Область техники
Настоящее изобретение относится к области биотехнологии разведения животных, а конкретно - к применению гипофизарного полипептида, активирующего аденилатциклазу (РАСАР), в культуре водных организмов. Введение пептида водным организмам путем наслаивания, инъекции или в качестве пищевой добавки приводит к повышению у этих организмов потребности в пище, большей скорости роста и выживаемости, более высокой иммунной активности и повышению выделения пролактина.
Предшествующий уровень техники
Гипофизарный полипептид, активирующий аденилатциклазу (PACAP), был впервые выделен в 1989 г. из гипоталамуса быка, и была показана его способность стимулировать секрецию гормона роста через активацию фермента аденилатциклазы (Miyata and col. (1989) Isolation of a novel 38 residue hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 164:567-574). PACAP принадлежит к семейству пептидов, которое включает в себя секретин, глюкагон и кишечный вазоактивный пептид (Arimura and Shioda (1995) Pituitary adenylate cyclase-activating polypeptide (PACAP) and its receptors: Neuroendocrine and endocrine interaction. Front. Neuroendocrinol. 16:53-88). У млекопитающих предшественники PACAP и релизинг-фактора гормона роста (GHRH) кодируются двумя различными генами (Hosoya and col. (1992) Structure of the human Pituitary adenylate cyclase-activating polypeptide (PACAP) gen. Biochim. Biophys. Acta. 1129:199-206). Во всех видах таксонов, более низких, чем млекопитающие, изученных к настоящему времени (птицы, рептилии и рыбы), пептиды GHRH и PACAP кодируются одним и тем же геном и содержатся в одном и том же предшественнике (Montero and col. (2000) Molecular evolution of the growth hormone-releasing hormone/ Pituitary adenylate cyclase-activating polypeptide gene family. Functional implication in the regulation of growth hormone secretion. Journal of Molec. Endocrinol. 25:157-168). Ген PACAP сильно экспрессируется в: центральной и периферической нервной системе, нервных волокнах, иннервирующих глаза, дыхательном тракте, слюнных железах, желудочно-кишечном тракте, органах системы размножения, поджелудочной железе и мочевыводящем тракте. Он также синтезируется в надпочечниках, гонадах и иммунных клетках (Sherwood and col. (2000) The origin and function of the Pituitary adenylate cyclase-activating polypeptide (PACAP)/Glucagon Superfamily. Endocrine Review 21:619-670). PACAP демонстрирует участие в различных биологических функциях, что соответствует его широкому распространению в различных тканях и его гипофизотропной, нейромедиаторной, нейромодуляторной и вазорегуляторной активности. (Chatterjee and col. (1997) Genomic organization of the rat Pituitary adenylate cyclase-activating polypeptide receptor gene. Alternative splicing within the 59-untranslated region. J. Biol. Chem. 272:12122-12131).
Он участвует в регуляции деления, дифференцировки и гибели клеток (Sherwood and col. (2000) The origin y function of the Pituitary adenylate cyclase-activating polypeptide (PACAP)/Glucagon Superfamily. Endocrine Review 21:619-670).
PACAP стимулирует выделение гормона роста (GH). Эффект пептида в выделении GH был показан in vitro на нескольких видах млекопитающих, птиц, амфибий (Hu and col. (2000) Characterization and messenger ribonucleic acid distribution of a cloned Pituitary adenylate cyclase-activating polypeptide type I receptor in the frog Xenopus laevis brain. Endocrinol. 141:657-665) и птиц (Anderson L. L. and col. (2004) Growth hormone Secretion: Molecular and Cellular Mechanisms and In Vivo Approaches. Society for Experim. Biol, and Med. 229:291-302). Существует небольшое количество исследований эффекта PACAP на секрецию и высвобождение in vivo. К настоящему времени известно, что этот пептид in vivo повышает уровень GH в плазме у крыс (Jarr and col. (1992) Contrasting effects of pituitary adenylate cyclase activating polypeptide (PACAP) on in vivo and in vitro prolactin and growth hormone release in male rats. Life Sci. 51:823-830) и в плазме быка (Radcliff and col. (2001) Pituitary adenylate cyclase-activating polypeptide induces secretion of growth hormone in cattle. Domestic. Animal. Endocrinol. 21:187-196). В то же время у овец (Sawangjaroen and Curlewis (1994) Effects of pituitary adenylate cyclase activating polypeptide (PACAP) and vasoactive intestinal polypeptide (VIP) on prolactin, luteinizing hormone and growth hormone secretion in the ewe. J. Neuroendocrinol. 6:549-555) и у человека (Chiodera and col. (1996) Effects of intravenously infused pituitary adenylate cyclase activating polypeptide on adenohypophyseal hormone secretion innormal men. Clin. Neuroendocrinol. 64:242-246) он не вызывает такого эффекта.
Эти данные позволяют предположить, что у млекопитающих эффект пептида на секрецию GH варьирует от вида к виду (Anderson and col. (2004) Growth hormone Secretion: Molecular and Cellular Mechanisms and In Vivo Approaches. Society for Experim. Biol. and Med. 229:291-302).
До настоящего времени на рыбах исследования in vivo, которые показывают функцию PACAP в регуляции GH, отсутствуют, кроме того, отсутствует предшествующий опыт применения этого пептида в стимуляции потребности в пище у водных организмов. У ракообразных пока отсутствуют доказательства присутствия этого пептида и неизвестен каскад передачи сигнала, который регулирует рост этих организмов.
PACAP стимулирует высвобождение пролактина гипофизарными клетками млекопитающих (Ortmann and col. (1999) Interactions of ovarian steroids with Pituitary adenylate cyclase-activating polypeptide and GnRH in anterior pituitary cells. Eur. J. Endocrinol. 140:207-214). Это активизирует высвобождение мелатропин -меланоцит-симулирующего гормона (MSH) меланотропными клетками гипофиза. (Vaudry and col. (2000) Pituitary adenylate cyclase-activating polypeptide and its receptors: from structure to functions. Pharmacol. Rev.s 52:269-364). Исследования in vivo на рыбах, которые бы показывали активацию этим пептидом высвобождение пролактина, отсутствуют. Также отсутствуют данные о его эффекте на развитие окраски рыб.
