Изобретение относится к области биотехнологии и может быть использовано для исчерпывающего сайт-специфического гидролиза по С5-метилированным последовательностям ДНК.
При проведении эпигенетического анализа ДНК, например при сравнении статуса метилирования ДНК малигнантных и здоровых клеток человека, анализируют преимущественно сайты узнавания метилзависимых сайт-специфических ДНК-эндонуклеаз (MD-эндонуклеаз), которые расщепляются с низкой эффективностью, что существенно ограничивает возможности использования данных ферментов.
Поскольку точность эпигенетического анализа ДНК, используемого, в частности, в онкодиагностике, напрямую зависит от полноты гидролиза С 5-метилированной ДНК [1], задача усовершенствования способов гидролиза С5-метилированной ДНК является актуальной.
К настоящему времени известны способы сайт-специфического гидролиза С5-метилированных последовательностей ДНК, включающие подготовку реакционной смеси, содержащей буфер (10-50 мМ Трис-ацетат или Трис-HCl, рН 7.8-8.5, 10 мМ магния ацетат или 10 мМ магния хлорид, 1 мМ дитиотреитол, NaCl или KCl в концентрации до 100 мМ) и образец С5-метилированной ДНК, добавление к смеси MD-эндонуклеазы, инкубирование смеси с последующим анализом результата гидролиза ДНК с помощью гель-электрофореза. В частном варианте для увеличения активности и специфичности расщепления в реакционную смесь добавляют спермидин в концентрации около 8 мМ [2].
Известно также, что для увеличения активности некоторых сайт-специфических ДНК-эндонуклеаз в реакционную смесь дополнительно добавляют олигонуклеотидные дуплексы, содержащие сайты узнавания данных ферментов [3].
Недостатком известных способов является низкая эффективность гидролиза С5-метилированной последовательности ДНК.
Наиболее близким к заявляемому способу - прототипом - является способ сайт-специфического гидролиза С5-метилированной последовательности ДНК, включающий подготовку реакционной смеси, содержащей буфер (33 мМ Трис-ацетат или Трис-HCl, рН 8.5, 10 мМ магния ацетат, 1 мМ дитиотреитол, 66 мМ калия ацетат) и образец С5-метилированной ДНК, добавление к смеси MD-эндонуклеазы Glal с удельной активностью 10 е.а./мкл, инкубирование смеси в течение часа при 30°C с последующим анализом результата гидролиза ДНК с помощью гель-электрофореза [4].
Недостатком данного способа является низкая эффективность гидролиза С5-метилированной последовательности ДНК.
Задачей изобретения является обеспечение высокоэффективного исчерпывающего сайт-специфического гидролиза по С5-метилированным последовательностям ДНК с помощью MD-эндонуклеаз.
Технический результат: повышение эффективности сайт-специфического гидролиза С5-метилированной последовательности ДНК.
Поставленная задача достигается предлагаемым способом, заключающимся в следующем.
К реакционной смеси, содержащей реакционный буфер (33 мМ Трис-ацетат, рН 7.9, 10 мМ магния ацетат, 66 мМ калия ацетат, 1 мМ дитиотреитол) и образец С5-метилированной ДНК, добавляют диметилсульфоксид (DMSO) до конечной концентрации 15-25% и препарат MD-эндонуклеазы. Смесь инкубируют в течение 1 часа при 30-37°C.
Конкретное количество реагентов и объем реакционной смеси зависят от количества С5-метилированной ДНК, которую необходимо подвергнуть сайт-специфическому гидролизу с целью определения уровня ее метилирования. В классическом (типовом) варианте к 20 мкл реакционной смеси, содержащей однократный реакционный буфер и образец С5-метилированной ДНК в количестве 0,1-1 мкг, добавляют 3-5 мкл диметилсульфоксида (до конечной концентрации 15-25%, соответственно) и 1 мкл препарата MD-эндонуклеазы. Смесь инкубируют в течение 1 часа при 30-37°C.
Результаты гидролиза анализируют с помощью электрофореза в 1% агарозном геле в буфере ТАЕ x1, содержащем 0,04 М Трис-HCl, рН 8.4, 0,02 М уксусной кислоты, 1 мМ ЭДТА.
