ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к модифицированным Fc-рецепторам IgE и к их применениям.
УРОВЕНЬ ТЕХНИКИ
Аллергические заболевания, такие как аллергический ринит, атопический дерматит и пищевая аллергия, включая астму, распространяются с высокой скоростью в индустриализированных и европизированных современных обществах, и также возрастает частота развития анафилаксии, тяжелого аллергического заболевания. Эти хронические иммунные заболевания значительно ухудшают качество жизни индивидуумов и социально-экономические затраты растут соответствующим образом. Таким образом, существует острая потребность в мерах по преодолению таких заболеваний.
Большинство аллергических заболеваний вызываются чрезмерным иммунным ответом вследствие иммуноглобулина E (IgE). IgE представляет собой антитело, которое присутствует в сыворотке в очень низкой концентрации в нормальных условиях. IgE также обычно продуцируется в результате воздействия безопасных антигенов. Существует случай, когда уровень IgE возрастает без какого-либо конкретного стимула. Такой случай может приводить к аллергическим заболеваниям. Аномально увеличенное количество IgE может связываться с высокоаффинными Fc-рецепторами IgE (FcεRI), которые экспрессируются на поверхности тучных клеток, базофилов и т.п.
Такое связывание вызывает высвобождение тучными клетками или базофилами химических медиаторов, таких как гистамин, лейкотриен, простагландин, брадикинин и активирующие тромбоциты факторы. Высвобождение этих химических медиаторов приводит к аллергическим симптомам. В частности, аллергические заболевания могут демонстрировать более тяжелые симптомы вследствие связывания между IgE и FcεRI. Известно, что уровень FcεRI-эспрессирующих клеток возрастает у пациентов с аллергией.
В настоящее время для лечения аллергических заболеваний предложены различные способы, такие как избегание аллергена, введение противоаллергических средств, модулирование синтеза IgE в организме и разработка антител против IgE. Однако терапевтические способы, известные на настоящий момент, имеют множество недостатков, таких как неспособность вылечить основную причину аллергии, недостаточную эффективность лекарственных средств и возникновение серьезных побочных эффектов.
Кроме того, были исследованы композиции иммуноглобулинов, способные связываться с IgE и FcγRIIb с высокой аффинностью и ингибировать клетки, экспрессирующие IgE (KR10-1783272B1). Описано, что такие композиции являются пригодными для лечения IgE-опосредуемых нарушений, включая аллергию и астму. Кроме того, омализумаб (торговое наименование: Xolair), который нацелен на Fc-часть IgE-антитела, разработан и используется в качестве терапевтического средства против не поддающейся лечению тяжелой астмы и не поддающейся лечению крапивницы.
Однако введение омализумаба в высокой дозе для поддержания терапевтических эффектов приводит к высокой стоимости, и побочным эффектам, таким как ангионевротический отек и анафилактическая реакция (The Journal of Clinical Investigation Volume 99, Number 5, March 1997, 915-925). Кроме того, на основании результатов постмаркетингового исследования, был описан аллергический гранулематозный васкулит и идиопатическая тяжелая тромбоцитопения. Таким образом, существует растущая необходимость в разработке терапевтических средств, способных эффективно лечит аллергические заболевания без побочных эффектов.
Хотя основной механизм, посредством которого омализумаб вызывает побочные эффекты, еще не идентифицирован, можно ожидать, что он вызван тем, что омализумаб представляет собой IgG1-антитело. Исследование в модели на мышах показало, что большое количество антигенспецифических IgG1-антител могут индуцировать пассивную системную анафилаксию (PSA) через FcγRIII, низкоаффинный рецептор IgG, и активирующие тромбоциты факторы в богатой антигенами среде. Кроме того, такая IgG-опосредуемая анафилаксия возникает в чрезмерной степени вследствие отсутствия передачи сигнала FcεRI. Таким образом, пассивная системная анафилаксия может возникать в случае, когда значительное количество омализумаба инъецируют пациенту с аллергическим заболеванием, который демонстрирует высокий уровень IgE. Недавно было описано, что FcγRIIA, низкоаффинный рецептор IgG, экспрессируемый у человека, также ассоциирован с IgG-опосредуемой анафилаксией. Кроме того, постмаркетинговые исследования показали аномальные реакции, такие как аллергический гранулематозный васкулит и идиопатическая тяжелая тромбоцитопения.
Техническая проблема
Задачей настоящего изобретения является предоставление полипептидного димерного белка для лечения IgE-опосредуемого аллергического заболевания. Другой задачей настоящего изобретения является предоставление молекулы нуклеиновой кислоты, кодирующей белок, экспрессирующего вектора, содержащего эту молекулу нуклеиновой кислоты, и клетки-хозяина, содержащего экспрессирующий вектор. Другой задачей настоящего изобретения является предоставление способа получения полипептидного димера.
Решение проблемы
Для достижения описанных выше задач предусматривается полипептидный димер, содержащий два мономера, каждый из которых содержит внеклеточный домен альфа-субъединицы Fc-рецептора IgE. Мономер содержит модифицированную Fc-область и модифицированная Fc-область и внеклеточный домен альфа-субъединицы Fc-рецептора IgE связаны через шарнирную область IgD-антитела. В другом аспекте предусматривается фармацевтическая композиция для лечения или предупреждения аллергического заболевания, содержащая полипептидый димер в качестве активного ингредиента.
Преимущественные эффекты изобретения
Полипептидный димерный белок в соответствии с настоящим изобретением не только обладает превосходной безопасностью и нахождением в организме по сравнению с традиционно используемыми антителами против IgE, но также очень сильно связывается с IgE вследствие наличия способности связываться с IgE, которая в 70 раз выше, чем у традиционно используемого антитела против IgE омализумаба, что позволяет более длительный курс введения. Кроме того, полипептидный димерный белок в соответствии с настоящим изобретением представляет собой вещество, полученное с использованием модифицированного Fc, который имеет только IgE в качестве единственной мишени и не связывается с рецептором Fc-гамма, и, таким образом, лишен функций антителозависимой клеточной цитотоксичности (ADCC) и комплементзависимой цитотоксичности (CDC). Таким образом, в отличие от общепринятых антител против IgE, содержащих Fc-область IgG1, полипептидный димерный белок не связывается с рецептором Fc-гамма и, таким образом, может ингибировать высвобождение медиаторов, вызываемое связыванием с рецептором Fc-гамма на поверхности тучных клеток, так что могут минимизироваться тяжелые побочные эффекты, такие как возникновение анафилаксии, которые могут вызываться связыванием между IgG1 и рецептором Fc-гамма III на тучных клетках. Таким образом, полипептидный димерный белок в соответствии с настоящим изобретением можно использовать в качестве новой фармацевтической композиции, которая может заменить терапевтические средства, содержащие общепринятое антитело против IgE.
Краткое описание чертежей
На фиг.1 представлены результаты SDS-PAGE и изоэлектрического фокусирования на геле (IEF) для белков, продуцированных в каждой клеточной линии. В данном случае, можно видеть, что укороченная форма не образовывалась ни в восстанавливающих, ни в не восстанавливающих условиях, и что содержание кислых белков возрастает вследствие повышения содержания сиаловых кислот, вызванного введением гена трансферазы сиаловых кислот.
На фиг.2 проиллюстрированы результаты SDS-PAGE для не восстановленных и восстановленных форм полипептидного димерного белка в соответствии с вариантом осуществления настоящего изобретения. В частности, можно видеть, что полипептидный димер обладает высокой чистотой даже в культуральном супернатанте, который соответствует материалу на входе.
На фиг.3 проиллюстрирован график, демонстрирующий способность связывания омализумаба с IgE. На графике показаны результаты, полученные путем иммобилизации омализумаба и анализа его связывающей способности в зависимости от использованных концентраций IgE.
На фиг.4 проиллюстрирован график, демонстрирующий способность связывания с IgE полипептидного димерного белка в соответствии с одним вариантом осуществления настоящего изобретения. На графике показаны результаты, полученные путем иммобилизации димерного белка и анализа его способности к связыванию в зависимости от используемых концентраций IgE.
На фиг.5 проиллюстрированы результаты, полученные путем идентификации взаимодействий полипептидного димерного белка (IgETRAP), являющегося вариантом осуществления настоящего изобретения, и омализумаба с IgG-рецепторами FcγRI (фиг.5А), FcγRIIA (фиг.5B), FcγRIIB (фиг.5C), FcγRIIIA (фиг.5D) и FcγRIIIB (фиг.5E) с использованием анализа на основе биослойной интерферометрии (BLI).
На фиг.6 проиллюстрирован график, полученный путем количественного определения связывающей способности между IgETRAP и рецепторами IgG, и между омализумабом и рецепторами IgG.
На фиг.7 проиллюстрирован график, демонстрирующий ингибиторную способность в отношении активности происходящих из мыши тучных клеток полипептидного димерного белка (IgETRAP) в соответствии с вариантом осуществления настоящего изобретения в зависимости от его концентраций.
На фиг.8 проиллюстрирован график, демонстрирующий сравнение между ингибиторной способностью в отношении активности экспрессирующих FcεRI человека происходящих из мыши тучных клеток полипептидного димерного белка (IgETRAP) согласно варианту осуществления настоящего изобретения и Xolair (омализумаб) в зависимости от их концентрации.
На фиг.9 проиллюстрирован график, демонстрирующий эффекты введения полипептидного димерного белка в соответствии с одним вариантом осуществления настоящего изобретения в модели пищевой аллергии.
Подробное описание изобретения
Настоящее изобретение относится к полипептидному димеру, содержащему два мономера, каждый из которых содержит внеклеточный домен (FcεRIa-ECD) альфа-субъединицы Fc-рецептора IgE, где мономер содержит модифицированную Fc-область и модифицированная Fc-область и FcεRIa-ECD связаны через шарнирную область IgD-антитела.
Как используют в рамках изобретения, термин "IgE" означает антительный белок, известный как иммуноглобулин E. IgE обладает аффинностью к тучным клеткам, базофилам крови и т.п. Кроме того, реакция между IgE-антителом и антигеном (аллергеном), соответствующим ему, вызывает воспалительную реакцию. Кроме того, известно, что IgE является антителом, которое вызывает анафилаксию.
Как используют в рамках изобретения, термин "Fc-рецептор IgE" также называют Fcε-рецептором, и он связывается с Fc-частью IgE. Существует два типа этого рецептора. Рецептор, обладающий высокой аффинностью к Fc IgE, называют Fcε-рецептором I (FcεRI). Рецептор, обладающий низкой аффинностью к Fc IgE, называют Fcε-рецептором II (FcεRII). FcεRI экспрессируется в тучных клетках и базофилах. В случае, когда IgE-антитела, связанные с FcεRI, связываются поливалентными антигенами, происходит дегрануляция тучных клеток или базофилов, тем самым высвобождая различные химические вещества-переносчики, включая гистамин. Это высвобождение приводит к немедленной аллергической реакции.
FcεRI представляет собой мембранный белок, состоящий из одной α-цепи, одной β-цепи и двух γ-цепей, связанных дисульфидной связью. Среди этих цепей часть, с которой связывается IgE, представляет собой α-цепь (FcεRIα). FcεRIα имеет размер приблизительно 60 кДа и состоит из гидрофобного домена, существующего внутри клеточной мембраны, и гидрофильного домена, существующего снаружи клеточной мембраны. В частности, IgE связывается с внеклеточным доменом α-цепи.
В частности, альфа-субъединица Fc-рецептора IgE может иметь аминокислотную последовательность, указанную в NP_001992.1. Кроме того, внеклеточный домен (FcεRIa-ECD) альфа-субъединицы Fc-рецептора IgE может иметь аминокислотную последовательность SEQ ID NO: 1. В настоящей заявке внеклеточный домен альфа-субъединицы Fc-рецептора IgE может представлять собой фрагмент или вариант внеклеточного домена альфа-субъединицы Fc-рецептора IgE при условии, что фрагмент или вариант способен связываться с IgE.
Вариант можно получать способом замены, делеции или вставки одного или нескольких белков в FcεRIa-ECD дикого типа (внеклеточный домен) при условии, что способ не изменяет функцию α-цепи FcεRI. Такие различные белки или пептиды могут быть на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичными аминокислотной последовательности SEQ ID NO: 1. Кроме того, FcεRia-ECD SEQ ID NO: 1 может кодироваться полинуклеотидом, имеющим последовательность SEQ ID NO: 5.
Кроме того, как используют в рамках изобретения, термин "модифицированная Fc-область" означает область, в которой часть Fc-области антитела модифицирована. В рамках настоящей заявки термин "Fc-область" относится к белку, который содержит константную область тяжелой цепи 2 (CH2) и константную область тяжелой цепи 3 (CH3) иммуноглобулина, и не содержит вариабельные области тяжелой и легкой цепей и константную область легкой цепи 1 (CH1) иммуноглобулина. В частности, модифицированная Fc-область означает область, полученную путем замены некоторых аминокислот в Fc-области или путем комбинирования различных типов Fc-областей. В частности, модифицированная Fc-область может иметь аминокислотную последовательность SEQ ID NO: 2. Кроме того, модифицированная Fc-область SEQ ID NO: 2 может кодироваться полинуклеотидом, имеющим последовательность SEQ ID NO: 6.
Кроме того, модифицированная Fc-область по настоящему изобретению может иметь форму, имеющую цепи сахаров в нативной форме, увеличенное количество цепей сахаров относительно нативной формы или уменьшенное количество цепей сахаров относительно нативной форме, или она может быть в форме, из которой удалена цепь сахаров. Цепи сахаров Fc иммуноглобулинов можно модифицировать общепринятыми способами, такими как химические способы, ферментативные способы и способы генной инженерии с использованием микроорганизмов.
