Изобретение относится к биотехнологии и касается способа определения гиперметилированных CpG островков, находящихся в области генов-супрессоров опухолевого роста в ДНК человека.
Известно, что метилирование ДНК млекопитающих по CG-динуклеотидам является одним из основных методов регуляции экспрессии генов. Метилирование CG-динуклеотида происходит с образованием 5-метилцитозинов (5mС) в обеих цепях ДНК, и эта модификация лежит в основе эпигенетической регуляции активности генов. В ходе эпигенетической регуляции происходит избирательное метилирование CG-богатых участков ДНК, называемых CpG островками и расположенных преимущественно в области промотора и первого экзона транскрибируемых генов. Подавление активности генов-супрессоров опухолевого роста в ДНК человека осуществляется путем метилирования CG-динуклеотидов преимущественно в последовательностях 5'-PuCGC-3', входящих в состав CpG островков [1]. CpG островки имеют размеры от 500 до 2000 пар нуклеотидов [2] и такая модификация CG-динуклеотидов приводит к возникновению так называемых гиперметилированых CpG островков. Переход CpG островков в гиперметилированное состояние резко снижает экспрессию генов-супрессоров опухолевого роста, что приводит к активации канцерогенеза. В связи с этим выявление гиперметилированных CpG островков генов-супрессоров опухолевого роста в ДНК человека является крайне важным и позволяет проводить раннюю диагностику заболевания.
Установление гиперметилированного состояния CpG островка проводят путем определения метилированных цитозинов в области промотора и первого экзона исследуемого гена-супрессора в опухолевых и здоровых клетках и сравнения полученных результатов.
Известен способ определения метилированных цитозинов путем проведения метилчувствительной полимеразной цепной реакции (ПЦР) [3], заключающийся в следующем. Образец ДНК обрабатывают рестриктазами HpaII и MspI, которые узнают последовательность ДНК 5'-CCGG-3'. При этом фермент MspI расщепляет метилированную последовательность 5'-C(5mC)GG-3', тогда как HpaII такой сайт не расщепляет. Затем фрагмент ДНК, содержащий последовательность 5'-CCGG-3', амплифицируют. В случае метилирования внутреннего CG-динуклеотида этого сайта он будет расщепляться рестриктазой MspI, и амплификация фрагмента ДНК происходить не будет, тогда как HpaII такой сайт не расщепляет и в ходе ПЦР будет происходить наработка фрагмента ДНК. В отсутствии метилированного цитозина оба фермента будут расщеплять ДНК и амплификации фрагмента не будет. Таким образом, наличие метилированного цитозина в сайте 5'-C(5mC)GG-3' устанавливают по появлению амплифицированного фрагмента после обработки ДНК ферментом HpaII.
Недостатком данного способа является его применимость только для последовательности ДНК 5'-CCGG-3', тогда как метилирование цитозинов в CpG островках генов-супрессоров происходит преимущественно по сайтам 5'-ACGC-3' и 5'-GCGC-3', что делает невозможным его использование для установления метилирования CG-динуклеотидов в CpG островках генов-супрессоров.
Наиболее близким к заявляемому способу - прототипом является способ определения статуса метилирования первого экзона гена-супрессора опухолевого роста р16 с применением метилчувствительных рестриктаз, таких как HpaII (5'-CCGG-3'), CfoI (5'-GCGC-3') и SmaI (5'-CCCGGG-3') [4].
Способ включает двукратное (для большей эффективности) расщепление образца геномной ДНК одной из этих чувствительных к метилированию рестриктаз и ПЦР-амплификацию продукта расщепления с праймерами, фланкирующими первый экзон гена р16.
Недостатком данного способа является его применимость только для одной из трех вышеперечисленных последовательностей ДНК, тогда как метилирование цитозинов в CpG островках генов-онкосупрессоров человека происходит преимущественно по сайтам 5'-ACGC-3' и 5'-GCGC-3' [1]. Однако метилчувствительные рестриктазы неспособны расщеплять свой сайт узнавания в том случае, если он содержит метилированный CG-динуклеотид, поэтому значительная часть метилированных последовательностей не может быть обнаружена этим способом.
Технической задачей изобретения является расширение функциональных возможностей известного способа.
