Область техники, к которой относится изобретение
Настоящее изобретение относится к микробиологической промышленности и, в частности, к способу получения полезных метаболитов ферментацией бактерии семейства Enterobacteriaceae, отличающийся тем, что генетическая экспрессионная система бактерии, регулируемая белком типа LysR, модифицирована таким способом, что функциональность указанной экспрессионной системы опосредована коиндуктором, и, как результат, повышен уровень экспрессии гена, регулируемого указанной системой экспрессии. Более точно, экспрессионная система и способ могут быть полезны для повышения продукции метаболитов биосинтетического пути L-аминокислот, таких как L-аминокислот с разветвленной цепью (разветвленных L-аминокислот).
Уровень техники
Традиционно L-аминокислоты в промышленности получают методом ферментации с использованием штаммов микроорганизмов, полученных из природных источников, или их мутантов, специально модифицированных для того, чтобы увеличить продукцию L-аминокислот.
Описано много способов повышения продукции L-аминокислот, включающие, например, трансформацию микроорганизма рекомбинантной ДНК (см., например, Патент США №4,278,765) и изменение регуляторных участков, таких как промотор, лидерная последовательность и/или аттенюаторы или другие, известные для специалиста данной области (см., например, Патентную заявку США №20060216796 A1 и WO 9615246 A1). Другие способы, повышающие выход, включают повышение активности ферментов, вовлеченных в биосинтез аминокислот, и/или снятие ингибирования ключевых ферментов продуцируемой L-аминокислотой по типу обратной связи (см., например, публикацию Патента Японии №56-18596 (1981), WO 95/16042 или Патенты США №5,661,012 и 6,040,160). Например, мутантная бактериальная ацетогидроксикислотасинтетаза I (также называемая как ацетолактатсинтаза I, в дальнейшем AHAS I), которая обладает устойчивостью к ингибированию по типу обратной связи L-валином, использовалась для увеличения выхода разветвленной L-аминокислоты в соответствующих штаммах, продуцирующих L-аминокислоту (Патент РФ №2355763).
Биосинтез разветвленных L-аминокислот (branched-chain L-amino acids, BCAAs), таких как L-валин, L-лейцин и L-изолейцин осуществляется в разветвленном биосинтетическом пути. Ацетолактатсинтаза (классификационный номер фермента (ЕС) 2.2.1.6) катализирует реакцию на первом этапе биосинтетического пути, который является общим для всех трех аминокислот. Реакция включает конденсацию активированного ацетальдегида (2-(α-гидроксиэтил)тиаминдифосфата), производного пирувата, с пируватом или 2-оксобутаноатом с образованием 2-ацетолактата (AΛ) или 2-ацето-2-гидроксибутаноата (АГБ), соответственно. AΛ является предшественником L-валина и L-лейцина, а АГБ - L-изолейцина. В Escherichia coli (далее E.coli), например, реакции между молекулами пирувата, а также между пируватом и 2-оксобутаноатом катализируются изоферментами AHAS, такими как AHAS I, AHAS II и AHAS III, кодируемыми генами ilvBN, ilvGM и ilvIH, соответственно. AHAS I и AHAS III ингибируются конечным продуктом (механизм называется как ингибирование по типу обратной связи или ретройнгибирование) - L-валином. Ингибирование по типу обратной связи целевым продуктом играет важную роль в физиологическом контроле биосинтетических путей в бактериях.
Продукты реакции, катализируемой AHAS, AΛ или АГБ, являются субстратами для 2-ацетогидроксикислотаизомероредуктазы IlvC (EC 1.1.1.86), кодируемой геном ilvC, принадлежащим к оперону ilvYC. Оперон ilvYC из E.coli является составной частью экспрессионной системы, регулируемой белком типа LysR, которая представляет собой наиболее широко распространенную положительно регулируемую экспрессионную систему бактерий и может быть найдена в бактериальных семействах прокариот от Enterobacteriaceae до Rhizobiaceae (Rhee K.Y. et al., Proc. Nat. Acad. Sci. USA, 1999, 96:14294-14299). Ген ilvY кодирует регуляторный белок IlvY типа LysR (транскрипционный регулятор), который связывается высококооперативным способом с двумя тандемными операторными участками в области ilvYC с разнонаправлено перекрывающимися промоторами (Фигура 1). При связывании с первым операторным участком регулятор IlvY отрицательно саморегулирует транскрипцию с промотора ilvY, ослабляя, таким образом, собственный синтез. Кроме этого, IlvY играет основную роль в активации транскрипции гена ilvC. Активация транскрипции ilvC требует связывания регулятора IlvY со вторым операторным участком и последующее связывание коиндуктора, такого как 2-ацетолактат (AΛ) или 2-ацето-2-гидроксибутаноат (АГБ) с ранее образованным комплексом IlvY/ДНК. При связывании коиндуктора происходит конформационное изменение в комплексе белок/ДНК, что вызывает изменение структуры промотора ilvC в области -35 и значительно повышает силу связывания PHK-полимеразы (Rhee K.Y. et al., J. Biol. Chem., 1998, 273:11257-11266).
В биосинтезе L-валина и L-лейцина 2-ацетолактат (AΛ) превращается под действием белка IlvC в 2,3-дигидрокси-3-метилбутаноат (другое название 2,3-дигидроксиизовалериат, ДГИВ) (Фигура 2). В биосинтезе L-изолейцина 2-ацето-2-гидроксибутаноат (АГБ) превращается под действием белка IlvC в 2,3-дигидрокси-3-метилпентаноат (другое название 2,3-дигидрокси-3-метилвалериат, ДГМВ).
Недавно самоиндуцируемые генетические системы экспрессии стали очень привлекательны для усиления экспрессии желаемого гена по сравнению с обычными генетическими подходами. Например, из литературы известна искусственно созданная система экспрессии генов, регулируемая по типу положительной обратной связи, которая может функционировать как усилитель генетического сигнала, что способствует повышению чувствительности к белку-индуктору, а также увеличению уровня экспрессии в отсутствии внешнего кофактора - ацилгомосеринлактона (АГΛ, также называемого как гомосеринлактон - ГСΛ) (Nistala G.J. et al., J. Biol. Eng., 2010, 4:4). Разработанная система использует постоянно активный вариант белка LuxR [lux оперон), регулятора кворум-сенсинга (КС) из Vibrio fischeri, который независим от самоиндуктора (АГΛ) ввиду точечной мутации Ala221Val (Sayut D.J. et al., Biochem. Biophys. Res. Commun., 2007, 363:667-673; Poellinger K.A. et al., FEMS Microbiol Lett., 1995, 129:97-101). Генетическая система экспрессии, похожая за исключением небольших изменений на приведенную, была использована для изменения кинетических параметров экспрессии модельного мембранного белка цитохром bd хинолоксидазы в E.coli (Bansal К. et al., J. Biol. Eng., 2010, 4:6).
Самоиндуцируемая регулируемая по типу положительной обратной связи система активации, основанная на аппарате кворум-сенсинга из V. fischery (lux-биолюминесцентные гены) и использующая внутриклеточный пул самоиндуктора (ГСΛ), применялась для экспрессии рекомбинантных белков, таких как антигены для приготовления фармацевтических композиций (WO 2010136897 A2).
Однако, в настоящее время нет данных, описывающих генетическую экспрессионную систему, регулируемую белком типа LysR, модифицированную таким образом, что функциональность указанной экспрессионной системы опосредована коиндуктором; и применение данной системы для получения полезных метаболитов пути биосинтеза L-аминокислот, таких как разветвленные L-аминокислоты, и/или его побочного пути.
Раскрытие сущности изобретения
Цель настоящего изобретения - предоставление генетической экспрессионной системы, включающей элементы транскрипционного аппарата, регулируемого белком типа LysR, модифицированной таким образом, что регуляция указанной системы, функционирующей по принципу самоиндукции с положительной обратной связью, опосредована коиндуктором.
Другая цель настоящего исследования - предоставление бактерии семейства Enterobacteriaceae, которая может принадлежать к роду Escherichia и, более точно, к виду Escherichia coli, которая модифицирована так, чтобы содержать указанную экспрессионную систему.
Другая цель настоящего изобретения - предоставление способа получения полезных метаболитов, например, L-аминокислот, таких как L-аминоки слоты с разветвленной цепью, или их солей, в частности, L-валина, L-лейцина и L-изолейцина или их солей. Эта цель была достигнута благодаря обнаружению того факта, что модификация генов, колирующих мутантную ацетолактатсинтазу, устойчивую к ингибированию по типу обратной связи L-валином, таким образом, что экспрессия этих генов регулируется транскрипционным аппаратом, регулируемым белком типа LysR и экспрессия которого опосредована коиндуктором, получаемым реакцией с участием ацетолактатсинтазы, приводит к увеличению продукции разветвленных L-аминокислот.
Цель настоящего изобретения - предоставление генетической экспрессионной системы, включающей транскрипционный аппарат, регулируемый белком типа LysR, который включает промотор и оператор, экспрессия которых положительно регулируется регуляторным белком типа LysR и коиндуктором, и целевой(ые) ген(ы), с которым(и) транскрипционный аппарат функционально связан, причем целевой (ые) ген(ы) кодирует(ют) белок(и), вовлеченный (е) в биосинтез коиндуктора, субстрата или предшественника коиндуктора, и самоиндуцируемая положительная регуляция по типу обратной связи такой экспрессионной системы опосредована коиндуктором.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что указанная система из бактерии, принадлежащей к семейству Enterobacteriaceae или Pseudomonadaceae.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что указанная система из бактерии, принадлежащей к семейству Enterobacteriaceae.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что указанная система из бактерии, принадлежащей к роду Escherichia.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что бактерия принадлежит к виду Escherichia coli.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что указанная система из биосинтетического пути L-аминокислоты, выбранной из группы, состоящей из разветвленных L-аминокислот, L-лизина, L-цистеина, L-метионина и L-триптофана.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что указанная система из биосинтетического пути L-аминокислот с разветвленной цепью.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что промотор есть промотор PilvC, регуляторный белок типа LysR есть белок IlvY, и коиндуктор есть 2-ацетомолочная кислота, 2-ацето-2-гидроксимаслянная кислота или их соли.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что коиндуктор есть 2-ацетомолочная кислота или ее соль.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что целевой(ые) ген(ы) кодирует(ют) ацетогидроксикислотасинтетазу.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что указанные целевые гены кодируют белки, выбранные из группы, состоящей из:
(A) комбинации из п.п.A1 и A2:
(A1) белка, содержащего аминокислотную последовательность SEQ ID NO: 2, или белка, содержащего аминокислотную последовательность SEQ ID NO: 2, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком A2; и
(A2) белка, содержащего аминокислотную последовательность SEQ ID NO:4, или белка, содержащего аминокислотную последовательность SEQ ID NO: 4, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком A1;
(B) комбинации из п.п.B1 и B2:
(B1) белка, содержащего аминокислотную последовательность SEQ ID NO: 6, или белка, содержащего аминокислотную последовательность SEQ ID NO: 6, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком B2; и
(B2) белка, содержащего аминокислотную последовательность SEQ ID NO: 8, или белка, содержащего аминокислотную последовательность SEQ ID NO: 8, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком B1; и
(C) комбинации из п.п.C1 и C2:
(C1) белка, содержащего аминокислотную последовательность SEQ ID NO: 32, или белка, содержащего аминокислотную последовательность SEQ ID NO: 32, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком C2; и
(C2) белка, включающего аминокислотную последовательность SEQ ID NO: 34, или белка, включающего аминокислотную последовательность SEQ ID NO: 34, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком C1.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что ацетогидроксикислотасинтетаза является мутантной ацетолактатсинтазой I, устойчивой к ингибированию по типу обратной связи L-валином.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что оператор включает участок, с которым связывается вышеописанный регуляторный белок типа LysR.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что регуляторный белок типа LysR выбран из группы, состоящей из:
(D) белка, содержащего аминокислотную последовательность SEQ ID NO: 10; и
(E) варианта белка, содержащего аминокислотную последовательность SEQ ID NO: 10, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и имеющего активность регуляторного белка типа LysR.
Также цель настоящего изобретения - предоставление вышеописанной экспрессионной системы, отличающейся тем, что промотор включает:
(F) ДНК, содержащую нуклеотидную последовательность SEQ ID NO: 30; или
(G) вариант ДНК, содержащий нуклеотидную последовательность SEQ ID NO: 30, которая содержит замену, делецию, вставку или добавление одного или нескольких нуклеотидных остатков, и имеющий активность нуклеотидной последовательности SEQ ID NO: 30.
Также цель настоящего изобретения - предоставление бактерии-продуцента L-аминокислоты, принадлежащей к семейству Enterobacteriaceae, отличающейся тем, что бактерия модифицирована таким образом, что она содержит экспрессионную систему как описано выше.
Также цель настоящего изобретения - предоставление вышеописанной бактерии, отличающейся тем, что бактерия содержит ген, кодирующий регуляторный белок типа LysR.
Также цель настоящего изобретения - предоставление вышеописанной бактерии, отличающейся тем, что бактерия принадлежит к роду Escherichia.
Также цель настоящего изобретения - предоставление вышеописанной бактерии, отличающейся тем, что бактерия принадлежит к виду Escherichia coli.
Также цель настоящего изобретения - предоставление вышеописанной бактерии, отличающейся тем, что L-аминокислота есть L-аминокислота с разветвленной цепью.
Также цель настоящего изобретения - предоставление вышеописанной бактерии, отличающейся тем, что L-аминокислота с разветвленной цепью выбрана из группы, состоящей из L-валина, L-лейцина и L-изолейцина.
Цель настоящего изобретения - предоставление способа получения L-аминокислоты с разветвленной цепью, включающего:
(i) выращивание вышеописанной бактерии в питательной среде, так что указанная L-аминокислота с разветвленной цепью накапливается в культуральной жидкости; и
(ii) выделение L-аминокислоты с разветвленной цепью из культуральной жидкости.
Также цель настоящего изобретения - предоставление способа получения вышеописанной L-аминокислоты с разветвленной цепью, отличающийся тем, что указанная L-аминоки слота с разветвленной цепью выбрана из группы, состоящей из L-валина, L-лейцина и L-изолейцина.
