Область техники
Настоящее изобретение относится к антителам, в частности к мышиным моноклональным антителам, связывающимся с белком F1 из Yersinia pestis, способу получения указанных антител с использованием дрожжей, применению указанных антител для детекции Yersinia pestis и к набору для указанного применения, содержащему указанные антитела.
Предшествующий уровень техники
Антитела обычно состоят из двух тяжелых цепей, связанных между собой дисульфидными связями, и легких цепей, ассоциированных с N-концом каждой из тяжелых цепей. Каждая тяжелая цепь содержит на N-конце вариабельный домен с константным доменом на другом конце. Каждая легкая цепь также содержит на N-конце вариабельный домен с константным доменом. Вариабельные домены каждой пары легкой и тяжелой цепей образуют антигенсвязывающий участок. Вариабельные домены легкой и тяжелой цепей обладают похожей общей структурой, и каждый домен включает каркас из четырех участков, последовательности которых являются относительно консервативными, связанных посредством трех участков, определяющих комплементарность (complementarity determining regions, CDRs). Четыре каркасных участка формируют конформацию типа бета-складчатого слоя, а CDRs образуют петли, связывающие конструкцию из бета-складчатого слоя. Участки CDRs расположены в непосредственной близости друг от друга благодаря каркасным участкам и вносят вклад в образование антигенсвязывающего участка. Участки CDRs и каркасные участки антител могут быть определены путем ссылки на нумерационную систему Кабата (Kabat numbering system, Kabat et al., (1987) "Sequences of Proteins of Immunological Interest", US Dept. of Health and Human Services, US Government Printing Office) в сочетании in conjunction with x-ray crystallography, as set forth in WO 91/09967.
Для получения антитела, которое может связываться с каким-либо специфическим антигеном, обычно используют методику Kohler и Milstein (Kohler et al., (1976) Nature 256:495-497). Она включает в себя главным образом иммунизацию мыши антигеном, слияние клеток селезенки из иммунизированной мыши с клетками миеломы мыши и селекцию среди полученных таким образом гибридом одной или нескольких гибридом, которые секретируют моноклональное антитело, специфическое для целевого антигена.
Капсульный антиген F1 является основным иммунохимическим компонентом антигена, расположенного на поверхности Y. pestis. Антиген F1 присоединен к внешней мембране микроба Y. pestis в виде олигомерного белка, образующего гранулярный слой и постепенно диффундирующего в окружающую среду. Капсульный полимер состоит из множества похожих субъединиц гидрофобного белка, агрегированных при физиологических условиях. Субъединица F1 антигена изначально синтезируется из 170 аминокислот и обладает молекулярной массой 17.6 кДа, затем, после удаления сигнального пептида в ходе секреции на поверхность, формируется белок с молекулярной массой 15.6 кДа и изоэлектрической точкой 4.1 (Karlyshev et al., FEBS Lett. 1992 Feb 3; 297(1-2):77-80).
Краткое описание изобретения
Целью настоящего изобретения было получение реагента для детекции Yersinia pestis.
Указанная цель была достигнута путем выделения мышиного антитела, обладающего способностью к связыванию антигена F1 из Yersinia pestis, определения его последовательности и демонстрацией высокой специфичности указанного антитела в связывании антигена F1 из Yersinia pestis. Также указанная цель была достигнута путем создания способа получения указанного функционального антитела с использованием клеток дрожжей.
Настоящее изобретение предоставляет новое моноклональное мышиное антитело, содержащее последовательность аминокислот SEQ ID NO:1 в качестве вариабельного участка тяжелой цепи (VH) указанного антитела, последовательность аминокислот SEQ ID NO:2 в качестве вариабельного участка легкой цепи (VL) указанного антитела и консервативные участки обеих цепей, необходимые для функционирования указанного антитела. Такое новое моноклональное мышиное антитело селективно связывается с антигеном F1 из Yersinia pestis.
Также настоящее изобретение предоставляет изолированный фрагмент ДНК, кодирующий описанное выше антитело.
Также настоящее изобретение предоставляет изолированный фрагмент с участком связывания антигена (fragment antigen binding, Fab) моноклонального мышиного антитела, который селективно связывается с антигеном F1 из Yersinia pestis и включает в себя вариабельный участок тяжелой цепи (VH) с последовательностью аминокислот SEQ ID NO:1 и вариабельный участок легкой цепи (VL) с последовательностью аминокислот SEQ ID NO:2.
Также настоящее изобретение предоставляет изолированный фрагмент ДНК, кодирующий фрагмент с антигенсвязывающим участком (fragment antigen binding, Fab), описанный выше.
Также настоящее изобретение предоставляет клетку дрожжей, трансформированную описанным выше фрагментом ДНК, обладающую способностью к продукции указанного Fab.
Также настоящее изобретение предоставляет клетку дрожжей, описанную выше, где указанной клеткой дрожжей является Pichia pastoris.
Также настоящее изобретение предоставляет способ получения Fab, описанного выше, включающий выращивание указанных дрожжей в питательной среде и выделение указанного Fab из культуральной жидкости.
Также настоящее изобретение предоставляет способ детекции Yersinia pestis, включающий: (а) получение образца от животного или человека; (b) взаимодействие указанного образца с антителом или Fab, описанными выше; (с) детектирование комплекса, образованного указанным антителом или Fab и антигена F1; и, необязательно, (d) определение количества антигена F1, присутствующего в указанном образце на основании детектированного количества комплекса, образованного антителом и антигеном F1, или комплекса, образованного Fab и антигеном F1.
Также настоящее изобретение предоставляет набор для детекции антигена F1 из Yersinia pestis, содержащего изолированные моноклональные антитела, которые селективно связываются с антигеном F1 из Yersinia pestis, и средства, позволяющие детектировать указанный антиген F1 с использованием указанного антитела.
Краткое описание чертежей
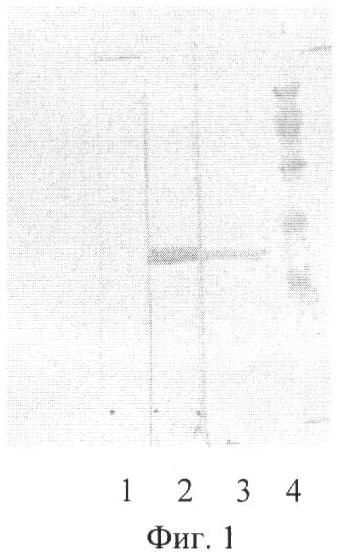
На Фиг.1 показан Вестерн-блоттинг (дорожки 1-2) и электрофорез в 12% SDS-ПААГ (восстановительные условия) антигена F1 из Y. pestis с мышиными антителами. Дорожки; 1 - конъюгат анти-IgG-POD (контроль), 2 - мышиные антитела (20 мкг/мл), 3 - антиген F1 из Y. pestis, 4 - стандарт молекулярных весов (97, 69, 45, 30, 20 кДа).
На Фиг.2 показана иммунореактивность антитела с использованием антигена F1 из Y. pestis, определенная с использованием метода ELISA.
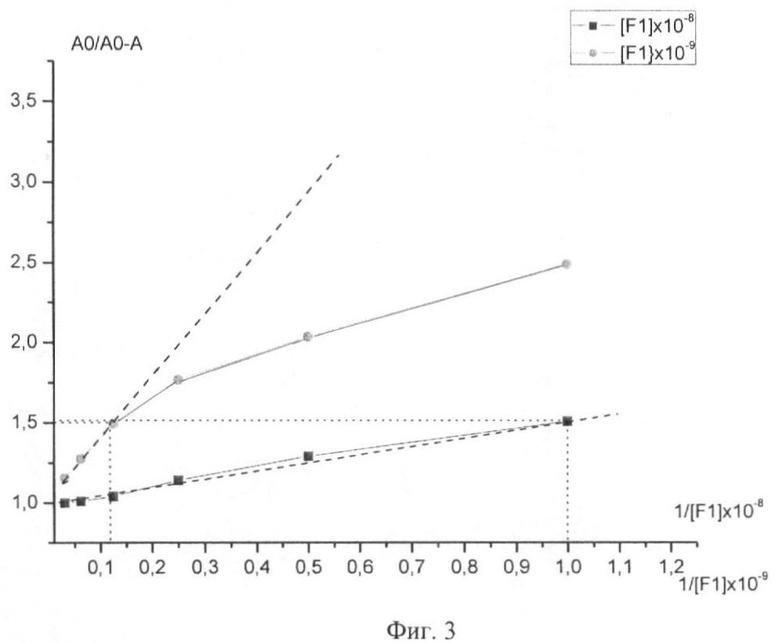
На Фиг.3 показаны калибровочные кривые для определения константы диссоциации антитела и антигена F1 из Y. pestis, полученные с использованием метода ELISA determined by ELISA method.