В качестве реакционного буфера в смеси может быть использован буфер, содержащий 33 мМ Трис-ацетат, рН 7.9, 10 мМ магния ацетат, 66 мМ калия ацетат, 1 мМ дитиотреитол, или буфер, содержащий 10-50 мМ Трис-HCl, рН 7.5-9.0, NaCl с концентрацией до 100 мМ, 1 мМ дитиотреитол.
В качестве MD-эндонуклеазы может быть использована GlaI [4] с концентрацией (удельной активностью) 4-8 е.а./мкл или PcsI [5] с концентрацией 1-2 е.а./мкл.
Предлагаемый способ позволяет провести исчерпывающий сайт-специфический гидролиз MD-эндонуклеазами по всем сайтам узнавания на конкретной ДНК, поскольку при использовании традиционных методов гидролиза часть сайтов не расщепляется полностью. Так, фермент PcsI становится способным эффективно расщеплять все последовательности вида 5′-(5mC)GNNNN^NNN(5mC)G-3′, а фермент GlaI - следующие С5-метилированные последовательности ДНК:
5′-G(5mC)G(5mC)-3′/3′-(5mC)G(5mC)G-5′;
5′-R(5mC)G(5mC)-3′/3′-YG(5mC)G-5′;
5′-A(5mC)GT-3′/3′-TG(5mC)A-5′;
5′-G(5mC)R(5mC)-3′/3′-(5mC)GYG-5′;
5′-G(5mC)GT-3′/3′-CG(5mC)A-5′;
5′-G(5mC)GC-3′/3′-CG(5mC)G-5′.
Определяющими отличиями заявляемого способа, по сравнению с прототипом, являются:
1. В реакционную смесь добавляют DMSO до конечной концентрации 15-25%, что позволяет повысить эффективность гидролиза С5-метилированных последовательностей ДНК MD-эндонуклеазами в 10-32 раза.
2. В реакционную смесь добавляют соответствующее количество MD-эндонуклеазы с определенной концентрацией (GlaI - 4-8 е.а./мкл, а PcsI - 1-2 е.а./мкл), что позволяет эффективно гидролизовать С5-метилированные последовательности ДНК без проявления MD-эндонуклеазами неспецифической активности. При гидролизе С5-метилированных последовательностей ДНК способом-прототипом данные MD-эндонуклеазы не способны расщеплять вышеуказанные последовательности полностью, что резко ограничивает применение этих ферментов в молекулярной биологии, генной инженерии, эпигенетике и онкодиагностике.
Изобретение иллюстрируется следующими примерами конкретного выполнения.
Пример 1. Сайт-специфический гидролиз С5-метилированной ДНК плазмиды pHspAI2 MD-эндонуклеазой GlaI в зависимости от концентрации DMSO.
Для сайт-специфического гидролиза использовали плазмиду pHspAI2, в которой во всех последовательностях 5′-GCGC-3′ С5-метилирован первый цитозин [4]. Таким образом, ДНК данной плазмиды содержит метилированные последовательности 5′-G(5mC)GC-3′/3′-CG(5mC)G-5′, которые должны расщепляться GlaI.
Для удобства анализа плазмида pHspAI2 была предварительно линеаризована эндонуклеазой рестрикции DriI.
Из исходного препарата GlaI с активностью 8 е.а./мкл путем последовательных разбавлений в буфере для хранения и разбавления ферментов «В 100» производства ООО «СибЭнзайм» (10 мМ Трис-HCl, рН 7.6, 50 мМ KCl, 0,1 мМ ЭДТА, 200 мкг/мл бычий сывороточный альбумин, 1 мМ дитиотреитол, 50% глицерин) получали препараты GlaI с концентрациями 4 е.а./мкл, 2 е.а./мкл, 1 е.а./мкл, 1/2 е.а./мкл и 1/4 е.а./мкл, то есть 6 различных растворов фермента, включая исходный и 5 разбавленных, отличающихся друг от друга концентрациями GlaI в 2 раза.
