Область техники, к которой относится изобретение
Изобретение в целом имеет отношение к лечению потери мышечной массы у млекопитающих, в особенности, к введению одной или нескольких аминокислоты(аминокислот) с разветвленной цепью (ВСАА), предшественника ВСАА, метаболита ВСАА, белка, богатого аминокислотами ВСАА, белка, искусственно обогащенного аминокислотами ВСАА, или к любым их комбинациям при лечении такой потери мышечной массы. Изобретение дополнительно имеет отношение к пищевым композициям, подходящим для такого введения.
Предшествующий уровень техники
Аминокислоты представляют собой мономерные строительные блоки белков, которые, в свою очередь, включают широкий диапазон биологических соединений, включающий ферменты, антитела, гормоны, транспортные молекулы для переноса ионов и небольших молекул, коллаген и мышечные ткани. Аминокислоты считаются гидрофобными или гидрофильными на основании их растворимости в воде, и, в особенности, на основании полярности их боковых цепей. Аминокислоты, имеющие полярные боковые цепи, представляют собой гидрофильные аминокислоты, тогда как аминокислоты, имеющие неполярные боковые цепи, представляют собой гидрофобные аминокислоты. Частично растворимость аминокислот определяет структуру белков. Гидрофильные аминокислоты имеют тенденцию формировать поверхности белков, тогда как гидрофобные аминокислоты имеют тенденцию формировать водонерастворимые расположенные в глубине участки белков.
У людей среди обычных 20 аминокислот девять считаются незаменимыми, так как организм не может синтезировать их сам. В значительной степени эти девять аминокислот должны поступать с пищей, которой питается индивидуум. Дефицит одной или нескольких аминокислот может привести к отрицательному балансу по азоту. Отрицательный баланс по азоту представляет собой, например, ситуацию, когда больше азота выводится, чем вводится. Такое состояние может привести к нарушению ферментативной активности и потере мышечной массы.
Был установлен ряд состояний, сопровождающихся истощением мышечной массы, для которых была доказано польза лечения аминокислотными добавками. Например, кахексия представляет собой состояние тяжелого истощения организма, характеризуемого заметной потерей массы, анорексией, астенией и анемией. Кахексия представляет собой общий признак ряда заболеваний, таких как рак, сепсис, хроническая сердечная недостаточность, ревматоидный артрит и синдром приобретенного иммунодефицита (AIDS). Известны другие заболевания и нарушения, связанные с истощением мышечной массы, включающие, например, саркопению, связанную с возрастом потерю мышечной массы.
Фактор, индуцирующий протеолиз (PIF)
Было обнаружено, что определенные опухоли могут индуцировать кахексию через образование 24 кДа гликопротеина, называемого фактором, индуцирующим протеолиз (PIF). Согласно одному из предложенных механизмов действие PIF заключается в снижении синтеза белка; согласно другому механизму действие PIF заключается в активации деградации белков; третий предполагаемый механизм представляет собой сочетание вышеупомянутого снижения в синтезе белка и активации деградации белков. Предполагается, что сниженный синтез белка, связанный с PIF, является результатом способности PIF блокировать процесс трансляции синтеза белка. Было показано, что другой фактор, ангиотензин II (Ang II), обладает похожим эффектом и может быть вовлечен в мышечное истощение, наблюдаемое в некоторых случаях при кахексии.
Известна природная роль PIF в системе убиквитин-протеосома. PIF вызывает увеличение выхода арахидоновой кислоты, которая затем метаболизирует до простагландинов и 15-гидроксиэйкозатетраеновая кислота (15-НЕТЕ). Было показано, что 15-НЕТЕ вызывает значительное усиление деградации белков и связывания в ядре транскрипционного фактора NF-κВ (ядерный фактор, который связывает усилитель гена легких каппа-цепей иммуноглобулина в В-клетках).
Регуляция синтеза белка через инициацию трансляции
Предполагается, что роль PIF в ингибировании синтеза белка связана с теоретической способностью фактора PIF блокировать трансляцию через активацию лежащих ниже факторов с помощью RNA-зависимой протеинкиназы (PKR). Ингибирование синтеза белка через фактор PIF ослабляется инсулином в физиологических концентрациях и ниже. Это означает, что PIF может ингибировать синтез белка на начальной стадии трансляции, так как инсулин регулирует синтез белка через активацию стадий связывания матричной РНК (мРНК) в инициации трансляции.
Существует две стадии в инициации трансляции, которые подчинены регуляции: (1) связывание инициирующей метионил-транспортной РНК (met-tRNA) с рибосомальной субъединицей 40s; и (2) связывание мРНК с комплексом преинициации 43s.
На первой стадии met-tRNA связывается с рибосомальной субъединицей 40s, как тройной комплекс с эукариотическим фактором инициации 2 (eIF2) и гуанозинтрифосфатом (GTP). Позднее GTP, связанный с eIF2 гидролизуется до гуанозиндифосфата (GDP) и eIF2 освобождается из рибосомальной субъединицы в комплексе GDP-eIF2. eIF2 должен затем обменять GDP на GTP для участия в другом цикле инициации. Это происходит посредством действия другого эукариотического фактора инициации, eIF2B, который опосредует обмен гуаниновых нуклеотидов на eIF2. eIF2B регулируется путем фосфорилирования фактора eIF2 по его альфа-субъединице, которое превращает его из субстрата в конкурентный ингибитор eIF2B.
На второй стадии для связывания мРНК с комплексом преинициации 43s необходима группа белков, в совокупности называемая eIF4F, представляющая собой полисубъединичный комплекс, состоящий из eIF4A (РНК-хеликазы), eIF4B (который функционирует вместе с eIF4A для расплетания вторичной структуры в нетранслируемой 5′-области мРНК), eIF4E (который связывает кэп m7GTP, присутствующий на 5′-конце мРНК) и eIF4G (который функционирует в качестве каркаса для eIF4E, eIF4A и мРНК). Вместе комплекс eIF4F служит для узнавания, развертывания и направления мРНК к комплексу преинициации 43s. Доступность eIF4E для образования комплекса eIF4F, по-видимому, регулируется репрессором трансляции еIF4Е-связывающим белком 1 (4Е-ВР1). 4Е-ВР1 конкурирует с eIF4G за связывание с eIF4E и способен перевести eIF4E в неактивный комплекс. Связывание 4Е-ВР1 регулируется фосфорилированием с помощью киназы мишени рапамицина млекопитающих (mTOR), при котором усиление фосфорилирования приводит к снижению сродства 4Е-ВР1 к eIF4E.