У млекопитающих функция PACAP в иммунной системе очень хорошо охарактеризована, и существует несколько патентов, в которых описано их применение на человеке в качестве модулятора иммунологического ответа. До настоящего времени в существующей литературе отсутствуют данные, которые бы объясняли функцию PACAP в иммунной системе водных организмов. Ген PACAP клонирован у нескольких видов позвоночных и одного вида оболочников (tunicate). У рыб этот ген выделен у некоторых видов лосося и сома (Sherwood and col. (2000) The Origin and Function of the Pituitary Adenylate Cyclase-Activating olypeptide (PACAP)/Glucagon Superfamily Endocrine Reviews 21 (6):619-670, золотой рыбки (Leung y col. (1999) Molecular cloning and tissue distribution of in pituitary adenylate cyclase-activating polypeptide (PACAP) the goldfish. Rec. Progr. Mol. Comp. Endocrinol. 338-388), данио (Fradinger and Sherwood (2000) Characterization of the gene encoding both growth hormone-releasing hormone (GRF) and pituitary adenylate cyclase-activating polypeptide. Mol. and Cell. Endocrinol. 165:211-219) и форели (Krueckl and Sherwood. (2001) Developmental expression, alternative splicing and gene copy number for the pituitary adenylate cyclase-activating polypeptide (PACAP) and growth hormone-releasing hormone (GRF) gene in rainbow trout. Molec. and Cell. Endocrinol. 182:99-108). Патент США 5695954 защищает выделение и очистку генной последовательности нуклеотидов, которая кодирует полипептид GHRH-PACAP рыбы, так же как и векторы и клетку-хозяина, экспрессирующие эти последовательности с целью использования для увеличения роста рыб путем трансгенеза, введением упомянутых генных конструкций в оплодотворенные яйцеклетки рыб. Он также защищает способ выявления трансгенных рыб, которые содержат эти последовательности.
В настоящем патенте сообщаются специфические генные последовательности, кодирующие полипептид GHRH-PACAP видов Oncorhynchus Nerka, Clarias macrocephalus и Acispenser transmontanus.
В данном изобретении использованы различные варианты аминокислотной последовательности PACAP с модификациями на N-конце, полученные в лаборатории авторов данного изобретения для видов Clarias gariepinus и Oreochromis niloticus. Эти варианты применяли на водных организмах в качестве стимулятора роста не через трансгенез, а путем введения способом погружения в аквариум супернатанта культуры, экспрессирующей их в E. coli и P. pastoris, без предварительной их очистки. Неожиданно авторами было обнаружено, что эти варианты способны в таких условиях стимулировать существенное увеличение иммунологической активности у этих организмов и повышать концентрацию пролактина в сыворотке. Эти свойства пептида у водных организмов описаны не были.
Некоторые авторы сообщали об эффекте стимуляции роста у рыб введением рекомбинантного гормона роста путем погружения в аквариум. Тем не менее, прямое применение гормона роста является предметом множества требований регулирующих документов, то же самое касается применения трансгенных рыб, которые экспрессируют гормон роста или фактор, высвобождающий гормон роста.
В настоящем изобретении описана нетрансгенная методология повышения роста и усиления иммунной системы водных организмов, включая в себя беспозвоночных.
На современном этапе водные организмы являются важным источником белка, однако их добыча в естественных условиях используется в полной мере. По этой причине для увеличения продукции необходимо культивирование таких водных видов (Pullin y col.; Conference Proceeding 7, 432 p. International Center for living Aquatic Resources Monagement. Manila, Philippines. 1982, ISSN 0115-4389).
Повышение эффективности аквакультуры путем стимуляции роста, повышения выживаемости организмов и улучшения качества личинок - это постоянно существующая, важная, требующая решения проблема аквакультуры.
Сущность изобретения
Настоящее изобретение предоставляет решение упомянутой выше проблемы получения вариантов гипофизарного полипептида, активирующего аденилатциклазу, с аминокислотной последовательностью, обозначенной как SEQ ID No 12, 13 и 14, которые повышают скорость роста водных организмов, включая в себя беспозвоночные организмы, за короткий период, который очень важен для аквакультуры, кроме того, эти пептиды повышают выживаемость личинок рыб и ракообразных, представляющих коммерческий интерес, при погружении в аквариум или при использовании в качестве кормовой добавки. Они стимулируют иммунную активность этих организмов, так же как и потребность в пище, развитие окраски рыб и высвобождение пролактина.
В предпочтительном варианте осуществления настоящего изобретения варианты PACAP давали рыбам и ракообразным периодическими инъекциями с интервалом в 3 дня до концентрации 0,1 мкг/г веса тела животного, путем погружения в аквариум каждые 4 дня в пресную или морскую воду до концентрации пептида между 100 и 200 мкг/на литр воды и как кормовую добавку - до концентрации 5 мг/кг комбикорма. Получены существенное повышение роста и превосходная иммунная активность. Аппликация вариантов PACAP дает преимущества вследствие их малого размера (5 кДа), что делает возможным его хорошую абсорбцию через кожу и слизистые этих организмов, когда эти вещества введены в аквариум, такое введение предоставляет преимущества в стоимости и простоте манипуляций с аквакультурой и с небольшим уровнем загрязнения, кроме того, механизм передачи сигнала PACAP запускается с активации аденилатциклазы, а не с активации гормона, и его активность в высвобождении гормона роста у млекопитающих, включая в себя млекопитающих, невелика - причина, по которой это изобретение выигрывает в общественном восприятии и встречает меньше требований регулирующих документов. Другие преимущества PACAP состоят в способности стимулировать врожденную и адаптивную иммунную активность у рыб и повышать сопротивляемость к инфицированию патогенными агентами.
При осуществлении настоящего изобретения варианты PACAP давали водным организмам, таким как тилапия Orechromis sp., сом Claria sp., лосось Salmon sp. и креветка Penaus sp.
В другом предпочтительном варианте осуществления настоящего изобретения варианты PACAP вводят рыбам и ракообразным для предупреждения или лечения инфицирования патогенными агентами.
Осуществление настоящего изобретения представляет препарат композиции для лечения рыб или ракообразных в культуре для стимуляции их роста и повышения их сопротивляемости заболеваниям, так же как и для профилактического или терапевтического лечения инфекции патогенными агентами, все это направлено на повышение продуктивности.
