Настоящее изобретение относится к клеточному способу идентификации веществ, которые способны влиять на активность молекулы-мишени. При разработке новых лекарственных средств, влияющих на молекулы-мишени, используют обычно биохимические или клеточные функциональные анализы, при помощи которых большое количество предположительных активных веществ может быть исследовано в отношении того, могут ли они оказывать влияние на исследуемую молекулу-мишень. Клеточные анализы работают обычно на основе базирующихся на росте тест-систем, при которых активность исследуемой молекулы-мишени влияет на пролиферацию клеток. Кроме того, известны тест-системы, в которых активность молекулы-мишени может быть детектирована и количественно определена посредством активности или экспрессии так называемых продуктов репортерных генов. Хотя оба вида анализов позволяют идентифицировать новые активные вещества, которые модулируют активность молекулы-мишени, также в высокопроизводительном скрининге (high throughput screen, HTS), однако, недостаток этих известных тест-систем заключается в том, что их отношение сигнал/фон, как правило, является относительно малым, так что их специфичность, в частности, при применении в HTS-формате, является очень низкой.
Ввиду указанных недостатков существующего уровня техники задача настоящего изобретения состоит в обеспечении тест-системы, которая делает возможным высокоэффективный скрининг в HTS-формате.
Эта задача решается при помощи клеточного способа идентификации веществ, которые способны влиять на активность молекулы-мишени, причем анализируемые клетки несут по меньшей мере один репортерный ген, и активность этой молекулы-мишени влияет на пролиферацию клеток, предусматривающего стадии:
а. контактирования по меньшей мере одной клетки с тестируемым веществом,
b. детекции пролиферации клеток,
с. детекции активности продукта репортерного гена.
При этом детекция пролиферации клеток и активности репортерного гена не должны обязательно иметь место в вышеуказанной последовательности.
При этом репортерный ген может быть интегрирован в геном клетки или быть стабильно или транзиторно трансфицирован в клетку. Понятие продукт репортерного гена включает в себя при этом как мРНК, так и белок. Подходящие репортерные гены и их продукты достаточно хорошо известны компетентному специалисту, и особенно пригодны здесь ферменты, такие как β-галактозидаза, β-глюкуронидаза, люцифераза, щелочная фосфатаза, кислая фосфатаза или флуоресцирующие белки, такие как зеленый флуоресцентный белок GFP, BFP, экворин и т.п. Подходящие промоторы для репортерных генов зависят при этом от типа и задачи конкретной тест-системы, а также от используемого вида клеток. Репортерные гены находятся предпочтительно под контролем промоторов, которые регулируются посредством пути трансдукции (передачи) сигнала, с которым молекула-мишень непосредственно или опосредованно связана. Предпочтительными являются репортерные гены, продукты которых являются ферментами, активность которых может детектироваться посредством превращения добавляемого извне субстрата.
Молекулы-мишени в рамках настоящего изобретения могут оказывать влияние прямо или опосредованно на пролиферацию используемой клетки. При этом молекула-мишень может сама оказывать влияние на пролиферацию используемой для этого способа клетки (таким образом, непосредственно активировать или ингибировать ее в активном состоянии). Например, молекула-мишень может быть конститутивно активной и ингибироваться тестируемым веществом или существовать в неактивном состоянии и активироваться тестируемым веществом. В соответствии с предпочтительным вариантом осуществления молекула-мишень оказывает влияние на пролиферацию используемой для этого способа клетки в активном состоянии, однако только посредством промежуточного включения непосредственно влияющей на пролиферацию молекулы (например, FUS1-HIS3 в клетках дрожжей, см. ниже). В принципе, в качестве молекулы-мишени рассматриваются все разновидности внеклеточных, мембраносвязанных или внутриклеточных биологических молекул, особенно предпочтительными являются биомолекулы человека, в частности, белки или нуклеиновые кислоты, среди них, в особенности, члены каскадов трансдукции сигналов клеточного деления, в частности, связанные с G-белком рецепторы (GPCR), протеинкиназы, протеинфосфатазы и т.д.
Влияние анализируемого вещества на молекулу-мишень может при этом влиять на ее активность стимулирующим или ингибирующим образом, например, посредством взаимодействия с самой молекулой-мишенью или влияния на молекулы, которые, в свою очередь, влияют на активность или экспрессию молекулы-мишени.
Детекция пролиферации клеток и активности продукта репортерного гена может при этом происходить чисто качественно, но также и количественно, и специалисту, компетентному в данной области, хорошо известны различные виды детекции (например, измерение плотности клеток непосредственно или опосредованно определением помутнения жидких культур в случае суспензионных клеток, колориметрические или люминометрические определения активности продукта репортерного гена и т.д.).
В соответствии с предпочтительным вариантом осуществления активность молекулы-мишени влияет на эту активность, предпочтительно на экспрессию продукта репортерного гена. При этом молекула-мишень может непосредственно (молекула-мишень сама влияет на активность/экспрессию продукта репортерного гена) или опосредованно (молекула-мишень влияет на активность или экспрессию продукта репортерного гена через активируемый молекулой-мишенью каскад клеточного обмена веществ или каскад сигналов) влиять на активность или экспрессию продукта репортерного гена.
Предпочтительно, молекула-мишень является гетерологичной молекулой (т.е. молекулой, не присутствующей или не экспрессирующейся природно в клетках, используемых в способе согласно изобретению), особенно предпочтительно она является олигонуклеотидом, полинуклеотидом, нуклеиновой кислотой, полипептидом, белком или фрагментом белка. При этом эта гетерологичная молекула может интегрироваться в геном клетки или может быть стабильно или транзиторно трансфицирована в эту клетку; экспрессия молекулы-мишени может быть конститутивной или индуцируемой.
Согласно предпочтительному варианту осуществления изобретения эта гетерологичная молекула-мишень производит свое действие на пролиферацию используемой в этом способе клетки посредством взаимодействия с химерной молекулой. Особенно предпочтительным является здесь способ, при котором гетерологичная молекула-мишень является молекулой человека, которая стабильно интегрирована в геном клетки не человека, в частности, дрожжевой клетки, и влияет на пролиферацию клеток через химерную молекулу, которая способна взаимодействовать с гетерологичной молекулой и интегрироваться в собственные каскады трансдукции сигналов или каскады обмена веществ дрожжевых клеток. При этом особенно предпочтительно, если эта химерная молекула является рекомбинантным белком, полипептидом или фрагментом белка, аминокислотная последовательность которого содержит части человеческого и дрожжевого происхождения. В частности, пригодной в рамках настоящего изобретения является комбинация человеческого связанного с G-белком рецептора (GPCR) в качестве гетерологичной молекулы-мишени с химерной субъединицей G-белка («трансплантат», см. ниже), причем в принципе каждая из субъединиц может существовать в химерной форме.
При использовании продукта репортерного гена, активность которого определяют по превращению субстрата, целесообразно добавлять этот субстрат со смещением во времени после добавления анализируемого вещества. При этом между добавлением подлежащего анализу вещества и добавлением субстрата существует, предпочтительно, интервал времени с продолжительностью прохождения по меньшей мере одного клеточного цикла используемой для этого способа клетки, особенно предпочтительно интервал 2-24 прохождений клеточного цикла. При использовании дрожжевых клеток этот интервал времени составляет предпочтительно приблизительно 4-48 часов, более предпочтительно 20-30 часов и особенно предпочтительно 24 часа.