Краткое описание фигур
Фигура 1 показывает схему регуляции транскрипции генов ilvY и ilvC. IlvY означает транскрипционный регулятор типа LysR, AΛ или АГБ - коиндуктор 2-ацетолактат или 2-ацето-2-гидроксибутаноат, O1 и O2 - операторы 1 и 2, TSilvC - старт транскрипции гена ilvC, TSilvY - старт транскрипции гена ilvY, знаки минус (-) и плюс (+) - отрицательное и положительное влияние на транскрипцию гена, соответственно.
Фигура 2 показывает схему биосинтеза L-валина из пирувата. AΛ означает 2-ацетолактат, ДГИВ - 2,3-дигидроксиизовалериат, AHAS -ацетолактатсинтазу, IlvC - изомероредуктазу
Описание последовательностей
SEQ ID NO: I - нуклеотидная последовательность гена ilvB.
SEQ ID NO: 2 - аминокислотная последовательность большой субъединицы ацетолактатсинтазы I дикого типа.
SEQ ID NO; 3 - нуклеотидная последовательность гена ilvN.
SEQ ID NO: 4 - аминокислотная последовательность малой субъединицы ацетолактатсинтазы I дикого типа.
SEQ ID NO: 5 - нуклеотидная последовательность гена ilvI.
SEQ ID NO: 6 - аминокислотная последовательность большой субъединицы ацетолактатсинтазы III дикого типа.
SEQ ID NO: 7 - нуклеотидная последовательность гена ilvH.
SEQ ID NO: 8 - аминокислотная последовательность малой субъединицы ацетолактатсинтазы III дикого типа.
SEQ ID NO: 9 - нуклеотидная последовательность гена ilvY.
SEQ ID NO: 10 - аминокислотная последовательность белка IlvY.
SEQ ID NOs: 11-29 - нуклеотидные последовательности праймеров P1-P19.
SEQ ID NO: 30 - нуклеотидная последовательность промотора PilvC.
SEQ ID NO: 31 - нуклеотидная последовательность гена ilvG.
SEQ ID NO: 32 - аминокислотная последовательность большой субъединицы ацетолактатсинтазы II дикого типа.
SEQ ID NO: 33 - нуклеотидная последовательность гена ilvM.
SEQ ID NO: 34 - аминокислотная последовательность малой субъединицы ацетолактатсинтазы II дикого типа.
Наилучший способ осуществления изобретения
1. Бактерия
Используемый в настоящем изобретении термин «полезный метаболит» не ограничен каким-либо образом при условии, что этот метаболит может быть получен ферментативной реакцией или биосинтетическим путем и может включать L-аминоки слоты, высшие спирты и D-пантотеновую кислоту.
Термин «бактерия-продуцент L-аминокислоты» может означать бактерию семейства Enterobacteriaceae, которая способна продуцировать и вызывать накопление L-аминокислоты в культуральной жидкости, когда указанная бактерия выращивается (культивируется) в питательной среде. Способность бактерии продуцировать L-аминокислоту может означать способность бактерии продуцировать L-аминокислоту в питательной среде или в бактериальных клетках, что приводит к накоплению L-аминокислоты в количествах, достаточных для ее выделения из культуральной жидкости или из бактериальных клеток, когда указанная бактерия выращивается (культивируется) в питательной среде.
Термин «L-аминокислота» может означать L-аланин, L-аргинин, L-аспарагин, L-аспарагиновую кислоту, L-цистеин L-глутаминовую кислоту, L-глутамин, глицин L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин.
Термин «разветвленная L-аминокислота» может включать L-валин, L-лейцин и L-изолейцин.
Бактерия может обладать способностью к продукции полезного метаболита, такого как L-аминокислота, изначально, в соответствии со своими природными характеристиками, или может быть модифицирована с помощью мутаций (мутагенеза) или технологий рекомбинантных ДНК так, чтобы она получила способность к продукции.
Бактерия, принадлежащая к семейству Enterobacteriaceae, может быть выбрана из бактерий, относящихся к родам, входящим в состав этого семейства, таких как Enterobacter, Erwinia, Escherichia, Klebsiella, Morganella, Pantoea, Photorhabdus, Providencia, Salmonella, Yersinia и т.д., и способных продуцировать L-аминокислоту. Более конкретно, могут быть использованы бактерии, классифицируемые как принадлежащие к семейству Enterobacteriaceae в соответствии с таксономией, используемой в базе данных NCBI (National Center for Biotechnology Information) (http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=543). Примеры бактерий, которые могут быть модифицированы согласно настоящему изобретению, включают бактерии, относящиеся к роду Escherichia, Enterobacter или Pantoea.
Выбор штаммов бактерий, принадлежащих к роду Escherichia, которые могут быть модифицированы в настоящем изобретении, не ограничен каким-либо образом, однако, в качестве примеров бактерии рода Escherichia, описанные в книге Neidhardt et al. (Bachmann, B.J., Derivations and genotypes of some mutant derivatives of E.coli K-12, p.2460-2488. In F.C. Neidhardt et al. (ed.), E.coli and Salmonella: cellular and molecular biology, 2nd ed. ASM Press, Washington, D.C., 1996), могут быть включены в число бактерий согласно настоящему изобретению. В качестве конкретного примера могут быть взяты штаммы E.coli., такие как E.coli W3110 (ATTC 27325), E.coli MG1655 (ATCC 47076) и т.д., которые происходят из исходного штамма дикого типа, т.е. штамма E.coli K-12. Этот штамм может быть получен, в частности, из Американской коллекции типовых культур «American Type Culture Collection, (ATCC)» (P.O. Box 1549, Manassas, VA 20108, United States of America). Каждому штамму присвоен индивидуальный регистрационный номер, и штаммы могут быть заказаны согласно регистрационному номеру (см. ссылку www.atec.org). Регистрационные номера штаммов находятся в списке каталога Американской коллекции типовых культур «American Type Culture Collection, ATCC».
Примеры бактерий Enterobacter включают Enterobacter agglomerans, Enterobacter aerogenes и т.д. Примеры бактерии Pantoea включают Pantoea ananatis и т.д. Недавно некоторые виды Enterobacter agglomerans были реклассифицированы как Pantoea agglomerans, Pantoea ananatis или Pantoea stewartii на основе анализа нуклеотидной последовательности 168 рРНК и других доказательств. Бактерии, относящиеся к роду Enterobacter или Pantoea, могут быть использованы в соответствии с настоящим изобретением, так как принадлежат к семейству Enterobacteriaceae. Полученные с использованием технологий генной инженерии штаммы Pantoea ananatis, такие как штамм Pantoea ananatis AJ 3355 (PERM BP-6614), штамм AJ 3356 (FERM BP-6615), штамм AJ 3601 (PERM BP-7207) и их производные могут быть использованы в соответствии с настоящим изобретением. Эти штаммы были классифицированы как Enterobacter agglomerans при выделении, и они депонированы как Enterobacter agglomerans. Однако позднее они были реклассифицированы как Pantoea ananatis на основе анализа нуклеотидной последовательности 16S pPHK и других доказательств.
Примеры бактерий, принадлежащих к семейству Pseudomonadaceae, могут происходить из рода Pseudomonas. Примеры бактерий рода Pseudomonas включают Pseudomonas putida, P. aeruginosa, and P. syringae.
Термин «бактерия, которая модифицирована так, чтобы содержать генетическую экспрессионную систему» может означать бактерию, например, бактерию-продуцент L-аминокислоты, принадлежащую к семейству Enterobacteriaceae, в которой генетическая экспрессионная система, регулируемая белком типа LysR, модифицирована таким образом, что функциональность указанной самоиндуцируемой системы с положительной обратной регуляцией опосредована коиндуктором. Например, термин «бактерия, которая модифицирована так, чтобы содержать генетическую экспрессионную систему», может означать бактерию семейства Enterobacteriaceae, обладающую способностью к продукции L-аминокислоты, такой как разветвленная L-аминокислота, которая модифицирована таким образом, что она имеет один или несколько генов указанной системы экспрессии.
Бактерия-продуцент L-аминокислоты с разветвленной цепью Термин «бактерия-продуцент L-аминокислоты с разветвленной цепью» может означать бактерию, которая способна продуцировать и вызывать накопление разветвленной L-аминокислоты в культуральной жидкости, такую как L-валин, L-лейцин и L-изолейцин, когда указанная бактерия выращивается (культивируется) в питательной среде.
Разветвленные L-аминокислоты помимо L-валина, L-лейцина и L-изолейцина также могут включать неприродные разветвленные L-аминокислоты, такие как L-гомолейцин и L-гомоизолейцин. Способность бактерии к продукции разветвленной L-аминокислоты может быть сообщена бактерии или повышена путем отбора бактерии при выращивании (breeding). Термин «бактерия-продуцент L-аминокислоты с разветвленной цепью» также может означать бактерию, которая способна продуцировать и вызывать накопление разветвленной L-аминоки слоты в культуральной жидкости в большем количестве, чем немодифицированный штамм, например, штамм дикого типа или родительский штамм, и также может означать бактерию, которая способна продуцировать и вызывать накопление разветвленной L-аминокислоты в культуральной жидкости в количестве, не менее чем 0,5 г/л или даже не менее чем 1,0 г/л.
Бактерия-продуцент L-аминокислоты с разветвленной цепью может быть бактерией семейства Enterobacteriaceae, которая содержит регуляторный участок, положительно влияющий на экспрессию генов ацетолактатсинтазы комплексом, который включает транскрипционный регуляторный белок и коиндуктор, являющийся продуктом реакции, катализируемой ацетолактатсинтазой. Более того, бактерия может быть бактерией семейства Enterobacteriaceae, в которой активность мутантной ацетолактатсинтазы повышена. Более точно, бактерия может быть бактерией-продуцентом разветвленной L-аминокислоты семейства Enterobacteriaceae, отличающейся тем, что продукция разветвленной L-аминокислоты повышена за счет увеличенной активности мутантной ацетолактатсинтазы введением регуляторного участка в бактерию. Бактерия является бактерией-продуцентом разветвленной L-аминокислоты, принадлежащей к роду Escherichia, отличающаяся тем, что продукция бактерией разветвленной L-аминокислоты повышена за счет увеличенной активности мутантной ацетолактатсинтазы, устойчивой к ингибированию по типу обратной связи L-валином.
Бактерия-продуцент разветвленной L-аминокислоты семейства Enterobacteriaceae не ограничивается бактерией, описанной выше. Более точно, бактерией-продуцентом разветвленной L-аминокислоты также может быть бактерия, продуцирующая L-валин, L-лейцин и/или L-изолейцин.
Бактерия-продуцент L-валина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-валина в соответствии с настоящим изобретением, включают, но не ограничиваются ими, штаммы, имеющие модификации, приводящие к сверхэкспрессии генов оперона ilvGMEDA (Патент США №5,998,178). Желательно удалить фрагмент оперона ilvGMEDA, отвечающий за ослабление экспрессии, с тем, чтобы экспрессия оперона не ослаблялась образующимся L-валином. Далее, в опероне может быть разрушен ген ilvA для того, чтобы снизить активность треониндезаминазы.
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-валина, также включают мутантные штаммы, имеющие мутацию в аминоацил-тРНК-синтетазе (Патент США №5,658,766). Например, может использоваться штамм Е.coli VL1970, который имеет мутацию в гене ileS, кодирующем изолейцин-тРНК-синтетазу. Штамм Е.coli VL1970 депонирован в Российской Национальной Коллекции Промышленных Микроорганизмов (ВКПМ) (Россия, 117545 Москва, 1-й Дорожный проезд, 1) 24 июня 1988 г. с инвентарным номером ВКПМ В-4411. Далее, в качестве родительских штаммов также могут использоваться мутантные штаммы, для роста которых требуется липоевая кислота, и/или с недостаточным количеством H+-АТФазы (WO 96/06926).
В качестве родительского штамма могут использоваться бактерии-продуценты L-валина, принадлежащие к роду Escherichia, такие как H-81 (ВКПМ В-8066), NRRL В-12287 и NRRL В-12288 (Патент США №4,391,907), ВКПМ В-4411 (Патент США №5,658,766), ВКПМ В-7707 (Европейская Патентная заявка EP 1016710 A2), NS1610 (ссылка на Патент ЕР1942183, Пример 7 или примеры, упомянутые там же) и подобные им.
Бактерия-продуцент L-лейцина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-лейцина, включают, но не ограничиваются ими, штаммы, принадлежащие к роду Escherichia, такие как штаммы Е.coli, устойчивые к лейцину (например, штамм 57 (ВКПМ В-7386, Патент США №6,124,121)) или аналогам лейцина, включающие р-2-тиенилаланин, 3-гидроксилейцин, 4-азалейцин, 5,5,5-трифторлейцин (JP 62-34397B и JP 8-70879A); штаммы E.coli, полученные с помощью генно-инженерных методов и описанные в WO 96/06926; Е.coli H-9068 (JP 8-70879 A), и подобные им.
Бактерия может быть улучшена путем усиления экспрессии одного или нескольких генов, вовлеченных в биосинтез L-лейцина. Примеры таких генов включают в себя гены оперона leuABCD, которые могут быть представлены мутантным геном leuA, кодирующим изопропилмалатсинтазу со снятым ретро-ингибированием L-лейцином (Патент США №6,403,342). Кроме того, бактерия согласно настоящему изобретению может быть улучшена путем усиления экспрессии одного или нескольких генов, кодирующих белки, которые экспортируют L-аминоки слоту из бактериальной клетки. Примеры таких генов включают в себя гены Ь2682 и Ь2б83 (гены удаZH) (Европейская Патентная заявка EP 1239041 A2).
В качестве родительского штамма могут использоваться бактерии-продуценты L-лейцина, принадлежащие к роду Escherichia, такие как H-9070 (FERM ВР-4704) и Н-9072 (FERM BP-4706) (Патент США №5744331), ВКПМ В-7386 и ВКПМ В-7388 (Патент РФ №2140450), W1485atpA401/pMWdAR6, W14851ip2/pMWdAR6, AJ12631/pMWdAR6 (Европейская Патентная заявка EP 0872547) и подобные им.
