Область изобретения
Изобретение относится к связывающим элементам, в частности к молекулам антитела, для IgE. Связывающие элементы в числе прочего являются полезными для лечения расстройств, опосредованных IgE, включая аллергии и астму.
IgE представляет собой члена семейства иммуноглобулинов и опосредует аллергические ответы, такие как астма, пищевые аллергии, гиперчувствительность типа 1 и воспаление носовых пазух.
IgE секретируется и экспрессируется на поверхности В-клеток. Вкратце, IgE прикрепляется к мембране В-клетки с помощью трансмембранного домена, который является связанным с молекулой зрелого IgE посредством короткого мембраносвязывающего участка. IgE может также быть связан с помощью своего Fc участка с В-клетками, моноцитами, эозинофилами и тромбоцитами посредством рецептора IgE с низкой аффинностью (FcεRII, который также является известным как CD23). При воздействии аллергена В-клетки, которые продуцируют специфический для аллергена IgE, клонально размножаются. Специфический для аллергена IgE потом высвобождается в системное русло крови В-клетками, где он, в свою очередь, связывается с В-клетками с помощью FcεRII, а также с тучными клетками и базофилами посредством рецептора с высокой аффинностью (FcεRI). Такие тучные клетки и базофилы, таким образом, сенсибилизируются с помощью аллергена. Последующее воздействие аллергена перекрестно связывает FcεRI на тучных клетках и базофилах, активируя таким образом высвобождение ими гистамина и других факторов, ответственных за клиническую гиперчувствительность и анафилаксию.
Связывающие элементы, которые ингибируют связывание и функциональную активность посредством FcERI с одновременным ингибированием FcERII или без его ингибирования являются полезными для ингибирования опосредованных IgE болезненных состояний, таких как аллергии и астма.
При этом является понятным, что FcεRI и FcεRII связываются с сайтом(ами) узнавания в константном (Fc) домене IgE. Были предприняты различные исследования для идентификации таких сайтов узнавания. Например, пептиды, соответствующие специфическим частям IgE молекулы, использовали либо в качестве конкурентных ингибиторов связывания IgE с рецептором (Burt и др., Eur. J. Immun, 17: 437-440 [1987]; Helm и др., Nature, 331:180-183 [1988]; Helm и др., Proc. Natl. Acad. Sci., 86: 9465-9469 [1989]; Vercelli и др., Nature, 338: 649-651 [1989]; Nio и др., Peptide Chemistry, 203-208 [1990]), либо для стимуляции выработки анти-IgE антител, которые могут блокировать взаимодействие IgE с рецептором (Burt и др., Molec. Immun. 24: 379-389 [1987]; Robertson и др., Molec. Immun., 25:103-113 [1988]; Baniyash и др., Molec. Immun. 25: 705-711 [1988]).
Недавно был получен Xolair® (Омализумаб), который продается для лечения пациентов с астмой. Xolair® представляет собой гуманизированное IgG1k моноклональное антитело, которое селективно связывается с IgE, снижая, таким образом, связывание IgE, по крайней мере, с FcεRI на поверхности тучных клеток и базофилов. Путем снижения уровня связанного с поверхностью IgE на клетках, которые несут FcεRI, Xolair® в некоторой мере снижает степень высвобождения медиаторов аллергического ответа. Xolair® является раскрытым в публикациях международных патентных заявок: WO 93/04173 и WO 97/04807.
Однако существует потребность в иных связывающих элементах для IgE, таких, которые обладают более высокой аффинностью и/или эффективностью по сравнению с Xolair®, для многообещающих терапевтических стратегий.
Изобретение
Путем использования приемлемых методик селекции и анализов мы усовершенствовали связывающие элементы, которые ингибируют связывание с и функциональную активность посредством FcERI (рецептор IgE, представленный на тучных клетках) с одновременным ингибированием FcERII или при отсутствии такого ингибирования.
Связывающий элемент в соответствии с изобретением ингибирует связывание с и функциональную активность посредством FcERI с одновременным ингибированием FcERII или при отсутствии такого ингибирования. Ингибирование связывания может представлять собой непосредственное ингибирование, например путем нейтрализации IgE. Связывающий элемент в соответствии с изобретением типично нейтрализует человеческий IgE со значением IC50, меньшим, чем приблизительно 10 нМ, как определяется с помощью, например, RBL-ER51 анализа передачи кальциевого сигнала. В некоторых воплощениях связывающий элемент в соответствии с изобретением нейтрализует человеческий IgE со , значением IC50, меньшим, чем приблизительно 1 нМ, или меньшим, чем приблизительно 0,5 нМ, или меньшим, чем приблизительно 0,2 нМ, как определяется, например, с помощью RBL-ER51 анализа передачи кальциевого сигнала.
Связывающие элементы в соответствии с изобретением могут также связываться с и нейтрализовать нечеловеческий IgE, означающий IgE ортологи, которые существуют в природе у видов, отличных от человека.
Связывающие элементы в соответствии с изобретением обычно являются специфическими для IgE по сравнению с другими иммуноглобулинами и, таким образом, связывают IgE селективно. Такая селективность может быть определена или продемонстрирована, например, в стандартном анализе конкурентного связывания.
Связывающие элементы являются полезными для лечения и/или предотвращения расстройств, которые являются опосредованными IgE, в частности различных аллергий и астмы.
Связывающие элементы являются полезными для снижения циркуляции свободного IgE у млекопитающих, а также полезными для ингибирования индуцированной аллергеном дегрануляции тучных клеток in vivo или in vitro.
Связывающие элементы также являются полезными для ингибирования биологических ответов, опосредованных связыванием IgE с FcERI с одновременньм ингибированием биологических ответов, опосредованных связыванием IgE с FcERII или при отсутствии такого ингибирования, либо in vivo, либо in vitro.
Связывающие элементы в соответствии с изобретением могут также обладать диагностической полезностью, как, например, для определения присутствия или количества IgE, или присутствия, или количества специфического для аллергена IgE в образце, который представляет интерес, таком как образец, полученный от пациента с астмой или аллергией.
Может использоваться любой приемлемый способ для определения последовательности остатков, связанных связывающим элементом. Например, может использоваться сканирование пептидного связывания, такое как иммуноферментный анализ (ELISA) на основе PEPSCAN, как описано где-либо в данной заявке. При сканировании пептидного связывания, таком как форма, которая обеспечивается PEPSCAN системами, короткие перекрывающиеся пептиды, имеющие происхождение от антигена, подвергаются системному скринингу на связывание со связывающим элементом. Пептиды могут быть ковалентно связаны с поверхностью основы с образованием набора для анализа пептидов. Пептиды могут быть линейными или иметь сжатую конформацию. Сжатая конформация может быть получена при использовании пептидов, содержащих терминальный Cys остаток на каждом конце пептидной последовательности. Cys остатки могут быть ковалентно связанными непосредственно или опосредовано с поверхностью основы так, что пептид удерживается в скрученной конформации. Таким образом, пептиды, используемые в способе, могут содержать Cys остатки, присоединенные к каждому концу пептидной последовательности, соответствующей фрагменту антигена. Также могут использоваться пептиды с двумя петлями, в которых Cys остаток дополнительно размещается при или поблизости средней части пептидной последовательности. Cys остатки могут быть ковалентно связанными непосредственно или опосредовано с поверхностью основы так, что пептиды образуют конформацию с двумя петлями, с одной петлей с каждой стороны центрального Cys остатка. Пептиды могут быть получены синтетически, и Cys остатки могут, таким образом, быть встроены в желаемых положениях, несмотря на то, что такая последовательность IgE не существует в природе. Необязательно, как линейные, так и сжатые пептиды могут подвергаться скринингу в анализе пептидного связывания. Сканирование пептидного связывания может вовлекать идентификацию (например, при использовании ELISA) множества пептидов, с которыми связывается связывающий элемент, где пептиды обладают аминокислотными последовательностями, соответствующими фрагментам IgE (например, пептиды, содержащие приблизительно 5, 10 или 15 смежных остатков IgE), и сравнение пептидов для того, чтобы определить отпечаток остатков, связанных связывающим элементом, где отпечаток включает остатки, общие для перекрывающихся пептидов.
Альтернативно или дополнительно способ пептидного сканирования может вовлекать идентификацию пептидов, с которыми связывается связывающий элемент, по крайней мере, с заданным соотношением сигнал: фоновый сигнал. Подробности приемлемого способа пептидного сканирования для определения связывания являются известными в области техники. Другие способы, являющиеся хорошо известными в области техники, и такие, которые могут использоваться для определения остатков, связанных антителом и/или для подтверждения результатов пептидного сканирования, включают сайт-направленный мутагенез, обмен водород-дейтерий, масс-спектрометрию, ЯМР и рентгеновскую кристаллографию.
Связывающий элемент в соответствии с изобретением может или не может связывать и/или нейтрализовать варианты IgE. Таким образом, связывающий элемент в соответствии с изобретением может или не может ингибировать связывание вариантов IgE с FcERIc одновременным ингибированием FcERII или при отсутствии такого ингибирования.
Последовательности линейного эпитопа IgE, например в виде изолированных пептидных фрагментов или полипептидов, которые их включают, могут использоваться для идентификации, получения, изоляции и/или анализа связывающих элементов в соответствии с настоящим изобретением.
Как описывается более подробно ниже, связывающие элементы в соответствии с изобретением были продемонстрированы как такие, которые нейтрализуют IgE с высокой эффективностью. Нейтрализация означает ингибирование биологической активности IgE. Связывающие элементы в соответствии с изобретением могут нейтрализовать одну или более биологических активностей IgE, но типично ингибируют связывание IgE с FcERI с одновременным ингибированием связывания с FcERII или при отсутствии такого ингибирования.
Нейтрализация связывания IgE с FcERI с одновременным ингибированием FcERII или при его отсутствии может необязательно измеряться как функция биологической активности рецептора, такая как индуцированная аллергеном дегрануляция тучных клеток.
Приемлемые анализы для измерения нейтрализации IgE связывающими элементами в соответствии с изобретением включают, например, биохимические анализы лиганда и рецептора и поверхностный плазменный резонанс (SPR) (например, BIACORE).
Ингибирование биологической активности может быть частичным или полным. Связывающие элементы могут ингибировать биологическую активность IgE, такую как связывание рецептора или дегрануляция тучных клеток, на 100%, или альтернативно, на: по крайней мере, 95 %, по крайней мере, 90 %, по крайней мере, 85 %, по крайней мере, 80 %, по крайней мере, 75 %, по крайней мере, 70 %, по крайней мере, 60 % или, по крайней мере, 50 % активности при отсутствии связывающего элемента.
Нейтрализующая эффективность связывающего элемента обычно выражается в виде значения IC50, в нМ, если не указано другое. В функциональных анализах IC50 представляет собой концентрацию связывающего элемента, которая снижает биологический ответ на 50% от его максимума. В исследованиях связывания лиганда IC50 представляет собой концентрацию, которая снижает связывание рецептора на 50 % уровня максимального специфического связывания. Значение IC50 может быть подсчитано путем выстраивания % максимального биологического ответа как функции log концентрации связывающего элемента и при использовании программного обеспечения, такого как Prism (GraphPad) для выстраивания сигмоидальной функции в соответствии с данными для получения значений IC50. Эффективность может быть определена или измерена при использовании одного или более анализов, известных квалифицированному специалисту, и/или описанных в данной заявке, или тех, на которые содержится ссылка в данной заявке.
Нейтрализующая эффективность связывающего элемента может быть выражена в виде среднегеометрического значения. Среднегеометрическое значение (которое также является известным как среднее геометрическое значение), как используется в данной заявке, означает средние логарифмические значения набора данных, превращенные обратно к основанию 10. Это требует, по крайней мере, двух измерений, например, по крайней мере, 2, предпочтительно, по крайней мере, 5, более предпочтительно, по крайней мере, 10 повторностей. Специалист в данной области техники может понять, что чем больше количество повторностей, тем надежнее будет среднегеометрическое значение. Выбор количества повторностей может быть отнесен к компетенции специалиста, квалифицированного в данной области техники.
Нейтрализация активности IgE связывающим элементом в анализе, описанном в данной заявке, показывает, что связывающий элемент связывает и нейтрализует IgE. Другие способы, которые могут использоваться для определения связывания связывающего элемента с IgE, включают ELISA, Вестерн-блоттинг, иммунопреципитацию, аффинную хроматографию и биохимические анализы.
В другом воплощении в соответствии с изобретением обеспечивается изолированный связывающий элемент, специфический для иммуноглобулина Е, при этом связывающий элемент имеет значение IC50 для связывания указанного связывающего элемента с иммуноглобулином Е в сыворотке, по крайней мере, в 10 раз ниже, чем для Xolair™, или альтернативно, по крайней мере, в 20 раз ниже, по крайней мере, в 50 раз ниже, по крайней мере, в 75 раз ниже, по крайней мере, в 100 раз ниже, по крайней мере, в 125 раз ниже, по крайней мере, в 150 раз ниже, по крайней мере, в 200 раз ниже, по крайней мере, в 300 раз ниже, по крайней мере, в 400 раз ниже или, по крайней мере, в 500 раз ниже.
Нейтрализующая эффективность связывающего элемента, как подсчитано в анализе при использовании IgE из первых видов (например, человека), может сравниваться с нейтрализующей эффективностью связывающего элемента в подобном анализе при подобных условиях, анализе при использовании IgE из вторых видов (например, обезьяны циномолгуса) для того, чтобы оценить степень перекрестной реактивности связывающего элемента для IgE двух видов. Альтернативно, перекрестная реактивность может быть оценена в конкурентном анализе связывания, как описано более подробно где-либо в данной заявке.
Связывающий элемент в соответствии с изобретением может иметь более высокую нейтрализующую эффективность в отношении связывании человеческого IgE или в биологическом анализе, чем при использовании IgE из видов, отличных от человека. Таким образом, нейтрализующая эффективность связывающего элемента в анализе с человеческим IgE может быть большей, чем в подобном анализе с IgE из видов, отличных от человека. Эффективность в связывании человеческого IgE в биологическом анализе может, например, быть приблизительно в 5 раз большей, чем в подобном анализе, использующем IgE обезьяны циномолгуса, или в другом воплощении может быть в 15 или 20 раз больше. В частности, эффективность в анализе передачи кальциевого сигнала для человеческого RBL-ER51 может быть определена для концентрации человеческого IgE 25 нг/мл, при этом она сравнивается с эффективностью при использовании 100 нг/мл IgE циномолгуса при других подобных условиях. Примеры данных, полученных в подобных анализах передачи кальциевого сигнала RBL-ER51 при использовании человеческого IgE и IgE циномолгуса, показаны в Таблице 2b.
Связывающий элемент в соответствии с изобретением может обладать более высокой аффинностью для человеческого IgE, чем для IgE других видов. Аффинность связывающего элемента для человеческого IgE может быть, например, приблизительно в 5 или 10 раз выше, чем для IgE обезьяны циномолгуса, или, в другом воплощении, может быть приблизительно в 100 раз выше. Примеры данных, полученных для IgE, как человека, так и обезьяны циномолгуса, показаны в Таблице 2а и b.
Связывающий элемент в соответствии с изобретением может иметь нейтрализующая эффективность в отношении IgE или IC50 приблизительно 10 нМ или менее, с концентрацией человеческого IgE 25 нг/мл, например, в RBL-ER51 анализе передачи кальциевого сигнала. Альтернативно, значение IC50 является меньшим, чем приблизительно 3 нМ. В других воплощениях IC50 является меньшим, чем приблизительно 1 нМ, или меньшим, чем приблизительно 0,5 нМ, или меньшим, чем приблизительно 0,2 нМ.
В другом воплощении в соответствии с изобретением обеспечивается изолированный связывающий элемент, специфический для иммуноглобулина Е, где связывающий элемент имеет среднегеометрическое значение IC50 для ингибирования передачи сигнала кальция, индуцированного 25 нг/мл IgE в RBL-ER51 клетках менее 1 нМ, или альтернативно менее 0,6 нМ, менее 0,5 нМ, менее 0,4 нМ, менее 0,3 нМ, менее 0,2 нМ или менее 0,1 нМ.
Кинетики связывания и аффинность (выраженная как константа равновесной диссоциации KD) элементов, связывающих IgE для r человеческого IgE, может быть определена, например, при использовании поверхностного плазмонного резонанса (BIACORE). Связывающие элементы в соответствии с изобретением обычно имеют аффинность для человеческого IgE (KD) меньшую, чем приблизительно 10 нМ, и в некоторых воплощениях имеют KD меньшую, чем приблизительно 5 нМ, в других воплощениях имеют KD меньшую, чем 2 нМ. Аффинность для IgE обезьяны циномолгуса обычно составляет менее приблизительно 20 нМ, в некоторых воплощениях значение KD составляет меньше, чем приблизительно 10 нМ.
Является доступным ряд методик для измерения связывающей аффинности антитела с его антигенами, одна из таких методик представляет собой KinExA. Кинетический эксклюзионный анализ (KinExA) представляет собой иммуноаналитическую платформу для общих целей (по существу с помощью проточного спектрофлуориметра), которая является способной к измерению констант равновесной диссоциации, констант скорости ассоциации и диссоциации для взаимодействий антиген/антитело. Поскольку KinExA осуществляют после достижения равновесного состояния, методика представляет собой преимущественную для использования при измерениях KD взаимодействия мультиваленый антиген/mAb. Связывание антитела с молекулой IgE представляет собой пример связывания мультивалентного антигена. Применение KinExA является особенно приемлемым, когда мультивалентный антиген означает, что мультимеры антитела и антигена, которые образуются, включают более чем одно антитело и более чем один антиген. В таких моделях комплексного взаимодействия определение точного значения KD может быть затруднено. Методика KinExA может быть осуществлена так, как описано Drake и др. (2004) Analytical Biochemistry 328, 35-43, как измеряется с помощью KinExA методики. Антитело 11 имеет значение KD 6,3 пМ, что существенно ниже, чем для Xolair™, которое имеет значение KD 353 пМ.
В другом воплощении в соответствии с изобретением обеспечивается изолированный связывающий элемент, специфический для иммуноглобулина Е, с KD 300 пМ или ниже, как измеряется при использовании методики KinExA. Альтернативно, KD составляет 200 пМ или ниже, 100 пМ или ниже, 50 пМ или ниже, 20 пМ или ниже, или 10 пМ или ниже.
In vivo эндогенный IgE может быть гликозилированным и, таким образом, гликозилированный человеческий IgE представляет собой терапевтическую модель терапии человека. Несмотря на то, что рекомбинантный человеческий IgE, который может иметь бактериальное происхождение и не являться гликозилированным, может использоваться в анализах, описанных в данной заявке. Связывающие элементы в соответствии с изобретением могут связывать гликозилированный человеческий IgE, такой как IgE, полученный с помощью миеломной линии клеток, такой как U266.B1. Это составляет значительное преимущество связывающих элементов в соответствии с изобретением, поскольку гликозилированный человеческий IgE представляет собой целевой антиген для in vivo применений у человека.
Связывающий элемент в соответствии с изобретением может включать молекулы антитела, предпочтительно человеческие молекулы антитела или гуманизированные молекулы антитела. В одном аспекте в соответствии с изобретением молекулы антитела представляют собой моноклональное антитело.
Сайт связывания антигена обычно образуется с помощью вариабельного тяжелого (VH) и вариабельного легкого (VL) доменов иммуноглобулина, с антигенсвязывающей областью, образованной шестью поверхностными полипептидными петлями, которые называются участками, определяющими комплементарность (CDR). Существует три CDR в каждом VH (HCDR1, HCDR2, HCDR3) и в каждом VL LCDR1, LCDR2, LCDR3), вместе с участками каркасного участка (FR).
Связывающий элемент в соответствии с изобретением обычно включает домен VH и/или VL антитела. VH домен включает набор HCDR, а VL домен включает набор LCDR. Молекула антитела может включать VH домен антитела, включающий VH CDR1, CDR2, и CDR3, и каркасный участок. Она может альтернативно или дополнительно включать VL домен антитела, содержащий VL CDR1, CDR2 и CDR3, и каркасный участок. Примеры VH доменов антитела (SEQ ID NOS:2, 12, 22, 32, 42, 52, 62, 72, 82, 92, 102, 112, 122, 132, 142, 152, 162, 172, 182, 192, 202, 212, 222, 232, 242, 252, 262, 272, 282, 288, 300, и 306) и VL доменов антитела (SEQ ID NOS:318, 320, 322, 324, 326, 328, 330, 332, 334, 336, 338, 340, 342, 344, 346, 348, 350, 352, 354, 356, 358, 360, 362, 364, 366, 368, 370, 372, 374, 376, 378 и 380) и CDR (SEQ ID NOS:3-5, 8-10, 13-15, 18-20, 23-25, 28-30, 33-35, 38-40, 43-45, 48-50, 53-55, 58-60, 63-65, 68-70, 73-75, 78-80, 83-85, 88-90, 93-95, 98-100, 103-105, 108-110, 113-115, 118-120, 123-125, 128-130, 133-135, 138-140, 143-145, 148-150, 153-155, 158-160, 163-165, 168-170, 173-175, 178-180, 183-185, 188-190, 193-195, 198-200, 203-205, 208-210, 213-215, 218-220, 223-225, 228-230, 233-235, 238-240, 243-245, 248-250, 253-255, 258-260, 263-265, 268-270, 273-275, 278-280, 283-285, 296-298, 289-291, 296-298, 301-303, 307-309, и 314-316) в соответствии с настоящим изобретением приведены в прилагаемом списке последовательностей, который образует часть настоящего раскрытия (смотри также Таблицу 3а). Дополнительные CDR раскрыты ниже и в Таблице 1. Все VH и VL последовательности, CDR последовательности, наборы CDR, наборы HCDR и наборы LCDR, раскрытые в данной заявке, представляют собой аспекты и воплощения в соответствии с изобретением.
Как описывается в данной заявке, "набор CDR" включает CDR1, CDR2 и CDR3. Таким образом, набор HCDR относится к HCDR1, HCDR2 и HCDR3, а набор LCDR относится к LCDR1, LCDR2 и LCDR3. Если не указано другое, то "набор CDR" включает HCDR и LCDR.
Альтернативно, связывающий элемент в соответствии с изобретением может включать сайт связывания антигена в пределах молекулы, отличной от антитела, обычно обеспеченный одним или более CDR, например, набором CDR в белковом каркасе молекулы, отличной от антитела, как обсуждается дополнительно ниже.
Как описывается в данной заявке, молекула исходного антитела была изолирована как такая, которая содержит набор CDR последовательностей, как показано в Таблице 1 (смотри Антитело 1), Посредством осуществления процесса оптимизации мы получили панель клонов антитела под номерами 2-28, с последовательностями CDR, которые имеют происхождение от исходных CDR последовательностей и содержат модификации в положениях, указанных в Таблице 1. Таким образом, например, из Таблицы 1 можно увидеть, что Антитело 2 имеет исходные последовательности HCDR1, HCDR2, LCDR1, LCDR2 и LCDR3, и содержит исходную HCDR3 последовательность в которой: остаток Кэбата 96 заменен на S, остаток Кэбата 97 заменен на L, остаток Кэбата 99 заменен на S, и остаток Кэбата 100 заменен на А.
Описанный в данной заявке связывающий элемент представляет собой такой включающий набор CDR, как показано в Таблице 1 (Антитело 1), где HCDR1 представляет собой SEQ ID NO:3 (остатки Кэбата 31-35), HCDR2 представляет собой SEQ ID NO:4 (остатки Кэбата 50-65), HCDR3 представляет собой SEQ ID NO:5 (остатки Кэбата 95-102), LCDR1 представляет собой SEQ ID NO:8 (остатки Кэбата 24-34), LCDR2 представляет собой SEQ ID NO:9 (остатки Кэбата 50-56) и LCDR3 представляет собой SEQ ID NO:10 (остатки Кэбата 89-97). Связывающий элемент в соответствии с изобретением может являться исходным связывающим элементом, как показано в Таблице 1, где один или более CDR содержат одно или более из аминокислотных добавлений, замен, делеций и/или инсерций. В некоторых воплощениях связывающий элемент включает набор CDR, содержащий от одного до десяти из добавлений, замен, делеций и/или инсерций по отношению к исходным последовательностям Антитела 11. В другом воплощении - от одной до десяти замен по сравнению с Антителом 11. В другом воплощении образуется от одного до одиннадцати прибавлений, замен, делеций и/или инсерций по отношению к исходным последовательностям Антитела 1. В другом воплощении - от одной до десяти замен по отношению к Антителу 1.
В некоторых воплощениях связывающий элемент в соответствии с изобретением включает HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3; где HCDR3 имеет аминокислотную последовательность SEQ ID NO:5, необязательно содержащую от 1 до 5 аминокислотных добавлений, замен, делеций и/или инсерций; а LCDR3 имеет аминокислотную последовательность SEQ ID NO:10, необязательно содержащую от 1 до 6 аминокислотных прибавлений, замен, делеций и/или инсерций. В этих воплощениях HCDR1 может содержать аминокислотную последовательность SEQ ID NO:3; HCDR2 может содержать аминокислотную последовательность SEQ ID NO:4; LCDR1 может содержать аминокислотную последовательность SEQ ID NO:8; и LCDR2 может содержать аминокислотную последовательность SEQ ID NO:9. Альтернативно, HCDR1, HCDR2, LCDR1 и LCDR2 могут также коллективно содержать одно или более аминокислотных прибавлений, замен, делеций и/или инсерций по отношению к исходным последовательностям (Антитело 1), как, например, от одной до десяти замен.
Связывающий элемент в соответствии с изобретением может включать один или комбинацию CDR, как описывается в данной заявке. Например, связывающий элемент в соответствии с изобретением может включать HCDR1, имеющий аминокислотную последовательность SEQ ID NO:3; HCDR2, имеющий аминокислотную последовательность SEQ ID NO:4; HCDR3 имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOS:5, 15, 25, 65, 75, 85, 95, 145, 155, 175 и 255; LCDR1, имеющий аминокислотную последовательность SEQ ID NO:8; LCDR2, имеющий аминокислотную последовательность SEQ ID NO:9; и LCDR3, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOS:10, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 210, 220, 230, 240, 250, 260, 270 и 280.
В некоторых воплощениях связывающий элемент или VH домен в соответствии с изобретением включает исходный HCDR3 (SEQ ID NO:5) с одной или более из следующих замен:
остаток Кэбата 96 заменен на S, М или Т;
остаток Кэбата 97 заменен на L или G;
остаток Кэбата 98 заменен на K;
остаток Кэбата 99 заменен на S, W, А, Т или Е;
остаток Кэбата 100 заменен на А или I.
В некоторых воплощениях связывающий элемент или его VL домен может включать исходный LCDR3 (SEQ ID NO 10) с остатком Кэбата 94, замененньм на Т, R, D, Р, Е, N, H, Q или А.
В некоторых воплощениях связывающий элемент или VL домен в соответствии с изобретением включает исходный LCDR3 (SEQ ID NO 10) с одной или более из следующих замен:
остаток Кэбата 94 заменен на Т, R, D, Р, Е, N, H, Q или А;
остаток Кэбата 95 заменен на Т, K, S, I, G, H, М, F, R, N, K или Q;
остаток Кэбата 95А заменен на L, H, D, G, R, N, Q, K или Е;
остаток Кэбата 95В заменен на Т, H, S, Y, L или N;
остаток Кэбата 96 заменен на G или А;
остаток Кэбата 97 заменен на Р, S или G.
В одном воплощении изобретение представляет собой связывающий элемент, в котором HCDR1 имеет аминокислотную последовательность SEQ ID NO:103, HCDR2 имеет аминокислотную последовательность SEQ ID NO:104, HCDR3 имеет аминокислотную последовательность SEQ ID NO:105, LCDR1 имеет аминокислотную последовательность SEQ ID NO:108, LCDR2 имеет аминокислотную последовательность SEQ ID NO:109, и LCDR3 имеет аминокислотную последовательность SEQ ID NO:110. Например, смотри Антитело 11 Таблицы 1.
Другие воплощения в соответствии с изобретением представляют собой связывающие элементы, такие как молекулы антитела, способные конкурировать с антителами в соответствии с изобретением, такими как Антитело 11 Таблицы 1, за связывание с человеческим IgE, при этом связывающие элементы нейтрализуют человеческий IgE со значением IC50 меньшим, чем приблизительно 1 нМ в анализе, описанном в данной заявке, или со значением IC50 меньшим, чем приблизительно 0,5 нМ. В некоторых воплощениях IC50 является меньшим приблизительно 0,2 нМ.
Изобретение обеспечивает связывающие элементы, включающие HCDR1, и/или HCDR2, и/или HCDR3 любого из антител 1-28, и/или LCDR1, и/или LCDR2, и/или LCDR3 любого из антител 1-28, например набор CDR любого из антител 1-28, показанных в Таблице 1. Связывающий элемент может включать набор VH CDR одного из этих антител. Необязательно, он может также включать набор VL CDR одного из этих антител, и VL CDR могут быть из того же или из различных антител, что и VH CDR. VH домен, включающий набор HCDR любого из антител 1-28, и/или VL домен, включающий набор LCDR любого из антител 1-28, также обеспечиваются изобретением.
Типично, когда VH домен спаривается с VL доменом с обеспечением антигенсвязывающего сайта антитела, хотя, как обсуждается в данной заявке ниже, для связывания антигена может использоваться только VH или VL домен. VH домен Антитела 1 (смотри Таблицу 1) может спариваться с VL доменом Антитела 1, так, что образуется антигенсвязывающий сайт антитела, включающий оба VH и VL домены антитела 1. Аналогичные воплощения обеспечиваются для других VH и VL доменов, раскрытых в данной заявке. В других воплощениях VH Антитела 1 спаривается с VL доменом, отличным от VL Антитела 1. Промискуитет легких цепей является хорошо установленным в области техники. Кроме того, аналогичные воплощения обеспечиваются изобретением для других VH и VL доменов, раскрытых в данной заявке. Таким образом, VH исходного или любого из антител 2-28 может спариваться с VL исходного или любого из антител 2-28.
Связывающий элемент может включать набор Н, и/или L CDR исходного антитела, или любого из антител 2-28 с двадцатью, шестнадцатью, десятью, девятью или менее, например, одной, двумя, тремя, четырьмя или пятью аминокислотными добавлениями, заменами, делениями, и/или инсерциями в пределах раскрытого набора Н и/или L CDR. Альтернативно, связывающий элемент может включать набор Н и/или L CDR исходного антитела или любого другого из антител 2-28 с двадцатью, шестнадцатью, десятью, девятью или менее, например одной, двумя, тремя, четырьмя или пятью аминокислотными заменами в пределах раскрытого набора Н и/или L CDR. Такие модификации могут потенциально быть осуществлены при любом остатке в пределах набора CDR. Например, модификации могут быть осуществлены в положениях, модифицированных в любом другом из антител 2-28, как показано в Таблице 1. Таким образом, одна или более модификаций могут включать одну или более замен в следующих остатках: остатках Кэбата 96, 97, 98, 99 и 100 в HCDR; остатках Кэбата 94, 95, 95А, 95В, 96 и 97 в LCDR.
Связывающий элемент может включать молекулу антитела, содержащую один или более CDR, например набор CDR, в каркасном участке антитела. Например, один или более CDR или набор CDR антитела может быть пересажен в каркасный участок (например, человеческий каркасный участок) для обеспечения молекулы антитела. Каркасные участки могут представлять собой эмбрионального гены последовательности человека. Таким образом, каркасный участок может быть герминизирован, в соответствии с чем один или более остатков в каркасном участке изменяют для соответствия участкам в эквивалентном положении в наиболее подобном эмбриональном каркасном участке человека. Таким образом, связывающий элемент в соответствии с изобретением может представлять собой изолированную молекулу антитела человека, содержащую VH домен, включающий набор HCDR в человеческом эмбриональном каркасном участке, например, человеческом эмбриональном каркасном участке VH IgG. Связывающий элемент также содержит VL домен, включающий набор LCDR, например, в человеческом эмбриональном каркасном участке IgG VL.
Остатки VH и/или VL каркасного участка могут быть модифицированы, как обсуждается и представлено в примерах в данной заявке, например при использовании сайт-направленного мутагенеза. VH или VL домен в соответствии с изобретением или связывающий элемент, включающий такой VL домен, предпочтительно имеет последовательность VH и/или VL домена антитела, как представлено в Таблице 3.
Негерминизированная молекула антитела содержит те же CDR, но отличные каркасные участки, по сравнению с герминизированной молекулой антитела. Герминизированное антитело может быть получено путем герминизации каркасных участков последовательностей VH и VL доменов, указанных в данной заявке для этих антител.
Связывающий элемент в соответствии с изобретением может быть таким, который конкурирует за связывание с IgE с любым связывающим элементом, которые оба связывают IgE и включают связывающий элемент, такой, как VH и/или VL домен, CDR, например, HCDR3, и/или набор CDR, раскрытые в данной заявке. Конкуренция между связывающими элементами может быть легко проанализирована in vitro, например, при использовании ELISA и/или с помощью маркирования при использовании специфической репортерной молекулы одного связывающего элемента, который может быть определен в присутствии одного или более других немаркированных связывающих элементов, что позволяет осуществить идентификацию связывающих элементов, которые связывают один и тот же эпитоп или перекрывающиеся эпитопы. Такие способы являются хорошо известными среднему специалисту в данной области техники и описаны более подробно в данной заявке. Таким образом, дополнительный аспект в соответствии с настоящим изобретением обеспечивает связывающий элемент, включающий сайт связывания антигена человеческого антитела, которое конкурирует с молекулой антитела, например, в частности, молекулой антитела, включающей VH и/или VL домен CDR, например HCDR3 или набор CDR исходного антитела или любого другого из антител 1-28, для связывания с человеческим IgE. В одном воплощении связывающий элемент в соответствии с изобретением конкурирует с Антителом 11, как представлено в Таблице 1.
Другое воплощение в соответствии с изобретением обеспечивает связывающие элементы, которые связываются со специфическим участком IgE. Связывание может быть определено, например, с помощью определения или наблюдения специфического взаимодействия между связывающим элементом и остатками IgE, например, в структуре связывающего элемента: IgE комплекс, который может быть определен, например, при использовании рентгеновской кристаллографии. Структура Антитела 11, связанного с Cε3-Cε4 доменами человеческого IgE определяется при использовании рентгеновской кристаллографии, обеспечивающей возможность изучения двух взаимодействий Fab антитела 11 с IgE в кристалле. IgE представляет собой бивалентный антиген, поскольку существуют две легких цепи и две тяжелых цепи. Рентгеновское кристаллографическое исследование показало, что Fab связывается с эпитопом, расположенным поперек двух тяжелых цепей IgE.
Первое взаимодействие указывает на то, что сайт взаимодействия Антитела 11 включает остатки от Glu390 до Asn394 включительно и остатки сахаров GlcNAc1 и Man6 в одной тяжелой цепи IgE и Leu340, Arg342, от Ala428 до Thr434 включительно, Thr436, Ser437 и Glu472 и остатки сахара Man 5 в другой тяжелой цепи IgE.
В одном воплощении в соответствии с изобретением обеспечивается изолированный связывающий элемент, специфический для иммуноглобулина Е, где указанный связывающий элемент связывается с эпитопом в иммуноглобулине Е, который включает: остатки от Glu390 до Asn394 включительно в первой тяжелой цепи IgE и Leu340, Arg342, от Ala428 до Thr434, включительно, Thr436, Ser437 и Glu472 во второй тяжелой цепи IgE; в дополнительном воплощении указанный эпитоп дополнительно включает остатки сахаров GlcNAc1 и Man6 первой тяжелой цепи IgE и остатки сахара Man 5 во второй тяжелой цепи IgE.
Второе взаимодействие указывает на то, что сайт взаимодействия Антитела 11 включает остатки Glu390 от Gln392 до Asn394 включительно, и остатки сахаров GlcNAc1 и Man6 в первой тяжелой цепи IgE и Leu340, Arg342, от Ala428 до Thr434 включительно, Thr436, Ser437 и Glu472 во второй тяжелой цепи IgE.
В дополнительном воплощении в соответствии с изобретением обеспечивается изолированный связывающий элемент, специфический для иммуноглобулина Е, где указанный связывающий элемент связывается с эпитопом в иммуноглобулине Е, который включает:
остатки Glu390 от Gln392 до Asn394, включительно, в первой тяжелой цепи IgE и Leu340, Arg342, от Ala428 до Thr434 включительно, Thr436, Ser437 и Glu472 во второй тяжелой цепи IgE;
в дополнительном воплощении указанный эпитоп дополнительно включает остатки сахаров GlcNAc1 и Man6 первой тяжелой цепи IgE.
В дополнительном воплощении в соответствии с изобретением обеспечивается изолированный связывающий элемент, специфический для иммуноглобулина Е, где указанный связывающий элемент связывается с эпитопом в иммуноглобулине Е, который включает:
остатки Glu390 от Gln392 до Asn393 включительно, в первой IgE тяжелой цепи и Leu340, Arg342 от Ala428 до Thr434 включительно, Thr436, Ser437 и Glu472 во второй тяжелой цепи IgE;
указанный эпитоп дополнительно включает остатки сахаров GlcNAc1 и Man6 первой тяжелой цепи IgE.
В дополнительном воплощении в соответствии с изобретением обеспечивается изолированный связывающий элемент, специфический для иммуноглобулина Е, который связывается с эпитопом, который включает элементы из первой тяжелой цепи IgE и элементы из второй тяжелой цепи IgE.
В дополнительных аспектах настоящее изобретение обеспечивает связывающий элемент, включающий сайт связывания антигена человеческого антитела, которое конкурирует с антигенсвязывающим сайтом антитела за связывание с человеческим IgE, где антигенсвязывающий сайт антитела состоит из VH домена и VL домена, и где VH и VL домены включают набор CDR исходного (Антитела 1) или любого из антител 2-28, как раскрыто в данной заявке.
