Настоящее изобретение относится к способам индукции летальности раковых клеток, в частности раковых клеток, которые являются дефектными по репарации двухнитевых разрывов (double strand break, DSB, ДНР) ДНК, зависимой от гомологической рекомбинации (homologous recombination, HR).
Эффективная репарация повреждений ДНК в клетках основана на механизме обнаружения повреждения с последующей передачей сигналов нарушения к эффекторам, расположенным далее по пути передачи сигнала (downstream effectors), которые останавливают клеточный цикл в контрольных точках и устраняют нарушение ДНК. Клетки содержат ряд различных сигнальных путей и эффекторов, которые опосредуют репарацию различных типов повреждений ДНК. Эти пути включают эксцизионную репарацию оснований (base excision repair, BER), репарацию двухнитевых разрывов (ДНР) ДНК, зависимую от гомологической рекомбинации (HR), негомологичное соединение концов (non-homologous end joining), эксцизионную репарацию нуклеотидов (nucleotide excision repair, NER) и репарацию неправильно спаренных оснований (mismatch repair, MMR). Взаимодействие и взаимозависимость между различными путями репарации ДНК до сих пор до конца не известны.
Авторы настоящего изобретения установили, что ингибирование эксцизионной репарации оснований, BER, например, путем ингибирования полиаденозиндифосфорибозы-(ADP-ribose)-полимеразы (PARP) является селективно летальным для раковых клеток, дефектных по репарации двухнитевых разрывов ДНК, зависимой от HR. Это имеет большое значение для лечения раковых заболеваний.
Одним из аспектов настоящего изобретения является применение ингибитора эксцизионной репарации оснований для получения медикамента, предназначенного для лечения ракового заболевания у индивидуума,
при котором раковые клетки являются дефектными по активности репарации двухнитевых разрывов ДНК, зависимой от HR.
Способ лечения рака у индивидуума может включать
введение указанному индивидууму ингибитора эксцизионной репарации оснований,
при этом указанный рак является дефектным по пути репарации двухнитевых разрывов ДНК, зависимой от HR.
Рак может содержать одну или несколько раковых клеток, обладающих пониженной способностью или утративших способность устранять повреждения ДНК указанным вторым путем репарации по сравнению с нормальными клетками.
Путь репарации двухнитевых разрывов ДНК, зависимой от HR, устраняет двухнитевые разрывы (double-strand breaks, DSB) в ДНК посредством гомологичного механизма восстановления непрерывной спирали ДНК (К.К.Khanna и S.P.Jackson, Nat. Genet. 27(3): 247-254 (2001)). Компоненты пути репарации двухнитевых разрывов ДНК, зависимой от HR, включают ATM (NM_000051), RAD51 (NM_002875), RAD51L1 (NM_002877), RAD51C (NM_002876), RAD51L3 (NM_002878), DMC1 (NM_007068), XRCC2 (NM_005431), XRCC3 (NM_005432), RAD52 (NM_002879), RAD54L (NM_003579), RAD54B (NM_012415), BRCA1 (NM_007295), BRCA2 (NM_000059), RAD50 (NM_005732), MRE11A (NM_005590) и NBS1 (NM_002485). Другие белки, участвующие в пути репарации двухнитевых разрывов ДНК, зависимой от HR, включают регуляторные факторы, в частности, EMSY (Hughes-Davies et al. Клетка, Vol 115, pp 523-535).
Путь эксцизионной репарации оснований (base excision repair, BER) устраняет однонитевые разрывы ДНК и участки отсутствия нескольких оснований (зазоры, gaps), а также удаляет специфические поврежденные основания. Зазоры в спирали ДНК заполняются последовательным действием полиаденозиндифосфорибозы-(ADP-ribose)-полимеразы (PARP) и лигазы (К.К.Khanna и S.P.Jackson, Nat. Genet., 27(3): 247-254 (2001); F.Dantzer et al. Biochemistry 39, 7559-69 2000; J.H.Hoeijmakers, Nature 411 366-74 (2001)). Ингибитор эксцизионной репарации оснований может ингибировать любой из компонентов пути эксцизионной репарации оснований. Компоненты пути BER включают: UNG (NM_003362), SMUG1 (NM_014311), MBD4 (NM_003925), TDG (NM_003211), OGG1 (NM_002542), MYH (NM_012222), NTHL1 (NM_002528), MPG (NM_002434), NEIL1 (NM_024608), NEIL2 (NM_145043), NEIL3 (NM_018248), APE1 (NM_001641), APE2 (NM_014481), LIG3 (NM_013975), XRCC1 (NM_006297), ADPRT (PARP1) (NM_0016718) и ADPRTL2 (PARP2) (NP_005475).
Ингибиторы BER можно использовать в сочетании с агентом, повреждающим ДНК, при лечении раковых заболеваний, которые связаны с дефектом репарации двухнитевых разрывов ДНК, зависимой от HR. Агент, повреждающий ДНК, предпочтительно используют в такой дозировке или в таком составе, которые не являются летальными для клеток при отсутствии ингибитора BER. Пригодные химиотерапевтические агенты, повреждающие ДНК, описаны ниже.
В некоторых предпочтительных вариантах реализации можно использовать ингибитор фермента млекопитающих полиаденозиндифосфорибоза-полимеразы (PARP) (D'Amours et al., (1999) Biochem. J. 342: 249-268). Таким образом, ингибитор PARP можно использовать для лечения рака, который является дефектным по репарации ДНР ДНК, зависимой от HR.
Способ лечения рака, клетки которого являются дефектными по репарации ДНР ДНК, зависимой от HR, у индивидуума может включать:
введение ингибитора PARP указанному индивидууму.
Ингибитор PARP может быть использован для получения медикамента, применяемого для лечения рака у индивидуума, при этом указанный рак является дефектным по репарации ДНР ДНК, зависимой от HR.
Ингибиторы PARP более подробно описаны ниже.
Раковое заболевание, которое является дефектным по репарации ДНР ДНК, зависимой от HR, может содержать или включать одну или несколько раковых клеток, которые имеют пониженную способность к репарации ДНР ДНК этим путем или полностью утратили эту способность по сравнению с нормальными клетками, т.е. активность пути репарации ДНР ДНК, зависимой от HR, может быть понижена или утрачена у одной или нескольких раковых клеток.
Активность одного или нескольких компонентов пути репарации ДНР ДНК, зависимой от HR, может быть утрачена у одной или нескольких раковых клеток индивидуума, страдающего раковым заболеванием, которое является дефектным по репарации ДНР ДНК, зависимой от HR. Компоненты пути репарации ДНР ДНК, зависимой от HR, хорошо описаны специалистами в данной области (см., например, Wood et al. (2001) Science 291 1284-1289) и включают компоненты, перечисленные выше.
В некоторых предпочтительных вариантах реализации настоящего изобретения раковые клетки могут иметь дефектный фенотип BRCA1 и/или BRCA2, т.е. в раковых клетках активность BRCA1 и/или BRCA2 может быть снижена или утрачена. Раковые клетки с таким фенотипом могут быть дефектными по BRCA1 и/или BRCA2, т.е. экспрессия и/или активность BRCA1 и/или BRCA2 в раковых клетках может быть снижена или утрачена, например, путем мутации или полиморфизма кодирующей нуклеиновой кислоты, или посредством мутации или полиморфизма гена, кодирующего регуляторный фактор, например, гена EMSY, который кодирует регуляторный фактор BRCA2 (Hughes-Davies et al. Cell, Vol 115, pp.523-535).
BRCA1 и BRCA2 являются известными суппрессорами опухолей, аллели дикого типа которых часто утрачены в опухолях гетерозиготных носителей (Jasin M. Oncogene. 2002 Dec 16; 21(58): 8981-93; Tutt et al. Trends Mol Med. (2002) 8(12): 571-6). Связь мутаций BRCA1 и/или BRCA2 с раком молочной железы подробно описана в литературе (Radice P J Exp Clin Рак Res. 2002 Sep; 21 (3 Suppl): 9-12). Также известно, что амплификация гена EMSY, который кодирует фактор, связующий ВРСА2, ассоциирована с раком молочной железы и раком яичников.
Кроме того, носители мутаций в BRCA1 и/или BRCA2 имеют повышенные риски возникновения рака яичника, простаты и поджелудочной железы.
В некоторых вариантах реализации раковое заболевание индивидуума может быть предварительно идентифицировано как рак, который является дефектным по репарации ДНР ДНК, зависимой от HR.
В других вариантах реализации способ согласно настоящему описанию может содержать операцию идентификации ракового заболевания у индивидуума как дефектного по репарации ДНР ДИК, зависимой от HR.
Рак можно идентифицировать как раковое заболевание, дефектное по репарации ДНР ДНК, зависимой от HR, например, путем определения активности пути репарации ДНР ДНК, зависимой от HR, в одной или нескольких раковых клетках образца, полученного от индивидуума, или путем определения активности одного или нескольких компонентов указанного пути. Активность можно определить по отношению к нормальным (т.е. нераковым) клеткам, предпочтительно из той же самой ткани.
Активность пути репарации ДНР ДНК, зависимой от HR, можно определить, измеряя образование фокусов, содержащих Rad51 в ядре, в ответ на действие повреждающих ДНК агентов, или ингибиторов PARP. В клетках, дефектных на пути репарации ДНР ДНК, зависимой от HR, не такие фокусы не образуются. Присутствие фокусов Rad51 можно определить при помощи стандартных иммунофлуоресцентных способов. Другие способы определения активности пути репарации ДНР ДНК, зависимой от HR, могут включать определение чувствительности к инфракрасному излучению, средствам химиотерапии, в частности реагентам, способствующим образованию межнитевых поперечных связей, агентам, индуцирующим ДНР (ингибиторы топоизомеразы I и II), а также применение вестерн-блоттинга, иммуногистологического анализа, анализа хромосомных аномалий, анализа ферментного связывания или связывания ДНК и анализа с использованием плазмид.
В некоторых вариантах реализации рак можно идентифицировать как дефектный по пути репарации ДНР ДНК, зависимой от HR, по наличию в раковых клетках индивидуума одного или нескольких видоизменений, например полиморфизмов или мутаций в нуклеиновой кислоте, кодирующей полипептид, который представляет собой компонент пути репарации ДНР ДНК, зависимой от HR.
Видоизменения последовательности, в частности мутации и полиморфизмы, могут включать делецию, вставку или замещение одного или нескольких нуклеотидов по сравнению с последовательностью нуклеотидов дикого типа. В некоторых вариантах реализации видоизменение может представлять собой амплификацию гена, например амплификацию гена EMSY (CAD22881; символ гена C11ORF30). Одно или несколько видоизменений может иметь место в кодирующей или некодирующей области последовательности нуклеиновых кислот и может снижать или устранять экспрессию или функцию полипептида - компонента пути репарации ДНР ДНК, зависимой от HR. Иными словами, видоизмененная нуклеиновая кислота может кодировать видоизмененный полипептид, у которого уменьшена или утрачена активность, или может кодировать полипептид дикого типа, который экспрессируется на низком уровне или не экспрессируется в клетке, например, из-за изменения активности регуляторного элемента. Видоизмененная нуклеиновая кислота может иметь одну, две, три, четыре или более мутаций или полиморфизмов по сравнению с последовательностью дикого типа.
Наличие одного или нескольких видоизменений в нуклеиновой кислоте, которая кодирует компонент пути репарации ДНР ДНК, зависимой от HR, можно определить по наличию в одной или нескольких клетках контрольного образца кодирующей последовательности нуклеиновой кислоты, которая содержит одну или несколько мутаций или полиморфизмов, или по наличию видоизмененного полипептидного компонента, который кодируется указанной последовательностью нуклеиновой кислоты.
Существуют различные способы определения наличия или отсутствия в образце, полученном от индивидуума, конкретной последовательности нуклеиновой кислоты, например последовательности нуклеиновой кислоты, которая содержит мутацию или полиморфизм, уменьшающие или подавляющие экспрессию или активность компонента пути репарации ДНР ДНК, зависимой от HR. Кроме того, выполнив секвенирование нуклеиновой кислоты индивидуума или образца, можно сохранить информацию и проводить последующие исследования без использования самой исходной нуклеиновой кислоты. Так, например, при сканировании базы данных по последовательностям нуклеиновых кислот с помощью программного обеспечения для анализа последовательностей можно определить наличие изменения или мутации последовательности.
Способы согласно некоторым аспектам настоящего изобретения могут включать определение связывания олигонуклеотидного зонда с нуклеиновой кислотой, полученной из образца, например, с геномной ДНК, РНК или кДНК. Зонд может содержать последовательность нуклеотидов, которая специфически связывается с последовательностью нуклеиновой кислоты, содержащей одну или несколько мутаций или полиморфизмов, и не связывается специфически с последовательностью нуклеиновых кислот, не содержащей одной или нескольких мутаций или полиморфизмов, или наоборот.
Олигонуклеотидный зонд может содержать метку, при этом связывание зонда можно определить по наличию указанной метки.
Способ может включать гибридизацию одного или нескольких (например, двух) олигонуклеотидных зондов или праймеров с получения нуклеиновой кислоты. Если нуклеиновая кислота представляет собой двухнитевую ДНК, гибридизации обычно предшествует денатурация, обеспечивающая получение однонитевой ДНК. Гибридизация может быть частью процедуры полимеразно-цепной реакции (ПЦР) или частью процедуры зондирования без применения ПЦР. Примером процедуры является комбинация ПЦР и гибридизации в условиях пониженной специфичности.
Связывание зонда с целевой нуклеиновой кислотой (например, с ДНК) можно определить при помощи множества способов, которые находятся в распоряжении специалистов в данной области техники. Так, например, зонды могут иметь радиоактивную, флуоресцентную или ферментную метку. Другие способы, не использующие мечение зонда, включают исследования полиморфизма длины рестрикционных фрагментов, амплификацию с помощью ПЦР, расщепление РНК-азой и аллельспецифическое олигонуклеотидное зондирование. Зондирование может использовать стандартный способ саузерн-блоттинга. Так, например, ДНК можно экстрагировать из клетки и расщепить с различными рестрикционными ферментами. Затем можно разделить рестрикционные фрагменты с помощью электрофореза на агарозовом геле, произвести их денатурацию и перенести на фильтр из нитроцеллюлозы. Меченый зонд можно гибридизовать с фрагментами ДНК на фильтре и определить связывание.
Специалисты в данной области техники могут использовать пригодные условия желаемой специфичности реакции для селективной гибридизации с учетом таких факторов, как длина олигонуклеотидов, состав оснований, температура и т.п.
Пригодные условия для селективной гибридизации олигонуклеотидов, содержащих 17-30 оснований, включают гибридизацию в течение ночи при 42°С в 6Х SSC и отмывку в 6Х SSC при последовательном повышении температуры от 42°С до 65°С.
Другие пригодные условия и протоколы описаны в работах Molecular Cloning: a Laboratory Manual: 3rd edition, Sambrook & Russell (2001) Cold Spring Harbor Laboratory Press NY и Current Protocols in Molecular Biology, Ausubel et al. eds. John Wiley & Sons (1992).
Нуклеиновая кислота, которая может представлять собой геномную ДНК, РНК, кДНК или их амплифицированную область, может быть секвенирована для идентификации или определения наличия полиморфизма или мутации. Полиморфизм или мутация могут быть идентифицированы путем сравнения секвенированной последовательности с последовательностью компонента (пути репарации), полученной из базы данных, как описано выше. В частности, можно определить присутствие одного или нескольких полиморфизмов или мутаций, которые вызывают изменение или потерю функции полипептидного компонента и, следовательно, пути репарации ДНР ДНК, зависимой от HR, в целом.
Секвенирование может быть выполнено одним из стандартных способов. Секвенирование амплифицированного продукта может включать, например, осаждение изопропанолом, ресуспендирование и секвенирование с помощью набора для секвенирования TaqFS+ Dye terminator. Дополнительные продукты могут быть подвергнуты электрофорезу в секвенаторе ДНК ABI 377, а данные - проанализированы с помощью программного обеспечения Sequence Navigator.
Для амплификации области, представляющей интерес, в последовательности нуклеиновых кислот, например, части последовательности, в которой предполагается наличие мутаций или полиморфизмов, можно использовать специфическую реакцию амплификации, в частности, ПЦР с использованием одной или нескольких пар праймеров. Затем можно секвенировать амплифицированную нуклеиновую кислоту, как описано выше, и/или проанализировать ее каким-либо иным способом, чтобы определить наличие или отсутствие мутации или полиморфизма, которые уменьшают или полностью подавляют экспрессию или активность компонента пути репарации ДНР ДНК, зависимой от HR.
Пригодные реакции амплификации включают полимеразно-цепную реакцию (ПЦР) (см., например, в "PCR protocols; A Guide to Methods and Applications", Eds. Innis et al., 1990, Academic Press, New York, Mullis et al. Cold Spring Harbor Symp. Quant. Biol, 51: 263, (1987), Ehrlich (ed), PCR technology, Stockton Press, NY, 1989, и Ehrlich et al. Science, 252: 1643-1650, (1991)).