К 20 мкл реакционной смеси, содержащей реакционный буфер (33 мМ Трис-ацетат, рН 7.9, 10 мМ магния ацетат, 66 мМ калия ацетат, 1 мМ дитиотреитол) и 1 мкг ДНК плазмиды pHspAI2, линеаризованной DriI (pHspAI2/DriI), добавляли 3 мкл, 4 мкл, 5 мкл или 6 мкл DMSO (до концентрации 15%, 20%, 25% или 30% соответственно), а затем добавляли по 1 мкл каждого из полученных разведений фермента GlaI.
Поскольку растворы фермента GlaI готовили последовательным разбавлением исходного препарата в 2 раза, использование в реакции расщепления метилированной ДНК 1 мкл каждого из полученных разбавлений фермента эквивалентно тому, что в разные реакционные смеси добавляются по 1 (8 е.а.), 1/2 (4 е.а.), 1/4 (2 е.а.), 1/8 (1 е.а.), 1/16 (0,5 е.а.) и 1/32 мкл (0,25 е.а.) фермента соответственно.
Расщепление ДНК проводили в течение 1 часа при 30°C. Результаты гидролиза анализировали с помощью электрофореза в 1% агарозном геле в буфере ТАЕ x1, содержащем 0,04 М Трис-HCl, рН 8.4, 0,02 М уксусной кислоты, 1 мМ ЭДТА.
Результаты анализа приведены на фиг. 1, где дорожка 1 - 1 мкл (8 е.а.) исходного препарата GlaI; дорожка 2 - 1/2 мкл (4 е.а.) исходного препарата GlaI; дорожка 3 - 1/4 мкл (2 е.а.) исходного препарата GlaI; дорожка 4 - 1/8 мкл (1 е.а.) исходного препарата GlaI; дорожка 5-1/16 мкл (0,5 е.а.) исходного препарата GlaI; дорожка 6 - 1/32 мкл (0,25 е.а.) исходного препарата GlaI; дорожка К - контрольная ДНК (pHspAI2/DriI); М - маркер молекулярного веса ДНК 1 kb (производства ООО «СибЭнзайм»).
Как видно из набора фрагментов ДНК на фиг. 1, наиболее полный гидролиз ДНК плазмиды pHspAI2/DriI MD-эндонуклеазой GlaI наблюдается в случае использования DMSO в конечной объемной концентрации 20%. В этом случае глубина гидролиза pHspAI2/DriI на 5 дорожке (0,5 мкл препарата фермента, 20% DMSO) заметно превосходит таковую на 1 дорожке (1 мкл препарата фермента, без DMSO), а набор продуктов гидролиза на 6 дорожке (0,25 мкл препарата фермента, 20% DMSO), близка к набору продуктов расщепления этой же ДНК на 1 дорожке (1 мкл препарата фермента, без DMSO). При увеличении же концентрации DMSO до 30% активность фермента начинает ингибироваться. Таким образом, можно заключить, что активность GlaI в присутствии 20% DMSO (оптимальная концентрация) увеличивается в 16-32 раза.
Из данных, приведенных на фиг. 1, также видно, что при обработке субстратной ДНК 1 мкл препарата GlaI (8 е.а.) в реакционной смеси без DMSO гидролиз этой ДНК оказывается неполным, тогда как при использовании в буфере 15-20% DMSO сайт-специфический гидролиз становится исчерпывающим.
Пример 2. Сайт-специфический гидролиз MD-эндонуклеазой GlaI ДНК фага лямбда, предварительно метилированной по последовательности 5′-CG-3′ в зависимости от концентрации DMSO.
Для иллюстрации возможности осуществления исчерпывающего сайт-специфического гидролиза С5-метилированной ДНК при использовании в реакционном буфере DMSO и препарата MD-эндонуклазы GlaI с концентрацией 4 е.а./мкл использовали ДНК фага лямбда, в которой все CG-пары С5-метилированы метилазой SssI. Именно такой тип метилирования характерен для геномной ДНК человека, поэтому данный анализ показывает, что эффективность расщепления GlaI CG-метилированной ДНК человека увеличивается при использовании 20% DMSO.