На Фиг.4 показаны кривые смертности мышей как в экспериментальной, так и в контрольной группах.
На Фиг.5 показаны последовательности нуклеотидов и аминокислот участка VH. 5'-Праймер, использованный для клонирования гена, кодирующего фрагмент Fab мышиного моноклонального антитела против антигена F1 из Y. pestis, указан голубым, каркасные участки показаны желтым, гипервариабельные петли (CDR) показаны зеленым, константный домен тяжелой (IgG1) цепи мышиного антитела показан красным.
На Фиг.6 показаны последовательности нуклеотидов и аминокислот участка VL. 5'-Праймер, использованный для клонирования гена, кодирующего фрагмент Fab мышиного моноклонального антитела против антигена F1 из Y. pestis, указан голубым, каркасные участки показаны желтым, гипервариабельные петли (CDR) показаны зеленым, константный домен легкой (kappa) цепи мышиного антитела показан красным.
На Фиг.7 показана конструкция плазмид pPICZαA-F1-mouse-H и pPICZαA-F1-mouse-L.
На Фиг.8 показана схематическая карта плазмиды pPICZαA-F1-mouse-H-L.
На Фиг.9 показан Вестерн-блоттинг культуральной жидкости штамма GS115/AOX-F1-mouse-H-L, продуцирующего фрагмент Fab, с анти-мышиными антителами. Дорожки с β-меркаптоэтанолом: 1 - маркеры веса; 2 - GS115/AOX-F1-mouse-H-L, 3-GS115.
На Фиг.10 показаны данные анализа белков (12% SDS-ПААГ) для 4 фракций препарата Fab после очистки методом гель-фильтрации на колонке с Sephadex G50. Дорожки: 1 - стандарт, 2-5 - различные фракции препарата антитела.
На Фиг.11 показана калибровочная кривая для Fab против антигена F1 из Y. pestis, полученная методом ELISA.
На Фиг.12 показана кривая связывания для природного (а) и рекомбинантного (b) Fab против антигена F1 из Y. pestis в координатах Клоца (Klotz coordinates).
Подробное описание настоящего изобретения
Подходящий способ для выделения антител, эффективных для использования в рамках настоящего изобретения, включает (а) назначение животному эффективного количества белка, пептида или его мимотопа с целью получения антител и (b) выделение указанных антител. Антитела, полученные против определенных белков или мимотопов, могут обладать некоторыми преимуществами, поскольку такие антитела не сильно загрязнены антителами против других соединений, что могло бы, в противном случае, повлиять на точность диагностического метода. Способы получения таких антител известны из предшествующего уровня техники и подробно описаны Harlow и др. (Antibodies, a Laboratory Manual, Cold Spring Harbor Labs Press, 1988) и включают в себя иммунизацию животных с целью получения препаратов антител, которые выделяют из, например, плазмы или асцитной жидкости и очищают способами, известными из уровня техники, с получением препаратов, дающих реакцию с антигеном. Множество видов имеют белки с близкими последовательностями и поэтому могут возникнуть сложности в ходе использования стандартных протоколов иммунизации при получении антител, которые узнают белок только одного вида. Поэтому также стоит рассмотреть модификации стандартных методов получения антител, таких как, например, разностные (вычитательные) гибридизационные методики. Такие модификации могут быть известны специалисту в данной области техники, также они описаны в настоящем описании. В других методах антитела, которые могут быть использованы в рамках настоящего изобретения, получают с использованием рекомбинантных методов, описанных Sambrook и др. (Molecular Cloning: A Laboratory Manual, (Cold Spring Harbor Labs Press, 1989)).
Вкратце, моноклональные антитела получают путем слияния клеток селезенки из иммунизированного животного и клеток миеломы с получением гибридомы. Гибридомы могут быть проверены на способность к продукции нужного антитела, затем гибридомы могут быть выращены, из них могут быть выделены указанные антитела. Термин "выращенные клетки", использованный здесь, означает гибридомы или другие линии клеток, которые производят антитела. Методы получения и проверки таких выращенных клеток описаны Harlow и др. (см. выше). Получение материала, использующегося в качестве антигена, для инъекций животных включают в себя методики, хорошо известные из уровня техники, например использование полноразмерного белка, использование пептида, выбранного из иммуногенных участков белка, модифицирование антигена такими способами, как, например, связывание с динитрофенолом, связывание с арсаниловой кислотой, денатурация антигена, связывание антигена с белком-переносчиком, таким как, например, keyhole limpet hemacyanin, с пептидами, содержащими участки связывания с рецепторами Т-клеток класса II, с бусинами, а также любыми другими методами, известными из уровня техники. Смотри Harlow и др. (см. выше).
В частности, антителами в соответствии с настоящим изобретением являются мышиные моноклональные антитела, обладающие способностью к связыванию антигена F1 из Yersinia pestis (Y. pestis). Указанные антитела являются мышиными моноклональными антителами, содержащими аминокислотную последовательность SEQ ID NO:1 в качестве вариабельного участка тяжелой цепи антитела (VH), аминокислотную последовательность SEQ ID NO:2 в качестве вариабельного участка легкой цепи антитела (VL) консервативных участков обеих цепей, необходимых для функционирования указанного антитела. Консервативные участки обеих цепей, необходимых для функционирования указанного антитела, представлены, но не ограничиваются аминокислотными последовательностями, представленными SEQ ID NO:54 и SEQ ID NO:55. Указанные новые мышиные моноклональные антитела селективно связываются с антигеном F1 из Y. pestis.
Далее, участком связывания антигена (fragment antigen binding, Fab) согласно настоящему изобретению является изолированный Fab, который селективно связывается с антигеном F1 из Yersinia pestis и включает в себя вариабельный участок тяжелой цепи (VH) с аминокислотной последовательностью SEQ ID NO:1 и вариабельный участок легкой цепи (VL) с аминокислотной последовательностью SEQ ID NO:2.
В настоящем изобретении фраза "антитело или Fab, обладающий способностью к связыванию с антигеном F1 из Yersinia pestis" означает молекулу, которая связывается с антигеном F1 и образует стабильный комплекс. Стабильным комплексом является комплекс, в котором связывание между партнерами происходит на период времени, достаточный для того, чтобы произвести детектирование указанного комплекса с использованием описанных здесь методов. Термин "селективно связывает антиген F1 из Yersinia pestis" означает способность указанной молекулы предпочтительно связываться с антигеном F1 в отличие от связывания с белками, не имеющими отношения к антигену F1, или связывания с небелковыми компонентами, присутствующими в образце. Антителом или Fab, которое предпочтительно связывается с антигеном F1, является антитело или Fab, которое связывается с антигеном F1, но не связывается в существенной степени с другими молекулами или компонентами, которые могут присутствовать в образце. Существенное связывание предполагает, например, связывание антитела, связывающегося с антигеном F1, с молекулой, не являющейся антигеном F1, с аффиностью или силой, достаточной для того, чтобы помешать способности антитела, связывающегося с антигеном F1, определить уровень антигена F1 из Y. pestis в образце. Примерами таких молекул и компонентов, которые могут присутствовать в образце, являются, но не ограничиваются ими, белки, не являющиеся антигеном F1, липиды и углеводы.
Способность антитела или Fab к связыванию с антигеном может быть определена специалистом в данной области с использованием методов, включающих, но не ограничивающимися методом ELISA и равновесным диализом. Методы определения аффинности и силы связывания хорошо известны специалисту в данной области техники, подробно описаны Janeway и др. (Immunobiology: The Immune System in Health and Disease (Garland Publishing Company, 1996)).
Fab, пригодный для осуществления настоящего изобретения, - это Fab, обладающий способностью к связыванию антигена F1 из Y. pestis, когда концентрация антигена F1 составляет от около 10 нг/мл и около 10 пг/мл. В частности, пригодные в рамках настоящего изобретения антитела связывают антиген F1 из Y. pestis, когда концентрация антигена F1 составляет 10 нг/мл, около 1 нг/лм или менее, предпочтительно 100 пг/мл. Такие антитела и Fab описаны в сопутствующих Примерах.
Изолированным фрагментом ДНК, кодирующим антитело согласно настоящему изобретению, является экспрессирующийся фрагмент ДНК, содержащий промотор, сигнальную последовательность, последовательность нуклеотидов, кодирующую структурные части тяжелой и легкой цепей антитела, участок терминации транскрипции.
