggccca tcggtcttcc ccctggcacc ctcctccaag 780
agcacctctg ggggcacagc ggccctgggc tgcctggtca aggactactt ccccgaaccg 840
gtgacggtgt cgtggaactc aggcgccctg accagcggcg tgcacacctt cccggctgtc 900
ctacagtcct caggactcta ctccctcagc agcgtggtga ccgtgccctc cagcagcttg 960
ggcacccaga cctacatctg caacgtgaat cacaagccca gcaacaccaa ggtggacaag 1020
agagttgagc ccaaatcttg tgacaaaact cacacatgcc caccgtgccc agcacctgaa 1080
ctcctggggg gaccgtcagt cttcctcttc cccccaaaac ccaaggacac cctcatgatc 1140
tcccggaccc ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc 1200
aagttcaact ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccgcgggag 1260
gagcagtaca acagcacgta ccgtgtggtc agcgtcctca ccgtcctgca ccaggactgg 1320
ctgaatggca aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag 1380
aaaaccatct ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca 1440
tcccgggatg agctgaccaa gaaccaggtc agcctgacct gcctggtcaa aggcttctat 1500
cccagcgaca tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc 1560
acgcctcccg tgctggactc cgacggctcc ttcttcctct acagcaagct caccgtggac 1620
aagagcaggt ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac 1680
aaccactaca cgcagaagag cctctccctg tctccgggta aa 1722
<210> 274
<211> 335
<212> Белок
<213> Искусственная последовательность
<220>
<223> H8B5VL-HBCRCVH M48I, M62K_(-4)LEIK_hKappa
<400> 274
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Lys Phe Lys Asp Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val
210 215 220
Leu Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
245 250 255
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
260 265 270
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
275 280 285
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
290 295 300
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
305 310 315 320
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 275
<211> 1005
<212> ДНК
<213> Искусственная последовательность
<220>
<223> H8B5VL-HBCRCVH M48I, M62K_(-4)LEIK_hKappa
<400> 275
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctgtgggaga tagagtcacc 60
atcacttgtc gggcaagtca ggaaattagt ggttacttaa gctggctgca gcagaaacca 120
ggcaaggccc ctagacgcct gatctacgcc gcatccactt tagattctgg tgtcccatcc 180
aggttcagtg gcagtgagtc tgggaccgag ttcaccctca ccatcagcag ccttcagcct 240
gaagattttg caacctatta ctgtctacaa tattttagtt atccgctcac gttcggaggg 300
gggaccaagg tggaaataaa aggaggcgga tccggaggcg gaggccaggt tcagctggtg 360
cagtctggag ctgaggtgaa gaagcctggc gcctcagtga aggtctcctg caaggcttct 420
ggttacacct ttaccagcta ctggatgaac tgggtgcgac aggcccctgg acaagggctt 480
gagtggatcg gaatgattga tccttcagac agtgaaactc actacaatca aaagttcaag 540
gacagagtca ccatgaccac agacacatcc acgagcacag cctacatgga gctgaggagc 600
ctgagatctg acgacacggc cgtgtattac tgtgcgagag ctatgggcta ctgggggcaa 660
gggaccacgg tcctggagat caagcgaact gtggctgcac catcggtctt catcttcccg 720
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 780
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 840
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 900
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 960
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgt 1005
<210> 276
<211> 566
<212> Белок
<213> Искусственная последовательность
<220>
<223> HBCRCVL R45N-h8B5VH_(-4)TVSS-hG1 = HBCRCVL R45N-h8B5VH_-hG1
<400> 276
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
225 230 235 240
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
245 250 255
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
260 265 270
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
275 280 285
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
290 295 300
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
305 310 315 320
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro
325 330 335
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
340 345 350
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
355 360 365
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
370 375 380
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
385 390 395 400
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
405 410 415
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
420 425 430
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
435 440 445
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
450 455 460
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
465 470 475 480
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
485 490 495