В результате обработки метилазой SssI метилируются в том числе и последовательности 5′-RCGY-3′ с образованием сайтов узнавания GlaI: 5′-R(5mC)GY-3′/3′-YG(5mC)R-5′.
К 20 мкл реакционной смеси, содержащей реакционный буфер (33 мМ Трис-ацетат, рН 7.9, 10 мМ магния ацетат, 66 мМ калия ацетат, 1 мМ дитиотреитол) и 1 мкг ДНК фага лямбда, предварительно метилированной метилазой SssI, добавляли 4 мкл DMSO (до конечной концентрации 20%), а затем добавляли 1 мкл препарата GlaI с концентрацией 4 е.а./мкл.
Расщепление ДНК проводили в течение 1 часа при 30°C. Результаты гидролиза анализировали в 1% агарозном геле после проведения электрофореза в буфере ТАЕ x1.
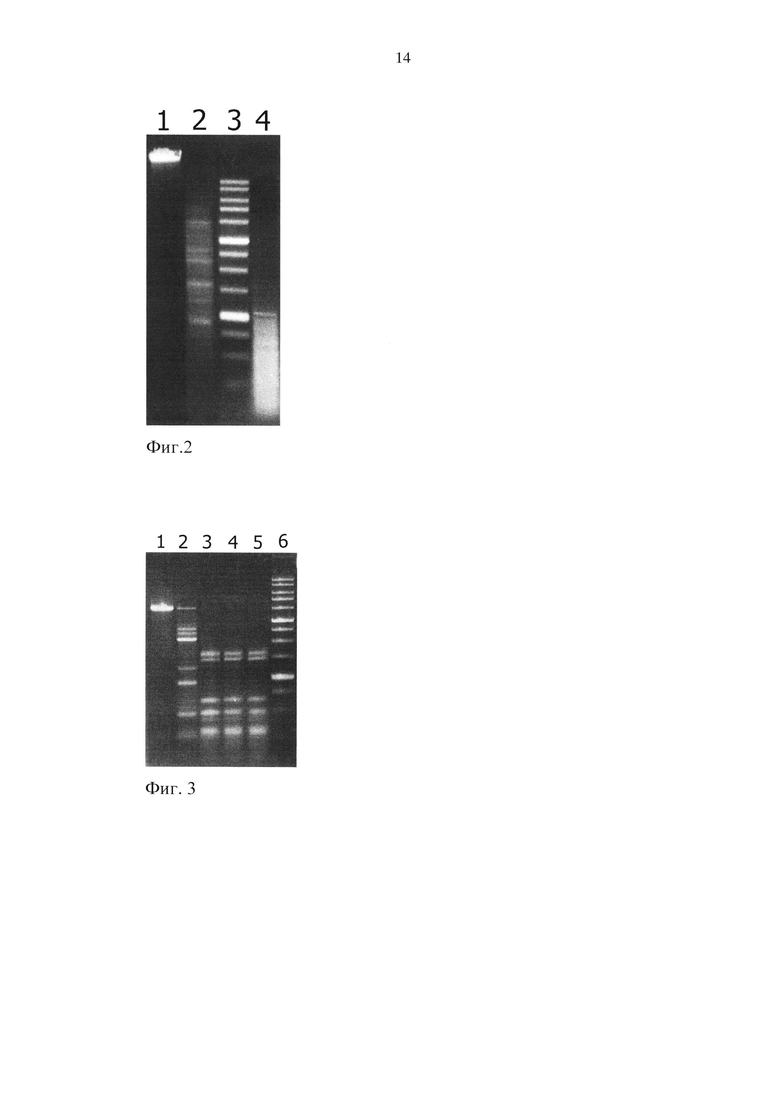
Результаты анализа приведены на фиг. 2.
Описание дорожек на электрофореграмме, изображенной на фиг. 2:
1 - контрольная ДНК фага λ+M.SssI;
2 - 1 мкл препарата (4 е.а.) GlaI в реакционной смеси без DMSO;
3 - маркер молекулярного веса ДНК 1 kb (производства ООО «СибЭнзайм»);
4-1 мкл препарата (4 е.а.) GlaI в реакционной смеси с 20% DMSO.