В частности, нуклеотидные последовательности, кодирующие структурные части тяжелой и легкой цепей антитела согласно настоящему изобретению, содержат фрагмент ДНК, кодирующий вариабельный участок тяжелой цепи (VH) (SEQ ID NO:3) и вариабельный участок легкой цепи (VL) (SEQ ID NO:4), связанный с фрагментом ДНК, кодирующим консервативные участки обеих цепей, необходимые для функционирования указанного антитела. Нуклеотидные последовательности, кодирующие консервативные участки обеих цепей, необходимые для функционирования указанного антитела, представлены, но не ограничиваются последовательностями, представленными в SEQ ID NO:56 и SEQ ID NO:57.
Также, нуклеотидные последовательности, кодирующие Fab согласно настоящему изобретению, содержат фрагмент ДНК, кодирующий вариабельный участок тяжелой цепи (VH) (SEQ ID NO:3) и вариабельный участок легкой цепи (VL) (SEQ ID NO:4).
Ввиду вырожденности трансляционного кода могут быть различия в последовательности ДНК. Фрагменты ДНК согласно настоящему изобретению не ограничены фрагментами, показанными в SEQ ID NO:3 или 4 при условии, что они кодируют участки цепей антитела с аминокислотной последовательностью, показанной в SEQ ID NO:1 или 2.
Клетками дрожжей согласно настоящему изобретению являются клетки дрожжей, трансформированные фрагментом ДНК, описанным выше, и обладающие способностью к продукции Fab против антигена F1 из Y. pestis.
Фраза «клетки дрожжей, трансформированные фрагментом ДНК» означает, что желаемый фрагмент ДНК был введен в клетку дрожжей с использованием методов, известных специалисту в данной области техники. Трансформация клетки дрожжей фрагментом ДНК приводит к увеличению экспрессии фрагмента ДНК, кодирующего антитело согласно настоящему изобретению. Присутствие сигнальной последовательности α-фактора приводит к секреции произведенного антитела в культуральную жидкость. Методы трансформации клеток дрожжей включают в себя все известные методы, например модифицированная версия процедуры, описанной для S. cerevisiae (Gietz and Schiesti, 1996).
Способом согласно настоящему изобретению является способ получения Fab против антигена F1 из Y. pestis, включающий выращивание дрожжей в питательной среде и выделение полученного Fab из культуральной жидкости.
Примером клетки дрожжей, пригодных для продукции Fab согласно настоящему изобретению, являются, но не ограничивается ими, клетки дрожжей Pichia pastoris. Фраза "дрожжи Pichia pastoris" означает, что указанные дрожжи классифицируют как Pichia pastoris (Р. pastoris) в соответствии с классификацией, известной специалисту в данной области микробиологии. Примерами дрожжей Р. pastoris, применимых в рамках настоящего изобретения, являются, но не ограничиваются ими, дрожжи Р. pastoris GS115 (Invitrogen).
В настоящем изобретении выращивание, накопление и очистка Fab из культуральной жидкости и других жидкостей может быть осуществлена методом, сходным с традиционными методами ферментации, когда некий белок производится с использованием микроорганизма.
Питательная среда для выращивания может быть как синтетической, так и натуральной при условии, что указанная среда содержит источник углерода, источник азота, минералы и, если это необходимо, подходящее количество питательных веществ, в которых нуждаются дрожжи для их роста. К источникам углерода относятся углеводы, такие как глюкоза и сахароза, и различные органические кислоты. В зависимости от способа ассимиляции у указанного микроорганизма может быть использован спирт, включая метанол, этанол, глицерин. В качестве источника азота могут быть использованы различные соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, источники природного азота, такие как пептон, гидролизат соевых бобов, расщепленные ферментированные микроорганизмы. В качестве источника минералов могут быть использованы монофосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут быть использованы тиамин, дрожжевой экстракт и подобные им соединения.
Выращивание предпочтительно осуществляют в аэробных условиях, таких как культивирование с перемешиванием, взбалтывание с аэрацией, при температуре от 20 до 40°С, предпочтительно от 28 до 30°С. рН среды обычно поддерживают в диапазоне от 2 до 9, предпочтительно в диапазоне от 6 до 7.5. рН среды может быть скорректирован с помощью аммиака, карбоната кальция, различных оснований и буферов. Обычно выращивание в течение от 1 до 7 дней приводит к накоплению Fab в культуральной жидкости.
После выращивания твердые компоненты, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрации с использованием мембраны, а затем антитела или Fab могут быть выделены и очищены методом осаждения с солями, с использованием сульфата натрия или сульфата аммония, аффинной хроматографии, ионообменной хроматографии и т.п.
Моноклональные мышиные антитела в соответствии с настоящим изобретением, которые селективно связываются с антигеном F1 из Y. pestis, также могут быть получены с использованием подходящей гибридомы. Гибридомы могут быть выращены в питательной среде и накопленные антитела могут быть выделены. Здесь и далее термин "выращиваемые клетки" относится к гибридомам или другим линиям клеток, которые производят антитела. Способы получения и проверки таких выращиваемых клеток описаны Harlow и др. (см. выше). Методы получения антигенного материала для инъекций в животное включают в себя методы, известные из уровня техники, например использование полноразмерного белка, пептидов, выбранных из иммуногенного участка этого белка, модифицирование антигена такими способами, как, например, связывание с динитрофенолом, связывание с арсаниловой кислотой, денатурация антигена, связывание антигена с белком-переносчиком, таким как, например, keyhole limpet hemacyanin, с пептидами, содержащими участки связывания с рецепторами Т-клеток класса II, с бусинами, а также любыми другими методам, известными из уровня техники. Смотри Harlow и др. (см. выше).
Другим способом согласно настоящему изобретению является способ детекции Yersinia pestis. Указанный способ включает в себя: (а) получение образца из животного или человека; (b) взаимодействие указанного образца с антителом или Fab, описанными выше; (с) детектирование комплекса, образованного указанным антителом или Fab и антигена F1; и, необязательно, (d) определение количества антигена F1, присутствующего в указанном образце на основании детектированного количества комплекса, образованного антителом и антигеном F1, или комплекса, образованного Fab и антигеном F1. Присутствие в образце любого количества антигена F1 из Н. pestis является свидетельством наличия заболевания.
В одном из вариантов осуществления настоящего изобретения образец получают или выделяют из животного или человека для того, чтобы протестировать на наличие Y. pestis. Животное может как быть под подозрением на наличие ранней стадии заболевания или нет. В качестве образца, который может быть использован для определения наличия Y. pestis, может быть использован образец любой ткани животного. Кровь, мокрота, содержание бубонов, образцы тканей являются подходящими образцами. Предпочтительным образцом является кровь. Специалист в данной области может без труда выбрать подходящие образцы.
Хотя это не является необходимым в рамках настоящего изобретения, образец может быть подвергнут желаемой предварительной обработке. Например, образец может быть нормирован до достижения желаемой степени специфической активности. Нормирование образца с использованием подходящего метода разведения, известного из уровня техники, позволяет определить количественное содержание антигена F1 из Y. pestis независимо от начальной концентрации (например, удельного веса) в образце.
После получения образца определяют уровень антигена F1 из Y. pestis в данном образце. Термины "определяют", "определяют уровень антигена F1 из Y. pestis", "определяют количество антигена F1 из Y. pestis" и подобные им, использованные здесь, используются для того, чтобы очертить круг методов, которые могут быть использованы для определения или измерения количества антигена F1 из Y. pestis в образце. Такие методы могут дать качественные или количественные результаты. Уровень содержания антигена F1 из Y. pestis может быть определен по содержанию целого белка антигена F1 из Y. pestis protein или по содержанию фрагментов, продуктов деградации или продуктов реакции антигена F1 из Y. pestis.
Антитела, связывающиеся с антигеном F1 из Y. pestis, пригодные в способах согласно настоящему изобретению, могут включать в себя многофункциональные антитела, например бифункциональные антитела, содержащие, по крайней мере, одну функциональную часть, которая специфически связывается с антигеном F1. Такие многофункциональные антитела могут включать в себя, например, химерные молекулы, включающие в себя молекулу, которая связывается с антигеном F1, и вторую часть, которая дает возможность данной химерной молекуле связываться с субстратом или позволяет детектировать ее способом, при котором связывание с антигеном F1 не ухудшается. Примерами таких вторых частей являются, но не ограничиваются ими, фрагменты молекулы иммуноглобулина, флуоресцентный белок или фермент.