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
500 505 510
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
515 520 525
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
530 535 540
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
545 550 555 560
Ser Leu Ser Pro Gly Lys
565
<210> 277
<211> 1698
<212> ДНК
<213> Искусственная последовательность
<220>
<223> HBCRCVL R45N-h8B5VH_(-4)TVSS-hG1
<400> 277
gatgttgtga tgactcagtc tccactctcc ctgcccgtca cccttggaca gccggcctcc 60
atctcctgca agtcaagtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttcagcaga ggccaggcca atctccaaac cgcctaattt atctggtgtc taaactggac 180
tctggggtcc cagacagatt cagcggcagt gggtcaggca ctgatttcac actgaaaatc 240
agcagggtgg aggctgagga tgttggggtt tattactgct ggcaaggtac acattttccg 300
ctcacgttcg gcggagggac caagcttgag atcaaaggag gcggatccgg aggcggaggc 360
gaagtgcagc ttgtggagtc tggaggaggc ttggtgcaac ctggaggatc cctgagactc 420
tcttgtgccg cctctggatt cacttttagt gacgcctgga tggactgggt ccgtcaggcc 480
ccaggcaagg ggcttgagtg ggttgctgaa attagaaaca aagctaaaaa tcatgcaaca 540
tactatgctg agtctgtgat agggaggttc accatctcaa gagatgacgc caaaaacagt 600
ctgtacctgc aaatgaacag cttaagagct gaagacactg ccgtgtatta ctgtggggct 660
ctgggccttg actactgggg ccaaggcacc ctggtgaccg tctccagcgc ctccaccaag 720
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 780
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 840
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 900
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 960
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 1020
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 1080
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 1140
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 1200
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 1260
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1320
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1380
cagccccgag aaccacaggt gtacaccctg cccccatccc gggatgagct gaccaagaac 1440
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1500
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1560
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1620
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1680
tccctgtctc cgggtaaa 1698
<210> 278
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 278
Phe Asn Arg Gly Glu Cys Gly Gly Gly Ser
1 5 10
<210> 279
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 279
Phe Asn Arg Gly Glu Cys Leu Gln Val Tyr Tyr Arg Met
1 5 10
<210> 280
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 280
Leu Glu Gly Glu Glu Gly
1 5
<210> 281
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 281
Leu Glu Gly Glu Glu Gly Cys
1 5
<210> 282
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 282
Leu Glu Ile Lys
1
<210> 283
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 283
Leu Gly Glu Glu Gly
1 5
<210> 284
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 284
Leu Gly Glu Glu Gly Cys
1 5
<210> 285
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 285
Leu Gly Gly Cys Gly Gly Gly Ser
1 5
<210> 286
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 286
Leu Gly Lys Lys Gly
1 5
<210> 287
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 287
Leu Gly Lys Lys Gly Cys
1 5
<210> 288
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 288
Leu Lys Gly Lys Lys Gly
1 5
<210> 289
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 289
Leu Lys Gly Lys Lys Gly Cys
1 5
<210> 290
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 290
Leu Gln Val Tyr Tyr Arg Met
1 5
<210> 291
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 291
Leu Gln Val Tyr Tyr Arg Met Cys
1 5
<210> 292
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 292
Thr Val Ser Ser
1
<210> 293
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 293
Val Glu Pro Lys Ser Cys Gly Gly Gly Ser
1 5 10
<210> 294
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 294
Val Glu Pro Lys Ser Cys Tyr Leu Tyr Leu Arg Ala Arg Val
1 5 10
<210> 295
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 295
Val Gln Val His Tyr Arg Met
1 5
<210> 296
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 296
Val Gln Val His Tyr Arg Met Cys
1 5
<210> 297
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 297
Tyr Leu Tyr Leu Arg Ala Arg Val
1 5
<210> 298
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 298
Tyr Leu Tyr Leu Arg Ala Arg Val Cys
1 5
<210> 299
<211> 28
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер Е-спирали