В некоторых вариантах реализации рак может быть идентифицирован как дефектный по репарации ДНР ДНК, зависимой от HR, путем определения уровня экспрессии или активности положительного или отрицательного регулятора компонента пути репарации ДНР ДНК, зависимой от HR, в частности, EMSY. Уровни экспрессии можно определить, например, с помощью вестерн-блоттинга, ELISA (твердофазного иммуноферментного анализа), RT-PCR (ревертазо-полимеразно-цепной реакции), гибридизации нуклеиновой кислоты или кариотипического анализа.
В некоторых предпочтительных вариантах реализации индивидуум является гетерозиготным по одному или нескольким видоизменениям, в частности, мутациям и полиморфизмам, в BRCA1 и/или BRCA2 или в их регуляторе. Способ обнаружения видоизменения в BRCA1 и BRCA2 хорошо известен специалистам в данной области техники и описан, например, в ЕР 699754, ЕР 705903, Neuhausen S.L. and Ostrander E.A. Genet. Test (1992) 1, 75-83; Chappnis, P.O. and Foulkes, W.D, Cancer Treat Res (2002) 107, 29-59; Janatova M et al. Neoplasma. 2003: 50(4): 246-50; Jancarkova N Ceska Gynekol. 2003 68(1): 11-6). Определение амплификации связующего фактора BRCA2 EMSY описали Hughes-Davies et al. Cell 115 523-535.
Мутации и полиморфизмы, связанные с раком, также могут быть обнаружены на уровне белка по присутствию вариантного (например, мутантного или аллельного варианта) полипептида.
Способ идентификации раковой клетки в образце, полученном у индивидуума, как дефектной по репарации ДНР ДНК, зависимой от HR, может включать контактирование образца со специфическим связывающим агентом, направленным против вариантной формы (например, мутантного варианта) полипептидного компонента пути, и определение связывания специфического связывающего агента с образцом. Связывание специфического связывающего агента с образцом может быть показателем присутствия видоизмененного полипептидного компонента пути репарации ДНР ДНК, зависимой от HR, в клетке образца.
Предпочтительные специфические связывающие молекулы для применения в аспектах настоящего изобретения включают антитела и их фрагменты или их производные ("антителоподобные молекулы").
Реакционную способность связывающего агента, в частности антитела по отношению к нормальным и опытным образцам, можно определить любыми пригодными средствами. Одна из возможностей заключается в мечении индивидуальными репортерными группами. Репортерные группы могут прямо или косвенно генерировать обнаруживаемые и предпочтительно измеряемые сигналы. Связь репортерных групп может быть прямой или косвенной, ковалентной, например посредством пептидной связи, или нековалентной. Соединение посредством пептидной связи может быть результатом экспрессии рекомбинантного гена, совместно кодирующего связующую молекулу (например, антитело) и репортерную группу.
Способ определения связывания не является предметом настоящего изобретения, и специалисты в данной области техники могут выбрать пригодный способ согласно их предпочтениям и общим знаниям.
Раковые клетки в общем случае отличаются от нормальных клеток аномальной пролиферацией и обычно образуют в организме индивидуума, страдающего раковым заболеванием, кластеры или опухоли.
Раковое заболевание, которое является дефектным по пути репарации ДНР ДНК, зависимой от HR, согласно данному описанию может включать любой тип плотного рака или злокачественной лимфомы, в частности лейкемию, саркому, рак кожи, рак мочевого пузыря, рак молочной железы, рак матки, рак яичника, рак простаты, рак легкого, рак ободочной и прямой кишки, рак шейки матки, рак печени, рак головы и шеи, рак пищевода, рак поджелудочной железы, рак почки, рак желудка и рак головного мозга. В некоторых предпочтительных вариантах реализации раковое заболевание может представлять собой рак молочной железы, яичника, поджелудочной железы или простаты. Раковые заболевания могут быть семейными или спорадическими.
Образец, полученный у индивидуума, может представлять собой образец ткани, содержащий одну или несколько клеток, например биопсию из раковой ткани, как описано выше, или нераковую ткань, например, для применения в качестве контрольного образца.
Способы согласно изобретению могут быть использованы для обследования индивидуума, страдающего раковым заболеванием, например, для определения терапевтического лечения. Способ обследования индивидуума, страдающего раковым заболеванием, может включать
идентификацию раковой клетки, полученной от индивидуума, как дефектной по репарации ДНР ДНК, зависимой от HR, по сравнению с нормальными клетками, и
выбор ингибитора пути BER, пригодного для введения указанному индивидууму.
В некоторых предпочтительных вариантах реализации ингибитор пути BER представляет собой ингибитор PARP. Ингибиторы PARP более подробно описаны ниже. Способ исследования ракового заболевания может включать
идентификацию раковой клетки, полученной у индивидуума, как дефектной по репарации ДНР ДНК, зависимой от HR, по сравнению с нормальными клетками, и
выбор ингибитора PARP, пригодного для введения указанному индивидууму.
В некоторых предпочтительных вариантах реализации раковая клетка, которая идентифицирована как дефектная по репарации ДНР ДНК, зависимой от HR, может иметь дефектный фенотип BRCA1 или BRCA2.
Индивидуум может иметь предрасположение к раку, который является дефектным по репарации ДНР ДНК, зависимой от HR. Способы и средства настоящего изобретения особенно полезны для таких индивидуумов.
Индивидуум может быть, например, гетерозиготным по мутации или полиморфизму нуклеиновой кислоты, кодирующей компонент пути репарации ДНР ДНК, зависимой от HR, например, нуклеиновой кислоты, кодирующей компоненты, описанные выше.
Способ лечения рака у индивидуума может включать
введение ингибитора пути BER указанному индивидууму,
при этом указанный индивидуум является гетерозиготным по мутации или полиморфизму в гене, кодирующем компонент пути репарации ДНР ДНК, зависимой от HR.
Ингибитор BER может быть использован для получения медикамента, применяемого для лечения рака у индивидуума, который является гетерозиготным по мутации в гене, кодирующем компонент пути репарации ДНР ДНК, зависимой от HR.
В некоторых предпочтительных вариантах реализации индивидуум, который является гетерозиготным по мутации или полиморфизму в гене, кодирующем компонент пути репарации ДНР ДНК, зависимой от HR, может быть гетерозиготным по мутации или полиморфизму в BRCA1 и/или BRCA2.
Ингибитор BER, пригодный для применения в описанном здесь способе, может представлять собой любое соединение или объект, в частности малую органическую молекулу, пептид или нуклеиновую кислоту, которые ингибируют, уменьшают или блокируют активность одного или нескольких компонентов пути BER.
В некоторых предпочтительных вариантах реализации ингибитор BER может уменьшать или блокировать активность фермента полиаденозиндифосфорибоза-полимеразы (PARP).
Термин PARP, используемый в настоящем описании, относится к PARP1 (ЕС 2.4.2.30, Genbank No: M32721) и/или PARP2 (Ame et al. J. Biol. Chem. (1999) 274 15504-15511; Genbank No: AJ 236912), если из контекста не следует иного.
Примеры соединений, которые являются известными ингибиторами PARP и которые могут быть использованы в соответствии с настоящим изобретением, включают:
1. Никотинамиды, в частности, 5-метилникотинамид и амидоксим O-(2-гидрокси-3-пиперидинопропил)-3-карбоновой кислоты, а также их аналоги и производные.
2. Бензамиды, включая 3-замещенные бензамиды, в частности, 3-аминобензамид, 3-гидроксибензамид, 3-нитрозобензамид, 3-метоксибензамид и 3-хлорпрокаинамид, 4-аминобензамид, 1,5-ди[(3-карбамоилфенил)аминкарбонилокси]-пентан, а также их аналоги и производные.
3. Изохинолины и дигидроизохинолины, включая 2Н-изохинолин-1-оны, 3Н-хиназолин-4-оны, 5-замещенные дигидроизохинолины, в частности, 5-гидрокси-дигидроизохинолинон, 5-метилдигидроизохинолинон и 5-гидроксиизохинолинон, 5-аминоизохинолин-1-он, 5-дигидроксиизохинолинон, 3,4-дигидроизохинолин-1(2Н)-оны, в частности, 3,4-дигидро-5-метоксиизохинолин-1(2Н)-он и 3,4-дигидро-5-метил-1(2Н)изохинолинон, изохинолин-1(2Н)-оны, 4,5-дигидроимидазо[4,5,1-i,j]хинолин-6-оны, 1,6-нафтиридин-5(6Н)-оны, 1,8-нафталимиды, в частности, 4-амино-1,8-нафталимид, изохинолинон, 3,4-дигидро-5-[4-1(1-пиперидинил)бутокси]-1(2Н)-изохинолинон, 2,3-дигидробензо[де]изохинолин-1-он, 1-11b-дигидро-[2Н]бензопирано[4,3,2-де]изохинолин-3-он и тетрациклические лактамы, включая бензопираноизохинолины, в частности, бензопирано[4,3,2-де]изохинолинон, а также их аналоги и производные.
4. Бензоимидазолы и индолы, включая бензоксазол-4-карбоксамиды, бензоимидазол-4-карбоксамиды, в частности, 2-замещенные бензоксазол-4-карбоксамиды и 2-замещенные бензоимидазол-4-карбоксамиды, в частности, 2-арилбензоимидазол-4-карбоксамиды и 2-циклоалкилбензоимидазол-4-карбоксамиды, включая 2-(4-гидроксифенил)-бензоимидазол-4-карбоксамид, хиноксалинкарбоксамиды, имидазопиридинкарбоксамиды, 2-фенилиндолы, 2-замещенные бензоксазолы, в частности, 2-фенилбензоксазол и 2-(3-метоксифенил)-бензоксазол, 2-замещенные бензоимидазолы, в частности, 2-фенилбензоимидазол и 2-(3-метоксифенил)-бензоимидазол, 1,3,4,5-тетрагидроазепино[5,4,3-cd]индол-6-он, азепининдолы и азепининдолоны, в частности, 1,5-дигидроазепин[4,5,6-cd]индолин-6-он и дигидродиазепининдолинон, 3-замещенные дигидродиазепининдолиноны, в частности, 3-(4-трифторметилфенил)-дигидродиазепининдолинон, тетрагидродиазепининдолинон и 5,6,-дигидроимидазо[4,5,1-j,k][1,4]бензодиазепин-7(4Н)-он, 2-фенил-5,6-дигидро-имидазо[4,5,1-j,k][1,4]бензодиазепин-7(4Н)-он и 2,3-дигидроизоиндол-1-он, а также их аналоги и производные.
5. Фталазин-1(2Н)-оны и хиназолиноны, в частности, 4-гидроксихиназолин, фталазинон, 5-метокси-4-метил-1(2)фталазиноны, 4-замещенные фталазиноны, 4-(1-пиперазинил)-1(2Н)-фталазинон, тетрациклические бензопирано[4,3,2-де]-фталазиноны и тетрациклические инден-[1,2,3-де]фталазиноны и 2-замещенные хиназолины, в частности, 8-гидрокси-2-метилхиназолин-4-(3Н)-он, трициклические фталазиноны и 2-аминофталгидразид, а также их аналоги и производные.
6. Изоиндолиноны, а также их аналоги и производные.
7. Фенантридины и фенантридиноны, в частности, 5[Н]фенантридин-6-он, замещенные 5[Н]-фенантридин-6-оны, в частности, 2-, 3-замещенные 5[Н]-фенантридин-6-оны и сульфонамидные/карбамидные производные 6(5Н)фенантридинонов, тиен-[2,3-с]-изохинолоны, в частности, 9-аминтиен-[2,3-с]-изохинолон и 9-гидрокситиен-[2,3-с]-изохинолон, 9-метокситиен-[2,3-с]-изохинолон и N-(6-оксо-5,6-дигидрофенантридин-2-ил)-2-(N,N-диметиламин)-ацетамид, замещенные 4,9-дигидроциклопента[lmn]-фенантридин-5-оны, а также их аналоги и производные.
8. Бензопироны, в частности, 1,2-бензопирон, 6-нитрозобензопирон, 6-нитрозо-1,2-бензопирон и 5-йодо-6-аминобензопирон, а также их аналоги и производные.
9. Производные ненасыщенных гидроксамовых кислот, в частности, O-(3-пиперидино-2-гидрокси-1-пропил)-никотиновый амидоксим, а также их аналоги и производные.
10. Пиридазины, включая конденсированные пиридазины, а также их аналоги и производные.
11. Другие соединения, в частности кофеин, аминофиллин и тимидин, а также их аналоги и производные.
Дополнительные ингибиторы PARP описаны, например, в US 6,635,642, US 5,587,384, WO 2003080581, WO 2003070707, WO 2003055865, WO 2003057145, WO 2003051879, US 6514983, WO 2003007959, US 6426415, WO 2003007959, WO 2002094790, WO 2002068407, US 6476048, WO 2001090077, WO 2001085687, WO 2001085686, WO 2001079184, WO 2001057038, WO 2001023390, WO 2001021615, WO 2001016136, WO 2001012199, WO 9524379, Banasik et al. J. Biol. Chem., 267: 3, 1569-75 (1992), Banasik et al. Molec. Cell. Biochem. 138: 185-97 (1994), Cosi (2002) Expert Opin. Ther. Patents 12(7), Southan & Szabo (2003) Curr Med Chem 10 321-340 и в приведенных там ссылках.
Один из предпочтительных классов пригодных ингибиторов PARP включает фталазиноны, в частности, 1(2H)-фталазинон, и их производные, как описано в WO 02/36576. В частности, соединения формулы:
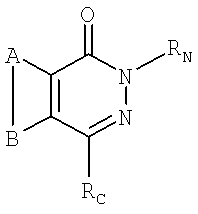
а также их изомеры, соли, сольваты, химически защищенные формы и пролекарства могут быть использованы для ингибирования PARP, где
А и В совместно представляют собой конденсированное ароматическое ядро, возможно, содержащее замещение,
RC представляет собой -L-RL, где L имеет формулу:
-(СН2)n1-On2-(СН2)n3-
где n1, n2 и n3 выбирают из группы, содержащей числа 0, 1, 2 и 3, таким образом, чтобы сумма n1, n2 и n3 была равна 1, 2 или 3, a Q выбирают из группы, включающей О, S, NH, С(=O) или -CR1R2-, где R1 и R2 независимо выбирают из группы, включающей водород, галоген или, возможно, содержащий замещение С1-7 алкил, или могут совместно с атомом углерода, к которому они присоединены, образовывать С3-7 циклическую алкильную группу, которая может быть насыщенной (С3-7 циклоалкильная группа) или ненасыщенной (С3-7 циклоалкенильная группа), или один из R1 и R2 может быть присоединен к атому RL, образуя ненасыщенную С3-7 циклоалкенильную группу, содержащую атомы углерода, к которым присоединены R1 и R2 в Q, -(СН2)n3- (если присутствует) и часть RL;
а RL представляет собой C5-20 арил, содержащий замещение, а
RN выбирают из группы, включающей водород, C1-7 алкил, возможно, содержащий замещение, С3-20 гетероциклил, и C5-20 арил, гидроксил, простой эфир, нитро, амин, амид, тиол, тиоэфир, сульфоксид и сульфон.
Предпочтительно соединение формулы:
или его изомер, соль, сольват, химически защищенную форму или пролекарство можно использовать для ингибирования PARP,
где
А и В совместно представляют собой конденсированное ароматическое ядро, возможно, содержащее замещение,
RC представляет собой -CH2-RL;
RL представляет собой фенил, возможно, содержащий замещение; а
RN представляет собой водород.
В некоторых предпочтительных вариантах реализации для ингибирования PARP можно использовать соединение, имеющее структуру KU-0058684 или KU-0058948, показанную на фигуре 2, или его изомер, соль, сольват, химически защищенную форму или пролекарство.
Пригодные ингибиторы PARP являются коммерчески доступными или могут быть синтезированы известными способами из исходных материалов, которые являются известными (см., например, Suto et al. Anticancer Drug Des. 6: 107-17 (1991)).
Другой класс ингибиторов эксцизионной репарации оснований включает пептидные фрагменты компонентов пути BER. Например, пептидные фрагменты последовательности PARP можно использовать для ингибирования PARP и тем самым понизить или подавить активность пути BER. Пептидные фрагменты можно генерировать полностью или частично с помощью химического синтеза, используя опубликованные последовательности компонентов, например опубликованную последовательность PARP (учетный номер: NM_001618). Пептидные фрагменты можно легко получить известными стандартными способами жидкостного или, предпочтительно, твердофазного пептидного синтеза, общие описания которых широко доступны (см., например, J.M.Stewart and J.D.Young, Solid Phase Peptide Synthesis, 2nd edition, Pierce Chemical Company, Rockford, Illinois (1984), M. Bodanzsky and A.Bodanzsky, The Practice of Peptide Synthesis, Springer Verlag, New York (1984); and Applied Biosystems 430A Users Manual, ABI Inc., Foster City, California), или их можно получить в растворе жидкофазным способом или комбинацией твердофазного способа, жидкофазного способа и химии растворов, например, получив вначале соответствующую пептидную часть, а затем, если желательно и требуется, после удаления присутствующих защитных групп ввести остаток Х путем реакции с соответствующей карбоновой или сульфоновой кислотой или ее реактивным производным.