В рамках настоящей заявки "модифицированная Fc-область" по настоящему изобретению может представлять собой область, которая лишена функций антителозависимой клеточной цитотоксичности (ADCC) и комплементзависимой цитотоксичности (CDC) вследствие отсутствия участков связывания для FcγR или C1q. Кроме того, модифицированная Fc-область и FcεRIα-ECD могут быть связаны через шарнирную область IgD-антитела. Шарнирная область IgD-антитела состоит из 64 аминокислот и может селективно содержать 20-60 последовательно расположенных аминокислот, 25-50 последовательно расположенных аминокислот, или 30-40 аминокислот. В одном варианте осуществления шарнирная область IgD-антитела может содержать 30 или 49 аминокислот, как показано ниже. Кроме того, шарнирная область IgD-антитела может представлять собой вариант шарнирной области, полученный посредством модификации шарнирной области, где шарнирная область может содержать по меньшей мере один остаток цистеина. В рамках настоящего изобретения вариант шарнирной области может быть получен путем модификации некоторых остатков в последовательности шарнирной области IgD-антитела для минимизации образования укороченных форм в ходе процесса продуцирования белка.
В одном варианте осуществления шарнирная область может содержать следующую последовательность:
Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Xaa1 Xaa2 Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro (SEQ ID NO: 17), где Xaa1 может представлять собой Lys или Gly, и Xaa2 может представлять собой Glu, Gly или Ser. В частности, шарнирная область может иметь аминокислотную последовательность SEQ ID NO: 3 или SEQ ID NO: 19, тем самым минимизируя образование укороченных форм в процессе продуцирования белка.
В другом варианте осуществления шарнирная область может содержать следующую последовательность:
Ala Gln Pro Gln Ala Glu Gly Ser Leu Ala Lys Ala Thr Thr Ala Pro Ala Thr Thr Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Xaa3 Xaa4 Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro (SEQ ID NO: 18), где Xaa3 может представлять собой Lys или Gly, и Xaa4 может представлять собой Glu, Gly или Ser. В частности, шарнирная область может иметь аминокислотную последовательность SEQ ID NO: 4, тем самым минимизируя образование укороченных форм в ходе продуцирования белка.
В частности, в шарнирной области, имеющей последовательность SEQ ID NO: 4, по меньшей мере один из Thr может быть гликозилирован. В частности, среди аминокислот SEQ ID NO: 18, 13-й, 14-й, 18-й или 19-й Thr могут быть гликозилированными. Предпочтительно, все четыре аминокислоты могут быть гликозилированными. В рамках настоящей заявки гликозилирование может представлять собой O-гликозилирование.
Кроме того, как описано выше, полипептидный димер в соответствии с настоящим изобретением может иметь форму, в которой два мономера связаны друг с другом и каждый мономер получен путем связывания между внеклеточным доменом альфа-субъединицы Fc-рецептора IgE и модифицированной Fc-областью. Полипептидный димер может иметь форму, в которой два одинаковых мономера связаны друг с другом посредством цистеина, находящегося в линкерной области. Кроме того, полипептидный димер может иметь форму, в которой два различных мономера связаны друг с другом. Например, в случае, когда два мономера отличаются друг от друга, полипептидный димер может иметь форму, в которой один мономер содержит внеклеточный домен альфа-субъединицы Fc-рецептора IgE, а другой мономер содержит фрагмент внеклеточного домена альфа-субъединицы Fc-рецептора IgE. В рамках настоящей заявки вариант осуществления мономера может иметь аминокислотную последовательность SEQ ID NO: 20, SEQ ID NO: 21 или SEQ ID NO: 22.
Кроме того, полипептидный димер в соответствии с настоящим изобретением демонстрирует способность связываться с IgE, которая в 10-100 раз, в 20-90 раз, в 20-70 раз, в 30-70 раз или в 40-70 раз выше, чем у омализумаба, антитела против IgE, и предпочтительно он может демонстрировать способность связываться с IgE, которая в 70 раз выше, чем у омализумаба.
В другом аспекте изобретения предусматривается полинуклеотид, кодирующий мономер, который содержит внеклеточный домен альфа-субъединицы Fc-рецептора IgE, с которым связана модифицированная Fc-область.
Между тем, полинуклеотид, кроме того, может содержать сигнальную последовательность или лидерную последовательность. Как используют в рамках изобретения, термин "сигнальная последовательность" означает нуклеиновую кислоту, кодирующую сигнальный пептид, который направляет секрецию белка-мишени. Сигнальный пептид транслируется, а затем отщепляется в клетке-хозяине. В частности, сигнальная последовательность по настоящему изобретению представляет собой нуклеотид, кодирующий аминокислотную последовательность, которая инициирует перемещение белка через мембрану эндоплазматической сети (ER). Пригодные в рамках настоящего изобретения сигнальные последовательности включают последовательности легких цепей антител, например, антитела 14.18 (Gillies et al., J. Immunol. Meth 1989. 125:191-202), сигнальные последовательности тяжелых цепей антител, например, сигнальную последовательность тяжелой цепи антитела MOPC141 (Sakano et al., Nature, 1980. 286:676-683), и другие сигнальные последовательности, известные в данной области (см., например, Watson et al., Nucleic Acid Research, 1984. 12:5145-5164).
Характеристики сигнальных последовательностей хорошо известны в данной области. Сигнальные последовательности, как правило, содержат 16-30 аминокислотных остатков, и могут содержать больше или меньше аминокислотных остатков. Типичный сигнальный пептид состоит из трех областей, которые представляют собой основную N-концевую область, центральную гидрофобную область и более полярную C-концевую область. Центральная гидрофобная область содержит 4-12 гидрофобных остатков, которые иммобилизуют сигнальную последовательность на липидном бислое мембраны в ходе перемещения незрелого полипептида.
После инициации сигнальная последовательность отщепляется в просвете ER клеточными ферментами, широко известными как сигнальные пептидазы. В рамках настоящего изобретения сигнальная последовательность может представлять собой секреторную сигнальную последовательность тканевого активатора плазминогена (tPA), гликопротеина D вируса простого герпеса (HSV gD), или гормона роста. Предпочтительно, можно использовать секреторные сигнальные последовательности, используемые в клетках высших эукариот, включая млекопитающих и т.п. Кроме того, может использоваться сигнальная последовательность, кодоны в которой замещены на кодоны, имеющие высокую частоту экспрессии в клетке-хозяине.
Между тем, внеклеточный домен мономер альфа-субъединицы Fc-рецептора IgE, с которым связана сигнальная последовательность и модифицированная Fc-область, может иметь аминокислотную последовательность SEQ ID NO: 11 или SEQ ID NO: 13. Белки SEQ ID NO: 11 и SEQ ID NO: 13 могут кодироваться полинуклеотидами, имеющими последовательности SEQ ID NO: 12 и SEQ ID NO: 14, соответственно.
В другом аспекте настоящего изобретения предусматривается экспрессирующий вектор, в который встроен полинуклеотид, кодирующий мономер. В рамках настоящего изобретения полинуклеотид может иметь последовательность SEQ ID NO: 12 или SEQ ID NO: 14.
Как используют в рамках изобретения, термин "вектор" предполагает структуру, предназначенную для введения в клетку-хозяина и способную рекомбинировать с геномом клетки-хозяина и встраиваться в него. Альтернативно вектор представляет собой эписому, и его понимают как элемент нуклеиновой кислоты, содержащий нуклеотидную последовательность, которая может автономно реплицироваться. Векторы включают линейные нуклеиновые кислоты, плазмиды, фагмиды, космиды, РНК-векторы, вирусные векторы и их аналоги. Примеры вирусных векторов включают, но не ограничиваются ими, ретровирусы, аденовирусы и аденоассоциированные вирусы. Кроме того, плазмиды могут содержать селективный маркер, такой как ген резистентности к антибиотикам, и клетки-хозяева, содержащие плазмиды, можно культивировать в селективных условиях.
Как используют в рамках изобретения, под термином "генетическая экспрессия" или "экспрессия" белка-мишени подразумевают транскрипцию последовательности ДНК, трансляцию мРНК-транскрипта и секрецию слитого белкового продукта или его фрагмента. Пригодным экспрессирующим вектором может быть RcCMV (Invitrogen, Carlsbad) или его вариант. Экспрессирующий вектор может содержать промотор цитомегаловируса человека (CMV) для обеспечения непрерывной транскрипции гена-мишени в клетках млекопитающих, и сигнальную последовательность поладенилирования бычьего гормона роста для повышения уровня стабильности РНК после транскрипции.
В следующем аспекте настоящего изобретения предусматривается клетка-хозяин, в которую введен экспрессирующий вектор. Как используют в рамках изобретения, термин "клетка-хозяин" относится к прокариотической или эукариотической клетке, в которую может быть введен рекомбинантный экспрессирующий вектор. Как используют в рамках изобретения, термины "трансдуцированный", "трансформированный" и "трансфицированный" означает введение нуклеиновой кислоты (например, вектора) в клетку с использованием способов, известных в данной области.
Предпочтительные клетки-хозяева, которые можно использовать в рамках настоящего изобретения, включают иммортализованные гибридомные клетки, клетки миеломы NS/0, клетки 293, клетки яичника китайского хомячка (клетки CHO), клетки HeLa, клетки человека, происходящие из амниотической жидкости (клетки CapT), или клетки COS. Предпочтительно, клетки-хозяева могут представлять собой клетки CHO. С другой стороны, клетка-хозяин может представлять собой клетку, в которую введен указанный вектор и вектор, нагруженный геном трансферазы сиаловых кислот. В рамках настоящего изобретения трансфераза сиаловых кислот может представлять собой трансферазу 2,3-сиаловой кислоты или трансферазу 2,6-сиаловой кислоты. В рамках настоящего изобретения трансфераза 2,6-сиаловой кислоты может иметь аминокислотную последовательность SEQ ID NO: 15.
В другом аспекте настоящего изобретения предусматривается фармацевтическая композиция для лечения или предупреждения аллергического заболевания, содержащая полипептидный димер в качестве активного ингредиента.
В настоящем описании термин "аллергическое заболевание" означает патологический симптом, вызванный аллергической реакцией, опосредуемой активацией тучных клеток, такой как дегрануляция тучных клеток. Такие аллергические заболевания включают пищевую аллергию, атопический дерматит, астму, аллергический ринит, аллергический конъюнктивит, аллергический дерматит, аллергический контактный дерматит, анафилаксию, крапивницу, зуд, аллергию на укусы насекомых, хроническую идиопатическую крапивницу, аллергию на лекарственные средства и т.п. В частности, аллергические заболевания могут быть IgE-опосредуемыми.
В композиции для лечения или предупреждения аллергического заболевания по настоящему изобретению, активный ингредиент может содержаться в любом количестве (эффективное количество) в зависимости от применения, состава, цели смешивания и т.п., при условии, что активный ингредиент может демонстрировать противоаллергическую активность. Типичное эффективное количество активного ингредиента находится в диапазоне от 0,001% по массе до 20,0% по массе в расчете на общую массу композиции. В настоящем описании "эффективное количество" относится к количеству активного ингредиента, которое способно индуцировать антиаллергический эффект. Такое эффективное количество может определить экспериментально специалист в данной области.
В рамках настоящего изобретения фармацевтическая композиция, кроме того, может содержать фармацевтически приемлемый носитель. В качестве фармацевтически приемлемого носителя может использоваться любой носитель при условии, что носитель представляет собой нетоксичное вещество, пригодное для доставки пациенту. В качестве носителей могут содержаться дистиллированная вода, спирт, жир, воск и инертное твердое вещество. Также в фармацевтической композиции могут содержаться фармацевтически приемлемые адъюванты (буферы и диспергирующие вещества).
В частности, фармацевтическая композиция по настоящему изобретению содержит, в дополнение к активному ингредиенту, фармацевтически приемлемый носитель, и она может быть изготовлена в форме перорального или парентерального состава в зависимости от пути введения общепринятым способом, известным в данной области. В рамках настоящей заявки, термин "фармацевтически приемлемый" означает имеющий не более высокую токсичность, чем может переноситься индивидуумом, который будет применять (которому будет назначено) средство, без ингибирования активности активного ингредиента.
В случае, когда фармацевтическую композицию по настоящему изобретению изготавливают в форме перорального состава, фармацевтическая композиция может быть изготовлена в форме составов, таких как порошки, гранулы, таблетки, пилюли, таблетки с сахарным покрытием, капсулы, жидкости, гели, сиропы, суспензии и вафли, вместе с подходящими носителями, в соответствии со способами, известными в данной области. В рамках настоящего изобретения, примеры подходящих фармацевтически приемлемых носителей могут включать сахара, такие как лактоза, глюкоза, сахароза, декстроза, сорбит, маннит и ксилит, крахмалы, такие как кукурузный крахмал, картофельный крахмал и пшеничный крахмал, целлюлозы, такие как целлюлоза, метилцеллюлоза, этилцеллюлоза, натрий карбоксиметилцеллюлоза, и гидроксипропилметилцеллюлоза, поливинилпирролидон, вода, метилгидроксибензоат, пропилгидроксибензоат, стеарат магния, минеральное масло, солод, желатин, тальк, полиол, растительное масло и т.п. В случае изготовления в виде препаратов, получение можно проводить, при необходимости, путем включения разбавителей и/или эксципиентов, таких как наполнитель, сухой разбавитель, связующее вещество, смачивающее вещество, разрыхлитель и поверхностно-активное вещество.
В случае, когда фармацевтическая композиция по настоящему изобретению изготовлена в виде парентерального состава, фармацевтическая композиция может быть изготовлена в виде препаратов в форме инъекций, трансдермальных лекарственных средств, назальных ингаляторов и суппозиториев, вместе с подходящими носителями, в соответствии со способами, известными в данной области. В случае получения в виде инъекций, в качестве подходящего носителя можно использовать стерилизованную воду, этанол, полиол, такой как глицерин и пропиленгликоль, или их смесь. В качестве носителя предпочтительно можно использовать изотонические растворы, такие как раствор Рингера, фосфатно-солевой буфер (PBS), содержащий триэтаноламин, стерильную воду для инъекций и 5% декстрозу, и т.п.