Бактерия-продуцент L-изолейцина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-изолейцина, включают, но не ограничиваются ими, мутантные штаммы с устойчивостью к 6-диметиламинопурину (заявка Японии 5-304969А), мутантные штаммы с устойчивостью к аналогу изолейцина, такому как тиаизолейцин и гидроксамат изолейцина, и мутантные штаммы, дополнительно имеющие устойчивость к DL-этионину и/или гидроксамату аргинина (заявка Японии 5-130882A). Кроме того, в качестве родительских штаммов также могут использоваться рекомбинантные штаммы, трансформированные генами, кодирующими белки, вовлеченные в биосинтез L-изолейцина, такие как треониндезаминаза и ацетогидроксатсинтаза (заявка Японии 2-458A, Европейская Патентная заявка EP 0356739 и Патент США №5,998,178).
Бактерия-продуцент высших спиртов и D-пантотеновой кислоты
Бактерия-продуцент L-аминокислоты семейства Enterobacteriaceae и, более точно, бактерия-продуцент разветвленной L-аминокислоты семейства Enterobacteriaceae может быть использована для получения высших спиртов и органических кислот, или их производных. Например, высшие спирты, такие как изобутанол, 2-метил-1-бутанол и 3-метил-1-бутанол, и органическая кислота, такая как D-пантотеновая кислота (витамин B5) могут быть получены с использованием подобной бактерии.
Известно из области техники, что в бактерии семейства Enterobacteriaceae биосинтетический путь L-валина, L-лейцина и L-изолейцина проходит через кето-кислотные интермедиаты. Более точно, 2-оксовалериат (2-кетовалериат, 2-КИВ) является предшественником для L-валина и L-лейцина; а 2-оксобутаноат является предшественником для L-изолейцина. Также, из уровня техники известно, что кето-кислотные интермедиаты или их производные могут быть предшественниками для высших спиртов с длиной углеродной цепи более чем 2 углеродных атома ввиду наличия пути деградации Эрлиха (Yan Y. and Liao J.C. J. Ind. Microbiol. Biotechnol., 2009, 36:471-479). Например, 2-оксоизовалериат является предшественником для изобутанола, а его производное 4-метил-2-оксопентаноат является предшественником для 3-метил-1-бутанола; 2-оксобутаноат является предшественником для пропан-1-ола, а его производное 3-метил-2-оксопентаноат является предшественником для 2-метил-1-бутанола.
Способность к продукции высших спиртов может быть сообщена штаммам E.coli, которые изначально не способны продуцировать высшие спирты, путем введения генов kivd и adh2 из донорных микроорганизмов, кодирующих, соответственно, кетокислотадекарбоксилазу (КДК) с широкой субстратной специфичностью и алкогольдегидрогеназу (АДГ). Например, могут использоваться ген kivd из Bacillus subtilis и ген adh2 из Lactococcus lactis. В этом аспекте можно сослаться на Connor M.R. and Liao J.C., Appl. Env. Mocrobiol., 2008, 74:5769-5775; и Патентную заявку WO 2009046370 A2.
Таким образом, способность бактерии продуцировать высшие спирты, такие как изобутанол, 3-метил-1-бутанол и 2-метил-1-бутанол может быть сообщена бактерии за счет введения в бактерию-продуцент L-аминокислоты семейства Enterobacteriaceae генетической экспрессионной системы и подходящих гетерологичных генов, например, kivd и adh2 или их вариантов из донорных микроорганизмов.
Также известно из уровня техники, что в бактерии семейства Enterobacteriaceae 2-оксовалериат является предшественником D-пантотеновой кислоты. Таким образом, способность к продукции D-пантотеновой кислоты может быть сообщена бактерии путем модификации бактерии семейства Enterobacteriaceae генетической экспрессионной системой.
2. Генетическая экспрессионная система
Экспрессионная система может быть регуляторной системой транскрипции одного или нескольких целевых генов, которая включает элементы транскрипционного аппарата, регулируемого белком типа LysR. Элементы транскрипционного аппарата, регулируемого белком типа LysR, могут включать промотор, оператор, экспрессия которых положительно регулируется регуляторным белком типа LysR и коиндуктором.
Система экспрессии также может включать один или несколько целевых генов, которые функционально связаны с вышеупомянутым транскрипционным аппаратом. Термин «функционально связан» может означать, что целевой ген связан с регуляторной последовательностью(ями), такой как промотор или оператор таким способом, что возможна экспрессия этого гена.
Один или несколько целевых генов могут кодировать белки, вовлеченные в биосинтез коиндуктора, субстрата или предшественника указанного коиндуктора.
В соответствии с вышеописанной схемой экспрессионной системы, экспрессия системы может быть самоиндуцируемой и положительно регулируемой по типу обратной связи, опосредованной коиндуктором, полученным с участием продукта(ов) экспрессии целевого(ых) гена(ов). Такая экспрессионная система может называться «самоиндуцируемая регулируемая по типу обратной связи экспрессионная система», но с целью упрощения также может называться здесь как «экспрессионная система» или «система экспрессии». Система экспрессии может саморегулироваться по принципу положительной обратной связи коиндуктором, который может быть субстратом или предшественником для полезного метаболита в биосинтетическом пути, более точно, в биосинтетическом пути разветвленных L-аминокислот и/или его побочном пути.
«Регуляторный белок типа LysR» также может иметь название «транскрипционный регулятор типа LysR (LTTR)», «транскрипционный регулятор семейства LysR» или просто «регулятор». Регуляторный белок типа LysR может принадлежать к разнообразным семействам олигомерных бактериальных транскрипционных факторов, которые регулируют широкий спектр элементов транскрипции в ответ на широкий спектр внешних сигналов. Представители этого семейства могут выступать в качестве транскрипционных активаторов и/или транскрипционных репрессоров и иметь несколько общих структурных характеристик, таких как i) ДНК-связывающий домен, имеющий мотив спираль-поворот-спираль (остатки могут быть из положений с 1 по 65 с N-конца LTTR), (ii) домены, вовлеченные в распознавание коиндуктора и/или отклика (остатки могут быть из положений с 100 по 173 и с 196 по 206), и (iii) домен, необходимый для связывания ДНК и отклика (остатки могут быть из положений с 227 по 253), как описано в работе Schell M.A., Ann. Rev. Microbiol., 1993, 47:597-626. В отсутствие коиндуктора, LTTR могут связываться с регулируемыми промоторами посредством двойного двуцепочечного участка длиной 15 п.н., имеющего общую структуру, и участком ДНК в позиции около -65, как описано в работе Schell M.A., Ann. Rev. Microbiol., 1993, 47:597-626. В присутствие коиндуктора могут происходить дополнительные взаимодействия LTTR с участками связывания РНК-полимеразы (около -35) и/или изгиб ДНК, что приводит к активации транскрипции (Schell M.A., Ann. Rev. Microbiol., 1993, 47:597-626). Более того, некоторые представители семейства регуляторных белков типа LysR могут иметь другие четыре функциональные характеристики, такие как: (i) быть коиндуктор-зависимым транскрипционным регулятором различного размера от 276 до 324 аминокислотных остатков, (ii) независимо от присутствия коиндуктора связываться с операторными участками ДНК, имеющими схожую позицию и структурный мотив, (iii) разнонаправлено транскрибироваться с промотора, который расположен близко или даже перекрывается с промотором регулируемого гена, и (iv) подавлять собственную транскрипцию в различной степени, например, от 3 до 10 раз, т.е. быть отрицательно саморегулируемыми, как описано в работе Schell M.A., Ann. Rev. Microbiol., 1993, 47:597-626.
В качестве примера, но не ограничиваясь им, система экспрессии может включать транскрипционный аппарат, регулируемый белком типа LysR, который содержит самоиндуцируемый промотор с положительной обратной регуляцией, оператор и целевой ген(ы). Экспрессионная система есть система с положительной регуляцией регуляторным белком типа LysR и коиндуктором. Пример самоиндуцируемого промотора с положительной обратной регуляцией может включать промотор PilvC. Пример регуляторного белка типа LysR может включать белок IlvY. Примеры коиндуктора могут включать представителя 2-ацето-2-гидроксикарбоновых кислот, такого как 2-ацетомолочная кислота (2-acetolactic acid, AL, AΛ), 2-ацето-2-гидроксимаслянная кислота (2-aceto-2-hydroxybutyric acid, AHB, АГБ) или их соли. Примеры целевых генов могут включать гены, кодирующие ацетолактатсинтазу I, II и/или III или их мутантные варианты. Пример системы экспрессии может быть описан схематически без ограничения типа, количества и расположения элементов системы экспрессии.
Гены ilvY и ilvC расположены рядом друг с другом на противоположных сторонах цепи как показано на Фигуре 1. Транскрипция генов ilvY and ilvC инициируется с дивергентно перекрывающихся промоторов ilvY и ilvC, соответственно. Однако, термины «ilvC промотор» или «PilvC промотор» могут означать промоторы генов ilvY и ilvC. В промоторной области находятся два тандемных оператора (Фигура 1).
Один или несколько целевых генов функционально связаны с транскрипционным аппаратом, более точно, с промотором. Например, в случае, если целевыми генами являются гены ilvBN, кодирующие ацетолактатсинтазу I, и когда промотор есть промотор ilvC, то нативный участок промотора, расположенного перед генами ilvBN, замещен на фрагмент ДНК, который включает индуцируемый промотор гена ilvC и второй оператор таким образом, что экспрессия указанных генов контролируется этим промотором. Например, гены оперона ilvBN4, кодирующие AHAS I, обладающую устойчивостью к ингибированию по типу обратной связи L-валином, помещены под промотор ilvC, который становится транскрипционно активированным только тогда, когда образуется комплекс между AΛ или АГБ, транкрипционным регулятором IlvY и вторым операторным участком. Кассета экспрессии, которая включает промотор PilvC и оперон генов ilvBN4, позволяет генам ilvBN4 транскрибироваться для продукции AHAS I. AHAS I превращает два моля пирувата в один моль 2-ацетолактата (AΛ) или один моль пирувата и один моль 2-оксобутаноата в один моль 2-ацето-2-гидроксибутаноата (АГБ) в зависимости от наличия 2-оксобутаноата.
Преимущество настоящего изобретения заключается в том, что часть продукта реакции (AΛ или АГБ), катализируемой AHAS I, действует как коиндуктор и связывается в комплекс с IlvY/ДНК, индуцируя транскрипцию генов оперона ilvBN4 и, таким образом, обеспечивая синтез AΛ или АГБ. Другая часть AΛ или АГБ может превращаться в целевой продукт - разветвленную L-аминокислоту (L-валин, L-лейцин или L-изолейцин) с помощью кето-кислотной редуктоизомеразы IlvC и других ферментов из биосинтетического пути разветвленных L-аминокислот как описано выше.
Непрерывное снабжение системы экспрессии 2-ацетолактатом или 2-ацето-2-гидроксибутаноатом может придавать системе свойство самоиндуцируемости; и циклический поток коиндуктора через самоиндуцируемую систему экспрессии с регуляцией по принципу положительной обратной связи может приводить к непрерывному снабжению пути биосинтеза разветвленных L-аминокислот предшественниками, такими как 3-гидрокси-3-метил-2-оксобутаноат для L-валина и L-лейцина или 3-гидрокси-3-метил-2-оксопентаноат для L-изолейцина.
Предложенный подход для экспрессии генов может быть назван как самоиндуцируемая генетическая экспрессионная система, имеющая положительную регуляцию по типу обратной связи и опосредованная коиндуктором AΛ или АГБ.
Экспрессионная система, функционирующая по принципу самоиндукции с положительной обратной связью, не ограничивается упомянутой выше системой экспрессии, включающей транскрипционный регулятор ilvY. Экспрессионная система также может включать другие системы экспрессии, использующие регуляторные белки типа LysR, которые описаны в Таблице 1.
Термин «ацетолактатсинтаза» может означать фермент, существующих в бактериях, таких как бактерии семейства Enterobacteriaceae, бактерии рода Corynebacterium, бактерии рода Bacillus и т.д. Примерами бактерий семейства Enterobacteriaceae могут быть бактерии, принадлежащие к роду Escherichia, Pantoea, Erwinia, Providencia и Serratia, такие как E.coli, Pantoea ananatis (P. ananatis) и подобные им. Примером коринеформной бактерии может быть бактерия, принадлежащая к роду Corynebacterium, такая как Corynebacterium glutamicum. Примерами бактерий, принадлежащих к роду Bacillus, могут быть Bacillus subtilis. Bacillus amyloliquefaciens FZB42 и Bacillus amyloliquefaciens DSM7. Термин «ацетолактатсинтаза» также может означать фермент, имеющий активность ацетолактатсинтазы.
Термин «активность ацетолактатсинтазы» может означать способность фермента к катализу реакции образования i) 2-ацетолактата и CO2 из двух молекул пирувата и/или ii) 2-ацето-2-гидроксибутаноата и CO2 из пирувата и 2-оксобутаноата при соответствующих условиях, таких как температура, ионная сила, кислотность (pH), концентрация кофакторов и субстратов и т.д. Активность ацетолактатсинтазы можно измерить, используя метод Stormer F.C. and Umbarger H.E., Biochem. Biophys. Res. Commun., 1964, 17(5):587-592.
Ацетолактатсинтаза I, II или III (AHAS I, AHAS II или AHAS III) (EC 2.2.1.6) также может быть названа как ацетолактатсинтаза. Ацетолактатсинтаза является гетеротетрамерным белком со структурой типа α2β2, состоящим из двух каталитических и двух регуляторных доменов (Weinstock О. et al., J. Bacterial., 1992, 174(17):5560-55бб). Считается, что большая субъединица (примерно 60-kDa) является каталитической, в то время как малая субъединица (примерно 11-kDa) - регуляторной.
AHAS I кодируется генами ilvB и ilvN, образующими оперон ilvBN. AHAS II кодируется генами ilvG and ilvM, образующими оперон ilvGMEDA. Гены, кодирующие AHAS II, обычно не экспрессируются в клетках E.coli K-12 (Guardiola J. et al., Mol. Gen. Genet, 1977, 156:17-25). AHAS III кодируется генами ilvI и ilvH, образующими оперон ilvIH.
AHAS I из E.coli, например, может иметь замены аминокислотных остатков, такие как N17K и/или A30P, которые сообщают устойчивость к ингибированию по типу обратной связи L-валином (Патент РФ №2355763, Патентная заявка США №2009197309 A1). Также, замещение аланина в положении 33 на любую аминокислоту или несколько аминокислот, такое как замещение его на 12 аминокислот, содержащих сайт терминации трансляции, приводит к белку IlvN33, укороченному на 45 аминокислотных остатка (Патент РФ №2355763, Патентная заявка США №2009197309 A1). Этот мутантный белок также обладает устойчивостью к ингибированию по типу обратной связи L-валином.