<400> 299
Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
1 5 10 15
Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys
20 25
<210> 300
<211> 28
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер K-спирали
<400> 300
Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val
1 5 10 15
Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
20 25
<210> 301
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 301
Gly Gly Gly Asn Ser
1 5
<210> 302
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельная область легкой цепи YA2B6
<400> 302
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 303
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельная область тяжелой цепи YA2B6
<400> 303
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Asp Pro Ser Asp Thr Tyr Pro Asn Tyr Asn Lys Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Gly Asp Ser Asp Tyr Tyr Ser Gly Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 304
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий домен
<400> 304
Leu Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly
1 5 10 15
Val Ser Asp Tyr Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu
20 25 30
Gly Val Lys Ala Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro
35 40 45
<210> 305
<211> 279
<212> Белок
<213> Искусственная последовательность
<220>
<223> h3G8VL1-G3SG4-h2B6VH4-Kcoil-GGGNS
<400> 305
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala
115 120 125
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
130 135 140
Gly Tyr Thr Phe Thr Asn Tyr Trp Ile His Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Gln Gly Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Thr Tyr
165 170 175
Pro Asn Tyr Asn Lys Lys Phe Lys Gly Arg Val Thr Met Thr Val Val
180 185 190
Val Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp
195 200 205
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Gly Asp Ser Asp Tyr Tyr
210 215 220
Ser Gly Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Gly Gly Cys Gly Gly Gly Lys Val Ala Ala Leu Lys Glu Lys Val Ala
245 250 255
Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu
260 265 270
Lys Glu Gly Gly Gly Asn Ser
275
<210> 306
<211> 837
<212> ДНК
<213> Искусственная последовательность
<220>
<223> h3G8VL1-G3SG4-h2B6VH4-Kcoil-GGGNS
<400> 306
gacatcgtga tgacccaatc tccagactct ttggctgtgt ctctagggga gagggccacc 60
atcaactgca aggccagcca aagtgttgat tttgatggtg atagttttat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctata ctacatccaa tctagaatct 180
ggggtcccag acaggtttag tggcagtggg tctgggacag acttcaccct caccatcagc 240
agcctgcagg ctgaggatgt ggcagtttat tactgtcagc aaagtaatga agatccgtac 300
acgttcggac aggggaccaa gcttgagatc aaaggaggcg gatccggcgg cggaggccag 360
gttcagctgg tgcagtctgg agctgaggtg aagaagcctg gggcctcagt gaaggtctcc 420
tgcaaggctt ctggttacac ctttaccaac tactggatac actgggtgcg acaggcccct 480
ggacaagggc ttgagtggat tggagtgatt gatccttctg atacttatcc aaattacaat 540
aaaaagttca agggcagagt caccatgacc gtagtcgtat ccacgagcac agcctacatg 600
gagctgagga gcctgagatc tgacgacacg gccgtgtatt actgtgcgag aaacggtgat 660
tccgattatt actctggtat ggactactgg gggcaaggga ccacggtcac cgtctcctcc 720
ggaggatgtg gcggtggaaa agtggccgca ctgaaggaga aagttgctgc tttgaaagag 780
aaggtcgccg cacttaagga aaaggtcgca gccctgaaag agggcggcgg gaattct 837
<210> 307
<211> 318
<212> Белок
<213> Искусственная последовательность
<220>
<223> h2B6VL5-G3SG4-h3G8VH5-Ecoil-GGGNS-ABD
<400> 307
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Phe Thr Cys Arg Thr Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Glu Val Ser Glu Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys
115 120 125
Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu
130 135 140
Ser Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys
145 150 155 160
Ala Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr
165 170 175
Asn Pro Ala Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys
180 185 190
Asn Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala
195 200 205
Thr Tyr Tyr Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly
210 215 220
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Cys Gly Gly Gly Glu
225 230 235 240
Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val Ala
245 250 255
Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Gly Gly Gly Asn Ser
260 265 270
Leu Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly
275 280 285
Val Ser Asp Tyr Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu
290 295 300
Gly Val Lys Ala Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro
305 310 315
<210> 308
<211> 954
<212> ДНК
<213> Искусственная последовательность
<220>
<223> h2B6VL5-G3SG4-h3G8VH5-Ecoil-GGGNS-ABD
<400> 308
gaaattgtgc tgactcagtc tccagacttt cagtctgtga ctccaaagga gaaagtcacc 60
ttcacctgca ggaccagtca gagcattggc acaaacatac actggtacca gcagaaacca 120
gatcagtctc caaagctcct catcaaggag gtttctgagt ctatctctgg agtcccatcg 180
aggttcagtg gcagtggatc tgggacagat ttcaccctca ccatcaatag cctggaagct 240
gaagatgctg caacgtatta ctgtcaacaa agtaatacct ggccgttcac gttcggcgga 300
gggaccaagg tggagatcaa aggaggcgga tccggcggcg gaggccaggt taccctgaga 360
gagtctggcc ctgcgctggt gaagcccaca cagaccctca cactgacttg taccttctct 420
gggttttcac tgagcacttc tggtatgggt gtaggctgga ttcgtcagcc tcccgggaag 480
gctctagagt ggctggcaca catttggtgg gatgatgaca agcgctataa tccagccctg 540
aagagccgac tgacaatctc caaggatacc tccaaaaacc aggtagtcct cacaatgacc 600
aacatggacc ctgtggatac tgccacatac tactgtgctc aaataaaccc cgcctggttt 660
gcttactggg gccaagggac tctggtcact gtgagctccg gaggatgtgg cggtggagaa 720
gtggccgcac tggagaaaga ggttgctgct ttggagaagg aggtcgctgc acttgaaaag 780
gaggtcgcag ccctggagaa aggcggcggg aattctctgg ccgaagcaaa agtgctggcc 840
aaccgcgaac tggataaata tggcgtgagc gattattata agaacctgat taacaacgca 900
aagaccgtgg aaggcgtgaa agcactgatt gatgaaattc tggccgccct gcct 954
<210> 309
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 309
Ala Pro Ser Ser Ser
1 5
<210> 310
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 310
Val Pro Ser Met Gly Ser Ser Ser
1 5
<210> 311
<211> 383
<212> Белок
<213> Homo sapiens
<400> 311
Met Gly Leu Gly Pro Val Phe Leu Leu Leu Ala Gly Ile Phe Pro Phe
1 5 10 15
Ala Pro Pro Gly Ala Ala Ala Glu Pro His Ser Leu Arg Tyr Asn Leu
20 25 30
Thr Val Leu Ser Trp Asp Gly Ser Val Gln Ser Gly Phe Leu Thr Glu
35 40 45
Val His Leu Asp Gly Gln Pro Phe Leu Arg Cys Asp Arg Gln Lys Cys
50 55 60
Arg Ala Lys Pro Gln Gly Gln Trp Ala Glu Asp Val Leu Gly Asn Lys
65 70 75 80
Thr Trp Asp Arg Glu Thr Arg Asp Leu Thr Gly Asn Gly Lys Asp Leu
85 90 95
Arg Met Thr Leu Ala His Ile Lys Asp Gln Lys Glu Gly Leu His Ser
100 105 110
Leu Gln Glu Ile Arg Val Cys Glu Ile His Glu Asp Asn Ser Thr Arg
115 120 125
Ser Ser Gln His Phe Tyr Tyr Asp Gly Glu Leu Phe Leu Ser Gln Asn
130 135 140
Leu Glu Thr Lys Glu Trp Thr Met Pro Gln Ser Ser Arg Ala Gln Thr
145 150 155 160
Leu Ala Met Asn Val Arg Asn Phe Leu Lys Glu Asp Ala Met Lys Thr
165 170 175
Lys Thr His Tyr His Ala Met His Ala Asp Cys Leu Gln Glu Leu Arg
180 185 190
Arg Tyr Leu Lys Ser Gly Val Val Leu Arg Arg Thr Val Pro Pro Met
195 200 205
Val Asn Val Thr Arg Ser Glu Ala Ser Glu Gly Asn Ile Thr Val Thr
210 215 220
Cys Arg Ala Ser Gly Phe Tyr Pro Trp Asn Ile Thr Leu Ser Trp Arg
225 230 235 240
Gln Asp Gly Val Ser Leu Ser His Asp Thr Gln Gln Trp Gly Asp Val
245 250 255
Leu Pro Asp Gly Asn Gly Thr Tyr Gln Thr Trp Val Ala Thr Arg Ile
260 265 270
Cys Gln Gly Glu Glu Gln Arg Phe Thr Cys Tyr Met Glu His Ser Gly
275 280 285
Asn His Ser Thr His Pro Val Pro Ser Gly Lys Val Leu Val Leu Gln
290 295 300
Ser His Trp Gln Thr Phe His Val Ser Ala Val Ala Ala Ala Ala Ile
305 310 315 320
Phe Val Ile Ile Ile Phe Tyr Val Arg Cys Cys Lys Lys Lys Thr Ser
325 330 335
Ala Ala Glu Gly Pro Glu Leu Val Ser Leu Gln Val Leu Asp Gln His
340 345 350
Pro Val Gly Thr Ser Asp His Arg Asp Ala Thr Gln Leu Gly Phe Gln
355 360 365
Pro Leu Met Ser Asp Leu Gly Ser Thr Gly Ser Thr Glu Gly Ala
370 375 380
<210> 312
<211> 305
<212> Белок
<213> Homo sapiens
<400> 312
Pro His Ser Leu Arg Tyr Asn Leu Met Val Leu Ser Gln Asp Gly Ser
1 5 10 15
Val Gln Ser Gly Phe Leu Ala Glu Gly His Leu Asp Gly Gln Pro Phe
20 25 30
Leu Arg Tyr Asp Arg Gln Lys Arg Arg Ala Lys Pro Gln Gly Gln Trp
35 40 45
Ala Glu Asp Val Leu Gly Ala Lys Thr Trp Asp Thr Glu Thr Glu Asp
50 55 60
Leu Thr Glu Asn Gly Gln Asp Leu Arg Arg Thr Leu Thr His Ile Lys
65 70 75 80
Asp Gln Lys Gly Gly Leu His Ser Leu Gln Glu Ile Arg Val Cys Glu
85 90 95
Ile His Glu Asp Ser Ser Thr Arg Gly Ser Arg His Phe Tyr Tyr Asp
100 105 110
Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu Thr Gln Glu Ser Thr Val
115 120 125
Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala Met Asn Val Thr Asn Phe
130 135 140
Trp Lys Glu Asp Ala Met Lys Thr Lys Thr His Tyr Arg Ala Met Gln
145 150 155 160
Ala Asp Cys Leu Gln Lys Leu Gln Leu Pro Pro Met Val Asn Val Ile
165 170 175
Cys Ser Glu Val Ser Glu Gly Asn Ile Thr Val Thr Cys Arg Ala Ser