Получение фармацевтической композиции известно в данной области, и, в частности, может быть упомянут Remington's Pharmaceutical Sciences (19th ed., 1995) и т.п. Этот документ считается частью настоящего описания.
Предпочтительная суточная дозировка фармацевтической композиции по настоящему изобретению находится в диапазоне от 0,01 мкг/кг до 10 г/кг, и предпочтительно от 0,01 мг/кг до 1 г/кг, в зависимости от состояния пациента, массы тела, пола, возраста, тяжести заболевания или пути введения. Введение можно проводить один раз или несколько раз в сутки. Такую дозировку никоим образом не следует интерпретировать как ограничивающую объем настоящего изобретения.
Индивидуумом, у которого может применяться (может быть назначена) композиция по настоящему изобретению, является млекопитающее и человек, причем особенно предпочтительным является человек. Композиция против аллергии по настоящему изобретению, кроме того, может содержать, в дополнение к активному ингредиенту, любое соединение или природный экстракт, для которого уже подтверждена безопасность и для которого известно, что оно обладает противоаллергической активностью, для повышения и усиления противоаллергической активности.
В другом аспекте настоящего изобретения предусматривается пищевая композиция для смягчения или облегчения симптома аллергии, содержащая в качестве активного ингредиента полипептидный димер.
В рамках настоящего изобретения полипептидный димер может быть связан с соответствующей доставляющей структурой для эффективной доставки в кишечник. Пищевая композиция по настоящему изобретению может быть изготовлена в любой форме и может быть получена, например, в форме напитков, таких как чай, сок, газированные напитки и ионные напитки, переработанные молочные продукты, такие как молоко и йогурт, диетические функциональные пищевые препараты, такие как таблетки, капсулы, пилюли, гранулы, жидкости, порошки, хлопья, пасты, сиропы, гели, желе и батончики, и т.п. Кроме того, пищевая композиция по настоящему изобретению может относиться к любой категории продуктов по официальной или функциональной классификации при условии, что пищевая композиция удовлетворяет предписаниям контролирующих органов на момент производства и распространения. Например, пищевая композиция может представлять собой диетический функциональный продукт питания в соответствии с Актом о функциональной диетической продукции, или может относиться к кондитерским изделиям, продуктам на основе бобовых, чаям, напиткам, продуктам питания специального назначения и т.п. в соответствии с каждым типом продуктов питания в Кодом продуктов питания Акта о пищевой санитарии (стандарты и спецификации для продуктов писания, зарегистрированные в Управление продовольствия и лекарственных препаратов). Что касается других пищевых добавок, которые могут содержаться в пищевой композиции по настоящему изобретению, может быть упомянут Код продуктов питания или Дополнительный код продуктов питания в соответствии с Актом о пищевой санитарии.
В другом аспекте настоящего изобретения предусматривается способ получения полипептидного димера, включающий стадию культивирования клетки-хозяина, в которую введен полинуклеотид, кодирующий мономер, и ген трансферазы сиаловых кислот; и стадию выделения полипептидного димера.
В рамках настоящего изобретения полинуклеотид, кодирующий мономер, может быть введен в клетку-хозяина будучи нагруженным на экспрессирующий вектор. Кроме того, ген трансферазы сиаловых кислот может быть введен в клетку-хозяина будучи нагруженным на вектор.
Сначала проводят стадию введения в клетку-хозяина вектора, в который встроен полинуклеотид, кодирующий мономер, и вектора, в который встроен ген трансферазы сиаловых кислот. В рамках настоязего изобретения трансфераза сиаловых кислот может представлять собой трансферазу 2,3-сиаловой кислоты или трансферазу 2,6-сиаловой кислоты.
Далее проводят стадию культивирования трансформированной клетки.
Наконец, проводят стадию выделения полипептидного димера. В рамках настоящей заявки полипептидный димер может быть очищен из культуральной среды или клеточного экстракта. Например, после получения супернатанта культуральной среды, в которой секретировался полипептидный димер, супернатант можно концентрировать с использованием коммерчески доступного фильтра для концентрирования белка, например, элемента для ультрафильтрации Amicon или Millipore Pellicon. Затем концентрат можно очищать способами, известными в данной области. Например, очистку можно проводить с использованием матрикса, связанного с белком A.
В другом аспекте настоящего изобретения предусматривается полипептидный димер, продуцированный описанным выше способом получения димера.
В рамках настоящего изобретения полипептидный димер имеет высокое содержание сиаловой кислоты и, таким образом, имеет очень высокую кислотность относительно теоретической величины pI.
В следующем аспекте настоящего изобретения предусматривается фармацевтическая композиция для лечения или предупреждения аллергического заболевания, содержащая в качестве активного ингредиента полипептидный димер, полученный описанным выше способом получения димера.
В другом аспекте настоящего изобретения предусматривается пищевая композиция для смягчения или облегчения симптома аллергии, содержащая в качестве активного ингредиента полипептидгый димер, полученный описанным выше способом получения димера.
В следующем аспекте настоящего изобретения предусматривается способ лечения или предупреждения аллергического заболевания, включающий стадию введения индивидууму полипептидного димера, который содержит два мономера, каждый из которых содержит внеклеточный домен (FcεRIa-ECD) альфа-субъединицы Fc-рецептора IgE.
Индивидуумом может быть млекопитающее, предпочтительно человека. В рамках настоящего изобретения введение можно проводить перорально или парентерально. В рамках настоящего изобретения парентеральное введение можно проводить такими способами, как подкожное введение, внутривенное введение, мукозальное введение и мышечное введение.
Далее настоящее изобретение описано более подробно с помощью следующих примеров. Однако следующие примеры предназначены только для иллюстрации настоящего изобретения и объем настоящего изобретения не ограничивается только ими.
Пример 1. Получение полипептида, содержащего FcεRIα-ECD и модифицированную Fc-область
C-концевой модифицированный полипептид внеклеточного домена (FcεRIα-ECD) альфа-субъединицы Fc-рецептора IgE получали способом, описанным в патенте IgE США № 7867491.
Сначала для экспрессии белка (FcεRIαECD-Fc1), белка (FcεRIαECD-Fc2) и белка (FcεR1αECD-Fc3), в котором внеклеточный домен α-цепи FcεRI, имеющий аминокислотную последовательность SEQ ID NO: 1 и модифицированный Fc иммуноглобулин SEQ ID NO: 2 связаны через шарнирную область SEQ ID NO: 19, шарнирную область SEQ ID NO: 3 и шарнирную область SEQ ID NO: 4, соответственно, кассеты, полученные путем связывания гена, кодирующего каждый белок, клонировали в векторы pAD15 (Genexin, Inc.), с конструкцией, экспрессирующей белок FcεRIαECD-Fc. Затем каждый из экспрессирующих векторов трансдуцировали в клетки CHO DG44 (от Dr. Chasin, Columbia University, США).
В данном случае, при трансдукции в клеточную линию экспрессирующий вектор, полученный посредством клонирования гена трансферазы α-2,6-сиаловой кислоты в вектор pCI Hygro (Invitrogen), одновременно трансдуцировали для получения отдельно клеточных линий, которые способны экспрессировать белки FcεRIαECD-Fc2ST и FcεRIα ECD-Fc3ST, к которым добавлена сиаловая кислота.
В качестве первичной скрининговой процедуры проводили селекцию с HT с использованием среды, свободной от 5-гидрокситриптамина (HT) с 10% dFBS (Gibco, США, 30067-334), среды MEMα (Gibco, 12561, США, каталожный номер № 12561-049) и среды HT+ (Gibco, США, 11067-030). Затем проводили амплификацию с метотрексатом (MTX) с использованием отобранных с помощью HT клонов для повышения продуктивности с использованием системы "дигидрофолатредуктаза (DHFR)-".
После завершения амплификации с MTX проводили субкультивирование приблизительно 1-5 раз для стабилизации клеток с целью оценки продуктивности. После этого проводили оценку продуктивности в единицах для амплифицированных с MTX клеток. Результаты представлены в таблице 1 ниже.
[Таблица 1]
Как показано в таблице 1, клеточная линия FcεRIαECD-Fc3 демонстриует продуктивность 16,9 мкг/106 клеток после амплификациии с метотрексатом в концентрации 2 мкМ. С другой стороны, клеточная линия FcεRIαECD-Fc3 (FcεRIαECD-Fc3ST), котрансдуцированная трансферазой 2,6-сиаловых кислот, демонстрировала продуктивность 17,5 мкг/106 клеток после амплификации с метотрексатом в концентрации 1 мкМ. Кроме того, клеточная линия FcεRIαECD-Fc2 демонстрировала продуктивность 20,9 мкг/106 клеток в условиях амплификации с метотрексатом в концентрации 0,5 мкМ. Кроме того, клеточная линия FcεRIαECD-Fc2 (FcεRIαECD-Fc2ST), котрансдуцированная трансферазой 2,6-сиаловых кислот, демонстрировала продуктивность 25,1 мкг/106 клеток после амплификации с метотрексатом в концентрации 0,1 мкМ. Следовательно, было идентифицировано, что клеточная линия FcεRIαECD-Fc2, котрансфицированная трансферазой 2,6-сиаловых кислот, которая была подвергнута селекции в условиях амплификации с метотрексатом в концентрации 0,1 мкМ, демонстрировала наилучшую продуктивность.
Пример 2. Очистка слитого белка FcεRIα ECD и определение его чистоты
Среди клеточных линий, отобранных согласно примеру 1 выше, i) FcεRIαECD-Fc3, ii) FcεRIαECD-Fc3ST и iii) FcεRIαECD-Fc2ST культивировали в масштабе 60 мл способом периодической культуры. Полученные культуры очищали с использованием аффинной колонки с белком A, а затем очищенные белки подвергали SDS-PAGE и эксклюзионной ВЭЖХ (SE-ВЭЖХ) для определения чистоты белков.
Как показано на фиг.1, было идентифицировано, что все соответствующие белки, очищенные способом SE-ВЭЖХ, имели чистоту 93% или выше. Кроме того, в результате анализа SDS-PAGE было идентифицировано, что белки размером приблизительно 150 кДа и приблизительно 75 кДа, выявлялись, соответственно, в не восстанавливающих и восстанавливающих условиях (фиг.1, дорожки 1-6). Исходя из этого было обнаружено, что связанный с Fc FcεRIαECD образует димер. Кроме того, в результатах SDS-PAGE не наблюдалось примесей, таких как укороченная форма. В частности, даже после процесса оттаивания/замораживания (фиг.1, дорожки 7-8), было идентифицировано, что все белки имеют чистоту 93% или выше и не имеют примесей. Из этого было обнаружено, что продуцирование укороченных белковых форм снижено относительно FcεRIαECD-Fc1, в котором использовалась шарнирная область IgD дикого типа.
В данном случае, гель-IEF проводили в приведенных ниже условиях тестирования для идентификации степени содержания сиаловых кислот в белках после введения трансферазы сиаловых кислот. Из этого было идентифицировано, что содержание кислых белков возрастало вследствие увеличения содержания сиаловых кислот.
[Таблица 2]
Для определения воспроизводимости выхода очистки клеточную линию FcεRIαECD-Fc2ST подвергали периодическому культивированию в 1-л колбе в масштабе 250 мл и очищали с использованием аффинной колонки с белком A. Затем культуральный супернатант и очищенный продукт подвергали разделению на 4%-15% геле TGXTM (Bio-Rad Laboratories, Inc.) в течение 30 минут в условиях буфера Tris-глицин SDS (TGS) и 200 В, а затем подвергали анализу SDS PAGE. В результате было идентифицировано что не только белки с очень высокой чистотой (98% или выше) очищались даже посредством только первой стадии очистки, но также белки экспрессировались с очень высокой чистотой даже в культуральном супернатанте. Это указывает на то, что стадии разработки процесса могут быть упрощены при разработке белка FcεRIαECD-Fc, который экспрессируется в рассматриваемой клеточной линии, в виде медицинского продукта, и в результате вероятно, что стоимость разработки медицинского продукта значительно снизится.
Пример 3. Определение способности связывания слитого белка FcεRIα ECD с IgE
Способность связывания с IgE подвергали сравнительному определению для четырех белков: i) FcεRIαECD-Fc2, ii) FcεRIαECD-Fc2ST, iii) FcεRIαECD-Fc3 и iv) FcεRIαECD-Fc3ST, которые были очищены способом согласно примеру 2 выше, и коммерчески доступного антитела против IgE омализумаба (торговое название: Xolair). В частности, способность связываться с IgE определяли путем нанесения IgE на канал сенсорного чипа Protein GLC (Bio-Rad Laboratories, Inc., каталожный номер № 176-5011) и позволения омализумабу или каждому белку FceR1αECD-Fc в различных концентрациях течь со скоростью 30 мкл в минуту.
Эксперименты проводили путем идентификации нулевого фона с использованием 25 мМ NaOH в качестве регенерирующего буфера, а затем повторения описанных выше стадий. После этого кривую связывания идентифицировали с использованием анализатора связывания белков (ProteOn XPR36, Bio-Rad Laboratories, Inc., США). Результаты представлены в таблице 3, и на фиг.3 и 4.
[Таблица 3]
Положения
Как показано в таблице 3, было определено, что величина скорости ассоциации (ka) полипептидного димера в соответствии с одним вариантом осуществления настоящего изобретения в 1,5-2,0 раза ниже, чем у омализумаба. Таким образом, было обнаружено, что его аффинность связывания с веществами, отличными от IgE, была в 1,5-2,0 ниже, чем у омализумаба. Кроме того, было определено, что величина скорости диссоциации (kd) полипептидного димера в соответствии с одним вариантом осуществления настоящего изобретения в 40-106 раз выше, чем у омализумаба. Кроме того, как показано на фиг.3 и 4, можно было идентифицировать, что омализумаб утрачивал свое связывание с IgE в случае, когда после связывания проходил определенный период времени, в то время как после связывания полипептидного димера слитого белка FcεRIαECD по настоящему изобретению с IgE полипептидный димер не отделялся от IgE.