Поставленная техническая задача достигается предлагаемым способом, заключающимся в получении высокоочищенной ДНК из больных и здоровых клеток (контроль), предварительной фрагментации выделенной ДНК эндонуклеазой рестрикции, не имеющей сайта узнавания в амплифицируемом районе, гидролизе фрагментированной ДНК метилзависимой сайт-специфической эндонуклеазой GlaI или BlsI с последующей амплификацией продуктов расщепления ДНК в ПНР с использованием пары праймеров, фланкирующих CpG островки в регуляторной области гена супрессора опухолевого роста в ДНК человека, с последующим составлением заключения о наличии дополнительных по сравнению с контрольным образцом метилированных CG динуклеотидов в исследуемых CpG островках.
Высокоочищенную ДНК выделяют из культур клеток человека как описано ранее в [5]. Предварительную фрагментацию ДНК проводят в реакционной смеси, содержащей ДНК и эндонуклеазу рестрикции TaqI в течение 2 часов при 65°С в буфере: 33 мМ трис-ацетат, рН 7,9; 10 мМ ацетат магния, 66 мМ ацетат калия, 1 мМ дитиотреитол.
Затем фрагментированную ДНК расщепляют путем добавления 10-20 единиц активности метилзависимой сайт-специфической эндонуклеазы, GlaI или ВЫ, в соответствующем реакционном буфере в течение 2 часов при 30°С. Метилзависимая сайт-специфическая эндонуклеаза GlaI узнает и расщепляет только метилированную ДНК по последовательности 5'-Pu(5mC)GPy-3'/3'-PyG(5mC)Pu-5' [6], которая включает в себя последовательность 5'-Pu(5mС)ОС-3'/3'-РуG(5mС)O-5', преимущественно метилируемую в ДНК человека, в том числе в гиперметилированных CpG островках генов-супрессоров. Метилзависимая сайт-специфическая эндонуклеаза BlsI расщепляет метилированную последовательность 5'-GCNGC-3', которая содержит как минимум один 5-метилцитозин (не включая центральный нуклеотид N) в каждой цепи [7]. В зависимости от нуклеотидов, находящихся в центре, до и после последовательности 5'-GCNGC-3', ее перекрывание с метилированным динуклеотидом CG может привести к образованию сайта узнавания фермента BIsI. Например, последовательности ДНК 5'-G(5mC)GG(5mC)G-3'/3'-CG(5mC)CG(5mC)-5' и 5'-(5mC)GC(5mC)GC-3'/3'-G(5mC)GG(5mC)G-5' являются субстратами BIsI.
Далее 1 мкл реакционной смеси отбирают для проведения ПНР с Hot Start Taq ДНК-полимеразой. Продукты ПНР разделяют путем электрофореза в 1,2% агарозном геле и делают заключение о наличии или отсутствии гиперметилированных по сравнению с контролем CpG островков в регуляторной области генов-онкосупрессоров человека.
Определяющими отличительными признаками предлагаемого способа от прототипа, являются:
- предварительную фрагментацию ДНК осуществляют рестриктазой TaqI, что позволяет провести более полное расщепление исходной ДНК;
- фрагментированную ДНК гидролизуют метилзависимыми сайт-специфическими эндонуклеазами GlaI или BlsI, что позволяет расщепить метилированные CG-динуклеотиды в CpG островках генов-супрессоров;
Предлагаемый способ прост в исполнении, обладает высокой чувствительностью, так как позволяет обнаружить максимальное количество сайтов метилирования, а также обеспечивает возможность анализировать протяженные (до нескольких тысяч пар оснований) участки ДНК.
Изобретение иллюстрируется следующими примерами конкретного выполнения.
Пример 1. Определение гиперметилированного CpG островка в промоторной области гена-супрессора DАРK1
Предварительную фрагментацию ДНК, выделенной из культур клеток человека L-68 (контроль-фибробласты легкого от здорового человека), HeLa, Jurkat, K-562, Raji, U-937 (малигнантные линии клеток) проводили в 6 отдельных пробирках, содержащих по 50 мкл реакционной смеси, включающей 5 мкг ДНК и 20 единиц рестриктазы TaqI в течение 2 часов при 65°С в буфере: 33 мМ трис-ацетат, рН 7,9; 10 мМ ацетат магния, 66 мМ ацетат калия, 1 мМ дитиотреитол. В седьмой пробирке аналогичным способом обрабатывали образец контрольной ДНК Drosophila melanogaster (контроль на отсутствие контаминации в ПЦР). Затем гидролизаты ДНК очищали фенол-хлороформной экстракцией и высаживали 96% этанолом по общепринятой методике [5], после чего осадки ДНК высушивали при комнатной температуре и растворяли в 40 мкл буфера ТЕ (10 мМ трис-HCl, рН 8,0; 1 мМ ЭДТА). Концентрацию фрагментированной ДНК измеряли при помощи спектрофотометра и доводили буфером ТЕ до 0,1 мкг/мкл.