Краткое описание чертежей
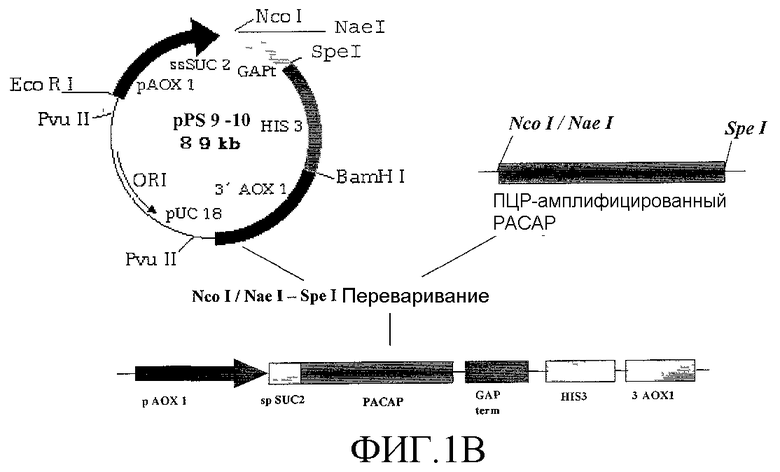
Фиг.1. Стратегия клонирования PACAP в бактериальном векторе экспрессии (Фиг.1A) и в векторе экспрессии дрожжей (Фиг.1B).
Фиг.2. Эксперимент по стимуляции роста в ювенильном Claria gariepinus путем интраперитонеальной инъекции рекомбинантного PACAP, очищенного с помощью аффинной хроматографии, до дозы 0,1 мкг/г веса животного. График представляет средние веса тела у группы, получавшей PACAP, по сравнению с контрольной группой.
Фиг.3. Эксперимент по стимуляции роста в ювенильном Claria gariepinus путем интраперитонеальной инъекции рекомбинантного PACAP, очищенного с помощью аффинной хроматографии, до дозы 0,1 мкг/г веса животного. График представляет средние гепатосоматического индекса и сухого веса мышц у группы, получавшей PACAP, по сравнению с контрольной группой.
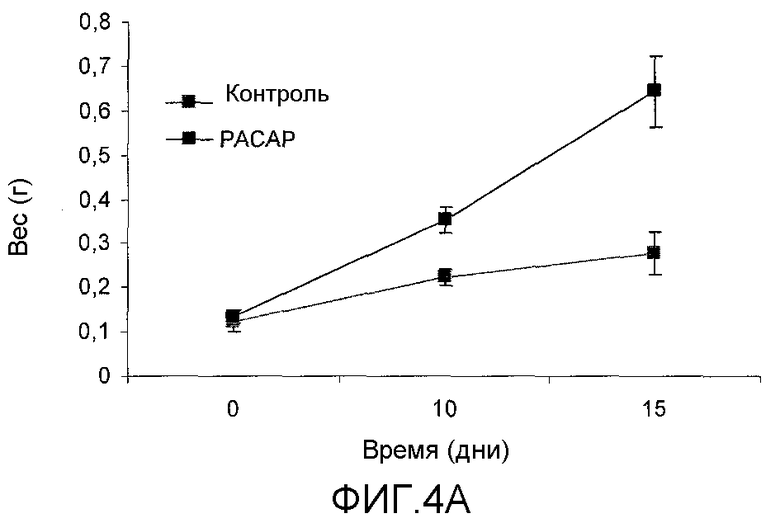
Фиг.4. Эксперимент по стимуляции роста у личинок тилапии при погружении в супернатант разрушенных E. coli, содержащий рекомбинантный PACAP в дозе 100 мкг/л воды. Графики 4A и 4B представляют средний вес тела и длину обработанных групп по сравнению с негативным контролем.
Фиг.5. Эксперимент по стимуляции роста у личинок тилапии при погружении в супернатант разрушенных E. coli, содержащий рекомбинантный PACAP в дозе 100 мкг/л воды. График представляет средний вес тела обработанных групп по сравнению с негативным контролем через 22 дня после начала обработки.
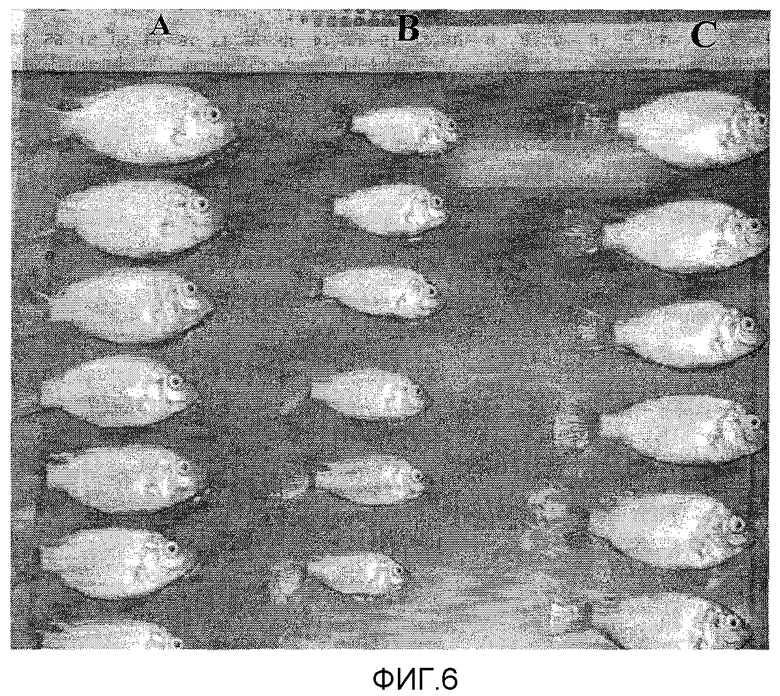
Фиг.6. Эксперимент по стимуляции роста у личинок тилапии при погружении в супернатант разрушенных E. coli, содержащий рекомбинантный PACAP в дозе 100 мкг/л воды. Чертеж показывает различие длины тела через 30 дней после последнего погружения рыб, обработанных PACAP (A и C) по отношению к контрольной группе (B).