Таким образом, можно видеть, что полипептидный димер по настоящему изобретению не легко отделяется от IgE, и он обладает значительно лучшей способностью сохранять его связанное состояние, чем омализумаб. В результате, было обнаружено, что полипептидный димер в соответствии с одним вариантом осуществления настоящего изобретения имеет равновесную величину константы диссоциации (KD <kd/ka>), которая в 22-69 раз выше, чем у омализумаба. Исходя из этого, можно видеть, что слитый белок FcεRIαECD по настоящему изобретению обладает значительно увеличенной способностью связываться с IgE по сравнению с омализумабом. В частности, было идентифицировано, что FcεRIαECD-Fc2 (FcεRIαECD-Fc2ST), к которому добавлена сиаловая кислота, демонстрирует наивысшую способность связывания IgE, которая была в 69 раз выше, чем у омализумаба.
Пример 4. Идентификации способности слитого белка FcεRIα ECD связываться с рецептором IgG
Степень связывания IgETRAP и омализумаба с рецепторами IgG определяли с использованием системы Octet RED384 (ForteBio, CA, США). Рекомбинантные белки FcγRI, FcγRIIA, FcγRIIB, FcγRIIIA и FcγRIIIB (R & D Systems Inc., 5 мкг/мл) иммобилизовывали в 300 мМ ацетатном буфере (pH 5) на активированном биосенсоре AR2G. В качестве подвижного буфера использовали PBS, содержавший 0,1% Tween-20 и 1% бычью сыворотку. Все эксперименты проводили при 30°C с использованием устройства для встряхивания планшетов со скоростью 1000 об./мин. Результаты представлены на фиг.5A-5E, и способность омализумаба и IgETRAP связываться с рецепторами IgG количественно определена и представлена на фиг.6.
Пример 5. Определение активности слитого белка FcεRIα ECD с использованием анализа с бета-гексозаминидазой в тучных клетках костномозгового происхождения мыши
Анализ с бета-гексозаминидазой проводили для исследования активности in vitro слитого бека FcεRIαECD по настоящему изобретению. В частности, белок FcεRIαECD-Fc2 в соответствии с одним вариантом осуществления настоящего изобретения смешивали в каждой концентрации с IgE мыши (1 мкг/мл) и инкубировали при комнатной температуре (20°C) в течение 30 минут с получением образцов. Тучные клетки костномозгового происхождения в культуре для активации тучных клеток промывали буфером на основе сбалансированного солевого раствора Хэнка (HBSS) для удаления среды, и определяли количество клеток. Затем проводили доведение таким образом, чтобы 5×105 клеток инжектировали в 40 мкл буфера HBSS.
Затем к активированным тучным клеткам добавляли 50 мкл раствора образца, полученного посредством предварительной инкубации. Затем полученный материал инкубировали в инкубаторе с 5% CO2 при 37°C в течение 30 минут. Затем после добавления в каждом случае по 10 мкл DNP (2,4-динитрофенол, 100 нг/мл), который является посторонним антигеном, вновь проводили инкубацию при 37°C в течение 30 минут в 5% CO2, а затем отделяли 30 мкл супернатанта. 30 мкл отделенного супернатанта и 30 мкл субстрата (4-нитрофенил N-ацетил-β-D-глюкозаминид, 5,84 мМ) хорошо перемешивали, а затем инкубировали при 37°C в течение 20 минут в 5% CO2. Затем добавляли 140 мкл 0,1 M натрий-карбонатного буфера (pH 10) в качестве стоп-раствора для завершения реакции. После этого определяли поглощение при 405 нм для идентификации секретируемого количества β-гексозаминидазы, секретируемой посредством постороннего антигена, в активированных тучных клетках. Результаты показаны на фиг.7.
Как показано на фиг.7, полипептидный димер согласно одному варианту осуществления настоящего изобретения демонстрировал долю ингибирования тучных клеток приблизительно 49,4% в случае половинной (0,5 мкг/мл) концентрации IgE мыши, и демонстрировал долю ингибирования тучных клеток приблизительно 99,4% в случае той же концентрации (1 мкг/мл) IgE мыши. Таким образом, можно видеть, что индуцируемая IgE активность происходящих из костного мозга тучных клеток значительно подавляется полипептидным димером FceRIa-ECD по настоящему изобретению.
Пример 6. Сравнение активности слитого белка FcεRIα ECD и антитела против IgE человека с использованием анализа с β-гексозаминидазой в экспрессирующих FcεRI человека тучных клетках костномозгового происхождения
Анализ с β-гексозаминидазой проводили для идентификации превосходства слитого белка FcεRIα ECD над Xolair посредством анализа активности in vitro. Соответствующие лекарственные средства, FcεRIαECD-Fc2ST (IgETRAP) и Xolair, получали в каждой концентрации, а затем смешивали с IgE человека (1 мкг/мл). Затем проводили инкубацию при комнатной температуре в течение 30 минут. В ходе предварительной инкубации с лекарственным средством вводили ген FcεRI человека и получали тучные клетки, происходящие и дифференцированные из костного мозга мыши, в которых ген FcεRI мыши удален. Полученные тучные клетки промывали буфером HBSS, а затем 5×105 клеток инжектировали в 60 мкл буфера HBSS. К полученным тучным клеткам добавляли 20 мкл предварительно инкубированного образца, а затем инкубировали в инкубаторе с 5% CO2 при 37°C в течение 30 минут.
Затем добавляли 20 мкл антитела против IgE человека (BioLegend, каталожный номер № 325502, 0,5 мкг/мл), а затем полученный материал вновь инкубировали в инкубаторе с 5% CO2 при 37°C в течение 30 минут. Затем после центрифугирования при 1500 об/мин при 4°C отделяли 30 мкл супернатанта. 30 мкл отделенного супернатанта и 30 мкл субстрата (4-нитрофенил N-ацетил-β-глюкозаминида, 5,84 мМ) хорошо перемешивали, а затем инкубировали в инкубаторе с 5% CO2 при 37°C в течение 25 минут. Затем добавляли 140 мкл 0,1 M натрий-карбонатного буфера (pH 10) для завершения реакции.
Затем определяли поглощение при 405 нм для сравнения относительных количеств секретируемой β-гексозаминидазы и идентифицировали эффект ингибирования тучных клеток в зависимости от каждой концентрации лекарственного средства. Результаты представлены на фиг.8. Как показано на фиг.8, было определено, что IC50 слитого белка FcεRIα ECD составляет приблизительно 11,16 нг/мл, и IC50 белка Xolair составляет приблизительно 649,8 нг/мл. Таким образом, было идентифицировано, что слитый белок FcεRIα ECD обладает в 58 раз более высокой ингибиторной активностью в отношении активности тучных клеток, чем Xolair.
Пример 7. Анализ in vivo слитого белка FcεRIα ECD (модель пищевой аллергии)
50 мкг овальбумина (OVA) и 1 мг квасцов внутрибрюшинно вводили мышам Balb/c (Orientbio Inc.) два раза с интервалом 14 суток для индукции сенсибилизации. После этого 50 мг OVA перорально вводили пять раз всего на 28, 30, 32, 34 и 36 сутки для индукции пищевой аллергии в кишечнике.
После перорального введения OVA два раза, т.е. на 31 сутки, мышей разделяли на три группы, каждая из которых содержала 7 мышей. Три разделенных группы были следующими: в первой группе вводили слитый белок FcεRIαECD-Fc2ST в высокой концентрации (200 мкг), во второй группе вводили слитый белок FcεRIαECD-Fc2ST в низкой концентрации (20 мкг), и в третьей группе ничего не вводили. При пероральном введении OVA определяли, происходит ли диарея в результате индукции пищевой аллергии. Мышей умерщвляли на 37 сутки и анализировали количество тучных клеток в тонком кишечнике, концентрацию IgE в крови и концентрацию фермента (протеаза-1 тучных клеток (MCPT-1)) с дегрануляцией тучных клеток в крови для мышей, относящихся к каждой группе.
Как показано на фиг.9, было идентифицировано, что мыши, относящиеся к группе, в которой вводили FcεRIαECD-Fc2ST, который представляет собой полипептидный димер, в высокой концентрации демонстрирует эффект смягчения пищевой аллергии зависимым от концентрации образом по сравнению с мышами, относящимися к группе, в которой ничего не вводили.
[Список последовательностей]
SEQ ID NO: 1
VPQKPKVSLN PPWNRIFKGE NVTLTCNGNN FFEVSSTKWF HNGSLSEETN
SSLNIVNAKF EDSGEYKCQH QQVNESEPVY LEVFSDWLLL QASAEVVMEG
QPLFLRCHGW RNWDVYKVIY YKDGEALKYW YENHNISITN ATVEDSGTYY
CTGKVWQLDY ESEPLNITVI KAPREKYWLQ
SEQ ID NO: 2
SHTQPLGVFL FPPKPKDTLM ISRTPEVTCV VVDVSQEDPE VQFNWYVDGV
EVHNAKTKPR EEQFNSTYRV VSVLTVLHQD WLNGKEYKCK VSNKGLPSSI
EKTISKAKGQ PREPQVYTLP PSQEEMTKNQ VSLTCLVKGF YPSDIAVEWE
SNGQPENNYK TTPPVLDSDG SFFLYSRLTV DKSRWQEGNV FSCSVMHEAL
HNHYTQKSLS LSLGK
SEQ ID NO: 3
RNTGRGGEEK KGSKEKEEQE ERETKTPECP
SEQ ID NO: 4
AQPQAEGSLA KATTAPATTR NTGRGGEEKK GSKEKEEQEE RETKTPECP
SEQ ID NO: 5
gtgccccaga agcccaaggt gagcctgaac cctccctgga acagaatctt
caagggcgag aacgtgaccc tgacctgcaa cggcaacaac ttcttcgagg
tgagcagcac caagtggttc cacaatggca gcctgagcga ggagaccaac
agctccctga acatcgtgaa cgccaagttc gaggacagcg gcgagtacaa
gtgccagcac cagcaggtga acgagagcga gcccgtgtac ctggaggtgt
tcagcgactg gctgctgctg caggccagcg ccgaggtggt gatggagggc
cagcccctgt tcctgagatg ccacggctgg agaaactggg acgtgtacaa
ggtgatctac tacaaggatg gcgaggccct gaagtactgg tacgagaacc
acaacatctc catcaccaac gccaccgtgg aggacagcgg cacctactac
tgcacaggca aggtgtggca gctggactac gagagcgagc ccctgaacat
caccgtgatc aaggctccca gagagaagta ctggctgcag
SEQ ID NO: 6
