Область техники
Изобретение относится к молекулярной биологии и медицине и рассматривает создание и применение биочипа для определения наличия точечных мутаций в гене галактоза-1-фосфат-уридилтрансферазы, вызывающих поражение печени у новорожденных детей.
Одним из наиболее частых нарушений метаболизма, выявляемых в период новорожденности, является повышение концентрации билирубина в сыворотке крови. Известно, что причинами гипербилирубинемии являются: повышенное образование билирубина при распаде гемоглобина в клетках ретикулоэндотелиальной системы, снижение конъюгации билирубина, нарушение его экскреции или сочетание некоторых из вышеперечисленных факторов. Частота гипербилирубинемии составляет около 55% у доношенных новорожденных. В большинстве случаев гипербилирубинемия новороженных является функциональной (связана с незрелостью ферментативной системы) и проходит в течение первых недель жизни. Тем не менее, повышение уровня билирубина может быть связано с патологией экскреторной функции гепатобилиарной системы и носить злокачественный характер, в частности приводить к необратимому поражению центральной нервной системы.
За последние десятилетия углубилось понимание основ и патофизиологии многих болезней печени, был установлен ряд новых нозологических форм, появились возможности эффективного лечения многих заболеваний, в частности стала применяться диетотерапия (при галактоземии и тирозинемии), стала возможной трансплантация печени. На сегодняшний день число распознанных наследственных болезней печени у новорожденных составляет около 20 различных форм. Гены всех этих заболеваний картированы, и во многих случаях определен спектр основных патогенных мутаций. Для некоторых генов (кодирующих в основном ферменты) также охарактеризованы полиморфные варианты, представленные преимущественно однонуклеотидными заменами, которые могут влиять на остаточную активность белка. Однако дифференциальная диагностика большинства из них крайне затруднительна ввиду схожести клинических проявлений. В нашей стране число пациентов, которым быстро и правильно устанавливается диагноз, сравнительно невелико. Большинство детей погибают в раннем возрасте, не получая адекватного лечения, а отягощенные семьи не имеют информации о наследственном заболевании.
Поскольку в ряде случаев причины злокачественной гипербилирубинемии новорожденных остаются невыясненными, изучение роли генетических факторов в формировании данного патологического состояния - актуальная научная задача, имеющая большое практическое значение.
Уже на первом месяце жизни могут выявляться первые клинические признаки, свидетельствующие о наследственной патологии печени и желчевыводящих протоков и проявляющиеся в виде синдрома холестаза. Одной из причин нарушения экскреторной функции гепатобилиарной системы в неонатальный период (неонатального холестаза) являются наследственные заболевания. По приблизительным подсчетам распространенность врожденных или наследственных заболеваний печени у детей составляет 1 случай на 2500 новорожденных детей. Именно у таких детей развиваются тяжелые формы печеночной патологии. Правильно и быстро установленный диагноз в данном случае является очень важным, т.к. позволяет прогнозировать течение болезни и проводить адекватную терапию. Однако дифференциальная диагностика этой группы заболеваний чрезвычайно сложна.
Одним из факторов, вызывающих тяжелейшие нарушения функционирования печени, является нарушение метаболизма галактозы, вызываемое мутациями в генах, кодирующих белки-ферменты. В результате галактоза, поступающая с пищей в составе молочного сахара - лактозы, подвергается превращению, но реакция превращения не завершается в связи с мутацией гена, кодирующего один из ключевых ферментов. Галактоза и ее производные накапливаются в крови и тканях, оказывая токсическое действие на центральную нервную систему, печень и хрусталик глаза.
На сегодняшний день известно три аутосомно-рецессивных наследственных заболевания, обусловленных недостаточностью ферментов, участвующих в метаболизме галактозы: галактоземия I типа (недостаточность галактоза-1-фосфат уридилтрансферазы), галактоземия II типа (недостаточность галактокиназы) и галактоземия III типа (недостаточность галактоза-4-эпимеразы).
Среди этих заболеваний галактоземия I типа является наиболее тяжелой, требующей неотложной коррекции патологией. Массовый скрининг новорожденных, проводимый во многих странах, направлен на выявление именно этой формы галактоземии.
Следствием недостаточности галактоза-1-фосфат уридилтрансферазы является накопление галактозы, галактоза-1-фосфата и их производных. Эти метаболиты токсически действуют на ткани мозга, печени, почек, кишечника. При отсутствии специфической диетотерапии (назначения безлактозных смесей) заболевание неуклонно прогрессирует и приводит к необратимым изменениям со стороны печени и нервной системы. Своевременная диагностика галактоземии позволяет вовремя начать диетотерапию и избежать серьезных последствий для здоровья ребенка, а также снизить экономические издержки, связанные с медицинской реабилитацией при поздней диагностике.
Основными методами подтверждения диагноза галактоземии I типа являются биохимические тесты, основанные на определении концентрации галактозы, галактоза-1-фосфата в крови и/или на измерении уровня активности фермента галактоза-1-фосфат уридиотрансферазы в эритроцитах.
Трудности интерпретации метаболических изменений связаны с тем, что при поражении печени другой этиологии (как наследственной, так и ненаследственной) возможно повышение уровня галактозы в крови. Описаны ложноположительные тесты при тирозинемии, врожденной атрезии желчевыводящих путей, неонатальном гепатите и многих других заболеваниях (Gitzelmann R, Arbenz UV, Willi UV, et al. Hypergalactosaemia and portosystemic encephalopathy due to persistence of ductus venosus Arantii. // Eur J Pediatr. 1992. V.151. P.564-568).
Определение активности фермента является более специфичным и надежным, однако на результаты теста влияет режим хранения и транспортировки образцов. Также ложноположительные результаты могут наблюдаться при недостаточной активности глюкозо-6-фосфат дегидрогеназы. Кроме того, некоторые полиморфные варианты гена и их сочетания с мутантными аллелями могут искажать результаты биохимической диагностики (Lin HC, Kirby LT, Ng WG, Reichardt JKV. On the nature of the Duarte variant of galactose-1-phosphate uridyl transferase. // Hum Genet. 1994. V.93. P.167-169; Elsas LJ, Dembure PP, Langley S, et al. A common mutation associated with the Duarte galactosemia allele. // Am J Hum Genet. 1994. V.54. P.1030-1036).
Ген галактоза-1-фосфат уридиотрансферазы человека (GALT) располагаеся на 9 хромосоме (локус 9p13) и состоит из 11 экзонов, разделенных 10 интронами. На сегодняшний день в гене описано более 180 различных мутаций, которые преимущественно представлены миссенс-мутациями.
Наиболее распространенными являются мутации Q188R и K285N, в совокупности они составляют в европейских популяциях более 70% от всех мутантных аллелей и обуславливают развитие классической формы галактоземии. Так, в популяции Чехии и Словении частоты мутаций Q188R и K285N составляют 46.0% и 25.7%, а в популяции белой расы Америки - 37% и 4% соответственно (Kozak L, Francova H, Fajkusova L, et al. Mutation analysis of the GALT gene in Czech and Slovak galactosemia populations: identification of six novel mutations, including a stop codon mutation (X380R). // Hum Mutat. 2000. V.15. P.206).
Для некоторых популяций характерны свои особенности профиля мутаций, так среди пациентов афроамериканского происхождения аллель S135L встречается в 62% случаев заболевания, обуславливая относительно легкое течение болезни. Большая делеция в гене GALT, протяженностью 5 т.п.н., является наиболее распространенной в популяции евреев-ашкенази (Elsas LJ, Lai K The molecular biology of galactosemia. // Genet Med. 1998. V.1. P.40-48).
В гене GALT также описано большое число нуклеотидных замен как внутри интронов, так и внутри экзонов, наличие которых отдельно или в различных комбинациях может оказывать влияние на остаточную активность фермента. Одной из наиболее изученных внутригенных замен является мутация N314D в экзоне 10 гена GALT, приводящая к развитию галактоземии Дуарте (Дуарте вариант). Мутация N314D в гомозиготном состоянии, как правило, не приводит к развитию заболевания, однако при этом меняется уровень активности фермента. Комбинации N314D/нормальный аллель, N314D/N314D и N314D/Q188R: обуславливают 75%, 50% и 25% от нормальной активности GALT соответственно. Частота встречаемости аллеля N314D среди здоровых индивидуумов в различных популяциях составляет по литературным данным 6-8% (Lukac-Bajalo J, Marc J, Mlinar B Frequencies of Q188R and N314D mutations and IVS5-24g>A intron variation in the galactose-1-phosphate uridyl transferase gene in the Slovenian population. // Clin Chem Lab Med. 2002. V.40. P.1109-1113). Также описано повышение активности фермента (20-150% от нормы) при наличии генотипа N314D+L218L (вариант Лос-Анжелес или Дуарте 1) (Langley SD et al. Molecular basis for Duarte and Los Angeles variant galactosemia. // Am J Hum Genet. 1997. V.60. P.366-372).