Полагают, что mТОR активируется путем фосфорилирования и ингибирования комплекса туберозного склероза (TSC), 1-TSC2 комплекса, посредством передачи сигнала через фосфатидилинозитол-3-киназный (PI3K) / серин/треонинкиназный путь (PI3K/AKT-путь). mTOR также фософорилирует p70S6 киназу, которая фосфорилирует рибосомальный белок S6, который, как считается, усиливает трансляцию мРНК с непрерывной нитью пиримидиновых остатков, расположенных рядом с 5'-кэп структурой. Белки, кодируемые такими мРНК, включают рибосомальные белки, факторы элонгации трансляции и поли-А-связывающие белки.
Анаболические факторы, вовлеченные в инициацию трансляции
В многочисленных исследованиях было показано, что анаболические факторы, такие как инсулин, инсулинподобные факторы роста (IGFs) и аминокислоты увеличивают синтез белка и приводят к гипертрофии мышц. Аминокислоты с разветвленной цепью (BCAAs), в особенности, лейцин, могут инициировать пути передачи сигнала, которые регулируют инициацию трансляции. Такие пути часто включают mTOR. Другие исследования показали, что митогенные стимулы, такие как инсулин и BCAAs, осуществляют передачу сигнала через eIF2. Как таковая, нехватка аминокислот приводит к увеличенному фосфорилированию eIF2-α и снижению синтеза белка.
Пути передачи сигнала, вовлеченные в синтез и деградацию белка
Как уже отмечалось, известно, что PIF индуцирует деградацию белков через NF-κВ путь. Следовательно, вероятно, что ингибирование синтеза белка с помощью PIF происходит через обычную точку инициации пути передачи сигнала, который затем разделяется на два отдельных пути, один, стимулирующий деградацию белков посредством фактора NF-κB, и другой, ингибирующий синтез белка посредством mTOR и/или eIF2.
АКТ представляет собой серин/треонинкиназу, также известную как протеинкиназа В (РКВ). Активации АКТ происходит через прямое связывание инозитольных липидных продуктов PI3K с его доменом гомологии к плекстрину. PI3K-зависимая активация АКТ также происходит через опосредуемое фосфоинозитид-зависимой киназой (PDK1) фосфорилирование треонина 308, которое приводит к автофосфорилированию серина 473. Хотя исходно считалось, что они функционируют как компоненты особого сигнального пути, некоторые исследования показали, что сигнальные пути NF-κВ и АКТ сходятся. Исследования показали, что передача сигнала через АКТ ингибирует in vitro апоптоз во множестве типов клеток, это опосредуется его способностью фосфорилировать регулирующие апоптоз компоненты, включающие IκК, киназу, вовлеченную в активацию фактора NF-κВ. Таким образом, активации АКТ стимулирует активацию фактора NF-κВ. Хотя это помещает АКТ выше по отношению к активации фактора NF-κB в последовательности событий при передаче сигнала, в одном исследовании сообщалось о том, что АКТ может быть расположенной ниже мишенью фактора NF-κB. В целом, на основании этого предположили, что АКТ вовлечена в катаболический путь. Однако на основании других данных, предположили, что АКТ также вовлечена в анаболические процессы, через активации mTOR и последующее фосфорилирование p70S6 киназы и 4Е-ВР1, приводящее к усилению синтеза белка.
PKR представляет собой интерферон-индуцируемую, RNA-зависимую серин/треонинпротеинкиназу, ответственную за контроль антивирусного защитного пути. PKR может быть индуцирована другими формами клеточного стресса, отличными от интерферона. Некоторые наблюдения позволили предположить, что фактор некроза опухолей (ТNF)-альфа также действует через PKR. Интересно, что как интерферон, так и TNF-альфа привлечены в качестве факторов, вызывающих кахектические состояния. Сообщалось, что при последующем взаимодействии с активирующими стимулами (например, с инсулином, IGF, BCAAs), происходит образование гомодимеров PKR и его автофософорилирование. В результате, PKR становится способным катализировать фосфорилирование субстратов-мишеней, наиболее хорошо охарактеризовано фосфорилирование серина 51 на субъединице eIF2-a. Затем еIF2 связывает eIF2B, скорость-лимитирующий компонент трансляции, что приводит к ингибированию синтеза белка. Последние исследования позволили заключить, что PKR физически связывается с комплексом IκК и стимулирует NF-κВ-индуцируемую киназу (NIK), тогда как форилирование 1кК приводит к его последующей деградации. В некоторых исследованиях было предположено, что фактор NF-κВ активируется посредством PKR с помощью механизма, не зависящего от активности его еIF2-киназы, тогда как результаты других исследований указывают на то, что фосфорилирование eIF2-α требуется для активации фактора NF-κВ.
PKR-подобная ER-резидент киназа (PERK) представляет собой другую киназу, которая фосфорилирует eIF2-α и активирует NF-κВ. Однако маловероятно, что PIF действует через этот путь, так как PERK вызывает освобождение IκК из фактора NF-κВ, но не его деградацию. Кроме того, было показано, что PIF вызывает деградацию IκК в ходе активации фактора NF-κВ.
Известные способы лечения потери мышечной массы
Лечение таких состояний, как кахексия, часто включает применение пищевых добавок, и, в особенности, дополнение диеты аминокислотами, при попытках увеличить синтез белка. Эти три BCAAs представляют собой валин, лейцин и изолейцин. Ранее было показано, что лейцин функционирует не только как блок для строительства белка, но также как индуктор пути передачи сигнала, который регулируют инициацию трансляции. Наши последние новые исследования позволяют предположить, что все эти три BCAAs обладают способностью снижать деградацию белков и сравнительно усиливать трансляцию белка.
Кахексия представляет собой только одно из состояний, нарушений и болезней, для которых была подтверждена польза дополнения диеты аминокислотами. Дополнение диеты аминокислотами также применяют для лечения диабета, гипертонии, высоких уровней сывороточного холестерина и триглицеридов, болезни Паркинсона, бессонницы, наркозависимости и хронического алкоголизма, болевых ощущений, бессонницы и гипогликемии. Дополнение аминокислотами BCAAs, в особенности, применяют при лечении расстройств печени, которые включают нарушения функции печени, включающие цирроз печени, нарушения функции желчного пузыря, хорею и дискинезию, и нарушение функции почек, включающее уремию. Было также подтверждено успешное применение дополнения аминокислотами ВСАА лечения пациентов, перенесших гемодиализ, приводившее к улучшению здоровья и настроения в целом.