Гибриды или слитые белки антител, которые сохраняют способность связываться с антигеном F1 из Y. pestis, также могут быть использованы. В таких гибридах часть, связывающая антиген F1, может быть связана со второй частью, которая может связываться с субстратом или может быть детектирована. Примеры таких вторых частей включают в себя, но не ограничиваются ими, фрагменты молекул иммуноглобулина, эпитопы, флуоресцентные белки или ферменты.
Антитело, обладающее способностью к связыванию с антигеном F1 из Y. pestis, использованное в рамках настоящего изобретения, может содержаться в составе для детекции. Например, антитело может содержаться в буфере, в котором это антитело растворено, и/или быть объединено с носителем. Подходящие буферы и носители известны для специалиста в данной области техники. Примеры подходящих буферов включают в себя любой буфер, в котором антитело может функционировать с селективным связыванием антигена F1, такой как, но не ограничивающийся, фосфатный солевой буфер, вода, буфер HEPES (буферная соль N-2-гидроксиэтилпиперазин-N'-2-этансульфоновой кислоты), буфер TES (солевой буфер Трис-ЭДТА), буфер Трис и буфер ТАЭ (Трис-ацетат-ЭДТА). Примеры носителей включают в себя, но не ограничиваются ими, полимерные матрицы, токсоиды, сывороточные альбумины, такой как бычий сывороточный альбумин. Носители могут быть комбинированы с антителом или конъюгированы (присоединены) к антителу способом, который существенно не изменяет способность антитела к связыванию антигена F1.
Термин "взаимодействие", использованный здесь, означает введение, добавление образца, предположительно содержащего антиген F1 из Y. pestis, к антителу, связывающему антиген F1, например, путем объединения или смешения образца с антителом. В случае, если антиген F1 из Y. pestis присутствует в образце, образуется комплекс антигена F1 с антителом; образование такого комплекса означает способность антитела селективно связываться с антигеном F1 с образованием стабильного комплекса, который может быть детектирован. Детектирование может быть качественным, количественным или полуколичественным. Связывание антигена F1 из образца с антителом происходит в условиях, подходящих для формирования комплекса. Такие условия (например, подходящие концентрации, буферы, температура, время реакции), а также методы оптимизации таких условий хорошо известны специалисту в данной области техники. Связывание может быть обнаружено и измерено с использованием множества методов, являющихся стандартными в данной области техники, включая, но не ограничиваясь ими, ферментативные методы иммунохимии (например, ELISA), иммунопреципитации, методы иммуноблотинга и другие методы иммунохимии, описанные, например, Sambrook и др. (см. выше) и Harlow и др. (см. выше). Эти ссылки также описывают примеры условий, в которых образуется комплекс.
В рамках одного из воплощений настоящего изобретения комплекс антигена F1 и антитела может образовываться в растворе. В рамках другого воплощения настоящего изобретения может образовываться комплекс антигена F1 с антителом, в котором антиген F1 или антитело иммобилизовано на подложке (например, нанесено на подложку). Методики иммобилизации хорошо известны специалисту в данной области техники. Подходящие материалы для подложки включают в себя, но не ограничиваются пластмассой, стеклом, гелем, целлулоидом, тканями, бумагой и другими материалами. Примеры материалов подложки включают в себя, но не ограничиваются латексом, полистиролом, нейлоном, нитроцеллюлозой, агарозой, хлопком, PVDF (поливинилиденфторидом) и магнитными смолами. Подходящие формы материала подложки включают в себя, но не ограничиваются углублениями (например, плашки для микротитрования), чашки для микротитрования, стержни, полоски, бусины, аппараты с ответвляющимся потоком, мембраны, фильтры, трубки, чашки, целлулоидный матрикс, магнитные частицы, другие макрочастицы. В частности, предпочтительными подложками являются, например, пластинки для ELISA, стержни, полоски для иммуноанализа, радиоиммунологические чашки, агарозные бусины, пластиковые бусины, бусины из латекса, гибки, хлопковые нити, пластиковые чипы, мембраны для иммуноблотинга, бумага для иммуноблотинга и проточные мембраны. В одном из вариантов воплощения изобретения подложка, указанная выше, может содержать маркер для детекции. Описание примеров материалов для подложки смотри, например, в Kemeny, D.М. (1991) A Practical Guide to ELISA, Pergamon Press, Elmsford, N.Y. pp 33-44, and Price, С.) и Newman, D. eds. (Principles and Practice of Immunoassay, 2nd edition (1997) Stockton Press, NY, N.Y.).
Антитело может быть иммобилизовано на подложке, такой как плашка для микротитрования, стержне, полоске для иммуноанализа, аппараты с ответвляющимся потоком. Образец, полученный из животного или человека, наносят на подложку и инкубируют в условиях, подходящих (то есть, достаточных) для образования комплекса антигена F1 с антителом, связанным с подложкой.
В соответствии с настоящим изобретением комплекс антигена F1 с антителом детектируют. Термин "детектирование образования комплекса", использованный здесь, означает идентификацию присутствия антитела, связанного с антигеном F1. Если комплекс образуется, то количество образованного комплекса может быть оценено количественно, но это не является обязательным. Образование комплекса или селективное связывание между предполагаемым антигеном F1 с антителом может быть измерено (то есть, детектировано, определено) с использованием множества методов, являющихся стандартными в данной области (смотри, например, Sambrook и др., выше), примеры которых описаны здесь. Комплекс может быть детектирован множеством методов, включающих в себя, но не ограниченных использованием одного или нескольких методов: ферментоподобным иммунометодом, конкурентным ферментоподобным иммунометодом, радиоиммунным, флуоресцентным иммунометодом, хемилюминесцентным иммунометодом, методом с использованием аппаратов с ответвляющимся потоком, проточным методом, методом агглютинации, методом с использованием макрочастиц (например, с использованием частиц, таких как, но не ограниченных магнитными частицами или пластиковыми полимерами, такими как латекс или полистироловые бусины), методом иммунопреципитации, методом BioCore™ (например, с использованием коллоидного золота), методом иммунодота (например, Heska AG's Immunodot System, Фрибург, Швейцария) и методом иммуноблотинга (например, Вестерн-блотинг), методом фосфоресценции, проточным методом, методом хроматографии, методом с использованием полиакриламидного геля, методом поверхностного резонанса плазмонов, методом спектрофотометрии, методом с использованием частиц, методом электронных сенсоров. Указанные способы хорошо известны специалисту в данной области техники.
Эти методы могут быть использованы для получения количественных или качественных результатов в зависимости от того, как эти методы будут использованы. Результаты методов могут базироваться на детектировании нативной молекулы антигена F1 или ее фрагментов, продуктов их деградации или продуктов реакции антигена F1. Некоторые методы, такие как агглютинация, разделение с использованием макрочастиц и иммунопреципитация, могут дать визуальный результат (например, как наблюдение невооруженным глазом или с использованием устройств, таких как денситометр или спектрофотометр), не требующий применения маркера для детектирования.
В других методах конъюгация (то есть, присоединение) антитела с маркером для детектирования или с реагентом, который селективно связывается с антителом (например, антитело на антитело, связывающее антиген F1), помогает в детектировании образования комплекса. Такие соединения и реагенты являются примерами молекул для детектирования (то есть молекул, способных детектировать комплекс антитела с антигеном F1). Маркер для детектирования может быть присоединен к части молекулы, которая не влияет на способность этой молекулы связываться с антигеном F1. Методы конъюгации хорошо известны специалисту в данной области техники. Примеры маркеров для детектирования включают в себя, но не ограничиваются радиоактивными, флуоресцентными, хемилюминесцентными, хромофорными, ферментативными, фосфоресцентными, электронными маркерами; метками из золей металлов, цветных бусин, физическими метками или лигандами. Бусинами считаются макрочастицы, состоящие из материала в виде матрикса, такого как латекс или полистирол, который может быть ковалентно или нековалентно связан с молекулой для детекции. К лигандам относятся молекулы, которые селективно связываются с другой молекулой. Предпочтительные маркеры для детектирования включают в себя, но не ограничиваются флуоресцеином, радиоактивными изотопами, фосфатазой (например, щелочной фосфатазой), биотином, авидином, пероксидазой (например, пероксидазой хрена), β-галактозидазой, соединениями, подобными биотину или авидину (например, стрептовидин или ImmunoPure® NeutrAvidin). Например, в наиболее предпочтительном варианте осуществления изобретения антитела, связывающие антиген F1, конъюгируют с бусинами их латекса.