Ранее также были обнаружены следующие мутации в гене GALT, приводящие к развитию галактаземии 1 типа: M142K (Seyrantepe V et al. Identification of mutations in the galactose-1-phosphate uridyltransferase (GALT) gene in 16 Turkish patients with galactosemia, including a novel mutation of F294Y. // Hum Mutat. 1999. V.13. P.339), M142V (Hirokawa H et al. Molecular basis for phenotypic heterogeneity in galactosaemia: prediction of clinical phenotype from genotype in Japanese patients. // Eur J Hum Genet. 1999. V.7. P.757-764), R333W, R333Q (Hirokawa H et al. Molecular basis for phenotypic heterogeneity in galactosaemia: prediction of clinical phenotype from genotype in Japanese patients. // Eur J Hum Genet. 1999.V.7. P.757-64.), R333G (Murphy M et al. Genetic basis of transferase-deficient galactosaemia in Ireland and the population history of the Irish Travellers. // Eur J Hum Genet. 1999. V.7. P.549-554), замена в четвертом интроне нуклеотидов g на c в положении -27 (IVS4-27 g->c) (Kozбk L et al. Mutation analysis of the GALT gene in Czech and Slovak galactosemia populations: identification of six novel mutations, including a stop codon mutation (X380R). // Hum Mutat. 2000. V.15. P.206), замена нуклеотидов а на c в третьем интроне в положении -2 (IVS3-2a->c) (Elsas LJ et al. Galactosemia: a strategy to identify new biochemical phenotypes and molecular genotypes. // Am J Hum Genet. 1995. V.57. P.978-981), возникновение стоп-кодона в 316 положении белка (W316X) (Sommer M et al. Mutations in the galactose-1-phosphate uridyltransferase gene of two families with mild galactosaemia variants. // J Inherit Metab Dis. 1995. V.18. P.567-576).
Для обнаружения точечных мутаций в различных генах применяются следующие методы:
1. экстенция цепи меченым дидезоксирибонуклеозидтрифосфатом (SNP-single nucleotide polymorphism);
2. аллель - специфичная ПЦР (Allele specific PCR);
3. изучение полиморфизма фрагментов после рестрикции (RFLP-restriction fragment length polymorphism);
4. анализ конформационного полиморфизма одно- и двухцепочечной ДНК (SSCP и DSCP- single- (double)-strand conformation polymorphism);
5. гибридизацией на микроматрицах (Hybridization on microaiTay);
6. секвенирование (sequencing);
7. дидезоксифингерпринтинг (ddF-method);
8. ПЦР-гетеродуплексный анализ (PCR-heteroduplex analysis);
9. метод несовершенного дуплекса с РНК (RNA mismatch analysis);
10. метод структурно-специфичного расщепления (Structure-specific endonuclease cleavage);
11. метод зондов (Line Probe assay (LiPA));
12. метод PhaB.
Однако все перечисленные выше методы имеют определенные недостатки, которые на практике могут существенно усложнить массовый скрининг пациентов. Так, методы экстенции цепи и аллель-специфичная ПЦР (1, 2) требуют постановки независимых реакций по числу изучаемых мутаций (т.е. несколько десятков проб для одного пациента в случае гена галактоза-1-фосфат уридилтрансферазы) и, соответственно, большого количества изучаемого образца.
Методы изучения полиморфизма (3, 4), ПЦР-гетеродуплексный анализ (8) и метод несовершенного дуплекса с РНК (9) трудоемки, занимают большое количество времени, дают косвенное заключение о типе мутации и требуют типовых стандартов (на каждую мутацию); кроме того, для метода несовершенного дуплекса предъявляются повышенные требования к отсутствию РНК-азы. Также данный метод трудоемкий и не приемлем для детекции дуплекса G-U.
Гибридизация на микроматрицах (5) требует сложного компьютерного обсчета полученных результатов и дорогостоящих расходных материалов (собственно микроматрица).
Прямое секвенирование (6) требует выделения последовательности гена, отличается высокой себестоимостью и требует секвенирующее устройство.
Методы структурно-специфического расщепления (10) и дидезоксифингерпринтинг (7) требуют предварительной стандартизации (подбора условий), что увеличивает трудоемкость, а также наличия типовых стандартов. Кроме того, дидезоксифингерпринтинг выполняется с радиоактивно-меченым зондом.
Метод зондов (11) отличается высокой себестоимостью и работает в случае небольшого количества мутаций.
Осуществление метода PhaB (12) занимает большое количество времени, поскольку связано с репликацией фагов и последующей регистрацией лизирования на культуре M. smegmatis, а также он очень трудоемок.
Данные недостатки устраняются в настоящем изобретении.
Метод гибридизации на биочипах выгодно отличается от всех вышеперечисленных методик возможностью определения нескольких присутствующих мутаций одновременно, низкой себестоимостью, малым временем получения результата. Также он не требует дорогостоящего оборудования и высококвалифицированного персонала.
В качестве ближайшего аналога заявленных изобретений может быть принят метод детекции точечных мутаций в гене GALT при помощи гибридизации различных последовательностей ДНК из гена GALT, описанный в патенте US 6207387, выданном патентным ведомством США. Однако представленное изобретение обладает рядом преимуществ по сравнению с методом, изложенным в US 6207387.
1. Заявленный биочип позволяет детектировать неизвестные ранее мутации в гене GALT, вызывающие развитие галактоземии I типа (E352Q, L358P).
2. При помощи заявленного биочипа можно детектировать большее число различных точечных мутаций в гене GALT, что позволит с большей вероятностью исключить ложноположительные результаты при постановке диагноза пациенту.
3. При реализации описанного в патенте US 6207387 способа детекции точечных мутаций в гене GALT был использован метод гибридизации на мембране, который более ресурсоемок, чем метод гибридизации на биочипах. Следовательно, заявленный биочип будет более приемлем для использования в рутинной диагностике галактоземии I типа в клинических лабораториях.
Таким образом, заявленное нами изобретение позволит более просто и точно определять наличие мутаций в гене GALT, что позволит в значительной мере улучшить диагностику галактоземии I типа.
Раскрытие изобретения.
Заявленные изобретения относятся к биочипу, который позволяет детектировать наличие точечных мутаций у пациента-человека; применению указанного биочипа для постановки диагноза пациенту-человеку, а также способу получения и использования указанного биочипа.
Краткое описание чертежей
Фигура 1. Нанесение олигонуклетидов, комплементарных различным мутациям в гене GALT, на биочип 1.
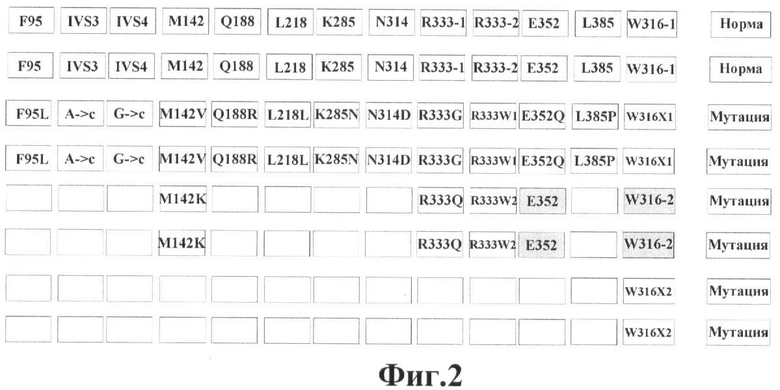
Фигура 2. Нанесение олигонуклетидов, комплементарных различным мутациям в гене GALT, на биочип 2.
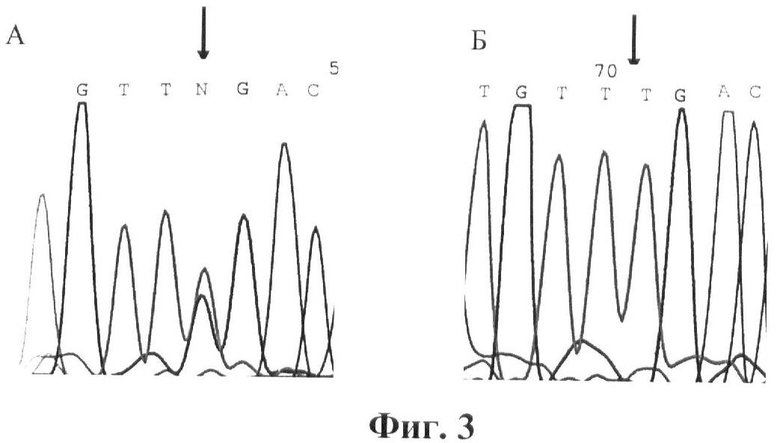
Фигура 3. Детекция мутации F95L (TTT->TTG). Прямое ДНК-секвенирование. Стрелками указана локализация мутации. А.Фрагмент экзона 3 гена GALT. Мутация F95L в гетерозиготном состоянии. Б. Фрагмент экзона 3 гена GALT. Норма.
Фигура 4. Детекция мутации IVS3-2a->c. ПДРФ анализ. При возникновении мутации IVS3-2a->c появляется сайт рестрикции для фермента рестрикции Msp I (C^CGG). Дорожки: 1 - маркер молекулярного веса, 2 - фрагмент экзонов 3-5 гена GALT до рестрикции, 3, 4 - фрагменты экзонов 3-5 гена GALT после обработки ферментом рестрикции Msp I (C^CGG): 3-4 - мутация IVS3-2a->c в гетерозиготном состоянии; 5 - норма.
Фигура 5. Детекция полиморфизма IVS4-27 g->c. ПДРФ анализ. При замене g->c исчезает сайт рестрикции для фермента рестрикции Msp I (C^CGG). Дорожки: 1, 2, 3 - фрагменты интрона 4 гена GALT после обработки ферментом рестрикции Msp I (C^CGG); 1 - гомозигота по полиморфизму IVS4-27 g; 2 - гомозигота по полиморфизму IVS4-27 c; 3 - гетерозигота по полиморфизму IVS4-27 g->c.
