ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
[0001] Настоящей заявке испрашивается приоритет по предварительной заявке США №62/597388, поданной 11 декабря 2017 года, содержание которой включено в настоящее описание в качестве ссылки в полном объеме.
УТВЕРЖДЕНИЕ, КАСАЮЩЕЕСЯ СПОНСИРУЕМОГО ПРАВИТЕЛЬСТВОМ ИССЛЕДОВАНИЯ
[0002] Настоящее изобретение было осуществлено при поддержке правительства по гранту №1747391, выданному Национальным научным фондом. Правительство имеет определенные права на изобретение.
ОБЛАСТЬ ТЕХНИКИ
[0003] Изобретение относится к клеточному дисплею и к способам высокопроизводительного скрининга белковых библиотек. В частности, изобретение относится к способам дисплея белков в периплазаматическом пространстве дрожжей и к применению таких способов для скрининга белковых библиотек в отношении специфического связывания или функциональных характеристик.
УРОВЕНЬ ТЕХНИКИ
[0004] Технология молекулярного дисплея зарекомендовала себя в качестве бесценной для открытия, продуцирования и оптимизации белков и пептидов для различных биотехнологических и биомедицинских применений. Различные подходы, включая фаговый дисплей (Smith (1985) Science 228:1315-1317), мРНК- (Wilson et al. (2001) Proc. Natl. Acad. Sci. USA 98:3750-3755) и ДНК-дисплей (Yonezawa et al. (2003) Nucleic Acids Res. 31:e118), рибосомальный дисплей (Hanes & Pluckthun (1997) Proc. Natl. Acad. Sci. USA 94:4937-4942), дисплей на эукариотических вирусах (Bupp & Roth (2002) Mol. Ther. 5:329-335; Muller et al. (2003) Nat. Biotechnol. 21:1040-1046), бактериальный дисплей (Lu et al. (1995) Biotechnology13:366-372) и дрожжевой дисплей (Boder & Wittrup (1997) Nat. Biotechnol. 15:553-557), были разработаны для скрининга комбинаторных библиотек рекомбинантных белков в отношении желаемых характеристик. Такие технологии дисплея широко используются в инженерии белков для идентификации белков, обладающих увеличенной стабильностью, и желаемой аффинностью связывания и ферментативной активностью, и они оказались применимыми в различных применениях, включая направленные изменения, созревание аффинности, инженерию терапевтических белков и антител, получение биотоплива, адсорбцию загрязнителей окружающей среды, картирование эпитопов и исследование белок-белковых взаимодействий.
[0005] В частности, дрожжевой дисплей используют для широкого множества прокариотических и эукариотических белков (Cherf et al. (2015) Methods Mol. Biol. 1319:155-175). Экспрессия в дрожжевых клетках обеспечивает преимущество в виде возможности надлежащего сворачивания и гликозилирования эукариотических белков. В общепринятом дрожжевом дисплее рекомбинантные белки экспонируют на поверхности дрожжевых клеток путем слияния с белком клеточной стенки. Хотя Saccharomyces cerevisiae является наиболее часто используемым видом для дисплея на клеточной поверхности, другие виды дрожжей, включая Pichia, Candida и Yarrowia, оказались применимыми для некоторых применений (Tanaka et al. (2012) Appl. Microbiol. Biotechnol. 95(3):577-591, Buerth et al. (2016) Appl. Microbiol. Biotechnol. 100(16):6981-6990, Madzak (2015) Appl. Microbiol. Biotechnol. 99(11):4559-4577).
[0006] Остается потребность в усовершенствованных способах, которые более эффективно экспонируют белки, в частности, для высокопроизводительного скрининга белок-белковых взаимодействий с мембранными белками.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0007] Настоящее изобретение относится к высокопроизводительному скринингу белковых библиотек в отношении специфического связывания или функциональных характеристик посредством дисплея белков в периплазматическом пространстве дрожжевых клеток.
[0008] В одном аспекте изобретение относится к библиотеке дрожжевого периплазматического дисплея, содержащей множество дрожжевых клеток-хозяев, где каждая дрожжевая клетка содержит: a) вариант белка для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где вариант экспонированного белка является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество вариантов белков; b) периплазматический якорный белок, где периплазматический якорный белок связан с вариантом белка, так что вариант белка экспонируется в периплазматическом пространстве; и c) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок находится на плазматической мембране дрожжевой клетки-хозяина и доступен для варианта белка, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина. Дрожжевая клетка-хозяин может быть гаплоидной или диплоидной.
[0009] В определенных вариантах осуществления вариант белка и периплазматический якорный белок ковалентно связаны вместе в слитом белке. В других вариантах осуществления вариант белка и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий в комплексе. В других вариантах осуществления вариант белка и периплазматический якорный белок связаны посредством непептидной связи в комплексе. В некоторых вариантах осуществления непептидная связь представляет собой дисульфидную связь.
[0010] В определенных вариантах осуществления периплазматический якорный белок содержит сигнальную последовательность, которая обеспечивает транспорт слитого белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или клеточную стенку, так что слитый вариант белка экспонируется в периплазме. Иллюстративная сигнальная последовательность, которую можно использовать, представляет собой сигнальную последовательность препро-альфа-фактора.
[0011] В определенных вариантах осуществления периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен, который выставляет слитый вариант белка в периплазму.
[0012] В определенных вариантах осуществления периплазматический якорный белок содержит связанный с клеточной мембраной белковый домен, который локализуется на внешней поверхности клеточной мембраны, так что экспонированный вариант белка выступает в периплазму. В определенных вариантах осуществления связанный с клеточной мембраной белковый домен представляет собой заякоривающий на плазматической мембране гликозилфосфатидилинозитол (GPI)-домен. Например, заякоривающий на плазматической мембране GPI-домен может представлять собой заякоривающий на плазматической мембране GPI-домен япсина, такой как, но не ограничиваясь ими, заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7.
[0013] В определенных вариантах осуществления периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что экспонируемый вариант белка выступает в периплазму.
[0014] В определенных вариантах осуществления периплазматический якорный белок содержит сигнальную последовательность, которая обеспечивает транспорт слитого белка в периплазму дрожжевой клетки-хозяина, и периплазматический якорный белок является достаточно большим, чтобы слитый белок оставался в периплазме.
[0015] В определенных вариантах осуществления якорный белок является компонентом периплазматического белкового комплекса, который является достаточно большим, чтобы образование комплекса в периплазме приводило к удержанию слитого белка в периплазме.
[0016] В другом варианте осуществления слитый белок дополнительно содержит метку.
[0017] В определенных вариантах осуществления варианты белка представляют собой антитела, миметики антител, аптамеры, антигены, ферменты, рецепторы, гормоны, субтраты, агонисты, антагонисты или лиганды.
[0018] В определенных вариантах осуществления варианты белка представляют собой антитела, выбранные из группы, состоящей из моноклональных антител, химерных антител, наноантител, рекомбинантных фрагментов антител, Fab-фрагментов, Fab'-фрагментов, F(ab')2-фрагментов, Fv-фрагментов и scFv-фрагментов.
[0019] В определенных вариантах осуществления каждая дрожжевая клетка-хозяин в библиотеке дрожжевого периплазматического дисплея, кроме того, содержит представляющий интерес белок-мишень, который экспрессируется в месте, доступном для экспонируемого варианта белка (например, достаточно близко для связывания экспонированного варианта белка с представляющим интерес белком-мишенью). Например, представляющий интерес белок-мишень может находиться на плазматической мембране или в периплазме дрожжевой клетки-хозяина. Представляющий интерес белок может представлять собой, например, мембранный белок, рецептор, ионный канал или переносчик. В одном варианте осуществления представляющий интерес белок-мишень представляет собой сопряженный с G-белком рецептор (GPCR).
[0020] В определенных вариантах осуществления каждая дрожжевая клетка-хозяин дополнительно содержит репортерную систему для детекции ответа представляющего интерес белка-мишени на белок-белковое взаимодействие с экспонированным вариантом белка. В определенных вариантах осуществления экспонированный вариант белка представляет собой антагонист представляющего интерес белка-мишени, и ответ представляет собой снижение активности представляющего интерес белка-мишени при связывании антагониста с представляющим интерес белком-мишенью, где репортерная система выявляет снижение активности представляющего интерес белка-мишени при связывании антагониста с представляющим интерес белком-мишенью. В других вариантах осуществления экспонированный вариант белка представляет собой агонист представляющего интерес белка-мишени и ответ представляет собой увеличение активности представляющего интерес белка-мишени при связывании агониста с представляющим интерес белком-мишенью, где репортерная система позволяет детекцию повышения активности представляющего интерес белка-мишени при связывании агониста с представляющим интересом белком-мишенью.
[0021] В определенных вариантах осуществления активация представляющего интерес белка-мишени увеличивает рост дрожжевых клеток-хозяев. В этом случае библиотеку дрожжевого периплазматического дисплея можно подвергать скринингу в отношении агониста представляющего интерес белка-мишени посредством культивирования по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея в среде, где рост дрожжевых клеток-хозяев в среде указывает на то, что вариант белка, экспонированный в дрожжевой клетке-хозяине, является агонистом представляющего интерес белка-мишени.
[0022] В других вариантах осуществления активация представляющего интерес белка-мишени снижает рост дрожжевых клеток-хозяев. В этом случае библиотеку дрожжевого периплазматического дисплея можно подвергать скринингу в отношении антагониста представляющего интерес белка-мишени посредством культивирования по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея в среде, где рост дрожжевых клеток-хозяев в среде указывает на то, что вариант белка, экспонированный в дрожжевой клетке-хозяине, является антагонистом представляющего интерес белка-мишени.
[0023] В другом варианте осуществления изобретение относится к библиотеке дрожжевого периплазматического дисплея, содержащей множество дрожжевых клеток-хозяев, где каждая дрожжевая клетка-хозяин содержит: a) слитый белок, содержащий периплазматический якорный белок, слитый с антителом, для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител; и b) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок находится на плазматической мембране дрожжевой клетки-хозяина и является доступным для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
[0024] Представляющий интерес мембранный белок-мишень может представлять собой, например, рецептор, ионный канал и переносчик. В некоторых вариантах осуществления представляющий интерес мембранный белок-мишень содержит мутацию, которая повышает или снижает его активность.
[0025] Антитела, которые могут экспонироваться с представляющим интерес мембранным белком-мишенью, могут включать, но не ограничиваются ими, моноклональные антитела, химерные антитела, гуманизированные антитела, наноантитела, рекомбинантные фрагменты антител, Fab-фрагменты, Fab'-фрагменты, F(ab')2-фрагменты, Fv-фрагменты и scFv-фрагменты.
[0026] В определенных вариантах осуществления библиотека дрожжевого периплазматического дисплея дополнительно содержит репортерную систему, содержащую репортерный ген, функционально связанный с индуцибельным промотором, который активируется, когда активируется представляющий интерес мембранный белок-мишень, чтобы позволить детекцию увеличения или снижения активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью. Например, репортерный ген может представлять собой алиментарный маркер (например, HIS3, HIS7, ARG6, LEU2, URA3 и TRP1), маркер резистентности к антибиотикам (например, сообщает резистентность к антибиотику, такому как генетицин (например, aphA1), зеоцин (например, ble), гигромицин B, ноурсеотрицин или биалафос), флуоресцентный маркер (например, зеленый флуоресцентный белок, красный флуоресцентный белок, синий флуоресцентный белок, голубой флуоресцентный белок, желтый флуоресцентный белок и оранжевый флуоресцентный белок), биолюминесцентный маркер (например, люцифераза или экворин), или маркер контрселекции (например, CAN1, URA3, MET15, TRP1 и TK). В определенных вариантах осуществления репортерный ген представляет собой селективный маркер, так что повышение активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью выявляется по росту дрожжевых клеток-хозяев на позитивной селективной среде. В других вариантах осуществления репортерный ген представляет собой маркер контрселекции, так что снижение активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью выявляется по росту дрожжевых клеток-хозяев на среде, содержащей агент контрселекции.
[0027] В определенных вариантах осуществления представляющий интерес мембранный белок-мишень представляет собой сопряженный с G-белком рецептор (GPCR), например, экзогенный GPCR, такой как GPCR млекопитающего (например, из человека или не являющегося человеком примата, грызуна, лабораторного животного, домашнего скота). В определенных вариантах осуществления GPCR млекопитающего представляет собой GPCR человека, выбранный из группы, состоящей из CXCR4, CXCR5, SSTR2, MOR, AVPR2, FPR2/ALX, ADORA2A, CHRM3, CGRP2, CCR2, CCR4, CCR5, CHRM4, PAC1, b2AR, CXCR2, CYSLTR2, KSHV vGPCR, PKR1, PKR2, CB1, CB2, A3AR и AT1R.
[0028] В определенных вариантах осуществления библиотека дрожжевого периплазматического дисплея дополнительно содержит сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяина.
[0029] В определенных вариантах осуществления сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен субъединицы G(дрожжей и C-концевой домен экзогенной субъединицы Gα. Например, дрожжевая субъединица G(может принадлежать семейству G-белков Gαi, Gαq, Gαs или Gαo. В химерной субъединице G(по меньшей мере пять C-концевых остатков субъединицы G(дрожжей могут быть заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего. В некоторых вариантах осуществления по меньшей мере 20 C-концевых остатков дрожжевой субъединицы G(заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего. В другом варианте осуществления химерная субъединица G(содержит по меньшей мере 41 N-концевой остаток дрожжевой субъединицы Gα.
[0030] Иллюстративные субъединицы G(млекопитающего включают G альфа-S, G альфа-I, G альфа-O, G альфа-T, G альфа-Z, G альфа-Q, G альфа-11, G альфа-12, G альфа-13 и трансдуцин.
[0031] В некоторых вариантах осуществления представляющий интерес GPCR-мишень обладает конститутивной независимой от лиганда активностью. В других вариантах осуществления лиганд добавляют для активации представляющего интерес GPCR-мишени.
[0032] В определенных вариантах осуществления дрожжевая клетка-хозяин представляет собой гаплоидную или диплоидную дрожжевую клетку-хозяина. В определенных вариантах осуществления дрожжевая клетка-хозяин представляет собой штамм Δfar1, Δsst2, Δste14, Δste3 или Δmat. Штамм Δmat может содержать, например, удаленный или инактивированный локус MAT(или удаленный или инактивированный локус MATa.
[0033] В другом варианте осуществления дрожжевая клетка-хозяин дополнительно содержит модифицированный белок CLN3, содержащий C-концевое укорочение, которое повышает содержание CLN3 в дрожжевой клетке-хозяине по сравнению с белком CLN3 дикого типа. Например, модифицированный белок CLN3 может сохранять по меньшей мере N-концевые аминокислоты 1-387 или 1-408 белка CLN3 дикого типа, или любое количество N-концевых аминокислот в этих диапазонах, такое как 1-388, 1-389, 1-390, 1-391, 1-392, 1-393, 1-394, 1-395, 1-396, 1-397, 1-398, 1-399, 1-400, 1-401, 1-402, 1-403, 1-404, 1-405, 1-406, 1-407 или 1-408, где C-концевое укорочение содержит делецию всех или некоторых из остальных остатков белка CLN3 дикого типа.
[0034] В другом варианте осуществления дрожжевая клетка-хозяин представляет собой штамм FAR1 для селекции антител-антагонистов GPCR.
[0035] В другом варианте осуществления дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном для селекции антител-агонистов GPCR.
[0036] В другом аспекте изобретение относится к библиотеке дрожжевого периплазматического дисплея, содержащей множество дрожжевых клеток-хозяев, где каждая дрожжевая клетка-хозяин содержит: a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител, где антитело связано с сигнальной последовательностью, которая обеспечивает транспорт в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или клеточную стенку, так что антитело экспонируется в периплазматическом пространстве дрожжевой клетки-хозяина; и b) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок находится на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
[0037] В другом аспекте изобретение относится к способу получения библиотеки дрожжевого периплазматического дисплея, описанной в настоящем описании, причем способ включает: a) предоставление множества рекомбинантных полинуклеотидов, кодирующих слитые белки, где каждый рекомбинантный полинуклеотид кодирует отличающийся слитый белок, содержащий периплазматический якорный белок, слитый с отличающимся антителом, для дисплея; b) трансфекцию множества дрожжевых клеток-хозяев множеством рекомбинантных полинуклеотидов, кодирующих слитые белки; c) трансфекцию множества дрожжевых клеток-хозяев рекомбинантным полинуклеотидом, кодирующим представляющий интерес мембранный белок-мишень; и d) культивирование множества дрожжевых клеток-хозяев в условиях, которые позволяют экспрессию слитых белков и представляющего интерес мембранного белка-мишени, где каждая дрожжевая клетка-хозяин экспонирует отличающееся антитело в периплазматическом пространстве, и представляющий интерес мембранный белок-мишень локализуется на плазматической мембране (т.е., где он доступен для связывания экспонированным антителом). В определенных вариантах осуществления в экспрессирующих векторах предоставлены рекомбинантные полинуклеотиды, кодирующие слитые белки или рекомбинантный полинуклеотид, кодирующий представляющий интерес мембранный белок-мишень. В других вариантах осуществления рекомбинантные полинуклеотиды, кодирующие слитые белки или представляющий интерес мембранный белок-мишень, встраиваются в геном дрожжевой клетки-хозяина в локусе-мишени.
[0038] В другом аспекте изобретение относится к способу получения библиотеки дрожжевого периплазматического дисплея, причем способ включает: a) предоставление множества рекомбинантных полинуклеотидов, кодирующих антитела, для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител; b) предоставление второго рекомбинантного полинуклеотида, кодирующего периплазматический якорный белок, где периплазматический якорный белок связан с антителом так, что антитело экспонируется в периплазматическом пространстве; c) трансфекцию множества дрожжевых клеток-хозяев первым множеством рекомбинантных полинуклеотидов и вторым рекомбинантным полинуклеотидом; d) трансфекцию множества дрожжевых клеток-хозяев рекомбинантным полинуклеотидом, кодирующим представляющий интерес мембранный белок-мишень; и e) культивирование множества дрожжевых клеток-хозяев в условиях, которые позволяют экспрессию антител, периплазматического якорного белка и представляющего интерес мембранного белка-мишени, где каждая дрожжевая клетка-хозяин экспонирует отличающееся антитело в периплазматическом пространстве и представляющий интерес мембранный белок-мишень локализуется на плазматической мембране, с получением библиотеки дрожжевого периплазматического дисплея.
[0039] Экспрессия слитых белков и представляющего интерес мембранного белка-мишени обычно зависит от присутствия промотора, который может быть включен в вектор или в хромосомный локус, в который встроены рекомбинантные полинуклеотиды. Промотор может представлять собой конститутивный или индуцибельный промотор. В определенных вариантах осуществления каждый рекомбинантный полинуклеотид содержит промотор, функционально связанный с полинуклеотидом, кодирующим слитый белок или представляющий интерес мембранный белок-мишень. Рекомбинантный полинуклеотид может быть предоставлен в векторе, содержащем промотор. В других вариантах осуществления хромосомный локус-мишень содержит промотор, который становится функционально связанным с полинуклеотидом, кодирующим слитый белок или представляющий интерес мембранный белок-мишень, который встраивается в хромосомный локус-мишень.
[0040] В другом варианте осуществления способ дополнительно включает введение в множество дрожжевых клеток-хозяев рекомбинантного полинуклеотида, кодирующего сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяине.
[0041] В другом варианте осуществления изобретение относится к нацеливающему в периплазму экспрессирующему вектору, содержащему: a) полинуклеотид, кодирующий сигнальный пептид; b) участок клонирования, пригодный для вставки в рамке считывания полинуклеотида, кодирующего вариант белка, после полинуклеотида, кодирующего сигнальный пептид; c) полинуклеотид, кодирующий заякоривающий на плазматической мембране гликофосфатидилинозитол (GPI)-домен, расположенный так, чтобы вектор был способен продуцировать слитый белок, содержащий сигнальный пептид и вариант белка, слитый с заякоривающим на плазматической мембране GPI-доменом; и d) промотор, функционально связанный с последовательностями, кодирующими слитый белок. В одном варианте осуществления сигнальный пептид содержит сигнальную последовательность препро-альфа-фактора. В другом варианте осуществления участок клонирования содержит один или более участков рестрикции. В определенных вариантах осуществления заякоривающий на плазматической мембране GPI-домен представляет собой заякоривающий на плазматической мембране GPI-домен япсина, такой как, но не ограничиваясь ими, заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7. В другом варианте осуществления нацеливающий в периплазму экспрессирующий вектор дополнительно содержит полинуклеотид, кодирующий линкер, где указанный полинуклеотид, кодирующий линкер, находится между участком клонирования и полинуклеотидом, кодирующим заякоривающий на плазматической мембране GPI-домен. Линкер может дополнительно содержать метку. В другом варианте осуществления нацеливающий на периплазму экспрессирующий вектор дополнительно содержит селективный маркер.
[0042] В другом аспекте изобретение относится к способу получения библиотеки дрожжевого периплазматического дисплея, описанной в настоящем описании, причем способ включает: a) предоставление множества рекомбинантных полинуклеотидов, кодирующих варианты антител, где каждый рекомбинантный полинуклеотид кодирует отличающийся вариант антитела; b) трансфекцию множества дрожжевых клеток-хозяев нацеливающим в периплазму экспрессирующим вектором, описанным в настоящем описании, c) трансфекцию множества дрожжевых клеток-хозяев множеством рекомбинантных полинуклеотидов, кодирующих варианты антител, где в каждой дрожжевой клетке-хозяине, рекомбинантный полинуклеотид, кодирующий вариант антитела, встроен в участок клонирования нацеливающего в периплазму экспрессирующего вектора посредством гомологичной рекомбинации, чтобы позволить экспрессию слитого белка, содержащего периплазматический якорный белок, слитый с вариантом антитела, для дисплея; c) трансфекцию множества дрожжевых клеток-хозяев рекомбинантным полинуклеотидом, кодирующим представляющий интерес мембранный белок-мишень; и d) культивирование множества дрожжевых клеток-хозяев в условиях, которые позволяют экспрессию слитых белков и представляющего интерес мембранного белка-мишени, где каждая дрожжевая клетка-хозяин экспонирует отличающееся антитело в периплазматическом пространстве и представляющий интерес мембранный белок-мишень локализуется на плазматической мембране (т.е., где он доступен для связывания экспонированным антителом). В другом варианте осуществления способ дополнительно включает введение в множество дрожжевых клеток-хозяев рекомбинантного полинуклеотида, кодирующего сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяине.
[0043] В другом аспекте изобретение относится к способу скрининга библиотеки дрожжевого периплазматического дисплея, содержащей репортерную систему, как описано в настоящем описании, в отношении антитела, которое модулирует активность представляющего интерес мембранного белка-мишени, причем способ включает культивирование по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея, описанной в настоящем описании, в селективной среде; и детекцию экспрессию репортерного гена, где увеличенная экспрессия репортерного гена указывает на то, что антитело увеличивает активность представляющего интерес мембранного белка-мишени, и сниженная экспрессия репортерного гена указывает на то, что антитело снижает активность представляющего интерес мембранного белка-мишени.
[0044] Иллюстративные репортерные гены включают алиментарный маркер (например, HIS3, HIS7, ARG6, LEU2, URA3 и TRP1), маркер резистентности к антибиотику (например, сообщает резистентность к антибиотику, такому как генетицин (aphA1), зеоцин (ble), гигромицин B, ноурсеотрицин и биалафос), флуоресцентный маркер (например, зеленый флуоресцентный белок, красный флуоресцентный белок, синий флуоресцентный белок, голубой флуоресцентный белок, желтый флуоресцентный белок и оранжевый флуоресцентный белок), биолюминесцентный маркер (например, люцифераза или экворин) и маркер контрселекции (например, CAN1, URA3, MET15, TRP1 и TK).
[0045] В другом варианте осуществления способ дополнительно включает положительную селекцию в отношении экспрессии алиментарного маркера, где рост дрожжевых клеток-хозяев в селективной среде с дефицитом питательных веществ указывает на то, что представляющий интерес мембранный белок-мишень активирован.
[0046] В другом варианте осуществления способ дополнительно включает положительную селекцию в отношении экспрессии маркера резситентности к антибиотику, где рост дрожжевых клеток-хозяев в селективной среде, содержащей антибиотик, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
[0047] В другом варианте осуществления способ дополнительно включает положительную селекцию для экспрессии флуоресцентного маркера, где детекция флуоресценции, испускаемой дрожжевыми клетками-хозяевами, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
[0048] В другом варианте осуществления способ дополнительно включает положительную селекцию в отношении экспрессии биолюминесцентного маркера, где детекция биолюминесценции, испускаемой дрожжевыми клетками-хозяевами, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
[0049] В другом варианте осуществления способ дополнительно включает отрицательную селекцию в отношении маркера контрселекции, где снижение активности представляющего интерес мембранного белка-мишени при связывании экспонированного антитела с представляющим интерес мембранным белком-мишенью определяют по росту дрожжевых клеток-хозяев в среде, содержащей средство, которое позволяет проводить селекцию против клеток, экспрессирующих маркер контрселекции.
[0050] В другом варианте осуществления изобретение относится к способу скрининга библиотеки дрожжевого периплазматического дисплея в отношении антитела, которое модулирует активность представляющего интерес GPCR-мишени, причем способ включает культивирование по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея в среде, где детекция активации или ингибирования ответа феромонов по меньшей мере в одной дрожжевой клетке-хозяине по сравнению с контрольной дрожжевой клеткой-хозяином, не имеющей антитела, экспонированного в периплазматическом пространстве, указывает на то, что экспонированное антитело в указанной по меньшей мере одной дрожжевой клетке-хозяине связывает и модулирует активность GPCR. В некоторых вариантах осуществления способ дополнительно включает приведение в контакт GPCR человека с лигандом. В других вариантах осуществления GPCR имеет конститутивную лиганд-независимую активность.
[0051] В определенных вариантах осуществления дрожжевая клетка-хозяин содержит сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяине. В определенных вариантах осуществления сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен дрожжевой субъединицы G(и C-концевой домен экзогенной субъединицы Gα. Например, дрожжевая субъединица G(может принадлежать к семейству G-белков Gαi, Gαq, Gαs или Gαo. В химерной субъединице G(по меньшей мере пять C-концевых остатков дрожжевой субъединицы G(могут быть заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающих. В некоторых вариантах осуществления по меньшей мере 20 C-концевых остатков дрожжевой субъединицы G(заменены соответствующими C-концевыми остатками субъединицы G(млекопитающих, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающих. В другом варианте осуществления химерная субъединица G(содержит по меньшей мере 41 N-концевой остаток дрожжевой субъединицы Gα. Иллюстративные субъединицы G(млекопитающих включают G альфа-S, G альфа-I, G альфа-O, G альфа-T, G альфа-Z, G альфа-Q, G альфа-11, G альфа-12, G альфа-13 и трансдуцин.
[0052] В определенных вариантах осуществления дрожжевая клетка-хозяин представляет собой штамм FAR1, где ингибирование ответа феромонов посредством антитела, действующего в качестве антагониста, который связывает и ингибирует GPCR в дрожжевой клетке-хозяине, приводит к прекращению остановки клеточного цикла и росту дрожжевой клетки-хозяина. В других вариантах осуществления дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном, где активация ответа феромонов посредством антитела, действующего в качестве агониста, который связывает и активирует GPCR в дрожжевой клетке-хозяине, приводит к повышению экспрессии репортерного гена.
[0053] В другом аспекте изобретение относится к дрожжевой клетке-хозяину, содержащей: a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, b) периплазматический якорный белок, где периплазматический якорный белок связан с антителом, так что антитело экспонируется в периплазматическом пространстве; и c) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок находится на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
[0054] В другом аспекте изобретение относится к антителу, связанному с периплазматическим якорным белком. В некоторых вариантах осуществления, в котором антитело продуцируется в дрожжевой клетке-хозяине, антитело локализуется в периплазмамтическом пространстве дрожжевой клетки-хозяина. В некоторых вариантах осуществления антитело и периплазматический якорный белок нековалентно связаны друг с другом посредством молекулярных связывающих взаимодействий в комплексе или связаны ковалентной непептидной связью в комплексе. В некоторых вариантах осуществления непептидная связь представляет собой дисульфидную связь. В некоторых вариантах осуществления антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
[0055] В другом аспекте изобретение относится к способу локализации антитела в периплазматическом пространстве дрожжевой клетки-хозяина, включающему связывание антитела с периплазматическим якорным белком, так чтобы антитело локализовалось в периплазматическом пространстве. В некоторых вариантах осуществления антитело и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий в комплексе или связаны посредством ковалентной непептидной связи в комплексе. В некоторых вариантах осуществления непептидная связь представляет собой дисульфидную связь. В некоторых вариантах осуществления антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
[0056] В другом аспекте изобретение относится к набору, содержащему библиотеку дрожжевого периплазматического дисплея, описанную в настоящем описании, и инструкции по скринингу множества вариантов белков в отношении их способности связывать и/или модулировать активность представляющего интерес белка-мишени.
[0057] Эти и другие варианты осуществления настоящего изобретения станут хорошо понятными специалистам в данной области с учетом настоящего описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0058] На фиг.1A-1D представлен новый способ дисплея дрожжевых клеток для скрининга антител, которые модулируют функцию GPCR. На фиг.1A представлена уникальная комбинация 1) функционального сопряжения GPCR человека-дрожжи с 2) секрецией аффинной молекулы и 3) локализацией аффинной молекулы вместе в высокопроизводимой в высокой степени модифицируемой дрожжевой клеточной платформы. Функциональный надлежащим образом свернутый GPCR обеспечивает ScFv (которые могут легко конвертироваться в IgG-антитела) или наноантитела, которые более вероятно будут функционировать в качестве терапевтических средств в контексте организма человека. На фиг.1B представлено использование "штамма для селекции антагонистов" для поиска антагонистов. На фиг.1C представлено применение "штамма для селекции агонистов" для поиска агонистов. Посредством изменения логики репортеров и селективных маркеров, сопряженных с выходом системы ответа феромонов, платформу можно использовать для селекции агонистов или антагонистов. На фиг.1D показано, что прямой функциональный скрининг обеспечивает терапевтические антитела-кандидаты, которые в норме были бы упущены при традиционном скрининге, что может обеспечить новые способы связывания и функциональное модулирование мишеней GPCR. Вследствие простоты проведения способов генной инженерии в дрожжах, можно корректировать как уровни экспрессии антител, так и уровни экспрессии GPCR, и приспособить селективные и поддающиеся скринингу репортеры так, чтобы они были очень чувствительными. Оба из них позволяют найти низкоаффинные, но функциональные кандидаты, которые можно без труда позднее подвергать созреванию аффинности.
[0059] На фиг.2 представлен способ снижения фонового уровня/ложноположительных результатов в "анализе гало". 107 клеток родительского штамма (слева) и текущего платформенного штамма (NIY326, справа) высевали на среду с агаром. На чашку помещали диск фильтровальной бумаги и на нее наносили в виде пятна 3 мкл 1 мМ альфа-фактора. Зону без роста в ответ на лиганд (желаемый фенотипический ответ) наблюдали в обоих случаях, однако в родительском штамме (слева) возникают мутанты-супрессоры и вырастают в колонии в присутствии феромона (колонии в области гало). В платформенном штамме NI326 (справа), фоновая скорость была снижена до ~10-7, что было продемонстрировано прозрачной зоной гало и отсутствием фоновых супрессорных мутаций, которые могли бы выступать в качестве ложноположительных результатов при селекции антагониста.