tgcgtggtcg tggatgtgag ccaggaagat cccgaagtgc agttcaactg
gtacgtggat ggcgtggaag tgcacaacgc caagaccaag cccagagaag
agcagttcaa ctccacctac agagtggtga gcgtgctgac cgtgctgcac
caggactggc tgaacggcaa ggagtacaag tgcaaggtgt ccaacaaagg
cctgcccagc tccatcgaga agaccatcag caaagccaaa ggccagccca
gagaacccca ggtgtacacc ctgcctccca gccaggaaga gatgaccaag
aaccaggtgt ccctgacctg cctggtgaaa ggcttctacc ccagcgacat
cgccgtggag tgggaaagca acggccagcc cgagaacaat tacaagacaa
cccctcccgt gctggatagc gatggcagct tctttctgta cagcagactg
accgtggaca agagcagatg gcaggaaggc aacgtgttca gctgcagcgt
gatgcacgaa gccctgcaca accactacac ccagaagagc ctgtccctga
gcctgggcaa g
SEQ ID NO: 7
aggaacaccg gcagaggagg cgaggaaaag aaaggaagca aggagaagga
ggagcaggag gaaagagaaa ccaagacccc cgagtgcccc agccacaccc
agcccctggg cgtgttcctg ttccccccca agcccaagga caccctgatg
atcagcagaa cccccgaggt gacc
SEQ ID NO: 8
gcccagcccc aggccgaggg cagcctggct aaggccacca cagctcccgc
caccaccagg aacaccggca gaggaggcga ggaaaagaaa ggaagcaagg
agaaggagga gcaggaggaa agagaaacca agacccccga gtgccccagc
cacacccagc ccctgggcgt gttcctgttc ccccccaagc ccaaggacac
cctgatgatc agcagaaccc ccgaggtgac c
SEQ ID NO: 9
MDAMLRGLCC VLLLCGAVFV SPSHA
SEQ ID NO: 10
atggacgcca tgctgagagg cctgtgctgt gtgctgctgc tgtgcggcgc
cgtgttcgtg tcccctagcc acgcc
SEQ ID NO: 11
MDAMLRGLCC VLLLCGAVFV SPSHAVPQKP KVSLNPPWNR IFKGENVTLT
CNGNNFFEVS STKWFHNGSL SEETNSSLNI VNAKFEDSGE YKCQHQQVNE
SEPVYLEVFS DWLLLQASAE VVMEGQPLFL RCHGWRNWDV YKVIYYKDGE
ALKYWYENHN ISITNATVED SGTYYCTGKV WQLDYESEPL NITVIKAPRE
KYWLQRNTGR GGEEKKGSKE KEEQEERETK TPECPSHTQP LGVFLFPPKP
KDTLMISRTP EVTCVVVDVS QEDPEVQFNW YVDGVEVHNA KTKPREEQFN
STYRVVSVLT VLHQDWLNGK EYKCKVSNKG LPSSIEKTIS KAKGQPREPQ
VYTLPPSQEE MTKNQVSLTC LVKGFYPSDI AVEWESNGQP ENNYKTTPPV
LDSDGSFFLY SRLTVDKSRW QEGNVFSCSV MHEALHNHYT QKSLSLSLGK
SEQ ID NO: 12
atggacgcca tgctgagagg cctgtgctgt gtgctgctgc tgtgcggcgc
cgtgttcgtg tcccctagcc acgccgtgcc ccagaagccc aaggtgagcc
tgaaccctcc ctggaacaga atcttcaagg gcgagaacgt gaccctgacc
tgcaacggca acaacttctt cgaggtgagc agcaccaagt ggttccacaa
tggcagcctg agcgaggaga ccaacagctc cctgaacatc gtgaacgcca
agttcgagga cagcggcgag tacaagtgcc agcaccagca ggtgaacgag
agcgagcccg tgtacctgga ggtgttcagc gactggctgc tgctgcaggc
cagcgccgag gtggtgatgg agggccagcc cctgttcctg agatgccacg
gctggagaaa ctgggacgtg tacaaggtga tctactacaa ggatggcgag
gccctgaagt actggtacga gaaccacaac atctccatca ccaacgccac
cgtggaggac agcggcacct actactgcac aggcaaggtg tggcagctgg
actacgagag cgagcccctg aacatcaccg tgatcaaggc tcccagagag
aagtactggc tgcagaggaa caccggcaga ggaggcgagg aaaagaaagg
aagcaaggag aaggaggagc aggaggaaag agaaaccaag acccccgagt
gccccagcca cacccagccc ctgggcgtgt tcctgttccc ccccaagccc
aaggacaccc tgatgatcag cagaaccccc gaggtgacct gcgtggtcgt
ggatgtgagc caggaagatc ccgaagtgca gttcaactgg tacgtggatg
gcgtggaagt gcacaacgcc aagaccaagc ccagagaaga gcagttcaac
tccacctaca gagtggtgag cgtgctgacc gtgctgcacc aggactggct
gaacggcaag gagtacaagt gcaaggtgtc caacaaaggc ctgcccagct
ccatcgagaa gaccatcagc aaagccaaag gccagcccag agaaccccag
gtgtacaccc tgcctcccag ccaggaagag atgaccaaga accaggtgtc
cctgacctgc ctggtgaaag gcttctaccc cagcgacatc gccgtggagt
gggaaagcaa cggccagccc gagaacaatt acaagacaac ccctcccgtg
ctggatagcg atggcagctt ctttctgtac agcagactga ccgtggacaa
gagcagatgg caggaaggca acgtgttcag ctgcagcgtg atgcacgaag
ccctgcacaa ccactacacc cagaagagcc tgtccctgag cctgggcaag
SEQ ID NO: 13
MDAMLRGLCC VLLLCGAVFV SPSHAVPQKP KVSLNPPWNR IFKGENVTLT
CNGNNFFEVS STKWFHNGSL SEETNSSLNI VNAKFEDSGE YKCQHQQVNE
SEPVYLEVFS DWLLLQASAE VVMEGQPLFL RCHGWRNWDV YKVIYYKDGE
ALKYWYENHN ISITNATVED SGTYYCTGKV WQLDYESEPL NITVIKAPRE
KYWLQAQPQA EGSLAKATTA PATTRNTGRG GEEKKGSKEK EEQEERETKT
PECPSHTQPL GVFLFPPKPK DTLMISRTPE VTCVVVDVSQ EDPEVQFNWY
VDGVEVHNAK TKPREEQFNS TYRVVSVLTV LHQDWLNGKE YKCKVSNKGL
PSSIEKTISK AKGQPREPQV YTLPPSQEEM TKNQVSLTCL VKGFYPSDIA
VEWESNGQPE NNYKTTPPVL DSDGSFFLYS RLTVDKSRWQ EGNVFSCSVM
HEALHNHYTQ KSLSLSLGK
SEQ ID NO: 14
atggacgcca tgctgagagg cctgtgctgt gtgctgctgc tgtgcggcgc
cgtgttcgtg tcccctagcc acgccgtgcc ccagaagccc aaggtgagcc
tgaaccctcc ctggaacaga atcttcaagg gcgagaacgt gaccctgacc
tgcaacggca acaacttctt cgaggtgagc agcaccaagt ggttccacaa
tggcagcctg agcgaggaga ccaacagctc cctgaacatc gtgaacgcca
agttcgagga cagcggcgag tacaagtgcc agcaccagca ggtgaacgag
agcgagcccg tgtacctgga ggtgttcagc gactggctgc tgctgcaggc
cagcgccgag gtggtgatgg agggccagcc cctgttcctg agatgccacg
gctggagaaa ctgggacgtg tacaaggtga tctactacaa ggatggcgag
gccctgaagt actggtacga gaaccacaac atctccatca ccaacgccac
cgtggaggac agcggcacct actactgcac aggcaaggtg tggcagctgg
actacgagag cgagcccctg aacatcaccg tgatcaaggc tcccagagag
aagtactggc tgcaggccca gccccaggcc gagggcagcc tggctaaggc
caccacagct cccgccacca ccaggaacac cggcagagga ggcgaggaaa
agaaaggaag caaggagaag gaggagcagg aggaaagaga aaccaagacc
cccgagtgcc ccagccacac ccagcccctg ggcgtgttcc tgttcccccc
caagcccaag gacaccctga tgatcagcag aacccccgag gtgacctgcg
tggtcgtgga tgtgagccag gaagatcccg aagtgcagtt caactggtac
gtggatggcg tggaagtgca caacgccaag accaagccca gagaagagca
gttcaactcc acctacagag tggtgagcgt gctgaccgtg ctgcaccagg
actggctgaa cggcaaggag tacaagtgca aggtgtccaa caaaggcctg
cccagctcca tcgagaagac catcagcaaa gccaaaggcc agcccagaga
accccaggtg tacaccctgc ctcccagcca ggaagagatg accaagaacc
aggtgtccct gacctgcctg gtgaaaggct tctaccccag cgacatcgcc
gtggagtggg aaagcaacgg ccagcccgag aacaattaca agacaacccc
tcccgtgctg gatagcgatg gcagcttctt tctgtacagc agactgaccg
tggacaagag cagatggcag gaaggcaacg tgttcagctg cagcgtgatg
cacgaagccc tgcacaacca ctacacccag aagagcctgt ccctgagcct
gggcaag
SEQ ID NO: 15
MIHTNLKKKF SCCVLVFLLF AVICVWKEKK KGSYYDSFKL QTKEFQVLKS
LGKLAMGSDS QSVSSSSTQD PHRGRQTLGS LRGLAKAKPE ASFQVWNKDS
SSKNLIPRLQ KIWKNYLSMN KYKVSYKGPG PGIKFSAEAL RCHLRDHVNV
SMVEVTDFPF NTSEWEGYLP KESIRTKAGP WGRCAVVSSA GSLKSSQLGR
EIDDHDAVLR FNGAPTANFQ QDVGTKTTIR LMNSQLVTTE KRFLKDSLYN
EGILIVWDPS VYHSDIPKWY QNPDYNFFNN YKTYRKLHPN QPFYILKPQM
PWELWDILQE ISPEEIQPNP PSSGMLGIII MMTLCDQVDI YEFLPSKRKT
DVCYYYQKFF DSACTMGAYH PLLYEKNLVK HLNQGTDEDI YLLGKATLPG
FRTIHC
SEQ ID NO: 16
atgatccaca ccaacctgaa gaagaagttc agctgctgcg tgctggtgtt
cctgctgttc gccgtgatct gcgtgtggaa ggagaagaag aaaggcagct
actacgacag cttcaagctg cagaccaagg agttccaggt gctgaagagc
ctgggcaagc tggccatggg cagcgacagc cagagcgtgt ccagctcctc
cacccaggat ccccacagag gcagacagac cctgggcagc ctgagaggcc
tggccaaggc caagcccgag gccagcttcc aggtgtggaa caaggacagc
agcagcaaga acctgatccc cagactgcag aagatctgga agaactacct
gagcatgaac aagtacaagg tgagctacaa aggacccgga cccggcatca
agttcagcgc cgaggccctg aggtgccacc tgagagacca cgtgaacgtg
agcatggtgg aagtgaccga cttccccttc aacaccagcg agtgggaagg
ctacctgccc aaggagagca tcaggaccaa ggctggcccc tggggcagat
gcgccgtggt gagcagcgct ggcagcctga agagctccca gctgggcaga
gagatcgacg accacgatgc cgtgctgagg ttcaatggcg ctcccaccgc
caacttccag caggacgtgg gcaccaagac cacaatccgg ctgatgaaca
gccagctggt gacaaccgag aagcggttcc tgaaggacag cctgtacaac
gagggcatcc tgatcgtgtg ggatcccagc gtgtaccaca gcgacatccc
caagtggtac cagaatcccg actacaactt cttcaacaac tacaagacct
atagaaagct gcaccccaac cagcccttct acatcctgaa gccccagatg
ccctgggagc tgtgggacat cctgcaggag atcagccctg aagagatcca
gcccaaccct ccctccagcg gcatgctggg cattatcatc atgatgaccc
tgtgcgacca ggtggacatc tacgagttcc tgcccagcaa gagaaagacc
gacgtgtgct actactatca gaagttcttc gacagcgcct gcaccatggg
cgcctaccac cccctgctgt acgagaagaa cctggtgaag cacctgaacc
agggcaccga cgaggacatc tacctgctgg gcaaagccac cctgcccggc
ttcagaacca tccactgc
SEQ ID NO: 17
RNTGRGGEEK KXXKEKEEQE ERETKTPECP
SEQ ID NO: 18
AQPQAEGSLA KATTAPATTR NTGRGGEEKK XXKEKEEQEE RETKTPECP
SEQ ID NO: 19
RNTGRGGEEK KKEKEKEEQE ERETKTPECP
SEQ ID NO: 20
VPQKPKVSLN PPWNRIFKGE NVTLTCNGNN FFEVSSTKWF HNGSLSEETN
SSLNIVNAKF EDSGEYKCQH QQVNESEPVY LEVFSDWLLL QASAEVVMEG
QPLFLRCHGW RNWDVYKVIY YKDGEALKYW YENHNISITN ATVEDSGTYY
CTGKVWQLDY ESEPLNITVI KAPREKYWLQ RNTGRGGEEK KKEKEKEEQE
ERETKTPECP SHTQPLGVFL FPPKPKDTLM ISRTPEVTCV VVDVSQEDPE
VQFNWYVDGV EVHNAKTKPR EEQFNSTYRV VSVLTVLHQD WLNGKEYKCK
VSNKGLPSSI EKTISKAKGQ PREPQVYTLP PSQEEMTKNQ VSLTCLVKGF
YPSDIAVEWE SNGQPENNYK TTPPVLDSDG SFFLYSRLTV DKSRWQEGNV
FSCSVMHEAL HNHYTQKSLS LSLGK
SEQ ID NO: 21
VPQKPKVSLN PPWNRIFKGE NVTLTCNGNN FFEVSSTKWF HNGSLSEETN
SSLNIVNAKF EDSGEYKCQH QQVNESEPVY LEVFSDWLLL QASAEVVMEG
QPLFLRCHGW RNWDVYKVIY YKDGEALKYW YENHNISITN ATVEDSGTYY
CTGKVWQLDY ESEPLNITVI KAPREKYWLQ RNTGRGGEEK KGSKEKEEQE
ERETKTPECP SHTQPLGVFL FPPKPKDTLM ISRTPEVTCV VVDVSQEDPE
VQFNWYVDGV EVHNAKTKPR EEQFNSTYRV VSVLTVLHQD WLNGKEYKCK
VSNKGLPSSI EKTISKAKGQ PREPQVYTLP PSQEEMTKNQ VSLTCLVKGF
YPSDIAVEWE SNGQPENNYK TTPPVLDSDG SFFLYSRLTV DKSRWQEGNV
FSCSVMHEAL HNHYTQKSLS LSLGK
SEQ ID NO: 22
VPQKPKVSLN PPWNRIFKGE NVTLTCNGNN FFEVSSTKWF HNGSLSEETN
SSLNIVNAKF EDSGEYKCQH QQVNESEPVY LEVFSDWLLL QASAEVVMEG
QPLFLRCHGW RNWDVYKVIY YKDGEALKYW YENHNISITN ATVEDSGTYY
CTGKVWQLDY ESEPLNITVI KAPREKYWLQ AQPQAEGSLA KATTAPATTR
NTGRGGEEKK GSKEKEEQEE RETKTPECPS HTQPLGVFLF PPKPKDTLMI
SRTPEVTCVV VDVSQEDPEV QFNWYVDGVE VHNAKTKPRE EQFNSTYRVV
SVLTVLHQDW LNGKEYKCKV SNKGLPSSIE KTISKAKGQP REPQVYTLPP
SQEEMTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS
FFLYSRLTVD KSRWQEGNVF SCSVMHEALH NHYTQKSLSL SLGK
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> PROGEN CO., LTD.
