Изобретение относится к антителам против глобуломера Aβ, их антигенсвязывающим частям, гибридомам, продуцирующим указанные антитела, нуклеиновым кислотам, кодирующим указанные антитела, векторам, содержащим указанные нуклеиновые кислоты, клеткам-хозяевам, содержащим указанные векторы, способам получения указанных антител, композициям, содержащим указанные антитела, терапевтическим и диагностическим применениям указанных антител и соответствующих способов, относящимся к болезни Альцгеймера и другим амилоидозам.
В 1907 врач Алоис Альцгеймер впервые описал невропатологические признаки формы деменции, впоследствии названной в его честь (Alzheimer 1907). Болезнь Альцгеймера (AD) является наиболее частой причиной деменции среди пожилых с заболеваемостью приблизительно 10% в популяции старше 65 лет. С увеличением возраста вероятность заболевания также возрастает. В мировом масштабе существует приблизительно 15 миллионов пораженных людей и ожидают, что с дальнейшим увеличением продолжительности жизни число заболевших людей увеличится приблизительно в три раза в течение следующих десятилетий.
С молекулярной точки зрения болезнь Альцгеймера (AD) характеризуется накоплением аномально агрегированных белков. В случае внеклеточных амилоидных бляшек эти отложения состоят главным образом из филаментов пептида β-амилоида, в случае внутриклеточных нейрофибриллярных узлов (NFT) - из белка tau. Пептид β-амилоида (Aβ) образуется из белка-предшественника β-амилоида посредством протеолитического расщепления. Это расщепление происходит благодаря кооперативной активности нескольких протеаз, обозначенных α-, β- и γ-секретазой. Расщепление приводит к ряду специфических фрагментов различной длины. Амилоидные бляшки состоят в основном из пептидов длиной 40 или 42 аминокислоты (Aβ40, Aβ42). Доминирующим продуктом расщепления является Aβ40; однако, Aβ42 обладает намного более сильным токсическим эффектом.
Церебральные отложения амилоида и когнитивные нарушения, очень похожие на нарушения, наблюдаемые при заболевании Альцгеймера, являются также признаками синдрома Дауна (трисомии 21), которая возникает с частотой приблизительно 1 на 800 рождений.
Гипотеза амилоидного каскада Харди и Хиггинса постулирует, что увеличенная продукция Aβ(1-42) будет приводить к формированию протофибрилл и фибрилл, главных компонентов бляшек Aβ, причем эти фибриллы отвечают за симптомы болезни Альцгеймера. Несмотря на слабую корреляцию между тяжестью деменции и уровнем накопления бляшек Aβ, этой гипотезе до недавнего времени отдавали предпочтение. Обнаружение растворимых форм Aβ в мозге при AD, которое лучше коррелирует с симптомами AD, чем уровень бляшек, привела к пересмотренной гипотезе амилоидного каскада.
Активная иммунизация пептидами Aβ приводит к уменьшению формирования бляшек, так же как к частичному разложению существующих бляшек. В то же самое время она приводит к облегчению когнитивных нарушений на моделях трансгенных мышей APP.
Для пассивной иммунизации антителами, направленными против пептидов Aβ, также обнаружили уменьшение уровня бляшек Aβ.
Результаты испытания фазы IIa (ELAN Corporation Plc, South San Francisco, CA, USA и Dublin, UK) для активной иммунизации AN-1792 (пептид Aβ(1-42) в состоянии фибриллярной агрегации) позволяют предполагать, что эта иммунотерапия, направленная против пептида Aβ, являлась успешной. В подгруппе из 30 пациентов прогрессирование заболевания было значительно снижено у пациентов с положительным титром антитела против Aβ, измеренным посредством индекса MMSE и DAD. Однако это исследование было остановлено из-за серьезных побочных эффектов в форме менингоэнцефалита (Bennett and Holtzman, 2005, Neurology, 64, 10-12).
Менингоэнцефалит характеризуется нейровоспалением и инфильтрацией T-клеток в мозг. Вероятно, это вызвано T-клеточным иммунным ответом, индуцированным инъекцией Aβ(1-42) в качестве антигена. Такого иммунного ответа не ожидали после пассивной иммунизации. До настоящего времени еще не существует доступных клинических данных относительно этого. Однако по отношению к такому пассивному способу иммунизации высказывали опасения относительно профиля побочных эффектов из-за доклинических исследований на очень старых мышах APP23, которым вводили антитело, направленное против N-концевого эпитопа Aβ(1-42) один раз в неделю в течение 5 месяцев. Для этих мышей показали увеличение количества и тяжести микрокровоизлияний по сравнению с контрольными животными, обработанными физиологическим раствором (Pfeifer et al., 2002, Science, 298, 1379). Описано также сравнительное увеличение микрокровоизлияний у очень старых (> 24 месяцев) мышей Tg2576 и PDAPP (Racke et al., 2005, J. Neurosci, 25, 629-636; Wilcock et al. 2004, J. Neuroinflammation, 1(1):24; De Mattos et al., 2004, Neurobiol. Aging 25(S2):577). Для обеих линий мышей инъекция антитела приводила к значительному увеличению микрокровоизлияний. В отличие от этого антитело, направленное против центральной области пептида Aβ(1-42), не индуцирует микрокровоизлияний (de Mattos et al., выше). Отсутствие индукции микрокровоизлияний являлось связанным с обработкой антителом, которое не связывалось с агрегированным пептидом Aβ при форме CAA (Racke et al., J. Neurosci, 25, 629-636). Однако точный механизм, приводящий к микрокровоизлияниям у трансгенных по APP мышей, неясен. Возможно, церебральная амилоидная ангиопатия (CAA) индуцирует или по меньшей мере усиливает кровоизлияние в мозг. CAA присутствует почти в каждом мозге при болезни Альцгеймера, и приблизительно 20% случаев оценивают как «тяжелую CAA». Пассивная иммунизация таким образом должна быть направлена на исключение микрокровоизлияний посредством выбора антитела, узнающего центральную или C-концевую область пептида Aβ.
В WO 2004/067561 описаны стабильные олигомеры Aβ(1-42) (глобуломеры Aβ(1-42)) и антитела, специфически направленные против глобуломеров. Расщеплением неспецифическими протеазами показали, что глобуломер Aβ можно расщеплять, начиная с гидрофильного N-конца, высунутого из глобулярной коровой структуры (Barghom et al., 2005, J. Neurochem, 95, 834-847). Такие усеченные с N-конца глобуломеры Aβ (Aβ(12-42) и глобуломеры Aβ(20-42)) представляют собой основную структурную единицу этого олигомерного Aβ. Они являются очень высокоиммунным антигеном для активной иммунизации кроликов и мышей, приводящим к высоким титрам антитела (WO 2004/067561). Предположительную патологическую роль усеченных с N-конца форм Aβ in vivo предположили по нескольким недавним сообщениям о их существовании в мозге при AD (Sergeant et al., 2003, J. Neurochem, 85, 1581-1591; Thai et al., 1999, J. Neuropathol. Exp Neurol, 58, 210-216). В ходе расщепления in vivo могут быть вовлечены конкретные протеазы, обнаруженные в мозге, например, неприлизин (NEP 24.11) или инсулиндеградирующий фермент (IDE) (Selkoe, 2001, Neuron, 32, 177-180).
Целью настоящего изобретения являлось предоставление антител, направленных против глобуломеров Aβ, улучшающих когнитивное поведение пациента при иммунотерапии, в то же самое время вступающих в реакцию только с малой частью полного количества пептида Aβ в мозге. Ожидают, что это будет предотвращать значительный дисбаланс церебрального Aβ и приводить к меньшим побочным эффектам. (Например, наблюдали терапевтически сомнительное уменьшение объема мозга при исследовании активной иммунизации пептидами Aβ в состоянии фибриллярной агрегации (испытание ELAN с AN1792). Более того, в этом испытании наблюдали тяжелые побочные эффекты в форме менингоэнцефалита.
Настоящее изобретение решает эту проблему предоставлением глобуломерспецифических антител, обладающих высокой аффинностью для усеченных форм глобуломеров Aβ. Эти антитела способны к дискриминации не только других форм пептидов Aβ, в частности мономеров и фибрилл, но также неусеченных форм глобуломеров Aβ.
Таким образом, настоящее изобретение относится к антителу, обладающему аффинностью связывания для глобуломера Aβ(20-42), превышающей аффинность связывания этого антитела для глобуломера Aβ(1-42).
Кроме того, настоящее изобретение относится к антителу, обладающему аффинностью связывания с глобуломером Aβ(20-42), превышающей аффинность связывания этого антитела для глобуломера Aβ(12-42).
Согласно конкретному варианту осуществления изобретение таким образом относится к антителам, обладающим аффинностью связывания для глобуломера Aβ(20-42), превышающей аффинность связывания этого антитела как для глобуломера Aβ(1-42), так и для глобуломера Aβ(12-42).
Термин «Aβ(X-Y)» здесь относится к аминокислотной последовательности от положения аминокислоты X до положения аминокислоты Y белка β-амилоида человека, включая как X, так и Y, в частности к аминокислотной последовательности от положения аминокислоты X до положения аминокислоты Y аминокислотной последовательности DAEFRHDSGY EVHHQKLVFF AEDVGSNKGA IIGLMVGGVV IAT (соответствующей положениям аминокислот 1-43) или любому из ее встречающихся в природе вариантов, в частности вариантов по меньшей мере с одной мутацией, выбранной из группы, состоящей из A2T, H6R, D7N, A21G («фламандская»), E22G («арктическая»), E22Q («голландская»), E22K («итальянская»), D23N («Айова»), A42T и A42V, где номера приведены относительно старта пептида Aβ, включая как положение X, так и положение Y, или последовательности, содержащей вплоть до трех дополнительных замен аминокислот, ни одна из которых не может предотвращать формирование глобуломера, предпочтительно без дополнительных замен аминокислот в части от аминокислоты 12 или X, в зависимости от того, какой номер выше, до аминокислоты 42 или Y, в зависимости от того, какой номер ниже, более предпочтительно, без дополнительных замен аминокислот в части от аминокислоты 20 или X, в зависимости от того, какой номер выше, до аминокислоты 42 или Y, в зависимости от того, какой номер ниже, и наиболее предпочтительно, без дополнительных замен аминокислот в части от аминокислоты 20 или X, в зависимости от того, какой номер выше, до аминокислоты 40 или Y, в зависимости от того, какой номер ниже, где «дополнительная» замена аминокислоты здесь представляет собой любое отклонение от канонической последовательности, не обнаруженное в природе.
Более конкретно, термин «Aβ(1-42)» здесь относится к аминокислотной последовательности от положения аминокислоты 1 до положения аминокислоты 42 в белке β-амилоида, включая как 1, так и 42, в частности к аминокислотной последовательности DAEFRHDSGY EVHHQKLVFF AEDVGSNKGA IIGLMVGGVV IA или любому из ее встречающихся в природе вариантов, в частности вариантов по меньшей мере с одной мутацией, выбранной из группы, состоящей из A2T, H6R, D7N, A21G («фламандская»), E22G («арктическая»), E22Q («голландская»), E22K («итальянская»), D23N («Айова»), A42T и A42V, где номера приведены относительно старта пептида Aβ, включая как 1, так и 42, или последовательности, содержащей вплоть до трех дополнительных замен аминокислот, ни одна из которых не может предотвращать формирование глобуломера, предпочтительно без дополнительных замен аминокислот в части от аминокислоты 20 до аминокислоты 42. Подобным образом, термин «Aβ(1-40)» здесь относится к аминокислотной последовательности от положения аминокислоты 1 до положения аминокислоты 40 белка β-амилоида человека, включая как 1, так и 40, в частности к аминокислотной последовательности DAEFRHDSGY EVHHQKLVFF AEDVGSNKGA IIGLMVGGVV или любому из ее встречающихся в природе вариантов, в частности вариантов по меньшей мере с одной мутацией, выбранной из группы, состоящей из A2T, H6R, D7N, A21G («фламандская»), E22G («арктическая»), E22Q («голландская»), E22K («итальянская»), и D23N («Айова»), где номера приведены относительно старта пептида Aβ, включая как 1, так и 40, или последовательности, содержащей вплоть до трех дополнительных замен аминокислот, ни одна из которых не может предотвращать формирование глобуломера, предпочтительно, без дополнительных замен аминокислот в части от аминокислоты 20 до аминокислоты 40.
Более конкретно, термин «Aβ(12-42)» здесь относится к аминокислотной последовательности от положения аминокислоты 12 до положения аминокислоты 42 белка β-амилоида человека, включая как 12, так и 42, в частности, к аминокислотной последовательности VHHQKLVFF AEDVGSNKGA IIGLMVGGVV IA или любому из ее встречающихся в природе вариантов, в частности вариантов по меньшей мере с одной мутацией, выбранной из группы, состоящей из A21G («фламандская»), E22G («арктическая»), E22Q («голландская»), E22K («итальянская»), D23N («Айова»), A42T и A42V, где номера приведены относительно старта пептида Aβ, включая как 12, так и 42, или последовательности, содержащей вплоть до трех дополнительных замен аминокислот, ни одна из которых не может предотвращать формирование глобуломера, предпочтительно, без дополнительных замен аминокислот в части от аминокислоты 20 до аминокислоты 42.
Более конкретно, термин «Aβ(20-42)» здесь относится к аминокислотной последовательности от положения аминокислоты 20 до положения аминокислоты 42 белка β-амилоида человека, включая как 20, так и 42, в частности к аминокислотной последовательности F AEDVGSNKGA IIGLMVGGVV IA или любому из ее встречающихся в природе вариантов, в частности вариантов по меньшей мере с одной мутацией, выбранной из группы, состоящей из A21G («фламандская»), E22G («арктическая»), E22Q («голландская»), E22K («итальянская»), D23N («Айова»), A42T и A42V, где номера приведены относительно старта пептида Aβ, включая как 20, так и 42, или последовательности, содержащей вплоть до трех дополнительных замен аминокислот, ни одна из которых не может предотвращать формирование глобуломера, предпочтительно, без каких-либо дополнительных замен аминокислот.
Термин «глобуломер Aβ(X-Y)» (глобулярный олигомер Aβ(X-Y)) здесь относится к растворимому, глобулярному, нековалентному объединению пептидов Aβ(X-Y), как определено выше, обладающему гомогенностью и четкими физическими характеристиками. Согласно одному аспекту глобуломеры Aβ(X-Y) представляют собой стабильные, нефибриллярные, олигомерные ансамбли пептидов Aβ(X-Y), которые можно получить инкубацией с анионными детергентами. В отличие от мономеров и фибрилл, эти глобуломеры характеризуются определенным числом субъединиц в ансамбле (например, формы ранних ансамблей, n=4-6, «олигомеры A», и формы поздних ансамблей, n=12-14, «олигомеры B», как описано в WO2004/067561). Глобуломеры обладают 3-мерной структурой глобулярного типа («расплавленная глобула», см. Barghorn et al., 2005, J. Neurochem, 95, 834-847). Они могут дополнительно характеризоваться одним или несколькими из следующих свойств:
- возможность отщепления N-концевых аминокислот X-23 неспецифическими протеазами (такими как термолизин или эндопротеаза GluC) с получением усеченных форм глобуломеров;
- недоступность C-концевых аминокислот 24-Y для неспецифических протеаз и антител;
- усеченные формы этих глобуломеров поддерживают 3-мерную коровую структуру указанных глобуломеров с лучшей доступностью корового эпитопа Aβ(20-Y) в его глобуломерной конформации.
Согласно изобретению и, в частности, для целей оценки аффинностей связывания антител по настоящему изобретению термин «глобуломер Aβ(X-Y)» здесь относится, в частности, к продукту, который можно получить способом, как описано в WO 2004/067561, содержание которого приведено здесь в качестве ссылки.
Указанный способ включает в себя разворачивание природного, рекомбинантного или синтетического пептида Aβ(X-Y) или его производного; подвергание по меньшей мере частично развернутого пептида Aβ(X-Y) или его производного воздействию детергента, уменьшение действия детергента и продолжение инкубации.
Для целей разворачивания пептида можно допускать воздействие на белок разрушающих водородную связь средств, например, таких как гексафторизопропанол (HFIP). Времени воздействия несколько минут, например приблизительно 10-60 минут, достаточно, когда температура воздействия составляет от приблизительно 20 до 50°C и в частности, приблизительно 35-40°C. Последующим растворением осадка, выпаренного до сухости, предпочтительно, в концентрированной форме, в подходящих органических растворителях, способных смешиваться с водными буферами, например, таких как диметилсульфоксид (DMSO), получают суспензию по меньшей мере частично развернутого пептида или его производного, которые можно затем использовать. Если необходимо, исходную суспензию можно хранить при низкой температуре, например, приблизительно при -20°C, в течение временного периода.
Альтернативно, пептид или его производное можно помещать в слабокислый, предпочтительно водный, раствор, например, в водный раствор приблизительно 10 мМ HCl. После времени инкубации, обычно нескольких минут, нерастворимые компоненты удаляют центрифугированием. Подходят несколько минут при 10000 g. Эти стадии способа предпочтительно проводят при комнатной температуре, т.е. температуре в диапазоне от 20 до 30°C. Супернатант, полученный после центрифугирования, содержит пептид Aβ(X-Y) или его производное, и его можно хранить при низкой температуре, например, приблизительно при -20°C, в течение временного периода.
Следующее воздействие детергента относится к олигомеризации пептида или его производного для получения промежуточного типа олигомеров (обозначенных в WO 2004/067561 как олигомеры A). Для этой цели детергенту позволяют действовать на по меньшей мере частично развернутый пептид или его производное до получения достаточного количества промежуточного олигомера.
Предпочтение оказывают применению ионных детергентов, в частности анионных детергентов.
Согласно конкретному варианту осуществления используют детергент формулы (I):
R-X,
в котором радикал R представляет собой неразветвленный или разветвленный алкил, обладающий от 6 до 20 и, предпочтительно, от 10 до 14 атомов углерода, или неразветвленный или разветвленный алкенил, обладающий от 6 до 20 и, предпочтительно, от 10 до 14 атомов углерода, радикал X представляет собой кислую группу или ее соль, с X, предпочтительно, выбранным из -COO-M+, -SO3 -M+, и особенно -OSO3 -M+, и M+ представляет собой катион водорода или неорганический или органический катион, предпочтительно, выбранный из катионов щелочного металла и щелочноземельного металла и катионов аммония.
Преимущественными являются детергенты формулы (I), в которых R представляет собой неразветвленный алкил, в котором, в частности, должны быть приведены алкильные радикалы. Особенное предпочтение отдают додецилсульфату натрия (SDS). Предпочтительно можно использовать также лауриновую кислоту и олеиновую кислоту. Натриевая соль детергента лаурилсаркозина (известная также как саркозил NL-30 или Gardol®) также является особенно предпочтительной.
Время воздействия детергента зависит, в частности, от того, является ли - и если является, до какой степени, - пептид или его производное, подвергаемое олигомеризации, развернутым. Если согласно стадии разворачивания пептид или его производное предварительно обрабатывают разрушающим водородные связи средством, т.е., в частности, гексафторизопропанолом, времени воздействия в диапазоне нескольких часов, преимущественно от приблизительно 1 до 20 и, в частности, от приблизительно 2 до 10 часов, достаточно при температуре воздействия приблизительно 20-50°C и, в частности, приблизительно 35-40°C. Если исходной точкой является несвернутый или в основном не свернутый пептид или его производное, соответственно, более длительное время воздействия является целесообразным. Если пептид или его производное предварительно обработан, например, согласно способу, указанному выше, в качестве альтернативы обработке HFIP, или указанный пептид или его производное непосредственно подвергают олигомеризации, достаточно времени воздействия в диапазоне от приблизительно 5 до 30 часов и, в частности, от приблизительно 10 до 20 часов, когда температура воздействия составляет приблизительно 20-50°C и, в частности, приблизительно 35-40°C. После инкубации нерастворимые компоненты предпочтительно удаляют центрифугированием. Несколько минут при 10000 g являются целесообразными.
Выбираемая концентрация детергента зависит от используемого детергента. Если применяют SDS, доказано, что концентрация в диапазоне от 0,01 до 1% по массе, предпочтительно от 0,05 до 0,5% по массе, например приблизительно 0,2% по массе, является целесообразной. Если применяют лауриновую кислоту или олеиновую кислоту, целесообразными являются несколько более высокие концентрации, например, в диапазоне от 0,05 до 2% по массе, предпочтительно от 0,1 до 0,5% по массе, например, приблизительно 0,5% по массе.
Действие детергента должно происходить при концентрации соли приблизительно в физиологическом диапазоне. Таким образом, целесообразными являются концентрации NaCl, в частности, в диапазоне от 50 до 500 мМ, предпочтительно от 100 до 200 мМ и, в частности, приблизительно 140 мМ.
Последующее уменьшение действия детергента и продолжение инкубации относится к дальнейшей олигомеризации для получения глобуломера Aβ(X-Y) по изобретению (в WO 2004/067561 обозначенного как олигомеры B). Поскольку композиция, полученная на предшествующих стадиях, обычно содержит детергент и концентрацию соли в физиологическом диапазоне, является целесообразным уменьшить действие детергента и, предпочтительно, также концентрацию соли. Это можно осуществлять уменьшением концентрации детергента и соли, например разведением, целесообразно водой или буфером с более низкой концентрации соли, например Трис-HCl, pH 7,3. Доказано, что подходят факторы разведения, лежащие в диапазоне от приблизительно 2 до 10, преимущественно в диапазоне от приблизительно 3 до 8 и, в частности, приблизительно 4. Уменьшения действия детергента можно также достигать добавлением веществ, которые могут нейтрализовать указанное действие детергента. Примеры веществ включают в себя вещества, способные к образованию комплексов с детергентами, такие как вещества, способные стабилизировать клетки в ходе способов очистки и выделения, например, конкретные блок-сополимеры EO/PO, в частности блок-сополимеры под торговым наименованием Pluronc® F 68. Равным образом, можно использовать алкоксилированные и, в частности, этоксилированные алкилфенолы, такие как этоксилированные t-октилфенолы серии Triton® X, в частности Triton® X100, 3-(3-хлорамидопропилдиметиламмоний)-1-пропансульфонат (CHAPS®) или алкоксилированные и, в частности, этоксилированные жирные сложные эфиры сорбитана, такие как эфиры серий Tween®, в частности Tween® 20, в диапазонах концентраций в районе или выше конкретной критической концентрации мицелл.
Затем раствор инкубируют до получения достаточного количества глобуломера Aβ(X-Y) по изобретению. Времени действия в диапазоне нескольких часов, предпочтительно в диапазоне от приблизительно 10 до 30 часов, и в частности, в диапазоне от приблизительно 15 до 25 часов, достаточно, когда температура действия составляет приблизительно 20-50°C и, в частности, приблизительно 35-40°C. Раствор затем можно концентрировать и возможные осадки можно удалять центрифугированием. Доказано, что здесь также являются целесообразными несколько минут при 10000 g. Супернатант, полученный после центрифугирования, содержит глобуломер Aβ(X-Y) по изобретению.
Глобуломер Aβ(X-Y) по изобретению можно окончательно выделять в чистом виде известным способом, например, ультрафильтрацией, диализом, преципитацией или центрифугированием.
Кроме того, является предпочтительным, если при электрофоретическом разделении глобуломеров Aβ(X-Y) в денатурирующих условиях, например, в SDS-PAGE, получают двойную полосу (например, с кажущейся молекулярной массой 38 / 48 кДа для Aβ(1-42)), и особенно предпочтительно, если при обработке глутардиальдегидом глобуломеров перед разделением эти две полосы сливаются в одну. Также является предпочтительным, если при эксклюзионной хроматографии глобуломеров получают отдельный пик (например, соответствующий молекулярной массе приблизительно 100 кДа для глобуломера Aβ(1-42) или приблизительно 60 кДа для перекрестно сшитого глутардиальдегидом глобуломера Aβ(1-42)), соответственно.
Начиная с пептида Aβ(1-42), пептида Aβ(12-42) и пептида Aβ(20-42), указанные способы, в частности, являются пригодными для получения глобуломеров Aβ(1-42), глобуломеров Aβ(12-42) и глобуломеров Aβ(20-42).
В конкретном варианте осуществления по изобретению глобуломеры Aβ(X-Y), где X выбран из группы, состоящей из номеров 2.. 24 и Y является таким, как определено выше, представляют собой такие, какие можно получить усечением глобуломеров Aβ(1-Y) до более коротких форм, где X выбран из группы, состоящей из номеров 2.. 24, где X предпочтительно представляет собой 20 или 12, и Y является таким, как определено выше, которые можно получить обработкой соответствующими протеазами. Например, глобуломер Aβ(20-42) можно получить, подвергая глобуломер Aβ(1-42) протеолизу термолизином, и глобуломер Aβ(12-42) можно получить, подвергая глобуломер Aβ(1-42) протеолизу эндопротеазой GluC. При достижении желаемой степени протеолиза протеазу инактивируют общеизвестным способом. Полученные глобуломеры затем можно выделять согласно способам, уже описанным здесь и, если необходимо, посредством дополнительной обработки посредством дополнительных стадий воздействия и очистки. Подробное описание указанных способов описано в WO 2004/067561, содержание которого приведено здесь в качестве ссылки.
Для целей по настоящему изобретению глобуломер Aβ(1-42), в частности, представляет собой глобуломер Aβ(1-42), как описано в примере 1a здесь; глобуломер Aβ(20-42), в частности, представляет собой глобуломер Aβ(20-42), как описано в примерах 1c здесь, и глобуломер Aβ(12-42), в частности, представляет собой глобуломер Aβ(12-42), как описано в примерах 1d здесь.