180 185 190
Ser Phe Tyr Pro Arg Asn Ile Thr Leu Thr Trp Arg Gln Asp Gly Val
195 200 205
Ser Leu Ser His Asn Thr Gln Gln Trp Gly Asp Val Leu Pro Asp Gly
210 215 220
Asn Gly Thr Tyr Gln Thr Trp Val Ala Thr Arg Ile Arg Gln Gly Glu
225 230 235 240
Glu Gln Arg Phe Thr Cys Tyr Met Glu His Ser Gly Asn His Gly Thr
245 250 255
His Pro Val Pro Ser Gly Lys Ala Leu Val Leu Gln Ser Gln Arg Thr
260 265 270
Asp Phe Pro Tyr Val Ser Ala Ala Met Pro Cys Phe Val Ile Ile Ile
275 280 285
Ile Leu Cys Val Pro Cys Cys Lys Lys Lys Thr Ser Ala Ala Glu Gly
290 295 300
Pro
305
<210> 313
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 313
Gly Phe Asn Arg Gly Glu Cys
1 5
<210> 314
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 314
Gly Val Glu Pro Lys Ser Cys
1 5
<210> 315
<211> 241
<212> Белок
<213> Искусственная последовательность
<220>
<223> TCRVL-HER2VH
<400> 315
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Thr Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Ser Gly Gly
100 105 110
Gly Gly Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro
115 120 125
Gly Ala Ser Leu Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile Lys
130 135 140
Asp Thr Tyr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu
145 150 155 160
Trp Ile Gly Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Asp Pro
165 170 175
Lys Phe Gln Asp Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr
180 185 190
Ala Tyr Leu Gln Val Ser Arg Leu Thr Ser Glu Asp Thr Ala Val Tyr
195 200 205
Tyr Cys Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp
210 215 220
Gly Gln Gly Ala Ser Val Thr Val Ser Ser Gly Phe Asn Arg Gly Glu
225 230 235 240
Cys
<210> 316
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TCRVL-HER2VH
<400> 316
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gtgccacctc aagtgtaagt tacatgcact ggtatcagca gaaaccaggg 120
aaagccccta agcgctggat ctatgacaca tccaaactgg cttctggggt cccatcaagg 180
ttcagcggca gtggatctgg gacagaattt actctcacaa tcagcagcct gcagcctgaa 240
gattttgcaa cttattactg tcagcagtgg agtagtaacc cgctcacgtt tggccagggg 300
accaagcttg agatcaaagg aggcggatcc ggcggcggag gccaggttca gctgcagcag 360
tctgggccag agcttgtgaa gccaggggcc tcactcaagt tgtcctgtac agcttctggc 420
ttcaacatta aagacaccta tatacactgg gtgaaacaga ggcctgaaca gggcctggaa 480
tggattggaa ggatttatcc tacgaatggt tatactagat atgacccgaa gttccaggac 540
aaggccacta taacagcaga cacatcctcc aacacagcct acctgcaggt cagccgcctg 600
acatctgagg acactgccgt ctattattgt tctagatggg gaggggacgg cttctatgct 660
atggactact ggggtcaagg agcctcggtc accgtgagct ccggattcaa caggggagag 720
tgt 723
<210> 317
<211> 242
<212> Белок
<213> Искусственная последовательность
<220>
<223> HER2VL-TCRVH
<400> 317
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly His Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Lys Phe
130 135 140
Thr Ser Tyr Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Tyr Ile Asn Pro Tyr Asn Asp Val Thr Lys Tyr Asn
165 170 175
Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
195 200 205
His Tyr Cys Ala Arg Gly Ser Tyr Tyr Asp Tyr Asp Gly Phe Val Tyr
210 215 220
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Val Glu Pro Lys
225 230 235 240
Ser Cys
<210> 318
<211> 726
<212> ДНК
<213> Искусственная последовательность
<220>
<223> HER2VL-TCRVH
<400> 318
gacatcgtga tgacccagtc ccacaagttc atgtccacct ctgtgggcga tagggtcagc 60
atcacctgca aggccagcca ggatgtgaat actgctgtag cctggtatca gcagaaacca 120
ggacattctc ccaaactgct gatttactcc gcatccttcc ggtacactgg agtccctgat 180
cgcttcactg gcagcagatc tgggacagat ttcactttca ccatcagcag tgtgcaggct 240
gaagacctgg cagtttatta ctgtcagcaa cattatacta cacctcccac cttcggaggg 300
ggtaccaagg tggagatcaa aggaggcgga tccggcggcg gaggccaggt tcagctggtg 360
cagtctggag ctgaggtgaa gaagcctggg gcctcagtga aggtctcctg caaggccagc 420
ggttacaagt ttaccagcta cgtgatgcac tgggtgcgac aggcccctgg acaagggctt 480
gagtggatcg gatatattaa tccttacaat gatgttacta agtacaatga gaagttcaaa 540
ggcagagtca cgattaccgc ggacaaatcc acgagcacag cctacatgga gctgagcagc 600
ctgagatccg aggacacggc cgtgcactac tgtgcgagag ggagctacta tgattacgac 660
gggtttgttt actggggcca agggactctg gtcactgtga gctccggagt tgagcccaaa 720
tcttgt 726
<210> 319
<211> 273
<212> Белок
<213> Искусственная последовательность
<220>
<223> TCRVL-HER2VH-E-спираль
<400> 319
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Thr Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Ser Gly Gly
100 105 110
Gly Gly Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro
115 120 125