[0060] На фиг.3 представлена структура и концепция нацеливающего вектора для аффинной молекулы. Авторы изобретения клонировали аффинную молекулу ниже сигнала секреции и выше линкера и заякоривающего на внеклеточной мембране GPI-домена. При экспрессии в клетках белок секретируется во внеклеточное пространство, а затем GPI-домен процессируется, оставляя домен с GPI, который связывает с мембраной, который привязывает аффинные молекулы к этой клетке и оставляет их свободными для взаимодействия с GPCR-мишенью на ее внеклеточной поверхности.
[0061] На фиг.4A и 4B представлено подтверждение экспрессии аффинной молекулы/нацеливающего вектора. На фиг.4A показано, что, если экспрессированное наноантитело против GFP надлежащим образом сворачивается и локализуется, GFP, примененный снаружи клетки (после расщепления клеточной стенки), должен метить клеточную мембрану. На фиг.4B представлены изображения дрожжей, экспрессирующих наноантитело против GFP с использованием нацеливающего вектора авторов изобретения, после расщепления клеточной стенки и применения очищенного белка GFP, которые указывают на связывание GFP с мембраной. Не наблюдали флуоресценции в контрольных клетках (данные не представлены). Слева, светлое поле; справа, канал GFP.
[0062] На фиг.5A и 5B представлено подтверждение зависимости от плазмиды клонов, резистентных к альфа-фактору. На фиг.5A представлена схема стратегии. На фиг.5B представлен пример клона-"кандидата", который демонстрировал резистентный к альфа-фактору рост при исследовании посредством анализа гало, а затем продемонстрировал отсутствие резистентности после выключения плазмиды.
[0063] На фиг.6 представлена схема технологического потока.
[0064] На фиг.7 представлено воздействие на скорость роста дрожжевых клеток посредством активации каннабиноидного рецептора типа 2 (рецептор CB2) с использованием агонистов домена VHH, экспонированных в периплазматическом пространстве различными способами.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0065] Множество терапевтических мишеней при таких заболеваниях, как злокачественная опухоль и воспаление, вовлекают связанные с клеточной мембраной белки. Однако многие связанные с клеточной мембраной белки, обладающие наибольшим терапевтическим потенциалом для оказывающих большое влияние заболеваний, трудно преобразовать в лекарственное средство. Хотя низкомолекулярные соединения, влияющие на функцию этих белков, найти легко, они часто являются неспецифическими. В отличие от низкомолекулярных соединений, антитела и родственные аффинные молекулы (например, наноантитела, и ScFv и Fab), являются привлекательным классом терапевтических средств вследствие их потенциально более высокой специфичности, функционального разнообразия и фармакологических свойств. Кроме того, антитела могут лучше взаимодействовать с внеклеточными доменами и петлями, которые могут модулировать структуру (и, таким образом, функцию) связанных с клеточной мембраной белков, таких как GPCR, более сложными путями, чем низкомолекулярные соединения. Однако на сегодняшний день в США не существует ни одного одобренного антительного терапевтического средства против GPCR, и существует только одно в мире, в Японии.
[0066] Современные технологии дрожжевого и фагового дисплея идентифицируют антитела, которые прочно связывают, но часто не влияют на функцию связанных с клеточной мембраной белков, таких как GPCR. Используемые антигены часто представляют собой фрагменты, которые не репрезентируют функциональный белок, доступный для антитела in vivo, или представляют собой гетерогенно структурированные полноразмерные белковые препараты. Эта технология также не учитывает огромную долю общего функционального разнообразия, поскольку большинство антител никогда не подвергают функциональному анализу. Необходима высокопроизводительная платформа для прямой селекции антител, которые модулируют функцию связанных с клеточной мембраной белков, таких как GPCR.
[0067] Является значительно менее простой разработка антител, которые изменяют функцию связанных с клеточной мембраной белков, таких как GPCR (Jo 2015, Hutchings 2010). Это в первую очередь является следствием следующих проблем с множеством существующих решений: 1) отсутствуют используемые антигены. Антигены, происходящие из внеклеточных пептидов или фрагментов могут быть хорошо пригодными для разработки антител для вестерн-блоттинга, но не отражают структурно имеющие терапевтическое значение мишени. Кроме того, однородно функционально свернутый полноразмерный белок в липидах или детергентах может быть трудно получить в достаточных количествах для иммунизации, фагового дисплея или дрожжевого дисплея. 2) Антитела, отобранные вследствие их высокой аффинности, по большей части являются нефункциональными; они связываются с областями в белке, которые не влияют на функцию. 3) Технологии утрачивают значительную часть разнообразия антител и, таким образом, функциональность отобранных антител. Посредством селекции сначала антител, которые прочно связываются, и исключения остальных утрачивается значительная часть функционального разнообразия. Клеточные системы млекопитающих были созданы для функционального скрининга подгрупп антител-кандидатов аутокринным образом (Zhang 2014), которые частично решают проблему 2, но вследствие эффективности трансформации (~104) и ограниченной возможности конструировать подвергаемые селекции/скринингу результаты, они ограничены скринингом небольших подгрупп кандидатов.
[0068] Инновация авторов настоящего изобретения включает комбинирование сопряжения связанного с клеточной мембраной белка с ответом феромонов дрожжей и экспрессии аффинных молекул, имеющих цис-действие в той же клетке, в высокопроизводительной платформе. Это позволяет прямую и высокопроизводительную функциональную селекцию аффинных молекул в периплазматическом пространстве дрожжей. Антитела и родственные аффинные молекулы представляют собой крупные молекулы с комплексными паттернами сворачивания, которые должны сохраняться для сохранения активности связывания. Хотя в неструктурированной форме короткие пептиды могут быть способны локализоваться в периплазматическом пространстве дрожжей, ранее не было известно, что антитела и родственные аффинные молекулы могут быть экспонированы в периплазматическом пространстве дрожжей и сохранять активность связывания.
[0069] Для применения на практике настоящего изобретения могут использоваться, если нет иных указаний, общепринятые способы фармакологии, химии, биохимии, способы рекомбинантных ДНК и иммунологии, относящиеся к данной области. Такие способы полностью объяснены в литературе. См., например, High Throughput Screening: Methods and Protocols (Methods in Molecular Biology, W.P. Janzen ed., Humana Press, 3rd edition, 2016); G Protein-Coupled Receptors: Structure, Signaling, and Physiology (S. Siehler and G. Milligan eds., Cambridge University Press, 2010); Handbook of Experimental Immunology, Vols. I-IV (D.M. Weir and C.C. Blackwell eds., Blackwell Scientific Publications); A.L. Lehninger, Biochemistry (Worth Publishers, Inc., current addition); Sambrook, et al., Molecular Cloning: A Laboratory Manual (3rd Edition, 2001); Methods In Enzymology (S. Colowick and N. Kaplan eds., Academic Press, Inc.).
[0070] Все цитированные в настоящем описании публикации, патенты и патентные заявки, как выше, так и ниже, включены в настоящее описание в качестве ссылок в полном объеме.
I. ОПРЕДЕЛЕНИЯ
[0071] При описании настоящего изобретения используются следующие термины, и подразумевается, что они определяются, как указано ниже.
[0072] Следует отметить, что, как используют в настоящем описании и в прилагаемой формуле изобретения, форма единственного числа включает множественное число, если контекст явно не указывает на иное. Таким образом, например, указание на "клетку" включает смесь двух или более клеток, и т.п.
[0073] Термин "приблизительно", в частности в отношении данного количества, охватывает отклонения в пределах плюс или минус пять процентов.
[0074] Термин "приблизительно", в частности, в отношении данного количества, охватывает и описывает само данное количество.
[0075] Термины "полипептид" и "белок" относятся к полимеру из аминокислотных остатков, и они не ограничиваются минимальной длиной. Таким образом, в определение входят пептиды, олигопептиды, димеры, мультимеры и т.п.Определение охватывает как полноразмерные белки, так и их фрагменты. Термины также включают модификации полипептида после экспрессии, например, гликозилирование, ацетилирование, фосфорилирование, гидроксилирование и т.п.Более того, для целей настоящего изобретения "полипептид" относится к белку, который включает модификации, такие как делеции, вставки и замены в нативной последовательности. Эти модификации могут быть преднамеренными, например, посредством сайт-направленного мутагенеза, или могут быть случайными, такими как путем мутаций у хозяев, которые продуцируют белки, или ошибок в ходе амплификации способом ПЦР.
[0076] Термин "антитело" охватывает моноклональные антитела, а также гибридные антитела, измененные антитела, химерные антитела и гуманизированные антитела. Термин "антитело" включает: гибридные (химерные) молекулы антител (см., например, Winter et al. (1991) Nature 349:293-299; и патент США №4816567); F(ab')2- и F(ab)-фрагменты; Fv-молекулы (нековалентные гетеродимеры, см., например, Inbar et al. (1972) Proc Natl Acad Sci USA 69:2659-2662; и Ehrlich et al. (1980) Biochem 19:4091-4096); одноцепочечные Fv-молекулы (scFv) (см., например, Huston et al. (1988) Proc Natl Acad Sci USA 85:5879-5883); наноантитела или однодоменные антитела (sdAb) (см., например, Wang et al. (2016) Int J Nanomedicine 11:3287-3303, Vincke et al. (2012) Methods Mol Biol 911:15-26; димерные и тримерные конструкции фрагментов антител; миниантитела (см., например, Pack et al. (1992) Biochem 31:1579-1584; Cumber et al. (1992) J Immunology 149B:120-126); гуманизированные молекулы антител (см., например, Riechmann et al. (1988) Nature 332:323-327; Verhoeyan et al. (1988) Science 239:1534-1536; и публикацию патента Великобритании №GB 2276169, опубликованную 21 сентября 1994 года); и любые функциональные фрагменты, получаемые из таких молекул, где такие фрагменты сохраняют свойства специфического связывания исходной молекулы антитела.
[0077] Выражение "специфически (или селективно) связывает" в отношении связывания антитела с антигеном (например, GPCR) относится к реакции связывания, которая определяет присутствие антигена в гетерогенной популяции белков и других биологических агентов. Таким образом, в предусматриваемых условиях иммуноанализа специфические антитела связываются с конкретным антигеном на уровне, по меньшей мере в два раза превышающем фоновый уровень, и по существу не связываются на значительном уровне с другими антигенами, присутствующими в образце. Специфическое связывание с антигеном в таких условиях может требовать антитела, которое отобрано по его специфичности в отношении конкретного антигена. Например, антитела, индуцированные в отношении антигена из конкретного вида, такого как крыса, мышь или человек, можно отбирать с получением только тех антител, которые являются специфически иммунореактивными в отношении антигена, но не других белков, за исключением полиморфных вариантов и аллелей. Эту селекцию можно осуществлять путем исключения антител, которые перекрестно реагируют с молекулами из другого вида. Для селекции антител, специфически иммунореактивных в отношении конкретного антигена, можно использовать различные форматы иммуноанализа. Например, твердофазный иммуноанализ ELISA обычно используют для селекции антител, специфически иммунореактивных в отношении белка (см., например, Harlow & Lane. Antibodies, A Laboratory Manual (1988), для описания форматов иммуноанализа и условий, которые можно использовать для определения специфической иммунореактивности). Как правило, специфическая или селективная реакция по меньшей мере в два раза превышает фоновый сигнал или шум и, более конкретно, более чем в 10-100 раз превышает фон.
[0078] Белок считают "взаимодействующим" с другим белком, если он связывается с ним специфически (например, по механизму типа "замок и ключ"), неспецифически или в некоторой комбинации специфического и неспецифического связывания. Первый белок "предпочтительно взаимодействует" со вторым белком, если он связывается (неспецифически и/или специфически) со вторым белком с большей аффинностью и/или большей специфичностью, чем с другими белками. Термин "аффинность" относится к силе связывания, и она может выражаться количественно в качестве константы диссоциации (Kd). Следует понимать, что специфическое связывание не обязательно требует взаимодействия между конкретными аминокислотными остатками и/или мотивами каждого белка. Например, в определенных вариантах осуществления первый белок может предпочтительно взаимодействовать со вторым белком, но, тем не менее, может быть способен связывать другие полипептиды на слабом, но, поддающемся обнаружению, уровне (например, 10% или менее от связывания, показанного для представляющего интерес полипептида). Как правило, слабое связывание или фоновое связывание без труда отличимо от предпочтительного взаимодействия с представляющим интерес соединением или полипептидом, например, с использованием надлежащих контролей.
[0079] Как используют в рамках изобретения, термин "связывающаяся пара" относится к первой и второй молекулам, которые специфически связываются друг с другом. На "специфическое связывание" первого представителя связывающейся пары со вторым представителем связывающейся пары указывает связывание первого представителя со вторым представителем, или наоборот, с более высокой аффинностью и специфичностью, чем с другими компонентами в образце. Связывание между представителями связывающейся пары, как правило, является нековалентным. Примеры включают связывающиеся пары антиген-антитело, рецептор-гормон, рецептор-лиганд, рецептор-агонист и рецептор-антагонист.
[0080] Как используют в рамках изобретения, термин "лиганд" относится к молекуле, которая связывается с другой молекулой, например, к антигену, связывающемуся с антителом, гормону, агонисту или антагонисту, связывающемуся с рецептором, нейротрансмиттеру, связывающемуся с ионным каналом, или субстрату, ингибитору или аллостерическому эффектору, связывающемуся с ферментом, и включает натуральные и синтетические биомолекулы, такие как белки, полипептиды, пептиды, молекулы нуклеиновых кислот, углеводы, сахара, липиды, липопротеины, низкомолекулярные соединения, натуральные и синтетические органические и неорганические материалы, синтетические полимеры, аптамеры и т.п.
[0081] Термин "полинуклеотид", как известно в данной области, обычно относится к молекуле нуклеиновой кислоты. "Полинуклеотид" может включать как двухцепочечные, так и одноцепочечные последовательности, и относится к, но не ограничиваясь ими, прокариотическим последовательностям, эукариотической мРНК, кДНК из вирусной, прокариотической или эукариотической мРНК, геномным последовательностям РНК и ДНК из вирусов (например, РНК- и ДНК-вирусов и ретровирусов), прокариотической ДНК или эукариотической (например, млекопитающих) ДНК, и особенно синтетическим последовательностям ДНК. Также термин охватывает последовательности, которые включают любые известные аналоги оснований ДНК и РНК, и включает модификации, такие как делеции, вставки и замены (как правило, консервативные по своей природе), в нативной последовательности. Эти модификации могут быть преднамеренными, например, посредством сайт-направленного мутагенеза, или могут быть случайными, например, посредством мутаций у хозяев, включающих полинуклеотиды, кодирующие варианты полипептидов для дисплея. Модификации полинуклеотидов могут иметь любое количество эффектов, включая, например, облегчение экспрессии полипептидного продукта в клетке-хозяине.
[0082] Полинуклеотид может кодировать биологически активный белок или полипептид. В зависимости от природы полипептида, кодируемого полинуклеотидом, полинуклеотид может включать только 10 нуклеотидов, например, где полинуклеотид кодирует антиген или эпитоп.Как правило, полинуклеотид кодирует пептиды по меньшей мере из 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30 или даже больше аминокислот.
[0083] Термины "вариант", "аналог" и "мутеин" относятся к биологически активным производным эталонной молекулы, которые сохраняют желаемую активность (например, эффективный полипептидный дисплей), как описано в настоящем описании. Как правило, термины "вариант" и "аналог" относятся к соединениям, имеющим нативную полипептидную последовательность и структуру с одной или более вставками, заменами и/или делециями аминокислот относительно нативной молекулы при условии, что модификации не нарушают биологическую активность, и которые являются "по существу гомологичными" эталонной молекуле, как определено ниже. Как правило, аминокислотные последовательности таких аналогов имеют высокую степень гомологии последовательности с эталонной последовательностью, например, гомологию аминокислотных последовательностей более 50%, как правило, более 60%-70%, еще более конкретно 80%-85% или более, например, по меньшей мере 90%-95% или более, когда эти две последовательности выравнивают. Часто, аналоги включают то же количество аминокислот, но включают замены, как объяснено в настоящем описании. Термин "мутеин", кроме того, включает полипептиды, имеющие одну или более подобных аминокислотам молекул, включая, но не ограничиваясь ими, соединения, содержащие только амино- и/или имино-молекулы, полипептиды, содержащие один или более аналогов аминокислот (включая, например, неприродные аминокислоты и т.д.), полипептиды с замещенными связями, а также другие модификации, известные в данной области, как встречающиеся в природе, так и не встречающиеся в природе (например, синтетические), циклизованные, разветвленные молекулы и т.п. Также термин включает молекулы, содержащие один или более N-замещенных остатков глицина ("пептоид") и других синтетических аминокислот или пептидов. (См., например, патенты США №5831005; 5877278 и 5977301; Nguyen et al., Chem. Biol. (2000) 7:463-473; и Simon et al., Proc. Natl. Acad. Sci. USA (1992) 89:9367-9371 для описания пептоидов). Способы получения аналогов полипептидов и мутеинов известны в данной области и дополнительно описаны ниже.
[0084] Аналоги, как правило, включают замены, которые являются консервативными по своей природе, т.е. замены, которые происходят в семействе аминокислот, которые являются сходными их боковыми цепями. В частности, аминокислоты, как правило, подразделяют на четыре семейства: (1) кислотные - аспартат и глутамат; (2) основные - лизин, аргинин, гистидин; (3) неполярные - аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженные полярные - глицин, аспарагин, глутамин, цистеин, серин треонин, тирозин. Фенилаланин, триптофан и тирозин иногда классифицируют как ароматические аминокислоты. Например, разумно предположить, что отдельная замена лейцина изолейцином или валином, аспартата глутаматом, треонина серином, или сходная консервативная замена аминокислоты структурно сходной аминокислотой, не будет иметь значительного эффекта на биологическую активность. Например, представляющий интерес полипептид может включать вплоть до приблизительно 5-10 консервативных или неконсервативных аминокислотных замен, или даже вплоть до приблизительно 15-25 консервативных или неконсервативных аминокислотных замен, или любое целое число в диапазоне 5-25, при условии, что желаемая функция молекулы остается интактной. Специалист в данной области может легко определить области представляющей интерес молекулы, которые могут допускать изменение согласно графикам Хоппа/Вудса и Кайта-Дулиттла, хорошо известным в данной области.
[0085] "Рекомбинантный", как используют в рамках изобретения для описания молекулы нуклеиновой кислоты, означает полинуклеотид геномного, кДНК, вирусного, полусинтетического или синтетического происхождения, который, вследствие его происхождения или посредством манипуляции, не ассоциирован со всем или частью полинуклеотида, с которым он ассоциирован в природе. Термин "рекомбинантный", используемый в отношении белка, полипептида или пептида, означает полипептид, получаемый посредством экспрессии рекомбинантного полинуклеотида. Как правило, представляющий интерес ген клонируют, а затем экспрессируют в трансформированных организмах, как дополнительно описано ниже. Организм-хозяин экспрессирует чужеродный ген для продуцирования белка в условиях экспрессии.
[0086] "Полинуклеотидная кодирующая последовательность" или последовательность, которая "кодирует" отдельный полипептид, представляет собой молекулу нуклеиновой кислоты, которая транскрибируется (в случае ДНК) и транслируется (в случае мРНК) в полипептид in vivo, когда она находится под контролем надлежащих регуляторных последовательностей (или "элементов контроля"). Границы кодирующей последовательности определяются инициирующим кодоном на 5'(амино)-конце и стоп-кодоном трансляции на 3'(карбокси)-конце. Последовательность терминации транскрипции может находиться с 3'-стороны от кодирующей последовательности. Типичные "элементы контроля" включают, но не ограничиваются ими, регуляторы транскрипции, такие как промоторы, транскрипционные энхансерные элементы, сигналы терминации транскрипции и последовательности полиаденилирования; и регуляторы трансляции, такие как последовательности для оптимизации инициации трансляции, например, последовательности Шайна-Дальгарно (участок связывания рибосом), последовательности Козака (т.е. последовательности для оптимизации трансляции, находящиеся, например, с 5'-стороны от кодирующей последовательности), лидерные последовательности (гетерологичные или нативные), кодон инициации трансляции (например, ATG) и последовательности терминации трансляции. Промоторы могут включать индуцибельные промоторы (где экспрессия полинуклеотидной последовательности, функционально связанной с промотором, индуцируется анализируемой молекулой, кофактором, регуляторным белком и т.д.), репрессируемые промоторы (где экспрессия полинуклеотидной последовательности, функционально связанной с промотором, индуцируется анализируемой молекулой, кофактором, регуляторным белком и т.д.) и конститутивные промоторы.
[0087] "Функционально связанный" относится к расположению элементов, где компоненты, описанные таким образом, организованы для выполнения их обычной функции. Таким образом, данный промотор, функционально связанный с кодирующей последовательностью, способен обеспечивать экспрессию кодирующей последовательности, когда присутствуют надлежащие ферменты. Промотор не должен быть смежным с кодирующей последовательностью, при условии, что он функционирует, запуская ее экспрессию. Таким образом, например, между промоторной последовательностью и кодирующей последовательностью могут присутствовать промежуточные нетранслируемые, но транскрибируемые последовательности, и промоторная последовательность все еще может считаться "функционально связанной" с кодирующей последовательностью.
[0088] Под "фрагментом" подразумевают молекулу, состоящую только из части интактной полноразмерной последовательности и структуры. Фрагмент может включать C-концевую делецию и N-концевую делецию, и/или внутреннюю делецию в пептиде. Активные фрагменты конкретного белка или пептида, как правило, включают по меньшей мере приблизительно 5-10 последовательно расположенных аминокислотных остатков полноразмерной молекулы, предпочтительно по меньшей мере приблизительно 15-25 последовательно расположенных аминокислотных остатков полноразмерной молекулы, и наиболее предпочтительно по меньшей мере приблизительно 20-50 или более последовательно расположенных аминокислотных остатков полноразмерной молекулы, или любое целое число от 5 аминокислот до полноразмерной последовательности, при условии, что рассматриваемый фрагмент сохраняет биологическую активность.
[0089] "По существу очищенный", главным образом, относится к выделению вещества (соединение, полинуклеотид, белок, полипептид, композиция полипептидов), так что вещество составляет основной процент образца, в котором оно находится. Как правило, в образце по существу очищенный компонент составляет 50%, предпочтительно 80%-85%, более предпочтительно 90-95% образца. Способы очистки представляющих интерес полинуклеотидов и полипептидов хорошо известны в данной области и включают, например, ионообменную хроматографию, аффинную хроматографию и седиментацию по плотности.
[0090] Под "выделенным" подразумевают, при указании на полипептид, что указанная молекула отделена и обособлена от целого организма, в котором она находится в природе, или присутствует по существу в отсутствии других биологических макромолекул того же типа. Термин "выделенный" в отношении полинуклеотида представляет собой молекулу нуклеиновой кислоты, лишенную, целиком или частично, последовательностей, обычно ассоциированных с ней в природе; или последовательность, как она существует в природе, но имеющую гетерологичные последовательности, связанные с ней; или молекулу, отделенную от хромосомы.
[0091] "Гомология" относится к процентной идентичности между двумя полинуклеотидными или двумя полипептидными молекулами. Две последовательности нуклеиновых кислот или две полипептидных последовательности являются "по существу гомологичными" друг другу, когда последовательности демонстрируют по меньшей мере приблизительно 50%, предпочтительно по меньшей мере приблизительно 75%, более предпочтительно по меньшей мере приблизительно 80%-85%, предпочтительно по меньшей мере приблизительно 90%, и наиболее предпочтительно по меньшей мере приблизительно 95%-98% идентичность последовательностей на протяжении определенной длины молекул. Как используют в рамках изобретения, по существу гомологичный также относится к последовательностям, демонстрирующим полную идентичность с указанной последовательностью.
[0092] Как правило, "идентичность" относится к точному соответствию нуклеотид-нуклеотид или аминокислота-аминокислота двух полинуклеотидных или полипептидных последовательностей, соответственно. Процентную идентичность можно определять посредством прямого сравнения информации о последовательности между двумя молекулами (эталонная последовательность и последовательность с неизвестной % идентичностью с эталонной последовательностью) путем выравнивания последовательностей, подсчета точного количества совпадений между двумя выровненными последовательностями, деления на длину эталонной последовательности и умножения результата на 100. Для облегчения анализа можно использовать хорошо доступные компьютерные программы, такие как ALIGN, Dayhoff, M.O., Atlas of Protein Sequence and Structure M.O. Dayhoff ed., 5 Suppl. 3:353-358, National biomedical Research Foundation, Washington, DC, в которой используется алгоритм локальной гомологии Smith и Waterman, Advances in Appl. Math. 2:482-489, 1981, для анализа пептидов. Программы для определения идентичностей нуклеотидных последовательностей доступны в Wisconsin Sequence Analysis Package, Version 8 (доступном от Genetics Computer Group, Madison, WI), например, программы BESTFIT, FASTA и GAP, которые также основаны на алгоритме Smith и Waterman. Эти программы можно без труда использовать с параметрами по умолчанию, рекомендованными изготовителем, и они описаны в Wisconsin Sequence Analysis Package, упоминаемом выше. Например, процентную идентичность конкретной нуклеотидной последовательности с эталонной последовательностью можно определять с использованием алгоритма гомологии Smith и Waterman с оценочной таблицей по умолчанию и штрафом за делецию, составляющим шесть нуклеотидных положений.
[0093] Другим способом установления процентной идентичности в контексте настоящего изобретения является использование пакета программ MPSRCH, авторские права на который имеет University of Edinburgh, разработанный John F. Collins и Shane S. Sturrok, и распространяемый IntelliGenetics, Inc. (Mountain View, CA). В этом пакете программ используется алгоритм Smith-Waterman, где используются параметры по умолчанию для оценочной таблицы (например, штраф за внесение пропуска 12, штраф за продолжение пропуска, равный одному, и пропуск, равный шести). Из полученных данных величина "Match" отражает "идентичность последовательностей". Другие подходящие программы для вычисления процентной идентичности или сходства между последовательностями широко известны в данной области, например, другой программой для выравнивания является BLAST, используемая с параметрами по умолчанию. Например, BLASTN и BLASTP можно использовать с использованием следующих параметров по умолчанию: генетический код=стандартный; фильтр=нет; цепь=оби; пороговое значение=60; ожидание=10; матрица=BLOSUM62; описания=50 последовательностей; сортировка по=наивысшей показатель; базы данных=неизбыточная GenBank+EMBL+DDBJ+PDB+GenBank CDS translations+Swiss protein+Spupdate+PIR. Детальное описание этих программ свободно доступно.
[0094] Альтернативно гомологию можно определять гибридизацией полинуклеотидов в условиях, которые образуют стабильные дуплексы между гомологичными областями с последующим расщеплением специфичной к одноцепочечной последовательности нуклеазой(ами), и определением размера расщепленных фрагментов. Последовательности ДНК, которые являются по существу гомологичными, можно идентифицировать в эксперименте с использованием Саузерн-гибридизации, например, в жестких условиях, как определяют для этой конкретной системы. Определение подходящих условий гибридизации входит в пределы способностей специалиста в данной области. См., например, Sambrook et al., выше; DNA Cloning, выше; Nucleic Acid Hybridization, выше.
[0095] "Экспрессирующая кассета" или "экспрессирующая конструкция" относится к системе, которая способна направлять экспрессию представляющей интерес последовательности(ей) или гена(ов). Экспрессирующая кассета, как правило, включает элементы контроля, как описано выше, такие как промотор, который функционально связан с (т.е. направляет транскрипцию) представляющей интерес последовательностью(ями) или геном(ами), и также часто включает последовательность полиаденилирования. В определенных вариантах осуществления изобретения экспрессирующая кассета, описанная в настоящем описании, может находиться в плазмиде или вирусной векторной конструкции. В дополнение к компонентам экспрессирующей кассеты, конструкция также может включать один или более селективных маркеров, сигнал, который позволяет конструкции существовать в качестве одноцепочечной ДНК (например, ориджин репликации M13), по меньшей мере один участок множественного клонирования и ориджин репликации (например, автономно реплицирующаяся последовательность в дрожжах).
[0096] Термин "трансфекция" используют для обозначения захвата чужеродной ДНК клеткой. Клетка является "трансфицированной", когда экзогенная ДНК введена внутрь клеточной мембраны. Ряд способов трансфекции широко известен в данной области. См., например, Graham et al. (1973) Virology, 52:456, Sambrook et al. (2001) Molecular Cloning, a laboratory manual, 3rd edition, Cold Spring Harbor Laboratories, New York, Davis et al. (1995) Basic Methods in Molecular Biology, 2nd edition, McGraw-Hill, and Chu et al. (1981) Gene 13:197. Такие способы можно использовать для введения одной или более экзогенных частей ДНК в подходящие клетки-хозяева. Термин относится как к стабильному, так и к временному захвату генетического материала, и он включает захват связанных с пептидом или антителом ДНК.
[0097] "Вектор" способен переносить последовательности нуклеиновых кислот в клетки-мишени (например, вирусные векторы, невирусные векторы, носители в форме частиц и липосомы). Как правило, "векторная конструкция", "экспрессирующий вектор" и "вектор для генного переноса" означают любую конструкцию нуклеиновой кислоты, которая способна направлять экспрессию представляющей интерес нуклеиновой кислоты и которая может переносить последовательности нуклеиновых кислот в клетки-мишени. Таким образом, термин включает клонирующие и экспрессирующие носители, а также плазмидные и вирусные векторы.
[0098] Термин "трансформация" относится к введению экзогенного полинуклеотида в клетку-хозяина независимо от способа, используемого для встраивания. Например, включены прямой захват, трансдукция или f-скрещивание. Экзогенный полинуклеотид может оставаться в качестве не встроенного вектора, например плазмиды, или альтернативно он может встраиваться в геном хозяина.
[0099] "Рекомбинантные клетки-хозяева", "клетки-хозяева", "клетки", "клеточные линии", "клеточные культуры" и другие такие термины, обозначающие микроорганизмы или линии эукариотических клеток, культивируемых в качестве единичных клеток, относятся к клеткам, которые могут быть использованы или используются в качестве реципиентов для рекомбинантного вектора или другой перенесенной ДНК, и включают оригинальное потомство оригинальной клетки, которая трансфицирована.
[0100] "Кодирующая последовательность" или последовательность, которая "кодирует" определенный полипептид, представляет собой молекулу нуклеиновой кислоты, которая транскрибируется (в случае ДНК) и транслируется (в случае мРНК) в полипептид in vivo, когда она находится под контролем соответствующих регуляторных последовательностей (или "элементов контроля"). Границы кодирующей последовательности могут определяться инициирующим кодоном на 5'(амино)-конце и стоп-кодоном трансляции на 3'(карбокси)-конце. Кодирующая последовательность может включать, но не ограничивается ими, кДНК из вирусной, прокариотической или эукариотической мРНК, геномные последовательности ДНК из вирусной или прокариотической ДНК, и даже синтетические последовательности ДНК. Последовательность терминации транскрипции может находиться с 3'-стороны от кодирующей последовательности.
[0101] Типичные "элементы контроля" включают, но не ограничиваются ими, промоторы транскрипции, энхансерные элементы транскрипции, сигналы терминации транскрипции, последовательности полиаденилирования (находящиеся с 3'-стороны от стоп-кодона трансляции), последовательности оптимизации инициации трансляции (находящиеся с 5'-стороны от кодирующей последовательности), и последовательности терминации трансляции.