Детекцию активности репортерного гена производят предпочтительно при разрушении клетки, особенно предпочтительно посредством добавления делающего клеточную стенку проницаемой или разрушающего клеточную стенку вещества (предпочтительно, детергента или комбинации двух или более детергентов; для этого особенно пригодны дигитонин, Тритон Х-100, Нонидет Р-40, Твин 20, CHAPS или ДСН). Особенно предпочтительными являются дигитонин в области концентраций 10-600, предпочтительно 20-400 и особенно предпочтительно 40-60 мкг/мл и Тритон Х-100 в области концентраций 0,005-0,4, предпочтительно 0,01-0,2 об.%, в каждом случае в расчете на конечную концентрацию. Предпочтительно, детергенты добавляют к исходной реакционной смеси в буферных растворах; особенно подходящие буферные условия достаточно хорошо известны компетентному специалисту (физиологические буферы, нейтральный рН, изотоническая концентрация соли и т.д.).
Для способа согласно изобретению могут использоваться самые разнообразные разновидности клеток; так, пригодны в принципе как прокариотические, так и эукариотические клетки, растительные клетки или животные клетки. Однако предпочтительными являются эукариотические клетки, особенно предпочтительно - клетки млекопитающих или клетки дрожжей, в частности штаммы Saccharomyces cerevisiae.
В соответствии со следующим предпочтительным вариантом осуществления изобретения используют в одной исходной реакционной смеси или в одном проведении опыта различные клетки и одновременно проводят скрининг, причем эти клетки отличаются друг от друга типом молекулы-мишени (так называемый «мультиплексный способ»).
В соответствии с особенно предпочтительным вариантом осуществления изобретения, настоящее изобретение относится к широко применяемому способу идентификации веществ, которые действуют в качестве лигандов для клонируемых G-белок-связанных рецепторов. При этом способ согласно изобретению является настолько чувствительным и надежным, что могут испытываться одновременно несколько GPCR в высокопроизводительном анализе в мультиплексном формате.
Настоящее изобретение будет объясняться далее с использованием примеров осуществления изобретения и фигур.
Примеры
Пример 1:
Скрининг веществ, которые влияют на активность связанных с G-белком рецепторов
Один из важнейших классов молекул-мишеней для промышленности лекарственных средств представляют связанные с G-белком рецепторы. Многочисленные представители этого семейства белков были постепенно клонированы и фармакологически охарактеризованы в прошлом. Поскольку геном человека теперь секвенирован полностью, в последнее время было идентифицировано большое количество GPCR на уровне их последовательности. Главная цель фармацевтической промышленности состоит в том, чтобы идентифицировать лиганды для этих рецепторов посредством скрининга обширных банков веществ.
К сожалению, в случае доступных в настоящее время методов и способов имеется существенное препятствие при поиске веществ, заключающееся в трудоемкости и расходах, необходимых в настоящее время для проведения поиска в этих банках на такие многочисленные молекулы-мишени. В Европейском патенте ЕР 0708922 В1 утверждается, что можно подвергать скринингу несколько GPCR одновременно в культуре клеток. Описанные в этом патенте клетки млекопитающих, которые сверхэкспрессируют GPCR, отвечают реакцией усиленного роста, если их приводят в контакт с активирующим рецептор веществом. Поскольку клетки, которые не экспрессируют активируемые этим веществом рецепторы, несмотря на это, продолжают расти далее, чувствительность этой тест-системы является не очень высокой. Кроме того, этот способ занимает много времени, так как периоды инкубации являются очень длительными и дорогостоящими, так как речь идет в этом случае о системе клеток млекопитающих.
Возможность скрининга на GPCR с меньшими затратами предоставляет использование тест-системы на основе дрожжей. Так как время является крайне важным фактором в фармацевтическом исследовании, цель настоящего изобретения состояла в том, чтобы разработать дрожжевую систему, которая делает возможным одновременный скрининг многочисленных GPCR. Для того чтобы сделать возможным высокопроизводительный скрининг, этот способ должен быть просто манипулируемым и обнаруживать по возможности большое окно измерения.
Связанные с G-белком рецепторы (GPCR) играют важную роль при многих физиологических процессах. Они представляют одно из важнейших известных до сих пор семейств белков, и предполагается, что в геноме человека приблизительно 1000 генов кодируют этот класс рецепторов. GPCR имеют характерную структуру. Речь идет об интегральных мембранных белках, которые семикратно закручены в форме  -спиралей через фосфолипидный двойной слой клеточной мембраны, причем они расположены круговым образом. Согласно оценке, приблизительно 50% имеющихся в распоряжении в настоящее время прописываемых лекарственных средств связываются с GPCR. Это подчеркивает важную роль этого класса рецепторов для фармацевтической промышленности. На основании размера и значения этого семейства белков и ввиду того факта, что для многих GPCR еще не известны химические партнеры связывания (orphan-GPCR), следует исходить из того, что этот класс рецепторов в будущем станет одним из важнейших резервуаров для подходящих белков-мишеней при поиске новых лекарственных веществ.
-спиралей через фосфолипидный двойной слой клеточной мембраны, причем они расположены круговым образом. Согласно оценке, приблизительно 50% имеющихся в распоряжении в настоящее время прописываемых лекарственных средств связываются с GPCR. Это подчеркивает важную роль этого класса рецепторов для фармацевтической промышленности. На основании размера и значения этого семейства белков и ввиду того факта, что для многих GPCR еще не известны химические партнеры связывания (orphan-GPCR), следует исходить из того, что этот класс рецепторов в будущем станет одним из важнейших резервуаров для подходящих белков-мишеней при поиске новых лекарственных веществ.
Все связанные с G-белком рецепторы функционируют согласно общему основному принципу: связывание внеклеточного лиганда приводит к конформационному изменению рецепторного белка, так что он может вступать в контакт со связывающим гуаниновый нуклеотид белком (G-белком). Расположенные на цитоплазматической стороне плазматической мембраны G-белки являются посредниками внеклеточных сигналов во внутреннем пространстве клетки. Они приводят в действие в зависимости от специфичности рецептора различные пути трансдукции сигналов, каждый из которых имеет следствием образование молекул-мессенджеров (вторичных мессенджеров), таких как цАМФ, цГМФ, Са2+ или другие молекулы, которые посредством активации или дезактивации внутриклеточных белков приводят в действие реакции в клетке.
Гетеротримерные G-белки состоят из трех субъединиц α, β и γ. G-субъединица связана в ГДФ гетеротримера G-белка. Взаимодействие с одним активируемым одним лигандом рецептором приводит к обмену ГДФ на ГТФ. Возникающие в результате конформационные изменения приводят к тому, что G-белок-тример диссоциируется на α-субъединицу и βγ-комплекс. Как активированная α-субъединица, так и βγ-комплекс могут влиять на внутриклеточные эффекторные белки. -субъединицы могут быть подразделены на четыре различных класса: Gαs, Gαi, Gαq и Gα12.
GPCR классифицируют в соответствии с участвующими в трансдукции сигналов G-белками. То есть GPCR Gs-семейства опосредуют через активацию посредством Gαs стимуляции аденилатциклазы и тем самым повышают внутриклеточную концентрацию цАМФ. GPCR Gi-семейства опосредуют через активацию Gαi ингибирование аденилатциклазы и понижают таким образом концентрацию внутриклеточного цАМФ. GPCR Gq-семейства опосредуют через активацию Gαq стимуляцию различных PLCβ-изоформ и приводят к гидролизу мембраносвязанного фосфатидилинозит-4,5-бифосфата до диацилглицерина и инозиттрифосфата (IP3). IP3 высвобождает внутриклеточные накопители Са2+. Gα12 взаимодействует с rho-специфическими факторами обмена гуанинового нуклеотида.
Сигнал держится до тех пор, пока Gα-субъединица, которая обладает ГТФ-азной активностью, не гидролизует связанный ГТФ. Члены семейства RGS-белков (регуляторов передачи сигнала G-белка) регулируют временную длительность сигнала, действуя в качестве активаторов на ГТФ-азную активность Gα-субъединицы. Эта регулируемая G-белками система трансдукции сигнала является, по-видимому, общей для всех эукариотических систем.