Другие соединения-кандидаты для ингибирования компонента пути BER, в частности, PARP, могут быть получены на основании моделирования трехмерной структуры указанного компонента с использованием рационального строения препарата, чтобы обеспечить соответствующие характеристики формы, размера и заряда молекул соединений-кандидатов. Ингибитор-кандидат может быть, например, "функциональным аналогом" пептидного фрагмента или другого соединения, которое ингибирует указанный компонент. Функциональный аналог имеет такую же функциональную активность, что и пептид или другое соответствующее соединение, т.е. может препятствовать взаимодействию или подавлять активность компонента пути репарации ДНК. Примеры таких аналогов включают химические соединения, которые моделируют таким образом, чтобы обеспечить сходство с трехмерной структурой компонента в зоне, контактирующей с другим компонентом, и в особенности с расположением остатков ключевых аминокислот в соответствии с их видом.
Следующий класс пригодных ингибиторов пути HER включает кодирующую часть нуклеиновых кислот или всю последовательность аминокислот компонента пути BER, в частности, PARP (учетный номер: NM_001618), или их комплемент, которые ингибируют активность или функцию путем отрицательной (понижающей) регуляции производства активного полипептида.
Ингибирование активности PARP можно определить с помощью обычных способов, включая, например, дот-блоттинг (Affar ЕВ et al. Anal Biochem. 1998; 259(2): 280-3), и анализы BER, которые измеряют прямую активность PARP для формирования цепей полиаденозиндифосфорибозы, например, с помощью радиоактивных анализов с применением субстрата NAD, меченого тритием, или антител, специфических для полимерных цепей, сформированных активностью PARP (K.J.Dillon et al. Journal of Biomolecular Screening, 8(3): 347-352 (2003)).
Так, например, экспрессия компонента пути BER может быть ингибирована с помощью антисмыслового метода или интерференции РНК (RNAi). Применение таких подходов к понижающей регуляции генной экспрессии в настоящее время хорошо известно в данной области техники.
Антисмысловые олигонуклеотиды могут быть сформированы для гибридизации с комплементарной последовательностью нуклеиновой кислоты, пред-мРНК или зрелой мРНК, чтобы оказывать влияние на производство компонента пути эксцизионной репарации оснований таким образом, чтобы уменьшить его экспрессию или предотвратить ее полностью или практически полностью. Кроме обеспечения кодирующей последовательности антисмысловые способы могут быть направлены на воздействие на контрольные последовательности гена, например, во фланкирующей последовательности 5', при этом антисмысловые олигонуклеотиды могут блокировать функцию контролирующих экспрессию последовательностей. Конструкция антисмысловых последовательностей и их применение описаны, например, в работах Peyman and Ulman, Chemical Reviews, 90: 543-584, (1990) и Crooke, Ann. Rev. Pharmacol. Toxicol. 32: 329-376, (1992).
Олигонуклеотиды можно получить in vitro или ex vivo для последующего введения, или антисмысловую РНК можно получить in vivo в клетках, в которых желательна понижающая регуляция. Так, например, двухнитевую ДНК можно поместить под контролем промотора в "обратной ориентации" таким образом, чтобы транскрипция антисмысловой нити ДНК образовывала РНК, комплиментарную к нормальной мРНК, транскрибированной из смысловой нити целевого гена. Предполагают, что затем комплементарная антисмысловая последовательность РНК связывается с мРНК, образуя дуплекс, который ингибирует трансляцию эндогенной мРНК, транскрибированной с целевого гена в белок. До настоящего времени не установлено, как реально действует этот механизм. Однако функционирование такого механизма представляет собой подтвержденный факт.
Нет необходимости использовать полную последовательность, соответствующую кодирующей последовательности, в обратной ориентации. Например, можно использовать фрагменты достаточной длины. Отбор фрагментов различных размеров и из различных частей кодирующих или фланкирующих последовательностей гена для оптимизации уровня антисмыслового ингибирования является стандартной задачей для специалиста в данной области техники. При этом может оказаться предпочтительным включение инициирующего метионинового кодона ATG и, возможно, одного или нескольких нуклеотидов, расположенных за инициирующим кодоном. Пригодный фрагмент может содержать около 14-23 нуклеотидов, например около 15, 16 или 17.
Альтернативой антисмысловому способу является использование полной или частичной копии целевого гена, инсертированной в «смысловом» положении, т.е имеющей такую же ориентацию, как целевой ген, чтобы уменьшить экспрессию целевого гена путем косупрессии (Angell & Baulcombe (1997) The EMBO Journal 16, 12: 3675-3684; и Voinnet & Baulcombe (1997) Nature 389: pg 553). Оказалось, что двухнитевая РНК (днРНК) является еще более эффективной для подавления генов, чем смысловая и антисмысловая нити по отдельности (Fire A. et al. Nature 391, (1998)). Подавление посредством днРНК является генспецифическим и часто называется интерференцией РНК (РНКи).
Интерференция РНК представляет собой двухэтапный процесс. Вначале днРНК расщепляют в клетке, чтобы получить короткие интерферирующие РНК (киРНК) длиной примерно 21-23 nt с концевым фосфатом 5' и короткими выступами 3' (~2 nt).
киРНК специфически направлены на разрушение соответствующей последовательности мРНК (Zamore P.D. Nature Structural Biology, 8, 9, 746-750, (2001)).
РНКи можно также эффективно индуцировать с помощью химически синтезированных дуплексов киРНК той же самой структуры с выступающими концами 3' (Zamore PD et al. Cell, 101, 25-33, (2000)). Показано, что синтетические дуплексы киРНК специфически подавляют экспрессию эндогенных и гетерогенных генов в широком спектре клеточных линий млекопитающих (Elbashir SM. et al. Nature, 411, 494-498, (2001)).
Другая возможность состоит в применении нуклеиновой кислоты, которая при транскрипции вырабатывает рибозим, способный разрезать нуклеиновую кислоту в специфическом сайте, что также оказывает полезное влияние на экспрессию гена. Ссылки на рибозимы приведены в работах Kashani-Sabet and Scanlon, 1995, Cancer Gene Therapy, 2(3): 213-223, и Mercola and Cohen, 1995, Cancer Gene Therapy, 2(1), 47-59.
Способы согласно изобретению могут включать введение индивидууму ингибитора BER, в частности ингибитора PARP. Это можно производить после идентификации индивидуума как страдающего раковым заболеванием, клетки которого дефектны по репарации ДНР ДНК, зависимой от HR.
Активное соединение можно вводить отдельно, однако предпочтительно представлять его в форме фармацевтического состава (например, препарата), содержащего по меньшей мере одно активное соединение, как описано выше, вместе с одним или несколькими фармацевтически допустимыми носителями, адъювантами, связующими, разбавителями, наполнителями, буферами, стабилизаторами, консервантами, смазками или иными материалами, хорошо известными специалистам, а также, возможно, с другими терапевтическими или профилактическими агентами.
Фармацевтические составы, содержащие ингибиторы эксцизионной репарации оснований, как определено выше, например, ингибитор, смешанный с одним или несколькими фармацевтически допустимыми носителями, связующими, буферами, адъювантами, стабилизаторами или иными материалами, указанными в данном описании, могут быть использованы в способах согласно настоящему изобретению.
Термин "фармацевтически допустимые", используемый в данном описании, относится к соединениям, материалам, составам и/или лекарственным формам, которые согласно тщательной медицинской оценке являются пригодными для применения в контакте с тканями субъекта (например, человека) без чрезмерной токсичности, раздражения, аллергической реакции или других проблем или осложнений в соответствии с разумным соотношением между выгодой и риском. Кроме того, все носители, связующие и т.п. должны быть "допустимыми" с точки зрения совместимости с другими ингредиентами состава.
Пригодные носители, связующие и т.п можно найти в стандартной фармацевтической литературе, например Remington's Pharmaceutical Sciences, 18th edition. Mack Publishing Company, Easton, Pa., 1990.
Составы можно удобно представить в одинарной дозированной форме и получать любыми способами, хорошо известными специалистам в области фармацевтики. Такие способы включают операцию объединения активного соединения с носителем, который может содержать один или несколько вспомогательных ингредиентов. В общем случае препараты получают путем равномерного и тщательного перемешивания активного соединения с жидкими носителями, с тонко измельченными твердыми носителями или с теми и другими, а затем, если требуется, формуют продукт.
Препараты могут иметь форму жидкостей, растворов суспензий, эмульсий, эликсиров, сиропов, таблеток, лепешек, гранул, порошков, капсул, облаток, пилюль, ампул, свечей, вагинальных суппозиториев, мазей, гелей, паст, кремов, спреев, пен, лосьонов, масел, болюсов, электуариев или аэрозолей.
Ингибитор BER или фармацевтический состав, содержащий указанный ингибитор, можно вводить субъекту любым удобным способом, системно/периферически или в область желаемого действия, включая, в частности, но без ограничения, перорально, (например, путем глотания), топически (в том числе, трансдермально, интраназально, окулярно, трансбуккально и сублингвально), пульмонально (например, путем ингаляционной или инсуффляционной терапии, в частности, при помощи введения аэрозоля, например, через рот или нос), ректально, вагинально, парентерально, например, путем инъекции, в том числе подкожно, внутрикожно, внутримышечно, внутривенно, внутриартериально, внутрисердечно, интратекально, интраспинально, интракапсулярно, субкапсулярно, интраорбитально, интраперитонеально, интратрахеально, субкутикулярно, интраартикулярно, субарахноидально и интрастернально, а также путем имплантации депо, например, подкожно или внутримышечно.
Препараты, пригодные для перорального введения (например, путем глотания), могут быть представлены как дискретные дозированные формы, в частности капсулы, облатки или таблетки, содержащие определенную дозу активного соединения, а также в форме гранул, раствора или суспензии в водной или неводной жидкости, жидкой эмульсия типа "масло-в-воде" или "вода-в-масле", болюса, электуария или пасты.
Таблетку можно изготовить обычными способами, например прессованием или литьем, возможно, с добавлением одного или нескольких дополнительных ингредиентов. Прессованные таблетки можно получить путем прессования в соответствующем оборудовании активного соединения в сыпучей форме, в частности в форме порошка или гранул, возможно, смешанного с одним или несколькими связующими (например, с повидоном, желатином, гуммиарабиком, сорбитом, трагакантом, гидроксипропилметилцеллюлозой), с наполнителями или разбавителями (например, с лактозой, микрокристаллической целлюлозой, гидрофосфатом кальция), со смазками (например, со стеаратом магния, тальком, кремнеземом), с дезинтегрантами (например, с крахмалом гликолята натрия, сшитым повидоном, сшитой карбоксиметилцеллюлозой), с поверхностно-активными, диспергирующими или смачивающими агентами (например, с лаурилсульфатом натрия) и с консервантами (например, метил-р-гидроксибензоатом, пропил-р-гидроксибензоатом, сорбиновой кислотой). Литые таблетки можно получить с помощью соответствующего оборудования путем литья порошкообразного соединения, увлажненного инертным жидким разбавителем. Таблетки могут иметь покрытие или рифление, а их рецептура может обеспечивать замедленное или регулируемое выделение активного соединения за счет использования, например, гидроксипропилметилцеллюлозы в изменяющихся пропорциях, чтобы обеспечить желаемый профиль выделения. На таблетки можно нанести энтеросолюбильное покрытие, чтобы обеспечить частичное выделение в кишечнике, а не в желудке.
Препараты, пригодные для парентерального введения (например, путем инъекции, в том числе кожной, подкожной, внутримышечной, внутривенной и внутрикожной инъекции), включают водные и неводные изотонические, непирогенные стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, консерванты, стабилизаторы, бактериостаты и растворенные вещества, которые делают препарат изотоническим с кровью потенциального реципиента, а также водные и неводные стерильные суспензии, которые могут включать суспендирующие агенты и загустители, липосомы или иные системы микрочастиц, которые обеспечивают введение соединения в компоненты крови или в один или несколько органов. Примеры пригодных изотонических носителей, предназначенных для применения в таких препаратах, включают физиологический раствор для инъекций, раствор Рингера или лактат Рингера для инъекций. Обычно концентрация вышеуказанного соединения в растворе составляет примерно от 1 нг/мл до примерно 10 мкг/мл, в частности, примерно от 10 нг/мл до примерно 1 мкг/мл. Препараты могут быть представлены в герметичных контейнерах, например в ампулах и флаконах, и могут храниться в сублимированном (лиофилизированном) состоянии, которое требует непосредственно перед применением добавления только стерильного жидкого носителя, например воды для инъекций. Растворы и суспензии для инъекций, приготавливаемые непосредственно перед введением, можно получить из стерильных порошков, гранул и таблеток. Препараты могут иметь форму липосом или систем микрочастиц, которые обеспечивают введение соединения в компоненты крови или в один или несколько органов.
Следует понимать, что соответствующие дозы активных соединений и составы, содержащие активные соединения, могут изменяться для различных пациентов. Определение оптимального дозирования в общем случае включает нахождение баланса между уровнем терапевтической пользы и риска или вредных побочных эффектов от лечения согласно настоящему изобретению. Выбор уровня дозирования зависит от множества факторов, включая, в частности, но без ограничения, активность конкретного соединения, способ введения, время введения, скорость выделения соединения, длительность лечения, другие лекарства, соединения и/или материалы, применяемые в комбинации, а также возраст, пол и вес пациента, его состояние, общее здоровье и историю болезни. Доза соединения и способ введения в конечном счете определяются по усмотрению лечащего врача, однако в общем случае дозирование должно обеспечить получение локальных концентраций в области действия, которые создают желаемый эффект, не вызывая существенных вредных или опасных побочных эффектов.
Составы, содержащие ингибиторы пути BER, можно использовать в описанных здесь способах в сочетании со стандартными химиотерапевтическими режимами, которые повреждают ДНК раковой клетки. Пригодные агенты могут включать ингибиторы активности топоизомеразы I и II, в частности, камптотецин, лекарственные препараты, в частности, иринотекан, топотекан и рубитекан, алкилирующие агенты, в частности, темозоломид и DTIC (дакарбазин), а также платиновые агенты, например, цисплатин, цисплатин-доксорубицин-циклофосфамид, карбоплатин и карбоплатин-паклитаксел.
Другие пригодные химиотерапевтические агенты включают доксорубицин-циклофосфамид, капецитабин, циклофосфамид-метотрексат-5-фторурацил, доцетаксел, 5-фторурацил-эпирубицин-циклофосфамид, паклитаксел, винорелбин, этопосид, пегилированный липосомный доксорубицин и топотекан.
Лечение индивидуумов при помощи таких агентов хорошо известно специалистам в данной области медицины.
В рамках курса лечения введение препарата in vivo можно производить в форме одной дозы, непрерывно или периодически (например, раздельными дозами через соответствующие интервалы времени). Способы определения наиболее эффективных средств и доз для введения хорошо известны специалистам в данной области медицины и зависят от препарата, применяемого для лечения, цели лечения, целевой клетки, на которую оказывают воздействие, и субъекта, подлежащего лечению. Однократные или многократные введения могут производиться при уровне дозы и схеме лечения, выбранной лечащим врачом.
В общем случае пригодная доза активного соединения лежит в пределах примерно от 100 мкг до примерно 250 мг на килограмм массы тела субъекта в сутки. Если активное соединение представляет собой соль, сложный эфир, пролекарство и т.п., вводимую дозу рассчитывают на основе родоначального соединения, поэтому фактически применяемую дозу пропорционально увеличивают.
Способы согласно изобретению можно также использовать для исследования и оценки ракового заболевания у индивидуума.
Способ определения активности пути репарации ДНР ДНК, зависимой от HR, при раковом заболевании может включать:
контактирование ингибитора эксцизионной репарации оснований с образцом раковых клеток, полученным у индивидуума, страдающего заболеванием, и
определение уровня смертности клеток в указанном образце по сравнению с контрольным образцом.
Повышение уровня смертности клеток в образце по отношению к контрольным клеткам, которые имеют нормальные уровни активности репарации ДНР ДНК, зависимой от HR, указывает на то, что рак является дефектным по репарации ДНР ДНК, зависимой от HR.
Индивидуум может иметь раковое заболевание, а образец может представлять собой образец раковых клеток, полученный, например, с помощью биопсии опухоли.
В предпочтительных вариантах реализации ингибитор эксцизионной репарации оснований представляет собой ингибитор PARP. При этом способ оценки репарации ДНР ДНК, зависимой от HR, при раковом заболевании может включать:
контактирование ингибитора PARP с образцом раковых клеток, полученных у иднивидуума, страдающего раковым заболеванием, и
определение уровня клеточной смертности в указанном образце по отношению к контрольному образцу.
Повышенная чувствительность ингибитора PARP в клетках образца по сравнению с контрольными клетками указывает на то, что рак представляет собой недостаток активности репарации ДНР ДНК, зависимой от HR.
Повышенная чувствительность к ингибиторам PARP может указывать на то, что раковые клетки имеют дефектный фенотип BRCA1 или BRCA2, например уменьшение или подавление экспрессии или активности BRCA1 или BRCA2.