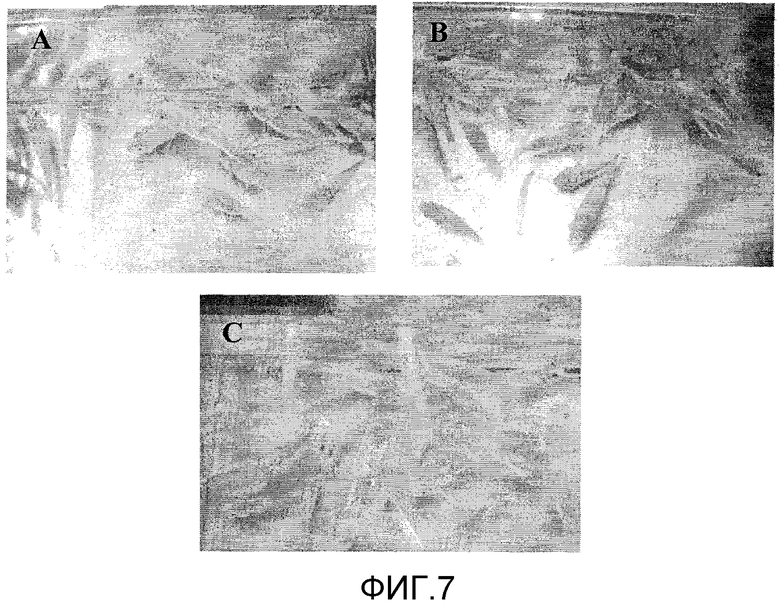
Фиг.7. Эксперимент по стимуляции роста у личинок тилапии при погружении в супернатант разрушенных E. coli, содержащий рекомбинантный PACAP в дозе 100 мкг/л воды. Чертеж показывает раннее развитие окраски у рыб, обработанных PACAP (A и B) по отношению к контрольной группе (C).
Фиг.8. Оценка влияния рекомбинантного PACAP, очищенного с помощью аффинной хроматографии, в дозе 0,5 мкг/г веса животного на потребность в пище у тилапии Orechromis niloticus. Чертеж показывает среднее количество пищи, потребленной рыбой через 6 часов и 22 часа после начала обработки.
Подробное описание конкретных вариантов осуществления / Примеры
Пример 1. Конструкция векторов экспрессии, содержащих последовательности, кодирующие PACAP, для внутриклеточной экспрессии в E. coli и внеклеточной продукции в супернатант культуры P. pastoris.
Ген РАСАР Claria gariepinus был выделен с помощью полимеразной цепной реакции с применением к ДНК GHRH-PACAP в качестве образца, предварительно клонированного в T-вектор. Авторы данного изобретения применяли специфические олигонуклеотиды, соответствующие последовательностям SEQ ID No 1 и SEQ ID No 2 для получения полной последовательности GHRH-PACAP, включающей в себя последовательность сигнального пептида, и специфические олигонуклеотиды SEQ ID No 3 и SEQ ID No 4 для амплификации только гена PACAP с рестрикционным сайтом, необходимым для клонирования в векторе экспрессии E. coli.
Ген PACAP у тилапии был выделен образом, сходным тому, как это описано выше, с применением специфических олигонуклеотидов SEQ ID No 3 и SEQ ID No 4. Настоящее изобретение представляет собой первое сообщение о выделении этого гена у тилапии.
Последовательность, кодирующая PACAP, была клонирована в векторе экспрессии pAR 3040 E. coli с использованием сайтов рестрикции Ndel и BamHI (Фиг.1A). Авторы отобрали один из них для рекомбинантной плазмиды для трансформации бактерии E.coli BL21D3, чтобы индуцировать экспрессию PACAP под управлением промотора T7 с использованием в качестве индуктора 0,5 мМ изопропил-d-тиогалактопиранозида (IPTG).
Экспрессию гена проводили при 28°С в течение 5 часов. Экспрессия рекомбинантного PACAP и его сохранность подтверждена масс-спектрометрией. Для экспрессии PACAP в P. pastoris авторы использовали векторы экспрессии дрожжей pPS9 и pPS10. Авторы использовали специфические олигонуклеотиды SEQ ID No 7 и SEQ ID No 6 для клонирования гена pPS9 и олигонуклеотиды SEQ ID No 5 и SEQ ID No 6 для клонирования pPS10. Авторы использовали для клонирования pPS7 сайты рестрикции NcoI и SpeI, такой подход к клонированию приводит к присоединению к интересующему белку метионина и глицина на N-конце. Для клонирования pPS10 авторы использовали сайты рестрикции NaeI и SpeI, такая стратегия клонирования не приводит к присоединению аминокислот к интересующему белку (Фиг.1B).
Перед трансформацией плазмиды линеаризовали с помощью фермента Sph I. Штамм MP36 Pichia pastoris трансформировали с помощью электропробоя мембраны с рекомбинантным вектором экспрессии. Этот штамм представляет собой ауксотрофный мутант his3, который приобретает после трансформации фенотип His+.
Идентифицированные с помощью Dot Blot трансформанты также анализировали с помощью сазерн-блота для определения того, в каких из них наблюдается интеграция плазмиды путем замещения гена AOX1 P. pastoris на полигенный экспрессирующий кластер рекомбинантной плазмиды. Это событие интеграции приводит к формированию фенотипов Muts (с низким уровнем утилизации метанола) и His+. Генетическое замещение AOX1 происходит путем рекомбинации между промоторными участками AOX1 и 3'AOX1 в векторе и геноме. В результате рекомбинации происходит делеция в кодирующем участке AOX1. Рекомбинантные штаммы с фенотипом Muts поддерживают продукцию алкогольоксидазы геном AOX2 и имеют низкий темп роста в метаноле.
Гены, кодирующие полипептиды, представляющие интерес, и гормон роста тилапии регулируются промотором AOX1, индуцируемым метанолом, и имеют один сигнальный пептид. Pichia pastoris имеет низкий уровень секреции собственных белков, а ее среда культивирования не нуждается в добавлении белков. Поэтому можно ожидать, что секретируемый гетерологический белок составит большой процент от общего белка в среде (более чем 80%) (Tschopp y col.; Bio/Technology 1987, 5: 1305-1308; Barr et al.; Pharm. Eng. 1992, 12: 48-51). Продукция рекомбинантных белков, описываемая в данном изобретении, выполнялась в биореакторах объемом 5 л с добавлением в культуру метанола.
Пример 2. Эксперимент по стимуляции роста ювенильного Claria gariepinus, определение гепатосоматического индекса и сухого веса мышц рыбы.