Предпочтительно, для глобуломера показывают аффинность для нейрональных клеток. Предпочтительно, для глобуломера также показывают нейромодулирующие эффекты.
Согласно другому аспекту изобретения глобуломер состоит из 11-16 и, наиболее предпочтительно, из 12-14 пептидов Aβ(X-Y).
Согласно другому аспекту изобретения термин «глобуломер Aβ(X-Y)» здесь относится к глобуломеру, состоящему в основном из субъединиц Aβ(X-Y), где является предпочтительным, если в среднем по меньшей мере 11 из 12 субъединиц принадлежат к типу Aβ(X-Y), более предпочтительно, если менее чем 10% глобуломеров содержат какие-либо не относящиеся к Aβ(X-Y) пептиды и, наиболее предпочтительно, если содержание не относящихся к Aβ(X-Y) пептидов ниже порога детекции.
Более конкретно, термин «глобуломер Aβ(1-42)» здесь относится к глобуломеру, состоящему в основном из единиц Aβ(1-42), как определено выше; термин «глобуломер Aβ(12-42)» здесь относится к глобуломеру, состоящему в основном из единиц Aβ(12-42), как определено выше; и термин «глобуломер Aβ(20-42)» здесь относится к глобуломеру, состоящему в основном из единиц Aβ(20-42), как определено выше.
Термин «перекрестно-сшитый глобуломер Aβ(X-Y)» здесь относится к молекуле, которую можно получить из глобуломера Aβ(X-Y), как описано выше, посредством перекрестного сшивания, предпочтительно, химического перекрестного сшивания, более предпочтительно, перекрестного сшивания альдегидом, наиболее предпочтительно, перекрестного сшивания глутардиальдегидом составляющих глобуломер единиц. В другом аспекте изобретения перекрестно-сшитый глобуломер в основном представляет собой глобуломер, в котором единицы являются по меньшей мере частично соединенными посредством ковалентных связей, а не удерживаемыми вместе посредством только нековалентных взаимодействий. Для целей настоящего изобретения перекрестно сшитый глобуломер Aβ(1-42) представляет собой, в частности, перекрестно-сшитый олигомер Aβ(1-42), как описано в примере 1b здесь.
Термин «производное глобуломера Aβ(X-Y)» здесь относится, в частности, к глобуломеру, который является меченым посредством ковалентного связывания с группой, облегчающей детекцию, предпочтительно флуорофором, например, флуоресцеинизотиоцианатом, фикоэритрином, флуоресцентным белком Aequorea victoria, флуоресцентным белком Dictyosoma или любым их сочетанием или флуоресцентно активным производным; хромофором; хемилюминофором, например люциферазой, предпочтительно люциферазой Photinus pyralis, люциферазой Vibrio fischeri или любым их сочетанием или хемилюминесцентно активным производным; ферментативно активной группой, например, пероксидазой, например, пероксидазой хрена или любым их ферментативно активным производным; электронноплотной группой, например, группой, содержащей тяжелый металл, например, группой, содержащей золото; гаптеном, например, полученным из фенола гаптеном; структурой сильного антигена, например пептидной последовательностью, как предсказано, являющейся антигенной, например, являющейся антигенной, как предсказано по алгоритму Kolaskar и Tongaonkar; аптамером для другой молекулы; хелатообразующей группой, например гексагистидинилом; природной или полученной из природной белковой структурой, опосредующей дополнительные белок-белковые взаимодействия, например, членом пары fos/jun; магнитной группой, например ферромагнитной группой; или радиоактивной группой, например группой, содержащей 1H, 14C, 32P, 35S или 125I, или любое их сочетание; или к глобуломеру, маркированному посредством ковалентного или нековалентного высокоаффинного взаимодействия, предпочтительно ковалентно связанному с группой, облегчающей инактивацию, секвестрацию, деградацию и/или преципитацию, предпочтительно маркированному группой, стимулирующей деградацию in vivo, более предпочтительно убиквитином, где особенно предпочтительным является, чтобы этот маркированный олигомер являлся собранным in vivo; или к глобуломеру, модифицированному любым сочетанием вышеуказанного. Такие группы для мечения и маркировки и способы для присоединения их к белкам известны в данной области. Мечение и/или маркировку можно выполнять до, во время или после глобуломеризации. В другом аспекте изобретения производное глобуломера представляет собой молекулу, которую можно получить из глобуломера посредством реакции мечения и/или маркировки.
Соответственно, термин «производное мономера Aβ(X-Y)» здесь относится, в частности, к мономеру Aβ, который является меченым или маркированным, как описано для глобуломера.
Целесообразно, если антитело по настоящему изобретению связывает глобуломер Aβ(20-42) с KD в диапазоне от 1×10-6 M до 1×10-12 M. Предпочтительно, антитело связывает глобуломер Aβ(20-42) с высокой аффинностью, например с KD 1×10-7 M или большей аффинностью, например с KD 3×10-8 M или большей аффинностью, с KD 1×10-8 M или большей аффинностью, например, с KD 3×10-9 M или большей аффинностью, с KD 1×10-9 M или большей аффинностью, например, с KD 3×10-10 M или большей аффинностью, с KD 1×10-10 M или большей аффинностью, например, с KD 3×10-11 M или большей аффинностью, или с KD 1×10-11 M или большей аффинностью.
Термин «большая аффинность» здесь относится к степени взаимодействия, где равновесие между несвязанным антителом и несвязанным глобуломером, с одной стороны, и комплексом антитело-глобуломер, с другой, больше сдвинуто в сторону комплекса антитело-глобуломер. Подобным образом, термин «меньшая аффинность» здесь относится к степени взаимодействия, где равновесие между несвязанным антителом и несвязанным глобуломером, с одной стороны, и комплексом антитело-глобуломер, с другой, больше сдвинуто в сторону несвязанного антитела и несвязанного глобуломера. Термин «большая аффинность» является синонимом термина «более высокая аффинность», а термин «меньшая аффинность» является синонимом термина «более низкая аффинность».
Согласно конкретному варианту осуществления изобретение относится к антителу, связывающему глобуломер Aβ(20-42) с KD в диапазоне от 1×10-6 M до 1×10-12 M, глобуломер Aβ(1-42) с KD 10-12 M или меньшей аффинностью, где аффинность связывания с глобуломером Aβ(20-42) больше, чем аффинность связывания с глобуломером Aβ(1-42).
Является предпочтительным, чтобы аффинность связывания антитела по настоящему изобретению с глобуломером Aβ(20-42) по меньшей мере в 2 раза, например, по меньшей мере в 3 раза или по меньшей мере в 5 раз, предпочтительно, по меньшей мере в 10 раз, например, по меньшей мере в 20 раз, по меньшей мере в 30 раз или по меньшей мере в 50 раз, более предпочтительно, по меньшей мере в 100 раз, например, по меньшей мере в 200 раз, по меньшей мере в 300 раз или по меньшей мере в 500 раз и даже, более предпочтительно, по меньшей мере в 1000 раз, например по меньшей мере в 2000 раз, по меньшей мере в 3000 раз или по меньшей мере в 5000 раз, даже более предпочтительно, по меньшей мере в 10000 раз, например, по меньшей мере в 20000 раз, по меньшей мере в 30000 или по меньшей мере в 50000 раз и, наиболее предпочтительно, по меньшей мере в 100000 раз превышала аффинность связывания антитела с глобуломером Aβ(1-42).
Согласно конкретному варианту осуществления изобретение относится к антителу, связывающему глобуломер Aβ(12-42) с KD 10-12 M или меньшей аффинностью, где аффинность связывания с глобуломером Aβ(20-42) превышает аффинность связывания с глобуломером Aβ(12-42).
Является предпочтительным также, чтобы аффинность связывания антитела по настоящему изобретению с глобуломером Aβ(20-42) по меньшей мере в 2 раза, например, по меньшей мере в 3 раза или по меньшей мере в 5 раз, предпочтительно по меньшей мере в 10 раз, например, по меньшей мере в 20 раз, по меньшей мере в 30 раз или по меньшей мере в 50 раз, более предпочтительно по меньшей мере в 100 раз, например, по меньшей мере в 200 раз, по меньшей мере в 300 раз или по меньшей мере в 500 раз, и даже, более предпочтительно, по меньшей мере в 1000 раз, например, по меньшей мере в 2000 раз, по меньшей мере в 3000 раз или по меньшей мере в 5000 раз, даже более предпочтительно по меньшей мере в 10000 раз, например, по меньшей мере в 20000 раз, по меньшей мере в 30000 или по меньшей мере в 50000 раз, и, наиболее предпочтительно, по меньшей мере в 100000 раз превышала аффинность связывания антитела с глобуломером Aβ(12-42).
Предпочтительно, антитела по настоящему изобретению связывают по меньшей мере один глобуломер Aβ, как определено выше, и обладают сравнительно меньшей аффинностью по меньшей мере для одной не относящейся к глобуломеру формой Aβ.
Антитела по настоящему изобретению, обладающие сравнительно меньшей аффинностью по меньшей мере для одной не относящейся к глобуломеру формы Aβ, чем по меньшей мере для одного глобуломера Aβ, включают в себя антитела, обладающие аффинностью связывания с глобуломером Aβ(20-42), превышающей аффинность для мономера Aβ(1-42). Кроме того, является предпочтительным, чтобы, альтернативно или дополнительно, аффинность связывания антитела с глобуломером Aβ(20-42) превышала аффинность для мономера Aβ(1-40).
В предпочтительном варианте осуществления по изобретению аффинность антитела для глобуломера Aβ(20-42) превышает его аффинность как к мономеру Aβ(1-40), так и к мономеру Aβ(1-42).
Термин «мономер Aβ(X-Y)» здесь относится к выделенной форме пептида Aβ(X-Y), предпочтительно, в форме пептида Aβ(X-Y), не вступающей в главным образом нековалентные взаимодействия с другими пептидами Aβ. На практике, мономер Aβ(X-Y) обычно получают в форме водного раствора. В особенно предпочтительном варианте осуществления по изобретению водный раствор мономера содержит 0,05%-0,2%, более предпочтительно, приблизительно 0,1% NH4OH. В другом особенно предпочтительном варианте осуществления изобретения водный раствор мономера содержит 0,05%-0,2%, более предпочтительно, приблизительно 0,1% NaOH. При использовании (например, для определения аффинностей связывания антител по настоящему изобретению), может являться целесообразным разводить указанный раствор подходящим образом. Кроме того, обычно целесообразно использовать указанный раствор в течение 2 часов, в частности в течение 1 часа, и особенно в течение 30 минут после его приготовления.
Более конкретно, термин «мономер Aβ(1-40)» здесь относится к препарату мономера Aβ(1-40), как описано в примере 2 здесь, и термин «мономер Aβ(1-42)» здесь относится к препарату Aβ(1-42), как описано в примере 2 здесь.
Целесообразно, чтобы антитело по настоящему изобретению связывало один или, более предпочтительно, оба мономера с низкой аффинностью, наиболее предпочтительно, с KD 1×10-8 M или с меньшей аффинностью, например, с KD 3×10-8 M или с меньшей аффинностью, с KD 1×10-7 M или с меньшей аффинностью, например, с KD 3×1-7 M или с меньшей аффинностью, или с KD 1×10-6 M или с меньшей аффинностью, например, с KD 3×10-5 M или с меньшей аффинностью, или с KD 1×10-5 M или с меньшей аффинностью.
Особенно предпочтительным является, чтобы аффинность связывания антитела по настоящему изобретению с глобуломером Aβ(20-42) по меньшей мере в 2 раза, например, по меньшей мере в 3 раза или по меньшей мере в 5 раз, предпочтительно по меньшей мере в 10 раз, например, по меньшей мере в 20 раз, по меньшей мере в 30 раз или по меньшей мере в 50 раз, более предпочтительно по меньшей мере в 100 раз, например по меньшей мере в 200 раз, по меньшей мере в 300 раз или по меньшей мере в 500 раз, и даже более предпочтительно, по меньшей мере в 1000 раз, например, по меньшей мере в 2000 раз, по меньшей мере в 3000 раз или по меньшей мере в 5000 раз, даже более предпочтительно, по меньшей мере в 10000 раз, например, по меньшей мере в 20000 раз, по меньшей мере в 30000 или по меньшей мере в 50000 раз, и наиболее предпочтительно, по меньшей мере в 100000 раз превышала аффинность связывания антитела с одним или, более предпочтительно, с обоими мономерами.
Кроме того, антитела по настоящему изобретению, обладающие сравнительно меньшей аффинностью по меньшей мере для одной не относящейся к глобуломеру формы Aβ, чем по меньшей мере для одного глобуломера Aβ, дополнительно включают в себя антитела, обладающие аффинностью связывания с глобуломером Aβ(20-42), превышающей аффинность связывания для фибрилл Aβ(1-42). Кроме того, является предпочтительным, чтобы, альтернативно или дополнительно, аффинность связывания антитела с глобуломером Aβ(20-42) превышала аффинность связывания для фибрилл Aβ(1-40).
Термин «фибрилла» здесь относится к молекулярной структуре, содержащей ансамбли нековалентно связанных отдельных пептидов Aβ(X-Y), для которой показана фибриллярная структура в электронном микроскопе, связывание Конго красного, и затем двойное лучепреломление под поляризованным светом и которая по характеру рентгенодифракции представляет собой сшитую β-структуру.
В другом аспекте по изобретению фибрилла представляет собой молекулярную структуру, которую можно получить способом, включающим в себя самоиндуцированную полимерную агрегацию соответствующего пептида Aβ в отсутствие детергентов, например, в 0,1 M HCl, приводящую к формированию агрегатов из более, чем 24, предпочтительно более чем 100 единиц. Эти способы хорошо известны в данной области. Целесообразно использовать фибриллы Aβ(X-Y) в форме водного раствора. В особенно предпочтительном варианте осуществления изобретения водный раствор фибрилл получают растворением пептида Aβ в 0,1% NH4OH, разведением его 1:4 в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, с последующим повторным доведением pH до 7,4, инкубацией раствора при 37°C в течение 20 час с последующим центрифугированием при 10000 g в течение 10 мин и ресуспендированием в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4.
Термин «фибрилла Aβ(X-Y)» здесь относится к фибрилле, состоящей в основном из субъединиц Aβ(X-Y), где предпочтительным является, если в среднем по меньшей мере 90% субъединиц являются субъединицами типа Aβ(X-Y), более предпочтительно, если по меньшей мере 98% субъединиц являются субъединицами типа Aβ(X-Y), и наиболее предпочтительно, если содержание не относящихся к Aβ(X-Y) пептидов ниже порога детекции.
Более конкретно, термин «фибрилла Aβ(1-42)» здесь относится к препарату фибриллы Aβ(1-42), как описано в примере 3 здесь.
Целесообразно, чтобы антитело по настоящему изобретению связывало одну или, более предпочтительно, обе фибриллы с низкой аффинностью, наиболее предпочтительно с KD 1×10-8 M или с меньшей аффинностью, например с KD 3×10-8 M или с меньшей аффинностью, с KD 1×10-7 M или с меньшей аффинностью, например с KD 3×10-7 M или с меньшей аффинностью, или с KD 1×10-6 M или с меньшей аффинностью, например, с KD 3×10-5 M или с меньшей аффинностью, или с KD 1×10-5 M или с меньшей аффинностью.
Особенно предпочтительным является, чтобы аффинность связывания антитела по настоящему изобретению с глобуломером Aβ(20-42) по меньшей мере в 2 раза, например по меньшей мере в 3 раза или по меньшей мере в 5 раз, предпочтительно, по меньшей мере в 10 раз, например, по меньшей мере в 20 раз, по меньшей мере в 30 раз или по меньшей мере в 50 раз, более предпочтительно, по меньшей мере в 100 раз, например, по меньшей мере в 200 раз, по меньшей мере в 300 раз или по меньшей мере в 500 раз, и даже более предпочтительно, по меньшей мере в 1000 раз, например, по меньшей мере в 2000 раз, по меньшей мере в 3000 раз или по меньшей мере в 5000 раз, даже более предпочтительно, по меньшей мере в 10000 раз, например, по меньшей мере в 20000 раз, по меньшей мере в 30000 или по меньшей мере в 50000 раз, и наиболее предпочтительно, по меньшей мере в 100000 раз превышала аффинность связывания антитела с одной или, более предпочтительно, с обеими фибриллами.
Согласно одному из конкретных вариантов осуществления изобретение относится к антителам, обладающим аффинностью связывания с глобуломером Aβ(20-42), превышающей его аффинность связывания как с фибриллами Aβ(1-40), так и с фибриллами Aβ(1-42).
Согласно особенно предпочтительному варианту осуществления настоящее изобретение относится к антителам, обладающим сравнительно меньшей аффинностью как для мономерной, так и для фибриллярной форм Aβ, чем по меньшей мере для одного глобуломера Aβ, в частности глобуломера Aβ(20-42). Эти антитела далее здесь обозначают как специфические для глобуломера антитела.
Кроме того, антитела по настоящему изобретению включают в себя антитела, обладающие аффинностью связывания с глобуломером Aβ(20-42), превышающей аффинность связывания для перекрестно сшитого глобуломера Aβ(1-42), в частности для перекрестно сшитого глутардиальдегидом глобуломера Aβ(1-42), такого как описанный в примере 1b здесь.
В особенно предпочтительном варианте осуществления по изобретению аффинность связывания антитела с глобуломером Aβ(20-42) по меньшей мере в 2 раза, например, по меньшей мере в 3 раза или по меньшей мере в 5 раз, предпочтительно, по меньшей мере в 10 раз, например, по меньшей мере в 20 раз, по меньшей мере в 30 раз или по меньшей мере в 50 раз, более предпочтительно, по меньшей мере в 100 раз, например по меньшей мере в 200 раз, по меньшей мере в 300 раз или по меньшей мере в 500 раз, и даже более предпочтительно по меньшей мере в 1000 раз, например, по меньшей мере в 2000 раз, по меньшей мере в 3000 раз или по меньшей мере в 5000 раз, даже более предпочтительно, по меньшей мере в 10000 раз, например, по меньшей мере в 20000 раз, по меньшей мере в 30000 или по меньшей мере в 50000 раз, и наиболее предпочтительно, по меньшей мере в 100000 раз превышает аффинность связывания антитела с перекрестно сшитым глобуломером Aβ(1-42).
Антитела по настоящему изобретению являются предпочтительно выделенными, в частности моноклональными и, более конкретно, рекомбинантными.
Настоящее изобретение также относится к моноклональному антителу (5F7), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7241.
Настоящее изобретение также относится к моноклональному антителу (10F11), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7239.
Настоящее изобретение также относится к моноклональному антителу (7C6), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7240.
Настоящее изобретение также относится к моноклональному антителу (4B7), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7242.
Настоящее изобретение также относится к моноклональному антителу (6A2), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7409.
Настоящее изобретение также относится к моноклональному антителу (2F2), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7408.
Настоящее изобретение также относится к моноклональному антителу (4D10), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7405.
Настоящее изобретение также относится к моноклональному антителу (7E5), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7809.
Настоящее изобретение также относится к моноклональному антителу (10C1), которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7810.
Настоящее изобретение также относится к моноклональному антителу (3B10) которое можно получить из гибридомы, обозначенной в Американской коллекции типовых культур номером депонирования PTA-7851.
Эти антитела по настоящему изобретению, 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10, характеризуют как обладающие аффинностью связывания с глобуломером Aβ(20-42), превышающей аффинность связывания этого антитела для глобуломера Aβ(1-42).
Настоящее изобретение также относится к антителам, обладающим таким же профилем связывания, как любое из указанных моноклональных антител, 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10. Антитела, обладающие таким же профилем связывания, как любое из указанных моноклональных антител, не следует рассматривать как ограниченные антителами, обладающими аффинностью связывания с глобуломером Aβ(20-42), превышающей аффинность связывания этого антитела для глобуломера Aβ(1-42).
Антитела, обладающие таким же профилем связывания, как любое из указанных моноклональных антител, 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10, включают в себя антитела, связывающие тот же самый эпитоп, что и моноклональные антитела 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10.
Все моноклональные антитела из группы, состоящей из 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10, связывают эпитоп, содержащийся внутри диапазона последовательности 20 и 42 Aβ, в частности внутри диапазона последовательности 20-30 Aβ. Без связи с теорией, считают, что указанный эпитоп представляет собой структурный, не линейный эпитоп между субъединицами в области аминокислот 20 и 42, в частности в области аминокислот 20 и 30.
Настоящее изобретение также относится к антителам, способным конкурировать по меньшей мере с одним, предпочтительно со всеми, антителами, выбранными из группы, состоящей из 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10.
Термин «конкурирующие антитела» здесь относится к любому числу антител, направленных против той же самой молекулярной или нековалентно связанной надмолекулярной сущности, предпочтительно, той же самой молекулы, где по меньшей мере одно является способным специфически снижать измеряемое связывание другого, предпочтительно, посредством стерических помех для доступа другого антитела к эпитопу, на который оно нацелено, или наведения, и/или стабилизации конформации группы-мишени, уменьшающей аффинность мишени для другого антитела, более предпочтительно, посредством непосредственного блокирования доступа второго антитела к его эпитопу-мишени посредством связывания с эпитопом в достаточно близком соседстве с эпитопом для первого, перекрывающимся с эпитопом для первого или идентичным с эпитопом для первого, наиболее предпочтительно, перекрывающимся или идентичным, особенно, идентичным. Здесь говорят, что два эпитопа являются «перекрывающимися», если они разделяют часть их химической структуры, предпочтительно их аминокислотой последовательности, и являются «идентичными», если их химические структуры, предпочтительно их аминокислотные последовательности, являются идентичными.
Таким образом, настоящее изобретение также относится к антителам, эпитопы-мишени которых являются перекрывающимися, предпочтительно идентичными с эпитопом-мишенью по меньшей мере одного антитела, выбранного из группы, состоящей из 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10.
Антитела, обладающие одинаковым профилем связывания с профилем любого из указанных моноклональных антител, 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10, таким образом, дополнительно включают в себя антитела, содержащие по меньшей мере часть антигенсвязывающей части любого из указанных моноклональных антител, 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10. Предпочтительно, указанная часть содержит по меньшей мере одну определяющую комплементарность область (CDR) любого из указанных моноклональных антител, 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10.
Таким образом, согласно дополнительному конкретному варианту осуществления, настоящее изобретение относится к антителам, содержащим аминокислотную последовательность CDR3 тяжелой цепи и/или аминокислотную последовательность CDR3 легкой цепи моноклонального антитела 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 или 3B10. Конкретные примеры таких антител включают в себя антитела, которые также содержат аминокислотную последовательность CDR2 тяжелой цепи и/или аминокислотную последовательность CDR2 легкой цепи моноклонального антитела 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 или 3B10, соответственно. Еще более конкретно, такие антитела включают в себя антитела, которые также содержат аминокислотную последовательность CDR1 тяжелой цепи и/или аминокислотную последовательность CDR1 легкой цепи моноклонального антитела 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 или 3B10, соответственно.
Таким образом, в одном аспекте настоящее изобретение относится к антителам, содержащим тяжелую цепь, где домен CDR3, CDR2 и/или CDR1 содержит аминокислотную последовательность CDR3, CDR2 и/или CDR1 тяжелой цепи моноклонального антитела 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 или 3B10.
Таким образом, в следующем аспекте настоящее изобретение относится к антителам, содержащим легкую цепь, где домен CDR3, CDR2 и/или CDR1 содержит аминокислотную последовательность CDR3, CDR2 и/или CDR1 легкой цепи, соответственно, моноклонального антитела 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 или 3B10.
Предпочтительно, антитело содержит по меньшей мере одну CDR, содержащую аминокислотную последовательность, выбранную из группы, состоящей из: аминокислотных остатков 31-35 из SEQ ID NO:3, аминокислотных остатков 50-66 из SEQ ID NO:3, аминокислотных остатков 99-109 из SEQ ID NO:3, аминокислотных остатков 24-39 из SEQ ID NO:4, аминокислотных остатков 55-61 из SEQ ID NO:4, аминокислотных остатков 94-102 из SEQ ID NO:4, аминокислотных остатков 31-35 из SEQ ID NO:7, аминокислотных остатков 50-66 из SEQ ID NO:7, аминокислотных остатков 97-109 из SEQ ID NO:7, аминокислотных остатков 24-39 из SEQ ID NO:8, аминокислотных остатков 55-61 из SEQ ID NO:8, аминокислотных остатков 94-102 из SEQ ID NO:8, аминокислотных остатков 31-35 из SEQ ID NO:11, аминокислотных остатков 50-65 из SEQ ID NO:11, аминокислотных остатков 98-107 из SEQ ID NO:11, аминокислотных остатков 24-39 из SEQ ID NO:12, аминокислотных остатков 55-61 из SEQ ID NO:12, аминокислотных остатков 94-102 из SEQ ID NO: 12, аминокислотных остатков 31-35 из SEQ ID NO: 15, аминокислотных остатков 50-66 из SEQ ID NO: 15, аминокислотных остатков 99-107 из SEQ ID NO: 15, аминокислотных остатков 24-40 из SEQ ID NO: 16, аминокислотных остатков 56-62 из SEQ ID NO: 16, аминокислотных остатков 95-103 из SEQ ID NO: 16, аминокислотных остатков 31-35 из SEQ ID NO: 19, аминокислотных остатков 50-66 из SEQ ID NO: 19, аминокислотных остатков 99-109 из SEQ ID NO: 19, аминокислотных остатков 24-39 из SEQ ID NO:20, аминокислотных остатков 55-61 из SEQ ID NO:20, аминокислотных остатков 94-102 из SEQ ID NO:20, аминокислотных остатков 31-35 из SEQ ID NO:23, аминокислотных остатков 50-66 из SEQ ID NO:23, аминокислотных остатков 99-109 из SEQ ID NO:23, аминокислотных остатков 24-39 из SEQ ID NO:24, аминокислотных остатков 55-61 из SEQ ID NO:24, аминокислотных остатков 94-102 из SEQ ID NO:24, аминокислотных остатков 31-35 из SEQ ID NO:27, аминокислотных остатков 50-65 из SEQ ID NO:27, аминокислотных остатков 98-101 из SEQ ID NO:27, аминокислотных остатков 24-39 из SEQ ID NO:28, аминокислотных остатков 55-61 из SEQ ID NO:28, аминокислотных остатков 94-102 из SEQ ID NO:28, аминокислотных остатков 31-35 из SEQ ID NO:31, аминокислотных остатков 50-66 из SEQ ID NO:31, аминокислотных остатков 99-107 из SEQ ID NO:31, аминокислотных остатков 24-40 из SEQ ID NO:32, аминокислотных остатков 56-62 из SEQ ID NO:32, аминокислотных остатков 95-103 из SEQ ID NO:32, аминокислотных остатков 31-35 из SEQ ID NO:35, аминокислотных остатков 50-66 из SEQ ID NO:35, аминокислотных остатков 99-107 из SEQ ID NO:35, аминокислотных остатков 24-40 из SEQ ID NO:36, аминокислотных остатков 56-62 из SEQ ID NO:36, аминокислотных остатков 95-103 из SEQ ID NO:36, аминокислотных остатков 31-35 из SEQ ID NO:38, аминокислотных остатков 50-66 из SEQ ID NO:38, и аминокислотных остатков 98-109 из SEQ ID NO:38.