По одному мкл каждого из шести гидролизатов ДНК переносили в 4 новые пробирки и в трех из них проводили обработку ДНК ферментами НаеIII (№1 контроль положительного гидролиза ДНК), GlaI (№2) и BlsI (№3), соответственно, в 20 мкл реакционной смеси. Четвертую пробирку оставляли в качестве положительного контроля. Дополнительная пробирка №5 содержала 20 мкл образца ДНК Drosophila melanogaster в буфере: 33 мМ трис-ацетат, рН 7,9; 10 мМ ацетат магния, 66 мМ ацетат калия, 1 мМ дитиотреитол в качестве отрицательного контроля. В первую добавляли 2 мкл буфера, содержащего 100 мМ трис-HCl, рН 7,6; 100 мМ MgCl2, 500 мМ Nacl, 10 мМ дитиотреитол, бидистиллированную воду до 20 мкл и 100 единиц активности рестриктазы НаеIII. Инкубировали смесь в течение 1 часа при 37°С. В пробирку №2 добавляли 2 мкл буфера, содержащего 100 мМ трис-HCl, рН 8,5; 50 мМ MgCl2, 100 мМ NaCl, 10 мМ 2-меркаптоэтанол, бидистиллированную воду до 20 мкл и 16 единиц активности эндонуклеазы Glal. Инкубировали смесь в течение 1 часа при 30°С. В каждую третью пробирку добавляли по 2 мкл буфера, содержащего 100 мМ трис-HCl, рН 8,5; 100 мМ MgCl2, 1 M NaCl, 10 мМ дитиотреитол, бидистиллированную воду до 20 мкл и 10 единиц активности эндонуклеазы BlsI. Инкубировали смесь в течение 1 часа при 30°С. В пробирку №4 (контроль гидролиза) и №5 (контроль ДНК) добавляли только 2 мкл буфера, содержащего 330 мМ трис-ацетат, рН 7,9; 100 мМ ацетат магния, 660 мМ ацетат калия, 10 мМ дитиотреитол и воду до 20 мкл. Инкубировали контрольную пробирку в течение 1 часа при 37°С.
По истечении времени инкубации проводили термоинактивацию ферментов, после чего из каждой пробирки отбирали по одному мкл реакционной смеси и переносили в 4 новые пробирки с 24 мкл буфера, содержащего 67 мМ трис-HCl, рН 8,8; 16,6 мМ (NH4)2SO4, 0,01% Tween-20, 3 мМ MgCl2; однократный стабилизатор из "Набора для GC-ПЦР"; 0,4 мкМ ДНК-праймер РК51 5'-GAACCGTGTTTCCCTAGAACCCAGTC-3' и 0,4 мкМ ДНК-праймер РК387 5'-CGGTCCGGCTGTCCTCCTCAC-3'; 0,2 мМ каждый dNTP, 60 единиц/мл Hot Start Taq ДНК-полимеразы. Поверх реакционной смеси наслаивали по 30 мкл минерального масла. Сбрасывали капли в настольной центрифуге в течение 30 сек при 4000 g. ПЦР проводили на амплификаторе «Терцик». Пробирки ставили в прибор, предварительно прогретый до 95°С.Профиль амплификации был следующим: 95°С 5 мин, далее 40 циклов 95°С 30 сек, 66°С 20 сек, 72°С 25 сек; завершение реакции 72°С 1 мин. После окончания ПЦР в каждую пробирку добавляли по 2 мкл буфера для нанесения, 12 мкл полученной смеси наносили на дорожку 1,2% агарозного геля и проводили электрофорез.