Очень хорошо охарактеризованный пример такой сигнальной системы представляет так называемый "путь ответной реакции на феромоны" пекарных дрожжей Saccharomyces cerevisiae. Дрожжевые клетки с типом спаривания МАТа экспрессируют рецептор, который кодируется геном STE2. Этот рецептор активируется связыванием α-фактора, пептидного феромона, который высвобождается дрожжевыми клетками другого типа спаривания (МАТα). Гетеротримерный G-белок дрожжей состоит из продуктов гена GPA1 (Gα), STE4 (Gβ) и STE18 (Gγ). Gβγ-комплекс высвобождается после активации рецептора Ste2p и переносит сигнал на активируемый митогеном каскад протеинкиназы. Он приводит к активации циклин-зависимого ингибитора киназы Far1p, что приводит к остановке клеточного цикла и транскипционной индукции ряда генов, которые участвуют в процессе спаривания (например, FUS1). Этот путь десенсибилизируется Sst2p, членом RGS-семейства. Дрожжевые клетки другого типа спаривания (МАТα) экспрессируют другой рецептор (Ste3p) и поэтому отвечают на другой феромон (а-фактор), который высвобождается МАТа-клетками. Несмотря на это сигнальный аппарат, который используют оба типа спаривания, является одним и тем же.
Было многократно показано, что GPCR млекопитающих могут связываться с G-белок-сигнальной системой дрожжей. Несколько рецепторов, в том числе соматостатин 2-рецепторы крысы (Price et al., Mol Cell Biol 15, 6188-6195 (1995)) и аденозин А2а-рецепторы крысы (Price et al., Molecular Pharmacology 50, 829-837 (1996)), могут непосредственно взаимодействовать с дрожжевым Gα-белком Gpa1p, тогда как другие рецепторы, в том числе рецепторы гормона, высвобождающего гормон роста (GHRHR) (Kajkowski et al., J Recept Signal Transduct Res 17, 293-303 (1997)), являются несовместимыми с Gpa1p. Для того чтобы, несмотря на это, осуществить связывание этих рецепторов, Gα-субъединица дрожжей может быть делетирована, и вместо нее будет экспрессироваться гетерологичный рецептор с полноразмерной Gα-субъединицей млекопитающего. Кроме того, альтернативно применяют гибридные Gα-субъединицы, у которых С-концевой домен (приблизительно одна треть пептидной последовательности) Gpa1p был заменен соответствующим районом Gα-субъединицы млекопитающего. См. WO 95/21925 в отношении обеих исходных реакционных смесей. Гибриды или другие модифицированные или гетерологичные субъединицы должны удовлетворять нескольким критериям, чтобы они могли быть связанными с системой трансдукции сигнала дрожжей. Самым важным является то, что они, с одной стороны, могут эффективно связывать Gβγ, чтобы таким образом сделать возможным ингибирование сигнала в отсутствие активированных GPCR, и, с другой стороны, могут эффективно связываться с активируемым агонистом рецептором, чтобы сделать возможной дальнейшую передачу сигнала. Conklin et al., Nature 363, 274-276 (1993) описали впервые гибрид, у которого С-концевые 5 аминокислот Gαq были заменены соответствующей последовательностью Gαi (Gαqi5), что делает возможным отщепление обычным образом Gαi-связанных рецепторов и путь трансдукции сигнала Gαq. В WO 99/14344 и в статье Brown et al., Yeast 16, 11-22 (2000) описано, что подобная реакционная смесь функционирует также в дрожжах. В этом случае С-концевые 5 аминокислот Gpa1p были заменены соответствующими аминокислотами всех Gα-белков человека. Применение этих названных «трансплантатами» гибридов делает возможными связывание многочисленных GPCR млекопитающих и путь спаривания дрожжей.
Используемые здесь штаммы дрожжей несут делеции в генах SST2, FAR1 и, в зависимости от типа спаривания клеток, STE2 или STE3. SST2, член семейства RGS-белков, делетируют для ингибирования понижающей регуляции сигнала. Делеция FAR1 делает возможным продолжение клеточного роста также и при условиях, при которых является включенным путь ответной реакции на феромоны. STE2 или STE3 выключаются для ингибирования нежелательной конкуренции за гетеротримерный G-белок. Ген GPA1 заменяли в геноме дрожжей описанными выше трансплантатами. Посредством экспрессии трансплантатов под контролем GPA1-промотора в природном генном локусе гарантируется то, что остается сохраненной стехиометрия гетеротримерного G-белка.
Модификация действия по меньшей мере одного зависимого от одного GPCR пути трансдукции сигнала биологического организма может происходить ингибирующим или стимулирующим образом. Ингибирующее действие химического соединения имеет место в том случае, если зависимый от пути трансдукции сигнала измеряемый сигнал в присутствии химического соединения оказывается более слабым, чем в отсутствие этого соединения. Соединения, которые будут вызывать подобное действие, называют также антагонистами. С другой стороны, стимулирующее действие химического соединения имеет место в том случае, если зависимый от пути трансдукции сигнала измеряемый сигнал в присутствии химического соединения оказывается более сильным, чем в отсутствие этого соединения. Подобные соединения называют также агонистами.
Для функциональных анализов в Saccharomyces cerevisiae описано использование промоторов генов FUS1, FUS2 (Cismowski et al., Nat Biotechnol 17, 878-883 (1999); Frederickson, Nat Biotechnol 17, 852-853 (1999)) и YNL279w (WO 02/40660). Эти гены экспрессируются в повышенной степени в ответ на стимуляцию путей ответной реакции на феромоны фактором спаривания. Если промоторный элемент одного из этих генов функционально связан со структурным геном, то экспрессия этого структурного гена (также называемого репортерным геном) регулируется через описанный путь трансдукции сигнала дрожжей. Такие репортерные гены являются обычно эндогенными маркерами роста, такими как HIS3 или другие ауксотрофные маркерные гены (например, URA3, LEU2, ADE2, LYS1 или TRP1), которые при стимулированном пути передачи сигнала делают возможным рост клеток в соответствующим образом истощенной среде, или гены, которые придают резистентность или чувствительность к определенным веществам (например, CYH2 или G418R). Однако могут использоваться также репортерные гены, которые кодируют внутриклеточные ферменты, такие как β-галактозидаза (LacZ), или «зеленый флуоресцентный белок» (GFP) или секретируемые ферменты, такие как фосфатазы (РНО5). При использовании в качестве репортера CAN1 клетки растут в канаванинсодержащей среде. В присутствии активаторов (агонистов) одного из гетерологично экспрессируемых GPCR будет экспрессироваться ген CAN1, так что клетки уже не будут расти в канаванинсодержащей среде. При добавлении ингибитора (антагониста) культуры растут в этой селективной среде.