[0102] Термины "метка" и "поддающаяся определению метка" относятся к молекуле, способной осуществляют детекцию, включая, но не ограничиваясь ими, радиоактивннвые изотопы, стабильные (нерадиоактивные) тяжелые изотопы, флуорофоры, хемилюминесцеры, ферменты, субстраты ферментов, кофакторы ферментов, ингибиторы ферментов, хромофоры, красители, ионы металлов, лиганды (например, биотин или гаптены) и т.п. Термин "флуорофор" относится к веществу или его части, которые способны к флуоресценции в поддающемся детекции диапазоне. Конкретные примеры меток, которые можно использовать в рамках изобретения, включают, но не ограничиваются ими, радиоактивные метки (например, 3H, 125I, 35S, 14C или 32P), стабильные (нерадиоактивные) тяжелые изотопы (например, 13C или 15N), фикоэритрин, флуоресцеин, 7-нитробензо-2-окса-1,3-диазол (NBD), YPet, CyPet, Cascade blue, аллофикоцианин, красители Alexa (например, Alexa 350, Alexa 430, Alexa 488, Alexa 532, Alexa 546, Alexa 555, Alexa 594, Alexa 647, Alexa 660, Alexa 680 и Alexa 750), красители Atto (например, Atto 488, Atto 532, Atto 550, Atto 565, Atto 590, Atto 610, Atto 620, Atto 635, Atto 647, Atto 655 и Atto 680), цианиновые красители (например, Cy3, Cy5 и Cy7), TYE 563, TYE 665, TYE 705, TEX 615, JOE, TET, HEX, TAMRA, ROX, родамин, дансил, умбеллиферон, техасский красный, люминол, сложные эфиры акрадимия, биотин или другие стрептавидин-связывающие белки, магнитные гранулы, электронноплотные реагенты, зеленый флуоресцентный белок (GFP), усиленный зеленый флуоресцентный белок (EGFP), желтый флуоресцентный белок (YFP), усиленный желтый флуоресцентный белок (EYFP), синий флуоресцентный белок (BFP), красный флуоресцентный белок (RFP), TagRFP, Dronpa, Padron, mApple, mCherry, rsCherry, rsCherryRev, люциферазу светлячка, люциферазу Renilla, NADPH, бета-галактозидазу, пероксидазу хрена, глюкозооксидазу, щелочную фосфатазу, хлорамфениколацетилтрансферазу и уреазу. Ферментные метки используют с их собственным субстратом. Как и в случае многих стандартных методик, ассоциированных с применением изобретения на практике, квалифицированному специалисту известны дополнительные метки, которые можно использовать.
II. СПОСОБЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
[0103] Перед подробным описанием настоящего изобретения следует понимать, что настоящее изобретение не ограничивается конкретными составами или параметрами процесса, поскольку они, безусловно, могут варьироваться. Также следует понимать, что терминология, приведенная в настоящем описании, приведена только для описания конкретных вариантов осуществления изобретения и не является ограничивающей.
[0104] Хотя при применении настоящего изобретения на практике можно использовать ряд способов и материалов, сходных или эквивалентных способам и материалам, описанным в настоящем описании, предпочтительные материалы и способы описаны в настоящем описании.
[0105] Настоящее изобретение основано на разработке способов дисплея рекомбинантных белков в периплазматическом пространстве дрожжевых клеток. В частности, рекомбинантные белки связаны с проходящим через клеточную мембрану трансмембранным доменом, связанным с клеточной мембраной белковым доменом, который находится на наружной поверхности мембраны дрожжевых клеток, белком, который связывается с внутренней поверхностью клеточной стенки дрожжей, или периплазматическим белком для дисплея белков в периплазматическом пространстве дрожжей. Рекомбинантные белки также могут быть нацелены на периплазму путем связывания рекомбинантного белка с секреторным сигналом. Кроме того, в дрожжах может быть коэкспрессирован представляющий интерес белок-мишень так, чтобы он локализовался на плазматической мембране или в периплазматическом пространстве, и был доступен для экспонированных белков. В конкретных вариантах осуществления авторы изобретения использовали их способ дрожжевого периплазматического дисплея для скрининга антител, которые связывают и модулируют функцию GPCR человека (см. примеры). Антитела, экспонированные в периплазматическом пространстве дрожжевой клетки, находятся достаточно близко для связывания GPCR, экспрессируемого на клеточной мембране. Авторы изобретения, кроме того, разработали способ высокопроизводительного скрининга GPCR в отношении антагонистов и агонистов с использованием периплазматического дисплея посредством сопряжения GPCR человека с каскадом ответа феромонов дрожжей.
[0106] Для дальнейшего понимания изобретения ниже приведено более детальное обсуждение, касающееся дрожжевого периплазматического дисплея и способов его применения для высокопроизводительного скрининга библиотек белков.
A. Периплазматический дисплей рекомбинантных белков в дрожжах
[0107] В одном аспекте изобретение относится к дисплею белка в периплазматическом пространстве дрожжевой клетки-хозяина. В некоторых вариантах осуществления дрожжевая клетка-хозяин содержит белок для дисплея в периплазматическом пространстве дрожжей, периплазматический якорный белок, связанный с белком, подлежащим дисплею в периплазматическом пространстве, и представляющий интерес мембранный белок-мишень, находящийся в периплазматическом пространстве дрожжевой клетки-хозяина. В некоторых вариантах осуществления дрожжевую клетку-хозяина можно использовать для определения того, связывается ли белок, подлежащий дисплею в периплазматическом пространстве, специфически с представляющим интерес мембранным белком-мишенью, или влияет ли он на его функцию. Белок, подлежащий дисплею в периплазматическом пространстве дрожжевой клетки-хозяина, можно получать путем связывания рекомбинантного белка с периплазматическим якорным белком, который обеспечивает локализацию рекомбинантного белка в периплазматическом пространстве дрожжевой клетки. Связь может быть ковалентной или нековалентной. Например, рекомбинантный белок может быть ковалентно связан с периплазматическим якорным белком в слитом белке. Альтернативно рекомбинантный вариант белка может образовывать комплекс с периплазматическим якорным белком, где рекомбинантный белок и периплазматический якорный белок нековалентно связаны посредством молекулярных связывающих взаимодействий в комплексе. Альтернативно рекомбинантный вариант белка и периплазматический якорный белок связаны ковалентно посредством непептидной связи в комплексе. В некоторых вариантах осуществления непептидная связь представляет собой дисульфидную связь. Белок, подлежащий дисплею в периплазматическом пространстве дрожжевой клетки-хозяина, также можно получать путем связывания белка, подлежащего дисплею, с секреторным сигналом. В некоторых вариантах осуществления род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia. В некоторых вариантах осуществления род дрожжевой клетки-хозяина представляет собой Saccharomyces. В некоторых вариантах осуществления вид дрожжевой клетки-хозяина представляет собой Saccharomyces cerevisiae.
[0108] В другом аспекте изобретение относится к антителу, связанному с периплазматическим якорным белком. В некоторых вариантах осуществления антитело, связанное с периплазматическим якорным белком, кроме того, содержит дополнительную модификацию, часть или взаимодействующий белок. В некоторых вариантах осуществления дополнительная модификация представляет собой посттрансляционную модификацию. В некоторых вариантах осуществления часть представляет собой аффинную метку, эпитоп, маркировку и т.п.В некоторых вариантах осуществления антитело локализуется в периплазматическом пространстве дрожжевой клетки-хозяина. В некоторых вариантах осуществления, когда антитело продуцируется в дрожжевой клетке-хозяине или введено в нее, антитело локализуется в периплазматическом пространстве дрожжевой клетки-хозяина. В некоторых вариантах осуществления антитело связано с периплазматическим якорным белком, так что антитело локализуется в периплазматическом пространстве. Связь антитела с периплазматическим якорным белком может быть ковалентной или нековалентной. Например, антитело может быть ковалентно связано с периплазматическим якорным белком в слитом белке. Альтернативно антитело может образовывать комплекс с периплазматическим якорным белком, где антитело и периплазматический якорный белок связаны нековалентно посредством молекулярных связывающих взаимодействий в комплексе. Альтернативно антитело и периплазматический якорный белок связаны ковалентно посредством непептидной связи в комплексе. В некоторых вариантах осуществления непептидная связь представляет собой дисульфидную связь. Также предусматриваются дрожжевые клетки-хозяева, содержащие антитело, как описано в настоящем описании. В некоторых вариантах осуществления род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia. В некоторых вариантах осуществления род дрожжевой клетки-хозяина представляет собой Saccharomyces. В некоторых вариантах осуществления вид дрожжевой клетки-хозяина представляет собой Saccharomyces cerevisiae.
[0109] В другом аспекте изобретение относится к способам локализации антитела в периплазматическом пространстве дрожжевой клетки-хозяина, включающим связывание антитела с периплазматическим якорным белком, так чтобы антитело было локализовано в периплазматическом пространстве. В некоторых вариантах осуществления антитело связано с периплазматическим якорным белком так, что антитело локализуется в периплазматическом пространстве. Связь антитела с периплазматическим якорным белком может быть ковалентной или нековалентной. Например, антитело может быть ковалентно связано с периплазматическим якорным белком в слитом белке. Альтернативно антитело может образовывать комплекс с периплазматическим якорным белком, где антитело и периплазматический якорный белок связаны нековалентно посредством молекулярных связывающих взаимодействий в комплексе. Альтернативно антитело и периплазматический якорный белок связаны ковалентно посредством непептидной связи в комплексе. В некоторых вариантах осуществления непептидная связь представляет собой дисульфидную связь. В некоторых вариантах осуществления род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia. В некоторых вариантах осуществления род дрожжевой клетки-хозяина представляет собой Saccharomyces. В некоторых вариантах осуществления вид дрожжевой клетки-хозяина представляет собой Saccharomyces cerevisiae.
[0110] В другом аспекте изобретение относится к способам высокопроизводительного скрининга белковых библиотек в отношении специфического связывания или функциональных характеристик посредством дисплея белков в периплазматическом пространстве дрожжей. Библиотеку дрожжевого периплазматического дисплея можно получать путем связывания рекомбинантных белков с периплазматическим якорным белком, который локализует рекомбинантные белки в периплазматическом пространстве дрожжевой клетки. Связь может быть ковалентной или нековалентной. Например, рекомбинантный белок может быть ковалентно связан с периплазматическим якорным белком в слитом белке. Альтернативно рекомбинантный вариант белка может образовывать комплекс с периплазматическим якорным белком, где рекомбинантный белок и периплазматический якорный белок связаны нековалентно посредством молекулярных связывающих взаимодействий в комплексе. Альтернативно рекомбинантный вариант белка и периплазматический якорный белок связаны ковалентно посредством непептидной связи в комплексе. В некоторых вариантах осуществления непептидная связь представляет собой дисульфидную связь. Библиотеку дрожжевого периплазматического дисплея также можно получать путем связывания рекомбинантных белков с секреторными сигналами. В некоторых вариантах осуществления род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia. В некоторых вариантах осуществления род дрожжевой клетки-хозяина представляет собой Saccharomyces. В некоторых вариантах осуществления вид дрожжевой клетки-хозяина представляет собой Saccharomyces cerevisiae.
[0111] Локализации в периплазматическом пространстве можно достигать различными способами. В определенных вариантах осуществления рекомбинантный белок направляют в периплазму путем связывания рекомбинантного белка с секреторным сигналом, который приводит к секреции рекомбинантного белка во внеклеточное пространство. В определенных вариантах осуществления периплазматический якорный белок содержит сигнальную последовательность, которая обеспечивает транспорт периплазматического якорного белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что связанный рекомбинантный экспонируется в периплазме. Например, периплазматический якорный белок может содержать сигнальную последовательность, которая направляет транспорт периплазматического якорного белка и связанного рекомбинантного белка в периплазму дрожжевой клетки-хозяина. Предпочтительно, периплазматический якорный белок является достаточно большим, чтобы периплазматический якорный белок и связанный рекомбинантный белок удерживались в периплазме. Альтернативно, периплазматический якорный белок может быть компонентом периплазматического белкового комплекса, который является достаточно большим, чтобы образование комплекса в периплазме приводило к задерживанию в периплазме. В других вариантах осуществления периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или ассоциированный с мембраной белковый домен, который локализуется на наружной поверхности клеточной мембраны, так что связанный рекомбинантный белок выступает в периплазму. Например, для этой цели можно использовать заякоривающий гликозилфосфатидилинозитол(GPI)-домен, который локализуется на плазматической мембране. В одном варианте осуществления заякоривающий на плазматической мембране GPI-домен представляет собой заякоривающий на плазматической мембране GPI-домен япсина, такой как, но не ограничиваясь ими, заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7. В другом варианте осуществления периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что экспонированный вариант белка выступает в периплазму. В определенных вариантах осуществления периплазматический якорный белок и рекомбинантный белок ковалентно связаны в слитом варианте белка, и периплазматический якорный белок содержит сигнальную последовательность, которая направляет транспорт слитого белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что слитый вариант белка экспонируется в периплазме. Например, периплазматический якорный белок может содержать сигнальную последовательность, которая направляет транспорт слитого белка в периплазму дрожжевой клетки-хозяина. Предпочтительно, периплазматический якорный белок является достаточно большим, чтобы слитый белок удерживался в периплазме. Альтернативно периплазматический якорный белок может быть компонентом периплазматического белкового комплекса, который является достаточно большим, чтобы образование комплекса в периплазме приводило к задерживанию слитого белка в периплазме. В других вариантах осуществления периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или ассоциированный с мембраной белковый домен, который локализуется на наружной поверхности клеточной мембраны, так что слитый вариант белка выступает в периплазму. Например, для этой цели можно использовать заякоривающий гликозилфосфатидилинозитол(GPI)-домен, который локализуется на плазматической мембране. В одном варианте осуществления заякоривающий на плазматической мембране GPI-домен представляет собой заякоривающий на плазматической мембране GPI-домен япсина, такой как, но не ограничиваясь ими, заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7. В другом варианте осуществления периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что экспонированный вариант белка выступает в периплазму. В определенных вариантах осуществления периплазматический якорный белок представляет собой фрагмент полноразмерного белка, который сохраняет способность к локализации в периплазме.
[0112] Любой тип белка может экспонироваться в периплазматическом пространстве дрожжевой клетки, включая, но не ограничиваясь ими, антитела, миметики антител, аптамеры, антигены, ферменты, рецепторы, переносчики, ионные каналы, гормоны, субстраты, агонисты, антагонисты или лиганды. Библиотека дрожжевого периплазматического дисплея презентирует множество таких белков, которые можно подвергать скринингу в отношении сворачивания и/или биологической активности в присутствии представляющей интерес молекулы-мишени. При внеклеточном расположении молекула-мишень должна быть способна проникать через клеточную стенку дрожжей для достижения экспонированных белков для скрининга. В определенных вариантах осуществления представляющий интерес белок-мишень коэкспрессируется с экспонированным вариантом белка в дрожжевой клетке в положении, доступном для экспонированного варианта белка (например, достаточно близко, чтобы экспонированный вариант белка связывал и/или модулировал активность представляющего интерес белка-мишени). Например, представляющий интерес белок-мишень может быть локализован на плазматической мембране дрожжевой клетки-хозяина или в периплазме вблизи места, где экспонируется вариант белка. В частности, этот способ применим для рецепторов, ионных каналов, переносчиков и других мембранных белков, которые локализуются на плазматической мембране. Таким образом, дрожжевой периплазматический дисплей можно использовать для презентации белков мембранному белку-мишени в среде, по существу сходной с нативной средой.
[0113] Любые полипептиды, включенные в слитую конструкцию, включая периплазматический якорный белок и экспонированный вариант белка, могут быть соединены прямо друг с другом пептидными связями или могут быть отделены промежуточными аминокислотными последовательностями или линкерами. Линкерные аминокислотные последовательности, как правило, являются короткими, например, 20 или менее аминокислот (т.е. 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1). Примеры включают короткие пептидные последовательности, которые облегчают клонирование, полиглициновые линкеры (Glyn, где n=2, 3, 4, 5, 6, 7, 8, 9, 10 или более), гистидиновые метки (Hisn, где n=3, 4, 5, 6, 7, 8, 9, 10 или более), линкеры, состоящие из остатков глицина и серина, где n=1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или более), линкеры GSAT, SEG и Z-EGFR. Линкеры могут включать участки рестрикции, которые облегчают клонирование и манипулирование. Другие подходящие линкерные аминокислотные последовательности известны специалистам в данной области (см., например, Argos (1990) J. Mol. Biol. 211(4):943-958; Crasto et al. (2000) Protein Eng. 13:309-312; George et al. (2002) Protein Eng. 15:871-879; Arai et al. (2001) Protein Eng. 14:529-532; и Registry of Standard Biological Parts (partsregistry.org/Protein_domains/Linker).
[0114] Необязательно, в слитые конструкции может быть включена метка. Метки, которые можно использовать для применения изобретения на практике, включают, но не ограничиваются ими, His-метку, Strep-метку, TAP-метку, S-метку, SBP-метку, Arg-метку, кальмодулин-связывающую пептидную метку, метку на основе целлюлоза-связывающего домена, DsbA-метку, метку c-myc, метку на основе глутатион S-трансферазы, FLAG-метку, HAT-метку, метку на основе мальтоза-связывающего белка, NusA-метку и метку на основе тиоредоксина.
B. Полинуклеотиды, кодирующие заякоренные в периплазме варианты белка и белки-мишени, и конструирование библиотек
[0115] Полинуклеотиды, кодирующие заякоренные в периплазме варианты белка и представляющие интерес белки-мишени, можно получать рядом способов, все из которых хорошо известны в данной области. Например, полинуклеотиды можно получать с использованием рекомбинантных способов, хорошо известных в данной области. Специалист в данной области может без труда определить нуклеотидные последовательности, которые кодируют желаемые белки, белки с использованием стандартной методологии и идей, описанных в настоящем описании.
[0116] Олигонуклеотидные зонды могут быть установлены на основе известных последовательностей генов и могут быть использованы для исследования геномных или кДНК-библиотек. Затем полинуклеотиды с желаемыми последовательностями можно далее выделять с использованием стандартных способов и, например, использовать ферменты рестрикции для укорочения гена в желаемых участках полноразмерной последовательности. Аналогично, полинуклеотиды с представляющими интерес последовательностями можно выделять прямо из клеток и тканей, содержащих их, с использованием известных способов, таких как экстракция фенолом, и последовательность можно далее подвергать манипулированию для получения желаемых вариантов белков. См., например, Sambrook et al., выше, для описания способов, используемых для получения и выделения ДНК.
[0117] Последовательности, кодирующие варианты белков, также можно получать синтетическим путем, например, на основе известных последовательностей. Нуклеотидную последовательность можно конструировать с соответствующими кодонами для конкретной желаемой аминокислотной последовательности. Полную последовательность, как правило, собирают из перекрывающихся олигонуклеотидов, полученных стандартными способами, и собирают в полную кодирующую последовательность. См., например, Edge (1981) Nature 292:756; Nambair et al. (1984) Science 223:1299; Jay et al. (1984) J. Biol. Chem. 259:6311; Stemmer et al. (1995) Gene 164:49-53.
[0118] Рекомбинантные способы без труда используют для клонирования последовательностей, кодирующих варианты белков (например, антитела), пригодные для заявленного изобретения, которые затем можно подвергать мутагенезу in vitro путем замены соответствующий пары(пар) оснований с получением кодона для желаемой аминокислоты. Такое изменение может включать всего одну пару оснований, обеспечивая изменение одной аминокислоты, или оно может охватывать изменение нескольких пар оснований. Альтернативно мутации можно осуществлять с использованием праймера с несоответствиями, который гибридизуется с исходной нуклеотидной последовательностью (как правило, кДНК, соответствующая последовательности РНК), при температуре ниже температуры плавления дуплекса с несоответствиями. Специфичность праймера можно обеспечивать путем поддержания длины праймера и композиции оснований в относительно узких пределах и путем расположения мутантного основания по центру. См., например, Innis et al, (1990) PCR Applications: Protocols for Functional Genomics; Zoller and Smith, Methods Enzymol. (1983) 100:468. Удлинения праймеров достигают с использованием ДНК-полимерзы, продукт клонируют и клоны, содержащие мутантную ДНК, полученную путем сегрегации удлиненной цепи праймера, отбирают. Селекцию можно проводить с использованием мутантного праймера в качестве зонда для гибридизации. Этот способ также применим для внесения множества точковых мутаций. См, например, Dalbie-McFarland et al. Proc. Natl. Acad. Sci. USA (1982) 79:6409.
[0119] Разнообразие библиотеки дисплея зависит от используемого способа мутагенеза. Для быстрого получения большого количества мутаций можно использовать кассетный мутагенез путем встраивания мутагенных кассет в нуклеиновую кислоту (см., например, Worrall (1994) Methods Mol. Biol. 30:199-210, Kegler-Ebo et al. (1994) Nucleic Acids Research. 22 (9):1593-1599). Кроме того, также для получения большого количества вариантов для библиотеки дисплея можно использовать случайный мутагенез. Подходящие способы случайного мутагенеза включают, но не ограничиваются ими, ПЦР с пониженной точностью, ПЦР по типу катящегося кольца с пониженной точностью, химический мутагенез, мутагенез в штамме-мутаторе с дефектом каскадов репарации ДНК, инсерционный мутагенез с использованием системы транспозонов, или шаффлинг ДНК (см., например, McCullum et al. (2010) Methods Mol. Biol. 634:103-109, Fujii et al. (2014) Methods Mol. Biol. 1179:23-29, Muteeb (2010) Methods Mol. Biol. 634:411-419, Bose (2016) Methods Mol. Biol. 1373:111-115, Labrou (2010) Curr Protein Pept Sci. 11(1):91-100, Wilson et al. (2011) Methods Mol. Biol. 765:359-371). Такие способы можно использовать для эффективного получения большого количества вариантов с модификациями исходной молекулы нуклеиновой кислоты. В некоторых вариантах осуществления конструируют библиотеку ДНК, содержащую по меньшей мере 106, предпочтительно по меньшей мере 108, и более предпочтительно по меньшей мере 1010 вариантов с уникальными последовательностями, с использованием способов, известных в данной области.
[0120] В некоторых вариантах осуществления библиотеки антител для дрожжевого периплазматического дисплея конструируют путем клонирования природных антител из B-лимфоцитов, полученных от доноров крови. Нуклеиновые кислоты, кодирующие легкие и тяжелые цепи антител или их фрагменты, содержащие определяющие комплементарность области вариабельных доменов (например, Fab), можно амплифицировать способом ПЦР и клонировать в векторы. ScFv-антитела можно получать путем клонирования в вектор, который соединяет легкую и тяжелую цепи через линкер в одной открытой рамке считывания. Донор крови может быть любого вида. В некоторых вариантах осуществления для получения библиотеки антител человека используют доноров крови-людей. В других вариантах осуществления для получения библиотеки верблюжьих антител используют доноров крови, относящихся к семейству верблюжьих. Верблюжьи антитела могут происходить, например, из дромадеров, двугорбых верблюдов, лам или альпак. Такие животные семейства верблюжьих продуцируют уникальный тип антител, которые лишены легкой цепи. Эти антитела с тяжелыми цепями (HCAb) или их фрагменты вариабельных доменов (например, однодоменные антитела или наноантитела) можно использовать для конструирования библиотеки антител (см., например, Vincke et al. (2012) Methods Mol. Biol. 911:15-26, Krah et al. (2016) Immunopharmacol. Immunotoxicol. 38(1):21-8; включенную в настоящее описание в качестве ссылки).
[0121] После выделения и/или синтеза кодирующих последовательностей для вариантов белков (например, антител), их можно клонировать в любой подходящий вектор или репликон для экспрессии в дрожжах. "Вектор" представляет собой композицию, которую можно использовать для доставки представляющей интерес нуклеиновой кислоты внутрь клетки. В данной области известны многочисленные векторы, включая, но не ограничиваясь ими, линейные полинуклеотиды, полинуклеотиды, ассоциированные с ионными или амфифильными соединениями, плазмиды и вирусы. Таким образом, термин "вектор" включает автономно реплицирующуюся плазмиду или вирус. Экспрессирующая конструкция может быть получена путем репликации в живой клетке, или ее можно получать синтетическим путем. Для целей настоящей заявки термины "экспрессирующая конструкция", "экспрессирующий вектор" и "вектор" используют взаимозаменяемо для демонстрации применения изобретения в общем иллюстративном значении, и они не предназначены для ограничения изобретения.
[0122] В определенных вариантах осуществления нуклеиновая кислота, кодирующая представляющий интерес полинуклеотид, находится под транскрипционным контролем промотора. "Промотор" относится к последовательности ДНК, распознаваемой синтетическим аппаратом клетки или введенным синтетическим аппаратом, требуемой для инициации специфической транскрипции гена. Термин "промотор" используется в настоящем описании для отнесения к группе модулей контроля транскрипции, которые кластеризуются вокруг участка инициации для РНК-полимеразы I, II или III. Совместно с промотором можно использовать энхансерные элементы для повышения уровня экспрессии конструкций. В определенных вариантах осуществления экспрессирующий вектор содержит промотор, "функционально связанный" с полинуклеотидом, кодирующим слитый белок или представляющий интерес белок-мишень. Выражение "функционально связанный" или "под контролем транскрипции", как используют в рамках изобретения, означает, что промотор находится в правильном положении и ориентации относительно полинуклеотида для контроля инициации транскрипции РНК-полимеразой и экспрессии слитого белка или представляющего интерес белка-мишени.
[0123] Как правило, также в экспрессирующей конструкции присутствует терминатор транскрипции/сигналы полиаденилирования. Примеры таких последовательностей включают, но не ограничиваются ими, последовательности, происходящие из SV40, как описано в Sambrook et al., выше, а также последовательность терминатора бычьего гормона роста (см., например, патент США №5122458). Кроме того, рядом с кодирующей последовательностью можно помещать последовательности 5'-UTR для усиления ее экспрессии. Такие последовательности могут включать UTR, содержащие участок внутренней посадки рибосомы (IRES).
[0124] Включение IRES позволяет трансляцию одной или более открытых рамок считывания с вектора. Элемент IRES привлекает эукариотический рибосомальный комплекс инициации трансляции и стимулирует инициацию трансляции. См., например, Kaufman et al., Nuc. Acids Res. (1991) 19:4485-4490; Gurtu et al., Biochem. Biophys. Res. Comm. (1996) 229:295-298; Rees et al., BioTechniques (1996) 20:102-110; Kobayashi et al., BioTechniques (1996) 21:399-402; и Mosser et al., BioTechniques (1997 22 150-161). Известно множество последовательностей IRES, и они включают последовательности, происходящие из широкого множества вирусов, такие как лидерные последовательности пикорнавирусов, такие как UTR вируса энцефаломиокардита (EMCV) (Jang et al. J. Virol. (1989) 63:1651-1660), лидерная пооследовательность полиомиелита, лидерная последовательность вируса гепатита A, IRES вируса гепатита C, IRES риновируса человека 2 типа (Dobrikova et al., Proc. Natl. Acad. Sci. (2003) 100(25):15125-15130), элемент IRES из вируса ящура (Ramesh et al., Nucl. Acid Res. (1996) 24:2697-2700), IRES гиардиавируса (Garlapati et al., J. Biol. Chem. (2004) 279(5):3389-3397) и т.п.В рамках настоящего изобретения также применимы различные невирусные последовательности IRES, включая, но не ограничиваясь ими, последовательности IRES из дрожжей, а также IRES рецептора ангиотензина II типа 1 человека (Martin et al., Mol. Cell Endocrinol. (2003) 212:51-61), IRES фибробластного фактора роста (IRES FGF-1 и IRES FGF-2, Martineau et al. (2004) Mol. Cell. Biol. 24(17):7622-7635), IRES сосудисто-эндотелиального фактора роста (Baranick et al. (2008) Proc. Natl. Acad. Sci. U.S.A. 105(12):4733-4738, Stein et al. (1998) Mol. Cell. Biol. 18(6):3112-3119, Bert et al. (2006) RNA 12(6):1074-1083), и IRES инсулиноподобного фактора роста 2 (Pedersen et al. (2002) Biochem. J. 363(Pt 1):37-44). Последовательность IRES может быть включена в вектор, например, для экспрессии слитого белка, содержащего периплазматический якорный белок, слитый с вариантом белка для дисплея в комбинации с представляющим интерес белком-мишенью из экспрессирующей кассеты.
[0125] Альтернативно полинуклеотид, кодирующий вирусный пептид T2A, можно использовать для обеспечения продукции множества белковых продуктов из одного вектора. Линкерные пептиды 2A встраивают между кодирующими последовательностями в мультицистронной конструкции. Пептид 2A, который является саморасщепляющимся, позволяет продуцирование коэкспрессируемых белков из мультицистронной конструкции в эквимолярных количествах. Можно использовать пептиды 2A из различных вирусов, включая, но не ограничиваясь ими, пептиды 2A, происходящие из вируса ящура, вируса лошадиного ринита A, вируса Thosea asigna и свиного тешовируса 1. См., например, Kim et al. (2011) PLoS One 6(4):e18556, Trichas et al. (2008) BMC Biol. 6:40, Provost et al. (2007) Genesis 45(10):625-629, Furler et al. (2001) Gene Ther. 8(11):864-873; включенные в настоящее описание в качестве ссылок в полном объеме.
[0126] В определенных вариантах осуществления экспрессирующая конструкция содержит плазмиду, пригодную для трансформации дрожжевой клетки. Дрожжевые экспрессирующие плазмиды, как правило, содержат специфический для дрожжей ориджин репликации (ORI) и алиментарные селективные маркеры (например, HIS3, URA3, LYS2, LEU2, TRP1, MET15, ura4+, leu1+, ade6+), маркеры для селекции с антибиотиками (например, aphA1 или ble), флуоресцентные маркеры (например, mCherry, зеленый флуоресцентный белок), биолюминесцентные маркеры (например, люцифераза), или другие маркеры для селекции трансформированных дрожжевых клеток. Дрожжевая плазмида, кроме того, может содержать компоненты для обеспечения переноса между бактериальным хозяином (например, E.coli) и дрожжевыми клетками. Доступен ряд различных типов дрожжевых плазмид, включая дрожжевые встраивающиеся плазмиды (YIp), которые лишены ORI и встраиваются в хромосомы хозяина посредством гомологичной рекомбинации; дрожжевые реплицирующиеся плазмиды (YRp), которые содержат автономно реплицирующуюся последовательность (ARS) и могут реплицироваться независимо; дрожжевые центромерные плазмиды (YCp), которые представляют собой низкокопийные векторы, содержащие часть ARS и часть последовательности центромеры (CEN); и дрожжевые эписомальные плазмиды (YEp), которые представляют собой высококопийные плазмиды, содержащие фрагмент из 2-микронного кольца (природная плазмида дрожжей), который позволяет стабильное получение 50 или более копий на клетку.
[0127] Также может быть желательным включение регуляторных последовательностей, которые позволяют регуляцию экспрессии последовательностей белков относительно роста клетки-хозяина. Такие регуляторные последовательности известны специалистам в данной области, и их примеры включают последовательности, которые вызывают включение или выключение экспрессии гена в ответ на химический или физический стимул, включая присутствие регуляторного соединения. Например, феромон-индуцируемый промотор, такой как промотор PRM1 или FUS2, можно использовать для достижения транскрипции, зависимой от активации каскада передачи сигнала феромонов. Последовательности контроля и другие регуляторные последовательности могут быть лигированы с кодирующей последовательностью перед встраиванием в вектор. Альтернативно кодирующую последовательность можно клонировать непосредственно в экспрессирующий вектор, который уже содержит последовательности контроля и соответствующий участок рестрикции.