В предпочтительном варианте осуществления антитело содержит по меньшей мере 3 CDR, выбранные из группы, состоящей из последовательностей, описанных выше. Более предпочтительно, 3 CDR выбраны из наборов CDR вариабельных доменов, выбранных из группы, состоящей из:
Остатки 31-35 из SEQ ID NO:3
Остатки 50-66 из SEQ ID NO:3
Остатки 99-109 из SEQ ID NO:3
Остатки 24-39 из SEQ ID NO:4
Остатки 55-61 из SEQ ID NO:4
Остатки 94-102 из SEQ ID NO:4
Остатки 31-35 из SEQ ID NO:7
Остатки 50-66 из SEQ ID NO:7
Остатки 97-109 из SEQ ID NO:7
Остатки 24-39 из SEQ ID NO:8
Остатки 55-61 из SEQ ID NO:8
Остатки 94-102 из SEQ ID NO:8
Остатки 31-35 из SEQ ID NO:11
Остатки 50-65 из SEQ ID NO:11
Остатки 99-107 из SEQ ID NO:11
Остатки 24-39 из SEQ ID NO:12
Остатки 55-61 из SEQ ID NO:12
Остатки 94-102 из SEQ ID NO:12
Остатки 31-35 из SEQ ID NO:15
Остатки 50-66 из SEQ ID NO:15
Остатки 99-107 из SEQ ID NO:15
Остатки 24-40 из SEQ ID NO:16
Остатки 56-62 из SEQ ID NO:16
Остатки 95-103 из SEQ ID NO:16
Остатки 31-35 из SEQ ID NO:19
Остатки 50-66 из SEQ ID NO:19
Остатки 99-109 из SEQ ID NO:19
Остатки 24-39 из SEQ ID NO:20
Остатки 55-61 из SEQ ID NO:20
Остатки 94-102 из SEQ ID NO:20
Остатки 31-35 из SEQ ID NO:23
Остатки 50-66 из SEQ ID NO:23
Остатки 99-109 из SEQ ID NO:23
Остатки 24-39 из SEQ ID NO:24
Остатки 55-61 из SEQ ID NO:24
Остатки 94-102 из SEQ ID NO:24
Остатки 31-35 из SEQ ID NO:27
Остатки 50-65 из SEQ ID NO:27
Остатки 98-101 из SEQ ID NO:27
Остатки 24-39 из SEQ ID NO:28
Остатки 55-61 из SEQ ID NO:28
Остатки 94-102 из SEQ ID NO:28
Остатки 31-35 из SEQ ID NO:31
Остатки 50-66 из SEQ ID NO:31
Остатки 99-107 из SEQ ID NO:31
Остатки 24-40 из SEQ ID NO:32
Остатки 56-62 из SEQ ID NO:32
Остатки 95-103 из SEQ ID NO:32
Остатки 31-35 из SEQ ID NO:35
Остатки 50-66 из SEQ ID NO:35
Остатки 99-107 из SEQ ID NO:35
Остатки 24-40 из SEQ ID NO:36
Остатки 56-62 из SEQ ID NO:36
Остатки 95-103 из SEQ ID NO:36
Остатки 31-35 из SEQ ID NO:38
Остатки 50-66 из SEQ ID NO:38
Остатки 98-109 из SEQ ID NO:38
В одном варианте осуществления антитело по изобретению содержит по меньшей мере два набора CDR вариабельных доменов. Более предпочтительно, два набора CDR вариабельных доменов выбраны из группы, состоящей из: набора CDR VH 5F7 и набора CDR VL 5F7; набора CDR VH 10F11 и набора CDR VL 10F11; набора CDR VH 7C6 и набора CDR VL 7C6; набора CDR VH 4B7 и набора CDR VL 4B7; набора CDR VH 2F2 и набора CDR VL 2F2; набора CDR VH 6A2 и набора CDR VL 6A2; набора CDR VH 4D10 и набора CDR VL 4D10; набора CDR VH 7E5 и набора CDR VL 7E5; и набора CDR VH 10C1 и набора CDR VL 10C1.
В другом варианте осуществления антитело, описанное выше, содержит человеческую акцепторную каркасную область.
В предпочтительном варианте осуществления антитело представляет собой CDR-привитое антитело. Предпочтительно, CDR-привитое антитело содержит одну или несколько CDR, описанных выше.
Предпочтительно, CDR-привитое антитело содержит человеческую акцепторную каркасную область.
В предпочтительном варианте осуществления антитело представляет собой гуманизированное антитело. Предпочтительно, гуманизированное антитело содержит одну или несколько CDR, описанных выше. Более предпочтительно, гуманизированное антитело содержит три или более CDR, описанных выше. Наиболее предпочтительно, гуманизированное антитело содержит шесть CDR, описанных выше. В конкретном варианте осуществления CDR встроены в вариабельный домен человеческой акцепторной каркасной области антитела человека. Предпочтительно, вариабельный домен антитела человека представляет собой консенсусный вариабельный домен человека. Более предпочтительно, человеческая акцепторная каркасная область содержит по меньшей мере одну замену ключевого аминокислотного остатка каркасной области, где ключевой остаток выбран из группы, состоящей из остатка, соседнего с CDR; остатка участка гликозилирования; редкого остатка; остатка, способного взаимодействовать с глобуломером Aβ(20-42); остатка, способного взаимодействовать с CDR; канонического остатка; остатка из области контакта между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи; остатка внутри зоны Верньера; остатка в области, перекрывающейся между CDR1 вариабельной тяжелой цепи, определенной по Chothia, и первой каркасной области тяжелой цепи, определенной по Kabat. Предпочтительно, человеческая акцепторная каркасная область содержит по меньшей мере одну замену аминокислоты в каркасной области, где аминокислотная последовательность каркаса по меньшей мере на 65% идентична последовательности указанного акцепторного каркаса и содержит по меньшей мере 70 аминокислотных остатков, идентичных указанной человеческой акцепторной каркасной области.
В дополнительном аспекте настоящее изобретение относится к антителам, содержащим как тяжелую, так и легкую цепь, как определено выше.
Предпочтительно, антитело содержит по меньшей мере один вариабельный домен, обладающий аминокислотной последовательностью, выбранной из группы, состоящей из: SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:15, SEQ ID NO:16, SEQ ID NO:19, SEQ ID NO:20, SEQ ID NO:23, SEQ ID NO:24, SEQ ID NO:27, SEQ ID NO:28, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:35, SEQ ID NO:36 и SEQ ID NO:38. Более предпочтительно, антитело содержит два вариабельных домена, где указанные два вариабельных домена обладают аминокислотными последовательностями, выбранными из группы, состоящей из: SEQ ID NO:3 и SEQ ID NO:4, SEQ ID NO:7 и SEQ ID NO:8 и SEQ ID NO:11 и SEQ ID NO:12, SEQ ID NO:15 и SEQ ID NO:16, SEQ ID NO: 19 и SEQ ID NO:20, SEQ ID NO:23 и SEQ ID NO:24, SEQ ID NO:27 и SEQ ID NO:28, SEQ ID NO:31 и SEQ ID NO:32, и SEQ ID NO:35 и SEQ ID NO:36.
В другом аспекте антитела по настоящему изобретению содержат константную область тяжелой цепи, выбранную из группы, состоящей из константных областей IgG1, IgG2, IgG3, IgG4, IgM, IgA, IgD, IgE и мутанта Ala234 Ala235 IgG1 человека. В частности, антитела содержат константную область человека. Более предпочтительно, антитела содержат аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:39-42. Предпочтительными являются антитела, содержащие константную область тяжелой цепи IgG1.
В другом варианте осуществления антитело является гликозилированным. Предпочтительно, характер гликозилирования представляет собой характер гликозилирования человека или характер гликозилирования, получаемый в любой из эукариотических клеток, описанных здесь, в частности в клетках CHO.
Настоящее изобретение относится также к антигенсвязывающей части антитела по настоящему изобретению. Такие антигенсвязывающие части включают в себя в качестве неограничивающих примеров, Fab-фрагменты, F(ab')2-фрагменты и одноцепочечные Fv-фрагменты антитела. Дополнительные антигенсвязывающие части представляют собой Fab'-фрагменты, Fv-фрагменты, и связанные дисульфидом Fv-фрагменты.
Изобретение относится также к выделенной нуклеиновой кислоте, кодирующей любое из антител, описанных здесь. Дополнительный вариант осуществления относится к вектору, содержащему выделенную нуклеиновую кислоту, описанную здесь. Указанный вектор может, в частности, являться выбранным из группы, состоящей из pcDNA; pTT (Durocher et al., Nucleic Acids Research 2002, Vol 30, No.2); pTT3 (pTT с дополнительным полилинкером; pEFBOS (Mizushima, S. and Nagata, S., (1990) Nucleic Acids Research Vol 18, No. 17); pBV; pJV; и pBJ.
В другом аспекте клетку-хозяин трансформируют вектором, описанным здесь. Предпочтительно, клетка-хозяин представляет собой прокариотическую клетку. Более предпочтительно, клетка-хозяин представляет собой E.coli. В родственном варианте осуществления клетка-хозяин представляет собой эукариотическую клетку. Предпочтительно, эукариотическая клетка выбрана из группы, состоящей из клетки протиста, клетки животного (например, клетки млекопитающих, клетки птиц и клетки насекомых), клетки растений и клетки грибов. Более предпочтительно, клетка-хозяин представляет собой клетку млекопитающих, включая, в качестве неограничивающих примеров, CHO и COS; или клетку грибов, например, клетку дрожжей, таких как Saccharomyces cerevisiae; или клетку насекомых, такую как Sf9.
Другой аспект изобретения относится к способу получения антитела по изобретению, включающему в себя культивирование любой из клеток-хозяев или гибридомы, описанной здесь, в культуральной среде в условиях, подходящих для продукции антитела. Другой вариант осуществления относится к антителу, которое можно получить способом, описанным здесь.
Антитела по настоящему изобретению можно получить способом, известным по существу.
B-лимфоциты, которые в целом содержат репертуар антител, составленный из сотен миллиардов различных специфичностей антител, являются частью иммунной системы млекопитающих. Нормальный иммунный ответ на конкретный антиген означает выбор одного или нескольких антител из указанного репертуара, которые специфически связываются с указанным антигеном, и успешный иммунный ответ основан, по меньшей мере частично, на способности указанных антител специфически распознавать (и, в конечном счете, удалять) стимулирующий антиген и игнорировать другие молекулы в окружении указанных антител.
Применимость антител, специфически узнающих один конкретный антиген-мишень, привела к развитию технологии моноклональных антител. Стандартизованный способ гибридом в настоящее время позволяет получение антител с единственной специфичностью к интересующему антигену. Позднее разработаны способы рекомбинантных антител, такие как скрининг библиотек антител in-vitro. Подобным образом, эти способы позволяют получать антитела, обладающие единственной специфичностью к интересующему антигену.
В способе по изобретению интересующему антигену можно позволять воздействовать на репертуар антител либо in vivo, либо in vitro.
Согласно одному варианту осуществления антигену позволяют воздействовать на репертуар посредством иммунизации животного указанным антигеном in vivo. Такой способ in vivo может, кроме того, включать в себя получение ряда гибридом из лимфоцитов животного и отбор конкретной гибридомы, секретирующей антитело, специфически связывающееся с указанным антигеном. Животное, подлежащее иммунизации, может представлять собой, например, мышь, крысу, кролика, курицу, верблюда или овцу или может представлять собой трансгенный вариант любого из упомянутых выше животных, например, трансгенную мышь с генами иммуноглобулина человека, продуцирующее антитела человека после антигенного стимула. Другие типы животных, которых можно иммунизировать, включают в себя мышей с тяжелым комбинированным иммунодефицитом (SCID), который восстанавливали с помощью мононуклеарных клеток периферической крови человека (химерные мыши hu-PBMC SCID) или с помощью лимфоидных клеток или их предшественников, так же как мышей, которых обрабатывали летальной дозой облучения, затем защищали против облучения клетками костного мозга от мышей с тяжелым комбинированным иммунодефицитом (SCID) с последующей трансплантацией функциональных лимфоцитов человека (система «Тримера»). Другой тип животного, подлежащего иммунизации, представляет собой животное (например, мышь), в геноме которого эндогенный ген, кодирующий интересующий антиген, выключают (нокаутируют), например, посредством гомологичной рекомбинации, так что после иммунизации антигеном указанное животное распознает указанный антиген как чужеродный. Специалисту в данной области очевидно, что поликлональные или моноклональные антитела, полученные этим способом, характеризуют и отбирают с использованием известных способов скрининга, которые включают в себя в качестве неограничивающих примеров способы ELISA и дот-блоттинга.
Согласно другому варианту осуществления антигену позволяют воздействовать на репертуар антител in vitro посредством скрининга библиотек рекомбинантных антител с помощью указанного антигена. Рекомбинантную библиотеку антител можно экспрессировать, например, на поверхности бактериофагов, или на поверхности клеток дрожжей, или на поверхности бактериальных клеток. Во множестве вариантов осуществления библиотека рекомбинантных антител представляет собой, например, библиотеку scFv или библиотеку Fab. Согласно другому варианту осуществления библиотеки антител экспрессируют как слитые РНК-белки.
Другой способ получения антител по изобретению включает в себя сочетание подходов in vivo и in vitro. Например, антигену можно позволять воздействовать на репертуар антител посредством иммунизации животного указанным антигеном in vivo с последующим скринингом in vitro с помощью указанного антигена библиотеки рекомбинантных антител, полученной из лимфоидных клеток указанного животного или библиотеки отдельного домена антител (например, содержащей тяжелые и/или легкие цепи). Согласно другому способу антигену позволяют воздействовать на репертуар антител посредством иммунизации животного указанным антигеном in vivo и затем подвергают библиотеку рекомбинантных антител или библиотеку отдельного домена, полученную из лимфоидных клеток указанного животного, аффинному созреванию. Согласно другому способу антигену позволяют воздействовать на репертуар антитела посредством иммунизации животного указанным антигеном in vivo, затем отбора отдельных продуцирующих антитела клеток, секретирующих интересующее антитело, получения из указанных отобранных клеток кДНК для вариабельной области тяжелой и легкой цепей (например, способами ПЦР), и экспрессии указанных вариабельных областей тяжелой и легкой цепей в клетках-хозяевах млекопитающих in vitro (это обозначено здесь способом антитела из отобранного лимфоцита или SLAM), таким образом получая возможность для дальнейшего отбора выбранных последовательностей гена антитела и манипуляций с ними. Более того, моноклональные антитела можно отбирать экспрессионным клонированием посредством экспрессии генов для тяжелых и легких цепей антитела в клетках млекопитающих и отбора клеток млекопитающих, секретирующих антитело, обладающее желаемой аффинностью связывания.
Настоящее изобретение относится к определенным антигенам для скрининга и контрскрининга. Таким образом, является возможным, согласно изобретению, отбирать такие поликлональные и моноклональные антитела, которые связываются с глобуломером Aβ(20-42) с аффинностями связывания, как определено выше.
Способы по изобретению для получения антител можно использовать для получения различных типов антител. Они включают в себя моноклональные, в частности рекомбинантные, антитела, особенно в основном человеческие антитела, химерные антитела, гуманизированные антитела и CDR-привитые антитела, а также их антигенсвязывающие части.
Кроме того, настоящее изобретение относится к гибридоме, способной продуцировать (секретировать) моноклональное антитело по настоящему изобретению. Гибридомы по настоящему изобретению включают в себя гибридомы, обозначенные в Американской коллекции типовых культур номером депонирования, выбранным из группы, состоящей из PTA-7241, PTA-7239, PTA-7240, PTA-7242, PTA-7408, PTA-7409, PTA-7405, PTA-7809, PTA-7810 и PTA-7851.
Отмечено, что антитела по настоящему изобретению могут также являться реакционноспособными, т.е. связываться с формами Aβ, отличными от глобуломеров Aβ, описанных здесь. Эти антигены могут являться или не являться олигомерными или глобуломерными. Таким образом, антигены, с которыми связываются антитела по настоящему изобретению, включают в себя любую форму Aβ, содержащую эпитоп глобуломера, для которого антитела по настоящему изобретению являются реакционно-способными. Такие формы Aβ включают в себя усеченные и не усеченные формы Aβ(X-Y) (с X и Y, определенными, как выше), такие как формы Aβ(20-42), Aβ(20-40), Aβ(12-42), Aβ(12-40), Aβ(1-42) и Aβ(1-40), при условии, что указанные формы содержат эпитоп глобуломера.
Настоящее изобретение также относится к композиции, содержащей антитело по изобретению или его антигенсвязывающую часть, как определено выше.
Согласно конкретному варианту осуществления указанная композиция представляет собой фармацевтическую композицию, содержащую антитело по изобретению или антигенсвязывающую часть и фармацевтически приемлемый носитель.
Антитело по изобретению или антигенсвязывающая часть, как определено выше, предпочтительно является способным нейтрализовывать, как in vitro и in vivo, активность глобуломера Aβ или его производного, с которым он связывается. Указанное антитело или антигенсвязывающую часть, таким образом, можно использовать для ингибирования активности указанного глобуломера или его производного, например, в препарате, содержащем указанный глобуломер или его производное, или у индивидуумов-людей, или у других млекопитающих, у которых присутствует указанный глобуломер или его производное.
Согласно одному из вариантов осуществления изобретение относится к способу ингибирования активности указанного глобуломера, или его производного, где способ включает в себя обеспечение возможности для антитела по изобретению или его антигенсвязывающей части воздействия на глобуломер, или его производное, так что они ингибируют активность указанного глобуломера или его производного. Указанную активность можно ингибировать, например, in vitro. Например, антитело по изобретению или его антигенсвязывающую часть можно добавлять к препарату, такому как образец, полученный из субъекта или культуры клеток, которые содержат, или предположительно содержат, указанный глобуломер или его производное, чтобы ингибировать активность указанного глобуломера или его производного в указанном образце. Альтернативно, активность глобуломера или его производного можно ингибировать у индивидуума in vivo.
Таким образом, кроме того, настоящее изобретение относится к использованию антитела или антигенсвязывающей части, как определено выше, для получения фармацевтической композиции для лечения или предупреждения амилоидоза, в частности амилоидоза, выбранного из группы, состоящей из болезни Альцгеймера и амилоидоза при синдроме Дауна. Одним из аспектов указанного применения по изобретению, таким образом, является способ лечения или предупреждения амилоидоза, в частности болезни Альцгеймера или амилоидоза при синдроме Дауна, у нуждающегося в этом субъекта, который включает в себя введение субъекту антитела или антигенсвязывающей части, как определено выше. Использование указанного антитела или антигенсвязывающей части для лечения и, особенно, предупреждения амилоидоза, в частности при болезнь Альцгеймера или амилоидоза при синдроме Дауна, предназначено, в частности, для пассивной иммунизации. Соответственно, в способе лечения или предупреждения амилоидоза, в частности болезни Альцгеймера, или амилоидоза при синдроме Дауна у нуждающегося в этом субъекта, одной из целей введения антитела или антигенсвязывающей части субъекту является пассивная иммунизация субъекта против амилоидоза, в частности болезни Альцгеймера или амилоидоза при синдроме Дауна.
Антитело по изобретению или антигенсвязывающая часть, как определено выше, предпочтительно, являются пригодными для детекции, как in vitro, так и in vivo, глобуломера Aβ или его производного, с которыми они связываются. Указанное антитело или антигенсвязывающую часть, таким образом, можно использовать для детекции указанного глобуломера или его производного, например, в препарате, содержащем указанный глобуломер или его производное, или у индивидуумов-людей, или других млекопитающих, у которых присутствует указанный глобуломер или его производные.
Согласно одному из вариантов осуществления изобретение относится к способу детекции указанного глобуломера или его производного, где способ включает в себя обеспечение возможности антителу по изобретению или его антигенсвязывающей части воздействовать на глобуломер или его производное, так что они связываются с указанным глобуломером или его производным (и, таким образом, предпочтительно формируют комплекс, содержащий антитело или его антигенсвязывающую часть и глобуломер или его производное). Указанный глобуломер можно детектировать, например, in vitro. Например, антитело по изобретению или антигенсвязывающую часть можно добавлять к препарату, например образцу, полученному из субъекта или культуры клеток, содержащему или предположительно содержащему указанный глобуломер или его производное, для детекции указанного глобуломера или его производного в указанном препарате. Альтернативно, глобуломер или его производное можно детектировать у индивидуума in vivo.
Таким образом, кроме того, настоящее изобретение относится к применению антитела или антигенсвязывающей части, как определено выше, для получения композиции для диагностики амилоидоза, в частности болезни Альцгеймера или амилоидоза при синдроме Дауна. Одним аспектом указанного применения по изобретению является способ диагностики амилоидоза, в частности болезни Альцгеймера или амилоидоза при синдроме Дауна, у субъекта, предположительно, страдающего амилоидозом, в частности болезнью Альцгеймера или амилоидозом при синдроме Дауна, включающий в себя введению субъекту антитела или антигенсвязывающей части, как определено выше, и детекцию формирования комплекса, содержащего антитело или антигенсвязывающую часть с антигеном, где присутствие комплекса указывает на амилоидоз, в частности болезнь Альцгеймера или амилоидоз при синдроме Дауна, у субъекта. Второй аспект указанного применения по изобретению представляет собой способ диагностики амилоидоза, в частности болезни Альцгеймера или амилоидоза при синдроме Дауна, у субъекта, предположительно страдающего амилоидозом, в частности болезнью Альцгеймера или амилоидозом при синдроме Дауна, включающий в себя получение образца от субъекта, контактирование образца с антителом или антигенсвязывающей частью, как определено выше, и детекцию формирования комплекса, содержащего антитело или антигенсвязывающую часть с антигеном, где присутствие комплекса указывает на амилоидоз, в частности болезнь Альцгеймера или амилоидоз при синдроме Дауна, у субъекта.
Подробное описание изобретения
Аффинности связывания антител по изобретению можно оценивать с использованием стандартизованных иммуноанализов in vitro, таких как анализы ELISA, дот-блот или BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden и Piscataway, NJ). Дополнительные описания см. в Jonsson, U., et al. (1993) Ann. Biol. Clin. 51:19-26; Jonsson, U., et al. (1991) Biotechniques 11:620-627; Johnsson, B., et al. (1995) J. Mol. Recognit. 8:125-131; и Johnsson, B., et al. (1991) Anal. Biochem. 198:268-277.
Согласно конкретному варианту осуществления аффинности, определяемые здесь, относятся к значениям, полученным проведением дот-блоттинга, как описано в примере 8, и оценкой его посредством денситометрии. Согласно конкретному варианту осуществления по изобретению определение аффинности связывания посредством дот-блоттинга включает в себя следующее: конкретное количество антигена (например, глобуломера Aβ(X-Y), мономера Aβ(X-Y) или фибрилл Aβ(X-Y), как определено выше) или, целесообразно, их соответствующего разведения, например, в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, 0,2 мг/мл BSA до концентрации антигена, например, 100 пмоль/мкл, 10 пмоль/мкл, 1 пмоль/мкл, 0,1 пмоль/мкл и 0,01 пмоль/мкл, наносят точками на нитроцеллюлозную мембрану, затем мембрану блокируют молоком для предупреждения неспецифического связывания и промывают, затем приводят в контакт с интересующим антителом с последующей детекцией последнего посредством конъюгированного с ферментом вторичного антитела и колориметрической реакции; при определенных концентрациях антитела количество связавшегося антитела позволяет определение аффинности. Таким образом, относительную аффинность двух различных антител для одной мишени или одного антитела для двух различных мишеней определяют здесь как отношение соответствующих количеств связанного с мишенью антитела, наблюдаемое для двух сочетаний антитело-мишень при в остальном идентичных условиях дот-блоттинга. В отличие от похожего способа, основанного на Вестерн-блоттинге, по способу дот-блоттинга будут определять аффинность антитела для данной мишени при естественной конформации последней; в отличие от способа ELISA способ дот-блоттинга не страдает от различий в аффинностях между различными мишенями и матрицей, таким образом, позволяя более точные сравнения между различными мишенями.
Термин «Kd», как применяют здесь, предназначен для обозначения константы диссоциации для конкретного взаимодействия антитело-антиген, как известно в данной области.