Фигура 6. Детекция мутации M142K (ATG->AAG). Прямое ДНК-секвенирование. Стрелками указана локализация мутации. А. Фрагмент экзона 5 гена GALT. Мутация M142K в гомозиготном состоянии. Б. Фрагмент экзона 5 гена GALT. Норма.
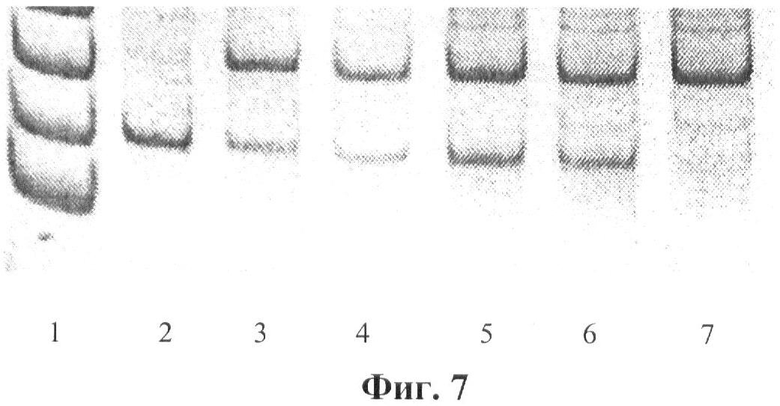
Фигура 7. Детекция мутации Q188R (CAG->CCG). ПДРФ анализ. При мутации Q188R появляется сайт рестрикции для фермента рестрикции Msp I (C^CGG) и исчезает сайт рестрикции для фермента рестрикции Bst2U I (CCWGG). Дорожки: 1 - маркер молекулярного веса, 2-7 - фрагменты экзона 6 гена GALT после обработки ферментом рестрикции Msp I (C^CGG); 2 - гомозигота по мутации Q188R; 3-6 - гетерозиготы по мутации Q188R; 7 - норма.
Фигура 8. Детекция полиморфизма L218L (CTA->TTA). Прямое ДНК-секвенирование. Обратная цепь. Стрелками указана локализация полиморфизма. А. Фрагмент экзона 7 гена GALT. Полиморфизм L218L в гетерозиготном состоянии. Б. Фрагмент экзона 7 гена GALT. Норма.
Фигура 9. Детекция мутации K285N (AAG->AAT). ПДРФ анализ. При мутации K285N появляется сайт рестрикции для фермента рестрикции Sse9 I (^AATT). Дорожки: 1 - маркер молекулярного веса, 2, 3 - фрагменты экзона 9 гена GALT после обработки ферментом рестрикции Sse9 I (^AATT): 2 - норма; 3 - мутация K285N в гетерозиготном состоянии.
Фигура 10. Детекция полиморфизма N314D (AAC->GAC). ПДРФ анализ. При замене aac->gac появляется сайт рестрикции для фермента рестрикции Bmu I (ACTGGG(N)5^). Дорожки: 1 - фрагмент 10 экзона гена GALT до рестрикции, 2, 3 - фрагменты экзона 10 гена GALT после обработки ферментом рестрикции Bmu I (ACTGGG(N)5^); 2 - полиморфизм в гетерозиготном состоянии, 3 - норма.
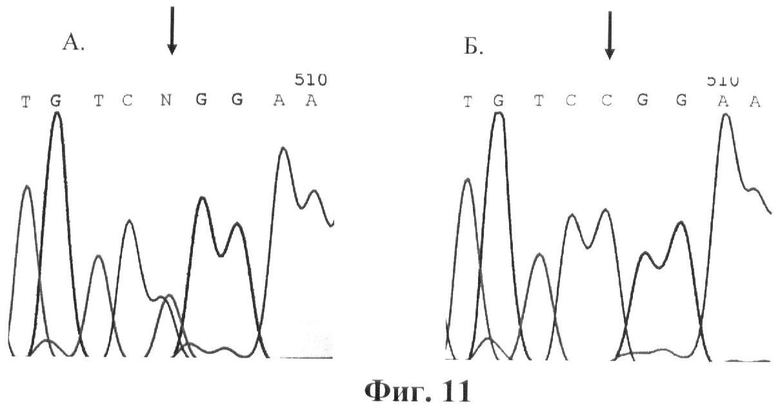
Фигура 11. Детекция мутации R333W (CGG->TGG). Прямое ДНК-секвенирование. Стрелками указана локализация мутации. А. Фрагмент экзона 10 гена GALT. Мутация R333W в гетерозиготном состоянии. Б. Фрагмент экзона 10 гена GALT. Норма.
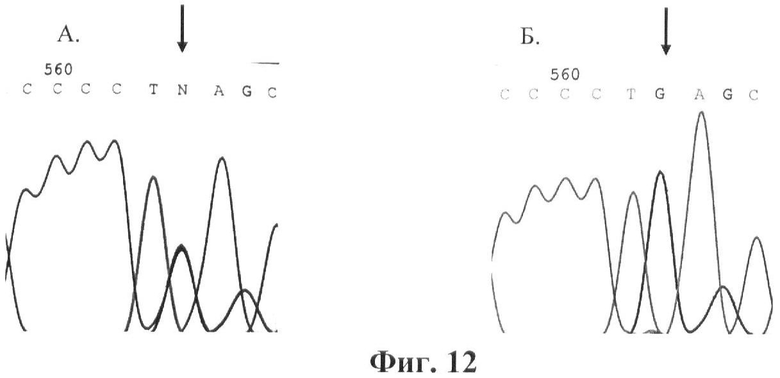
Фигура 12. Детекция мутации E352Q (GAG->CAG). Прямое ДНК-секвенирование. Стрелками указана локализация мутации. А. Фрагмент экзона 10 гена GALT. Мутация E352Q в гетерозиготном состоянии. Б. Фрагмент экзона 10 гена GALT. Норма.
Фигура 13. Детекция мутации L358P (CTA->CCA). ПДРФ анализ. При мутации L358P появляется сайт рестрикции для фермента рестрикции BssT1 I (C^CWWGG). Дорожки: 1, 2 - фрагменты экзона 11 гена GALT после обработки ферментом рестрикции BssT1 I (C^CWWGG); 1 - гетерозигота по мутации L358P; 2 - норма.
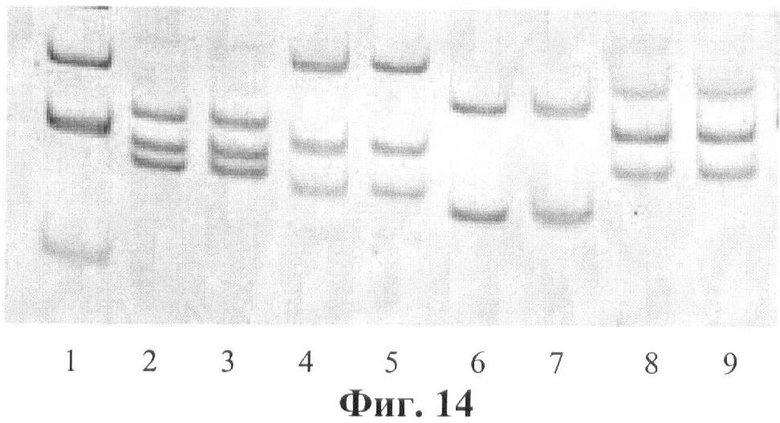
Фигура 14. Электрофореграмма мультиплексных ПЦР для второго раунда.
Дорожки: 1 - маркер молекулярного веса,
2-3 - второй раунд ПЦР: фрагмент экзона 3 гена GALT
(мутация F95L) - 119 пн+фрагмент экзона 10 гена GALT
(мутация R333W) - 104 пн+фрагмент экзона 6 гена GALT
(мутация Q188R) - 96 пн;
4-5 - второй раунд ПЦР: фрагмент экзона 10 гена GALT
(полиморфизм N314D) - 151 пн+фрагмент интрона 3 гена
GALT (мутация IVS3-2a->c) - 107 пн+фрагмент экзона 7 гена
GALT (полиморфизм L218L) - 89 пн;
6-7 - второй раунд ПЦР: фрагмент интрона 4 гена GALT
(полиморфизм IVS4-27 g->c) - 130 пн+фрагмент экзона 10
гена GALT (мутация E352Q) - 84 пн;
8-9 - второй раунд ПЦР: фрагмент экзона 11 гена GALT
(мутация L358P) - 139 пн+фрагмент экзона 9 гена GALT
(мутация K285N) - 112 пн+фрагмент экзона 5 гена GALT
(мутация M142K) - 100 пн.
Фигура 15. Гибридизационная картина образца ДНК, полученного из человека, не несущего мутации в гене GALT с биочипом-1. Расположение точек соответствует фиг.1.