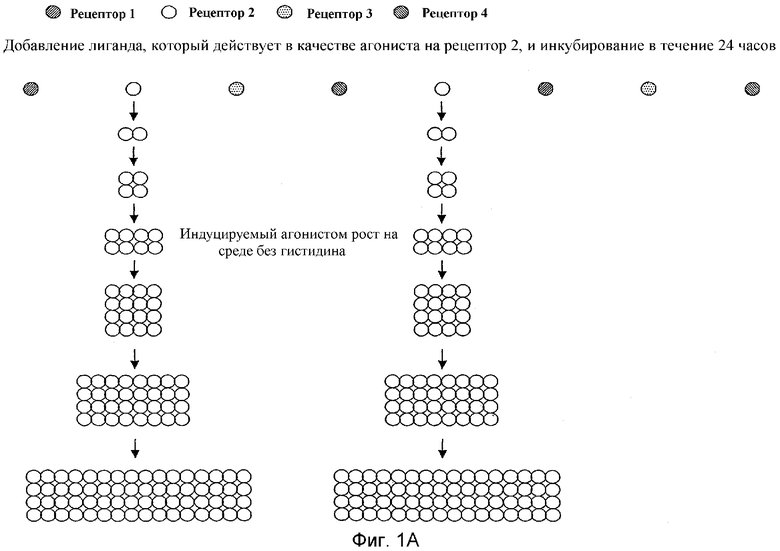
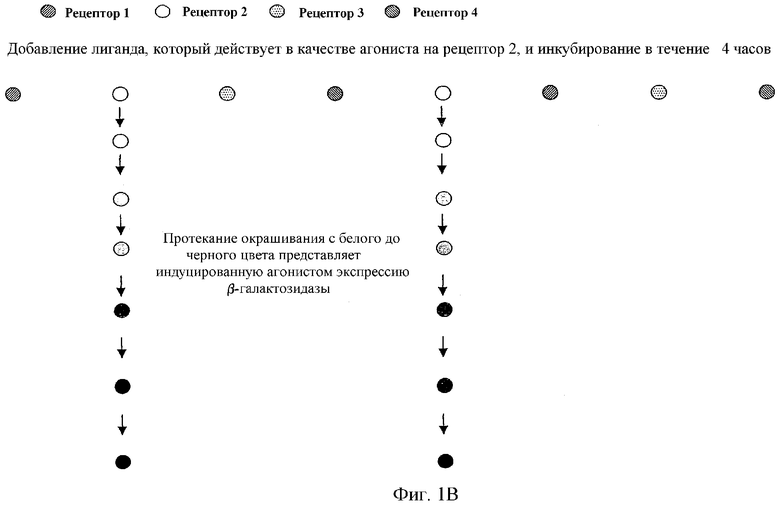
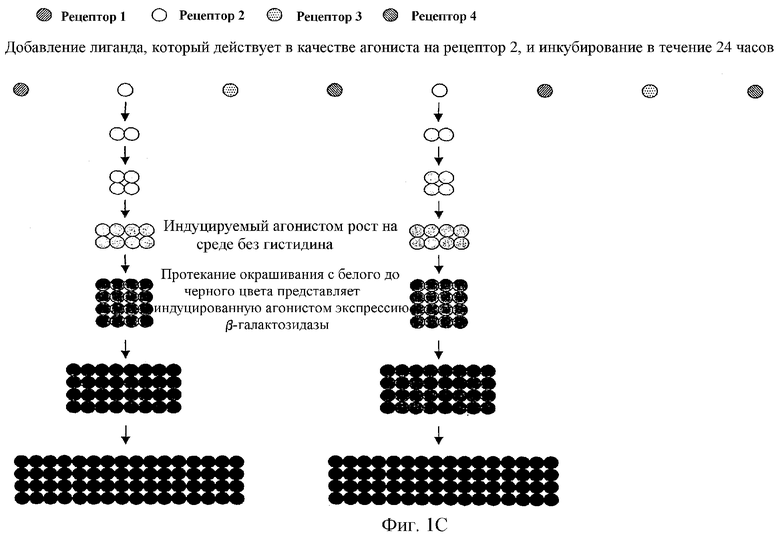
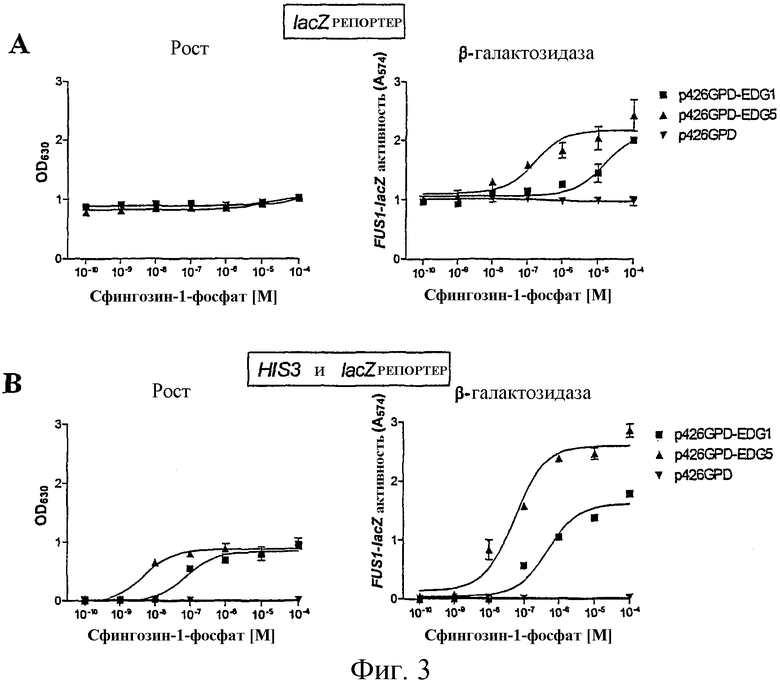
Описанные в литературе анализы на основе GPCR дрожжей используют большей частью только один репортерный ген, в основном HIS3 или LacZ (Price et al., Mol Cell Biol 15, 6188-6195 (1995); Price et al., Molecular Pharmacology 50, 829-837 (1996); Campbell et al., Bioorg. Med. Chem. Lett. 9, 2413-2418 (1999); Pausch, Trends Biotechnol 15, 487-494 (1997)) под контролем FUS1-промотора (FUS1-HIS3 или FUS1-lacZ). При использовании FUS1-HIS3 активацию пути трансдукции сигнала измеряют как помутнение культуры дрожжей в жидкой среде без гистидина. Эксперименты авторов настоящего изобретения показали, что определение только показателя роста дает в жидкой культуре отношение сигнала к фону приблизительно 30-50:1 (см. фиг.1а и 3). Анализы в жидкости β-галактозидазы при использовании хлорфеноловый красный-β-D-галактопиранозида (CPRG) в качестве субстрата для фермента показали после стимуляции сигнал, повышенный в 2-3-раза над фоном (см. также фиг.1b). В попытке дальнейшего увеличения окна измерения использовали одновременно оба репортерных гена в дрожжевой клетке, так как это должно было привести к усилению (умножению) обоих измеряемых сигналов. На фиг.1с этот принцип представлен для наглядного пояснения. Этот анализ с двумя репортерными генами приводил затем также к улучшению отношения сигнала к фону до приблизительно 100-150:1. Brown et al., Yeast 16, 11-22 (2000) описывает подобный анализ. Здесь также использовали одновременно FUS1-HIS3 и FUS1-lacZ в анализе β-галактозидазы в жидкости с использованием CPRG в качестве субстрата. CPRG добавляют здесь во время всего периода времени стимуляции рецептора лигандом. В противоположность этому, в данном эксперименте CPRG добавляют вместе с детергентом в забуференном растворе лишь после стимуляции рецептора лигандом, что приводит к явному улучшению результата измерения β-галактозидазы. Дело в том, что, во-первых, если CPRG присутствует во время индуцируемого лигандом роста, он легко ингибируется, во-вторых, CHRG лишь с трудом проникает через плазматическую мембрану во внутреннее пространство клетки. Обе проблемы могут быть устранены, если CHRG добавляется лишь после произошедшего роста вместе с детергентом, который может разрушать плазматическую мембрану. Этот способ используется в предпочтительном варианте осуществления анализа с двумя репортерными генами, в котором один репортерный ген является маркером роста, а другой репортерный ген является ферментом или GFP (зеленым флуоресцентным белком). Только комбинирование роста, логарифмического события, и более или менее линейно индуцируемой экспрессии измеряемого фермента или флуоресцирующего белка приводит к описанному усилению сигнала, т.е. к большому окну измерения. В ЕР 0708922 В1 (Acadia Pharmaceuticals) описан также способ, который основан на росте как ответе на стимуляцию рецептора. Однако при этом стимулируемые лигандом экспрессирующие рецептор клетки растут только быстрее, чем нестимулированные клетки (см. фиг.2 и фиг.10 в ЕР 0708922 В1). В описанном здесь изобретении такие нестимулированные дрожжевые клетки вообще не растут (см. приведенные примеры: например, фиг.3В, левый график). В ЕР 0708922 В1 используют в качестве измеряемого сигнала также активность гетерологично экспрессируемого фермента β-галактозидазы. Однако здесь LacZ экспрессируется конститутивно, т.е. измеренная активность фермента является только мерой количества клеток, которые выросли в виде ответной реакции на стимуляцию лигандом пути трансдукции сигнала, а не мерой эффективности стимуляции пути трансдукции сигнала. В описанном здесь изобретении экспрессия LacZ, в противоположность этому, находится под контролем индуцируемого посредством пути ответной реакции на феромоны промотора (например, FUS1 или YNL279w). На фиг.3А показано, что также и при одном и том же количестве клеток дрожжей (левый график) измеряемая активность β-галактозидазы зависит от добавленного количества лиганда, т.е. от эффективности стимуляции пути трансдукции сигнала (правый график).