Антитела по настоящему изобретению предпочтительно представляют собой выделенные антитела. «Выделенное антитело» означает антитело, обладающее аффинностями связывания, как описано выше, и в основном свободное от других антител, обладающих отличными аффинностями связывания. Термин «в основном свободный» здесь относится к препарату антитела, в котором по меньшей мере 95% антител, предпочтительно, по меньшей мере 98% антител и, более предпочтительно, по меньшей мере 99% антител обладают желательной аффинностью связывания. Более того, выделенное антитело может являться по существу свободным от другого клеточного материала и/или химических веществ.
Выделенные антитела по настоящему изобретению включают в себя моноклональные антитела. «Моноклональное антитело», как применяют здесь, предназначено для обозначения препарата молекул антитела, антитела в котором разделяют аминокислотную последовательность общей тяжелой цепи и общей легкой цепи, в отличие от препаратов «поликлонального» антитела, содержащих смесь антител различной аминокислотной последовательности. Моноклональные антитела можно получать посредством нескольких новых способов, таких как фаговый, бактериальный, дрожжевой или рибосомный дисплей, так же как классическими способами, проиллюстрированными полученными из гибридомы антителами (например, антителом, секретируемым гибридомой, полученной посредством способа гибридом, таким как общепринятый способ гибридом Kohler и Milstein ((1975) Nature 256:495-497). Таким образом, антитело, полученное не из гибридомы, с однородной последовательностью, еще обозначают здесь как моноклональное антитело, хотя его можно получить посредством неклассических способов, и термин «моноклональные» не ограничивают полученными из гибридомы антителами, но используют как относящийся ко всем антителам, полученным из одного клона нуклеиновой кислоты.
Таким образом, моноклональные антитела по настоящему изобретению включают в себя рекомбинантные антитела. Термин «рекомбинантный» здесь относится к любому искусственному объединению двух иначе разделенных фрагментов последовательности, например, посредством химического синтеза или манипуляций с выделенными фрагментами нуклеиновых кислот способами генетической инженерии. В частности, термин «рекомбинантное антитело» относится к антителам, которые продуцируют, экспрессируют, получают или выделяют рекомбинантными способами, таким как антитела, которые экспрессируют с использованием рекомбинантного экспрессирующего вектора, трансфицированного в клетку-хозяина; антителам, выделенным из рекомбинантной комбинаторной библиотеки антител; антителам, выделенным из животного (например, мыши), которое является трансгенным по генам иммуноглобулина человека (см., например, Taylor, L.D., et al. (1992) Nucl. Acids Res. 20:6287-6295); или антителам, которые продуцируют, экспрессируют, получают или выделяют любым другим способом, в котором конкретные последовательности генов иммуноглобулинов (такие как последовательности генов иммуноглобулинов человека) объединяют с другими последовательностями ДНК. Рекомбинантные антитела включают в себя, например, химерные, CDR-привитые и гуманизированные антитела. Специалисты в данной области сознают, что для экспрессии моноклонального антитела, полученного из общепринятых гибридом, в гетерологичной системе необходимо получение рекомбинантного антитела, даже если аминокислотную последовательность полученного белка-антитела не изменяют или не предполагают изменять.
В конкретном варианте осуществления изобретения антитело представляет собой гуманизированное антитело.
Согласно множеству вариантов осуществления антитело может содержать аминокислотную последовательность, полностью полученную из отдельных видов, таких как антитело человека или антитело мыши. Согласно другим вариантам осуществления, антитело может представлять собой химерное антитело, или CDR-привитое антитело, или другую форму гуманизированного антитела.
Термин «антитело» предназначен для обозначения молекул иммуноглобулина, состоящих из 4 полипептидных цепей, двух тяжелых (H) цепей и двух легких (L) цепей. Цепи обычно связаны друг с другом посредством дисульфидных связей. Каждая тяжелая цепь состоит из вариабельной области указанной тяжелой цепи (обозначенной здесь как HCVR или VH) и константной области указанной тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области указанной легкой цепи (обозначенной здесь как LCVR или VL) и константной области указанной легкой цепи. Константная область легкой цепи состоит из домена CL. Области VH и VL можно далее разделить на гипервариабельные области, обозначенные определяющими комплементарность областями (CDR) и перемежающиеся консервативными областями, обозначенными каркасными областями (FR). Каждая VH и VL область, таким образом, состоит из трех CDR и четырех FR, расположенных от N-конца к C-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Эта структура хорошо известна специалистам в данной области.
Термин «антигенсвязывающая часть» антитела (или просто «часть антитела») относится к одному или нескольким фрагментам антитела по изобретению, где указанный фрагмент(ы) еще обладает аффинностями связывания, как определено выше. Показано, что фрагменты полного антитела способны выполнять антигенсвязывающую функцию антитела. В соответствии с термином «антигенсвязывающая часть» антитела, примеры связывающих фрагментов включают в себя (i) Fab-фрагмент, т.е. моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2 фрагмент, т.е. бивалентный фрагмент, содержащий два Fab-фрагмента, связанные друг с другом в шарнирной области посредством дисульфидного мостика; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов FL и VH одного плеча антитела; (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), состоящий из домена VH или из VH, CH1, CH2, DH3, или VH, CH2, CH3; и (vi) выделенную определяющую комплементарность область (CDR). Хотя два домена Fv-фрагмента, а именно VL и VH, кодированы отдельными генами, их можно дополнительно связать друг с другом с использованием синтетического линкера, например аминокислотной последовательности поли-G4S, и рекомбинантными способами, позволяющими получать их как одну белковую цепь, в которой области VL и VH объединяют для формирования моновалентных молекул (известных как одноцепочечные Fv (ScFv); см., например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Термин «антигенсвязывающая часть» антитела также предназначен, чтобы включать в себя такие одноцепочечные антитела. Другие формы одноцепочечных антител, такие как «диатела» подобным образом включены сюда. Диатела представляют собой бивалентные, биспецифические антитела, в которых домены VH и VL экспрессированы на одной полипептидной цепи, но с использованием линкера, который является слишком коротким, чтобы позволять спаривание между двумя доменами на одной и той же цепи, таким образом заставляя домены спариваться с комплементарными доменами на другой цепи и создавая два антигенсвязывающих участка (см., например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak, R.J., et al. (1994) Structure 2:1121-1123). Константный домен иммуноглобулина относится к константному домену тяжелой или легкой цепи. Аминокислотные последовательности константного домена тяжелой цепи и легкой цепи IgG человека известны в данной области и представлены в таблице 1.

Более того, антитело по настоящему изобретению или его антигенсвязывающая часть может являться частью более крупной молекулы иммуноадгезии, сформированной посредством ковалентного или нековалентного объединения указанного антитела или части антитела с одним или несколькими дополнительными белками или пептидами. Подходящими для таких молекул иммуноадгезии являются использование стрептавидиновой коровой области для получения тетрамерной молекулы scFv (Kipriyanov, S. M., et al. (1995) Human Antibodies and Hybri- domas 6:93-101) и использование остатка цистеина, маркерного пептида и C-концевой полигистидиниловой, например гексагистидиниловой, метки для получения бивалентных и биотинилированных молекул scFv (Kipriyanov, S.M., et al. (1994) Mol. Immunol. 31:1047-1058).
Термин «антитело человека» относится к антителам, вариабельные и константные области которых соответствуют последовательностям иммуноглобулина зародышевой линии человека или получены из них, как описано, например, Kabat et al. (см. Kabat, et al. (1991) Sequences of Protein of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242). Однако антитела человека по изобретению могут содержать аминокислотные остатки, не кодируемые последовательностями иммуноглобулина зародышевой линии человека (например, мутации, введенные посредством случайного или сайт-специфического мутагенеза in vitro или посредством соматической мутации in vivo), например в CDR, и, в частности, в CDR3. Рекомбинантные антитела человека по изобретению обладают вариабельными областями и могут также содержать константные области, полученные из иммуноглобулиновых последовательностей зародышевой линии человека (см. Kabat, E. A., et al. (1991) Sequences of Protein of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242). Однако, согласно конкретным вариантам осуществления, такие рекомбинантные антитела человека подвергают мутагенезу in-vitro (или соматическому мутагенезу in-vivo, если используют животное, трансгенное по последовательностям Ig человека), так что аминокислотные последовательности областей VH и VL рекомбинантных антител представляют собой последовательности, которые, хотя относятся к последовательностям VH и VL из зародышевой линии человека или получены из них, не существуют в природе in vivo в репертуаре антител зародышевой линии человека. Согласно конкретным вариантам осуществления рекомбинантные антитела этого вида представляют собой результат избирательного мутагенеза, или обратной мутации, или того и другого. Предпочтительно, мутагенез приводит к аффинности к мишени, которая превышает аффинность исходного антитела, и/или к аффинности к не относящимся к мишени структурам, которая ниже аффинности исходного антитела.
Термин «химерное антитело» относится к антителам, содержащим последовательности вариабельных областей тяжелых и легких цепей из одних видов и последовательности константных областей из других видов, таким как антитела, обладающие вариабельными областями тяжелой и легкой цепи мыши, связанными с константными областями человека.
Термин «CDR-привитое антитело» относится к антителам, содержащим последовательности вариабельных областей тяжелых и легких цепей из одних видов, но в которых последовательности одной или нескольких областей CDR VH и/или VL заменены последовательностями CDR из других видов, таким как антитела, обладающие вариабельными областями тяжелых и легких цепей, в которых одна или несколько CDR мыши (например, CDR3) заменены последовательностями CDR человека.
Термин «гуманизированное антитело» относится к антителам, содержащим последовательности вариабельных областей тяжелых и легких цепей из не относящихся к человеку видов (например, мыши, крысы, кролика, курицы, верблюда, овцы или козы), в которых, однако, по меньшей мере одну часть последовательности VH и/или VL изменяют, чтобы они являлись более «подобными человеческим», т.е. являлись более сходными с вариабельными последовательностями зародышевой линии человека. Одним типом гуманизированного антитела является CDR-привитое антитело, в котором последовательности CDR человека вставлены в не относящиеся к человеку последовательности VH и VL для замены соответствующих не относящихся к человеку последовательностей CDR.
Термины «нумерация Kabat», «определения Kabat» и «маркировка Kabat» используют здесь как взаимозаменяемые. Эти термины, которые известны в данной области, относятся к системе нумерации аминокислотных остатков, которые являются более вариабельными (т.е. гипервариабельными), по сравнению с другими аминокислотными остатками в вариабельных областях тяжелых и легких цепей антитела или его антигенсвязывающей части (Kabat et al. (1971) Ann. NY Acad, Sci 190:382-391 и Kabat, E. A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242). В случае вариабельной области тяжелой цепи гипервариабельная область лежит в диапазоне положений аминокислот от 31 до 35 для CDR1, положений аминокислот от 50 до 65 для CDR2 и положений аминокислот от 95 до 102 для CDR3. В случае вариабельной области тяжелой цепи гипервариабельная область лежит в диапазоне положений аминокислот от 24 до 34 для CDR1, положений аминокислот от 50 до 56 для CDR2 и положений аминокислот от 89 до 97 для CDR3.
Как применяют здесь, термины «акцептор» и «акцепторное антитело» относятся к последовательности антитела или нуклеиновой кислоты, представляющим или кодирующим по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100% аминокислотных последовательностей одной или нескольких каркасных областей. В некоторых вариантах осуществления термин «акцептор» относится к аминокислотной последовательности антитела или последовательности нуклеиновой кислоты, представляющих или кодирующих константную область(области). В другом варианте осуществления термин «акцептор» относится к аминокислотой последовательности антитела или последовательности нуклеиновой кислоты, представляющей или кодирующей одну или несколько каркасных областей и константную область(области). В конкретном варианте осуществления термин «акцептор» относится к аминокислотной последовательности антитела человека или последовательности нуклеиновой кислоты, представляющей или кодирующей по меньшей мере 80%, предпочтительно, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100% аминокислотных последовательностей одной или нескольких каркасных областей. Согласно этому варианту осуществления акцептор может содержать по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5 или по меньшей мере 10 аминокислотных остатков, не присутствующих в одном или нескольких конкретных положениях антитела человека. Акцепторную каркасную область и/или акцепторную константную область(области) можно, например, вывести или получить из зародышевого гена антитела, зрелого гена антитела, функционального антитела (например, антител, хорошо известных в данной области, разрабатываемых антител или коммерчески доступных антител).
Как применяют здесь, термин «CDR» относится к определяющей комплементарность области внутри вариабельных последовательностей антитела. Существуют три CDR в каждой из вариабельных областей тяжелой цепи и легкой цепи, обозначенные CDR1, CDR2 и CDR3 для каждой вариабельной области. Термин «набор CDR», как применяют здесь, относится к группе из трех CDR, присутствующих в одной вариабельной области, способной связывать антиген. Точные границы этих CDR определяют различно согласно различным системам. Система, описанная Kabat (Kabat et al., Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987) и (1991)) не только предоставляет однозначную систему нумерации остатков, применимую к любой вариабельной области антитела, но также предоставляет точные границы остатков, определяющих три CDR. Эти CDR можно обозначать CDR Kabat. Chothia и соавторы (Chothia & Lesk, J. Mol. Biol. 196:901-917 (1987) и Chothia et al., Nature 342:877-883 (1989)) обнаружили, что конкретные субфрагменты внутри CDR Kabat принимают почти идентичные конформации пептидного остова, несмотря на большое разнообразие на уровне аминокислотной последовательности. Эти субфрагменты обозначают L1, L2 и L3 или H1, H2 и H3, где «L» и «H» обозначают области легкой цепи и тяжелых цепей соответственно. Эти области можно обозначать CDR Chothia, которые облают границами, перекрывающимися с CDR Kabat. Другие границы, определяющие CDR, перекрывающиеся с CDR Kabat, определены Padlan (FASEB J. 9:133-139 (1995)) и MacCallum (J. Mol. Biol. 262(5):732-45 (1996)). Другие определения границ CDR могут не следовать строго одной из вышеуказанных систем, однако, тем не менее, будет перекрываться с CDR Kabat, хотя они могут являться укороченными или удлиненными в свете предсказания или экспериментального обнаружения того, что конкретные остатки или группы остатков или даже целые CDR не вносят значительного вклада в связывание антигена. В способах, применяемых здесь, можно использовать CDR, определяемые согласно любой из этих систем, хотя в предпочтительных вариантах осуществления используют CDR, определенные по Kabat или Chothia.
Как применяют здесь, термин «канонический» остаток относится к остатку в CDR или каркасе, который определяет конкретную каноническую структуру CDR, как определено Chothia et al. (J. Mol. Biol. 196:901-907 (1987); Chothia et al., J. Mol. Biol. 227:799 (1992), оба приведены здесь в качестве ссылок). Согласно Chothia et al. критические части CDR многих антител обладают почти идентичными конформациями пептидного остова несмотря на большое разнообразие на уровне аминокислотной последовательности. Каждая каноническая структура задает в первую очередь набор торсионных углов пептидного остова для непрерывного фрагмента аминокислотных остатков, формирующих петлю.
Как применяют здесь, термины «донор» и «донорное антитело» относятся к антителу, предоставляющему одну или несколько CDR. В предпочтительном варианте осуществления донорное антитело представляет собой антитело из видов, отличных от вида для антитела, из которого получены или выведены каркасные области. В контексте гуманизированного антитела термин «донорное антитело» относится к не относящемуся к человеку антителу, предоставляющему одну или несколько CDR.
Как применяют здесь, термин «каркас» или «каркасная последовательность» относится к оставшимся последовательностям вариабельной области минус CDR. Поскольку точное задание границ последовательности CDR можно определить с использованием различных систем, значение каркасной последовательности является субъектом соответственно различных интерпретаций. Шесть CDR (CDR-L1, -L2 и -L3 из легкой цепи и CDR-H1, -H2, и -H3 из тяжелой цепи) также разделяют каркасные области легкой цепи и тяжелой цепи на четыре субфрагмента (FR1, FR2, FR3 и FR4) на каждой цепи, в которых CDR1 расположен между FR1 и FR2, CDR2 между FR2 и FR3, и CDR3 между FR3 и FR4. Без определения конкретных субфрагментов как FR1, FR2, FR3 или FR4 каркасная область, как определяют другие, представляет собой объединенные FR внутри вариабельной области отдельной, существующей в природе цепи иммуноглобулина. Как применяют здесь, FR представляет собой один из четырех субфрагментов, а FR во множественном числе представляет собой два или более из четырех субфрагментов, составляющих каркасную область.
Акцепторные последовательности тяжелой цепи и легкой цепи человека известны в данной области.
Как применяют здесь, термин «зародышевый ген антитела» или «фрагмент гена» относится к последовательности иммуноглобулина, кодируемой нелимфоидными клетками, которые не подвергались процессу созревания, приводящему к генетической реаранжировке и мутации для экспрессии конкретного иммуноглобулина (см., например, Shapiro et al., Crit. Rev. Immunol. 22(3): 183-200 (2002); Marchаlоnis et al., Adv Exp Med Biol. 484:13-30 (2001)). Одно из преимуществ, предоставленных различными вариантами осуществления настоящего изобретения, вытекает из обнаружения, что зародышевые гены антитела более вероятно, чем зрелые гены антитела, сохраняют необходимые структуры аминокислотной последовательности, характерные для индивидуумов в видах, таким образом, менее вероятно, узнаваемые как не свое при использовании в этих видах.
Как применяют здесь, термин «ключевые» остатки относится к конкретным остаткам внутри вариабельной области, которые вносят больший вклад в специфичность связывания и/или аффинность антитела, в частности гуманизированного антитела. Ключевые остатки включают в себя, без ограничения, одно или несколько из следующего: остатка, соседнего с CDR, потенциального участка гликозилирования (который может представлять собой участок либо N-, либо O-гликозилирования), редкого остатка, остатка, способного взаимодействовать с антигеном, остатка, способного взаимодействовать с CDR, канонического остатка, остатка из области контакта между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, остатка внутри зоны Верньера и остатка в области перекрывания между определенной по Chothia CDR1 вариабельной тяжелой цепи и определенного по Kabat каркаса первой тяжелой цепи.
Как применяют здесь, термин «гуманизированное антитело» конкретно относится к антителу или его варианту, производному, аналогу или фрагменту, которое иммуноспецифически связывается с интересующим антигеном и которое содержит каркасную (FR) область, обладающую в основном аминокислотной последовательностью антитела человека и определяющую комплементарность область (CDR), обладающую в основном аминокислотной последовательностью не относящегося к человеку антитела. Как применяют здесь, термин «в основном» в контексте CDR относится к CDR, обладающей аминокислотной последовательностью, по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98% или по меньшей мере на 99% идентичной аминокислотной последовательности CDR не относящегося к человеку антитела. Гуманизированное антитело содержит в основном все из по меньшей мере одного и, как правило, двух, вариабельных доменов (Fab, Fab', F(ab')2, FabC, Fv), в которых все или в основном все из областей CDR соответствуют областям не относящегося к человеку иммуноглобулина (т.е. донорного антитела) и все или по существу все из каркасных областей представляют собой области с консенсусной последовательностью иммуноглобулина человека. Предпочтительно, гуманизированное антитело содержит также по меньшей мере часть константной области (Fc) иммуноглобулина, как правило, области иммуноглобулина человека. В некоторых вариантах осуществления гуманизированное антитело содержит как легкую цепь, так и по меньшей мере вариабельный домен тяжелой цепи. Антитело может содержать также CH1, шарнирную, CH2, CH3, и CH4 области тяжелой цепи. В некоторых вариантах осуществления гуманизированное антитело содержит только гуманизированную легкую цепь. В некоторых вариантах осуществления гуманизированное антитело содержит только гуманизированную тяжелую цепь. В конкретных вариантах осуществления гуманизированное антитело содержит только гуманизированный вариабельный домен легкой цепи и/или гуманизированную тяжелую цепь.
Гуманизированное антитело можно выбирать из любого класса иммуноглобулинов, включая IgM, IgG, IgD, IgA и IgE, и любого подкласса, включая без ограничения IgG1, IgG2, IgG3 и IgG4.
Каркасные и CDR области гуманизированного антитела не обязательно точно соответствуют исходным последовательностям, например, CDR или консенсусный каркас донорного антитела можно подвергать мутагенезу посредством замены, вставки и/или делеции по меньшей мере одного аминокислотного остатка, так что остаток CDR или каркаса в этом участке не соответствует точно ни донорному антителу, ни консенсусному каркасу. В предпочтительном варианте осуществления такие мутации, однако, не будут обширными. Обычно по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90%, и наиболее предпочтительно по меньшей мере 95% остатков гуманизированного антитела будет соответствовать остаткам исходных последовательностей FR и CDR. Как применяют здесь, термин «консенсусный каркас» относится к каркасной области консенсусной последовательности иммуноглобулина. Как применяют здесь, термин «консенсусная последовательность иммуноглобулина» относится к последовательности, сформированной из наиболее часто встречающихся аминокислот (или нуклеотидов) в семействе родственных последовательностей иммуноглобулинов (см., например, Winnaker, From Genes to Clones (Verlagsgesellschaft, Weinheim, Germany 1987). В семействе иммуноглобулинов каждое положение в консенсусной последовательности занято аминокислотой, наиболее часто встречающейся в этом положении в семействе. Когда две аминокислоты встречаются одинаково часто, любую можно включать в консенсусную последовательность.
Как применяют здесь, зона «Верньера» обозначает подгруппу каркасных остатков, которые могут регулировать структуру CDR и осуществлять тонкую настройку для соответствия антигену, как описано Foote and Winter (1992, J. Mol. Biol. 224:487-499, содержание которой приведено здесь в качестве ссылки). Остатки зоны Верньера формируют слой, подстилающий CDR, и могут вносить вклад в структуру CDR и аффинность антитела.
Термин «эпитоп» включает в себя любую полипептидную детерминанту, способную специфически связываться с иммуноглобулином. В конкретных вариантах осуществления детерминанты эпитопа включают в себя химически активные поверхностные группировки таких молекул, как аминокислоты, боковые цепи сахаров, фосфорил или сульфонил, и, в конкретных вариантах осуществления, могут обладать конкретными трехмерными структурными характеристиками и/или конкретными характеристиками заряда. Эпитоп представляет собой область антигена, связываемую антителом. В конкретных вариантах осуществления указывают, что антитело специфически связывается с антигеном, когда оно преимущественно распознает свой антиген-мишень в сложной смеси белков и/или макромолекул.
Термин «полинуклеотид», как обозначают здесь, обозначает полимерную форму двух или более нуклеотидов, либо рибонуклеотидов, либо дезоксинуклеотиды, либо модифицированной формы любого типа нуклеотидов. Термин включает в себя одно- и двухцепочечные формы ДНК, но предпочтительно ДНК является двухцепочечной ДНК.
Термин «выделенный полинуклеотид», как применяют здесь, должен обозначать полинуклеотид (например, геномного, кДНК, или синтетического происхождения, или любое их сочетание), такой, что, в силу его происхождения, «выделенный полинуклеотид» не является связанным со всем или частью полинуклеотида, с которым «выделенный полинуклеотид» обнаружен в природе; является функционально связанным с полинуклеотидом, с которым он не связан в природе; или не встречается в природе как часть большей последовательности.
Термин «вектор», как применяют здесь, предназначен для обозначения молекулы нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она связана. Одним типом вектора является «плазмида», которая обозначает кольцевую петлю двухцепочечной ДНК, в которую можно лигировать дополнительные фрагменты ДНК. Другим типом вектора является вирусный вектор, где дополнительные фрагменты ДНК можно лигировать в вирусный геном. Конкретные векторы являются способными к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, обладающие бактериальной точкой начала репликации и эписомальные векторы млекопитающих). Другие векторы (например, не эписомальные векторы млекопитающих) можно интегрировать в геном клетки-хозяина при введении в клетку-хозяина, и таким образом они реплицируются вместе с геномом хозяина. Кроме того, конкретные векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы обозначены здесь «рекомбинантные экспрессирующие векторы» (или просто «экспрессирующие векторы»). Как правило, экспрессирующие векторы для использования в способах рекомбинантной ДНК часто присутствуют в форме плазмид. В настоящем описании «плазмиду» и «вектор» можно использовать как взаимозаменяемые, так как плазмида является наиболее общепринятой формой вектора. Однако изобретение предназначено, чтобы включать в себя другие такие формы экспрессирующих векторов, такие как вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), выполняющие эквивалентные функции.
Термин «функционально связанный» обозначает непосредственное соседство, где описанные компоненты находятся во взаимосвязи, позволяющей им функционировать предназначенным для них способом. Контрольная последовательность, «функционально связанная» с кодирующей последовательностью, присоединена таким образом, что экспрессию кодирующей последовательности получают в условиях, соответствующих контрольным последовательностям. «Функционально связанные» последовательности включают в себя как контролирующие экспрессию последовательности, смежные с интересующим геном, так и контролирующие экспрессию последовательности, которые для контроля интересующего гена действуют в трансположении или на расстоянии. Термин «контролирующая экспрессию последовательность», как применяют здесь, обозначает полинуклеотидные последовательности, необходимые для воздействия на экспрессию и процессинг кодирующих последовательностей, с которыми они лигированы. Контролирующие экспрессию последовательности включают в себя подходящие последовательности инициации транскрипции, терминации, промотор и энхансер; сигналы эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, стабилизирующие цитоплазматическую мРНК; последовательности, увеличивающие эффективность трансляции (т.е. консенсусная последовательность Козака); последовательности, увеличивающие стабильность белка; и, когда желательно, последовательности, усиливающие секрецию белка. Природа таких контрольных последовательностей отличается в зависимости от организма-хозяина; для прокариот такие контрольные последовательности, как правило, включают в себя промотор, участок связывания рибосомы и последовательность терминации транскрипции; для эукариот, как правило, такие контрольные последовательности включают в себя промотор и последовательность терминации транскрипции. Термин «контрольные последовательности» предназначен, чтобы включать в себя компоненты, присутствие которых является необходимым для экспрессии и процессинга, и может также включать в себя дополнительные компоненты, присутствие которых является выгодным, например, лидирующие последовательности и последовательности партнера по слиянию.