В дополнительных аспектах изобретение обеспечивает изолированную нуклеиновую кислоту, которая включает последовательность, кодирующую связывающий элемент, VH домен и/или VL домен в соответствии с настоящим изобретением. Например, SEQ ID NOS:1, 11, 21, 31, 41, 51, 61, 71, 81, 91, 101, 111, 121, 131, 141, 151, 161, 171, 181, 191, 201, 211, 221, 231, 241, 251, 261, 271, 281, 287, 299 и 305 кодируют типичные VH домены в соответствии с настоящим изобретением, a SEQ ID NOS:317, 319, 321, 323, 325, 327, 329, 331, 333, 335, 337, 339, 341, 343, 345, 347, 349, 351, 353, 355, 357, 359, 361, 363, 365, 367, 369, 371, 373, 375, 377 и 379 кодируют типичные VL домены в соответствии с настоящим изобретением. Изобретение также включает способы получения связывающего элемента, VH домена и/или VL домена в соответствии с изобретением, которые включают экспрессию указанной нуклеиновой кислоты при условиях, которые обеспечивают получение указанного связывающего элемента, VH домена и/или VL домена и восстановление его с помощью изоляции или очистки связывающего элемента.
Другой аспект в соответствии с настоящим изобретением обеспечивает нуклеиновую кислоту, обычно изолированную, которая кодирует VH CDR или VL CDR
последовательности, раскрытые в данной заявке.
Дополнительный аспект обеспечивает хозяйскую клетку, содержащую или трансформированную нуклеиновой кислотой в соответствии с изобретением.
Дополнительные аспекты в соответствии с настоящим изобретением обеспечивают композиции, содержащие связывающие элементы в соответствии с изобретением, и их применение в способах ингибирования и/или нейтрализации IgE, включая способы лечения организма человека или животного с помощью терапии.
Например, связывающие элементы в соответствии с изобретением могут использоваться в способе лечения или предотвращения или использоваться в способе диагностики биологического ответа, заболевания, расстройства или состояния организма человека или животного (например, у пациента, который представляет собой человека) или in vitro.
Способ лечения и/или предотвращения может включать введение указанному пациенту связывающего элемента в соответствии с изобретением в количестве, достаточном для того, чтобы вызвать способную к измерению нейтрализацию IgE. Состояния, которые могут подвергаться лечению в соответствии с настоящим изобретением, включают любое, в котором IgE играет определенную роль, такие как различные виды аллергии и астма.
Эти и другие аспекты в соответствии с изобретением описаны более подробно ниже.
Является приемлемым указать здесь на то, что когда в данной заявке используется "и/или", то это выражение взято как специфическое раскрытие каждого из двух указанных признаков или компонентов с другими или при отсутствии других. Например, "А и/или В" взято как специфическое раскрытие каждого из (i) A, (ii) В и (iii) А и В, точно так же, как если бы каждый предлагался индивидуально в данной заявке.
IgE представляет собой иммуноглобулин Е. Аминокислотная последовательность константного участка человеческого IgE находится в свободном доступе. В некоторых воплощениях IgE может быть IgE человека или обезьяны циномолгуса. Как описывается где-либо в данной заявке, IgE может быть рекомбинантным, и/или может быть либо гликозилированным, либо негликозилированным. IgE экспрессируется естественно in vivo в гликозилированной форме, например в U266.B1 клетках. Гликозилированный IgE может также экспрессироваться в рекомбинантных системах.
Связывающий элемент обычно относится к одному члену пары молекул, которые связываются друг с другом. Члены пары связывания могут быть существующими в природе или полностью или частично полученными синтетически. Один член пары молекул имеет область на своей поверхности, или полость, связывается с и является, таким образом, комплементарной определенной пространственной и полярной организации другого члена пары молекул. Примеры типов пар связывания представляют собой антиген - антитело, биотин - авидин, гормон - рецептор гормона, рецептор - лиганд и фермент - субстрат. Настоящее изобретение относится к типу реакций антиген - антитело.
Связывающий элемент обычно включает молекулу, имеющую сайт связывания антигена. Например, связывающий элемент может представлять собой молекулу антитела или белок, который не происходит от антитела, включающий сайт связывания антигена.
Сайт связывания антигена может обеспечиваться с помощью перестройки CDR белковых каркасных структур, отличных от антитела, таких как фибронектин или цитохром В и др. [1. Haan & Maggos (2004) BioCentury, 12(5): A1-A6; 2. Koide et al. (1998) Journal of Molecular Biology, 284: 1141-1151; 3. Nygren et al. (1997) Current Opinion in Structural Biology, 7: 463-469], или путем рандомизации или мутации аминокислотных остатков петли внутри белковой каркасной структуры для обеспечения связывающей специфичности для желаемой мишени. Каркасные структуры для сконструированных новых сайтов связывания в белках подробно рассматривались Nygren и др. [3. Nygren et al. (1997) Current Opinion in Structural Biology, 7: 463-469]. Белковые каркасные структуры для имитаторов антитела раскрываются в заявке WO/0034784, ссылка на которую введена в данную заявку в своей целостности. В этом документе авторы описывают белки (имитаторы антитела), которые включают домен фибронектина типа III, содержащий, по крайней мере, одну рандомизированную петлю. Приемлемая каркасная структура, в которую пересаживают один или более CDR, например набор HCDR, может обеспечиваться любым доменным членом надсемейства гена иммуноглобулина. Каркасная структура может представлять собой человеческий или нечеловеческий белок. Преимущество белковой каркасной структуры, отличной от антитела, заключается в том, что она может обеспечивать сайт связывания антигена в молекуле каркасной структуры, который имеет меньшие размеры и/или является более легким для производства, чем, по крайней мере, некоторые молекулы антитела. Небольшой размер связывающего элемента может придавать полезные физиологические свойства, такие как способность вводиться в клетки, глубоко проникать в ткани или достигать мишеней, которые находятся в других структурах, или связываться в белковых полостях целевого антигена. Применение сайтов связывания антигена в каркасных структурах белка, отличного от антитела, обсуждается у Wess, 2004 [4. Wess, L. In: BioCentury, The Bernstein Report on BioBusiness, 12(42), A1-A7, 2004]. Типичными являются белки, обладающие стабильным скелетом и одной или более вариабельными петлями, в которых аминокислотная последовательность петли или петель является специфически или случайно мутированной с образованием сайта связывания антигена, который связывается с целевым антигеном. Такие белки включают IgG-связывающие домены белка А из S. aureus, трансферрин, тетранектин, фибронектин (например, 10-й домен фибронектина типа III), липокалины, а также гамма-кристаллины и другие каркасные структуры Affilin™ (Scil белки). Примеры других подходов включают синтетические "микротела" на основе циклотидов - небольшие белки, обладающие внутримолекулярными дисульфидными связями, микробелки (Versabodies™, Amunix) и белки анкириновых повторов (DARРины, Molecular Partners). Такие белки также включают маленькие, сконструированные белковые домены, такие как, например, иммунодомены (смотри, например, патентные публикации US №2003082630 и №2003157561.
В дополнение к последовательностям антитела и/или сайту связывания антигена связывающий элемент в соответствии с настоящим изобретением может включать другие аминокислоты, например образующие пептид или полипептид, такие как складчатый домен, или для придания молекуле другой функциональной характеристики в дополнение к способности связывать антиген. Связывающие элементы в соответствии с изобретением могут нести способную к определению метку или могут быть конъюгированными с токсином или нацеливающим остатком или ферментом (например, с помощью пептидильной связи или линкера). Например, связывающий элемент может включать каталитический сайт (например, в ферментном домене), а также сайт связывания антигена, где сайт связывания антигена связывается с антигеном и, таким образом, нацеливает каталитический сайт на антиген. Каталитический сайт может ингибировать биологическую функцию антигена, например, путем отщепления.
Хотя, как было отмечено ранее, CDR могут содержаться в каркасных структурах, отличных от антитела, структуры для перенесения CDR или набора CDR в соответствии с изобретением будут в общем случае представлять собой последовательность тяжелой или легкой цепи антитела или ее существенную часть, в которой CDR или набор CDR размещается в области, соответствующей CDR или набору CDR существующих в природе вариабельных доменов VH и VL антитела, кодируемых перестроенными генами иммуноглобулина. Структуры и расположение иммуноглобулиновых вариабельных доменов могут быть определены со ссылкой на Kabat и др., 1987 [5. Kabat, E.A. et al. Sequences of Proteins of Immunological Interest. 4th Edition. US Department of Health and Human Services. 1987] и на их уточнения, которые могут быть найдены как "Kabat (Кэбат)" при использовании любой поисковой системы Internet.
С помощью участка CDR или CDR предполагается идентифицировать гипервариабельные участки тяжелой и легкой цепей иммуноглобулина, как определено Kabat и др. 1991 [6. Kabat, Е.А. et al. (1991) Sequences of Proteins of Immunological Interest, 5th Edition. US Department of Health and Human Services, Public Service, NIH, Washington] и более поздними изданиями. Антитело, как правило, содержит 3 CDR тяжелой цепи и 3 CDR легкой цепи. Термин CDR или несколько CDR используется в данной заявке для того, чтобы указать, в соответствии с конкретным случаем, один из этих участков или несколько участков, или даже все эти участки, которые содержат большинство аминокислотных остатков, ответственных за связывание за счет аффинности антитела к антигену или эпитопу, который оно распознает.
Среди шести коротких последовательностей CDR третий CDR тяжелой цепи (HCDR3) имеет большую степень вариабельности (большее разнообразие, существенно благодаря механизмам перестройки генов, которые дают ему начало). Он может быть коротким, например размером 2 аминокислоты, несмотря на то, что самый большой размер, который является известным, составляет 26. Длина CDR может также варьировать в соответствии с длиной, которая может быть приспособлена с помощью специфического лежащего в основе каркасного участка. Функционально HCDR3 отчасти играет определенную роль в определении специфичности антитела [7. Segal et al., PNAS, 71:4298-4302, 1974; 8. Amit et al., Science. 233:747-753, 1986; 9. Chothia et al., J. Mol. Biol., 196:901-917, 1987; 10. Chothia et al., Nature, 342:877-883, 1989; 11. Caton et al., J. Immunol., 144:1965-1968, 1990; 12. Sharon et al., PNAS, 87:4814-4817, 1990; 13. Sharon et al., J. Immunol., 144:4863-4869, 1990; 14. Kabat et al., J. Immunol., 147:1709-1719, 1991]. В другом воплощении в соответствии с изобретением обеспечивается изолированный связывающий элемент, включающий последовательность HCDR3, выбранную из Таблицы 3а.
Термин "молекулы антитела" относится к иммуноглобулину либо природному, либо частично или полностью полученному синтетическим путем. Термин также охватывает любой полипептид или белок, включающий антигенсвязывающий сайт антитела. При этом в данной заявке понятно, что изобретение не относится к антителу в своей природной форме, иными словами, они не находятся в своем природном окружении, но они могут быть изолированы или получены путем очистки из природных источников или же получены с помощью генетической рекомбинации или путем химического синтеза, включая модификацию с помощью неприродных аминокислот. Фрагменты антитела, которые включают антигенсвязыващий сайт антитела, содержат, но без ограничения, молекулы, такие как Fab, Fab', Fab'-SH, scFv, Fv, dAb и Fd. Были сконструированы различные другие молекулы антитела, включающие один или более антигенсвязывающих сайтов антитела, включая, например, Fabz, РаЬз, диатела, триатела, тетратела и минитела. Молекулы антитела и способы их получения, а также их применение являются описанньми в [15. Holliger & Hudson, Nature Biotechnology 23(9):1126-1136 2005].
Также является возможным использовать моноклональные и другие антитела и методики технологии рекомбинантной ДНК для получения других антител или химерных молекул, которые связывают целевой антиген. Такие методики могут также вовлекать введение ДНК, кодирующей вариабельный участок иммуноглобулина, или CDR антитела к константным участкам или константным участкам плюс каркасные участки, отличного иммуноглобулина. Смотри, например, ЕР-А-184187, GB 2188638A или ЕР-А-239400 и целый ряд другой литературы. Гибридома или другая клетка, продуцирующая антитело, может представлять собой объект генетической мутации или других изменений, которые могут или не могут изменять связывание специфичности полученных антител.
Поскольку антитела могут быть модифицированы с помощью ряда способов, термин "молекула антитела" будет трактоваться как такой, который охватывает любой связывающий элемент или вещество, имеющее антигенсвязывающий сайт антитела с требуемыми специфичностью и/или связыванием с антигеном. Таким образом, этот термин охватывает фрагменты антитела и производные, включая любой полипептид, включающий какой-либо антигенсвязывающий сайт антитела, как природный, так и полностью или частично синтетический. Сюда, таким образом, также включаются химерные молекулы, включающие антигенсвязывающий сайт антитела или эквивалент, слитый с другим полипептидом (например, тот, который происходит от других видов или принадлежит к другому классу или подклассу антител). Клонирование и экспрессия химерных антител являются описанными в ЕР-А-0120694 и ЕР-А-0125023, а также в целом ряде другой литературы.
Дополнительные методики, доступные в области конструирования антител, позволили изолировать человеческие и гуманизированные антитела. Например, человеческие гибридомы могут быть получены так, как описано Kontermann и Dubel [16. Kontermann, R & Dubel, S, Antibody Engineering, Springer-Verlag New York, LLC; 2001, ISBN: 3540413545]. Фаговый дисплей, другая общепринятая методика для получения связывающих элементов, был подробно описан во многих публикациях, таких как Kontermann и Dubel и WO92/01047 (дополнительно обсуждается ниже), и патентах US5969108, US5565332, US5733743, US5858657, US5871907. US5872215, US5885793, US5962255, US6140471, US6172197, US6225447, US6291650, US6492160, US6521404.
Трансгенные мыши, у которых гены мышиного антитела являются инактивированными и функционально заменяются генами человеческого антитела, в то время как остаются интактными другие компоненты мышиной иммунной системы, могут использоваться для изоляции человеческих антител [17. Mendez, M. et al. (1997) Nature Genet, 15(2): 146-156]. Гуманизированные антитела могут быть получены при использовании методик, известных в области техники, таких как те, что раскрыты, например, в WO91/09967, US 5,585,089, EP592106, US 565,332 и WO93/17105. Кроме того, WO2004/006955 описывает способы для гуманизации антител, основанные на селекции каркасных последовательностей вариабельного участка из человеческих генов антитела путем сравнения канонических типов структуры CDR для CDR последовательности вариабельного участка антитела вида, отличного от человека, с каноническими типами структуры CDR для соответствующих CDR из библиотеки последовательностей человеческого антитела, например эмбриональных сегментов генов антитела. Вариабельные участки человеческого антитела, обладающими подобными каноническими типами структуры CDR по сравнению с CDR видов, отличных от человека, образуют поднабор членов последовательностей человеческого антитела, из которого выбирают последовательности человеческого каркасного участка. Поднабор членов может быть дополнительно упорядочен при использовании аминокислотной подобности между последовательностями CDR человека и CDR видов, отличных от человека. В способе в соответствии с WO2004/006955 выбирают высоко котирующиеся человеческие последовательности для того, чтобы обеспечить последовательности каркасного участка для конструирования химерного антитела, которое функционально заменяет последовательности CDR человека эквивалентами CDR видов, отличных от человека, при использовании выбранного поднабора членов человеческих каркасных участков, обеспечивая, таким образом, гуманизированное антитело с высокой аффинностью и низкой иммуногенностью при отсутствии необходимости сравнения последовательностей каркасного участка между антителами человека и видов, отличных от человека. Химерные антитела, полученные в соответствии с этим способом, также являются раскрытыми.
Синтетические молекулы антитела могут представлять собой такие, которые созданы путем экспрессии из генов, полученных с помощью олигонуклеотидов, синтезированных и объединенных в рамках приемлемых экспрессионных векторов, например, как описано Knappik и др. [18. Knappik et al. J. Mol. Biol. (2000) 296, 57-86] или Krebs и др. [19. Krebs et al. Journal of Immunological Methods 254 2001 67-84].
Было показано, что фрагменты цельного антитела могут выполнять функцию связывания антигенов. Примеры связывающих фрагментов представляют собой (i) Fab фрагмент, состоящий из VL, VH, CL и СН1 доменов; (ii) Fd фрагмент, состоящий из VH и СН1 доменов; (iii) Fv фрагмент, состоящий из VL и VH доменов одного антитела; (iv) dAb фрагмент [20 Ward, E.S. et al., Nature 341, 544-546 (1989); 21. McCafferty et al (1990) Nature, 348, 552-554; 22. Holt et al (2003) Trends in Biotechnology 21, 484-490], который состоит из VH или VL домена; (v) изолированные CDR участки; (vi) F(ab')2 фрагменты, бивалентный фрагмент, включающий два связанных Fab фрагмента (vii) одноцепочечные Fv молекулы (scFv), где VH домен и VL домен являются соединенными с помощью пептидного линкера, который позволяет двум доменам объединяться с образованием сайта связывания антигена [23. Bird et al. Science, 242, 423-426, 1988; 24. Huston et al, PNAS USA, 85, 5879-5883, 1988]; (viii) биспецифические одноцепочечные Fv димеры (PCT/US92/09965) и (ix) "диатела", мультивалентные или мультиспецифические фрагменты, сконструированные с помощью слияния генов (WO94/13804; [25. Holliger, P. et al, Proc. Natl. Acad. Sci. USA 90 6444-6448, 1993]). Молекулы Fv, scFv или диатело могут быть стабилизированы с помощью введения дисульфидных мостиков, связывающих VH и VL домены [26. Reiter, Y. et al. Nature Biotech, 14, 1239-1245, 1996]. Могут также быть получены минитела, включающие scFv, соединенный с СН3 доменом [27. Hu, S. et al. Cancer Res., 56, 3055-3061, 1996]. Другие примеры связывающих фрагментов представляют собой Fab', который отличается от Fab фрагментов присоединением нескольких остатков на карбокситерминальном конце СН1 домена тяжелой цепи, включая один или более цистеинов из шарнирного участка антитела, и Fab'-SH, который представляет собой Fab' фрагмент, в котором цистеиновый(ые) остаток(остатки) константных доменов несут свободную группу тиола.
Фрагменты антитела в соответствии с изобретением могут быть получены исходя из молекул антитела 1-28, с помощью способов, таких как переваривание ферментами, например, пепсином или папаином и/или путем отщепления дисульфидных мостиков с помощью химического восстановления. В соответствии с другим способом фрагменты антитела, которые включены в настоящее изобретение, могут быть получены с помощью методик генетической рекомбинации, также хорошо известных специалисту в данной области техники, или также путем пептидного синтеза с помощью, например, автоматических пептидных синтезаторов, таких как те, что поставляются компанией Applied Biosystems и др., или путем синтеза и экспрессии нуклеиновой кислоты.
Функциональные фрагменты антитела в соответствии с настоящим изобретением включают любой функциональный фрагмент, чей период полураспада является увеличенным с помощью химической модификации, в частности путем ПЭГилирования, или с помощью встраивания в липосому.
dAb (домен антитела) представляет собой небольшой мономерный связывающий антиген фрагмент антитела, в частности вариабельный участок тяжелой или легкой цепи антитела [22]. VH dAb образуется в природе у верблюжьих (например, у верблюда, ламы) и может быть получен путем иммунизации верблюжьих с помощью целевого антигена, изоляции специфических для антигена В клеток и непосредственного клонирования dAb генов из индивидуальных В клеток. dAb также является возможным получать в культуре клеток. Его небольшой размер, хорошая растворимость и температурная стабильность делают его особенно физиологически полезным и приемлемым для селекции и аффинного созревания. Верблюжьи VH dAb были усовершенствованы для терапевтического применения под названием "нанотела™". Связывающий элемент в соответствии с настоящим изобретением может представлять собой dAb, включающий VH или VL домен, существенно такой, как установлено в данной заявке, или VH, или VL домен, включающий набор CDR, существенно такой, как установлено в данной заявке.
Биспецифические или бифункциональные антитела образуют вторую генерацию моноклональных антител, в которых два различных вариабельных участка соединены в одной и той же молекуле [28. Holliger and Bohlen 1999 Cancer and metastasis rev. 18:411-419]. Их применение было продемонстрировано как в области диагностики, так и в области терапии благодаря их способности привносить новые эффекторные функции или нацеливать некоторые молекулы на поверхность опухолевой клетки. Если используются биспецифические антитела, то они могут представлять собой традиционные биспецифические антитела, которые могут быть получены с помощью различных способов [29. Holliger, P. and Winter G. Current Opinion Biotechnol. 4, 446-449 1993], например, получены химически или из гибридных гибридом, или могут представлять собой любой из биспецифических фрагментов антитела, упомянутых выше. Эти антитела могут быть получены с помощью химических способов [30. Glennie M J et al., 1987 J. Immunol. 139, 2367-2375; 31. Repp R. et al., 1995 J. Hemat. 377-382] или соматических способов [32. Staerz U.D. and Bevan M. J. 1986 PNAS 83; 33. Suresh M.R. et al., 1986 Method Enzymol. 121: 210-228], но также и предпочтительно с помощью методик генетического конструирования, которые позволяют индуцировать гетеродимеризацию и, таким образом, улучшить процесс очистки желаемого антитела [34. Merchand et al., 1998 Nature Biotech. 16:677-681]. Примеры биспецифических антител включают такие в соответствии с технологией BiTE™, в которой можно использовать связывание доменов двух антител с различными специфичностями и непосредственно связывать их с помощью коротких гибких пептидов. Это позволяет объединять два антитела на одной короткой полипептидной цепи. Диатела и scFv могут быть сконструированы без Fc участка, при использовании только вариабельных доменов, что потенциально снижает эффекты антиидиотипической реакции.
Биспецифические антитела могут быть сконструированы как цельный IgG, как биспецифический Fab'2, как Fаb'ПЭГ, как диатела или также как биспецифические scFv. Кроме того, два биспецифических антитела могут быть связаны при использовании обычных способов, известных в области техники, с образованием тетравалентных антител.
Биспецифические диатела, в отличие от биспецифических цельных антител, могут также быть особенно полезными, поскольку они могут быть легко сконструированы и экспрессированы в E.coli. Диатела (и многие другие полипептиды, такие как фрагменты антитела) с приемлемыми связывающими специфичностями могут быть легко отобраны при использовании фагового дисплея (WO94/13804) из библиотек. Если одно плечо диатела остается константным, например, со специфичностью, направленной против IgE, то библиотека может быть получена, когда другое плечо подвергается вариации, и отбирают антитело приемлемой специфичности. Биспецифические цельные антитела могут представлять собой такие, полученные с помощью альтернативных способов конструирования, как описано у Ridgeway и др., 1996 [35. Ridgeway, J. B. B. et al. Protein Eng., 9, 616-621, 1996].
Различные способы являются доступными в области техники для получения антител против IgE. Антитела могут представлять собой моноклональные антитела, в частности человеческого, мышиного, химерного или гуманизированного происхождения, которые могут быть получены в соответствии со стандартными способами, хорошо известными специалисту в данной области техники.
В общем случае, для получения моноклональных антител или их функциональных фрагментов, в частности таких мышиного происхождения, является возможным обратиться к методикам, которые являются описанными, в частности в руководстве "Antibodies" [36. Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor N.Y., pp.726, 1988] или к методике получения из гибридом, описанной Kohler и Milstein [37. Kohler and Milstein, Nature, 256:495-497, 1975].
Моноклональные антитела могут быть получены, например, из клетки животного, иммунизированной IgE или одним из его фрагментов, содержащим эпитоп, который узнается указанным моноклональным антителом. Приемлемые фрагменты и пептиды или полипептиды, включающие их, описываются в данной заявке и могут использоваться для иммунизации животных с целью получения антител против IgE. Указанный IgE или один из его фрагментов может, в частности, быть получен в соответствии с обычными рабочими способами, путем генетической рекомбинации, исходя из последовательности нуклеиновой кислоты, содержащейся в последовательности кДНК, кодирующей IgE или его фрагмент, с помощью пептидного синтеза исходя из последовательности аминокислот, содержащейся в пептидной последовательности IgE и/или ее фрагменте.
Моноклональные антитела могут быть, например, очищены на аффинной колонке, на которой был предварительно иммобилизован IgE или один из его фрагментов, содержащий эпитоп, который узнается указанным моноклональным антителом. В частности, моноклональные антитела могут быть очищены с помощью хроматографии на белке А и/или G, после чего осуществляют или не осуществляют ионообменную хроматографию, направленную на устранение остаточной белковой контаминации, а также ДНК и LPS, как таковых, после чего осуществляют или не осуществляют вытеснительную хроматографию на сефарозном геле для того, чтобы устранить потенциальные агрегаты благодаря присутствию димеров или других мультимеров. В одном воплощении все эти методики могут использоваться одновременно или последовательно.
Антигенсвязывающий сайт представляет собой часть молекулы, которая связывается с и является комплементарной всему или части целевого антигена. В молекуле антитела этот термин относится к антигенсвязывающему сайту антитела и включает часть антитела, которая связывается с и является комплементарной всему или части целевого антигена. Если антиген является большим, то антитело может связываться только с определенной частью антигена, при этом такая часть называется эпитопом. Антигенсвязывающий сайт антитела может обеспечиваться одним или более вариабельными доменами антитела. Антигенсвязывающий сайт антитела может включать вариабельный участок легкой цепи антитела (VL) и вариабельный участок тяжелой цепи антитела (VH).
Изолированный относится к состоянию, в котором связывающие элементы в соответствии с изобретением или нуклеиновая кислота, кодирующая такие связывающие элементы, будет в общем случае представлять собой такую в соответствии с изобретением. Таким образом, связывающие элементы, VH и/или VL домены, кодирующие молекулы нуклеиновой кислоты и векторы в соответствии с настоящим изобретением могут обеспечиваться изолированными и/или очищенными, например из их природной среды, в существенно чистой или гомогенной форме или, в случае нуклеиновой кислоты, свободными или существенно свободными от нуклеиновой кислоты или генов, имеющих происхождение, отличное от последовательности, кодирующей полипептид с требуемыми функциями. Изолированные элементы и изолированная нуклеиновая кислота будут свободными или существенно свободными от материала, с которым они являются природным образом ассоциированными, такого как другие полипептиды или нуклеиновые кислоты, с которыми их обнаруживают в естественном окружении, или окружающей среды, в которой они получены (например, культура клеток), когда такое получение осуществляют с помощью технологии рекомбинантной ДНК, осуществляемой in vitro или in vivo. Элементы и нуклеиновая кислота могут представлять собой такие, рецептированные с разбавителями или вспомогательными веществами, и все еще оставаться для практических целей изолированными, например элементы будут обычно смешиваться с желатином или другими носителями, если используются для покрытия микротитровальных планшетов для применения в иммуноанализах или будут смешиваться с фармацевтически приемлемыми носителями или разбавителями, когда используются для диагностики или терапии. Связывающие элементы могут быть гликозилированными либо естественным путем, либо с помощью систем гетерологичных эукариотических клеток (например, СНО или NS0 (ЕСАСС 85110503) клеток), или они могут быть (например, тогда, когда получены с помощью экспрессии в прокариотической клетке) негликозилированными.
Гетерогенные препараты, включающие молекулы антитела анти-IgE, также образуют часть в соответствии с изобретением. Например, такие препараты могут представлять собой смеси антител с тяжелыми цепями полной длины и тяжелыми цепями с отсутствующим С-терминальным лизином, с различной степенью гликозилирования и/или с дериватизованными аминокислотами, такими как циклизация N-терминальной глутаминовой кислоты с образованием остатка пироглутаминовой кислоты.
Как используется в данной заявке, фраза "существенно так, как изложено" относится к характеристике(ам) релевантных CDR VH или VL домена связывающих элементов, описанных в данной заявке, которые будут либо идентичными, либо в высокой степени подобными указанным участкам, последовательность которых представлена в данной заявке. Как описано в данной заявке, фраза "в высокой степени подобные" в отношении указанного(ых) участка(ов) одного или более вариабельных доменов подразумевает, что от 1 до приблизительно 6, например от 1 до 5, включая от 1 до 3, или 1, или 2, или 3, или 4 аминокислотные замены, могут представлять собой такие, которые осуществлены в CDR и/или VH или VL домене.
Подробное описание
Как было указано выше, связывающий элемент в соответствии с настоящим изобретением модулирует и может нейтрализовать биологическую активность IgE. Как описывается в данной заявке, связывающие IgE элементы в соответствии с настоящим изобретением могут быть оптимизированы по нейтрализующей эффективности. В общем случае оптимизация активности вовлекает мутирование последовательности выбранного связывающего элемента (обычно последовательности вариабельного домена антитела) для получения библиотеки связывающих элементов, которые потом подвергают анализу на активность и отбирают более активные связывающие элементы. Таким образом, отобранные "оптимизированные по эффективности" связывающие элементы имеют тенденцию к более высокой эффективности, чем связывающий элемент, из которого была получена библиотека. Тем не менее, высокая эффективность связывающих элементов может представлять собой такую, полученную без оптимизации, например высокая эффективность связывающего элемента может быть получена непосредственно при осуществлении исходного скрининга, например биохимического анализа нейтрализации. "Оптимизированная активность" связывающего элемента относится к связывающему элементу с оптимизированной активностью нейтрализации определенной активности или с понижающей функцией. Анализы и эффективности являются описанными более подробно где-либо в данной заявке. Настоящее изобретение обеспечивает как оптимизированные по эффективности, так и не оптимизированные связывающие элементы, а также способы для оптимизации эффективности для выбранного связывающего элемента. Настоящее изобретение, таким образом, позволяет квалифицированному в данной области специалисту получить связывающие элементы, обладающие высокой эффективностью.
Несмотря на то, что оптимизация эффективности может использоваться для получения связывающих элементов с более высокой эффективностью из данного связывающего элемента, также следует отметить, что связывающие элементы с высокой эффективностью могут быть получены даже без оптимизации эффективности.
В дополнительном аспекте настоящее изобретение обеспечивает способ получения одного или более связывающих элементов, способных связываться с антигеном, при этом способ включает приведение в контакт библиотеки связывающих элементов в соответствии с изобретением и указанного антигена, а также отбор одного или более связывающих элементов библиотеки, способных связываться с указанным антигеном.
Библиотека может быть просмотрена на частицах или молекулярных комплексах, например на способных к репликации генетических модулях, таких как дрожжи, бактерии или частицы бактериофагов (например, Т7), вирусы, клетки или ковалентные рибосомальные или другие in vitro системы дисплея, каждая частица или молекулярный комплекс, содержащие нуклеиновую кислоту, кодирующую VH вариабельный домен антитела просматриваются на ней, и необязательно просматривается также VL домен, в случае его присутствия. Фаговый дисплей является описанным в WO92/01047 и, например, патентах US5969108, US5565332, US5733743, US5858657, US5871907, US5872215, US5885793, US5962255, US6140471, US6172197, US6225447, US6291650, US6492160 и US6521404, каждый из которых введен как ссылка в данную заявку в своей целостности.
После селекции связывающих элементов, способных связываться с антигеном и просмотренных на бактериофаге или библиотеке других частиц или молекулярных комплексов, нуклеиновая кислота может быть заимствована из бактериофага или другой частицы или молекулярного комплекса, показывающего указанный выбранный связывающий элемент. Такая нуклеиновая кислота может использоваться при последующем получении связывающего элемента или VH или VL вариабельного домена антитела путем экспрессии нуклеиновой кислоты с последовательностью нуклеиновой кислоты, заимствованной из бактериофага, или другой частицы, или молекулярного комплекса, показывающего указанный выбранный связывающий элемент.
Вариабельный домен VH антитела с аминокислотной последовательностью вариабельного домена VH антитела указанного выбранного связывающего элемента может обеспечиваться в изолированной форме как связывающий элемент, включающий такой VH домен.
Способность связываться с IgE может быть дополнительно проанализирована, может также быть определена способность конкурировать, например, с исходной молекулой антитела или молекулой антитела 2-28 (например, в scFv формате и/или IgG формате, например, IgG1) за связывание с IgE. Способность нейтрализовать IgE может быть проанализирована, как обсуждается дополнительно в каком-либо месте в данной заявке.
Связывающий элемент в соответствии с настоящим изобретением может нейтрализовать биологическую активность IgE с эффективностью исходной молекулой антитела или другой молекулы антитела, одного из антител 2-28, например scFv или IgG1, или с более высокой эффективностью.
Связывающая аффинность и эффективность нейтрализации различных связывающих элементов может сравниваться при приемлемых условиях.
Варианты VH и VL доменов и CDR в соответствии с настоящим изобретением, включая те, для которых аминокислотные последовательности являются изложенными в данной заявке и которые могут использоваться в связывающих элементах для IgE, могут быть получены с помощью способов изменения последовательности или мутации и скрининга для антигенсвязывающих элементов с желаемыми характеристиками. Примеры желаемых характеристик включают, но не ограничены, следующие:
- Повышенная связывающая аффинность для антигена по сравнению с известными антителами, которые являются специфическими для антигена.
- Повышенная нейтрализация активности антигена по сравнению с известными антителами, которые являются специфическими для антигена, если такая является известной.
- Предусмотренная конкурентоспособность с известным антителом или лигандом за антиген при определенном молярном соотношении.
- Способность к иммунопреципитации комплекса.
- Способность связываться с указанным эпитопом.
- Линейным эпитопом, например, пептидной последовательностью, идентифицированной при использовании сканирования пептидного связывания, как описано в данной заявке, например, при использовании просеянных пептидов, в линейной и/или изогнутой конформации.
- Конформационным эпитопом, образованным разомкнутыми остатками.
- Способность модулировать новую биологическую активность IGE или молекулы с понижающей регуляцией. Такие способы также обеспечиваются в данной заявке.
Варианты молекулы антитела, раскрытые в данной заявке, могут быть получены и использованы в настоящем изобретении. Следуя руководству по вычислительной химии при применении методик анализа многовариантных данных к взаимоотношениям структура/отличительная особенность - активность [38. Wold, et al. Multivariate data analysis in chemistry. Chemometrics-Mathematics and Statistics in Chemistry (Ed.: B. Kowalski), D. Reidel Publishing Company, Dordrecht, Holland, 1984 (ISBN 90-277-1846-6)] могут быть выведены количественные взаимоотношения отличительной особенности - активности антител при использовании хорошо известных математических методик, таких как статистическая регрессия, распознание и классификация изображений [39. Norman et al. Applied Regression Analysis. Wiley-Interscience; 3rd edition (April 1998) ISBN: 0471170828; 40. Kandel, Abraham & Backer, Eric. Computer-Assisted Reasoning in Cluster Analysis. Prentice Hall PTR, (May 11,1995), ISBN: 0133418847; 41. Krzanowski, Wojtek. Principles of Multivariate Analysis: A User's Perspective (Oxford Statistical Science Series, No 22 (Paper)). Oxford University Press; (December 2000), ISBN: 0198507089; 42. Witten, Ian H. & Frank, Eibe. Data Mining: Practical Machine Learning Tools and Techniques with Java Implementations. Morgan Kaufmann; (October 11, 1999), ISBN: 1558605525; 43. Denison David G. T. (Editor), Christopher C. Holmes, Bani K. Mallick, Adrian F. M. Smith. Bayesian Methods for Nonlinear Classification and Regression (Wiley Series in Probability and Statistics). John Wiley & Sons; (July 2002), ISBN: 0471490369; 44. Ghose, Arup K. & Viswanadhan, Vellarkad N.. Combinatorial Library Design and Evaluation Principles, Software, Tools, and Applications in Drug Discovery. ISBN: 0-8247-0487-8]. Свойства антител могут быть выведены из эмпирических и теоретических моделей (например, анализа вероятно контактирующих остатков или рассчитанного физико-химического свойства) последовательности антитела, функциональной и пространственной структур и их свойств, которые могут рассматриваться отдельно или в комбинации.
Антигенсвязывающий сайт антитела, состоящий из VH домена и VL домена, является, как правило, состоящим из шести петель полипептида: три из вариабельного домена легкой цепи (VL) и три из вариабельного домена тяжелой цепи (VH). Анализ антител с известной атомной структурой определяет взаимоотношения между последовательностью и пространственной структурой антигенсвязывающих активных центров антитела [45. Chothia C. et al. Journal Molecular Biology (1992) 227, 799-817, 46 Al-Lazikani, et al. Journal Molecular Biology (1997) 273(4), 927-948]. Эти взаимоотношения предполагают, что, за исключением третьего участка (петли) в VH доменах, петли связывающего сайта имеют одну из небольшого числа основных конформаций цепи: канонические структуры. Канонические структуры, образовавшиеся в определенной петле, как было показано, определяются своими размерами и присутствием определенных остатков в ключевых сайтах как в петле, так и в каркасных участках [45. Chothia C. et al. Journal Molecular Biology (1992) 227, 799-817; 46. Al-Lazikani, et al. Journal Molecular Biology (1997) 273(4), 927-948].
Это исследование взаимоотношения последовательности - структуры могут использоваться для предсказания тех остатков в антителе известной последовательности, но с неизвестной пространственной структурой, которые являются важными для поддержания пространственной структуры его петель CDR и, таким образом, поддерживают связывающую специфичность. Эти предсказания могут подкрепляться с помощью сравнения предсказаний и результата, полученного путем экспериментов по оптимизации. В структурном подходе может быть создана модель молекулы антитела [47. Chothia, et al. Science, 223, 755-758 (1986)] при использовании любого свободно доступного или коммерческого пакета программ, такого как WAM [48. Whitelegg, N.R.u. and Rees, A.R (2000). Prot. Eng., 12, 815-824]. Программное обеспечение для визуализации и анализа белка, такое, как Insight II (Accelrys, Inc.) или Deep View [49. Guex, N. and Peitsch, M.C. Electrophoresis (1997) 18, 2714-2723] может потом использоваться для оценки возможных замен в каждом положении CDR. Эта информация может потом использоваться для осуществления замен, возможно имеющих минимальное или благоприятное влияние на активность.
Методики, необходимые для осуществления замен в аминокислотной последовательности CDR, VH или VL доменах антитела и связывающих элементах в общем случае являются доступными в области техники. Вариант последовательностей может быть получен с помощью замен, которые могут или не могут быть предсказаны как такие, которые имеют минимальное или благоприятное влияние на активность, и анализируются на способность связывать и/или нейтрализовать IgE и/или на любое другое желаемое свойство.