Настоящее изобретение также может включать в себя один или несколько слоев и/или типов вторичных молекул или других связывающих молекул, способных детектировать присутствие комплекса антитела с антигеном F1. Например, немодифицированные (то есть не связанные с молекулой для детектирования) вторичные антитела, которые селективно связываются с антителом, связывающим антиген F1, могут связываться модифицированным (то есть связанным с молекулой для детектирования) третичным антителом, которое селективно связывает вторичные антитела, что позволяет детектировать антиген F1. Подходящие вторичные антитела, третичные антитела и другие вторичные и третичные молекулы могут быть легко выбраны специалистом в данной области техники. Предпочтительные третичные молекулы также могут быть выбраны специалистом в данной области техники на основании характеристик вторичной молекулы. Подобная стратегия может быть применена для последовательных слоев.
Также могут быть использованы не иммунологические методы. Для того чтобы детектировать антиген F1, могут быть использованы методы, такие как предварительное концентрирование образца, для повышения концентрации антигена F1 с целью повышения чувствительности к белку. Такие не иммунологические методы также включают в себя, например, электрофорез, где детектирование антигена F1 может быть произведено методами, известными из уровня техники, например окрашивание белка. В другом варианте осуществления изобретения тест на наличие антигена F1, основанный на использовании белков, может быть использован для определения антигена F1 в предварительно концентрированном образце из животного или человека.
Как только уровень антигена F1 измерен, может быть сделано заключение о наличии заболевания. Заключение о наличии заболевания означает сравнение уровня антигена F1 в образце для тестирования с уровнем, обнаруженным у здорового животного или человека. В рамках настоящего изобретения заключение о наличии заболевания делается, когда детектируется любое количество антигена F1.
Настоящее изобретение также включает в себя набор, подходящий для детекции (детектирования) антигена F1 из Y. pestis, использующий способ, описанный выше. Термин «подходящий» означает, что детекция включает в себя методы, описанные здесь, использующие антитела, связывающие антиген F1, в соответствии с настоящим изобретением. Такие антитела могут быть конъюгированы с маркером для детектирования. В другом варианте осуществления изобретения набор может содержать немеченые антитела в соответствии с настоящим изобретением, также как и меченые антитела с или без маркеров для детекции, обладающие способностью к детектированию антигена F1. Набор также может содержать сопутствующие компоненты, такие как, но не ограниченные буферами, метками, контейнерами, вставками, тюбингами, флаконами, шприцами и подобными предметами.
Методы получения плазмидной ДНК, расщепления и лигирования ДНК, трансформации, подбора олигонуклеотидов в качестве праймеров и подобные методы могут быть стандартными методами, хорошо известными специалисту в данной области техники. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, Т., ("Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)).
Последующие примеры приведены для целей объяснения и не ограничивают каким-либо образом рамки настоящего изобретения.
Пример 1. Очистка антигена F1.
Выращивание.
Выращивание микроорганизма Y. pestis EB76 (штамм с живой вакциной) осуществляли в плотной питательной среде, на агаре Хоттингера (Hottinger's agar) с добавлением сульфата магния до 0.25%, хлорида кальция до 0.025%, гемолизированной крови до 0.1% при 37°С в течение 48 часов.
Очистка антигена F1.
Культуру смывали с поверхности агара натрий-фосфатным буфером рН 8.5 (PBS 8.5). Клетки осаждали центрифугирование при 5000 g в течение 15 минут. Супернатант отбирали, осадок тщательно ресуспендировали в буфере PBS рН 8.5 и дважды промывали (на этой стадии капсульный антиген смывался в раствор с поверхности клеток). Супернатанты от промывок объединяли и использовали в качестве источника антигена F1 (грубый экстракт F1). Присутствие антигена F1 в растворе контролировали путем сбора данных ELISA с использованием приготовленных самостоятельно конъюгатов кроличьих антител против антигена F1 с пероксидазой и мышиных моноклональных антител против антигена F1.
Осаждение белков из грубых экстрактов антигена F1 осуществляли с помощью сульфата аммония с 30% насыщением. После инкубирования в течение 12 часов при 4°С образовавшийся осадок центрифугировали при 10000 g в течение 20 минут, полученный осадок (осадок SA30) растворяли в буфере PBS и снова осаждали в изоэлектрической точке при рН 4.05±0.05 титрованием с использованием разбавленного раствора HCl. Полученные агрегаты отделяли центрифугированием при 10000 g в течение 20 минут, растворяли в фосфатном буфере, а затем для подготовки к хранению полученные белки диализировали против деионизированной воды и лиофилизировали.
Определение количества белков в образцах с антигеном F1
Определение количества белков осуществляли методом Лоури. Раствор альбумина 1 мг/мл использовали в качестве стандарта.
Электрофорез образцов с антигеном F1
Электрофорез капсульного белка был произведен в 12.0% SDS-ПААГ методом Лемли (Laemmly). В качестве стандарта использовали белок F1 (Государственный научно-исследовательский центр прикладной микробиологии, Оболенск, Россия). Анализ с помощью электрофореза подтвердил, что очищенный образец F1 обладал такой подвижностью в геле, как и внутренний стандарт F1. Были получены препаративные количества антигена F1 из Y. pestis.
Пример 2. Накопление и характеристика мышиного моноклонального антитела к антигену F1 из Y. pestis
Иммунизация
Мыши линии Balb/c в возрасте 8 недель были трижды с интервалом в 2 недели внутрибрюшинно иммунизированы 50 мкг антигена. Для первой иммунизации использовали смесь 1:1 антигена и полным адъювантом Фройнда (complete Freund's adjuvant) в форме гомогенной эмульсии. Антиген для второй иммунизации был приготовлен похожим образом, но только с неполным адъювантом Фройнда. Третью, окончательную иммунизацию осуществляли с использованием 50 мкг антигена в буфере PBS.
Получение гибридом
Мыши были терминированы на 4-й день после третьей иммунизации, из них было извлечена кровь и получена сыворотка, которую использовали в качестве положительного контроля. Селезенку гомогенизировали, половину полученных лимфоцитов замораживали в жидком азоте с использованием 10% ДМСО, с использованием остальных лимфоцитов получали клетки, слитые с миеломой SP2/0, в соответствии со стандартным методом с применением 50% ПЭГ 4000.
Указанные клетки высевали в плашки с 96 лунками, содержащими питательную среду DMEM (Gibco, cat.No. 52100-039) с добавлением L-глутамина, пирувата натрия, гентамицина, HAT (100 мкМ гипоксантин, 0.4 мкМ аминоптерин, 16 мкМ тимидин) и 8% эмбриональной телячьей сыворотки. Сначала вносили в лунки суспензию клеток подкормки - перинатальных макрофагов, полученных путем промывки брюшной полости здоровой мыши. Рост первичной культуры определяли визуально с использованием микроскопа. По истечении 10-12 дней после гибридизации супернатанты из лунок проверяли на содержание специфических антител с использованием ELISA.
Первичные культуры из лунок, содержащие антитела, дважды клонировали с использованием метода ограниченных разведений в питательной среде DMEM с теми же добавками, за исключением HAT, на слой клеток подкормки. Проверку супернатантов осуществляли на 10-12 день. Если требовалось, положительные клоны переклонировали еще 1-3 раза, а затем определяли их изотипы с использованием EIA.
Получение асцитов
Выбранные клоны инокулировали в мышей, в которых предварительно ввели пристан (Pristane). Асциты выделяли на 7-10 день после инокуляции клетками.
Очистка моноклональных антител против антигена F1 из Y. pestis
Асциты (20 мл) разводили в буфере MES (150 мМ хлорид натрия, 50 мМ 2-[N-морфолино]этансульфоновая кислота, рН 5.7) в соотношении 1:4 и наносили на колонку Protein G-column (Protein G-Sepharose 4B Fast Flow (Pharmacia, 17-0618-03)). Выход мышиного изотипа IgG1 составлял 3.8 мг/мл асцитов. После очистки степень чистоты антител проверяли в SDS-ПААГ и с помощью иммунореактивности (непрямой метод ELISA). Чистота антител была около 95%.
В соответствии результатами иммуноблотинга (Фиг.1) антитела F19 специфически окрашивали одну полосу в образце F1, полоса с высоким молекулярным весом соответствует димеру F1.
Проверка кросс-реактивности с антигенами F. tularensis, В. anthracis PA и SP, Т. gondii, Salmonella О Ag, Legionella pn., VAg Y. pestis, SEB, E.coli была отрицательной.
Иммунореактивность очищенных антител
Иммунореактивность антител проверяли непрямым методом ELISA с использованием антигена F1 из Y. pestis (покрытие F1 - 5 мкг/мл в 0.1 М карбонатном буфере, антитела - 1 мкг/мл, конъюгат - мышиные анти-IgG, субстрат - 3,3',5,5'- тетраметил-[1,1'-бифенил]-4,4'-диамин (ТМВ). Данные были получены на многоканальном фотометре "Multiscan" при длине волны 450 нм. Результаты показаны на Фиг.2. Указанные моноклональные антитела демонстрировали высокую специфическую активность и чувствительность до 0.5 нг/мл.