Под "амплификацией" клеток понимают согласно ЕР 0708922 В1 (см. стр. 10) "рост трансфицированных рецептором клеток, в частности, в сравнении с ростом не трансфицированных рецептором клеток". То есть как трансфицированные рецептором, так и не трансфицированные рецептором клетки могут расти, только трансфицированные клетки растут более быстро после стимуляции лигандом. Это наглядно поясняется на фигурах стр. 33 и 44. В описанных клеточных линиях не присутствует никакая репортерная конструкция, только экспрессия которой делает возможным рост. Единственная модификация, которая делает возможным рост, представляет собой стимулируемый лигандом сверхэкспрессируемый рецептор.
Однако в соответствии с предпочтительным вариантом осуществления проводят двойной отбор: в питательной среде отсутствуют урацил и гистидин, вещества, которые являются необходимыми для используемых здесь штаммов дрожжей. Поскольку авторы настоящего изобретения используют URA3-ген в качестве селективного маркера на применяемых ими рецепторных плазмидах, клетки, у которых отсутствует эта рецепторная плазмида, вообще не могут расти.
Трансфицированные ДНК-рецептором клетки в принципе также не могут расти так долго на этой питательной среде, пока они не стимулируются присутствием лиганда. Лишь при связывании лиганда экспрессируется репортерный ген HIS3, и клетки становятся способными расти на этой питательной среде. В принципе, дрожжевой штамм, который трансфицирован рецептором, но не несет маркера роста, такого как HIS3, в качестве репортерного гена, не реагирует с изменением роста.
Описанный способ может применяться как в формате отдельного рецептора, так и в формате множественных рецепторов (мультиплексном формате). Преимущества данного анализа проявляются особенно в формате множественных рецепторов. Теоретически должно быть достаточным наличие клетки дрожжей, которая экспрессирует определенный рецептор, для того чтобы, если эта клетка приходит в контакт с подходящим лигандом (химическим соединением или природным лигандом), она в качестве ответа на стимуляцию, так сказать, «превышала рост» в сравнении с фоном не реагирующих других дрожжевых клеток. Это создает преимущества для высокопроизводительного скрининга, так как этот способ позволяет одновременно подвергать скринингу несколько GPCR. Как раз для orphan-GPCR, значение которых для фармацевтической промышленности до сих пор является неясным, описанный здесь способ минимизирует применение отнимающих много времени и требующих больших расходов операций. Поскольку в отношении orphan-GPCR неизвестно также, с какой Gα-субъединицей они связываются, с использованием этого способа существует также возможность одновременного тестирования одного или нескольких orphan-GPCR в нескольких штаммах-трансплантатах.
Описание фигур
Фиг.1 наглядно объясняет принцип анализа с двумя репортерными генами в формате множественных рецепторов: фиг.1а показывает результат индуцированного агонистом роста. Фиг.1b представляет результат окрашивания, опосредованный индуцированной агонистом β-галактозидазы. Наконец, фиг.1с показывает двойной результат индуцированного агонистом роста и окрашивания.
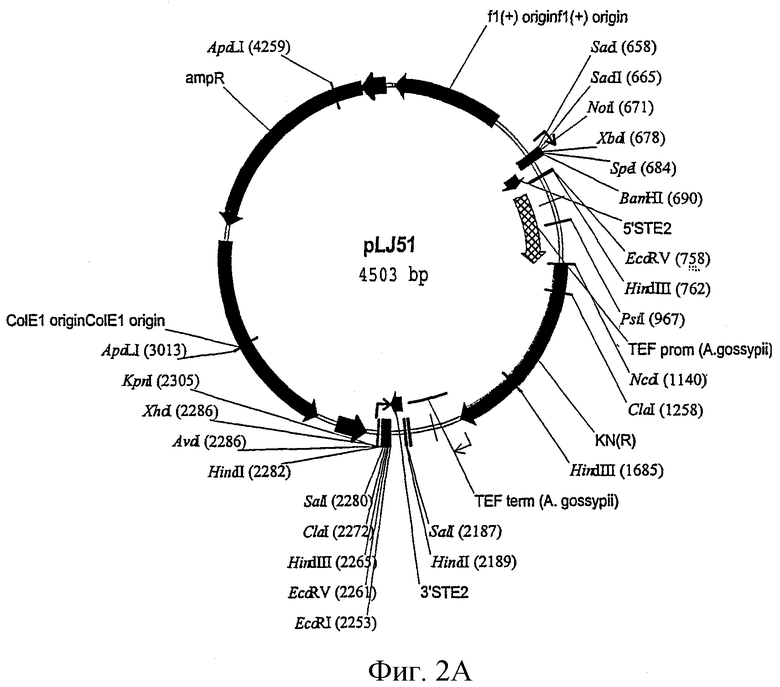
Фиг.2А-D показывают плазмиды, которые использовали для конструирования штаммов, которые основаны на промоторе YNL279w.
На фиг.3 показано, как анализ с двумя репортерными генами улучшает производительность жидкостного анализа дрожжей, в сравнении с использованием только одного репортерного гена.
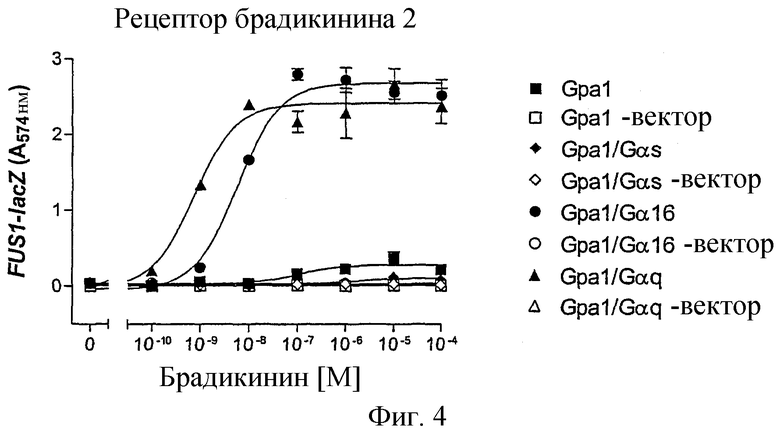
Фиг.4 показывает связывание рецептора брадикинина В2 человека с путем трансдукции сигнала дрожжей в зависимости от используемого Gα-трансплантата. В качестве контролей всегда использовали пустой вектор р426GPD.
Фиг.5 показывает, что анализ с двумя репортерными генами может быть также использован для скрининга антагонистов. В качестве примера изображены рецептор брадикинина В2 человека или пустой векторный контроль.