<120> ВНЕКЛЕТОЧНЫЙ ДОМЕН АЛЬФА-СУБЪЕДИНИЦЫ FC-РЕЦЕПТОРА IGE,
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ЕГО, И СПОСОБ ЕГО ПОЛУЧЕНИЯ
<130> PCB812140PRG/PCT
<150> KR 10-2018-0002248
<151> 2018-01-08
<160> 22
<170> KopatentIn 2.0
<210> 1
<211> 180
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FCeRI1 ECD
<400> 1
Val Pro Gln Lys Pro Lys Val Ser Leu Asn Pro Pro Trp Asn Arg Ile
1. 5 10 15
Phe Lys Gly Glu Asn Val Thr Leu Thr Cys Asn Gly Asn Asn Phe Phe
20 25 30
Glu Val Ser Ser Thr Lys Trp Phe His Asn Gly Ser Leu Ser Glu Glu
35 40 45
Thr Asn Ser Ser Leu Asn Ile Val Asn Ala Lys Phe Glu Asp Ser Gly
50 55 60
Glu Tyr Lys Cys Gln His Gln Gln Val Asn Glu Ser Glu Pro Val Tyr
65 70 75 80
Leu Glu Val Phe Ser Asp Trp Leu Leu Leu Gln Ala Ser Ala Glu Val
85 90 95
Val Met Glu Gly Gln Pro Leu Phe Leu Arg Cys His Gly Trp Arg Asn
100 105 110
Trp Asp Val Tyr Lys Val Ile Tyr Tyr Lys Asp Gly Glu Ala Leu Lys
115 120 125
Tyr Trp Tyr Glu Asn His Asn Ile Ser Ile Thr Asn Ala Thr Val Glu
130 135 140
Asp Ser Gly Thr Tyr Tyr Cys Thr Gly Lys Val Trp Gln Leu Asp Tyr
145 150 155 160
Glu Ser Glu Pro Leu Asn Ile Thr Val Ile Lys Ala Pro Arg Glu Lys
165 170 175
Tyr Trp Leu Gln
180
<210> 2
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Модифицированный Fc
<400> 2
Ser His Thr Gln Pro Leu Gly Val Phe Leu Phe Pro Pro Lys Pro Lys
1. 5 10 15
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
20 25 30
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
35 40 45
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
50 55 60
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
65 70 75 80
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
85 90 95
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
100 105 110
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
115 120 125
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
130 135 140
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
145 150 155 160
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
165 170 175
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
180 185 190
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
195 200 205
Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 3
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgD
<400> 3
Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Gly Ser Lys Glu Lys
1. 5 10 15
Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro
20 25 30
<210> 4
<211> 49
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgD
<400> 4
Ala Gln Pro Gln Ala Glu Gly Ser Leu Ala Lys Ala Thr Thr Ala Pro
1. 5 10 15
Ala Thr Thr Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Gly Ser
20 25 30
Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys
35 40 45
Pro
<210> 5
<211> 540
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность FCeRI1 ECD
<400> 5
gtgccccaga agcccaaggt gagcctgaac cctccctgga acagaatctt caagggcgag 60
aacgtgaccc tgacctgcaa cggcaacaac ttcttcgagg tgagcagcac caagtggttc 120
cacaatggca gcctgagcga ggagaccaac agctccctga acatcgtgaa cgccaagttc 180
gaggacagcg gcgagtacaa gtgccagcac cagcaggtga acgagagcga gcccgtgtac 240
ctggaggtgt tcagcgactg gctgctgctg caggccagcg ccgaggtggt gatggagggc 300
cagcccctgt tcctgagatg ccacggctgg agaaactggg acgtgtacaa ggtgatctac 360
tacaaggatg gcgaggccct gaagtactgg tacgagaacc acaacatctc catcaccaac 420
gccaccgtgg aggacagcgg cacctactac tgcacaggca aggtgtggca gctggactac 480
gagagcgagc ccctgaacat caccgtgatc aaggctccca gagagaagta ctggctgcag 540
540
<210> 6
<211> 561
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность Модифицированный Fc
<400> 6
tgcgtggtcg tggatgtgag ccaggaagat cccgaagtgc agttcaactg gtacgtggat 60
ggcgtggaag tgcacaacgc caagaccaag cccagagaag agcagttcaa ctccacctac 120
agagtggtga gcgtgctgac cgtgctgcac caggactggc tgaacggcaa ggagtacaag 180
tgcaaggtgt ccaacaaagg cctgcccagc tccatcgaga agaccatcag caaagccaaa 240
ggccagccca gagaacccca ggtgtacacc ctgcctccca gccaggaaga gatgaccaag 300
aaccaggtgt ccctgacctg cctggtgaaa ggcttctacc ccagcgacat cgccgtggag 360
tgggaaagca acggccagcc cgagaacaat tacaagacaa cccctcccgt gctggatagc 420
gatggcagct tctttctgta cagcagactg accgtggaca agagcagatg gcaggaaggc 480
aacgtgttca gctgcagcgt gatgcacgaa gccctgcaca accactacac ccagaagagc 540
ctgtccctga gcctgggcaa g 561
<210> 7
<211> 174
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность варианта шарнирной области IgD
<400> 7
aggaacaccg gcagaggagg cgaggaaaag aaaggaagca aggagaagga ggagcaggag 60
gaaagagaaa ccaagacccc cgagtgcccc agccacaccc agcccctggg cgtgttcctg 120
ttccccccca agcccaagga caccctgatg atcagcagaa cccccgaggt gacc 170
<210> 8
<211> 231
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность варианта шарнирной области IgD
<400> 8
gcccagcccc aggccgaggg cagcctggct aaggccacca cagctcccgc caccaccagg 60
aacaccggca gaggaggcga ggaaaagaaa ggaagcaagg agaaggagga gcaggaggaa 120
agagaaacca agacccccga gtgccccagc cacacccagc ccctgggcgt gttcctgttc 180
ccccccaagc ccaaggacac cctgatgatc agcagaaccc ccgaggtgac c 231
<210> 9
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный пептид
<400> 9
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1. 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala
20 25
<210> 10
<211> 75
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность сигнального пептида
<400> 10
atggacgcca tgctgagagg cctgtgctgt gtgctgctgc tgtgcggcgc cgtgttcgtg 60
tcccctagcc acgcc 75
<210> 11
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FceRIa ECD-шарнирная область-Fc2
<400> 11
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1. 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Pro Gln Lys Pro Lys Val
20 25 30
Ser Leu Asn Pro Pro Trp Asn Arg Ile Phe Lys Gly Glu Asn Val Thr
35 40 45
Leu Thr Cys Asn Gly Asn Asn Phe Phe Glu Val Ser Ser Thr Lys Trp
50 55 60
Phe His Asn Gly Ser Leu Ser Glu Glu Thr Asn Ser Ser Leu Asn Ile
65 70 75 80
Val Asn Ala Lys Phe Glu Asp Ser Gly Glu Tyr Lys Cys Gln His Gln
85 90 95
Gln Val Asn Glu Ser Glu Pro Val Tyr Leu Glu Val Phe Ser Asp Trp
100 105 110
Leu Leu Leu Gln Ala Ser Ala Glu Val Val Met Glu Gly Gln Pro Leu
115 120 125
Phe Leu Arg Cys His Gly Trp Arg Asn Trp Asp Val Tyr Lys Val Ile
130 135 140
Tyr Tyr Lys Asp Gly Glu Ala Leu Lys Tyr Trp Tyr Glu Asn His Asn
145 150 155 160
Ile Ser Ile Thr Asn Ala Thr Val Glu Asp Ser Gly Thr Tyr Tyr Cys
165 170 175
Thr Gly Lys Val Trp Gln Leu Asp Tyr Glu Ser Glu Pro Leu Asn Ile
180 185 190
Thr Val Ile Lys Ala Pro Arg Glu Lys Tyr Trp Leu Gln Arg Asn Thr
195 200 205
Gly Arg Gly Gly Glu Glu Lys Lys Gly Ser Lys Glu Lys Glu Glu Gln
210 215 220
Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro Ser His Thr Gln Pro
225 230 235 240
Leu Gly Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
435 440 445
Gly Lys
450
<210> 12
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность FceRIa ECD-шарнирная область-Fc2
<400> 12
atggacgcca tgctgagagg cctgtgctgt gtgctgctgc tgtgcggcgc cgtgttcgtg 60
tcccctagcc acgccgtgcc ccagaagccc aaggtgagcc tgaaccctcc ctggaacaga 120
atcttcaagg gcgagaacgt gaccctgacc tgcaacggca acaacttctt cgaggtgagc 180
agcaccaagt ggttccacaa tggcagcctg agcgaggaga ccaacagctc cctgaacatc 240
gtgaacgcca agttcgagga cagcggcgag tacaagtgcc agcaccagca ggtgaacgag 300
agcgagcccg tgtacctgga ggtgttcagc gactggctgc tgctgcaggc cagcgccgag 360
gtggtgatgg agggccagcc cctgttcctg agatgccacg gctggagaaa ctgggacgtg 420
tacaaggtga tctactacaa ggatggcgag gccctgaagt actggtacga gaaccacaac 480
atctccatca ccaacgccac cgtggaggac agcggcacct actactgcac aggcaaggtg 540
tggcagctgg actacgagag cgagcccctg aacatcaccg tgatcaaggc tcccagagag 600
aagtactggc tgcagaggaa caccggcaga ggaggcgagg aaaagaaagg aagcaaggag 660
aaggaggagc aggaggaaag agaaaccaag acccccgagt gccccagcca cacccagccc 720
ctgggcgtgt tcctgttccc ccccaagccc aaggacaccc tgatgatcag cagaaccccc 780
gaggtgacct gcgtggtcgt ggatgtgagc caggaagatc ccgaagtgca gttcaactgg 840
tacgtggatg gcgtggaagt gcacaacgcc aagaccaagc ccagagaaga gcagttcaac 900
tccacctaca gagtggtgag cgtgctgacc gtgctgcacc aggactggct gaacggcaag 960
gagtacaagt gcaaggtgtc caacaaaggc ctgcccagct ccatcgagaa gaccatcagc 1020
aaagccaaag gccagcccag agaaccccag gtgtacaccc tgcctcccag ccaggaagag 1080
atgaccaaga accaggtgtc cctgacctgc ctggtgaaag gcttctaccc cagcgacatc 1140
gccgtggagt gggaaagcaa cggccagccc gagaacaatt acaagacaac ccctcccgtg 1200
ctggatagcg atggcagctt ctttctgtac agcagactga ccgtggacaa gagcagatgg 1260
caggaaggca acgtgttcag ctgcagcgtg atgcacgaag ccctgcacaa ccactacacc 1320
cagaagagcc tgtccctgag cctgggcaag 1350
<210> 13
<211> 469
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FceRIa ECD-шарнир-Fc3
<400> 13
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1. 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Pro Gln Lys Pro Lys Val
20 25 30
Ser Leu Asn Pro Pro Trp Asn Arg Ile Phe Lys Gly Glu Asn Val Thr
35 40 45
Leu Thr Cys Asn Gly Asn Asn Phe Phe Glu Val Ser Ser Thr Lys Trp
50 55 60
Phe His Asn Gly Ser Leu Ser Glu Glu Thr Asn Ser Ser Leu Asn Ile
65 70 75 80
Val Asn Ala Lys Phe Glu Asp Ser Gly Glu Tyr Lys Cys Gln His Gln
85 90 95
Gln Val Asn Glu Ser Glu Pro Val Tyr Leu Glu Val Phe Ser Asp Trp
100 105 110
Leu Leu Leu Gln Ala Ser Ala Glu Val Val Met Glu Gly Gln Pro Leu
115 120 125
Phe Leu Arg Cys His Gly Trp Arg Asn Trp Asp Val Tyr Lys Val Ile
130 135 140
Tyr Tyr Lys Asp Gly Glu Ala Leu Lys Tyr Trp Tyr Glu Asn His Asn
145 150 155 160