В настоящее время лечение потери мышечной массы, содержащее лечения, включающие дополнение питания аминокислотами, сфокусировано на стимуляции анаболизма мышц. Например, в патенте US Patent Application Publication No. 2004/0122097 Verlaan et al. описали питательные добавки, содержащие как лейцин, так и белок стимуляции формирования мышечных тканей. Предшественники лейцина, такие как пируват, и метаболиты, такие как β-гидрокси-β-метилбутират и α-кетоизокапроат, проявляют свойства подобные лейцину. Следует заметить, что β-гидрокси-β-метилбутират не производится человеком в каких-либо клинически значимых количествах и, следовательно, должен пополняться.
Другие исследователи показали, что инсулин, анаболический гормон, способен активировать синтез белка при введении в больших дозах. Таким образом, известные подходы в лечении, хотя и обеспечивают некоторую пользу людям, страдающим от потери мышечной массы, путем увеличения образования мышечной ткани, не воздействуют на потерю самой мышечной массы. Это означает, что известные способы лечения потери мышечной массы в большей степени направлены на увеличение мышечного анаболизма, чем на снижение мышечного катаболизма.
Аминокислоты, включаемые в скелетную мышцу, находятся в состоянии постоянного протока, тогда как новые аминокислоты, или поступающие при введении энтеральным или парентеральным путем, или рециркулирующие, откладываются в виде белка и данные белки деградируют. Потеря мышечной массы может быть результатом многих факторов, включающих сниженную скорость синтеза белка с нормальной деградацией, усиленную деградацию с нормальным синтезом или усиление как сниженного синтеза и увеличенной деградации. В результате лечение, направленное на увеличение синтеза, адресовано только к половине проблемы заболевания (заболеваний), связанного (связанных) с мышечным истощением.
Следовательно, в этой области техники существует необходимость в способе лечения потери мышечной массы, который снижает мышечный катаболизм и, необязательно, увеличивает мышечный анаболизм.
Раскрытие изобретения
Изобретение обеспечивает способы лечения потери мышечной массы у индивидуума. В одном воплощении, изобретение включает введение индивидууму эффективного количества аминокислоты с разветвленной цепью (ВСАА), предшественника ВСАА, метаболита ВСАА, белка, богатого аминокислотой ВСАА, белка, искусственно обогащенного по содержанию ВСАА, или любые их сочетания. Изобретение дополнительно обеспечивает пищевые продукты для такого введения, включающие вводимые перорально пищевые продукты.
В первом аспекте изобретение обеспечивает способ лечения потери мышечной массы у индивидуума, включающий: введение индивидууму эффективного количества, по меньшей мере, одного из соединений: аминокислоты с разветвленной цепью (ВСАА); предшественника ВСАА; и метаболита ВСАА, белка, богатого аминокислотой ВСАА, белка, искусственно обогащенного аминокислотой ВСАА, в котором, по меньшей мере, ВСАА, предшественник ВСАА, метаболит ВСАА, белок, богатый аминокислотой ВСАА, и белок, искусственно обогащенный по содержанию ВСАА, противодействует катаболизму белков.