«Трансформация», как определено здесь, обозначает любой способ, посредством которого экзогенная ДНК входит в клетку-хозяина. Трансформацию можно осуществлять в естественных или искусственных условиях с использованием различных способов, хорошо известных в данной области. Трансформация может являться основанной на любом известном способе введения последовательностей чужеродной нуклеиновой кислоты в прокариотическую или эукариотическую клетку-хозяина. Способ выбирают на основании подлежащей трансформации клетки-хозяина, и он может включать в себя в качестве неограничивающих примеров вирусную инфекцию, электропорацию, липофекцию и бомбардировку частицами. Такие «трансформированные» клетки включают в себя стабильно трансформированные клетки, в которых вставленная ДНК способна реплицироваться либо как автономно реплицирующаяся плазмида, либо как часть хромосомы хозяина. Они включают в себя также клетки, временно экспрессирующие вставленную ДНК или РНК в течение ограниченных периодов времени.
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин»), как применяют здесь, предназначен для обозначения клетки, в которую введена экзогенная ДНК. Следует понимать, что такие термины предназначены для обозначения не только конкретной рассматриваемой клетки, но и потомства такой клетки. Поскольку в последующих поколениях могут присутствовать определенные модификации, в результате либо мутаций, либо воздействия внешней среды, такое потомство фактически может не являться идентичным родительской клетке, однако еще входит в объем термина «клетка-хозяин», как применяют здесь. Предпочтительно, клетки-хозяева включают в себя прокариотические и эукариотические клетки, выбранные из любого из царств живых организмов. Предпочтительные эукариотические клетки включают в себя клетки протистов, грибов, растений и животных. Наиболее предпочтительно, клетки-хозяева включают в себя в качестве неограничивающих примеров линию прокариотических клеток E.coli; линии клеток млекопитающих CHO, HEK 293 и COS; линию клеток насекомых Sf9; и клетки грибов Saccharomyces cerevisiae.
Общепринятые способы можно использовать для рекомбинантной ДНК, синтеза олигонуклеотидов, культивирования клеток и трансформации (например, электропорации, липофекции). Ферментативные реакции и способы очистки можно осуществлять согласно инструкциям производителя, или как является общепринятым в данной области, или как описано здесь. Вышеупомянутые методы и способы можно в основном осуществлять согласно общепринятым способам, хорошо известным в данной области, и как описано в различных общих и более конкретных ссылках, которые цитируют и обсуждают на всем протяжении настоящего описания. См., например, Sambrook et al. Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)), приведенный здесь в качестве ссылки для всех целей.
«Трансгенный организм», как известно в данной области и как применяют здесь, обозначает организм, обладающий клетками, содержащими трансген, где с трансгена, введенного в организм (или в предка организма), экспрессируют полипептид, естественным образом не экспрессирующийся в организме. «Трансген» представляет собой конструкцию ДНК, которая является стабильно и функционально интегрированной в геном клетки, из которой развивается трансгенный организм, управляя экспрессией закодированного продукта гена в одном или нескольких типах клеток или тканей трансгенного организма.
Способы получения антител по изобретению описаны ниже. Проведено разделение между способами in-vivo, способами in-vitro или сочетанием обоих.
Некоторые способы получения антител по изобретению описаны ниже. Проведено разделение между способами in-vivo, способами in-vitro или сочетанием обоих.
Способы in-vivo
В зависимости от типа желаемого антитела, различных животных-хозяев можно использовать для иммунизации in-vivo. Можно использовать хозяина, который сам экспрессирует эндогенный вариант интересующего антигена. Альтернативно, можно использовать хозяина, дефицитного по эндогенному варианту интересующего антигена. Например, показано, что мыши, сделанные дефицитными по конкретному эндогенному белку посредством гомологичной рекомбинации в соответствующем эндогенном гене (т.е. нокаутированные мыши) производят гуморальный ответ на белок, которым их иммунизируют и таким образом их можно использовать для получения высокоаффинных моноклональных антител к белку (см., например, Roes, J. et al. (1995) J. Immunol. Methods 183:231-237; Lunn, MP. et al. (2000) J. Neurochem. 75:404-412).
Множество не относящихся к человеку млекопитающих являются подходящими хозяевами для продукции антител для получения не относящихся к человеку антител по изобретению. Они включают в себя мышей, крыс, кур, верблюдов, кроликов, овец и коз (и их нокаутированные варианты), хотя предпочтение отдают мышам для получения гибридом. Кроме того, не относящегося к человеку животного-хозяина, экспрессирующего репертуар антител человека, можно использовать для получения необходимых антител человека с двойной специфичностью. Не относящиеся к человеку животные этого вида включают в себя трансгенных животных (например, мышей), несущих трансгены иммуноглобулинов человека (химерные мыши hu-PBMC SCID) и облученные химеры человек/мышь, более подробно описанные ниже. Согласно одному варианту осуществления, животное, иммунизированное глобуломером Aβ(20-42) или его производным, представляет собой не относящееся к человеку животное, предпочтительно мышь, которое является трансгенным по генам иммуноглобулинов человека, так что указанное не относящееся к человеку млекопитающее производит антитела человека при антигенной стимуляции. Как правило, трансгены иммуноглобулинов для тяжелых и легких цепей с конфигурацией зародышевой линии человека вводят таким животным, которые являются измененными так, что их эндогенные локусы тяжелых и легких цепей являются неактивными. Если таких животных стимулируют антигеном (например, антигеном человека), они продуцируют антитела, полученные из последовательностей иммуноглобулина человека (антитела человека). Можно получить из лимфоцитов таких животных моноклональные антитела человека посредством стандартизованного способа гибридомы. Дополнительное описание трансгенных мышей с иммуноглобулинами и их использование для получения антител человека, см., например, в US 5939598, WO 96/33735, WO 96/34096, WO 98/24893 и WO 99/53049 (Abgenix Inc.), и US 5545806, US 5569825, US 5625126, US 5633425, US 5661016, US 5770429, US 5814318, US 5877397 и WO 99/45962 (Genpharm Inc.); см. также MacQuitty, J.J. and Kay, R.M. (1992) Science 257:1188; Taylor, L.D. et al. (1992) Nucleic Acid Res. 20:6287-6295; Lonberg, N. et al. (1994) Nature 368:856-859; Lonberg, N. and Huszar, D. (1995) Int. Rev. Immunol. 13:65-93; Harding, F.A. and Lonberg, N. (1995) Ann. N Y. Acad. Sci. 764:536-546; Fishwild, D. M. et al. (1996) Nature Biotechnology 14:845-851; Mendez, M. J. et al. (1997) Nature Genetics 15:146-156; Green, L.L. and Jakobovits, A. (1998; J. Exp. Med. 188:483-495; Green, L.L. (1999) J. Immunol. Methods 231:11-23; Yang, X.D. et al. (1999) J. Leukoc. Biol. 66:401-410; Gallo, M.L. et al. (2000) Eur. J. Immunol. 30:534-540.
Согласно другому варианту осуществления животное, которое иммунизируют глобуломером Aβ(20-42) или его производным, может представлять собой мышь с тяжелым комбинированным иммунодефицитом (SCID), который восстанавливали мононуклеарными клетками периферической крови человека или лимфоидными клетками, или их предшественниками. Такие мыши, обозначенные химерными мышами hu-PBMC SCID, как было показано, производят ответы иммуноглобулинов человека при антигенной стимуляции. Дополнительное описание этих мышей и их использования для получения антител, см., например, в Leader, K.A. et al. (1992) Immunology 76:229-234; Bombil, F. et al. (1996) Immunobiol. 195:360-375; Murphy, W.J. et al. (1996) Semin. Immunol. 8:233-241; Herz, U. et al. (1997) Int. Arch. Allergy Immunol. 113:150-152; Albert, S.E. et al. (1997) J. Immunol. 159:1393-1403; Nguyen, H. et al. (1997) Microbiol. Immunol. 41:901-907; Arai, K. et al. (1998) J. Immunol. Methods 217:79-85; Yoshinari, K. and Arai, K. (1998) Hybridoma 17:41-45; Hutchins, W.A. et al. (1999) Hybridoma 18:121-129; Murphy, W.J. et al. (1999) Clin. Immunol. 90:22-27; Smithson, S. L. et al. (1999) Mol. Immunol. 36:113-124; Chamat, S. et al. (1999) J. Infect. Diseases 180:268-277; и Heard, C. et al. (1999) Molec. Med. 5:35-45.
Согласно другому варианту осуществления животное, которое иммунизируют глобуломером Aβ(20-42) или его производным, представляет собой мышь, которую обработали летальной дозой общего облучения организма, затем защищали от облучения клетками костного мозга от мыши с тяжелым комбинированным иммунодефицитом (SCID) и которой затем трансплантировали функциональные лимфоциты человека. Этот тип химеры, обозначенный системой тримерa, использовали для получения моноклональных антител человека посредством иммунизации указанных мышей интересующим антигеном и получения затем моноклональных антител посредством использования стандартизованного способа гибридомы. Дополнительное описание этих мышей и их применения для получения антител см., например, в Eren, R. et al. (1998) Immunology 93:154-161; Reisner, Y and Dagan, S. (1998) Trends Biotechnol. 16:242- 246; Nan, E. et al. (1999) Hepatology 29:553-562; и Bocher, W.O. et al. (1999) Immunology 96:634-641.
Начиная с полученных in-vivo продуцирующих антитело клеток, моноклональные антитела можно получать посредством стандартизованных способов, таких как способ гибридомы, первоначально описанный Kohler и Milstein (1975, Nature 256:495-497) (см. также Brown et al. (1981) J. Immunol 127:539-46; Brown et al. (1980) J Biol Chem 255:4980-83; Yeh et al. (1976) PNAS 76:2927-31; и Yeh et al. (1982) Int. J. Cancer 29:269-75). Способ получения гибридом для моноклональных антител достаточно известен (см. в общем R. H. Kenneth, in Monoclonal Antibodies: A New Dimension In Biological Analyses, Plenum Publishing Corp., New York, New York (1980); E. A. Lerner (1981) Yale J. Biol. Med., 54:387-402; M. L. Gefter et al. (1977) Somatic Cell Genet, 3:231-36). Коротко, иммортализованную линию клеток (как правило, миеломы) сливают с лимфоцитами (как правило, спленоцитами или клетками лимфатических узлов или лимфоцитами периферической крови) млекопитающего, иммунизированного глобуломером Aβ по изобретению или его производным, и супернатанты культуры полученных клеток гибридомы скринируют, чтобы идентифицировать гибридому, продуцирующую моноклональное антитело по настоящему изобретению. Любые из множества хорошо известных способов для слияния лимфоцитов и иммортализованных линий клеток можно применять для этой цели (см. также G. Galfre et al. (1977) Nature 266:550-52; Gefter et al. Somatic Cell Genet, процитированный выше; Lerner, Yale J. Biol. Med., процитированный выше; Kenneth, Monoclonal Antibodies, процитированный выше). Более того, опытный специалист в данной области будет признавать, что существуют различные варианты этих способов, которые являются применимыми подобным образом. Как правило, иммортализованную линию клеток (например, линию клеток миеломы) получают из того же самого вида млекопитающих, что и лимфоциты. Например, можно стабилизировать мышиные гибридомы посредством слияния лимфоцитов от мыши, иммунизированной иммуногенным препаратом по изобретению, с иммортализованной линией клеток мыши. Предпочтительными иммортализованными линиями клеток являются линии клеток миеломы мыши, которые являются чувствительными к культуральной среде, содержащей гипоксантин, аминоптерин и тимидин (среда HAT). Любую из ряда линий клеток миеломы можно использовать по умолчанию в качестве партнера по слиянию, например, линии миеломы P3-NS1/1-Ag4-1, P3-x63-Ag8.653 или Sp2/O-Ag14. Эти линии клеток миеломы доступны в Американской коллекции типовых культур (ATCC), Rockville, MD. Как правило, HAT-чувствительные клетки миеломы мыши сливают со спленоцитами мыши с использованием полиэтиленгликоля (PEG). Затем клетки гибридомы, полученные в результате слияния, отбирают с использованием среды HAT, таким образом убивая неслитые и непродуктивно слитые клетки миеломы (неслитые спленоциты погибают спустя несколько суток, поскольку они не являются трансформированными). Клетки гибридомы, продуцирующие моноклональные антитела по изобретению, идентифицируют скринингом супернатантов культуры гибридомы по таким антителам, например, посредством использования анализа дот-блоттингом, как описано выше и в примере 8, чтобы отобрать такие антитела, которые обладают аффинностями связывания, как определено выше.
Моноклональные антитела 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 и 3B10 все получены с использованием описанного выше способа in-vivo, и их можно получить из гибридомы, как определено здесь.
Подобным образом, указанную гибридому можно использовать как источник нуклеиновой кислоты, кодирующей легкие и/или тяжелые цепи, для рекомбинантного получения антител по настоящему изобретению, как более подробно описано ниже.
Способы in-vitro
В качестве альтернативы получению антител по изобретению посредством иммунизации и селекции, антитела по изобретению можно идентифицировать и выделять посредством скрининга рекомбинантных комбинаторных библиотек иммуноглобулинов с помощью глобуломера Aβ(20-42) или его производного, чтобы таким образом выделить члены библиотеки иммуноглобулинов, обладающие требуемой аффинностью связывания. Наборы для получения и скрининга библиотек дисплея являются коммерчески доступными (например, Pharmacia Recombinant Phage Antibody System, каталожный No. 27-9400-01; и Stratagene SurfZAP® Phage Display Kit, каталожный No. 240612). Во многих вариантах осуществления библиотека дисплея представляет собой библиотеку scFv или библиотеку Fab. Способ фагового дисплея для скрининга библиотек рекомбинантных антител описан в достаточной мере. Примеры способов и соединений, которые можно использовать особенно предпочтительно для получения и скрининга библиотек дисплея антител, можно найти, например, в McCafferty et al. WO 92/01047, US 5969108 и EP 589877 (описан, в частности, дисплей scFv), Ladner et al. US 5223409, US 5403484, US 5571698, US 5837500 и EP 436597 (описан, например слитый белок с pIII); Dower et al. WO 91/17271, US 5427908, US 5580717 и EP 527839 (описан, в частности, Fab дисплей); Winter et al. Международная публикация WO 92/20791 и EP 368684 (описано, в частности, клонирование последовательностей для вариабельных доменов иммуноглобулинов); Griffiths et al. US 5885793 и EP 589877 (описано, в частности, выделение антител человека против антигенов человека с использованием рекомбинантных библиотек); Garrard et al. WO 92/09690 (описаны, в частности, способы экспрессии фагов); Knappik et al. WO 97/08320 (описана библиотека рекомбинантных антител человека HuCal); Salfeld et al. WO 97/29131, (описано получение рекомбинантного антитела человека против антигена человека (фактора некроза опухоли альфа человека), а также созревание аффинности рекомбинантного антитела in-vitro) и Salfeld et al. Предварительная патентная заявка США № 60/126603 и основанные на ней патентные заявки (подобным образом описано получение рекомбинантных антител человека против антигена человека (интерлейкина-12 человека), а также созревание аффинности рекомбинантного антитела in-vitro).
Дополнительные описания скрининга библиотек рекомбинантных антител можно найти в научных публикациях, таких как Fuchs et al. (1991) Bio/Technology 9: 1370-1372; Hay et al. (1992) Hum Antibod Hybridomas 3:81-85; Huse et al. (1989) Science 246:1275-1281; Griffiths et al. (1993) EMBO J 12:725-734; Hawkins et al. (1992) J Mol Biol 226:889-896; Clarkson et al. (1991) Nature 352:624-628; Gram et al. (1992) PNAS 89:3576-3580; Garrard et al. (1991) Bio/Technology 9: 1373-1377; Hoogenboom et al. (1991) Nuc Acid Res 19:4133-4137; Barbas et al. (1991) PNAS 88:7978-7982; McCafferty et al. Nature (1990) 348:552-554; и Knappik et al. (2000) J. Mol. Biol. 296:57-86.
В качестве альтернативы использованию систем бактериофаговых дисплеев, библиотеки рекомбинантных антител можно экспрессировать на поверхности клеток дрожжей или бактериальных клеток. В WO 99/36569 описаны способы получения и скрининга библиотек, экспрессированных на поверхности клеток дрожжей. В WO 98/49286 более подробно описаны способы получения и скрининга библиотек, экспрессированных на поверхности бактериальных клеток.
Во всех способах in vitro процесс отбора для обогащения рекомбинантными антителами с желательными свойствами формирует интегральную часть процесса, обычно обозначаемого «пэннинг» и часто принимает форму аффинной хроматографии на колонках, к носителю которых присоединена структура-мишень. Затем перспективные молекулы-кандидаты подвергают индивидуальному определению их абсолютной и/или относительной аффинностей, предпочтительно посредством стандартизованных анализов дот-блоттингом, как описано выше и в примере 8.
После идентификации и достаточной характеризации интересующего антитела из комбинаторной библиотеки последовательности ДНК, кодирующие легкие и тяжелые цепи указанного антитела, выделяют посредством стандартизованных молекулярно-биологических способов, например посредством амплификации ПЦР ДНК из упаковки дисплея (например, фага), выделенной во время скрининга библиотеки. Нуклеотидные последовательности генов для легких и тяжелых цепей антитела, которые можно использовать для получения праймеров для ПЦР, известны специалистам в данной области. Множество таких последовательностей описано, например, в Kabat, E. A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242 и в базе данных последовательностей зародышевой линии человека VBASE.
Антитело или часть антитела по изобретению можно получать посредством рекомбинантной экспрессии генов легких и тяжелых цепей иммуноглобулина в клетке-хозяине. Для рекомбинантной экспрессии антитела клетку-хозяина трансфицируют одним или несколькими рекомбинантными экспрессирующими векторами, несущими фрагменты ДНК, кодирующие легкие и тяжелые иммуноглобулиновые цепи указанного антитела, таким образом экспрессируя легкие и тяжелые цепи в клетке-хозяине и, предпочтительно, секретируя их в среду, в которой культивируют указанные клетки-хозяева. Антитела можно выделять из этой среды. Стандартизованные способы рекомбинантной ДНК используют, чтобы получить гены для тяжелой и легкой цепей антитела, вставить указанные гены в рекомбинантные экспрессирующие векторы и ввести указанные векторы в клетки-хозяева. Способы этого вида указаны, например, в Sambrook, Fritsch and Maniatis (eds.), Molecular Cloning; A Laboratory Manual, Second Edition, Cold Spring Harbor, N. Y., (1989), Ausubel, F.M. et al. (eds.) Current Protocols in Molecular Biology, Greene Publishing Associates, (1989) и в US 4816397 Boss et al.
После получения фрагментов ДНК, кодирующих фрагменты VH и VL интересующего антитела, с указанными фрагментами ДНК можно дополнительно манипулировать с использованием стандартизованных способов рекомбинантной ДНК, например, чтобы превратить гены для вариабельных областей в гены для полноразмерных цепей антитела, в гены для Fab-фрагментов или в ген scFv. Эти манипуляции включают в себя функциональное связывание VL- или VH-кодирующего фрагмента ДНК с другим фрагментом ДНК, кодирующим другой белок, например константную область антитела или гибкий линкер. Термин «функционально связанный» следует понимать в том смысле, что два фрагмента ДНК связаны таким образом, что аминокислотные последовательности, кодируемые указанными двумя фрагментами ДНК, остаются в рамке.
Выделенную ДНК, кодирующую область VH, можно превратить в ген для полноразмерной тяжелой цепи посредством функционального связывания ДНК, кодирующей VH-область, с другой молекулой ДНК, кодирующей константные области тяжелой цепи (CH1, CH2 и CH3). Последовательности генов константной области тяжелой цепи человека хорошо известны (см., например, Kabat, E. A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242), и фрагменты ДНК, перекрывающие указанную область, можно получить посредством стандартизованной амплификации ПЦР. Константная область тяжелой цепи может представлять собой константную область из IgG1, IgG2, IgG3, IgG4, IgM, IgA, IgE или IgD, где предпочтение отдают константной области из IgG, в частности IgG1 или IgG4. Для получения гена для Fab-фрагмента тяжелой цепи VH-кодирующую ДНК можно функционально связывать с другой молекулой ДНК, кодирующей только константную область CH1 тяжелой цепи.
Выделенную ДНК, кодирующую область VL, можно превратить в ген для полноразмерной легкой цепи (и в ген для Fab легкой цепи) посредством функционального связывания VL-кодирующей ДНК с другой молекулой ДНК, кодирующей константную область CL легкой цепи. Последовательности генов константной области легкой цепи человека хорошо известны (см. Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242), и фрагменты ДНК, перекрывающие указанную область, можно получить посредством стандартизованной амплификации ПЦР. Константная область легкой цепи может представлять собой константную область каппа или лямбда, где предпочтение отдают константной области каппа.
Для получения гена scFv VH- и VL-кодирующие фрагменты ДНК можно функционально связывать с другим фрагментом, кодирующим гибкий линкер, например, аминокислотной последовательностью (Gly4-Ser)3, так что последовательности VH и VL экспрессированы как непрерывный одноцепочечный белок, где области VL и VH связаны друг с другом посредством указанного гибкого линкера (см. Bird et al. (1988) Science 242:423-426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883; McCafferty et al., Nature (1990) 348:552-554).
Отдельный домен VH и VL, обладающий аффинностями связывания, как описано выше, можно выделить из библиотек отдельного домена посредством описанных выше способов. Две однодоменные цепи VH (с или без CH1) или две цепи VL, или пару из одной цепи VH и одной цепи VL с желаемой аффинностью связывания можно использовать, как описано в настоящем описании, для антител по изобретению.
Чтобы экспрессировать рекомбинантные антитела или части антитела по изобретению, ДНК, кодирующие частичные или полноразмерные легкие и тяжелые цепи, можно встраивать в экспрессирующие векторы, так чтобы функционально связывать гены с подходящими последовательностями для контроля транскрипции и трансляции. В этом контексте термин «функционально связанный» следует понимать в том смысле, что ген антитела лигирован в вектор таким образом, что последовательности для контроля транскрипции и трансляции внутри вектора выполняют предназначенную им функцию регуляции транскрипции и трансляции указанного гена антитела.
Целесообразно выбирать экспрессирующий вектор и контролирующие экспрессию последовательности так, чтобы они являлись совместимыми с используемой клеткой-хозяином. Ген для легкой цепи антитела и ген для тяжелой цепи антитела можно вставлять в отдельные векторы или оба гена вставляют в один и тот же экспрессирующий вектор, что является обычным случаем. Гены антитела вставляют в экспрессирующий вектор посредством стандартизованных способов (например, посредством лигирования комплементарных участков рестрикционного расщепления в фрагменте гена антитела и в векторе, или посредством лигирования тупых концов, если не присутствуют участки рестрикционного расщепления). Экспрессирующий вектор может уже нести последовательности для константных областей антитела перед вставкой последовательностей для легкой и тяжелой цепей. Например, один способ представляет собой превращение последовательностей VH и VL в гены полноразмерного антитела посредством их вставки в экспрессирующие векторы, уже кодирующие константные области тяжелой и, соответственно, легкой цепи, таким образом, функционального связывания фрагмента VH с фрагментом(фрагментами) CH в векторе, а также функционального связывания фрагмента VL с фрагментом CL в векторе.
Дополнительно или альтернативно, рекомбинантный экспрессирующий вектор может кодировать сигнальный пептид, облегчающий секрецию цепи антитела из клетки-хозяина. Ген для указанной цепи антитела можно клонировать в вектор, таким образом связывая сигнальный пептид в рамке с N-концом гена для цепи антитела. Сигнальный пептид может представлять собой сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (т.е. сигнальный пептид из не относящегося к иммуноглобулину белка). Помимо генов для цепи антитела, экспрессирующие векторы по изобретению могут содержать регуляторные последовательности, контролирующие экспрессию генов цепи антитела в клетке-хозяине.
Термин «регуляторная последовательность» предназначен, чтобы включать в себя промоторы, энхансеры и дополнительные элементы для контроля экспрессии (например, сигналы полиаденилирования), контролирующие транскрипцию или трансляцию генов для цепи антитела. Регуляторные последовательности этого вида описаны, например, в Goeddel; Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Специалисту в данной области понятно, что дизайн экспрессирующего вектора, включающий в себя выбор регуляторных последовательностей, может зависеть от таких факторов, как подлежащая трансформации клетка-хозяин, желательная сила экспрессии белка и т.д. Предпочтительные регуляторные последовательности для экспрессии в клетках-хозяевах млекопитающих включают в себя вирусные элементы, обеспечивающие сильную и конститутивную экспрессию белка в клетках млекопитающих, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV) (такие как промотор/энхансер CMV), вируса обезьян 40 (SV40) (такие как промотор/энхансер SV40), аденовируса (например, главный поздний промотор аденовируса (AdMLP)) и полиомы. Дополнительное описание вирусных регуляторных элементов и их последовательностей см., например, в US 5168062 Stinski, US 4510245 Bell et al. и US 4968615 Schaffner et al.