Определение константы диссоциации для антител против антигена F1 из Y. pestis Константу диссоциации определяли методом Клотца (Klotz I.M., The Proteins, Vol.1, eds H.Neurath and K.Bailey, Academic Press, New York, 1953, p.727) в модифицированном варианте B.Friguet, A.F.Chaffotte, L.Djavadi-Ohaniance и M.E. Goldberg (Journal of Immunological Methods, 77 (1985), 305-319).
Для восстановления равновесия в системе антитело-антиген плашки, которые предварительно блокировали 1% бычьим сывороточным альбумином (БСА), покрывали 50 мкл раствора антигена F1 в буфере PBS-AT в концентрациях 10-8 и 10-9 M, 3 ряда для каждой, и титровали серийными разведениями по 1/2. Затем все лунки из этих 6 рядов покрывали 50 мкл антител в выбранной концентрации. Стандартные растворы антител 1; 0,8; 0,5; 0,2; 0,1 вносили в 3 ряда той же плашки, где 1 соответствовала концентрации антител для покрытия слоем оттитрованного антигена, несколько лунок покрывали 50 мкл антитела для определения Ао (оптическая плотность без антигена). Все лунки, содержащие антитела, также были покрыты слоем 50 мкл буфера PBS-AT (0.05 М фосфатный буфер (рН 7,5) с добавлением 0,2% БСА и 0,1% Tween 20). Инкубацию проводили в течение 2 часов при комнатной температуре в мешалке. После инкубации содержимое всех лунок переносили на чашку, предварительно покрытую слоем антигена концентрации 5 мкг/мл в буфере PBS. Инкубацию проводили в течение 1 часа, чашки отмывали буфером PBS-T (0.05 М фосфатный буфер (рН 7,5) с добавлением 0,1% Tween 20) и покрывали слоем анти-мышиного IgG конъюгата. После 1 часа инкубации чашку омывали и наносили ТМВ, по истечении 20 минут получали результаты путем измерения на многоканальном фотометре "Multiscan" при 450 нм. Если стандарты антител имели линейные значения, эксперименты считались действительными. Ao рассчитывали как среднее значение во всех лунках, в которых антитела были нанесены без антигена. Результаты были представлены на Фиг.2. Константы диссоциации определяли на основе калибровочной кривой.
Таким образом, Kd для моноклональных антител к антигену F1 из Y. pestis равно 4,5×10-9 M.
Пример 3. Изучение in vivo нейтрализующей активности мышиных моноклональных антител против антигена F1 из Y. pestis по способности к протекции животных от вирулентного штамма Y. pestis F1 (+).
Изучение способности мышиных антител против антигена F1 из Y. pestis к протекции мышей от вирулентного штамма Y. pestis F1 (+) проводили in vivo. Антитела вводили в мышей внутривенно перед и после заражения. Способность антител к протекции оценивали, сравнивая LD50 (с антителами) и LD50 (без антител). Параметр "средняя продолжительность жизни" оценивали в случае минимальной разницы значений LD50.
Мыши М. musculus Balb/c (18-20 г, десять в каждой экспериментальной группе) были подкожно инокулированы штаммом Y. pestis 231(F1+) (Государственный научно-исследовательский центр прикладной микробиологии, Оболенск, Россия) дозой в 30 клеток в 0.2 мл физиологического раствора на одну мышь. Обработку антителами проводили за 2 часа перед заражением. Антитела в дозировке, равной 1 мг (в 0,2 мл физиологического раствора) на одну мышь, вводили в брюшную область. Эффективность лечения оценивали по времени выживаемости животным в течение 21 дня и по количеству погибших животных. Специфику гибели определяли путем выделения единиц образования колоний (cfu) Y. pestis из органов после препарирования. Результаты представлены в Таблице.
Выжившие животные на 21-й день после инфицирования распределились на три группы, получавшие иммунотерапию с антителами Mab анти F1 C1 F19, анти VAg C1 Va48 и Va49. В первом случае терапевтический эффект был высоким: 80% мышей выжило, в других случаях - только 20%. Необходимо отметить, что бактериологические исследования органов и крови выживших мышей на присутствие культуры Y. pestis 231 дало негативные результаты. Все погибшие животные содержали specific специфическую культуру патогена чумы. Кривые смертности мышей как в экспериментальной, так и в контрольной группах представлены на Фиг.4.
Таким образом, результаты испытаний показали, что Mab "анти F1 C1 F19" оказались высокоэффективными в протекции мышей против заражения вирулентным штаммом Y. pestis F1 (+).
Пример 4. Клонирование генов, кодирующих Fab фрагменты мышиных моноклональных антител против антигена F1 из Y. pestis, используя мРНК из подходящих линий клеток в качестве матриц.
Тотальная РНК была очищена из 1×107 клеток подходящих гибридом, содержащих антитела согласно настоящему изобретению Mab анти F1 C1 F19. Клетки (107 клеток) два раза промывали однократным буфером PBS. В пробирку со 100 мкл клеток добавляли 400 мкл раствора GTB (4 М гуанидинтиоцинат, 30 мМ цитрат натрия, 1% N-лаурилсаркозил натрия, 120 мМ β-меркалтоэтанол). Клетки суспендировали пипетированием в течение 2-3 минут и помещали на лед. Затем добавляли 50 мкл 3 М ацетата натрия (рН 5.2) и 500 мкл фенола, насыщенного водой, и полученную суспензию тщательно перемешивали на мешалке Vortex. Затем добавляли 100 мкл смеси хлороформ - изоамиловый спирт и суспензию тщательно перемешивали на мешалке Vortex в течение 1 мин с перерывами в 1 минуту. Суспензию откручивали в течение 10 мин и супернатант переносили в новую пробирку. Процедуру повторяли. Добавляли равный объем изопропанола (500 мкл) и хранили при -20°С. Образец откручивали в течение 10 минут, осадок промывали этанолом (70%) и растворяли в воде (50 мкл). Выделили в целом 7.5 мкг РНК (0.15 мкг/мкл), которые использовали для дальнейшего синтеза кДНК.
кДНК синтезировали с использованием набора RevertAid™ H Minus First Strand cDNA Synthesis (Fermentas) в соответствии с "протоколом синтеза кДНК, подходящего для ПЦР", рекомендованного производителем.
Дизайн праймеров для клонирования гена, кодирующего тяжелую цепь.
Для того чтобы идентифицировать пару праймеров, пригодных для клонирования гена, кодирующего тяжелую цепь, проводили ПЦР для клонирования с использованием ДНК полимеразы Tag (реакционная смесь содержала 10X буфер для ДНК полимеразы Tag (67 мМ Tris-HCl (pH 8.8 при 25°С), 166 мМ (NH4)2SO4, 100 мМ 2-меркаптоэтанола)+20 мМ MgCl2, 1.25 мМ dNTP, 5'-концевой праймер (25 пмоль), 3'-концевой праймер (25 пмоль), ДНК полимераза Taq (2.5 U), кДНК 1 мл, бидистиллированная вода до 50 мкл). Тотальную кДНК использовали в качестве матрицы. Программа ПЦР была следующей: 95°С в течение 3 минут; 35 циклов 94°С в течение 1 минуты, 45°С в течение 1 минуты и 72°С в течение 1 минуты; в конце 72°С в течение 10 минут.
5'-Концевой праймер соответствовал набору из 16 дегенеративных праймеров (SEQ ID NO:5-20), узнающих 95% последовательностей сигнальных пептидов в генах, кодирующих тяжелую цепь, выложенных в базе данных Кабат (Kabat, E., Wu, Т., Perry, Н. 1991. Sequences of proteins oflmmunological Interest, 5th edn. US Departament of Health and Human Services, Public Health Service, NIH). В качестве 3'-концевого праймера в ПЦР, соответствовавшего 3'-концу тяжелой цепи фрагмента Fab (IgG1), с введенным на 5'-конец сайтом XhoI использовали праймер А26 (SEQ ID NO:21). С использованием электрофореза продуктов ПЦР было показано, что использование единственной пары праймеров (SEQ ID NO:16 и 21) приводило к появлению фрагмента ПЦР. Размер полученного продукта ПЦР (≈700 пар оснований) находился в хорошем соответствии с предсказанной последовательностью кДНК гена тяжелой цепи IgG Mus musculus из баз данных Gene-Bank и IMGT-GENE-DB.