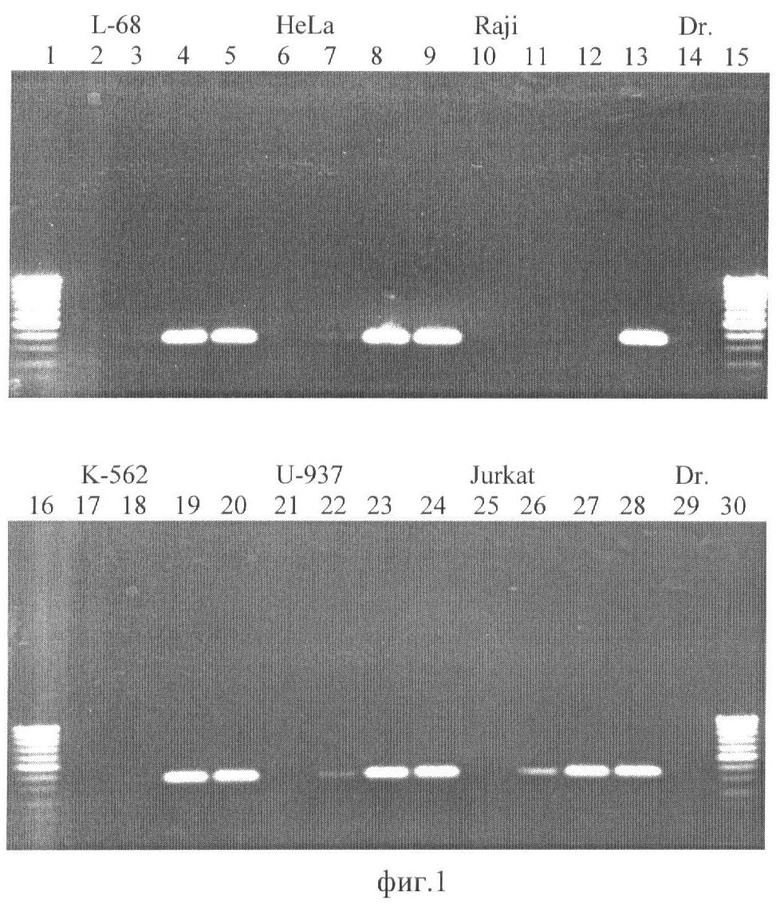
На фиг.1 приведена фотография агарозного геля после разделения продуктов ПЦР, где: ДНК L-68 (дор. 2-5); HeLa (дор. 6-9); Raji (дор. 10-13); К-562 (дор. 17-20); U-937 (дор. 21-24); Jurkat (дор. 25-28), расщепление НаеIII (дор. 2, 6, 10, 17, 21, 25); расщепление Glal (дор. 3, 7, 11, 18, 22, 26); расщепление BlsI (дор. 4, 8, 12, 19, 23, 27); TaqI-контроль (5, 9, 13, 20, 24, 28). Дорожки 14 and 29 - ПЦР с контрольной ДНК (Drosophila melanogaster). Дорожки 1, 15, 16, 30-100 bp SE-маркер длин ДНК (100, 200, 300,… 1000 пар оснований).
Из фиг.1 видно, что в отрицательном контроле с ДНК дрозофилы (Dr.) не наблюдается амплифицированных фрагментов (дор. 14 и 29), а фермент НаеIII, имеющий сайт узнавания внутри амплифицируемого фрагмента, расщепляет геномную ДНК, вследствие чего также не происходит амплификация фрагмента ДНК (дор. 2, 6, 10, 17, 21, 25). В то же время с исследуемыми ДНК человека после предварительного гидролиза TaqI (дор. 5, 9, 13, 20, 24, 28) в реакции ПНР образуется фрагмент длиной 357 пар оснований.
Полученные результаты показывают, что эндонуклеаза GlaI в значительной степени расщепляет все ДНК (дор. 3, 7, 11, 18, 22, 26), тогда как BlsI расщепляет только ДНК из клеток Raji (дор. 12). Таким образом, CpG островок в промоторной области гена-супрессора DAPK1 в ДНК из клеток Raji содержит дополнительные метилированные CG-динуклеотиды и является гиперметилированным по сравнению с препаратами ДНК из здоровых клеток линии L-68 и ДНК из остальных клеточных линий.
Пример 2. Определение гиперметилированного CpG островка в области промотора и первого экзона гена-супрессора RARB
Предварительную фрагментацию и очистку шести ДНК культур клеток человека и контрольной ДНК дрозофилы рестриктазой TaqI проводили аналогично примеру 1. Последующее расщепление полученных фрагментов также проводили в условиях, описанных в примере 1, за исключением того, что для положительного контроля гидролиза вместо 100 единиц активности рестриктазы HaeIII использовали 20 единиц активности рестриктазы FatI, a GlaI брали в количестве 10 единиц активности, BlsI - в количестве 16 единиц активности.