Помимо генов для цепи антитела и регуляторных последовательностей, рекомбинантные экспрессирующие векторы по изобретению могут содержать дополнительные последовательности, такие как последовательности, регулирующие репликацию вектора в клетках-хозяевах (например, точки начала репликации) и гены селективных маркеров. Гены селективных маркеров облегчают отбор клеток-хозяев, в которые был введен вектор (см., например, патенты США No 4399216, 4634665 и 5179017, все Axel et al.). Например, общим для генов селективных маркеров является то, что они обеспечивают клетке-хозяину, в которую введен вектор, устойчивость к цитотоксическим лекарственным средствам, таким как G418, гигромицин или метотрексат. Предпочтительные гены селективных маркеров включают в себя ген для дигидрофолатредуктазы (DHFR) (для использования в dhfr- клетках-хозяевах с отбором/амплификацией с метотрексатом) и ген neo (для отбора с G418).
Для экспрессии легких и тяжелых цепей экспрессирующий вектор(ы), кодирующий(ие) указанные тяжелые и легкие цепи, трансфицируют в клетку-хозяина посредством стандартизованных способов. Различные формы термина «трансфекция» предназначены, чтобы включать в себя множество способов, общепринятых для введения экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например, электропорацию, кальций-фосфатную преципитацию, DEAE-декстрановую трансфекцию и т.п. Хотя теоретически является возможным экспрессировать антитела по изобретению либо в прокариотических, либо в эукариотических клетках-хозяевах, предпочтение отдают экспрессии антител в эукариотических клетках и, в частности, в клетках-хозяевах млекопитающих, поскольку вероятность правильного сворачивания, сборки и секреции иммунологически активного антитела выше в таких эукариотических клетках и, в частности, в клетках млекопитающих, чем в прокариотических клетках. Опубликовано, что прокариотическая экспрессия генов антитела является неэффективной для получения высокого выхода активного антитела (Boss, M. A. and Wood, C. R. (1985) Immunology Today 6:12-13).
Предпочтительные клетки-хозяева млекопитающих для экспрессии рекомбинантных антител по изобретению включают в себя клетки CHO (включая dhfr- клетки CHO, описанные в Urlaub and Chasin, (1980) Proc. Natl. Acad. Sci. USA 77:4216-4220, которые использовали вместе с DHFR-селективным маркером, как описано, например, в R.J. Kaufman and P.A. Sharp (1982) Mol. Biol. 159:601-621), клетки миеломы NS0, клетки COS и клетки SP2. При введении рекомбинантных экспрессирующих векторов, кодирующих гены антитела, в клетки-хозяева млекопитающих, антитела получают посредством культивирования клеток-хозяев, пока антитело не экспрессируется в указанных клетках-хозяевах, или, предпочтительно, пока антитело не секретируется в культуральную среду, в которой выращивают клетки-хозяева. Затем антитела можно выделять из культуральной среды с использованием стандартизованных способов очистки белка.
Подобным образом, является возможным использовать клетки-хозяева для получения частей интактных антител, таких как Fab-фрагменты или scFv-молекулы. Конечно, варианты описанного выше способа включены в изобретение. Например, может являться желательным трансфицировать клетку-хозяина ДНК, кодирующую либо легкую цепь, либо тяжелую цепь (но не обе) антитела по изобретению. Если присутствуют либо легкие, либо тяжелые цепи, которые не требуются для связывания интересующего антигена, тогда ДНК, кодирующую либо такую легкую, либо такую тяжелую цепь, либо обе, можно удалить частично или полностью посредством способов рекомбинантной ДНК. Молекулы, экспрессируемые с таких усеченных молекул ДНК, подобным образом включены в антитела по изобретению. Кроме того, является возможным получить бифункциональные антитела, в которых тяжелая цепь и легкая цепь представляют собой антитело по изобретению, а другая тяжелая цепь и другая легкая цепь обладают специфичностью к антигену, отличному от интересующего антигена, посредством перекрестного связывания антитела по изобретению со вторым антителом посредством стандартизованных химических способов.
В предпочтительной системе для рекомбинантной экспрессии антитела по изобретению или его антигенсвязывающей части рекомбинантный экспрессирующий вектор, кодирующий как тяжелую цепь антитела, так и легкую цепь антитела, вводят в клетки dhfr- CHO посредством трансфекции, опосредованной фосфатом кальция. В рекомбинантном экспрессирующем векторе гены для тяжелых и легких цепей антитела в каждом случае являются функционально связанными с регуляторными элементами CMV энхансер/AdMLP-промотор, чтобы обеспечивать сильную транскрипцию указанных генов. Рекомбинантный экспрессирующий вектор несет также ген DHFR, который можно использовать для отбора клеток dhfr- CHO, трансфицированных вектором, с использованием селекции/амплификации с метотрексатом. Отобранные трансформированные клетки-хозяева культивируют, так что экспрессируются тяжелые и легкие цепи антитела, и интактное антитело выделяют из культуральной среды. Стандартизованные молекулярно-биологические способы используют для получения рекомбинантного экспрессирующего вектора, для трансфекции клеток-хозяев, для отбора трансформантов, для культивирования указанных клеток-хозяев и для получения антитела из культуральной среды. Таким образом, изобретение относится к способу синтеза рекомбинантного антитела по изобретению посредством культивирования клетки-хозяина по изобретению в подходящей культуральной среде, пока не синтезируется рекомбинантное антитело по изобретению. Способ, кроме того, может включать в себя выделение указанного рекомбинантного антитела из указанной культуральной среды.
В качестве альтернативы скринингу библиотек рекомбинантных антител посредством фагового дисплея другие способы, известные специалистам в данной области, можно использовать для скрининга больших комбинаторных библиотек для идентификации антител по изобретению. В принципе, можно применять любую систему экспрессии, в которой достигают близкой физической связи между нуклеиновой кислотой и кодируемой ею антителом и которую можно использовать для отбора подходящей последовательности нуклеиновой кислоты на основании свойств кодируемого ею антитела.
В одном типе альтернативной системы экспрессии библиотеку рекомбинантных антител экспрессируют в слитой форме РНК-белок, как описано в WO 98/31700 Szostak and Roberts, и в Roberts, R.W. and Szostak, J.W. (1997) Proc. Natl. Acad. Sci USA 94:12297-12302. В этой системе трансляцией in-vitro синтетических мРНК, несущих на их 3'-конце пуромицин, пептидил-акцепторный антибиотик, получают ковалентно слитые мРНК и кодируемый ею пептид или белок. Таким образом, специфическую мРНК в сложной смеси мРНК (например, комбинаторной библиотеке) можно концентрировать на основании свойств кодируемого пептида или белка (например, антитела или его части), таких как связывание указанного антитела или указанной его части с глобуломером Aβ(20-42) или его производным. Последовательности нуклеиновой кислоты, которые кодируют антитела или их части и которые получены скринингом таких библиотек, можно экспрессировать посредством рекомбинантных способов указанным выше образом (например, в клетках-хозяевах млекопитающих) и можно, кроме того, подвергать дополнительному созреванию аффинности посредством либо скрининга слитых мРНК-пептидов в дополнительных циклах, либо введения мутаций в исходно отобранную последовательность(последовательности), либо с использованием других способов созревания аффинности рекомбинантных антител in-vitro описанным выше образом.
Сочетания способов in-vivo и in-vitro
Подобным образом, антитела по изобретению можно получать с использованием сочетания способов in-vivo и in-vitro, например, способами, в которых глобуломеру Aβ(20-42) или его производному сначала позволяют воздействовать на репертуар антител в организме-хозяине in vivo для стимуляции продукции антител, связывающих глобуломер Aβ(20-42) или его производное, и затем дополнительный отбор антитела и/или созревание антитела (т.е. оптимизацию) осуществляют с помощью одного или нескольких способов in-vitro. Согласно одному варианту осуществления комбинированный способ этого вида может включать в себя, во-первых, иммунизацию не относящегося к человеку животного (например, мыши, крысы, кролика, курицы, верблюда, овцы или козы, или их трансгенного варианта, или химерной мыши) указанным глобуломером Aβ(20-42) или его производным для стимуляции ответа антител на антиген и затем получение и скрининг библиотеки фагового дисплея антител с использованием последовательностей иммуноглобулинов лимфоцитов, стимулированных in vivo посредством воздействия указанного глобуломера Aβ(20-42) или производного. Первую стадию этого комбинированного способа можно проводить описанным выше применительно к способам in-vivo образом, в то время как вторую стадию этого способа можно проводить описанным выше применительно к способам in-vitro образом. Предпочтительные способы гипериммунизации не относящихся к человеку животных с последующим скринингом библиотек фагового дисплея, полученных из указанных стимулированных лимфоцитов, включают в себя способы, описанные BioSite Inc., см., например, WO 98/47343, WO 91/17271, US 5427908 и US 5580717.
Согласно другому варианту осуществления комбинированный способ включает в себя, во-первых, иммунизацию не относящегося к человеку животного (например, мыши, крысы, кролика, курицы, верблюда, овцы, или козы, или их нокаутированного и/или трансгенного варианта, или химерной мыши) глобуломером Aβ(20-42) по изобретению или его производным для стимуляции ответа антител на указанный глобуломер Aβ(20-42) или его производное и отбора лимфоцитов, продуцирующих антитела, обладающие желаемой специфичностью, посредством скрининга гибридом (полученных, например, из иммунизированных животных). Гены для антител или однодоменных антител выделяют из отобранных клонов (посредством стандартизованных способов клонирования, таких как полимеразная цепная реакция с обратной транскриптазой) и подвергают созреванию аффинности in-vitro, чтобы таким образом улучшить связывающие свойства отобранного антитела или отобранных антител. Первую стадию этого способа можно проводить описанным выше применительно к способам in-vivo образом, в то время как вторую стадию этого способа можно проводить описанным выше применительно к способам in-vitro образом, в частности, с использованием способов созревания аффинности in-vitro, таких как способы, описанные в WO 97/29131 и WO 00/56772.
В дополнительном комбинированном способе рекомбинантные антитела получают из отдельных выделенных лимфоцитов с использованием способа, известного специалистам в данной области как способ антитела из отобранного лимфоцита (SLAM) и описанного в US 5627052, WO 92/02551 и в Babcock, J. S. et al. (1996) Proc. Natl. Acad. Sci. USA 93:7843-7848. В этом способе не относящееся к человеку животное (например, мышь, крысу, кролика, курицу, верблюда, овцу, козу, или их трансгенный вариант, или химерную мышь), во-первых, иммунизируют in vivo глобуломером Aβ(20-42) или его производным для стимуляции иммунного ответа на указанный олигомер или производное, и затем отдельные клетки, секретирующие интересующие антитела, отбирают с использованием антигенспецифического анализа гемолитических бляшек. С этой целью глобуломер, или его производное, или интересующие структурно родственные молекулы можно объединять с эритроцитами овцы с использованием линкера, такого как биотин, таким образом получая возможность идентифицировать отдельные клетки, секретирующие антитела с подходящей специфичностью, с использованием анализа гемолитических бляшек. После идентификации клеток, секретирующих интересующие антитела, кДНК для вариабельных областей легких и тяжелых цепей получают из клеток посредством ПЦР с обратной транскриптазой, и затем указанные вариабельные области можно экспрессировать совместно с подходящими константными областями иммуноглобулина (например, константными областями человека) в клетках-хозяевах млекопитающих, таких как клетки COS или CHO. Затем клетки-хозяева, трансфицированные амплифицированными иммуноглобулиновыми последовательностями, полученными из отобранных in vivo лимфоцитов, можно подвергать дальнейшему анализу in-vitro и отбору in-vitro посредством рассредоточения трансфицированных клеток, например, для выделения клеток, экспрессирующих антитела с аффинностью связывания. Кроме того, с иммуноглобулиновыми последовательностями можно манипулировать in vitro.
Антитела, обладающие требуемыми аффинностями, определенными здесь, можно отбирать проведением дот-блоттинга, в основном, как описано выше. Коротко, антиген присоединяют к твердой матрице, предпочтительно, наносят точками на нитроцеллюлозную мембрану, в серийных разведениях. Затем проводят контактирование иммобилизованного антигена с интересующим антителом с последующей детекцией последнего посредством конъюгированного с ферментом вторичного антитела и колориметрической реакции; при определенных концентрациях антитела и антигена количество связавшегося антитела позволяет определение аффинности. Таким образом, относительную аффинность двух различных антител для одной мишени или одного антитела для двух различных мишеней, определяют здесь как отношение соответствующих количеств связанного с мишенью антитела, наблюдаемое для двух сочетаний антитело-мишень при в остальном идентичных условиях дот-блоттинга.
Антитела, связывающиеся с тем же самым эпитопом, как моноклональное антитело 5F7, 10F11, 7C6, 4B7, 6A2, 2F2, 4D10, 7E5, 10C1 или 3B10, можно получить по существу известным способом.
Таким же образом, описанным выше, каким антитела могут конкурировать, различные структуры мишени называют здесь «конкурирующими» за определенное антитело, если по меньшей мере одна из этих структур способна специфически снижать измеряемое связывание другой, предпочтительно посредством предоставления перекрывающегося или идентичного эпитопа, более предпочтительно, идентичного эпитопа.
Конкурирующие молекулы-мишени можно применять для прямого отбора антител на основании их относительной аффинности для таких структур-мишеней. Относительные аффинности, таким образом, можно определить непосредственно с использованием конкурентного анализа, в котором различимые формы конкурирующих молекул, например дифференциально меченные конкурирующие структуры приводят в контакт с интересующим антителом и относительную аффинность антитела для каждой из этих групп выводят из относительных количеств этих молекул, которые являются связанными с антителом.
Такую конкуренцию можно использовать для прямого обогащения антителами, обладающими желательной относительной аффинностью к молекуле-мишени, посредством присоединения молекулы-мишени, для которой желательна большая аффинность, к твердой подложке-матрице и добавления соответствующего количества, предпочтительно молярного избытка, конкурирующей молекулы, для которой желательна меньшая аффинность, в среду. Таким образом, антитела, обладающие желательными относительными аффинностями, будут проявлять тенденцию связываться с матрицей более сильно, чем другие, и их можно получить после отмывки от менее желательных форм, например, посредством отмывки при низких концентрациях соли и последующего сбора связанного антитела посредством его обратимого отделения от его мишени с использованием высоких концентраций соли. Если желательно, можно проводить несколько циклов обогащения. В конкретном варианте осуществления изобретения, где генотип, лежащий в основе антитела, физически связан с этим антителом, например, в пуле гибридом или антиген-экспонирующих фагов или клеток дрожжей, можно сохранить соответствующий фенотип.
В другом варианте осуществления изобретения используют модифицированный дот-блоттинг, где иммобилизованный антиген конкурирует с растворенной молекулой за связывание с антителом, так что относительную аффинность антитела можно вывести из процента, связанного с иммобилизованным антигеном.
Части антитела, такие как Fab- и F(ab')2-фрагменты можно получать из целых антител с использованием общепринятых способов, таких как расщепление папаином или пепсином. Кроме того, антитела, части антител и молекулы иммуноадгезии можно получать с использованием стандартизованных способов рекомбинантной ДНК.
Настоящее изобретение также относится к фармацевтическим средствам (композициям), содержащим антитело по изобретению и, необязательно, фармацевтически пригодный носитель. Фармацевтические композиции по изобретению могут, кроме того, содержать по меньшей мере одно дополнительное лекарственное средство, например, одно или несколько дополнительных лекарственных средств для лечения заболевания, для облегчения которого являются применимыми антитела по изобретению. Если, например, антитело по изобретению связывается с глобуломером по изобретению, фармацевтическая композиция может, кроме того, содержать одно или несколько дополнительных лекарственных средств, применимых для лечения нарушений, при которых активность указанного глобуломера является важной.
Фармацевтически приемлемые носители включают в себя любые растворители, диспергенты, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие всасывание средства и т.п., пока они являются физиологически совместимыми. Фармацевтически приемлемые носители включают в себя, например, воду, физиологический раствор, фосфатно-солевой буфер, глюкозу, глицерин, этанол и т.п. и их сочетания. Во многих случаях предпочтение отдают использованию изотонических средств, например сахаров, полиспиртов, таких как маннит или сорбит, или дополнительно, хлорида натрия. Фармацевтически приемлемые носители могут дополнительно содержать относительно небольшие количества вспомогательных веществ, таких как смачиватели или эмульгаторы, консерванты или буферы, увеличивающие время полужизни или эффективность антител.
Фармацевтические композиции могут являться пригодными, например, для парентерального введения. Здесь антитела получают предпочтительно в форме пригодных для инъекции растворов с содержанием антитела 0,1-250 мг/мл. Пригодные для инъекции растворы можно получать в жидкой или лиофилизированной форме, где лекарственная форма находится в сосуде из флинтгласа или флаконе, ампуле или заполненном шприце. Буфер может содержать L-гистидин (1-50 мМ, предпочтительно 5-10 мМ) и обладать pH 5,0-7,0, предпочтительно, 6,0. Дополнительные подходящие буферы включают в себя, в качестве неограничивающих примеров, натрий-сукцинатные, натрий-цитратные, натрий-фосфатные или калий-фосфатные буферы. Чтобы регулировать тоничность раствора, можно использовать хлорид натрия в концентрации 0-300 мМ (предпочтительно 150 мМ для жидкой лекарственной формы). Криопротектор, например сахарозу (например, 0-10%, предпочтительно, 0,5-1,0%), можно также включать в лиофилизированную лекарственную форму. Другими подходящими криопротекторами являются трегалоза и лактоза. Наполнители, например маннит (например, 1-10%, предпочтительно, 2-4%), можно также включать в лиофилизированную лекарственную форму. Стабилизаторы, например L-метионин (например, 51-50 мМ, предпочтительно, 5-10 мМ), можно использовать как в жидкой, так и в лиофилизированной лекарственной форме. Дополнительными подходящими наполнителями являются глицин и аргинин. Можно использовать также поверхностно-активные вещества, например полисорбат 80 (например, 0-0,05%, предпочтительно, 0,005-0,01%). Дополнительными поверхностно-активными веществами являются полисорбат 20 и поверхностно-активные вещества BRIJ.
Композиции по изобретению могут обладать множеством форм. Они включают в себя жидкие, полужидкие и твердые лекарственные формы, такие как жидкие растворы (например, пригодные для инъекции и пригодные для инфузии растворы), дисперсии или суспензии, таблетки, пилюли, порошки, липосомы и суппозитории. Предпочтительная форма зависит от предназначенного типа введения и от терапевтического применения. Как правило, ссылаются на композиции в форме пригодных для инъекции или пригодных для инфузии растворов, например, композиции, сходные с композициями других антител для пассивной иммунизации людей. Предпочтительным способом введения является парентеральный (например, внутривенный, подкожный, внутрибрюшинный или внутримышечный). Согласно предпочтительному варианту осуществления антитело вводят внутривенной инфузией или инъекцией. Согласно другому предпочтительному варианту осуществления антитело вводят посредством внутримышечной или подкожной инъекции.
Терапевтические композиции, как правило, должны являться стерильными и стабильными в условиях получения и хранения. Композиции можно составлять в форме растворов, микроэмульсий, дисперсий, липосом или других упорядоченных структур, пригодных для высоких концентраций активного вещества. Стерильные пригодные для инъекции растворы можно получать посредством введения активного соединения (т.е., антитела) в требуемом количестве в подходящий растворитель, когда это целесообразно, с одним или с сочетанием вышеупомянутых ингредиентов, как необходимо, и затем стерилизации фильтрованием указанного раствора.
Дисперсии обычно получают введением активного соединения в стерильный носитель, содержащий основную дисперсионную среду и, когда это целесообразно, другие необходимые ингредиенты. В случае стерильного лиофилизированного порошка для получения стерильных пригодных для инъекции растворов сушка в вакууме и сушка распылением являются предпочтительными способами получения, которыми получают порошок активного ингредиента и, когда это целесообразно, дополнительных желательных ингредиентов из предварительно стерилизованного фильтрованием раствора. Правильную текучесть раствора можно поддерживать с использованием, например, покрытия, такого как a лецитин, посредством поддержания, в случае дисперсий, требуемого размера частиц или посредством использования поверхностно-активных веществ. Кроме того, можно достигать замедленного всасывания пригодной для инъекции композиции введением в композицию средства, замедляющего всасывание, например, солей моностеаратов и желатина.
Антитела по изобретению можно вводить множеством способов, известных специалисту в данной области, хотя предпочтительным типом введения для многих терапевтических применений является подкожная инъекция, внутривенная инъекция или вливание. Специалисту в данной области понятно, что способ и/или тип введения зависит от желаемого результата. Согласно конкретным вариантам осуществления активное соединение можно получать с носителем, который защищает соединение против быстрого высвобождения, например, таким как составы с замедленным или контролируемым высвобождением, включающие в себя импланты, чрескожные пластыри и микроинкапсулированные системы высвобождения. Можно использовать биологически разлагаемые биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевую кислоту, коллаген, полиортоэфиры и полимолочную кислоту. Способы получения таких составов хорошо известны специалистам в данной области; см., например, Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Согласно конкретным вариантам осуществления, антитело по изобретению можно вводить перорально, например, в инертном разбавителе или в преобразующемся в ходе обмена веществ съедобном носителе. Антитело (и дополнительные ингредиенты, если желательно) можно также заключать в твердую или мягкую желатиновую капсулу, прессовать в таблетки или добавлять непосредственно в пищу. Для перорального терапевтического введения антитела можно смешивать с наполнителями и использовать в форме пероральных таблеток, буккальных таблеток, капсул, эликсиров, суспензий, сиропов и т.п. Если подразумевают вводить антитело по изобретению посредством способа, отличного от парентерального, может являться необходимым выбирать покрытие из материала, предупреждающего его инактивацию.
Настоящее изобретение также относится к способу ингибирования активности глобуломеров по изобретению у индивидуумов, страдающих от нарушения, в которое вовлечен белок β-амилоида, и при котором особенно важной является активность указанных глобуломеров по изобретению. Указанный способ включает в себя введение по меньшей мере одного антитела по изобретению индивидууму с целью ингибирования активности глобуломера, с которым связывается антитело. Указанный индивидуум предпочтительно является человеком. Антитело по изобретению можно вводить индивидууму-человеку для терапевтических целей. Кроме того, антитело по изобретению можно вводить не относящемуся к человеку млекопитающему для ветеринарных целей или в рамках модели конкретного нарушения на животных. Такие модели на животных могут являться применимыми для оценки терапевтической эффективности антител по изобретению (например, для тестирования доз и периодов введения).
Нарушения, при которых играют роль глобуломеры по изобретению, включают в себя, в частности, нарушения, в которые вовлечены развитие и/или прогрессирование глобуломера по изобретению. Это, в частности, те нарушения, при которых глобуломеры по изобретению являются явно или предположительно ответственными за патофизиологию указанного нарушения или являются фактором, вносящим вклад в развитие и/или прогрессирование указанного нарушения. Соответственно, сюда включены те нарушения, при которых ингибирование активности глобуломеров по изобретению может облегчать симптомы и/или прогрессирование нарушения. Такие нарушения можно верифицировать, например, по увеличенной концентрации глобуломеров по изобретению в жидкости организма индивидуума, страдающего от конкретного нарушения (например, увеличенной концентрации в сыворотке, плазме, CSF, моче и т.д.). Это можно детектировать, например, с использованием антитела по изобретению. Глобуломеры по изобретению играют важную роль в патологии, связанной с множеством нарушений, в которые вовлечены нейродегенеративные элементы, когнитивные нарушения, нейротоксические элементы и воспалительные элементы.
В другом аспекте изобретения нарушения, которые можно лечить или предупреждать, включают в себя нарушения, связанные с амилоидозами. Термин «амилоидозы» здесь обозначает ряд нарушений, характеризующихся аномальным сворачиванием, агглютинацией, агрегацией и/или накоплением определенных белков (амилоиды, фиброзные белки и их предшественники) в различных тканях организма. При болезни Альцгеймера и синдроме Дауна повреждена нервная ткань, а при церебральной амилоидной ангиопатии (CAA) повреждены кровеносные сосуды.
Фармацевтические композиции по изобретению могут содержать «терапевтически эффективное количество» или «профилактически эффективное количество» антитела или части антитела по изобретению. «Терапевтически эффективное количество» обозначает количество, эффективное в дозах и в течение периодов времени, необходимых для достижения желательного терапевтического результата. Специалист в данной области может определить терапевтически эффективное количество антитела или части антитела, и оно может меняться в соответствии с такими факторами, как стадия заболевания, возраст, пол и масса индивидуума, и способность антитела или части антитела вызывать желаемый ответ у индивидуума. Терапевтически эффективным количеством является также количество, при котором терапевтически благоприятные эффекты антитела или части антитела превосходят любые токсические или неблагоприятные эффекты. «Профилактически эффективное количество» обозначает количество, эффективное в дозах и в течение периодов времени, необходимых для достижения желательного профилактического результата. Как правило, поскольку профилактическую дозу используют для субъектов до заболевания или на более ранней стадии заболевания, профилактически эффективное количество будет меньше, чем терапевтически эффективное количество.
Кроме того, настоящее изобретение относится к дополнительному способу предупреждения или лечения болезни Альцгеймера у пациента, нуждающегося в таком предупреждении или лечении. Этот способ включает в себя стадию введения пациенту упомянутой выше вакцины в количестве, достаточном для обеспечения предупреждения или лечения.
Кроме того, настоящее изобретение относится к способу идентификации соединений, подходящих для активной иммунизации пациента, для которого предсказано развитие амилоидоза, например болезни Альцгеймера. Этот способ включает в себя: 1) подвергание одного или нескольких интересующих соединений воздействию одного или нескольких антител, описанных выше, в течение времени и при условиях, подходящих для связывания одного или нескольких соединений с антителом или антителами; 2) идентификацию соединений, связавшихся с антителом или антителами, где идентифицированные соединения подлежат использованию для активной иммунизации пациента, для которого предсказано развитие амилоидоза, например болезни Альцгеймера.
В рамках диагностического использования антител качественное или количественное определение специфического глобуломера служит, в частности, для диагностики связанных с заболеванием форм β-амилоида. В этом контексте специфичность означает возможность детекции конкретного глобуломера, или его производного, или их смеси, с достаточной чувствительностью. Антитела по изобретению преимущественно обладают концентрациями порога детекции менее чем 10 нг/мл образца, предпочтительно, менее чем 1 нг/мл образца и особенно предпочтительно, менее чем 100 пг/мл образца, что означает, что по меньшей мере концентрацию глобуломера на мл образца, указанную в каждом случае, преимущественно, также более низкие концентрации, можно детектировать посредством антител по изобретению.
Детекцию проводили иммунологически. Ее, в принципе, можно проводить с использованием любого аналитического или диагностического способа анализа, в котором используют антитела, включая способы агглютинации и преципитации, иммуноанализы, иммуногистохимические способы и способы иммуноблоттинга, например, Вестерн-блоттинг или, предпочтительно, способы дот-блоттинга. Сюда включены также способы in vivo, например, способы визуализации.
Использование иммуноанализов является преимущественным. Подходят как конкурентные иммуноанализы, т.е. анализы, где антиген и меченый антиген (индикатор) конкурируют за связывание с антителом, так и сэндвич-иммуноанализы, т.е. анализы, где связывание специфических антител с антигеном детектируют посредством второго, обычно, меченого антитела. Эти анализы могут являться либо гомогенными, т.е. без разделения на твердую и жидкую фазы, либо гетерогенными, т.е. связанные метки отделяют от несвязанных меток, например, посредством связанных с твердой фазой антител. В зависимости от мечения и способа измерения различные гетерогенные и гомогенные форматы иммуноанализа можно классифицировать на конкретные классы, например, RIA (радиоиммунные анализы), ELISA (твердофазный иммуноферментный анализ), FIA (флуоресцентный иммуноанализ), LIA (люминесцентный иммуноанализ), TRFIA (FIA с временным разрешением), IMAC (иммуноактивация), EMIT (ферментно-мультиплицируемый иммунный тест), TIA (турбидиметрический иммуноанализ), I-ПЦР (иммуно-ПЦР).
Для количественного определения глобуломера по изобретению предпочтение отдают конкурентным иммуноанализам, в которых определенное количество меченного производного глобуломера, служащего индикатором, конкурирует с глобуломером из образца (содержащего неизвестное количество немеченых глобуломеров), подлежащего количественному определению, за связывание с используемым антителом. Количество антигена, т.е. количество глобуломера в образце, можно определить по количеству вытесненного индикатора с помощью стандартной кривой.
Показано, что из меток, доступных для этих целей, ферменты являются предпочтительными. Можно использовать, например, системы, основанные на пероксидазах, в частности пероксидазе хрена, щелочной фосфатазе и β-D-галактозидазе. Специфические субстраты, превращение которых можно мониторировать, например фотометрически, доступны для этих ферментов. Подходящие субстратные системы основаны на p-нитрофенилфосфате (p-NPP), 5-бром-4-хлор-3-индолилфосфате/нитросинем тетразолии (BCIP/NPT), прочном красном/нафтол-AS-TS фосфате для щелочной фосфатазы; 2,2-азинобис(3-этилбензотиазолин-6-сульфоновой кислоте) (ABTS), o-фенилендиамине (OPT), 3,3',5,5'-тетраметилбензидине (TMB), o-дианизидине, 5-аминосалициловой кислоте, 3-диметиламинобензойной кислоте (DMAB) и 3-метил-2-бензотиазолингидразоне (MBTH) для пероксидаз; o-нитрофенил-β-D-галактозиде (o-NPG), p-нитрофенил-β-D-галактозиде и 4-метилумбеллифенил-β-D-галактозиде (MUG) для β-D-галактозидазы. Во многих случаях эти субстратные системы являются коммерчески доступными в готовой к использованию форме, например в форме таблеток, которые могут также содержать дополнительные реагенты, такие как подходящие буферы и т.п..
Используемые индикаторы могут представлять собой меченые глобуломеры. В этом смысле, конкретный глобуломер можно определять посредством мечения подлежащего определению глобуломера и использования его в качестве индикатора.
Присоединение меток к глобуломерам для получения индикаторов можно выполнять способом, известным по существу. Комментарии выше относительно дериватизации глобуломеров по изобретению относятся к аналогии. Кроме того, доступен ряд меток, соответствующим образом модифицированных для конъюгации с белками, например, биотин-, авидин-, экстравидин- или стрептавидин-конъюгированные ферменты, малеинимид-активированные ферменты и т.п. Для получения индикатора можно проводить реакцию этих меток непосредственно с олигомером или, глобуломером. Если, например, используют конъюгат стрептавидин-пероксидаза, тогда это, во-первых, требует биотинилирования глобуломера. Это применяют, соответственно, в обратном порядке. Подходящие для этой цели способы также известны специалисту в данной области.
Если выбран формат гетерогенного иммуноанализа, комплекс антиген-антитело можно отделить посредством его связывания с подложкой, например через антиидиотипическое антитело, присоединенное к указанной подложке, например, антитело, направленное против IgG кролика. Подходящие подложки, в частности микропланшеты для титрования, покрытые соответствующими антителами, известны и частично коммерчески доступны.
Кроме того, настоящее изобретение относится к наборам для иммуноанализа, содержащим по меньшей мере одно антитело, как описано выше, и дополнительные компоненты. Указанные наборы обычно представлены в форме упакованной единицы, сочетания средств для проведения определения глобуломера по изобретению. Для целей настолько простых манипуляций, насколько возможно, указанные средства предпочтительно предоставляют в готовой к использованию форме. При предпочтительной компоновке иммуноанализ предлагают в форме набора. Набор обычно содержит множество контейнеров для отдельной компоновки компонентов. Все компоненты можно предоставлять в готовом к использованию растворе, в форме концентрата для разведения или сухого вещества или лиофилизата для растворения или суспендирования; отдельные или все компоненты можно замораживать или хранить при комнатной температуре до использования. Сыворотка предпочтительно является быстрозамороженной, например при -20°C, так что в этих случаях компоненты для иммуноанализа перед использованием необходимо хранить, предпочтительно при температурах ниже температуры замерзания.
Дополнительные компоненты, поставляемые с компонентами для иммуноанализа, зависят от типа указанного иммуноанализа. Обычно стандартный белок, индикатор, который может являться или не являться необходимым, и контрольную сыворотку поставляют вместе с антисывороткой. Кроме того, можно включать также микропланшеты для титрования, предпочтительно, покрытые антителом, буферы, например, для тестирования, для промывки или для превращения субстрата, и собственно субстрат для фермента.
Общие принципы иммуноанализов, а также получения и использования антител в качестве вспомогательных средств в лаборатории и лечебном учреждении можно найти, например, в Antibodies, A Laboratory Manual (Harlow, E., and Lane, D., Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1988).
Таким образом, настоящее изобретение также относится к способу диагностики амилоидоза, например болезни Альцгеймера, у пациента, предположительно страдающего данным заболеванием. Этот способ включает в себя стадии: 1) выделения биологического образца от пациента; 2) контактирования биологического образца по меньшей мере с одним из описанных выше антител в течение периода времени и в условиях, подходящих для формирования комплексов антиген/антитело; и 3) детекции присутствия комплексов антиген/антитело в указанном образце, где присутствие комплексов указывает на диагноз амилоидоз, например болезнь Альцгеймера, у пациента. Антиген может представлять собой, например, глобуломер или его часть, или фрагмент, обладающий теми же функциональными свойствами, что и полноразмерный глобуломер (например, активность связывания).
Кроме того, настоящее изобретение относится к другому способу диагностики амилоидоза, например болезни Альцгеймера, у пациента, предположительно страдающего данным заболеванием. Этот способ включает в себя стадии: 1) выделения биологического образца от пациента; 2) контактирования биологического образца с антигеном в течение периода времени и в условиях, подходящих для формирования комплексов антитело/антиген; 3) добавление конъюгата к полученным комплексам антитело/антиген на период времени и в условиях, подходящих, чтобы позволить конъюгату связаться со связанным антителом, где конъюгат содержит одно из описанных выше антител, присоединенное к производящему сигнал соединению, способному производить поддающийся детекции сигнал; 4) детекции присутствия антитела, которое может присутствовать в биологическом образце, посредством детекции сигнала, производимого производящим сигнал соединением, где сигнал указывает на диагноз амилоидоз, например болезнь Альцгеймера, у пациента. Антиген может представлять собой глобуломер или его часть или фрагмент, обладающие теми же функциональными свойствами, что и полноразмерный глобуломер (например, активность связывания).
Настоящее изобретение относится к дополнительному способу диагностики амилоидоза, например болезни Альцгеймера, у пациента, предположительно страдающего амилоидозом, например болезнью Альцгеймера. Этот способ включает в себя стадии: 1) выделения биологического образца от указанного пациента; 2) контактирования биологического образца с анти-антителом, где анти-антитело является специфическим для одного из описанных выше антител, в течение периода времени и в условиях, подходящих, чтобы позволить формирование комплексов анти-антитело/антитело, где комплексы содержат антитело, присутствующее в биологическом образце; 2) добавление конъюгата к полученным комплексам анти-антитело/антитело на период времени и в условиях, подходящих, чтобы позволить конъюгату связаться со связанным антителом, где конъюгат содержит антиген, связанный с производящим сигнал соединением, способным производить поддающийся детекции сигнал; 3) детекции сигнала, производимого производящим сигнал соединением, где сигнал указывает на диагноз амилоидоз, например болезнь Альцгеймера, у пациента.
Настоящее изобретение относится также к набору, содержащему: a) по меньшей мере одно из описанных выше антител и b) конъюгат, содержащий антитело, присоединенное к производящему сигнал соединению, где антитело из конъюгата отличается от выделенного антитела.
Настоящее изобретение относится также к набору, содержащему: a) анти-антитело против одного из описанных выше антител и b) конъюгат, содержащий антиген, присоединенный к производящему сигнал соединению. Антиген может представлять собой глобуломер или его фрагмент или часть, обладающие теми же функциональными характеристиками, что и полноразмерный глобуломер (например, активность связывания).
В одном из диагностических вариантов осуществления настоящего изобретения антителом по настоящему изобретению или его частью покрывают твердую фазу (или оно присутствует в жидкой фазе). Тогда тестируемый или биологический образец (например, цельную кровь, цереброспинальную жидкость, сыворотку и т.д.) приводят затем в контакт с твердой фазой. Если в образце присутствует антиген (например, глобуломер), такие антигены связываются с антителами на твердой фазе и затем их детектируют либо прямым, либо непрямым способом. Прямой способ включает в себя простую детекцию собственно комплекса и, таким образом, присутствия антигенов. По непрямому способу, к связанному антигену добавляют конъюгат. Конъюгат содержит второе антитело, которое связывается со связанным антигеном, присоединенным к производящему сигнал соединению или метке. Если второе антитело связывается со связанным антигеном, производящее сигнал соединение производит поддающийся измерению сигнал. Тогда такой сигнал указывает на присутствие антигена в тестируемом образце.
Примерами твердых фаз, применяемых для диагностических иммуноанализов, являются пористые и непористые материалы, частицы латекса, магнитные частицы, микрочастицы (см. патент США No. 5705330), бусины, мембраны, лунки для микротитрования и пластиковые пробирки. Выбор материала твердой фазы и способ мечения антигена или антитела, присутствующего в конъюгате, если желательно, определяют на основании характеристик производительности желательного формата анализа.
Как указано выше, конъюгат (или реагент-индикатор) содержит антитело (или, возможно, анти-антитело, в зависимости от анализа), присоединенное к производящему сигнал соединению или метке. Такое производящее сигнал соединение или «метку» можно детектировать само по себе или оно может реагировать с одним или несколькими дополнительными соединениями для получения поддающегося детекции продукта. Примеры производящих сигнал соединений включают в себя хромогены, радиоактивные изотопы (например, 125I, 131I, 32P, 3H, 35S и 14C), хемилюминесцентные соединения (например, акридин), частицы (видимые или флуоресцентные), нуклеиновые кислоты, комплексообразующие средства или катализаторы, такие как ферменты (например, щелочная фосфатаза, кислая фосфатаза, пероксидаза хрена, бета-галактозидаза и рибонуклеаза). В случае использования фермента (например, щелочной фосфатазы или пероксидазы хрена), добавление хромо-, флуоро- или люмогенного субстрата приводит к получению поддающегося детекции сигнала. Другие системы детекции, такие как флуоресценция с разрешением по времени, флуоресценция внутреннего отражения, амплификация (например, полимеразная цепная реакция) и рамановская спектроскопия, также являются применимыми.
Примеры биологических жидкостей, которые можно тестировать посредством описанных выше иммуноанализов, включают в себя плазму, цельную кровь, высушенную цельную кровь, сыворотку, цереброспинальную жидкость или водные или органически-водные экстракты тканей и клеток.
Настоящее изобретение относится также к способу детекции присутствия антител в тестируемом образце. Этот способ включает в себя стадии: (a) контактирования тестируемого образца, предположительно содержащего антитела, с анти-антителом, специфическим для антитела в образце от пациента, в течение периода времени и в условиях, подходящих, чтобы позволить формирование комплексов анти-антитело/антитело, где анти-антитело представляет собой антитело по настоящему изобретению, которое связывается с антителом в образце от пациента; (b) добавления конъюгата к полученным комплексам анти-антитело/антитело, где конъюгат содержит антиген (связывающийся с анти-антителом), присоединенный к производящему сигнал соединению, способному производить поддающийся детекции сигнал; (d) детекции присутствия антител, которые могут присутствовать в тестируемом образце, посредством детекции сигнала, производимого производящим сигнал соединением. Можно использовать контроль или калибратор, содержащий антитело против анти-антитела.
Наборы также входят в объем настоящего изобретения. Более конкретно, настоящее изобретение относится к наборам для определения присутствия антигенов (например, глобуломеров) у пациента, предположительно страдающего от болезни Альцгеймера или другого состояния, характеризующегося когнитивным нарушением. В частности, набор для определения присутствия антигенов в тестируемом образце содержит: a) антитело, как определено здесь, или его часть; b) конъюгат, содержащий второе антитело (обладающее специфичностью к антигену), присоединенное к производящему сигнал соединению, способному производить поддающийся детекции сигнал. Набор может содержать также контроль или калибратор, содержащий реагент, связывающийся с антигеном.
Настоящее изобретение относится также к набору для детекции антител в тестируемом образце. Набор может содержать: a) анти-антитело (например, одно из рассматриваеимого изобретения), специфическое для интересующего антитела, b) антиген или его часть, как определено выше. Можно включать также контроль или калибратор, содержащий реагент, связывающийся с антигеном. Более конкретно, набор может содержать: a) анти-антитело (такое как одно из настоящего изобретения), специфическое для антитела b) конъюгат, содержащий антиген (например, глобуломер), присоединенный к производящему сигнал соединению, способному производить поддающийся детекции сигнал. И в этом случае набор может также содержать контроль или калибратор, содержащий реагент, связывающийся с антигеном.
Набор может также содержать один контейнер, такой как флакон, бутылки или полоска, каждый контейнер с подготовленной твердой фазой, и другие контейнеры, содержащие соответствующие конъюгаты. Эти наборы могут также содержать флаконы или контейнеры других реагентов, необходимых для проведения анализа, таких как реагенты для отмывки, обработки и реагенты-индикаторы.
Следует также заметить, что рассматриваемое изобретение относится не только к полноразмерным антителам, описанным выше, но также к их частям или фрагментам, например, их части Fab. Кроме того, рассматриваемое изобретение относится к любому антителу, обладающему теми же свойствами, что и настоящие антитела, например, с точки зрения специфичности связывания, структуры и т.д.
Преимущества по изобретению:
Посредством иммунизации глобуломером Aβ(20-42) можно получить различные моноклональные антитела, отличающиеся по их толерантности или распознаванию для различных олигомеров Aβ(1-42) и олигомеров Aβ(X-42), как определяют посредством конкурентного дот-блоттинга, как описано выше. Это позволяет разработку антитела, направленного против усеченных по N-концу олигомеров Aβ, обладающего оптимальным соотношением между эффектом улучшения когнитивности, желательной специфичностью для других форм Aβ и минимальным профилем побочных эффектов. Неожиданно, Aβ(1-42) и глобуломеры Aβ(12-42), несмотря на содержание структурного элемента, являются только частично узнаваемыми антителами, полученными с использованием дополнительно усеченного глобуломера Aβ(20-42) в качестве антигена.
Посредством ограничения специфических олигомерных форм Aβ на основании структурного принципа (т.е. использования усеченных по N-концу глобуломеров Aβ), получили профиль антител, активная иммунизация которыми является высокоспецифической для олигомерных форм Aβ. То же самое остается в силе для моноклональных антител для использования для пассивной иммунизации. Преимущество такой специфической стратегии иммунизации (активной и пассивной) состоит в том, что она не будет вызывать иммунный ответ против мономеров Aβ, пептидов Aβ в состояниях фибриллярной агрегации или sAPPα. Это обеспечивает преимущество несколькими путями.
1) В форме нерастворимых бляшек Aβ пептиды Aβ в состояниях фибриллярной агрегации составляют главную часть полного пула пептидов Aβ в мозге при AD. Массовое высвобождение Aβ посредством растворения бляшек Aβ, индуцированного реакцией антител против Aβ с этими бляшками, следует рассматривать как неблагоприятное. Затем это массовое высвобождение Aβ будет пересекать гематоэнцефалический барьер, входить в кровоток и потенциально увеличивать риск микрокровоизлияний. Кроме того, в испытании ELAN та же самая стратегия иммунизации фибриллярными формами пептида Aβ потребовала прекращение испытания из-за 6% случаев дебюта менингоэнцефалита.
2) Иммунные ответы, направленные против мономерных форм пептидов Aβ, являются нежелательными, так как являлось возможным показать, что позднее они могут влиять на эффекты улучшения когнитивности.
3) Иммунные ответы, направленные против sAPPα, таким же образом являются нежелательными, так как они могут приводить к реакции против физиологически существующего белка-предшественника APP и, таким образом, к аутоиммунной реакции. Более того, показано также, что sAPPα также может влиять на эффекты улучшения когнитивности.
4) Ответ, направленный против васкулярного пептида Aβ в форме CAA, также следует прекращать, чтобы избежать нежелательного побочного эффекта микрокровоизлияний (антитела против центральной части Aβ, которые, кроме того, не связываются с пептидами Aβ, агрегированными в форме CAA, индуцируют меньше микрокровоизлияний по сравнению с антителами против N-конца, см. выше).
5) Антитела, специфически реагирующие с олигомерами Aβ, обладают более высокой биодоступностью по отношению к патофизиологически релевантным видам Aβ, так как они не будут связываться, например, с фибриллярным Aβ и, таким образом, становиться недоступными для терапевтического эффекта.
Информация по депонированию: гибридома, продуцирующая моноклональное антитело 5F7, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 20110, 01 декабря 2005 г. в рамках Будапештского договора и получила обозначение PTA-7241. Кроме того, гибридома, продуцирующая моноклональное антитело 10F11, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801, 01 декабря 2005 г. в рамках Будапештского договора и получила обозначение PTA-7239. Кроме того, гибридома, продуцирующая моноклональное антитело 4B7, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801 01 декабря 2005 г. в рамках Будапештского договора и получила обозначение PTA-7242, и гибридома, продуцирующая моноклональное антитело 7C6, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801, 01 декабря 2005 г. в рамках Будапештского договора и получила обозначение PTA-7240. Кроме того, гибридома, продуцирующая моноклональное антитело 6A2, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801, 28 февраля 2006 г. в рамках Будапештского договора и получила обозначение PTA-7409, и гибридома, продуцирующая моноклональное антитело 2F2, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801, 28 февраля 2006 г. в рамках Будапештского договора и получила обозначение PTA-7408. Гибридома, продуцирующая моноклональное антитело 4D10, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801, 28 февраля 2006 г. в рамках Будапештского договора и получила обозначение PTA-7405. Гибридома, продуцирующая моноклональное антитело 7E5, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801, 16 августа 2006 г. в рамках Будапештского договора и получила обозначение PTA-7809. Гибридома, продуцирующая моноклональное антитело 10C1, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801, 16 августа 2006 г. в рамках Будапештского договора и получила обозначение PTA-7810. Гибридома, продуцирующая моноклональное антитело 3B10, депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, Virginia 10801, 01 сентября 2006 г. в рамках Будапештского договора и получила обозначение PTA-7851. Все депонирования выполнены от имени Abbott Laboratories, 100 Abbott Park Road, Abbott Park, Illinois 60064 (US).
На чертежах
На фигуре 1 показаны эксклюзионные хроматограммы Aβ(1-42) и Aβ(1-40). Мономер Aβ(1-42) растворяли в A) 0,1% NH4OH, B) 70% муравьиной кислоте C) 0,1% NaOH и в D) Aβ(1-40) растворяли в 0,1% NaOH. Затем образцы дополнительно разводили 1:10 в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4. Эти образцы инкубировали в течение 5 мин (левая колонка) или 1 часа (правая колонка) после растворения при температуре окружающей среды, затем наносили на колонку для эксклюзионной хроматографии;
На фигуре 2 A) показан SDS PAGE стандартных белков (белки маркера молекулярной массы, дорожка 1); препарата фибрилл Aβ(1-42); контроля (дорожка 2); препарата фибрилл Aβ(1-42) + mAb 5F7, 20 час, 37°C, супернатанта (дорожка 3); препарата фибрилл Aβ(1-42) + mAb 5F7, 20 час, 37°C, осадка (дорожка 4); препарата фибрилл Aβ(1-42) + mAb 6E10, 20 час, 37°C, супернатанта (дорожка 5); препарата фибрилл Aβ(1-42) + mAb 6E10, 20 час, 37°C, осадка (дорожка 6);
На B) показаны результаты количественного анализа mAb, связанных с фибриллами Aβ в процентах от общего антитела;
На фигуре 3 представлена гистограмма, где показаны результаты теста распознавания объекта трансгенными мышами APP/L после активной иммунизации мономерами Aβ(1-42) в 0,1% NH4OH, глобуломерами Aβ(1-42) и глобуломерами Aβ(20-42) по сравнению с мышами дикого типа (положительный контроль) и мышами APP/L, обработанными PBS (отрицательный контроль), где кругами показаны значимые различия с обработанными PBS мышами APP/L, а звездочками показаны высокозначимые различия со случайным уровнем (50%) согласно вторичному t-тесту после P<0,05 в ANOVA для различий между группами;
На фигуре 4 показаны дот-блоты для реактивности 100 пмоль/мкл (ряд A); 10 пмоль/мкл (ряд B); 1 пмоль/мкл (ряд C); 0,1 пмоль/мкл (ряд D) и 0,01 пмоль/мкл (ряд E) глобуломера Aβ(1-42) (столбец 1); предварительно обработанного HFIP мономера Aβ(1-42) в Плюроник F68 (столбец 2); глобуломера Aβ(20-42) (столбец 3); глобуломера Aβ(12-42) (столбец 4); предварительно обработанного HFIP мономера Aβ(1-40) в DMSO (столбец 5); мономера Aβ(1-42), NH4OH (столбец 6); препарата фибрилл Aβ(1-42) (столбец 7); и sAPPα из Sigma (столбец 8) с различными антисыворотками, полученными после активной иммунизации мышей APP/L Tg глобуломером Aβ(20-42);
На фигуре 5 показана гистограмма, где показаны концентрации растворимого и нерастворимого Aβ(1-42) и Aβ(1-40) пептида в экстрактах мозга APP/PS1 Tg-мышей, активно иммунизированных либо мономером Aβ(1-42) (0,1% NH4OH), либо глобуломером Aβ(1-42), либо глобуломером Aβ(20-42), либо носителем в качестве контроля;
На фигуре 6 показана гистограмма, где показаны результаты теста распознавания объекта трансгенными мышами APP/L после пассивной иммунизации антителами 5F7, 10F11 и 7C6 против глобуломера Aβ(20-42) по сравнению с контрольными мышами для A) каждого антитела отдельно и B) для всех антител, взятых вместе;
На фигуре 7 A) показан анализ дот-блоттингом специфичности различных антител против Aβ (-6E10, -5F7, -4B7, -10F11, -6A2, -4D10, -3B10, -2F2, -7C6, -7E5, -10C1). Моноклональные антитела, тестированные здесь, получены посредством активной иммунизации мышей глобуломером Aβ(20-42) с последующим отбором слитых клеток гибридомы (за исключением коммерчески доступного 6E10, Signet No 9320). Выделенные формы Aβ применяли в серийных разведениях и инкубировали с соответствующими моноклональными антителами для иммунной реакции:
1) мономер Aβ(1-42), 0,1 % NH4OH;
2) мономер Aβ(1-40), 0,1% NH4OH;
3) мономер Aβ(1-42), 0,1% NaOH;
4) мономер Aβ(1-40), 0,1% NaOH;
5) глобуломер Aβ(1 -42);
6) глобуломер Aβ(12-42);
7) глобуломер Aβ(20-42);
8) препарат фибрилл Aβ(1-42);
9) sAPPα (Sigma) (первая точка: 1 пмоль).