Авторы использовали 18 сомов вида Clarias gariepinus, без различения пола, приблизительно одного и того же возраста со средним весом тела 30-40 граммов. Были сформированы две экспериментальные группы, каждая из девяти особей. Группы были акклиматизированы в раздельных емкостях при постоянной рециркуляции воды к температуре 28°С и фотопериодичности 14 часов света и 10 часов темноты. Животных кормили 2 раза в день одинаковыми рационами, составляющими 5% общего веса тел в каждом танке. Животных перед экспериментом идентифицировали. Одну из групп обрабатывали полуочищенным (чистота 70%) SEQ ID No 13, тогда как другая, использовавшаяся как контрольная группа, получала белки E.coli, содержащиеся в растворе фосфатного буфера (PBS) 1X (белки E. coli получали такой же процедурой очистки пептида, представляющего интерес, в количестве, эквивалентном загрязнениям, присутствующим в очищенном образце PACAP). Рыбы, обрабатываемые PACAP, получали интраперитонеальные инъекции в дозе 0,1 мкг пептида на грамм веса тела животного 2 раза в неделю. Контрольную группу инъецировали сходным с описанным выше образом. Через 22 дня после начала эксперимента животные, которым инъецировали в брюшную полость PACAP, демонстрировали существенное увеличение веса тела (p<0,05) по сравнению с негативным контролем (Фиг.2).
Гепатосоматический индекс и сухой вес мышц определяли, чтобы продемонстрировать, что увеличение веса тела рыб происходит не вследствие увеличения веса внутренних органов или увеличения содержания воды в мышцах. Существенных различий между значениями гепатосоматических индексов и сухого веса мышц в экспериментальных группах не наблюдалось (Фиг.3).
Сходные результаты были получены при использовании рекомбинантного PACAP с последовательностью SEQ ID No 12.
Пример 3. Эксперимент по стимуляции роста, резистентности к патогенным агентам и высвобождения пролактина у личинок тилапии с помощью погружения в аквариум супернатанта разрушенных E. сoli, содержащих рекомбинантный PACAP.
Авторы выполнили эксперимент по оценке функции рекомбинантного PACAP из Claria gariepinus, присутствующего в супернатанте разрушенных E. сoli, на рост личинок тилапии.
Сформировали две экспериментальные группы из 60 особей в каждой, одна из групп была обработана нейропептидом PACAP (SEQ ID No 13), а другая группа использовалась как контрольная. Группы личинок акклиматизировались в отдельных емкостях при постоянной рециркуляции воды к температуре 28°С с фотопериодичностью 14 часов света и 10 часов темноты, и животные получали корм в количестве, вычисленном по нижеследующему уравнению: Количество корма = количество животных × средний вес тела (г) × 40%/100. Обработка состояла в погружении в аквариум 2 л воды три раза в неделю в течение 60 мин в течение 20 дней, доза составляла 200 мкг белка, представляющего интерес/литр воды. В результате через 10 дней после начала эксперимента в обработанной PACAP группе получено достоверное увеличение веса и длины тела по сравнению с контрольной группой (p<0,01), через 15 дней после начала эксперимента различие между группами было еще более достоверным (p<0,001) (Таблица 1 и Фиг.4A и 4B). Через 20 дней после начала погружения в аквариумы различие между группой, обработанной PACAP, и контрольной группой было статистически достоверным (p<0,001) (Фиг.5).
Средние веса и длины тела личинок тилапии через 10 дней и 15 дней с начала эксперимента
Было обнаружено, что эффект PACAP на рост сохраняется со временем, поскольку через 30 дней после последнего погружения в аквариум различия веса и длины тела в экспериментальных группах животных оставались вполне достоверными (p<0,01) (Фиг.6). Кроме того, было обнаружено, что обработанные PACAP рыбы демонстрируют более развитую окраску кожи на стадии раннего развития по сравнению с негативным контролем (Фиг.7).
В данном эксперименте авторы также изучали присутствие кожного простейшего Trichodina sp., было выбрано в случайном порядке по 10 животных из каждой экспериментальной группы, и определяли интенсивность заражения патогенным агентом. Величину интенсивности заражения определяли в соответствии с уравнением:
(I: # общее число паразитов у рыб) I=N/n-F0 и E=n-F0×100/n
I: (средняя интенсивность заражения)
E: (# число рыб с паразитами от общего числа)
N: (общее число обнаруженных паразитов)
F0: (число незараженных рыб)
n: (число проверенных рыб)
Рыбы, обработанные PACAP, демонстрируют достоверно более низкую интенсивность заражения (среднее I=2,20) простейшим Trichodinas sp. (p<0,01) по отношению к контрольной группе (среднее I=5,56).
Рыб обрабатывали через 45 дней с начала эксперимента в таких же условиях, как и описаны выше, и через 24 часа после обработки получали кровь от 10 особей из каждой группы для измерения пролактина в сыворотке с помощью вестерн-блота и http://www.multitran.ru/c/m.exe?a=sa&t=3256842_1_2&sc=517 твердофазного иммуноферментного анализа (ELISA). Для этих исследований использовали поликлональные антитела к пролактину тилапии. Наблюдали статистически достоверные различия между группой, обработанной PACAP, и контрольной группой p<0,01 (Таблица 2). Это очень привлекательный результат у коммерческих водных организмов, поскольку это касается лососевых, чей жизненный цикл распределен между пресноводным и морским периодами и у которых пролактин выполняет важную функцию в осморегуляции.
Концентрация пролактина (нг/мл) в сыворотке тилапии к 45-му дню с начала эксперимента
* показывает наличие достоверной разницы p<0,01
Пример 4. Эксперимент по оценке эффекта рекомбинантного PACAP на потребность в пище у ювенильных тилапий Orechromis niloticus.
До настоящего времени биологические эффекты PACAP на потребность в пище у рыб не были исследованы. У позвоночных, относящихся к таксонам, низшим, чем млекопитающие, активность этих пептидов в сфере потребности в пище охарактеризована мало (Jensen, 2001, Regulatory peptides and control of food intake in non-mammalian vertebrates. Comp. Biochem. And Phisiol. Part A 128:471-479).
Чтобы проанализировать эффект PACAP на потребность в пище у рыб, были использованы тилапии вида Oreochromis niloticus без различения пола и с приблизительно одинаковым средним весом тела. Были сформированы три экспериментальные группы, каждая из 3 особей в 3 повторах на группу. Группы акклиматизировали в отдельных емкостях при постоянной рециркуляции воды к температуре 28°С и фотопериодичности 14 часов освещения и 10 часов темноты.