Малая субъединица мутантной AHAS I, содержащая замену N17K (Asn в положение 17 заменен на Lys, т.е. соответствующий кодон AAC заменен на AAG) в диком типе AHAS I, может быть кодирована мутантным геном ilvN, который может быть обозначен как ген ilvN4 или ген ilvN ValR4, как описано в EP 1942183. AHAS I, имеющая аминокислотную последовательность, представленную в SEQ ID NOs: 2 и 4, может быть названа как «ацетолактатсинтаза I дикого типа».
На основании модели валин-связывающего участка регуляторной (малой) субъединицы AHAS III из Е.coli были сделаны укорочения карбоксильного конца малой субъединицы. Подобная модификация привела к тому, что укороченный фермент AHAS III потерял чувствительность к валину (Mendel S. et al., J. Mol. Biol., 2003, 325(2):275-284).
Ацетолактатсинтаза III, которая укорочена с карбоксильного конца малой субъединицы на 35, 48, 80 или 95 аминокислотных остатков, может быть названа как «мутантная ацетолактатсинтаза III». ДНК, кодирующая малую субъединицу мутантной AHAS III, может быть названа как «мутантный ген ilvH». Оперон ilvIH, содержащий мутантный ген ilvH, может быть назван как «мутантный оперон ilvIH». AHAS III, имеющая аминокислотную последовательность, представленную в SEQ ID NOs: 6 и 8, может быть названа как «ацетолактатсинтаза III дикого типа».
Мутантная AHAS I может содержать делецию, замену, вставку или добавление одного или нескольких аминокислотных остатков в одном или нескольких положениях помимо 17, 30 и/или 33 в исходной аминокислотной последовательности при условии, что активность ацетолактатсинтазы сохранена. Кроме того, мутантная AHAS III также может содержать делецию, замену, вставку или добавление одного или нескольких аминокислотных остатков в одном или нескольких положениях в исходной аминокислотной последовательности при условии, что активность ацетолактатсинтазы сохранена. Кроме того, карбоксильный конец мутантной AHAS III может быть укорочен на один или несколько аминокислотных остатков, не ограничиваясь 35, 48, 80 или 95 аминокислотными остатками, при условии, что активность ацетолактатсинтазы не уменьшилась.
Количество «нескольких» аминокислотных остатков различается в зависимости от положения в третичной структуре белка или типа аминокислотного остатка. Это возможно потому, что некоторые аминокислоты подобны друг другу по структуре и функции внутри белка, и взаимные перестановки таких аминокислот не сильно влияют на третичную структуру или функцию белка. AHAS III может быть укорочена последовательностью аминокислотных остатков различной длины до тех пор, пока укороченный фермент способен связывать и активировать каталитическую (большую) субъединицу, так чтобы мутантная AHAS III сохраняла активность. Таким образом, мутантные AHAS I, II и III могут иметь гомологию не менее чем 65%, не менее чем 80%, не менее чем 90%, не менее чем 95%, не менее чем 97% или не менее чем 99% по сравнению с изначальной аминокислотной последовательностью ацетолактатсинтазы при условии, что активность ацетолактатсинтазы сохранена. В связи с этим, в настоящем описании термин «гомология» может означать «идентичность».
Мутантная AHAS I может быть получена введением мутаций в ген ilvN дикого типа с использованием известных методов. Например, мутантный оперон ilvBN4, который содержит ген ilvB и мутантный ген ilvN4, может быть получен методом ПЦР (полимеразная цепная реакция), как описано в White T.J. et al., Trends Genet., 1989, 5:185-189, с использованием праймеров, подобранных на основании нуклеотидной последовательности гена UvN (SEQ ID NO: 3). Гены, кодирующие ацетолактатсинтазу из других микроорганизмов, могут быть получены похожим способом.
Мутантная AHAS III с укороченным карбоксильным концом малой субъединицы может быть получена сайт-направленным мутагенезом с использованием ПЦР, позволяющей получать ДНК-фрагменты с перекрывающимися участками, путем введения стоп-кодонов в желаемое положение гена UvH (Но S.N. et al., Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene, 1989, 77:51-59). Праймеры для ПЦР, пригодные для синтеза генов оперона ilvIH, могут быть подобраны на основании нуклеотидной последовательности гена ilvH (SEQ ID NO: 7).
Ген ilvB кодирует большую субъединицу ацетолактатсинтазы I (KEGG, Kyoto Encyclopedia of Genes and Genomes, Киотская энциклопедия генов и геномов, No.b3671). Ген ilvB (инвентарный номер в GenBank NC_000913.2; нуклеотиды, комплементарные нуклеотидам в положении с 3849119 по 3850807; Gene ID: 948182) расположен между генами ilvN и ivbL на хромосоме E.coli K-12. Нуклеотидная последовательность гена ilvB и аминокислотная последовательность большой субъединицы ацетолактатсинтазы I, кодируемой геном ilvB, приведены в последовательностях SEQ ID NO: 1 и SEQ ID NO: 2, соответственно.
Ген ilvN кодирует малую субъединицу ацетолактатсинтазы I (KEGG, No.b3670). Ген ilvN (инвентарный номер в GenBank NC_000913.2; нуклеотиды, комплементарные нуклеотидам в положении с 3848825 по 3849115; Gene ID: 948183) расположен между генами uhpA и ilvB на хромосоме E.coli K-12. Нуклеотидная последовательность гена ilvN и аминокислотная последовательность малой субъединицы ацетолактатсинтазы I, кодируемой геном ilvN, приведены в последовательностях SEQ ID NO: 3 и SEQ ID NO: 4, соответственно.
Ген ilvI кодирует большую субъединицу ацетолактатсинтазу III (KEGG, No.b0077). Ген ilvI (инвентарный номер в GenBank NC_000913.2; положения нуклеотидов с 85630 по 87354; Gene ID: 948793) расположен между генами leoO и ilvH на хромосоме E.coli K-12. Нуклеотидная последовательность гена ilvI и аминокислотная последовательность большой субъединицы ацетолактатсинтазы III, кодируемой геном ilvI, приведены в последовательностях SEQ ID NO: 5 и SEQ ID NO: 6, соответственно.
Ген ilvH кодирует малую субъединицу ацетолактатсинтазы III (KEGG, No.b0078). Ген ilvI (инвентарный номер в GenBank NC_000913.2; положения нуклеотидов с 87357 по 87848 Gene ID: 947267) расположен между генами ilvI и fruR на хромосоме E.coli K-12. Нуклеотидная последовательность гена ilvH и аминокислотная последовательность малой субъединицы ацетолактатсинтазы III, кодируемой геном ilvH, приведены в последовательностях SEQ ID NO: 7 и SEQ ID NO: 8, соответственно.
Ген ilvY кодирует ДНК-связывающий двойной транскрипционный регулятор ilvY (транскрипционный регулятор семейства LysR, положительный регулятор для ilvC) (KEGG, No.B3773). Ген ilvY (инвентарный номер в GenBank NC_000913.2; нуклеотиды, комплементарные нуклеотидам в положении с 3954950 по 3955843; Gene ID: 948284) расположен между генами ilvA и ilvC на хромосоме Е.coli K-12. Нуклеотидная последовательность гена ilvI и аминокислотная последовательность белка IlvY, кодируемого геном ilvY, приведены в последовательностях SEQ ID NO: 9 и SEQ ID NO: 10, соответственно.
Ген ilvG является псевдогеном (KEGG, No.b4488). Ген ilvG (инвентарный номер в GenBank NC_000913.2; положения нуклеотидов с 3948583 по 3950227; Gene ID: 2847699) расположен между генами ilvX и ilvM на хромосоме E.coli K-12. Нуклеотидная последовательность гена ilvG приведена в SEQ ID NO: 31. Ген ilvG может кодировать большую субъединицу ацетолактатсинтазы II при условии, что он содержит замену кодона TGA на кодон AAT в положении с 982 по 984 от начала гена, или мутации, как описано в Lawther R.P. et al., J. Bacteriol., 1982, 159:294-298. Аминокислотная последовательность большой субъединицы ацетолактатсинтазы II, кодируемой геном ilvG, в котором кодон TGA замещен на кодон AAT в положениях от 982 до 984, приведена в SEQ ID No: 32.
Ген ilvM кодирует малую субъединицу ацетолактатсинтазы II (KEGG, No.b3769). Ген ilvM (GenBank accession No. NC_000913.2; положения нуклеотидов с 3950224 по 3950487; Gene ID: 948279) расположен между генами ilvG и ilvE на хромосоме E.coli K-12. Нуклеотидная последовательность гена ilvM и аминокислотная последовательность малой субъединицы ацетолактатсинтазы II, кодируемой геном ilvM, приведена в последовательностях SEQ ID NO: 33 и SEQ ID NO: 34, соответственно.
Поскольку могут существовать некоторые различия в последовательностях ДНК между родами или штаммами бактерий семейства Enterobacteriaceae, гены ilvB, ilvN, ilvI, ilvH, ilvG и ilvM, кодирующие ацетолактатсинтазу, и ген ilvY, кодирующий транскрипционный регулятор, не ограничиваются генами, представленными в последовательностях SEQ ID NOs: 1, 3, 5, 7, 31, 33 и 9, но могут включать гены, которые являются вариантами или гомологами последовательностей, представленных в SEQ ID NOs: I, 3, 5, 7, 31, 33 и 9, и которые кодируют варианты белков IlvB, IlvN, IlvI, IlvH, IlvG, IlvM и IlvY.
Термин «вариант белка», используемый в настоящем изобретении, может означать белок, который содержит одно или несколько изменений в аминокислотных последовательностях, представленных в SEQ ID NOs: 2, 4, 6, 8, 32, 34 и 10, которыми могут быть замены, делении, вставки и/или добавления одного или нескольких аминокислотных остатков при условии, что активность такого белка такая же, как и у белков IlvB, IlvN, Ilvl, IlvH, IlvG, IlvM и IlvY, соответственно. Количество изменений в вариантах белков зависит от вида или положения аминокислотного остатка в третичной структуре белка. Оно может быть от 1 до 30, или от 1 до 15, или от 1 до 5 в SEQ ID NO: 2, 4, 6, 8, 32, 34 и 10
Замены, делении, вставки и/или добавления одного или нескольких аминокислотных остатков могут быть консервативными мутациями при условии, что активность и/или функция варианта белка сохранена или подобна активности белков IlvB, IlvN, IlvI, IlvH, IlvG, IlvM и IlvY. Примером консервативной мутации может быть консервативная замена. Консервативными заменами могут быть взаимные замены между Ser или Thr на Ala, замены Gln, His или Lys на Arg; замена Glu, Gln, Lys, His или Asp на Asn; замена Glu или Gln на Asp; замена Ser или Ala на Cys; замена Asn, Glu, Lys, His, Asp или Arg на Gln; замена Gly, Asn, Gln, Lys или Asp на Glu; замена Pro на Gly; замена Asn, Lys, Gln, Arg или Trp на His; замена Leu, Met, Val или Phe на Ile; замена Ile, Met, Val или Phe на Leu; замена Asn, Glu, Gln, His или Arg на Lys; замена Ile, Leu, Val или Phe на Met; замена Trp, Tyr, Met, Ile или Leu на Phe; замена Thr или Ala на Ser; замена Ser или Ala на Thr; замена Phe или Tyr на Trp; замена His, Phe или Trp на Tyr; и замена Met, Ile или Leu на Val.
Гомология между белками или ДНК может быть определена с использованием нескольких известных подходов, например, компьютерных алгоритмов BLAST и FASTA и метода ClustalW. Алгоритм BLAST (Basic Local Alignment Search Tool, www.ncbi.nlm.nih.gov/BLAST/), позволяющий проводить иерархический поиск, заложен в программах blastp, blastn, blastx, megablast, tbiastn и tbiastx; эти программы присваювают уровень значимости найденным объектам, используя статистические методы, описанные в Samuel К. and Altschul S.F. («Methods for assessing the statistical significance of molecular sequence features by using general scoring schemes» Proc. Natl. Acad. Sci. USA, 1990, 87:2264-2268; «Applications and statistics for multiple high-scoring segments in molecular sequences». Proc. Natl. Acad. Sci. USA, 1993, 90:5873-5877). Алгоритм BLAST вычисляет три параметра: число аминокислотных остатков, идентичность и сходство. Поисковый алгоритм FASTA описан в Pearson W.R. («Rapid and sensitive sequence comparison with FASTP and FASTA», Methods Enzymol., 1990, 183:63-98). Метод ClustalW описан в Thompson J.D. et al. («CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice». Nucleic Acids Res., 1994, 22:4673-4680).
Более того, гены ilvB, ilvN, ilvI, ilvH, ilvG, ilvM и ilvY могут быть вариантами нуклеотидных последовательностей.
Термин «вариант нуклеотидной последовательности» может означать нуклеотидную последовательность, кодирующую «вариант белка». Термин «вариант нуклеотидной последовательности» также может означать нуклеотидную последовательность, гибридизуюшуюся в жестких условиях с нуклеотидной последовательностью, комплементарной нуклеотидным последовательностям, приведенным в Перечне последовательностей под номерами SEQ ID NOs: 1, 3, 5, 7, 31, 33 или 9, или с зондом, который может быть синтезирован на основе указанных нуклеотидных последовательностей в жестких условиях, при условии, что он кодирует функциональные ацетолактатсинтазу или регуляторный белок до инактивации. Под «жесткими условиями» понимаются такие условия, в которых образуется специфический гибрид, например, гибрид, имеющий гомологию не менее чем 70%, не менее чем 80%, не менее чем 90%, не менее чем 95%, не менее чем 96%, не менее чем 97% или не менее чем 99%, и в которых не образуется неспецифический гибрид, например, гибрид, имеющий гомологию меньшую, чем указанно выше. Практическим примером жестких условий является однократная, двух- или трехкратная отмывка при концентрации солей 1×SSC, 0.1% SDS или 0.1×SSC, 0.1% SDS, при 60°C или 65°C. Продолжительность отмывки зависит от типа используемой для блоттинга мембраны и, как правило, такова, как рекомендовано производителем. Например, рекомендуемая продолжительность отмывки для положительно заряженной нейлоновой мембраны Hybond™-N+(GE Healthcare) при жестких условиях составляет 15 минут. Промывка может быть произведена двух- или трехкратно. В качестве зонда может быть использована часть последовательности, комплементарная последовательности, приведенной в SEQ ID NOs: 1, 3, 5, 7, 31, 33 или 9. Подобный зонд может быть получен с помощью ПЦР с использованием олигонуклеотидных праймеров, выбранных на основе последовательностей, приведенных в SEQ ID NOs: I, 3, 5, 7, 31, 33 или 9, и фрагмента ДНК, содержащего нуклеотидную последовательность в качестве матрицы. Рекомендуемая длина зонда может быть подобрана в зависимости от условий гибридизации и составляет >50 п.н., обычно около 100-1000 п.н. Например, при использовании в качестве зонда фрагмента ДНК длиной около 300 п.н. условия отмывки могут быть следующие: 2×SSC, 0.1% SDS при 50°C, 60°C или 65°C.