ПНР осуществляли, как описано в примере 1, за исключением того, что использовали ДНК-праймеры AR118 5'- CCG GGT AGG GTT CAC CGA AAG TTC ACT CGC -3' и AR941 5'- ТСА GCA AAG GGA АТС ААТ ATG CAT GCC AGC -3', а профиль амплификации был следующим: 95°С 5 мин, далее 40 циклов 95°С 30 сек, 68°С 20 сек, 72°С 50 сек; завершение реакции 72°С 1 мин. После окончания ПЦР анализ проводили аналогично примеру 1.
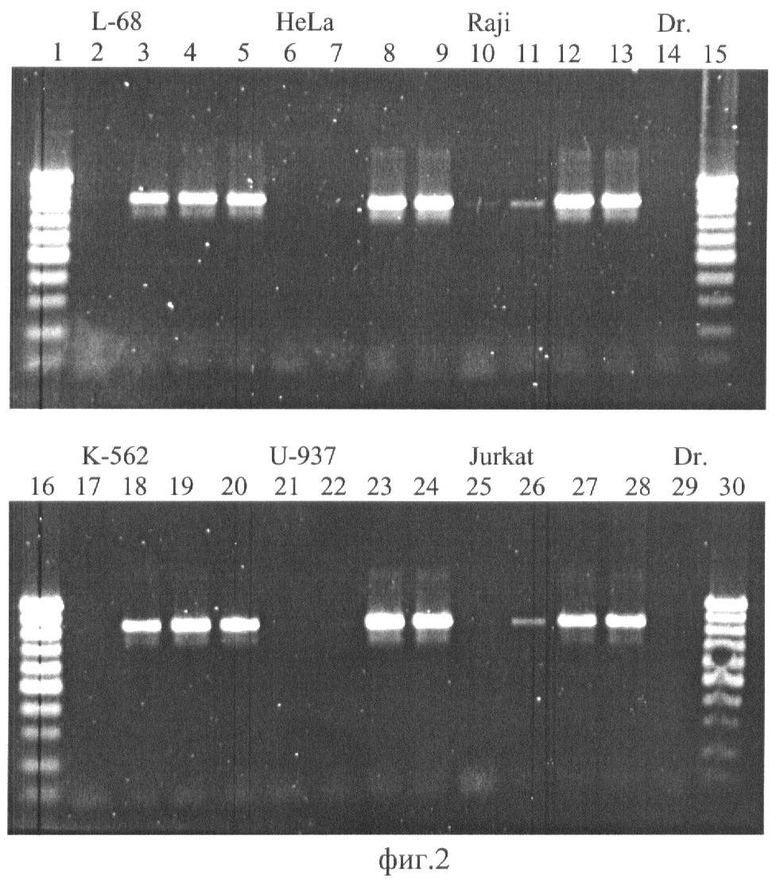
На фиг.2 приведена фотография агарозного геля после разделения продуктов ПЦР, где: ДНК L-68 (дор. 2-5); HeLa (дор. 6-9); Raji (дор. 10-13); К-562 (дор. 17-20); U-937 (дор. 21-24); Jurkat (дор. 25-28), расщепление FatI (дор. 2, 6, 10, 17, 21, 25); расщепление GlaI (дор. З, 7, 11, 18, 22, 26); расщепление BlsI (дор. 4, 8, 12, 19, 23, 27); TaqI-контроль (5, 9, 13, 20, 24, 28). Дорожки 14 and 29 - ПЦР с контрольной ДНК (Drosophila melanogaster). Дорожки 1, 15, 16, 30-100 bp SE-маркер длин ДНК (100, 200, 300,… 1000 пар оснований).
Из фиг.2 видно, что в контроле с ДНК дрозофилы (Dr.) не наблюдается амплифицированных фрагментов (дор. 14 и 29), а фермент FatI, имеющий сайт узнавания внутри амплифицируемого фрагмента, расщепляет геномную ДНК, вследствие чего также не происходит амплификация фрагмента ДНК (дор. 2, 6, 10, 17, 21, 25). В то же время с исходной ДНК после гидролиза TaqI (дор. 5, 9, 13, 20, 24, 28) в реакции ПЦР образуется фрагмент длиной 853 пары оснований. Полученные результаты показывают, что эндонуклеаза GlaI в значительной степени расщепляет ДНК HeLa, Raji, U-937, Jurkat (дор. 7, 11, 22, 26), тогда как BlsI не расщепляет ни одну из шести ДНК (4, 8, 12, 19, 23, 27), что объясняется тем, что в исследуемом районе не встречается такой последовательности, когда два сайта 5'-PuCGPy-3' разделены одним нуклеотидом и, соответственно, BlsI не может гидролизовать данный участок ДНК. Таким образом, ДНК из клеток HeLa, Jurkat, Raji, U-937 имеет в рассматриваемом фрагменте дополнительные метилированные CG-динуклеотиды по сравнению с препаратами ДНК из клеточных линий К-562 и L-68.
Пример 3. Определение гиперметилированных CG-динуклеотидов в области первого экзона гена-супрессора RASSF1
Предварительную фрагментацию и очистку шести ДНК культур клеток человека и контрольной ДНК дрозофилы рестриктазой TaqI проводили аналогично примеру 1. Последующее расщепление полученных фрагментов также проводили в условиях, описанных в примере 1, за исключением того, что вместо 100 единиц активности рестриктазы НаеIII использовали 20 единиц активности рестриктазы Bme18I в буфере, содержащем 50 мМ трис-HCl, рН 7,6; 10 мМ MgCl2, 100 мМ Nacl, 1 мМ дитиотреитол, а GlaI и BlsI брали в количестве 20 единиц активности.
ПЦР осуществляли, как описано в примере 1, за исключением того, что использовали ДНК-праймеры SF104 5'- CCG GAT GTG GGG ACC CTC TTC CTC TAG С-3' и SF879 5'- CTG TGG CCC AGA TAC GAG TGG AGT GCG AC-3', а профиль амплификации был следующим: 95°С 5 мин, далее 40 циклов 95°С 30 сек, 70°С 20 сек, 72°С 50 сек; завершение реакции 72°С 1 мин. После окончания ПЦР анализ проводили аналогично примеру 1.
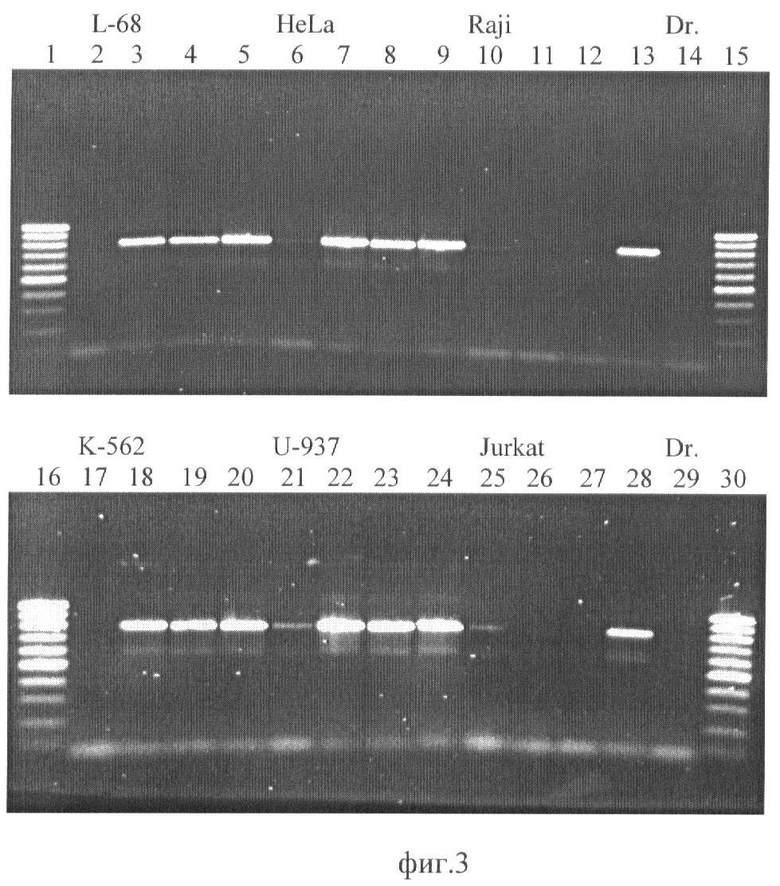
На фиг.3 приведена фотография агарозного геля после разделения продуктов ПЦР, где: ДНК L-68 (дор. 2-5); HeLa (дор. 6-9); Raji (дор. 10-13); К-562 (дор. 17-20); U-937 (дор. 21-24); Jurkat (дор. 25-28), расщепление Bme18I (дор. 2, 6, 10, 17, 21, 25); расщепление GlaI (дор. 3, 7, 11, 18, 22, 26); расщепление BlsI (дор. 4, 8, 12, 19, 23, 27); TaqI-контроль (5, 9, 13, 20, 24, 28). Дорожки 14 and 29 - ПЦР с контрольной ДНК (Drosophila melanogaster). Дорожки 1, 15, 16, 30-100 bp SE-маркер длин ДНК (100, 200, 300,… 1000 пар оснований).
Из фиг.3 видно, что в контроле с ДНК дрозофилы (Dr.) не наблюдается амплифицированных фрагментов (дор. 14 и 29), а фермент Bmel8I, имеющий сайт узнавания внутри амплифицируемого фрагмента, расщепляет геномную ДНК, вследствие чего также не происходит амплификация фрагмента ДНК (дор. 2, 6, 10, 17, 21, 25). В тоже время с исходной ДНК после гидролиза TaqI (дор. 5, 9, 13, 20, 24, 28) в реакции ПНР образуется фрагмент длиной 805 пар оснований. Полученные результаты показывают, что эндонуклеазы GlaI и BlSI расщепляют только ДНК Jurkat и Raji (дор. 11,12 и 26, 27), то есть образцы ДНК из клеток Jurkat и Raji имеют в рассматриваемом фрагменте дополнительные метилированные CG-динуклеотиды по сравнению с препаратами ДНК из клеточных линий К-562, L-68, HeLa и U-937.
Использование заявляемого способа позволит более полно и эффективно определелять гиперметилированные CpG островки, находящиеся в области промотора и первого экзона генов-супрессоров опухолевого роста в ДНК человека.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Kim, S., Li, M., Paik,H., Nephew, K., Shi, H., Kramer, R., Xu, D., Huang, T-H. Predicting DNA methylation susceptibility using CpG flanking sequences. // Pacific Symposium on Biocomputing. - 2008. - V.13. - P.135-326.
2. Sulewska, A., Niklinska, W., Kozlowski, M., Minarowski, L., Naumnik, W., Niklinski, J., Dabrowska, K., Chyczewski, L. DNA methylation in states of cell physiology and pathology. // Folia Histochemica et Cytobiologica. - 2007. - V.45.-P.149-158.
3. Bird, A. P. and Southern, E. M. Use of restriction enzymes to study eukaryotic DNA methylation: I. The methylation pattern in ribosomal DNA from Xenopus laevis. // J. Mol. Biol. - 1978. - V.118. - P.27-47.
4. Miracca, E.C., Kowalski L.P. and Nagai M.A. High prevalence of p 16 genetic alterations in head and neck tumors. // British J. Of Cancer. - 1999. -V.81.-P.677-683.
5. Sambrook, J. and Russell, D. / Molecular cloning: A laboratory manual, 3rd edition. Cold Spring Harbor Laboratory Press, - Cold Spring Harbor, New York., 2001.
6. Tarasova, G.V., Nayakshina, T.N., Degtyarev, S.Kh. Substrate specificity of new methyl-directed DNA endonuclease GlaI. // BMC Molecular Biology - 2008. - V.9:7.
7. Чернухин В.А., Томилова Ю.Э., Чмуж Е.В., Соколова О.О., Дедков B.C., Дегтярев С.Х. // Вестник биотехнологии и физико-химической биологии имени Ю. А. Овчинникова. - 2007. -№3. - С.28-33.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ Pu(5mC)GPy В ЗАДАННОМ ПОЛОЖЕНИИ ПРОТЯЖЕННОЙ ДНК | 2013 |
|
RU2525710C1 |
| СПОСОБ КАРТИРОВАНИЯ ПОЛОЖЕНИЙ РЯДА МЕТИЛИРОВАННЫХ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ Pu(5mC)GPy В ПРОТЯЖЕННОЙ ДНК ДЛЯ ПОСТРОЕНИЯ ЭПИГЕНЕТИЧЕСКОГО ПРОФИЛЯ И ВЫЯВЛЕНИЯ АНОМАЛЬНО МЕТИЛИРОВАННЫХ УЧАСТКОВ ДНК | 2015 |
|
RU2586502C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ МЕТИЛИРОВАНИЯ САЙТОВ PuCGPy РЕГУЛЯТОРНЫХ ОБЛАСТЕЙ ГЕНОВ-ОНКОМАРКЕРОВ КОЛОРЕКТАЛЬНОГО РАКА МЕТОДОМ GLAD-ПЦР-АНАЛИЗА И ОЛИГОНУКЛЕОТИДНЫЕ ПРАЙМЕРЫ И ФЛУОРЕСЦЕНТНО-МЕЧЕНЫЕ ЗОНДЫ ДЛЯ ОСУЩЕСТВЛЕНИЯ УКАЗАННОГО СПОСОБА | 2015 |
|
RU2596404C1 |
| Способ определения метилирования сайтов PuCGPy регуляторных областей генов-онкомаркеров колоректального рака методом GLAD-ПЦР-анализа и набор олигонуклеотидных праймеров и флуоресцентно-меченых зондов для осуществления указанного способа | 2016 |
|
RU2630669C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ R(5mC)GY В ЗАДАННОМ ПОЛОЖЕНИИ ПРОТЯЖЕННОЙ ДНК | 2015 |
|
RU2587631C1 |
| Способ обнаружения и анализа метилирования геномных участков ДНК в биологических образцах перифирической крови больных неходжкинской лимфомой | 2022 |
|
RU2804962C1 |
| ШТАММ БАКТЕРИИ Kocuria rosea - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ KroI | 2009 |
|
RU2394099C1 |
| СПОСОБ САЙТ-СПЕЦИФИЧЕСКОГО ГИДРОЛИЗА С5-МЕТИЛИРОВАННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК | 2015 |
|
RU2597985C1 |
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
Изобретение относится к биотехнологии и может быть использовано в медицинской практике. Предложен способ определения гиперметилированных CpG островков в области генов-супрессоров опухолевого роста в ДНК человека, который предусматривает получение образцов высокоочищенной ДНК; фрагментацию выделенной ДНК эндонуклеазой рестрикции, не имеющей сайта узнавания в амплифицируемом районе, в частности TaqI; гидролиз фрагментированной ДНК метилзависимыми сайт-специфическими эндонуклеазами GlaI и BlsI, проводимый в двух раздельных пробах; последующую амплификацию продуктов гидролиза с использованием пары праймеров, фланкирующих содержащую CpG островки область гена и составление заключения на основании изменений по сравнению с контролем, определяемых по меньшей мере в одной из двух опытных проб при их электрофоретическом разделении. Способ по изобретению обеспечивает возможность эффективной диагностики раковых заболеваний. 1 з.п. ф-лы, 3 ил.
1. Способ определения гиперметилированных CpG островков в последовательности ДНК гена-супрессора опухолевого роста человека, включающий расщепление образца геномной ДНК, чувствительной к метилированию эндонуклеазой, ПЦР-амплификацию продуктов расщепления с использованием пары праймеров, фланкирующих содержащую CpG островки область гена, анализ ПЦР-продуктов методом электрофореза и составление заключения на основе его результатов, отличающийся тем, что исходную ДНК предварительно фрагментируют эндонуклеазой рестрикции, не имеющей сайта узнавания в амплифицируемом районе, последующее расщепление фрагментированной ДНК проводят параллельно в двух образцах, которые обрабатывают метилзависимыми сайт-специфическими эндонуклеазами GlaI и BlsI, а при составлении заключения по результатам электрофоретического анализа учитывают изменения по сравнению с контролем, определяемые, по меньшей мере, в одном из двух указанных опытных образцов.
2. Способ по п.1, отличающийся тем, что для предварительного фрагментирования исходной ДНК используют эндонуклеазу рестрикции TaqI.
| MIRACCA E | |||
| et al | |||
| British Journal of Cancer, 1999, 81(4), 677-683 | |||
| SINGER-SAM J | |||
| et al | |||
| Molecular and Cellular Biology | |||
| Способ приготовления консистентных мазей | 1919 |
|
SU1990A1 |
| ZEBALA J | |||
| et al | |||
| J | |||
| Biol.Chem | |||
| Тепловой измеритель силы тока | 1921 |
|
SU267A1 |
| ЧЕРНУХИН В.А | |||
| и др | |||
| Вестник биотехнологии и физико-химической биологии им | |||
| Ю.А.Овчинникова, 2007, 3, №4, 19-27. | |||