[0128] В некоторых случаях может быть необходимой модификация кодирующей последовательности так, чтобы она была связана с последовательностями контроля в надлежащей ориентации; т.е. для сохранения надлежащей рамки считывания. Мутанты или можно получать путем делеции части последовательности, кодирующей белок, путем встраивания последовательности и/или путем замены одного или более нуклеотидов в последовательности. Способы модификации нуклеотидных последовательностей, такие как сайт-направленный мутагенез, хорошо известны специалистам в данной области. См., например, Sambrook et al., выше; DNA Cloning, Vols. I and II, выше; Nucleic Acid Hybridization, выше.
[0129] В одном варианте осуществления рекомбинантные полинуклеотиды, кодирующие варианты белка, клонируют в нацеливающий в периплазму экспрессирующий вектор, содержащий: a) полинуклеотид, кодирующий сигнальный пептид; b) участок клонирования, пригодный для встраивания в рамке считывания полинуклеотида, кодирующего вариант белка, после полинуклеотида, кодирующего сигнальный пептид; c) полинуклеотид, кодирующий заякоривающий на плазматической мембране гликофосфатидилинозитол(GPI)-домен, расположенный так, чтобы вектор был способен продуцировать слитый белок, содержащий сигнальный пептид и вариант белка, слитый с заякоривающим на плазматической мембране GPI-доменом; и d) промотор, функционально связанный с последовательностями, кодирующими слитый белок. В определенных вариантах осуществления заякоривающий на плазматической мембране GPI-домен представляет собой заякоривающий на плазматической мембране GPI-домен япсина, такой как, но не ограничиваясь ими, заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7. Нацеливающий в периплазму экспрессирующий вектор, кроме того, может содержать полинуклеотид, кодирующий линкер, находящийся между участком клонирования и полинуклеотидом, кодирующим заякоривающий на плазматической мембране GPI-домен. Участок клонирования может содержать один или более участков рестрикции. Нацеливающий в периплазму экспрессирующий вектор, кроме того, может содержать селективный маркер.
[0130] В некоторых вариантах осуществления к варианту белка добавляют аффинную метку, эпитоп, маркировку и т.п., для обеспечения возможности определения общего уровня дисплея в дрожжевых клетках. Как используют в рамках изобретения, термин "аффинная метка" относится к биомолекуле, такой как полипептидный сегмент, которая может быть связана со второй биомолекулой для обеспечения очистки или детекции второй биомолекулы или предоставления участков для присоединения второй биомолекулы к субстрату. Примеры аффинных меток включают поли-гистидиновый участок, белок A (Nilsson et al. (1985) EMBO J. 4:1075; Nilsson et al. (1991) Methods Enzymol. 198:3, глутатион S трансферазу (Smith and Johnson (1988) Gene 67:31), аффинную метку Glu-Glu (Grussenmeyer et al., (1985) PNAS USA 82:7952), вещество P, пептид FLAG (Hopp et al. (1988) Biotechnology 6:1204), стрептавидин-связывающий пептид, или другой антигенный эпитоп или связывающий домен, и т.п.(Ford et al. (1991) Protein Expression and Purification 2:950), все из которых включены в настоящее описание в качестве ссылок. Как используют в рамках изобретения, "маркировка" представляет собой молекулу или атом, которые могут быть конъюгированы с биомолекулой, чтобы сделать биомолекулу или форму биомолекулы, такую как конъюгат, поддающейся детекции или количественному определению. Примеры маркировок включают флуоресцентные агенты, биолюминесцентные белки, фотоактивные агенты, радиоизотопы, парамагнитные ионы, хелаторы и т.п.
[0131] Экспрессирующий вектор используют для трансформации подходящей дрожжевой клетки-хозяина. В данной области известен ряд дрожжевых хозяев, включая, но не ограничиваясь ими, Saccharomyces arboricolus, Saccharomyces bayanus, Saccharomyces boulardii, Saccharomyces bulderi, Saccharomyces cariocanus, Saccharomyces cariocus, Saccharomyces cerevisiae, Saccharomyces chevalieri, Saccharomyces dairenensis, Saccharomyces ellipsoideus, Saccharomyces eubayanus, Saccharomyces exiguus, Saccharomyces florentinus, Saccharomyces fragilis, Saccharomyces kluyveri, Saccharomyces kudriavzevii, Saccharomyces martiniae, Saccharomyces mikatae, Saccharomyces monacensis, Saccharomyces norbensis, Saccharomyces paradoxus, Saccharomyces pastorianus, Saccharomyces spencerorum, Saccharomyces turicensis, Saccharomyces unisporus, Saccharomyces uvarum, Saccharomyces zonatus, Candida albicans, Candida ascalaphidarum, Candida amphixiae, Candida antarctica, Candida argentea, Candida atlantica, Candida atmosphaerica, Candida auris, Candida blattae, Candida bromeliacearum, Candida carpophila, Candida carvajalis, Candida cerambycidarum, Candida chauliodes, Candida corydali, Candida dosseyi, Candida dubliniensis, Candida ergatensis, Candida fructus, Candida glabrata, Candida fermentati, Candida guilliermondii, Candida haemulonii, Candida humilis, Candida insectamens, Candida insectorum, Candida intermedia, Candida jeffresii, Candida kefyr, Candida keroseneae, Candida krusei, Candida lusitaniae, Candida lyxosophila, Candida maltosa, Candida marina, Candida membranifaciens, Candida mogii, Candida oleophila, Candida oregonensis, Candida parapsilosis, Candida quercitrusa, Candida rugosa, Candida sake, Candida shehatea, Candida temnochilae, Candida tenuis, Candida theae, Candida tolerans, Candida tropicalis, Candida tsuchiyae, Candida sinolaborantium, Candida sojae, Candida subhashii, Candida viswanathii, Candida utilis, Candida ubatubensis, Candida zemplinina, Pichia farinosa, Pichia anomala, Pichia heedii, Pichia guilliermondii, Pichia kluyveri, Pichia membranifaciens, Pichia norvegensis, Pichia ohmeri, Pichia pastoris, Pichia methanolica, Pichia subpelliculosa, Kluyveromyces aestuarii, Kluyveromyces africanus, Kluyveromyces bacillisporus, Kluyveromyces blattae, Kluyveromyces dobzhanskii, Kluyveromyces hubeiensis, Kluyveromyces lactis, Kluyveromyces lodderae, Kluyveromyces marxianus, Kluyveromyces nonfermentans, Kluyveromyces piceae, Kluyveromyces sinensis, Kluyveromyces thermotolerans, Kluyveromyces waltii, Kluyveromyces wickerhamii, Kluyveromyces yarrowii, Yarrowia bubula, Yarrowia deformans, Yarrowia lipolytica, Yarrowia porcina и Yarrowia yakushimensis, которые могут использоваться в экспрессирующих конструкциях по настоящему изобретению. В некоторых вариантах осуществления дрожжи представляют собой вид Saccharomyces. В некоторых вариантах осуществления дрожжи представляют собой Saccharomyces cerevisiae.
[0132] Для обеспечения экспрессии смысловых или антисмысловых генных конструкций, экспрессирующую конструкцию необходимо доставить в дрожжевую клетку. Эту доставку можно проводить in vitro с использованием лабораторных методик трансформации дрожжевых клеток, хорошо известных в данной области, таких как трансформация сферопластов, обработка щелочными ионами (например, Cs+или Li+), электропорация, конъюгация между царствами, электропорация и биолистические и опосредуемые стеклянными гранулами способы (см., например, Kawai et al. (2010) Bioeng. Bugs. 1(6):395-403, Gietz et al. (1995) Yeast 11(4):355-360, Gietz et al. (2007) Nat. Protoc. 2(1):38-41, Hinnen et al. (1978) Proc. Natl. Acad. Sci. USA 75:1929-1933, Avery et al. (1995) Mol. Med. 1(4):344-365, Ito et al. (1983) J. Bacteriol. 153:163-168, Johnston et al. (1988) Science 240:1538-1541, Dohmen et al. (1991) Yeast 7(7):691-246, Hayama et al. (2002) J. Biosci. Bioeng. 94(2):166-171, и Wang et al. (2001) Crit. Rev. Biotechnol. 21(3):177-218; включенные в настоящее описание в качестве ссылок).
[0133] После доставки экспрессирующей конструкции в клетку нуклеиновая кислота, кодирующая представляющий интерес ген, может помещаться и экспрессироваться в различных участках. В определенных вариантах осуществления нуклеиновая кислота, кодирующая ген, может стабильно встраиваться в геном клетки посредством гомологичной рекомбинации. Это встраивание может осуществляться в сходном положении и ориентации (замена гена), в гене (прерывание гена) или в случайном неспецифическом положении (увеличение гена). Встраивание конструкции в локусе-мишени, которое прерывает ген, может быть приемлемым при условии, что прерывание гена не препятствует росту клеток или скринингу библиотеки дрожжевого периплазматического дисплея (например, не происходит нарушения ответа феромонов, если его используют при скрининге). В других вариантах осуществления нуклеиновую кислоту можно стабильно поддерживать в клетке в качестве отдельного эписомального сегмента ДНК. Такие сегменты нуклеиновых кислот или "эписомы" кодируют последовательности, достаточные для того, чтобы позволить поддержание и репликацию независимо от или синхронно с клеточным циклом хозяина. Способ доставки экспрессирующей конструкции в клетку и то, где остается нуклеиновая кислота в клетке, зависит от типа используемой экспрессирующей конструкции.
[0134] В определенных вариантах осуществления экспрессирующая конструкция может просто состоять из голой рекомбинантной ДНК или плазмид. Перенос конструкции можно проводить любыми способами, упомянутыми выше, которые физически или химически обеспечивают проницаемость клеточной мембраны.
[0135] В другом варианте осуществления экспрессирующую конструкцию голой ДНК можно переносить в клетки посредством бомбардировки частицами. Этот способ основан на способности ускорять покрытые ДНК микроснаряды до высокой скорости, что позволяет им проходить через клеточные стенки и мембраны дрожжей и входить в клетки, не убивая их (Armaleo et al. (1990) Curr. Genet. 17(2):97-103). Было разработано несколько устройств для ускорения мелких частиц. Одно такое устройство основано на высоковольтном разряде для генерирования электрического тока, который в свою очередь обеспечивает движущую силу (Yang et al. (1990) Proc. Natl. Acad. Sci. USA 87:9568-9572). Микроснаряды могут состоять из биологически инертных веществ, таких как вольфрамовые или золотые гранулы.
[0136] В некоторых вариантах осуществления получают набор линейных молекул ДНК, кодирующих варианты белка. Вместо клонирования линейных молекул ДНК в вектор перед трансформацией дрожжевые клетки трансформируют пустым вектором вместе с набором линейных молекул ДНК, кодирующих варианты белков, которые затем встраиваются в вектор in vivo, например, посредством гомологичной рекомбинации в дрожжевых клетках-хозяевах.
C. Наборы
[0137] Набор может включать библиотеку дрожжевого периплазматического дисплея, как описано в настоящем описании, или средства для получения библиотеки дрожжевого периплазматического дисплея, такие как подходящие векторы для клонирования нуклеиновых кислот, кодирующих варианты белка, для получения вариантов белка, связанных с периплазматическим якорным белком, дрожжевые клетки, агенты трансфекции, среду, пригодную для выращивания дрожжевых клеток, средства для позитивной и негативной селекции клеток и другие реагенты, которые необходимы. В набор, как правило, включают инструкции (например, в печатной форме, CD-ROM, DVD, Blu-ray, флэш-память, цифровая загружаемая информация, и т.д.) по получению и/или скринингу библиотеки дрожжевого периплазматического дисплея, как описано в настоящем описании.
[0138] Набор может включать библиотеку дрожжевого периплазматического дисплея, как описано в настоящем описании, или средства для получения библиотеки дрожжевого периплазматического дисплея, такие как подходящие векторы для клонирования нуклеиновых кислот, кодирующих варианты белка, для получения слитых конструкций с периплазматическим якорным белком, дрожжевые клетки, агенты трансфекции, среду, пригодную для выращивания дрожжевых клеток, средства для позитивной или негативной селекции клеток и другие реагенты, которые требуются. В набор, как правило, включают инструкции (например, в печатной форме, CD-ROM, DVD, Blu-ray, флэш-память, цифровая загружаемая информация, и т.д.) по получению и/или скринингу библиотеки дрожжевого периплазматического дисплея, как описано в настоящем описании.
[0139] В одном варианте осуществления набор содержит нацеливающий в периплазму экспрессирующий вектор, содержащий: a) полинуклеотид, кодирующий сигнальный пептид; b) участок клонирования, пригодный для встраивания в рамке считывания полинуклеотида, кодирующего вариант белка (например, антитело) после полинуклеотида, кодирующего сигнальный пептид; c) полинуклеотид, кодирующий заякоривающий на плазматической мембране гликофосфатидилинозитол (GPI)-домен, расположенных так, чтобы вектор был способен продуцировать слитый белок, содержащий сигнальный пептид и вариант белка, слитый с заякоривающим на плазматической мембране GPI-доменом; и d) промотор, функционально связанный с последовательностями, кодирующими слитый белок. В другом варианте осуществления сигнальный пептид содержит сигнальную последовательность препро-альфа-фактора. В определенных вариантах осуществления заякоривающий на плазматической мембране GPI-домен представляет собой заякоривающий на плазматической мембране GPI-домен япсина, такой как, но не ограничиваясь ими, заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7. В другом варианте осуществления нацеливающий в периплазму экспрессирующий вектор дополнительно содержит полинуклеотид, кодирующий линкер, где указанный полинуклеотид, кодирующий линкер, находится между участком клонирования и полинуклеотидом, кодирующим заякоривающий на плазматической мембране GPI-домен. Линкер, кроме того, может содержать метку. В другом варианте осуществления нацеливающий в периплазму экспрессирующий вектор, кроме того, содержит селективный маркер. В другом варианте осуществления участок клонирования содержит один или более участков рестрикции.
[0140] В другом варианте осуществления набор включает библиотеку дрожжевого периплазматического дисплея, содержащую множество дрожжевых клеток-хозяев, причем каждая дрожжевая клетка-хозяин содержит: a) слитый белок, содержащий периплазматический якорный белок, слитый с вариантом белка (например, антитело), для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированный вариант белка является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество вариантов белков; b) представляющий интерес сопряженный с G-белком рецептор (GPCR), являющийся мишенью, где представляющий интерес GPCR-мишень находится на плазматической мембране дрожжевой клетки-хозяина и является доступным для варианта белка, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина; и c) сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяине. В определенных вариантах осуществления сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен дрожжевой субъединицы G(и C-концевой домен экзогенной субъединицы Gα. Например, дрожжевая субъединица G(может принадлежать семейству G-белков Gαi, Gαq, Gαs или Gαo. В химерной субъединице G(по меньшей мере пять C-концевых остатков дрожжевой субъединицы G(могут быть заменены соответствующими C-концевыми остатками субъединицы G(млекопитающих, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего. В некоторых вариантах осуществления по меньшей мере 20 C-концевых остатков дрожжевой субъединицы G(заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего. В другом варианте осуществления химерная субъединица G(содержит по меньшей мере 41 N-концевой остаток дрожжевой субъединицы Gα. В определенных вариантах осуществления GPCR млекопитающих представляет собой GPCR мыши. В определенных вариантах осуществления GPCR млекопитающих представляет собой GPCR человека, выбранный из группы, состоящей из CXCR4, CXCR5, SSTR2, MOR, AVPR2, FPR2/ALX, ADORA2A, CHRM3, CGRP2, CCR2, CCR4, CCR5, CHRM4, PAC1, b2AR, CXCR2, CYSLTR2, KSHV vGPCR, PKR1, PKR2, CB1, CB2, A3AR и AT1R.
[0141] В другом варианте осуществления набор включает библиотеку дрожжевого периплазматического дисплея, содержащую: множество дрожжевых клеток-хозяев, причем каждая дрожжевая клетка-хозяин содержит слитый белок, содержащий периплазматический якорный белок, слитый с вариантом белка для дисплея, где периплазматический якорный белок является достаточно большим, чтобы слитый белок удерживался в периплазме, и экспонированный вариант белка является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество вариантов белков.
[0142] В другом варианте осуществления набор включает библиотеку дрожжевого периплазматического дисплея, содержащую: множество дрожжевых клеток-хозяев, причем каждая дрожжевая клетка-хозяин содержит слитый белок, содержащий вариант белка для дисплея, связанный с представляющим интерес мембранным белком-мишенью, где экспонированный вариант белка является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество вариантов белка с представляющим интерес мембранным белком-мишенью.
D. Применения
[0143] Настоящее изобретение можно в общем применять для скрининга белков, которые выполняют полезные или желаемые функции, включая связывание, катализ, сборку, транспорт и т.п. Например, дрожжевой периплазматический дисплей можно использовать для идентификации агонистов и антагонистов рецепторов, конструирования терапевтических белков и антител, идентификации белок-белковых взаимодействий и картирования эпитопов.
[0144] Дрожжевой периплазматический дисплей является особенно пригодным для скрининга библиотеки белков в отношении кандидатов, которые связывают и/или модулируют функцию белка-мишени, который является мембранным белком, такого как рецептор, ионный канал или переносчик. Локализация белка на мембране делает его доступным для экспонированных вариантов белков в периплазматическом пространстве (например, достаточно близко, чтобы экспонированный вариант белка связывался с представляющим интерес белком-мишенью).
[0145] В определенных вариантах осуществления активация представляющего интерес белка-мишени повышает рост дрожжевых клеток-хозяев. В этом случае библиотеку дрожжевого периплазматического дисплея можно подвергать скринингу в отношении агониста представляющего интерес белка-мишени посредством культивирования по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея в среде, где рост дрожжевой клетки-хозяина в среде указывает на то, что вариант белка, экспонированный в дрожжевой клетке-хозяине, является агонистом представляющего интерес белка-мишени.
[0146] В других вариантах осуществления активация представляющего интерес белка-мишени снижает рост дрожжевых клеток-хозяев. В этом случае библиотеку дрожжевого периплазматического дисплея можно подвергать скринингу в отношении антагониста представляющего интерес белка-мишени посредством культивирования по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея в среде, где рост дрожжевой клетки-хозяина в среде указывает на то, что вариант белка, экспонированный в дрожжевой клетке-хозяине, является антагонистом представляющего интерес белка-мишени.
[0147] В определенных вариантах осуществления каждая дрожжевая клетка-хозяин дополнительно содержит репортерную систему, содержащую репортерный ген, функционально связанный с индуцибельным промотором, который активируется, когда активируется представляющий интерес белок-мишень, позволяя детецию повышения или снижения активности представляющего интерес белка-мишени при связывании экспонированного варианта белка с представляющим интерес белком-мишенью. Например, репортерный ген может представлять собой алиментарный маркер (например, HIS3, HIS7, ARG6, LEU2, URA3 и TRP1), маркер резистентности к антибиотику (например, сообщает резистентность к антибиотику, такому как генетицин (например, aphA1), зеоцин (например, ble), гигромицин B, ноурсеотрицин или биалафос), флуоресцентный маркер (например, зеленый флуоресцентный белок, красный флуоресцентный белок, синий флуоресцентный белок, голубой флуоресцентный белок, желтый флуоресцентный белок и оранжевый флуоресцентный белок), биолюминесцентный маркер (например, люцифераза или экворин) или маркер контрселекции (например, CAN1, URA3, MET15, TRP1 и TK).
[0148] Например, позитивную селекцию (т.е. селекцию в отношении активации экспрессии репортерного гена) можно использовать для детекции увеличения активности представляющего интерес мембранного белка-мишени при связывании экспонированного варианта белка с представляющим интерес мембранным белком-мишенью. Детекцию экспрессии алиментарного маркера можно проводить, например, по росту дрожжевых клеток-хозяев в селективной среде с дефицитом питательных веществ. Детекцию экспрессии маркера резистентности к антибиотику можно проводить, например, по росту дрожжевых клеток-хозяев в селективной среде, содержащей антибиотик. Детекцию экспрессии флуоресцентного маркера можно проводить, например, по флуоресценции, испускаемой дрожжевыми клетками-хозяевами. Детекцию экспрессии биолюминесцентного маркера можно проводить, например, по биолюминесценции, испускаемой дрожжевыми клетками-хозяевами.
[0149] Альтернативно контрселекцию (т.е. селекцию на основе роста в отношении потери экспрессии репортерного гена) можно использовать для детекции снижения активности представляющего интерес мембранного белка-мишени при связывании экспонированного варианта белка с представляющим интерес мембранным белком-мишенью. Маркер контрселекции может убивать клетки путем индукции апоптоза, конвертировать нетоксичное лекарственное средство в токсичное соединение или транспортировать токсичную молекулу в клетку. Контрселекцию можно проводить путем культивирования дрожжевых клеток-хозяев в среде, содержащей средство, которое селективно убивает клетки, экспрессирующие маркер контрселекции. Иллюстративные маркеры контрселекции включают CAN1 (контрселекция с канаванином), URA3 (контрселекция с 5-фтороротовой кислотой (5-FOA)), MET15 (контрселекция с метилртутью), TRP1 (контрселекция с 5-фторантраниловой кислотой (5-FAA)) и тимидинкиназу TK вируса герпеса человека (контрселекция с флоксуридином (FUDR)).
[0150] В частности, библиотеку дрожжевого периплазматического дисплея можно использовать для скрининга в отношении антител, которые связывают и модулируют функцию GPCR. В некоторых вариантах осуществления GPCR в дрожжевой клетке-хозяине заменен на GPCR млекопитающего, например, GPCR человека. В некоторых вариантах осуществления дрожжевая клетка-хозяин экспрессирует GPCR млекопитающего, например, GPCR человека, и эндогенный GPCR дрожжей. В некоторых вариантах осуществления антагонисты и агонисты идентифицируют с использованием репортерной системы, которая сопрягает ответ GPCR на связывание экспонированного антитела с уровнями секреции феромонов дрожжей (см. примеры). Для этой цели дрожжевую клетку-хозяин можно генетически модифицировать для экспрессии сконструированной субъединицы Gα, способной активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяине. В определенных вариантах осуществления сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен дрожжевой субъединицы G(и C-концевой домен экзогенной субъединицы Gα. Например, дрожжевая субъединица G(может принадлежать семейству G-белков Gαi, Gαq, Gαs или Gαo. В химерной субъединице G(по меньшей мере пять C-концевых остатков дрожжевой субъединицы G(могут быть заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего. В некоторых вариантах осуществления по меньшей мере 20 C-концевых остатков дрожжевой субъединицы G(заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего. В другом варианте осуществления химерная субъединица G(содержит по меньшей мере 41 N-концевой остаток дрожжевой субъединицы Gα. Иллюстративные субъединицы G(млекопитающих включают G альфа-S, G альфа-I, G альфа-O, G альфа-T, G альфа-Z, G альфа-Q, G альфа-11, G альфа-12, G альфа-13 и трансдуцин.
[0151] В частности, штаммы дрожжей Δfar1, Δsst2, Δste14, Δste3 или Δmat являются пригодными для скрининга антагонистов и агонистов GPCR. Штамм Δmat может содержать, например, удаленный или инактивированный локус MAT(или удаленный или инактивированный локус MATa. Дрожжевая клетка-хозяин, кроме того, может содержать модифицированный белок CLN3, содержащий C-концевое укорочение, которое повышает содержание CLN3 в дрожжевой клетке-хозяине по сравнению с белком CLN3 дикого типа. Например, модифицированный белок CLN3 может сохранять по меньшей мере N-концевые аминокислоты 1-387 или 1-408 белка CLN3 дикого типа, или любое количество N-концевых аминокислот в этих диапазонах, такое как 1-388, 1-389, 1-390, 1-391, 1-392, 1-393, 1-394, 1-395, 1-396, 1-397, 1-398, 1-399, 1-400, 1-401, 1-402, 1-403, 1-404, 1-405, 1-406, 1-407 или 1-408, где C-концевое укорочение содержит делецию всех или некоторых из остальных остатков белка CLN3 дикого типа. Дрожжевые клетки-хозяева, используемые для получения библиотеки периплазматического дисплея, могут быть гаплоидными или диплоидными. Иллюстративные штаммы дрожжей, предназначенные для применения в селекции антагониста и агониста, описаны в примерах 2 и 5, соответственно.
[0152] В определенных вариантах осуществления дрожжевая клетка-хозяин представляет собой штамм FAR1, где ингибирование ответа феромонов посредством антитела, выступающего в качестве антагониста, который связывает и ингибирует GPCR в дрожжевой клетке-хозяине, приводит к прекращению остановки клеточного цикла и росту дрожжевой клетки-хозяина. В других вариантах осуществления дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном, где активация ответа феромонов посредством антитела, выступающего в качестве агониста, который связывает и активирует GPCR в дрожжевой клетке-хозяине, приводит к повышению экспрессии репортерного гена.
[0153] Любой тип GPCR из любого вида можно подвергать скринингу с использованием периплазматического дисплея, как описано в настоящем описании. В некоторых вариантах осуществления представляющий интерес GPCR-мишень представляет собой GPCR млекопитающего (например, из человека или не являющегося человеком примата. грызуна, лабораторного животного, домашнего скота). Например, GPCR млекопитающего может представлять собой GPCR человека (например, CXCR4, CXCR5, SSTR2, MOR, AVPR2, FPR2/ALX, ADORA2A, CHRM3, CGRP2, CCR2, CCR4, CCR5, CHRM4, PAC1, b2AR, CXCR2, CYSLTR2, KSHV vGPCR, PKR1, PKR2, CB1, CB2, A3AR и AT1R). Представляющий интерес GPCR-мишень может обладать конститутивной лиганд-независимой активностью. Альтернативно можно добавлять лиганд для активации представляющего интерес GPCR-мишени в ходе скрининга на агонисты или антагонисты. В некоторых вариантах осуществления дрожжевая клетка-хозяин экспрессирует представляющий интерес GPCR-мишень, например, представляющий интерес GPCR человека, и эндогенный GPCR дрожжей.
[0154] В некоторых вариантах осуществления варианты белков представляют собой антитела. С использованием способов дрожжевого периплазматического дисплея, описанных в настоящем описании, можно проводить скрининг любого типа антител, включая моноклональные антитела, гибридные антитела, измененные антитела, химерные антитела и гуманизированные антитела, а также: гибридные (химерные) молекулы антител (см., например, Winter et al. (1991) Nature 349:293-299; и патент США №4816567); F(ab')2- и F(ab)-фрагменты; Fv-молекулы (нековалентные гетеродимеры, см., например, Inbar et al. (1972) Proc Natl Acad Sci USA 69:2659-2662; и Ehrlich et al. (1980) Biochem 19:4091-4096); одноцепочечные Fv-молекулы (scFv) (см., например, Huston et al. (1988) Proc Natl Acad Sci USA 85:5879-5883); наноантитела или однодоменные антитела (sdAb) (см., например, Wang et al. (2016) Int J Nanomedicine 11:3287-3303, Vincke et al. (2012) Methods Mol Biol 911:15-26; димерные и триерные конструкции фрагментов антител; миниантитела (см., например, Pack et al. (1992) Biochem 31:1579-1584; Cumber et al. (1992) J Immunology 149B:120-126); гуманизированные молекулы антител (см., например, Riechmann et al. (1988) Nature 332:323-327; Verhoeyan et al. (1988) Science 239:1534-1536; и публикацию патента Великобритании №GB 2276169, опубликованную 21 сентября 1994 года); и любые функциональные фрагменты, получаемые из таких молекул, где такие фрагменты сохраняют свойства специфического связывания исходной молекулы антитела.
[0155] В других вариантах осуществления варианты белков представляют собой аптамеры. Аптамеры можно выделять из комбинаторной библиотеки и усовершенствовать посредством направленной мутации или повторяющихся раундов мутагенеза и селекции. Для описания способов получения аптамеров см., например, Aptamers: Tools for Nanotherapy and Molecular Imaging (R.N. Veedu ed., Pan Stanford, 2016), Nucleic Acid and Peptide Aptamers: Methods and Protocols (Methods in Molecular Biology, G. Mayer ed., Humana Press, 2009), Aptamers Selected by Cell-SELEX for Theranostics (W. Tan, X. Fang eds., Springer, 2015), Cox et al. (2001) Bioorg. Med. Chem. 9(10):2525-2531; Cox et al. (2002) Nucleic Acids Res. 30(20): e108, Kenan et al. (1999) Methods Mol. Biol. 118:217-231; Platella et al. (2016) Biochim. Biophys. Acta Nov 16 pii: S0304-4165(16)30447-0, and Lyu et al. (2016) Theranostics 6(9):1440-1452; включенные в настоящее описание в качестве ссылок в полном объеме.
[0156] В других вариантах осуществления варианты белков представляют собой миметики антител. Можно использовать любой тип миметиков антител, включая, но не ограничиваясь ими, молекулы аффиантител (Nygren (2008) FEBS J. 275 (11):2668-2676), аффилины (Ebersbach et al. (2007) J. Mol. Biol. 372 (1):172-185), аффимеры (Johnson et al. (2012) Anal. Chem. 84 (15):6553-6560), аффитины (Krehenbrink et al. (2008) J. Mol. Biol. 383 (5):1058-1068), альфатела (Desmet et al. (2014) Nature Communications 5:5237), антикалины (Skerra (2008) FEBS J. 275 (11):2677-2683), авимеры (Silverman et al. (2005) Nat. Biotechnol. 23 (12):1556-1561), дарпины (Stumpp et al. (2008) Drug Discov. Today 13 (15-16):695-701), финомеры (Grabulovski et al. (2007) J. Biol. Chem. 282 (5):3196-3204) и монотела (Koide et al. (2007) Methods Mol. Biol. 352:95-109).
[0157] Кроме того, можно проводить направленную эволюцию посредством ммногократных раундов мутагенеза и скрининга посредством дрожжевого периплазматического дисплея для обогащения библиотек вариантами белков (например, антителами), которые связывают представляющий интерес белок-мишень с высокой аффинностью. Направленная эволюция, в частности, может быть пригодной для улучшения характеристик связывания кандидатов с желаемой функциональной активностью, но слабой аффинностью связывания.
III. Иллюстративные варианты осуществления
[0158] Варианты осуществления, описанные в настоящем описании, представляют собой следующие:
1. Библиотека дрожжевого периплазматического дисплея, содержащая множество дрожжевых клеток-хозяев, где каждая дрожжевая клетка-хозяин содержит:
a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител;
b) периплазматический якорный белок, где периплазматический якорный белок связан с антителом, так что антитело экспонируется в периплазматическом пространстве; и
c) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок находится на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
2. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 1, где антитело и периплазматический якорный белок нековалентно связаны друг с другом посредством молекулярных связывающих взаимодействий в комплексе или связаны посредством ковалентной непептидной связи в комплексе.
3. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 1, где антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
4. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-3, где периплазматический якорный белок дополнительно содержит сигнальную последовательность, которая направляет транспорт периплазматического якорного белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что антитело экспонируется в периплазме.
5. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-3, где периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или связанный с мембраной белковый домен, который выставляет антитело в периплазму.
6. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 5, где связанный с клеточной мембраной белковый домен представляет собой заякоривающий на плазматической мембране гликозилфосфатидилинозитол (GPI)-домен.
7. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 6, где заякоривающий на плазматической мембране GPI-домен представляет собой заякоривающий на плазматической мембране GPI-домен япасина.
8. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 7, где заякоривающий на плазматической мембране GPI-домен япсина представляет собой заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7.
9. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 2, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что экспонируемый вариант белка выступает в периплазму.
10. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 3, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что экспонируемый вариант белка выступает в периплазму.
11. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 2, где периплазматический якорный белок является достаточно большим, чтобы такой периплазматический якорный белок и связанное антитело удерживались в периплазме.
12. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 3, где периплазматический якорный белок является достаточно большим, чтобы слитый белок удерживался в периплазме.
13. Библиотека дрожжевого периплазматического дисплея, содержащая множество дрожжевых клеток-хозяев, где каждая дрожжевая клетка-хозяин содержит:
a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител, где антитело связано с сигнальной последовательностью, которая направляет транспорт антитела в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или клеточную стенку, так что антитело экспонируется в периплазматическом пространстве дрожжевой клетки-хозяина; и
b) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок находится на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
14. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-13, дополнительно содержащая репортерную систему, содержащую репортерный ген, функционально связанный с индуцибельным промотором, который активируется, когда активируется представляющий интерес мембранный белок-мишень, позволяя детекцию увеличения или снижения активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью.
15. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 14, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотику, флуоресцентный маркер, биолюминесцентный маркер или маркер контрселекции.
16. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 15, где алиментарный маркер выбран из группы, состоящей из HIS3, HIS7, ARG6, LEU2, URA3 и TRP1.
17. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 15, где маркер резистентности к антибиотику сообщает резистентность к антибиотику, выбранному из группы, состоящей из генетицина, зеоцина, гигромицина B, ноурсеотрицина и биалафоса.
18. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 15, где флуоресцентный маркер выбран из группы, состоящей из зеленого флуоресцентного белка, красного флуоресцентного белка, синего флуоресцентного белка, голубого флуоресцентного белка, желтого флуоресцентного белка и оранжевого флуресцентного белка.
19. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 15, где биолюминесцентным маркером является люцифераза или экворин.
20. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 15, где маркер контрселекции выбран из группы, состоящей из CAN1, URA3, MET15, TRP1 и TK.
21. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 14, где репортерный ген представляет собой селективный маркер, так что указанное повышение активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью поддается обнаружению посредством роста дрожжевых клеток-хозяев на среде для позитивной селекции.
22. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 14, где репортерный ген представляет собой маркер контрселекции, так что указанное снижение активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью поддается обнаружению по росту дрожжевых клеток-хозяев на среде для негативной селекции.
23. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-22, где представляющий интерес мембранный белок-мишень выбран из группы, состоящей из рецептора, ионного канала и переносчика.
24. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 23, где рецептор представляет собой сопряженный с G-белком рецептор (GPCR).
25. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 24, где GPCR представляет собой экзогенный GPCR.
26. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 25, где дрожжевая клетка-хозяин дополнительно содержит эндогенный GPCR.
27. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 25 или 26, дополнительно содержащая сконструированную субъединицу Gα, способную активироваться экзогенным GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяине.
28. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 27, где сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен субъединицы G(дрожжей и C-концевой домен экзогенной субъединицы Gα.
29. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 28, где субъединица G(дрожжей принадлежит семейству G-белков G(i, G(q, G(s или G(o.
30. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 25-29, где экзогенный GPCR представляет собой GPCR млекопитающего.
31. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 30, где по меньшей мере пять C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
32. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 31, где по меньшей мере 20 C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
33. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 31 или 32, где субъединица G(млекопитающего выбрана из группы, состоящей из G альфа-S, G альфа-I, G альфа-O, G альфа-T, G альфа-Z, G альфа-Q, G альфа-11, G альфа-12, G альфа-13 и трансдуцина.
34. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 28-33, где химерная субъединица G(содержит по меньшей мере 41 N-концевой остаток субъединицы G(дрожжей.
35. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 30-34, где GPCR млекопитающего представляет собой GPCR человека.
36. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 35, где GPCR человека выбран из группы, состоящей из CXCR4, CXCR5, SSTR2, MOR, AVPR2, FPR2/ALX, ADORA2A, CHRM3, CGRP2, CCR2, CCR4, CCR5, CHRM4, PAC1, b2AR, CXCR2, CYSLTR2, KSHV vGPCR, PKR1, PKR2, CB1, CB2, A3AR и AT1R.
37. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 24-36, где представляющий интерес мембранный белок-мишень GPCR обладает конститутивной лиганд-независимой активностью.
38. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 24-37, где дрожжевая клетка-хозяин представляет собой штамм FAR1 для селекции антител-антагонистов представляющего интерес мембранного белка-мишени GPCR.
39. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 24-36, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном для селекции антител-агонистов представляющего интерес мембранного белка-мишени GPCR.
40. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-39, где антитела выбраны из группы, состоящей из моноклональных антител, химерных антител, гуманизированных антител, наноантител, рекомбинантных фрагментов антител, Fab-фрагментов, Fab'-фрагментов, F(ab')2-фрагментов, Fv-фрагментов и scFv-фрагментов.
41. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-40, где представляющий интерес мембранный белок-мишень содержит мутацию, которая повышает или снижает его активность.
42. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-40, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, Δsst2, Δste14, Δste3 или Δmat.
43. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 42, где дрожжевая клетка-хозяин представляет собой штамм Δmat, содержащий удаленный или инактивированный локус MAT(или удаленный или инактивированный локус MATa.
44. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-43, где дрожжевая клетка-хозяин дополнительно содержит модифицированный белок CLN3, содержащий C-концевое укорочение, которое повышает содержание CLN3 в дрожжевой клетке-хозяине по сравнению с белком CLN3 дикого типа.
45. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 44, где в модифицированном белке CLN3 сохранены по меньшей мере N-концевые аминокислоты 1-387 белка CLN3 дикого типа.
46. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 44, где в модифицированном белке CLN3 сохранены по меньшей мере N-концевые аминокислоты 1-408 белка CLN3 дикого типа.
47. Библиотека дрожжевого периплазматического дисплея согласно любому из вариантов осуществления 1-46, где дрожжевая клетка-хозяин представляет собой гаплоидную или диплоидную дрожжевую клетку-хозяина.
48. Способ получения библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 1, причем способ включает:
a) предоставление первого множества рекомбинантных полинуклеотидов, кодирующих антитела, для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител;
b) предоставление второго рекомбинантного полинуклеотида, кодирующего периплазматический якорный белок, где периплазматический якорный белок связан с антителом, так что антитело экспонируется в периплазматическом пространстве;
c) трансфецкию множества дрожжевых клеток-хозяев первым множеством рекомбинантных полинуклеотидов и вторым рекомбинантным полинуклеотидом;
d) трансфекцию множества дрожжевых клеток-хозяев рекомбинантным полинуклеотидом, кодирующим представляющий интерес мембранный белок-мишень; и
e) культивирование множества дрожжевых клеток-хозяев в условиях, которые позволяют экспрессию антител, периплазматического якорного белка и представляющего интерес мембранного белка-мишени, где каждая дрожжевая клетка-хозяин экспонирует отличающееся антитело в периплазматическое пространство и представляющий интерес мембранный белок-мишень локализуется на плазматической мембране, с получением библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 1.
49. Способ согласно варианту осуществления 48, где рекомбинантные полинуклеотиды, кодирующие антитела, или рекомбинантный полинуклеотид, кодирующий периплазматический якорный белок или представляющий интерес мембранный белок-мишень, предоставлены посредством экспрессирующих векторов.
50. Способ согласно варианту осуществления 48, где рекомбинантные полинуклеотиды, кодирующие антитела, или рекомбинантный полинуклеотид, кодирующий периплазматический якорный белок или представляющий интерес мембранный белок-мишень, встраиваются в геном дрожжевой клетки-хозяина в локусе-мишени.
51. Способ получения библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 3, причем способ включает:
a) предоставление множества рекомбинантных полинуклеотидов, кодирующих слитые белки, где каждый рекомбинантный полинуклеотид кодирует отличающийся слитый белок, содержащий периплазматический якорный белок, связанный с отличающимся антителом, для дисплея;
b) трансфекцию множества дрожжевых клеток-хозяев множеством рекомбинантных полинуклеотидов, кодирующих слитые белки;
c) трансфекцию дрожжевых клеток-хозяев рекомбинантным полинуклеотидом, кодирующим представляющий интерес мембранный белок-мишень; и
d) культивирование множества дрожжевых клеток-хозяев в условиях, которые позволяют экспрессию слитых белков и представляющего интерес мембранного белка-мишени, где каждая дрожжевая клетка-хозяин экспонирует отличающееся антитело в периплазматическом пространстве, и представляющий интерес мембранный белок-мишень локализуется на плазматической мембране, с получением библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 3.
52. Способ согласно варианту осуществления 51, где рекомбинантные полинуклеотиды, кодирующие слитые белки, или рекомбинантный полинуклеотид, кодирующий представляющий интерес мембранный белок-мишень, предоставлены посредством экспрессирующих векторов.
53. Способ согласно варианту осуществления 51, где рекомбинантные полинуклеотиды, кодирующие слитые белки или представляющий интерес мембранный белок-мишень, встраиваются в геном дрожжевой клетки-хозяина в локусе-мишени.
54. Способ согласно любому из вариантов осуществления 48-53, где представляющий интерес мембранный белок-мишень выбран из группы, состоящей из рецептора, ионного канала и переносчика.
55. Способ согласно варианту осуществления 54, где рецептор представляет собой сопряженный с G-белком рецептор (GPCR).
56. Способ согласно любому из вариантов осуществления 48-55, включающий введение в множество дрожжевых клеток-хозяев рекомбинантного полинуклеотида, кодирующего сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяине.
57. Способ согласно варианту осуществления 56, где сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен субъединицы G(дрожжей и C-концевой домен экзогенной субъединицы Gα.
58. Способ согласно варианту осуществления 57, где субъединица G(дрожжей принадлежит семейству G-белков G(i, G(q, G(s или G(o.
59. Способ согласно варианту осуществления 57 или 58, где экзогенная субъединица G(представляет собой субъединицу G(млекопитающего.
60. Способ согласно варианту осуществления 59, где по меньшей мере пять C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
61. Способ согласно варианту осуществления 60, где по меньшей мере 20 C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
62. Способ согласно любому из вариантов осуществления 55-61, где дрожжевая клетка-хозяин представляет собой штамм FAR1 для селекции антител-антагонистов представляющего интерес GPCR-мишени.
63. Способ согласно любому из вариантов осуществления 55-61, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном для селекции антител-агонистов GPCR.
64. Способ скрининга библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 14 в отношении антитела, которое модулирует активность представляющего интерес мембранного белка-мишени, причем способ включает культивирование по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 14 в селективной среде; и детекцию экспрессии репортерного гена, где увеличенная экспрессия репортерного гена указывает на то, что антитело повышает активность представляющего интерес мембранного белка-мишени, и сниженная экспрессия репортерного гена указывает на то, что антитело снижает активность представляющего интерес мембранного белка-мишени.
65. Способ согласно варианту осуществления 64, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотику, флуоресцентный маркер, биолюминесцентный маркер или маркер контрселекции.
66. Способ согласно варианту осуществления 65, дополнительно включающий проведение позитивной селекции в отношении экспрессии алиментарного маркера, где рост дрожжевых клеток-хозяев в селективной среде с дефицитом питательных веществ указывает на то, что представляющий интерес мембранный белок-мишень активирован.
67. Способ согласно варианту осуществления 66, где алиментарный маркер представляет собой HIS3, HIS7, ARG6, LEU2, URA3 и TRP1.
68. Способ согласно варианту осуществления 65, дополнительно включающий проведение позитивной селекции в отношении экспрессии маркера резистентности к антибиотикам, где рост дрожжевых клеток-хозяев в селективной среде, содержащей антибиотик, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
69. Способ согласно варианту осуществления 68, где маркер резистентности к антибиотику сообщает резистентность к антибиотику, выбранному из группы, состоящей из генетицина, зеоцина, гигромицина B, ноурсеотрицина и биалафоса.
70. Способ согласно варианту осуществления 65, дополнительно включающий проведение позитивной селекции в отношении экспрессии флуоресцентного маркера, где детекция флуоресценции, испускаемой дрожжевыми клетками-хозяевами, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
71. Способ согласно варианту осуществления 70, где флуоресцентный маркер выбран из группы, состоящей из зеленого флуоресцентного белка, красного флуоресцентного белка, синего флуоресцентного белка, голубого флуоресцентного белка, желтого флуоресцентного белка и оранжевого флуоресцентного белка.
72. Способ согласно варианту осуществления 65, дополнительно включающий проведение позитивной селекции в отношении экспрессии биолюминесцентного маркера, где детекция биолюминесценции, испускаемой дрожжевыми клетками-хозяевами, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
73. Способ согласно варианту осуществления 72, где биолюминесцентный маркер представляет собой люциферазу или экворин.
74. Способ согласно варианту осуществления 65, дополнительно включающий проведение негативной селекции в отношении экспрессии маркера контрселекции, где снижение активности представляющего интерес мембранного белка-мишени при связывании экспонированного антитела с представляющим интерес мембранным белком-мишенью поддается обнаружению по росту дрожжевых клеток-хозяев в среде, содержащей агент, который осуществляет селекцию против клеток, экспрессирующих маркер контрселекции.
75. Способ согласно варианту осуществления 74, где маркер контрселекции выбран из группы, состоящей из CAN1, URA3, MET15, TRP1 и TK.
76. Способ скрининга библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 27 в отношении антитела, которое модулирует активность представляющего интерес GPCR-мишени, причем способ включает культивирование по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 27 в среде, где детекция активации или ингибирования ответа феромонов по меньшей мере в одной дрожжевой клетке-хозяине по сравнению с контрольной дрожжевой клеткой-хозяином, не имеющей антитела, экспонированного в периплазматическом пространстве, указывает на то, что экспонированное антитело в указанной по меньшей мере одной дрожжевой клетке-хозяине связывает и модулирует активность GPCR.
77. Способ согласно варианту осуществления 76, где представляющий интерес GPCR-мишень представляет собой GPCR человека.
78. Способ согласно варианту осуществления 77, дополнительно включающий приведение в контакт GPCR человека с лигандом.
79. Способ согласно варианту осуществления 78, где GPCR обладает конститутивной лиганд-независимой активностью.
80. Способ согласно любому из вариантов осуществления 76-79, где дрожжевая клетка-хозяин представляет собой штамм FAR1, где ингибирование ответа феромонов антителом, выступающим в качестве антагониста, который связывает и ингибирует GPCR в дрожжевой клетке-хозяине, приводит к прекращению остановки клеточного цикла и росту дрожжевой клетки-хозяина.
81. Способ согласно любому из вариантов осуществления 76-79, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном, где активация ответа феромонов антителом, выступающим в качестве агониста, который связывает и активирует GPCR в дрожжевой клетке-хозяине, приводит к увеличенной экспрессии репортерного гена.
82. Способ согласно варианту осуществления 73, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотику, флуоресцентный маркер, биолюминесцентный маркер и маркер контрселекции.
83. Способ согласно любому из вариантов осуществления 1-82, где род дрожжевых клеток-хозяев выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia.
84. Способ согласно варианту осуществления 83, где род дрожжевых клеток-хозяев представляет собой Saccharomyces.
85. Способ согласно варианту осуществления 84, где вид Saccharomyces представляет собой Saccharomyces cerevisiae.
86. Дрожжевая клетка-хозяин, содержащая:
a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина,
b) периплазматический якорный белок, где периплазматический якорный белок связан с антителом, так что антитело экспонируется в периплазматическом пространстве; и
c) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок находится на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
87. Дрожжевая клетка-хозяин согласно варианту осуществления 86, где антитело и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий в комплексе или связаны посредством ковалентной непептидной связи в комплексе.
88. Дрожжевая клетка-хозяин согласно варианту осуществления 86, где антитело и периплазматический якорный белок ковалентно связаны в слитом белке.
89. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-88, где периплазматический якорный белок дополнительно содержит сигнальную последовательность, которая обеспечивает транспорт периплазматического якорного белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что антитело экспонируется в периплазме.
90. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-88, где периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или связанный с мембраной белковый домен, который выставляет антитело в периплазму.
91. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-88, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что антитело выступает в периплазму.
92. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-88, где периплазматический якорный белок является достаточно большим, чтобы периплазматический якорный белок и связанное антитело удерживались в периплазме.
93. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-92, где представляющий интерес мембранный белок-мишень выбран из группы, состоящей из рецептора, ионного канала и переносчика.
94. Дрожжевая клетка-хозяин согласно варианту осуществления 93, где рецептор представляет собой сопряженный с G-белком рецептор (GPCR).
95. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-94, дополнительно включающая введение в дрожжевую клетку-хозяина рекомбинантного полинуклеотида, кодирующего сконструированную субъединцу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся обнаружению ответ феромонов в дрожжевой клетке-хозяине.
96. Дрожжевая клетка-хозяин согласно варианту осуществления 95, где сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен субъединицы G(дрожжей и C-концевой домен экзогенной субъединицы Gα.
97. Дрожжевая клетка-хозяин согласно варианту осуществления 96, где субъединица G(дрожжей принадлежит семейству G-белков G(i, G(q, G(s или G(o.
98. Дрожжевая клетка-хозяин согласно варианту осуществления 96 или 97, где экзогенная субъединица G(представляет собой субъединицу G(млекопитающего.
99. Дрожжевая клетка-хозяин согласно варианту осуществления 98, где по меньшей мере пять C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
100. Дрожжевая клетка-хозяин согласно варианту осуществления 99, где по меньшей мере 20 C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
101. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-100, где дрожжевая клетка-хозяин представляет собой штамм FAR1 для селекции антител-антагонистов представляющего интерес GPCR-мишени.
102. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-101, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном для селекции антител-агонистов GPCR.
103. Дрожжевая клетка-хозяин согласно любому из вариантов осуществления 86-101, где род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia.
104. Дрожжевая клетка-хозяин согласно варианту осуществления 103, где род дрожжевой клетки-хозяина представляет собой Saccharomyces.
105. Дрожжевая клетка-хозяин согласно варианту осуществления 104, где веди Saccharomyces представляет собой Saccharomyces cerevisiae.
106. Антитело, связанное с периплазматическим якорным белком.
107. Антитело согласно варианту осуществления 106, где антитело локализуется в периплазматическом пространстве дрожжевой клетки-хозяина.
108. Антитело согласно варианту осуществления 106, где когда антитело продуцируется в дрожжевой клетке-хозяине, антитело локализуется в периплазматическом пространстве дрожжевой клетки-хозяина.
109. Антитело согласно любому из вариантов осуществления 106-108, где антитело и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий в комплексе или связаны посредством ковалентной непептидной связи в комплексе.
110. Антитело согласно любому из вариантов осуществления 106-108, где антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
111. Антитело согласно любому из вариантов осуществления 107-110, где периплазматический якорный белок дополнительно содержит сигнальную последовательность, которая обеспечивает транспорт периплазматического якорного белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что антитело экспонируется в периплазме.
112. Антитело согласно любому из вариантов осуществления 107-110, где периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или связанный с мембраной белковый домен, который выставляет антитело в периплазму.
113. Антитело согласно варианту осуществления 112, где связанный с мембраной белковый домен представляет собой заякоривающий на плазматической мембране гликозилфосфатидилинозитол (GPI)-домен.
114. Антитело согласно любому из вариантов осуществления 107-110, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что антитело выступает в периплазму.
115. Антитело согласно любому из вариантов осуществления 107-110, где периплазматический якорный белок является достаточно большим, чтобы периплазматический якорный белок и связанное антитело удерживались в периплазме.
116. Антитело согласно любому из вариантов осуществления 106-115, где антитело выбрано из группы, состоящей из моноклонального антитела, химерного антитела, гуманизированного антитела, наноантитела, рекомбинантного фрагмента антитела, Fab-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента, Fv-фрагмента и scFv-фрагмента.
117. Антитело согласно любому из вариантов осуществления 107-116, где род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia.
118. Антитело согласно варианту осуществления 117, где род дрожжевой клетки-хозяина представляет собой Saccharomyces.
119. Антитело согласно варианту осуществления 118, где вид Saccharomyces представляет собой Saccharomyces cerevisiae.
120. Дрожжевая клетка-хозяин, содержащая антитело согласно любому из вариантов осуществления 106-119.
121. Способ локализации антитела в периплазматическом пространстве дрожжевой клетки-хозяина, включающий связывание антитела с периплазматическимм якорным белком, так чтобы антитело локализовалось в периплазматическом пространстве.
122. Способ согласно варианту осуществления 121, где антитело и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий в комплексе или связаны посредством ковалентной непептидной связи в комплексе.
123. Способ согласно варианту осуществления 122, где антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
124. Способ согласно любому из вариантов осуществления 121-123, где периплазматический якорный белок дополнительно содержит сигнальную последовательность, которая обеспечивает транспорт периплазматического якорного белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так чтобы антитело экспонировалось в периплазме.
125. Способ согласно любому из вариантов осуществления 121-123, где периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или связанный с мембраной белковый домен, который выставляет антитело в периплазму.
126. Способ согласно варианту осуществления 125, где связанный с мембраной белковый домен представляет собой заякоривающий на плазматической мембране гликозилфосфатидилинозитол (GPI)-домен.
127. Способ согласно любому из вариантов осуществления 121-123, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что антитело выступает в периплазму.
128. Способ согласно любому из вариантов осуществления 120-123, где периплазматический якорный белок является достаточно большим, чтобы периплазматический якорный белок и связанное антитело удерживались в периплазме.
129. Способ согласно любому из вариантов осуществления 121-128, где антитело выбрано из группы, состоящей из моноклонального антитела, химерного антитела, гуманизированного антитела, наноантитела, рекомбинантного-фрагмента антитела, Fab-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента, Fv-фрагмента и scFv-фрагмента.
130. Способ согласно любому из вариантов осуществления 121-129, где род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia.
131. Способ согласно варианту осуществления 130, где род дрожжевой клетки-хозяина представляет собой Saccharomyces.
132. Способ согласно варианту осуществления 131, где вид Saccharomyces представляет собой Saccharomyces cerevisiae.
133. Библиотека дрожжевого периплазматического дисплея, содержащая множество дрожжевых клеток-хозяев, где каждая дрожжевая клетка-хозяин содержит:
a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител;
b) периплазматический якорный белок, где периплазматический якорный белок связан с антителом так, что антитело экспонируется в периплазматическом пространстве; и
c) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок локализуется на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
134. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
135. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где антитело и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий.
136. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, дополнительно содержащая репортерную систему, содержащую репортерный ген, функционально связанный с индуцибельным промотором, который активируется, когда активируется представляющий интерес мембранный белок-мишень, позволяя детекцию увеличения или снижения активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью.
137. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 136, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотику, флуоресцентный маркер, биолюминесцентный маркер или маркер контрселекции.
138. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 137, где алиментарный маркер выбран из группы, состоящей из HIS3, HIS7, ARG6, LEU2, URA3 и TRP1.
139. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 137, где маркер резистентности к антибиотику сообщает резистентность к антибиотику, выбранному из группы, состоящей из генетицина, зеоцина, гигромицина B, ноурсеотрицина и биалафоса.
140. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 137, где флуоресцентный маркер выбран из группы, состоящей из зеленого флуоресцентного белка, красного флуоресцентного белка, синего флуоресцентного белка, голубого флуоресцентного белка, желтого флуоресцентного белка и оранжевого флуоресцентного белка.
141. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 137, где биолюминесцентным маркером является люцифераза или экворин.
142. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 137, где маркер контрселекции выбран из группы, состоящей из CAN1, URA3, MET15, TRP1 и TK.
143. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 136, где репортерный ген представляет собой селективный маркер, так что увеличение активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью поддается детекции посредством роста дрожжевых клеток-хозяев на среде для позитивной селекции.
144. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 136, где репортерный ген представляет собой маркер контрселекции, так что указанное снижение активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью поддается обнаружению посредством роста дрожжевых клеток-хозяев на среде для негативной селекции.
145. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где представляющий интерес мембранный белок-мишень выбран из группы, состоящей из рецептора, ионного канала и переносчика.
146. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 145, где рецептор представляет собой сопряженный с G-белком рецептор (GPCR).
147. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 146, где GPCR представляет собой экзогенный GPCR.
148. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 147, дополнительно содержащая сконструированную субъединицу G(способную активироваться посредством экзогенного GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся детекции ответ феромонов в дрожжевой клетке-хозяине.
149. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 148, где сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен субъединицы G(дрожжей и C-концевой домен экзогенной субъединицы Gα.
150. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 149, где субъединица G(дрожжей принадлежит семейству G-белков G(i, G(q, G(s или G(o.
151. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 149, где экзогенный GPCR представляет собой GPCR млекопитающего.
152. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 151, где по меньшей мере пять C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
153. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 152, где по меньшей мере 20 C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
154. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 151, где субъединица G(млекопитающего выбрана из группы, состоящей из G альфа-S, G альфа-I, G альфа-O, G альфа-T, G альфа-Z, G альфа-Q, G альфа-11, G альфа-12, G альфа-13 и трансдуцина.
155. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 149, где химерная субъединица G(содержит по меньшей мере 41 N-концевой остаток субъединицы G(дрожжей.
156. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 151, где GPCR млекопитающих представляет собой GPCR человека.
157. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 156, где GPCR человека выбран из группы, состоящей из CXCR4, b2AR, CXCR2, CYSLTR2, KSHV vGPCR, PKR1, PKR2, CB2, A3AR и AT1R.
158. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 146, где GPCR обладает конститутивной лиганд-независимой активностью.
159. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 146, где дрожжевая клетка-хозяин представляет собой штамм FAR1 для селекции антител-антагонистов GPCR.
160. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 146, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном для селекции антител-агонистов GPCR.
161. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где антитела выбраны из группы, состоящей из моноклональных антител, химерных антител, гуманизированных антител, наноантител, рекомбинантных фрагментов антител, Fab-фрагментов, Fab'-фрагментов, F(ab')2-фрагментов, Fv-фрагментов и scFv-фрагментов.
162. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где представляющий интерес мембранный белок-мишень содержит мутацию, которая повышает или снижает его активность.
163. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, Δsst2, Δste14, Δste3 или Δmat.
164. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 163 где штамм Δmat содержит удаленный или инактивированный локус MAT(или удаленный или инактивированный локус MATa.
165. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где дрожжевая клетка-хозяин дополнительно содержит модифицированный белок CLN3, содержащий C-концевое укорочение, которое повышает содержание CLN3 в дрожжевой клетке-хозяине по сравнению с белком CLN3 дикого типа.
166. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 165, где модифицированный белок CLN3 сохраняет по меньшей мере N-концевые аминокислоты 1-387 белка CLN3 дикого типа.
167. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 166, где модифицированный белок CLN3 сохраняет по меньшей мере N-концевые аминокислоты 1-408 белка CLN3 дикого типа.
168. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где дрожжевая клетка-хозяин представляет собой гаплоидную или диплоидную дрожжевую клетку-хозяина.
169. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где периплазматический якорный белок дополнительно содержит сигнальную последовательность, которая обеспечивает транспорт слитого белка в периплазму дрожжевой клетки-хозяина на плазматическую мембрану или на клеточную стенку, так что слитый вариант белка экспонируется в периплазме.
170. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или связанный с мембраной белковый домен, который выставляет слитый вариант белка в периплазму.
171. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 170, где связанный с мембраной белковый домен представляет собой заякоривающий на плазматической мембране гликозилфосфатидилинозитол (GPI)-домен.
172. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 171, где заякоривающий на плазматической мембране GPI-домен представляет собой заякоривающий на плазматической мембране GPI-домен япсина.
173. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 172, где заякоривающий на плазматической мембране GPI-домен япсина представляет собой заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7.
174. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что экспонированный вариант белка выступает в периплазму.
175. Библиотека дрожжевого периплазматического дисплея согласно варианту осуществления 133, где периплазматический якорный белок является достаточно большим, чтобы слитый белок удерживался в периплазме.
176. Способ получения библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 134, причем способ включает:
a) предоставление множества рекомбинантных полинуклеотидов, кодирующих слитые белки, где каждый рекомбинантный полинуклеотид кодирует отличающийся слитый белок, содержащий периплазматический якорный белок, связанный с отличающимся антителом, для дисплея;
b) трансфекцию множества дрожжевых клеток-хозяев множеством рекомбинантных полинуклеотидов, кодирующих слитые белки;
c) трансфекцию множества дрожжевых клеток-хозяев рекомбинантным полинуклеотидом, кодирующим представляющий интерес мембранный белок-мишень; и
d) культивирование множества дрожжевых клеток-хозяев в условиях, которые позволяют экспрессию слитых белков и представляющего интерес мембранного белка-мишени, где каждая дрожжевая клетка-хозяин экспонирует отличающееся антитело в периплазматическое пространство и представляющий интерес мембранный белок-мишень локализуется на плазматической мембране, с получением библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 134.
177. Способ согласно варианту осуществления 176, где рекомбинантные полинуклеотиды, кодирующие слитые белки, или рекомбинантный полинуклеотид, кодирующий представляющий интерес мембранный белок-мишень, предоставлен посредством экспрессирующих векторов.
178. Способ согласно варианту осуществления 176, где рекомбинантные полинуклеотиды, кодирующие слитые белки или представляющий интерес мембранный белок-мишень, встраиваются в геном дрожжевой клетки-хозяина в заданном локусе.
179. Способ согласно варианту осуществления 176, где представляющий интерес мембранный белок-мишень выбран из группы, состоящей из рецептора, ионного канала и переносчика.
180. Способ согласно варианту осуществления 179, где рецептор представляет собой сопряженный с G-белком рецептор (GPCR).
181. Способ согласно варианту осуществления 180, дополнительно включающий введение в множество дрожжевых клеток-хозяев рекомбинантного полинуклеотида, кодирующего сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица G(способна активировать поддающийся детекции ответ феромонов в дрожжевой клетке-хозяине.
182. Способ согласно варианту осуществления 181, где сконструированная субъединица G(представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен дрожжевой субъединицы G(и C-концевой домен экзогенной субъединицы Gα.
183. Способ согласно варианту осуществления 182, где субъединица G(дрожжей принадлежит семейству G-белков G(i, G(q, G(s или G(o.
184. Способ согласно варианту осуществления 182, где экзогенная субъединица G(представляет собой субъединицу G(млекопитающего.
185. Способ согласно варианту осуществления 184, где по меньшей мере пять C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
186. Способ согласно варианту осуществления 185, где по меньшей мере 20 C-концевых остатков субъединицы G(дрожжей заменены соответствующими C-концевыми остатками субъединицы G(млекопитающего, так что химерная субъединица G(способна активироваться посредством GPCR млекопитающего.
187. Способ согласно варианту осуществления 181, где дрожжевая клетка-хозяин представляет собой штамм FAR1 для селекции антител-антагонистов представляющего интерес GPCR-мишени.
188. Способ согласно варианту осуществления 181, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном для селекции антител-агонистов GPCR.
189. Способ скрининга библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 136 в отношении антитела, которое модулирует активность представляющего интерес мембранного белка-мишени, причем способ включает культивирование по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 136 в селективной среде; и детекцию экспрессии репортерного гена, где увеличенная экспрессия репортерного гена указывает на то, что антитело повышает активность представляющего интерес мембранного белка-мишени, и сниженная экспрессия репортерного гена указывает на то, что антитело снижает активность представляющего интерес мембранного белка-мишени.