Варианты аминокислотной последовательности вариабельного домена любого из VH и VL доменов, чьи последовательности являются, в частности, раскрытыми в данной заявке, могут использоваться в соответствии с настоящим изобретением, как обсуждалось выше.
Дополнительный аспект в соответствии с изобретением представляет собой молекулу антитела, включающую VH домен, который имеет, по крайней мере, 60, 70, 80, 85, 90, 95, 98 или 99%-ную идентичность с аминокислотной последовательностью VH домена антитела 1, показанного в прилагаемом списке последовательностей, и/или, включающий VL домен, который имеет, по крайней мере, 60, 70, 80, 85, 90, 95, 98 или 99%-ную идентичность с аминокислотной последовательностью VL домена антитела 1, показанного в прилагаемом списке последовательностей. Алгоритмы, которые могут использоваться для подсчета % идентичности двух аминокислотных последовательностей, включают например, BLAST [50. Altschul et al. (1990) J. Mol. Biol. 215:405-410], FASTA [51. Pearson and Lipman (1988) PNAS USA 85:2444-2448], или алгоритм Смита-Ватермана [52. Smith and Waterman (1981) J. Mol Biol. 147:195-197], например, при использовании параметров по умолчанию.
Особые варианты могут включать одно или более изменений аминокислотной последовательности (добавление, делению, замену и/или инсерцию аминокислотного остатка). В некоторых воплощениях варианты имеют менее чем приблизительно 20 изменений.
Изменения могут представлять собой такие, которые осуществлены в одном или более каркасных участках и/или одном или более CDR. Изменения обычно не приводят к потере функции, так что связывающий элемент, включающий таким образом измененную аминокислотную последовательность, может сохранять способность связывать и/или нейтрализовать IgE. Они могут сохранять то же количественное значение связывающей и/или нейтрализующей способности, что и связывающий элемент, в котором не было осуществлено изменение, например, как измеряется в анализе, описанном в данной заявке. Связывающий элемент, включающий измененную таким образом аминокислотную последовательность, может иметь улучшенную способность связывать и/или нейтрализовать IgE.
Изменение может включать замену одного или более аминокислотного (ых) остатка(ов) несуществующими в природе или нестандартными аминокислотами, модификацию одного или более аминокислотных остатков на несуществующую в природе или нестандартную форму или встраивание одной или более несуществующих в природе или нестандартных аминокислот в последовательность. Примеры номеров и локализации изменений в последовательности в соответствии с изобретением являются описанными где-либо в данной заявке. Существующие в природе аминокислоты включают 20 "стандартных" L-аминокислот, идентифицированных как G, А, V, L, I, M, P, F, W, S, Т, N, Q, Y, С, K, R, Н, D, Е в соответствии с их стандартными однобуквенными кодами. Нестандартные аминокислоты включают любой другой остаток, который может представлять собой встроенный в полипептидный скелет или результат модификации существующих аминокислотных остатков. Нестандартные аминокислоты могут представлять собой существующие в природе или несуществующие в природе. Некоторые существующие в природе нестандартные аминокислоты являются известными в области техники, такие как 4-гидроксипролин, 5-гидроксилизин, 3-метилгистидин, N-ацетилсерин и т.д. [53. Voet & Voet, Biochemistry, 2nd Edition, (Wiley) 1995.]. Те аминокислотные остатки, которые являются дериватизованными в своих N-альфа положениях, будут располагаться только на N-терминальном конце аминокислотной последовательности. Обычно в настоящем изобретении аминокислота представляет собой L-аминокислоту, но она может представлять собой D-аминокислоту. Изменение может, таким образом, включать модификацию L-аминокислоты на или замену ее на D-аминокислоту. Метилированные, ацетилированные и/или фосфорилированные формы аминокислот являются также известными, аминокислоты в настоящем изобретении могут представлять собой предмет такой модификации.
Аминокислотные последовательности в доменах антитела и связывающих элементах в соответствии с изобретением могут включать неприродные или нестандартные аминокислоты, описанные выше. Нестандартные аминокислоты (например, D-аминокислоты) могут представлять собой встроенные в аминокислотную последовательность во время синтеза или полученные путем модификации или замены "исходных" стандартных аминокислот после синтеза аминокислотной последовательности.
Применение нестандартных и/или не существующих в природе аминокислот повышает структурное и функциональное разнообразие и может, таким образом, повышать потенциал для достижения желаемых свойств связывания IgE и нейтрализации в связывающем элементе в соответствии с изобретением. Кроме того, D-аминокислоты и аналоги были продемонстрированы как такие, которые обладают различными фармакокинетическими профилями по сравнению со стандартными L-аминокислотами, благодаря in vivo разложению полипептидов, содержащих L-аминокислоты, после введения животному, например человеку.
Новые VH или VL участки, несущие производные CDR последовательностей в соответствии с изобретением, могут быть получены при использовании случайного мутагенеза одного или более выбранных VH и/или VL генов для получения мутаций в пределах вариабельного домена. Такая методика описана Gram и др. [54. Gram et al., 1992, Proc. Natl. Acad. Sci., USA. 89:3576-3580], которые использовали подверженную ошибкам ПЦР. В некоторых воплощениях одна или две аминокислотные замены осуществляют в пределах цельного вариабельного домена или набора CDR.
Другой способ, который может использоваться, представляет собой направленный на CDR участки VH или VL генов мутагенез. Такие методики являются раскрытыми Barbas и др. [55. Barbas et al., 1994, Proc. Natl. Acad. Sci., USA, 91:3809-3813] и Schier и др. [56. Schier et al., 1996, J. Mol. Biol. 263:551-567].
Все описанные выше методики являются известными в данной области техники, и квалифицированный специалист будет в состоянии использовать эти методики для обеспечения связывающих элементов в соответствии с изобретением при использовании обычной методологии в уровне техники.
Дополнительный аспект в соответствии с изобретением обеспечивает способ для получения антигенсвязывающего сайта антитела для IgE, способ, включающий обеспечение с помощью прибавления, делеции, замены или инсерции одной или более аминокислот в аминокислотной последовательности VH домена, представленного в данной заявке, VH домена, который представляет собой вариант аминокислотной последовательности VH домена, необязательное соединение VH домена, обеспеченного таким образом, с одним или более VL доменами и анализ VH домена, или VH/VL комбинации, или комбинаций для идентификации связывающего элемента или антигенсвязывающего сайта антитела для IgE и необязательно с одной или более желаемыми характеристиками, например способностью к нейтрализации активности IgE. Указанный VL домен может иметь аминокислотную последовательность, которая является существенно представленной в данной заявке. Может использоваться аналогичный способ, в котором один или более вариантов последовательности VL домена, раскрытых в данной заявке, объединяют с одним или более VH доменами.
Как указано выше, аминокислотная последовательность CDR существенно такая, как представлено в данной заявке, может содержаться в виде CDR в вариабельном домене человеческого антитела или его существенной части. HCDR3 последовательности, существенно такие, как представлено в данной заявке, представляют собой воплощения в соответствии с настоящим изобретением и каждая из них может содержаться в виде HCDR3 в вариабельном домене тяжелой цепи человека или его существенной части.
Вариабельные домены, используемые в данном изобретении, могут представлять собой полученные, или происходящие из любой эмбриональной линии, или полученные путем перестройки человеческого вариабельного домена, или могут представлять собой синтетический вариабельный домен на основе консенсусных или фактически существующих последовательностей известных вариабельных доменов человека. Вариабельный домен может быть получен из антитела вида, отличного от человека. CDR последовательность в соответствии с изобретением (например, CDR3) может представлять собой встроенную в репертуар вариабельных доменов, у которых отсутствует CDR (например, CDR3), при использовании технологии рекомбинантной ДНК. Например, Marks и др. [57. Marks et al Bio/Technology, 1992, 10:779-783] описывают способы получения репертуаров вариабельных доменов антитела, в которых консенсусные праймеры, направленные к или соседствующие с 5' концом участка вариабельного домена используются в соединении с консенсусными праймерами к третьему каркасному участку человеческих VH генов для обеспечения репертуара VH вариабельных доменов, в которых отсутствует CDR3. Marks и др. дополнительно описали, как этот репертуар может быть соединен с CDR3 определенного антитела. При использовании аналогичных методик происходящие от CDR3 последовательности в соответствии с настоящим изобретением могут быть перетасованы с репертуарами VH или VL доменов, в которых отсутствует CDR3, и перетасованные полные VH или VL домены соединяют с родственными VL или VH доменами для обеспечения связывающих элементов в соответствии с изобретением. Репертуары могут потом просматриваться в подходящей системе хозяина, такой как фаговая система дисплея в соответствии с заявкой WO92/01047, которая введена в данную заявку как ссылка в своей целостности, или с любым последующим литературным источником, включая Kay, Winter и McCafferty [58. Kay, В.K., Winter, J., and McCafferty, J. (1996) Phage Display of Peptides and Proteins: A Laboratory Manual, San Diego: Academic Press], так, что могут быть отобраны приемлемые связывающие элементы. Репертуар может состоять из любого числа от 104 индивидуальных членов и выше, например, по крайней мере, 105, по крайней мере, 106, по крайней мере, 107, по крайней мере, 108, по крайней мере, 109, или, по крайней мере, 1010 членов или более. Другие приемлемые системы хозяина включают, но не ограничены, дрожжевой дисплей, бактериальный дисплей, Т7 дисплей, вирусный дисплей, клеточный дисплей, рибосомальный дисплей и ковалентный дисплей.
Обеспечивается способ получения связывающего элемента для IgE антигена, который включает:
(a) обеспечение исходного репертуара нуклеиновых кислот, кодирующих VH домен, который либо включает CDR3, подвергающийся замене, либо в котором отсутствует участок, кодирующий CDR3;
(b) соединение указанного репертуара с донорной нуклеиновой кислотой, кодирующей аминокислотную последовательность, существенно такую, как представлено в данной заявке для VH CDR3, так, что указанная донорная нуклеиновая кислота встраивается в CDR3 участок в репертуаре с целью обеспечения репертуарного продукта нуклеиновых кислот, кодирующих VH домен;
(c) экспрессию нуклеиновых кислот указанного репертуарного продукта;
(d) селекцию связывающего элемента для IgE;
(e) восстановление указанного связывающего элемента или нуклеиновой кислоты, которая кодирует его.
Кроме того, может использоваться аналогичный способ, в котором VL CDR3 в соответствии с изобретением соединяют с репертуаром нуклеиновых кислот, кодирующих VL домен, который либо включает CDR3, подвергающийся замене, либо в котором отсутствует участок, кодирующий CDR3.
Подобно этому один, или более, или все три CDR могут вводиться в репертуар VH или VL доменов, которые потом подвергают скринингу на связывающий элемент или связывающие элементы для IgE.
Например, могут использоваться один или более HCDR1, HCDR2 и HCDR3 исходного антитела или антитела 2-28, или набор HCDR исходного антитела или антитела 2-28, и/или могут использоваться один или более LCDR1, LCDR2 и LCDR3 исходного антитела или антитела 2-28, или набор LCDR исходного антитела или антитела 2-28.
Подобно этому могут использоваться VH и VL домены, наборы CDR и наборы HCDR и/или наборы LCDR, раскрытые в данной заявке.
Существенная часть вариабельного домена иммуноглобулина может включать, по крайней мере, три CDR участка, вместе с их промежуточными каркасными участками. Часть может также включать, по крайней мере, приблизительно 50 % любого из двух либо обоих первого и четвертого каркасных участков, при этом 50 % являются С-терминальными 50 % первого каркасного участка, а N-терминальные 50 % являются такими четвертого каркасного участка. Дополнительные остатки на N-терминальном или С-терминальном конце существенной части вариабельного домена могут представлять собой такие, которые обычно не ассоциируются с существующими в природе участками вариабельного домена. Например, конструирование связывающих элементов в соответствии с настоящим изобретением, осуществленное с помощью методик рекомбинантной ДНК, может приводить к встраиванию N- или С-терминальных остатков, кодируемых линкерами, введенными для способствования клонированию и другим этапам манипуляции. Другие этапы манипуляции включают введение линкеров для соединения вариабельных доменов в соответствии с изобретением с дополнительными белковыми последовательностями, включая константные участки антитела, другие вариабельные домены (например, при получении диатела) или способные к выявлению /функциональные метки, как обсуждается более подробно где-либо в данной заявке.
Несмотря на то, что в некоторых аспектах в соответствии с изобретением связывающие элементы включают пару VH и VL доменов, одиночные связывающие домены на основе либо последовательности VH, либо VL домена, образуют дополнительные аспекты в соответствии с изобретением. Является известным, что одиночные иммуноглобулиновые домены, в частности VH домены, способны к связыванию целевых антигенов специфическим образом. Например, смотри обсуждение dAb, приведенное выше.
В случае какого-либо одиночного из связывающих доменов эти домены могут использоваться для скрининга на комплементарные домены, способные к образованию двухдоменного связывающего элемента, способного к связыванию с IgE. Это может быть получено способами скрининга из фагового дисплея при использовании так называемой иерархиального двойного комбинаторного подхода, как раскрыто в заявке WO92/01047, введенной в данную заявку в качестве ссылки в своей целостности, при котором индивидуальная колония, содержащая какой-либо из клонов Н или L цепи, используется для инфицирования полной библиотеки клонов, кодирующих другие цепи (L или Н), и полученный двухцепочечный связывающий элемент подвергают селекции в соответствии с методиками фагового дисплея, такими, как те, что описаны в данной ссылке. Эта методика является также раскрытой у Marks и др., выше.
Связывающие элементы в соответствии с настоящим изобретением могут дополнительно включать константные участки антитела или их части, например константные участки человеческого антитела или их части. Например, VL домен может присоединяться на своем С-терминальном конце к константным доменам легкой цепи антитела, включая человеческие Cκ или Cλ цепи. Подобно этому связывающий элемент на основе VH домена может присоединяться на своем C-терминальном конце ко всей или части (например, СН1 домену) тяжелой цепи иммуноглобулина, имеющего происхождение от любого изотипа антитела, например IgG, IgA, IgE и IgM и любого из подклассов изотипа, в частности IgG1 и IgG4. IgG1 является предпочтительным благодаря своей эффекторной функции и простоте производства. Любой синтетический или другой вариант константного участка, который обладает такими свойствами и стабилизирует вариабельные участки, может также быть полезным в настоящем изобретении.
Связывающие элементы в соответствии с изобретением могут представлять собой меченные с помощью способной к определению или функциональной метки. Таким образом, связывающий элемент или молекула антитела могут присутствовать в форме иммуноконъюгата так, чтобы получить способный к определению и/или к количественному определению сигнал. Иммуноконъюгат может включать молекулу антитела в соответствии с изобретением, конъюгированную со способной к определению или функциональной меткой. Метка может представлять собой любую молекулу, которая вырабатывает или может индуцировать выработку сигнала, включая, но без ограничения, флуоресцирующие вещества, радиоизотопные метки, ферменты, хемилюминисцентные вещества или фотосенсибилизирующие вещества. Таким образом, связывание может представлять собой обнаруживаемую и/или измеряемую путем определения флуоресценцию или люминисценцию, радиоактивность, ферментативную активность или поглощение света.
Приемлемые метки включают, например, но без ограничения,
- ферменты, такие, как щелочная фосфатаза, глюкоза-6-фосфат дегидрогеназа ("G6PDH"), альфа-D-галактозидаза, глюкозаоксидаза, глюкозамилаза, карбоангидраза, ацеталхолинэстераза, лизоцим, малатдегидрогеназа и пероксидаза, например пероксидаза хрена;
- красители;
- флуоресцентные метки и флуоресцирующие вещества, такие как флуоресцеин и его производные, флуорохром, соединения родамина и производные, GFP (GFP является сокращением для "зеленого флуоресцентного белка"), данзил, умбеллиферон, фикоэритрин, фикоцианин, аллофикоцианин, о-фталевый альдегид и флуорескамин; флуорофоры, такие как лантанидкриптаты и хелаты, например, европий и другие (Perkin Elmer и Cis Biointernational),
- хемолюминисцентные метки или хемилюминисцирующие вещества, такие как изолюминол, люминол и диоксетаны;
- биолюминисцентные метки, такие как люцифераза и люциферин;
- сенсибилизирующие вещества;
- коферменты;
- ферментные субстраты;
- радиоактивные метки, включая, но без ограничения, бром77, углерод14, кобальт57, фтор8, галлий67, галлий68, водород3 (тритий), индий111, индий113m, йод123m, йод125, йод126, йод131, йод133, ртуть107, ртуть203, фосфор32, рений99m, рений101, рений105, рутений95, рутений97, рутений103, рутений105, скандий47, селен75, сера35, технеций99, технеций99m, теллур121m, теллур122m, теллур125m, тулий165, тулий167, тулий168, иттрий 199 и другие радиоактивные метки, упомянутые в данной заявке;
- частицы, такие как латексные и углеродные частицы; соли металлов; кристаллоиды; липосомы; клетки, и т.д., которые могут быть дополнительно меченными с помощью красителя, катализатора или других способных к определению групп;
- молекулы такие, как биотин, дигоксигенин или 5-бромдиоксиуредин;
- остатки токсинов, такие как, например, остаток токсина, выбранный из группы, состоящей из экзотоксина Pseudomonas (РЕ или его цитотоксического фрагмента или мутанта), токсина Diptheria или его цитотоксического фрагмента или мутанта, ботулинического токсина А, В, С, D, Е или F, рицина или его цитотоксического фрагмента, например рицина А, абрина или его цитотоксического фрагмента, сапорина или его цитотоксического фрагмента, антивирусного токсина фитолакки американской или его цитотоксического фрагмента и бриодина 1 или его цитотоксического фрагмента.
Приемлемые ферменты и коферменты являются раскрытыми в Litman, и др., US4275149, и Boguslaski, и др., US4318980, каждый из которых введен в данную заявку в качестве ссылки в своей целостности. Приемлемые флуоресцирующие вещества и хемолюминисцирующие вещества являются раскрытыми у Litman, и др., патент US4275149, который введен в данную заявку в качестве ссылки в своей целостности. Метки дополнительно включают химические остатки, такие как биотин, которые могут определяться с помощью связывания со специфическим родственным, способным к определению остатком, например, меченным авидином или стрептавидином. Способные к определению метки могут присоединяться к антителам в соответствии с изобретением при использовании традиционных химических реакций, известных в области техники.
Иммуноконъюгаты или их функциональные фрагменты могут быть получены с помощью способов, известных квалифицированному специалисту в данной области техники. Они могут быть слиты с ферментами или флуоресцентными метками непосредственно или при использовании вспомогательной спейсерной группы или связывающей группы, такой как полиальдегид, подобный глутаральдегиду, этилендиаминтетрауксусная кислота (ЭДТА), диэтилентриаминпентауксусная кислота (DPTA), или в присутствии конденсирующих агентов, таких как те, что упомянуты выше для терапевтических конъюгатов. Конъюгаты, содержащие метки флуоресцентного типа, могут быть получены путем реакции с изотиоцианатом.
Способы, известные квалифицированному специалисту в данной области техники, существующие для конденсации терапевтических радиоизотопов к антителам либо непосредственно, либо при использовании хелатирующего агента, такого как ЭДТА, DTPA, упомянутые выше, могут использоваться для радиоактивных элементов, которые могут использоваться в диагностике. Является также возможным осуществлять мечение натрием125 с помощью способа на основе хлорамина Т [59. Hunter W. M. and Greenwood F. С. (1962) Nature 194:495] или технеция99m с помощью методики Crockford и др. (US4424200, введенный в данную заявку в качестве ссылки в своей целостности) или присоединять DTPA так, как описано Hnatowich (US4479930, введенный в данную заявку в качестве ссылки в своей целостности).
Существует множество способов, с помощью которых метка может вырабатывать сигнал, определяемый с помощью внешних средств, например с помощью визуального наблюдения, электромагнитного излучения, нагревания и химических реагентов. Метка может также связываться с другим связывающим элементом, который связывается с антителом в соответствии с изобретением или с носителем.
Метка может непосредственно вырабатывать сигнал, и, таким образом, для выработки сигнала не требуются дополнительные компоненты. Многочисленные органические молекулы, например флуоресцентные вещества, являются способными поглощать ультрафиолетовый и видимый свет, где поглощение света передает энергию этим молекулам и поднимает их до возбужденного энергетического состояния. Эта поглощенная энергия потом рассеивается путем излучения света при второй длине волны. Это излучение света со второй длиной волны может также передавать энергию меченой акцепторной молекуле, и полученная энергия рассеивается из акцепторной молекулы путем излучения света, например резонансного переноса энергии флуоресценции (FRET). Другие метки, которые непосредственно вырабатывают сигнал, включают радиоактивные изотопы и красители.
Альтернативно, метка может требовать других компонентов для выработки сигнала, и сигнал, выработанный системой, будет потом включать все компоненты, необходимые для выработки измеряемого сигнала, который может включать субстраты, коферменты, энхансеры, дополнительные ферменты, вещества, которые реагируют с ферментативными продуктами, катализаторы, активаторы, кофакторы, ингибиторы, скэвенджеры, ионы металлов и специфическое связывающее вещество, необходимое для связывания веществ, генерирующих сигнал. Подробное обсуждение приемлемых систем, вырабатывающих сигнал, может быть найдено у Ullman, и др. US5185243, который введен в данную заявку в качестве ссылки в своей целостности.
Настоящее изобретение обеспечивает способ, включающий индукцию или разрешение связывания связывающего элемента, как обеспечивается в данной заявке, с IgE. Как было отмечено, такое связывание может иметь место in vivo, например, после введения связывающего элемента или нуклеиновой кислоты, кодирующей связывающий элемент, или оно может иметь место in vitro, например, в анализах ELISA, вестерн-блоттинга, иммуноцитохимии, иммунопреципитации, аффинной хроматографии и биохимических анализах или анализах на основе клеток.
В общем случае, комплексы, образованные между связывающим элементом в соответствии с изобретением и IgE, могут быть определены, среди прочих, с помощью иммуноферментного анализа, радиоанализа, иммунопреципитации, флуоресцентного иммуноанализа, хемилюминисцентного анализа, анализа иммуноблоттинга, анализа на основе растекания в радиальном направлении, анализа агглютинации или анализа на основе частиц.
Настоящее изобретение также предусматривает непосредственное измерение уровней антигена путем использования связывающего элемента в соответствии с изобретением, например, в биосенсорной системе. Например, настоящее изобретение включает способ определения и/или измерения связывания с IgE, включающий (i) взаимодействие указанного связывающего элемента с IgE и (ii) определение связывания указанного связывающего элемента с IgE, где связывание определяют при использовании любого способа или определяемой метки, описанных в данной заявке. Этот и любой другой способ определения связывания, описанные в данной заявке, могут интерпретироваться непосредственно лицом, осуществляющим данный способ, например, с помощью визуального наблюдения определяемой метки. Альтернативно, этот способ или любой другой способ определения связывания, описанные в данной заявке, могут обеспечивать отчет в форме ауторадиографии, фотографии, компьютерной распечатки, отчета проточной цитометрии, графика, схемы, исследуемой пробирки, или контейнера, или ячейки, содержащей этот результат, или любого другого визуального или физического представления результата способа.
Может быть определено количество связывания связывающего элемента с IgE. Количественная оценка связана с количеством антигена в исследуемом образце, которое может представлять диагностический интерес. Скрининг на IgE связывание и/или его количественная оценка могут быть полезными, например, при отборе пациентов на заболевания и расстройства, на которые ссылаются в данной заявке, и/или на любое другое заболевание или расстройство, вовлекающее абберантную экспрессию и/или активность IgE, экспрессию и/или активность.
Диагностический способ в соответствии с изобретением может включать (i) получения образца ткани или жидкости от субъекта, (ii) воздействие на указанный образец ткани или жидкости одним или более связывающими элементами в соответствии с настоящим изобретением; (iii) определение связанного IgE по сравнению с контрольным образцом, где повышение количества связывания IgE по сравнению с контролем может свидетельствовать об абберантном уровне экспрессии или активности IgE. Образцы ткани или жидкости, которые подвергают анализу, включают кровь, сыворотку, мочу, материал биопсии, опухоль, или любую ткань, которая подозревается в содержании абберантных уровней IgE. Позитивные по результатам тестирования на абберантные уровни или активность IgE субъекты могут также получить выгоду от способов лечения, раскрытых далее в данной заявке.
Диагностический способ в соответствии с изобретением может дополнительно включать захват комплекса связывающего элемента и IgE с помощью иммобилизованного антигена. Например, антиген может быть иммобилизован на боковой полоске аналитического устройства для захвата антигенспецифического IgE в образце, который представляет интерес.
Специалист в данной области техники способен выбрать приемлемый способ определения связывания связывающего элемента с антигеном в соответствии с его предпочтением и общими знаниями в свете способов, представленных в данной заявке.
Реактивности связывающих элементов в образце могут быть определены с помощью любого приемлемого способа. Радиоиммуноанализ (RIA) представляет собой один из возможных. Радиоактивно меченный антиген смешивают с немеченым антигеном (исследуемый образец) и осуществляют связывание со связывающим элементом. Связанный антиген физически отделяют от несвязанного антигена и определяют количество радиоактивного антигена, связанного со связывающим элементом. Чем больше антигена в исследуемом образце, тем меньше радиоактивного антигена будет связываться со связывающим элементом. Анализ конкурентного связывания может также применяться при использовании нерадиоактивного антигена при использовании антигена или аналога, связанного с репортерной молекулой. Репортерная молекула может представлять собой флуорохром, фосфор и лазерный краситель со спектрально изолированными характеристиками поглощения или эмиссии. Приемлемые флуорохромы включают флуоресцеин, родамин, фикоэритрин и Texas Red, а также хелаты и криптаты лантаноидов. Приемлемые хромогенные красители включают диаминобензидин.
Другие репортеры включают макромолекулярные коллоидные частицы или зернистый материал, такой как латексные шарики, которые являются окрашенными, магнитными и парамагнитными, и биологически или химически активные агенты, которые могут непосредственно или опосредованно вызывать способные к определению сигналы, которые подвергаются визуальному наблюдению, электронно определяются или регистрируются иным путем. Такие молекулы могут, например, представлять собой ферменты, которые катализируют реакции, которые развиваются, или изменять цвет, или вызывать изменения в электрических свойствах. Они могут представлять собой молекулярно возбуждаемые, так что электронные переходы между энергетическими состояниями приводят к характерному спектральному поглощению или эмиссии. Они могут включать химические вещества, используемые в сочетании с биосенсорами. Системы определения на основе биотина/авидина или биотина/стрептавидина и щелочной фосфатазы также могут использоваться.
Сигналы, генерированные конъюгатами индивидуальный связывающий элемент-репортер, могут использоваться для получения способного к количественной оценки абсолюта или относительных данных связывания релевантного связывающего элемента в образцах (нормальный и исследуемый).
Набор, включающий связывающий элемент согласно с любым из аспектов или воплощений в соответствии с настоящим изобретением, может также обеспечиваться как аспект настоящего изобретения. В этом наборе связывающий элемент может быть меченым для того, чтобы было возможным осуществлять его реактивность в образце, например так, как описано в данной заявке ниже. Кроме того, связывающий элемент может или не может быть прикрепленным к твердой основе. Компоненты набора в общем случае являются стерильными и содержаться в запечатанных флаконах или других контейнерах. Наборы могут использоваться в диагностическом анализе или других способах, для которых связывающие элементы являются полезными. Набор может содержать инструкции для применения компонентов в способе, например способе в соответствии с настоящим изобретением. Вспомогательные материалы, которые помогают в способе или позволяют осуществить такой способ, могут также включаться в набор в соответствии с изобретением. Вспомогательные материалы включают второй, отличный связывающий элемент, который связывается с первым связывающим элементом и является конъюгированным с определяемой меткой (например, флуоресцентной меткой, радиоактивным изотопом или ферментом). Наборы на основе антитела могут также включать шарики для осуществления иммунопреципитации. Каждый компонент наборов в общем случае представлен в своем собственном контейнере. Таким образом, эти наборы в общем случае включают различные контейнеры, приемлемые для каждого связывающего элемента. Кроме того, эти наборы могут включать инструкции для осуществления анализа и способы для интерпретации и анализа данных, полученных в результате проведения анализа.
Настоящее изобретение также обеспечивает применение связывающего элемента, как описано выше, для измерения уровней антигена в конкурентном анализе, иными словами, способе измерения уровня антигена в образце путем использования связывающего элемента, как обеспечивается настоящим изобретением в конкурентном анализе. Такой способ может быть таким, в котором не требуется физического разделения связанного и несвязанного антигена. Присоединение репортерной молекулы к связывающему элементу так, что при этом происходит физическое или оптическое изменение, представляет собой одну из возможностей. Репортерная молекула может непосредственно или опосредованно генерировать определяемые сигналы, которые могут быть оценены количественно. Связь репортерной молекулы может осуществляться непосредственно или опосредованно, ковалентно, например, с помощью пептидной связи или нековалентно. Связь с помощью пептидной связи может представлять собой результат рекомбинантной экспрессии генного слияния, кодирующего антитело и репортерную молекулу.
В различных аспектах и воплощениях настоящее изобретение охватывает связывающий элемент, который конкурирует за связывание с IgE, например человеческим IgE, с любым связывающим элементом, определенным в данной заявке, например антителом 1, например, в IgG1 формате. Конкуренция между связывающими элементами может быть легко проанализирована in vitro, например, с помощью маркировки специфической репортерной молекулой одного связывающего элемента, который может быть определен в присутствии другого(их) немаркированного(ых) связывающего(их) элемента(ов) для того, чтобы позволить осуществлять определение связывающих элементов, которые связываются с одним и тем же эпитопом или перекрывающимся эпитопом. Конкуренция может определяться, например, при использовании ELISA, в котором IgE является иммобилизованным на планшете, а первый маркированный или меченный связывающий элемент вместе с одним или более другими немаркированными или немеченными связывающими элементами прибавляются к планшету. Присутствие немаркированного связывающего элемента, который конкурирует с маркированным связывающим элементом, наблюдают при снижении сигнала, испускаемого маркированным связывающим элементом. В одном примере может использоваться HTRF® эпитопный конкурентный анализ.
Например, настоящее изобретение включает способ идентификации соединения, связывающего IgE, включающий (i) иммобилизацию IgE на основе, (ii) контакт указанного иммобилизованного IgE одновременно или поэтапно, по крайней мере, с одним маркированным или меченным связывающим элементом в соответствии с изобретением и одним или более немаркированным или немеченным исследуемым связывающим соединением и (iii) идентификацию нового связывающего IgE соединения путем наблюдения снижения количества связанного маркера маркированного связывающего элемента. Такие способы могут быть осуществлены с высоким выходом при использовании в формате многолуночного планшета или в формате массивов. Такие анализы могут также осуществляться в растворе. Смотри, например, патент US5814468, который введен в данную заявку в качестве ссылки в своей целостности. Как описано выше, определение связывания может интерпретироваться непосредственно лицом, осуществляющим способ, например, с помощью визуального наблюдения определяемой метки или снижения ее присутствия. Альтернативно, способы связывания в соответствии с изобретением могут представлять отчет в форме ауторадиограммы, фотографии, компьютерной распечатки, отчета результатов проточной цитометрии, графика, схемы, исследуемой пробирки, или контейнера, или ячейки, содержащей этот результат, или любого другого визуального или физического представления результата способа.
Конкурентные анализы могут также использоваться в картировании эпитопов. В одном случае картирование эпитопов может использоваться для идентификации эпитопа, связанного с элементом, связывающим IgE, который необязательно может обладать оптимизированными характеристиками нейтрализации и/или модуляции. Такой эпитоп может быть линейным или конформационным. Конформационный эпитоп может включать, по крайней мере, два различных фрагмента IgE, где указанные фрагменты размещаются поблизости друг к другу, когда IgE сворачивается в свою третичную или четвертичную структуру с образованием конформационного эпитопа, который узнается ингибитором IgE, таким как связывающий IgE элемент. В анализе на конкуренцию может использоваться пептидный фрагмент антигена, в частности пептид, включающий или существенно состоящий из эпитопа, представляющего интерес. Может использоваться пептид, содержащий последовательность эпитопа плюс одна или более аминокислот на любом конце. Связывающие элементы в соответствии с настоящим изобретением могут быть такими, что их связывание с антигеном ингибируется пептидом, имеющим последовательность или включающим приведенную последовательность.
Настоящее изобретение также обеспечивает изолированную нуклеиновую кислоту, кодирующую связывающий элемент в соответствии с настоящим изобретением. Нуклеиновая кислота может включать ДНК и/или РНК. В одном аспекте настоящее изобретение обеспечивает нуклеиновую кислоту, которая кодирует CDR, или набор CDR, или VH домен, или VL домен, или антигенсвязывающий сайт антитела, или молекулу антитела, например, scFv или IgG1, в соответствии с изобретением, как определено выше.
Настоящее изобретение также обеспечивает конструкции в форме плазмид, векторов, транскрипционных или экспрессионных кассет, которые включают, по крайней мере, один полинуклеотид, как представлено выше.
Настоящее изобретение также обеспечивает рекомбинантную хозяйскую клетку, которая включает одну или более конструкций, как описано выше. Нуклеиновая кислота, кодирующая любой CDR или набор CDR, или VH домен, или VL домен, или антигенсвязывающий сайт антитела, или молекулу антитела, например, scFv или IgG1, как обеспечивается в данной заявке, сама по себе образует аспект в соответствии с настоящим изобретением, как и способ получения кодируемого продукта, где способ соответственно включает экспрессию при использовании кодирующей нуклеиновой кислоты. Экспрессия может быть достигнута путем культивирования при приемлемых условиях рекомбинантной хозяйской клетки, содержащей нуклеиновую кислоту. После получения путем экспрессии VH или VL домен или связывающий элемент может быть изолирован и/или очищен при использовании любой приемлемой методики, а потом может использоваться так, как это является приемлемым.
Нуклеиновая кислота в соответствии с настоящим изобретением может включать ДНК или РНК и может представлять собой полностью или частично синтетическую. Ссылка на нуклеотидную последовательность, как представлено в данной заявке, охватывает ДНК молекулу с указанной последовательностью, и охватывает РНК молекулу с указанной последовательностью, в которой U является замещенным на Т, если в контексте не требуется другое.
Еще один дополнительный аспект обеспечивает способ получения VH вариабельного домена антитела, где указанный способ включает индукцию экспрессии кодирующей нуклеиновой кислоты. Такой способ может включать культивирование хозяйской клетки в условиях, подходящих для получения указанного VH вариабельного домена антитела.
Аналогичные способы получения VL вариабельных доменов и связывающих элементов, включающих VH и/или VL домен, обеспечиваются как дополнительные аспекты в соответствии с настоящим изобретением.
Способ получения может включать этап изоляции и/или очистки продукта. Способ получения может включать рецептирование продукта в виде композиции, включающей, по крайней мере, один дополнительный компонент, такой как фармацевтически приемлемый наполнитель.
Системы для клонирование и экспрессии полипептида во множестве различных хозяйских клеток являются хорошо известными. Приемлемые хозяйские клетки включают бактерии, клетки млекопитающих, растительные клетки, мицелиальные грибы, дрожжи и бакуловирусные системы и трансгенных растений и животных. Экспрессия антител и фрагментов антител в прокариотических клетках является хорошо изученной в области техники. Для обзора смотри, например, Plilckthun [60 Pluckthun, A. Bio/Technology 9:545-551 (1991)]. Обычный бактериальный хозяин представляет собой Е.coli.
Экспрессия в эукариотических клетках в культуре является также доступной квалифицированному специалисту в данной области техники как возможность получения связывающего элемента [61. Chadd HE and Chamow SM (2001) Current Opinion in Biotechnology 12:188-194; 62. Andersen DC and Krummen L (2002) Current Opinion in Biotechnology 13:117; 63. Larrick JW and Thomas DW (2001) Current Opinion in Biotechnology 12:411-418]. Линии клеток млекопитающих, доступные в области техники для экспрессии гетерологичного полипептида, включают клетки яичника китайского хомячка (СНО), клетки HeLa, клетки почки детеныша хомяка, NS0 клетки мышиной меланомы, YB2/0 клетки крысиной меланомы, эмбриональные клетки почки человека, человеческие зародышевые клетки сетчатки и многие другие.
Могут быть выбраны или сконструированы приемлемые векторы, содержащие приемлемые регуляторные последовательности, включая промоторные последовательности, последовательности терминатора, последовательности полиаденилирования, энхансерные последовательности, маркерные гены и другие последовательности, которые являются приемлемыми. Векторы могут быть плазмидами, например фагмидами, или быть вирусными, например фагом, если это является приемлемым [64. Sambrook and Russell, Molecular Cloning: a Laboratory Manual: 3rd edition, 2001, Cold Spring Harbor Laboratory Press]. Любые известные методики и прописи для работы с нуклеиновой кислотой, например, при получении конструкций на основе нуклеиновой кислоты, мутагенезе, секвенировании, введении ДНК в клетки и экспрессии генов, а также при анализе белков являются описанными подробно у Ausubel и др. [65. Ausubel et al. eds., Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, John Wiley & Sons, 4th edition 1999].
Дополнительный аспект настоящего изобретения обеспечивает хозяйскую клетку, содержащую нуклеиновую кислоту, как раскрыто в данной заявке. Такая хозяйская клетка может быть in vitro и может находиться в культуре. Такая хозяйская клетка может быть in vivo. In vivo присутствие хозяйской клетки может позволить осуществлять внутриклеточную экспрессию связывающих элементов в соответствии с настоящим изобретением как "интрател" или внутриклеточных антител. Интратела могут использоваться для генной терапии.