Раковое заболевание, идентифицированное как дефект активности репарации ДНР ДНК, зависимой от HR, например, заболевание, связанное с дефектным фенотипом BRCA1 или BRCA2, может быть подвергнуто терапии, специально направленной на такие заболевания. Пригодная терапия может включать применение агентов, сшивающих ДНК, в частности митомицина С, цисплатина или карбоплатина.
Определенные способы можно использовать для прогнозирования реакции ракового заболевания у индивидуума на лечение, направленное на HR, например, на лечение, специфическое для раковых заболеваний, имеющих дефектный фенотип BRCA1 или BRCA2.
Способ прогнозирования реакции ракового заболевания у индивидуума на лечение рака, связанного с дефектом репарации ДНР ДНК, зависимой от HR, может включать:
контактирование ингибитора BER, например ингибитора PARP, с образцом раковых клеток, полученным у пациента, страдающего раковым заболеванием, и
определения уровня клеточной смертности в указанном образце по сравнению с контрольным образцом.
Увеличение клеточной смертности в образце по сравнению с контрольными клетками, которые имеют нормальные уровни активности репарации ДНР ДНК, зависимой от HR (т.е. повышенная чувствительность к ингибиторам PARP), указывает на то, что рак может быть восприимчивым к указанному лечению.
Средства для лечения раковых заболеваний, клетки которых имеют дефект репарации ДНР ДНК, зависимой от HR, например, раковых заболеваний, связанных с дефектом BRCA1 или BRCA2, могут включать, например, агенты, сшивающих ДНК, в частности митомицин С, цисплатин или карбоплатин.
Другие аспекты изобретения относятся к применению ингибитора репарации ДНР ДНК, зависимой от HR, для лечения рака, который является дефектным по эксцизионной репарации оснований.
Способ лечения ракового дефекта эксцизионной репарации оснований у индивидуума может включать:
введение указанному индивидууму ингибитора пути репарации ДНР ДНК, зависимой от HR.
Ингибитор пути репарации ДНР ДНК, зависимой от HR, можно использовать для получения медикамента, предназначенного для лечения рака у индивидуума, при этом указанный рак является дефектным по эксцизионной репарации оснований.
Ингибитор пути репарации ДНР ДНК, зависимой от HR, может включать ингибитор одного или нескольких компонентов пути, как указано выше. Пригодные ингибиторы включают ингибиторы ATM.
Ингибитор ATM может представлять собой, например, соединение формулы I:
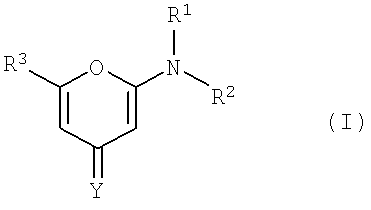
или его изомер, соль, сольват, химически защищенную форму или пролекарство, где
Y представляет собой О или S,
R1 и R2 представляют собой независимо водород, С1-7 алкильную группу, возможно, содержащую замещение, С3-20 гетероциклильную группу или C5-20 арильную группу, или могут совместно с атомом азота, к которому они присоединены, образовывать гетероциклическое ядро, возможно, содержащее замещение, и от 4 до 8 атомов в ядре, а R3 представляет собой фенильную группу, присоединенную к C5-20 карбоарильной группе, возможно содержащей замещение, с помощью эфирного или тиоэфирного мостика, при этом фенильная группа и, возможно, содержащая замещение C5-20 карбоарильная группа могут быть связаны другой мостиковой группой, которая связана соседним эфирным или тиоэфирным мостиком с обеими группами таким образом, чтобы образовывать C5-7 гетероцикл, возможно, содержащий замещение кислородом и серой, слитый с фенильной группой и C5-20 карбоарильной группой, а фенильная группа может также содержать замещения.
Более подробно такие ингибиторы описаны в патентной заявке WO 03/070726.
Другие ингибиторы включают пептидильные фрагменты компонентов репарации ДНР ДНК, зависимой от HR, и кодирующие нуклеиновые кислоты, как описано выше.
Раковое заболевание может быть идентифицировано с помощью вышеописанного способа как дефектное по активности BER.
Далее аспекты настоящего изобретения иллюстрируются с помощью ссылок на прилагаемые фигуры, описанные ниже, и экспериментальных примеров, которые не являются ограничивающими. Другие аспекты и варианты реализации будут очевидными для специалистов в данной области техники.
Различные параметры и характеристики изобретения описаны выше. Во избежание неопределенности заявляется, что все комбинации и субкомбинации этих параметров и характеристик включены в объем настоящего изобретения.
Все документы, упомянутые в данном описании, включены в него в качестве ссылки.
Фигура 1 показывает, что снижение уровня Parp1 уменьшает жизнеспособность мутантных клеток BRCA1 и BRCA2 по сравнению с клетками дикого типа.
Фигура 2 показывает ингибиторы PARP KU 0058684, KU 0058948 и KU 0051529 и значения их IC50 по отношению к ферментной активности PARP-1.
Фигуры 3 и 4 показывают кривые клоногенного выживания клеток под действием ингибиторов PARP.
Фигура 3 показывает Brca1 дикого типа (11СО:■), гетерозиготные (Cre6:▲) и дефектные (Cre10:●) клетки ES при непрерывном действии ингибиторов PARP (KU0058684, верх; KU 0058948, середина; KU 0051529, низ). Отрезки прямой показывают стандартную ошибку среднего значения.
Фигура 4 показывает Brca2 дикого типа (D3:■), гетерозиготные (Cre6:▲) и дефектные (Cre24:●) клетки ES при непрерывном действии ингибиторов PARP (KU 0058684, верх; KU 0058948, середина; KU 0051529, низ). Отрезки прямой показывают стандартную ошибку среднего значения.
Фигуры 5 и 6 показывают кривые клоногенного выживания после действия KU 0058684 в течение 1, 4 и 24 часов.
Фигура 5 показывает Brca1 дикого типа (11СО:■), гетерозиготные (Cre6:▲) и дефектные (Cre10:●) клетки ES после действия KU 0058684 в течение 1 (верх), 4 (середина) и 24 часов (низ). Отрезки прямой показывают стандартную ошибку среднего значения.
Фигура 6 показывает Brca2 дикого типа (D3:■), гетерозиготные (Cre6:▲) и дефектные (Cre24:●) клетки ES после действия KU 0058684 в течение 1 (верх), 4 (середина) и 24 часов (низ). Отрезки прямой показывают стандартную ошибку среднего значения.
Фигуры 7 и 8 показывают, что ингибирование PARP в мутантных клетках BRCA-1 и BRCA-2, обработанных ингибитором PARP, приводит к усилению ареста на этапе G2/M.
Фигура 7 показывает Brca1 дикого типа (11СО:верх) и мутантные (Cre10:низ) клетки, обработанные в течение 24 часов KU 0058684 с концентрацией 0 нМ (слева), 10 нМ (середина) или 1 мкМ (справа) и проанализированные с помощью FACS (fluorescent-activated cell sorter, клеточный сортер с возбуждением флуоресценции).
Фигура 8 показывает Brca2 дикого типа (D3) и мутантные (Cre24) клетки, обработанные в течение 24 часов KU 0058684 с концентрацией 0 нМ (слева), 10 нМ (середина) или 1 мкМ (справа) и проанализированные с помощью FACS.
Фигура 9 показывает результаты количественного анализа очагов Rad51, индуцированных ингибированием PARP в клетках дикого типа, но не в дефектных клетках Brca1 или Brca2.
Фигура 10 показывает результаты нейтрального гельэлектрофореза одиночных клеток (comet analysis) BRCA2-/- VC8 и BRCA2, комплементарных VC8-BAC. Обработка KU 0058684 (1 мкМ) в течение 30 часов индуцирует значительное увеличение ДНР ДИК, которое оценивается по увеличению хвостового момента в клетках BRCA2-/-, в то время как в комплементарной клеточной линии BRCA2 существенного увеличения хвостового момента не наблюдается. Усредненные данные 3 независимых опытов показаны с +/- SEM (Standard Error of Mean, стандартная ошибка среднего значения) при подсчете хвостового момента для 50 одиночных клеток в каждом опыте.
Фигура 11 показывает возможную модель селективного влияния ингибирования PARP на мутантные клетки BRCA1 и BRCA2.
Фигуры 12 и 13 показывают результаты анализа FACS фосфогистона Н3 для клеток ES.
Фигура 12 показывает результаты анализа FACS фосфогистона Н3 для Brca1 дикого типа (11СО:верх) и мутантных (Cre10:) клеток ES, а также для Brca2 дикого типа (D3) и мутантных (Cre24) клеток, обработанных в течение 24 часов KU 0058684 с концентрацией 0 нМ (слева), 10 нМ (середина) или 1 мкМ (справа) и проанализированных с помощью FACS.
Фигура 13 показывает результаты анализа FACS фосфогистона Н3 для клеток VC8 и VC8BAC, обработанных в течение 24 часов KU 0058684 с концентрацией 0 мкМ, 100 мкМ, 1 нМ и 10 нМ (слева направо соответственно).
Фигуры 14 и 15 показывают анализ влияния ингибирования PARP в других клетках с недостатком функции BRCA1 и BRCA2.
Фигура 14 показывает кривые клоногенного выживания Brca2 дефектных (V-C8:■) и комплементарных (V-C8 ВАС+:▲) клеток при непрерывном действии ингибиторов PARP (KU 0058684: верх, KU 0058948: середина, и KU 0051529: низ).
Фигура 15 показывает кривые клоногенного выживания Brca2 дефектных (V-C8:■) и комплементарных (V-C8 ВАС+:▲) клеток после действия KU 0058684 в течение 1 часа (верх), 4 часов (середина) и 24 часов (низ). Отрезки прямой показывают стандартную ошибку среднего значения.
Фигура 16 показывает образование опухоли в ксенотрансплантатах ES и влияние обработки KU 0058684; пунктирная линия - дикий тип с наполнителем, жирная сплошная линия - дикий тип с препаратом KU 0058684, сплошная линия - дефектные Brca2 с наполнителем, штриховая линия - дефектные по Brca2 с KU 0058684.
Фигура 17 показывает клональные кривые выживания клеток с BRCA1 дикого типа (МСF7-скремблированные) и клеток с подавленной экспрессией BRCA1 (MCF7-3.23) при непрерывном действии различных концентраций ингибитора PARP KU 0058684 в течение 12-14 дней. Показана зависимость логарифма доли выживания клеток от логарифма концентарции ингибитора. Отрезки прямой показывают стандартную ошибку среднего значения.
Фигура 18 показывает клональные кривые выживания клеток BRCA1 дикого типа (MCF7-скремблированные) и клеток с подавленной экспрессией BRCA1 (MCF7-3.23) при непрерывном действии различных концентраций ингибитора PARP KU 0051529 в течение 12-14 дней. Показана зависимость логарифма доли выживания клеток от логарифма концентрации ингибитора. Отрезки прямой показывают стандартную ошибку среднего значения.
Примеры
Материалы и методики
Интерференция РНК
Генноспецифические конструкции pSUPER (Т.R.Brummelkamp et al. Science 296, 550-3 (2002)) генерировали путем экспрессии следующих целевых последовательностей мРНК: (1) мышиная Parp1 5'-GCGGAGUACGCCAAGUCCA-3', (2) скремблированная контрольная 5'- CAUGCCUGAUCCGCUAGUC-3'.
Фрагмент 1,6 т.п.н., содержащий промотор CMV IE и eCFP (enhanced cyan fluorescent pritein, усовершенствованный циан-флуоресцентный белок) субклонировали из pECFP-Mito (Invitrogen) в сайте SapI генерированных конструкций pSUPER, получив pSUPER-eCFP-Parp1 и pSUPER-eCFP-control. Клетки ES D3 трансфицировали этими плазмидами при помощи липофектамина 2000 (Invitrogen) согласно инструкциям изготовителя. Через сорок восемь часов после трансфекции получили суммарные лизаты клеток с помощью буфера, содержащего 20 мМ трис-буфера с рН 8, 200 мМ NaCl, 1 мМ EDTA (этилендиаминтетрауксусная кислота), 0,5% (по объему) NP40, 10% (по объему) глицерина и ингибиторов протеазы. 30 мкг каждого лизата подвергли электрофорезу на бис-трис ацетатакриламидных готовых гелях (Novex) и блотировали на транс-блот-нитроцеллюлозу (Biorad). Блоты зондировали с поликлональным анти-PARP-1 антителом кролика (Cell Signalling, Cat No. 9542) или анти-GFP/CFP антисывороткой кролика (Invitrogen, Cat. No. R970-01), затем провели вторичную гибридизацию с анти-кроличьим IgG-HRP с последующим хемилюминесцентным анализом с помощью ECL™ (Amersham, UK).
Синтетические ингибиторы PARP
Ингибиторы PARP синтезировали согласно патентной заявке WO 02/36576. Химические ингибиторы растворили в DMSO (диметилсульфоксид) с концентрацией 10 мМ и хранили при -20°С в темноте.
Клеточные линии
Клетки VC8 и их производные, дополненные мышиным ВАС Brca2, соответствовали описанию в работе М.Kraakman-van der Zwet et al., Mol Cell Biol 22, 669-79 (2002). Клетки ES с дефектной функцией Brca2 были описаны ранее (Tutt et al. (2002) ЕМВО Rep 3, 255-60). Конструкция клеток ES с дефектом Brca1 будет описана в другом месте, однако ранее она была подтверждена (Foray et al. (2003) Embo J 22 2860-71). Клетки HBL100 трансфицировали плазмидой РНКи BRCA1 pSUPER и отбирали с помощью генетицина в течение 3 недель. Клоны отбирали на основе их экспрессии BRCA1 в соответствии с анализом назерн-блоттинг.
Клоногенные анализы
Для определения чувствительности к разрушению РНК Parp1 клетки ES, культивируемые в кюветах с тканевой культурой, покрытых 0,1% желатином, трансфицировали, как описано выше, pSVPER-eCFP-Parp1 или pSUPER-eCFP-control совместно с вектором, экспрессирующим резистентность к антибиотику бластицидину (pEF-Bsd, Invitrogen). Через двадцать четыре часа после трансфекции клетки обработали трипсином и посеяли в планшеты с 6 лунками. Через сорок восемь часов после трансфекции начали обработку бластицидином, и производили питание клеток один раз в три дня. Через 10-14 дней клетки промыли PBS (забуференный фосфатом физиологический раствор), фиксировали метанолом и окрасили кристаллвиолетом. Произвели подсчет колоний, содержащих более чем примерно 50 клеток.
Для определения чувствительности к химическим ингибиторам клеточные культуры в стадии экспоненциального роста обработали трипсином и посеяли с различной плотностью в планшеты с 6 лунками, содержащие фибробласты мышиных эмбрионов, дезактивированные митомицином С, и по мере необходимости обработали ингибиторами через 18 часов. Для непрерывного воздействия в клетки один раз в 4 дня добавляли свежую питательную среду и ингибитор. Для периодического воздействия ингибитор добавляли через определенный период времени, а затем клетки промывали и добавляли свежую питательную среду. Через 10-14 дней клетки промыли PBS, фиксировали метанолом и окрасили кристаллвиолетом. Произвели подсчет колоний, содержащих более чем примерно 50 клеток. Опыты проводили по меньшей мере три раза по три серии.
Анализ FACS
Для определения содержания ДНК клетки фиксировали в 70% этаноле, инкубировали с рибонуклеазой и йодидом пропидия (PI) и проанализировали с помощью FACS Calibur (Becton Dickinson). Для анализа фосфогистона Н3 клетки фиксировали 70% этанолом, пермеабилизовали 0,25% Тритоном Х-100, инкубировали с антителом против фосфорилированной формы гистона Н3 (Upstate Biotechnology) в течение 3 часов, а затем - с FITC (флуоресцинизотиоцианатом)-антикроличьим IgG (Serotec) в течение 30 минут. Анализ FACS проводили, как описано выше.
Анализ апоптоза
Клетки обработали трипсином, сохранив надосадочный слой культуры и промывную среду. Их объединили, клетки промыли в холодном PBS-A, а затем ресуспендировали при концентрации 1×106 клетки/мл в буфере для связывания (10 мМ HEPES (N-2-гидроксиэтилпиперазин-N-2-этансульфоновая кислота), 140 мМ NaCl, 2.5 мМ CaCl2 (рН 7,4)). 100 мкл суспензии инкубировали в темноте с 5 мкл Annexin V-FITC (BD Biosciences) / 0,1 мкг йодида пропидия в течение 15 минут при комнатной температуре, добавили 400 мкл буфера для связывания и незамедлительно провели анализ на FACS Calibur (BD Biosciences).
Образование фокусов Rad 51
Клетки ES культивировали в течение 48 часов при различных концентрациях ингибитора PARP, фиксировали 4% раствором параформальдегида в PBS и пермеабилизовали 0,2% Тритоном X100 в PBS. Клетки окрасили разведением 1:100 кроличьего анти-Rad51 поликлонального антитела (Аb 551922, BD-Pharmingen, Oxford, UK). После промывки первичное антитело визуализировали с помощью Alexa Fluor-555 IgG козы против кроличьего (Alexa) и ядер с TO-PRO-3 йодидом (Molecular Probes). Фокусы Rad51 визуализировали и определили их количество с помощью конфокального микроскопа Leica TCS-SP2.