Во втором аспекте изобретение обеспечивает вводимый перорально пищевой продукт, включающий, по меньшей мере, одно из следующих соединений: аминокислоту с разветвленной цепью (ВСАА); предшественника ВСАА, метаболита ВСАА, белок, богатый аминокислотой ВСАА, белок, искусственно обогащенный по содержанию ВСАА, в котором, по меньшей мере, ВСАА, предшественник ВСАА, метаболит ВСАА, белок, богатый аминокислотой ВСАА, и белок, искусственно обогащенный по содержанию ВСАА, противодействует катаболизму белков.
Иллюстративные аспекты настоящего изобретения предназначены для решения проблем, описанных в этом документе, и других, не обсуждавшихся проблем, которые могут быть обнаружены специалистом в этой области техники.
Краткое описание чертежей
Эти и другие отличительные признаки изобретения будут легко понятны из следующего подробного описания различных аспектов изобретения, взятых вместе с сопроводительными чертежами, на которых изображены различные воплощения изобретения, в которых:
на фиг.1 показан график подавления синтеза белка фактором, индуцирующим протеолиз (PIF), в различных концентрациях;
на фиг.2 показан график влияния аминокислот на фосфорилирование eIF2-α и PIF;
на фиг.3 показан график влияния инсулина и инсулин-подобного фактора роста 1 (IGF) на фосфорилирование eIF2-α фактора PIF;
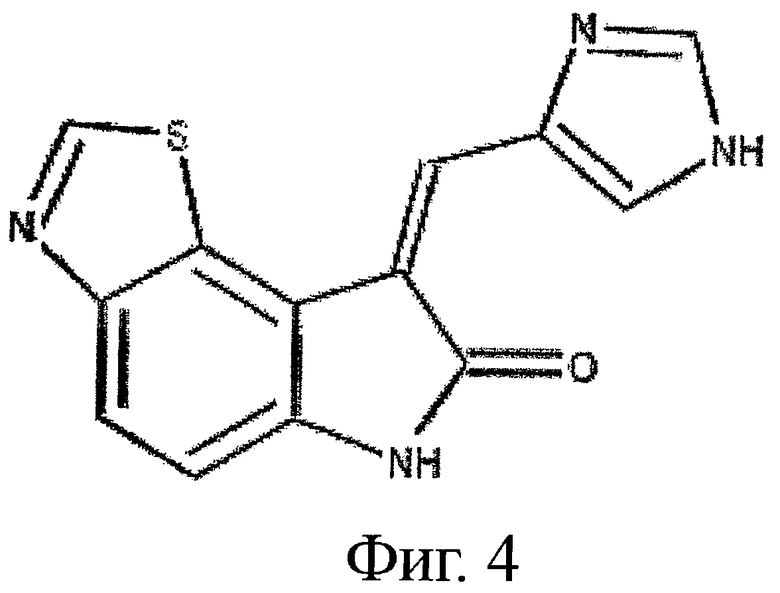
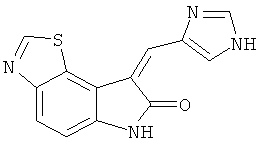
на фиг.4 показана структура ингибитора RNА-зависимой протеинкиназы (PKR), подходящего для применения в настоящем изобретении;
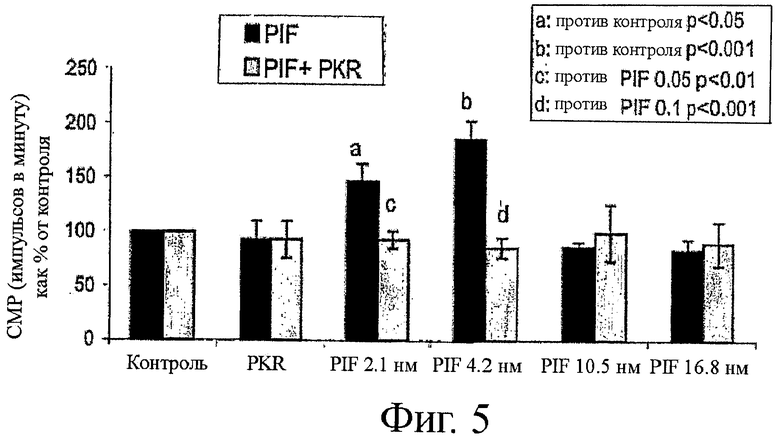
на фиг.5 показан график влияния ингибитора PKR, приведенного на фиг.4, на протеолитическую активность PIF;
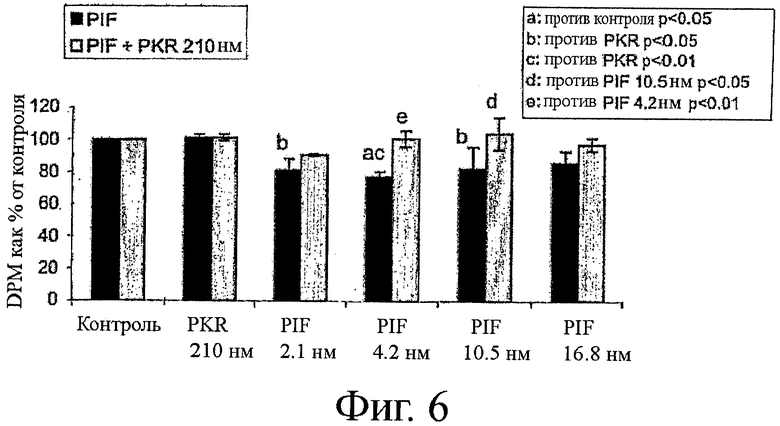
на фиг.6 показан график влияния ингибитора PKR, приведенного на фиг.4, в обращении PIF-опосредуемого восстановления синтеза белка;
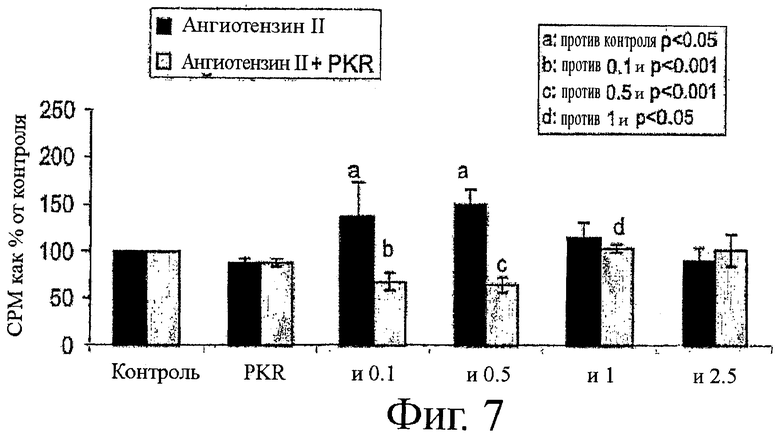
на фиг.7 показан график влияния ингибитора PKR, приведенного на фиг.4, на протеолитическую активность ангиотензина II;
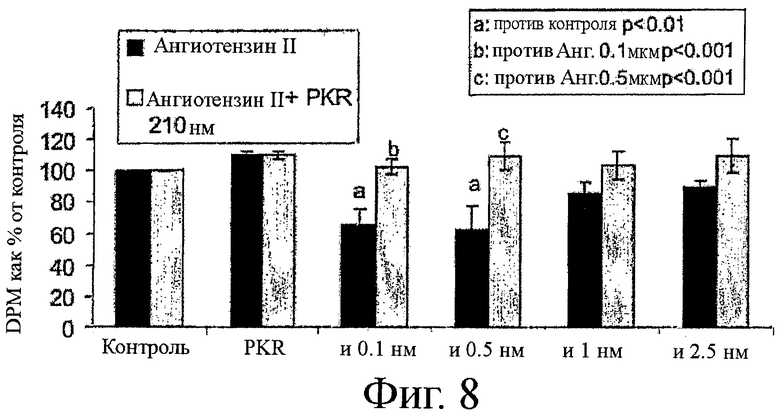
на фиг.8 показан график влияния ингибитора PKR, приведенного на фиг.4, в обращении ангиотензин II-опосредуемого восстановления синтеза белка;
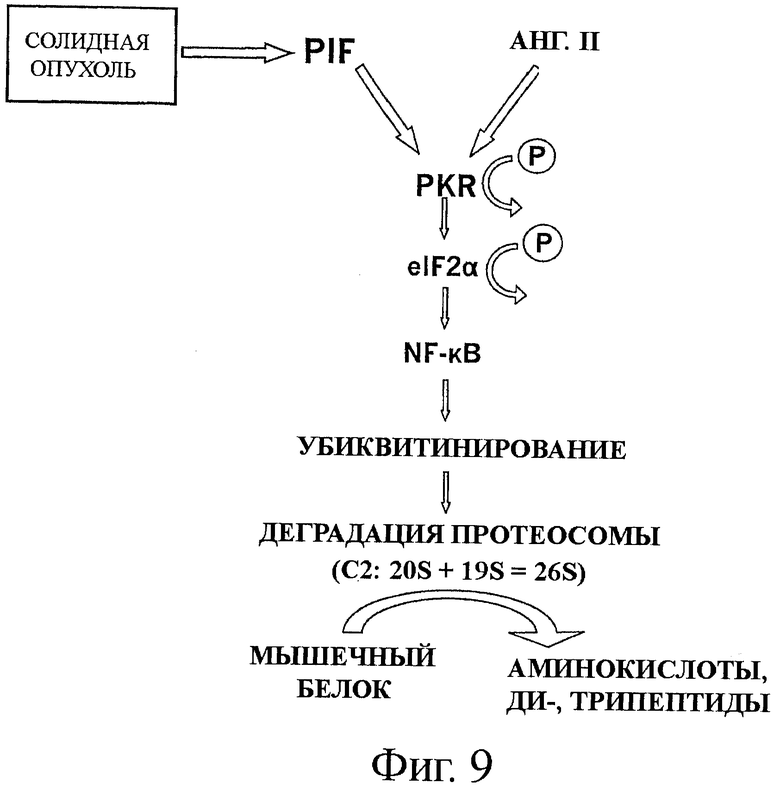
на фиг.9 показан альтернативный механизм деградации белков, вызванной фактором, индуцирующим протеолиз (PIF), и ингибируемый аминокислотами с разветвленной цепью, инсулином и IGF-1;
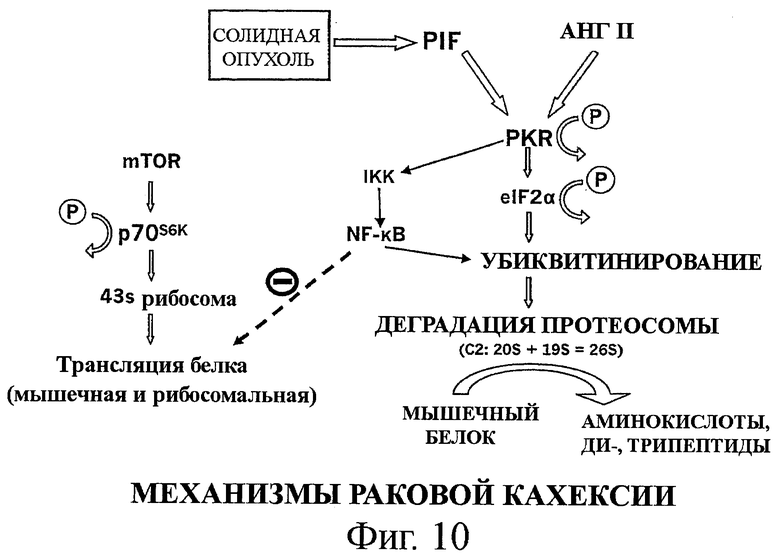
на фиг.10 показан дополнительный альтернативный механизм деградации белков, вызванной фактором, индуцирующим протеолиз (PIF), через активацию PKR и еIF2α, который ингибируется аминокислотами с разветвленной цепью, инсулином и IGF-1.