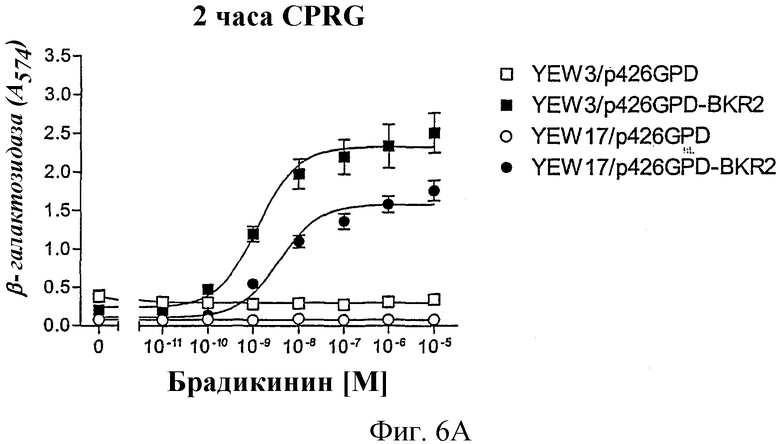
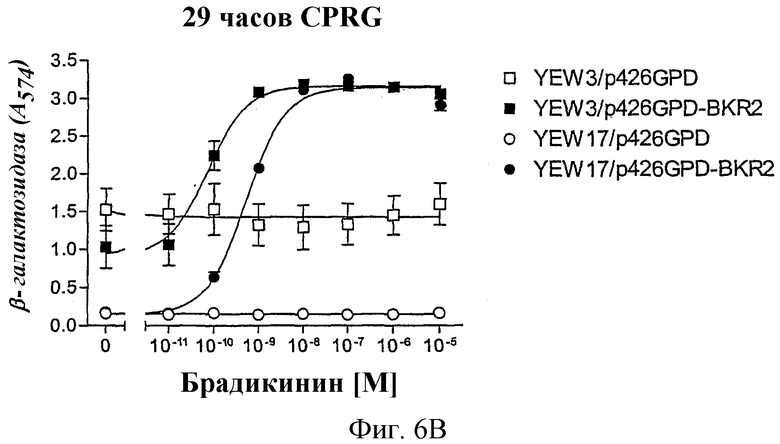
На фиг.6А и В показано, что применение промотора YNL279w также после 29-часовой инкубации с субстратом фермента явно вызывает меньший сигнал фона, чем применение FUS1-промотора.
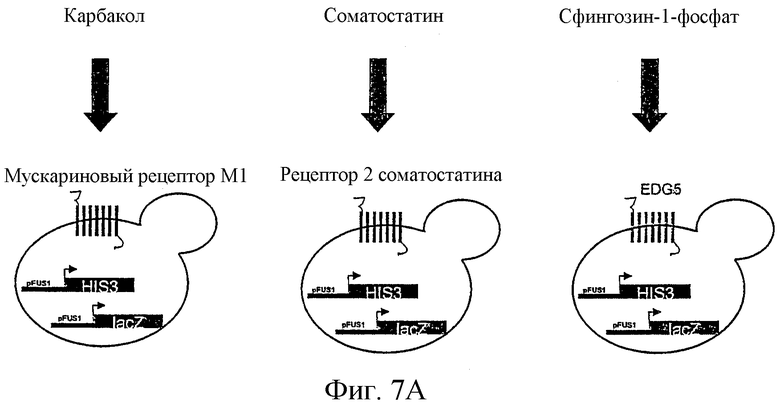
Фиг.7 наглядно объясняет проведение анализа в формате множественных рецепторов. На фиг.7А ясно показано, что различные GPCR в каждом отдельном случае экспрессируются в собственном штамме дрожжей, а не все вместе в одном штамме. Фиг.7В показывает производительность одного анализа в формате множественных рецепторов в сравнении с простым форматом рецептора в микротитрационном планшете.
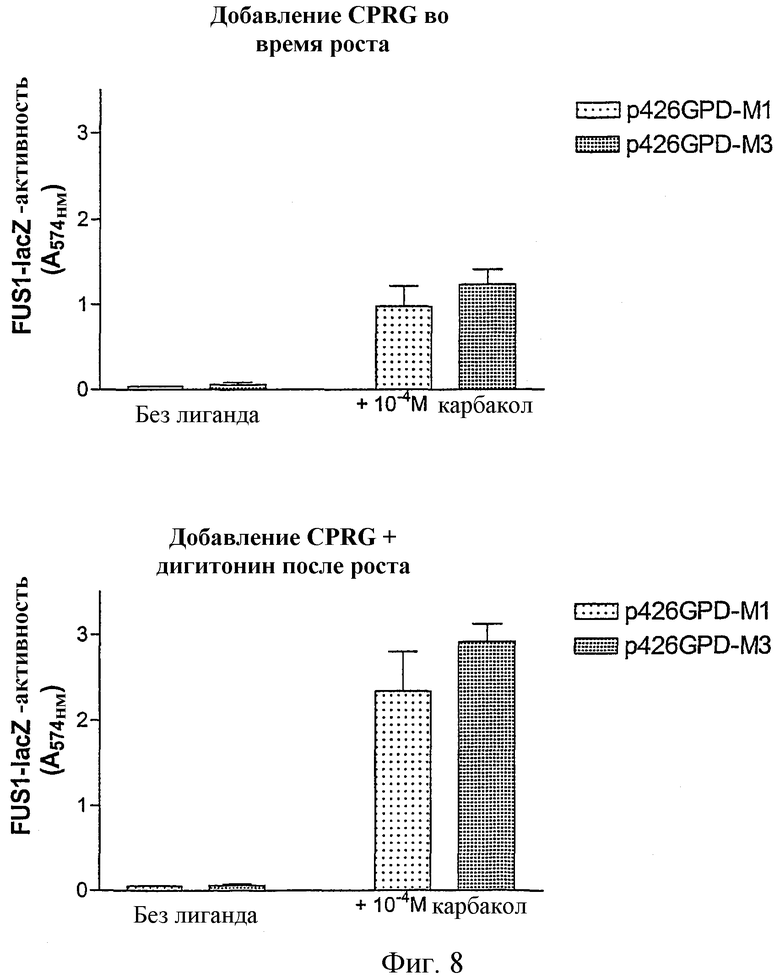
Фиг.8 показывает, что производительность анализа повышается, если субстрат фермента CPRG добавляют вместе с детергентом лишь после инкубирования с лигандом.
Материал и способы
Плазмиды и генетика дрожжей:
Все молекулярно-биологические и генетические манипуляции проводили в соответствии со стандартными способами (Ausubel et al., Current Protocols in Molecular Biology, Wiley & Sons, New York; Guthrie and Fink, Guide to Yeast Genetics and Molecular Biology, Methods in Enzymology, Academic Press, San Diego).
Экспрессионные конструкции для рецепторов:
Все экспрессионные конструкции основываются на эписомном челночном векторе 2-дрожжи-E. coli p426GPD (Mumberg et al., Gene 156, 119-122 (1995)). кДНК-последовательности, которые кодируют связанные с G-белком рецепторы, клонируют в этом векторе между GPD-промотором и CYC-терминатором для достижения сильной конститутивной экспрессии в клетках дрожжей. Следующие GPCR человека были клонированы в этот вектор:
EDG1-рецептор (GenBank NM_001400), EDG5-рецептор (GenBank NM_004230), рецептор брадикинина В2 (GenBank NM_000623), мускариновый рецептор M1 (GenBank NM_000738), рецептор соматостатина SSTR2 (GenBank NM_001050), мускариновый рецептор М3 (GenBank NM_000740).
Штаммы дрожжей:
Все штаммы дрожжей произведены из штамма дикого типа Saccharomyces cerevisiae W303-1a, описанного под АТСС-номером 208352.
Генотип: МАТа, ade2-1, ura3-1, his3-11, trp1-1, leu2-3, leu2-112, can1-100.