Gly Ala Ser Leu Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile Lys
130 135 140
Asp Thr Tyr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu
145 150 155 160
Trp Ile Gly Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Asp Pro
165 170 175
Lys Phe Gln Asp Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr
180 185 190
Ala Tyr Leu Gln Val Ser Arg Leu Thr Ser Glu Asp Thr Ala Val Tyr
195 200 205
Tyr Cys Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp
210 215 220
Gly Gln Gly Ala Ser Val Thr Val Ser Ser Gly Gly Cys Gly Gly Gly
225 230 235 240
Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
245 250 255
Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Gly Gly Gly Asn
260 265 270
Ser
<210> 320
<211> 819
<212> Белок
<213> Искусственная последовательность
<220>
<223> TCRVL-HER2VH-E-спираль
<400> 320
Gly Ala Ala Ala Thr Thr Gly Thr Gly Thr Thr Gly Ala Cys Ala Cys
1 5 10 15
Ala Gly Thr Cys Thr Cys Cys Ala Gly Cys Cys Ala Cys Cys Cys Thr
20 25 30
Gly Thr Cys Thr Thr Thr Gly Thr Cys Thr Cys Cys Ala Gly Gly Gly
35 40 45
Gly Ala Ala Ala Gly Ala Gly Cys Cys Ala Cys Cys Cys Thr Cys Thr
50 55 60
Cys Cys Thr Gly Cys Ala Gly Thr Gly Cys Cys Ala Cys Cys Thr Cys
65 70 75 80
Ala Ala Gly Thr Gly Thr Ala Ala Gly Thr Thr Ala Cys Ala Thr Gly
85 90 95
Cys Ala Cys Thr Gly Gly Thr Ala Thr Cys Ala Gly Cys Ala Gly Ala
100 105 110
Ala Ala Cys Cys Ala Gly Gly Gly Ala Ala Ala Gly Cys Cys Cys Cys
115 120 125
Thr Ala Ala Gly Cys Gly Cys Thr Gly Gly Ala Thr Cys Thr Ala Thr
130 135 140
Gly Ala Cys Ala Cys Ala Thr Cys Cys Ala Ala Ala Cys Thr Gly Gly
145 150 155 160
Cys Thr Thr Cys Thr Gly Gly Gly Gly Thr Cys Cys Cys Ala Thr Cys
165 170 175
Ala Ala Gly Gly Thr Thr Cys Ala Gly Cys Gly Gly Cys Ala Gly Thr
180 185 190
Gly Gly Ala Thr Cys Thr Gly Gly Gly Ala Cys Ala Gly Ala Ala Thr
195 200 205
Thr Thr Ala Cys Thr Cys Thr Cys Ala Cys Ala Ala Thr Cys Ala Gly
210 215 220
Cys Ala Gly Cys Cys Thr Gly Cys Ala Gly Cys Cys Thr Gly Ala Ala
225 230 235 240
Gly Ala Thr Thr Thr Thr Gly Cys Ala Ala Cys Thr Thr Ala Thr Thr
245 250 255
Ala Cys Thr Gly Thr Cys Ala Gly Cys Ala Gly Thr Gly Gly Ala Gly
260 265 270
Thr Ala Gly Thr Ala Ala Cys Cys Cys Gly Cys Thr Cys Ala Cys Gly
275 280 285
Thr Thr Thr Gly Gly Cys Cys Ala Gly Gly Gly Gly Ala Cys Cys Ala
290 295 300
Ala Gly Cys Thr Thr Gly Ala Gly Ala Thr Cys Ala Ala Ala Gly Gly
305 310 315 320
Ala Gly Gly Cys Gly Gly Ala Thr Cys Cys Gly Gly Cys Gly Gly Cys
325 330 335
Gly Gly Ala Gly Gly Cys Cys Ala Gly Gly Thr Thr Cys Ala Gly Cys
340 345 350
Thr Gly Cys Ala Gly Cys Ala Gly Thr Cys Thr Gly Gly Gly Cys Cys
355 360 365
Ala Gly Ala Gly Cys Thr Thr Gly Thr Gly Ala Ala Gly Cys Cys Ala
370 375 380
Gly Gly Gly Gly Cys Cys Thr Cys Ala Cys Thr Cys Ala Ala Gly Thr
385 390 395 400
Thr Gly Thr Cys Cys Thr Gly Thr Ala Cys Ala Gly Cys Thr Thr Cys
405 410 415
Thr Gly Gly Cys Thr Thr Cys Ala Ala Cys Ala Thr Thr Ala Ala Ala
420 425 430
Gly Ala Cys Ala Cys Cys Thr Ala Thr Ala Thr Ala Cys Ala Cys Thr
435 440 445
Gly Gly Gly Thr Gly Ala Ala Ala Cys Ala Gly Ala Gly Gly Cys Cys
450 455 460
Thr Gly Ala Ala Cys Ala Gly Gly Gly Cys Cys Thr Gly Gly Ala Ala
465 470 475 480
Thr Gly Gly Ala Thr Thr Gly Gly Ala Ala Gly Gly Ala Thr Thr Thr
485 490 495
Ala Thr Cys Cys Thr Ala Cys Gly Ala Ala Thr Gly Gly Thr Thr Ala
500 505 510
Thr Ala Cys Thr Ala Gly Ala Thr Ala Thr Gly Ala Cys Cys Cys Gly
515 520 525
Ala Ala Gly Thr Thr Cys Cys Ala Gly Gly Ala Cys Ala Ala Gly Gly
530 535 540
Cys Cys Ala Cys Thr Ala Thr Ala Ala Cys Ala Gly Cys Ala Gly Ala
545 550 555 560
Cys Ala Cys Ala Thr Cys Cys Thr Cys Cys Ala Ala Cys Ala Cys Ala
565 570 575
Gly Cys Cys Thr Ala Cys Cys Thr Gly Cys Ala Gly Gly Thr Cys Ala
580 585 590
Gly Cys Cys Gly Cys Cys Thr Gly Ala Cys Ala Thr Cys Thr Gly Ala
595 600 605
Gly Gly Ala Cys Ala Cys Thr Gly Cys Cys Gly Thr Cys Thr Ala Thr
610 615 620
Thr Ala Thr Thr Gly Thr Thr Cys Thr Ala Gly Ala Thr Gly Gly Gly
625 630 635 640
Gly Ala Gly Gly Gly Gly Ala Cys Gly Gly Cys Thr Thr Cys Thr Ala
645 650 655
Thr Gly Cys Thr Ala Thr Gly Gly Ala Cys Thr Ala Cys Thr Gly Gly
660 665 670
Gly Gly Thr Cys Ala Ala Gly Gly Ala Gly Cys Cys Thr Cys Gly Gly
675 680 685
Thr Cys Ala Cys Cys Gly Thr Gly Ala Gly Cys Thr Cys Cys Gly Gly
690 695 700
Ala Gly Gly Ala Thr Gly Thr Gly Gly Cys Gly Gly Thr Gly Gly Ala
705 710 715 720
Gly Ala Ala Gly Thr Gly Gly Cys Cys Gly Cys Ala Cys Thr Gly Gly
725 730 735
Ala Gly Ala Ala Ala Gly Ala Gly Gly Thr Thr Gly Cys Thr Gly Cys