190. Способ согласно варианту осуществления 189, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотикам, флуоресцентный маркер, биолюминесцентный маркер и маркер контрселекции.
191. Способ согласно варианту осуществления 190, дополнительно включающий проведение позитивной селекции в отношении экспрессии алиментарного маркера, где рост дрожжевых клеток-хозяев в селективной среде с дефицитом питательных веществ указывает на то, что представляющий интерес мембранный белок-мишень активирован.
192. Способ согласно варианту осуществления 191, где алиментарный маркер представляет собой HIS3, HIS7, ARG6, LEU2, URA3 и TRP1.
193. Способ согласно варианту осуществления 190, дополнительно включающий проведение позитивной селекции в отношении экспрессии маркера резистентности к антибиотику, где рост дрожжевых клеток-хозяев в селективной среде, содержащей антибиотик, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
194. Способ согласно варианту осуществления 193, где маркер резистентности к антибиотику сообщает резистентность к антибиотику, выбранному из группы, состоящей из генетицина, зеоцина, гигромицина B, ноурсеотрицина и биалафоса.
195. Способ согласно варианту осуществления 190, дополнительно включающий проведение позитивной селекции в отношении экспрессии флуоресцентного маркера, где детекция флуоресценции, испускаемой дрожжевыми клетками-хозяевами, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
196. Способ согласно варианту осуществления 195, где флуоресцентный маркер выбран из группы, состоящей из зеленого флуоресцентного белка, красного флуоресцентного белка, синего флуоресцентного белка, голубого флуоресцентного белка, желтого флуоресцентного белка и оранжевого флуоресцентного белка.
197. Способ согласно варианту осуществления 190, дополнительно включающий проведение позитивной селекции в отношении экспрессии биолюминесцентного маркера, где детекция биолюминесценции, испускаемой дрожжевыми клетками-хозяевами, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
198. Способ согласно варианту осуществления 197, где биолюминесцентным маркером является люцифераза или экворин.
199. Способ согласно варианту осуществления 190, дополнительно включающий проведение негативной селекции в отношении экспрессии маркера контрселекции, где снижение активности представляющего интерес мембранного белка-мишени при связывании экспонированного антитела с представляющим интерес мембранным белком-мишенью поддается детекции посредством роста дрожжевых клеток-хозяев в среде, содержащей средство, которое осуществляет селекцию против клеток, экспрессирующих маркер контрселекции.
200. Способ согласно варианту осуществления 199, где маркер контрселекции выбран из группы, состоящей из CAN1, URA3, MET15, TRP1 и TK.
201. Способ скрининга библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 148 в отношении антитела, которое модулирует активность представляющего интерес GPCR-мишени, причем способ включает культивирование по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея согласно варианту осуществления 148 в среде, где детекция активации или ингибирования ответа феромонов по меньшей мере в одной дрожжевой клетке-хозяине по сравнению с контрольной дрожжевой клеткой-хозяином, не имеющей антитела, экспонированного в периплазматическом пространстве, указывает на то, что экспонированное антитело в указанной по меньшей мере одной дрожжевой клетке-хозяине связывает и модулирует активность GPCR.
202. Способ согласно варианту осуществления 201, дополнительно включающий приведение в контакт GPCR человека с лигандом.
203. Способ согласно варианту осуществления 201, где GPCR обладает конститутивной лиганд-независимой активностью.
204. Способ согласно варианту осуществления 201, где дрожжевая клетка-хозяин представляет собой штамм FAR1, где ингибирование ответа феромонов посредством антитела, выступающего в качестве антагониста, которое связывает и ингибирует GPCR в дрожжевой клетке-хозяине, приводит к прекращению остановки клеточного цикла и росту дрожжевой клетки-хозяина.
205. Способ согласно варианту осуществления 201, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном, где активация ответа феромонов посредством антитела, выступающего в качестве агониста, который связывает и активирует GPCR в дрожжевой клетке-хозяине, приводит к повышенной экспрессии репортерного гена.
206. Способ согласно варианту осуществления 205, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотику, флуоресцентный маркер, биолюминесцентный маркер или маркер контрселекции.
IV. Экспериментальная часть
[0159] Изобретение станет более понятным с помощью следующих примеров конкретных вариантов осуществления настоящего изобретения. Эти примеры приведены только для иллюстративных целей, и они не предназначены для ограничения объема настоящего изобретения никоим образом. Различные модификации или изменения ввиду этого могут прийти в голову специалистам в данной области, и они входят в сущность и границы настоящей заявки и объем прилагаемой формулы изобретения.
[0160] Были предприняты усилия для обеспечения точности в отношении использованных чисел (например, количества, температуры и т.д.), однако, безусловно, допустима некоторая экспериментальная ошибка и отклонение.
Пример 1
Дрожжевой дисплей для селекции антител, которые модулируют функцию GPCR
Обзор
[0161] Множество терапевтических мишеней при таких заболеваниях, как злокачественная опухоль и воспаление, вовлекают сопряженные с G-белком рецепторы (GPCR). Однако многие GPCR с наибольшим терапевтическим потенциалом в отношении оказывающих большое влияние заболеваний, трудно преобразовать в лекарственное средство. Хотя низкомолекулярные соединения, влияющие на функцию этих белков, найти легко, они часто являются неспецифическими вследствие структурного сходства между лиганд-связывающими карманами GPCR, потенциально вызывая значительные неспецифические побочные эффекты. В отличие от низкомолекулярных соединений, антитела и родственные аффинные молекулы (например, наноантитела, и ScFv и Fab), являются привлекательным классом терапевтических средств вследствие их потенциально более высокой специфичности, функционального разнообразия и фармакологических свойств. Кроме того, антитела могут лучше взаимодействовать с внеклеточными доменами и петлями, которые могут модулировать структуру (и, таким образом, функцию) GPCR, более сложными путями, чем низкомолекулярные соединения. Однако на сегодняшний день в США не существует ни одного одобренного антительного терапевтического средства против GPCR, и существует только одно в мире, в Японии.
[0162] Современные технологии дрожжевого и фагового дисплея идентифицируют антитела, которые прочно связывают, но часто не влияют на функцию GPCR. Используемые антигены часто представляют собой фрагменты GPCR, которые не репрезентируют функциональный белок, доступный для антитела in vivo, или представляют собой гетерогенно структурированные полноразмерные белковые препараты. Эта технология также не учитывает огромную долю общего функционального разнообразия, поскольку большинство антител никогда не подвергают функциональному анализу. Необходима высокопроизводительная платформа для прямой селекции антител, которые модулируют функцию GPCR.
[0163] Однако является значительно менее простой разработка антител, которые изменяют функцию GPCR (Jo 2015, Hutchings 2010). Это в первую очередь является следствием следующих проблем с множеством существующих решений: 1) отсутствуют используемые антигены. Антигены, происходящие из внеклеточных пептидов или фрагментов GPCR, могут быть хорошо пригодными для разработки антител для вестерн-блоттинга, но не отражают структурно имеющие терапевтическое значение мишени. Кроме того, однородно функционально свернутый полноразмерный белок в липидах или детергентах может быть трудно получить в достаточных количествах для иммунизации, фагового дисплея или дрожжевого дисплея. 2) Антитела, отобранные вследствие их высокой аффинности, по большей части являются нефункциональными; они связываются с областями в GPCR, которые не влияют на функцию. 3) Технологии утрачивают значительную часть разнообразия антител и, таким образом, функциональность отобранных антител. Посредством селекции сначала антител, которые прочно связываются, и исключения остальных, утрачивается значительная часть функционального разнообразия. Клеточные системы млекопитающих были созданы для функционального скрининга подгрупп антител-кандидатов аутокринным образом (Zhang 2014), которые частично решают проблему 2, но вследствие эффективности трансформации (~104) и ограниченной возможности конструировать подвергаемые селекции/скринингу результаты, они ограничены скринингом небольших подгрупп кандидатов.
[0164] Инновация авторов настоящего изобретения включает комбинирование сопряжения GPCR с ответом феромонов дрожжей и экспрессии аффинных молекул, имеющих цис-действие в той же клетке, в высокопроизводительной платформе. Это позволяет прямую и высокопроизводительную функциональную селекцию аффинных молекул (фиг.1). Для этого авторы изобретения объединили две стратегии, которые являются оптимальными для дрожжей:
1. Функциональная экспрессия GPCR человека, сопряженная с данными об ответе феромонов дрожжей
[0165] Дрожжи представляют собой предпочтительную для выбора систему для экспрессии полноразмерных функциональных GPCR человека для биохимических и структурных исследований in vitro. Примечательно, что дрожжи также успешно используют для функционального сопряжения GPCR человека с каскадом ответа феромонов дрожжей и, в зависимости от полученной информации, скрининга/селекции низкомолекулярных пептидных или белковых лигандов, которые функционально взаимодействуют с GPCR (King 1990, Brown 2000, Erlenbach 2001, Minic 2005, Dowell 2009, Dong 2010, Liu 2016). Наиболее "универсальным" способом сопряжения является модификация субъединицы G(дрожжей, Gpa1, для связывания с GPCR человека и передачи сигналов посредством пересадки 5 C-концевых остатков соответствующего G(человека для замены нативных 5 C-концевых аминокислот в Gpa1 с получением "трансплантата Gα" (Conklin 1993, Brown 2000, Erlenbach 2001). Другие способы включают полную замену Gpa1 на полноразмерный соответствующий G(рецептора человека (King 1990), или конструирование более комплексной химеры Gpa1/Gα, в которой различные части комбинированы в гибридный G(человека/дрожжей (Klein 1998, Price 1995).
2. Дрожжевой дисплей аффинных молекул - антител, наноантител и ScFv
[0166] Также существует фундаментальное знание об экспрессии и секреции аффинных молекул, таких как IgG, ScFv, Fab и наноантитела, в дрожжах (Horwitz 1988, Hamilton 2006, Jeong 2011). Было разработано множество антител, являющихся клиническими кандидатами, компаниями, проводящими дрожжевой дисплей, такими как Adimab. Многие академические лаборатории и компании усовершенствовали способы экспрессии этих молекул и "дисплея" их на дрожжах путем слияния аффинных молекул с белками, которые ковалентно связаны с внеклеточной поверхностью клеточной стенки, например, Aga1 и Aga2 (Boder 1997, Bidlingmaier 2015). Для дрожжей были на рабочем уровне разработаны библиотеки, имеющие комплексность 109, которые являются достаточными для обычных исследований связывания и более чем достаточными для платформы авторов настоящего изобретения для функциональной селекции (по причинам, описанным на фиг.1).
[0167] Кроме того, авторы изобретения приложили усилия для того, чтобы привязать секретируемые аффинные молекулы не к клеточной стенке снаружи, как в случае традиционного дрожжевого дисплея, а чтобы связать их с внеклеточной поверхностью клеточной мембраны. Таким образом, аффинная молекула может функционально взаимодействовать в цис-формате с GPCR в мембране. Авторы протестировали четыре разных доменов и определили, что заякоривающий GPI-домен YPS1 (Frieman 2003) является наилучшим, исходя из оценки флуоресценции мембраны клеток, экспрессирующих слитые конструкции GFP с протестированными доменами.
Пример 2
Штамм дрожжей с низким фоновым уровнем
[0168] Авторы изобретения создали штамм дрожжей, который при обработке лигандом GPCR не растет и не склонен к частому возникновению мутаций, которые могут обеспечить рост.Целью авторов изобретения был фоновый/ложноположительный рост 10-7.
[0169] Гаплоидные дрожжи типа скрещивания "a" (клетки "MATa") претерпевают остановку клеточного цикла, когда феромон скрещивания, альфа-фактор (α-фактор) активирует распознающий его рецептор GPCR Ste2. Этот фенотип остановки роста можно использовать для селекции антагонистов Ste2. К сожалению, появляются спонтанные "резистентные к феромонам мутанты с огромной скоростью. Фоновый или "ложноположительный" уровень в родительском гаплоидном штамме авторов изобретения составляет ~10-4, т.е. ~100 колоний вырастают, когда 106 клеток наносят на чашки с α-фактором. Для снижения фонового уровня авторы изобретения сконструировали отвечающий на феромоны диплоидный основной штамм (Herskowitz 1989). Ложноположительные результаты, вызываемые мутациями с потерей функции, встречаются значительно менее часто в диплоидах. Однако нормальные диплоидные дрожжи содержат гены как MATa, и так МАТальфа; клетки a/альфа не экспрессируют Ste2 и не отвечают на феромоны. Авторы изобретения сконструировали диплоидный организм, который ведет себя как гаплоид MATa, путем делеции всего локуса МАТальфа из его генома. Диплоид MATa/Δmat-альфа имел в ~100 раз более низкий фоновый уровень.
[0170] Практически ни одна из "ложноположительных" колоний в диплоидном штамме не отвечала на феромоны вообще. Предположительно, они содержали мутации с приобретением функции, которые инактивировали сигнальный каскад. Таким образом, авторы изобретения разработали селективный маркер, который является активным только в клетках с функционирующим каскадом ответа феромонов. Этот маркер зависит на базального уровня передачи сигнала каскада для его экспрессии - эта передача сигнала не требует феромона или рецептора феромонов и, вместо этого, зависит от стохастической "исходной" активации распознающего G-белка (Hagen 1991; Oehlen 1995). Эта низкая базальная передача сигнала является недостаточной для запуска остановки клеточного цикла; этот ответ требует активации GPCR Ste2 посредством α-фактора. Некоторые конститутивно экспрессируемые гены дрожжей зависят от базальной активности каскада ответа феромонов, подлежащего экспрессии. Авторы изобретения сконструировали свой селективный маркер путем вставки промотора одного такого гена, MFA1, направляющего экспрессию HIS3 - гена, требуемого для синтеза клетками своего собственного гистидина (Daniel 2006). Конструкция P(MFA1)-HIS3 обеспечивает рост клеток в среде, лишенной гистидина (среда H-), только если они могут передавать сигнал от G-белков далее к транскрипционному фактору ответа феромонов Ste12. Сконструированный диплоидный штамм (NIY326), содержащий P(MFA1)-HIS3 и посеянный в среду H- с α-фактором, имел фоновый уровень 10-7.
Таблица 1. Обобщение разработки штамма для снижения фонового уровня. Фоновый уровень представляет собой колонии, образованные на 107 клеток, посеянных на содержащий феромоны агар
Пример 3
Конструирование локализованной в периплазме библиотеки наноантител
[0171] Авторы изобретения сконструировали набор нацеливающих в периплазму экспрессирующий векторов, обеспечивающих экспрессию химеры с N-концевым сигналом секреции, MFalpha PrePro (Brake 1984), за которым следовал линкер из 18 аминокислот, содержавший один эпитоп FLAG (DYKDDDDK) и оканчивающийся заякоривающим гликофосфатидилинозитол (GPI)-доменом YPS1. Сигнал PrePro направляет белок на перемещение в эндоплазматическую сеть и секрецию и позднее отщепляется, в то время как процессинг GPI-домена YPS1 в ER обеспечивает N-концевой GPI-якорь, который удерживает химеру связанной с плазматической мембраной (Frieman 2003). Участки рестрикции сразу после кодирующей последовательности MFalphaPrePro позволяют клонирование аффинных молекул (например, для создания библиотеки кДНК наноантител). Векторы содержат селективный маркер URA3, который позволяет позитивную селекцию в среде с дефицитом урацила, а также контрселекцию (см. ниже).
[0172] Авторы изобретения подтвердили, что эти экспрессирующие векторы локализовали наноантитело на внеклеточной поверхности мембраны путем клонирования наноантитела против GFP (Kirchhoffer 2009). Когда авторы изобретения расщепили клеточные стенки этих клеток и применили GFP внеклеточно, они наблюдали зеленую флуоресценцию, совпадающую с их клеточными мембранами (фиг.4), что подтвердило периплазматическую локализацию антитела против GFP.
[0173] Далее авторы изобретения сконструировали библиотеку наноантител. Авторы использовали в качестве источника библиотеку наноантител с титром клонов 106, предварительно клонированную в вектор E.coli (Salema 2013). Авторы изобретения амплифицировали кодирующие наноантитела последовательности посредством ПЦР, и клонировали их в нацеливающий на плазматическую мембрану вектор посредством гомологичной рекомбинации дрожжей, с использованием платформенного штамма с низким фоновым уровнем NIY326 (van Leeuwen 2015). Авторы изобретения получали ~1×106 независимых колоний на агарозной среде с дефицитом урацила и гистидина (U-/H-). Авторы изобретения соскребли колонии с использованием среды для криогенного хранения и хранили взвесь аликвотами при -80°C (Library 001).
Пример 4
Селекция наноантител, которые действуют в качестве аутокринных антагонистов Ste2
A. Селекция резистентных к феромонам клонов из библиотеки наноантител:
[0174] Авторы изобретения отобрали резистентные к феромонам клоны из библиотеки 001, описанной в примере 3. Авторы изобретения посеяли 108 клеток на несколько чашек U-/H- с α-фактором и инкубировали их в течение 5 суток. Они проанализировали случайную выборку из 90 колоний приблизительно из 300, которые выросли. Далее авторы изобретения определили, была ли их способность расти на α-факторе зависимой от плазмиды. Они отобрали клоны, которые спонтанно утратили плазмиду, путем посева их на среду, содержавшую 5-FOA, которая является токсичной для клеток, экспрессирующих URA3. Затем авторы изобретения протестировали клоны, которые выросли до селекции 5-FOA, но не росли после нее. Они провели анализы гало и обнаружили, что 12 клонов из 90 первоначально выделенных утрачивали их способность расти на агарозной среде, содержавшей α-фактор, после селекции с 5-FOA в анализах гало (фиг.5).
B. Испытание специфичности и участка действия:
[0175] Для тестирования того, являются ли аффинные молекулы наноантител-кандидаты специфическими (т.е. требуют рецептора-мишени Ste2 для блокирования ответа феромонов), авторы изобретения экспрессируют кандидаты в штамме МАТальфа. Клетки МАТальфа экспрессируют рецептор a-фактора Ste3 и не экспрессируют Ste2. Ste3 является GPCR, только отдаленно родственным Ste2, и его лиганд, феромон a-фактор, представляет собой гликозилированный пептид, полностью отличающийся от α-фактора. Для обоих из них каскады передачи сигнала феромонов MATa и МАТальфа после рецептора являются идентичными. Клетки как MATa, так и МАТальфа останавливают их клеточный цикл и активируют индуцируемые феромонами гены в ответ на распознаваемый ими феромон. Тестирующие клетки MATa и МАТальфа авторов изобретения содержат феромон-индуцируемый транскрипционный репортер, P(PRM1)-YFP, который можно использовать для тестирования функции каскада в отсутствии и в присутствии антагонистов-кандидатов. Таким образом, авторы изобретения оценивают специфичность путем сравнения эффекта антагонистов-кандидатов посредством определения остановки клеточного цикла и индукции YFP в клетках MATa и МАТальфа, подвергнутых воздействию распознаваемых ими феромонов. Авторы изобретения отбирают кандидатов, если они выступают в качестве антагонистов только в штамме MATa (Ste2).
C. Оценка участка действия с использованием очищенного наноантитела внеклеточно
[0176] Хотя и маловероятно, антагонизм GPCR мог быть результатом внутриклеточного связывания и нарушения локализации Ste2 на плазматической мембране. Поскольку любое "связывающее" антитело потенциально может действовать таким путем, авторы изобретения тестируют способность кандидатов модулировать функцию GPCR при экзогенном добавлении.
[0177] Авторы изобретения экспрессируют и очищают белки наноантител, добавляют их к клеткам в присутствии феромона и используют рост в присутствии феромонов и анализ индукции репортера для определения их эффекта на функцию Ste2 (и Ste3). Авторы изобретения экспрессируют кандидатов в бактериях с C-концевой 6xHis-меткой с использованием вектора pET28b и клеток BL21(DE3), и очищают их с использованием неденатурирующей аффинной очистки с 6His (Bornhorst 2000). Вследствие их небольшого размера (15 кДа), наноантитела способны диффундировать через клеточную стенку дрожжей (Ries 2012). В дополнение к этому функциональному тесту, авторы изобретения метят рекомбинантные наноантитела с использованием флуоресцентного красителя (Alexa 488, совместимый с длинами волн GFP; набор #A20181, Thermo-Fisher Scientific) и тестируют их способность окрашивать плазматическую мембрану экспрессирующих Ste2 клеток, но не контрольных экспрессирующих Ste3 клеток. Эксперименты по иммунофлуоресценции также позволяют сравнить окрашивание фиксированных клеток с или без их клеточных стенок (которые можно без труда удалить обработкой ферментом литиказой), и, таким образом, подтвердить, что наноантитела могут эффективно диффундировать через клеточную стенку. Наконец, иммунофлуоресценцию используют для подтверждения того, что антагонисты-кандидаты прямо связывают рецептор, а не лиганд, чтобы проявлять их эффекты.
Пример 5
Штаммы для селекции агонистов
[0178] Авторы изобретения конструируют штаммы для селекции агонистов, которые требуют стимуляции Ste2 для роста. В версии платформенного штамма авторов изобретения, лишенного маркера P(MFA1)-HIS3, они выключают функцию остановки индуцированного феромонами клеточного цикла путем делеции обеих геномных копий FAR1 с использованием подхода CRISPR-Cas9 (Horwitz 2015). Далее авторы изобретения заменяют открытую рамку считывания PRM1 в одном из аллелей PRM1 этого диплоидного штамма на HIS3 ORF, получая селективный маркер P(PRM1)-HIS3. Авторы изобретения включали P(PRM1)-HIS3 в другие штаммы и наблюдали отсутствие значительной "просачивания", т.е. эти клетки росли на чашках H- только в присутствии α-фактора. Авторы изобретения создали версии MATa и MAT(этого штамма для тестов специфичности. Как и ожидалось, частота ложноположительных колоний штаммов P(PRM1)-HIS3 является значительно более низкой, чем для штамма для селекции антагонистов, поскольку мутации, которые включают каскад феромонов, являются более редкими, чем те, которые выключают его (Brown 2000). В случае, когда вторы наблюдают большое количество слабо "растущих колоний" на чашках H- при посеве клеток, экспрессирующих библиотеку наноантител, авторы изобретения используют минимальную эмпирически определенную концентрацию ингибитора His3 3-аминотриазола, который блокирует их рост вследствие "просачивания" экспрессии HIS3 (стандартная стратегия для применений селекции с HIS3 на дрожжах).
[0179] Аналогично скринингу антагонистов, авторы изобретения проводят селекцию клонов, экспрессирующих наноантитела-агонисты Ste2, являющиеся кандидатами, на основе роста на чашках H-. Авторы изобретения демонстрируют, является ли рост зависимым от плазмиды/экспрессии и подтверждают специфичность посредством тестирования наилучших результатов в штамме для селекции агонистов Ste3 (с использованием репортера P(PRM1)-HIS3 и P(PRM1)-YFP). Также, как описано выше, авторы изобретения продуцируют наноантитела в бактериях и добавляют их прямо к клеткам для тестирования их эффективности.
Пример 6
Сопряжение GPCR человека с каскадом феромонов дрожжей
A. Панель трансплантатов Gpa1
[0180] Широко используемым способом сопряжения GPCR человека с каскадом феромонов дрожжей является модификация G(дрожжей, Gpa1, чтобы его 5 C-концевых аминокислот были заменены на 5 C-концевых аминокислот G(человека, с получением "трансплантата GPA1" (Brown 2000, Erlenbach 2001). Для каждого рецептора подходящий G(часто находят эмпирическим путем (Dowell 2009). В большинстве случаев сопряжения достигают с использованием трансплантатов для Gαi, Gαq, Gαs или Gαo (Dong 2010). С использованием подхода CRISPR (Horwitz 2015), авторы изобретения создают панель диплоидных штаммов с трансплантатами GPA1, являющихся агонистами и антагонистами, для этих 4 штаммов с трансплантатами Gα.
B. Тестирование GPCR человека в системе авторов изобретения
[0181] В большинстве случаев GPCR человека экспрессируются лучше всего в дрожжах с встроенной в геном конструкции, контролируемой умеренным промотором, подобным P(ACT1) (Shiroishi 2012, Schutz 2016). Часто, хорошо экспрессируемые GPCR представляют собой химеры с N-концевым отщепляемым секреторным сигналом (как правило, MFalpha PrePro), за которым следует эпитопная метка FLAG для иммунодетекции и иногда C-концевой GFP. Авторы изобретения сконструировали векторы с этими признаками, и они могут быть без труда модифицированы в зависимости от случая. Авторы изобретения клонируют рецепторы, приведенные в таблице 2, в эти векторы и тестируют уровень их экспрессии и локализацию на плазматической мембране посредством флуоресцентной микроскопии и/или иммунофлуоресценции с антителом против FLAG.
[0182] Авторы изобретения трансформируют штаммы с трансплантатами конструкциями, экспрессирующими GPCR. Для каждого GPCR проводят тестирование силы его сопряжения с ответом феромонов с использованием репортеров P(PRM1)-HIS3 и P(PRM1)-YFP в штамме для селекции агонистов.
Таблица 2. Терапевтические GPCR человека-мишени, являющиеся кандидатами для сопряжения и выявления антагонистов/агонистов
конститутивный мутант
Ангиогенез (33)
Ангиогенез (33)
Пример 7
Скрининг антител, которые модулируют функцию GPCR
A. Проведение процесса селекции антител с отобранными GPCR
[0183] Авторы изобретения следуют подходу, сходному с их предшествующей работой с Ste2, за исключением следующего. Авторы изобретения используют диверсифицированную библиотеку ScFv человека (гарантированное разнообразие 108-109, Oak Biosciences). Авторы изобретения клонируют эту библиотеку в тот же вектор с аффинной молекулой, что и выше, экспрессирующий ScFv на плазматической мембране в качестве заякоренных на GPI YPS1 химер. Ожидается, что для селекции антагонистов передача сигнала от сопряженных рецепторов не будет достаточно сильной для запуска остановки клеточного цикла. Таким образом, авторы изобретения используют маркер контрселекции P(FUS2)-CAN1 (Erlenbach 2001). P(FUS2) представляет собой феромон-индуцируемый промотор, такой как P(PRM1). CAN1 кодирует переносчик плазматической мембраны для аргинина, а также для токсического аналога аргинина канаванина. Клетки, содержащие P(FUS2)-CAN1, не могут расти на чашках с канаванином, если они содержат активированный GPCR человека, сопряженный с ответом феромонов. Этот фенотип позволяет проводить селекцию антител-антагонистов GPCR человека на чашках с канаванином. Для селекции агонистов авторы изобретения используют маркер P(PRM1)-HIS3, как указано выше. После селекции кандидатов, как описано выше, выделяют свободные от плазмиды производные этих клонов для тестирования зависимости от плазмиды их фенотипов блокирования феромонов.
B. Тестирование специфичности и участка действия антител-кандидатов
1) Тестирование на других GPCR
[0184] Авторы изобретения подтверждают специфичность зависимых от плазмиды кандидатов посредством трансформации плазмид в штаммы, экспрессирующие другие рецепторы. В этом случае, авторы тестируют их на штаммах MATa (Ste2) и MAT((Ste3), а также на штаммах, экспрессирующих по меньшей мере 2 других GPCR человека, сопряженных с ответом феромонов через трансплантат GPA1 (т.е. для проверки того, что агонист или антагонист не действует на другие GPCR).
2) Тестирование с использованием очищенного белка наноантитела внеклеточно
[0185] В то время как наноантитела (15 кДа) могут диффундировать через клеточную стенку дрожжей (Ries 2012), более крупные ScFv (27 кДа) могут быть значительно ограничены. Авторы изобретения проводят иммунофлуоресценцию с меченными ScFv-кандидатами в дрожжах с и без расщепления клеточной стенки.
Пример 8
Тестирование влияния презентации VHH-домена, являющегося агонистом CB2, на скорость роста
[0186] Для определения эффекта презентации агониста на скорость роста дрожжевых клеток авторы изобретения сконструировали штаммы дрожжей, экспрессирующие белок GPCR человека, рецептор каннабиноидов 2 типа человека (рецептор CB2), и трансформировали их либо пустой плазмидой (без VHH), либо плазмидами, экспрессирующими агонист, в которых однодоменное VHH-антитело Ab101 презентируется различными путями. Авторы протестировали четыре различных плазмиды, экспрессирующих VHH-агонист, в которых VHH-домен презентируется посредством 1) короткого линкера, соединенного с якорем плазматической мембраны Yps1, 2) длинного линкера, соединенного с якорем плазматической мембраны Yps1, 3) N-концевой слитой конструкции, соединенной с растворимым периплазматическим ферментом Suc2, и 4) прямой секреции немеченого VHH в периплазму. От двух до пяти независимых клонов выращивали до насыщения в отсутствии селекции. Насыщенные клетки использовали для начала культур с техническими дублями в 96-луночном микропланшете для титрования для автоматизированного измерения поглощения (OD630) в устройстве для считывания планшетов. Использовали два состава культуральной среды. Первый состав культуральной среды не требует экспрессии с репортера ответа феромонов (без селекции). Второй состав культуральной среды был таким же, как и первый, но лишен одной аминокислоты, которая может продуцироваться только в результате экспрессии репортера ответа феромонов (селекция). Измерение поглощения проводили каждые пять минут в течение 48 часов. Для каждой технической реплики проводили компьютерное вычисление максимальной скорости роста из исходных данных поглощения. Все полученные данные наносили на график, и срединная скорость роста указана горизонтальной чертой (фиг.7). Рисунки под графиком изображают каждый способ презентации (фиг.7).
[0187] Каждая из четырех плазмид, экспрессирующих VHH-агонист, увеличивала скорость роста во втором составе культуральной среды по сравнению с клетками, трансформированными пустой плазмидой (без VHH). Это демонстрируют, что для активации рецептора CB2 белка GPCR можно использовать различные способы презентации агониста в периплазме. В частности, авторы изобретения продемонстрировали, что рецептор CB2 может активироваться антителами с VHH-доменом, которые ковалентно связаны с якорным белком плазматической мембраны, либо через длинный, либо через короткий линкер. Авторы изобретения также продемонстрировали, что функциональные антитела-агонисты с VHH-доменом могут быть локализиованы на периплазме путем слияния антитела с периплазматическим белком, Suc2, который является достаточно большим, чтобы слитый белок удерживался в периплазме. Suc2 образует олигомеры, содержащие несколько белков Suc2, связанных посредством нековалентных взаимодействий, и эта мультимеризация требуется для удержания Suc2 в периплазме. Таким образом, эти условиях также демонстрируют задерживание в периплазме частично посредством нековалентных взаимодействий между антителом и якорным белком. Наконец, этот эксперимент демонстрирует, что прямая секреция немеченых антител с VHH-доменом в периплазму может активировать рецептор CB2.