Использовали два различных набора штаммов дрожжей. Один набор происходит из YLJ21 и использует промотор гена FUS1 для экспрессии репортерного гена, другой набор происходит из YSG13 и использует промотор гена YNL279w.
Штамм дрожжей YLJ21 был предоставлен Эккехардом Леберером.
Генотип: МАТа, ste2::KanR sst2::ura3FOA far1::hisG FUS1::HIS3 mfa2::FUS1-lacZ::ura3FOA ade2-1, ura3-1, his3-11, trp1-1, leu2-3, leu2-112, can1-100.
Активация пути реакции на феромоны может измеряться с использованием двух репортерных генов FUS1::HIS3 и FUS1-lacZ, которые интегрированы в генном локусе HIS3 или MFA2. Ген FAR1 заменяли hisG-повтором, так что клетки могли продолжать расти также и в том случае, если активирован путь реакции на феромоны. Ген SST2 заменяли геном URA3, чтобы понижающая регуляция сигнала G-белка подавлялась ГТФазной функцией Sst2p. Маркер ura3 в каждом случае опять возвращали отбором на содержащей 5-фтороротовую кислоту среде. Ген STE2, который кодирует рецептор α-фактора, заменяли геном KanR.
Штамм дрожжей YSG13 получали следующим образом:
Генотип:
МАТа, ste2::KanR sst2::pYNL279w-HIS3 far1::pYNL279w-N136FUS1-lacZ::ADE2 ade2-1, ura3-1, his3-11, trp1-1, leu2-3, leu2-112, can1-100.
Конструирование штамма:
ste2::KanR
Чтобы заменить ген STE2 дрожжей геном резистентности к канамицину, плазмиду pLJ51 разрезали BamHI и EcoRI и трансформировали в штамм дрожжей дикого типа W303-1a. Отбор проводили на среде YPD+G418.
sst2::pYNL279w-HIS3
В следующей стадии ген SST2 дрожжей заменяли кассетой, которая делает возможной экспрессию гена HIS3 под контролем промотора YNL270w. Для этого плазмиду sst2::279LHIS3/pCR-BluntII разрезали BamHI и NotI и трансформировали. Отбор проводили на среде SC/Gluc-Ura-His+α-фактор.
far1::pYNL279w-N136FUS1-lacZ::ADE2
Наконец, ген FAR1 заменяли кассетой, которая делает возможной экспрессию N-концевых 136 аминокислот Fus1p, слитых с β-галактозидазой, под контролем промотора YNL279w. Для этого плазмиду pBSfar1::YNL279w-N136FUS1-lacZ::ADE2 w/o разрезали SacI и XhoI и трансформировали. Отбор проводили на среде SC/Gluc-Ade.
Правильную интеграцию всех фрагментов в геном всегда проверяли при помощи ПЦР.
Встраивание трансплантата в YLJ21 и YSG13:
На основе штаммов YLJ21 и YSG13 в конце концов заменяли последние 5 аминокислот α-субъединицы G-белка Gpa1 в геноме дрожжей последними 5 аминокислотами α-субъединиц G-белка человека. Для этого, например, для конструирования штамма дрожжей YEW3 плазмиду интеграции GPA1-C5-Galpha q разрезали SacI и трансформировали в штамм дрожжей YLJ21. Отбор проводили на среде SC/Gluc-Trp. Интеграцию других трансплантатов проводили подобным образом. В таблице 1 приведены различные трансплантаты и полученные из них штаммы дрожжей.
5-аминокислот
Анализ с двумя репортерными генами:
Клонированные в векторе p426GPD GPCR человека трансформируют в каждом отдельном случае в штамм дрожжей и на чашках для SC-отбора инкубируют без урацила и с 2% глюкозой в качестве источника углевода (SC/Gluc-Ura) в течение 3 дней при 30°С. Затем полученные таким образом отдельные колонии используют для засева ночных культур 2 мл в SC/Gluc-Ura. На следующий день клетки разводят 1:100 в SC/Gluc-Ura-His рН 6,8. В случае штаммов дрожжей, которые используют для экспрессии репортерного гена FUS1-промотор, к этой среде добавляют еще 2-10 мМ 3-аминотриазол (3-АТ, Sigma). По 90 мкл этой разбавленной клеточной суспензии пипетируют в каждую лунку 96-луночного микротитрационного планшета, в которой находятся уже 10 мкл подлежащего исследованию лиганда. Планшеты инкубируют при встряхивании или без встряхивания в течение 5-24 часов при 30°С. Затем в каждую лунку добавляют 50 мкл смеси для анализа. Смесь для анализа состоит из 150 мкг/мл дигитонина (Sigma), 300 мкг/мл смеси хлорфеноловый красный-β-D-галактопиранозид (CPRG, Roche), 300 мМ натрий-фосфатного буфера рН 6,7. После инкубирования в течение 2 часов при 30°С при встряхивании или без встряхивания измеряют активность β-галактозидазы в виде поглощения при 574 нм в спектрофотометре (Spectramax Plus, Molecular Devices). Обработку данных и построение кривых зависимости действия от дозы производят с использованием компьютерной программы Graphpad Prism 3.0. Все точки измерений являются средними показателями из трехкратных определений.
Пример 2:
Сравнение между применением отдельно одного репортерного гена или двух репортерных генов
Штамм дрожжей YLJ21 трансформировали либо пустым вектором p424GPD, либо GPCR EDG1 и EDG5 человека, которые были клонированы в p426GPD. Затем трансформированные дрожжи выращивали в 2 мл SC/Gluc-Ura в течение ночи при 30°С. На следующий день эти культуры разбавляли 1:100 в среде SC/Gluc-Ura-His рН 6,8 без (фиг.3А) или с 2 мМ 3-АТ (фиг.3В). По 90 мкл разбавленной клеточной суспензии пипетировали в каждую лунку 96-луночного микротитрационного планшета, в которой уже находились 10 мкл серийного разведения сфингозин-1-фосфата (Biomol) или в качестве контроля чистой воды. Планшеты инкубировали в течение 23 часов при 30°С при встряхивании (700 об/мин). Помутнение, которое происходило в результате роста клеток дрожжей, измеряли при 630 нм в фотометре (фиг.3А и В, в каждом случае левый график). После этого добавляли по 50 мкл в каждую лунку. После 2-часового инкубирования при 30°С при встряхивании измеряли активность β-галактозидазы в виде поглощения при 574 нм в фотометре.
Левый график фиг.3А показывает, что при использовании FUS1-HIS в качестве репортерной конструкции без добавления 3-АТ не наблюдается кривая «доза-ответ». FUS1-промотор приводит также без стимуляции пути трансдукции сигнала к довольно высокому сигналу фона на среде без гистидина, т.е. этот промотор не регулируется строгим образом. В отличие от этого, при добавлении 2 мМ 3-АТ, конкурентного ингибитора His3p, сигнал фона подавляется, и высота измеряемого сигнала зависит от количества сфингозин-1-фосфата (Ancellin et al., J Biol Chem 274, 18997-19002 (1999)) в среде, если экспрессируется EDG1 или EDG5 (фиг.3В, левый график). Из правого графика фиг.3А видно, что LacZ в качестве репортерного гена также приводит к приемлемой кривой «доза-ответ», однако окно измерений является очень малым. Как показывает правый график 3В, применение HIS3 и LacZ одновременно приводит к многократно улучшенному отношению сигнала к фону. В целом этот эксперимент показывает, что GPCR EDG1 и EDG5 человека могут быть сопряжены через собственную дрожжевую Gα-субъединицу GPA1p с путем реакции на феромоны.