Ile Ser Ile Thr Asn Ala Thr Val Glu Asp Ser Gly Thr Tyr Tyr Cys
165 170 175
Thr Gly Lys Val Trp Gln Leu Asp Tyr Glu Ser Glu Pro Leu Asn Ile
180 185 190
Thr Val Ile Lys Ala Pro Arg Glu Lys Tyr Trp Leu Gln Ala Gln Pro
195 200 205
Gln Ala Glu Gly Ser Leu Ala Lys Ala Thr Thr Ala Pro Ala Thr Thr
210 215 220
Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Gly Ser Lys Glu Lys
225 230 235 240
Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro Ser His
245 250 255
Thr Gln Pro Leu Gly Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
260 265 270
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
275 280 285
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
290 295 300
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
305 310 315 320
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
325 330 335
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
340 345 350
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
355 360 365
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
370 375 380
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
385 390 395 400
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
405 410 415
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
420 425 430
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
435 440 445
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
450 455 460
Leu Ser Leu Gly Lys
465
<210> 14
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность FceRIa ECD-шарнир-Fc3
<400> 14
atggacgcca tgctgagagg cctgtgctgt gtgctgctgc tgtgcggcgc cgtgttcgtg 60
tcccctagcc acgccgtgcc ccagaagccc aaggtgagcc tgaaccctcc ctggaacaga 120
atcttcaagg gcgagaacgt gaccctgacc tgcaacggca acaacttctt cgaggtgagc 180
agcaccaagt ggttccacaa tggcagcctg agcgaggaga ccaacagctc cctgaacatc 240
gtgaacgcca agttcgagga cagcggcgag tacaagtgcc agcaccagca ggtgaacgag 300
agcgagcccg tgtacctgga ggtgttcagc gactggctgc tgctgcaggc cagcgccgag 360
gtggtgatgg agggccagcc cctgttcctg agatgccacg gctggagaaa ctgggacgtg 420
tacaaggtga tctactacaa ggatggcgag gccctgaagt actggtacga gaaccacaac 480
atctccatca ccaacgccac cgtggaggac agcggcacct actactgcac aggcaaggtg 540
tggcagctgg actacgagag cgagcccctg aacatcaccg tgatcaaggc tcccagagag 600
aagtactggc tgcaggccca gccccaggcc gagggcagcc tggctaaggc caccacagct 660
cccgccacca ccaggaacac cggcagagga ggcgaggaaa agaaaggaag caaggagaag 720
gaggagcagg aggaaagaga aaccaagacc cccgagtgcc ccagccacac ccagcccctg 780
ggcgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagcag aacccccgag 840
gtgacctgcg tggtcgtgga tgtgagccag gaagatcccg aagtgcagtt caactggtac 900
gtggatggcg tggaagtgca caacgccaag accaagccca gagaagagca gttcaactcc 960
acctacagag tggtgagcgt gctgaccgtg ctgcaccagg actggctgaa cggcaaggag 1020
tacaagtgca aggtgtccaa caaaggcctg cccagctcca tcgagaagac catcagcaaa 1080
gccaaaggcc agcccagaga accccaggtg tacaccctgc ctcccagcca ggaagagatg 1140
accaagaacc aggtgtccct gacctgcctg gtgaaaggct tctaccccag cgacatcgcc 1200
gtggagtggg aaagcaacgg ccagcccgag aacaattaca agacaacccc tcccgtgctg 1260
gatagcgatg gcagcttctt tctgtacagc agactgaccg tggacaagag cagatggcag 1320
gaaggcaacg tgttcagctg cagcgtgatg cacgaagccc tgcacaacca ctacacccag 1380
aagagcctgt ccctgagcct gggcaag 1407
<210> 15
<211> 406
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> трансфераза a-2,6 сиаловой кислоты человека
<400> 15
Met Ile His Thr Asn Leu Lys Lys Lys Phe Ser Cys Cys Val Leu Val
1. 5 10 15
Phe Leu Leu Phe Ala Val Ile Cys Val Trp Lys Glu Lys Lys Lys Gly
20 25 30
Ser Tyr Tyr Asp Ser Phe Lys Leu Gln Thr Lys Glu Phe Gln Val Leu
35 40 45
Lys Ser Leu Gly Lys Leu Ala Met Gly Ser Asp Ser Gln Ser Val Ser
50 55 60
Ser Ser Ser Thr Gln Asp Pro His Arg Gly Arg Gln Thr Leu Gly Ser
65 70 75 80
Leu Arg Gly Leu Ala Lys Ala Lys Pro Glu Ala Ser Phe Gln Val Trp
85 90 95
Asn Lys Asp Ser Ser Ser Lys Asn Leu Ile Pro Arg Leu Gln Lys Ile
100 105 110
Trp Lys Asn Tyr Leu Ser Met Asn Lys Tyr Lys Val Ser Tyr Lys Gly
115 120 125
Pro Gly Pro Gly Ile Lys Phe Ser Ala Glu Ala Leu Arg Cys His Leu
130 135 140
Arg Asp His Val Asn Val Ser Met Val Glu Val Thr Asp Phe Pro Phe
145 150 155 160
Asn Thr Ser Glu Trp Glu Gly Tyr Leu Pro Lys Glu Ser Ile Arg Thr
165 170 175
Lys Ala Gly Pro Trp Gly Arg Cys Ala Val Val Ser Ser Ala Gly Ser
180 185 190
Leu Lys Ser Ser Gln Leu Gly Arg Glu Ile Asp Asp His Asp Ala Val
195 200 205
Leu Arg Phe Asn Gly Ala Pro Thr Ala Asn Phe Gln Gln Asp Val Gly
210 215 220
Thr Lys Thr Thr Ile Arg Leu Met Asn Ser Gln Leu Val Thr Thr Glu
225 230 235 240
Lys Arg Phe Leu Lys Asp Ser Leu Tyr Asn Glu Gly Ile Leu Ile Val
245 250 255
Trp Asp Pro Ser Val Tyr His Ser Asp Ile Pro Lys Trp Tyr Gln Asn
260 265 270
Pro Asp Tyr Asn Phe Phe Asn Asn Tyr Lys Thr Tyr Arg Lys Leu His
275 280 285
Pro Asn Gln Pro Phe Tyr Ile Leu Lys Pro Gln Met Pro Trp Glu Leu
290 295 300
Trp Asp Ile Leu Gln Glu Ile Ser Pro Glu Glu Ile Gln Pro Asn Pro
305 310 315 320
Pro Ser Ser Gly Met Leu Gly Ile Ile Ile Met Met Thr Leu Cys Asp
325 330 335
Gln Val Asp Ile Tyr Glu Phe Leu Pro Ser Lys Arg Lys Thr Asp Val
340 345 350
Cys Tyr Tyr Tyr Gln Lys Phe Phe Asp Ser Ala Cys Thr Met Gly Ala
355 360 365
Tyr His Pro Leu Leu Tyr Glu Lys Asn Leu Val Lys His Leu Asn Gln
370 375 380
Gly Thr Asp Glu Asp Ile Tyr Leu Leu Gly Lys Ala Thr Leu Pro Gly
385 390 395 400
Phe Arg Thr Ile His Cys
405
<210> 16
<211> 1218
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность трансферазы a-2,6-сиаловой
кислоты человека
<400> 16
atgatccaca ccaacctgaa gaagaagttc agctgctgcg tgctggtgtt cctgctgttc 60
gccgtgatct gcgtgtggaa ggagaagaag aaaggcagct actacgacag cttcaagctg 120
cagaccaagg agttccaggt gctgaagagc ctgggcaagc tggccatggg cagcgacagc 180
cagagcgtgt ccagctcctc cacccaggat ccccacagag gcagacagac cctgggcagc 240
ctgagaggcc tggccaaggc caagcccgag gccagcttcc aggtgtggaa caaggacagc 300
agcagcaaga acctgatccc cagactgcag aagatctgga agaactacct gagcatgaac 360
aagtacaagg tgagctacaa aggacccgga cccggcatca agttcagcgc cgaggccctg 420
aggtgccacc tgagagacca cgtgaacgtg agcatggtgg aagtgaccga cttccccttc 480
aacaccagcg agtgggaagg ctacctgccc aaggagagca tcaggaccaa ggctggcccc 540
tggggcagat gcgccgtggt gagcagcgct ggcagcctga agagctccca gctgggcaga 600
gagatcgacg accacgatgc cgtgctgagg ttcaatggcg ctcccaccgc caacttccag 660
caggacgtgg gcaccaagac cacaatccgg ctgatgaaca gccagctggt gacaaccgag 720
aagcggttcc tgaaggacag cctgtacaac gagggcatcc tgatcgtgtg ggatcccagc 780
gtgtaccaca gcgacatccc caagtggtac cagaatcccg actacaactt cttcaacaac 840
tacaagacct atagaaagct gcaccccaac cagcccttct acatcctgaa gccccagatg 900
ccctgggagc tgtgggacat cctgcaggag atcagccctg aagagatcca gcccaaccct 960
ccctccagcg gcatgctggg cattatcatc atgatgaccc tgtgcgacca ggtggacatc 1020
tacgagttcc tgcccagcaa gagaaagacc gacgtgtgct actactatca gaagttcttc 1080
gacagcgcct gcaccatggg cgcctaccac cccctgctgt acgagaagaa cctggtgaag 1140
cacctgaacc agggcaccga cgaggacatc tacctgctgg gcaaagccac cctgcccggc 1200
ttcagaacca tccactgc 1218
<210> 17
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgD
<400> 17
Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Xaa Xaa Lys Glu Lys
1. 5 10 15
Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro
20 25 30
<210> 18
<211> 49
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgD
<400> 18
Ala Gln Pro Gln Ala Glu Gly Ser Leu Ala Lys Ala Thr Thr Ala Pro
1. 5 10 15
Ala Thr Thr Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Xaa Xaa
20 25 30
Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys
35 40 45
Pro
<210> 19
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> шарнирная область IgD
<400> 19
Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Lys Glu Lys Glu Lys
1. 5 10 15
Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro
20 25 30
<210> 20
<211> 425
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FceRIa ECD-шарнир-Fc1
<400> 20
Val Pro Gln Lys Pro Lys Val Ser Leu Asn Pro Pro Trp Asn Arg Ile
1. 5 10 15
Phe Lys Gly Glu Asn Val Thr Leu Thr Cys Asn Gly Asn Asn Phe Phe
20 25 30
Glu Val Ser Ser Thr Lys Trp Phe His Asn Gly Ser Leu Ser Glu Glu
35 40 45
Thr Asn Ser Ser Leu Asn Ile Val Asn Ala Lys Phe Glu Asp Ser Gly
50 55 60
Glu Tyr Lys Cys Gln His Gln Gln Val Asn Glu Ser Glu Pro Val Tyr
65 70 75 80
Leu Glu Val Phe Ser Asp Trp Leu Leu Leu Gln Ala Ser Ala Glu Val
85 90 95
Val Met Glu Gly Gln Pro Leu Phe Leu Arg Cys His Gly Trp Arg Asn
100 105 110
Trp Asp Val Tyr Lys Val Ile Tyr Tyr Lys Asp Gly Glu Ala Leu Lys
115 120 125
Tyr Trp Tyr Glu Asn His Asn Ile Ser Ile Thr Asn Ala Thr Val Glu
130 135 140
Asp Ser Gly Thr Tyr Tyr Cys Thr Gly Lys Val Trp Gln Leu Asp Tyr
145 150 155 160
Glu Ser Glu Pro Leu Asn Ile Thr Val Ile Lys Ala Pro Arg Glu Lys
165 170 175
Tyr Trp Leu Gln Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Lys
180 185 190
Glu Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu
195 200 205
Cys Pro Ser His Thr Gln Pro Leu Gly Val Phe Leu Phe Pro Pro Lys
210 215 220
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
225 230 235 240
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
245 250 255
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
260 265 270
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
275 280 285
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
290 295 300
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
305 310 315 320
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
325 330 335