Еще один аспект обеспечивает способ, включающий введение нуклеиновой кислоты в соответствии с изобретением в хозяйскую клетку. При введении можно использовать любую доступную методику. Для эукариотических клеток приемлемые методики могут включать трансфекцию, опосредованную фосфатом кальция, DEAE-декстраном, электропорацию, опосредованную липосомами трансфекцию и трансдукцию при использовании ретровирусов и других вирусов, например, вакцины, или для клеток насекомых - бакуловирусов. При введение нуклеиновой кислоты в хозяйскую клетку, в частности в эукариотическую клетку, можно использовать вирусную систему или систему на основе плазмид. Плазмидная система может поддерживаться эписомально или может вводиться в хозяйскую клетку или в искусственную хромосому. Введение может представлять собой либо случайную, либо нацеленную интеграцию одной или более копий в единичный локус или множественные локусы. Для бактериальных клеток приемлемые методики могут включать опосредованную хлоридом кальция трансформацию, электропорацию и трансфекцию при использовании бактериофага.
Введение может сопровождаться индукцией или разрешением экспрессии нуклеиновой кислоты, например путем культивирования хозяйской клетки при условиях, приемлемых для экспрессии гена. Очистка экспрессированного продукта может быть достигнута с помощью способов, известных специалисту в данной области техники.
Нуклеиновая кислота в соответствии с изобретением может представлять собой интегрированную в геном (например, хромосому) хозяйской клетки. Интеграция может быть промотирована путем включения последовательности, которая способствует рекомбинации в геноме в соответствии со стандартными методиками.
Настоящее изобретение также обеспечивает способ, который включает использование конструкции, как описано выше, в экспрессионной системе для того, чтобы экспрессировать связывающий элемент или полипептид, как описано выше.
Связывающие элементы в соответствии с настоящим изобретением могут использоваться в способах диагностики или лечения человека или животных, в частности человека. Связывающие элементы для IgE могут использоваться для лечения расстройств, характеризующихся биологическими эффектами, опосредованными IgE, в частности различных видов аллергии и астмы. Например, связывающие элементы в соответствии с изобретением могут использоваться для лечения аллергического ринита, аллергического контактного дерматита, атопического дерматита, анафилактической реакции, пищевой аллергии, крапивницы, воспалительного заболевания кишечника, эозинофильного гастроэнтерита, индуцированной лекарственными средствами сыпи, аллергической офтальмопатии или аллергического конъюнктивита.
Связывающие элементы для IgE могут использоваться для ингибирования индуцированной аллергеном дегрануляции тучных клеток in vivo или in vitro, снижения опосредованных FcεR1 биологических ответов in vivo или in vitro, а также для снижения уровня циркулирующего IgE у пациента, который представляет собой человека или животное.
В соответствии с этим изобретение обеспечивает способ для ингибирования индуцированной аллергеном дегрануляции тучных клеток у млекопитающих, включающий введение указанному млекопитающему связывающего элемента, например антитела, VH домена, или VL домена в соответствии с изобретением, в количестве, достаточном для нейтрализации IgE.
Изобретение дополнительно обеспечивает способ снижения FcεRI-биологических ответов с одновременным снижением опосредованных FcεRII биологических ответов или без них, включающий контакт клетки, экспрессирующей FcεRI и/или FcεRII со связывающим элементом, например антителом, VH доменом, или VL доменом в соответствии с изобретением, в присутствии IgE.
Изобретение дополнительно обеспечивает способ снижения опосредованных FcERI биологических ответов с одновременным снижением опосредованных FcERII биологических ответов или без них, включающий контакт клетки, экспрессирующей FcεRI и/или FcεRII со связывающим элементом, например антителом, VH доменом, или VL доменом в соответствии с изобретением, в присутствии IgE.
Когда исследуемые клетки контактируют со связывающим элементом в соответствии с изобретением in vitro, контрольная(ые) клетка (клетки) также могут использоваться для позитивных контролей (например, реакции, не содержащие связывающего элемента) и/или негативных контролей (например, реакции, не содержащие IgE и/или антигена).
Когда клетки подвергают контакту со связывающим элементом in vivo, например, путем введения связывающего элемента в соответствии с изобретением млекопитающему, демонстрирующему опосредованные FcεRI- и/или FcεRII-биологические ответы, связывающий элемент в соответствии с изобретением вводится в количестве, достаточном для нейтрализации IgE.
Кроме того, изобретение обеспечивает способ снижения уровня циркулирующего IgE у млекопитающих, таких как человек, включающий введение связывающего элемента, такого как антитела, VH домен или VL домен в соответствии с изобретением, в количестве, достаточном для нейтрализации м снижения циркулирующего IgE.
Связывающие элементы в соответствии с изобретением могут использоваться в диагностике и лечении заболеваний или расстройств, включая, но не ограничиваясь, одно или более из следующих: аллергический ринит, аллергический контактный дерматит, атонический дерматит, анафилактическая реакция, пищевая аллергия, крапивница, воспалительное заболевание кишечника, эозинофильный гастроэнтерит, индуцированная лекарственными средствами сыпь, аллергическая офтальмопатия риноконъюнктивит, аллергический конъюнктивит, бронхиальная астма, гиперчувствительность дыхательных путей, косметическая аллергия, аллергия, индуцированная лекарственными средствами, синдром гиперчувствительности, индуцированный лекарственными средствами, аллергия на металлы, профессиональный гиперчувствительный пневмонит, хронический гиперчувствительный пневмонит, холодовая гиперчувствительность, гиперчувствительность, индуцированная гельминтами, аллергия на латекс и аллергия на пыльцу.
Данные, представленные в данной заявке в отношении связывания и нейтрализации IgE, таким образом, показывают, что связывающие элементы в соответствии с изобретением могут использоваться для лечения или предотвращения таких расстройств, включая снижение тяжести расстройств. В соответствии с этим, изобретение обеспечивает способ лечения или снижения тяжести, по крайней мере, одного симптома любого расстройства, упомянутого в данной заявке, включающий введение пациенту, который в этом нуждается, эффективного количества одного или более связывающих элементов в соответствии с настоящим изобретением отдельно или в сочетанием терапевтическом режиме с другим приемлемым лекарственным средством, известным в области техники или описанным в данной заявке, так, что тяжесть, по крайней мере, одного симптома любого из перечисленных выше расстройств снижается.
Связывающие элементы в соответствии с изобретением могут использоваться у приемлемых животных и в животных моделях заболевания, в частности у обезьян.
Таким образом, связывающие элементы в соответствии с настоящим изобретением являются полезными в качестве терапевтических агентов в лечении заболеваний или расстройств, вовлекающих IgE, например, продукцию IgE, экспрессию и/или активность, в частности абберантную продукцию, экспрессию или активность. Способ лечения может включать введение эффективного количества связывающего элемента в соответствии с изобретением пациенту, который в этом нуждается, где продукция, экспрессия и/или активность IgE таким образом снижается. Способ лечения может включать (i) идентификацию пациента, демонстрирующего повышенные уровни или активность IgE, например, при использовании диагностических способов, описанных выше, и (ii) введение эффективного количества связывающего элемента в соответствии с изобретением пациенту, где повышенная продукция, экспрессия и/или активность IgE снижается. Альтернативный способ лечения может включать (i) идентификацию пациента, который не имеет очевидного повышения уровней IgE, но который, как предполагается, может получить выгоду от введения связывающего элемента в соответствии с изобретением, и (ii) введение эффективного количества связывающего элемента в соответствии с изобретением такому пациенту. Эффективное количество в соответствии с изобретением представляет собой количество, которое снижает повышенную продукцию, экспрессию и/или активность IgE так, что снижается или уменьшается тяжесть, по крайней мере, одного симптома определенного заболевания или расстройства, которое подвергают лечению, но без обязательного излечивания заболевания или расстройства.
Изобретение также обеспечивает способ антагонизации, по крайней мере, одного эффекта IgE, включающий контакт с или введение эффективного количества одного или более связывающих элементов в соответствии с настоящим изобретением так, что, по крайней мере, один эффект IgE подвергается антагонизации. Эффекты IgE, которые могут подвергаться антагонизации с помощью способов в соответствии с изобретением, включают биологические ответы, опосредованные FcεRI и/или FcεRII, и любые снижающие эффекты, которые возникают как следствие таких реакций связывания.
В соответствии с этим дополнительные аспекты в соответствии с изобретением обеспечивают применение изолированного связывающего элемента, такого как антитело, VH домен или VL домен в соответствии с изобретением для производства лекарственного средства для лечения расстройства, ассоциированного с или опосредованного IgE, как обсуждается в данной заявке. Такое применение или способы получения лекарственного средства или фармацевтической композиции включают рецептирование связывающего элемента с фармацевтически приемлемым наполнителем.
Фармацевтически приемлемый носитель может представлять собой соединение или комбинацию соединений, введенных в состав фармацевтической композиции, не вызывающей вторичных реакций, и позволяющей, например, улучшить введение активного(ых) соединений), увеличить его (их) период жизни и/или его (их) эффективность в организме, повысить растворимость в растворе или также улучшить его превращение. Такие фармацевтически приемлемые носители являются хорошо известными и будут адаптироваться квалифицированным специалистом в данной области техники как функция природы и способов введения выбранного(ых) активного(ых) соединениями).
Связывающие элементы в соответствии с настоящим изобретением будут обычно вводиться в форме фармацевтической композиции, которая может включать, по крайней мере, один компонент в дополнение к связывающему элементу. Таким образом, фармацевтические композиции в соответствии с настоящим изобретением, и для применения в соответствии с настоящим изобретением могут включать, в дополнение к активному ингредиенту, фармацевтически приемлемый наполнитель, носитель, буфер, стабилизатор или другие материалы, хорошо известные квалифицированному специалисту в данной области техники. Такие материалы не должны быть токсическими и не должны препятствовать эффективности активного ингредиента. Точная природа носителя или другого материала будет зависеть от способа введения, который может представлять собой пероральный, путем ингаляции, интратрахеальный, местный, интравезикулярный или путем инъекции, как обсуждается ниже.
Фармацевтические композиции для перорального введение, такие, как например, для единичного домена молекулы антитела (например, "нанотела™") и др. также предусматриваются в настоящем изобретении. Такие пероральные композиции могут быть в виде таблетки, капсулы, порошка, содержаться в жидкой или полужидкой форме. Таблетка может включать твердый носитель, такой как желатин, или адъювант. Жидкие фармацевтические композиции в общем случае включают жидкий носитель, такой как вода, углеводороды, животные и растительные масла, минеральное масло или синтетические масла. Могут также включаться физиологический солевой раствор, декстроза или раствор другого сахарида, или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Для внутривенной инъекции или инъекции в место повреждения активный ингредиент будет находиться в форме парентерально доступного водного раствора, который является свободным от пирогена и имеет приемлемое значение рН, изотоничность и стабильность. Квалифицированные специалисты в данной области техники будут в состоянии приготовить приемлемые растворы при использовании, например, изотонических носителей, таких как инъекционный раствор хлорида натрия, инъекционный раствор Рингера, инъекционный раствор Рингера с лактатом. Консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки могут использоваться, если это является необходимым, включая буферы, такие как фосфатный, цитратный и других органических кислот; антиоксиданты, такие как аскорбиновая кислота и метионин; консерванты (такие как октадецилдиметилбензил аммоний хлорид; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3'-пентанол; и м-крезол); полипептиды с низкой молекулярной массой; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагины, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (ПЭГ).
Связывающие элементы в соответствии с настоящим изобретением могут быть рецептированы в виде жидкой, полутвердой или твердой формы, в зависимости от физико-химических свойств молекулы и способа доставки. Композиции могут включать наполнители или комбинации наполнителей, например: сахара, аминокислоты и поверхностно-активные вещества. Жидкие композиции могут иметь широкий спектр концентраций антитела и рН. Твердые композиции могут быть получены путем лиофилизации, высушивания распылением или высушены, например, при использовании технологии надкритической жидкости. Рецептуры анти-CXCL13 будут зависеть от предназначенного способа доставки: например, рецептуры для легочной доставки могут состоять из частиц с физическими свойствами, которые обеспечивают глубокое проникновение в легкие при ингаляции; рецептуры для местного введения (например, для лечения рубцевания, например, дермального рубцевания) могут включать агенты, модифицирующие вязкость, который продлить время, в течение которого лекарственное средство находится в месте действия. Связывающий элемент может быть приготовлен с носителем, который будет защищать связывающий элемент от быстрого высвобождения, так, как в композиции контролированного высвобождения, включая имплантаты, трансдермальные пластыри, и микроинкапсулированные системы доставки. Могут использоваться биоразлагаемые, биосовместимые полимеры, такие, как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэстеры, и полимолочная кислота. Многочисленные способы для получения таких композиций являются известными квалифицированному специалисту в данной области техники [66. Robinson, J. R. ed., Sustained and Controlled Release Drug Delivery Systems, Marcel Dekker, Inc., New York, 1978].
Анти-IgE лечение может обеспечиваться перорально (как, например, для одного домена молекулы антитела (например, "нанотела™"), путем инъекции (например, подкожно, интраартикулярно, внутривенно, интраперитонеально, интраартериально или внутримышечно), с помощью ингаляции, интратрахеально, с помощью интравезикулярного пути (инсталляция в мочевой пузырь) или местно (например, интраокулярно, интарназально, ректально, в раны, на кожу). Лечение может вводиться в режиме импульсной инфузии, в частности с помощью убывающих доз связывающего элемента. Способ введения может определяться физико-химическими характеристиками лечения, специальными соображениями для заболевания или требованием оптимизации эффективности или для минимизации побочных эффектов. Частный способ введения представляет собой внутривенный. Другой способ введения фармацевтической композиции в соответствии с настоящим изобретением представляет собой подкожное. При этом предусматривается, что анти-IgE лечение не будет ограничиваться применением его в клинике. Таким образом, подкожная инъекция при использовании безигольного устройства является также предпочтительной.
Примеры внутривенных композиций включают:
Композиция (1) включает
Изолированный связывающий элемент в соответствии с изобретением (необязательно 10, 50, 100 или 150 мг/мл указанного связывающего элемента, например, антитела)
50 мМ ацетата натрия
100 мМ NaCl
рН 5,5
Композиция (2) включает
Изолированный связывающий элемент в соответствии с изобретением (необязательно 10, 50, 100 или 150 мг/мл указанного связывающего элемента, например, антитела)
20 мМ сукцината
105 мМ NaCl
10 мМ аргинина
рН 6,00
Композиция может вводиться отдельно или в комбинации с другими лечениями, либо одновременно, либо последовательно, в зависимости от состояния, которое подвергается лечению.
Связывающий элемент для IgE может использоваться как часть комбинационной терапии в сочетании с дополнительным медицинским компонентом. Способы комбинационного лечения могут использоваться для обеспечения значительных синергетических эффектов, в частности комбинации анти-IgE связывающего элемента с одним или более другими лекарственными средствами. Связывающий элемент для IgE может вводиться одновременно или последовательно или в виде сочетанного препарата с другим терапевтическим агентом или агентами, для лечения одного или более состояний, приведенных в данной заявке.
Связывающий элемент в соответствии с настоящим изобретением может обеспечиваться в комбинации или в дополнение к одному или более из следующих агентов:
- цитокин, или агонист, или антагонист цитокиновой функции (например, агент, который действует на пути передачи цитокинового сигнала, такой как модулятор SOCS системы), такой как альфа-, бета- и/или гамма-интерферон; инсулинподобный фактор роста типа I (IGF-1), его рецепторы и ассоциированные связывающие белки; интерлейкины (IL), например, один или более IL-1 - -33, и/или интерлейкиновый антагонист или ингибитор, такой, как анакинра; ингибиторы рецепторов членов семейства интерлейкинов или ингибиторы специфических субъединиц таких рецепторов, ингибитор фактора некроза опухоли альфа (TNF-α), такой как анти-TNF моноклональные антитела (например, инфликсимаб, адалимумаб и/или CDP-870) и/или антагонист рецептора TNF, например молекула иммуноглобулина (такой, как этанерцепт) и/или агент с низким молекулярным весом, такой как пентоксифиллин;
- модулятор В клеток, например, моноклональное антитело, направленное против В-лимфоцитов (такое как CD20 (ритуксимаб, MRA-aIL16R или белимумаб) или Т-лимфоцитов (например, CTLA4-Ig (Абатацепт), HuMax I1-15);
- модулятор активации, созревания или выживания В клеток, (такой как Атацицепт);
- модулятор иммунной функции, например, антагонист лимфотоксинового LIGHT пути передачи (такой как LTBR-Fc)
- модулятор, который ингибирует остеокластную активность, например, антитело к RANKL;
- модулятор функции хемокина или хемокинового рецептора, такой как антагонист CCR1, CCR2, CCR2A, CCR2B, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10 и CCR11 (для С-С семейства); CXCR1, CXCR2, CXCR3, CXCR4 и CXCR5 и CXCR6 (для С-Х-С семейства) и CX3CR1 для С-Х3-C семейства;
- ингибитор матриксных металлопротеаз (ММР), то есть один или более из стромелизинов, коллагеназы и желатиназы, а также агреканазы, в частности коллагеназа-1 (ММР-1), коллагеназа-2 (ММР-8), коллагеназа-3 (ММР-13), стромелизин-1 (ММР-3), стромелизин-2 (ММР-10) и/или стромелизин-3 (ММР-11) и/или ММР-9 и/или ММР-12, например, агент, такой как доксициклин;
- ингибитор биосинтеза лейкотриена, ингибитор 5-липоксигеназы (5-LO) или антагонист белка, активирующего 5-липоксигеназу (FLAP), такой как зилейтон; АВТ-761; фенлейтон; тепоксалин; Abbott-79175; Abbott-85761; N-(5-замещенные)-тиофен-2-алкилсульфонамиды; 2,6-ди-трет-бутилфенолгидразоны; метокситетрагидропираны, такие как Zeneca ZD-2138; соединение SB-210661; замещенное пиридинилом соединение 2-цианонафталина, такое как L-739,010; соединение 2-цианохинолина, такое как L-746,530; индольное и/или хинолиновое соединение, такое как MK-591, MK-886 и/или BAY x 1005;
- антагонист рецептора для лейкотриенов (LT) B4, LTC4, LTD4, и LTE4, выбранный из группы, состоящей из фенотиазинов-3-1, таких как L-651,392; амидиновых соединений, таких как CGS-25019c; бензоксаламинов, таких как онтазоласт; бензолкарбоксиимидамидов, таких как BIIL 284/260; и соединений, таких как зафирлукаст, аблукаст, монтелукаст, пранлукаст, верлукаст (MK-679), RG-12525, Ro-245913, иралукаст (CGP 45715A) и BAY x 7195;
- ингибитор фосфодиэстеразы (PDE), такой как метилксантанин, например, теофиллин и/или аминофиллин; и/или селективный ингибитор изофермента PDE, например, PDE4 ингибитор и/или ингибитор изоформы PDE4D, и/или ингибитор PDE5;
- антагонист рецептора гистамина типа 1, такой как цетиризин, лоратадин, дезлоратадин, фексофенадин, акривастин, терфенадин, астемизол, азеластин, левокабастин, хлорфенирамин, прометазин, циклизин, и/или мизоластин (в общем случае применяется перорально, местно или парентерально);
- ингибитор протонного насоса (такой как омепразол) или гастропротективный антагонист рецептора гистамина типа 2;
- антагонист гистаминового рецептора типа 4;
- сосудосужающий симпатомиметический агент агониста альфа-1/альфа-2 адренорецептора, такой как пропилгекседрин, фенилэфрин, фенилпропаноламин, эфедрин, псевдоэфедрин, нафалозолин гидрохлорид, оксиметазолин гидрохлорид, тетрагидрозолин гидрохлорид, ксилометазолин гидрохлорид, трамазолин гидрохлорид и этилнорэфедрин гидрохлорид;
- антихолинергический агент, например, антагонист мускаринового рецептора (М1, М2, и М3), такой как атропин, гиосцин, гликопирролат, бромид ипратропия, бромид тиотропия, бромид окситропия, пирензепин и телензепин;
- агонист бета-адренергического рецептора (включая подтипы 1-4 бета-рецептора), такой как изопреналин, сальбутамол, формотерол, сальметерол, тербуталин, орципреналин, битолтерол мезилат и/или пирбутерол, например его хиральный энантиомер;
- хромон, например, кромогликат натрия и/или недокромил натрия;
- глюкокортикоид, такой как флунизолид, триамцинолон ацетонид, беклометазон дипропионат, будезонид, флутиказон пропионат, циклезонид, и/или мометазон фуроат;
- агент, который модулирует ядерные рецепторы гормонов, такой как PPAR;
- иммуноглобулин (Ig) или Ig препарат, или антагонист, или антитело, модулирующее функцию Ig, такое как анти-IgE (например, омализумаб);
- другой, применяемый системно или местно противовоспалительный агент, например, талидомид или его производная, ретиноид, дитранол и/или кальципотриол;
- комбинации аминосалицилатов и сульфапиридина, такие как сульфазалазин, мезалазин, бальсалазид, и олсалазин; и иммуномодуляторные агенты, такие как тиопурины; и кортикостероиды, такие как будезонид;
- антибактериальный агент, например, производная пенициллина, тетрациклин, макролид, бета-пактам, фторхинолоны, метронидазол и/или вводимые с помощью ингаляции аминогликозиды; и/или противовирусный агент, например ацикловир, фамцикловир, валацикловир, ганцикловир, цидофовир; амантадин, римантадин; рибавирин; занамавир и/или оселтамавир; ингибитор протеазы, такой как индинавир, нелфинавир, ретинавир и/или саквинавир; ингибитор обратной нуклеозид транскриптазы, такой как диданозин, ламивудин, ставудин, зальцитабин, зидовудин; ингибитор ненуклеозидной обратной транскриптазы, такой как невирапин, эфавиренц;
- сердечно-сосудистый агент, такой как блокатор кальциевого канала, блокатор бета-адренорецептора, ингибитор ангиотензин-превращающего фермента (АСЕ), антагонист рецептора ангиотензина-2; агент, снижающий уровень липидов, такой как статин и/или фибрат; модулятор морфологии клеток крови, такой как пентоксифиллин; тромболитик и/или антикоагулянт, например, ингибитор оседания тромбоцитов;
- агент, который воздействует на центральную нервную систему, такой как антидепрессант (такой как сертралин), лекарственное средство против болезни Паркинсона (такое как депренил, L-допа, ропинирол, прамипексол; ингибитор МАОВ, такой как селегин и разагилин; ингибитор comP, такой как тасмар; А-2 ингибитор, ингибитор повторного поглощения допамина, NMDA антагонист, агонист никотиновой кислоты, допаминовый агонист и/или ингибитор нейронной синтазы окисла азота) и лекарственное средство против болезни Альцгеймера, такое как донепезил, ривастигмин, такрин, СОХ-2 ингибитор, пропентофиллин или метрифонат;
- агент для лечение острой и хронической боли, например действующие на центральную и периферическую нервную систему анальгетики, такие как опиоидный аналог или производная, карбамазепин, фенитоин, валпроат натрия, амитриптилин или другой антидепрессантный агент, парацетамол, или нестероидный противовоспалительный агент;
- парентерально или местно применяемые (включая путем ингаляции) локальные анальгетические агенты, такие как лигнокаин или его аналог;
- агент против остеопороза, например, гормональный агент, такой как ралоксифен, или бифосфонат, такой как алендронат;
- (i) ингибитор триптазы; (ii) антагонист фактора активации тромбоцитов (PAF); (iii) ингибитор интерлейкин превращающего фермента (ICE); (iv) IMPDH ингибитор; (v) ингибиторы адгезионной молекулы, включая VLA-4 антагонист; (vi) катепсин; (vii) ингибитор киназы, например, ингибитор тирозинкиназ (такой как Btk, Itk, Jak3 MAP примеры ингибиторов могут включать гефетиниб, иматиниб мезилат), серин/треонин киназы (например, ингибитор MAP киназы, такой как р38, JNK, протеинкиназы А, В и С и IKK), или киназы, являющейся втянутой в регуляцию клеточного цикла (например, циклинзависимой киназы); (viii) ингибитор глюкоза-6 фосфатдегидрогеназы; (ix) антагонист рецептора кинина-B.sub1. - и/или B.sub2.; (x) антиподагрический агент, например колхицин; (xi) ингибитор ксантиноксидазы, например алопуринол; (xii) агент, способствующий выведению мочевой кислоты, например пробенецид, сульфинпиразон и/или бензбромарон; (xiii) средство, усиливающее секрецию гормона роста; (xiv) трансформирующий фактор роста (TGFβ); (xv) тромбоцитарный фактор роста (PDGF); (xvi) фактор роста фибробластов, например основной фактор роста фибробластов (bFGF); (xvii) колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF); (xviii) капсаициновый крем; (xix) антагонист рецептора ташикинина NK.sub1. и/или NK.sub3., такой как NKP-608С, SB-233412 (талнетант) и/или D-4418; (хх) ингибитор эластазы, например, UT-77 и/или ZD-0892; (xxi) ингибитор фермента, превращающего TNF-альфа (ТАСЕ); (xxii) ингибитор индуцированной синтазы окисла азота (iNOS) или (xxiii) хемоаттрактант гомологичной рецептору молекулы, которая экспрессируется на ТН2 клетках (такой, как CRTH2 антагонист); (xxiv) ингибитор Р38 (xxv) агента, модулирующего функцию Toll-подобных рецепторов (TLR) и (xxvi) агент, модулирующий активность пуринергических рецепторов, такой как Р2Х7; (xxvii) ингибитор фактора активации транскрипции, такой как NFkB, API, и/или STATS.
Ингибитор может быть специфическим или может представлять собой смешанный ингибитор, например ингибитор, нацеленный на более чем одну молекулу (например, рецепторы), или класс молекул, упомянутых выше.
Связывающий элемент может также использоваться в ассоциации с химиотерапевтическим агентом или другим ингибитором тирозинкиназы при совместном введении или в форме иммуноконъюгата. Фрагменты указанного антитела могут также использоваться в биспецифических антителах, полученных с помощью механизмов рекомбинации или биохимического слияния и потом ассоциации специфичности описанного выше антитела со специфичностью другого антитела, способного распознавать другие молекулы, вовлеченные в активность, для которой IgE ассоциируется.
Для лечения воспалительного заболевания, например ревматоидного артрита, остеоартрита, астмы, аллергического ринита, хронического обструктивного легочного заболевания (COPD), или псориаза, связывающий элемент в соответствии с изобретением может представлять собой такой, соединенный с одним или более агентами, такими как нестероидные противовоспалительные агенты (в данной заявке называются НПВП), включая неселективные ингибиторы циклооксигеназы(СОХ)-1/СОХ-2, которые применяются либо местно, либо системно, такие как пироксикам, диклофенак, пропионовые кислоты, такие как напроксен, флурбипрофен, фенопрофен, кетопрофен и ибупрофен, фенаматы, такие как мефенаминовая кислота, индометацин, сулиндак, азапропазон, пиразолоны, такие как фенилбутазон, салицилаты, такие как аспирин); селективные ингибиторы СОХ-2 (такие как мелоксикам, целекоксиб, рофекоксиб, валдекоксиб, лумарококсиб, парекоксиб и эторикоксиб); доноры окисла азотной кислоты, ингибирующей циклогеназу (CINOD); глюкокортикостероиды (вводимые либо пероральным, внутримышечным, внутривенным или внутрисуставным путем); метотрексат, лефлуномид; гидроксихлороквин, d-пеницилламин, ауранофин или другие парентеральные или пероральные препараты золота; анальгетики; диацереин; внутрисуставные терапевтические средства, такие как производные гиалуроновой кислоты; и пищевые добавки, такие как глюкозамин.
Связывающий элемент в соответствии с изобретением и один или более из перечисленных выше дополнительных медицинских компонентов могут использоваться в производстве лекарственного средства. Лекарственное средство может представлять собой такое для отдельного или сочетанного введения индивидууму, и в соответствии с этим может включать связывающий элемент и дополнительный компонент в виде комбинированного препарата или в виде отдельных препаратов. Отдельные препараты могут использоваться для улучшения отдельного и последовательного или одновременного введения, и позволяют вводить компоненты различными путями, например с помощью перорального и парентерального введения.
В соответствии с настоящим изобретением обеспечиваемые композиции могут вводиться млекопитающим. Введение обычно осуществляют в "терапевтически эффективном количестве", которое является достаточным для того, чтобы быть полезным пациенту. Такая польза может представлять собой, по крайней мере, облегчение, по крайней мере, одного симптома. Фактически вводимое количество, а также скорость и динамика введения, будут зависеть от природы и тяжести состояния, которое подвергают лечению, определенного млекопитающего, которого подвергают лечению, клинического состояния индивидуального пациента, причины расстройства, места доставки композиции, типа связывающего элемента, способа введения, режима введения и других факторов, известных лечащим врачам. Пропись лечения, например решения в отношении дозы, находятся в пределах компетенции лечащих врачей и других медицинский докторов и могут зависеть от тяжести симптомов и/или развития заболевания, которое подвергается лечению. Приемлемые дозы антитела являются хорошо известными в области техники [67. Ledermann J.A. et al. (1991) Int. J. Cancer 47:659-664; 68. Bagshawe K.D. et al. (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4:915-922]. Могут использоваться специфические дозировки, указанные в данной заявке или в во врачебных кабинетных рекомендациях (2003), как является приемлемым для типа лекарственного средства, которое вводится. Терапевтически эффективное количество или приемлемая доза связывающего элемента в соответствии с изобретением может быть определена путем сравнения его in vitro активности и in vivo активности на животной модели. Способы для экстраполяции эффективных дозировок у мышей и других исследуемых животных на человека являются известными. Точная доза будет зависеть от ряда факторов, включая является ли антитело таковым для диагностики, предотвращения или для лечения, размера и локализации участка, который подвергается обработке, точной природы антитела (например, цельное антитело, фрагмент или диатело) и природы любой используемой для выявления метки или другой молекулы, присоединенной к антителу. Типичная доза антитела будет находиться в интервале от 100 мкг до 1 г для системных применений и от 1 мкг до 1 мг для местных применений. Может вводиться начальная более высокая ударная доза, после которой следуют одна или больше более низких доз. Как правило, антитело будет представлять собой цельное антитело, например IgG1 изотипа. Это представляет собой дозу для одного лечения взрослого пациента, которая может быть пропорционально подогнана для детей и младенцев, а также подогнана для других форматов антитела пропорционально молекулярному весу. Лечения могут повторяться при ежедневных, два раза в неделю, еженедельных или ежемесячных, интервалах, по усмотрению лечащего врача. Лечения могут повторяться каждые от двух до четырех недель для подкожного введения и каждые от четырех до восьми недель для внутривенного введения. Лечение может быть периодическим, и период между введениями составляет приблизительно две недели или более, например приблизительно три недели или более, приблизительно четыре недели или более или приблизительно один раз в месяц. Лечение может быть проведено перед и/или после хирургического лечения, и/или может вводиться или применяться непосредственно к анатомическому сайту хирургического лечения.
Краткое описание таблиц и фигур
Таблица 1 список аминокислотных последовательностей CDR тяжелой цепи и CDR легкой цепи каждого из антител 1-28.
Таблица 2а показывает пример эффективностей, идентифицированных из библиотек направленного мутагенеза при исследовании с помощью HTRF® анализов связывания рецептор-лиганд. Таблица 2b показывает связывающую аффинность (KD) для типичных связывающих элементов в соответствии с изобретением по отношению к человеческому IgE и IgE обезьяны циномолгуса при использовании SPR (BIACORE). Таблица 2b также показывает эффективность, выраженную как IC50, для типичных связывающих элементов в соответствии с изобретением в RBL-ER51 анализе передачи кальциевого сигнала (время - 4 часа с 25 нг/мл IgE человека или 100 нг/мл IgE обезьяны циномолгуса).
Таблица 3а показывает последовательности типичных связывающих элементов в соответствии с изобретением, приведенные в списке последовательностей, где SEQ ID NOS соответствуют таким, как показано в Таблице 3а.
Таблица 3b показывает ДНК VL и аминокислотные последовательности VL типичных связывающих элементов в соответствии с изобретением из предварительной заявки, которые показаны в приложенном списке последовательностей, где SEQ ID NOS соответствуют таким, как показано в Таблице 3b.
Таблица 4. Пример подсчета связывающей аффинности при использовании анализа BIAcore и измерений эффективности при использовании RBL-ER51 анализа передачи кальциевого сигнала для герминизированных антител.
Таблица 5 показывает непосредственные взаимодействия между IgE Cε3-Cε4 и Fab Антитела 11 в рентгеновских кристаллографических исследованиях для взаимодействия 1.
Таблица 6 непосредственные взаимодействия между IgE Cε3-Cε4 и Fab Антитела 11 в рентгеновских кристаллографических исследованиях для взаимодействия 2.
Таблица 7 показывает кристаллические параметры и обработку данных рентгеноскопического исследования и уточняющую статистику из рентгеновских кристаллографических исследований.
Таблица 8 показывает модель заключительного исследования изучения безопасности (Пример 8).
Фигура 1: относится к Примеру 2.7 и показывает молярную концентрацию антитела, выраженную как log на оси Х и пик высоты в RBL-ER51 анализе передачи кальциевого сигнала на оси Y. Незаштрихованные квадраты относятся к антителу 11, заштрихованные - к нерелевантному IgG1 контрольному антителу, а перевернутые незаштрихованные треугольники - к анти-IgE перекрестно связывающему антителу (Biosource). Следует отметить, что заштрихованные и незаштрихованные фигуры перекрываются на этой фигуре.
Фигура 2 показывает последовательность Сε3-4 FLAG His10 циномолгуса.
Фигура 3 показывает последовательность вариабельной тяжелой цепи, которая кодирует scFv (D12_VH) человеческого антиэстрадиола и один гаплотип гена IgHE циномолгуса (cyIgHE TQ).
Фигура 4 показывает последовательность вариабельной тяжелой цепи, которая кодирует scFv (D12_VH) человеческого антиэстрадиола и один гаплотип гена IgHE циномолгуса (cyIgHE ME).
Фигура 5 показывает последовательность вариабельной легкой цепи scFv (D12_VL) человеческого антиэстрадиола и гены cyIGLC 4 лямбда константного участка циномолгуса, ряд последовательностей: от 1 до 708.
Фигура 6 показывает последовательность вариабельной легкой цепи scFv (D12_VL) человеческого антиэстрадиола и гены D12_VL cyIGLC 7 лямбда константного участка циномолгуса.
Фигура 7 относится к Примеру 3 и показывает процент ингибирования максимальной экспрессии IgE в В клетках, не обработанных с помощью анти-IgE, где ось Х представляет собой концентрацию Антитела 11 в нМ, а ось Y представляет собой процент ингибирования. Верхняя диаграмма относится к Антителу 11, а нижняя диаграмма - к контрольному антителу.
Фигура 8 относится к Примеру 4 и показывает процентное ингибирование общего высвобождения β-гексозаминидазы +/- стандартная ошибка среднего, где ось Х представляет собой концентрацию Антитела 11 в нМ, а ось Y - процент ингибирования. Верхняя диаграмма относится к Антителу 11, а нижняя диаграмма - к контрольному антителу.
Фигура 9 относится к Примеру 5. Фигура показывает процент связанного IgE в человеческой сыворотке с повышением концентраций анти-IgE антитела, Антитела 11. Ось Х измеряет концентрацию Антитела 11 в микрограммах/мл, а ось Y показывает количество связанного IgE как процент от свободного IgE, проанализированный и выстроенный как функция от общего IgE.
Фигура 10 относится к Примеру 7 и показывает
a) представление IgE Сε3-Сε4 димера в виде полоски, где два мономера обозначаются как IgE1 и IgE2 и взаимодействуют с двумя молекулами Fab Антитела 11. Гликозилирование в Asn394 показано как шаро-стержневая модель; и
b) повернутый на 90 градусов вид сверху с вершины, показывающий, что большинство взаимодействий IgE с Fab фрагментом обеспечивается тяжелой цепью. Фигура получена при использовании программы PyMOL (DeLano Scientific LLC, San Carlos, California, U.S.A)
Фигура 11 относится к Примеру 7 и показывает гликозилирование в положении Asn394 IgE.
Фигура 12 относится к Примеру 8 и показывает среднее значение токсикокинетических профилей Антитела 11 IgG1, Антитела 11 IgG2 и Е48 после введения дозировок 1 мг/кг (День 1), 30 мг/кг (День 8) и 100 мг/кг (День 16 и выше) обезьянам циномолгус. Строка ошибки представляет стандартные отклонения. Ось Y представляет сывороточную концентрацию антитела, а ось Y - время в днях после введения первой дозы. Группа 1 (Антитело 11 IgG1) показана как заштрихованные кружочки. Группа 2 (Антитело 11 IgG2) показана как незаштрихованные треугольники, а Группа 3 (различные анти-IgE молекулы Е48) показана как заштрихованные квадраты.
Фигура 13 относится к Примеру 8 и показывает профили среднего значения свободного IgE у обезьян циномолгус, которые получали еженедельные дозы Антитела 11 IgG1, Антитела 11 IgG2 и Е48 (1 мг/кг в день 1,30 мг/кг в день 8, и 100 мг/кг в день 16 и выше). Строка ошибки представляет стандартные отклонения. Ось Y представляет концентрацию IgE в нг/мл, а ось Х представляет собой время в днях. Группа 1 (Антитело 11 IgG1) показана как заштрихованные кружочки. Группа 2 (Антитело 11 IgG2) показана как незаштрихованные треугольники и Группа 3 (различные анти-IgE молекулы Е48) показана как заштрихованные квадраты.
Фигура 14 относится к Примеру 8 и показывает кривую количества тромбоцитов (х109/л), выраженную как процентное изменение по сравнению со средним значением объемов 2 предварительных доз против концентрации в плазме крови животных в Группе 1 (обработанные Антителом 11). Эта кривая является репрезентативной для других 16 животных в пределах 3 групп, что демонстрирует отсутствие значимого влияния на тромбоциты. Ось Х показывает время в часах, левая ось Y - уровень тромбоцитов как процент изменения по сравнению со средним уровнем перед обработкой, а левая ось Y - концентрацию анти-IgE антитела в нмол/л. Заштрихованные квадраты показывают концентрацию тромбоцитов, а заштрихованные ромбы - концентрацию анти-IgE антитела. Заштрихованные треугольники показывают дозу анти-IgE антитела в мг/кг.