Одну из групп обрабатывали полуочищенным PACAP (чистота 87%) SEQ ID No 13 путем интраперитонеальной инъекции 0,5 мкг/г веса тела животного. Вторую группу обрабатывали GHRP-6 (фирма Lipotec, S.A., Испания) таким же путем введения дозы 0,1 мкг пептида на грамм веса тела животного. Контрольную группу обрабатывали белками E. coli, содержащимися в PBS 1X (белки E. coli получали такой же процедурой очистки пептида, представляющего интерес, в количестве, эквивалентном загрязнителям, присутствующим в образце очищенного PACAP).
После обработок одинаковые количества корма давали трем экспериментальным группам, собирали остаток непотребленного корма через 6 часов и добавляли корм снова. Потребность в пище снова определяли через 22 часа после начала эксперимента. Непотребленный корм из каждой емкости высушивали в печке (100°С в течение 24 ч) и взвешивали на аналитических весах. Потребление пищи рассчитывали, определяя разницу между весом корма, добавленного в емкости (10 граммов с 20% влажностью), и кормом, непотребленным рыбами.
Тилапии, обработанные PACAP и GHRP-6, демонстрировали достоверное увеличение потребления пищи (p<0,05) по сравнению с контрольной группой (Фиг.8).
Пример 5. Оценка рекомбинантного PACAP в иммунной системе сома Claria gariepinus.
Использовали ювенильных Claria gariepinus. Были сформированы две группы из 10 особей в каждой группе. Группы акклиматизировали в отдельных емкостях при постоянной рециркуляции воды к температуре 28°С и фотопериодичности 14 часов освещения и 10 часов темноты. Животных кормили 2 раза в день рационами, эквивалентными 5% общего веса тел животных в каждой емкости. Животных перед экспериментом идентифицировали. Рыбы, обрабатывавшиеся PACAP (SEQ ID No 13), получали интраперитонеальные инъекции в дозе 0,1 мкг пептида на грамм веса тела животного 2 раза в неделю.
Через двадцать дней после начала эксперимента получали кровь рыб для измерения уровней лизоцима и лектина в сыворотке. Активность лизоцима в сыворотке измеряли с использованием способа, основанного на способности лизоцима лизировать бактерии Micrococcus lysodeikticus. В 96-луночном микропланшете 100 мкл образца в четырех последовательных разведениях в двух повторах в фосфатном буфере (0,05 M, pH 6,2) смешивали с 100 мкл суспензии 3 мг/мл Micrococcus lysodeikticus (фирма Sigma). Микропланшет инкубировали при 22°С и определяли оптическую плотность (O.D.) на длине волны 450 нм через 0, 2, 3, 5, 10, 15, 25, 35 и 45 мин. В качестве позитивного контроля сыворотку рыб заменяли на лизоцим белка из куриного яйца (последовательное разведение, начиная с 8 мкг/мл), а в качестве негативного контроля сыворотку заменяли на буфер. Единица активности лизоцима была определена как количество гомогената личинок, вызывающее снижение величины O.D. на 0,001 мин-1. Наблюдалась статистически достоверная разница (p<0,01) между группами, обработанными PACAP, по сравнению с контрольной группой (Таблица 3).
Концентрация лизоцима (мкг/мл) в сыворотке рыб через 20 дней после начала эксперимента
* указывает достоверную разность p<0,01
Чтобы определить присутствие лектина в сыворотке крови, было выполнено исследование гемагглютинации. Последовательные разведения сыворотки в двух повторах делали с использованием PBS pH 7,2 круглодонных микротитровальных лунках (96 лунок, фирма Greiner, Microlon), в которые добавляли равный объем свежеприготовленной 2% суспензии эритроцитов (кроличьих в PBS). Лунки инкубировали в течение 1 ч при комнатной температуре и титр определяли визуально - он был равен тому разведению, которое было в последней лунке, в которой наблюдалась агглютинация (что определяется равномерно распределенным слоем клеток по всему дну лунки). Проверяли активность гемагглютинации в образцах и для каждого из них определяли значение титра. Активность выражали как титр, т.е. обратную величину высшего разведения, демонстрирующего полную агглютинацию.
Рыбы, обработанные PACAP, демонстрируют достоверное повышение уровня лектина в сыворотке по сравнению с контрольной группой (p<0,05) (Таблица 4).
Титр активности гемагглютинина (обратная величина к высшему разведению, демонстрирующему полную агглютинацию) в сыворотке рыб на 45 день с начала эксперимента
Пример 6. Эксперимент по стимуляции роста личинок тилапии путем погружения в супернатант культуры P. рastoris, содержащий рекомбинантный PACAP.
Эксперимент выполняли с целью оценки функции PACAP из Clarias gariepinus (SEQ ID No 14), содержащегося в супернатанте культуры P. рastoris, при росте личинок тилапии.
Были сформированы три экспериментальные группы, каждая из 50 личинок. Одна из групп была обработана рекомбинантным PACAP (SEQ ID No 14), содержащимся в супернатанте культуры P. pastoris. Вторая группа была обработана рекомбинантным гормоном роста тиляпии (GH), содержащимся в супернатанте культуры P. pastoris. Контрольная группа была обработана супернатантом нетрансформированной культуры P. pastoris. Личинок кормили дважды в день, количество корма получали из нижеследующего уравнения: Количество корма = # число животных × средний вес тела (г) × 40%/100. Обработку выполняли с помощью погружения в объем 30 л три раза в неделю на 90 мин. Доза составляла 100 мкг представляющего интерес белка/литр воды.
Получен результат, согласно которому через 5 недель (35 дней) после начала эксперимента группа, обработанная PACAP, демонстрирует достоверное увеличение веса тела по сравнению с контрольной группой (p<0,01). Группа, обработанная гормоном роста, демонстрирует достоверное увеличение веса тела по сравнению с контрольной группой (p<0,05) (Таблица 5).
Вес личинок тилапии в граммах
Пример 7. Эксперимент по стимуляции роста и улучшению качества личинок креветки Litopenaeus schmitti, обработанных супернатантом культуры Pichia pastoris, содержащей рекомбинантный PACAP.
Использовали личинок креветки вида Litopenaeus schmitti. Были сформированы две экспериментальные группы из 100 личинок каждая. Одна группа была обработана рекомбинантным PACAP (SEQ ID No 14), содержащимся в супернатанте культуры P. pastoris, а другую, использовавшуюся в качестве контрольной группы, обрабатывали супернатантом нетрансформированной культуры P. pastoris.