Как видно из фиг. 2, в случае использования DMSO в конечной концентрации 20% эффективность гидролиза CG-метилированной ДНК фага лямбда существенно возрастает (дорожка 4) по сравнению со случаем, когда реакционная смесь не содержит DMSO (дорожка 2). Согласно теоретическим расчетам, проведенным в программе Vector NTI Suite 7.1, при 100% гидролизе С5-метилированной ДНК фага лямбда по сайтам узнавания GlaI 5′-R(5mC)GY-3′/3′-CG(5mC)R-5′ максимальная длина образующихся фрагментов ДНК не превышает 1083 п.н., что соответствует картине гидролиза при использовании в реакционной смеси, содержащей DMSO в конечной объемной концентрации 20%.
Таким образом, использование в реакционной смеси 20% DMSO и 4-8 е.а. GlaI позволяет осуществить исчерпывающий сайт-специфический гидролиз С5-метилированной ДНК, тогда как без использования DMSO гидролиз оказывается неполным.
Пример 3. Сайт-специфический гидролиз С5-метилированной ДНК плазмиды pMTL-MSssI MD-эндонуклеазой PcsI в зависимости от концентрации DMSO.
Для гидролиза использовали плазмиду pMTL-MSssI, в которой во всех последовательностях 5-CG-3′ С5-метилирован цитозин. Данная плазмида была получена путем лигирования фрагмента, несущего ген метилазы SssI [6] и вектора pMTL22 [7] по сайтам узнавания эндонуклеаз рестрикции FauNDI и BamHI.
Таким образом, ДНК данной плазмиды содержит метилированные последовательности 5′-(5mC)G-3′/3′-G(5mC)-5′, которые образуют несколько сайтов узнавания фермента PcsI - последовательностей 5′-(5mC)GNNNNNNN(5mC)G-3′/3′-G(5mC)NNNNNNNG(5mC)-5′.
Для удобства анализа плазмида pMTL-MSssI была предварительно переведена в линейную форму расщеплением эндонуклеазой рестрикции DriI по уникальному сайту.
К 20 мкл реакционной смеси, содержащей реакционный буфер (33 мМ Трис-ацетат, рН 7.9, 10 мМ магния ацетат, 66 мМ калия ацетат, 1 мМ дитиотреитол) и 1 мкг ДНК плазмиды pMTL-MSssI, линеаризованной DriI (pMTL-MSssI/DriI), добавляли 3 мкл, 4 мкл или 5 мкл DMSO (до концентрации 15%, 20% или 25% соответственно), а затем добавляли по 1 мкл препарата MD-эндонуклеазы PcsI с концентрацией 1 е.а./мкл.
Расщепление ДНК проводили в течение 1 часа при 37°C. Результаты гидролиза анализировали в 1% агарозном геле после проведения электрофореза в буфере ТАЕ x1.
Результаты анализа приведены на фиг. 3.
Описание дорожек на электрофореграмме, изображенной на фиг. 3:
1 - pMTL-MSssI/DriI;
2 - pMTL-MSssI/DriI+1 мкл препарата (1 е.а.) PcsI в реакционной смеси без DMSO;
3 - pMTL-MSssI/DriI+1 мкл препарата (1 е.а.) PcsI в реакционной смеси с 15% DMSO;
4 - pMTL-MSssI/DriI+1 мкл препарата (1 е.а.) PcsI в реакционной смеси с 20% DMSO;
5 - pMTL-MSssI/DriI+1 мкл препарата (1 е.а.) PcsI в реакционной смеси с 25% DMSO;
6 - маркер молекулярного веса ДНК 1 kb (производства ООО «СибЭнзайм»).
Как видно из набора фрагментов ДНК на фиг. 3, в случае использования реакционной смеси без DMSO (дорожка 2) сайт-специфический гидролиз линеаризованной формы pMTL-MSssI (pMTL-MSssI/DriI) является неполным. В то же время при добавлении в реакционную смесь DMSO до конечных объемных концентраций 15, 20 и 25% (дорожки 3, 4 и 5 соответственно) наблюдается исчерпывающий гидролиз метилированной ДНК при ее обработке препаратом PcsI с активностью 1 е.а.