Фигура 15 относится к Примеру 8 и показывает кривую количества тромбоцитов (х109/л), выраженную как процентное изменение по сравнению со средним значением объемов 2 предварительных доз против концентрации в плазме крови животных в Группе 1 (обработанные Антителом 11 IgG1), которая демонстрирует временно падение (35% ниже базовой линии) в количестве тромбоцитов на день 29. Ось Х показывает время в часах, левая ось Y - уровень тромбоцитов как процент изменения по сравнению со средним уровнем перед обработкой, а левая ось Y - концентрацию анти-IgE антитела. Заштрихованные треугольники показывают дозу анти-IgE антитела в мг/кг.
Фигура 16 относится к Примеру 9 и показывает ингибирование Антителом 11 опосредованной IgE/FceRI цитотоксичности. Ось Х представляет собой молярную концентрацию Антитела 11, а ось Y - процент цитотоксичности. На диаграмме незаштрихованные треугольники и заштрихованные кружочки относятся к Mov18 IgE эксперименту, где незаштрихованные треугольники представляют собой контроль изотипа, а заштрихованные кружочки - Антитело 11, на диаграмме также выделенный жирным шрифтом незаштрихованный треугольник (который скрыт под точками с правой стороны диаграммы) и незаштрихованный кружочек относятся к NIP IgE контролю, где незаштрихованный, выделенный жирным шрифтом треугольник представляет собой контроль изотипа, а незаштрихованный кружочек является Антителом 11.
Фигура 17 относится к Примеру 9 и показывает ингибирование Антителом 11 опосредованного IgE/CD23 фагоцитоза. Ось Х представляет собой молярную концентрацию Антитела 11, а ось Y - процент фагоцитоза. На диаграмме незаштрихованные треугольники и заштрихованные кружочки относятся к Mov18 IgE эксперименту, где незаштрихованные треугольники представляют собой контроль изотипа, а заштрихованные кружочки - Антитело 11, на диаграмме также выделенный жирным шрифтом незаштрихованный треугольник и незаштрихованный кружочек относятся к NIP IgE контролю, где незаштрихованный, выделенный жирным шрифтом треугольник представляет собой контроль изотипа, а незаштрихованный кружочек является Антителом 11.
Примеры
Библиотеки фагового дисплея интактного человеческого одноцепочечного Fv (scFv) клонировали в фагмидный вектор на основе нитчатого фага М13 и использовали для селекции [69. Vaughan, Т. J., et al. (1996). Nature Biotechnology 14, 309-314; 70. Hutchings, C. Generation of Naive Human Antibody Libraries, in Antibody Engineering, R. Kontermann and S. Dubel, Editors. 2001, Springer Laboratory Manuals, Berlin, p.93]).
Специфическое анти-IgE scFv антитело изолировали из библиотек фагового дисплея при использовании серий циклов селекции на рекомбинантном человеческом IgE.
Отобранные scFv антитела оптимизировали в отношении связывания с человеческим IgE и/или в отношении эффективности и подвергали переформатированию в качестве IgG антител.
ПОСЛЕДОВАТЕЛЬНОСТИ
Последовательности типичных связывающих элементов в соответствии с изобретением показаны в приложенном списке последовательностей, где SEQ ID NOS соответствует такому, как показано в Таблице 3а, приведенной ниже, где:
i) если номер антитела сопровождается указанием GL, например 8GL, то это относится к антителу, в котором один или более остатков подверглись обратным мутациям до эмбриональной конфигурации, в общем случае если используется GL, то это означает, что все негерминизированные остатки, которые могут подвергаться обратной мутации до эмбриональных без способной к определению потери активности, были герминизированы; и
ii) если номер антитела сопровождается указанием PGL, например 11PGL, то это относится к антителу, в котором один или более остатков подверглись обратным мутациям до эмбриональной конфигурации, в общем случае если используется PGL, то это означает, что больше остатков было подвергнуто обратной мутации до эмбриональных, чем в случае GL, но это приводило к некоторой потере активности по сравнению с негерминизированным антителом.
В списке последовательностей, поданном с временной заявкой (временная заявка US с номером: 60/901304) последовательности 3' ggt кодона и соответствующий остаток глицина, показанные в нуклеотидной и аминокислотной последовательности для ДНК VL и соответствующей аминокислотной последовательности VL, были включены в экспрессированные последовательности scFv и IgG этого антитела. С-терминальный остаток глицина последовательности соответствует остатку 108 Кэбата. Этот терминальный глицин не является частью VL последовательности и был удален из последовательности, приведенной в Таблице 3а. Последовательности для ДНК VL и аминокислотной VL из временной заявки включены в список последовательностей и приведены в Таблице 3b ниже. Происхождение этого остатка и кодируемого им триплета ggt объясняется ниже.
Для экспрессии легкой цепи IgG обеспечивается нуклеотидная последовательность, кодирующая легкую цепь антитела, включающая первый экзон, кодирующий VL домен, и второй экзон, кодирующий CL домен, а также интрон, отделяющий первый экзон от второго экзона. При нормальных условиях интрон выщепляется с помощью механизма процессинга мРНК, соединяя 3' конец первого экзона с 5' концом второго экзона. Таким образом, когда ДНК, содержащая указанную нуклеотидную последовательность, экспрессируется как РНК, первый и второй экзоны сплайсируются вместе. Трансляция сплайсированной РНК обеспечивает получение полипептида, включающего VL домен и CL домен. После сплайсинга Gly при остатке Кэбата 108 кодируется последним основанием (g) последовательности VL домена каркасного участка 4 и первыми двумя основаниями (gt) CL домена.
Таким образом, остаток глицина при остатке Кэбата 108 был включен в список последовательностей VL последовательностей во временной заявке, но как описано выше, не может считаться С-терминальным остатком VL домена молекулы антитела и, таким образом, был удален из списка последовательностей в Таблице 3а.
В списке последовательностей временной заявки последовательности, приведенные как Антитела 29-34, приведены в Таблице 3а как Антитела 8GL, 8PGL, 11GL и 11PGL. Некоторые из этих антител имеют общий VL домен и, в результате этого, некоторые идентификационные номера последовательностей в списке последовательностей, обеспечиваемые во временной заявке, являются пустыми. Правильный состав антител является следующим:
Антитело 29 соответствует 8GL VH домену.
Антитело 30 соответствует 8PGL VH домену.
Антитело 31 соответствует VL домену, общему для 8GL и 8PGL.
Антитело 32 соответствует 11GL VH домену.
Антитело 33 соответствует 11PGL VH домену.
Антитело 34 соответствует VL домену, общему для 11GL и 11PGL.
Последовательности с идентификационными номерами 286, 292, 293, 304, 310 и 311 являются пустыми. Это было исправлено в Таблице 3а.
Изобретение будет далее представлено следующими примерами, которые не ограничивают его:
Пример 1. Пропись для изоляции
1.1 Селекция
Библиотеки фагового дисплея интактного человеческого одноцепочечного Fv (scFv), клонированные в фагмидный вектор на основе нитчатого фага, использовали для отбора (Vaughan и др., Nature Biotechnology 14:309-314 (1996), Hutchings, Antibody Engineering, R-Kontermann и S. Dubel, ред. 2001, Springer Laboratory Manuals, Berlin. P93). Анти-IgE специфические scFv антитела изолировали из библиотек фагового дисплея при использовании серий циклов селекции либо на плазматическом очищенном человеческом IgEK (Calbiochem), либо на плазматическом очищенном человеческом IgEλ (Biodesign), существенно так, как ранее описано Vaughan и др. (Vaughan и др., Nature Biotechnology 14:309-314 (1996). Вкратце, для осуществления селекционного пэннинга человеческий IgE в PBS (PBS Дюльбекко, рН 7,4) адсорбировали на микротитровальном планшете Maxisorp (Nunc) в течение ночи при 4°С. Ячейки промывали с помощью PBS, а потом блокировали в течение одного часа при использовании PBS-Marvel (3% вес./об.). Очищенный фаг в PBS-Marvel (3% вес./об.) прибавляли к ячейкам и осуществляли связывание с антигеном, адсорбированным в ячейках в течение 1 часа. Несвязанный фаг удаляли с помощью серий циклов промывания при использовании PBS-Твин (0,1% об./об.) и PBS. Частицы связанного фага элюировали, вводили в бактерию и «спасали» для следующего цикла селекции (Vaughan и др., Nature Biotechnology 14:309-314 (1996)). Осуществляли попеременные циклы селекции при использовании каппа и лямбда форм IgE.
1.2 Ингибирование связывания IgE с FcεRI при использовании неочищенного scFv
Репрезентативное число индивидуальных scFv из второго цикла селекции выращивали в планшетах на 96 ячеек. ScFv экспрессировали в периплазме бактерий и подвергали скринингу на их ингибиторную активность в гомогенном анализе на основе FRET (резонансный перенос энергии флуоресценции) связывания человеческий IgE/человеческий FceRI. В этом анализе образцы конкурировали за связывание человеческого IgE (Calbiochem 401152), меченного с помощью хелата европия (Perkin Elmer 1244-302), с человеческим FcεRI-Fc (клетки NSO, полученные на фирме). Подробный способ анализа обеспечивается в разделе "Материалы и способы".
1.3 Ингибирование связывания IgE с FcεRI при использовании очищенного scFv
ScFv, который продемонстрировал значительное ингибиторное влияние на взаимодействие IgE:FcεRI, в виде неочищенных периплазматических экстрактов подвергали секвенированию ДНК (Vaughan и др. 1996, Nature Biotechnology 14: 309-314), (Osboum 1996; Immunotechnology. 2, 181-196). Уникальные scFv вновь экспрессировали в бактериях и очищали с помощью аффинной хроматографии (как описано Bannister и др. (2006) Biotechnology and bioengineering, 94. 931-937). Эффективности этих образцов определяли путем конкуренции серийных разведении очищенного препарата против FcεRI (клетки NS0, полученные на фирме), за связывание с человеческим IgE (Calbiochem 401152), меченным хелатом европия (Perkin Elmer 1244-302). Очищенные препараты scFv, например. Антитело 1 являются способными ингибировать взаимодействие IgE-FcεRI. Подробные прописи обеспечивается в разделе "Материалы и способы".
1.4 Переформатирование scFv до IgG1
Клоны переводили из scFv формата в IgG формат с помощью субклонирования VH и VL доменов в векторы, экспрессирующие тяжелые и легкие цепи антитела, соответственно. VH домен клонировали в вектор (pEU15.1 или pEU9.2), содержащий константные домены тяжелой цепи человека и регуляторные элементы для экспрессии цельной тяжелой цепи IgG1 или IgG2 в клетках млекопитающих, соответственно. Подобно этому VL домен клонировали либо в вектор pEU3.4 для экспрессии каппа легкой цепи человека, либо в pEU4.4 для экспрессии константных доменов лямбда легкой цепи человека, с регуляторными элементами для экспрессии цельной легкой цепи IgG в клетках млекопитающих. Векторы для экспрессии тяжелых цепей и легких цепей были описаны Persic и др. (Persic, L., и др. (1997) Gene 187, 9-18). Векторы Cambridge Antibody Technology конструировали для включения EBV OriP элемента, который в сочетании с EBNA1 белком позволяет осуществлять эписомальную репликацию плазмиды. Для получения IgG векторы, экспрессирующие тяжелую и легкую цепь IgG, трансфицировали в EBNA-HEK293 клетки млекопитающих. IgG экспрессировали и секретировали в среду. Полученные продукты объединяли и фильтровали перед очисткой. IgG очищали при использовании хроматографии на основе белка А. Супернатанты культур загружали на керамическую колонку с белком А (BioSepra) и промывали с помощью 50 мМ Трис-HCl рН 8,0,250 мМ NaCl. Связанный IgG элюировали из колонки при использовании 0,1 М цитрата натрия (рН 3,0) и нейтрализовали путем прибавления Трис-HCl (рН 9,0). Заменяли буфер в элюированном материале на PBS при использовании Nap10 колонок (Amersham, #17-0854-02) и определяли концентрацию IgG спектрофотометрически при использовании коэффициента экстинкции на основе аминокислотной последовательности IgG (Mach и др. Anal. Biochem. 200(1):20-26, 1992). Очищенный IgG подвергали анализу на агрегацию или деградацию при использовании SEC-ВЭЖХ и с помощью SDS-ПАГЭ.
1.5 Ингибирование передачи кальциевого сигнала в RBL-ER51 клетках с помощью очищенного scFv и IgG
Эффективность нейтрализации для очищенных препаратов scFv и IgG против биоактивности человеческого IgE, опосредованной через FcεRI, оценивали при использовании RBL-ER51 анализа передачи кальциевого сигнала. RBL-2H3 клетки (линия базофилов крысы) стабильно трансфицировали с помощью человеческого FcεRI (RBL-ER51 клетки). Свободный IgE, находящийся поблизости клеток, связывается с FcεRI на поверхности клеток, и последующее перекрестное связывание связанного с рецептором IgE приводит к мобилизации кальция, что может быть определено при использовании устройства для считывания флуоресцентного изображения (FLIPR). Подробное описание прописи обеспечивается в разделе "Материалы и способы".
Очищенные препараты scFv Антитела 1 являются способными к ингибированию индуцированной IgE передачи сигнала RBL-ER51 клетками при максимальной исследуемой концентрации. При анализе очищенного IgG значения IC50 для Антитела 1 было подсчитано как такое, которое равно 34 нМ.
1.6 Селективность и видовая перекрестная реактивность антител в DELFIA ® анализах конкуренции эпитопов
Видовую перекрестную реактивности и селективность антител по отношению к IgE и структурно родственным молекулам, IgA, IgM, IgD и IgG, устанавливали при использовании DELFIA® анализов конкуренции эпитопов. Анализ определяет относительную перекрестную реактивность путем измерения ингибирования биотинилированного IgE (очищенный, из плазмы крови, BIODESIGN International), связывающего каждое иммбилизованное анти-IgE антитело.
Титрования очищенного IgA, IgM, IgD, и IgG (все Calbiochem) анализировали в каждом анализе для установления профиля специфичности для каждого структурно родственного белка, как измеряется с помощью значений IC50 в этом анализе.
Титрования видовых IgE, включая IgE Сε3-Сε4 домен циномолгуса (полученный на фирме из HEK-EBNA клеток), человеческий IgE Cε3-Cε4 домен (полученный на фирме из HEK-EBNA клеток) и человеческий IgE лямбда (BIODESIGN International), подвергали тестированию в каждом анализе для установления видовой перекрестной реактивности антител. Полноразмерный человеческий IgEλ, вместе с IgE Сε3-Сε4 доменами человека и циномолгуса, обеспечивал кривые ингибирования. Не наблюдали никакого ингибирования для любого из структурно родственных белков. Эти данные демонстрируют тот факт, что Антитело 1 связывается с человеческим IgEλ, Сε3-Сε4 доменом IgE и является перекрестно реактивным с IgE циномолгуса. Однако Антитело 1 не связывается с каким-либо другими наиболее родственными IgE белками человека. Подробное описание прописи обеспечивается в разделе "Материалы и способы".
1.7 Ингибирование связывания IgE с CD23 с помощью очищенного IgG
IM9 клетки (линия В клеток человека) были продемонстрированы как такие, которые экспрессируют CD23 но не FcεRI, при основных условиях. IgE связывается с CD23 на поверхности IM9 клеток. Связанный с CD23 IgE может потом связываться с анти-IgE-фикоэритрином (Caltag) и определяться с помощью проточной цитометрии (FACSCalibur, BD Biosciences).
Антитела могут быть оценены на ингибирование взаимодействия IgE/CD23. Подробное описание прописи обеспечивается в разделе "Материалы и способы". Вкратце, титрования исследуемого IgG смешивали с IgE перед инкубацией с IM9 клетками. После одночасовой инкубации клетки промывали и связанный IgE определяли с помощью комплекса анти-IgE-фикоэритрин (Caltag). Антитело 1 ингибировало взаимодействие IgE/CD23 со значением IC50 16 нМ (n=3).
1.8 Перекрестное связывание связанного с FCεRI IgE
Антитела подвергали оценке на потенциал перекрестного связывания связанного с FcεRI IgE при использовании RBL-ER51 анализа кальциевого сигнала. RBL-ER51 клетки, описанные в разделе "Материалы и способы", нагружали IgE. Антитела инкубировали с нагруженными IgE клетками и оценивали их способность стимулировать кальциевый ответ. Антитело 1 было не способным индуцировать способный к определению кальциевый ответ.
Пример 2. Оптимизация антитела
2.1 Оптимизация исходного клона с помощью направленного мутагенеза
Антитело 1 подвергали оптимизации с помощью подхода направленного мутагенеза с селекцией фагового дисплея на основе аффинности. Для подхода направленного мутагенеза большие scFv фаговые библиотеки, которые имеют происхождение от ведущего клона, создавали с помощью олигонуклеотид-направленного мутагенеза участков 3, определяющих комплементарность (CDR3), вариабельной тяжелой (VH) и легкой (VL) цепи, как описано Clackson и Lowman 2004 (A Practical Approach, 2004. Oxford University Press).
Библиотеки подвергали селекции фагового дисплея на основе аффинности для того, чтобы отобрать варианты с более высокой аффинностью для IgE. Вследствие этого они будут демонстрировать улучшенную ингибиторную активностьв отношении связывания IgE с FcεRI. Такая селекция может быть осуществлена существенно так, как описано ранее (Thompson 1996; Journal of Molecular Biology. 256. 77-88). Вкратце, scFv фаговые частицы инкубировали в растворе с биотинилированным человеческим IgE λ (получен из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167] и модифицирован на фирме). ScFv-фаг, связанный с антигеном, потом захватывали с помощью парамагнитных гранул, покрытых стрептавидином (Dynabeads® M280) в соответствиями с рекомендациями производителя. Отобранные частицы scFv-фаг потом восстанавливали так, как описано ранее (Vaughan и др., Nature Biotechnology 14:309-314 (1996)), и процесс селекции повторяли в присутствии повышающихся концентраций био-человеческого IgE (от 250 нМ до 25 пМ в течение 5 циклов).
После завершения 5 циклов селекции VH и VL рандомизированные библиотеки подвергали рекомбинации с образованием одной библиотеки, в которой клоны содержали случайным образом спаренные индивидуально рандомизированные VH и VL последовательности. Селекцию потом продолжали так, как описано ранее в присутствии повышающихся концентраций биочеловеческого IgE (от 100 пМ до 1 пМ в течение дополнительных 3 циклов).
2.2 Идентификация улучшенных клонов, полученных в результате направленного мутагенеза, при использовании биохимического анализа антитело-лиганд
ScFv, полученные в результате селекции продуктов направленного мутагенеза экспрессировали в бактериальной периплазме и подвергали скринингу в формате эпитопного конкурентного HTRF® (однородная флуоресценция с разрешением во времени) анализа на ингибирование связывания человеческого IgE (получен из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]), меченного с помощью криптата европия (CIS bio International 62EUSPEA), с античеловеческим IgE (Антитело 1, изолированное в Примере 1). Подробный способ анализа обеспечивается в разделе "Материалы и способы". ScFv, который продемонстрировал значительный ингибиторный эффект, подвергали секвенированию ДНК, и единственный scFv получали в виде очищенных препаратов.
2.3 Ингибирование связывания IgE с FcεRI с помощью очищенного scFv
Очищенный scFv исследовали в формате HTRF® анализа связывания рецептор-лиганд (гомогенная флуоресценция с разрешением во времени) для ингибирования связывания либо человеческого IgE (получен из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]) либо IgE циномолгуса (рекомбинантный, смотри раздел "Материалы и способы"), меченного с помощью криптата европия (CIS bio International 62EUSPEA), с человеческим FcεR1-Fc (получен на фирме из NSO клеток). Пример данных относительно эффективности scFv включен в Таблицу 2а
2.4 Ингибирование передачи кальциевого сигнала в RBL-ER51 клетках с помощью очищенного IgG
После переформатирования в качестве IgG эффективности оптимизированных клонов определяли при использовании модифицированного RBL-ER51 анализа передачи кальциевого сигнала. Этот анализ был адаптирован из способа, используемого при направленной изоляции для улучшения чувствительности с целью определения более эффективных антител. Подробное описание прописи обеспечивается в разделе "Материалы и способы". Данные относительно IC50 эффективности в отношении IgE человека и циномолгуса приведены в Таблице 2b.
2.5. Германизация
Аминокислотные последовательности VH и VL доменов оптимизированных анти-IgE эмбриональной линии в базе данных VBASE (Tomlinson 1997; Journal of Molecular biology. 224. 487-499), и самую близкую зародышевую линию идентифицировали в отношении подобности последовательности. Для VH доменов Антитела 1 такая линия клеток представляла собой Vh1 DP-3 (1-f). Для VL доменов такая линия представляла собой VU DPL8 (1е).
Без учета остатков Верньера (Foote & Winter 1992), которые оставались без изменений, было осуществлено 10 изменений по сравнению с эмбриональной линией в каркасных участках VH домена и 2 изменения в VL домене Антитела 1. Пять изменений в VH домене и оба изменения в VL домене подвергались обратному превращению до ближайшей эмбриональной последовательности в отношении идентично выровненных человеческих антител. Изменения в остатках Кэбата с номерами 1, 20, 82а, 83 и 89 VH домена оставляли без изменений для сохранения эффективности (Антитело 8 GL и Антитело 11 GL). Герминизацию этих аминокислотных остатков осуществляли при использовании стандартных методик сайт-направленного мутагенеза с приемлемыми мутагенными праймерами. Герминизированный IgG потом подвергали повторному анализу для подтверждения отсутствия снижения аффинности или эффективности. Пример аффинностей и эффективностей для герминизированных (GL) антител обеспечивается в Таблице 4.
2.6 Ингибирование связывания IgE c CD23 с помощью очищенного IgG
Некоторые оптимизированные антитела оценивали на ингибирование взаимодействия IgE/CD23 при использовании IM9 анализа связывания, как было описано ранее. Антитела, исследованные в этой системе, были определены как такие, которые ингибируют взаимодействие IgE/CD23. Значения IC50 для Антитела 8 и Антитела 11 составляли 16,5 нМ и 23 нМ, соответственно.
2.7 Перекрестное связывание IgE, связанного с FcεRI
Некоторые оптимизированные антитела оценивали на способность перекрестно связываться с IgE, связанным с FcεRI, при использовании RBL-ER51 анализа передачи кальциевого канала. RBL-ER51 клетки, описанные в разделе "Материалы и способы", максимально нагружали IgE. Оптимизированные антитела инкубировали с нагруженными IgE клетками и подвергали оценке на их способность стимулировать кальциевый ответ. Не определяли никакого сигнала (Фигура 1).
2.8. Селективность и видовая перекрестная реактивность оптимизированных антител в DELFIA® анализах конкуренции эпитопов
Селективность и видовую перекрестную реактивность направленных антител повторно оценивали при использовании DELFIA® анализа конкуренции эпитопов, как описано ранее (смотри раздел 1.6 и "Материалы и способы")
Титрования очищенных IgA, IgM, IgD и IgG (все Calbiochem) анализировали в каждом анализе для установления профиля специфичности для каждого структурно родственного белка, как измеряется с помощью значений IC50 в этом анализе.
Титрования видовых IgE, включая человеческий IgEλ (полученный из U266), человеческий IgEκ (Calbiochem), IgE Сε3-Сε4 домен циномолгуса (полученный на фирме из HEK-EBNA клеток) и человеческий IgE Сε3-Сε4 домен (полученный на фирме из HEK-EBNA клеток) подвергали тестированию в каждом анализе для установления видовой перекрестной реактивности антител. Полноразмерный человеческий IgEλ и κ, вместе с IgE Сε3-Сε4 доменами обеспечивали кривые ингибирования. Не наблюдали никакого ингибирования для любого из структурно родственных белков человека (IgA, IgM, IgD и IgG). Эти данные демонстрируют тот факт, что панель исследуемых антител связывается с человеческим IgEλ и κ, Сε3-Сε4 доменом IgE и является перекрестно реактивной по отношению к IgE циномолгуса. Однако антитела не связывается с наиболее родственными IgE белками.
2.9 Подсчет связывающей аффинности на основе данных аффинности для оптимизированных клонов при использовании BIAcore
Связывающую аффинность очищенных образцов IgG репрезентативного количества клонов IgE человека и циномолгуса определяли с помощью поверхностного плазменного резонанса при использовании BIAcore 2000 биосенсора (BIAcore AB), существенно так, как описано Karlsson и др. 1991; Journal oflmmunological Methods 145 (1-2) 229-240. Вкратце, белок G' (Sigma Aldrich, P4689) ковалентно связывали с поверхностью СМ5 сенсорного чипа при использовании стандартных реагентов для аминного связывания в соответствии с инструкциями производителя (BIAcore). Этот поверхностный белок G' использовали для захвата очищенных анти-IgE антител с помощью Fc домена с обеспечением поверхностной плотности 50RU. Человеческий IgEX или IgE циномолгуса, приготовленный в HBS-EP буфере (BIAcore AB), в диапазоне концентраций от 125 нМ до 7,6 нМ, пропускали над поверхностью сенсорного чипа. Восстанавливали поверхность при использовании 10 мМ глицина, рН 1,75 между каждым введением антитела. Полученные сенсограммы оценивали при использовании программного обеспечения BIA evaluation 3.1 и подгоняли к бивалентной аналитической модели для обеспечения относительных данных связывания. Примеры аффинностей для анализируемого IgG показаны в Таблице 2b и Таблице 4.
Материалы и способы - Пример 1 и 2
Ингибирование связывания IgE с FcεRI с помощью неочищенного scFv
Результаты селекции подвергали скринингу в анализе FRET (резонансного переноса энергии флуоресценции) гомогенного связывания рецептор-лиганд на основе аналитического формата для ингибирования связывания человеческого IgE (Calbiochem 401152), меченного хелатом европия (Perkin Elmer 1244-302), с человеческим FcεRI-Fc (полученный из NS0 клеток на фирме).
Результаты, полученные во время направленной изоляции, подвергали скринингу в виде неразведенных периплазматических экстрактов, содержащих неочищенный scFv, приготовленный в: 50 мМ MOPS буфер рН 7,4,0,5 мМ ЭДТА и 0,5 М сорбита.
15 мкл неочищенного образца scFv прибавляли к аналитическому планшету на 384 ячейки (Perkin Elmer 6006280). После этого осуществляли прибавление 15 мкл 11 нМ человеческого FceRI-Fc (на основе молекулярного веса 260 кДа), 15 мкл 40 нМ IgG к человеческому Fc, меченному XL665 (CIS Bio International 61HFCXLA), а потом прибавляли 15 мкл 0,75 нМ меченного европием человеческого IgE. Контроль неспецифического связывания определяли при использовании 300 нМ человеческого IgE (Calbiochem). Все разведения готовили в 50 мМ Трис-HCl (рН 7,8), содержащем 250 мМ хлорида натрия и 0,05% Твин-20 (аналитический буфер).
Потом инкубировали аналитические планшеты в течение 1,5 часа при комнатной температуре перед считыванием разрешенной во времени флуоресценции при длинах волн 615 нм и 665 нм последовательно при использовании устройства для считывания планшетов VICTOR2 (Perkin Elmer).
Данные нормализовали с помощью программного обеспечения VICTOR2 для подсчета числа импульсов за секунду (CPS). Значения CPS последовательно использовали для подсчета %специфического связывания, как описано в уравнении 1.
Уравнение 1:

Ингибирование связывания IgE с FcεRI с помощью очищенного scFv
Очищенный scFv из позитивных клонов, идентифицированных с помощью скрининга, подвергали исследованию в анализе FRET (резонансного переноса энергии флуоресценции) гомогенного связывания рецептор-лиганд на основе аналитического формата для ингибирования связывания человеческого IgE (Calbiochem 401152), меченного хелатом европия (Perkin Elmer 1244-302), с человеческим FcεRI-Fc (полученный из NSO клеток на фирме).
Титрования scFv концентраций использовали для того, чтобы установить эффективность scFv, как измеряется с помощью значений IC50 в анализе. 15 мкл для титрования очищенного образца scFv прибавляли к аналитическому планшету на 384 ячеек (Perkin Elmer 6006280). После этого осуществляли прибавление 15 мкл 11 нМ человеческого FcεRI-Fc (на основе молекулярного веса 260 кДа) и 15 мкл 40 нМ IgG к человеческому Fc, меченному XL665 (CIS Bio International 61HFCXLA), а потом прибавляли 15 мкл 0,75 нМ меченного европием человеческого IgE. Контроль неспецифического связывания определяли при использовании 300 нМ человеческого IgE (Calbiochem). Все разведения готовили в 50 мМ Трис-HCl (рН 7,8), содержащем 250 мМ хлорида натрия и 0,05% Твин-20 (аналитический буфер).
Потом инкубировали аналитические планшеты в течение 1,5 часа при комнатной температуре, перед считыванием разрешенной во времени флуоресценции при длинах волн 615 нм и 665 нм последовательно при использовании устройства для считывания планшетов VICTOR2 (Perkin Elmer).
Данные нормализовали с помощью программного обеспечения VICTOR2 для подсчета числа импульсов за секунду (CPS). Значения CPS последовательно использовали для подсчета % специфического связывания, как описано в уравнении 1.
Уравнение 1:

Определяли IC50 значения при использовании программного обеспечения GraphPad Prism путем подгонки кривой при использовании логистического уравнения с четырьмя параметрами (Уравнение 2).
Уравнение 2:
Y=Дно+(Bepx-Дно)/(1+10^((LogEC50-X)*уклон))
X представляет собой логарифм концентрации. Y представляет собой специфическое связывание.
Y начинается со Дна и поднимается до Верха в виде сигмоидальной функции.
Ингибирование передачи кальциевого сигнала с помощью очищенного scFv и IgG в RBL-2H3 клетках, стабильно трансфицированных человеческим FcεR1 (RBL-ER51 клетки)
Эффективность нейтрализации очищенных препаратов scFv и IgG относительно биоактивности человеческого IgE, опосредованной через FcεR1, оценивали при использовании RBL-ER51 анализа передачи кальциевого сигнала. Человеческий FcεRI клонировали из лимфоцитов периферической крови человека в рсДНК3.1 вектор и трансфицировали в RBL-2H3 клетки (линия клеток крысиных базофилов) при использовании стандартной методики электропорации. Трансфицированные клетки клонировали методом серийных разведении и анализировали на экспрессию поверхностного FcεRI. Полученные RBL-ER51 клетки поддерживали в среде, содержащей G418 (Invitrogen 10131-027) для поддержания стабильной экспрессии рецептора. Свободный IgE, находящийся поблизости клеток, связывается с FcεRI на поверхности клеток, и последующее перекрестное связывание связанного с рецептором IgE приводит к мобилизации кальция, что может быть определено при использовании устройства для считывания флуоресцентного изображения (FLIPR).
RBL-ER51 клетки высевали при концентрации 5×104/100 мкл/ячейка в культуральную среду [DMEM (Invitrogen 41966) с 9% об./об. неинактивированного с помощью прогревания FBS (Invitrogen 10100-147) и 400 мкг/мл G418 (Invitrogen 10131-027)] в черные планшеты на 96 ячеек с плоским дном, содержащие культуру ткани (Costar) и инкубировали при 37°C, 5% CO2 в течение 18-24 часов. После этого изымали среду, оставляя интактный монослой клеток и заменяли ее 100 мкл/ячейка FLUO-4AM загружающим буфером [DMEM с 0,1% FBS, 20 мМ HEPES, 2,5 мМ пробеницида и 2 мкг/мл FLUO-4AM (TeffLabs)] в течение 1-2 часов при 37°C, 5% CO2. Удаляли загружающий буфер и клетки трижды промывали с помощью 200 мкл/ячейка PBS. Удаляли заключительный промывочный раствор и заменяли его 70 мкл/ячейка FLIPR буфера [125 мМ NaCl2 5 нМ KCl, 1 мМ MgCl2, 1,5 мМ CaCl2, 30 мМ Hepes, 2,5 мМ пробеницида 5 мМ глюкозы, 0,01% об./об. FCS]. Планшеты инкубировали при 37°C, 5% CO2 в течение 5-45 минут.
Исследуемые растворы очищенного scFv или IgG (в двукратной повторности) разводили до желаемой концентрации в FLIPR буфере в планшетах с V-образным дном (Greiner). Нерелевантные антитела, ненаправленные на IgE, использовали в качестве негативного контроля. IgE (полученный от Calbiochem или из U266 клеток [Ikeyama и др.. 1986. Molecular Immunology 23 (2); стр. 159-167]) получали в FLIPR буфере и смешивали с приемлемым исследуемым антителом для получения IgE концентрации 3,33 мкг/мл в общем объеме 40 мкл/ячейка. Используемую в этом анализе концентрацию IgE выбирали как дозу, которая при заключительной концентрации анализа обеспечивает приблизительно 80% от максимального кальциевого ответа. Все образцы инкубировали в течение 30 минут при комнатной температуре, перед перенесением 30 мкл смеси IgE / антитело к клеткам, нагруженным красителем, приготовленным так, как описано выше. Аналитические планшеты инкубировали при 37°C в течение 10 минут для того, чтобы позволить свободному IgE связаться с RBL-ER51 клетками.
Для измерения мобилизации кальция после прибавления перекрестно связывающего анти-IgE FLIPR (Molecular Devices) откалибровывали для приемлемого воздействия в соответствии с инструкциями производителя. Анти-IgE (Biosource AHI0501), разведенный в FLIPR буфере, прибавляли к аналитическим планшетам до получения заключительной концентрации 10 мкг/мл. Регистрировали флуоресценцию FLUO-4AM красителя при 1-секундных интервалах на протяжении 80 измерений, после чего использовали 8-секундные интервалы на протяжении 40 измерений. Выводили пик ответа для каждой ячейки, и данные потом подвергали анализу при использовании программного обеспечения Graphpad Prism.
Измерение перекрестного связывания анти-IgE в RBL-ER51 клетках
Для измерения способности очищенных IgG перекрестно связывать IgE, связанный с FcεRI, готовили RBL-ER51 клетки и загрузку красителем так, как описано в анализе ингибирования. Клетки инкубировали в течение 10 минут в 100 мкл 1 мкг/мл человеческого IgE (полученный от Calbiochem или из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]), разведенного в FLIPR буфере, для того, чтобы позволить IgE связаться с FcεRI на поверхности клеток. Используемую в этом анализе концентрацию IgE выбирали как дозу, которая при заключительной концентрации анализа обеспечивает приблизительно 80% от максимального кальциевого ответа. Для измерения мобилизации кальция после прибавления перекрестно связывающего анти-IgE, FLIPR (Molecular Devices) откалибровывали для приемлемого воздействия в соответствии с инструкциями производителя. 30 мкл исследуемых антител, разведенных до приемлемой концентрации в FLIPR буфере, прибавляли к аналитическим планшетам, загруженным IgE. Анти-IgE (Biosource AHI0501) использовали как позитивный контроль. Регистрировали флуоресценцию красителя FLUO-4AM (TeffLabs) при 1-секундных интервалах на протяжении 80 измерений, после чего использовали 8-секундные интервалы на протяжении 40 измерений. Выводили пик ответа для каждой ячейки, и данные потом подвергали анализу при использовании программного обеспечения Graphpad Prism.
Селективность и видовая перекрестная реактивность антител в DELFIA® анализах конкуренции эпитопов
Очищенный IgG адсорбировали на микротитровальных планшетах на 96 ячеек Maxisorp (Nunc) в PBS при концентрации, которая обеспечивает значимый сигнал, когда прибавляется биотинилированный человеческий IgE при концентрации, которая приблизительно равна оцененному значению KD для частного IgG. Избыток IgG вымывали с помощью PBS-Твин (0,1% об ./об.) и блокировали ячейки с помощью PBS-Marvel (3% вес./об.) в течение 1 часа. Готовили серийные разведения для каждого из следующих конкурирующих соединений в PBS, начиная от концентрации, приблизительно равной 1000-кратному значению KD взаимодействия между биотинилированным человеческим IgE и соответствующим IgG; человеческим IgE лямбда (полученный из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]), человеческим IgE каппа (Calbiochem), человеческим IgE Cε3-Cε4 доменом (полученный из HEK-EBNA клеток на фирме), IgE Cε3-Cε4 доменом циномолгуса (полученный из HEK-EBNA клеток на фирме), человеческими IgA, IgM, IgD и IgD (все от Calbiochem). К этим серийным разведениям прибавляли равный объем биотинилированного человеческого IgE при концентрации, приблизительно равной KD (что приводило к получению серий, начинающихся от соотношения конкурирующий антиген:биотинилированный человеческий IgE приблизительно 1000:1). Эти смеси потом переносили на блокированный IgG и оставляли для уравновешивания на 1 час. Несвязанный антиген удаляли вымыванием с помощью PBS-Твин (0,1% об./об.), в то время как оставшийся биотинилированный человеческий IgE определяли с помощью конъюгата стрептавидин-европий3+ (DELFIA® определение, PerkinElmer). Измеряли флуоресценцию с временным разрешением при 620 нм на устройстве для считывания планшетов EnVision (PerkinElmer). Данные флуоресценции подвергали анализу при использовании либо программного обеспечения Graphpad Prism, либо Microsoft™ Excel.
Идентификация улучшенных клонов при использовании биохимического анализа антитело-лиганд
Продукты, полученные в результате селекции, подвергали скринингу в формате эпитопного конкурентного HTRF® (однородная флуоресценция с разрешением во времени) анализа на ингибирование связывания меченного с помощью криптата европия человеческого IgE (получен из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); p159-167]), (CIS bio International 62EUSPEA) с анти-человеческим IgE (Антитело 1).
Во время направленной оптимизации продукты селекции подвергали скринингу в виде неразведенных и разведенных периплазматических экстрактов, содержащих неочищенный scFv, приготовленный в 50 мМ MOPS буфера рН 7,4,0,5 мМ ЭДТА и 0,5 М сорбита.