Личинки культивировали в фиберглассовых емкостях объемом в 100 л. Корм основывался на диатомеях (Chaetoceros gracilis), жгутиковых водорослях (Tetraselmis suecica) и Artemia nauplius (фирма Aquatic Eco-Systems Inc.). Абиотические факторы роста были нижеследующими:
Освещение (24:00 свет/темнота (L/D)).
Постоянная аэрация.
Соленость 34 промилле.
Растворенный кислород 5,2 ± 0,5 (в личиночном цикле).
Замена при рециркуляции PZIII 80%.
Использовали четыре аквариума для погружения экспериментальных групп длительностью в 1 час 1 раз каждые 3 дня.
Получен следующий результат, что группа, обработанная PACAP, демонстрирует достоверное повышение веса тела по сравнению с контрольной группой (p<0,01) (Таблица 6).
Вес личинок креветки в миллиграммах
В группе, обработанной PACAP, наблюдались высокая однородность и лучшее качество личинок (с более развитым ветвлением жабр и ростральными модификациями), что очень важно в разведении креветки. Различие в выживаемости на стадии PL9 было более 40% в группе, обработанной PACAP.
Пример 8. Стимуляция роста ювенильных Clarias gariepinus путем включения рекомбинантного PACAP в формулу диеты рыб.
Супернатант культуры Pichia pastoris, содержащий рекомбинантный PACAP (SEQ ID No 14), концентрировали и вводили в питательную диету рыб в концентрации приблизительно 5 мг/кг корма.
Были сформированы две экспериментальные группы по 100 личинок в каждой со средним весом тела 0,1 г. Одна группа была обработана рекомбинантным PACAP (SEQ ID No 14), содержащимся в супернатанте культуры P. pastoris, а другую, использовавшуюся как контрольную, обрабатывали супернатантом нетрансформированной культуры P. pastoris. Эксперимент проводили в течение 30 дней.
Рекомбинантный PACAP (SEQ ID No 14), включенный в диету в дозе 5 мг/кг корма, увеличивал рост на 30% по сравнению с контрольной группой с высокой степенью статистической достоверности разницы (p<0,01).
Пример. Эффекты синтетического РАСАР С.gariepinus на гуморальную врожденную иммунную систему у мальков Salmo salar.
Эксперименты с выращиванием проводили с использованием мальков атлантического лосося (0,2±0,03 г; n=1000 на группу). Перед экспериментом группы в течение одной недели подвергали акклиматизации в емкостях объемом 500 л с проточной свежей водой при 12°С. Животных кормили до сытости два раза в сутки.
Синтетический РАСАР С.gariepinus вводили с помощью ванн для погружения два раза в неделю в течение 5 недель в дозировке 100 мкг на литр воды. Было сформировано две экспериментальные группы, в одной группе вводили РАСАР (SEQ ID No 14), а другую группу использовали в качестве контрольной группы.
Уровень лизоцима в гомогенатах мальков рыб (двадцать пулов по пять животных в каждом) измеряли на 14 и 28 сутки, когда индуцируемое РАСАР увеличение массы тела было статистически значимым. Данные выражали в качестве среднего значения ± стандартное отклонение (SD).
В группе с обработкой РАСАР было показано статистически значимое повышение концентрации лизоцима по сравнению с контрольной группой как через 14, так и через 28 суток после введения. Введение считали значимо отличающимся при p<0,05.
Пример. Эффекты РАСАР на иммунную систему креветок Penaeus.
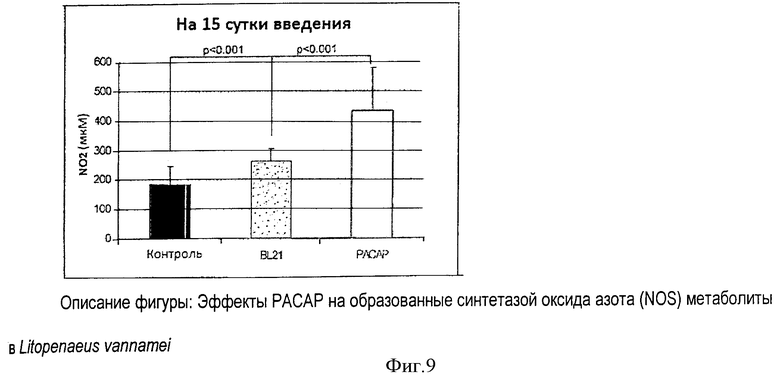
Эксперимент проводили в Litopenaeus vannamei на постличиночной стадии (0,021±0,002 г) по массе (n=250 на экспериментальную группу). Перед экспериментом креветок в течение одной недели подвергали акклиматизации в емкостях объемом 80 л, по одной емкости для каждой экспериментальной группы, содержащих проточную свежую воду. Их кормили без ограничений два раза в сутки. Схема введения включала три экспериментальные группы. Перед введением емкости очищали с использованием сифона и количество воды снижали до 2 л. Затем добавляли 10 мл очищенного рекомбинантного РАСАР (SEQ No 13) до конечной концентрации в воде, составляющей 100 мкг на литр (группа введения РАСАР). Каждое введение проводили в течение 90 мин без рециркуляции воды. В группе отрицательного контроля вводили количества белков E. coli, эквивалентные введению РАСАР (группа названа как группа BL21). Авторы изобретения включили другую контрольную группу, в которой не проводили введение, с единственным воздействием, заключавшимся в снижении уровня воды до объема 2 л (группа названа контрольной группой). Введение проводили три раза в неделю в течение четырех недель. Забор образцов (десять пулов по пять животных в каждом) для тестирования образованных синтетазой оксида азота (NOS) метаболитов и пектинов проводили на 15 сутки после начала эксперимента. Введение считали значимо отличающимся при p<0,05. Данные выражали в качестве арифметического среднего значения ± SD (фиг.9 и 10).
Пример. Эффекты перорального введения PACAP C.gariepinus на гуморальную приобретенную иммунную систему незрелых тиляпий.
Эксперименты по выращиванию проводили с использованием мальков нильской тиляпии (Oreochromis niloticus) (2±0,05 г; n=500 на группу). Перед экспериментом группы в течение одной недели подвергали акклиматизации в емкостях объемом 500 л с проточной свежей водой при 28°С. Животных кормили (два раза в сутки) питанием в количестве, вычисленном из следующего уравнения: Количество корма = # животных × средняя масса тела (г) × 40%/100. РАСАР С.gariepinus (SEQ ID No 12) вводили в рацион рыб в течение четырех недель в дозе 0,25 мг/кг корма. Группу с нормальным кормом (без PACAP) использовали в качестве группы отрицательного контроля.