[0188] Хотя предпочтительные варианты осуществления рассматриваемого изобретения описаны подробно, понятно, что можно вносить очевидные изменения без отклонения от сущности и объема изобретения, как описано в настоящем описании.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИДЕНТИФИКАЦИИ ВЕЩЕСТВ | 2003 |
|
RU2325652C2 |
| ДИСПЛЕЙ ИНТЕГРАЛЬНОГО МЕМБРАННОГО БЕЛКА НА ВНЕКЛЕТОЧНЫХ ОБОЛОЧЕЧНЫХ ВИРИОНАХ ПОКСВИРУСА | 2017 |
|
RU2759846C2 |
| СПОСОБ ИДЕНТИФИКАЦИИ ИММУННЫХ СВЯЗЫВАЮЩИХ ВЕЩЕСТВ ПОВЕРХНОСТНЫХ АНТИГЕНОВ КЛЕТКИ | 2010 |
|
RU2577965C2 |
| СОПРЯЖЕННЫЕ С G-БЕЛКОМ РЕЦЕПТОРЫ DROSOPHILA, НУКЛЕИНОВЫЕ КИСЛОТЫ И СПОСОБЫ, ИМЕЮЩИЕ К НИМ ОТНОШЕНИЕ | 2003 |
|
RU2326385C2 |
| СПОСОБЫ ИДЕНТИФИКАЦИИ ИММУННЫХ СВЯЗУЮЩИХ ВЕЩЕСТВ ПОВЕРХНОСТНЫХ АНТИГЕНОВ КЛЕТКИ | 2016 |
|
RU2635186C2 |
| УЛУЧШЕННЫЕ СПОСОБЫ ПОЛУЧЕНИЯ БИБЛИОТЕК ПОЛИНУКЛЕОТИДОВ В ВИРУСАХ ОСПОВАКЦИНЫ/ЭУКАРИОТИЧЕСКИХ КЛЕТКАХ | 2017 |
|
RU2747557C2 |
| СПОСОБ ЭКСПРЕССИИ БЕЛКА CRM197 | 2019 |
|
RU2803949C1 |
| СПОСОБ СВЯЗЫВАНИЯ ИНТЕРЕСУЮЩИХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2004 |
|
RU2392324C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2015 |
|
RU2748026C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2828630C2 |
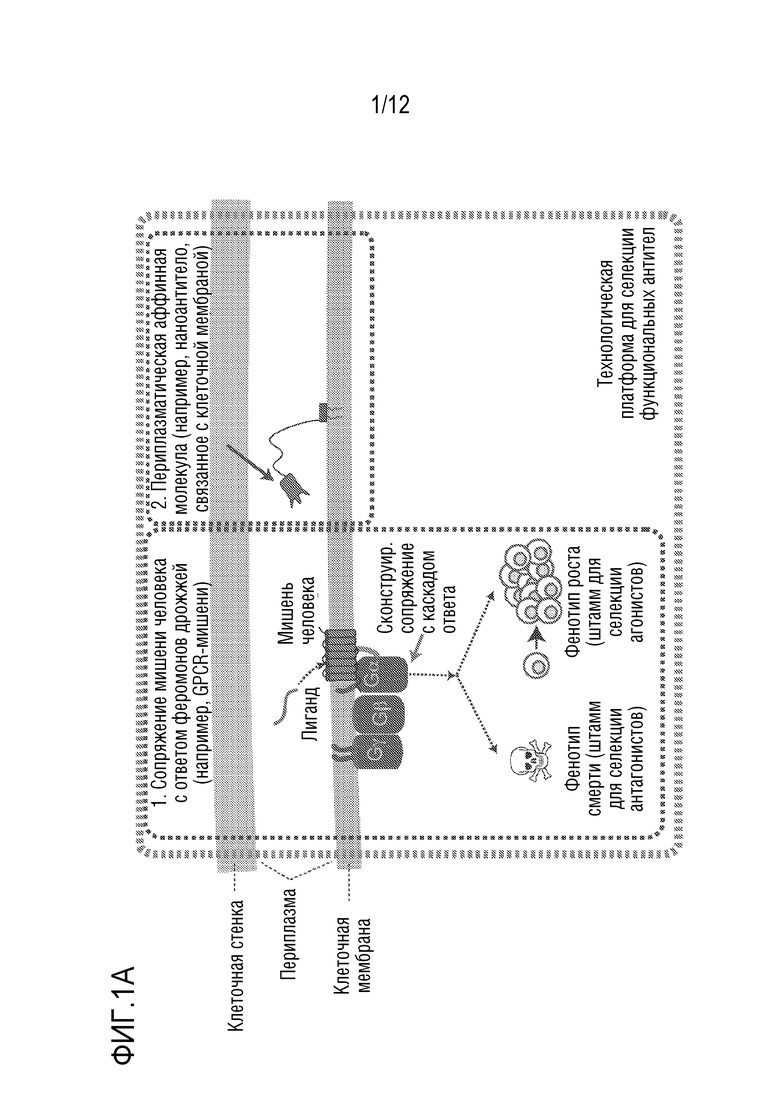
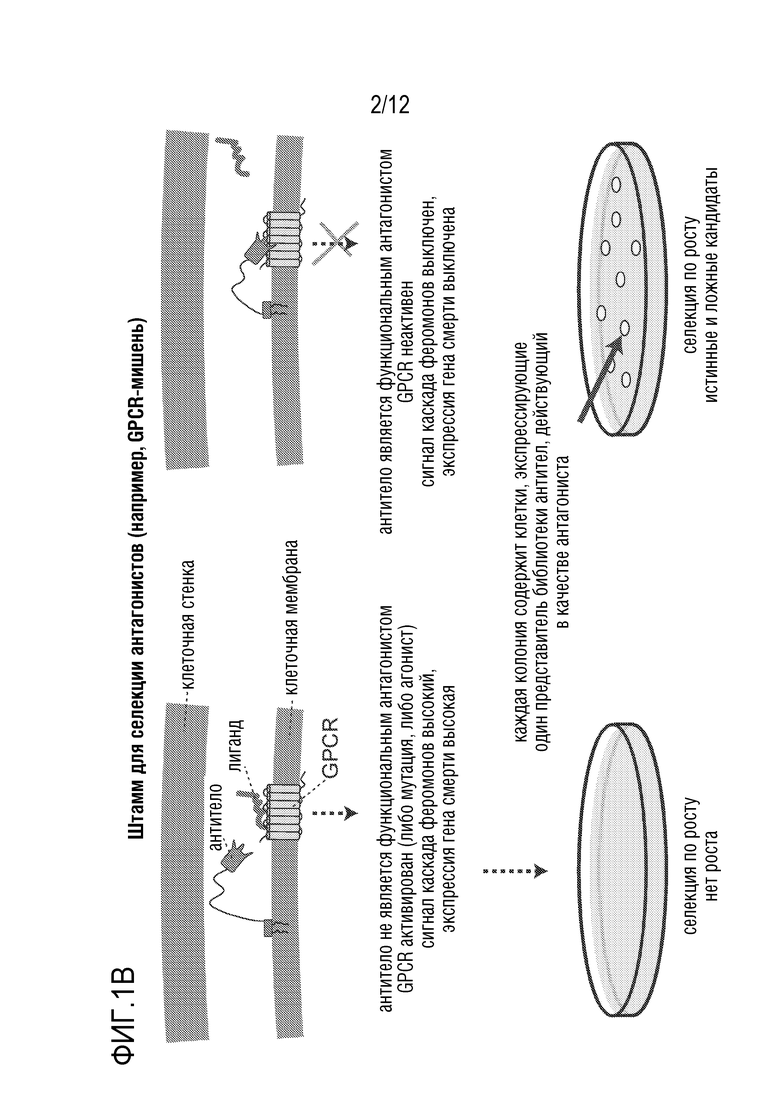
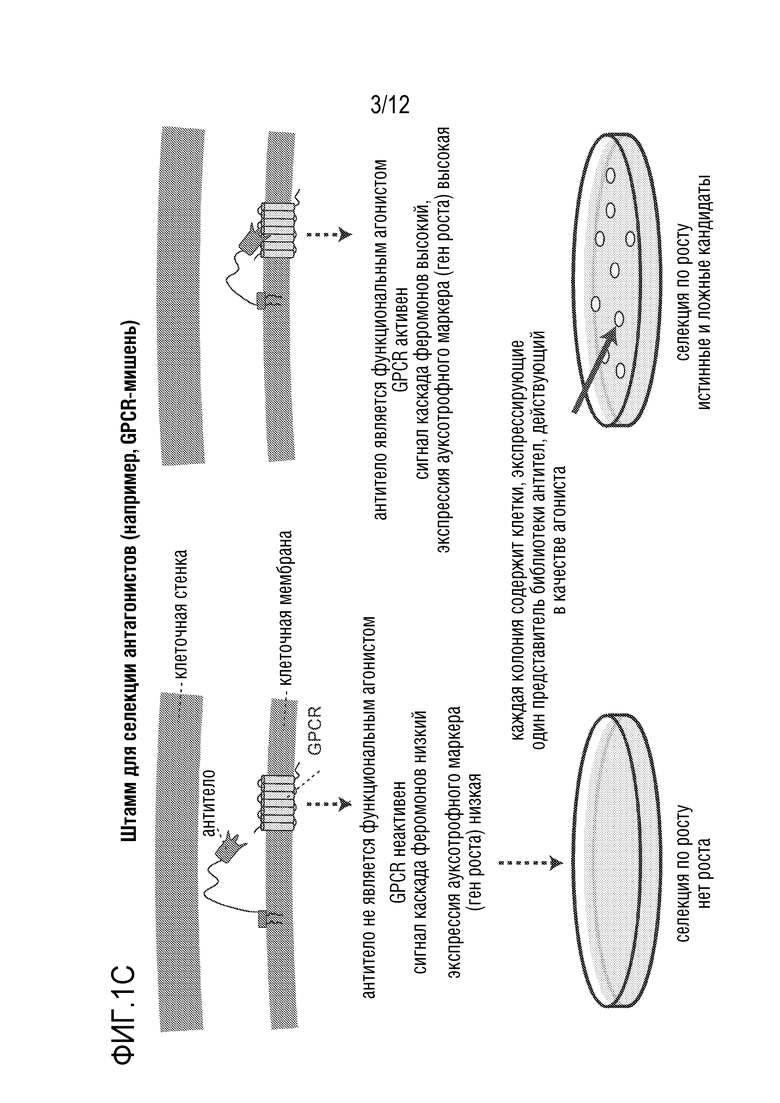
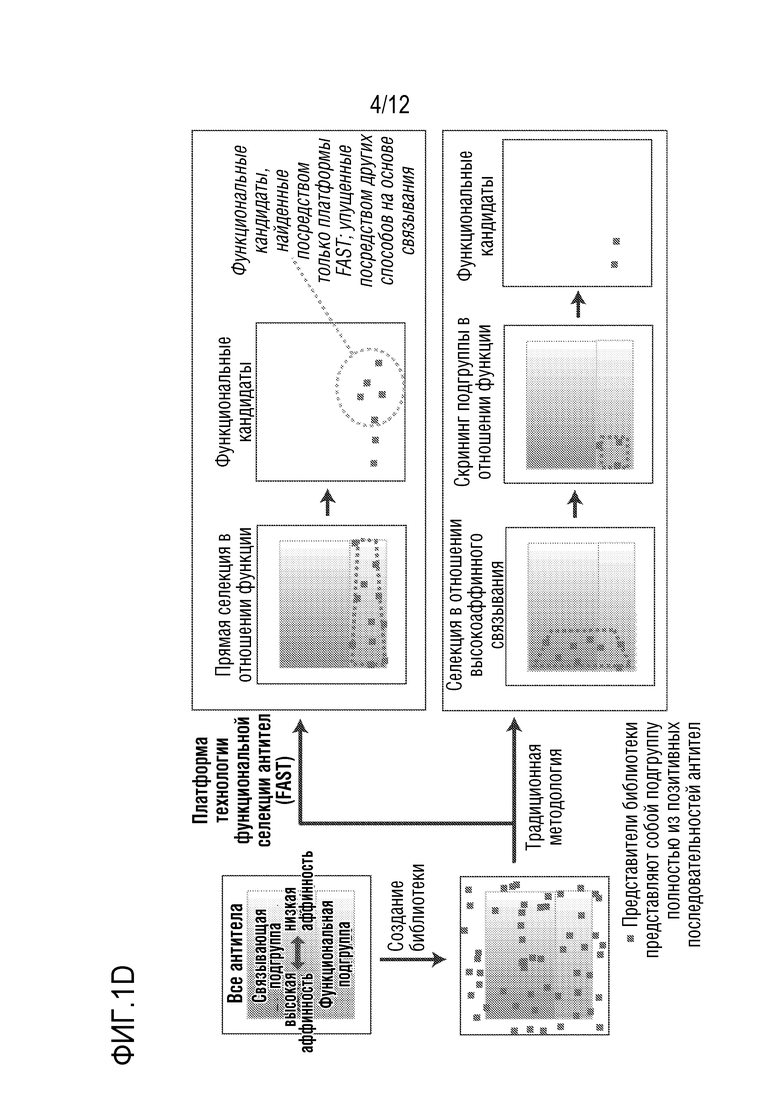
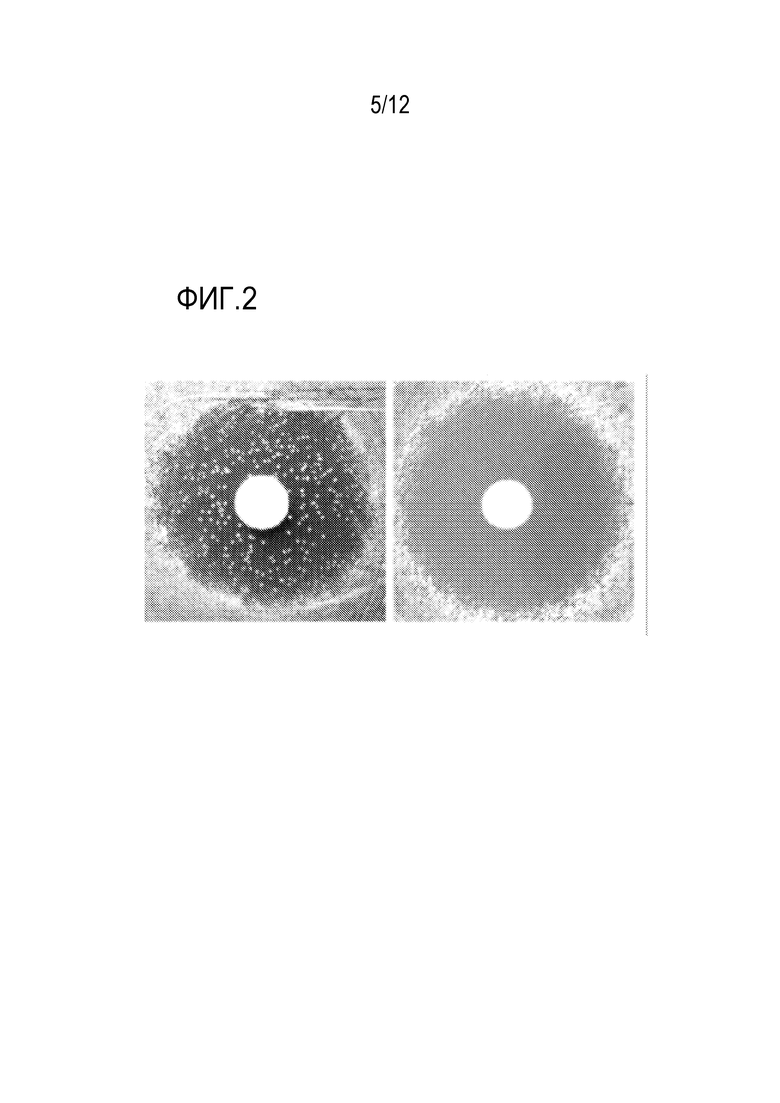
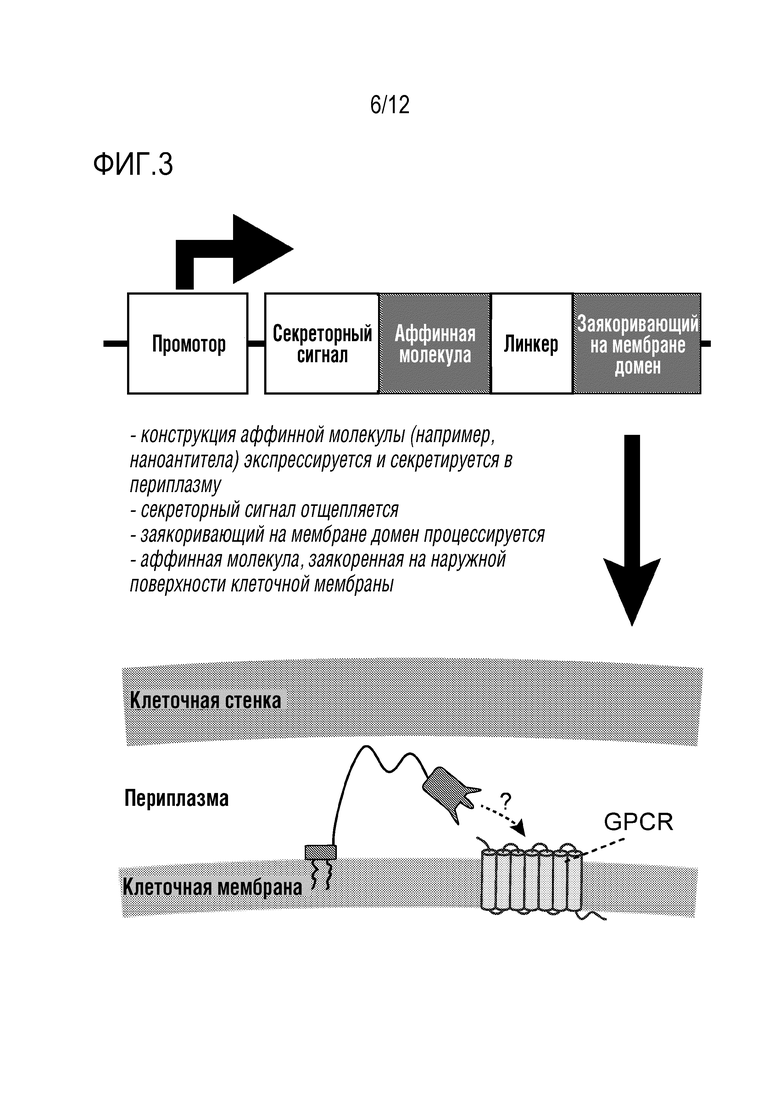
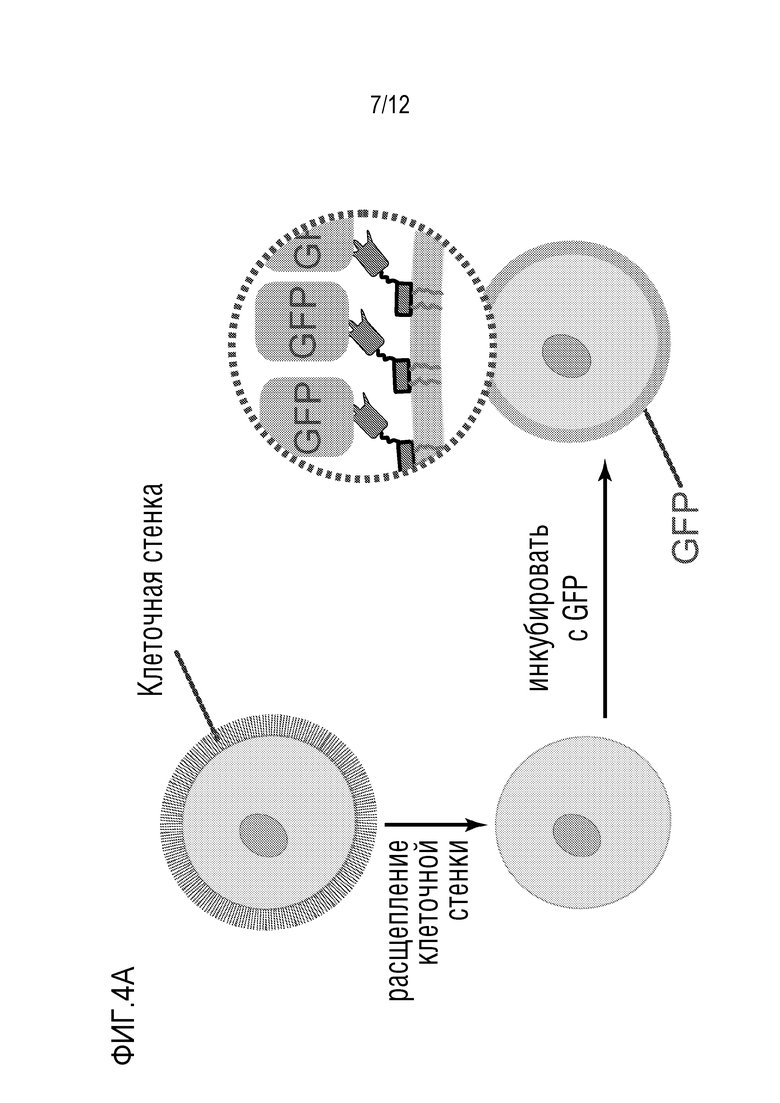
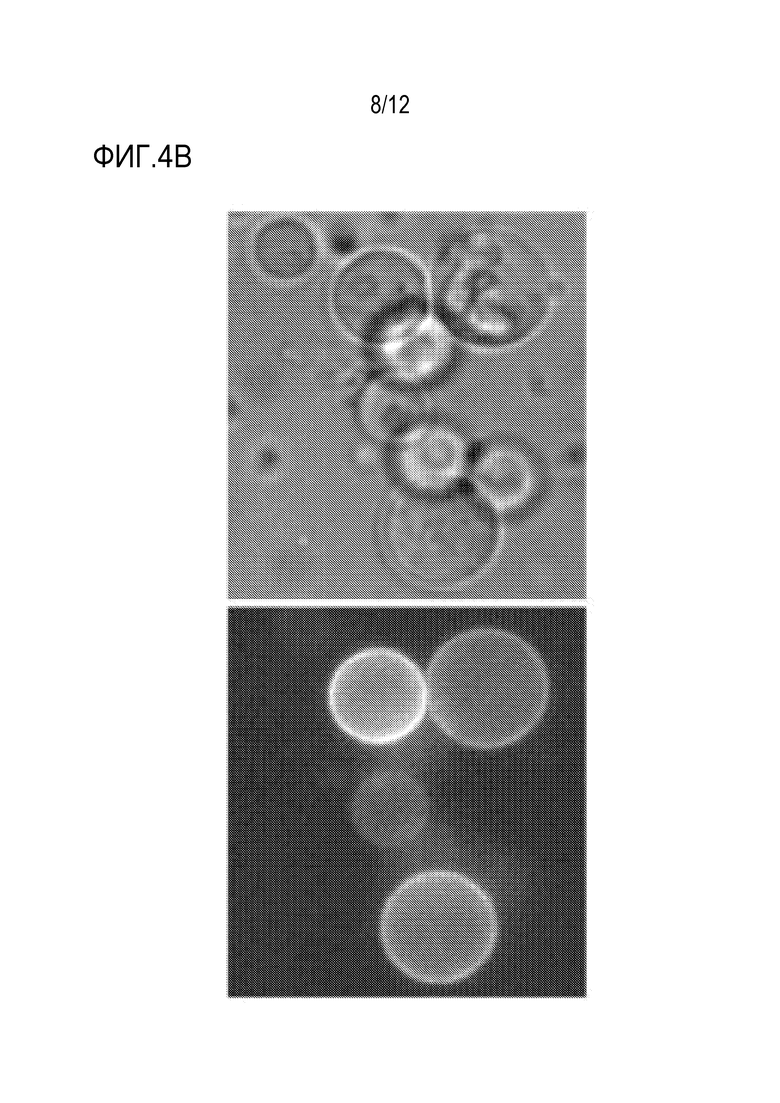
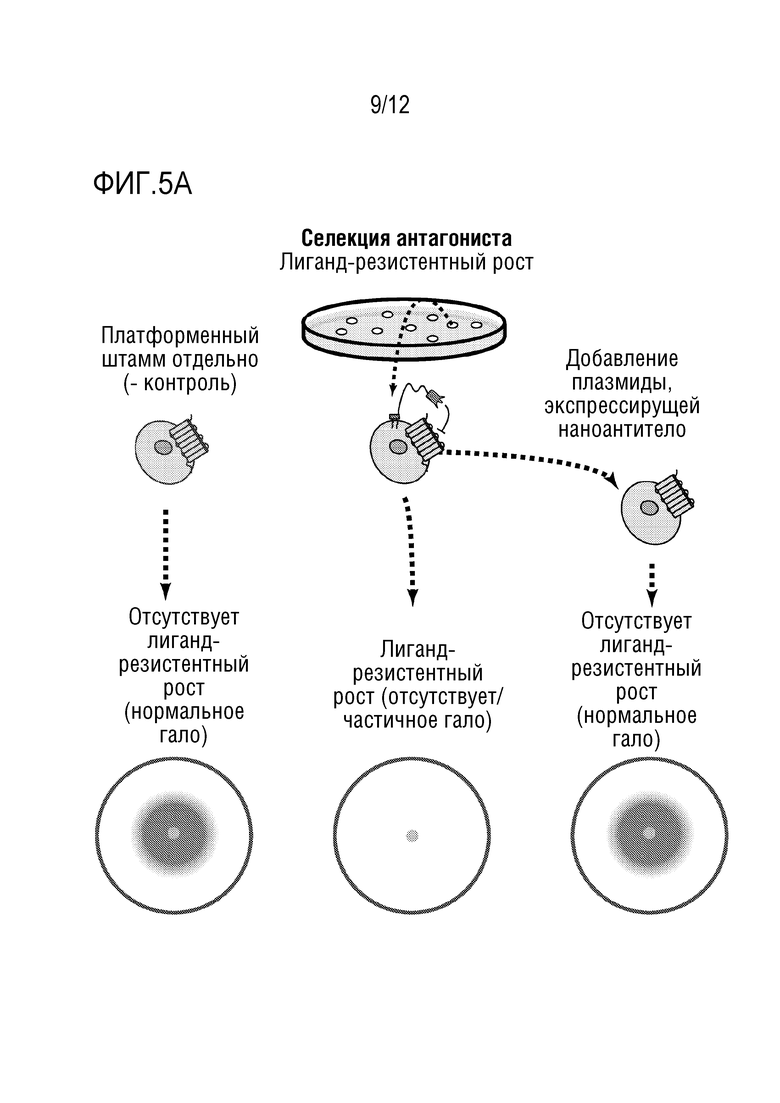
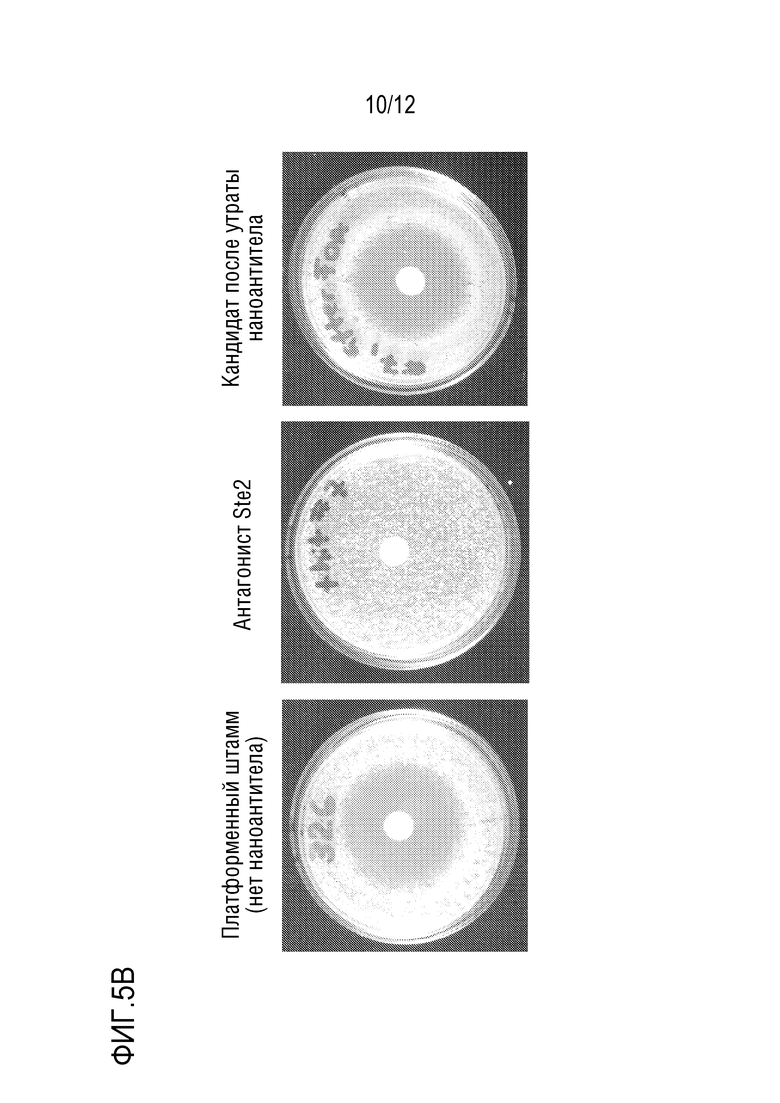
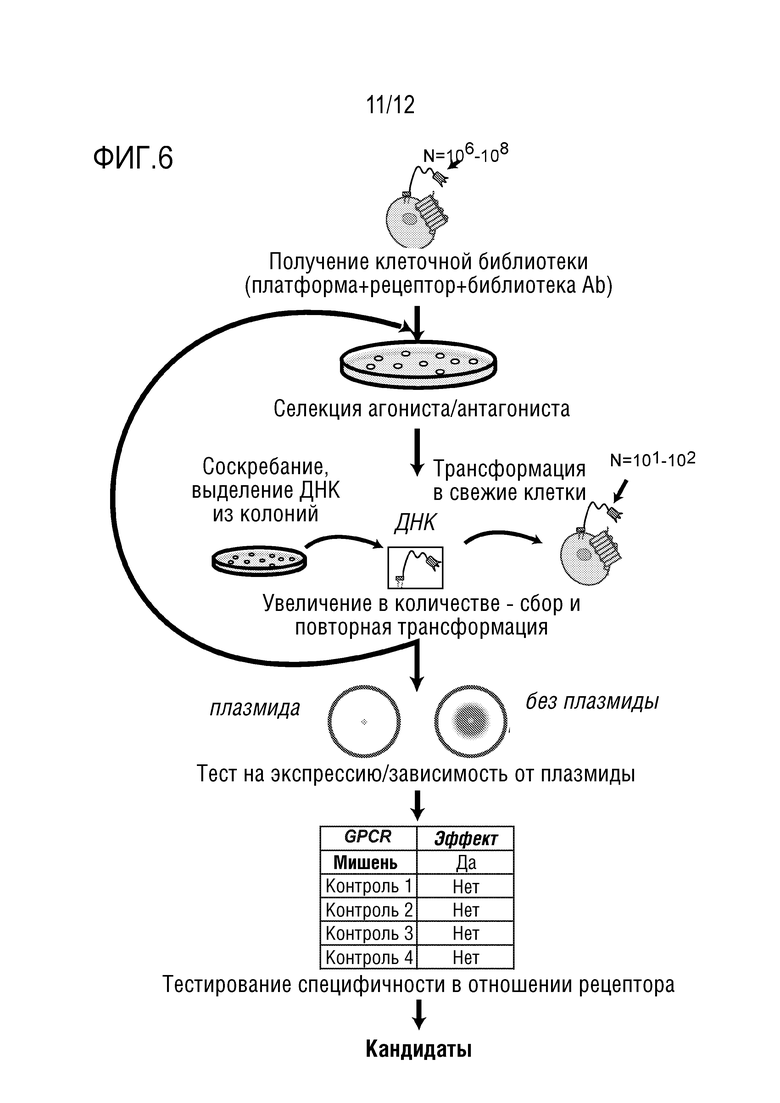
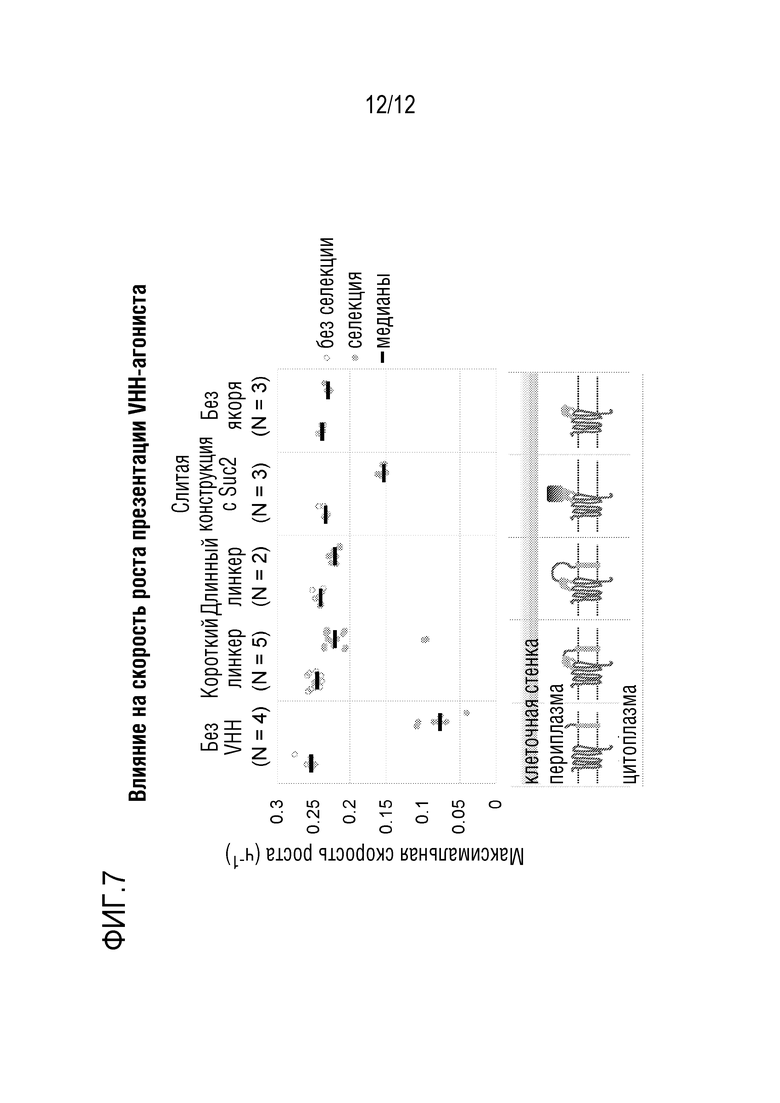
Изобретение относится к биотехнологии. Описаны композиции и способы для дисплея антител в периплазматическом пространстве дрожжевых клеток. Кроме того, изобретение относится к высокопроизводительному скринингу библиотек антител с использованием периплазматического дисплея в дрожжевых клетках. Антитела связаны с проходящим через мембрану трансмембранным доменом, связанным с клеточной мембраной белковым доменом, который находится на наружной поверхности клеточной мембраны дрожжей, белком, который связывается с внутренней поверхностью клеточной стенки дрожжей, или периплазматическим белком для дисплея антител в периплазматическом пространстве дрожжей. В дрожжах можно коэкспрессировать представляющий интерес белок-мишень, так что он локализуется на плазматической мембране или в периплазматическом пространстве и доступен для связывания экспонированными антителами. 8 н. и 109 з.п. ф-лы, 7 ил., 2 табл., 8 пр.