Поскольку гены, кодирующие белки IlvB, IlvN, IlvI, IlvH, IlvG, IlvM и IlvY из E.coli, известны (см. выше), варианты нуклеотидных последовательностей, кодирующих белки IlvB, IlvN, IlvI, IlvH, IlvG, IlvM и IlvY, могут быть получены методом ПЦР (полимеразная цепная реакция), как описано в White T.J. et al., Trends Genet., 1989, 5:185-189 с использованием праймеров, подобранных на основе нуклеотидной последовательности генов ilvB, ilvN, ilvI, ilvH, ilvG, ilvM и ilvY. Гены, кодирующие белки IlvB, IlvN, IlvI, IlvH, IlvG, IlvM и IlvY или их варианты, могут быть получены подобным способом.
Термин «самоиндуцируемый положительно регулируемый промотор» может означать промотор, известный для специалиста в данной области. Традиционно самоиндуцируемый положительно регулируемый промотор может означать промотор, который имеет: a) повышенную активность, если уровень транскрипционного фактора или активатора увеличен, т.е. имеет место положительная регуляция; b) отсутствие или низкая активность в отсутствии транскрипционного фактора или активатора, однако есть наличие активации промотора, если он связан с транскрипционным фактором или активатором, т.е. имеет место индукция; и c) может быть индуцирован продуктом экспрессии кодирующего гена, контролируемого этим промотором, включая любой пост-трансляционно модифицированный продукт экспрессии, а также его аналоги и производные или образуемые им комплексы, включая продукты, такие как индивидуальные вещества или их комплексы, получающиеся в результате активности продукта экспрессии кодирующего гена, т.е. имеет место самоиндукция.
В качестве примера самоиндуцируемого положительно регулируемого промотора можно привести промотор Puvc (SEQ ID NO: 30) (Wek R.C. and Hatfield G.W., J. Biol. Chem., 1986, 261(5): 2441-2450; Opel M.L. and Hatfield G.W., Mol. Microbiol, 2001, 39(1):191-198), расположенный перед структурной частью гена ilvC, промотор PcysP, расположенный перед транскрипционной единицей cysPUWAM, промотор PcysK, расположенный перед структурной частью гена cysK, и промотор PmetR, расположенный перед структурной частью гена metE. Промотор PilvC может регулироваться комплексом индукции IlvY/AΛ или IlvY/АГБ. Промоторы PcysP и PcysK могут регулироваться комплексом самоиндукции CysB/O-ацетил-L-серин. Промотор PmetR может регулироваться комплексом самоиндукции MetR/L-гомоцистеин. Самоиндуцируемые положительно регулируемые промоторы не ограничены, и могут включать замену, делецию, вставку или добавление одного или нескольких нуклеотидных остатков при условии, что функциональность промотора сохранена.
Термин «самоиндуцируемый отрицательно регулируемый промотор» может означать промотор, известный для специалиста в данной области. Традиционно самоиндуцируемый отрицательно регулируемый промотор может означать промотор, который имеет: a) пониженную активность, если уровень транскрипционного фактора или активатора понижен, т.е. имеет место отрицательная регуляция; b) отсутствие или низкая активность в присутствии транскрипционного фактора или активатора, однако есть наличие активации промотора, если достигнута граничная концентрация транскрипционного фактора или активатора, т.е. имеет место индукция; и c) может быть дерегулирован продуктом экспрессии кодирующего гена, контролируемого этим промотором, включая любой пост-трансляционно модифицированный продукт экспрессии, а также его аналоги и производные или образуемые им комплексы, включая продукты, такие как индивидуальные вещества или их комплексы, получающиеся в результате активности продукта экспрессии кодирующего гена, т.е. имеет место самоиндукция.
Термин «самоиндуцируемый промотор» также может означать «саморегулируемый промотор», «авторегулируемый промотор» или им подобные вне зависимости от положительного или отрицательного типа регулирования.
Термин «система экспрессии модифицирована таким образом, что регуляция указанной системы, функционирующей по принципу самоиндукции с положительной обратной связью, опосредована коиндуктором» может означать, что система экспрессии отвечает за синтез коиндуктора за счет регуляции экспрессии гена, кодирующего фермент, который отвечает за синтез коиндуктора. Указанный коиндуктор может связываться с регуляторным комплексом белок/ДНК, чтобы активировать транскрипцию гена, кодирующего белок, который отвечает за синтез коиндуктора. Таким образом, очевидно, что функциональность, или регуляция указанной экспрессионной системы может быть опосредована коиндуктором.
Термин «индуцируемая экспрессионная система» может означать систему экспрессии, в которой уровень транскрипции может быть изменен не менее чем в 1,5 раза, или не менее чем в 2 раза, или не менее чем в 3 раза, или не менее чем в 5 раз, или не менее чем в 10 раз, или не менее чем в 15 раз, или не менее чем в 20 раз, или не менее чем в 30 раз, или не менее чем в 100 раз, или более. Термин «индуцируемая экспрессионная система» может включать любую систему экспрессии, которая может быть индуцирована, например, коиндуктором, таким как 2-ацетолактат, 2-ацетогидрокси-2-бутаноат, арабиноза, лактоза, ИПТГ и т.д.; индуктором, таким как регуляторный белок, принадлежащим к белкам семейства типа LysR; внешним воздействием, например, нагреванием, охлаждением и т.д.; или ростовыми условиями, такими как плотность клеток, кислотность (pH) и т.д. Термин «индуцируемая экспрессионная система» также может включать любую систему экспрессии, которая может быть репрессирована, например, систему экспрессии, которая может быть репрессирована при добавлении химических соединений, белков, под воздействием внешних факторов и т.п.
Термин «целевой ген» может означать ген, уровень экспрессии которого должен быть дерегулирован с использованием экспрессионной системы. В качестве примеров целевых генов можно привести ген(ы), кодирующий(е) белок(ки), вовлеченный(е) в биосинтез коиндуктора, или субстрата или предшественника коиндуктора, или синтазу коиндуктора. Более точно, примерами целевых генов могут быть гены, кодирующие большую и/или малую субъединицу(ы) AHAS I, II и/или III, или их мутантные варианты. Примером целевых генов может быть оперон ilvBN4, кодирующий мутантную AHAS I, устойчивую к ингибированию по типу обратной связи L-валином.
Термин «дерегуляция уровня экспрессии» целевого(ых) гена(ов) может означать ослабление, инактивацию или повышение экспрессии указанного(ых) гена(ов); повышение экспрессии является одним из примеров.
Термин «операторный участок» может означать фрагмент ДНК, функционально связанный с промоторным участком и влияющий на транскрипцию гена, расположенного под промотором. Примером операторного участка может быть фрагмент ДНК, функционально связанный с промоторами PilvC или PilvY и с которым может связываться транскрипционный регулятор IlvY.
Термин «регулятор гена(ов) системы экспрессии», который также может быть назван как «регулятор самоиндуцируемой системы с положительной обратной регуляцией» или «регулятор», может означать белок, принадлежащий к семейству регуляторных белков типа LysR, способный напрямую или опосредованно регулировать транскрипцию гена. Примеры регуляторных белков приведены в Таблице 1.
Термин «инактивация гена» может означать, что модифицированный ген кодирует полностью неактивный или нефункциональный белок. Также возможно, что модифицированный участок ДНК не способен к естественной экспрессии гена благодаря делеции части гена или целого гена, сдвигу рамки считывания гена, введению миссенс/нонсенс мутации(ий) или модификации смежного участка гена, включению последовательностей, контролирующих экспрессию гена, таких как промотор(ы), энхансер(ы), аттенуатор(ы), рибосома-связывающий(ие) сайт(ы) (RBS, ribosome-binding site) и т.д. Инактивация гена также может быть проведена обычными способами, такими как мутагенез путем обработки микроорганизмов, например, УФ-облучением или нитрозогуанидином (N-метил-N'-нитро-N-нитрозогуанидин), сайт-направленный мутагенез, нарушение структуры гена с использованием гомологичной рекомбинации и/или мутагенеза за счет вставки-делеции (Yu D. et al., Proc. Natl. Acad. Sci. USA, 2000, 97(12):5978-83; Datsenko K.A. и Wanner B.L., Proc. Natl. Acad. Sci. USA 2000, 97(12):6640-45), также называемого как «λRed-зависимая интеграция».
Термин «повышение экспрессии гена» или «экспрессия гена повышена» может означать, что благодаря генетической модификации уровень экспрессии мутантного гена выше, чем уровень экспрессии гена в немодифицированном штамме, например, в штамме дикого типа или родительском штамме, таком как штамм E.coli MG1655, E.coli K-12 (VKPM B-7) или E.coli B7 ΔilvGM ΔilvIH PL-ilvBN4. Примеры таких модификаций включают увеличение числа копий экспрессированных генов в клетке и/или увеличение уровня экспрессии гена модификацией смежного с геном участка, включающего последовательности, контролирующие экспрессию гена, такие как промоторы, энхансеры, аттенюаторы, рибосома-связывающие сайты и т.д., и другие примеры. Способы, которые могут использоваться для увеличения экспрессии гена, также могут включать введение гена в вектор, который способен повышать копийность гена в бактерии семейства Enterobacteriaceae. В качестве примеров таких векторов можно привести, но не ограничиться этим, векторы с широким кругом хозяев (broad-host-range vectors), например, pCM110, pRK310, pVK101, pBBR122, pBHR1 и подобные им. Увеличение экспрессии гена также может быть достигнуто введением множества копий гена в хромосомную ДНК бактерии, например, с помощью гомологичной рекомбинации, Mu интеграции или подобными методами.
Усиление экспрессии гена(ов) и/или генов оперона также может быть достигнуто путем подстановки фрагмента ДНК под контроль сильного промотора. Примерами сильных промоторов являются lac промотор, trp промотор, trc промотор, tac промотор, PR или PL промоторы фага λ. В данном изобретении могут быть использованы сильные промоторы, обеспечивающие высокий уровень экспрессии гена в бактерии, принадлежащей к семейству Enterobacteriaceae. С другой стороны, действие промотора может быть усилено, например, введением мутации в промоторный участок гена на бактериальной хромосоме, что приводит к увеличению уровня транскрипции гена, находящегося под промотором. Кроме того, известно, что замена нескольких нуклеотидов в области между сайтом связывания рибосомы и стартовым кодоном, особенно в последовательности непосредственно перед стартовым кодоном, существенно влияет на трансляционную способность мРНК. Например, обнаружен 20-кратный разброс в уровне экспрессии гена в зависимости от природы трех нуклеотидов, предшествующих стартовому кодону (Gold L. et al., Annu. Rev. Microbiol., 1981, 35:365-403; Hui A. et al., EMBO J., 1984, 3:623-629). Использование сильного промотора можно сочетать с увеличением числа копий гена.
Количество копий, присутствие или отсутствие гена и/или генов оперона может быть измерено, например, рестрикцией хромосомной ДНК с последующим блоттингом по-Саузерну (Southern blotting), используя зонд, подобранный на основе последовательности гена, флуоресцентную гибридизацию in situ (fluorescence in situ hybridization, FISH) и подобные им методы. Уровень экспрессии гена и/или генов оперона можно определить с помощью различных известных методов, включая измерение количества транскрибируемой мРНК с применением хорошо известных методов, например, Нозерн-блоттинга, количественную ОТ-ПЦР (RT-PCR) и т.п. Количество белка, кодируемого геном, можно определить известными методами, включая электрофорез в SDS-ПААГ (SDS-PAGE) с последующим иммуноблоттингом (Вестерн-блоттинг) (Western blotting analysis), и подобными.
Методы приготовления плазмидной ДНК, рестрикции и лигирования ДНК, трансформации, выбора нуклеотидов в качестве праймера и т.п. могут быть обычными методами, известными специалисту в этой области. Эти методы описаны, например, в Sambrook J., Fritsch E.F. и Maniatis Т., «Molecular Cloning: A Laboratory Manual, 2nd ed.». Cold Spring Harbor Laboratory Press (1989). Методы молекулярного клонирования и экспрессии гетерологичных генов описаны в Bernard R. Glick, Jack J. Pasternak и Cheryl L. Patten, «Molecular Biotechnology: principles and applications of recombinant DNA», 4th ed., Washington, D.C: ASM Press (2009); Evans Jr., T.C. and Xu M. - Q., «Heterologous gene expression in E.coli», 1st ed., Humana Press (2011).
Термин «активность фермента, кодируемого геном, повышена» может означать, что общая активность фермента в расчете на одну клетку или специфическая активность в расчете на одну молекулу фермента выше, чем общая и/или специфическая активность фермента в немодифицированном штамме, например, штамме дикого типа или родительском штамме. Для сравнения активности фермента в качестве немодифицированного штамма можно взять штамм E.coli K-12, содержащий ацетолактатсинтазу дикого типа.
Термин «биосинтетический путь», который также может называться как «метаболический путь» или «биохимический путь», может означать набор анаболических или катаболических (био)химических реакций для превращения биомолекулы одного вида в биомолекулу другого вида, например, более сложного вида по направлению к целевому конечному продукту или полезному метаболиту. Термин «биосинтетический путь» обычно понятен для специалиста в данной области. Требуемым конечным продуктом может быть L-аминокислота, нуклеозид, нуклеотид, кофактор, низший или высший спирт, органическая кислота, их производные, белок и т.д.