Гельэлектрофорез одиночных клеток (Commet assay)
Клетки VC8 и VC8-BAC культивировали в течение 24 часов перед обработкой 1 мкМ KU0058684 в течение 30 часов. Все последующие операции выполняли в темноте. Клетки промыли и соскоблили в PBS перед проведением гельэлектрофореза одиночных клеток, как описано в литературе (Lemay and Wood, 1999). Клетки, суспендированные в LMP агарозе (0,5% в PBS), нанесли на предметные стекла Comet (Trevigen, Gaithersburg) и выдеражали при 4°С до осаждения, а затем провели лизис в течение 45 минут в составе, содержащем 2,5 М NaCl, 100 мМ EDTA, 10 мМ Трис основания, 1% натрия лаурил саркозината, 0,01% Тритон Х-100. Предметные стекла перенесли на 5 минут в ТВЕ (trichloro-bis ethan - метоксихлор), а затем провели электрофорез при 18 В в течение 15 минут. После этого предметные стекла фиксировали 100% этанолом в течение 5 минут и высушили на воздухе, добавили зеленый краситель SYBR и визуализировали с помощью эпифлуоресценции, используя флуоресцеиновые фильтры (Nikon). Анализ одиночных клеток проводили с помощью программного модуля Comet пакета визуализации Lucia G, поставляемого компанией Nikon. Для каждого из трех независимых экспериментов контролировали по 50 отдельных клеток на одну точку и рассчитывали средний хвостовой момент.
Митотический хромосомный анализ
Клетки ES посеяли на желатин, обработали химическими индикаторами в течение 24 часов, а затем обработали колхамином в течение 1 часа. Затем клетки отобрали, фиксировали, перенесли на предметные стекла, высушили, окрасили DAPI (4,6-диамидино-2-фенилиндолдигидрохлоридом) и провели хромосоный анализ под микроскопом.
Ксенотрансплантаты клеток ES и обработка KU 0058684
Опухоли (тератомы) из клеток ES получили путем подкожной инъекции 2×108 клеток ES бестимусным мышам породы BALB (Bagg Albino)/c с мутацией по гену nude (nu/nu) в возрасте 6-8 недель. Двадцати мышам ввели инъекцию клеток ES с дефектом Brca2, а идентичной группе - изогенные клетки дикого типа. Через два дня после инъекции клеток начали лечение KU 0058684 или наполнителем. В течение трех последовательных дней вводили интраперитонеально две дозы KU 0058684 (или наполнителя) с интервалом в шесть часов при дозировке 15 мг/кг массы животного. Затем это лечение приостановили на пять дней и возобновили (как ранее) в течение трех последующих дней. Рост опухолей контролировали от минимального объема 0,3 см3. Данные на фигуре 16 представляют два отдельных эксперимента с участием в общей сложности 40 животных.
Получение клеточных линий дефектных по BRCA1
Скремблированную клеточную линию MCF7 и клеточную линию MCF7-3.23 получили путем стабильной трансфекции клеток аденокарциномы молочной железы MCF7 генноспецифическими конструкциями pSUPER. Генноспецифические конструкции pSUPER получили путем экспрессии следующих целевых последовательностей мРНК: (1) последовательности ВRСА1 человека 5'-GGAACCTGTCTCCACAAAG-3' и (2) скремблированной контрольной последовательности 5'-CATGCCTGATCCGCTAGTC-3'. Фрагмент 1,8 т.п.н., содержащий промотор человека EF1a и ген устойчивости к бластицидину (bsd), субклонировали из pEFBsd (Invitrogen) в сайт SapI конструкций pSUPER, получив pSUPER-Bsd-BRCA1 и pSUPER-Bsd-скремблированную. Клетки MCF7 трансфицировали этими плазмидами с помощью FuGene6 (Roche) согласно инструкциям изготовителя. После селекции на бластицидине подавление мРНК BRCA1 определили в резистентных клонах с использованием полимеразно-цепной реакции в режиме реального времени (Egawa et al. Oncology. 2001; 61(4): 293-8; Egawa et al. Int J Cancer. 2001 Jul 20; 95(4): 255-9). Клоны с пониженным уровнем BRCA1 мРНК культивировали (при селекции в бластицидине) в течение 8 пассажей и повторили анализ в режиме реального времени. Клеточная линия MCF7-3.23 показала только 30% экспрессии BRCA1 по сравнению с клонами MCF7, содержащими конструкцию pSUPER-Bsd-скремблированную.
Результаты
Снижение уровней белка Parp1 при действии киРНК
Плазмиду (pSUPER-eCFP-Parp1), экспрессирующую киРНК, специфическую для Parp1, под контролем промотора H1 (Т.R.Brummelkamp et al. Science 296, 550-3 (2002)) и усовершенствованный циан-флуоресцентный белок (Enhanced Cyan Fluorescent Protein, eCFP) под контролем промотора CMV IE трансфицировали в мышиные эмбриональные стволовые клетки D3. В качестве контроля отдельно трансфицировали плазмиду pSUPER-eCFP-control, экспрессирующую несвязанную скремблированную киРНК. Через сорок восемь часов после трансфекции приготовили клеточные лизаты и провели их анализ способом вестерн-блоттинг. Блоты зондировали с поликлональным анти-PARP-1 антителом или с анти-GFP/CFP антисывороткой.
Наблюдаемые уровни PARP1 в клетках, экспрессирующих киРНК, специфическую для Parp1, были гораздо ниже, чем уровни PARP1 в контрольных клетках. Уровни eCFP были аналогичными в клетках, экспрессирующих киРНК Parp1, и в контрольных клетках.
Снижение жизнеспособности клеток, дефектных по BRCA1 и BRCA2, после нокдауна с использованием киРНК, специфической для Parp1
Мышиные эмбриональные стволовые клетки (ES) дикого типа Brca1-/- и Brca2-/- трансфицировали pSUPER-eCFP-Parp1 или pSUPER-eCFP-control совместно с плазмидой pEF-Bsd, кодирующей резистентность бластицидина, в отношении 10:1. Отобрали клоны, устойчивые к бластицидину, и определили их количество. Результаты представлены на фигуре 1, где показана зависимость количества колоний pSUPER-eCFP-Parp1 в зависимости от количества pSUPER-eCFP-control после трансфекции. Отрезки прямой указывают стандартное отклонение от среднего значения.
После корректировки эффективности трансфекции с помощью контрольной киРНК оказалось, что выживание клеток ES, дефектных по Brca1 и Brca2, существенно уменьшилось при ингибировании экспрессии Parp1.
Снижение жизнеспособности клеток, дефектных по BRCA1 и BRCA2, после применения химических ингибиторов PARP
Химические ингибиторы активности Parp применили для подтверждения селективного ингибирования клеток, дефектных по Brca1 и Brca2, наблюдавшегося выше. Использовали два различных ингибитора PARP - KU 0058684 и KU 0058948, а также слабо активное, но химически родственное соединение KU 0051529 (фиг.2). Эти новые ингибиторы PARP имеют в основе фталазин-1-оновое ядро и являются конкурентными ингибиторами по отношению к субстрату NAD+ PARP. KU 0058684 и KU 0058948 являются сильнодействующими специфическими ингибиторами активности полиаденозиндифосфорибозы-полимеразы (PARP) белков PARP-1 и PARP-2 и не ингибируют PARP свода, танкиразу или PARP-3 при концентрации до 1 мкМ. В отличие от этого соединение KU 0051529, несмотря на химическое родство, примерно в 250 раз менее эффективно для ингибирования этих ферментов.
KU 0058684, KU 0058948 и KU 0051529 использовали для определения чувствительности клеток, дефектных по Brca1 или Brca2, к ингибированию активности PARP. Клоногенные анализы показали, что клеточные линии, дефектные Brca1 и Brca2, чрезвычайно чувствительны к KU 0058684 и KU 0058948 по сравнению с другими изогенными клетками (фиг.3, 4). Величина SF50 (доза, при которой выживают 50% клеток) для KU 0058684 составила 3,5×10-8 М для Brca1 и 1,5×10-8 М для Brca2, а для клеток дикого типа - около 3,5×10-8 М. Это соответствует повышению чувствительности в 57 и 133 раза для клеток, мутантных по Brca1 и Brca2 соответственно, по сравнению с клетками дикого типа. Аналогичные результаты получили с клетками яичников китайского хомячка, дефектными по Brca2, которые показали повышение чувствительности более, чем в 1000 раз по сравнению с комплементарными производными Brca2 (фиг.14 и 15). Чувствительность клеток, мутантных по Brca1 и Brca2, к KU 0058948 была даже большей, чем к KU 0058684. В отличие от этого соединение KU 0051529 не оказывало селективного влияния на клетки с недостатком Brca1 или Brca2 дикого типа по сравнению с клетками дикого типа. Это в сочетании с данными для киРНК показывает, что механизм чувствительности действует специфически посредством ингибирования PARP. Примечательно, что ни один из ингибиторов не оказывал влияния клеткам, гетерозиготным по мутации Brca1 или Brca2.
Влияние времени действия KU0058684 на клоногенное выживание клеток, дефектных no Brca1 и Brca2
Клетки подвергли воздействию различных концентраций KU0058684 в течение определенных периодов времени. Затем ингибитор удалили и определили результат воздействия с помощью клоногенного анализа. Ингибирующее влияние KU 0058684 на клоногенный рост проявлялось после относительно короткого времени воздействия - 4 часа - и практически заканчивалось через 24 часа (фигуры 5 и 6). Влияние ингибирования PARP оказалось необратимым, поскольку кратковременное воздействие предотвращает рост в следующие 10-14 дней при отсутствии ингибитора.
Влияние ингибирования PARP на остановку клеточного цикла
Анализ FACS использовали для определения влияния ингибирования PARP на остановку клеточного цикла. Клетки подвергли действию KU 0058684 в течение различных периодов времени, а затем пометили BrdU и определили долю клеток в каждой фазе клеточного цикла. Результаты показаны на фигурах 7 и 8. Согласно наблюдениям KU0058684 обеспечивает полную остановку роста клеток, содержащих тетраплоидную ДНК, что указывает на остановку клеточного цикла в фазе G2 или М. Для того чтобы дополнительно охарактеризовать эту остановку, в клетках с помощью маркера фазы М анализировали содержание ДНК и фосфорилированного гистона Н3 (фигуры 12 и 13). Большинство остановленных клеток не было мечено антителами анти-фосфогистона Н3. Это означает, что большинство клеток было остановлено в фазе G2.
Образование фокусов Rad51
Одним из признаков репарации двухнитевого разрыва, зависимой от Brca, является образование в ядре фокусов, содержащих Rad51. Исследовали способность KU 0058684 создавать фокусы Rad51 в клетках дикого типа, а также в дефектных клетках Brca1 и Brca2.
Клетки ES дикого типа и дефектные клетки Brca1 и Brca2 подвергли действию различных концентраций KU 0058684 в течение 48 часов. Затем клетки фиксировали и окрасили для определения фокусов RAD51, как описано в Tarsounas (Tarsounas М et al. Oncogene. 2003 22(8): 1115-23).
В клетках ES дикого типа KU0058684 вызвал образование фокусов Rad51, зависимых от дозы (фигура 9). В отличие от этого в клетках, дефектных по Brca1 или Brca2, не было индукции фокусов. Этот последний результат согласуется с предшествующими наблюдениями, согласно которым агенты, повреждающие ДНК, не могут вызывать образования фокусов Rad51 в клетках, дефектных по Brca1 или Brca2.
Таким образом, показано, что KU 0058684 вызывает повреждения, в частности двухнитевые разрывы ДНК или повреждения, перерастающие в двухнитевые разрывы ДНК, которые репарирует комплекс, включающий Rad51, и требует наличия Brca1 и Brca2. Важно отметить, что KU 0051529 не вызывал образования фокусов Rad51 при сравнимых дозах, что подчеркивает специфичность механизма сенсибилизации.
Гельэлектрофорез одиночных клеток (Commet assay)
Чтобы определить, приводит ли ингибирование активности PARP к образованию двухнитевых разрывов ДНК, провели нейтральный гельэлектрофорез одиночных клеток на Brca2-мутантных клетках и их изогенных аналогах. Результаты показаны на фигуре 10. После 30-часового воздействия 1 мкМ KU 0058684 имело место 4,7-кратное увеличение хвостового момента Brca2-дефектных клеток VC8 и отсутствовало существенное увеличение хвостового момента комплементированной линии VC8-BAC. Этот результат показывает, что двухнитевые разрывы ДНК, индуцированные PARPi, остаются нерепарированными в линии, дефектной по Brca2.
Анализ митотических хромосом
Контроль митотических хромосом клеток, дефектных по Brca1 и Brca2, показал, что обработка KU0058684 приводит к частым крупным аберрациям. Они включают разрывы хроматид, а также трех- и четырехрадиальных хромосом. Эти фенотипы указывают на невозможность репарации двухнитевых разрывов путем конверсии генов сестринских хромосом и расширенное применение альтернативных, подверженных ошибкам путей.
Ксенотрансплантаты клеток ES и обработка KU 0058684
Опухоли (тератомы) получили из изогенных и клеток ES, дефектных по Brca2, бестимусных мышей породы BALB/c с мутацией по гену nude (nu/nu), как описано выше.
Определили влияние KU 0058684 на рост опухолей дикого типа и ксенотрансплантатов, дефектных по Brca2. Результаты показаны на фигуре 16.
Согласно наблюдениям KU0058684 резко уменьшает рост опухолей, дефектных по Brca2, по сравнению с опухолями дикого типа.
Эффект ингибирования PARP в BRCA1-дефектных клеточных линиях
Клеточные линии МСF7-скремблированную и MCF7-3.23 получили, как описано выше. MCF7-3.23 обеспечивала 30% экспрессии BRCA1 по сравнению с MCF7-скремблированной.
Клетки линий MCF7-скремблированной и MCF7-3.23 обработали KU 0058684 и KU 0051529 и определили выживание клеток (фигуры 17 и 18). KU 0058684 является сильным ингибитор PARP, в то время как KU 0051529 менее эффективен.
Клетки линий MCF7-скремблированной и MCF7-3.23 не проявили существенной чувствительности к KU 0051529 (фигура 18). Однако обе клеточные линии были чувствительны к KU 0058684.
Клетки линии MCF7-3.23 гораздо более чувствительны к KU0058684, чем клетки линии МСF7-скремблированной.
Взаимодействие PARP и пути репарации ДНР ДНК, зависимой от HR
Без какого-либо ограничения области распространения изобретения одна из возможных моделей взаимодействия PARP и пути репарации ДНР ДНК, зависимой от HR, показана на фигуре 11.
Однонитевые разрывы (single-strand breaks, SSB) ДНК образуются вследствие окислительного повреждения и его репарации. Ингибирование активности полимеразы PAR Parp-1 препятствует рекрутированию каркасного белка XRCC1 и последующему заполнению SSB участка ДНК- полимеразами (фигура 11, А).
SSB сохраняются в большом количестве и сталкиваются с репликативными вилками ДНК. Отсутствие матричной нити в SSB приводит к образованию ДНР и в зависимости от позиции может вызывать разрушение репликативной вилки (фигура 11, В).
Близость неповрежденной сестринской хроматидной матрицы обеспечивает инвазию сестринской хроматиды посредством одинарной нити ДНК, покрытой RAD51, и позволяет инициировать рекомбинационную репарацию сестринской хроматиды. Этот процесс зависит от BRCA1 и BRCA2 и образования в ядре множества фокусов RAD51. Разрушенные репликативные вилки можно восстановить при помощи аналогичного механизма (фигура 11, С).
Если структуры Холлидея, образующиеся в промежуточных стадиях рекомбинации, разрушаются, может происходить обмен сестринскими хроматидами (sister chromatid exchange, SCE). Чрезмерное количество SSB, сталкивающихся с репликативными вилками при ингибировании Раrр-1, приводит к увеличению SCE (фигура 11, D).
При отсутствии функциональных BRCA1 или BRCA2 образование фокусов RAD51 и рекомбинация сестринских хроматид резко подавляются. Во время репликации ДНК избыток нерепарированных SSB образует ДНР, однако сестринской рекомбинации не происходит. Они остаются нерепарированными, поскольку хроматиды разрушаются, или репарируются посредством механизмов, независимых от RAD51, подверженных ошибкам и вызывающих сложные перестройки хромосом. Такие клетки останавливаются в цикле, когда они сталкиваются с контрольной точкой повреждения ДНК в G2/M, что приводит к полной остановке цикла или апоптозу (фигура 11, Е).
Приведенные здесь данные о РНКи (интерференции РНК) вместе с наблюдаемой неэффективностью KU 0051529 показывают, что ингибирование PARP ответственно за наблюдаемые эффекты сенсибилизации.