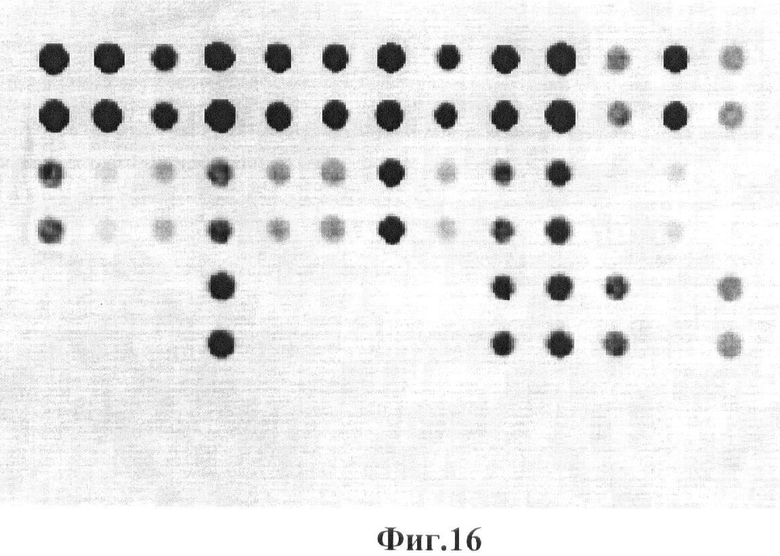
Фигура 16. Гибридизационная картина образца ДНК, полученного из человека не несущего мутации в гене GALT с биочипом-2. Расположение точек соответствует фиг.2.
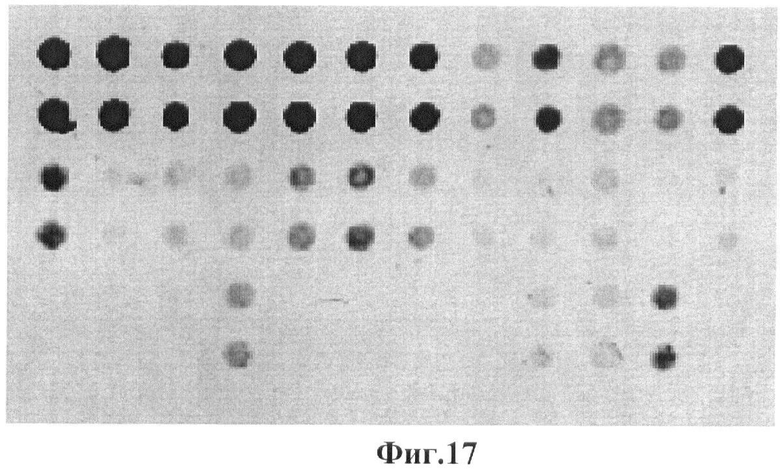
Фигура 17. Гибридизационная картина образца ДНК, полученного из человека не несущего мутации в гене GALT с биочипом-2. Расположение точек соответствует фиг.2.
Фигура 18. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию IVS3-2a->c в гене GALT с биочипом-1. Расположение точек соответствует фиг.1.
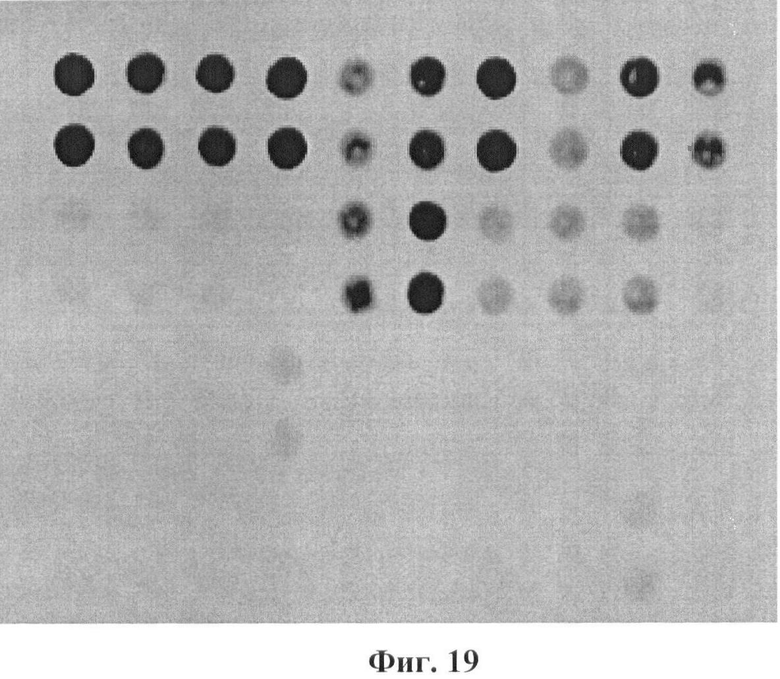
Фигура 19. Гибридизационная картина образца ДНК, полученного из человека несущего мутации L218L/Q188R/N314D в гене GALT с биочипом-1. Расположение точек соответствует фиг.1.
Фигура 20. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию Q188R в гомозиготном состоянии в гене GALT с биочипом-1. Расположение точек соответствует фиг.1.
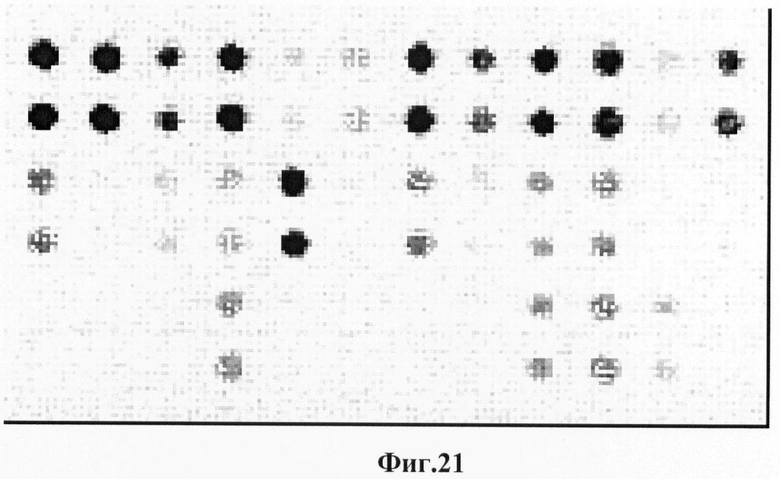
Фигура 21. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию Q188R в гомозиготном состоянии в гене GALT с биочипом-2. Расположение точек соответствует фиг.2.
Фигура 22. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию Q188R в гетерозиготном состоянии в гене GALT с биочипом-1. Расположение точек соответствует фиг.1.
Фигура 23. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию K285N в гене GALT в гетерозиготном состоянии с биочипом-1. Расположение точек соответствует фиг.1.
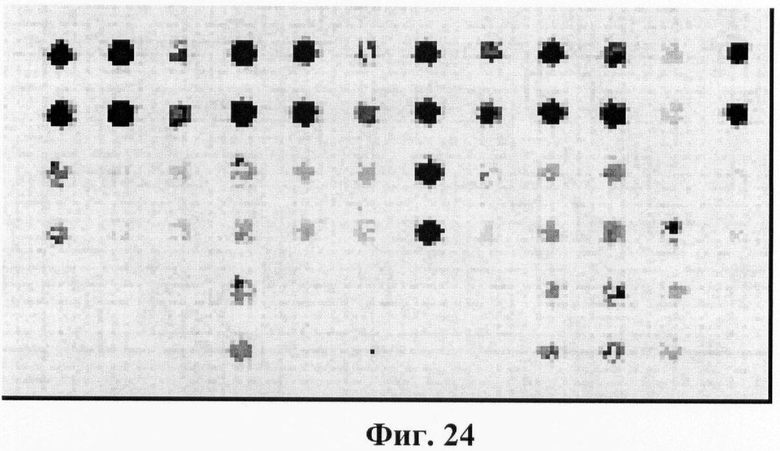
Фигура 24. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию K285N в гене GALT в гетерозиготном состоянии с биочипом-2. Расположение точек соответствует фиг.2.
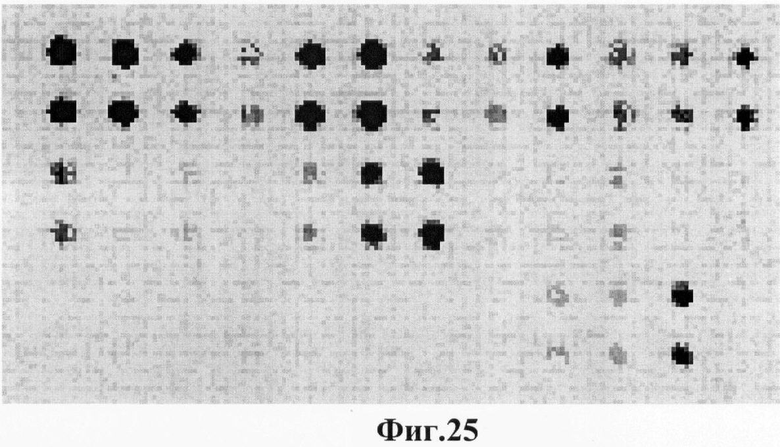
Фигура 25. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию K285N в гене GALT в гомозиготном состоянии с биочипом-2. Расположение точек соответствует фиг.2.