Иммуноглобулин М (lgM) в сыворотке рыб (12 животных на группу) измеряли, когда индуцируемое PACAP увеличение массы тела было статистически значимым.
Данные выражали в качестве среднего значения ± стандартное отклонение (SD).
В группе с введением РАСАР было показано статистически значимое повышение общей концентрации lgM в сыворотке по сравнению с группой отрицательного контроля на 21 сутки.
Введение считали статистически отличающимся при p<0,05.





| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ РАСАР В КАЧЕСТВЕ МОЛЕКУЛЯРНОГО АДЪЮВАНТА ДЛЯ ВАКЦИН | 2012 |
|
RU2580294C2 |
| ПРИМЕНЕНИЕ РАСАР ДЛЯ ЛЕЧЕНИЯ ВИРУСНЫХ ИНФЕКЦИЙ У ВОДНЫХ ОРГАНИЗМОВ | 2011 |
|
RU2567802C2 |
| АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ДЛЯ КОНТРОЛЯ ПАТОГЕНОВ | 2012 |
|
RU2636001C2 |
| СОЕДИНЕНИЯ, ПРЕДСТАВЛЯЮЩИЕ СОБОЙ ПЕПТИДНЫЕ АНАЛОГИ СТИМУЛЯТОРОВ СЕКРЕЦИИ ГОРМОНА РОСТА, И СОДЕРЖАЩИЕ ИХ ПРЕПАРАТЫ | 2007 |
|
RU2416618C2 |
| ПРИМЕНЕНИЕ ПЕПТИДА, УСИЛИВАЮЩЕГО СЕКРЕЦИЮ СОМАТОТРОПНОГО ГОРМОНА, В КАЧЕСТВЕ АДЪЮВАНТА ВАКЦИНЫ | 2017 |
|
RU2768134C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ДЛЯ БОРЬБЫ С ЗАРАЖЕНИЯМИ ЭКТОПАРАЗИТАМИ | 2011 |
|
RU2585226C2 |
| МУТАНТНЫЕ ПОЛИПЕПТИДЫ СУАА И ПРОИЗВОДНЫЕ ПОЛИПЕПТИДОВ, ПОДХОДЯЩИЕ ДЛЯ ДОСТАВКИ ИММУНОГЕННЫХ МОЛЕКУЛ В КЛЕТКУ | 2010 |
|
RU2585216C2 |
| АГОНИСТЫ РЕЦЕПТОРА-3(R3) ПЕПТИДА-АКТИВАТОРА ГИПОФИЗАРНОЙ АДЕНИЛАТЦИКЛАЗЫ(РАСАР) И СПОСОБЫ ИХ ФАРМАКОЛОГИЧЕСКОГО ПРИМЕНЕНИЯ | 2000 |
|
RU2269354C2 |
| ЭМУЛЬГИРОВАННАЯ ВАКЦИНА ДЛЯ ПОЛУЧЕНИЯ КОНЦЕНТРИРОВАННЫХ КОМПОЗИЦИЙ ИММУНОГЛОБУЛИНОВ IgY; СПОСОБЫ И ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2689386C2 |
| ФРАГМЕНТЫ АНТИТЕЛА, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ ЧЕЛОВЕЧЕСКОГО РАКОВО-ЭМБРИОНАЛЬНОГО АНТИГЕНА (CEA) | 2003 |
|
RU2294939C2 |







Изобретение относится к области биотехнологии. Способ повышения сопротивляемости рыб или ракообразных заболеваниям включает кормление или введение рыбам или ракообразным эффективного количества нейропептида РАСАР с последовательностями, соответствующими SEQ ID No 12, SEQ ID No 13 и SEQ ID No 14. Применение вариантов нейропептида приводит к повышению выживаемости и усилению иммунной активности водных организмов. 2 н. и 11 з.п. ф-лы, 10 ил., 8 табл.
1. Способ повышения сопротивляемости рыб или ракообразных заболеваниям, который включает кормление указанных рыб или ракообразных эффективным количеством нейропептида РАСАР с последовательностями, соответствующими SEQ ID No 12, SEQ ID No 13 и SEQ ID No 14, или введение рыбам или ракообразным эффективного количества нейропептида РАСАР с последовательностями, соответствующими SEQ ID No 12, SEQ ID No 13 и SEQ ID No 14, причем указанный способ является пригодным для применения в аквакультуре.
2. Способ по п.1, в котором нейропептид РАСАР получают путем химического синтеза.
3. Способ по п.1, в котором нейропептид РАСАР получают с помощью рекомбинантной технологии.
4. Способ по п.3, в котором нейропептид РАСАР, содержащийся в супернатанте разрушенных Е. coli, используют без очистки.
5. Способ по п.3, в котором нейропептид РАСАР, содержащийся в супернатанте культуры Р. pastoris, используют без очистки.
6. Способ по п.3, в котором нейропептид РАСАР используется в очищенном виде после получения в рекомбинантных системах продукции.
7. Способ по п.1, в котором нейропептид РАСАР вводят рыбам или ракообразным путем периодических инъекций каждые 3 дня в концентрации 0,1 мкг/г веса животного.
8. Способ по п.1, в котором нейропептид РАСАР доставляется рыбам или ракообразным путем погружения в аквариум с интервалами 1-4 дня в пресной воде или в морской воде в концентрации от 100 до 200 мкг РАСАР на литр воды.
9. Способ по пп.1-8, в котором нейропептид РАСАР доставляется тилапии Orechromis sp.
10. Способ по п.п.1-8, в котором нейропептид РАСАР доставляется сому Claria sp.
11. Способ по п.п.1-8, в котором нейропептид РАСАР доставляется Salmon sp.
12. Способ по п.п.1-8, в котором нейропептид РАСАР доставляется креветке Penaus sp.
13. Применение нейропептида РАСАР в соответствии с последовательностями SEQ ID No 12, SEQ ID No 13 и SEQ ID No 14 в аквакультуре для повышения сопротивляемости рыб и ракообразных заболеваниям.
| ЕР 1477181 А1, 17.11.2004 | |||
| Экономайзер | 0 |
|
SU94A1 |