740 745 750
Thr Thr Thr Gly Gly Ala Gly Ala Ala Gly Gly Ala Gly Gly Thr Cys
755 760 765
Gly Cys Thr Gly Cys Ala Cys Thr Thr Gly Ala Ala Ala Ala Gly Gly
770 775 780
Ala Gly Gly Thr Cys Gly Cys Ala Gly Cys Cys Cys Thr Gly Gly Ala
785 790 795 800
Gly Ala Ala Ala Gly Gly Cys Gly Gly Cys Gly Gly Gly Ala Ala Thr
805 810 815
Thr Cys Thr
<210> 321
<211> 274
<212> Белок
<213> Искусственная последовательность
<220>
<223> HER2VL-TCRVH-K-спираль
<400> 321
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly His Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Lys Phe
130 135 140
Thr Ser Tyr Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Tyr Ile Asn Pro Tyr Asn Asp Val Thr Lys Tyr Asn
165 170 175
Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
195 200 205
His Tyr Cys Ala Arg Gly Ser Tyr Tyr Asp Tyr Asp Gly Phe Val Tyr
210 215 220
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Cys Gly Gly
225 230 235 240
Gly Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys
245 250 255
Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Gly Gly Gly
260 265 270
Asn Ser
<210> 322
<211> 822
<212> ДНК
<213> Искусственная последовательность
<220>
<223> HER2VL-TCRVH-K-спираль
<400> 322
gacatcgtga tgacccagtc ccacaagttc atgtccacct ctgtgggcga tagggtcagc 60
atcacctgca aggccagcca ggatgtgaat actgctgtag cctggtatca gcagaaacca 120
ggacattctc ccaaactgct gatttactcc gcatccttcc ggtacactgg agtccctgat 180
cgcttcactg gcagcagatc tgggacagat ttcactttca ccatcagcag tgtgcaggct 240
gaagacctgg cagtttatta ctgtcagcaa cattatacta cacctcccac cttcggaggg 300
ggtaccaagg tggagatcaa aggaggcgga tccggcggcg gaggccaggt tcagctggtg 360
cagtctggag ctgaggtgaa gaagcctggg gcctcagtga aggtctcctg caaggccagc 420
ggttacaagt ttaccagcta cgtgatgcac tgggtgcgac aggcccctgg acaagggctt 480
gagtggatcg gatatattaa tccttacaat gatgttacta agtacaatga gaagttcaaa 540
ggcagagtca cgattaccgc ggacaaatcc acgagcacag cctacatgga gctgagcagc 600
ctgagatccg aggacacggc cgtgcactac tgtgcgagag ggagctacta tgattacgac 660
gggtttgttt actggggcca agggactctg gtcactgtga gctccggagg atgtggcggt 720
ggaaaagtgg ccgcactgaa ggagaaagtt gctgctttga aagagaaggt cgccgcactt 780
aaggaaaagg tcgcagccct gaaagagggc ggcgggaatt ct 822
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| БИСПЕЦИФИЧЕСКИЕ 2+1 КОНТОРСТЕЛА | 2018 |
|
RU2797305C2 |
| ИММУНОКОНЪЮГАТЫ IL2 И МУТАНТНОГО TNF | 2017 |
|
RU2758139C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ ГЕПАТИТА В И СВЯЗАННЫХ СОСТОЯНИЙ | 2016 |
|
RU2733496C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ КОСТИМУЛЯТОРНОГО TNF-РЕЦЕПТОРА | 2016 |
|
RU2761115C1 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |

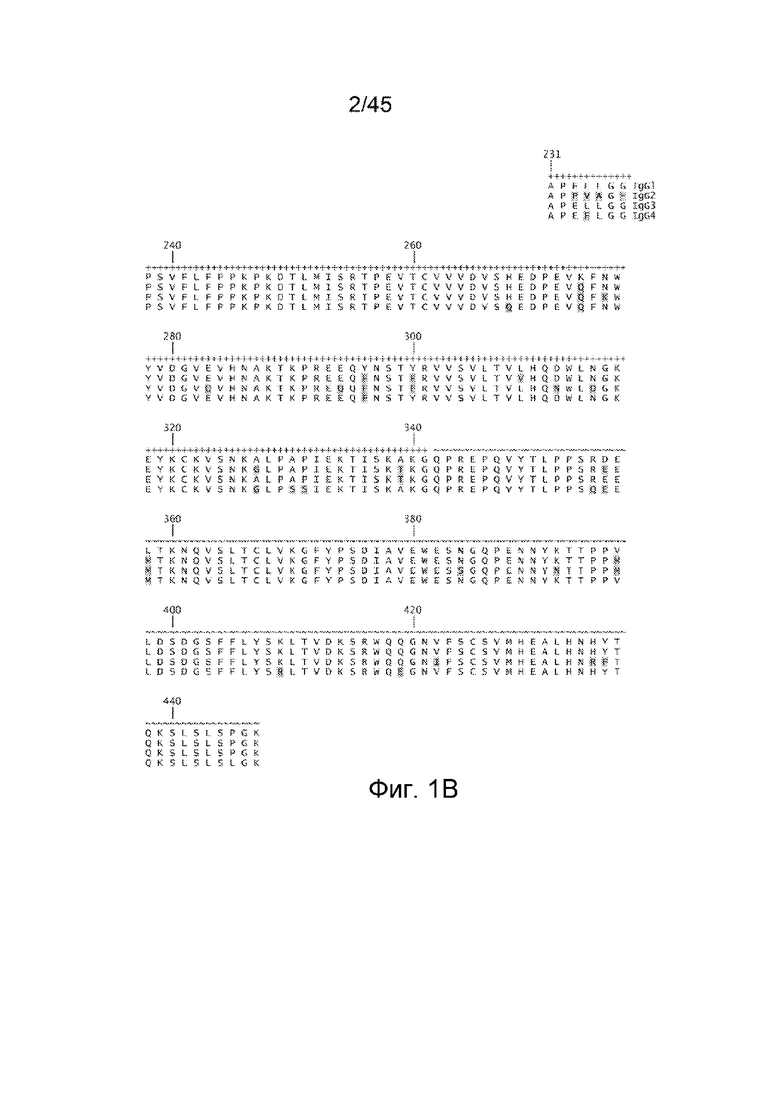
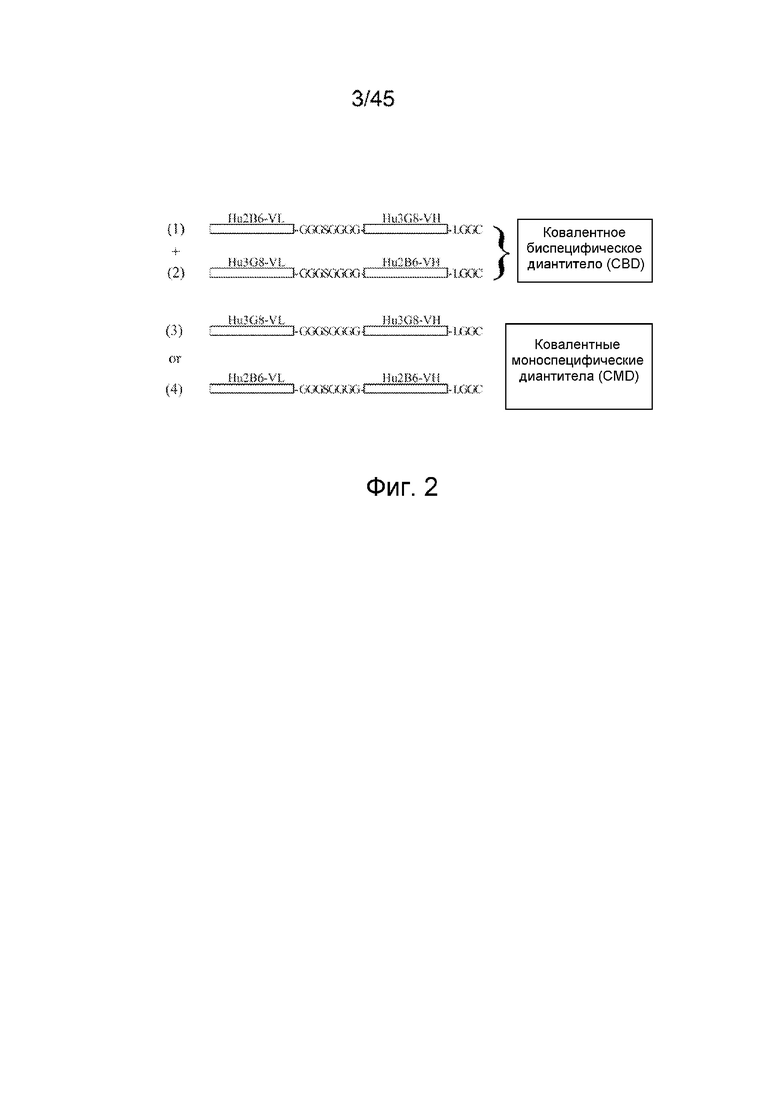

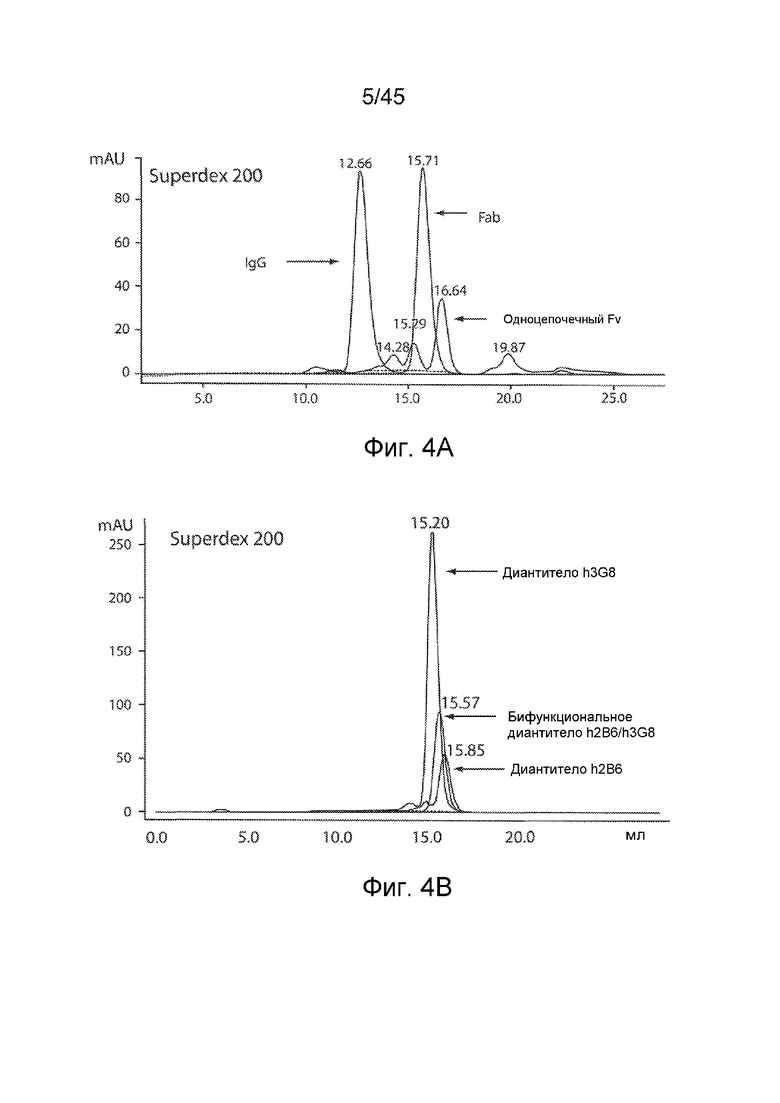

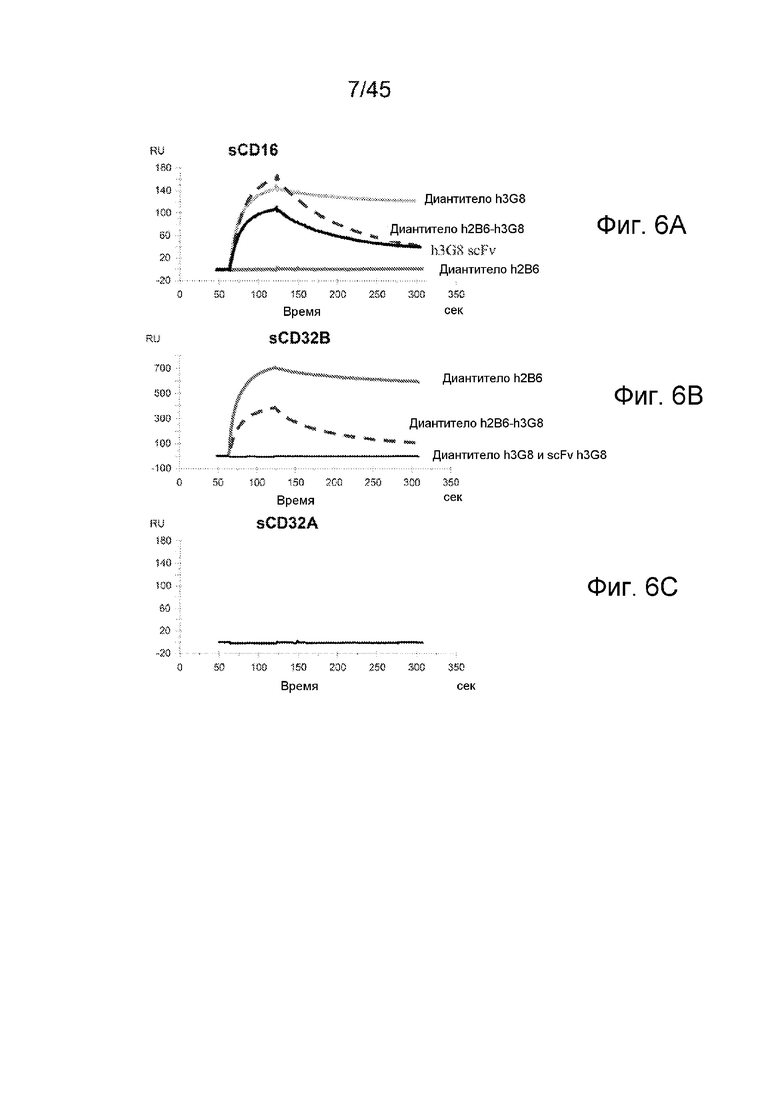
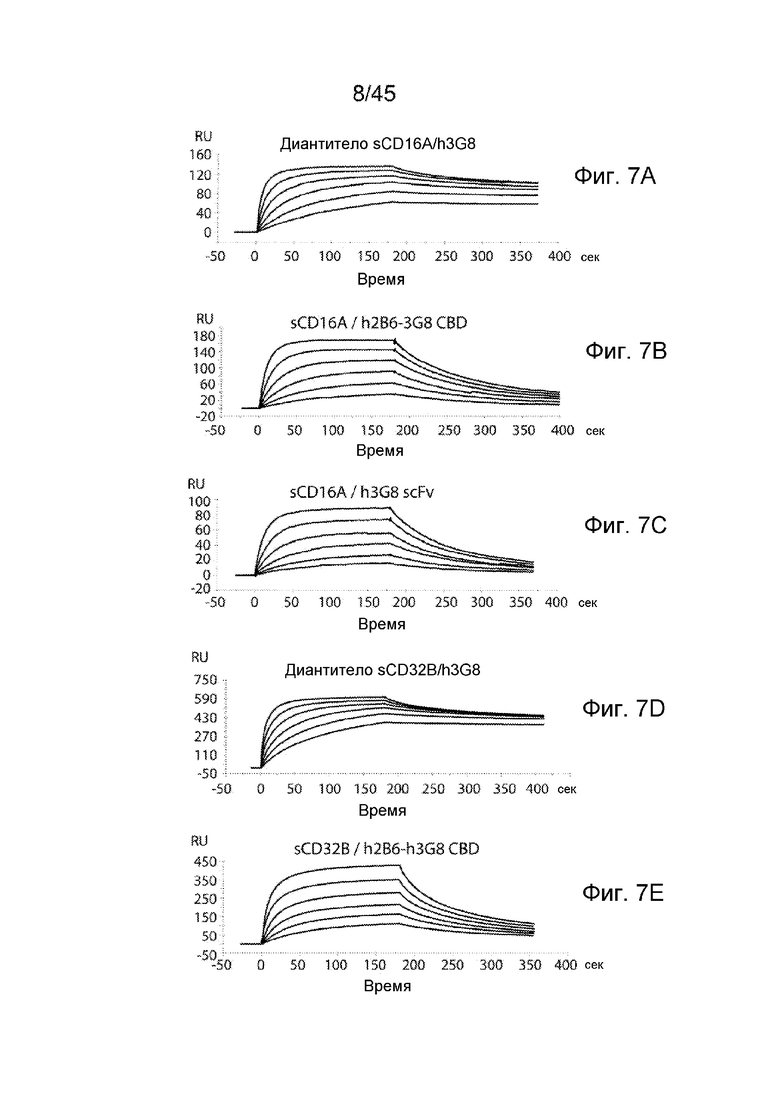
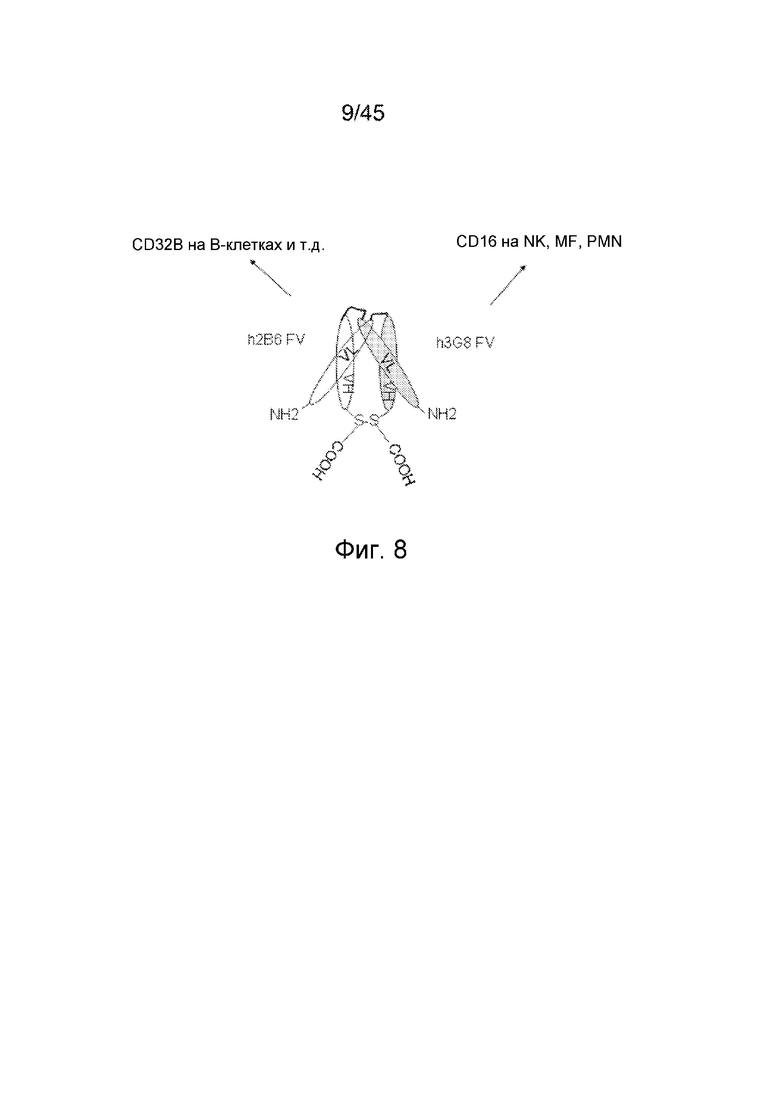


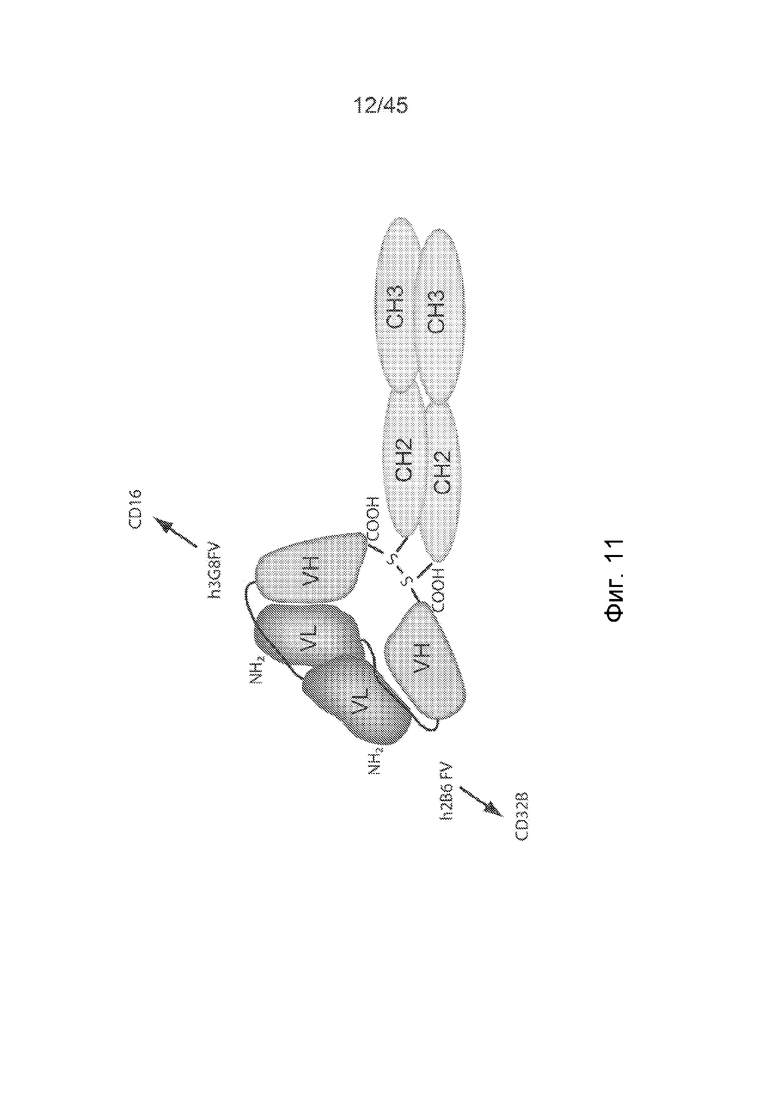
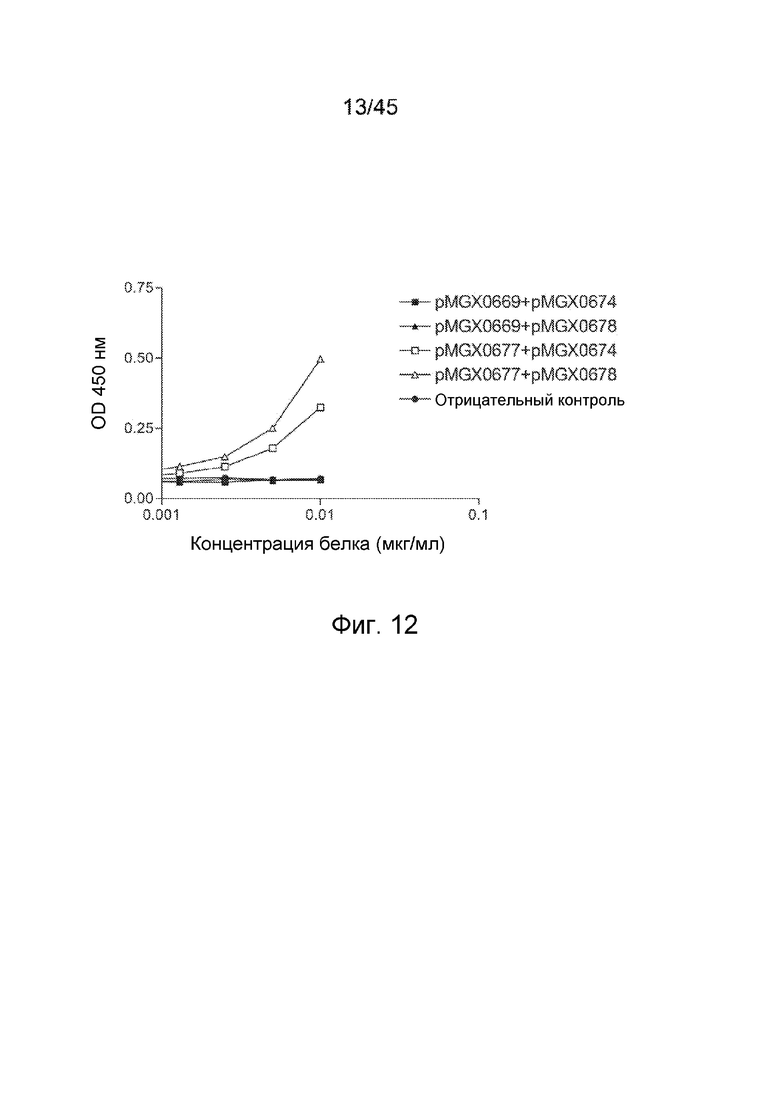

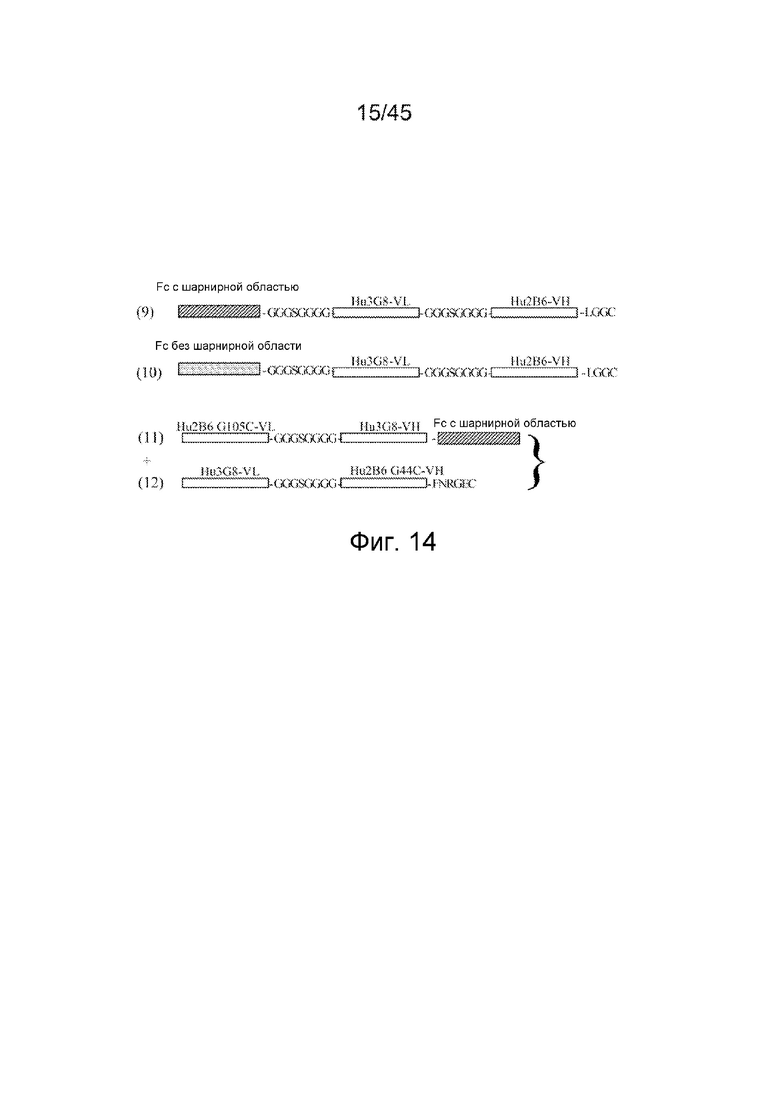
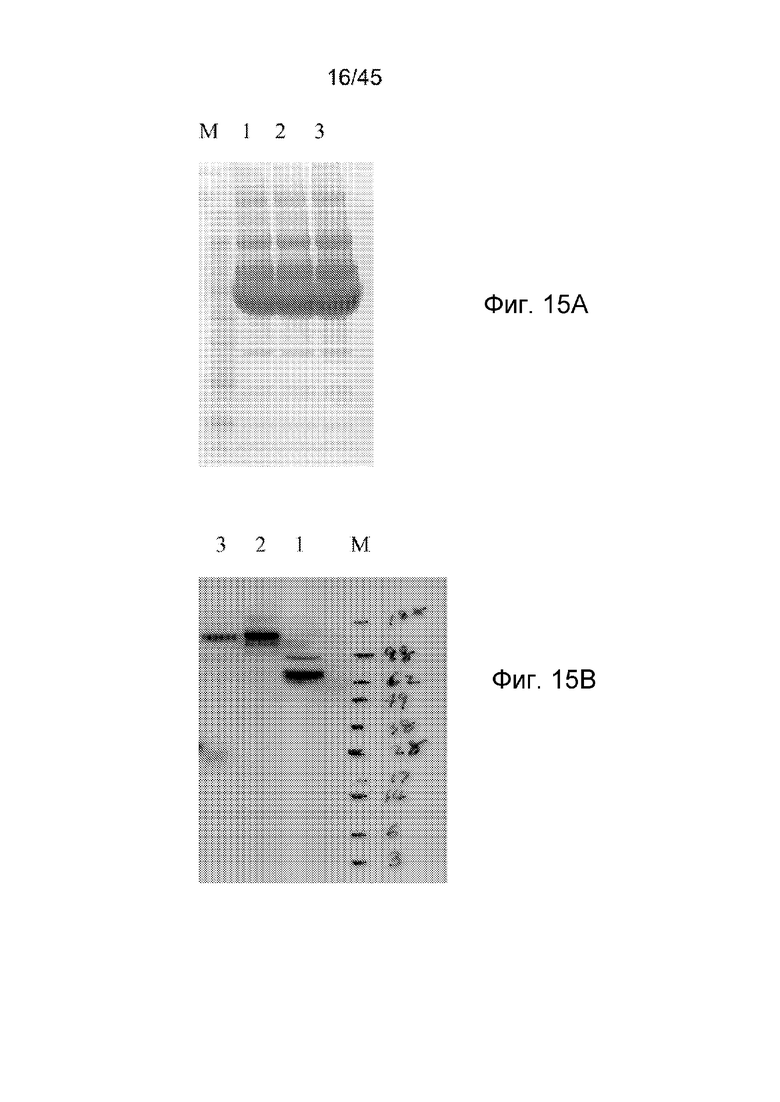





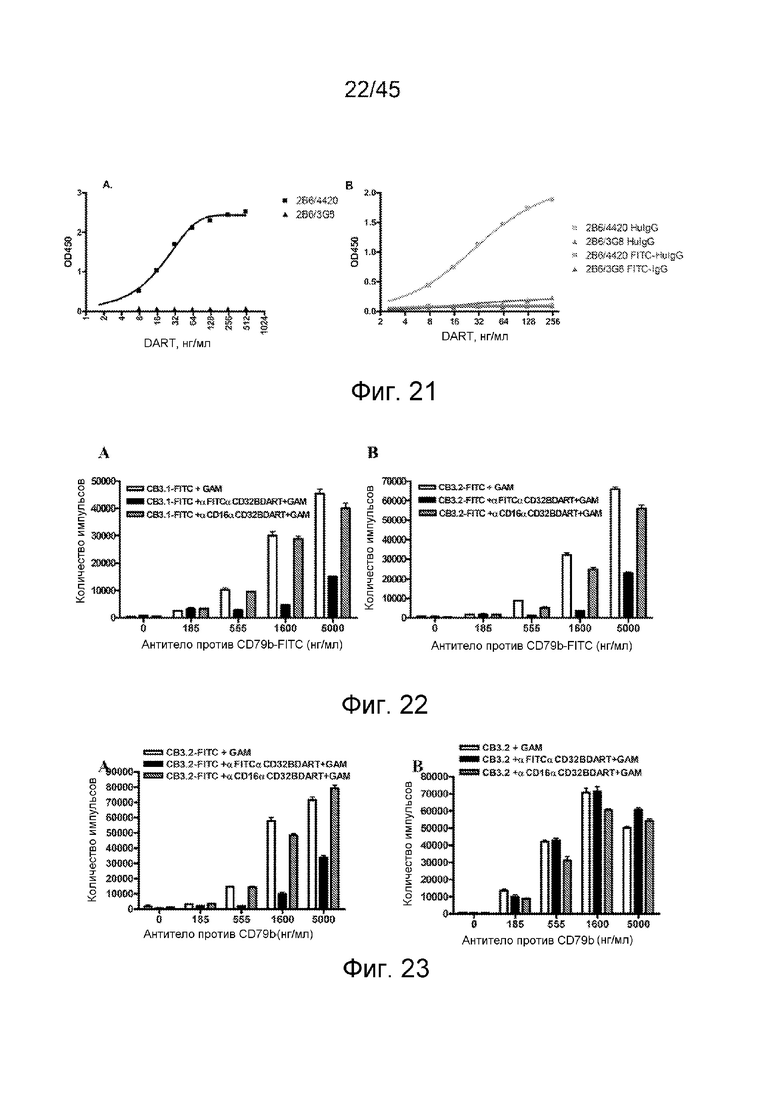
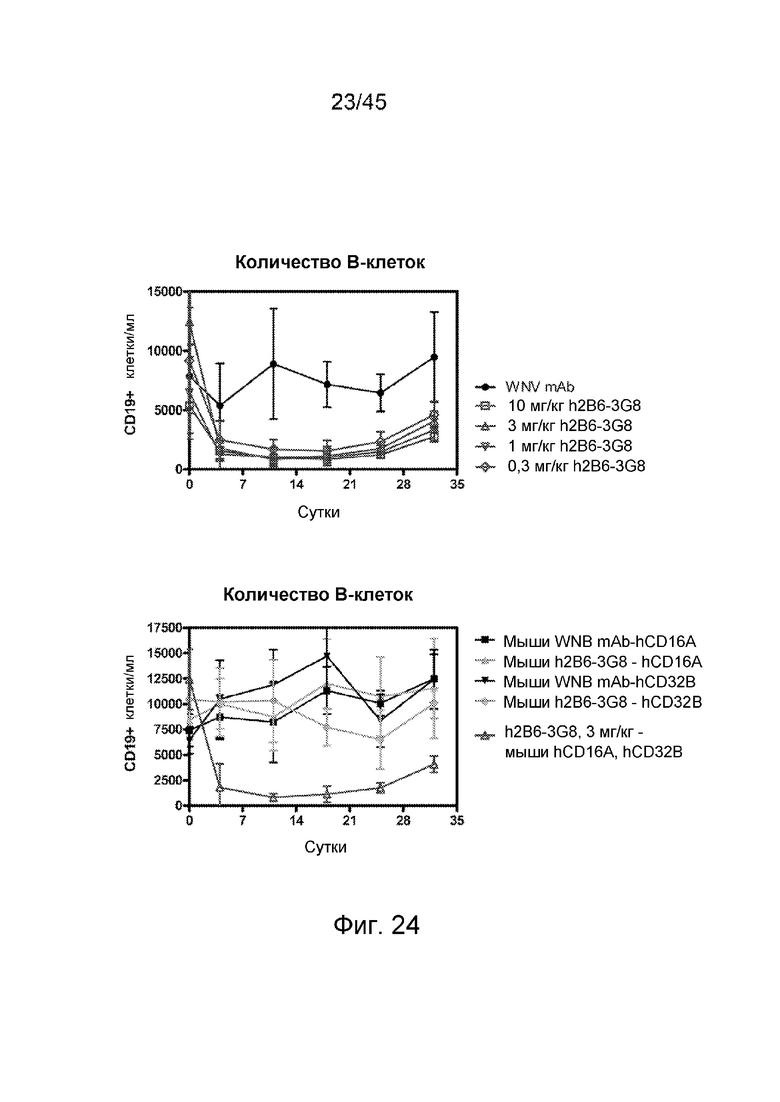
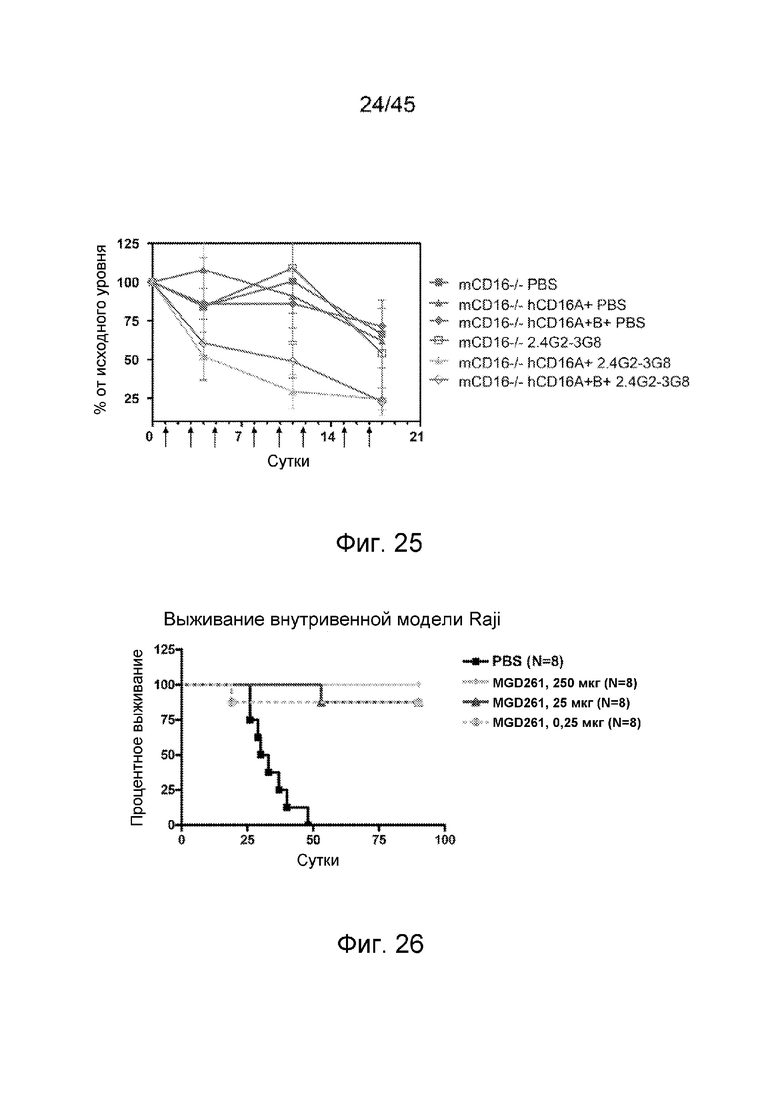
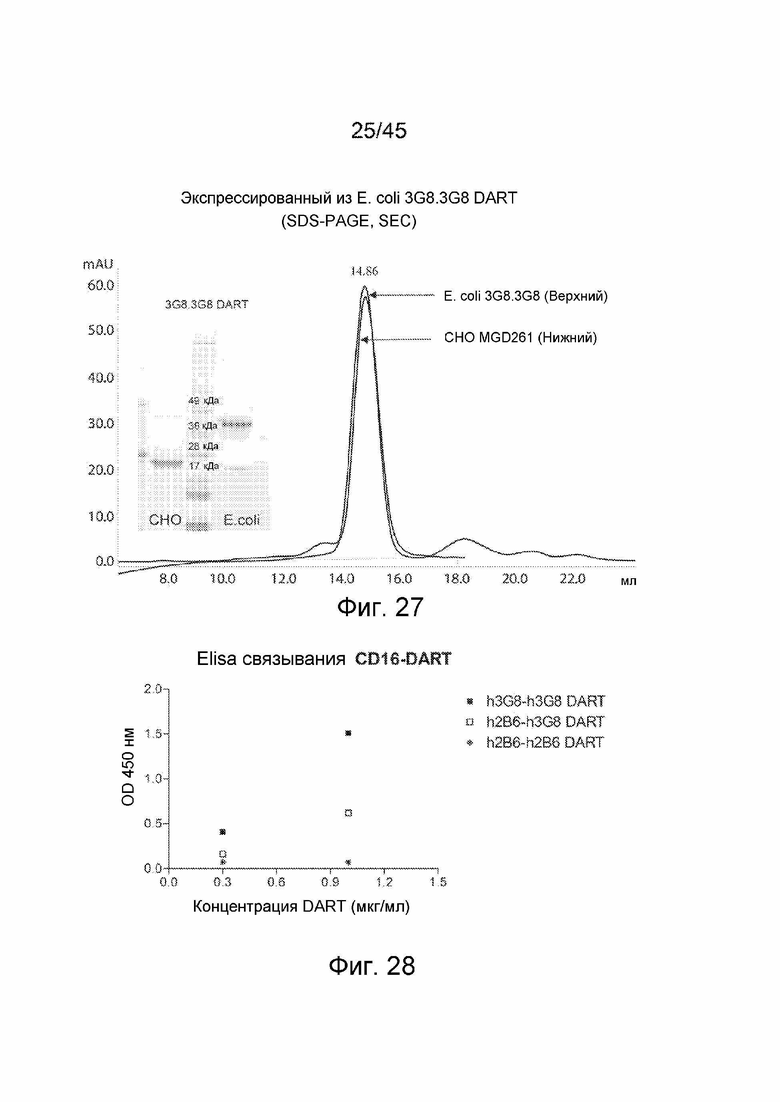

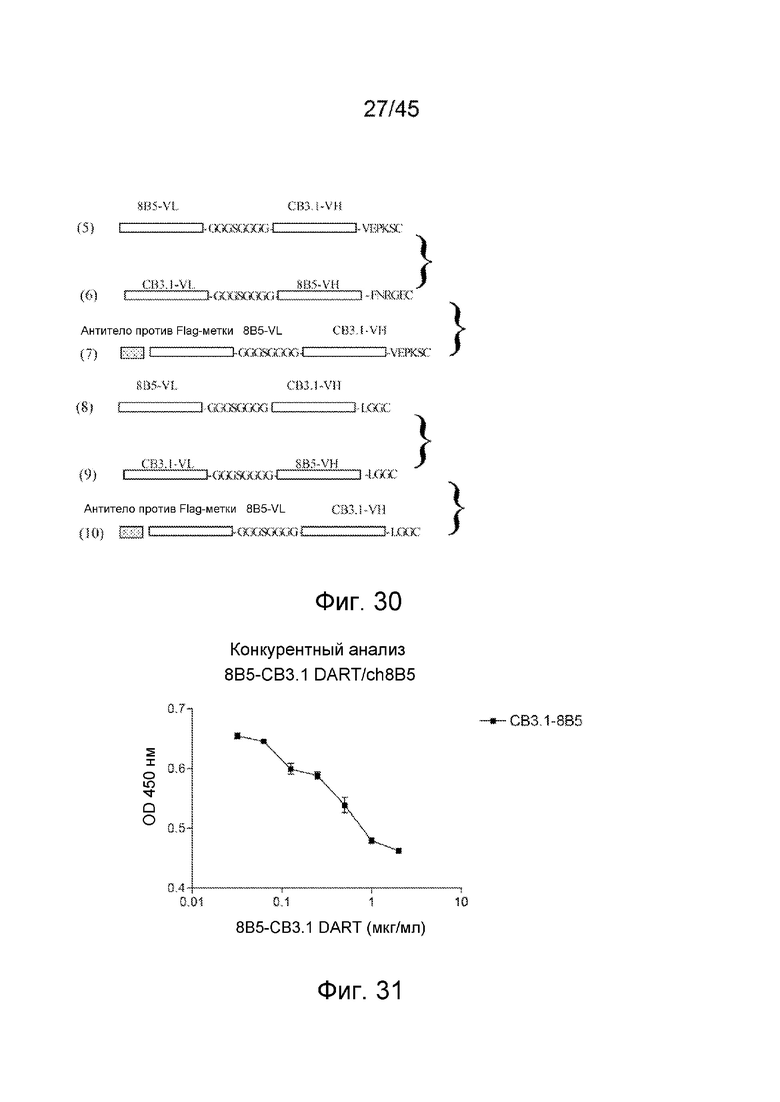
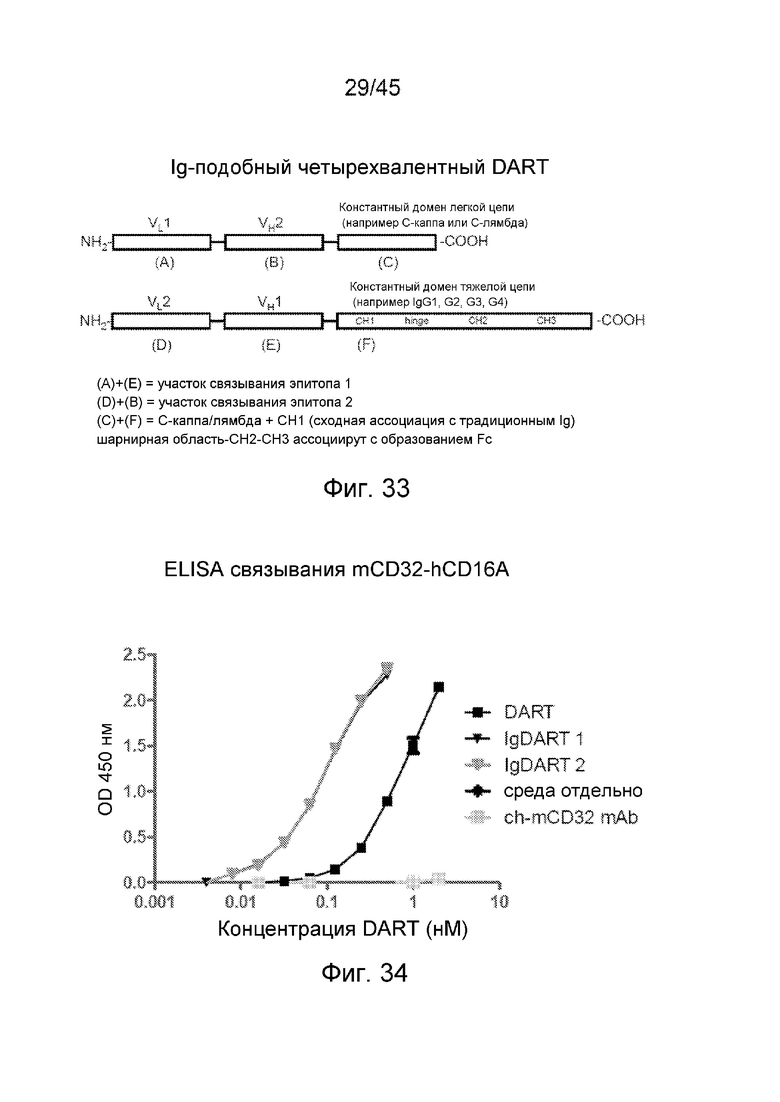
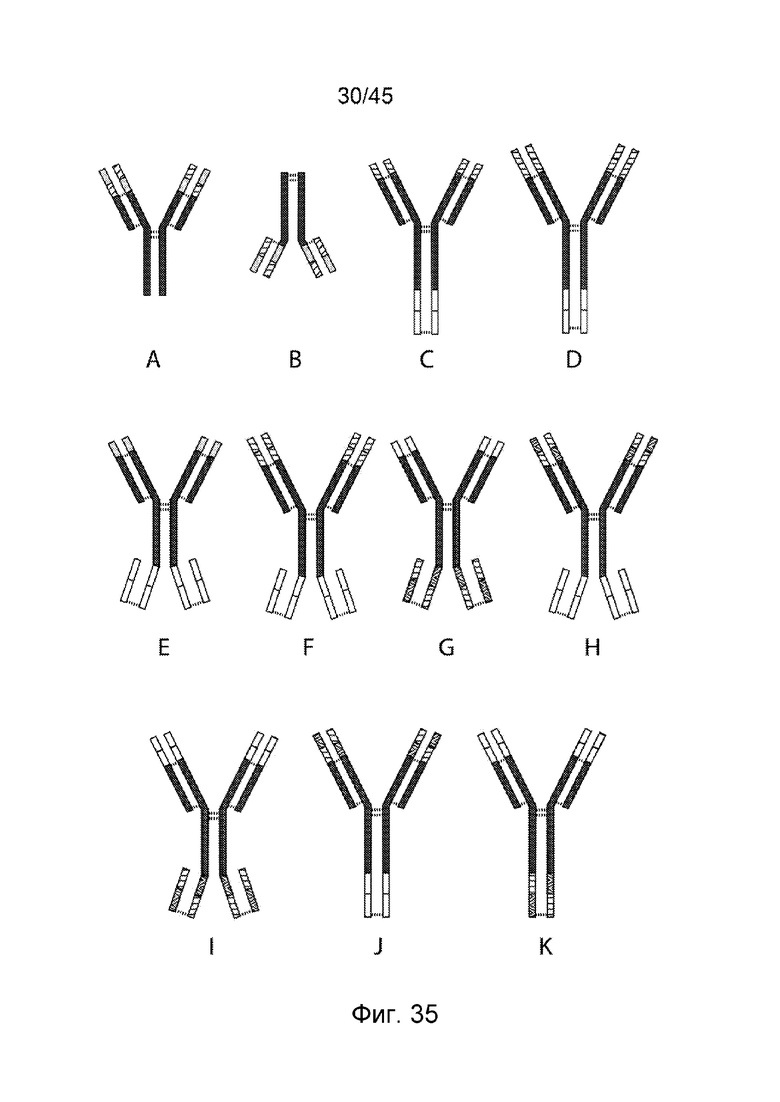
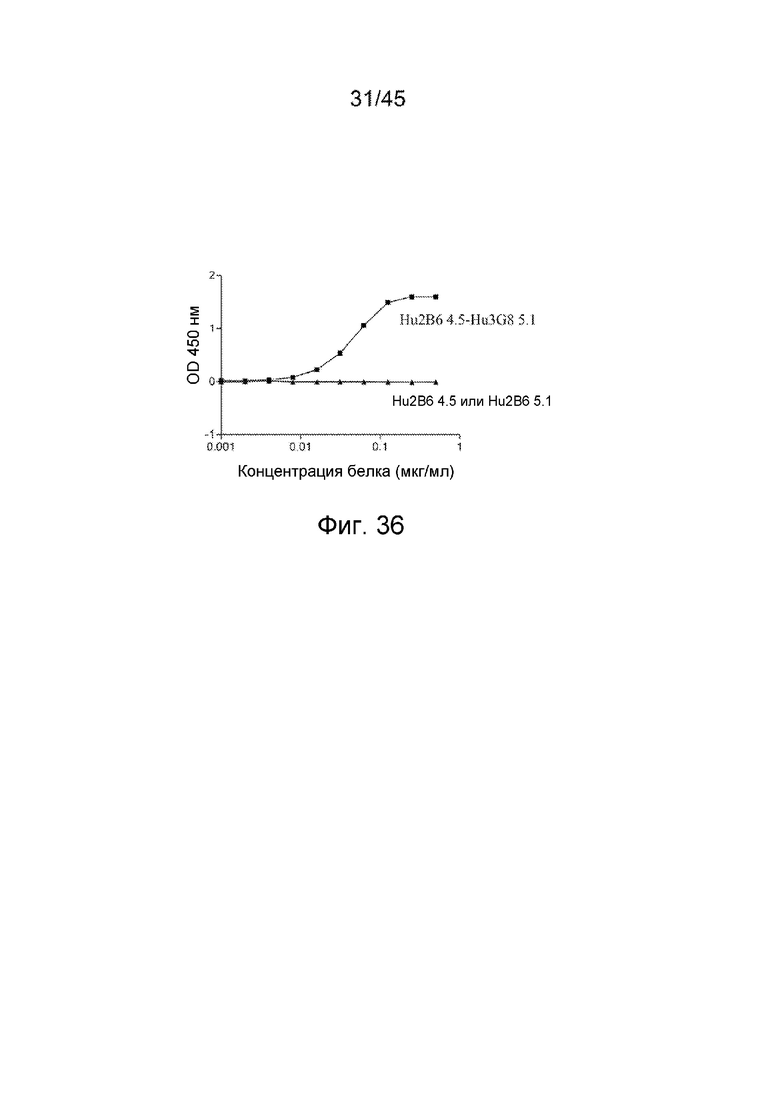
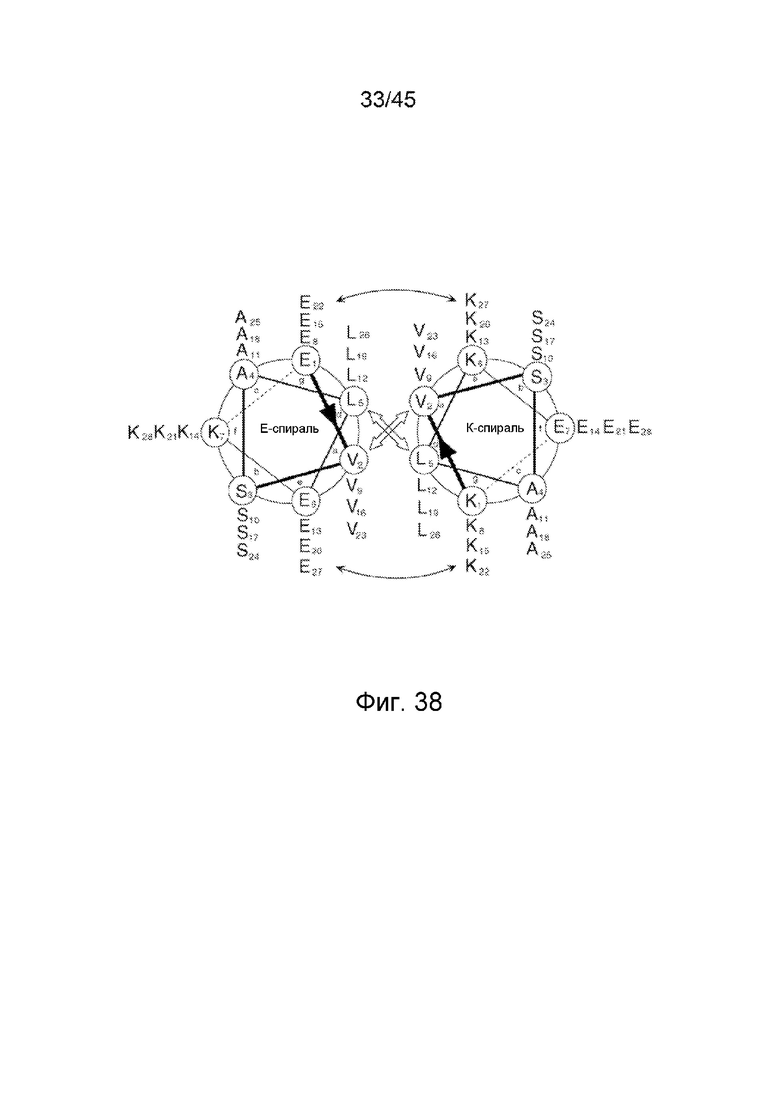

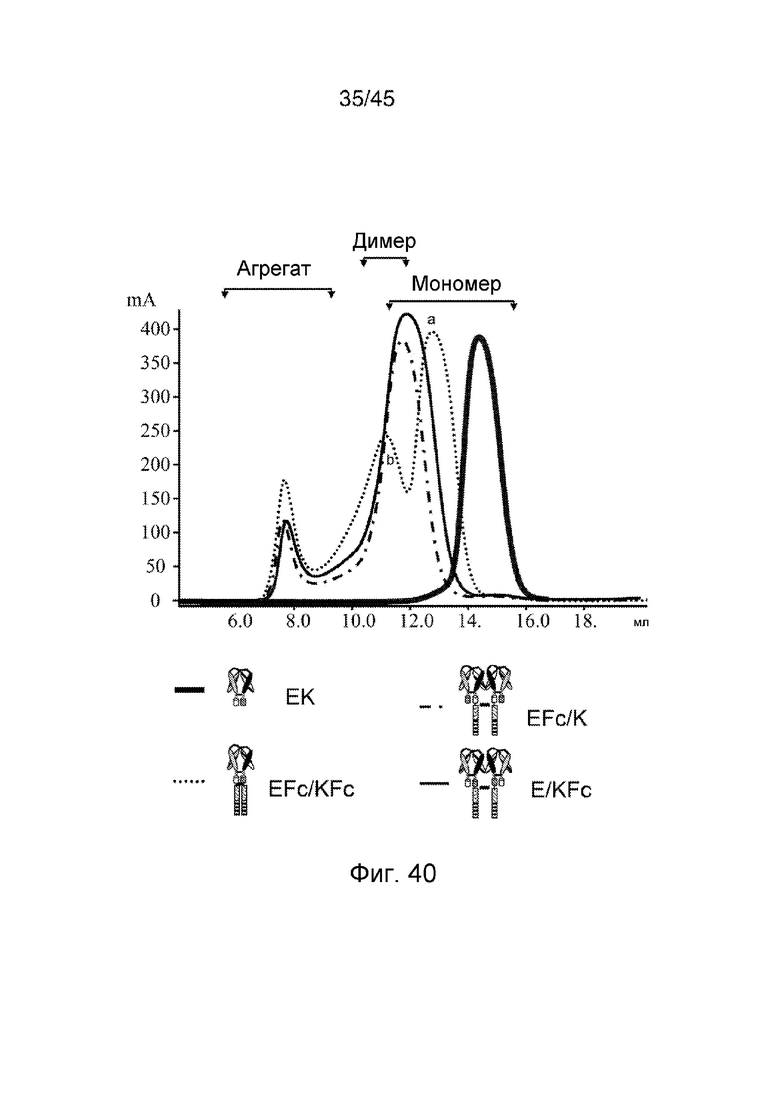
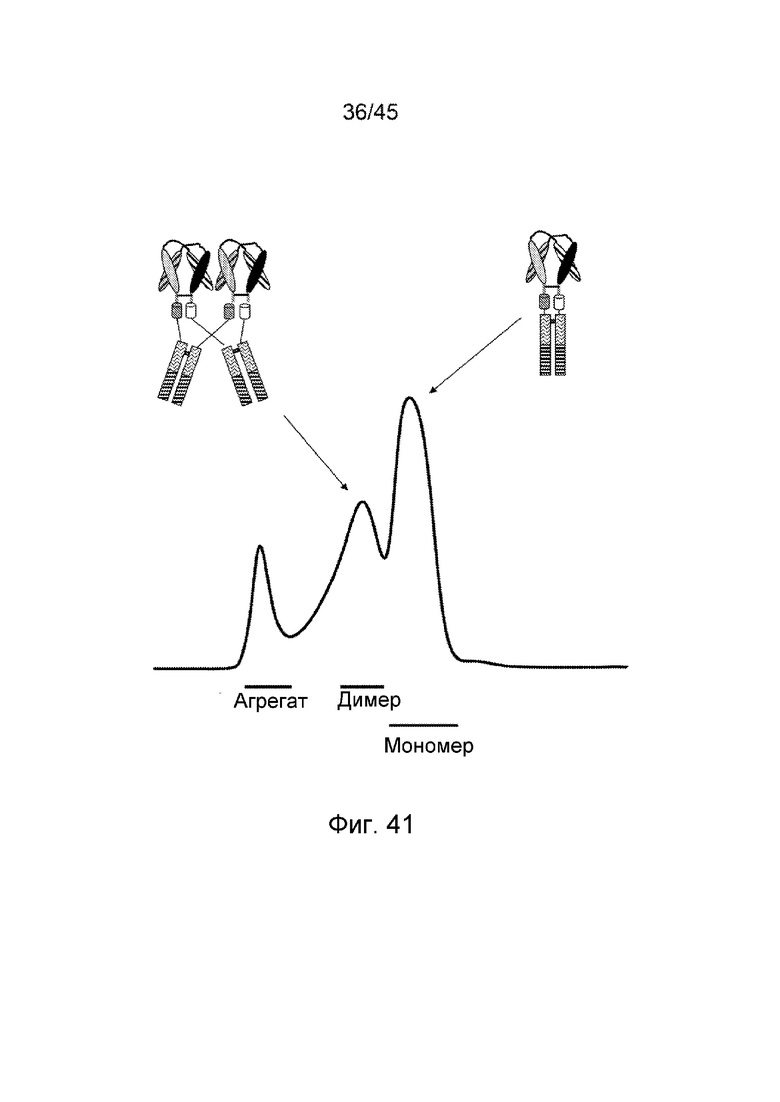
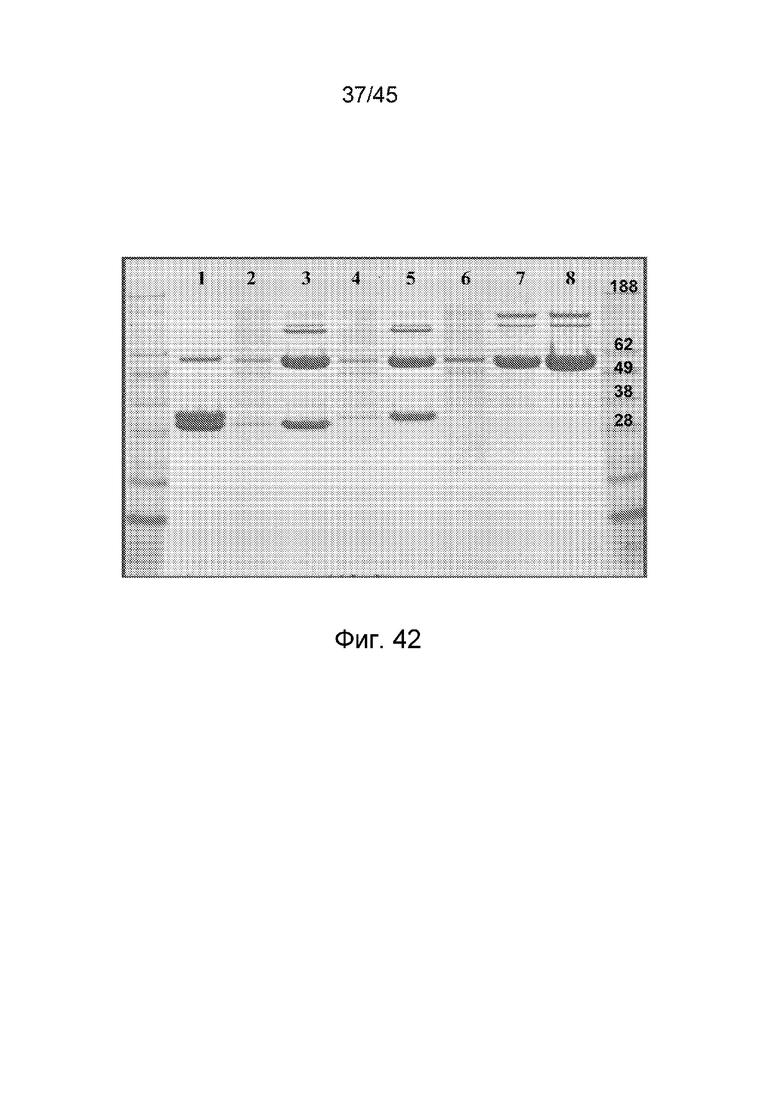
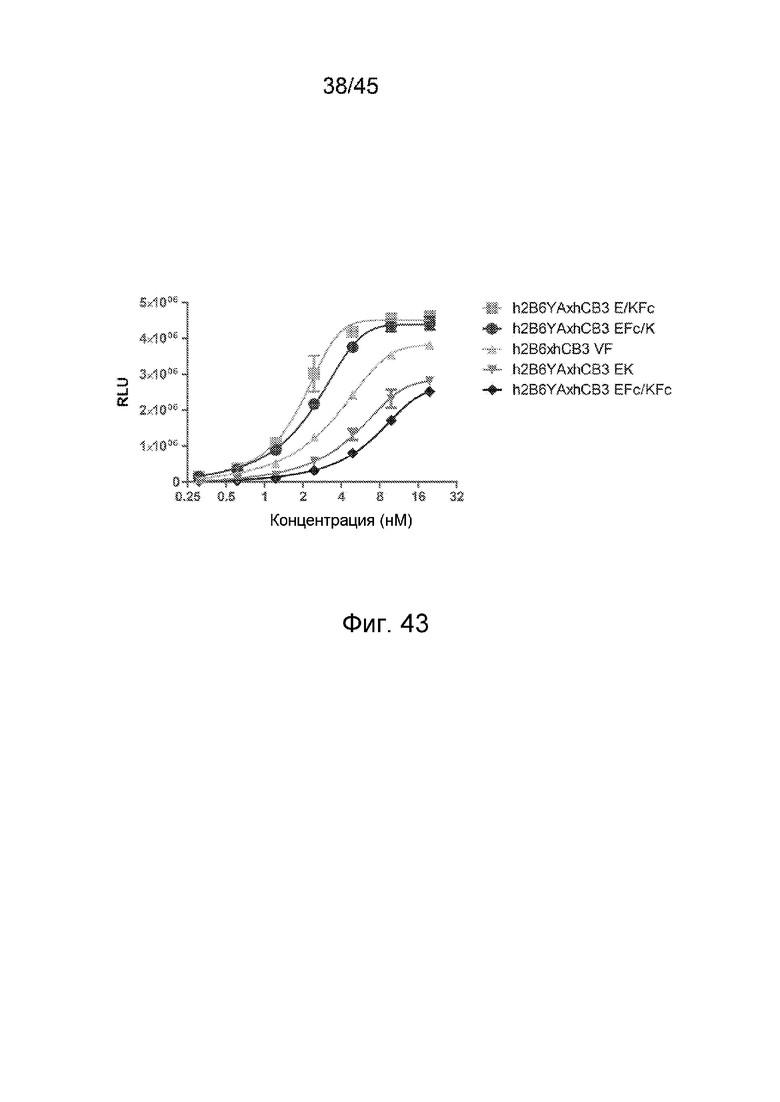
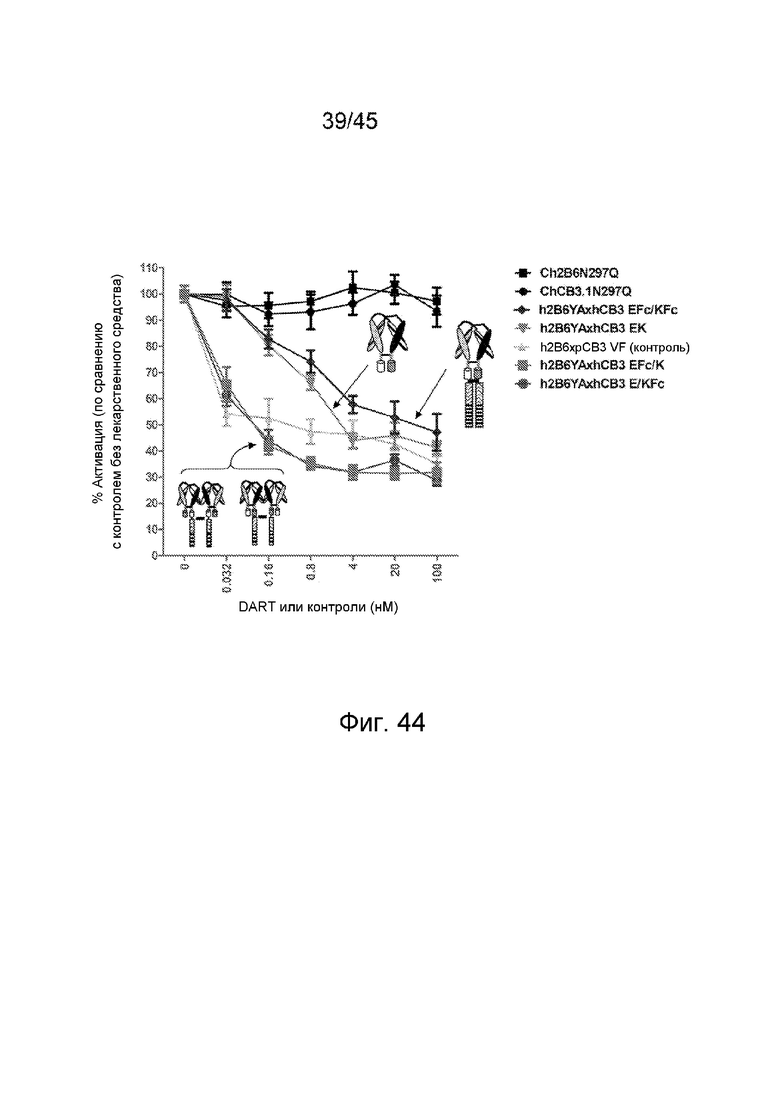
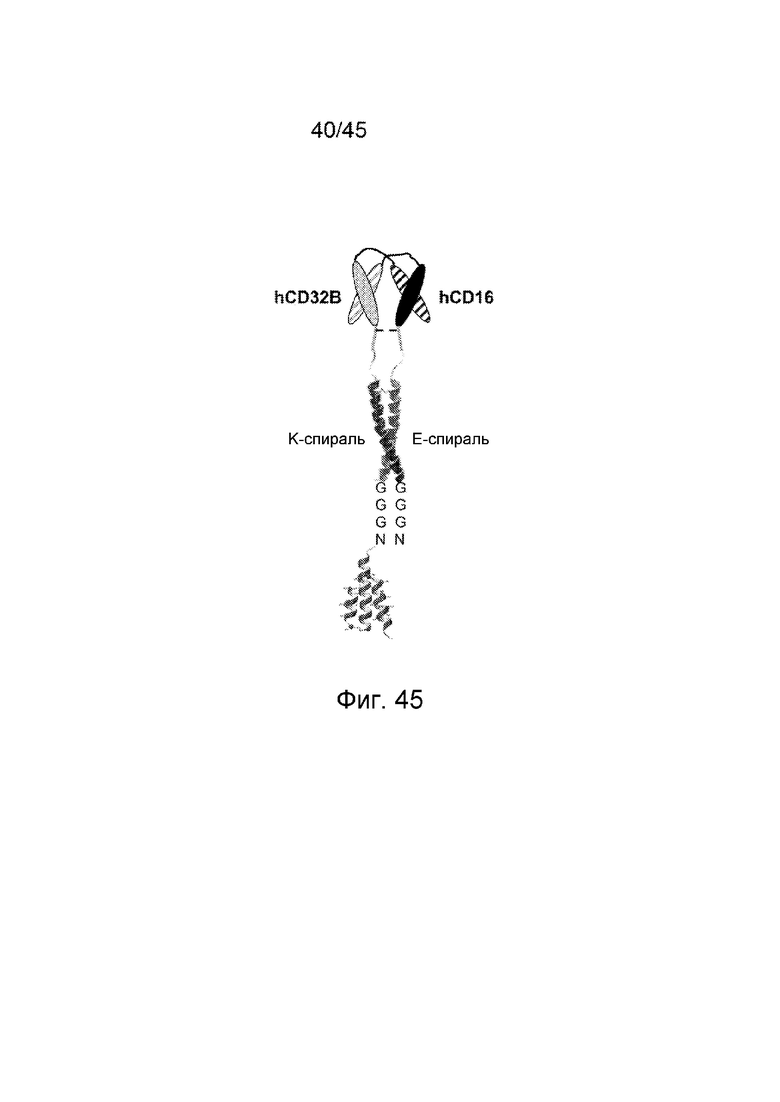
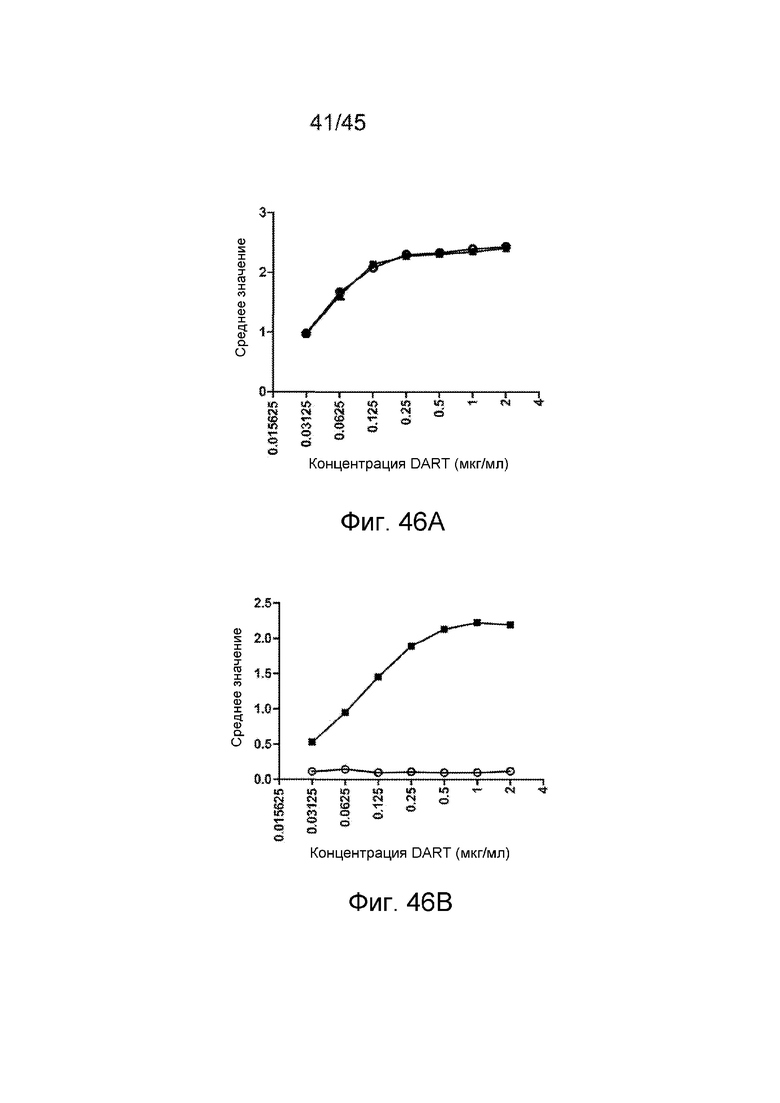

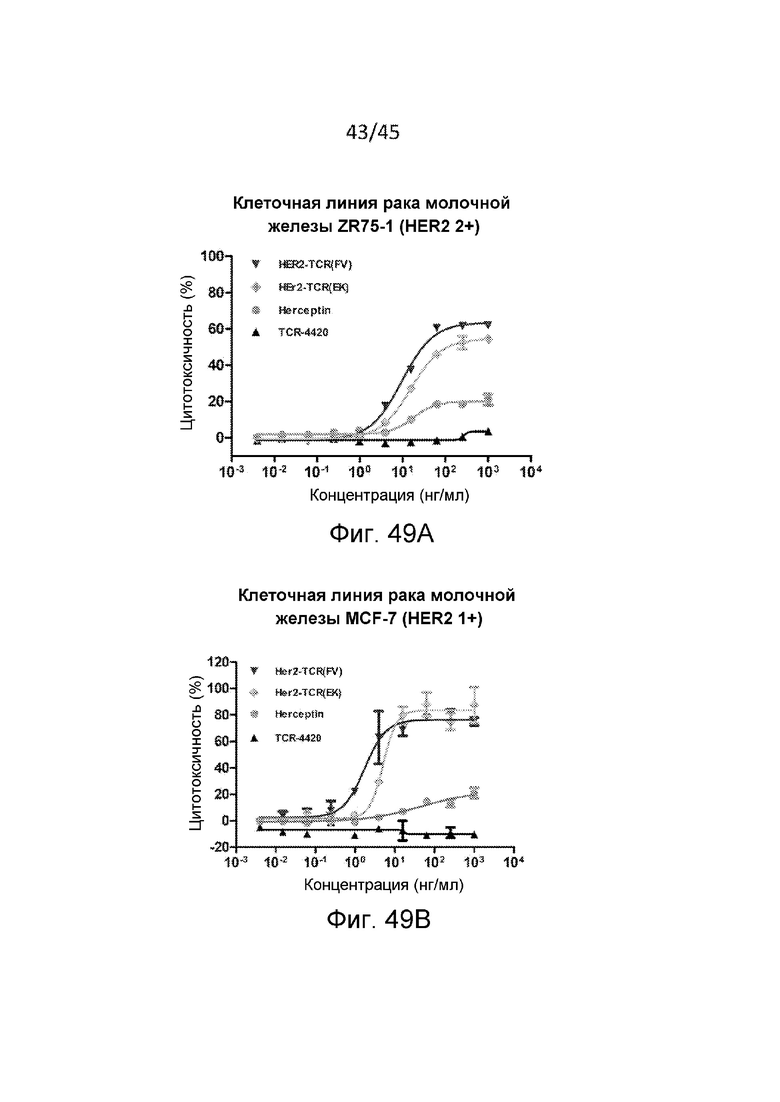
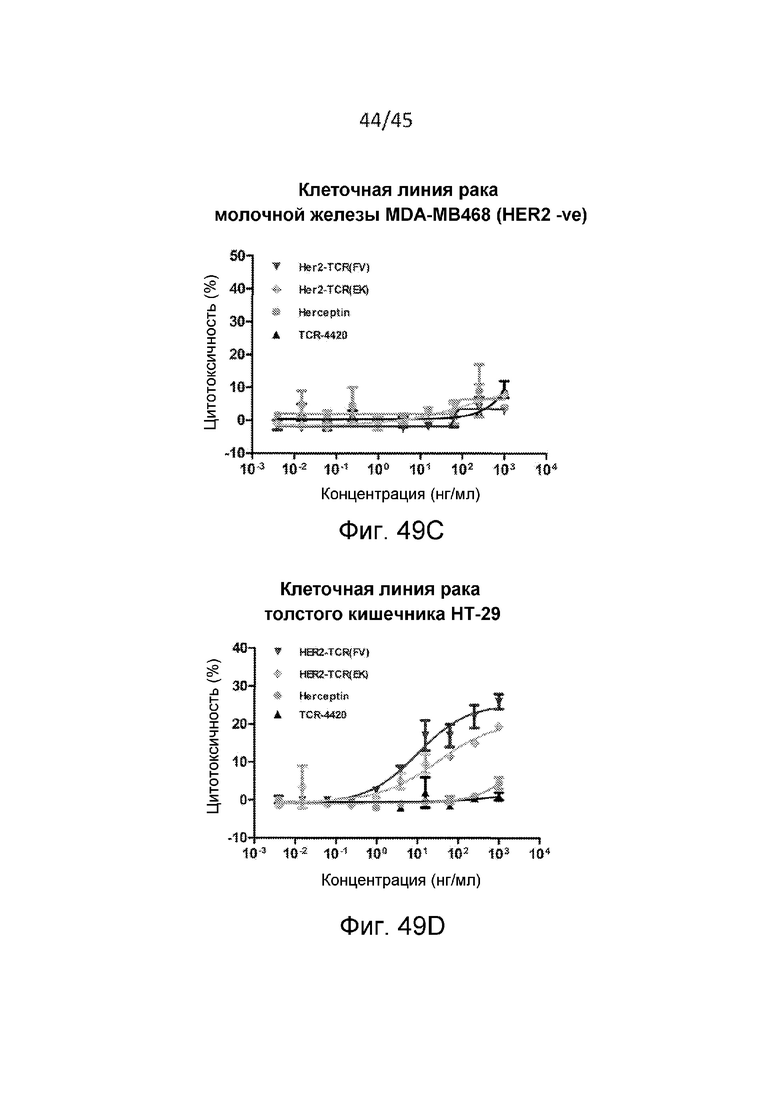
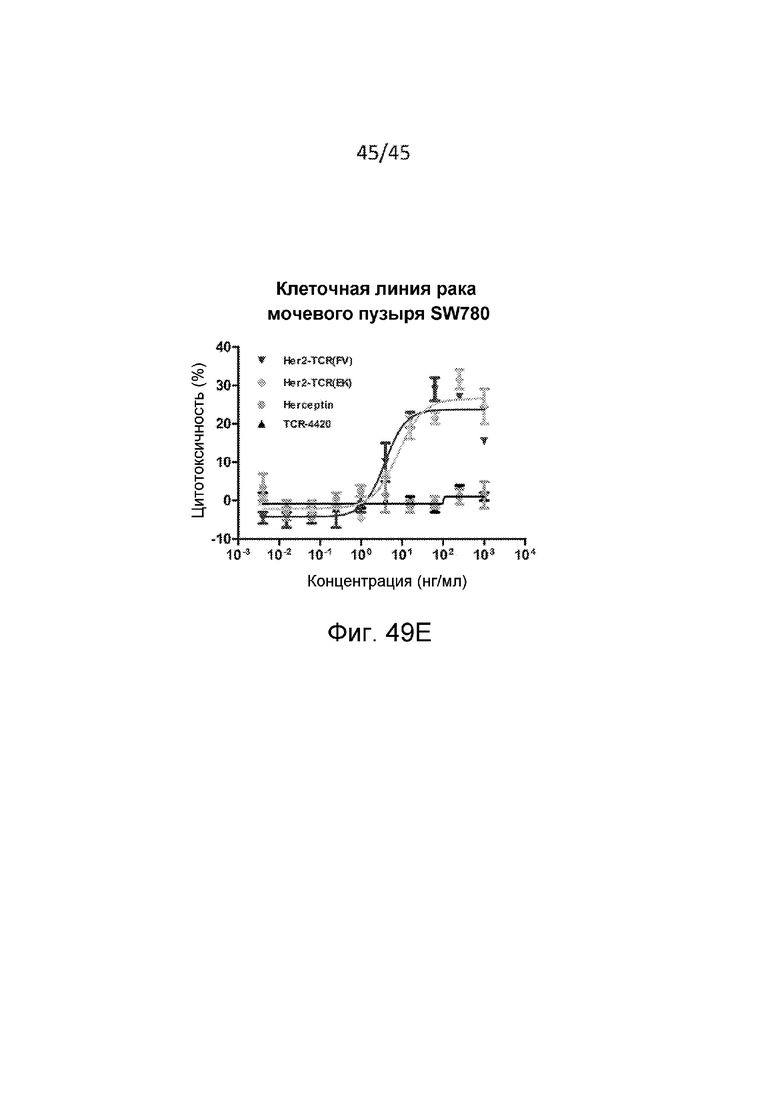
Изобретение относится к биотехнологии и медицине, в частности к молекулам диантител и к их применению для лечения различных заболеваний и нарушений, включая иммунологические нарушения, инфекционное заболевание, интоксикацию и злокачественные опухоли. Молекулы диантител по изобретению содержат две полипептидные цепи, которые ассоциируют с образованием по меньшей мере двух участков связывания эпитопа, которые могут распознавать один или различные эпитопы на одном или различных антигенах. Изобретение можно использовать для эффективного лечения аутоиммунного расстройства. 12 н. и 17 з.п. ф-лы, 17 табл., 1 пр., 49 ил.
1. Молекула диантитела, которая связывает эпитоп CD32B и эпитоп CD79b, где указанное диантитело содержит первую полипептидную цепь и вторую полипептидную цепь, ковалентно связанные друг с другом, при условии, что ковалентная связь не является пептидной связью, где:
указанная первая полипептидная цепь содержит, в направлении от N-конца к C-концу: (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичного к CD32B, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичного к CD79b, и (iii) третий домен, содержащий, по меньшей мере, один остаток цистеина, и разделитель в виде E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, где первый домен и второй домен ковалентно связаны, так что первый домен и второй домен не ассоциируют с образованием сайта связывания эпитопа;
указанная вторая полипептидная цепь содержит, в направлении от N-конца к C-концу: (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), специфичного к CD79b, (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), специфичного к CD32B, и (iii) шестой домен, содержащий, по меньшей мере, один остаток цистеина, и разделитель в виде E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, где четвертый домен и пятый домен ковалентно связаны, так что четвертый домен и пятый домен не ассоциируют с образованием сайта связывания эпитопа;
где указанный остаток цистеина указанных третьего и шестого доменов образует дисульфидную связь между указанной первой и второй полипептидными цепями;
где, когда указанная первая полипептидная цепь содержит указанный разделитель в виде E-спирали, указанная вторая полипептидная цепь содержит указанный разделитель в виде K-спирали; и когда указанная первая полипептидная цепь содержит указанный разделитель в виде K-спирали, указанная вторая полипептидная цепь содержит указанный разделитель в виде E-спирали;
где первый домен и пятый домен ассоциируют с образованием первого сайта связывания (VL1) (VH1), который связывает CD32B; и
где второй домен и четвертый домен ассоциируют с образованием второго сайта связывания (VL2) (VH2), который связывает CD79b.
2. Молекула диантитела по п.1, где указанная первая полипептидная цепь или указанная вторая полипептидная цепь дополнительно содержат Fc-домен или его часть.
3. Молекула диантитела по п.2, где указанный Fc-домен или его часть связаны с указанным разделителем в виде E-спирали или указанным разделителем в виде K-спирали.
4. Молекула диантитела по п.2, где указанный Fc-домен или его часть связаны с N-концом указанного первого домена или указанного четвертого домена.
5. Молекула диантитела, которая связывает эпитоп CD32B и эпитоп CD79, и которая представляет собой димер двух молекул диантитела, где:
(A) первая из указанных двух молекул диантитела содержит первую полипептидную цепь и вторую полипептидную цепь, где указанная первая полипептидная цепь и указанная вторая полипептидная цепь указанной первой молекулы диантитела ковалентно связаны друг с другом, при условии, что ковалентная связь не является пептидной связью, и где:
указанная первая полипептидная цепь указанной первой молекулы диантитела содержит, в направлении от N-конца к C-концу: (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичного к CD32B, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичного к CD79b, и (iii) третий домен, содержащий, по меньшей мере, один остаток цистеина, разделитель в виде E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, и домен CH2 и домен CH3; где первый домен и второй домен ковалентно связаны, так что первый домен и второй домен не ассоциируют с образованием сайта связывания эпитопа;
указанная вторая полипептидная цепь указанной первой молекулы диантитела содержит, в направлении от N-конца к C-концу: (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), специфичного к CD79b, (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), специфичного к CD32B, и (iii) шестой домен, содержащий, по меньшей мере, один остаток цистеина, и разделитель в виде E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, где четвертый домен и пятый домены ковалентно связаны таким образом, что четвертый домен и пятый домены не ассоциируют с образованием сайта связывания эпитопа;
где указанная первая полипептидная цепь указанной первой молекулы диантитела содержит указанный разделитель в виде E-спирали, указанная вторая полипептидная цепь указанной первой молекулы диантитела содержит указанный разделитель в виде K-спирали; и когда указанная первая полипептидная цепь указанной первой молекулы диантитела содержит указанный разделитель в виде K-спирали, указанная вторая полипептидная цепь указанной первой молекулы диантитела содержит указанный разделитель в виде E-спирали;
где указанный остаток цистеина указанных третьего и шестого доменов указанной первой молекулы диантитела образует дисульфидную связь между указанной первой и второй полипептидными цепями указанной первой молекулы диантитела;
где первый домен и пятый домен указанной первой молекулы диантитела ассоциируют с образованием первого сайта связывания (VL1) (VH1), который связывает CD32B;
где второй домен и четвертый домен указанной первой молекулы диантитела связываются с образованием второго сайта связывания (VL2) (VH2), который связывает CD79b; и
(B) вторая из указанных двух молекул диантитела содержит первую полипептидную цепь и вторую полипептидную цепь, где указанная первая полипептидная цепь и указанная вторая полипептидная цепь указанной второй молекулы диантитела ковалентно связаны друг с другом, при условии, что ковалентная связь не является пептидной связью, и где:
указанная первая полипептидная цепь указанной второй молекулы диантитела содержит, в направлении от N-конца к C-концу: (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичного к CD32B, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичного к CD79b, и (iii) третий домен, содержащий, по меньшей мере, один остаток цистеина, разделитель в виде E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, и домен CH2 и домен CH3; где первый домен и второй домен ковалентно связаны, так что первый домен и второй домен не ассоциируют с образованием сайта связывания эпитопа;
указанная вторая полипептидная цепь указанной второй молекулы диантитела содержит, в направлении от N-конца к C-концу: (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), специфичного к CD79b, (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), специфичного к CD32B, и (iii) шестой домен, содержащий, по меньшей мере, один остаток цистеина, и разделитель в виде E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, где четвертый домен и пятый домен ковалентно связаны таким образом, что четвертый домен и пятый домен не ассоциируют с образованием сайта связывания эпитопа;
где, когда указанная первая полипептидная цепь указанной второй молекулы диантитела содержит указанный разделитель в виде E-спирали, указанная вторая полипептидная цепь указанной второй молекулы диантитела содержит указанный разделитель в виде K-спирали; и когда указанная первая полипептидная цепь указанной молекулы второго диантитела содержит указанный разделитель в виде K-спирали, указанная вторая полипептидная цепь указанной молекулы второго диантитела содержит указанный разделитель в виде E-спирали;
где указанный остаток цистеина указанных третьего и шестого доменов указанной молекулы второго диантитела образует дисульфидную связь между указанной первой и второй полипептидными цепями указанной молекулы второго диантитела;
где первый домен и пятый домен указанной второй молекулы диантитела ассоциируют с образованием первого сайта связывания (VL1) (VH1), который связывает CD32B;
где второй домен и четвертый домен указанной молекулы второго диантитела ассоциируют с образованием второго сайта связывания (VL2) (VH2), который связывает CD79b; и
где указанные домен CH2 и домен CH3 указанной первой полипептидной цепи указанной первой молекулы диантитела ассоциируют с указанным доменом CH2 и доменом CH3 указанной первой полипептидной цепи указанной молекулы второго диантитела с образованием Fc-области.
6. Молекула диантитела, которая связывает эпитоп CD32B и эпитоп CD79b и которая представляет собой димер двух молекул диантитела, где:
(A) первая из указанных двух молекул диантитела содержит первую полипептидную цепь и вторую полипептидную цепь, где указанная первая полипептидная цепь и указанная вторая полипептидная цепь указанной первой молекулы диантитела ковалентно связаны друг с другом, при условии, что ковалентная связь не является пептидной связью, и где:
указанная первая полипептидная цепь указанной первой молекулы диантитела содержит, в направлении от N-конца к C-концу: (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичного к CD79b, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичного к CD32B, и (iii) третий домен, содержащий, по меньшей мере, один остаток цистеина, разделитель в виде E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, и домен CH2 и домен CH3; где первый домен и второй домен ковалентно связаны, так что первый домен и второй домен не ассоциируют с образованием сайта связывания эпитопа;
указанная вторая полипептидная цепь указанной первой молекулы диантитела содержит, в направлении от N-конца к C-концу: (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), специфичного к CD32B, (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), специфичного к CD79b, и (iii) шестой домен, содержащий, по меньшей мере, один остаток цистеина, и разделитель E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, где четвертый домен и пятый домен ковалентно связаны таким образом, что четвертый домен и пятый домен не ассоциируют с образованием сайта связывания эпитопа;
где указанная первая полипептидная цепь указанной первой молекулы диантитела содержит указанный разделитель в виде E-спирали, указанная вторая полипептидная цепь указанной первой молекулы диантитела содержит указанный разделитель в виде K-спирали; и когда указанная первая полипептидная цепь указанной первой молекулы диантитела содержит указанный разделитель в виде K-спирали, указанная вторая полипептидная цепь указанной первой молекулы диантитела содержит указанный разделитель в виде E-спирали;
где указанный остаток цистеина указанных третьего и шестого доменов указанной первой молекулы диантитела образует дисульфидную связь между указанной первой и второй полипептидными цепями указанной первой молекулы диантитела;
где первый домен и пятый домен указанной первой молекулы диантитела ассоциируют с образованием первого сайта связывания (VL1) (VH1), который связывает CD79b;
где второй домен и четвертый домен указанной первой молекулы диантитела ассоциируют с образованием второго сайта связывания (VL2) (VH2), который связывает CD32B; и
(B) вторая из указанных двух молекул диантитела содержит первую полипептидную цепь и вторую полипептидную цепь, где указанная первая полипептидная цепь и указанная вторая полипептидная цепь указанной второй молекулы диантитела ковалентно связаны друг с другом, при условии, что ковалентная связь не является пептидной связью, и где:
указанная первая полипептидная цепь указанной молекулы второго диантитела содержит, в направлении от N-конца к C-концу: (i) первый домен, содержащий связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), специфичного к CD79b, (ii) второй домен, содержащий связывающую область вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), специфичного к CD32B, и (iii) третий домен, содержащий, по меньшей мере, один остаток цистеина, разделитель в виде E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, и домен CH2 и домен CH3; где первый домен и второй домен ковалентно связаны, так что первый домен и второй домен не ассоциируют с образованием сайта связывания эпитопа;
указанная вторая полипептидная цепь указанной молекулы второго диантитела содержит, в направлении от N-конца к C-концу: (i) четвертый домен, содержащий связывающую область вариабельного домена легкой цепи второго иммуноглобулина (VL2), специфичного к CD32B, (ii) пятый домен, содержащий связывающую область вариабельного домена тяжелой цепи первого иммуноглобулина (VH1), специфичного к CD79b, и (iii) шестой домен, содержащий, по меньшей мере, один остаток цистеина, и разделитель E-спирали, содержащий аминокислотную последовательность SEQ ID NO: 299, или разделитель в виде K-спирали, содержащий аминокислотную последовательность SEQ ID NO: 300, где четвертый домен и пятый домен ковалентно связаны таким образом, что четвертый домен и пятый домен не ассоциируют с образованием сайта связывания эпитопа;
где, когда указанная первая полипептидная цепь указанной второй молекулы диантитела содержит указанный разделитель в виде E-спирали, указанная вторая полипептидная цепь указанной второй молекулы диантитела содержит указанный разделитель в виде K-спирали; и когда указанная первая полипептидная цепь указанной молекулы второго диантитела содержит указанный разделитель в виде K-спирали, указанная вторая полипептидная цепь указанной молекулы второго диантитела содержит указанный разделитель в виде E-спирали;
где указанный остаток цистеина указанных третьего и шестого доменов указанной молекулы второго диантитела образует дисульфидную связь между указанной первой и второй полипептидными цепями указанной молекулы второго диантитела;
где первый домен и пятый домен указанной второй молекулы диантитела ассоциируют с образованием первого сайта связывания (VL1) (VH1), который связывает CD79b;
где второй домен и четвертый домен указанной молекулы второго диантитела ассоциируют с образованием второго сайта связывания (VL2) (VH2), который связывает CD32B;
где указанные домен CH2 и домен CH3 указанной первой полипептидной цепи указанной первой молекулы диантитела ассоциируют с указанным доменом CH2 и доменом CH3 указанной первой полипептидной цепи указанной молекулы второго диантитела с образованием Fc-области.
7. Молекула диантитела по п.1, где как упомянутый разделитель в виде E-спирали, так и упомянутый разделитель в виде K-спирали связаны с Fc-доменом или его частью.
8. Молекула диантитела по п.1, где указанный разделитель в виде E-спирали или указанный разделитель K-coil связаны с полипептидной частью связываемого сывороткой белка, где указанная полипептидная часть способна связываться с указанным связываемым сывороткой белком.
9. Молекула диантитела по п.8, где указанный связываемый сывороткой белок представляет собой альбумин-связывающий белок.
10. Молекула диантитела по п.9, где указанный альбумин-связывающий белок представляет собой стрептококковый белок G, а указанная полипептидная часть представляет собой альбумин-связывающий домен (ABD) указанного стрептококкового белка G.
11. Молекула диантитела по п.10, где указанный альбумин-связывающий домен (ABD) указанного стрептококкового белка G представляет собой альбумин-связывающий домен 3 (ABD3) белка G штамма Streptococcus G148.
12. Молекула диантитела по любому из пп.8-11, где указанная молекула диантитела показывает период полужизни в сыворотке in vivo более 2 часов.
13. Молекула диантитела по п.12, где указанное диантитело показывает период полувыведения из сыворотки in vivo более 10 часов.
14. Молекула диантитела по п.13, где указанное диантитело показывает период полувыведения из сыворотки in vivo более 20 часов.
15. Молекула диантитела по любому из пп.1-5 и 7-14, где:
(A) указанная первая полипептидная цепь содержит вариабельный домен легкой цепи, промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и вариабельный домен тяжелой цепи SEQ ID NO: 252; и
(В) указанная вторая полипептидная цепь содержит:
(i) вариабельный домен легкой цепи, промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и вариабельный домен тяжелой цепи SEQ ID NO: 254; или
(ii) вариабельный домен легкой цепи, промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и вариабельный домен тяжелой цепи SEQ ID NO: 256.
16. Молекула диантитела по п.15, где указанная вторая полипептидная цепь содержит указанный вариабельный домен легкой цепи, указанный промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и указанный вариабельный домен тяжелой цепи SEQ ID NO: 254.
17. Молекула диантитела по п.15, где указанная вторая полипептидная цепь содержит указанный вариабельный домен легкой цепи, указанный промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и указанный вариабельный домен тяжелой цепи SEQ ID NO: 256.
18. Молекула диантитела по п.6, где:
(А) указанная первая полипептидная цепь содержит:
(i) вариабельный домен легкой цепи, промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и вариабельный домен тяжелой цепи SEQ ID NO: 254; или
(ii) вариабельный домен легкой цепи, промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и вариабельный домен тяжелой цепи SEQ ID NO: 256; и
(B) указанная вторая полипептидная цепь содержит вариабельный домен легкой цепи, промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и вариабельный домен тяжелой цепи SEQ ID NO: 252.
19. Молекула диантитела по п.18, где указанная первая полипептидная цепь содержит указанный вариабельный домен легкой цепи, указанный промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и указанный вариабельный домен тяжелой цепи SEQ ID NO: 254.
20. Молекула диантитела по п.18, где указанная первая полипептидная цепь содержит указанный вариабельный домен легкой цепи, указанный промежуточный линкер GGGSGGGG, содержащий аминокислотную последовательность SEQ ID NO: 10, и указанный вариабельный домен тяжелой цепи SEQ ID NO: 256.
21. Фармацевтическая композиция для ингибирования пролиферации B-клеток, содержащая молекулу диантитела по любому из пп.1-20 и фармацевтически приемлемый носитель.
22. Применение молекулы диантитела по любому из пп.1-20 или фармацевтической композиции по п.21 для производства лекарственного средства для ингибирования пролиферации B-клеток.
23. Фармацевтическая композиция для ингибирования активации B-клеток, содержащая молекулу диантитела по любому из пп.1-20 и фармацевтически приемлемый носитель.
24. Применение молекулы диантитела по любому из пп.1-20 или фармацевтической композиции по п.23 для производства лекарственного средства для ингибирования активации В-клеток.
25. Фармацевтическая композиция для лечения аутоиммунного расстройства, содержащая молекулу диантитела по любому из пп.1-20 и фармацевтически приемлемый носитель.
26. Применение молекулы диантитела по любому из пп.1-20 или фармацевтической композиции по п.25 для производства лекарственного средства для лечения аутоиммунного расстройства.
27. Способ ингибирования пролиферации B-клеток у субъекта, нуждающегося в этом, включающий введение указанному субъекту молекулы диантитела по любому из пп.1-20 или фармацевтической композиции по п.21.
28. Способ ингибирования активации B-клеток у субъекта, нуждающегося в этом, включающий введение указанному субъекту молекулы диантитела по любому из пп.1-20 или фармацевтической композиции по п.23.
29. Способ лечения аутоиммунного расстройства у субъекта, нуждающегося в этом, включающий введение указанному субъекту молекулы диантитела по любому из пп.1-20 или фармацевтической композиции по п.25.
| WO 2006113665 A2, 26.10.2006 | |||
| US 20070274998 A1, 29.11.2007 | |||
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО, ЗАМЕНЯЮЩЕЕ ФУНКЦИОНАЛЬНЫЕ БЕЛКИ | 2004 |
|
RU2339696C2 |