Термин «субстрат» может означать химическое или биохимическое соединение (вещество) любого вида, которое может быть превращено или предполагается, что может быть превращено в вещество или соединение другого вида под действием фермента. Термин «субстрат» может означать не только единичное вещество, но также комбинацию соединений, такие как растворы, смеси и другие вещества, которые содержат не менее чем один субстрат или его производное(ые). Термин «субстрат» также может означать соединения, выступающие в качестве приемлемого источника углерода для синтеза химических и биохимических веществ, которыми могут быть, например, происходящий из биомассы сахар, интермедиаты или конечные метаболиты, которые используются в биосинтетическом пути микроорганизма с измененным метаболизмом. Различные виды химических или биохимических веществ и соединений могут быть превращены в требуемый конечный продукт в результате единственной реакции, катализируемой ферментом. Примеры субстратов включают пируват и 2-оксобутаноат.
Термин «предшественник» может означать химическое или биохимическое соединение (вещество) любого вида, из которого может быть получено другое, более сложное химическое или биохимическое соединение (вещество) по направлению к целевому конечному продукту или полезному метаболиту. В биосинтезе L-валина примерами предшественников могут быть пируват, 2-ацетолактат, 3-гидрокси-3-метил-2-оксобутаноат, 2,3-дигидрокси-3-метилбутаноат и 2-оксоизовалериат. В биосинтезе L-лейцина примерами предшественников могут быть пируват, 2-ацетолактат, 3-гидрокси-3-метил-2-оксобутаноат, 2,3-дигидрокси-3-метилбутаноат и 2-оксоизовалериат, 2-изопропилмалат и т.д. В биосинтезе L-изолейцина примерами предшественников могут быть L-треонин, пируват, 2-ацето-2-гидроксибутаноат, 3-гидрокси-3-метил-2-оксопентаноат, 2,3-дигидрокси-3-метилпентаноат и т.д.
Термины «субстрат» и «предшественник» могут быть взаимозаменяемы.
Термин «биомолекула» может означать химическое или биохимическое соединение (вещество) любого вида, продукт, получаемый с помощью микроорганизма. Биомолекулами могут быть белки, полисахариды, липиды, нуклеиновые кислоты и небольшие молекулы, такие как первичные метаболиты, вторичные метаболиты и природные вещества.
Термин «полезный метаболит» может означать химическое или биохимическое соединение (вещество) любого вида, получаемое с помощью микроорганизма для промышленных, кормовых, пищевых, фармакологических или других целей. В качестве полезных метаболитов можно привести разветвленные L-аминокислоты, такие как L-валин, L-изолейцин и L-лейцин; высшие спирты, такие как изобутанол, 2-метил-1-бутанол, и 3-метил-1-бутанол; и органическую кислоту, такую как D-пантотеновая кислота.
Экспрессионная система может быть введена в бактерию вышеописанным способом для усиления экспрессии гена, например, с использованием вектора, содержащего экспрессионную систему. Экспрессионная система может быть также введена в бактерию замещением нативного промотора целевого гена(ов) самоиндуцируемым промотором. Бактерия может содержать ген, кодирующий регуляторный белок типа LysR, положительно регулирующий промотор. Экспрессия гена, кодирующего регуляторный белок типа LysR, может быть усилена, однако, это усиление не существенно. В случае, когда промотор есть промотор ilvC регуляторный белок типа LysR есть белок ilvY, эндогенной экспресси гена ilvY обычно бывает достаточно.
3. Способ получения полезных метаболитов, таких как L-аминокислоты, высшие спирты и D-пантотеновая кислота Способ получения полезных метаболитов, более точно, L-аминокислот, особенно, разветвленных L-аминокислот, таких как L-валин, L-лейцин и L-изолейцин, высших спиртов, таких как изобутанол, 2-метил-1-бутанол и 3-метил-1-бутанол и органических кислот, таких как D-пантотеновая кислота может включать стадии выращивания бактерии в питательной среде с целью образования, экскреции и накопления полезного метаболита в питательной среде и извлечения L-аминокислоты, высшего спирта и/или органической кислоты из культуральной жидкости.
Выращивание (культивирование), выделение и очистка метаболитов из культуральной жидкости могут быть осуществлены способами, подобными традиционным способам ферментации, в которых аминокислота, высший спирт или органическая кислота продуцируется микроорганизмом. Питательная среда, используемая для получения полезных метаболитов, может быть обычной питательной средой, которая содержит источники углерода, азота, неорганические ионы и другие необходимые органические компоненты. В качестве источника углерода могут использоваться различные углеводы, такие как глюкоза, лактоза, галактоза, фруктоза, арабиноза, мальтоза, ксилоза, трегалоза, рибоза и гидролизаты крахмала; спирты, такие как глицерин, маннитол и сорбитол; органические кислоты, такие как глюконовая кислота, фумаровая кислота, лимонная кислота, яблочная кислота и янтарная кислота; и подобные им. В качестве источника азота могут использоваться различные неорганические аммонийные соли, такие как сульфат аммония, хлорид аммония и фосфат аммония; органический азот такой, как гидролизаты соевых бобов; водный раствор аммиака; и подобные им. Желательно, чтобы витамины, такие как витамин B1, необходимые вещества, например, органические добавки, такие как нуклеиновые кислоты, аденин и РНК, или дрожжевой экстракт и т.п. могли присутствовать в соответствующих или даже в минимальных количествах. Кроме того, при необходимости могут быть добавлены малые количества фосфата кальция, сульфата магния, ионы железа, ионы марганца и т.п.
Выращивание может быть выполнено в аэробных условиях в течение от 16 до 72 ч при температуре в интервале 30-45°C или в интервале 30-37°C при кислотности (pH) питательной среды в интервале 5-8 или 6,5-7,2. pH можно регулировать с использованием неорганических или органических кислот или оснований, таких как, например, газообразный аммиак. Обычно выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки и клеточный дебрис могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем целевая L-аминокислота или органическая кислота может быть выделена и очищена с использованием любой комбинации традиционных методов, таких как ионообменная хроматография, концентрация и кристаллизация. Высшие спирты могут быть выделены из культуральной жидкости, например, дистилляцией с последующей очисткой повторной дистилляцией или хроматографией.
Примеры
Настоящее изобретение более подробно будет описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение Примеры.
Пример 1. Конструирование экспрессионной кассеты, регулируемой ацетогидроксикислотой, и анализ ее экспрессии с использованием гена-репортера lacZ при различных генетических окружениях
Транскрипция генов ilvYC хорошо известна (Rhee К. et al., Proa. Natl. Acad. Sci. USA, 1999, 96(25):14294-14299; Opel M.L. and Hatfield G.W. Mol. Microbiol., 2001, 39(1):191-198). Авторами настоящего изобретения была изучена возможность использования промотора гена ilvC для метаболически регулируемой экспрессионной системы. С этой целью сначала вводили ген cat за геном ilvY на хромосоме штамма E.coli MG1655 (ATCC 47076) с использованием λRed-зависимой интеграции. Фрагмент ДНК, несущий кассету λattL-cat-λattR, получали методом ПЦР (полимеразная цепная реакция) с использованием олигонуклеотидных праймеров P1 (SEQ ID NO: 11) для участка ilvY-attL и P2 (SEQ ID NO: 12) для участка attR-ilvYn плазмиды pMW118-λattL-cat-λattR (Katashkina Zh.I. et al., Mol. Biol. (Mosk.), 2005, 39(5):823-831) в качестве матрицы. Полученный фрагмент ДНК вводили в штамм E.coli MG1655/pKD46 электротрансформацией с использованием электропоратора «Bio-Rad» (США) (№165-2098, версия 2-89) в соответствии с инструкциями производителя. В результате был получен трансформант E.coli MG1655cat-ilvY, содержащий на хромосоме выше гена ilvY маркер устойчивости к хлорамфениколу (λattL-cat-λattR, CmR). Рекомбинантная плазмида pKD46 (Datsenko K.A. and Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97:6640-6645) с репликоном, обладающим температурной чувствительностью, была использована в качестве донора генов фага λ, ответственных за ARed-зависимую рекомбинационную систему. Штамм E.coli MG1655, содержащий рекомбинантную плазмиду pKD46, можно получить из E.coli Genetic Stock Center, Yale University, New Haven, USA, инвентарный номер CGSC7669. После интеграции плазмиды pKD46 в штамм E.coli MG1655 был получен штамм E.coli MG1655/pKD46.
Фрагмент cat-ilvY-Puvc, содержащий λattL-cat-λattR, ген ilvY и межгенный участок ilvY-ilvC с промотором PilvC, получали методом ПЦР с использованием олигонуклеотидных праймеров P3 (SEQ ID NO: 13) для участка attL-lacZ и праймера P4 (SEQ ID NO: 14) для участка ilvCp-lacZ и хромосомной ДНК штамма Е.coli MG1655 cat-ilvY в качестве матрицы. Полученный фрагмент ДНК вставляли в хромосому штамма E.coli MG1655/pKD46 выше гена lacZ с использованием λRed-зависимой интеграции. В результате был получен штамм Е.coli MG1655 cat-ilvY-PilvC-lacZ. Устойчивые к хлорамфениколу (CmR) колонии отбирали на чашках, содержащих лизогенную среду (lysogenic broth, LB) (Sambrook, J. and Russell, D.W. (2001) Molecular Cloning: A Laboratory Manual (3rd ed.). Cold Spring Harbor Laboratory Press), агар 1,5% и хлорамфеникол 40 мг/л.
Вставку фрагмента проверяли методом ПЦР. Для этой цели колонии, которые выращивали в течение 24 ч, проверяли на наличие фрагмента cat-ilvY-PilvC-lacZ вместо нативного гена lacZ методом ПЦР с использованием олигонуклеотидных праймеров Р13 (SEQ ID NO: 23) и Р14 (SEQ ID NO: 24). Для этого свежую изолированную колонию суспендировали в 20 мкл воды, и 1 мкл полученной суспензии использовали для ПЦР. Температурный профиль был следующим: начальная денатурация ДНК в течение 5 мин при 94°C; затем 30 циклов: денатурация в течение 30 сек при 94°C, отжиг в течение 30 сек при 53°C, элонгация в течение 3 мин при 72°C; и финальная элонгация в течение 7 мин при 72°C. Несколько анализируемых CmR-колоний должны содержать необходимый фрагмент ДНК длиной 3053 п.н., что подтверждает присутствие фрагмента cat-ilvY-PilvC-lacZ вместо нативного гена lacZ длиной 288 п.н. Один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем культивирования при 37°C. Так был получен штамм E.coli MG1655 cat-ilvY-PilvC-lacZ.
Для того чтобы получить данные об уровне экспрессии генов ilvBN и ilvBNA, кодирующих AHAS I, оценивали уровень экспрессии гена-репортера lacZ под контролем промотора PilvC при различных генетических окружениях (Таблица 2). Также анализировали штаммы, дополнительно содержащие делецию генов ilvAYC. Модификацию ΔilvAYC-KmR вводили с помощью P1-трансдукции (Miller J.H. (1972) Experiments in molecular genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor).
Все модификации комбинировали с помощью P1-трансдукции и полученные штаммы вылечивали от «вырезаемых» маркеров, как описано в EP 1942183.
Делецию ilvAYC конструировали в два этапа с использованием ARed-зависимой интеграции. Сначала получали ПЦР-фрагмент с использованием олигонуклеотидных праймеров P5 (SEQ ID NO: 15) и P6 (SEQ ID NO: 16) и плазмиды pMW118-λattL-kan-λattR (Katashkina Zh.I. (2002) Development of the methods for targeted modification of E.coli genetic loci for the construction of amino-acids-producing strains. PhD Thesis. Moscow) в качестве матрицы, содержащей маркер устойчивости к канамицину (λattL-kan-λattR, KmR). Затем полученный фрагмент вводили в E.coli MG1655/pKD46 электротрансформацией, как описано выше. Канамицин-устойчивые клоны с удаленными генами ilvAYC отбирали на содержащих канамицин чашках, как описано выше. Так был получен штамм E.coli MO1655ΔilvAYC::KmR.
Вставку фрагмента проверяли методом ПЦР. Для этой цели колонии, которые выращивали в течение 24 ч, проверяли на наличие фрагмента ΔilvAYC::KmR вместо нативных генов ilvAYC методом ПЦР с использованием праймеров P15 (SEQ ID NO: 25) и P16 (SEQ ID NO: 26). Для этой цели свежую изолированную колонию суспендировали в 20 мкл воды, и 1 мкл полученной суспензии использовали для ПЦР. Температурный профиль был следующим: начальная денатурация ДНК в течение 5 мин при 94°C; затем 30 циклов: денатурация в течение 30 сек при 94°C, отжиг в течение 30 сек при 57°C, элонгация в течение 2 мин при 72°C; и финальная элонгация в течение 7 мин при 72°C. Несколько анализируемых KmR-колоний должны содержать необходимый фрагмент ДНК длиной 1761 п.н., что подтверждает присутствие фрагмента ΔilvAYC::KmR вместо нативных генов ilvAYC длиной 2926 п.н. Один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем культивирования при 37°C. Так был получен штамм E.coli MG1655 ΔilvAYC::KmR.
Модификации cat-ilvY-PilvC-lacZ, ΔilvAYC::KmR, ΔilvGM, ΔilvIH и ΔilvBN, PL-ilvBN и PL-ilvBN4 вводили в штамм E.coli К-12 (ВКПМ В-7) с помощью P1-трансдукции. Конструкция ΔilvBN, ΔilvGM, ΔilvH и PL-ilvBN описана в EP 1942183. Штамм ВКПМ В-7 (другое название B7) может быть замещен другим подштаммом K-12, таким как K-12 MG1655, который доступен из Американской коллекции типовых культур «American Type Culture Collection, (ATCC)» (P.O. Box 1549, Manassas, VA 20108, United States of America). Полученные штаммы приведены в Таблице 2.
Для измерения активности β-галактозидазы штаммы выращивали до средне-логарифмической фазы роста на среде M9:LB (9:1, об./об.), содержащей глюкозу (0,4%, вес/об). Среда для штаммов, имеющих делецию ilvAYC, и штаммов с недостаточным уровнем AHAS дополнительно содержала Ile (25 мг/л) и Val (25 мг/л). Активность β-галактозидазы измеряли в соответствии с методом Миллера (Miller J.H. (1972) Experiments in molecular genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor). Представленные в Таблице 2 данные указывают на то, что уровень экспрессии гена-репортера lacZ под промотором Pave изменяется в пределах более чем двух порядков в зависимости от генетического окружения. Максимальный уровень экспрессии достигается в штаммах E.coli, в которых сверхэкспрессированы гены ilvBN4, кодирующие устойчивую к ингибированию L-валином AHAS I, или содержащих инактивированную изомероредуктазу IlvC, кодируемую геном ilvC.