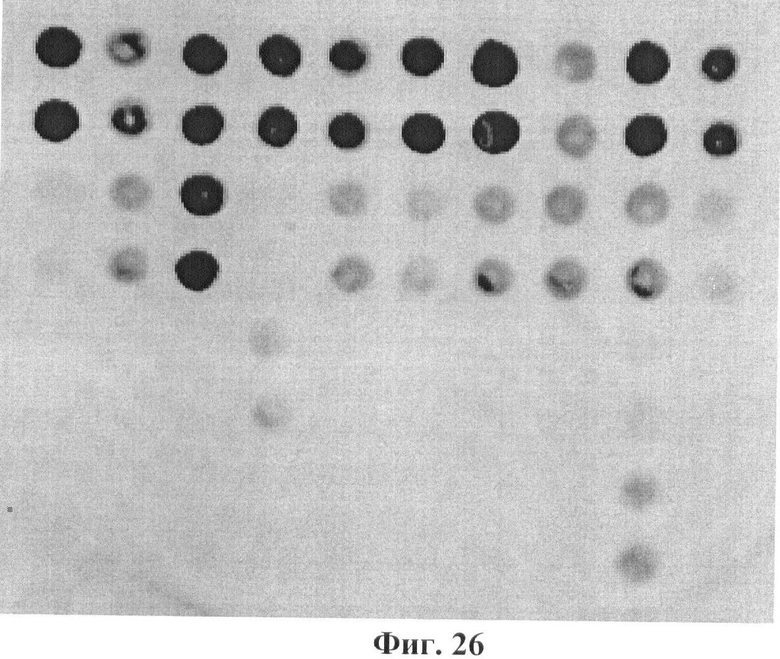
Фигура 26. Гибридизационная картина образца ДНК, полученного из человека несущего мутации IVS4-27 g->c и N314D в гене GALT в гетерозиготном состоянии с биочипом-1. Расположение точек соответствует фиг.1.
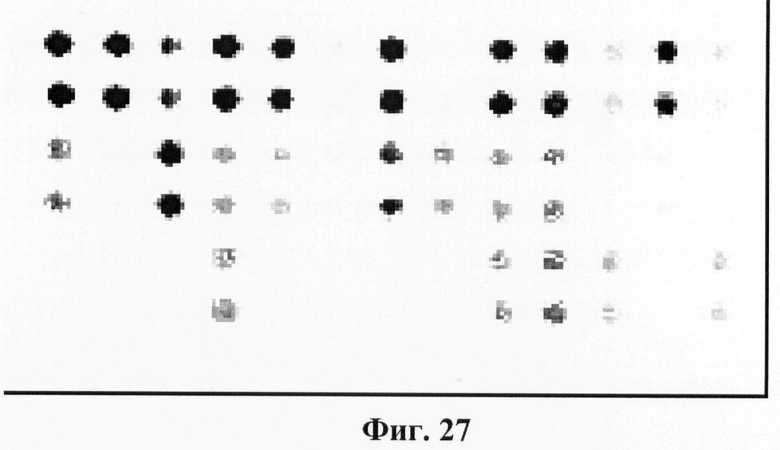
Фигура 27. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию N314D и полиморфизм IVS4-27 g->c в гомозиготном состоянии в гене GALT с биочипом-2. Расположение точек соответствует фиг.2.
Фигура 28. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию M142K в гене GALT в гетерозиготном состоянии с биочипом-1. Расположение точек соответствует фиг.1.
Фигура 29. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию M142K в гене GALT в гомозиготном состоянии с биочипом-2. Расположение точек соответствует фиг.2.
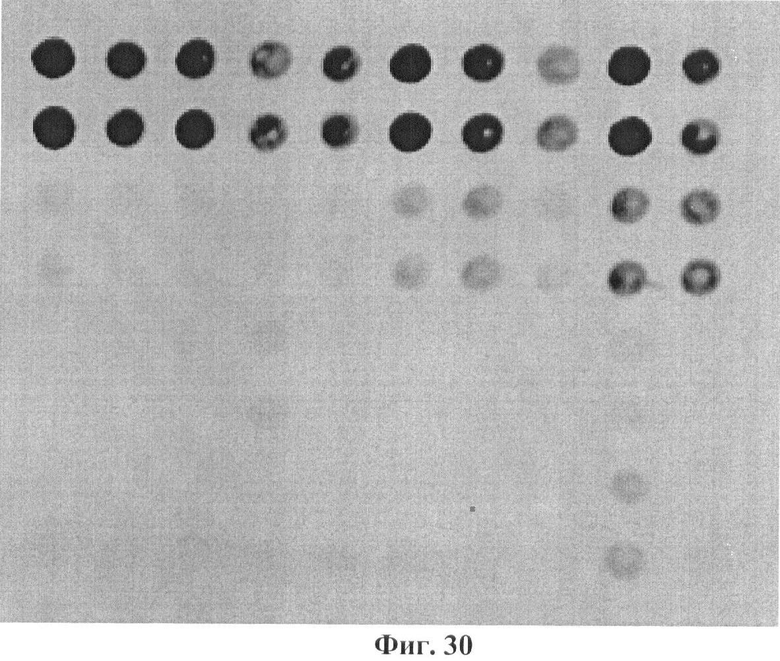
Фигура 30. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию L358P в гене GALT в гетерозиготном состоянии с биочипом-1. Расположение точек соответствует фиг.1.
Фигура 31. Гибридизационная картина образца ДНК, несущего мутацию F95L и нормальную последовательность гена GALT с биочипом-2. Расположение точек соответствует расположению олигонуктеотидов столбца F95 на фиг.2.
Фигура 32. Гибридизационная картина образца ДНК, несущего мутацию R333W/R333Q/R333G и нормальную последовательность гена GALT с биочипом-2. Расположение точек соответствует расположению олигонуктеотидов столбцов R333-1 и R333-2 на фиг.2.
Фигура 33. Гибридизационная картина образца ДНК, несущего мутацию E352Q и нормальную последовательность гена GALT с биочипом-2. Расположение точек соответствует расположению олигонуктеотидов столбца E352 на фиг.2.
Фигура 34. Гибридизационная картина образца ДНК, полученного из человека несущего мутацию несущего мутацию Q188R в гене GALT в гомозиготном состоянии с биочипом-2. Расположение точек соответствует фиг.2.
Осуществление изобретения
Осуществление изобретения проиллюстрировано следующими примерами.
Пример 1. Установление мутаций, использованных для создания биочипа.
С целью разработки биочипа были проанализированы частоты мутаций в гене GALT у пациентов с галактоземией 1 типа из Российской Федерации. Частоты мутаций Q188R и K285N составляют на данный момент исследования 60% и 16% соответственно. Среди редких мутаций, обнаруженных у пациентов, четыре уже были описаны ранее (IVS3-2a->g (обнаружена у двух пациентов), M142K (обнаружена у двух пациентов), R333W (обнаружена у одного пациента), W316X (обнаружена у одного пациента)) и две новые: E352Q (обнаружена у двух пациентов), L358P (обнаружена у двух пациентов). В результате массового скрининга было выявлено 25 носителей мутантных аллелей (Q188R, K285N, или редких мутаций); носителей только полиморфизма N314D (вариант Дуарте) - 5 человек, носителей полиморфизма N314D в сочетании с какой-либо мутацией в транс-положении - 40 человек. В случаях сочетанного носительства мутации и полиморфизма N314D был также обнаружен полиморфизм IVS-27g->c в гетерозиготном состоянии, в цис-положении с N314D. При этом активность фермента у таких пациентов была снижена по сравнению с нормой.
При анализе литературных данных также были выбраны мутации и полиморфизмы, которые с наибольшей частотой встречались в других популяциях.
Учитывая информацию, изложенную выше, для создания биочипа были выбраны следующие мутации и полиморфизмы в гене GALT: F95L, IVS3-2a->c, IVS4-27 g->c, M142K, M142V, Q188R, L218L, K285N, N314D, R333W, R333G, R333Q, E352Q, L358P, W316X.
Пример 2. Дизайн и оптимизация последовательностей олигонуклеотидов для иммобилизации на микрочипе.
Дизайн олигонуклеотидных последовательностей для иммобилизации на микрочипе проводили с учетом анализа нуклеотидной последовательности гена галактоза-1-фосфат уридилтрансферазы в областях пяти мутаций и пяти значимых полиморфизмов. Для этой цели использовали программу Oligo 6 (США).
Пример 3. Конструирование микрочипа для детекции полиморфизма в гене галактоза-1-фосфат уридилтрансферазы.
При выборе полиморфных локусов, анализ которых будет проводиться при помощи биочипа по изобретению, руководствовались следующими подходами: 1) в гене выбирались функционально значимые аллели, встречающиеся с наибольшей частотой; 2) включали только те нуклеотидные замены, полиморфизм которых уже изучен в ряде популяций России.
Последовательности нуклеотидов, использованных для создания биочипа по изобретению, приведены в таблицах 1 и 2.
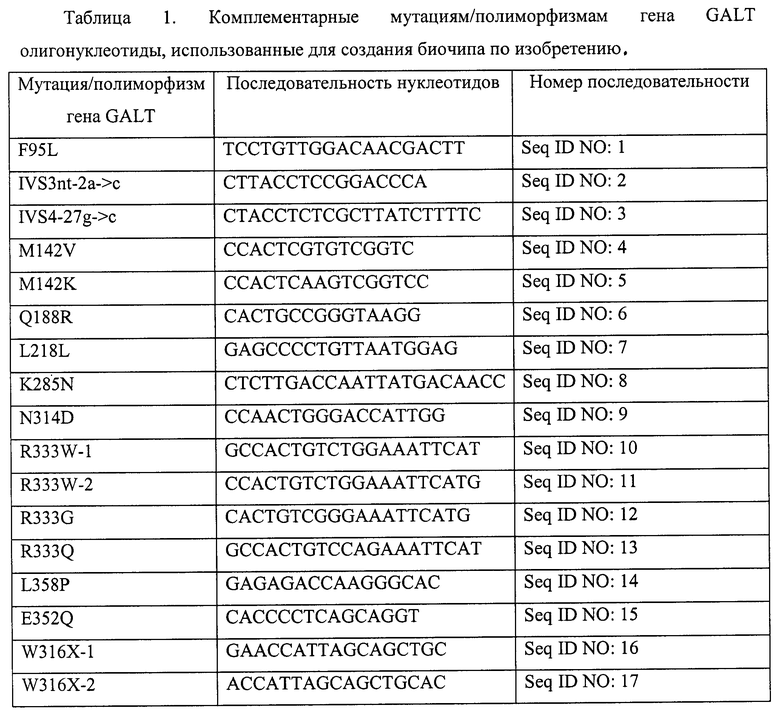

Олигонуклеотиды, приведенные в таблице 1, 2, синтезированы с использованием стандартной фосфоамитидной процедуры на автоматическом синтезаторе 394 DNA/RNA Synthesizer ("Applied Biosystems", США) На 3'-конце олигонуклеотидов находится спейсер со свободной аминогруппой, который вводили при синтезе с помощью 3'-Amino-Modifier C7 CPG 500 ("Glen Reseach", США) с целью проведения фотоиндуцируемой совместной полимеризации олигонуклеотидов и компонентов полиакриламидного геля.