Ингибирование PARP вызывает повреждения, которые обычно репарируются обменом сестринских хроматид (sister chromatid exchange, SCE) (Wang ZQ, et al. (1997) Genes Dev. Vol.11(18): 2347-58 и 'From DNA damage and stress signalling to cell death' G. de Murcia и S.Shall eds. Oxford University Press (2000)). Известно, что ингибирование PARP увеличивает SCE, не вызывая сопутствующего увеличения генной конверсии ДНР и, следовательно, глобального увеличения пути рекомбинации, зависимого от Rad51 (Schultz N et al. 2003, Nucleic Acids Research; vol.31 (17): 4959-4964).
Описанный здесь синтетической подход обеспечения клеточной летальности может быть использован для лечения: а) опухолей в носителях рака молочной железы, б) опухолей, в которых другие компоненты репарации ДНР ДНК, зависимой от HR, являются дефектными.
Несмотря на то что существуют описания различий во времени начала симптомов и патологии опухолей в носителях мутаций рака молочной железы, в настоящее время их лечение является точно таким же, как и для пациентов со спорадическим заболеванием. Данное изобретение обеспечивает новый подход к этим опухолям, изложенный в этом описании.
Важно отметить отсутствие наблюдений гетерозиготного эффекта. Это является существенным фактором для терапевтического применения описанных способов для гетерозиготных носителей мутаций на пути репарации ДНР ДНК, зависимой от HR, включая, например, мутации BRCA1/2.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНГИБИТОРЫ РЕПАРАЦИИ ПОВРЕЖДЕНИЙ ДНК ДЛЯ ЛЕЧЕНИЯ РАКА | 2017 |
|
RU2755865C2 |
| 4-[3-(4-ЦИКЛОПРОПАНКАРБОНИЛ-ПИПЕРАЗИН-1-КАРБОНИЛ)-4-ФТОР-БЕНЗИЛ]-2Н-ФТАЛАЗИН-1-ОН | 2008 |
|
RU2487868C2 |
| ПОЛИМОРФНАЯ ФОРМА 4-[3-(4-ЦИКЛОПРОПАНКАРБОНИЛПИПЕРАЗИН-1-КАРБОНИЛ)-4-ФТОРБЕНЗИЛ]-2Н-ФТАЛАЗИН-1-ОНА | 2007 |
|
RU2465270C2 |
| СПОСОБЫ НА ОСНОВЕ ДЕТЕКТИРОВАНИЯ ОЧАГОВ RAD51 В ОПУХОЛЕВЫХ КЛЕТКАХ | 2018 |
|
RU2825699C2 |
| УЛУЧШЕННЫЕ ЭКСПРЕССИЯ И ПРОЦЕССИНГ ТРАНСГЕНА | 2014 |
|
RU2808756C1 |
| ФАРМАЦЕВТИЧЕСКИ ПРИЕМЛЕМЫЕ СОЛИ 2-{4-[(3S)-ПИПЕРИДИН-3-ИЛ]ФЕНИЛ}-2Н-ИНДАЗОЛ-7-КАРБОКСАМИДА | 2009 |
|
RU2495035C2 |
| ТРИЦИКЛИЧЕСКИЕ ИНГИБИТОРЫ ПАРП | 2004 |
|
RU2404183C2 |
| ЛЕЧЕНИЕ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ С ПОМОЩЬЮ СОЕДИНЕНИЯ 4-ИОД-3-НИТРОБЕНЗАМИД В КОМБИНАЦИИ С ПРОТИВООПУХОЛЕВЫМИ СРЕДСТВАМИ | 2008 |
|
RU2480211C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ТЕТРАЦИКЛИЧЕСКИМИ АНАЛОГАМИ ХИНОЛОНА ДЛЯ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2752506C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ТЕТРАЦИКЛИЧЕСКИМИ АНАЛОГАМИ ХИНОЛОНА ДЛЯ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2786570C2 |
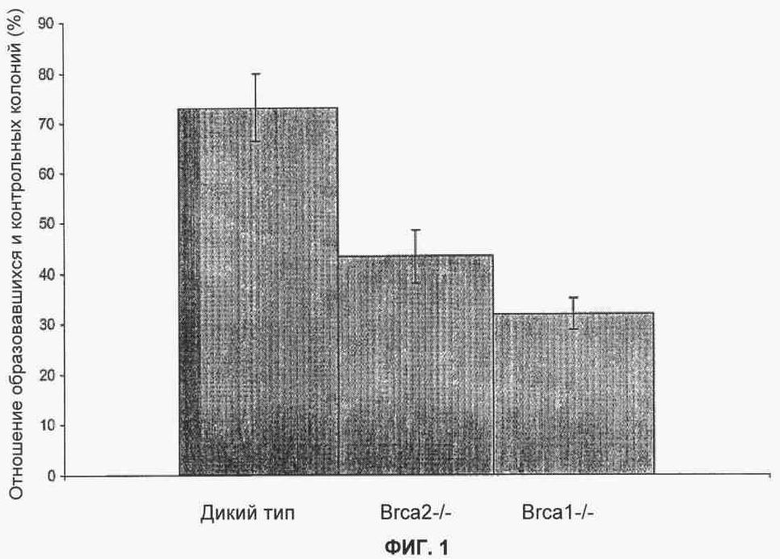
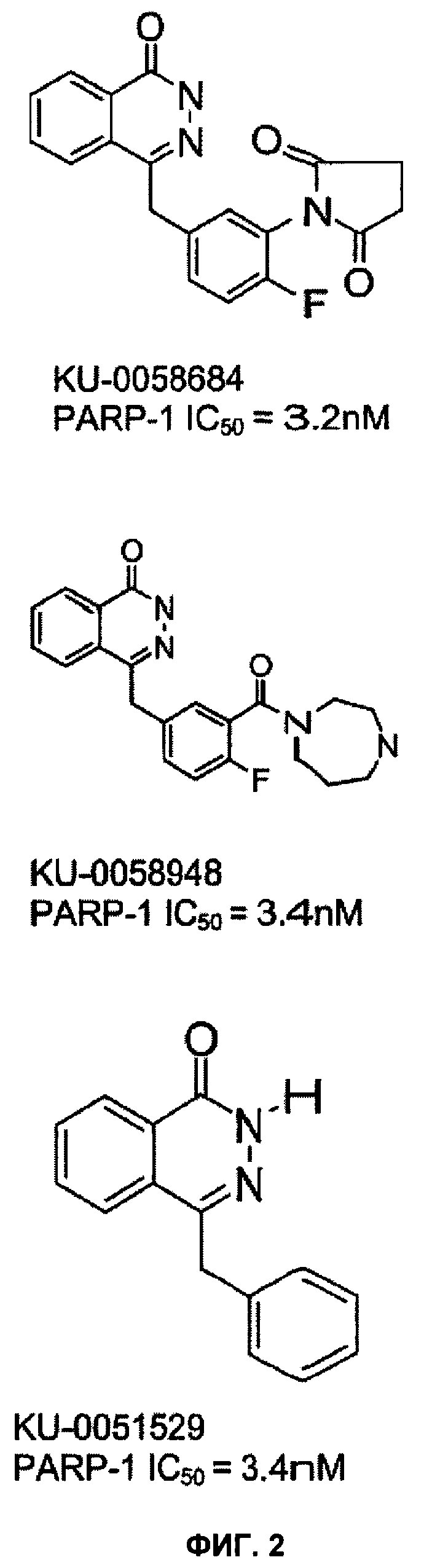
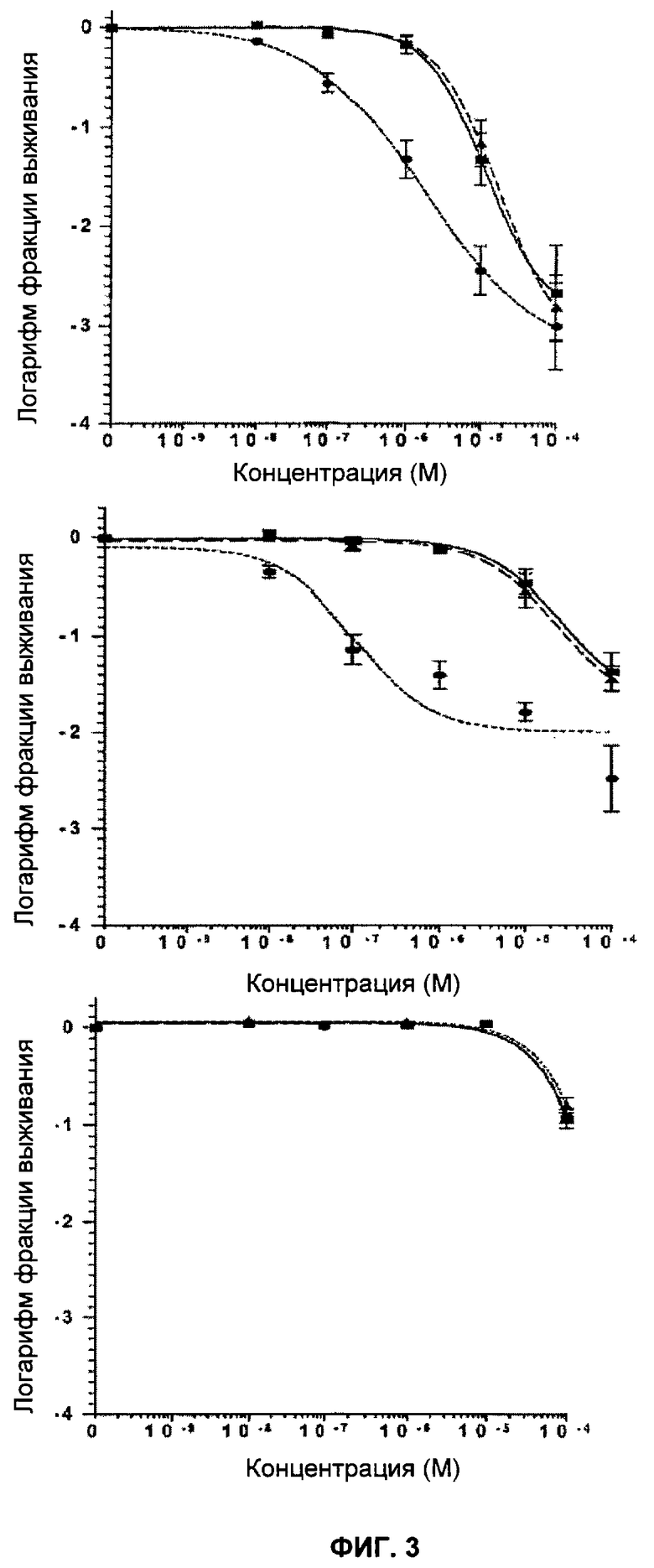
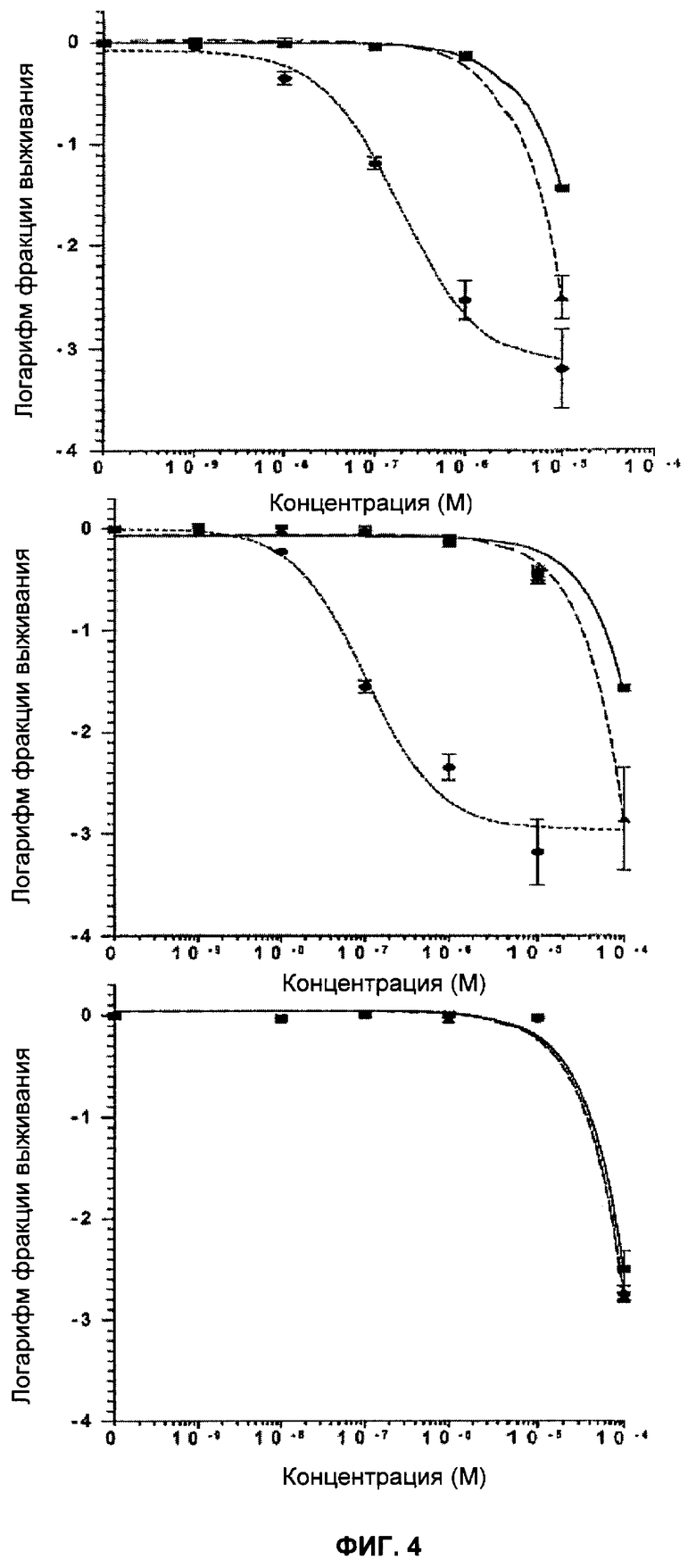
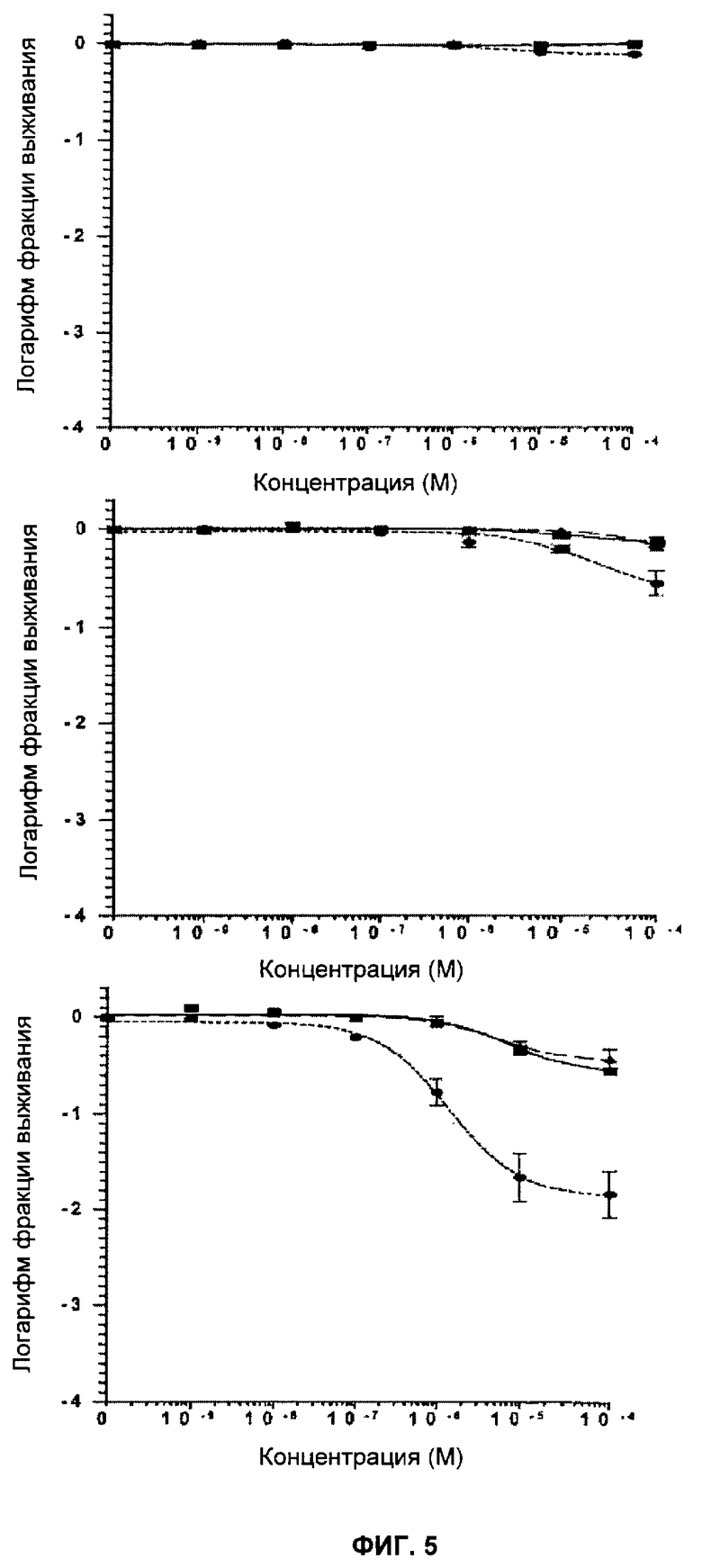
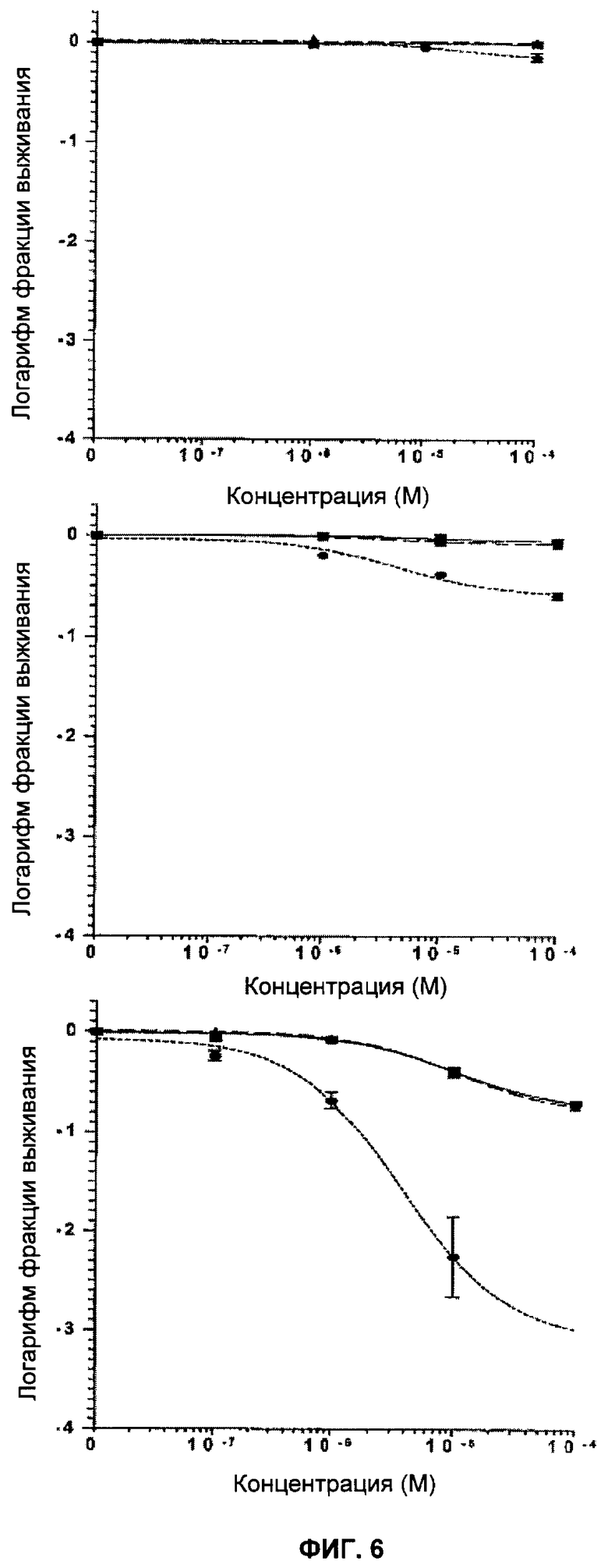
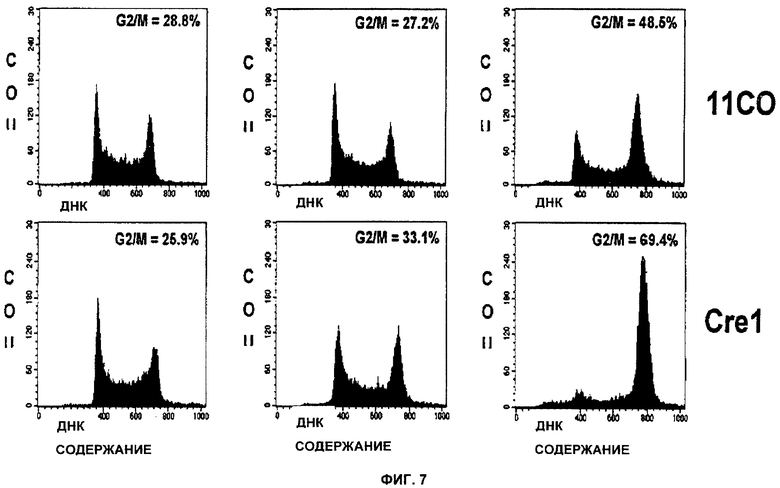
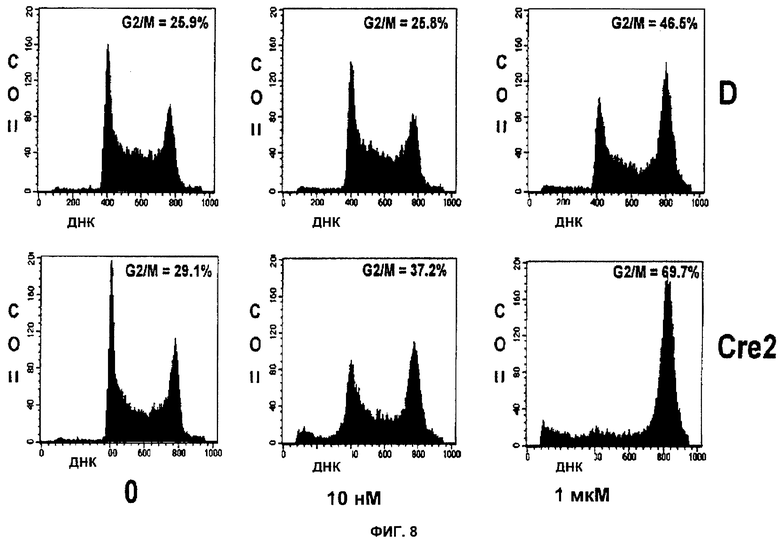
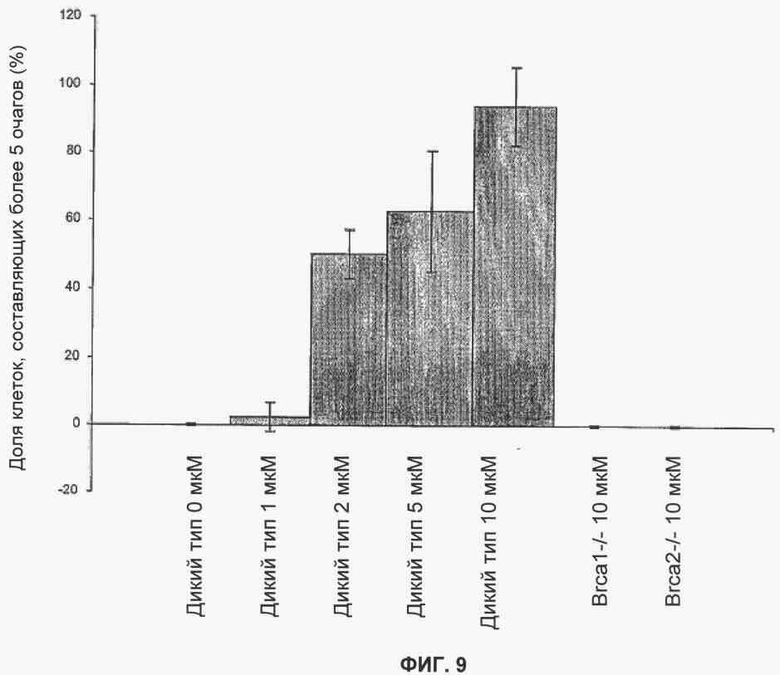
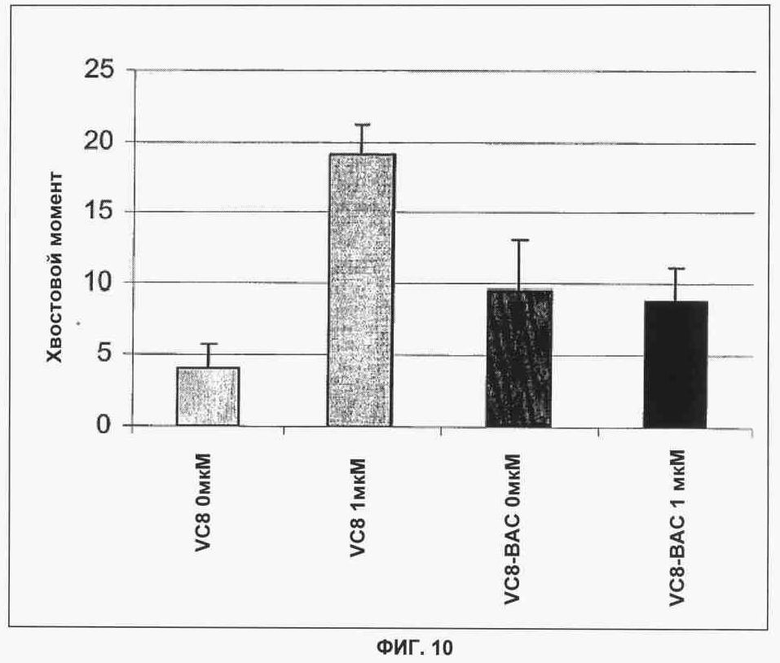
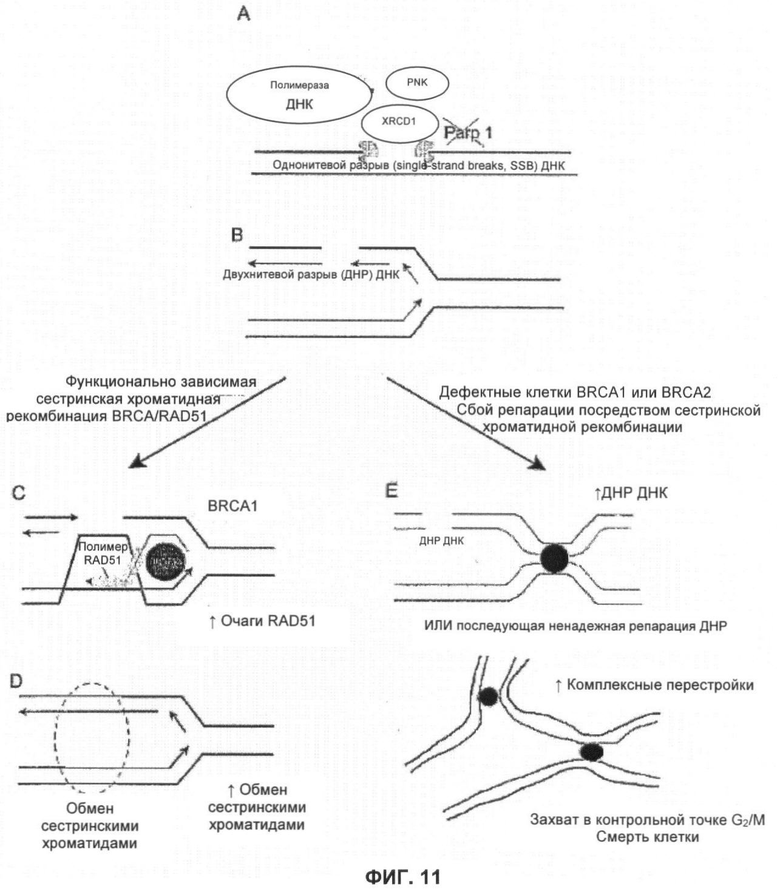
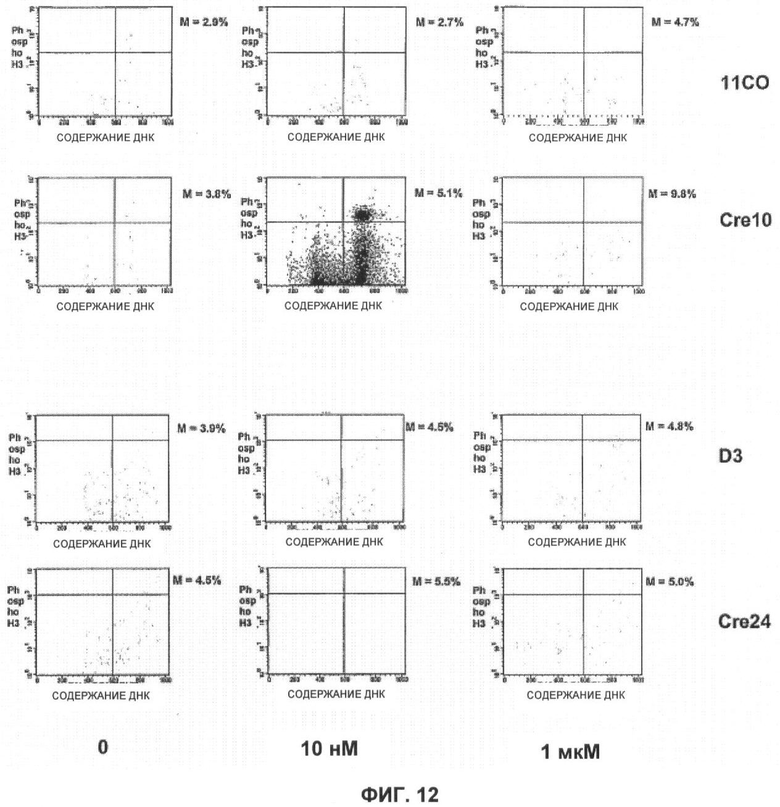
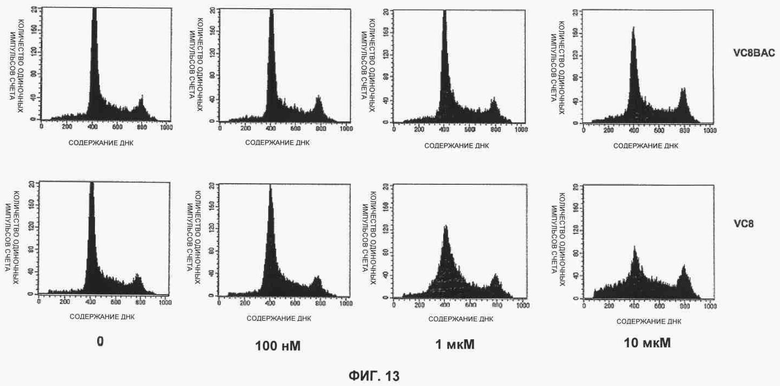