Пример 3:
Анализ с двумя репортерными генами при подключении рецептора брадикинина В2 человека к пути трансдукции сигнала дрожжей через Gα-трансплантаты
Штаммы дрожжей YLJ21 (Gpa1), YEW1 (Gpa1/Gαs), YEW2 (Gpa1/Gα16) и YEW3 (Gpa1/Gαq) трансформировали либо пустым вектором p426GPD, либо рецептором брадикинина В2 человека, который был клонирован в p426 GPD. Этот анализ проводили при стандартных условиях, как описано выше, в присутствии 2 мМ 3-АТ. В качестве лиганда служил природный агонист брадикинин (Sigma); инкубирование проводили в течение 20 часов. Из фиг.4 видно, что рецептор брадикинина В2 почти совсем не мог подключаться к пути реакции на феромоны, если присутствовал только штамм дрожжей Gpa1p. В отличие от этого, если дрожжи экспрессируют Gα-трансплантаты Gpa1/Gα16 или Gpa1/Gαq, подключение этого рецептора к пути трансдукции сигнала является успешным. Gpa1/Gαq обеспечивает самое эффективное подключение, что можно было ожидать, так как рецептор брадикинина В2 человека в его природном клеточном окружении сопряжен с Gαq (Hall, Pharmacol. Ther. 56, 131-190 (1992)).
Пример 4:
Использование анализа с двумя репортерными генами для анализов антагонистов
Дрожжевой штамм YEW3 (Gpa1/Gαq) трансформировали либо пустым вектором p426GPD, либо рецептором брадикинина В2 человека, который был клонирован в p426GPD. Этот анализ проводили аналогично примеру 2. Единственным различием было то, что в каждой лунке тест-планшета находился 1 нМ агонист брадикинина, к которому добавляли разведения антагониста НОЕ140 (Sigma; Hall, Gen. Pharmacol. 28, 1-6 (1996)). Фиг.5 показывает, что увеличивающиеся концентрации НОЕ140 подавляли индуцируемый брадикинином сигнал до уровня фона. Таким образом, анализ с двумя репортерными генами пригоден для анализов антагонистов.
Пример 5:
Сравнение между промоторами FUS1 и YNL279w
Штаммы дрожжей YEW3 (Gpa1/Gαq, FUS1-промотор) и YEW17 (Gpa1/Gαq, YNL279w-промотор) трансформировали либо пустым вектором p426GPD, либо рецептором брадикинина В2 человека, который был клонирован в p426 GPD. Этот анализ проводили при стандартных условиях, как описано выше, в присутствии 2 мМ 3-АТ в случае YEW3 и без 3-АТ в случае YEW17. После добавления смеси для анализа проводили измерение активности β-галактозидазы после 2 часов и еще раз после 29 часов.
Как показано на фиг.6А и еще более выразительно на фиг.6В, в случае FUS1-промотора повышается значительно на протяжении времени сигнал фона трансформированного рецептором штамма или также трансформированного пустым контрольным вектором штамма, даже хотя в среде присутствовал 3-АТ. Добавление 3-АТ не является необходимым. Отсюда можно сделать вывод, что регуляция YNL279w является очень строгой. Это оказывается особенно выгодным при высокопроизводительных анализах, так как временная точка измерения не должна точно соблюдаться, что делает возможным гибкий режим работы.
Пример 6:
Проведение анализа в формате множественных рецепторов (мультиплексном формате)
Штамм YEW3 (Gpa1/Gαq) трансформировали мускариновым рецептором М1 человека, а штамм YLJ21 (Gpa1) трансформировали рецептором 2 соматостатина и EDG5. Эти рецепторы были клонированы в p426GPD. Проведение анализа происходило согласно вышеописанному стандартному способу. Рецепторы тестировали либо по отдельности, либо в смеси. В случае смеси использовали такие же ночные культуры, какие использовали в отдельных тестах. Их смешивали сначала при разведении 1:100 в SC/Gluc-Ura-His рН 6,8, т.е. эта смесь содержит в результате в целом в три раза больше клеток, чем отдельные тесты. Фиг.7А должна объяснять, что все эти рецепторы экспрессировались индивидуально, т.е. не совместно в одной клетке. Инкубирование с лигандами карбаколом (Sigma; разведение 10-8-10-2 М), соматостатином-14 (Bachem; 10-10-10-4 М) и сфингозин-1-фосфатом (Biomol, 10-10-10-4 М) продолжалось в течение 24 часов. Из фиг.7В видно, что детекция агонистов является возможной также и в смеси.
Пример 7:
Сравнение двух способов детектирования β-галактозидазы
Обычно анализ CPRG дрожжей проводят таким образом, что CPRG на протяжении всего времени, в течение которого трансформированные рецептором клетки дрожжей находятся в контакте с лигандом, присутствует в среде (Brown et al., Yeast 16, 11-22 (2000) и WO99/14344). Фиг.8 показывает сравнение между этим способом и способом, предложенным в настоящем изобретении.
Штамм YEW3 (Gpa1/Gαq) трансформировали мускариновыми рецепторами М1 и М3 человека, которые были клонированы в p426GPD. Ночные культуры выращивали, как описано выше, и затем разводили в двух различных средах 1:100 до OD600 0,02. В одном случае (фиг.8А) клетки разводили в SC/Gluc-Ura-His, 2 мМ 3-АТ, 0,1 мг/мл CPRG, 0,1 М натрий-фосфатном буфере рН 7. В другом случае анализ проводили, как описано выше, в присутствии 2 мМ 3-АТ (фиг.8В). В качестве лиганда для мускариновых рецепторов использовали карбакол. Анализ проводили для данных, представленных на фиг.8А, в течение 28 часов перед измерением тест-планшета в фотометре. В случае фиг.8В инкубирование проводили в течение 26 часов и затем добавляли тест-смесь. Измерение выполняли спустя 2 часа.
Как видно из фиг.8, добавление CPRG и детергента, в данном случае дигитонина, лишь после инкубирования с лигандом, улучшает значительно производительность этого анализа. Следующим преимуществом описанного здесь способа является тот факт, что исключается возможное взаимодействие CPRG во время продолжительного инкубирования с химическими соединениями, прежде всего во время скрининга.


Настоящее изобретение относится к клеточному способу идентификации веществ, которые способны влиять на активность молекулы-мишени, причем анализируемые дрожжевые клетки несут по меньшей мере один репортерный ген, и активность этой молекулы-мишени влияет на пролиферацию клеток и на активность продукта репортерного гена, предусматривающему стадии контактирования по меньшей мере одной дрожжевой клетки с тестируемым веществом, детекции пролиферации клеток, детекции активности продукта репортерного гена. При этом указанная дрожжевая клетка содержит указанную молекулу-мишень, репортерный ген маркера роста и репортерный ген, выбранный из гена, кодирующего фермент или флуоресцирующий белок, причем активность указанного фермента может быть детектирована посредством превращения субстрата, который добавляют со смещением во времени после добавления анализируемого вещества в месте с веществом, делающим клеточную стенку проницаемой или разрушающим ее. Преимущество изобретения заключается в повышении специфичности. 8 з.п ф-лы, 8 ил.
a) контактирования по меньшей мере одной дрожжевой клетки с тестируемым веществом, причем эта дрожжевая клетка содержит указанную молекулу-мишень, репортерный ген маркера роста и репортерный ген, выбранный из гена, кодирующего фермент или флуоресцирующий белок, причем активность указанного фермента может быть детектирована посредством превращения субстрата, который добавляют со смещением во времени после добавления анализируемого вещества вместе с веществом, делающим клеточную стенку проницаемой или разрушающим ее,
b) детектирования пролиферации клеток и
c) детектирования активности продуктов репортерных генов.
| US 5691188 A, 25.11.1997 | |||
| US 5885769 A, 23.03.1999 | |||
| ЕР 0708922 А, 01.05.1996 | |||
| PAUSCH MN | |||
| et al., G-protein-coupled receptors in Saccharomyces cerevisiae high-throughput screening assays for drug discovery, Trend | |||
| Biotechnol., 1997, dec., 15(12), pp.487-494. |