С целью повышения надежности определения сигнала каждую олигонуклеотидную пробу продублировали. Для подбора последовательности олигонуклетидов, обеспечивающих максимальную специфичность, в биочипе использовались несколько типов олигонуклеотидов, комплементарных одной и той же мутации/полиморфизму или немутантной последовательности.
Факт наличия немутантных и мутантных аллелей в геноме пациента установлен при анализе интенсивности флюоресценции соответствующих ячеек на чипе. Нанесение олигонуклетидов, комплементарных нормальной последовательности и различным мутациям/полиморфизмам гена GALT, на биочип показано на фигурах 1 и 2 (обозначение олигонуклеотидов такое же, как и в таблице 1 и 2).
Таким образом, сконструированный биочип позволяет определять наличие мутаций в гене галактоза-1-фосфат уридилтрансферазы, приводящие к возникновению заболевания.
Пример 4. Подготовка образцов ДНК от здоровых доноров и больных галактоземией 1 типа для тестирования биочипа.
В результате работы были отобраны образцы ДНК больных галактоземией детей для каждой из десяти исследуемых мутаций. Эти образцы ДНК подготовлены для тестирования полученного ДНК-чипа.
Все выбранные нами для создания биочипа мутации были подтверждены методом прямого нерадиоактивного секвенирования и/или анализом полиморфизма длины рестрикционных фрагментов. В качестве примеров на фигурах 3-13 приведены результаты части экспериментов. Как видно на приведенных фигурах мутации и полиморфизмы хорошо детектируются с помощью данных методов. Таким образом, в результате данной работы были отобраны образцы ДНК, которые можно применить для тестирования полученного биочипа.
Пример 5. Гибридизация биочипа с ДНК-зондами с известной структурой и с ДНК-зондами, полученными от пациентов.
В результате исследования подобраны олигонуклеотидные праймеры и условия проведения реакции для проведения второго раунда ПЦР на последовательности гена GALT, полученной в первом раунде ПЦР, для дальнейшей детекции патогенно-значимых нуклеотидных замен гена GALT на микрочипе. Для проведения второго раунда мультиплексной ПЦР были подобраны следующие олигонуклеотидные праймеры (Таблица 3).

Так как экзоны гена GALT располагаются плотным кластером, а размер интронов гена GALT не превышает 740 bp, то чтобы избежать перекрестной ПЦР, второй раунд проводили следующим образом: аликвота от первого раунда ПЦР объемом 5 мкл с последующей мультиплексной ПЦР:
1. Фагмент экзона 3 гена GALT (мутация F95L)+фрагмент экзона 10 гена GALT (мутация R333W)+фрагмент экзона 6 гена GALT (мутация Q188R);
2. Фагмент экзона 10 гена GALT (полиморфизм N314D, W316X)+фрагмент интрона 3 гена GALT (мутация IVS3-2a->c)+фрагмент экзона 7 гена GALT (полиморфизм L218L);
3. Фагмент интрона 4 гена GALT (полиморфизм IVS4-27 g->c)+фрагмент экзона 10 гена GALT (мутация E352Q);
4. Фагмент экзона 11 гена GALT (мутация L358P)+фрагмент экзона 9 гена GALT (мутация K285N)+фрагмент экзона 5 гена GALT (мутация M142K).
Реакционная смесь (25 мкл) на втором этапе ПЦР содержала 0,5 пмоль каждого праймера, 2,5 мкл буфера для ПЦР с 2,5 мМ MgCl2 («Силекс», Россия), 6,25 нМ каждого из dNTP («Силекс», Россия), 0,2 нМ флюоресцентно меченого dUTP-Cy5 и 1 ед.акт. Taq-полимеразы («Силекс», Россия). Для амплификации использовали программируемый ДНК-амплификатор фирмы «ДНК-технология» (Россия). После предварительной денатурации ДНК (3 минуты при 94°С) проводили амплификацию в режиме: 33 цикла амплификации: 94°С 30 сек, 59°С 30 сек, 72°С 1 мин. Заключительный синтез 72°С - 3 минуты.
Электрофореграмма мультиплексных ПЦР представлена на фиг.14.
Гибридизацию на микрочипе проводили с использованием флюоресцентно меченых образцов, полученных на второй стадии реакции ПЦР, в 32 мкл смеси следующего состава: 8 мкл формамида ("Serva", США), 8 мкл 20XSSPE ("Promega", США), 4 мкл амплификата и 12 мкл Н2О. Гибридизационную смесь денатурировали при 95°С (5 мин), охлаждали на льду (3 мин), наносили на биочип и оставляли на ночь при температуре 37°С.Далее чип отмывали в 1XSSPE в течение 5 мин при 20°С и высушивали.
Флюоресцентный сигнал от ячеек регистрировали с помощью широкопольного люминисцентного микроскопа, снабженного камерой ПЗС и программным обеспечением Imageware («Биочип-ИМБ», Россия). Использовали нормированные сигналы флюоресценции Jm=(Im-I0)/(Bm-I0), где Im - интенсивность сигнала на единицу площади внутренней части составляющей ячейки, Bm - фоновый сигнал, отражающий распределение освещенности микрочипа, I0- темновой ток ПЗС матрицы, а m - номер ячейки чипа. При анализе интенсивности флюоресценции использовали среднее значение между двумя дублирующими ячейками. Сигналы Jm сортировали по возрастанию. Ячейкой сравнения считали ячейку, после которой наблюдали резкое увеличение нормированного сигнала. Считали, что ячейка имеет положительный сигнал, если ее сигнал превышал сигнал ячейки сравнения. Затем сравнивали между собой ячейки, содержащие нуклеотидную последовательность дикого типа и соответствующую мутантную последовательность, выявляя доминирующий сигнал, который превышал пороговое значение. Пороговое значение сигнала определяли статистически, обработав более 20 гибридизационных картин. Если в паре положительных сигналов «проба дикого типа - мутантная проба» ни один из сигналов не доминировал, значимыми считали сигналы с обеих ячеек, что соответствовало гетерозиготному состоянию анализируемого локуса.
Пример 6. Анализ данных, полученных при гибридизации на биочипе по изобретению.
6.1. Результат трех различных гибридизаций продуктов ПЦР-реакции, полученных с использованием геномной ДНК доноров, не болеющих галактаземией 1 типа, с биочипом по изобретению приведен на фигурах 15-17 и таблицах 4-6.
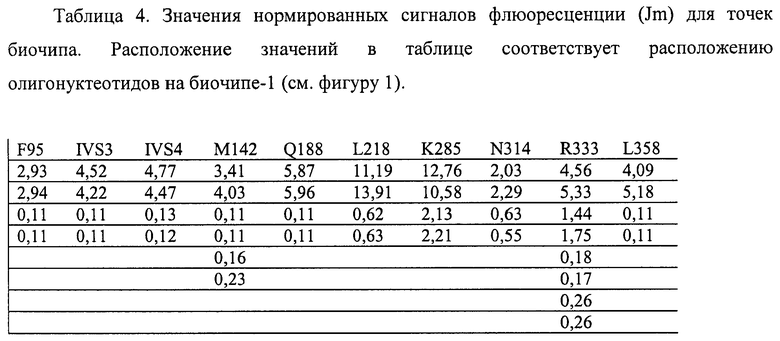


Согласно сведениям, приведенным в таблицах 4-6 и на фигурах 15-17, уровень нормированных сигналов флюоресценции (Jm) в точках, соответствующих нормальной последовательности гена GALT, в несколько раз выше, чем в точках соответствующих мутированным последовательностям. Таким образом, заявленный биочип не дает ложноположительных результатов в случае пациента, не несущего указанных мутаций в последовательности гена GALT.
6.2. Результат трех различных гибридизаций продуктов ПЦР-реакции, полученных с использованием геномной ДНК доноров, несущих полиморфизм IVS3-2a->c, с биочипом по изобретению приведен на фигуре 18 и таблице 7.

Согласно сведениям, приведенным в таблице 7 и на фигуре 18, уровень сигналов флюоресценции в точках, соответствующих мутации IVS3-2a->c в гене GALT, такой же, как и в точках, соответствующих последовательности, не содержащей данную замену в интроне 3. Данный факт говорит о том, что мутация находится в гетерозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Таким образом, заявленный биочип позволяет установить наличие мутации IVS3-2a->c в гене GALT с высокой специфичностью (даже в гетерозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.3. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутации L218L/Q188R/N314D в гетерозиготном состоянии, был получен результат, приведенный на фигуре 19 и таблице 8.