4 нМ античеловеческого IgE предварительно смешивали с 20 нМ IgG против человеческого Fc, меченного с помощью XL665 (CIS Bio International 61HFCXLA). Прибавляли 10 мкл образца неочищенного scFv к планшету низкого объема на 384 ячейки (Costar 3676). После этого прибавляли 5 мкл смеси антитела к человеческому IgE и анти-Fc-XL665, и потом прибавляли 5 мкл 1/245 разведения меченного криптатом человеческого IgE (приблизительно 2,3 нМ меченного криптатом человеческого IgE). Контроль неспецифического связывания определяли при использовании 300 нМ человеческого IgE (полученный из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]). Все разведения осуществляли в фосфатно-буферном солевом растворе (PBS), содержащем 0,4 М фторида калия и 0,1% BSA (аналитический буфер).
Аналитические планшеты потом центрифугировали при 1000 об/мин при комнатной температуре в течение 1 минуты и инкубировали в течение 3 часов при комнатной температуре, перед считыванием флуоресценции с временным разрешением при длинах волн 615 нм и 665 нм последовательно при использовании устройства для считывания планшетов EnVision (Perkin Elmer).
Данные подвергали анализу путем подсчета значений % Дельта F для каждого образца. Дельта F% определяли в соответствии с уравнением 1.
Уравнение 1:
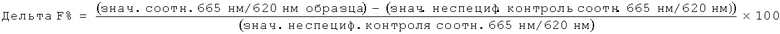
Дельта F% значения последовательно использовали для подсчета % специфического связывания, как описано в уравнении 2
Уравнение 2:
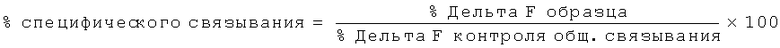
Ингибирование связывания IgEc FcεRIc помощью улучшенного scFv (очищенного)
Очищенный scFv исследовали в формате HTRF® анализа связывания рецептор-лиганд (гомогенная флуоресценция с разрешением во времени) для ингибирования связывания либо человеческого IgE (получен из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]), либо IgE циномолгуса (рекомбинантный, смотри раздел "Материалы и способы"), меченных с помощью криптата европия (CIS bio International 62EUSPEA), с человеческим FcER1-Fc (получен на фирме из NSO клеток).
Титрования scFv концентраций использовали для того, чтобы установить эффективность scFv, как измеряется с помощью значений IC50 в анализе. 1,9 нМ человеческого FcεR1-Fc (на основе молекулярного веса 260 кДа) предварительно смешивали с 20 нМ IgG против человеческого Fc, меченного XL665 (CIS Bio International 61HFCXLA). 10 мкл для титрования очищенного образца scFv прибавляли к аналитическому планшету низкого объема на 384 ячейки (Costar 3676). После этого осуществляли прибавление 5 мкл смеси FcεR1-Fc анти-Fc-XL665, а потом 5 мкл разведения 1/197 меченного криптатом IgE человека или циномолгуса). Контроль неспецифического связывания определяли при использовании 300 нМ IgE человека или циномолгуса (получены на фирме). Все разведения готовили в забуференном фосфатом солевом растворе (PBS), содержащем 0,4 М фторида калия и 0,1% BSA (аналитический буфер).
Аналитические планшеты потом центрифугировали при 1000 об/мин при комнатной температуре в течение 1 минуты и инкубировали в течение 3 часов при комнатной температуре перед считыванием флуоресценции с временным разрешением при длинах волн излучения 615 нм и 665 нм последовательно при использовании устройства для считывания планшетов EnVision (Perkin Elmer).
Данные подвергали анализу путем подсчета значений % Дельта F для каждого образца. Дельта F% определяли в соответствии с уравнением 1.
Уравнение 1:
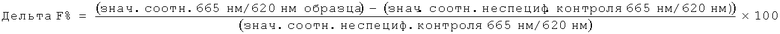
Дельта F% значения последовательно использовали для подсчета % специфического связывания, как описано в уравнении 2
Уравнение 2:
Определяли IC50 значения при использовании программного обеспечения GraphPad Pnsm путем подгонки кривой при использовании логистического уравнения с четырьмя параметрами (Уравнение 3).
Уравнение 2:
Y=Дно+(Верх-Дно)/(1+10^((LogEC50-Х)*уклон))
Х представляет собой логарифм концентрации. Y представляет собой специфическое связывание.
Y начинается со Дна и поднимается до Верха в виде сигмоидальной функции.
Идентификация улучшенных клонов в RBL-ER51 анализе передачи кальциевого сигнала
Эффективность нейтрализации очищенных препаратов IgG из улучшенных антител оценивали в модифицированной версии анализа передачи кальциевого сигнала при использовании RBL-ER51, так как описано в прописи для изоляции.
RBL-ER51 клетки высевали при концентрации 5×104/100 мкл/ячейка в культуральную среду [DMEM (Invitrogen 41966) с 9% об./об. неинактивированного с помощью прогревания FBS (Invitrogen 10100-147) и 400 мкг/мл G418 (Invitrogen 10131-027)] в черные планшеты на 96 ячеек с плоским дном, содержащие культуру ткани (Costar), и инкубировали при 37°C, 5% СО2 в течение 18-24 часов. После этого изымали среду и заменяли ее разведениями исследуемых антител (от 6,67 нМ до 1,33 пМ) в среде для анализа [DMEM (Invitrogen 41966) с 9% об./об. неинактивированного с помощью прогревания FBS (Invitrogen 10100-147) и 400 мкг/мл G418 (Invitrogen 10131-027) и 1,6% смеси пенициллин / стрептомицин (Invitrogen 15140-122)], после чего прибавляли IgE [человеческий получен из (U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]) или циномолгуса (рекомбинантный, смотри раздел "Материалы и способы")], разведенный в аналитической среде с получением заключительной концентрации IgE 25 нг/мл и 100 нг/мл, соответственно. Планшеты инкубировали при 37°C, 5% СО2 в течение 4 часов.
После этого изымали смесь антитело/IgE, оставляя интактный монослой клеток, и заменяли на 100 мкл/ячейка FLUO-4AM загружающего буфера [DMEM с 0,1% FBS, 20 мМ HEPES, 2,5 мМ пробеницида и 2 мкг/мл FLUO-4AM (Invitrogen)] в течение 1-2 часов при 37°C, 5% СО2. Удаляли загружающий буфер, и клетки трижды промывали с помощью 200 мкл/ячейка PBS. Удаляли заключительный промывочный раствор и заменяли его 100 мкл/ячейка FLIPR буфера [125 мМ NaCl, 5 нМ KCl, 1 мМ MgCl2, 1,5 мМ CaCl, 30 мМ Hepes, 2,5 мМ пробеницида 5 мМ глюкозы, 0,01% об./об. FCS]. Планшеты инкубировали при 37°C, 5% СО2 в течение 5-45 минут.
Для измерения мобилизации кальция после прибавления перекрестно связанного анти-IgE FLIPR (Molecular Devices) откалибровывали для приемлемого воздействия в соответствии с инструкциями производителя. Анти-IgE (Biosource AHI0501), разведенный в FLIPR буфере, прибавляли к аналитическим планшетам до получения заключительной концентрации 2,3 мкг/мл. Регистрировали флуоресценцию FLUO-4AM красителя при 1-секундных интервалах на протяжении 80 измерений, после чего использовали 3-секундные интервалы на протяжении 40 измерений. Выводили пик ответа для каждой ячейки, и данные потом подвергали анализу при использовании программного обеспечения Graphpad Prism.
Измерение перекрестного связывания Ig, связанного с FceRI, с помощью улучшенных антител
Для измерения способности очищенных IgG перекрестно связывать IgE, связанный с FcεRI, готовили RBL-ER51 клетки так, как описано в анализе ингибирования для оценки улучшенных антител. RBL-ER51 клетки высевали при концентрации 5х104/100 мкл/ячейка в культуральную среду [DMEM (Invitrogen 41966) с 9% об./об. неинактивированного с помощью прогревания FBS (Invitrogen 10100-147) и 400 мкг/мл G418 (Invitrogen 10131-027)] в черные планшеты на 96 ячеек с плоским дном, содержащие культуру ткани (Costar) и инкубировали при 37°C, 5% CO2 в течение 18-24 часов. После этого удаляли среду и заменяли ее 100 мкл/ячейка человеческого IgE, разведенного (получен из U266 [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]), разведенный до концентрации 1 мкг/мл в среде для анализа [DMEM (Invitrogen 41966) с 9% об./об. неинактивированного с помощью прогревания FBS (Invitrogen 10100-147) и 400 мкг/мл G418 (Invitrogen 10131-027) и 1,6% пенициллин / стерптомицин (Invitrogen 15140-122). Концентрация IgE была выбрана для обеспечения максимальной загрузки RBL-ER51 клеток. Планшеты инкубировали в течение 4 часов при 37°C, 5% СО2.
После этого удаляли раствор IgE, оставляя интактный монослой клеток, и заменяли его на 100 мкл/ячейка FLUO-4AM загружающего буфера [DMEM с 0,1% FBS, 20 мМ HEPES, 2,5 мМ пробеницида и 2 мкг/мл FLUO-4AM (Invitrogen)] в течение 1-2 часов при 37°C, 5% СО2. Удаляли загружающий буфер, и клетки трижды промывали с помощью 200 мкл/ячейка PBS. Удаляли заключительный промывочный раствор и заменяли его 100 мкл/ячейка FLIPR буфера [125 мМ NaCl2, 5 нМ KCl, 1 мМ MgCl2, 1,5 мМ CaCl2, 30 мМ Hepes, 2,5 мМ пробеницида 5 мМ глюкозы, 0,01% об./об. FCS]. Планшеты инкубировали при 37°C, 5% CO2 в течение 5-45 минут.
Для измерения мобилизации кальция после прибавления перекрестно связанного анти-IgE, FLIPR (от 1,53 мкМ до 2,33 нМ) FLIPR (Molecular Devices) откалибровывали для приемлемого воздействия в соответствии с инструкциями производителя. 30 мкл исследуемых антител, разведенных до приемлемых концентраций в FLIPR буфере, прибавляли к аналитическим планшетам. Анти-IgE (Biosource AHI0501) использовали в качестве позитивного контроля. Регистрировали флуоресценцию FLUO-4AM красителя при 1-секундных интервалах на протяжении 80 измерений, после чего использовали 3-секундные интервалы на протяжении 40 измерений. Выводили пик ответа для каждой ячейки, и данные потом подвергали анализу при использовании программного обеспечения Graphpad Prism.
Ингибирование связывания IgE с CD23 на IM9 клетках с помощью очищенного IgG
Антитела оценивали на ингибирование IgE / CD23 взаимодействия при использовании анализа связывания на IM9 клетках. IM9 клетки (линия В-клеток человека) поддерживали в культуральной среде [RPMI 1640 глутамакс (Invitrogen 61870-010); 9% об./об. инактивированного прогреванием FBS (Invitrogen 10100-147)] при использовании стандартных процедур культуры тканей
Для исследования оптимизированного IgG IM9 клетки предварительно обрабатывали с помощью 25 нг/мл человеческого IL-4 (Peprotech, 200-04) в течение 3 дней при 37°C/5%CO2 для того, чтобы повысить экспрессию CD23.
Собирали IM9 клетки и ресуспендировали в проточном буфере [PBS с 1% козьей]. Блокирование Fc рецептора осуществляли путем прибавления Fc фрагментов (TEBU-bio) до получения заключительной концентрации 5 мкг/мл. Эту клеточную суспензию высаживали в количестве 100 мкл/ячейка в полипропиленовый планшет, содержащий ячейки с U-образным дном (Greiner) и инкубировали на льду в течение 30 минут.
Разведения антител (от 667 нМ до 1 нМ) готовили в полипропиленовых планшетах, содержащих ячейки с U-образным дном (Greiner) и смешивали с IgE (полученный из U266 клеток [Ikeyama и др. 1986. Molecular Immunology 23 (2); стр. 159-167]) до получения заключительной концентрации IgE 10 мкг/мл в течение 30 минут при комнатной температуре. Планшеты с клетками центрифугировали при 2000 об/мин в течение 2 минут, удаляли супернатанты, оставляя клеточный осадок. Клетки ресуспендировали в смеси, содержащей 100 мкл/ячейка антитела и IgE, и инкубировали на льду в течение 1 часа. Осадок клеток центрифугировали при 2000 об/мин в течение 2 минут и удаляли супернатанты антитело/IgE. Клетки промывали путем ресуспендирования в 200 мкл/ячейка проточного буфера и центрифугировали так, как описано выше.
IgE, связанный с клеточной поверхностью, определяли с помощью комплекса анти-IgE-фикоэритрин (Caltag), разведенного до 1/30, об./об., 100 мкл/ячейка. Аналитические планшеты инкубировали на льду в течение 20 минут в темноте перед центрифугированием при 2000 об/мин в течение 2 минут и промывали с помощью 2×200 мкл проточного буфера, как описано выше. Клетки ресуспендировали в 100 мкл Cell Fix (BD biosciences) и подвергали анализу при использовании FACSCalibur (BD Biosciences) для определения FL2 окрашивания.
Данные анализировали при использовании программного обеспечения CellQuest Software (BD biosciences). Выводили среднее геометрическое значение флуоресценции FL2, и данные потом анализировали при использовании Microsoft Excel и Graphpad Prism программного обеспечения.
Получение человеческого IgE Сε3-4-С, терминально меченного с помощью FLAG His 10. Фрагмент человеческого IgE Сε3-4 был таким, как описано ранее у Wurzburg и др. (2000). Структура человеческого IgE-Fc Cε3-C4 выявила конформационную гибкость в эффекторынх доменах антитела. Фрагмент кДНК, который охватывает нуклеотиды 2135-2868 (GenBank депозитный номер J00222), амплифицировали при использовании RT-ПЦР из общей РНК стимулированных с помощью IL13 человеческих РВМС (мононуклеарных клеток периферической крови). Этот продукт ПЦР клонировали в pCR2.1 ТА (Invitrogen).
Для осуществления секреции экспрессированного белка и получения последовательности, которая включает находящийся в рамке С-терминальный FLAG эпитоп и полигистидиновую метку (His10), IgE Сε3-4 фрагмент амплифицировали с помощью ПЦР при использовании праймеров, которые включают 5' BssHII сайт, 3' FLAG эпитоп, полигистидиновую метку (His10) и XbaI сайт, и последовательно встраивали в pEU8.2 вектор. Модифицированный pEU8.2 вектор содержит EF-1 промотор, геномную последовательность для мышиного лидерного пептида IgG1, oriP источник репликации для того, чтобы позволить осуществить эписомальную репликацию плазмиды при трансфекции в клеточные линии, экспрессирующие EBNA-1 генный продукт (такие, как HEK293-EBNA клетки).
Белок очищали от кондиционной среды при использовании IMAC хроматографии, после чего осуществляли гель-фильтрацию (SEC).
Получение IgE Сε3-4 циномолгуса, С-терминально меченного с помощью FLAG His10
Константный участок IgE циномолгуса определяется с помощью непосредственного секвенирования продуктов ПЦР, амплифицированных из геномной ДНК при использовании праймеров, которые охватывают нуклеотиды 1174-2989 человеческого локуса IgHE (тяжелой цепи IgE) (номер доступа GenBank J00222). Экзоны идентифицировали путем гомологии с последовательностью человека и, таким образом, является возможным предсказать последовательность кДНК для константного участка тяжелой цепи IgE циномолгуса.
Синтезировали кДНК, кодирующую последовательность мышиного лидерного пептида IgG1 циномолгуса, Сε3-4 (Фигура 2), С-терминальный FLAG эпитоп и полигистидиновую метку (ДНК2.0), и клонировали в pDONR221 (Invitrogen). Потом при использовании LR Gateway® реакции (Invitrogen) ген, который представляет интерес, переносили в экспрессионный вектор pDEST12.2 (Invitrogen), модифицированный путем инсерции oriP источника репликации из рСЕР4 вектора (Invitrogen) для того, чтобы позволить осуществить эписомальную плазмидную репликацию при трансфекции в линии клеток, экспрессирующие продукт гена EBNA-1 (такие, как HEK293-EBNA клетки).
Белок очищали от кондиционной среды при использовании IMAC хроматографии, после чего осуществляли гель-фильтрацию (SEC).
Получение химерного D12 вариабельного участка и константного участка IgE циномолгуса
Синтезировали кДНК, кодирующую вариабельный участок тяжелой цепи scFv анти-эстрадиола человека (D12_VH) и один из двух различных гаплотипов гена IgHE циномолгуса (cyIGHE TQ и cyIGHE ME) (ДНК2.0) и клонировали ее в вектор pDONR221 (Invitrogen). Также синтезировали кДНК, представляющую вариабельный участок легкой цепи scFv анти-эстрадиола человека (D12_VL) и один из двух генов лямбда константного участка циномолгуса (cyIGLC4 и cyIGLC7) (ДНК2.0) и клонировали ее в вектор pDONR221 (Invitrogen).
Потом при использовании LR Gateway® реакции (Invitrogen) ген, который представляет интерес, переносили в экспрессионный вектор pDEST12.2 (Invitrogen), модифицированный путем инсерции oriP источника репликации из рСЕР4 вектора (Invitrogen) для того, чтобы позволить осуществить эписомальную плазмидную репликацию при трансфекции в линии клеток, экспрессирующие продукт гена EBNA-1 (такие как HEK293-EBNA клетки).
Рекомбинантный химерный IgE белок, представляющий вариабельный участок тяжелой цепи scFv человеческого анти-эстрадиола, слитого с Сε1-4 IgE циномолгуса (Фигуры 3 и 4), и вариабельный участок легкой цепи scFv человеческого анти-эстрадиола, слитого с лямбда константным участком циномолгуса (Фигуры 5 и 6), экспрессированным из HEK293 EBNA клеток, очищали при использовании способа, как описано Ikeyam и др. (1986) Mol Immunol 23 стр. 159-67.
Человеческий FcεRI-Fc (полученный из клеток NS0 на фирме) FcεRI, охватывающий нуклеотиды 67-711 (номер доступа GenBank NM_002001), клонировали выше от геномного участка Fc человеческого IgGI, так же, как из pEU1.2, впервые описанного Persic и др. (1997) Gene 187; 9-18. Продукт клонировали в рсДНК3.1 EcoRI - XbaI (SEQ ID NO:391 и 392). Экспрессию рекомбинантного слитого белка FcεRI_Fc осуществляли путем стабильной трансфекции NS0 клеток с помощью рсДНК3.1 FcεRI_Fc конструкции. Стабильную экспрессию устанавливали путем селекции при использовании G418, изоляции клонов с помощью серийного разведения и идентификации клонов с высоким уровнем экспрессии. FcεRI_Fc слитый белок потом очищали от кондиционный среды при использовании аффинной хроматографии с белком А, после чего опроводили гель-фильтрацию.
Пример 3. В-клетки человека - ингибирование внутриклеточного IgE
Мононуклеарные клетки периферической крови (РВМС) изолировали из гепаринизированной цельной крови человека путем центрифугирования с использованием фиколл-пак градиента (Phannacia). В-клетки последовательно изолировали из популяции РВМС при использовании позитивной анти-CD19 селекции с использованием магнитных частиц (Miltenyi). Как позитивные, так и негативные фракции В-клеток, собирали как клетки, которые прошли через магнитную колонку. Клетки из негативной фракции В-клеток, содержащие все РВМС клетки, за исключением В-клеток, обрабатывали митомицином С для предотвращения пролиферации. Клетки инкубировали с 50 мкг/мл митомицина С в течение 30 минут, потом промывали с помощью культуральной среды (RPMI 1640 с глутамаксом (Gibco) /10% FCS (Gibco) /50 ед./мл пенициллина /50 мкг/мл стрептомицина (Gibco)) и дополнительно инкубировали в PBS в течение 70 минут перед заключительным промыванием для удаления всего митомицина С. Для индукции дифференциации В-клеток, 4×104 В-клеток и 9,2×105 клеток из негативной фракции В-клеток высаживали на планшет из 96 ячеек в среду для культуры клеток, дополненную 3,5 мкМ бета-меркаптоэтанола (Sigma) и 20 мкг/мл трансферрина (Chemicon), и предварительно инкубировали с анти-IgE моноклональными антителами или контрольным антителом при концентрации 0,001-100 нМ в течение 30 минут перед прибавлением человеческого интерлейкина-4 (IL-4) в концентрации 10 нг/мл. Клетки последовательно инкубировали в увлажненной камере с CO2 в течение 12-14 дней. Через 12-14 дней планшеты подвергали короткому циклу центрифугирования, собирали супернатанты, и клетки окрашивали для обнаружения внутриклеточного IgE при использовании следующей прописи.
Сначала клетки инкубировали с PBS и 1% человеческой сывороткой в течение 10 минут для блокирования связывания Fc рецептора. Потом клетки фиксировали и подвергали нарушению проницаемости мембраны на льду при использовании набора Cytofix/Cytoperm от Becton Dickinson. Потом клетки промывали перед добавлением поликлонального античеловеческого IgE-FITC антитела кроля (DAKO) при конечном разведении 1:6, и моноклонального мышиного анти-человеческого CD19 - RPE/Cy5 антитела (DAKO) при конечном разведении 1:10. Является важным, чтобы клетки были тщательно промытыми для того, чтобы избежать воздействия остаточных анти-IgE MAb (моноклональных антител). Через 30 минут инкубации клетки промывали и образцы анализировали на FACS Calibur при использовании HTS устройства для загрузки планшетов на 96 ячеек. Потом регистрировали процентное содержание клеток в CD19+ популяции, которая совместно экспрессирует IgE, и экспрессию внутриклеточного IgE представляли в виде % ингибирования максимальной IgE экспрессии в клетках, не обработанных с помощью блокирующего анти-IgE моноклонального антитела. Антитело 11 ингибировало индукцию IgE позитивных клеток со значением IC50 1,6 нМ (Фигура 7 - верхняя диаграмма). Нерелевантное антитело того же формата использовали в качестве негативного контроля (CAT-002), оно не ингибировало индукции IgE позитивных В-клеток(Фигура 7 - нижняя диаграмма).
Пример 4. Линия тучных клеток (LAD2) - ингибирование высвобождения β-гексозаминидазы
LAD2 клетки [A.S.Kirshenbaum и др. Leukemia research 27 (2003)] культивировали при плотности клеток 0,25-0,6×106 клеток/мл в среде, свободной от сыворотки (StemPro-34, Life Technologies), дополненной питательной добавкой StemPro-34,2 мМ L-глутамина и 100 нг/мл рекомбинантного фактора стволовых клеток человека (rhSCF, R&D). Для анализа определения β-гексозаминидазы клетки высевали при плотности 2,5×104 клеток/ячейка и предварительно инкубировали в полипропиленовом планшете на 96 ячеек вместе с блокирующими анти-IgE МАb в интервале концентраций 0,0001-100 нМ. Клетки инкубировали при 37°C в течение 30 минут перед прибавлением IgE при концентрации 0,15 нМ, после чего клетки инкубировали в течение дополнительных 4 часов. После инкубирования с IgE клетки промывали с помощью буфера для удаления остатка IgE, и после этого IgE, связанный с FceR на LAD2 клетках, перекрестно связывали с αIgE (600 мкг/мл козьего античеловеческого IgE, Sigma) в течение 30 минут при 37°C. Инкубацию останавливали с помощью холодного центрифугирования, собирали клеточные супернатанты и переносили в планшет на 96 ячеек. Содержание β-гексозаминидазы подвергали анализу при использовании незначительно модифицированной версии способа, опубликованного Smith и др. [Smith J и др. Biochem. J. (1997)223, 321-328]. Вкратце, 2 мМ р-нитрофенил-N-ацетил-D-глюкозаминида в 0,2 М цитратном буфере, рН 4,5 использовали в качестве субстрата для гексозаминидазы. Реакцию останавливали путем прибавления 1М Трио-буфера рН 9,0. Оптическую плотность измеряли спектрофотометрически при длине волны 405 нм (минус значение при длине волны 570 нм) при использовании считывающего устройства Spectramax от Molecular Devices. Подсчитывали эффект анти-IgE MAb в отношении ингибирования β-гексозаминидазы и представляли его в виде процента ингибирования общего высвобождения +/- стандартная ошибка среднего. Антитело 11 ингибировало β-гексозаминидазу со значением IC50 0,04 нМ (Фигура 8 - верхняя диаграмма), в то время как нерелевантное MAb того же формата (CAT-002) не ингибировало β-гексозаминидазу (Фигура 8 - нижняя диаграмма).
Пример 5. Связывание анти-IgE антитела с IgE в сыворотке крови при использовании ELISA
Описание анализа
Образцы сывороток готовили из образцов крови, полученной от человеческих доноров. Планшеты ELISA на 96 ячеек (Nunc Maxisorp, No. 442404) покрывали 150 мкл/ячейка 1 мкг/мл FcERI-Fc-His, разведенным в PBS, и инкубировали при 4°C в течение ночи. После инкубации в течение ночи планшеты трижды промывали с помощью PBS, содержащего 0,05% Твин 20 (PBST, Medicago 09-9401-100). Для снижения фонового связывания планшеты последовательно инкубировали с 200 мкл/ячейка блокирующего буфера, состоящего из PBS, содержащего 0,5% BSA, инкубировали при комнатной температуре в течение 2 часов и трижды промывали с помощью PBST, как описано выше. Образцы (человеческая сыворотка или плазма с варьирующими концентрациями анти-IgE антитела. Антитело 11) и стандарты (ImmunoCAP общий IgE (человеческий) калибровочный стандарт, Phadia, Uppsala) разводили в PBS, содержащем 0,05% Твин 20, и выдерживали на льду до тех пор, пока их не использовали для аналитических планшетов в объеме 150 мкл/ячейка. Планшеты закрывали, и образцы инкубировали при комнатной температуре в течение 2 часов. Для удаления несвязанного образца планшеты трижды промывали с помощью PBST, как описано ранее. После этого прибавляли 150 мкл/ячейка анти-человеческого IgE кролика (D€1, 30-1917-00, 420036-02, 841204, 9911302 от Phadia, Uppsala) при концентрации 0,25 мкг/мл, разведенного в PBST, для определения связанного человеческого IgE. Потом планшеты снова закрывали и инкубировали в течением 1 часа при комнатной температуре. Для удаления несвязанных античеловеческих IgE антител кролика, планшеты трижды промывали с помощью PBST так, как описано выше. Конъюгированное с HRP-вторичное антитело (античеловеческий IgE козы). Конъюгат разводили 1:25000 в PBST, прибавляли 150 мкл на ячейку, закрывали планшеты и инкубировали в течение 1 часа при комнатной температуре. Потом планшеты трижды промывали с помощью PBST, как описано выше. Потом прибавляли раствор ТМВ-субстрата (DAKO субстрат-хромоген. No. S1599) к каждой ячейке в количестве 150 мкл/ячейка и планшеты инкубировали в течение 10 минут при комнатной температуре. Реакцию останавливали путем прибавления 150 мкл/ячейка раствора для остановки реакции (2М H2SO4) и считывали поглощение при 450 нм на устройстве Tecan SAFIRE. Поскольку измерение IC50 является зависимым от концентрации лиганда (то есть IgE) в анализе, в настоящем анализе IC50 будет зависеть от количества IgE лиганда, присутствующего в образце сыворотки человека. В репрезентативном эксперименте Антитело 11 имело значение IC50 202 пМ (Фигура 9). В том же эксперименте Xolair™ имел значение IC50 57 нМ.
Пример 6. Измерение образования комплекса между IgE и очищенным IgG
Характеристику иммунных комплексов, которые образовались между очищенным , человеческим IgE и очищенным анти-IgE IgG (Антитело 11), осуществляли с помощью высокоэффективной вытеснительной жидкостной хроматографии. В дополнение, непрерывное рассеивание света при различных углах (MALS) использовали для оценки размера комплекса. Комплексы образовывались путем инкубации IgE и IgG вместе при трех различных молярных соотношениях (3:1, 1:1 и 1:3, соответственно) в PBS Дюльбекко при 18°C в течение 1 часа. Для молярного соотношения 1:1 концентрация каждого белка составляла 2,5 мкМ. Более высокие соотношения достигались путем повышения концентрации релевантного белка до 7,5 мкМ. Эти образцы подвергали анализу на двух колонках Bio-Sep-SEC-S 4000 (300×7,8 мм), соединенных вместе. Уравновешивали колонки и образцы анализировали в PBS Дюльбекко при скорости потока 1 мл/мин, на системе Agilent HP1100 HPLC. Пики определяли при 220 и 280 нм при использовании детектора на диодной матрице, и элюат также пропускали через детекторы Wyatt Technologies DAWN EOS (MALS) и Optilab rEX (показатель рефракции).
Хроматография образца с молярным соотношением 1:1 давала дублет пиков (определяли с помощью поглощения УФ), которые полностью не разделялись и соответствовали времени удержания 13,88 минут и 14,9 минуты. Эти значения удержания показывают образование нековалентных комплексов IgE с IgG. MALS давал молекулярные массы 1,085 кДа (пик 13,88 мин) и 702 кДа (пик 14,9 мин). Эти массы являются согласованными с комплексами, соответствующими гетеротетрамеру (предсказанная масса 674 кДа, 2IgE:2IgG) и гетерогексамеру (предсказанная масса 1010 кДа, 3IgE:3IgG). Хроматографический анализ и MALS анализ для образцов с молярным соотношением как 3:1, так и 1:3 (IgE:IgG) обеспечивал подобный профиль с образцом 1:1 с пиками, соответствующими гетеротетрамеру и гетерогексамеру, определяемым с помощью поглощения УФ. Дополнительные пики определяли как соответствующие избытку IgE или IgG в образцах.
Пример 7. Определение связывания эпитопов с помощью герминизированного Антитела 11
Применение рентгеновской кристаллографии для определения точной трехмерной структуры белков при атомном разрешении является хорошо известным специалистам в данной области техники. Этот способ был использован для подробной визуализации частей белков, которые взаимодействуют с антителами (Padavattan и др., 2007; Karpusas и др., 2001). Таковая является наиболее точной методикой картирования эпитопов, однако требует значительных усилий и базируется на возможности получения кристаллов удовлетворительного качества, что, в свою очередь, зависит от чистоты и качества образца белка и компетентности в том, чтобы отыскать приемлемые условия кристаллизации. После получения кристаллов белок-антитело их облучали с помощью рентгеновских лучей с получением дифракционной модели, которая зависит от точного распределения атомов. Дифракционная модель может быть проанализирована с помощью кристаллографов для определения трехмерных позиционных координат атомов в структуре. Это позволяет тщательно исследовать сайты взаимодействия между белком и антителом.
7.1 Определение рентгеновской кристаллической структуры комплекса IgE Сε3-Сε4 антитело
IgE домен Сε2-Сε3 клонировали, экспрессировали и очищали с целью структурного определения. Подобным образом полученный Fab фрагмент получали путем переваривания и очистки цельного Антитела 11, усовершенствованного для связывания с IgE. Комплекс получали путем смешивания и очищали с помощью эксклюзионной хроматографии для удаления IgE доменов и Fab молекул, которые не образовывали комплексы. Получаемые кристаллы комплекса IgE Сε3-Сε4/Fab относились к трехгранной пространственной группе Р3221. Их подвергали анализу в европейской лаборатории синхротронного излучения (ESRF) в Гренобле, Франция. Были получены полные данные дифракции до разрешения 2,85 Å. Структура может быть разрешена с помощью молекулярного замещения (Rossman, 1972) при использовании вариабельной и константной части Fab фрагмента в качестве отдельных исследовательских моделей, посредством чего ориентировали и позиционировали Fab фрагменты в кристаллографических ассиметричных единицах. В общей сложности были идентифицированы три Fab фрагмента в ассиметричной единице. После этого три IgE Сε3-Сε4 молекулы могут быть размещены в ассиметричной единице. Каждый IgE димер связывается с двумя Fab молекулами и, таким образом, в общей сложности ассиметричная единица включает один и половину полного комплекса IgE/Fab.
7.1.1 Общее описание комплекса IgE Cε3-Cε4/Fab Антитела 11
Кристаллическая структура показывает, что каждый IgE Cε3-Cε4 димер является связанным симметричным или ассиметричным образом с двумя Fab фрагментами (Фигура 10). Поскольку ассиметричная единица кристалла включает один с половиной IgE/Fab комплекса, неполный комплекс образует димер с симметрично связанным партнером в прилегающей ассиметричной единице с помощью двойной оси.
Обе молекулы IgE димера, обозначаемые как IgE1 и IgE2, взаимодействуют с Fab фрагментом Антитела 11. Большая часть взаимодействий обеспечивается с помощью Fab тяжелой цепи, который взаимодействует как с IgE1, так и с IgE2, в то время как легкая цепь, как наблюдается, взаимодействует только с IgE1. Эпитоп антигена размещается, в основном, в домене Сε3, со вкладом одной аминокислоты, которая размещается близко к шарнирному участку домена Сε4.
Три сайта взаимодействия между IgE Cε3-Cε4 и Fab в ассиметричной единице кристалла являются весьма подобными. Однако после уточнения стало понятно, что одна из Fab молекул является значительно менее упорядоченной, чем две другие молекулы Fab. Это обычно наблюдается в кристаллических структурах и объясняется тем фактом, что определенный участок является гибким и приобретает различные ориентации в кристалле так, что электронная плотность является менее определенной. Такая Fab молекула и взаимодействие, осуществляемое ею с IgE Cε3-Cε4 молекулой, таким образом, не учитываются в этом анализе.
IgE, как известно, является гликозилированным на участке Fc при остатке Asn394 (Wurzburg и др.).
Характеристика гликозилирования Fc, осуществляемая с помощью масс-спектрометрического анализа после переваривания трипсином, показала три различных варианта гликана, связанные с Asn394, состоящего из коровой структуры Man3GlcNAc2 с удлинением 2,3 или 4 гексоз, предположительно всех манноз (Фигура 11). Фактически, от остатка Asn394 во всех трех IgE Сε3 доменах расширенная электронная плотность распространяется в полость между двумя IgE молекулами в димере. Электронная плотность предполагает высокоманнозный тип структуры с двумя N-ацетилглюкозаминами (GlcNAc) и тремя или четырьмя маннозными единицами, которые наблюдаются в каждой цепи, что согласуется с данными масс-спектрометрического анализа. Только одна из гексоз вне коровой структуры, Man6, которая является слитой с Man4, является видимой в электронной плотности, что свидетельствует о том, что остальные 1-3 гексозы являются гибкими.
7.1.2 Описание эпитопа и паратопа
Такая кристаллическая структура позволяет исследовать эпитопное взаимодействие между IgE Сε3-Сε4 и Fab с атомными деталями. Существуют два независимых взаимодействия IgE/Fab в разрешенной структуре, за исключением третьей Fab молекулы по причине ее слабо определенной карты электронной плотности, которая будет описана далее. Они являются весьма подобными, о чем свидетельствует среднее квадратическое отклонение между двумя эквивалентными Fab фрагментами вариабельной цепи 031 Å, подсчитанное при использовании Сε положений (наложение, ССР4 1994) и между эквивалентными IgE мономерами 0,82 и 0,96 Å, соответственно. Несмотря на это высокая подобность двух взаимодействий будет описана отдельно и будет обозначаться как IgE/Fab1 и IgE/Fab2. Детали взаимодействий приведены в Таблице 5 и Таблице 6, где номер остатка содержит указатель цепи (НС: Fab тяжелой цепи, LC: Fab легкой цепи, IgE1, IgE2). Нумерация аминокислотных остатков Антитела 11 является такой в соответствии с системой Кэбата (Kabat и др. 1991). Расстояния получали при использовании ССР4 программы CONTACT (ССР4, 1994).
Оба взаимодействия вовлекают участки, определяющие комплементарность (CDR) как из тяжелой, так и из легкой цепи фрагмента антитела, остатки каркасного участка (участок за пределами CDR Fab) и аминокислотные остатки из двух мономеров в IgE Сε3-Сε4 мономере. Легкая цепь антитела взаимодействует с IgE1 в IgE Сε3-Сε4 димере, в то время как тяжелая цепь взаимодействует с обоими мономерами. Однако большая часть контактов являются такими между тяжелой цепью и мономером IgE2 антигена. Эти два взаимодействия описаны подробно ниже.
7.1.3 Подробное описание взаимодействия между Fab1 и IgE, взаимодействие 1
Взаимодействие сайта, определяющего эпитоп IgE Сε3-Сε4, охватывает площадь 1100 Å2 (подсчитана при использовании программы Areaimol, смотри ссылку ССР4, 1994) и осуществляется с помощью аминокислотных остатков от Glu390 до Asn394, включительно для IgE1 и Leu340, Arg342, от А1а428 до Thr434, включительно Thr436, Ser437 и Glu472 в IgE2 антигена. Кроме того, остатки сахара GlcNAc1 и Man6 в IgE1 и Man5 в IgE находятся в контакте с Fab молекулами. Аминокислотные остатки из тяжелой цепи, которые взаимодействуют с антигеном, включая остатки сахара, являются такими из CDR1: Tyr32, из CDR2: Asp53 и Asn54, из CDR3: Val95, Met96, Ile100, Gly100b, Gly100c, Asp101 и Tyr102 и из каркасного участка: Glu1, Lys23, Thr30, от Ala71 до Arg77 включительно, и Tyr79. Остатки, осуществляющие свой вклад из легкой цепи Fab, представляют собой Asp50 и Ser56 из CDR2 и Leu 46 и Tyr49 из каркасного участка. Взаимодействие включает 19 водородных связей в дополнение к неполярным контактам Ван-дер-Ваальса.
7.1.4 Подробное описание взаимодействия между Fab1 и IgE, взаимодействие 2
Взаимодействие сайта, определяющего эпитоп IgE Сε3-Сε4, охватывает площадь 1165 Å2 (подсчитана при использовании программы Areaimol, смотри ссылку ССР4, 1994) и осуществляется с помощью аминокислотных остатков Glu390, от Gln392 до Asn394, включительно IgE1 и Leu340, Arg342, от Ala428 до Thr434, включительно Thr436, Ser437 и Glu472 в IgE2 антигена. Кроме того, остатки сахара GlcNAc1 и Man6 в IgE1 находятся в контакте с тяжелой цепью Fab. Аминокислотные остатки из тяжелой цепи, которые взаимодействуют с антигеном, включая остатки сахара, являются такими из CDR1: Tyr32, из CDR2: Pro52a, Asp53 и Asn54, из CDR3: Val95, Met96, Ile100, Gly100b, Gly100c, Asp101 и Tyr102, и из каркасного участка: Glu1, Lys19, Lys23, Thr30, от Ala71 до Ser75 включительно, Arg77 и Tyr79, и из каркасного участка: Glu1, Lys23, Thr30, от Ala71 до Arg77 включительно и Tyr79. Остатки, осуществляющие свой вклад из легкой цепи Fab, представляют собой Ser56 из CDR2 и Tyr49 из каркасного участка. Взаимодействие включает 19 водородных связей в дополнение к неполярным контактам Ван-дер-Ваальса.