Следует отметить, что чертежи изобретения не имеют шкалы. Чертежи предназначены для изображения только типичных аспектов изобретения, и следовательно, не должны рассматриваться, как ограничивающие объем изобретения.
Осуществление изобретения
Как указано выше, изобретение обеспечивает способы и полученные этими способами продукты для лечения потери мышечной массы у индивидуума. В особенности, способы и продукты изобретения снижают мышечный катаболизм, в особенности, мышечный катаболизм, опосредуемый фактором, индуцирующим протеолиз (PIF).
Примененные в этом документе термины «лечение» и «лечить» означают, как профилактическое или предупредительное лечение, так и терапевтическое лечение или лечение, изменяющее течение заболевания, включающие лечение пациентов рискующих заболеть, или тех, что имеют подозрение на заболевание, а также пациентов уже заболевших или имеющих диагноз, как страдающие от заболевания или по медицинским показаниям. Термины «лечение» и «лечить» также означают поддержание и/или улучшение здоровья у индивидуума, не страдающего от болезни, но который может быть восприимчивым к развитию болезненного состояния, такого как нарушение азотного баланса или потеря мышечной массы. Следовательно, термин «эффективное количество» представляет собой такое количество, которое лечит заболевание или требующее терапевтического вмешательства состояние у индивидуума или, в общем, обеспечивает питательную, физиологическую или лечебную пользу индивидууму. Лечение может быть связано с пациентом или с врачом. Кроме того, так как термины «индивидуум» и «пациент», часто применяемые в этом документе, относятся к человеку, изобретение не ограничено человеком. Соответственно, термины «индивидуум» и «пациент» означают любое млекопитающее, страдающее от или рискующее попасть в такое требующее терапевтического вмешательства состояние, как потеря мышечной массы.
Экспериментальные данные
С целью определить эффективность аминокислот с разветвленной цепью (BCAAs) и других агентов в снижении мышечного катаболизма, мышиные C2C12 мышечные трубочки подвергали воздействию PIF или ангиотензина II в комбинации с аминокислотами (включающими BCAAs), инсулином, инсулинподобным фактором роста-1 (IGF-1) и ингибитора PKR. PIF экстрагируют и очищают из опухоли MAC 16, как описано в работе Smith et al., Effect of a Cancer Cachectic Factor on Protein Synthesis/Degradation in Murine C2C12 Myoblasts: Modulation by Eicosapentaenoic Acid, Cancer Research, 59:5507-13 (1999), которая таким образом включена путем отсылки. Деградацию белков определяют, применяя способ, описанный в работе Whitehouse et al., Increased Expression of the Ubiquitin-Proteasome Pathway in Murine Myotubes by Proteolysis-Inducing Factor (PIF) is Associated with Activation of the Transcription Factor NF-кВ, British Journal of Cancer, 89:1116-22 (2003), которая также, таким образом, включена путем отсылки.
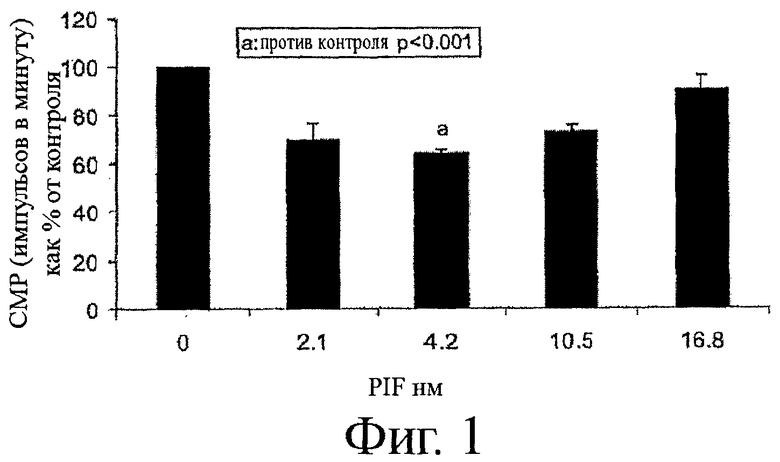
На фиг.1 показан график подавления синтеза белка при увеличении концентрации PIF, измеренный в импульсах в минуту (СРМ) как процент от контроля, не содержащего PIF. Было отмечено значительное снижение в синтезе белка, с максимальным подавлением синтеза белка, происходящим при концентрации PIF равным 4,2 нМ. Измеренная протеолитическая активность PIF, в особенности, может быть описана как активность убиквитинподобной деградации.