Дизайн праймеров для клонирования гена, кодирующего легкую цепь.
Ранее было показано, что изучаемое антитело содержит легкую цепь типа каппа. На основании этих данных в качестве 5'-концевого праймера использовали набор из 23 дегенеративных праймеров (SEQ ID NO:22-44). Этот набор узнает 95% последовательностей, соответствующих сигнальным пептидам в генах, кодирующих каппа-цепь, опубликованных в базе данных Кабат (Kabat, E., Wu, Т., Perry, Н. 1991. Sequences of proteins of Immunological Interest, 5th edn. US Departament of Health and Human Services, Public Health Service, NIH). В качестве 3'-концевого праймера в ПЦР, соответствовавшего 3'-концу тяжелой цепи фрагмента Fab (IgG1), с введенным на 5'-конец сайтом XhoI использовали А31 (SEQ ID NO:45). С использованием электрофореза продуктов ПЦР было показано, что использование единственной пары праймеров (SEQ ID NO:39 и 45) приводило к появлению фрагмента ПЦР. Размер полученного продукта ПЦР (≈700 пар оснований) находился в хорошем соответствии с предсказанной последовательностью кДНК гена легкой цепи IgG Mus musculus из баз данных Gene-Bank и IMGT-GENE-DB.
Клонирование генов, кодирующих Fab фрагменты антител против антигена F1 из Y. pestis
Pfu ДНК полимеразу (Fermentas) использовали для клонирования генов, кодирующих цепи антитела. Условия ПЦР оптимизировали подбором температуры отжига и концентрации Mg2+ (реакционная смесь содержала 10Х буфер для Pfu ДНК полимеразы (200 мМ Tris-HCl (pH 8.8 при 25°С), 100 мМ (NH4)2SO4, 100 мМ KCl, 1% Тритон Х-100, 1 мг/мл БСА)+20 мМ MgCl2, 1.25 мМ dNTP, 5'-концевой праймер (25 пмоль), 3'-концевой праймер (25 пмоль), Pfu ДНК полимераза (1.25 U), кДНК 1 мкл, бидистиллированная вода до 50 мкл). Каждый их продуктов двух независимых ПЦР элюировали из агарозного геля и фосфорилировали с использованием Т4 ДНК полинуклеотидкиназы. Программа для ПЦР была следующая: 95°С в течение 3 минут; 35 циклов 94°С в течение 1 минуты, 45°С в течение 1 минуты и 72°С в течение 2 минут; в конце 72°С в течение 10 минут. Полученные фрагменты клонировали в плазмидный вектор pUC18, линеаризованный рестриктазой SmaI. Рекомбинантные плазмиды, содержащие гены тяжелой и легкой цепей Fab указанного антитела, выделяли и назвали pUC18-F1-H и pUC18-F1-L, соответственно.
Пример 5. Секвенирование фрагментов генов, кодирующих вариабельные части Fab-фрагмента мышиного моноклонального антитела против антигена F1 из Y. pestis.
Рекомбинантная плазмида pUC18-F1-Н, содержащая независимый фрагмент ПЦР, кодирующий тяжелую цепь антитела против антигена F1 из Y. pestis, и плазмида pUC18-F1-L, содержащая независимый фрагмент ПЦР, кодирующий легкую цепь указанного антитела, выделяли и фрагменты ПЦР секвенировали в обоих направлениях. Использовали стандартные праймеры для секвенирования SEQ ID NO:46 и 47. В соответствии с результатами WAMPredict (https://antibody.bath.ac.uk/WAMpredict.html), канонические классы гипервариабельных петель следующие: CDR-L1 - 1, CDR-L2 - 1, CDR-L3 - 1, CDR-H1 - 1, CDR-H2 - 2, CDR-H3, Длина: 6, Статус перегиба: Изогнуты.
Пример 6. Конструирование экспрессионной системы для продукции Fab фрагмента мышиного моноклонального антитела против антигена F1 из Y. pestis и получение указанного антитела в дрожжах.
Конструирование экспрессионных кассет.
Гены, кодирующие цепи Fab фрагмента мышиного антитела против антигена F1 из Y. pestis, амплифицировали с помощью ПЦР для их введения в экспрессионные кассеты. Плазмиды pUC18-F1-L и pUC18-F1-H использовали в качестве матриц.
Для амплификации фрагмента ДНК, кодирующего вариабельный домен легкой цепи, использовали 5'-праймер Lf (SEQ ID NO:48) и 3'-праймер Lr (SEQ ID NO:50. 5'-Праймер Lf содержит на 5'-конце сайт рестрикции XhoI, необходимый для дальнейшего присоединения к сигнальной последовательности α-фактора, 3'-праймер Lr содержит на 5'-конце сайт рестрикции XbaI. Фрагмент ДНК XhoI - XbaI с легкой цепью лигировали в модифицированную плазмиду pPICZαA (Invitrogen), в которой сайт рестрикции PmeI был разрушен методом сайт-направленного мутагенеза.
Для амплификации фрагмента ДНК, кодирующего вариабельный домен тяжелой цепи, использовали 5'-праймер Hf (SEQ ID NO:51) и 3'-праймер Hr (SEQ ID NO:53). 5'-Праймер Hf содержит на 5'-конце сайт рестрикции XhoI, необходимый для дальнейшего присоединения к сигнальной последовательности α-фактора, 3'-праймер Hr содержит на 5'-конце сайт рестрикции XbaI. Фрагмент ДНК XhoI - XbaI с тяжелой цепью лигировали в оригинальную версию плазмиды pPICZαA.
Объединение генов
Объединение генов обеих цепей фрагмента Fab на одной плазмиде осуществляли путем двойного расщепления вектора с легкой цепью рестриктазами BglII и BamHI с последующим введением в уникальный сайт BamHI вектора с тяжелой цепью.
Таким образом был сконструирован экспрессионный вектор pPICZαA-F1-mouse-H-L, содержащий последовательности, кодирующие обе цепи фрагмента Fab мышиного антитела против антигена F1 из Y. pestis, каждая из которых непосредственно соединена в одной рамке считывания с сигнальной последовательность α-фактора из Saccharomyces cerevisiae под контролем промотора АОХ1 (Фиг.8).
Получение трансформантов дрожжей и анализ продукции Fab.
Экспрессионную плазмиду pPICZαA-F1-mouse-H-L, линеаризованную с помощью рестриктазы PmeI, вводили в клетки GS115 в соответствии с протоколом INVITROGEN с помощью методики, использующей L1C1. Полученные трансформанты обозначили как GS115/AOX-F1-mouse-H-L.
12 независимых трансформантов клонов GS115/AOX-F1-mouse-H-L выращивали в пробирках (V=4 мл) в течение 45 часов при 30°С и рН 6.0 с использованием питательной среды BMMY (Invitrogen) (конечная оптическая плотность OD 40). Измерение количества фрагментов Fab в культуральной жидкости было произведено методом ELISA. В соответствии с результатами этого метода отбирали штаммы, продуцирующие фрагмент Fab мышиного антитела против антигена F1 из Y. pestis. Отобранные клоны штаммов-продуцентов GS115/AOX-F1-mouse-H-L выращивали в колбах (V=100 мл) в течение 45 часов при 30°С и рН 6.0 на среде YPM.
Культуральные жидкости штаммов-продуцентов анализировали с помощью Вестерн-блотинга. Полученные результаты представлены на Фиг.9. Из этих данных специалист может сделать заключение, что некоторые штаммы-продуценты GS115/АОХ-F1-mouse-H-L секретируют в культуральную жидкость белки, которые могут быть детектированы за счет взаимодействия с анти-мышиными антителами с использованием Вестерн-блотинга. Предположительно эти белки являются отдельными цепями фрагмента Fab мышиного антитела против антигена F1 из Y. pestis.
Метод ELISA для анализа фрагментов антител в культуральной жидкости:
- Сорбция белков культуральной жидкости 100 мкл на лунку в течение 2 часов при комнатной температуре;
- Промывка лунок три раза буфером PBS;
- Блокировка 3% БСА в PBS (200 мкл на лунку) в течение 2 часов при комнатной температуре;
- Промывка лунок три раза буфером PBS с 0,05% Tween 20;
- Инкубирование с козьими анти-мышиными антителами;
- Промывка 5 раз буфером PBS с 0,05% Tween 20;
- Добавление о-фенилендиамина 100 мкл (4 мг/10 мл) с H2O2 в буфере рН 5.0;
- Остановка реакции добавлением 100 мкл 10% H2SO4.