Пример 4. Сайт-специфический гидролиз С5-метилированной ДНК плазмиды pMTL-MSssI MD-эндонуклеазой PcsI.
Для гидролиза использовали плазмиду pMTL-MSssI, в которой во всех последовательностях 5-CG-3′ С5-метилирован цитозин.
К 20 мкл реакционной смеси, содержащей реакционный буфер (33 мМ Трис-ацетат, рН 7.9, 10 мМ магния ацетат, 66 мМ калия ацетат, 1 мМ дитиотреитол) и 1 мкг ДНК плазмиды pMTL-MSssI, линеаризованной DriI (pMTL-MSssI/DriI), добавляли 4 мкл DMSO (до конечной концентрации 20%), а затем добавляли 1 мкл препарата MD-эндонуклеазы PcsI с концентрацией 2 е.а./мкл.
Расщепление ДНК проводили в течение 1 часа при 37°C. Результаты гидролиза анализировали в 1% агарозном геле после проведения электрофореза в буфере ТАЕ x1. При этом наблюдался исчерпывающий гидролиз метилированной ДНК pMTL-MSssI/DriI в присутствии 20% DMSO, то есть картина гидролиза субстратной ДНК препаратом PcsI с концентрацией 2 е.а./мкл оказалась идентичной таковой, приведенной на фиг. 3, дорожка 4.
Таким образом, предложенный способ позволяет осуществить высокоэффективный исчерпывающий сайт-специфический гидролиз С5-метилированных последовательностей ДНК с помощью MD-эндонуклеаз.
Источники информации
1. Д.А. Гончар, А.Г. Акишев, С.Х. Дегтярев. BlsI- и GlaI-ПЦР-анализ - новый метод исследования метилированных участков ДНК // Вестник биотехнологии и физико-химической биологии имени Ю.А. Овчинникова. - 2010. - Т. 6. - №1. - С. 5-12.
2. Pingoud A. Spermidine increases the accuracy of type II restriction endonucleases. Suppression of cleavage at degenerate, non-symmetrical sites // Eur. J. Biochem. - 1985. - V. 147. - P. 105-109.
3. Kupper D., Reuter M., Meisel A., Krtiger D.H. Reliable detection of DNA CpG methylation profiles by the isoschizomers MspI / HpaII using oligonucleotide stimulators // Biotechniques. - 1997. - V. 23. - P. 843-847.
4. Trasova G.V., Nayakshina T.N., Degtyarev S. Kh. Substrate specificity of new methyl-directed DNA endonuclease GlaI // BMC Molecular Biology. 2008, 9:7.
5. Чернухин B.A., Наякшина Т.Н., Тарасова Г.В., Голикова Л.Н., Акишев А.Г., Дедков B.C., Михненкова Н.А., Дегтярев С.Х. Штамм бактерии Paracoccus carotinifaciens 3К - продуцент сайт-специфической эндонуклеазы Pes I // Патент на изобретение RU 2377294 С1 (2008).
6. Renbaum P. et al. Cloning, characterization, and expression in Escherichia coli of the gene coding for the CpG DNA methylase from Spiroplasma sp. strain MQ1(M.SssI) // Nucleic Acids Res. - 1990. - V.18. - P. 1145-1152.
7. Chambers S.P., Prior S.E., Barstow D.A., Minton N.P. The pMTL nic-cloning vectors. I. Improved pUC polylinker regions to facilitate the use of sonicated DNA for nucleotide sequencing // Gene. - 1988. - V. 68. - P. 139-149.