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
340 345 350
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
355 360 365
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
370 375 380
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
385 390 395 400
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
405 410 415
Lys Ser Leu Ser Leu Ser Leu Gly Lys
420 425
<210> 21
<211> 425
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FceRIa ECD-шарнир-Fc2
<400> 21
Val Pro Gln Lys Pro Lys Val Ser Leu Asn Pro Pro Trp Asn Arg Ile
1. 5 10 15
Phe Lys Gly Glu Asn Val Thr Leu Thr Cys Asn Gly Asn Asn Phe Phe
20 25 30
Glu Val Ser Ser Thr Lys Trp Phe His Asn Gly Ser Leu Ser Glu Glu
35 40 45
Thr Asn Ser Ser Leu Asn Ile Val Asn Ala Lys Phe Glu Asp Ser Gly
50 55 60
Glu Tyr Lys Cys Gln His Gln Gln Val Asn Glu Ser Glu Pro Val Tyr
65 70 75 80
Leu Glu Val Phe Ser Asp Trp Leu Leu Leu Gln Ala Ser Ala Glu Val
85 90 95
Val Met Glu Gly Gln Pro Leu Phe Leu Arg Cys His Gly Trp Arg Asn
100 105 110
Trp Asp Val Tyr Lys Val Ile Tyr Tyr Lys Asp Gly Glu Ala Leu Lys
115 120 125
Tyr Trp Tyr Glu Asn His Asn Ile Ser Ile Thr Asn Ala Thr Val Glu
130 135 140
Asp Ser Gly Thr Tyr Tyr Cys Thr Gly Lys Val Trp Gln Leu Asp Tyr
145 150 155 160
Glu Ser Glu Pro Leu Asn Ile Thr Val Ile Lys Ala Pro Arg Glu Lys
165 170 175
Tyr Trp Leu Gln Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Gly
180 185 190
Ser Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu
195 200 205
Cys Pro Ser His Thr Gln Pro Leu Gly Val Phe Leu Phe Pro Pro Lys
210 215 220
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
225 230 235 240
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
245 250 255
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
260 265 270
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
275 280 285
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
290 295 300
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
305 310 315 320
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
325 330 335
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
340 345 350
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
355 360 365
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
370 375 380
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
385 390 395 400
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
405 410 415
Lys Ser Leu Ser Leu Ser Leu Gly Lys
420 425
<210> 22
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FceRIa ECD-шарнир-Fc3
<400> 22
Val Pro Gln Lys Pro Lys Val Ser Leu Asn Pro Pro Trp Asn Arg Ile
1. 5 10 15
Phe Lys Gly Glu Asn Val Thr Leu Thr Cys Asn Gly Asn Asn Phe Phe
20 25 30
Glu Val Ser Ser Thr Lys Trp Phe His Asn Gly Ser Leu Ser Glu Glu
35 40 45
Thr Asn Ser Ser Leu Asn Ile Val Asn Ala Lys Phe Glu Asp Ser Gly
50 55 60
Glu Tyr Lys Cys Gln His Gln Gln Val Asn Glu Ser Glu Pro Val Tyr
65 70 75 80
Leu Glu Val Phe Ser Asp Trp Leu Leu Leu Gln Ala Ser Ala Glu Val
85 90 95
Val Met Glu Gly Gln Pro Leu Phe Leu Arg Cys His Gly Trp Arg Asn
100 105 110
Trp Asp Val Tyr Lys Val Ile Tyr Tyr Lys Asp Gly Glu Ala Leu Lys
115 120 125
Tyr Trp Tyr Glu Asn His Asn Ile Ser Ile Thr Asn Ala Thr Val Glu
130 135 140
Asp Ser Gly Thr Tyr Tyr Cys Thr Gly Lys Val Trp Gln Leu Asp Tyr
145 150 155 160
Glu Ser Glu Pro Leu Asn Ile Thr Val Ile Lys Ala Pro Arg Glu Lys
165 170 175
Tyr Trp Leu Gln Ala Gln Pro Gln Ala Glu Gly Ser Leu Ala Lys Ala
180 185 190
Thr Thr Ala Pro Ala Thr Thr Arg Asn Thr Gly Arg Gly Gly Glu Glu
195 200 205
Lys Lys Gly Ser Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys
210 215 220
Thr Pro Glu Cys Pro Ser His Thr Gln Pro Leu Gly Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ПРОБИОТИКИ И ПОЛИПЕПТИД, ОБЛАДАЮЩИЙ АФФИННОСТЬЮ СВЯЗЫВАНИЯ В ОТНОШЕНИИ IGE, И ЕЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2786578C2 |
| ДИМЕР ПОЛИПЕПТИДА С ВЫСОКИМ СОДЕРЖАНИЕМ СИАЛОВОЙ КИСЛОТЫ, ВКЛЮЧАЮЩЕГО ЭКСТРАКЛЕТОЧНЫЙ ДОМЕН АЛЬФА-СУБЪЕДИНИЦЫ РЕЦЕПТОРА ДЛЯ Fc-ОБЛАСТИ IgE, И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2020 |
|
RU2833598C2 |
| СЛИТЫЕ МОЛЕКУЛЫ ЭКТОДОМЕНА РЕЦЕПТОРА TGF-БЕТА И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2797464C2 |
| АНТИ-IgE АНТИТЕЛА | 2017 |
|
RU2816207C2 |
| Новые гибридные ActRIIB белки-ловушки лигандов для лечения заболеваний, связанных с мышечной атрофией | 2016 |
|
RU2834707C1 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2728861C1 |
| ВАРИАНТЫ FC-ОБЛАСТИ С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С НЕОНАТАЛЬНЫМ FC-РЕЦЕПТОРОМ (FCRN) ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2830231C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2698655C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2746550C2 |
| АНТАГОНИСТЫ NGF ДЛЯ МЕДИЦИНСКОГО ИСПОЛЬЗОВАНИЯ | 2020 |
|
RU2829812C2 |


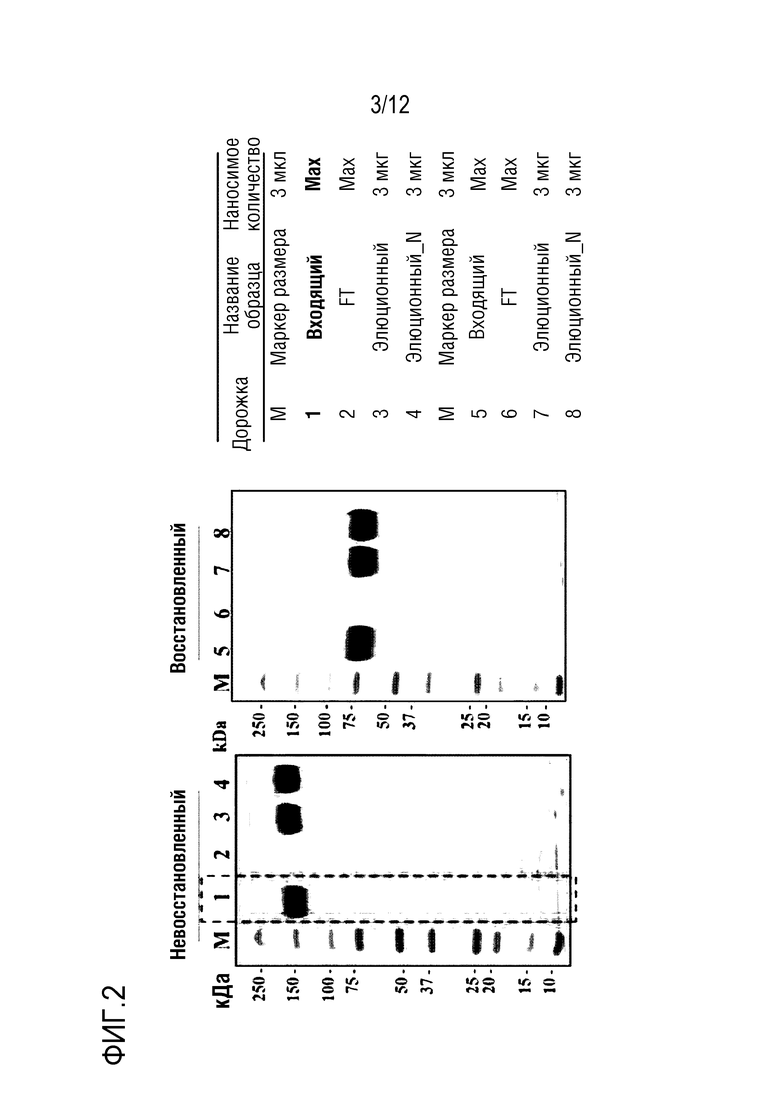
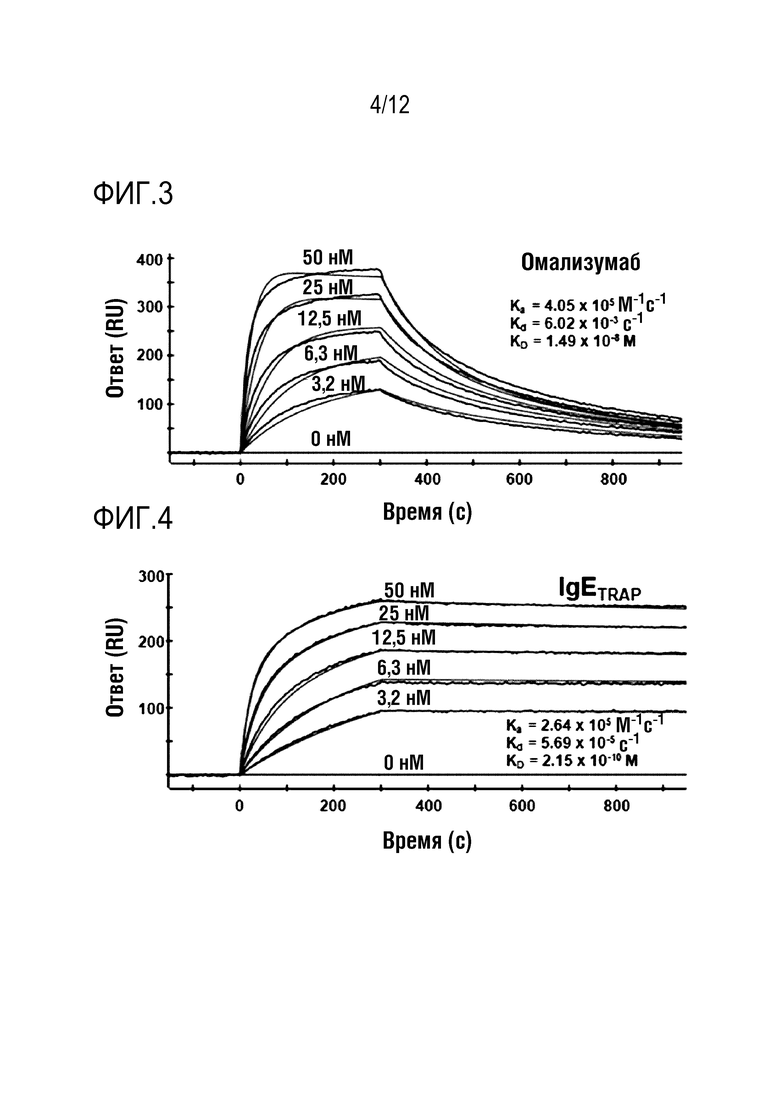





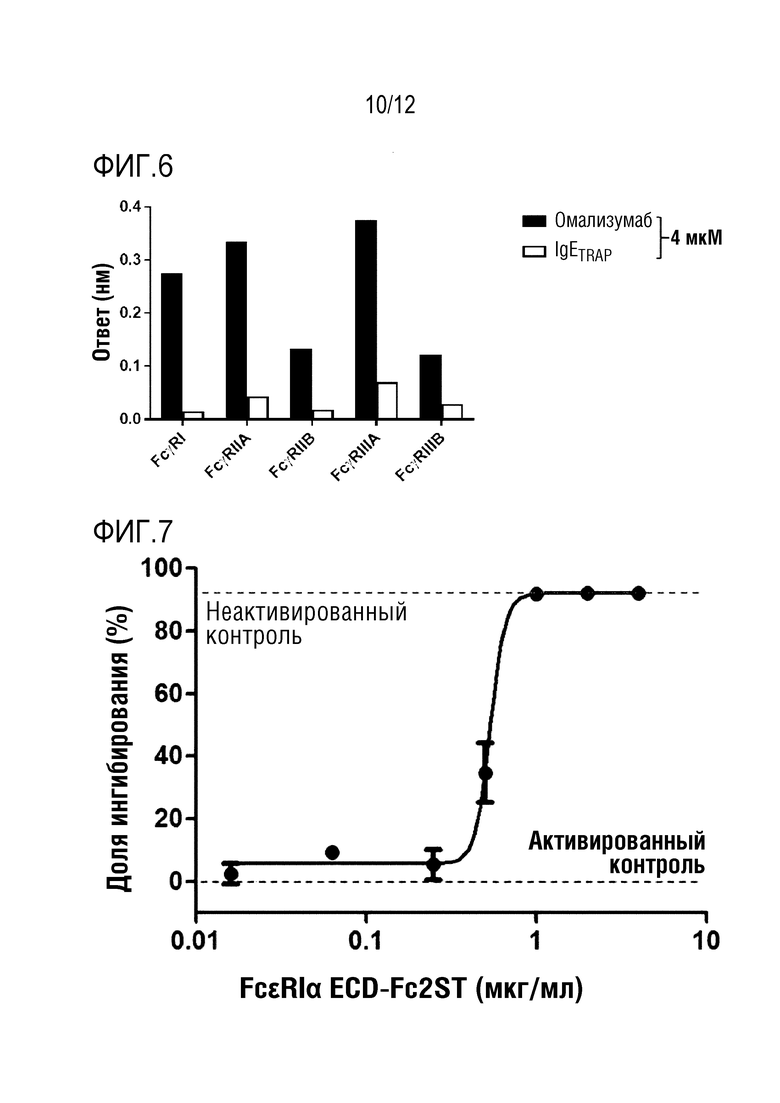


Изобретение относится полипептидному димеру, обладающему способностью связываться с IgE. Полипептидный димер содержит два мономера, каждый из которых содержит внеклеточный домен альфа-субъединицы Fc-рецептора IgE (FcεRIa-ECD). Также предложен полинуклеотид, который кодирует мономер, содержащийся в указанном полипептидном димере, при этом мономер содержит внеклеточный домен альфа-субъединицы Fc-рецептора IgE (FcεRIa-ECD). Также предложены экспрессирующий вектор, клетка-хозяин, фармацевтическая композиция, пищевая композиция, способ получения полипептидного димера и способ лечения или предупреждения аллергического заболевания. Полипептидный димер связывается с IgE и проявляет меньше побочных эффектов вследствие отсутствия функций ADCC и CDC. Изобретение может быть использовано для лечения или предупреждения IgE-опосредуемого аллергического заболевания. 8 н. и 2 з.п. ф-лы, 9 ил., 3 табл., 7 пр.
1. Полипептидный димер, обладающий способностью связываться с IgE, содержащий:
два мономера, каждый из которых содержит внеклеточный домен альфа-субъединицы Fc-рецептора IgE (FcεRIa-ECD),
где FcεRIa-ECD состоит из аминокислотной последовательности SEQ ID NO: 1,
где мономер содержит модифицированную Fc-область, состоящую из аминокислотной последовательности SEQ ID NO: 2, и
модифицированная Fc-область и FcεRIa-ECD связаны через шарнирную область,
где шарнирная область, происходящая из иммуноглобулина IgD, представляет собой Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Xaa1 Xaa2 Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro (SEQ ID NO: 17) или Ala Gln Pro Gln Ala Glu Gly Ser Leu Ala Lys Ala Thr Thr Ala Pro Ala Thr Thr Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Xaa3 Xaa4 Lys Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro (SEQ ID NO: 18), где
Xaa1 и Xaa3 представляют собой Gly и
Xaa2 и Xaa4 представляют собой Gly или Ser.
2. Полипептидный димер по п.1,
где шарнирная область имеет любую из аминокислотных последовательностей, выбранных из группы, состоящей из SEQ ID NO: 3 и SEQ ID NO: 4.
3. Полинуклеотид, который кодирует мономер, содержащийся в полипептидном димере по п.1,
где мономер содержит внеклеточный домен альфа-субъединицы Fc-рецептора IgE (FcεRIa-ECD),
где мономер состоит из аминокислотной последовательности SEQ ID NO: 21 или SEQ ID NO: 22.
4. Экспрессирующий вектор, в который встроен полинуклеотид по п.3.
5. Клетка-хозяин, которая содержит полинуклеотид, кодирующий полипептидный димер по п.1, и продуцирует полипептидный димер по п.1.
6. Фармацевтическая композиция для лечения или предупреждения аллергического заболевания, содержащая любой из полипептидных димеров по пп.1, 2 в качестве активного ингредиента и фармацевтически приемлемый носитель,
где полипептидный димер присутствует в эффективном количестве от 0,001% по массе до 20,0% по массе.
7. Фармацевтическая композиция по п.6,
где аллергическое заболевание представляет собой любое заболевание, выбранное из группы, состоящей из пищевой аллергии, атопического дерматита, астмы, аллергического ринита, аллергического конъюнктивита, аллергического дерматита, хронической идиопатической крапивницы и аллергического контактного дерматита.
8. Пищевая композиция для смягчения симптома аллергии, содержащая любой из полипептидных димеров по пп.1-2 в качестве активного ингредиента и приемлемый носитель,
где полипептидный димер присутствует в эффективном количестве от 0,001% по массе до 20,0% по массе,
где аллергический симптом представляет собой симптом любого из заболеваний, выбранного из группы, состоящей из пищевой аллергии, атопического дерматита, астмы, аллергического ринита, аллергического конъюнктивита, аллергического дерматита, хронической идиопатической крапивницы и аллергического контактного дерматита.
9. Способ получения полипептидного димера, включающий:
стадию культивирования клетки, в которую введен полинуклеотид, кодирующий полипептидный димер по п.1, и ген трансферазы сиаловых кислот; и
стадию выделения полипептидного димера.
10. Способ лечения или предупреждения аллергического заболевания, включающий:
стадию введения индивидууму полипептидного димера по п.1.
| EP 3260469 A1, 27.12.2017 | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| АПОПТОТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ IgE | 2008 |
|
RU2500686C2 |