Пример 2. Конструирование мутантного штамма E.coli B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBN4
Штамм E.coli B7 ΔilvGM ΔilvIH PL-ilvBN4 модифицировали так, чтобы штамм содержал фрагмент cat-ilvY-PilvC, полученный как описано в Примере 1. Фаговый промотор PL, расположенный перед генами ilvBN4, замещали на регуляторный участок cat-ilvY-PilvC с использованием λRed-зависимой интеграции в штамме E.coli B7 ΔilvIH ΔilvGM PL-ilvBN4 (конструирование этого штамма описано в ЕР 1942183). Для этой цели фрагмент ДНК, содержащий кассету экспрессии cat-ilvY-PilvC, фланкированную короткими участками, смежными с геном ilvB, получали методом ПЦР с использованием олигонуклеотидных праймеров P7 (SEQ ID NO: 17) и P8 (SEQ ID NO: 18) и хромосомной ДНК штамма Е.coli MG1655 cat-ilvY-PilvC-lacZ в качестве матрицы. Полученный ПЦР-фрагмент вводили в штамм E.coli B7 ΔilvGM ΔilvIH PL-ilvBN4/pKD46 электротрансформацией как описано выше. Так был получен штамм Е.coli B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBN4, в котором λ-фаговый промотор PL, расположенный выше генов оперона ilvBN4, кодирующего устойчивую к ингибированию по типу обратной связи AHAS I, заменен на самоиндуцируемый промотор PilvC.
Замену проверяли методом ПЦР. С этой целью свежую изолированную колонию суспендировали в 20 мкл воды, и 1 мкл полученной суспензии использовали для ПЦР. Температурный профиль был следующим: начальная денатурация ДНК в течение 5 мин при 94°C; затем 30 циклов: денатурация в течение 30 сек при 94°C, отжиг в течение 30 сек при 59°C, элонгация в течение 2 мин при 72°C; и финальная элонгация в течение 7 мин при 72°C. Несколько анализируемых CmR-колоний должны содержать необходимый фрагмент ДНК длиной 2865 п.н., что подтверждает присутствие фрагмента cat-ilvY-PilvC-ilvBN4 вместо исходного фрагмента PL-ilvBN4 длиной 382 п.н. Один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем культивирования при 37°C. Так был получен штамм E.coli B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBN4.
Пример 3. Свойства мутантного штамма E.coli B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBN4
Клетки выращивали до средне-логарифмической фазы роста на среде M9:LB (9:1, об./об.), содержащей глюкозу (0,4%, вес/об). Активность AHAS I измеряли в неочищенных клеточных экстрактах с добавлением или без добавления 10 мМ L-Val, как описано в Stormer F. и Umbarger Н., Biochem. Biophys. Res. Commun., 1964, 17(5):587-592. Средние результаты трех экспериментов представлены в Таблице 3. Как следует из Таблицы 3, регуляторный участок cat-ilvY-PilvC положительно влияет на уровень экспрессии генов, кодирующих AHAS I.
Пример 4. Продукция L-валина штаммом E.coli B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBN4
Модифицированный штамм E.coli B7 ΔlvGM ΔilvIH cat-ilvY-PilvC-ilvBN4 и контрольный штамм B7 ΔlvGM ΔluIH PL-ilvBN4 отдельно выращивали при 32°C в течение 18 часов в среде Luria-Bertani (также называемой лизогенная среда, как описано в Sambrook, J. and Russell, D.W. (2001) Molecular Cloning: A Laboratory Manual (3rd ed.). Cold Spring Harbor Laboratory Press). Затем 0.2 мл полученной среды вносили в 2 мл ферментационной среды в пробирках 20×200 мм и выращивали при 30°C в течение 60 часов на роторной качалке при 250 об/мин.
Состав ферментативной среды (г/л):
с добавлением среды LB: 10% (об./об.).
Ферментационную среду стерилизовали при 116°C в течение 30 мин, глюкозу и CaCO3 стерилизовали отдельно при следующих условиях: глюкозу - при 110°C в течение 30 мин, CaCO3 - при 116°C в течение 30 мин, pH поддерживали в районе 7.0 раствором KOH.
После выращивания накопленный L-валин измеряли методом тонкослойной хроматографии (ТСХ). Пластинки ТСХ (10×20 см) были покрыты слоем (0.11 мм) силикагеля Sorbfil, содержащим нефлуоресцентный индикатор («Сорбполимер», Краснодар, Российская Федерация). Образцы наносили на пластины с помощью аппликатора Camag Linomat 5. В качестве подвижной фазы использовали смесь iso-пропанол: этилацетат: 25% водный аммиак: вода (16:16:5:10, об./об.). Для визуализации использовали раствор нингидрина (2%) в ацетоне. После прокрашивания пластинки высушивали и сканировали с помощью Camag TLC Scanner 3 в режиме абсорбции с детекцией при 520 нм, данные обрабатывали с помощью программы winCATS (версия 1.4.2).
Средние результаты четырех независимых ферментации в пробирках приведены в Таблице 4. Как следует из Таблицы 4, модифицированный штамм E.coli B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBN4 вызывал большее накопление L-валина по сравнению с исходным штаммом E.coli B7 ΔilvGM ΔilvIH PL-ilvBN4.
Пример 5. Конструирование штамма E.coli B7 ΔilvGM ΔilvIH ilvYinactive-PilvC-ilvBN4
Кассета экспрессии ilvY-PilvC-ilvBN4 содержит ген ilvY, кодирующий регуляторный белок типа LysR. Таким образом, штамм, содержащий эту кассету, имеет две копии гена ilvY: одна копия содержится в нативном локусе на хромосоме, а другая копия расположена перед опероном ilvBN4. Чтобы выяснить, связан ли положительный эффект от кассеты ilvY-PilvC-ilvBN4 с дупликацией гена ilvY, ген ilvY инактивировали в кассете экспрессии ilvY-PilvC-ilvBN4. Нуклеотидную последовательность гена ilvY модифицировали таким образом, чтобы при инактивации гена ilvY не затронуть регуляторный участок генов оперона ilvBN4. Более точно, фрагмент ДНК длиной 31 нуклеотид был вставлен в ген ilvY кассеты экспрессии ilvY-PilvC-ilvBN4 для замены пятого кодона (GAT) структурной части гена ilvY «стоп» - кодоном TGA.
Инактивацию гена ilvY проводили следующим образом. Сначала получали фрагмент ПЦР, содержащий кассету λattL-cat-λattR с участками, смежными с внутренней частью гена ilvY, используя олигонуклеотидные праймеры P9 (SEQ ID NO: 19) и P10 (SEQ ID NO: 20) и плазмиду pMIV5-JS в качестве матрицы. Плазмиду pMIV5-JS конструировали, как описано в EP 1942183. Клетки E.coli MG1655/pKD46 электротрансформировали полученным фрагментом ПЦР. Устойчивые к хлорамфениколу трансформанты отбирали, как описано выше. Так был получен штамм E.coli MG1655 ilvY::cat, содержащий маркер устойчивости к хлорамфениколу (CmR) в структурной части гена ilvY. Вставку фрагмента проверяли методом ПЦР.
Колонии, выросшие в течение 24 ч, проверяли на наличие фрагмента ilvY::cat, интегрированного вместо нативного гена ilvY, с помощью ПЦР, используя олигонуклеотидные праймеры P11 (SEQ ID NO: 21) и P12 (SEQ ID NO: 22). С этой целью свежую изолированную колонию суспендировали в 20 мкл воды, и 1 мкл полученной суспензии использовали для ПЦР. Температурный профиль был следующим: начальная денатурация ДНК в течение 5 мин при 94°C; затем 30 циклов: денатурация при 94°C в течение 30 сек, отжиг при 55°C в течение 30 сек и элонгация при 72°C в течение 1 мин 30 сек; конечная элонгация в течение 7 мин при 72°C. Несколько анализируемых CmR-колоний должны содержать фрагмент ДНК длиной 1603 п.н., что подтверждает присутствие фрагмента ilvY::cat вместо нативного участка ilvY длиной 297 п.н. Один из полученных штаммов излечивали от термочувствительной плазмиды pKD46 путем культивирования при 37°C. Так был получен штамм E.coli MG1655 ilvY::cat.
Штамм E.coli B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBM (см. Пример 2} избавляли от CmR-маркера (cat) введением переходной плазмиды pMWts-λInt/Xis (Katashkina Zh.I. et al., Mol. Biol. (Mosk.), 2005, 39(5):823-831). Так получили немаркированный штамм E.coli B7 ΔilvGM ΔilvIH ilvY-PilvC-ilvBN4. Гены ilvAYC удаляли из штамма E.coli B7 ΔilvGM ΔilvIH ilvY-PilvC-ilvBN4 с помощью P1-трансдукции с использованием E.coli MG1655 ΔilvAYC::Km (см. Пример 1) в качестве донора, как описано выше. После клонирования λRed-генов на плазмиду pKD46 штамм E.coli B7 ΔilvGM ΔilvIH ilvY-PilvC-ilvBN4 ΔilvAYC::KmR электротрансформировали фрагментом ПЦР, содержащим кассету λattL-cat-λattR с участками, смежными с внутренней частью гена ilvY. Этот фрагмент был получен методом ПЦР с использованием олигонуклеотидных праймеров P11 (SEQ ID NO: 21) и Р12 (SEQ ID NO: 22) и хромосомной ДНК штамма E.coli MG1655 ilvY::cat в качестве матрицы. Так был получен штамм E.coli B7 ΔilvIH ΔilvIH ΔilvAYC::KmR ilvY::cat-PilvC-ilvBN4, который использовали в дальнейшем в качестве донорного штамма для трансдукции кассеты ilvY::cat-PilvC-ilvBN4 в штамм E.coli B7 ΔilvGM ΔilvIH ilvY-PilvC-ilvBN4. P1-Трансдукцию проводили как описано выше.
Проверку вставки кассеты ilvY::cat-PilvC-ilvBN4 проводили методом ПЦР. С этой целью колонии, выросшие в течение 24 ч, проверяли на наличие фрагмента ДНК, интегрированного вместо кассеты ilvY-PilvC-ilvBN4, с помощью ПЦР, используя праймеры P11 (SEQ ID NO: 21) и P19 (SEQ ID NO: 29). С этой целью свежую изолированную колонию суспендировали в 20 мкл воды, и 1 мкл полученной суспензии использовали для ПЦР. Температурный профиль был следующим: начальная денатурация ДНК в течение 5 мин при 94°C; затем 30 циклов: денатурация при 94°C в течение 30 сек, отжиг при 58°C в течение 30 сек и элонгация при 72°C в течение 2 мин; конечная элонгация в течение 7 мин при 72°C. Несколько анализируемых CmR-колоний должны содержать фрагмент ДНК длиной 1654 п.н., что подтверждает присутствие фрагмента ilvY::cat-PilvC-ilvBN4 вместо исходной конструкции ilvY-PilvC-ilvBN4 длиной 339 п.н. Один из полученных штаммов излечивали от термочувствительной плазмиды pKD46 путем культивирования при 37°C. Так был получен штамм E.coli B7 ΔilvGM ΔilvIH ΔilvAYC::KmR ilvY::cat-PilvC-ilvBN4. Этот штамм использовали в дальнейшем в качестве донора для трансдукции кассеты ilvY::cat-PilvC-iluBN4 в штамм E.coli B7 ΔilvGM ΔilvIH ilvY-PilvC-ilvBN4, как описано выше. Так был получен штамм E.coli B7 ΔilvGM ΔilvIH ilvY::cat-PilvC-ilvBN4, обладающий только одной копией гена ilvY, расположенной в его нативном локусе благодаря инактивации копии гена ilvY в кассете ilvY::cat-PilvC-ilvBN4. Ген cat удаляли введением переходной плазмиды pMWts-λInt/Xis. Так был получен немаркированный штамм E.coli B7 ΔilvGM ΔilvIH ilvYinactive-PilvC-ilvBN4.
Пример 6. Продукция L-валина штаммом E.coli B7 ΔilvGM ΔilvIH ilvYinactive-PilvC-ilvBN4
Модифицированный штамм E.coli B7 ΔilvGM ΔilvIH ilvYinactive-PilvC-ilvBN4 и контрольный штамм B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBN4 отдельно выращивали при 32°C в течение 18 часов в среде Luria-Bertani (также называемой лизогенная среда, как описано в Sambrook, J. and Russell, D.W. (2001) Molecular Cloning: A Laboratory Manual (3rd ed.). Cold Spring Harbor Laboratory Press). Затем 0,2 мл полученной среды вносили в 2 мл ферментационной среды в пробирках 20×200 мм и выращивали при 30°C в течение 60 часов на роторной качалке при 250 об/мин.
Состав ферментативной среды (г/л):
с добавлением среды LB: 10% (об./об.).
Ферментационную среду стерилизовали при 116°C в течение 30 мин, глюкозу и CaCO3 стерилизовали отдельно при следующих условиях: глюкозу - при 110°C в течение 30 мин, CaCO3 - при 116°C в течение 30 мин. pH поддерживали в районе 7,0 раствором КОН.
После выращивания накопленный L-валин измеряли методом тонкослойной хроматографии (ТСХ). Пластинки ТСХ (10×20 см) были покрыты слоем (0,11 мм) силикагеля Sorbfil, содержащим нефлуоресцентный индикатор («Сорбполимер», Краснодар, Российская Федерация). Образцы наносили на пластины с помощью аппликатора Camag Linomat 5. В качестве подвижной фазы использовали смесь iso-пропанол: этилацетат: 25% водный аммиак: вода (16:16:5:10, об./об.). Для визуализации использовали раствор нингидрина (2%) в ацетоне. После прокрашивания пластинки высушивали и сканировали с помощью Camag TLC Scanner 3 в режиме абсорбции с детекцией при 520 нм, данные обрабатывали с помощью программы winCATS (версия 1.4.2).