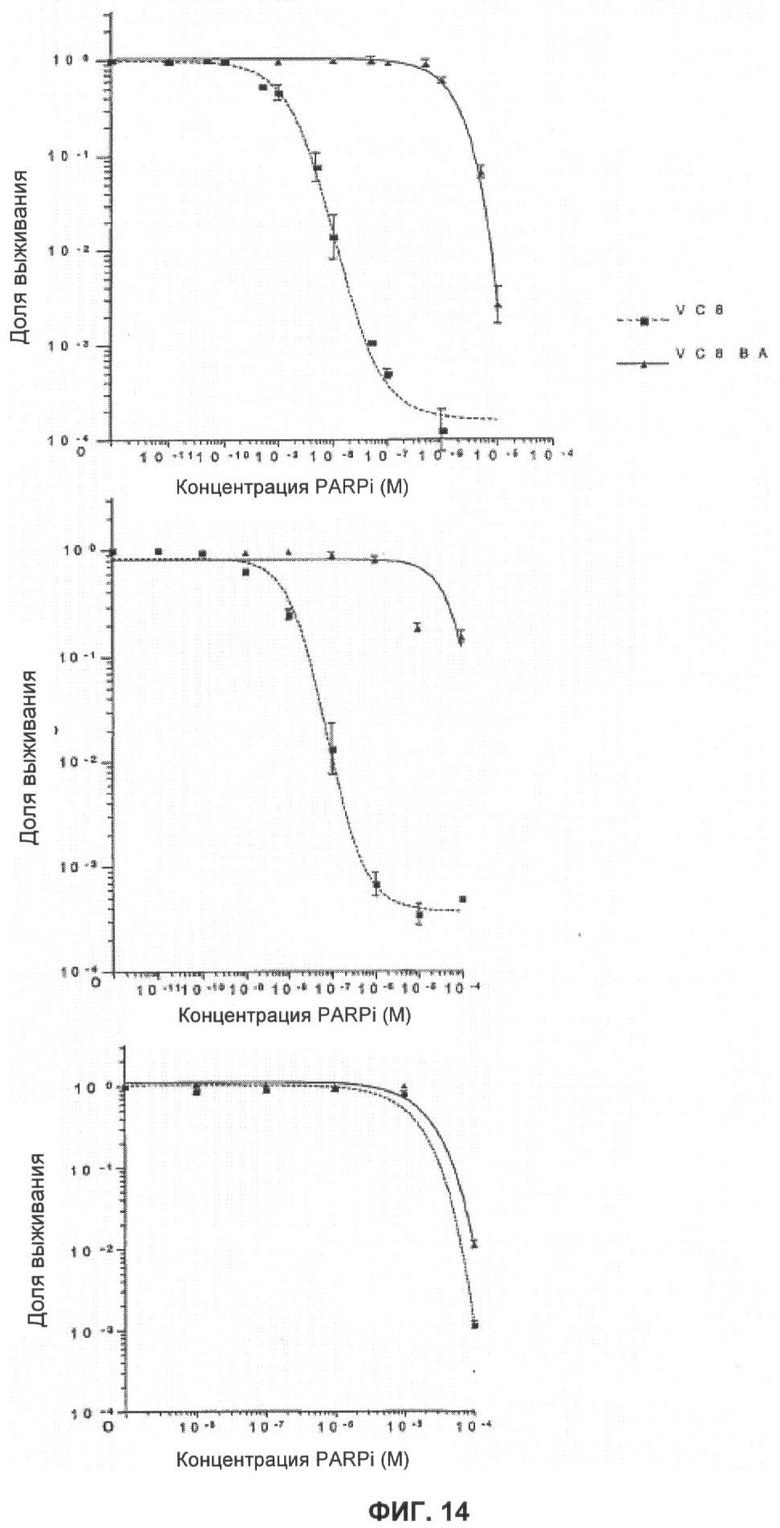
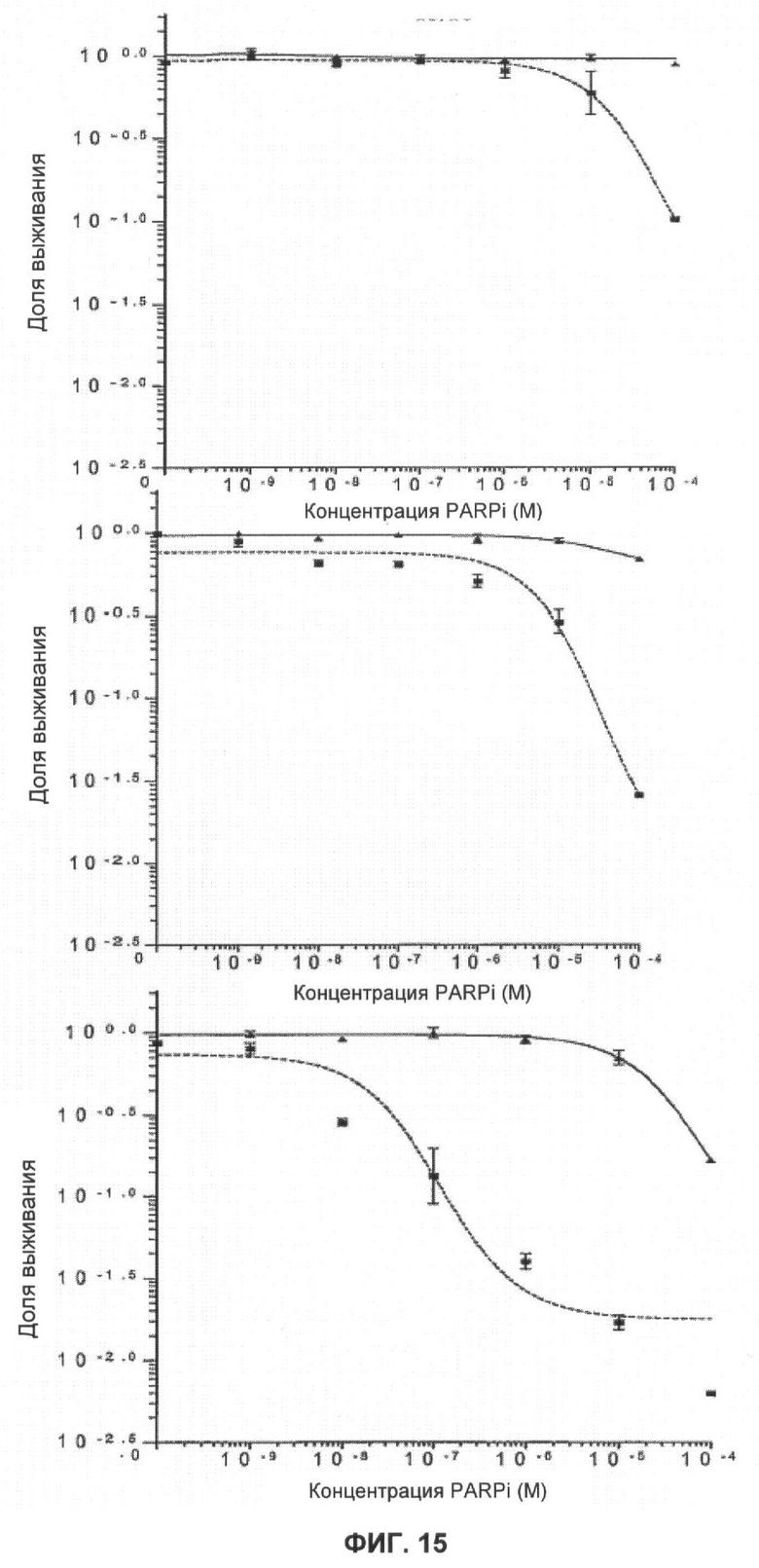
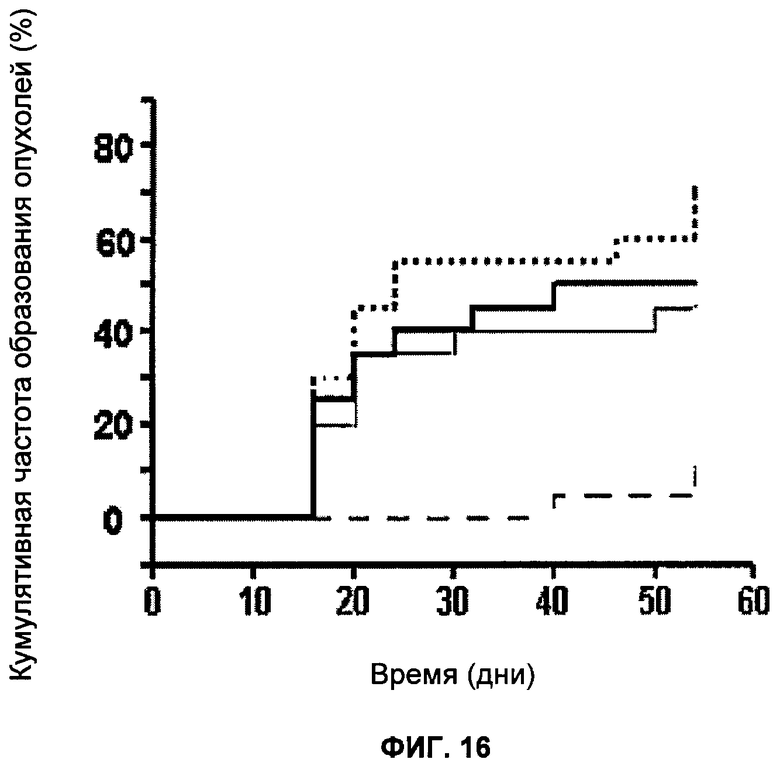
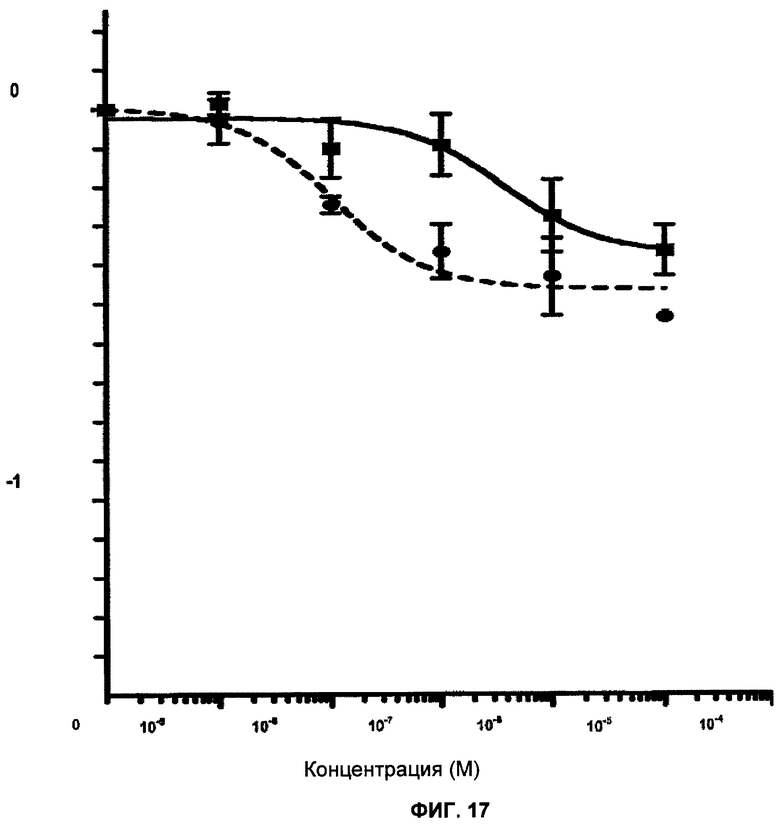
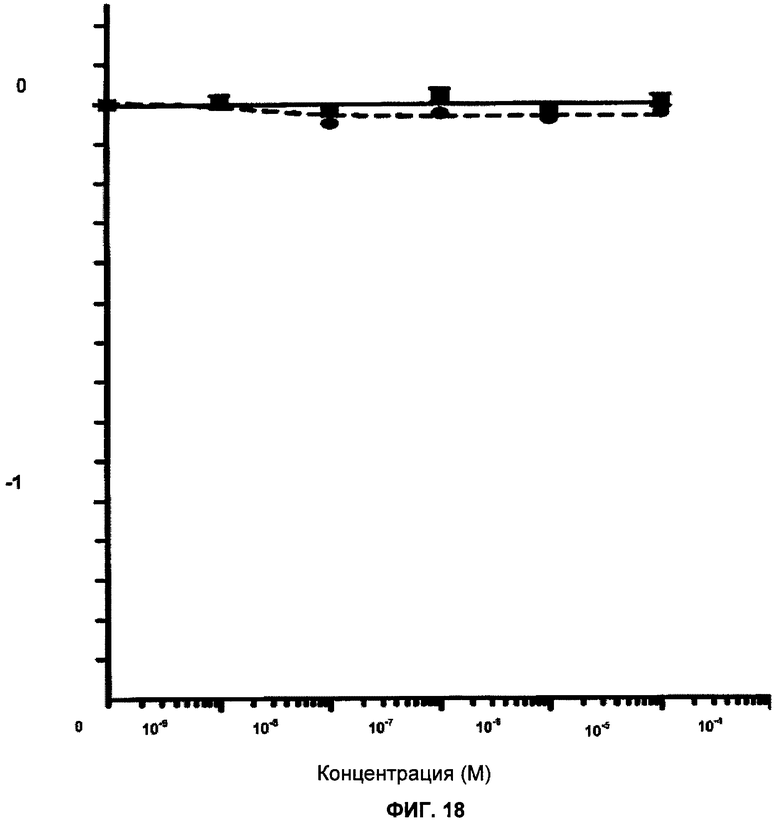
Изобретение относится к области медицины и касается ингибиторов репарации повреждения ДНК для лечения рака. Сущность изобретения включает установление того факта, что ингибирование пути эксцизионной репарации оснований является избирательно летальным в клетках, которые являются дефектными при репарации ДНР ДНК, зависимой от гомологической рекомбинации. При этом обеспечиваются способы и средства для лечения раковых заболеваний, которые являются дефектными по репарации ДНР ДНК, зависимой от гомологической рекомбинации, с использованием ингибиторов PARP, в частности ингибитор PARP представляет собой фтализин-1 (2Н)-он. Преимущество изобретения заключается в разработке медикамента, предназначенного для лечения рака, который является дефектным по пути репарации двунитевого разрыва ДНК, зависимой от гомологичной рекомбинации. 5 н. и 12 з.п. ф-лы, 18 ил.
1. Применение ингибитора PARP для получения медикамента, предназначенного для лечения у индивидуума рака, который является дефектным по пути репарации двухнитевого разрыва (ДНР) ДНК, зависимой от гомологической рекомбинации, причем ингибитор PARP представляет собой фталазин-1 (2Н)-он.
2. Применение по п.1, отличающееся тем, что указанный рак содержит одну или несколько раковых клеток, имеющих пониженную способность или утративших способность к репарации ДНР ДНК посредством гомологической рекомбинации по сравнению с нормальными клетками.
3. Применение по п.2, отличающееся тем, что указанные раковые клетки имеют фенотип, дефектный по BRCA1 или BRCA2, или указанные раковые клетки являются дефектными по BRCA1 или BRCA2, или указанные раковые клетки являются гомозиготными по мутации в BRCA1 или BRCA2.
4. Применение по пп.1-3, отличающееся тем, что указанный индивидуум является гетерозиготным по мутации в гене, кодирующем компонент пути репарации ДНР ДНК, зависимой от гомологической рекомбинации или является гетерозиготным по мутации в BRCA1 и/или BRCA2.
5. Применение по пп.1-3, отличающееся тем, что указанный рак представляет собой рак молочной железы, яичника, поджелудочной железы или простаты.
6. Применение по любому из пп.1-5, отличающееся тем, что указанный медикамент дополнительно содержит химиотерапевтический агент, повреждающий ДНК или указанное лечение включает также введение химиотерапевтического агента, повреждающего ДНК.
7. Способ лечения рака у индивидуума, включающий:
введение ингибитора PARP указанному индивидууму,
при этом указанный рак является дефектным по пути репарации ДНР ДНК, зависимой от гомологической рекомбинации, причем ингибитор PARP представляет собой фталазин-1 (2Н)-он.
8. Способ по п.7, отличающийся тем, что включает операцию идентификации индивидуума, как страдающего раковым заболеванием, которое дефектно по пути репарации ДНР ДНК, зависимой от гомологической рекомбинации.
9. Способ лечения рака у индивидуума, включающий:
введение указанному индивидууму ингибитора PARP, при этом указанный индивидуум является гетерозиготным по мутации в гене, который кодирует компонент пути репарации ДНР ДНК, зависимой от гомологической рекомбинации, при этом ингибитор PARP представляет собой фталазин-1 (2Н)-он.
10. Способ оценки состояния индивидуума, страдающего раковым заболеванием, включающий:
идентификацию раковой клетки, полученной у индивидуума, как дефектной по пути репарации ДНР ДНК, зависимой от гомологической рекомбинации, по сравнению с нормальными клетками, и
обеспечение ингибитора PARP, пригодного для введения указанному индивидууму, при этом ингибитор PARP представляет собой фталазин-1 (2Н)-он.
11. Способ по любому из пп.7-10, отличающийся тем, что включает обеспечение химиотерапевтического агента, повреждающего ДНК и пригодного для введения указанному индивидууму.
12. Способ определения активности пути репарации ДНР ДНК, зависимой от гомологической рекомбинации, при раковом заболевании индивидуума или прогнозирования реакции ракового заболевания у индивидуума на лечение, которое воздействует на раковые заболевания, дефектные по репарации ДНР ДНК, зависимой от гомологической рекомбинации, включающий:
приведение в контакт ингибитора PARP с образцом раковых клеток, полученных у индивидуума, страдающего указанным заболеванием, при этом ингибитор PARP представляет собой фтализин-1 (2Н)-он, и
определение уровня клеточной смертности в указанном образце.
13. Способ по любому из пп.7-10, 12, отличающийся тем, что указанный рак содержит одну или несколько раковых клеток, которые имеют пониженную способность или утратили способность к репарации ДНР ДНК посредством гомологической рекомбинации по сравнению с нормальными клетками.
14. Способ по п.13, отличающийся тем, что указанные раковые клетки имеют фенотип, дефектный по BRCA1 или BRCA2, или указанные раковые клетки являются дефектными по BRCA1 или BRCA2, или указанные раковые клетки являются гомозиготными по мутации в BRCA1 или BRCA2.
15. Способ по любому из пп.7-10 и 12, отличающийся тем, что указанный рак идентифицирован как рак, дефектный по репарации ДНР ДНК, зависимой от гомологической рекомбинации, путем определения активности репарации ДНР ДНК, зависимой от гомологической рекомбинации, раковых клеток, полученных у индивидуума, по сравнению с нормальными клетками, или зависимой от гомологической рекомбинации, путем определения наличия в раковых клетках, полученных от индивидуума, одной или нескольких мутаций или полиморфизмов в последовательности нуклеиновой кислоты, кодирующей компонент пути репарации ДНР ДНК, зависимой от гомологической рекомбинации.
16. Способ по любому из пп 7-10, 12 и 14, отличающийся тем, что указанный рак представляет собой рак молочной железы, яичника, поджелудочной железы или простаты.
17. Способ по любому из пп.7-10, 12 и 14, отличающийся тем, что указанный индивидуум является гетерозиготным по мутации в гене, кодирующем компонент пути репарации ДНР ДНК, зависимой от гомологической рекомбинации, или является гетерозиготным по мутации в BRCA1 и/или BRCA2.
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| SCHULTZ N | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |