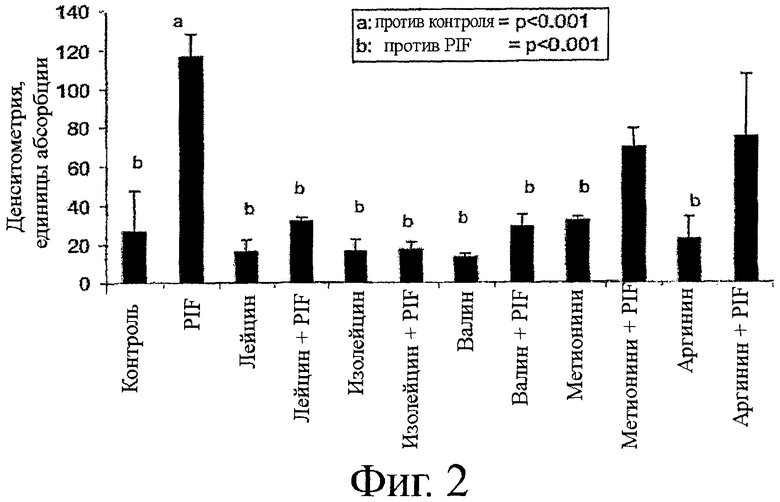
На фиг.2 показан график денситометрического анализа вестерн-блотов фосфорилированного eIF2-α в мышечных трубочках С2С12, инкубируемых с PIF, лейцином, изолейцином, валином, метионином и аргинином, как по одному, и в комбинации с PIF. Контрольный образец инкубируют только в фосфатном буферном солевом растворе (PBS). Как можно видеть на фиг.2, PIF значительно увеличивает фосфорилирование eIF2-α, по сравнению с контролем. Каждая из аминокислот снижает фосфорилирование elF2-α в присутствии PIF, по сравнению с одним PIF. Однако BCAAs (т.е. лейцин, изолейцин, и валин) снижают такое фосфорилирование примерно до уровня контроля или ниже, тогда как уровни метионин- и аргинин-индуцированного фосфорилирования были выше, чем в контроле. Удивительно, что в отличие от известных способов лечения, направленных на увеличение синтеза белка, и в которых лейцин проявлял более высокую эффективность, чем другие BCAAs, эти данные показали, что все BCAAs обладают примерно одинаковой эффективностью в снижении PIF-индуцированного фосфорилирования eIF2-α. Фактически, уровни фосфорилирования, возникшие в результате инкубации с изолейцином и валином, не отличались от тех, что наблюдали при инкубации с лейцином.
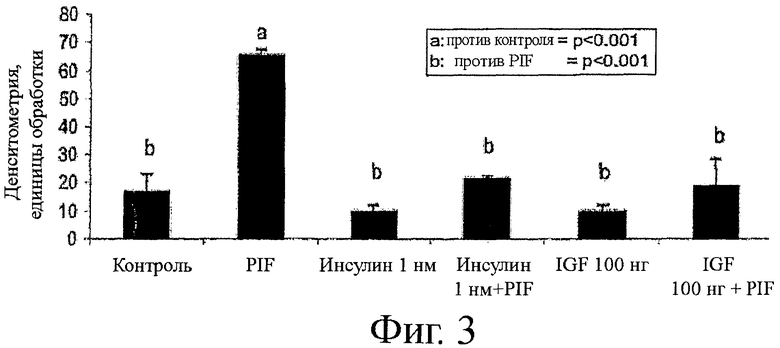
На фиг.3 показаны результаты похожих экспериментов, включающие инкубацию с инсулином и IGF-1, по одному и в комбинации с PIF. Как инсулин, так и IGF-1 значительно снижает фосфорилирование eIF2-α в присутствии PIF, по сравнению с одним PIF. Таким образом, способность BCAAs снижать PIF-опосредуемую деградацию белков может быть дополнена или усилена добавлением инсулина и/или IGF-1 или лечениями, которые увеличивают уровень инсулина и/или IGF-1.
На фиг.4 показана структура ингибитора PKR, применимого как в снижении PIF-индуцированной деградации белков, так и в увеличении синтеза белка, который применяли в качестве позитивного контроля ингибирования PKR. На фиг.5-8 показаны результаты экспериментов, включающие инкубацию ингибитора PKR в комбинации или с PIF, или с ангиотензином II. На фиг.5 можно видеть, что поскольку PIF увеличивает деградацию белков вплоть до 87%, при инкубации с одним PIF, то добавление ингибитора PKR обращает уровни деградации белка обратно примерно до уровней контроля. Подобным образом, на фиг.6 можно видеть, что поскольку PIF снижает синтез белка примерно до 25%, при инкубации с одним PIF, то добавление ингибитора PKR обращает уровни синтеза белка обратно примерно до уровней контроля.
На фиг.7 и 8 показаны похожие результаты, полученные в результате инкубации ингибитора PKR с ангиотензином II. На фиг.7 ангиотензин увеличивал деградацию белков до примерно 51%, по сравнению с контролем. Добавление ингибитора PKR обращало эту тенденцию, поддерживая уровни деградации белков примерно на уровне контроля. Подобным образом, на фиг.8 ангиотензин II снижает синтеза белка примерно на 40% по сравнению с контролем, так как добавление ингибитора PKR поддерживает уровни синтез белка примерно на уровне контроля.
Ингибитор PKR ослабляет действие PIF и ангиотензина II как на деградацию белка, так и на синтез белка. Этот факт позволяет предположить, что как PIF, так и ангиотензин II, опосредуют свои эффекты через похожие механизмы и через общий медиатор, вероятно, включающий PKR. В особенности, эти результаты позволяют предположить, что PIF активирует PKR, которая, в свою очередь, вызывает фосфорилирование eIF2-α, ингибирующее связывание инициатора метионил-tRNA (met-tRNA) с рибосомальной субъединицей 40s. BCAAs, инсулин и IGF-1 ослабляли фосфорилирование eIF2-α, вызванное PIF, что служит дополнительным подтверждением гипотезы, согласно которой для ингибирования синтеза белка PIF осуществляет позитивную регуляцию фосфорилирования eIF2-α. Поскольку PKR может ингибировать синтез белка и активировать фактор NF-kВ, который приводит к деградации белка, вероятно, PKR представляет собой ранний компонент в пути передачи сигнала PIF.
Существует также наблюдение, что PKR вовлечена в регуляцию фосфорилирования 4Е-ВР1. Таким образом, если фактор PIF действительно передает сигнал через PKR, вероятно, что он может также снижать синтез белка через PKR-опосредуемую активацию серин/треонинфосфатазы РР2А, которая может осуществлять дефосфорилирование 4Е-ВР1, который, в свою очередь, переводит elF4E в неактивный комплекс, предотвращающий образование комплекса преинициации 43s.
На фиг.9 показан альтернативный механизм. Оба фактора, индуцирующий протеолиз (PIF) и ангиотензин II (Ang II), снижают синтез белка на 40%, и концентрации обоих агентов, которые максимально эффективны в подавлении синтеза белка были одинаковыми, как те, что были максимально эффективны в индукции деградации белков. Результаты позволяют предположить, что как инсулин, так и IGF1, по меньшей мере, частично, ослабляют деградацию белков, индуцированную фактором PIF, через ингибирование PKR и/или фосфорилирование eIF2α. Механизм активации факторами PIF и Ang II может происходить через PACT (белок-активатор интерферон-индуцированной протеинкиназы), клеточный белок-активатор PKR, хотя PIF также представляет собой полианионную молекулу, и, таким образом, может активировать напрямую. Независимо, кажется, что фосфорилирование eIF2α факторами PIF и Ang II происходит через PKR, так как ингибитор PKR ослабляет ингибиторный эффект обоих факторов на синтез белка. Вероятно, что эффект как PIF, так и Ang II на трансляцию белка возникает в результате увеличенного фосфорилирования eIF2α.
Ингибирование синтеза белка в апоптозе фактором некроза опухолей-α (TNF-α) также связано с усиленным фосфорилированием eIF2α. Дополнительную поддержку роли фосфорилирования eIF2α в ингибировании синтеза белка фактором PIF и Ang II обеспечивается таким наблюдением, что инсулин и IGF1, которые были эффективны в подавлении ингибирования синтеза белка, полностью ослабляют индукцию фосфорилирования eIF2α. Собранные данные позволяют предположить, что BCAAs также действуют через похожий механизм ингибирования пути деградации, который инициируется фактором PIF. Это исследование предоставило первое наблюдение взаимосвязи между подавлением синтеза белка в скелетной мышце фактором PIF (и Ang II), через активацию PKR, и фосфорилирование eIF2α, и усиленной деградацией миофибриллярного белка миозина, через активацию фактора NF-κB, приводящей к увеличению экспрессии и активности убиквитин-протеосомного протеолитического пути. Это означает, что факторы, которые нацелены на PKR (например, BCAAs), могут быть эффективны в лечении мышечной атрофии в кахексии при раке.
На фиг.10 дополнительно показан альтернативный механизм. Как отмечалось ранее, оба фактора, индуцирующий протеолиз (PIF) и ангиотензин II (Ang II), усиливают деградацию белков через фосфорилирование PKR и/или eIF2α. NF-κВ может быть активирован фактором PIF или находящимся ниже медиатором фактора PIF (PKR и/или eIF2α), который появляется посредством освобождения фактора NF-κВ. В этом дополнительном альтернативным механизмом, NF-κВ является частью того же каскада фосфорилирования, несмотря на существование той же мишени для усиления деградации помеченных убиквитином белков.
В совокупности, данные, приведенные выше, подтверждают ряд новых аспектов настоящего изобретения. Во-первых, BCAAs могут быть применены для лечения потери мышечной массы у индивидуума путем противодействия катаболизму белков, опосредуемому фактором PIF и/или ангиотензином II через ингибирование активации PKR и/или eIF2α. Во-вторых, каждая из BCAAs в одинаковой степени эффективна в таком противодействии. В-третьих, совместное введение инсулина, IGF-1 и/или ингибитора PKR, или применение лечений для увеличения уровня или инсулина, или IGF-1, или их обоих, может увеличивать эффективность лечения с помощью ВСАА путем дополнительного противодействия катаболизму белка, усиления синтеза белка, или того и другого.
Пищевые продукты в соответствии с изобретением могут, следовательно, включать BCAAs, одни или в комбинации с инсулином, IGF-1 и/или ингибитором PKR. BCAAs могут быть введены или в свободной форме, или как дипептиды, как трипептиды, как полипептиды, как белок, богатый аминокислотой ВСАА, и/или как белок, искусственно обогащенный по содержанию ВСАА. Дипептиды, трипептиды и полипептиды могут включать две или несколько BCAAs. Если в дипептид, трипептид или полипептид включают аминокислоты, не представляющие собой BCAAs, то предпочтительные аминокислоты включают аланин и глицин, но аминокислоты, не представляющие собой BCAAs, могут представлять собой любую из обязательных или необязательных (заменимых или незаменимых) аминокислот. Например, предпочтительные дипептиды включают, но не ограничиваются, аланил-лейцин, аланил-изолейцин, аланил-валин, глицил-лейцин, глицил-изолейцин и глицил-валин.
Пищевые продукты в соответствии с изобретением могут, подобным образом, включать предшественников и/или метаболитов BCAAs, в особенности, предшественников и/или метаболитов лейцина, в дополнении к BCAAs или вместо BCAAs. Такие продукты могут дополнительно включать любое число дополнительных ингредиентов, включающих, например, белок, волокно, жирную кислоту, витамин, минерал, сахар, углевод, вкусовую добавку, лекарственные препараты и терапевтические средства.
Пищевые продукты настоящего изобретения могут быть введены перорально, посредством зонда для искусственного кормления или парентерально. Такие продукты могут быть применены в лечении человека, страдающего от ряда заболеваний, нарушений и состояний, связанных с истощением мышечной массы, или с любым заболеванием, нарушением или состоянием, с которым связана потеря мышечной массы, включающим, например, кахексию, рак, индуцированную опухолью потерю массы, сепсис, хроническую сердечную недостаточность, ревматоидный артрит, синдром приобретенного иммунодефицита (AIDS), саркопению, диабет, гипертонию, высокие уровни сывороточного холестерина, высокие уровни триглицеридов, болезнь Паркинсона, бессонницу, наркозависимость, хронический алкоголизм, болевые ощущения, бессонницу, гипогликемию, нарушение функции печени, включающее цирроз печени, нарушение функции желчного пузыря, хорею, дискинезию и нарушение функции почек, включающее уремию.
Приведенное выше описание различных аспектов изобретения было представлено с целью иллюстрации и описания. Оно не предназначалось для того, чтобы быть исчерпывающим или для ограничения изобретения точными формами раскрытия, и понятно, что возможны многочисленные модификации и изменения. Такие модификации и изменения, вероятно, очевидные специалисту в этой области техники, предназначены для включения в объем изобретения, как определено в прилагаемой формуле изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОВЫШЕНИЯ ЭКСПРЕССИИ РНК В КЛЕТКЕ | 2018 |
|
RU2784654C2 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ АМИНОКИСЛОТЫ ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ ИНСУЛЬТА У ПАЦИЕНТОВ С ДИСФАГИЕЙ | 2015 |
|
RU2668373C1 |
| СПОСОБЫ БИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ С ИСПОЛЬЗОВАНИЕМ КЛЕТОК ДЛЯ КОНТРОЛЯ КАЧЕСТВА НУТРИЦЕВТИЧЕСКИХ ПРОДУКТОВ И ЛЕКАРСТВЕННЫХ СРЕДСТВ | 2013 |
|
RU2662358C2 |
| ИСПОЛЬЗОВАНИЕ МЕТАБОЛИТОВ ТРИПТОФАНА ДЛЯ ЛЕЧЕНИЯ МЫШЕЧНОЙ АТРОФИИ | 2017 |
|
RU2806346C2 |
| ФЕНИЛ-2-ГИДРОКСИ-АЦЕТИЛАМИНО-2-МЕТИЛ-ФЕНИЛОВЫЕ СОЕДИНЕНИЯ | 2018 |
|
RU2804280C2 |
| ПРОЛЕКАРСТВЕННЫЕ МОДУЛЯТОРЫ ИНТЕГРИРОВАННОГО ПУТИ СТРЕССА | 2019 |
|
RU2824500C2 |
| СПОСОБ УСКОРЕНИЯ ВОССТАНОВЛЕНИЯ СКЕЛЕТНЫХ МЫШЦ ОТ АТРОФИИ ПОСЛЕ ДЛИТЕЛЬНОЙ АЛКОГОЛЬНОЙ ИНТОКСИКАЦИИ | 2014 |
|
RU2578463C1 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ МЫШЕЧНОЙ АТРОФИИ | 2020 |
|
RU2814282C2 |
| ПРИМЕНЕНИЕ (R)-АРИЛПРОПИОНОВЫХ КИСЛОТ ДЛЯ ПОЛУЧЕНИЯ ЛЕКАРСТВЕННЫХ СРЕДСТВ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ЛЮДЕЙ И ЖИВОТНЫХ, НА КОТОРЫЕ МОЖНО ОКАЗЫВАТЬ ТЕРАПЕВТИЧЕСКОЕ ВОЗДЕЙСТВИЕ ПУТЕМ ИНГИБИРОВАНИЯ АКТИВАЦИИ NF-κВ | 2000 |
|
RU2250103C2 |
| Новая композиция, содержащая аминокислоты с разветвленными боковыми цепями | 2020 |
|
RU2817963C2 |
Изобретение относится к медицине, а именно к терапии, и касается лечения потери мышечной массы. Для этого вводят пищевой продукт, включающий аминокислоту с разветвленной цепью (ВСАА), предшественник ВСАА или метаболит ВСАА, в сочетании с ингибитором РНК-зависимых протеинкиназ (PKR) в эффективном количестве. Это обеспечивает эффективное лечение потери мышечной массы при различных заболеваниях за счет способности ВСАА и PKR воздействовать на катаболизм белка по различным механизмам, и тем самым дополнять и усиливать способность друг друга противодействовать катаболизму белка. 16 з.п. ф-лы, 10 ил.
1. Пищевой продукт, включающий эффективное количество, по меньшей мере, одного из соединений: аминокислоту с разветвленной цепью (ВСАА), предшественника ВСАА или метаболита ВСАА, и эффективное количество ингибитора РНК-зависимых протеинкиназ (PKR), используемый в качестве антагониста катаболизма белков при лечении потери мышечной массы у индивидуума.
2. Пищевой продукт по п.1, в котором одну или несколько аминокислот ВСАА выбирают из группы, состоящей из лейцина, изолейцина и валина.
3. Пищевой продукт по п.1, в котором ВСАА находится в виде, по меньшей мере, одного из следующего: дипептида, трипептида или полипептида.
4. Пищевой продукт по п.3, в котором дипептид содержит две аминокислоты ВСАА.
5. Пищевой продукт по п.3, в котором трипептид или полипептид содержит две или несколько аминокислот ВСАА.
6. Пищевой продукт по п.3, в котором дипептид выбирается и аланин-лейцина, аланин-изолейцина, аналин-валина, глицил-лейцина, глицил-изолейцина и глицил-валина.
7. Пищевой продукт по п.1, в котором предшественник ВСАА представляет собой пируват.
8. Пищевой продукт по п.1, в котором метаболит ВСАА выбирается из β-гидрокси- β-метилбутирата и α-кетоизокапроата.
9. Пищевой продукт по п.1, дополнительно включающий, по меньшей мере, одно соединение из инсулина и инсулиноподобного фактора роста 1 (IGF-1).
10. Пищевой продукт по п.1, в котором ингибитор PKR имеет следующую структуру:
11. Пищевой продукт по п.1, дополнительно включающий, по меньшей мере, одно соединение из: белок, волокно, жирную кислоту, витамин, минерал, сахар, углевод, вкусовую добавку, лекарственный препарат и лечебное средство.
12. Пищевой продукт по п.1, в котором, по меньшей мере, одно соединение, выбираемое из аминокислоты ВСАА, предшественника ВСАА или метаболита ВСАА, находится в виде принимаемого внутрь пищевого продукта.
13. Пищевой продукт по п.12, дополнительно включающий, по меньшей мере, одно из следующего: белок, волокно, жирную кислоту, витамин, минерал, сахар, углевод, вкусовую добавку, лекарственный препарат и лечебное средство.
14. Пищевой продукт по п.1, в котором, по меньшей мере, одно соединение из аминокислоты ВСАА, предшественника ВСАА или метаболита ВСАА вводится через зонд для искусственного кормления.
15. Пищевой продукт по п.1, в котором, по меньшей мере, одно соединение из аминокислоты ВСАА, предшественника ВСАА, метаболита ВСАА и дипептида или трипептида, содержащих, по меньшей мере, одну аминокислоту ВСАА, вводится парентерально.
16. Пищевой продукт по п.1, предназначенный для индивидуума, имеющего, по меньшей мере, одно из следующих заболеваний: кахексию, рак, индуцированную раком потерю массы, сепсис, хроническую сердечную недостаточность, ревматоидный артрит, синдром приобретенного иммунодефицита (AIDS), саркопению, диабет, гипертонию, высокий уровень сывороточного холестерина, высокий уровень триглицеридов, болезнь Паркинсона, бессонницу, наркозависимость, алкоголизм, боли, гипогликемию, нарушение функции печени, в том числе цирроз, заболевание желчного пузыря, хорею, дискинезию, почечное заболевание, в том числе уремию.
17. Пищевой продукт по п.1, дополнительно предназначенный для усиления синтеза белка при лечении потери мышечной массы у индивидуума.
| US 4780475 А, 25.10.1988 | |||
| 0 |
|
SU156402A1 | |
| КАРЛОВ В.А | |||
| Неврология | |||
| - М., 2002, МИА, с.518-530 | |||
| SMITH J et al | |||
| "Mechanism of the attenuation of proteolysis-inducing factor stimulated protein degradation in muscle by β-hydroxy-β-methylbutyrate" | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Нефтяной конвертер | 1922 |
|
SU64A1 |