Материалы и способы для эксперимента 7
Сверхэкспрессия IgE Сε3-Сε4.
Линии клеток и культуральная среда.
В этой работе использовалась исходная адгезивная линия клеток HEK293-EBNA (Invitrogen, Stockholm, Sweden), стабильно экспрессирующая ядерный антиген-1 вируса Эпштейна-Барра. Клетки адаптировали к росту в суспензии перед перенесением в среду DHI с помощью поэтапной замены среды (Davies и др. 2005). Используемая DHI среда немного отличалась от исходного описания тем, что была свободна от СА2+. После адаптации создавали рабочий банк клеток и обе линии клеток выращивали обычным образом в среде DHI, свободной от СА2+, с добавкой 4 мМ глутамина, 2% об./об., IgG фетальной сыворотки теленка со сверхнизким молекулярньм весом, 250 мкг/мл G418 (все от Invitrogen, Stockholm, Sweden) и 0,1% вес./об. Плюроник F68 (Sigma-Aldrich, Stockholm, Sweden) максимально до 20 пассажей.
Процедура трансфекции
Готовили в воде маточный раствор 1 мг/мл линейного 25 кДа полиэтиленимина (Polysciences Europe, Eppenheim, Germany), значение рН доводили до 7,0, стерильно фильтровали и хранили в виде небольших аликвот при температуре -80°C до использования. Смесь для трансфекции получали непосредственно перед трансфекцией в среде DHI без добавок в объеме, эквивалентном 1/10 трансфекционного объема. Для получения смеси для трансфекции DHI разделяли на две половины. Прибавляли 0,8 мкг ДНК на мл трансфекционного объема к одной половине DHI среды, а в другую половину прибавляли 2 мкг PEI на мл трансфекционного объема. После короткого встряхивания двух растворов и инкубации их в течение 5 минут медленно прибавляли раствор ДНК к раствору PEI. Трансфекционную смесь инкубировали 20-30 минут при комнатной температуре перед внесением в волновой биореактор (Wave Biotech AG, Tagelswangen, Switzerland). Через 4 часа после трансфекции культуру прибавляли к объему для заключительной продукции, содержащему DHI с добавками и НурРер1510 (Kerry Bio-Sciences, Almere, the Netherlands) до получения заключительной концентрации 0,3% (вес./об.).
Культуры для посева
Для размножения культуры для посева клетки выращивали в пластиковых флаконах для встряхивания при 37°C в атмосфере 5% СО2, помещенных в термостат, оснащенный орбитальным встряхивателем (Infers AG, Bottmingen, Switzerland). Клетки пересаживали два раза в неделю обычным способом при достижении концентрации приблизительно 2×106 клеток/мл перед разделением. Плотность и жизнеспособность клеток оценивали при использовании автоматического устройства для подсчета клеток (Innovatis AG, Bielefeld, Germany). Для волновой культуры клетки разделяли до 1×106 клеток/мл за один день до трансфекции для того, чтобы убедиться, что они находятся в логарифмической фазе роста на начало эксперимента. Волновые культуры инокулировали непосредственно из встряхивателей. Все культуры для посева концентрировали путем центрифугирования и ресуспендировали в свежей культуральной среде перед прибавлением в биореакторы.
Волновые культуры
Экспрессию осуществляли в волновых биореакторах (Wave Biotech AG, Tagelswangen, Switzerland) при рабочем объеме 10 л. Волновые биореакторы засевали до плотности 1×106 клеток/мл в 4,5 л обогащенной DHI среды. Через 2 часа после осуществления фазы адаптации культуры трансфицировали 0,5 л смеси для трансфекции. Через 4 часа после транфекции культуру подпитывали 10 л общего объема обогащенной DHI среды и НуРер1510 до получения заключительной концентрации 0,3% (вес./об.). Ежедневно отбирали образцы для определения плотности клеток, жизнеспособности и концентрации белка.
Экспрессионный вектор
Вектор, экспрессирующий человеческий IgE Cε3-Cε4, с C-терминальной Flag меткой и 10-гистидиновой меткой, получали из вектора, описанного Persic и др. (1997). Система находилась под контролем EF1-a промотора.
Очистка IgE Cε3-Cε4
20 л клеточного супернатанта пять раз концентрировали и подвергали диафильтрации до 2xPBS (308 мМ NaCl, 20 мМ фосфата, рН 7,4) при использовании фильтра с поперечным потоком при отсечении молекулярного веса 10 кДа (Pellicon 2, Millipore). Среду подвергали связыванию в общем объеме с 30 мл NiSepharose (GE Healthcare) в течение 2 часов при температуре 4°C, промывали с помощью пяти объемов 2xPBS и загружали в XK26 колонку. Потом колонку промывали при использовании пяти объемов колонки с помощью 40 мМ имидазола в 2xPBS для вымывания контаминирующих белков. В завершение, IgE элюировали при использовании 400 мМ имидазола в 2xPBS. Объединенные фракции, содержащие IgE с высокой степенью чистоты, концентрировали приблизительно четыре раза (до ~5 мг/мл) перед тем, как пропустить через колонку Superdex 200 50/60 SEC (1200 мл, GE Healthcare) при использовании 2xPBS в качестве подвижного буфера. Отделяли некоторые большие белки, и IgE обнаруживали в основном пике. Собирали только фракции основного пика, так как в других фракциях присутствовала контаминация. Этот этап повышал чистоту образца до ~99%. Общее количество продукта составляло 42 мг, и очищенный IgE имел концентрацию 2 мг/мл.
Анализ гликозилирования IgE Сε3-Сε4
Переваривание в растворе при использовании трипсина
Человеческий минимальный домен IgE, IgE Сε3-Сε4, 2 мг/мл, в 2xPBS (состав: 308 мМ NaCl, 20 мМ фосфата, рН 7,4) смешивали с 100 мкл трипсина 0,02 мг/мл в 25 мМ NH4HCO3. Переваривание осуществляли в течение ночи при 37°C и останавливали его с помощью 2 мкл муравьиной кислоты (67%) в Н2О.
Нано-ЖХ МС/МС:
Анализ осуществляли на колонке 20 см х 50 мкм (внутренний диаметр) кварцевой капиллярной колонки, заполненной ReproSil-Pur C18-AQ пористыми частицами размером 3 мкм, связанной с LTQ-Orbitrap масс-спектрометром (Thermo). Осуществляли инъекцию 8 мкл образца (автоматический самплер Agilent), и пептиды подвергали улавливанию на предварительную колонку 4,5 см х 100 мкм (внутренний диаметр )перед разделением. Через 5 минут линейного прогона при использовании 0,1% муравьиной кислоты, градиент составлял 10-50% ацетонитрила в течение 5-30 минут (Agilent), 200 нл/мин и элюент подвергали электрораспылению с помощью испускающего наконечника. Устройство эксплуатировали в зависимом от данных режиме для переключения между Orbitrap (FT-MS) контрольным сканированием и ионной ловушкой (IT-MS/MS) трех наиболее избыточно протонированных ионов.
Статическое электрораспыление МС/МС:
Для проверки заряженного состояния гликопептидных фрагментов подвергали анализу выбранные предшественники с помощью ESI иглы 1,6 кВ, фрагментировали и определяли в устройстве Orbitrap, противоположно направленном к линейной ионной ловушке, в нано-ЖХ анализе.
Анализ данных в отношении гликопептидов:
Подсчитанные МН+ массы возможных гликопептидов анализировали на присутствие гликозилирования с помощью устройства GlycoMod (http:expasy.org/tools/glycomod) (Cooper и др. 2001). Вводили белковую последовательность и допуск по массе 10 част. на млн. Все предложенные гликопептиды проверяли на присутствие фрагментов, содержащих гликан.
Получение Fab Антитела 11.
Очистка IgG
Антитело 11 очищали от CHO-EBNA переходного материала при использовании MabSelect SuRe (GE Healthcare) хроматографической среды с белком А. Заменяли буфер в элюате белка А на PBS, рН 7,2 при использовании PD-10 колонок (GE Healthcare), а потом фильтровали через фильтр 0,22 мкм при использовании Millex-GP фильтра шприцевого типа (Millipore).
Переваривание и очистка Fab
Готовили буфер для переваривания, содержащий 30 мМ гидрохлорида DL-цистеина, растворенного в GIBCO PBS (Invitrogen). Папаин, полученный из латекса папайи (Sigma), восстанавливали в буфере для переваривания с получением раствора с концентрацией 10 мг/мл, который выдерживали при комнатной температуре в течение минимум 30 минут перед использованием. Прибавляли цистеин к IgG Антитела 11 с получением 30 мМ раствора, после чего прибавляли папаин в соотношении 1 мг папаина на 100 мг IgG. Переваривание останавливали через 90 минут путем прибавления 0,5 М йодацетамида (Sigma) с получением 50 мМ йодацетамида в заключительной смеси для переваривания. Fab очищали от среды для переваривания при использовании MabSelect SuRe (GE Healthcare) хроматографической среды с белком А в несвязывающем режиме. Фракцию Fab, полученную из MabSelect SuRe этапа подвергали смене буфера на 50 мМ ацетат натрия/100 мМ NaCl, рН 5,5 при использовании PD-10 колонок (GE Healthcare) и подвергали концентрированию до ~10 мг/мл при использовании Amicon Ultra-15 5 кДа MWCO устройства для фильтрования на центрифуге (Millipore). Заключительный продукт дополнительно очищали при использовании Mustang Q acrodisc (Pall) и потом фильтровали через фильтр 0,22 мкм при использовании Millex-GP фильтра шприцевого типа (Millipore).
Получение комплекса IgE Cε3-Cε4 Fab Антитела 11
Раствор, содержащий 2 мг/мл IgE Cε3-Cε4 в буфере 308 мМ NaCl и 20 мМ фосфата, рН 7,4, смешивали с раствором, содержащим 10,6 мг/мл Fab Антитела 11 в 50 мМ ацетата натрия, 100 мМ NaCl, рН 5,5 при стехиометрическом соотношении от 1 до 1,1 IgE Fc3-4 гомодимера и гетеродимера Fab Антитела 11, соответственно. Смесь оставляли на льду на ночь, после чего осуществляли гель-фильтрацию на HiLoad Superdex200 16/60 колонке (GE Healthcare), уравновешенной с помощью 20 мМ Трис HCl рН 7,6 и 0,15 М NaCl. Собирали основной пик, содержащий комплекс, и концентрировали до 10,4 мг/мл перед использованием в экспериментах по кристаллизации.
Кристаллизация комплекса IgE Cε3-Cε4 Fab Антитела 11.
Кристаллизацию осуществляли в соответствии со способом капельной диффузии паров. Капли, содержащие равный объем белка и раствора емкости (150+150 нл) помещали в планшет Crystal Quick на 96 ячеек (Greiner Bio-one) с объемами резервуара 80 мкл. Комплекс кристаллов выращивали в капле с раствором емкости 100 мМ MgCl2, 100 мМ цитрата натрия рН 5,0 и 15% PEG 4000 в течение периода времени 2-3 недели при 4°C. Кристаллы собирали в криопротекторном растворе (100 мМ MgCl2, 100 мМ цитрата натрия рН 5,0 и 15% ПЭГ 4000, который доводили до содержания 20% глицерина путем прибавления 100% глицерина) и медленно охлаждали с помощью жидкого азота.
Сбор данных и структурный раствор комплекса IgE Cε3-Cε4 Fab Антитела 11.
Собирали данные дифракции от единичных кристаллов в европейской лаборатории синхротронного излучения (ESRF) в Гренобле при использовании плоского пучка ID-29. Исходный массив данных (массив данных 1, Таблица 7) регистрировали до разрешения 3 Å, позднее собирали данные при более высоком разрешении 2,85 Å, при этом оба принадлежали к пространственной группе Р3221. Данные обрабатывали с помощью autoPROC (Global Phasing Limited GPhL, Cambridge, UK). Статистическая обработка данных представлена в Таблице 7. Ассиметричная единица содержит три Fab молекулы и три молекулы IgE Cε3-Cε4, что соответствует содержанию растворителя 54 %. Структуру разрешали с помощью способа молекулярного замещения при использовании программы PHASER (Read 2001, Storoni и др. 2004, МсСоу и др. 2005). Исходные модели для Fab фрагмента и IgE Fc домена получали из ранее описанных структур 1AQK (Faber и др. 1998) и 1FP5 (Wurzburg и др. 2000).
Применение цельного Fab в качестве модели исследования было неудачным по причине вариаций углов в шарнирном участке между вариабельным доменом и константными доменами. Вместо этого готовили и идентифицировали в начальном прогоне PHASER две отдельные модели исследования, состоящие из вариабельных доменов и константных доменов, соответственно (Read 2001, Storoni и др. 2004, МсСоу и др. 2005). Эти последние присоединяли до завершения Fab фрагментов. В общей сложности два Fab фрагмента и один вариабельный домен идентифицировали таким способом. Последующие прогоны молекулярного замещения размещали три IgE Cε3-Cε4 молекулы, одна из которых должна быть опять укорочена для включения одного домена 4. На этой стадии были собраны данные лучшего качества (массив данных 2, Таблица 7), и эта модель оптимизировалась с учетом новых данных при использовании программы autoBUSTER (Global PHMeering Limited GPhL, Cambridge, UK). В дальнейшем аминокислоты Fab молекул подвергали изменению в ручном режиме до корректировки последовательности Антитела 11 при использовании графической программы COOT (Emsley & Cowtan 2004). После второго цикла ограниченной макисмально правдоподобной оптимизации с использованием изотропных В факторов оптимизации в Refmac (CCP4 1994) оставшиеся домены последних Fab и IgE молекул вручную встраивали в электронную плотность. Два дополнительных признака расширенной электронной плотности наблюдали как выступающие из аминокислотного остатка Asn394 в полость между IgE молекулами. Это интерпретировали как гликозилирование и, таким образом, прибавляли N-ацетилглюкозамин и от трех до четырех единиц маннозы к моделям IgE. Дополнительные циклы оптимизации включали выстраивание в ручном режиме участков петли в COOT (Emsley & Cowtan 2004), осуществляемые с помощью оптимизации либо в autoBUSTER (Global Phasing Limited GPhL, Cambridge, UK), либо в Refmac5 (Murshudov и др. 1997) при использовании TLS для индивидуальных доменов и ограничений некристаллографической симметрии (NCS) для IgE молекул. В общей сложности были встроены 213 воды при использовании опции отнятия воды в Refmac5 (Murshudov и др. 1997), после чего осуществляли визуальную проверку. В заключительном цикле оптимизации ликвидировали NCS ограничения, что приводило к получению заключительной модели с R=20,0 % и Rсвоб.=27,0%.
Ссылки, цитируемые в Примере 7
Davies A. Greene A. Lullau E. Abbott WM. Optimisation and evaluation of a high-throughput mammalian protein expression system. Protein Expression & Purification. 42(1):111-21. (2005).
CCP4 (Collaborative Computational Project, Number 4) (1994) The CCP4 suite: programs for protein crystallography. Acta Crystallogr D 50:760-763.
Cooper C.A., Gasteiger E., Packer N.
GlycoMod - A software Tool for Determining Glycosylation Compositions from Mass.
Spectrometric Data.
Proteomics 1:340-349 (2001).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr D60:2126-2132 (2004).
Faber, C., Shan, L., Fan, Z., Guddat, L.W., Furebring, С., Ohlin, M., Borrebaeck, C.A., Edmundson, A.B. Three-dimensional structure of a human Fab with high affinity for tetanus toxoid. Immunotechnology: 253-270 (1998).
Kabat, E.A., Wu, T.T., Perry, H., Gottesman, K. and Foeller, C. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition. NIH Publication No. 91-3242.
Karpusas, M., Lucci, J., Ferrant, J., Benjamin, C., Taylor, F.R., Strauch, K., Garber, E., Hsu, Y.M. (2001) Structure ofCD40 ligand in complex with the Fab fragment of a neutralizing humanized antibody. Structure 9, 321, (2001).
Leslie A. (1991) Macromolecular data processing. In Moras, D., Podjarny, A.D. and Thierry, J.C. (eds), Crystallographic Computing V. Oxford University Press, Oxford, UK, pp.27-38.
McCoy, A.J., Grosse-Kunstleve, R.W., Storoni, L.C. & Read, R.J. Likelihood-enhanced fast translation functions. Acta Cryst D61, 458-464 (2005).
Murshudov, G.N., Vagin, A.A., and Dodson, E.J. (1997) Refinement of Macromolecular structures by the maximum-likelihood method, Acta Crystallogr D53:240-255.
Padavattan, S., Schirmer, Т., Schmidt, M., Akdis, C., Valenta, R., Mittermann, I., Soldatova, L., Slater, J., Mueller, U. & Markovic-Housley, Z. Identification of a B-cell Epitope of Hyaluronidase, a Major Bee Venom Allergen, from its Crystal Structure in Complex with a Specific Fab. J Mol Biol 368, 742-752. (2007).
Persic L. Roberts A. Wilton J. Cattaneo A. Bradbury A. Hoogenboom HR. An integrated vector system for the eukaryotic expression of antibodies or their fragments after selection from phage display libraries. Gene. 187(1):9-18. (1997).
Read, R.J. Pushing the boundaries of molecular replacement with maximum likelihood. Acta Cryst. D57, 1373-1382 (2001).
Rossmann, M.G. (edt): "The Molecular Replacement Method" Gordon & Breach, New York (1972).
Storoni, L.C., McCoy, A.J. & Read, R.J. Likelihood-enhanced fast rotation functions. Acta Cryst D60, 432-438 (2004).
Wurzburg, B.A., Garman, S.C. and Jerdetzky, H.S. Structure of the human IgE-Fc С epsilon 3-C epsilon 4 reveals conformational flexibility in the antibody effector domains. Immunity, 13, 375-385 (2000).
Пример 8. Оценка общей безопасности и способности герминизированных анти-IgE mAb индуцировать снижения количества тромбоцитов у неполовозрелых животных обезьян циномолгус
Изучающее (не соответствующее GLP) исследование осуществляли у неполовозрелых обезьян циномолгус для оценки общей безопасности и относительной возможности антител в соответствии с изобретением анти-IgE mAb, Антитела 11 IgG1, Антитела 11 IgG2 и другого анти-IgE антитела Е48 вызывать снижение количества тромбоцитов крови.
Задачи исследования состояли: 1) определить общую безопасность и относительные , способности трех кандидатных анти-IgE mAb индуцировать снижение количества тромбоцитов/ТСР и ассоциированные эффекты у неполовозрелых обезьян циномолгус; 2) определить предварительные фармакокинетические параметры для mAb у обезьян; 3) оценить способность трех кандидатных mAb вызывать снижение уровня свободного IgE и определять (PK/PD) взаимоотношение между концентрацией mAb и уровнями свободного IgE.
Материалы и способы для Примера 8
Восемнадцать выведенных для данной цели обезьян циномолгус (Масаса fascicularis) получали из Bioculture, Mauritius. Животные имели возраст от 63 до 67 недель на начало введения дозы. Обезьян подвергали предварительному отбору (из большого пула, состоящего из 100 животных) на наличие высоких уровней IgE (ед./мл) и разделяли на 3 группы, каждая и которых содержала 3 самца и 3 самки обезьян, которые получали Антитело 11 IgG1 (Группа 1), Антитело 11 IgG2 (Группа 2) или Е48 (Группа 3). Каждое из 3 mAb рецептировали в 50 мМ ацетата натрия, 100 мМ NaCl, pH 5,5 и вводили животным в объеме дозы 2 мл/кг путем медленной внутривенной инъекции (при использовании механизированного шприца/насоса для инфузии) при скорости введения 1 мл/мин.
Животные получали дозу один раз в неделю (в течение 5 недель/5 доз) с увеличивающимися уровнями доз 1 мг/кг, 30 мг/кг и 100 мг/кг (×3) в Дни 1, 8, 16, 22 и 29 (Таблица 8). Дополнительные дозы Антитела 11 IgG1 и Антитела 11 IgG2 вводили Группам 1 и 2, соответственно в День 37. 2 самых высоких уровня доз были теоретически определены как такие, которые приводят к получению концентраций в сыворотке крови, которые ранее были показаны в качестве приводящих к тромбоцитопении (TCP) с Xolair у неполовозрелых обезьян циномолгус. Низкая доза, как ожидалось, позволяла определить способность mAb вызывать снижение уровней IgE.
За животными наблюдали в течение 8 недель после введения дозы в День 28 и исследовали на излечение от каких-либо токсикологических эффектов
Всех животных подвергали ежедневному исследованию на признаки недомогания или очевидной токсичности, а также регистрировали вес тела и потребление пищи. Кроме того, каждое животное проходило тщательную проверку физического состояния ежедневно в течение периодов введения дозы и, по крайней мере, в течение одной недели во время периодов без введения дозы. Всех животных также подвергали наблюдению перед введением каждой дозы и через 0, 5, 2, 6, 24, 48 и 168 часов после введения дозы.
Образцы крови для анализа стандартных гематологических параметров (включая количество тромбоцитов, собранных в ЭДТА) и параметров коагуляции (собранных в цитрате тринатрия) брали из бедренной вены/артерии дважды в период, предваряющий лечение (недели -2 и -1). Дополнительные образцы для подсчета тромбоцитов и стандартной гематологии собирали через 24 часа и 144 часа после каждого введения дозы (Дни 2, 7, 9, 14, 17, 22, 23, 26, 30 и 35; образцы из Групп 1 и 2 только в Дни 38 и 43) и каждые 2 недели в течение 8-недельного периода выздоровления (Дни 43, 57, 71 и 82). Образцы для определения коагуляции собирали через 144 часа после каждого введения дозы (Дни 7, 14, 22, 26 и 35) и в конце периода выздоровления (День 82). Образцы для определения коагуляции также собирали в День 57. Образцы крови для активации комплимента (С3а, C3b и ВВ фрагменты) брали один раз в период, предваряющий лечение(неделя -1) и примерно через 24 часа после завершения периода лечения (День 30).
Образцы сыворотки для TK анализа брали у всех трех групп в День 1 перед введением дозы, через 0, 5, 6, 12, 24, 48, 144 часа после введения дозы, в Дни 8, 16, 22 и 29 через 0, 5, 24 и 144 часов после введения дозы, в День 29 (только Группы 3 и 4) через 336, 672, 1008, 1272 часа после введения дозы, в День 37 (только Группы 1 и 2) через 0, 5, 24, 144, 480, 816, 1080 часов после введения дозы. Образцы анализировали на mAb при использовании генерического сэндвич-иммуноанализа (при использовании биотинилированного человеческого IgE для mAb и меченного Аlеха-647 мышиного античеловеческого IgG агента для определения) и Gyrolab Bioaffy оборудования (включающего колонки с гранулами стрептавидина). Дополнительные образцы для анализа IgE собирали в День 1 перед введением дозы, через 0, 5, 6, 12, 24, 48, 144 часа после введения дозы, в Дни 8, 16, 22 и 29 через 0, 5, 144 часа после введения дозы и в конце исследования (День 82) через 1272 часа (Группы 3 и 4) или 1080 часов (Группы 1 и 2) после введения заключительной дозы. Образцы анализировали на свободный IgE с помощью иммуноанализа при использовании системы ImmunoCap (Phadia АВ, Uppsala, Sweden) при использовании человеческого IgG-FcεRIa для захвата свободного IgE и античеловеческого IgE кролика (PCS-конъюгат) для определения.
По окончании эксперимента в День 85 осуществляли полное макроскопическое исследование под общим наблюдением патоморфолога и регистрировали все повреждения. Определяли абсолютный вес органов и соотношение вес органа: вес тела. Ткани из ряда органов собирали и хранили в замороженном виде, однако не осуществляли никакого микроскопического исследования (за исключением макроскопических патологий или непредусмотренной смерти, смотри выше).
Результаты для Примера 8
Наблюдения в отношении общей безопасности
Все 3 mAb были хорошо переносимыми при отсутствии каких-либо клинических признаков недомогания на протяжении исследования, за исключением одного животного, получавшего Антитело 11, которое было отослано на вскрытие трупа досрочно по причине ухудшения общего клинического состояния и сниженного веса тела. Поскольку состояние этого животного ухудшилось в период восстановления и не было отмечено никаких данных во время патологического или гематологического исследования этих животных, наблюдаемые эффекты не предполагаются как такие, которые связаны mAb. Во всех группах отмечались случаи мягких или жидких испражнений, однако поскольку эти наблюдения не были связаны с дозой и не наблюдались у всех животных или во все точки времени у одного и того же животного, а также наблюдались многократно в период введения дозы и в восстановительные периоды, то является маловероятным, что они связаны с mAb. Среднее значение веса тела и среднее значение прибавки веса тела показали некоторую индивидуальную вариацию у животных в пределах каждой группы в течение всего периода исследования. Однако все животные набирали вес, как и ожидалось, в течение периода лечения (за исключением одного животного, которое обсуждалось выше) и не было четкого отличия между группами. Не было отмечено явственных эффектов, связанных с лечением, в отношении абсолютного или относительного веса органов в какой-либо из групп. В общем случае патологическое исследование и микроскопическое исследование патологии не обнаружили никаких наблюдений в ограниченном спектре исследованных тканей, что дало бы возможность предположить какой-либо эффект лечения mAb.
Токсикокинетики (TK) и уровни IgE
Не наблюдали зависимых от пола отличий в TK в этом исследовании. В общем случае воздействие было подобным для этих 3 mAb и оказалось линейным в интервале дозы 1-100 мг/кг. Средние значение профилей TK Антитела 11 IgG1, Антитела 11 IgG2 и Е48 показаны на Фигуре 12. Не наблюдали явного влияния падения уровня IgE на TK, даже при самом низком уровне дозы. TK этих 3 антител оказались типичными для человеческого IgG у обезьянах циномолгус.
Среднее значение максимума наблюдаемой концентрации (Cmax) после введения последней дозы 100 мг/кг составляло 18700, 15900 и 24000 нМ для Антитела 11 IgG1, Антитела 11 IgG2 и Е48 соответственно. Среднее значение заключительного периода полураспада для TK после введения последней дозы 100 мг/кг составляло приблизительно 10-13 дней. Не было подтверждения сниженного TK воздействия по причине потенциального развития античеловеческих антител примата у этих животных.
Средние значения профилей свободного IgE после еженедельного введения дозы Антитела 11 IgG1, Антитела 11 IgG2 и Е48 при различных уровнях дозы у обезьян циномолгус показаны на Фигуре 13. Усредненная базовая линия IgE перед тем, как животные получали первую дозу, составляла 514, 414 и 690 нг/мл для групп, получающих Антитело 11 IgG1, Антитело 11 IgG2 и Е48 соответственно. На День 1 доза 1 мг/кг индуцировала 75-80% снижение уровня свободного IgE через 1 час после введения дозы. По причине низкого воздействия после введения дозы 1 мг/кг уровень свободного IgE возвращался к уровню базовой линии в течение 1 недели. Более высокие дозы приводили к постоянной супрессии уровня свободного IgE в течение периода лечения. Уровень свободного IgE возвращался к уровню базовой линии для двух групп 2 Антитела 11 в конце исследования, в то время как уровень свободного IgE в Е48 группе оставался супрессированным.
Влияние на тромбоциты
Ни одно из 3 mAb (Антитело 11 IgG1, Антитело 11 IgG2 и Е48) не индуцировал достоверного снижения количества тромбоцитов в любой момент времени у любого животного, за исключением одного животного, которое получало Антитело 11 IgG1, которое имело снижение уровня тромбоцитов (34,9%) в единственный момент времени День 29 (спустя 24 часа после введения дозы 100 мг/кг в День 28). Четвертая доза Антитела 11 IgG1 и Антитела 11 IgG2 в День 37 не индуцировала какого-либо дополнительного снижения уровня тромбоцитов у этого или другого животного в этих 2 группах.
Фигура 14 показывает диаграмму количества тромбоцитов (×109/л), выраженного как процентное отличие от среднего значения двух предварительных доз против отличия концентрации от животных в Группые 1 (обработанные Антителом 11 IgG1). Эта диаграмма является характерной для 16 животных из 3 групп, которые не продемонстрировали достоверного влияния на тромбоциты [изменение для животного группы Антитело 11]. Фигура 15 показывает подобную диаграмму для животных Группы 1 (обработанные Антителом 11 IgG1), которые продемонстрировали временное достоверное снижение (35% ниже базовой линии) количества тромбоцитов на День 29.
Интересно отметить, что обезьяны, обработанные Антителом 11 IgG1, которые продемонстрировали временное достоверное снижение количества тромбоцитов после введения дозы на День 29 имели самое высокое значение Cmax (29400 нмол/л) (но не воздействие) в это время. Уровни в плазме крови последовательно резко падали, и количество тромбоцитов возвращалось к значениям, которые наблюдали до введения предварительной дозы. Это указывает на возможность того, что может требоваться более высокое пороговое значение концентрации в плазме крови для того, чтобы индуцировать снижение количества тромбоцитов. Однако одно животное, обработанное Е48, достигало подобных уровней (28500 нм/л) при отсутствии соответствующего влияния на тромбоциты (Фигура 14).
Другие гематологические эффекты
За исключением количества тромбоцитов (смотри ниже), не отмечали соответствующего влияния лечения с помощью mAb на большинство гематологических параметров (концентрация гемоглобина, показатель гематокрита, средний объем эритроцитов, среднее содержание гемоглобина в эритроците, ширина распределения эритроцитов по объемам, тромбоцитокрит, ширина распределения тромбоцитов по объемам, количество эритроцитов, среднее содержание гемоглобина в эритроците, ширина распределения гемоглобина, средний объем тромбоцита, количество ретикулоцитов, общая и дифференциальная лейкоцитарная формула) и параметров коагуляции крови (протромбиновое время, активированное частичное тромбопластиновое время). Повышение количества ретикулоцитов наблюдали во всех группах, однако изменения не были связаны взаимоотношением доза/воздействие, не были согласованны в пределах группы (животные в группе имели более высокий, более низкий или неизмененные уровни по сравнению со значениями периода до введения доз) или у одного и того же животного (значения для одного и того же животного поднимались и опускались в период между различными моментами времени независимо от воздействия), и при отсутствии параллельной контрольной группы взаимосвязь с лечением mAb не могла быть полностью определена в этот момент времени. Любое из таких изменений имело обратное развитие к концу периода восстановления. Не отмечали никаких достоверных воздействий, связанных с лечением, на активацию комплемента (С5а, С3а или ВВ фрагменты).
Обсуждение и выводы
Исследование показало, что анти-IgE mAb, Антитело 11 IgG1, Антитело 11 IgG2 и Е48 были хорошо переносимыми при введении при высоких уровнях повторяющихся доз (вплоть до 100 мг/кг) неполовозрелым обезьянам циномолгус при отсутствии достоверных вредных токсикологических эффектов. Только одно животное из 18 обезьян показало снижение количества тромбоцитов в единственный момент времени после введения 100 мг/кг Антитела 11 IgG1, когда концентрации mAb в плазме крови достигали значения почти 30000 нмоль/л. Значения Cmax концентрации в плазме крови, которые достигались при использовании всех 3 mAb в этом исследовании были намного больше тех, которые будут достигнуты при клиническом использовании (например, 200 нмоль/л).
Пример 9. Функциональное ингибирование влияния IgE на FcεRI и CD23
Способность оптимизированного Антитела 11 ингибировать функционально взаимодействие IgE с FcεRI и CD23 оценивали в опосредованном IgE анализе уничтожения клеток, адаптированном по сравнению с Bracher и др. (Journal Immunol. Methods 2007 323:160-171). U937, предварительно обработанные с помощью IL-4, были продемонстрированы как такие, которые вырабатывают как FcεRI, так и CD23. При совместном культивировании с опухолевыми клетками яичника IGROV1 в присутствии IgE, специфического для антигена, который экспрессируется на IGROV1 клетках, U937 клетки были способными уничтожать опухолевые клетки. Это уничтожение было опосредовано как механизмами цитотоксичности, так и фагоцитоза, которые, как было показано, запускаются с помощью взаимодействия IgE с FcεRI и CD23, соответственно, на U937 эффекторных клетках.
Антитело 11 и контроль изотипа оценивали в этом анализе на ингибирование опосредованного уничтожения с помощью либо FcεRI, либо CD23. Подробная пропись для этой процедуры обеспечивается в разделе "Материалы и способы". Вкратце, титрования исследуемого IgG смешивали с целевым специфическим (MOv18) или нерелевантным (NIP) IgE перед инкубацией с U937 эффекторными клетками, стимулированными с помощью IL-4, и меченными IGROV1 целевыми клетками. После 2,5 часов инкубации промывали клетки, окрашивали анти-CD89-фикоэритрин антителом (BD Biosciences) и йодидом пропидия (Molecular Probes). После промывания флуоресценцию клеток анализировали при использовании FACSCalibur проточного цитометра (BD Biosciences). Флуоресцентные красители, указанные выше, использовали для разделения живых клеток, клеток, уничтоженных с помощью цитотоксичности, и клеток, уничтоженных путем фагоцитоза. В отличие от изотипического контроля антитела, антитело 11 является способным ингибировать опосредованную IgE/FcεRI цитотоксичность (Фигура 16) и опосредованный IgE/CD23-фагоцитоз (Фигура 17).
Материалы и способы - Пример 9
Антитела оценивали на ингибирование опосредованного IgE уничтожения опухолевых клеток IGROV1 с помощью U937 клеток. IGROV1 клетки (линия клеток карциномы яичника человека) и U937 клетки (миеломоноцитарная линия клеток человека) поддерживали в культуральной среде [RPMI1640,10% об./об. FCS, 2 мМ глутамина, 5000 ед./мл пенициллина, 100 мкг/мл стрептомицина (все от Invitrogen)] при использовании стандартных процедур культуры тканей. MOv18 IgE, направленный против FBP (связывающий фолат белок), который экспрессируется на IGROV1 клетках, использовали в качестве специфического для опухоли антитела. NIP (гаптен 4-гидрокси-3-нитрофенацетил)-специфический IgE использовали в качестве контроля нерелевантного антитела. MOv18 и NIP антитела получали так, как описано выше у Gould и др., (1999) Eur. J. Immunol. 29:3527-3537.
U937 клетки предварительно обрабатывали в течение 4 дней перед экспериментом по уничтожению клеток с помощью 10 нг/мл рекомбинантного человеческого IL-4 (R&D Systems) для того чтобы повысить экспрессию CD23. За день до проведения эксперимента по уничтожению клеток IGROV1 целевые клетки метили с помощью флуоресцентного красителя CFSE (карбокифлуоресцеин диацетат сукцинимидильный эстер. Molecular Probes). Вкратце, клетки подвергали трипсинизации (трипсин/ЭДТА, Gibco), промывали в культуральной среде и ресуспендировали в PBS при концентрации 50×106 клеток/мл. Потом клетки инкубировали при 37°C в течение 10 минут с CFSE в концентрации 0,01 мМ. После осуществления мечения клетки промывали один раз в ледяной культуральной среде и потом инкубировали в течение ночи при 37°C, 5% CO2.
Для оценки ингибиторного эффекта Антитела 11 получали разведения антитела в пробирках 12×75 мм (Falcon, BD Biosciences) и 2 мкг MOv1 или NIP IgE прибавляли с получением заключительного объема 80 мкл. Эту смесь инкубировали без клеток в течение 30 минут. Стимулированные с помощью IL-4 U937 клетки один раз промывали в среде и ресуспендировали при концентрации 1,33×106 клеток/мл. Меченные CFSE IGROV1 клетки трипсинизировали, один раз промывали в среде и ресуспендировали при концентрации 4×105 клеток/мл. Прибавляли клетки к пробиркам, содержащим антитела (120 мкл для U937 клеток и 200 мкл для IGROV1 клеток), смешивали и инкубировали в течение 2,5 часов при 37°C, 5% CO2. Потом клетки промывали в ледяном FACS буфере (PBS, свободный от кальция/магния, 5% козья сыворотка) и инкубировали в течение 25 минут с анти-CD89-фикоэритрин антителом (BD Biosciences, 10 мкг/мл) для мечения U937 эффекторных клеток. Клетки еще раз промывали в ледяном FACS буфере и погибшие клетки окрашивали путем прибавления 0,25 мкг/мл йодида пропидия (Molecular Probes). Через 15 минут пребывания при 4°C клетки промывали дегяным FACS буфером, ресуспендировали в 250 мкл ледяного FACS буфера и анализировали флуоресценцию при использовании FACSCalibur проточного устройства для подсчета клеток (BD Biosciences) в соответствии с инструкциями производителя. Клетки с релевантным однообразным окрашиванием использовали для установления напряжения и компенсации каналов определения (FL1, FL2 и FL3). Комбинация красителей, используемая в этом анализе, позволяла осуществлять селекцию популяций различных клеток [живые эффекторные клетки (позитивные по фикоэритрину), фагоцитированные IGROV1 опухолевые клетки (позитивные по фикоэритрину и CFSE), живые опухолевые клетки (позитивные по CFSE), мертвые опухолевые клетки (позитивные по CFSE и йодиду пропидия), мертвые эффекторные клетки (позитивные по фикоэритрину и йодиду пропидия)]. Такую селекцию использовали для подсчета целевых клеток, уничтоженных путем опосредованной FcεRI цитотоксичности (уравнение 1) и путем опосредованного CD23 фагоцитоза (уравнение 2).
Уравнение 1: %цитотоксичности={[(R1SLконтроль-R1)+R3]/R1SL}×100
где R1 = общее количество CFSE позитивных опухолевых клеток
R3 = количество уничтоженных, но интактных опухолевых клеток (без фрагментации или фагоцитоза)
R1SL контроль = Средняя величина R1 3 контрольных образцов эффектора и целевых клеток без антитела (R1 контроль спонтанной гибели).