Согласно сведениям, приведенным в таблице 8 и на фигуре 19, уровень сигналов флюоресценции в точках, соответствующих мутации L218L/Q188R/N314D в гене GALT, такой же, как и в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутации находятся в гетерозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Таким образом, заявленный биочип позволяет установить наличие мутаций L218L/Q188R/N314D в гене GALT с высокой специфичностью (даже в гетерозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.4. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутацию Q188R в гомозиготном состоянии, был получен результат, приведенный на фигурах 20 и 21 и таблицах 9 и 10.
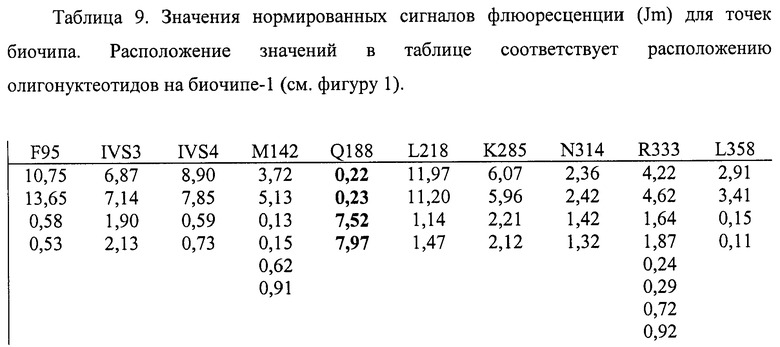
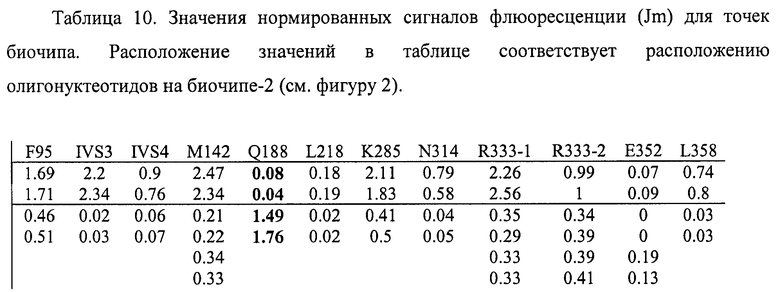
Согласно сведениям, приведенным в таблицах 9 и 10 и на фигурах 20 и 21, уровень сигналов флюоресценции в точках, соответствующих мутации Q188R в гене GALT, в несколько десятков раз сильнее, чем в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гомозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Данный факт показывает, что заявленный биочип позволяет установить наличие мутации Q188R в гене GALT с высокой специфичностью (в гомозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.5. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутацию Q188R в гетерозиготном состоянии, был получен результат, приведенный на фигуре 22 и таблице 11.
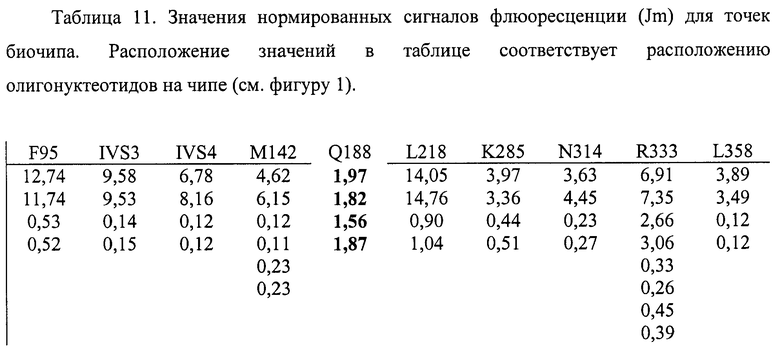
Согласно сведениям, приведенным в таблице 11 и на фигуре 22, уровень сигналов флюоресценции в точках, соответствующих мутации Q188R в гене GALT, такой же, как и в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гетерозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Таким образом, биочип позволяет установить наличие мутации Q188R в гене GALT с высокой специфичностью (в гетерозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.6. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутации K285N в гетерозиготном состоянии, был получен результат, приведенный на фигурах 23 и 24 и таблицах 12 и 13.

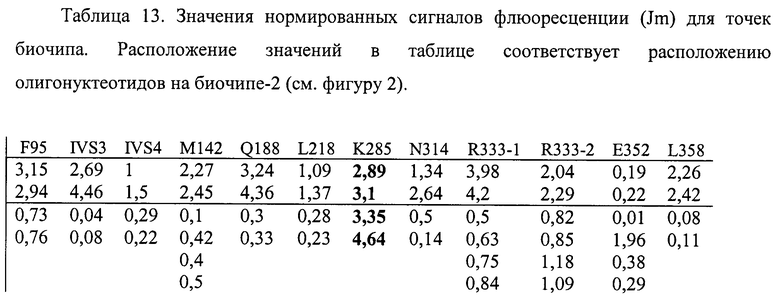
Согласно сведениям, приведенным на фигурах 23 и 24 и таблицах 12 и 13, уровень сигналов флюоресценции в точках, соответствующих мутации K285N в гене GALT, такой же, как и в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гетерозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Данный факт показывает, что заявленный биочип позволяет установить наличие мутаций K285N в гене GALT с высокой специфичностью (даже в гетерозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.7. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутацию K285N в гомозиготном состоянии, был получен результат, приведенный на фигуре 25 и таблице 14.

Согласно сведениям, приведенным в таблице 14 и на фигуре 25, уровень сигналов флюоресценции в точках, соответствующих мутации K285N в гене GALT, в несколько раз сильнее, чем в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гомозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Данный факт показывает, что заявленный биочип позволяет установить наличие мутации K285N в гене GALT с высокой специфичностью (в гомозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.8. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутации IVS4-27 g->c и N314D в гетерозиготном состоянии, был получен результат, приведенный на фигуре 26 и таблице 15.

Согласно сведениям, приведенным в таблице 15 и на фигуре 26, уровень сигналов флюоресценции в точках, соответствующих мутациям IVS4-27 g->c и N314D в гене GALT, такой же, как и в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гетерозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Таким образом, биочип позволяет установить наличие мутаций IVS4-27 g->c и N314D в гене GALT с высокой специфичностью (в гетерозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.9. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего полиморфизм IVS4-27 g->c в и мутацию N314D в гомозиготном состоянии, был получен результат, приведенный на фигуре 27 и таблице 16.
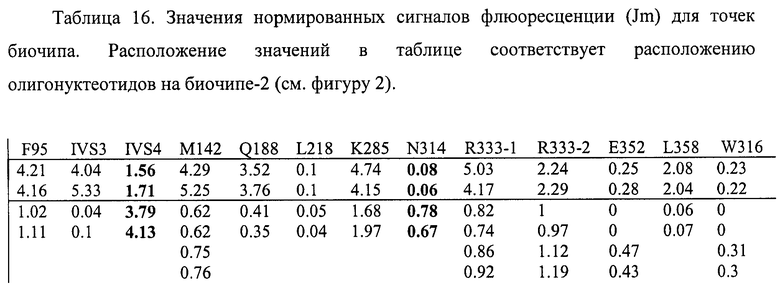
Согласно сведениям, приведенным в таблице 16 и на фигуре 27, уровень сигналов флюоресценции в точках, соответствующих мутации N314D и полиморфизму IVS4-27 g->c в гене GALT в несколько раз сильнее, чем в точках соответствуюших нормальной последовательности гена GALT. Данный факт говорит о том, что полиморфизм IVS4-27 g->c и мутация N314D находятся в гомозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Таким образом, биочип позволяет установить наличие полиморфизма IVS4-27 g->c и мутации N314D в гене GALT с высокой специфичностью. При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.10. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутации M142K в гетерозиготном состоянии, был получен результат, приведенный на фигуре 28 и таблице 17.
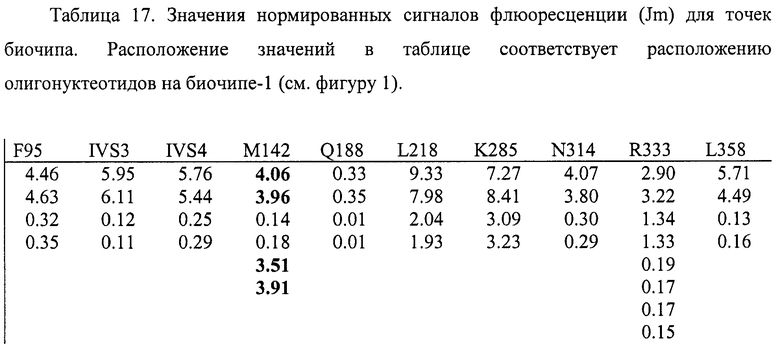
Согласно сведениям, приведенным в таблице 17 и на фигуре 28, уровень сигналов флюоресценции в точках, соответствующих мутации M142K в гене GALT, такой же, как и в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гетерозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Таким образом, биочип позволяет установить наличие мутаций M142K в гене GALT с высокой специфичностью (в гетерозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.11. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутацию M142K в гомозиготном состоянии, был получен результат, приведенный на фигуре 29 и таблице 18.