Пример 7. Характеристика аффинности фрагментов Fab мышиных моноклональных антител против антигена F1 из Y. pestis, произведенных дрожжами.
Продукция и характеристика природного мышиного фрагмента Fab из MoAb F19. Протеолиз MoAb F19 проводили пепсином в соотношении 1:100 к тотальному IgG в присутствии 10 мМ цистеина в течение 20 часов при 37°С, в результате бы получен фрагмент Fab. Как следует из данных электрофореза, полученный препарат содержал определенное количество других продуктов гидролиза, которые затрудняли определение количества фрагмента Fab, поэтому очистку успешно осуществили методом гель-фильтрации на колонке с Sephadex G50. Анализ 4 полученных фракций приведен на Фиг.10. Все 4 фракции содержали как фрагменты Fab, так и фрагменты Fab'. Фрагменты Fab, преобладающие в четвертой фракции, выбрали для последующих экспериментов.
Разработка метода ELISA для оценки количества рекомбинантного фрагмента Fab.
Для разработки метода ELISA для детекции рекомбинантного фрагмента Fab была предпринята попытка использовать конъюгат с «хвостом» анти-His6, которая не увенчалась успехом из-за неполной доступности всех 6 гистидинов для связывания. Был приготовлен конъюгат с пероксидазой хрена и MoAb против мышиных Ig каппа-легких цепей. Для этого 0.5 мл асцитов MoAb против мышиных каппа-легких цепей, клон ЕМ-34.1 (Sigma, cat. No K-2132), очищали троекратным осаждением в сульфате натрия и конъюгировали с пероксидазой хрена. Природные мышиные фрагменты Fab MoAb F19 использовали в качестве стандарта. Результаты показаны на Фиг.11.
Определение констант диссоциации рекомбинантных и природных фрагментов Fab.
Константы диссоциации (Kd) определяли в ходе двухступенчатого метода ELISA, как описано в Примере 2. Результаты представлены на Фиг.12.
Таким образом, Kd для природного моноклонального фрагмента Fab природного мышиного антитела MoAb F19 против антигена F1 из Y. pestis составляет
Kd для рекомбинантного фрагмента Fab против антигена F1 из Y. pestis составляет
Таким образом был получен рекомбинантный фрагмент Fab, обладающий высокой специфичностью против антигена F1 из Y. pestis и высокой константой диссоциации.
Хотя настоящее изобретение было подробно описано со ссылкой на предпочтительные варианты его осуществления, для специалиста в данной области техники ясно, что могут быть сделаны различные замены и применены эквиваленты, которые не выходят за рамки настоящего изобретения. Все процитированные здесь документы являются частью настоящей заявки, включенные путем ссылки.




















| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА И Fab, СВЯЗЫВАЮЩИЕСЯ С АНТИГЕНОМ F1 ИЗ Yersinia pestis, И СПОСОБ ИХ ПОЛУЧЕНИЯ С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ | 2009 |
|
RU2420587C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ (Fab) ПРОТИВ ВИРУСА БЕШЕНСТВА, ИЗОЛИРОВАННЫЙ ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА, КЛЕТКА ДРОЖЖЕЙ, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ | 2010 |
|
RU2440412C2 |
| УЧАСТОК СВЯЗЫВАНИЯ АНТИГЕНА (Fab), В ТОМ ЧИСЛЕ ГУМАНИЗИРОВАННЫЙ Fab, ПРОТИВ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА С (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ Fab С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ, СПОСОБ И НАБОР ДЛЯ ДЕТЕКЦИИ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА С | 2016 |
|
RU2623157C1 |
| ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ЛЕГКОЙ И ТЯЖЕЛОЙ ЦЕПИ МЫШИНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ АЛЬФА (ФНО-α) ЧЕЛОВЕКА (ВАРИАНТЫ), АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab) ПРОТИВ ФНО-α ЧЕЛОВЕКА, СОДЕРЖАЩИЙ УКАЗАННЫЕ ДОМЕНЫ (ВАРИАНТЫ) | 2012 |
|
RU2499000C1 |
| Моноклональное антитело к интерферону бета-1а человека | 2018 |
|
RU2728696C2 |
| Моноклональное антитело, способное нейтрализовать биологическую активность интерферона бета-1а человека | 2018 |
|
RU2729391C2 |
| Вариабельные домены лёгкой и тяжёлой цепи мышиного моноклонального антитела против интерферона альфа (IFN-α) человека, антигенсвязывающий фрагмент (Fab) против IFN-α человека, содержащий указанные домены | 2018 |
|
RU2694672C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab), СВЯЗЫВАЮЩИЕСЯ С ИНТЕРФЕРОНОМ- γ ЧЕЛОВЕКА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КЛЕТКА, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО АНТИТЕЛА И АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2013 |
|
RU2539752C2 |
| Вариабельные домены лёгкой и тяжёлой цепи мышиного моноклонального антитела против кадгерина-17 человека, антигенсвязывающий фрагмент (Fab) против кадгерина-17 человека, содержащий указанные домены | 2018 |
|
RU2703949C1 |
| СПОСОБ ДЕТЕКЦИИ БАКТЕРИЙ РОДА Yersinia И ДИФФЕРЕНЦИАЦИИ ПАТОГЕННЫХ ДЛЯ ЧЕЛОВЕКА ВИДОВ ИЕРСИНИЙ МЕТОДОМ МУЛЬТИПЛЕКСНОЙ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2008 |
|
RU2385941C1 |
Изобретение относится к биотехнологии. Описаны изолированное мышиное антитело, селективно связывающееся с антигеном F1 из Yersinia pestis, и антигенсвязывающий фрагмент (Fab). Представлена клетка дрожжей, продуцирующая описанный антигенсвязывающий фрагмент (Fab). Предложен способ получения описанного антигенсвязывающего фрагмента. Также предложен способ детекции Yersinia pestis и набор для детекции антигена F1 из Yersinia pestis. Изобретение предоставляет реагент для детекции Yersinia pestis. 8 н. и 1 з.п. ф-лы, 12 ил., 1 табл.
1. Изолированное мышиное антитело, селективно связывающееся с антигеном F1 из Yersinia pestis, отличающееся тем, что вариабельный домен тяжелой цепи (VH) указанного антитела включает в себя последовательность аминокислот SEQ ID NO:1, а вариабельный домен легкой цепи (VL) указанного антитела включает в себя последовательность аминокислот SEQ ID NO:2.
2. Изолированный фрагмент ДНК, кодирующий антитело по п.1.
3. Антигенсвязывающий фрагмент (Fab), селективно связывающийся с антигеном F1 из Yersinia pestis, который включает в себя вариабельный домен тяжелой цепи (VH) с последовательностью аминокислот SEQ ID NO:1 и вариабельный домен легкой цепи (VL) с последовательностью аминокислот SEQ ID NO:2.
4. Изолированный фрагмент ДНК, кодирующий антигенсвязывающий фрагмент (Fab) по п.3.
5. Клетка дрожжей, трансформированная фрагментом ДНК по п.4 и обладающая способностью к продукции Fab по п.3.
6. Клетка дрожжей по п.5, отличающаяся тем, что указанными дрожжами являются дрожжи Pichia pastoris.
7. Способ получения Fab по п.3, включающий в себя выращивание клеток дрожжей по п.5 в питательной среде и выделение указанного Fab из культуральной жидкости.
8. Способ детекции Yersinia pestis, включающий в себя: (а) получение образца от животного или человека; (b) взаимодействие указанного образца с антителом по п.1 или Fab по п.3; (с) детектирование комплекса, образованного указанным антителом или Fab, и антигена F1; и, необязательно, (d) определение количества антигена F1, присутствующего в указанном образце, на основании детектированного количества комплекса, образованного антителом и антигеном F1, или комплекса, образованного Fab и антигеном F1.
9. Набор для детекции антигена F1 из Yersinia pestis, содержащий изолированные моноклональные антитела по п.1 или Fab по п.3, которые селективно связываются с антигеном F1 из Yersinia pestis, и средства, позволяющие детектировать указанный антиген F1 с использованием указанного антитела или Fab.
| РЫБАЛЬСКИЙ Н.Г., СЕРОВА М.А., ИГНАТЬЕВА Г.А., СТАРЧЕУС А.П | |||
| Моноклональные антитела и гибридомы | |||
| - М.: ВАСХНИЛ, 1989 | |||
| ШТАММ БАКТЕРИЙ YERSINIA PESTIS - ПРОДУЦЕНТ ФРАКЦИИ I ЧУМНОГО МИКРОБА | 1991 |
|
RU2031938C1 |