Уравнение 2: %фагоцитоза=(R2/R1SLконтроль)×100
где R2 = количество опухолевых клеток, фагоцитированных эффекторными клетками
R1SL контроль = Средняя величина R1 3 контрольных образцов эффектора и целевых клеток без антитела (R1 контроль спонтанной гибели).
Ссылки
Все ссылки, процитированные где-либо в этом описании, включая те, которые цитируются где-либо выше, введены в данную заявку в качестве ссылок в своей целостности и для всех целей.
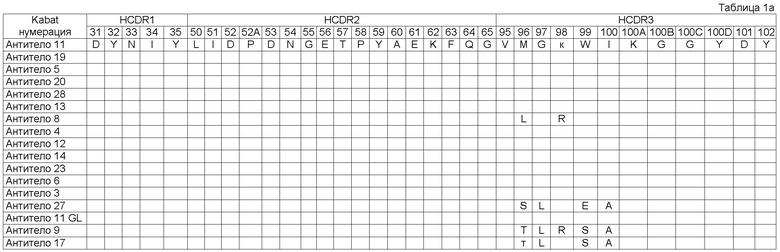
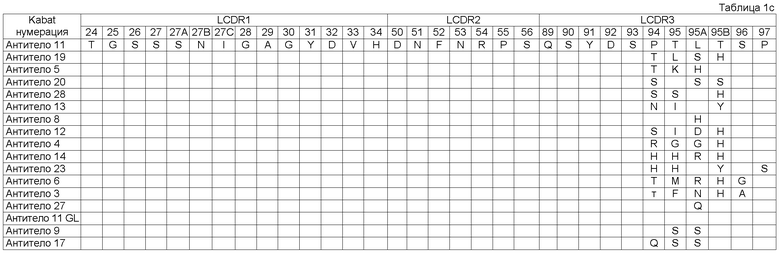
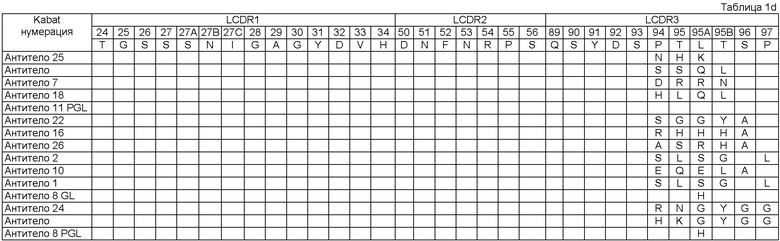
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab), СВЯЗЫВАЮЩИЕСЯ С ИНТЕРФЕРОНОМ- γ ЧЕЛОВЕКА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КЛЕТКА, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО АНТИТЕЛА И АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2013 |
|
RU2539752C2 |
| АНТИТЕЛО ПРОТИВ В7-Н3, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2765306C2 |
| СПОСОБ ИНГИБИРОВАНИЯ АКТИВНОСТИ ЧЕЛОВЕЧЕСКОГО TNF (ВАРИАНТЫ), ПРИМЕНЕНИЕ ВЫДЕЛЕННОГО АНТИТЕЛА ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА В КАЧЕСТВЕ КОМПОНЕНТА ДЛЯ ПРОИЗВОДСТВА ЛЕКАРСТВЕННОГО СРЕДСТВА (ВАРИАНТЫ) И ВЫДЕЛЕННОЕ ЧЕЛОВЕЧЕСКОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 1997 |
|
RU2270030C2 |
| АНТИ-IgE АНТИТЕЛА | 2017 |
|
RU2816207C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ (ВАРИАНТЫ), СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ОПОСРЕДОВАННЫХ hTNFα С ИХ ПОСРЕДСТВОМ (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, РЕКОМБИНАНТНОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА (ВАРИАНТЫ) | 2005 |
|
RU2458704C9 |
| МОЛЕКУЛЫ АНТИТЕЛ ЧЕЛОВЕКА К IL-13 | 2004 |
|
RU2387667C2 |
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |
| СКОНСТРУИРОВАННЫЕ АНТИ-TGF-БЕТА АНТИТЕЛА И АНТИГЕН-СВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ | 2014 |
|
RU2681502C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО 5D3HU, СВЯЗЫВАЮЩЕЕСЯ С ОПУХОЛЕВЫМ АНТИГЕНОМ PRAME, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ АНТИТЕЛА | 2020 |
|
RU2761876C1 |
| АНТИТЕЛА, НЕ СОДЕРЖАЩИЕ FC-ФРАГМЕНТА, ВКЛЮЧАЮЩИЕ ДВА FAB-ФРАГМЕНТА, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2617970C2 |






























































































































































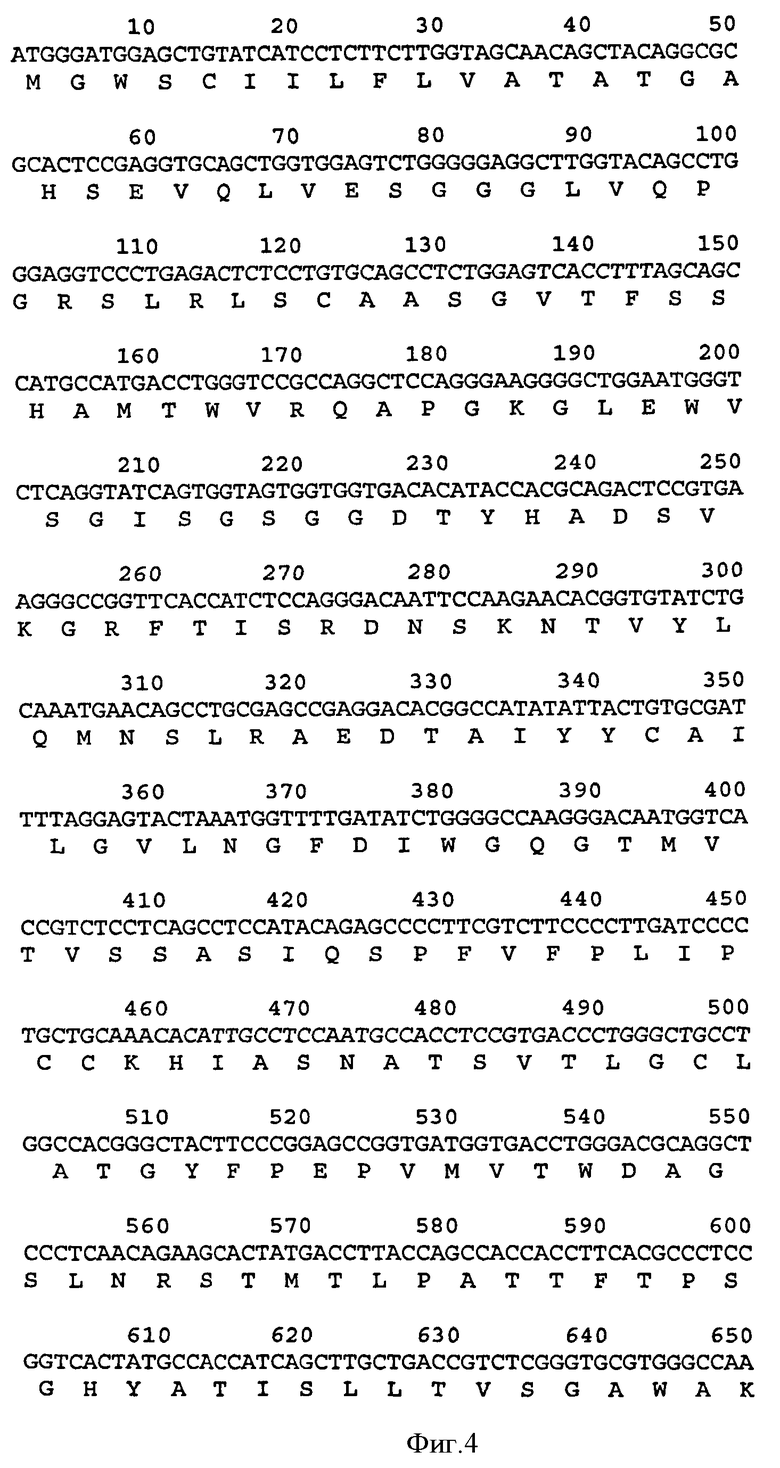

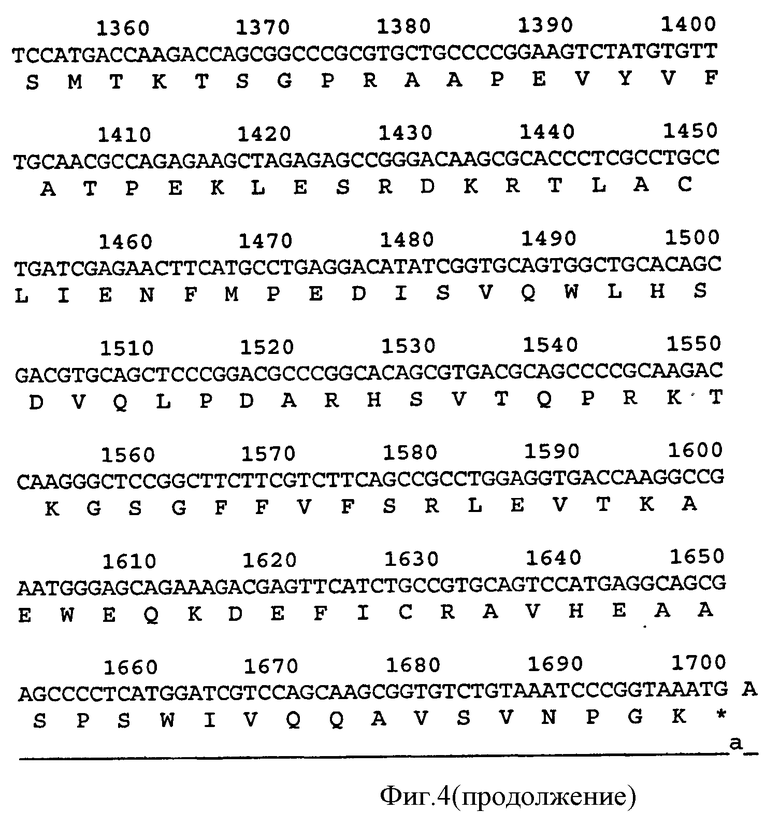
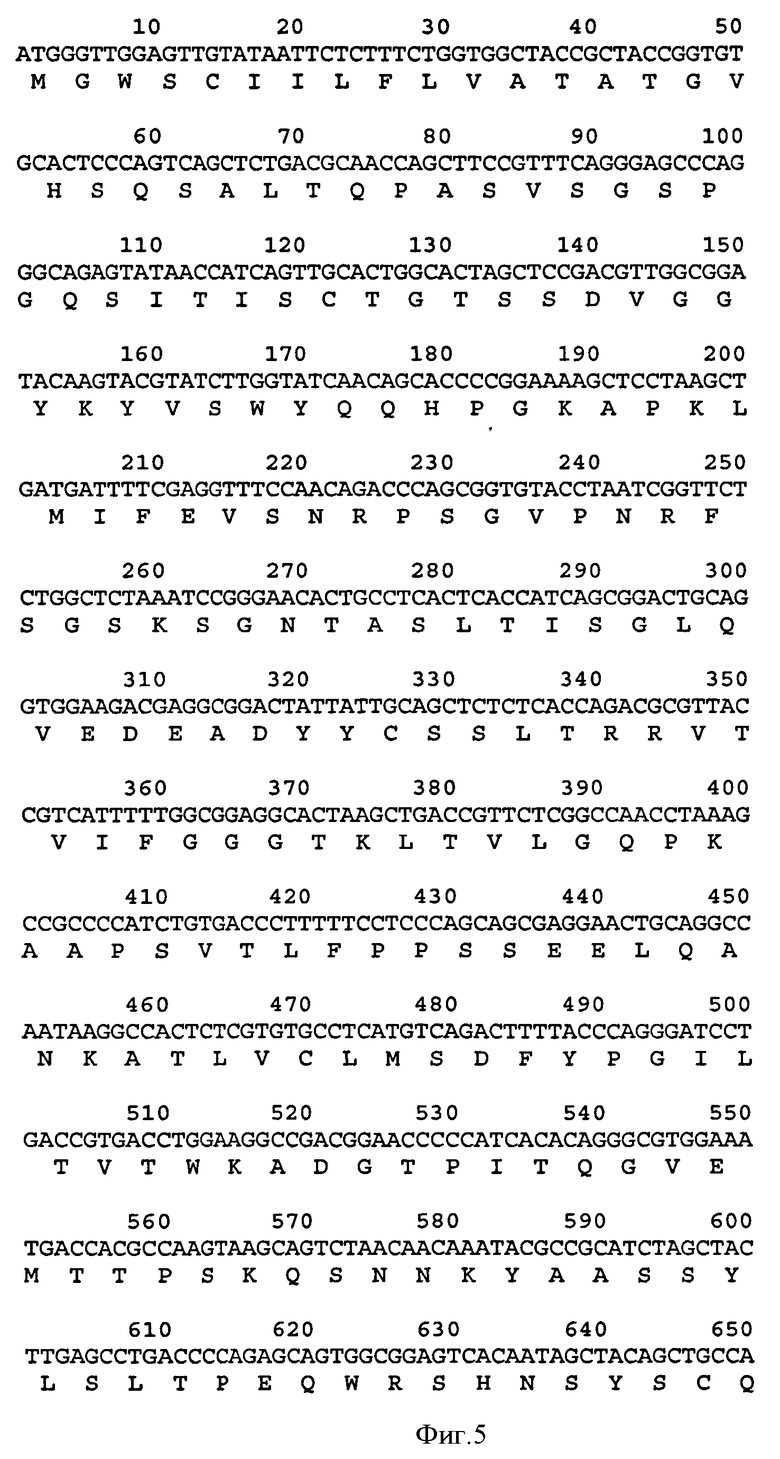
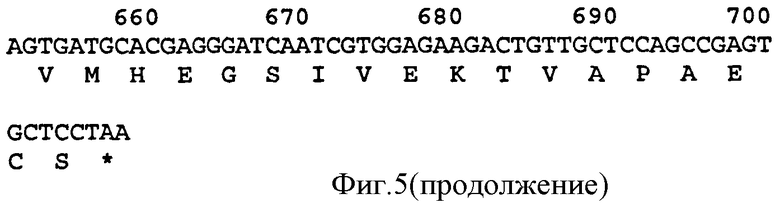
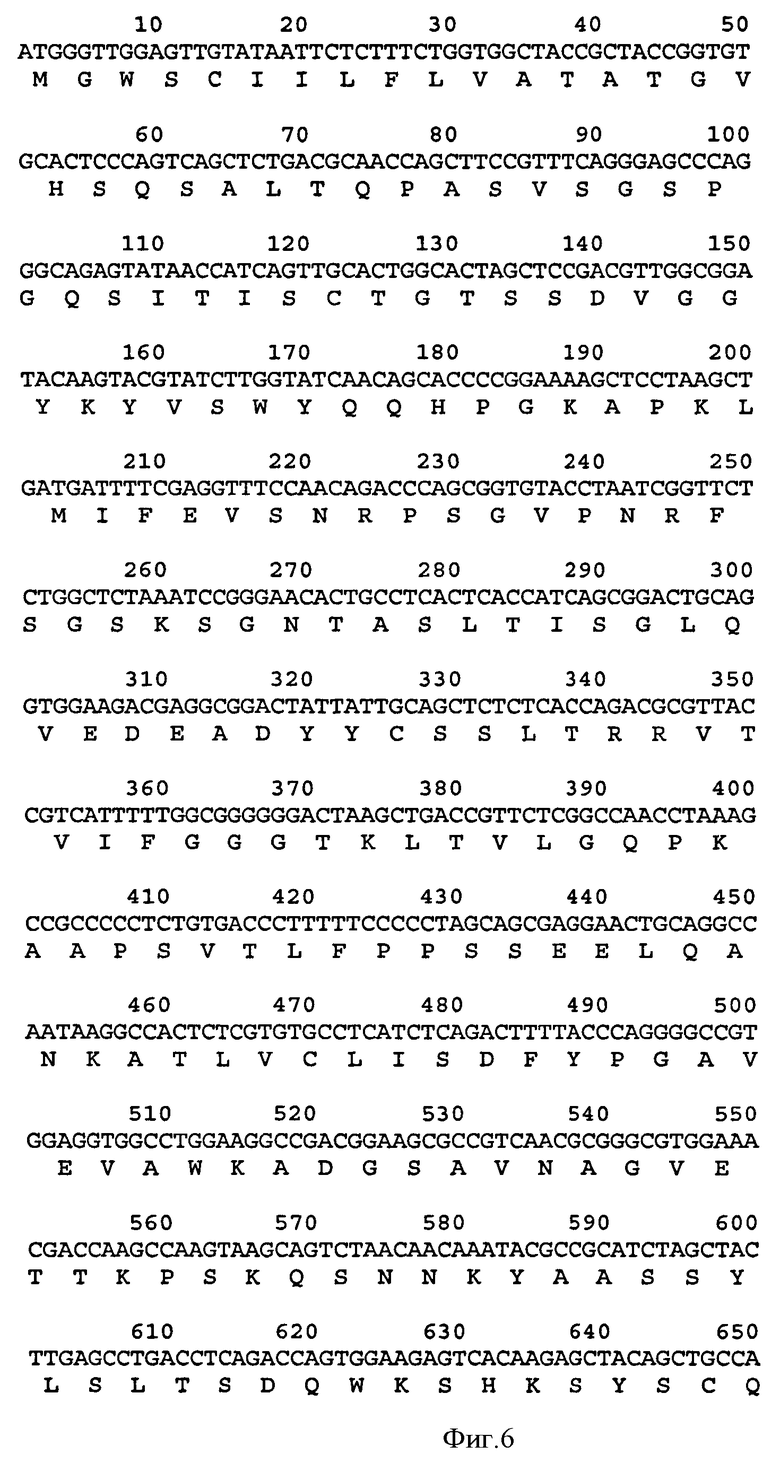
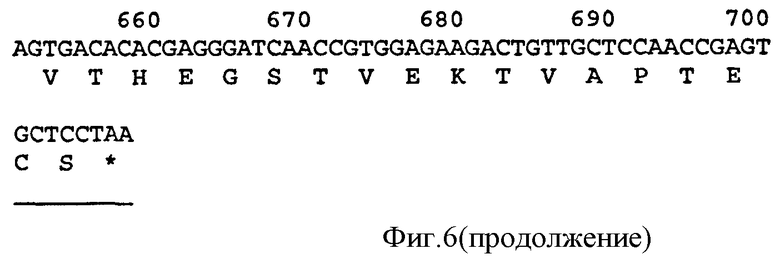
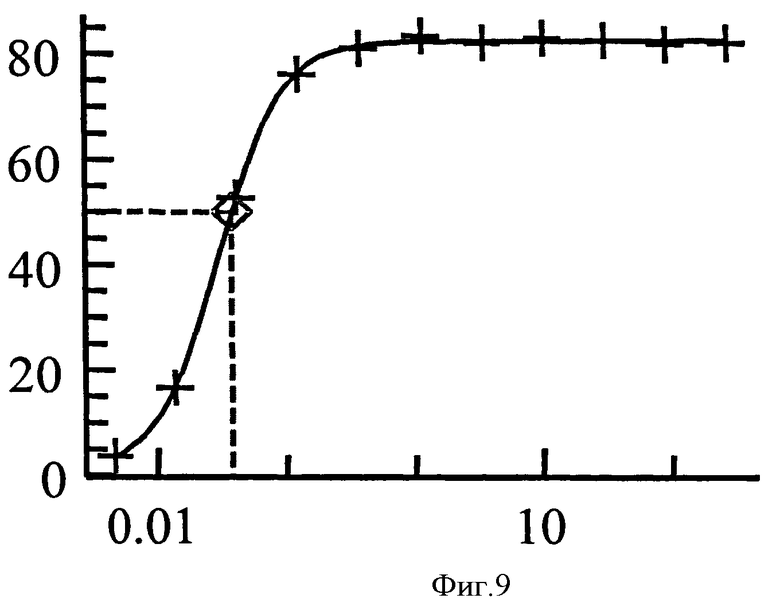
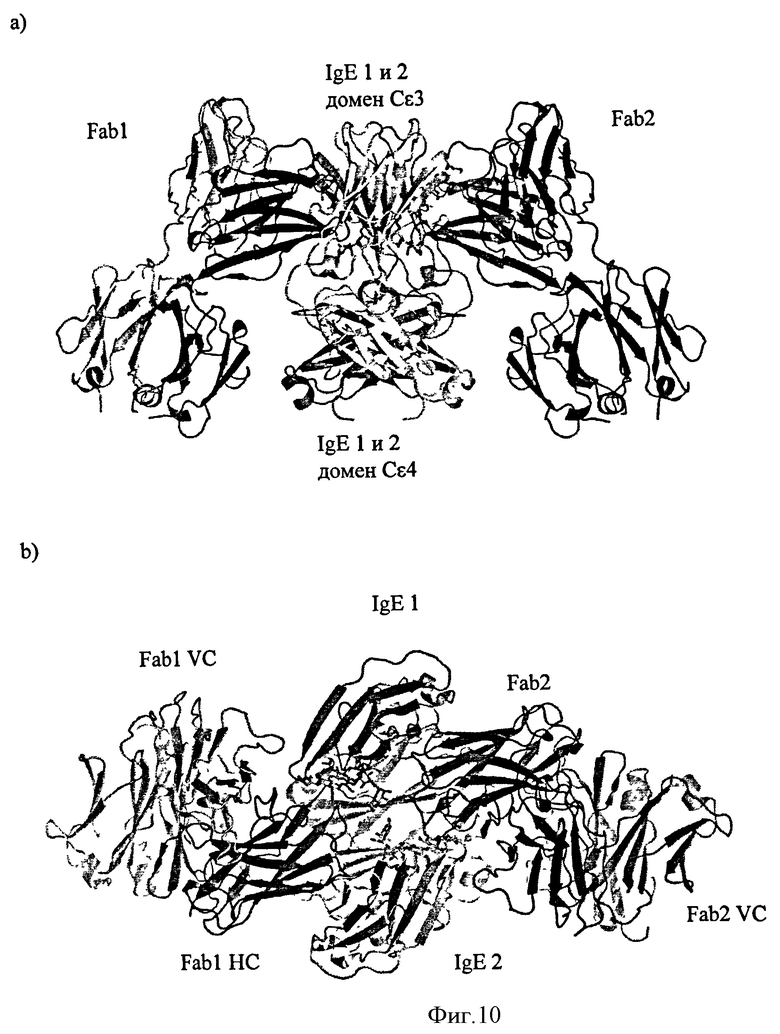
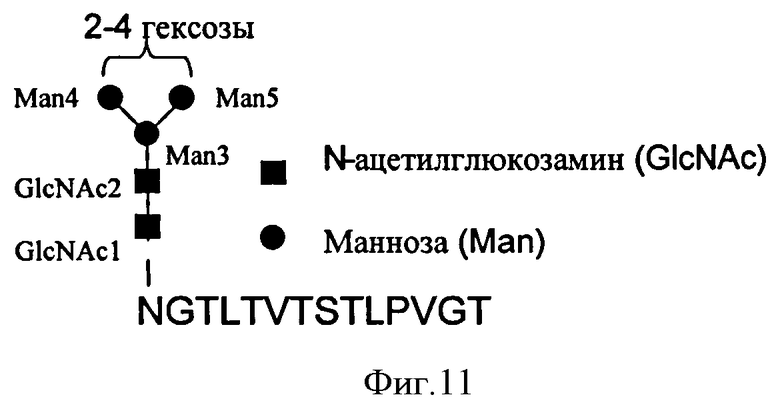
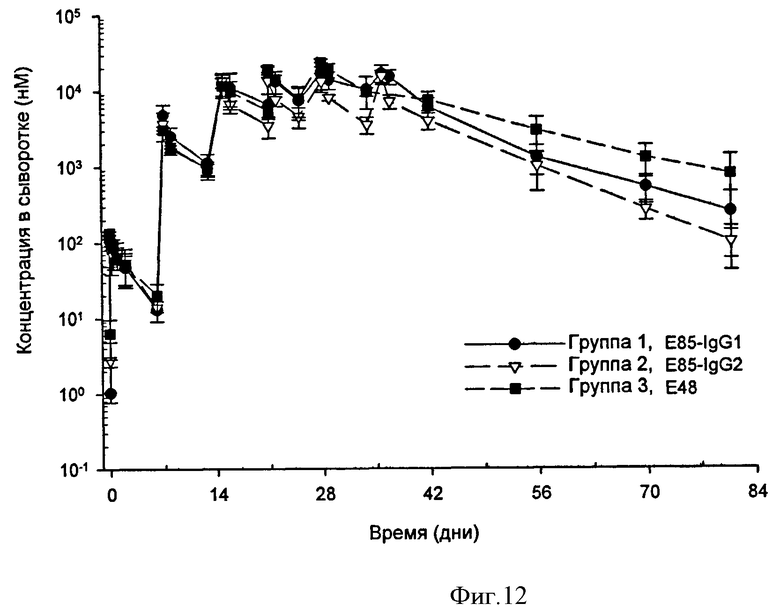
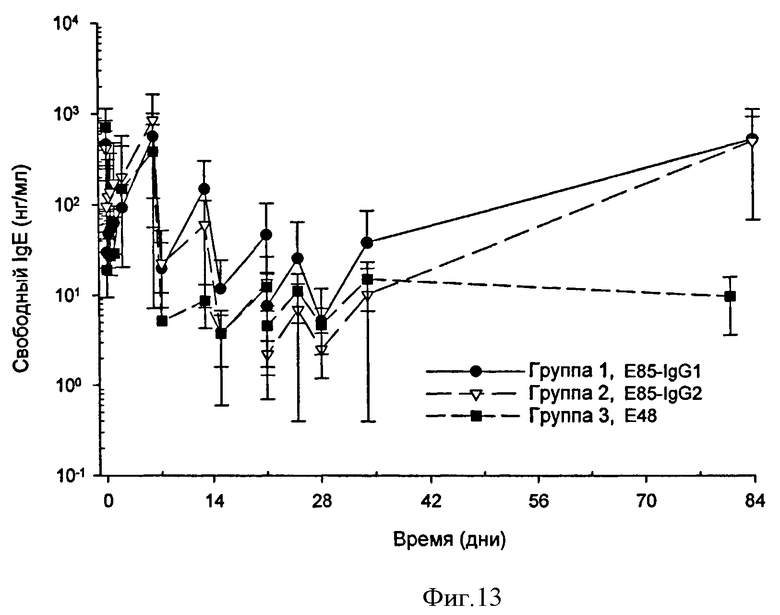
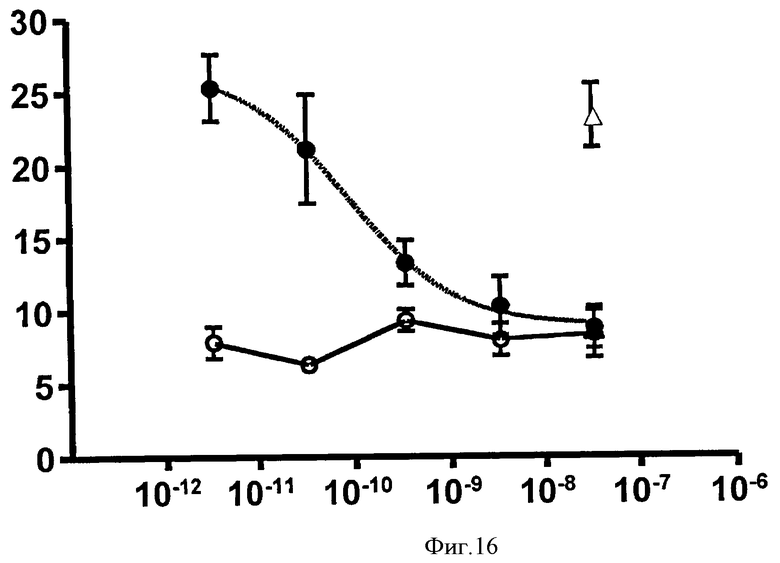
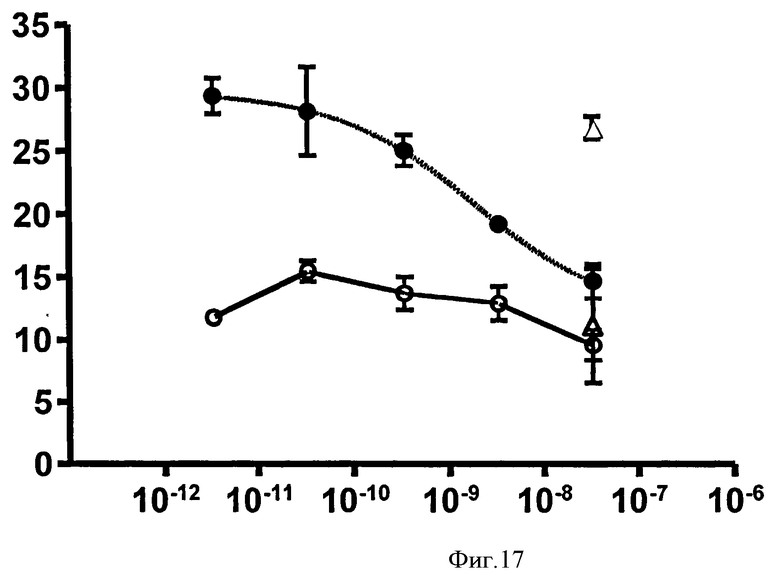
Изобретение относится к области биотехнологии, в частности к таким связывающим элементам, специфичным для IgE, как молекулам антитела и его антигенсвязывающим, фрагментам. Антитело и его варианты, специфичные для IgE, являются полезными для лечения расстройств, опосредованных IgE, включая аллергии и астму. 8 н. и 9 з.п. ф-лы, 17 ил., 8 табл., 9 пр.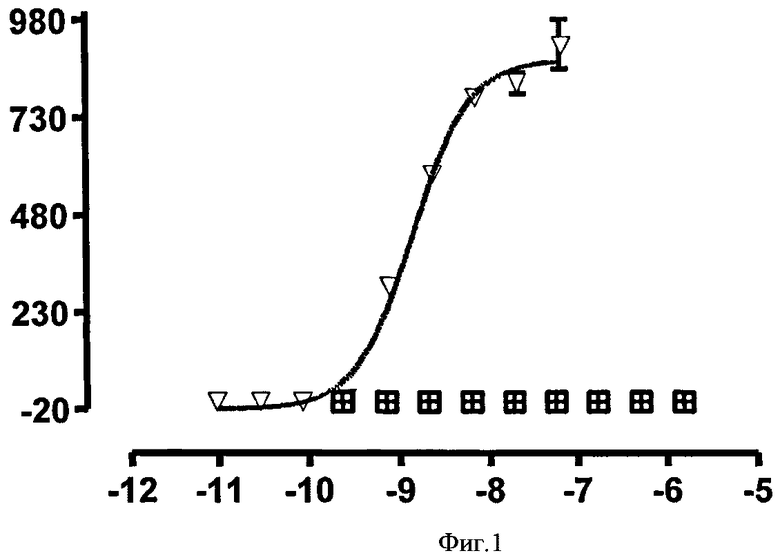
1. Изолированное антитело или его антигенсвязывающий фрагмент, специфическое(ий) для человеческого иммуноглобулина Е, где указанное антитело или его антигенсвязывающий фрагмент включает
набор CDR: HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, где в котором:
HCDR1 имеет аминокислотную последовательность SEQ ID NO: 103;
HCDR2 имеет аминокислотную последовательность SEQ ID NO: 104;
HCDR3 имеет аминокислотную последовательность SEQ ID NO: 105;
LCDR1 имеет аминокислотную последовательность SEQ ID NO: 108;
LCDR2 имеет аминокислотную последовательность SEQ ID NO: 109;
LCDR3 имеет аминокислотную последовательность SEQ ID NO: 110.
2. Изолированное антитело или его антигенсвязывающий фрагмент, специфическое(ий) для человеческого иммуноглобулина Е, молекула антитела включает набор CDR: HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, в котором:
HCDR1 имеет аминокислотную последовательность SEQ ID NO: 103;
HCDR2 имеет аминокислотную последовательность SEQ ID NO: 104;
HCDR3 имеет аминокислотную последовательность SEQ ID NO: 105, где SEQ ID NO: 105 может включать одну или более следующих замен: М в положении 2 заменен на L, S или Т; G в положении 3 заменен L или Н; K в положении 4 заменен R; W в положении 5 заменен Е, S, A, R или Т и I в положении 6 заменен А или L;
LCDR1 имеет аминокислотную последовательность SEQ ID NO: 108;
LCDR2 имеет аминокислотную последовательность SEQ ID NO: 109;
LCDR3 имеет аминокислотную последовательность SEQ ID NO: 110 где SEQ ID NO: 110 может включать одну или более следующих замен Р в положении 6 Т, S, N, R, В, Q, D, А или Е; Т в положении 7 заменен L, К, S, I, G, H, M, F, R, Q или N; L в положении 8 заменен S, Н, D, R, N или G; Т в положении 9 заменен Н, S, Y, L, N или G; S в положении 10 заменен G или А и Р в положении 11 заменен S, L или G.
3. Изолированное антитело или его антигенсвязывающий фрагмент по п.1 или 2, где указанное антитело или его антигенсвязывающий фрагмент специфично(ен) для иммуноглобулина Е и связывается с эпитопом, включающим элементы из первой тяжелой цепи IgE и элементы из второй тяжелой цепи IgE.
4. Изолированное антитело или его антигенсвязывающий фрагмент по п.3, где указанное антитело или его антигенсвязывающий фрагмент специфично(ен) для иммуноглобулина Е и связывается с эпитопом в иммуноглобулине Е, включающим:
остатки от Glu390 до Asn394, включительно, в первой тяжелой цепи IgE и Leu340, Arg342, от А1а428 до Thr434, включительно, Thr436, Ser437 и Glu472 во второй тяжелой цепи IgE.
5. Изолированное антитело или его антигенсвязывающий фрагмент, специфическое(ий) для иммуноглобулина Е в соответствии с п.4, где эпитоп дополнительно включает остатки сахара GlcNAcl и Man6 в первой тяжелой цепи IgE и остаток сахара Man 5 во второй тяжелой цепи IgE.
6. Изолированное антитело или его антигенсвязывающий фрагмент по п.3, где указанное антитело или его антигенсвязывающий фрагмент специфично(ен) для иммуноглобулина Е и связывается с эпитопом в иммуноглобулине Е, включающим:
остатки Glu390, от Gln392 до Asn394, включительно, в первой тяжелой цепи IgE и Leu340, Arg342, от Аlа428 до Thr434, включительно, Thr436, Ser437 и Glu472 во второй тяжелой цепи IgE.
7. Изолированное антитело или его антигенсвязывающий фрагмент, специфическое(ий) для иммуноглобулина Е в соответствии с п.5, где эпитоп дополнительно включает остатки сахара GlcNAcl и Man6 в первой тяжелой цепи IgE.
8. Антитело по любому из пп.1, 2 и 3-7, где связывающий элемент представляет собой моноклональное антитело.
9. Изолированная молекула нуклеиновой кислоты, кодирующая изолированное антитело или его антигенсвязывающий фрагмент по любому из пп.1, 2 и 3-8.
10. Хозяйская клетка, трансформированная молекулой нуклеиновой кислоты в соответствии с п.9 для получения антитела или его антигенсвязывающего фрагмента по пп.1, 2 и 3-8.
11. Способ получения изолированного антитела или его антигенсвязывающего фрагмента по любому из пп.1, 2 и 3-8 для лечения расстройства, связанного с IgE, включающий культивирование хозяйских клеток по п.10 при условиях, подходящих для продукции указанного антитела или его антигенсвязывающего фрагмента.
12. Фармацевтическая композиция для лечения расстройства, ассоциированного с IgE, включающая антитело или его антигенсвязывающий фрагмент по любому из пп.1, 2 и 3-8 и фармацевтически приемлемый наполнитель.
13. Фармацевтическая композиция по п.12, включающая:
антитело или его антигенсвязывающий фрагмент по пп.1, 2 и 3-8
20 мМ сукцината
105 мМ NaCl
10 мМ аргинина
рН 6,00.
14. Композиция по п.12 или 13 для применения в качестве лекарственного средства.
15. Применение композиции по п.14 для лечения расстройства, ассоциированного с IgE.
16. Применение композиции по п.12 или 13, где расстройство представляет собой одно или более из аллергии, астмы и бронхита.
17. Применение композиции по п.12 или 13, где расстройство представляет собой одно или более из аллергического ринита, аллергического контактного дерматита, атопического дерматита, анафилактической реакции, пищевой аллергии, крапивницы, воспалительного заболевания кишечника, эозинофильного гастроэнтерита, индуцированной лекарственными средствами сыпи, аллергической офтальмопатии, аллергического конъюнктивита, бронхиальной астмы, гиперчувствительности дыхательных путей, косметической аллергии, аллергии, индуцированной лекарственными средствами, синдрома гиперчувствительности, индуцированного лекарственными средствами, аллергии на металлы, профессионального гиперчувствительного пневмонита, хронического гиперчувствительного пневмонита, холодовой гиперчувствительности, гиперчувствительности, индуцированной гельминтами, аллергии на латекс и аллергии на пыльцу.
| WO 2006045049, 27.04.2006 | |||
| RU 2000102666, 30.06.1998 | |||
| Устройство для возведения земляных сооружений намывным способом | 1936 |
|
SU50460A1 |