1. Библиотека дрожжевого периплазматического дисплея для выбора антитела из библиотеки антител в отношении специфического связывания или функциональных характеристик, содержащая множество дрожжевых клеток-хозяев, где каждая дрожжевая клетка-хозяин содержит:
a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител;
b) периплазматический якорный белок, где периплазматический якорный белок связан с антителом, так что антитело экспонируется в периплазматическом пространстве; и
c) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок локализуется на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
2. Библиотека дрожжевого периплазматического дисплея по п. 1, где антитело и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий в комплексе или связаны посредством ковалентной непептидной связи в комплексе.
3. Библиотека дрожжевого периплазматического дисплея по п. 1, где антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
4. Библиотека дрожжевого периплазматического дисплея по п. 1, где периплазматический якорный белок дополнительно содержит сигнальную последовательность, которая обеспечивает транспорт периплазматического якорного белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что антитело экспонируется в периплазме.
5. Библиотека дрожжевого периплазматического дисплея по п. 1, где периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или связанный с мембраной белковый домен, который выставляет антитело в периплазму.
6. Библиотека дрожжевого периплазматического дисплея по п. 5, где связанный с мембраной белковый домен представляет собой заякоривающий на плазматической мембране гликозилфосфатидилинозитол (GPI)-домен.
7. Библиотека дрожжевого периплазматического дисплея по п. 6, где заякоривающий на плазматической мембране GPI-домен представляет собой заякоривающий на плазматической мембране GPI-домен япсина.
8. Библиотека дрожжевого периплазматического дисплея по п. 7, где заякоривающий на плазматической мембране GPI-домен япсина представляет собой заякоривающий на плазматической мембране GPI-домен япсина YPS1, YPS2, YPS3, YPS4, YPS5, YPS6 или YPS7.
9. Библиотека дрожжевого периплазматического дисплея по п. 2, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что антитело выступает в периплазму.
10. Библиотека дрожжевого периплазматического дисплея по п. 3, где периплазматический якорный белок представляет собой белок, связывающийся с внутренней поверхностью клеточной стенки, который выставляет слитый белок в периплазму.
11. Библиотека дрожжевого периплазматического дисплея по п. 2, где периплазматический якорный белок является достаточно большим, так что периплазматический якорный белок и связанное антитело удерживаются в периплазме.
12. Библиотека дрожжевого периплазматического дисплея по п. 3, где периплазматический якорный белок является достаточно большим, так что слитый белок удерживается в периплазме.
13. Библиотека дрожжевого периплазматического дисплея для выбора антитела из библиотеки антител в отношении специфического связывания или функциональных характеристик, содержащая множество дрожжевых клеток-хозяев, где каждая дрожжевая клетка-хозяин содержит:
a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител, где антитело связано с сигнальной последовательностью, которая обеспечивает транспорт антитела в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что антитело экспонируется в периплазматическом пространстве дрожжевой клетки-хозяина; и
b) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок локализуется на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
14. Библиотека дрожжевого периплазматического дисплея по п. 1, где каждая дрожжевая клетка-хозяин дополнительно содержит репортерную систему, содержащую репортерный ген, функционально связанный с индуцибельным промотором, который активируется, когда активируется представляющий интерес мембранный белок-мишень, позволяя детекцию увеличения или снижения активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью.
15. Библиотека дрожжевого периплазматического дисплея по п. 14, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотику, флуоресцентный маркер, биолюминесцентный маркер или маркер контрселекции.
16. Библиотека дрожжевого периплазматического дисплея по п. 15, где алиментарный маркер выбран из группы, состоящей из HIS3, HIS7, ARG6, LEU2, URA3 и TRP1.
17. Библиотека дрожжевого периплазматического дисплея по п.15, где маркер резистентности к антибиотику сообщает резистентность к антибиотику, выбранному из группы, состоящей из генетицина, зеоцина, гигромицина B, ноурсеотрицина и биалафоса.
18. Библиотека дрожжевого периплазматического дисплея по п. 15, где флуоресцентный маркер выбран из группы, состоящей из зеленого флуоресцентного белка, красного флуоресцентного белка, синего флуоресцентного белка, голубого флуоресцентного белка, желтого флуоресцентного белка и оранжевого флуоресцентного белка.
19. Библиотека дрожжевого периплазматического дисплея по п. 15, где биолюминесцентным маркером является люцифераза или экворин.
20. Библиотека дрожжевого периплазматического дисплея по п. 15, где маркер контрселекции выбран из группы, состоящей из CAN1, URA3, MET15, TRP1 и TK.
21. Библиотека дрожжевого периплазматического дисплея по п. 14, где репортерный ген представляет собой селективный маркер, так что увеличение активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью поддается детекции посредством роста дрожжевых клеток-хозяев на среде для позитивной селекции.
22. Библиотека дрожжевого периплазматического дисплея по п. 14, где репортерный ген представляет собой маркер контрселекции, так что указанное снижение активности представляющего интерес мембранного белка-мишени при связывании антитела с представляющим интерес мембранным белком-мишенью поддается детекции по росту дрожжевых клеток-хозяев на среде для негативной селекции.
23. Библиотека дрожжевого периплазматического дисплея по любому из пп. 1-22, где представляющий интерес мембранный белок-мишень выбран из группы, состоящей из рецептора, ионного канала и переносчика.
24. Библиотека дрожжевого периплазматического дисплея по п. 23, где рецептор представляет собой сопряженный с G-белком рецептор (GPCR).
25. Библиотека дрожжевого периплазматического дисплея по п. 24, где GPCR представляет собой экзогенный GPCR.
26. Библиотека дрожжевого периплазматического дисплея по п. 25, где дрожжевые клетки-хозяева дополнительно содержат эндогенный GPCR.
27. Библиотека дрожжевого периплазматического дисплея по п. 25, где каждая дрожжевая клетка-хозяин дополнительно содержит сконструированную субъединицу Gα, способную активироваться посредством экзогенного GPCR, где активированная сконструированная субъединица Gα способна активировать поддающийся детекции ответ феромонов в дрожжевой клетке-хозяине.
28. Библиотека дрожжевого периплазматического дисплея по п. 27, где сконструированная субъединица Gα представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен субъединицы Gα дрожжей и C-концевой домен экзогенной субъединицы Gα.
29. Библиотека дрожжевого периплазматического дисплея по п. 28, где субъединица Gα дрожжей принадлежит семейству G-белков Gα i, Gα q, Gα s или Gα o.
30. Библиотека дрожжевого периплазматического дисплея по п.25, где экзогенный GPCR представляет собой GPCR млекопитающего.
31. Библиотека дрожжевого периплазматического дисплея по п. 30, где по меньшей мере пять C-концевых остатков субъединицы Gα дрожжей заменены соответствующими C-концевыми остатками субъединицы Gα млекопитающего, так что химерная субъединица Gα способна активироваться посредством GPCR млекопитающего.
32. Библиотека дрожжевого периплазматического дисплея по п. 31, где по меньшей мере 20 C-концевых остатков субъединицы Gα дрожжей заменены соответствующими C-концевыми остатками субъединицы Gα млекопитающего, так что химерная субъединица Gα способна активироваться посредством GPCR млекопитающего.
33. Библиотека дрожжевого периплазматического дисплея по п. 31, где субъединица Gα млекопитающего выбрана из группы, состоящей из G альфа-S, G альфа-I, G альфа-O, G альфа-T, G альфа-Z, G альфа-Q, G альфа-11, G альфа-12, G альфа-13 и трансдуцина.
34. Библиотека дрожжевого периплазматического дисплея по п. 28, где химерная субъединица Gα содержит по меньшей мере 41 N-концевой остаток субъединицы Gα дрожжей.
35. Библиотека дрожжевого периплазматического дисплея по п. 30, где GPCR млекопитающего представляет собой GPCR человека.
36. Библиотека дрожжевого периплазматического дисплея по п. 35, где GPCR человека выбран из группы, состоящей из CXCR4, CXCR5, SSTR2, MOR, AVPR2, FPR2/ALX, ADORA2A, CHRM3, CGRP2, CCR2, CCR4, CCR5, CHRM4, PAC1, b2AR, CXCR2, CYSLTR2, KSHV vGPCR, PKR1, PKR2, CB1, CB2, A3AR и AT1R.
37. Библиотека дрожжевого периплазматического дисплея по п. 24, где представляющий интерес мембранный белок GPCR, являющийся мишенью, обладает конститутивной лиганд-независимой активностью.
38. Библиотека дрожжевого периплазматического дисплея по п. 24, где дрожжевая клетка-хозяин представляет собой штамм FAR1, содержащий обе геномные копии FAR1, для селекции антител-антагонистов представляющего интерес мембранного белка GPCR, являющегося мишенью.
39. Библиотека дрожжевого периплазматического дисплея по п. 24, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, у которого отсутствуют обе геномные копии FAR1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном для селекции антител-агонистов представляющего интерес мембранного белка GPCR, являющегося мишенью.
40. Библиотека дрожжевого периплазматического дисплея по любому из пп. 1-22, где антитела выбраны из группы, состоящей из моноклональных антител, химерных антител, гуманизированных антител, наноантител, рекомбинантных фрагментов антител, Fab-фрагментов, Fab'-фрагментов, F(ab')2-фрагментов, Fv-фрагментов и scFv-фрагментов.
41. Библиотека дрожжевого периплазматического дисплея по любому из пп. 1-22, где представляющий интерес мембранный белок-мишень содержит мутацию, которая повышает или снижает его активность.
42. Библиотека дрожжевого периплазматического дисплея по любому из пп. 1-22, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, у которого отсутствуют обе геномные копии FAR1, Δsst2, содержаший инактивированный ген SST2, Δste14, у которого отсутствуют обе геномные копии STE14, Δste3, у которого отсутствуют обе геномные копии STE3, или Δmat, содержащий удаленный или инактивированный локус MATα или удаленный или инактивированный локус MATa.
43. Библиотека дрожжевого периплазматического дисплея по п. 42, где дрожжевая клетка-хозяин представляет собой штамм Δmat, содержащий удаленный или инактивированный локус MATα или удаленный или инактивированный локус MATa.
44. Библиотека дрожжевого периплазматического дисплея по любому из пп. 1-22, где дрожжевая клетка-хозяин дополнительно содержит модифицированный белок CLN3, содержащий C-концевое укорочение, которое повышает содержание CLN3 в дрожжевой клетке-хозяине по сравнению с белком CLN3 дикого типа.
45. Библиотека дрожжевого периплазматического дисплея по п. 44, где в модифицированном белке CLN3 сохранены по меньшей мере N-концевые аминокислоты 1-387 белка CLN3 дикого типа.
46. Библиотека дрожжевого периплазматического дисплея по п. 44, где в модифицированном белке CLN3 сохранены по меньшей мере N-концевые аминокислоты 1-408 белка CLN3 дикого типа.
47. Библиотека дрожжевого периплазматического дисплея по любому из пп. 1-22, где дрожжевая клетка-хозяин представляет собой гаплоидную или диплоидную дрожжевую клетку-хозяина.
48. Способ получения библиотеки дрожжевого периплазматического дисплея по п. 1, причем способ включает:
a) предоставление первого множества рекомбинантных полинуклеотидов, кодирующих антитела для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина, где экспонированное антитело является отличающимся в каждой дрожжевой клетке-хозяине, так что множество дрожжевых клеток-хозяев экспонирует множество антител;
b) предоставление второго рекомбинантного полинуклеотида, кодирующего периплазматический якорный белок;
c) трансфекцию множества дрожжевых клеток-хозяев первым множеством рекомбинантных полинуклеотидов и вторым рекомбинантным полинуклеотидом;
d) трансфекцию множества дрожжевых клеток-хозяев рекомбинантным полинуклеотидом, кодирующим представляющий интерес мембранный белок-мишень; и
e) культивирование множества дрожжевых клеток-хозяев в условиях, которые позволяют экспрессию антител, периплазматического якорного белка и представляющего интерес мембранного белка-мишени, где периплазматический якорный белок связан с антителом так, что антитело экспонируется в периплазматическом пространстве, где каждая дрожжевая клетка-хозяин экспонирует отличающееся антитело в периплазматическое пространство и представляющий интерес мембранный белок-мишень локализуется на плазматической мембране, с получением библиотеки дрожжевого периплазматического дисплея по п. 1.
49. Способ по п. 48, где рекомбинантные полинуклеотиды, кодирующие антитела, или рекомбинантный полинуклеотид, кодирующий периплазматический якорный белок или представляющий интерес мембранный белок-мишень, предоставлены посредством экспрессирующих векторов.
50. Способ по п. 48, где рекомбинантные полинуклеотиды, кодирующие антитела, или рекомбинантный полинуклеотид, кодирующий периплазматический якорный белок или представляющий интерес мембранный белок-мишень, встраиваются в геном дрожжевой клетки-хозяина в локусе-мишени.
51. Способ получения библиотеки дрожжевого периплазматического дисплея по п. 3, причем способ включает:
a) предоставление множества рекомбинантных полинуклеотидов, кодирующих слитые белки, где каждый рекомбинантный полинуклеотид кодирует отличающийся слитый белок, содержащий периплазматический якорный белок, связанный с отличающимся антителом, для дисплея;
b) трансфекцию множества дрожжевых клеток-хозяев множеством рекомбинантных полинуклеотидов, кодирующих слитые белки;
c) трансфекцию множества дрожжевых клеток-хозяев рекомбинантным полинуклеотидом, кодирующим представляющий интерес мембранный белок-мишень; и
d) культивирование множества дрожжевых клеток-хозяев в условиях, которые позволяют экспрессию слитых белков и представляющего интерес мембранного белка-мишени, где каждая дрожжевая клетка-хозяин экспонирует отличающееся антитело в периплазматическое пространство и представляющий интерес мембранный белок-мишень локализуется на плазматической мембране, с получением библиотеки дрожжевого периплазматического дисплея по п. 3.
52. Способ по п. 51, где рекомбинантные полинуклеотиды, кодирующие слитые белки, или рекомбинантный полинуклеотид, кодирующий представляющий интерес мембранный белок-мишень, предоставлены посредством экспрессирующих векторов.
53. Способ по п. 51, где рекомбинантные полинуклеотиды, кодирующие слитые белки или представляющий интерес мембранный белок-мишень, встраиваются в геном дрожжевой клетки-хозяина в заданном локусе.
54. Способ по любому из пп. 48-53, где представляющий интерес мембранный белок-мишень выбран из группы, состоящей из рецептора, ионного канала и переносчика.
55. Способ по п. 54, где рецептор представляет собой сопряженный с G-белком рецептор (GPCR).
56. Способ по любому из пп. 48-53, дополнительно включающий введение в множество дрожжевых клеток-хозяев рекомбинантного полинуклеотида, кодирующего сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица Gα способна активировать поддающийся детекции ответ феромонов в дрожжевой клетке-хозяине.
57. Способ по п. 56, где сконструированная субъединица Gα представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен субъединицы Gα дрожжей и C-концевой домен экзогенной субъединицы Gα.
58. Способ по п. 57, где субъединица Gα дрожжей принадлежит семейству G-белков Gα i, Gα q, Gα s или Gα o.
59. Способ по п. 57, где экзогенная субъединица Gα представляет собой субъединицу Gα млекопитающего.
60. Способ по п. 59, где по меньшей мере пять C-концевых остатков субъединицы Gα дрожжей заменены соответствующими C-концевыми остатками субъединицы Gα млекопитающего, так что химерная субъединица Gα способна активироваться посредством GPCR млекопитающего.
61. Способ по п. 60, где по меньшей мере 20 C-концевых остатков субъединицы Gα дрожжей заменены соответствующими C-концевыми остатками субъединицы Gα млекопитающего, так что химерная субъединица Gα способна активироваться посредством GPCR млекопитающего.
62. Способ по п. 55, где дрожжевая клетка-хозяин представляет собой штамм FAR1, содержащий обе геномные копии FAR1, для селекции антител-антагонистов представляющего интерес GPCR-мишени.
63. Способ по п. 55, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, у которого отсутствуют обе геномные копии FAR1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном, для селекции антител-агонистов GPCR.
64. Способ скрининга библиотеки дрожжевого периплазматического дисплея по п. 14 в отношении антитела, которое модулирует активность представляющего интерес мембранного белка-мишени, причем способ включает культивирование по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея по п. 14 в селективной среде; и детекцию экспрессии репортерного гена, где увеличенная экспрессия репортерного гена указывает на то, что антитело повышает активность представляющего интерес мембранного белка-мишени, и сниженная экспрессия репортерного гена указывает на то, что антитело снижает активность представляющего интерес мембранного белка-мишени.
65. Способ по п. 64, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотику, флуоресцентный маркер, биолюминесцентный маркер или маркер контрселекции.
66. Способ по п. 65, дополнительно включающий проведение позитивной селекции в отношении экспрессии алиментарного маркера, где рост дрожжевых клеток-хозяев в селективной среде с дефицитом питательных веществ указывает на то, что представляющий интерес мембранный белок-мишень активирован.
67. Способ по п. 66, где алиментарный маркер представляет собой HIS3, HIS7, ARG6, LEU2, URA3 и TRP1.
68. Способ по п. 65, дополнительно включающий проведение позитивной селекции в отношении экспрессии маркера резистентности к антибиотику, где рост дрожжевых клеток-хозяев в селективной среде, содержащей антибиотик, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
69. Способ по п. 68, где маркер резистентности к антибиотику сообщает резистентность к антибиотику, выбранному из группы, состоящей из генетицина, зеоцина, гигромицина B, ноурсеотрицина и биалафоса.
70. Способ по п. 65, дополнительно включающий проведение позитивной селекции в отношении экспрессии флуоресцентного маркера, где детекция флуоресценции, испускаемой дрожжевыми клетками-хозяевами, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
71. Способ по п. 70, где флуоресцентный маркер выбран из группы, состоящей из зеленого флуоресцентного белка, красного флуоресцентного белка, синего флуоресцентного белка, голубого флуоресцентного белка, желтого флуоресцентного белка и оранжевого флуоресцентного белка.
72. Способ по п. 65, дополнительно включающий проведение позитивной селекции в отношении экспрессии биолюминесцентного маркера, где детекция биолюминесценции, испускаемой дрожжевыми клетками-хозяевами, указывает на то, что представляющий интерес мембранный белок-мишень активирован.
73. Способ по п. 72, где биолюминесцентным маркером является люцифераза или экворин.
74. Способ по п. 65, дополнительно включающий проведение негативной селекции в отношении экспрессии маркера контрселекции, где снижение активности представляющего интерес мембранного белка-мишени при связывании экспонированного антитела с представляющим интерес мембранным белком-мишенью поддается обнаружению по росту дрожжевых клеток-хозяев в среде, содержащей средство, которое осуществляет селекцию против клеток, экспрессирующих маркер контрселекции.
75. Способ по п. 74, где маркер контрселекции выбран из группы, состоящей из CAN1, URA3, MET15, TRP1 и TK.
76. Способ скрининга библиотеки дрожжевого периплазматического дисплея по п. 27 в отношении антитела, которое модулирует активность представляющего интерес GPCR-мишени, причем способ включает культивирование по меньшей мере подгруппы дрожжевых клеток-хозяев библиотеки дрожжевого периплазматического дисплея по п. 27 в среде, где детекция активации или ингибирования ответа феромонов по меньшей мере в одной дрожжевой клетке-хозяине по сравнению с контрольной дрожжевой клеткой-хозяином, не имеющей антитела, экспонированного в периплазматическом пространстве, указывает на то, что экспонированное антитело в указанной по меньшей мере одной дрожжевой клетке-хозяине связывает и модулирует активность GPCR.
77. Способ по п. 76, где представляющий интерес GPCR-мишень представляет собой GPCR человека.
78. Способ по п. 77, дополнительно включающий приведение в контакт GPCR человека с лигандом.
79. Способ по п. 78, где GPCR обладает конститутивной лиганд-независимой активностью.
80. Способ по любому из пп. 76-79, где дрожжевая клетка-хозяин представляет собой штамм FAR1, содержащий обе геномные копии FAR1, где ингибирование ответа феромонов посредством антитела, действующего в качестве антагониста, который связывает и ингибирует GPCR в дрожжевой клетке-хозяине, приводит к прекращению остановки клеточного цикла и росту дрожжевой клетки-хозяина.
81. Способ по любому из пп. 76-79, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, у которого отсутствуют обе геномные копии FAR1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном, где активация ответа феромонов посредством антитела, действующего в качестве агониста, который связывает и активирует GPCR в дрожжевой клетке-хозяине, приводит к повышению экспрессии репортерного гена.
82. Способ по п. 81, где репортерный ген представляет собой алиментарный маркер, маркер резистентности к антибиотику, флуоресцентный маркер, биолюминесцентный маркер или маркер контрселекции.
83. Библиотека дрожжевого периплазматического дисплея по любому из пп. 1-22 или способ по любому пп. 48-53 или 64-75, где род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia.
84. Способ по п. 83, где род дрожжевой клетки-хозяина представляет собой Saccharomyces.
85. Способ по п. 84, где вид Saccharomyces представляет собой Saccharomyces cerevisiae.
86. Дрожжевая клетка-хозяин для применения в библиотеке дрожжевого дисплея, содержащая:
a) антитело для дисплея в периплазматическом пространстве дрожжевой клетки-хозяина,
b) периплазматический якорный белок, где периплазматический якорный белок связан с антителом так, что антитело экспонируется в периплазматическом пространстве; и
c) представляющий интерес мембранный белок-мишень, где представляющий интерес мембранный белок локализуется на плазматической мембране дрожжевой клетки-хозяина и доступен для антитела, экспонированного в периплазматическом пространстве дрожжевой клетки-хозяина.
87. Дрожжевая клетка-хозяин по п. 86, где антитело и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий в комплексе или связаны посредством ковалентной непептидной связи в комплексе.
88. Дрожжевая клетка-хозяин по п. 86, где антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
89. Дрожжевая клетка-хозяин по любому из пп. 86-88, где периплазматический якорный белок дополнительно содержит сигнальную последовательность, которая обеспечивает транспорт периплазматического якорного белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что антитело экспонируется в периплазме.
90. Дрожжевая клетка-хозяин по любому из пп. 86-88, где периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или связанный с мембраной белковый домен, который выставляет антитело в периплазму.
91. Дрожжевая клетка-хозяин по любому из пп. 86-88, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что антитело выступает в периплазму.
92. Дрожжевая клетка-хозяин по любому из пп. 86-88, где периплазматический якорный белок является достаточно большим, так что периплазматический якорный белок и связанное антитело удерживаются в периплазме.
93. Дрожжевая клетка-хозяин по любому из пп. 86-88, где представляющий интерес мембранный белок-мишень выбран из группы, состоящей из рецептора, ионного канала и переносчика.
94. Дрожжевая клетка-хозяин по п. 93, где рецептор представляет собой сопряженный с G-белком рецептор (GPCR).
95. Дрожжевая клетка-хозяин по любому из пп. 86-88, дополнительно включающая рекомбинантный полинуклеотид, кодирующий сконструированную субъединицу Gα, способную активироваться посредством GPCR, где активированная сконструированная субъединица Gα способна активировать поддающийся детекции ответ феромонов в дрожжевой клетке-хозяине.
96. Дрожжевая клетка-хозяин по п. 95, где сконструированная субъединица Gα представляет собой химерную субъединицу G-белка альфа (Gα), содержащую N-концевой домен субъединицы Gα дрожжей и C-концевой домен экзогенной субъединицы Gα.
97. Дрожжевая клетка-хозяин по п. 96, где субъединица Gα дрожжей принадлежит семейству G-белков Gα i, Gα q, Gα s или Gα o.
98. Дрожжевая клетка-хозяин по п. 96, где экзогенная субъединица Gα представляет собой субъединицу Gα млекопитающего.
99. Дрожжевая клетка-хозяин по п. 98, где по меньшей мере пять C-концевых остатков субъединицы Gα дрожжей заменены соответствующими C-концевыми остатками субъединицы Gα млекопитающего, так что химерная субъединица Gα способна активироваться посредством GPCR млекопитающего.
100. Дрожжевая клетка-хозяин по п. 99, где по меньшей мере 20 C-концевых остатков субъединицы Gα дрожжей заменены соответствующими C-концевыми остатками субъединицы Gα млекопитающего, так что химерная субъединица Gα способна активироваться посредством GPCR млекопитающего.
101. Дрожжевая клетка-хозяин по любому из пп. 86-88, где дрожжевая клетка-хозяин представляет собой штамм FAR1, содержащий обе геномные копии FAR1, для селекции антител-антагонистов представляющего интерес GPCR-мишени.
102. Дрожжевая клетка-хозяин по любому из пп. 86-88, где дрожжевая клетка-хозяин представляет собой штамм Δfar1, у которого отсутствуют обе геномные копии FAR1, содержащий феромон-индуцируемый промотор PRM1, функционально связанный с репортерным геном.
103. Дрожжевая клетка-хозяин по любому из пп. 86-88, где род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia.
104. Дрожжевая клетка-хозяин по п. 103, где род дрожжевой клетки-хозяина представляет собой Saccharomyces.
105. Дрожжевая клетка-хозяин по п. 104, где вид Saccharomyces представляет собой Saccharomyces cerevisiae.
106. Способ локализации антитела в периплазматическом пространстве дрожжевой клетки-хозяина, включающий связывание антитела с периплазматическимм якорным белком, так что антитело локализуется в периплазматическом пространстве.
107. Способ по п. 106, где антитело и периплазматический якорный белок нековалентно связаны вместе посредством молекулярных связывающих взаимодействий в комплексе или связаны посредством ковалентной непептидной связи в комплексе.
108. Способ по п. 106, где антитело и периплазматический якорный белок ковалентно связаны вместе в слитом белке.
109. Способ по любому из пп. 106-108, где периплазматический якорный белок дополнительно содержит сигнальную последовательность, которая обеспечивает транспорт периплазматического якорного белка в периплазму дрожжевой клетки-хозяина, на плазматическую мембрану или на клеточную стенку, так что антитело экспонируется в периплазме.
110. Способ по любому из пп. 106-108, где периплазматический якорный белок содержит проходящий через мембрану трансмембранный домен или связанный с мембраной белковый домен, который выставляет антитело в периплазму.
111. Способ по п. 110, где связанный с мембраной белковый домен представляет собой заякоривающий на плазматической мембране гликозилфосфатидилинозитол (GPI)-домен.
112. Способ по любому из пп. 106-108, где периплазматический якорный белок представляет собой белок, который связывается с внутренней поверхностью клеточной стенки, так что антитело выступает в периплазму.
113. Способ по любому из пп. 106-108, где периплазматический якорный белок является достаточно большим, так что периплазматический якорный белок и связанное антитело удерживаются в периплазме.
114. Способ по любому из пп. 106-108, где антитело выбрано из группы, состоящей из моноклонального антитела, химерного антитела, гуманизированного антитела, наноантитела, рекомбинантного фрагмента антитела, Fab-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента, Fv-фрагмента и scFv-фрагмента.
115. Способ по любому из пп. 106-108, где род дрожжевой клетки-хозяина выбран из группы, состоящей из Saccharomyces, Candida, Pichia, Kluyveromyces и Yarrowia.
116. Способ по п. 115, где род дрожжевой клетки-хозяина представляет собой Saccharomyces.
117. Способ по п. 116, где вид Saccharomyces представляет собой Saccharomyces cerevisiae.
| US 2011076752 A1, 31.03.2011 | |||
| US 2016060642 A1, 03.03.2016 | |||
| ISHII J | |||
| et al | |||
| "Cell wall trapping of autocrine peptides for human Gprotein-coupled receptors on the yeast cell surface", PLoS ONE, May 2012, V.7, No.5, e37136, P.1-10 | |||
| WO 2013023251 А, 21.02.2013 | |||
| GRAY S.A | |||
| et al | |||
| "Flow cytometry-based methods for assessing soluble scFv activities and |