Средние результаты четырех независимых ферментации в пробирках приведены в Таблице 5. Как видно из Таблицы 5, модифицированный штамм E.coli B7 ΔilvGM ΔilvIH ilvYinactive-PilvC-ilvBN4 вызывал большее накопление L-валина по сравнению с исходным штаммом E.coli B7 ΔilvGM ΔilvIH PL-ilvBN4. Кроме того, инактивация гена ilvY в штамме E.coli B7 ΔilvGM ΔilvIH cat-ilvY-PilvC-ilvBN4 не сказывалась отрицательно на продукции L-валина штаммом.
Хотя указанное изобретение описано в деталях со ссылкой на наилучшие способы осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения. Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| МУТАНТНАЯ АЦЕТОЛАКТАТСИНТАЗА И СПОСОБ ПРОДУКЦИИ РАЗВЕТВЛЕННЫХ L-АМИНОКИСЛОТ | 2006 |
|
RU2355763C2 |
| СПОСОБ ПОЛУЧЕНИЯ НОРЛЕЙЦИНА И НОРВАЛИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ РОДА Escherichia, В КОТОРОЙ ВСЕ СИНТАЗЫ АЦЕТОГИДРОКСИКИСЛОТ ИНАКТИВИРОВАНЫ | 2004 |
|
RU2315807C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2007 |
|
RU2364628C2 |
| МУТАНТНАЯ АДЕНИЛАТЦИКЛАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ СЕМЕЙСТВА ENTEROBACTERIACEAE, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2010 |
|
RU2471868C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛЕЙЦИНА, L-ВАЛИНА, L-ИЗОЛЕЙЦИНА, АЛЬФА-КЕТОИЗОВАЛЕРАТА, АЛЬФА-КЕТО-БЕТА-МЕТИЛВАЛЕРАТА ИЛИ АЛЬФА-КЕТОИЗОКАПРОАТА С ИСПОЛЬЗОВАНИЕМ РЕКОМБИНАНТНЫХ КОРИНЕБАКТЕРИЙ, СОДЕРЖАЩИХ ОПЕРОН ILVBN, КОТОРЫЙ МОЖНО ИНДУЦИРОВАТЬ ПРОПИОНАТОМ | 2014 |
|
RU2675506C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |
| МАЛАЯ СУБЪЕДИНИЦА ИЗОЗИМА III И ИЗОЗИМ III СИНТЕТАЗЫ АЦЕТОГИДРОКСИКИСЛОТ ИЗ ESCHERICHIA COLI, ФРАГМЕНТ ДНК (ВАРИАНТЫ), ШТАММ БАКТЕРИИ ESCHERICHIA COLI - ПРОДУЦЕНТ L-ВАЛИНА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ L-ВАЛИНА | 2000 |
|
RU2209246C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ РОДА Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ОПЕРОН astCADBE | 2010 |
|
RU2482188C2 |
| МУТАНТЫ СИНТЕТАЗЫ АЦЕТООКСИКИСЛОТ, УСТОЙЧИВЫЕ К ОБРАТНОЙ СВЯЗИ | 2004 |
|
RU2325439C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2010 |
|
RU2460793C2 |


















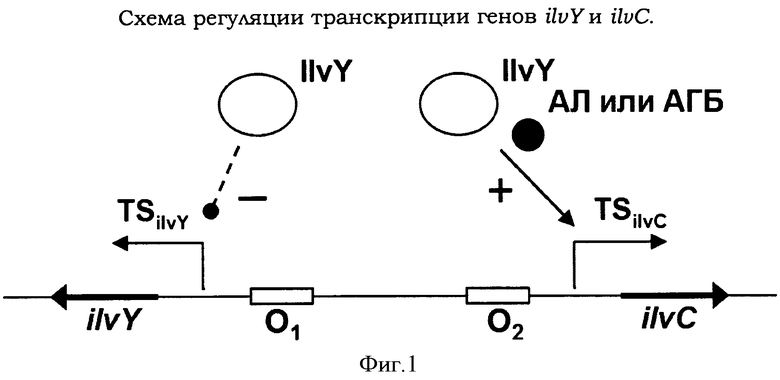
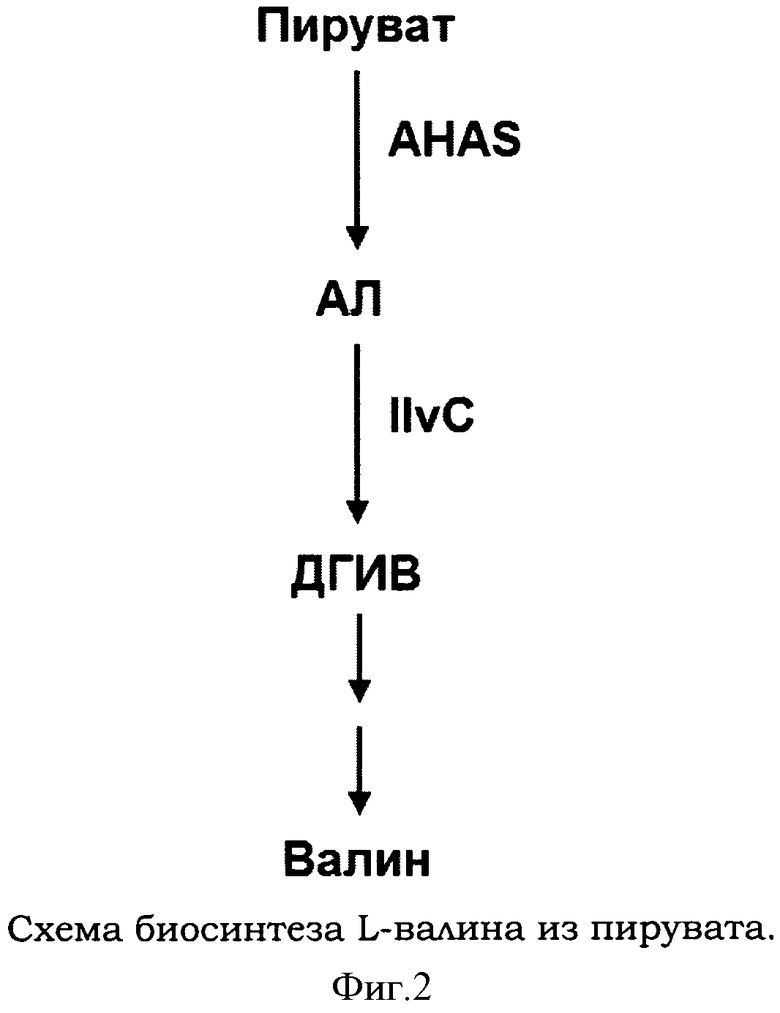
Изобретение относится к биотехнологии и предоставляет собой способ получения полезных метаболитов с использованием бактерии семейства Enterobacteriaceae, в частности бактерии, принадлежащей к роду Escherichia, которая модифицирована таким образом, что она содержит генетическую экспрессионную систему, включающую транскрипционный аппарат, регулируемый белком типа LysR, и модифицированную таким образом, что самоиндуцируемая положительная регуляция по типу обратной связи указанной системы опосредована коиндуктором. Данный способ пригоден для продукции L-аминокислот с разветвленной цепью, в частности L-валина, L-изолейцина и L-лейцина, высших спиртов и D-пантотеновой кислоты. Изобретение позволяет получать L-аминокислоты с высокой степенью эффективности. 3 н. и 20 з.п. ф-лы, 2 ил., 5 табл., 6 пр.
1. Генетическая экспрессионная система, включающая:
транскрипционный аппарат, регулируемый белком типа LysR, который включает промотор и оператор, экспрессия которых положительно регулируется регуляторным белком типа LysR и коиндуктором, и
целевой(ые) ген(ы), с которым(и) указанный транскрипционный аппарат функционально связан, причем целевой(ые) ген(ы) кодирует(ют) белок(ки), вовлеченный(е) в биосинтез указанного коиндуктора, субстрата или предшественника указанного коиндуктора,
характеризующаяся тем, что самоиндуцируемая положительная регуляция по типу обратной связи указанной экспрессионной системы опосредована коиндуктором.
2. Экспрессионная система согласно п.1, отличающаяся тем, что указанная система из бактерии, принадлежащей к семейству Enterobacteriaceae или Pseudomonadaceae.
3. Экспрессионная система согласно п.1, отличающаяся тем, что указанная система из бактерии, принадлежащей к семейству Enterobacteriaceae.
4. Экспрессионная система согласно п.3, отличающаяся тем, что указанная система из бактерии, принадлежащей к роду Escherichia.
5. Экспрессионная система согласно п.4, отличающаяся тем, что указанная бактерия принадлежит к виду Escherichia coli.
6. Экспрессионная система согласно любому из пп.1-4, отличающаяся тем, что указанная система из биосинтетического пути L-аминокислоты, выбранной из группы, состоящей из разветвленных L-аминокислот, L-лизина, L-цистеина, L-метионина и L-триптофана.
7. Экспрессионная система согласно п.6, отличающаяся тем, что указанная система из биосинтетического пути L-аминокислот с разветвленной цепью.
8. Экспрессионная система согласно любому из п.п.1-5 и 7, отличающаяся тем, что промотор есть промотор PilvC, указанный регуляторный белок типа LysR есть белок IlvY и коиндуктор есть 2-ацетомолочная кислота, 2-ацето-2-гидроксимаслянная кислота или их соли.
9. Экспрессионная система согласно п.8, отличающаяся тем, что коиндуктор есть 2-ацетомолочная кислота или ее соль.
10. Экспрессионная система согласно любому из п.п.1-5 и 7, отличающаяся тем, что целевой(ые) ген(ы) кодирует(ют) ацетогидроксикислотасинтетазу.
11. Экспрессионная система согласно п.10, отличающаяся тем, что указанные целевые гены кодируют белки, выбранные из группы, состоящей из:
(A) комбинации из п.п.A1 и A2:
(A1) белка, содержащего аминокислотную последовательность SEQ ID NO: 2, или белка, содержащего аминокислотную последовательность SEQ ID NO: 2, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком A2; и
(A2) белка, содержащего аминокислотную последовательность SEQ ID NO: 4, или белка, содержащего аминокислотную последовательность SEQ ID NO: 4, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком A1;
(B) комбинации из п.п.B1 и B2:
(B1) белка, содержащего аминокислотную последовательность SEQ ID NO: 6, или белка, содержащего аминокислотную последовательность SEQ ID NO: 6, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком B2; и
(B2) белка, содержащего аминокислотную последовательность SEQ ID NO: 8, или белка, содержащего аминокислотную последовательность SEQ ID NO: 8, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком B1; и
(C) комбинации из п.п.C1 и C2:
(C1) белка, содержащего аминокислотную последовательность SEQ ID NO: 32, или белка, содержащего аминокислотную последовательность SEQ ID NO: 32, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком C2; и
(C2) белка, включающего аминокислотную последовательность SEQ ID NO: 34, или белка, включающего аминокислотную последовательность SEQ ID NO: 34, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и обладающего активностью ацетолактатсинтазы с белком C1.
12. Экспрессионная система согласно п.11, отличающаяся тем, что указанная ацетогидроксикислотасинтетаза является мутантной ацетолактатсинтазой I, устойчивой к ингибированию по типу обратной связи L-валином.
13. Экспрессионная система согласно любому из п.п.1-5 и 7, отличающаяся тем, что указанный оператор включает участок, с которым связывается указанный регуляторный белок типа LysR.
14. Экспрессионная система согласно п.13, отличающаяся тем, что указанный регуляторный белок типа LysR выбран из группы, состоящей из:
(D) белка, содержащего аминокислотную последовательность SEQ ID NO: 10; и
(E) варианта белка, содержащего аминокислотную последовательность SEQ ID NO: 10, которая содержит замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и имеющего активность регуляторного белка типа LysR.
15. Экспрессионная система согласно любому из п.п.1-5 и 7, отличающаяся тем, что указанный промотор включает:
(F) ДНК, содержащую нуклеотидную последовательность SEQ ID NO: 30; или
(G) вариант ДНК, содержащий нуклеотидную последовательность SEQ ID NO: 30, которая содержит замену, делецию, вставку или добавление одного или нескольких нуклеотидных остатков, и имеющий активность нуклеотидной последовательности SEQ ID NO: 30.
16. Бактерия-продуцент L-аминокислоты, принадлежащая к семейству Enterobacteriaceae, отличающаяся тем, что указанная бактерия модифицирована таким образом, что она содержит экспрессионную систему согласно любому из п.п.1-15.
17. Бактерия согласно п.16, отличающаяся тем, что указанная бактерия содержит ген, кодирующий регуляторный белок типа LysR.
18. Бактерия согласно п.16, отличающаяся тем, что указанная бактерия принадлежит к роду Escherichia.
19. Бактерия согласно п.18, отличающаяся тем, что указанная бактерия принадлежит к виду Escherichia coli.
20. Бактерия согласно любому из п.п.16-19, отличающаяся тем, что указанная L-аминокислота есть L-аминокислота с разветвленной цепью и указанная бактерия модифицирована таким образом, что она содержит экспрессионную систему согласно п.7.
21. Бактерия согласно п.20, отличающаяся тем, что L-аминокислота с разветвленной цепью выбрана из группы, состоящей из L-валина, L-лейцина и L-изолейцина.
22. Способ получения L-аминокислоты с разветвленной цепью, включающий:
(i) выращивание бактерии согласно п.20 в питательной среде, так что указанная L-аминокислота с разветвленной цепью накапливается в культуральной жидкости; и
(ii) выделение указанной L-аминокислоты с разветвленной цепью из культуральной жидкости.
23. Способ получения L-аминокислоты с разветвленной цепью согласно п.22, отличающийся тем, что указанная L-аминокислота с разветвленной цепью выбрана из группы, состоящей из L-валина, L-лейцина и L-изолейцина.
| МУТАНТНАЯ АЦЕТОЛАКТАТСИНТАЗА И СПОСОБ ПРОДУКЦИИ РАЗВЕТВЛЕННЫХ L-АМИНОКИСЛОТ | 2006 |
|
RU2355763C2 |
| WO2010136897 A2, 12.02.2010 | |||
| NISTALA GJ et.al | |||
| A modular positive feedback-based gene amplifier, J Biol Eng | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |