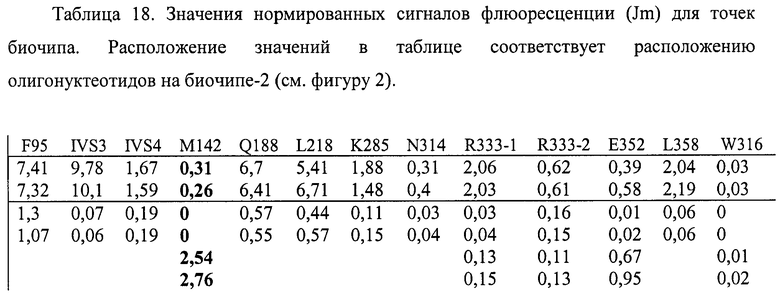
Согласно сведениям, приведенным в таблице 18 и на фигуре 29, уровень сигналов флюоресценции в точках, соответствующих мутации M142K в гене GALT, в несколько десятков раз сильнее, чем в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гомозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Данный факт показывает, что заявленный биочип позволяет установить наличие мутации M142K в гене GALT с высокой специфичностью (в гомозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.12. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутацию L358P в гетерозиготном состоянии, был получен результат, приведенный на фигуре 30 и таблице 19.

Согласно сведениям, приведенным в таблице 19 и на фигуре 30, уровень сигналов флюоресценции в точках, соответствующих мутации L358P в гене GALT, такой же, как и в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гетерозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Таким образом, биочип позволяет установить наличие мутаций L358P в гене GALT с высокой специфичностью (даже в гетерозиготном состоянии). При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
Поскольку в распоряжении авторов не было пациентов, несущищих мутации F95, R333, E352, то зонды для детекции специфичности биочипа по изобретению были синтезированны искусственно (соответствовали мутированным последовательностям гена GALT).
6.13. В результате гибридизации искусственных зондов, содержащих мутацию F95L и нормальную последовательность гена, c биочипом-2 был получен результат, представленный на фигуре 31 и таблице 20.
Согласно сведениям, приведенным в таблице 20 и на фигуре 31, уровень сигналов флюоресценции в точках, соответствующих мутации F95L в гене GALT, такой же, как и в точках, соответствующих последовательностям, не содержащим данные замены. Со всеми остальными точками биочипа-2 искусственный зонд не взаимодействовал. Таким образом, приведенные данные показывают, что заявленный биочип позволяет установить наличие мутации F95L в гене GALT с высокой специфичностью. При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.14. В результате гибридизации искусственных зондов, содержащих мутации R333W/R333Q/R333G и нормальную последовательность гена GALT, c биочипом-2 был получен результат, представленный на фигуре 32 и таблице 21.
Согласно сведениям, приведенным в таблице 21 и на фигуре 32, уровень сигналов флюоресценции в точках, соответствующих мутациям R333W/R333Q/R333G в гене GALT, такой же, как и в точках, соответствующих последовательностям, не содержащим данные замены. Со всеми остальными точками биочипа-2 искусственный зонд не взаимодействовал. Также наиболее предпочтительными для создания биочипа по изобретению были выбраны последовательноть Seq ID NO: 26, соответствующая немутированному гену, и последовательность Seq ID NO: 10, соответствующая мутации R333W. Таким образом, приведенные данные показывают, что заявленный биочип позволяет установить наличие мутации R333W в гене GALT с высокой специфичностью. При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.15. В результате гибридизации искусственных зондов, содержащих мутацию E352Q c биочипом-2 был получен результат, представленный на фигуре 33 и таблице 22.
Согласно сведениям, приведенным в таблице 23 и на фигуре 33, уровень сигналов флюоресценции в точках, соответствующих мутации E352Q в гене GALT, в несколько десятков раз сильнее, чем в точках, соответствующих последовательностям, не содержащих мутации. Со всеми остальными точками биочипа-2 искусственный зонд не взаимодействовал. Таким образом, приведенные данные показывают, что заявленный биочип позволяет установить наличие мутации E352Q в гене GALT с высокой специфичностью. При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
6.16. В результате гибридизации продуктов ПЦР-реакции, полученных с использованием геномной ДНК пациента, несущего мутацию Q188R в гомозиготном состоянии, был получен результат, приведенный на фигуре 34 и таблице 23.

Согласно сведениям, приведенным в таблице 23 и на фигуре 33, уровень сигналов флюоресценции в точках, соответствующих мутации Q188R в гене GALT, в несколько десятков раз сильнее, чем в точках, соответствующих последовательностям, не содержащим данные замены. Данный факт говорит о том, что мутация находится в гомозиготном состоянии. Во всех остальных точках уровень сигнала, соответствующий нормальным последовательностям, в несколько раз сильнее сигнала, соответствующего мутированным последовательностям. Данный факт показывает, что заявленный биочип позволяет установить наличие мутации Q188R в гене GALT с высокой специфичностью. При этом не возникает ложноположительных сигналов, соответствующих другим мутациям.
Исходя из всего изложенного выше можно сделать вывод, что биочип по изобретению позволяет:
1. детектировать точечные мутации в гене GALT с высокой специфичностью и надежностью, достаточной для постановки клинического диагноза при заболевании галактаземии 1 типа;
2. установить, находится ли мутация, вызывающая заболевание в гомо- или гетерозиготном состоянии. Данная информация является существенной при подборе лечения заболевания.
Таким образом, все приведенные сведения убедительно показывают, что биочип по изобретению соответствует условию патентоспособности «промышленная применимость».


































Изобретение относится к медицине и биотехнологии. Получен биочип, позволяющий диагностировать заболевание галактоземия 1 типа. Также предложен способ использования полученного биочипа для детекции точковых мутаций в гене GALT. Данный способ включает двухраундный мультиплексный ПЦР с получением одноцепочечного флюоресцентно меченого фрагмента ДНК; гибридизацию на биочипе, содержащем набор олигонуклеотидов; анализ результатов. Данное изобретение расширяет ассортимент арсенала технических средств, используемых при диагностике галактоземии 1 типа. 3 н.п.ф-лы, 23 табл., 34 ил.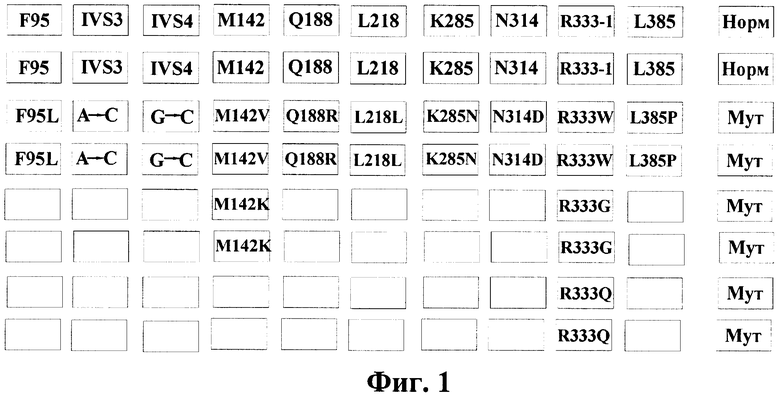
1. Биочип для детекции точковых мутаций в гене GALT, вызывающих галактоземию 1 типа человека, и содержащий последовательности Seq ID NO: 1, Seq ID NO: 2, Seq ID NO: 3, Seq ID NO: 4, Seq ID NO: 5, Seq ID NO: 6, Seq ID NO: 7, Seq ID NO: 8, Seq ID NO: 9, Seq ID NO: 10, Seq ID NO: 12, Seq ID NO: 13, Seq ID NO: 14, Seq ID NO: 15, Seq ID NO: 16, Seq ID NO: 18, Seq ID NO: 19, Seq ID NO: 20, Seq ID NO: 21, Seq ID NO: 22, Seq ID NO: 23, Seq ID NO: 24, Seq ID NO: 25, Seq ID NO: 26, Seq ID NO: 28, Seq ID NO: 29, Seq ID NO: 31.
2. Применение биочипа по п.1 для детекции точковых мутаций в гене GALT, вызывающих галактоземию 1 типа человека.
3. Способ детекции точковых мутаций в гене GALT, вызывающих галактоземию 1 типа человека, состоящий в:
1) постановке двухраундного мультиплексного ПЦР на геномной ДНК пациента с использованием праймеров, содержащих последовательности, Seq ID NO: 32, Seq ID NO: 33, Seq ID NO: 34, Seq ID NO: 35, Seq ID NO: 36, Seq ID NO: 37, Seq ID NO: 38, Seq ID NO: 39, Seq ID NO: 40, Seq ID NO: 41, Seq ID NO: 42, Seq ID NO: 43, Seq ID NO: 44, Seq ID NO: 45, Seq ID NO: 46, Seq ID NO: 47, Seq ID NO: 48, Seq ID NO: 49, Seq ID NO: 50, Seq ID NO: 51, Seq ID NO: 52, Seq ID NO: 53, Seq ID NO: 54, Seq ID NO: 55, Seq ID NO: 56, Seq ID NO: 57, Seq ID NO: 58, Seq ID NO: 59,
2) гибридизации полученных продуктов ПЦР с биочипом по п.1,
3) анализе результатов гибридизации с установлением наличия мутации и ее гомозиготного или гетерозиготного состояния.
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ТОЧЕЧНЫХ НУКЛЕОТИДНЫХ ЗАМЕН В ДНК МИКОБАКТЕРИЙ, СПОСОБ ДИАГНОСТИКИ УСТОЙЧИВОСТИ МИКОБАКТЕРИЙ К РИФАМПИЦИНУ, БИОЧИП ДЛЯ ОСУЩЕСТВЛЕНИЯ ЭТИХ СПОСОБОВ | 2000 |
|
RU2175015C1 |