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм бактерий Micrococcus luteus 805 - продуцент сайт-специфической метилзависимой эндонуклеазы MluVI | 2016 |
|
RU2614262C1 |
| ШТАММ БАКТЕРИИ Paracoccus carotinifaciens 3K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PcsI. | 2008 |
|
RU2377294C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli N42 (pElmI) - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ ElmI | 2015 |
|
RU2597987C1 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ Pu(5mC)GPy В ЗАДАННОМ ПОЛОЖЕНИИ ПРОТЯЖЕННОЙ ДНК | 2013 |
|
RU2525710C1 |
| СПОСОБ САЙТ-СПЕЦИФИЧЕСКОГО ГИДРОЛИЗА ДНК | 2015 |
|
RU2597984C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ГИПЕРМЕТИЛИРОВАННЫХ CpG ОСТРОВКОВ В ОБЛАСТИ ГЕНОВ-СУПРЕССОРОВ ОПУХОЛЕВОГО РОСТА В ДНК ЧЕЛОВЕКА | 2009 |
|
RU2413773C1 |
| СПОСОБ КАРТИРОВАНИЯ ПОЛОЖЕНИЙ РЯДА МЕТИЛИРОВАННЫХ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ Pu(5mC)GPy В ПРОТЯЖЕННОЙ ДНК ДЛЯ ПОСТРОЕНИЯ ЭПИГЕНЕТИЧЕСКОГО ПРОФИЛЯ И ВЫЯВЛЕНИЯ АНОМАЛЬНО МЕТИЛИРОВАННЫХ УЧАСТКОВ ДНК | 2015 |
|
RU2586502C1 |
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ R(5mC)GY В ЗАДАННОМ ПОЛОЖЕНИИ ПРОТЯЖЕННОЙ ДНК | 2015 |
|
RU2587631C1 |

Изобретение относится к области биотехнологии и представляет собой способ сайт-специфического гидролиза С5-метилированной последовательности ДНК. Готовят реакционную смесь, содержащую буферный раствор и образец С5-метилированной ДНК, добавляют к смеси диметилсульфоксид до конечной концентрации 15-25%, а затем MD-эндонуклеазу, смесь инкубируют в течение часа при 30-37°C с последующим анализом результата гидролиза ДНК методом гель-электрофореза, при этом в качестве MD-эндонуклеазы используют GlaI с концентрацией 4-8 е.а./мкл или PcsI с концентрацией 1-2 е.а./мкл. Изобретение позволяет повысить эффективность сайт-специфического гидролиза С5-метилированной последовательности ДНК. 3 з.п. ф-лы, 3 ил., 4 пр.
1. Способ сайт-специфического гидролиза C5-метилированной последовательности ДНК, включающий подготовку реакционной смеси, содержащей буферный раствор и образец C5-метилированной ДНК, добавление к смеси MD-эндонуклеазы, инкубирование смеси в течение часа с последующим анализом результата гидролиза ДНК с помощью гель-электрофореза, отличающийся тем, что в реакционную смесь добавляют диметилсульфоксид до конечной концентрации 15-25%, а в качестве MD-эндонуклеазы используют GlaI с концентрацией 4-8 е.а./мкл или PcsI с концентрацией 1-2 е.а./мкл.
2. Способ по п. 1, отличающийся тем, что в качестве буферного раствора используют буфер, содержащий 10-50 мМ Трис-HCl, pH 7.5-9.0, NaCl с концентрацией до 100 мМ, 1 мМ дитиотреитол.
3. Способ по п. 1, отличающийся тем, что в качестве буферного раствора используют буфер, содержащий 33 мМ Трис-ацетат, pH 7.9, 10 мМ магния ацетат, 66 мМ калия ацетат, 1 мМ дитиотреитол.
4. Способ по п. 1, отличающийся тем, что гель-электрофорез проводят в буфере, содержащем 0,04 М Трис-HCl, pH 8.4, 0,02 М уксусную кислоту, 1 мМ ЭДТА.
| TARASOVA GV et.al | |||
| Substrate specificity of new methil-direcred DNA endonuclease GlaI, BMC Molecular Biology, 2008, 9:7 | |||
| СПОСОБ ОПРЕДЕЛЕНИЯ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ Pu(5mC)GPy В ЗАДАННОМ ПОЛОЖЕНИИ ПРОТЯЖЕННОЙ ДНК | 2013 |
|
RU2525710C1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ШТАММ БАКТЕРИИ Paracoccus carotinifaciens 3K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PcsI. | 2008 |
|
RU2377294C1 |