B) Количественную оценку проводили с использованием денситометрического анализа интенсивности. Для каждой формы Aβ оценивали только одну точку, соответствующую только самой низкой концентрации антигена, при условии, что она обладала относительной плотностью большей, чем 20% от относительной плотности последней однозначно определенной точки глобуломера Aβ(20-42) (порог). Значение порога определяли для каждого дот-блота независимо. Значение указывает на соотношение между узнаванием глобуломера Aβ(20-42) и соответствующей формой Aβ для данного антитела;
На фигуре 8 показано связывание антител в различных концентрациях с поперечными срезами неокортекса пациентов с болезнью Альцгеймера (AD) или старых трансгенных мышей APP:
A) Верификация отложения амилоида посредством окрашивания Конго красным в форме бляшек в ткани мозга и в форме церебральной амилоидной ангиопатии (CAA) в сосудах мозга трансгенной мыши APP линии Tg2576 и пациента с AD (RZ55);
B) Сильное окрашивание паренхимальных отложений Aβ (амилоидных бляшек) у пациента с AD (RZ16) присутствует только с 6G1 и с коммерчески доступным антителом 6E10 (левый столбец), в то время как для антител 5F7, 2F2 и 6A2 (второй столбец), 4D10, 10F11 и 3B10 (третий столбец) и 7C6, 7E5 и 10C1 (правый столбец) не показали окрашивания. Все антитела использовали в концентрации 0,7 мкг/мл;
C) Сильное окрашивание паренхимальных отложений Aβ (амилоидных бляшек) у Tg2576 в возрасте 19 месяцев присутствовало только с 6G1 и с коммерчески доступным антителом 6E10 (левый столбец), в то время как для антител 5F7, 2F2, и 6A2 (второй столбец), 4D10, 10F11 и 3B10 (второй столбец) и 7C6, 7E5, и 10C1 (правый столбец) не показали окрашивания. Все антитела использовали в концентрации 0,7 мкг/мл;
D)-G) Количественная оценка анализа окрашивания бляшек Aβ на гистологических изображениях с использованием анализа изображений. Значения оптической плотности (0% = отсутствие окрашивания) рассчитывали по значениям для бляшек в шкале серого, из которых вычитали значения для фоновой ткани в шкале серого: D) Окрашивание 0,7 мкг/мл антитела для старых мышей Tg2576, E) окрашивание 3 различными концентрациями антител для мышей APP/L, F) окрашивание 0,7 мкг/мл антитела для пациента с AD (RZ55), G) окрашивание 3 различными концентрациями антител для пациента с AD (RZ16). Различие между окрашиванием коммерчески доступными антителами 6E10 (звездочки) и 4G8 (круги) и всеми другими антителами (три звездочки/круга: p<0,001 по сравнению с контролем; вторичный t-тест Бонферрони после ANOVA с p<0,001) оценивали статистически (D1F). В E) и G) для всех антител, за исключением 6G1 всегда показывали значимо меньшее окрашивание, чем для коммерчески доступных антител 6E10 и 4G8 (p<0,001 во вторичном t-тесте после p<0,001 в ANOVA). H) Сильное окрашивание отложений Aβ в сосудах (стрелки) присутствует только с 6G1 и коммерчески доступным антителом 6E10 (левый столбец), в то время как для антител 5F7, 2F2 и 6A2 (второй столбец), 4D10, 10F11 и 3B10 (третий столбец) и 7C6, 7E5, и 10C1 (правый столбец) не показали окрашивания. Все антитела использовали в концентрации 0,7 мкг/мл. Количественно сходную ситуацию обнаружили для мышей Tg2576 (здесь не показано);
Фигура 9. Титр антитела против Aβ и профиль избирательности при дот-блоттинге в плазме мышей TG2576 спустя приблизительно один год после активной иммунизации. В образцах плазмы мышей Tg2576 спустя приблизительно один год после последней иммунизации A) глобуломером Aβ (20-42), B) глобуломером Aβ (12-42), C) мономером Aβ (1-42) и D) носителем, посредством дот-блоттинга оценивали, что антитела против Aβ продуцированы и еще присутствуют.
1) глобуломер Aβ (1-42);
2) мономер Aβ (1-42), предварительно обработанный HFIP, в 0,1% Плюроник F68;
3) глобуломер Aβ (20-42);
4) глобуломер Aβ (12-42);
5) мономер Aβ (1-40), предварительно обработанный HFIP, 5 мМ в DMSO;
6) мономер Aβ (1-42), 0,1% NH4OH;
7) препарат фибрилл Aβ (1-42);
8) sAPPα (Sigma); (первая точка: 1 пмоль).
На фигуре 10 показана таблица, где суммированы уровни глобуломера Aβ(20-42) в ткани мозга людей, страдающих болезнью Альцгеймера, и контроля без деменции;
На фигуре 11 показаны нуклеотидные и аминокислотные последовательности вариабельных тяжелых и легких цепей моноклональных антител (mAb) следующим образом (определяющие комплементарность области (CDR) подчеркнуты в каждой аминокислотной последовательности):
Следующие примеры предназначены для иллюстрирования изобретения, без ограничения его объема.
Пример 1: Получение глобуломеров
a) глобуломер Aβ(1-42)
Синтетический пептид Aβ(1-42) (H-1368, Bachem, Bubendorf, Switzerland) суспендировали в 100% 1,1,1,3,3,3-гексафтор-2-пропаноле (HFIP) при 6 мг/мл и инкубировали для полного растворения при покачивании при 37°C в течение 1,5 ч. HFIP действует как разрушитель водородных связей, и его используют для удаления предсуществующей структурной негомогенности в пептиде Aβ. HFIP удаляли выпариванием в Speed Vac и Aβ(1-42) ресуспендировали до концентрации 5 мМ в диметилсульфоксиде и обрабатывали ультразвуком в течение 20 с. Предварительно обработанный HFIP Aβ(1-42) разводили в фосфатно-солевом буфере (PBS) (20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4) до 400 мкМ и добавляли 1/10 объема 2% додецилсульфата натрия (SDS) (в H2O) (конечная концентрация 0,2% SDS). Инкубацией в течение 6 ч при 37°C получали промежуточный глобуломер Aβ(1-42) 16/20-кДа (короткую форму глобулярного олигомера). Глобуломер Aβ(1-42) 38/48-кДа получали дополнительном разведением тремя объемами H2O и инкубацией в течение 18 ч при 37°C. После центрифугирования при 3000g в течение 20 мин образец концентрировали ультрафильтрацией (отсечение 30-кДа), диализовали против 5 мМ NaH2PO4, 35 мМ NaCl, pH 7,4, центрифугировали при 10000 g в течение 10 мин и отбирали супернатант, содержащий 38/48-кДа глобуломер Aβ(1-42). В качестве альтернативы диализу, 38/48-кДа глобуломер Aβ(1-42) можно также осаждать девятикратным избытком (об./об.) ледяного раствора метанол/уксусная кислота (33% метанол, 4% уксусная кислота) в течение 1 ч при 4°C. Затем 38/48-кДа глобуломер Aβ(1-42) осаждали (10 мин при 16200g), ресуспендировали в 5 мМ NaH2PO4, 35 мМ NaCl, pH 7,4 и доводили pH до 7,4.
b) Перекрестно-сшитый глобуломер Aβ(1-42)
Синтетический пептид Aβ(1-42) (H-1368, Bachem, Bubendorf, Switzerland) суспендировали в 100% 1,1,1,3,3,3-гексафтор-2-пропаноле (HFIP) при 6 мг/мл и инкубировали для полного растворения при покачивании при 37°C в течение 1,5 ч. HFIP действует как разрушитель водородных связей, и его используют для удаления предсуществующей структурной негомогенности в пептиде Aβ. HFIP удаляли выпариванием в SpeedVac и Aβ(1-42) ресуспендировали до концентрации 5 мМ в диметилсульфоксиде и обрабатывали ультразвуком в течение 20 с. Предварительно обработанный HFIP Aβ(1-42) разводили в фосфатно-солевом буфере (PBS) (20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4) до 400 мкМ и добавляли 1/10 объема 2% додецилсульфата натрия (SDS) (в H2O) (конечная концентрация 0,2% SDS). Инкубацией в течение 6 ч при 37°C получали промежуточный глобуломер 16/20-кДа Aβ(1-42) (короткую форму глобулярного олигомера). 38/48-кДа глобуломер Aβ(1-42) получали дополнительным разведением тремя объемами H2O и инкубацией в течение 18 ч при 37°C. Теперь проводили перекрестное сшивание 38/48-кДа глобуломера Aβ(1-42) инкубацией с 1 мМ глутаральдегидом в течение 2 ч при комнатной температуре (RT) 21°C с последующей обработкой этаноламином (5 мМ) в течение 30 мин при RT.
c) глобуломер Aβ(20-42)
1,59 мл препарата глобуломера Aβ(1-42), полученного согласно примеру 1a, смешивали с 38 мл буфера (50 мМ MES/NaOH, pH 7,4) и 200 мкл раствора термолизина 1 мг/мл (Roche) в воде. Реакционную смесь перемешивали при RT в течение 20 ч. Затем добавляли 80 мкл 100 мМ раствора ЭДТА, pH 7,4, в воде и, кроме того, доводили содержание SDS в смеси до 0,01% с помощью 400 мкл раствора SDS концентрации 1%. Реакционную смесь концентрировали приблизительно до 1 мл посредством пробирки Centriprep 15 мл 30 кДа. Концентрат смешивали с 9 мл буфера (50 мМ MES/NaOH, 0,02% SDS, pH 7,4) и опять концентрировали до 1 мл. Концентрат диализовали при 6°C против 1 л буфера (5 мМ фосфат натрия, 35 мМ NaCl) в диализной трубке в течение 16 ч. Затем доводили содержание SDS в диализате до 0,1% с помощью раствора SDS в воде с концентрацией 2%. Образец центрифугировали при 10000g в течение 10 мин, и отбирали супернатант глобуломера Aβ(20-42).
d) глобуломер Aβ(12-42)
2 мл препарата глобуломера Aβ(1-42), полученного согласно примеру 1a смешивали с 38 мл буфера (5 мМ фосфат натрия, 35 мМ хлорид натрия, pH 7,4) и 150 мкл 1 мг/мл эндопротеазы GluC (Roche) в воде. Реакционную смесь перемешивали в течение 6 ч при RT, и затем добавляли дополнительные 150 мкл 1 мг/мл эндопротеазы GluC (Roche) в воде. Реакционную смесь перемешивали при RT в течение 16 ч, с последующим добавлением 8 мкл 5 M раствора DIFP. Реакционную смесь концентрировали приблизительно до 1 мл посредством пробирки Centriprep 15 мл 30 кДа Centriprep. Концентрат смешивали с 9 мл буфера (5 мМ фосфат натрия, 35 мМ хлорид натрия, pH 7,4) и опять концентрировали до 1 мл. Концентрат диализовали при 6°C против 1 л буфера (5 мМ фосфат натрия, 35 мМ NaCl) в диализной трубке в течение 16 ч. Затем доводили содержание SDS до 0,1% с помощью раствора SDS в воде с концентрацией 1%. Образец центрифугировали при 10000g в течение 10 мин и отбирали супернатант глобуломера Aβ(12-42).
Пример 2: Эксклюзионная хроматография различных препаратов мономера Aβ(1-42) и мономера Aβ(1-40)
Aβ(1-42), 0,1% NH4OH:
1 мг Aβ(1-42) (Bachem, каталожный № H-1368) растворяли в 500 мкл 0,1% NH4OH в H2O и перемешивали 1 мин при температуре окружающей среды. Образец центрифугировали в течение 5 мин при 10000g. Собирали супернатант. Концентрацию Aβ(1-42) в супернатанте определяли согласно способу Брэдфорд (BIO-RAD).
Образец для 5 мин:
20 мкл супернатанта, содержащего Aβ(1-42) в 0,1% NH4OH, разводили 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 до концентрации Aβ(1-42) 0,2 мг/мл. Образец инкубировали в течение 5 мин при температуре окружающей среды. Затем 100 мкл анализировали эксклюзионной хроматографией (SEC).
Образец для 1 часа:
20 мкл супернатанта, содержащего Aβ(1-42) в 0,1% NH4OH, растворяли в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 до концентрации Aβ(1-42) 0,2 мг/мл. Образец инкубировали в течение 1 часа при температуре окружающей среды. Затем 100 мкл анализировали эксклюзионной хроматографией (SEC).
Aβ(1-42), 70% HCOOH:
1 мг Aβ(1-42) растворяли в 50 мкл 70% HCOOH в H2O и перемешивали 1 мин при температуре окружающей среды. Образец центрифугировали 5 мин при 10000 g. Собирали супернатант. Концентрацию Aβ(1-42) в супернатанте определяли согласно способу Брэдфорд (BIO-RAD).
Образец для 5 мин:
2 мкл Aβ(1-42) в 70% HCOOH разводили до концентрации 0,2 мг/мл Aβ(1-42) с помощью 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 и доводили pH до 7,4 с помощью 1 M NaOH. Образец инкубировали в течение 5 мин при температуре окружающей среды. Затем 100 мкл анализировали эксклюзионной хроматографией.
Образец для 1 часа:
2 мкл Aβ(1-42) в 70% HCOOH разводили до концентрации 0,2 мг/мл Aβ(1-42) с помощью 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 и доводили pH до 7,4 с помощью 1 M NaOH. Образец инкубировали в течение 1 часа при температуре окружающей среды. Затем 100 мкл анализировали эксклюзионной хроматографией.
Aβ(1-42), 0,1% NaOH:
1 мг Aβ(1-42) (Bachem, каталожный № H-1368) растворяли в 500 мкл 0,1% NaOH в H2O и перемешивали 1 мин при температуре окружающей среды. Образец центрифугировали в течение 5 мин при 10000g. Собирали супернатант. Концентрацию Aβ(1-42) в супернатанте определяли согласно способу Брэдфорд (BIO-RAD).
Образец для 5 мин:
20 мкл Aβ(1-42) в 0,1 % NaOH разводили до концентрации Aβ(1-42) 0,2 мг/мл с помощью 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4. Образец инкубировали в течение 5 мин при температуре окружающей среды. Затем 100 мкл анализировали эксклюзионной хроматографией.
Образец для 1 часа:
20 мкл Aβ(1-42) в 0,1% NaOH разводили до концентрации 0,2 мг/мл Aβ(1-42) с помощью 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4. Образец инкубировали в течение 1 часа при температуре окружающей среды. Затем 100 мкл анализировали эксклюзионной хроматографией.
Aβ(1-40), 0,1% NaOH:
1 мг Aβ(1-40) (Bachem, каталожный № H-1194) разводили в 500 мкл 0,1% NaOH в H2O и перемешивали 1 мин при температуре окружающей среды. Затем образец центрифугировали в течение 5 мин при 10000g. Собирали супернатант. Концентрацию Aβ(1-42) в супернатанте определяли согласно способу Брэдфорд (BIO-RAD).
Образец для 5 мин:
20 мкл Aβ(1-40) в 0,1 % NaOH разводили до концентрации 0,2 мг/мл Aβ(1-40) с помощью 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4. Образец инкубировали в течение 5 мин при температуре окружающей среды. Затем 100 мкл анализировали эксклюзионной хроматографией.
Образец для 1 часа:
20 мкл Aβ(1-40) в 0,1% NaOH разводили до концентрации 0,2 мг/мл Aβ(1-40) с помощью 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4. Образец инкубировали в течение 1 часа при температуре окружающей среды. Затем 100 мкл анализировали эксклюзионной хроматографией.
Условия эксклюзионной хроматографии (SEC):
Колонка SEC:
Superose 12 HR 10/300 GL (Amersham, каталожный № 17-5173-01)
Проток: 0,5 мл/мин
Загрузка бумаги: 0,2 см/мин
Поглощение при 214 нм: 0 - 0,2 единицы поглощения
Подвижная фаза: 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4
Результаты показаны на фигуре 1.
Получение раствора чистого мономерного Aβ является сложной задачей из-за сильной тенденции пептида Aβ, особенно мономера Aβ(1-42), агрегировать в фибриллы. Тем не менее, для скрининга и характеризации антиглобуломеров Aβ(20-42), различающих мономеры Aβ(1-42) и мономеры Aβ(1-40), следует использовать наилучший из технически достижимых препаратов мономерного Aβ. Здесь тестировали эффект первоначального растворителя пептида Aβ на эффект агрегации после дальнейшего растворения в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4. Поставщик пептида Aβ (Bachem) указывает в своей технической информации, что Aβ(1-42) следует растворять в 0,1% NH4OH. Через пять минут при комнатной температуре (RT) после растворения Aβ(1-42) в NH4OH и немедленного дальнейшего разведения 1:10 в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 эксклюзионной хроматографией показали первые признаки агрегации Aβ(1-42) до предшественников фибрилл с минорным пиком 74кДа. Мономерный Aβ(1-42) проходит как главный пик 11 кДа и плечо 6кДа. После инкубации в течение одного часа при комнатной температуре (RT) пептид Aβ(1-42) в NH4OH уже агрегирует до высокого содержания фибрилл Aβ(1-42), что приводит к потере подлежащего детекции материала, который не входит в колонку для эксклюзионной хроматографии. Если 70% муравьиную кислоту используют в качестве первоначального растворителя для пептида Aβ(1-42), агрегация до высокой степени спустя 1 час при RT происходит только для минорной фракции оставшегося мономера Aβ(1-42) (следует заметить, что муравьиная кислота сама по себе приводит к высокому фоновому поглощению при длине волны для детекции белка). Наилучшим первоначальным растворителем для Aβ(1-42) для предупреждения агрегации является 0,1% NaOH, для которого даже спустя 1 час инкубации при растворении и дальнейшем разведении показали только минорную фракцию агрегированного Aβ(1-42) при большинстве Aβ(1-42), еще остающимся мономерным. Для Aβ(1-40), первоначально растворенного в 0,1% NaOH, вообще не показали признаков агрегации даже после 1 час инкубации при RT.
Пример 3: Полуколичественный анализ дискриминации фибрилл Aβ(1-42) антителами, избирательными для глобуломера Aβ(20-42), с визуализацией в SDS-PAGE
Получение фибрилл Aβ(1-42):
1 мг Aβ(1-42) (Bachem, кат. no.: H-1368) растворяли в 500 мкл 0,1% NH4OH в H2O и перемешивали в течение 1 мин при температуре окружающей среды. Образец центрифугировали в течение 5 мин при 10000 g. Собирали супернатант. Концентрацию Aβ(1-42) в супернатанте определяли согласно способу Брэдфорд (BIO-RAD).
100 мкл Aβ(1-42) в 0,1% NH4OH смешивали с 300 мкл 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 и доводили pH до 7,4 с помощью 2% HCl. Затем образец инкубировали при 37°C в течение 20 часов. После этого образец центрифугировали в течение 10 мин при 10000 g. Супернатант сливали, и осадок смешивали с 400 мкл 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, ресуспендировали энергичным перемешиванием («встряхиванием») в течение 1 мин и центрифугировали 10 мин при 10000g. Супернатант сливали, и осадок смешивали с 400 мкл 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, ресуспендировали энергичным перемешиванием («встряхиванием») в течение 1 мин и центрифугировали 10 мин при 10000g еще раз. Супернатант сливали. Осадок ресуспендировали в 380 мкл 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 и взбалтывали энергичным перемешиванием («встряхиванием»).
Связывание антител против Aβ с фибриллами Aβ(1-42):
40 мкл препарата фибрилл Aβ(1-42) разводили 160 мкл 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% Tween 20, pH 7,4 и перемешивали 5 мин при температуре окружающей среды, затем образец центрифугировали 10 мин при 10000g. Супернатант сливали, и осадок ресуспендировали в 95 мкл 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% Tween 20, pH 7,4. Повторную суспензию взбалтывали энергичным перемешиванием («встряхиванием»).
Каждую аликвоту по 10 мкл препарата фибрилл смешивали с:
a) 10 мкл 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4;
b) 10 мкл 0,5 мкг/мкл 5F7 в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4;
c) 10 мкл 0,5 мкг/мкл 6E10 (Signet Nr.: 9320) в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4.
Образцы инкубировали при 37°C в течение 20 часов, затем центрифугировали 10 мин при 10000g. Супернатанты собирали и смешивали с 20 мкл буфера для образцов для SDS-PAGE. Остатки смешивали с 50 мкл 20 мМ NaH2PO4, 140 мМ NaCl, 0,025% Tween 20, pH 7,4 и ресуспендировали «встряхиванием», затем образцы центрифугировали 10 мин при 10000 g. Супернатанты сливали и осадки смешивали с 20 мкл 20 мМ NaH2PO4, 140 мМ NaCl, 0,025% Tween 20, pH 7,4, затем с 20 мкл буфера для образцов для SDS-PAGE. Образцы наносили на 4-20% Трис/глициновый гель для электрофореза.
Параметры для SDS-PAGE:
Буфер для образцов с SDS: 0,3 г SDS
4 мл 1 M Трис/HCl pH 6,8
8 мл глицерина
1 мл 1% бромфенолового синего в этаноле
Дополнить H2O до 50 мл
4-20% Трис/глициновый гель: (Invitrogen, кат. no.: EC6025BOX)
Буфер для электрофореза: 7,5 г Трис
36 г глицина
2,5 г SDS
Дополнить H2O до 2,5 л
Электрофорез в геле выполняли при постоянной силе тока 20 мА.
Окрашивание гелей: Кумасси синий R250
Результаты показаны на фигуре 2.
Полуколичественный анализ различных антител Aβ и их разделения фибрилл Aβ(1-42). Положения антител, фибрилл Aβ(1-42) и мономеров Aβ(1-42) помечены по краю геля. Из-за их размера фибриллы Aβ(1-42) не могут входить в гель для SDS-PAGE, и их можно видеть в лунке геля.
1. Маркер.
2. Препарат фибрилл Aβ(1-42); контроль.
3. Препарат фибрилл Aβ(1-42); + mAb 5F7; 20 час 37°C; супернатант.
4. Препарат фибрилл Aβ(1-42); + mAb 5F7; 20 час 37°C; осадок.
5. Препарат фибрилл Aβ(1-42); + mAb 6E10; 20 час 37°C; супернатант.
6. Препарат фибрилл Aβ(1-42); + mAb 6E10; 20 час 37°C; осадок.
Относительное связывание с типом фибрилл Aβ оценивали по анализу в SDS-PAGE посредством измерения значений оптической плотности (OD) для тяжелых цепей антител в связанной с фибриллами (фракции осадка) и фракциях супернатанта после центрифугирования. Антитела, которые связываются с фибриллами Aβ, должны осаждаться с фибриллами Aβ и, таким образом, их обнаруживают во фракциях осадка, в то время как не связанные с фибриллами Aβ (свободные) антитела обнаруживают в супернатанте. Процент антитела, связанного с фибриллами Aβ, рассчитывают по следующей формуле:
Процент антитела, связанного с фибриллами Aβ =
ODфракции фибрилл × 100%/(ODфракции фибрилл + OD фракции супернатанта).
Этот способ выполняли для mAb 6E10 (Signet, кат. no.: 9320), 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1.
В мозге при болезни Альцгеймера фибриллы Aβ представляют собой главный компонент общего пула пептидов Aβ. Из-за атаки этих фибрилл антителами против Aβ возрастает риск отрицательных побочных эффектов из-за высвобождения больших количеств Aβ, которые затем могут увеличивать риск микрокровоизлияний. Увеличение риска микрокровоизлияний наблюдали для способа активной иммунизации фибриллярными агрегатами пептида Aβ (Bennett and Holtzman, 2005, Neurology, 64, 10-12; Orgogozo J, Neurology, 2003, 61,46-54; Schenk et al., 2004, Curr Opin Immunol, 16, 599-606).
В отличие от коммерчески доступного антитела 6E10 (Signet 9320), распознающего линейный Aβ-эпитоп между AA1-17, избирательное для глобуломера Aβ(20-42) антитело 5F7 (которое в действительности обладает самой низкой избирательностью для глобуломеров Aβ(20-42) по сравнению с другими формами Aβ) не связывается с фибриллами Aβ(1-42) в эксперименте соосаждения. Это показали по тому факту, что антитело 5F7 после инкубации с фибриллами Aβ(1-42) после стадии осаждения остается в супернатанте и не соосаждается из-за связывания с фибриллами Aβ(1-42).
Пример 4: Анализ когнитивного поведения у мышей посредством теста распознавания объекта после активной иммунизации мономером Aβ(1-42) (0,1% NH 4 OH), глобуломером Aβ(1-42) или глобуломером Aβ(20-42) по сравнению с диким типом.
В этих экспериментах использовали мышей, сверэкспрессирующих APP человека с точечной мутацией. Точечная мутация относится к аминокислоте 717 (замене валина на изолейцин) и обнаружена в семье из Лондона, где она приводила к дебюту AD до начала шестого десятилетия жизни (Mullan et al., Nature Genetics 2 (1992) 340-342). Трансгенных мышей, обозначенных здесь как APP/L, получили и впервые описали в Leuven (Moechars et al., J. Biol. Chem. 274 (1999) 6483-6492). Самок мышей APP/L подвергали активной иммунизации в возрасте 6 недель.
Мышам вводили 100 мкг мономера Aβ(1-42) (0,1 % NH4OH), или глобуломера Aβ(1-42), или глобуломера Aβ(20-42) в фосфатно-солевом буфере (PBS), смешанных с равным количеством полного адъюванта Фрейнда внутрибрюшинно, с последующими бустер-инъекциями того же количества антигена в неполном адъюванте Фрейнда каждую третью неделю в течение трех месяцев. В течение периода времени эксперимента мышей содержали в стандартных условиях с обращенным циклом день/ночь (14 часов света, начиная с 19-00/10 часов темноты). Увеличение массы тела в течение времени эксперимента являлось таким, как ожидали, и не отличалось от контрольной группы, которой вводили только PBS/адъювант, что позволяет предполагать хорошую переносимость обработок антигеном.
В возрасте 4,5 месяцев когнитивную способность мышей тестировали посредством теста распознавания объектов, как описано в данной области (Dewachter et al. Journal of Neuroscience 22 (2002) 3445-3453). С этой целью мышей приучали к площадке и затем в течение 10 минут подвергали фазе обучения, в течение которой их по отдельности помещали на площадку, теперь содержащую два одинаковых элемента (голубые пирамиды, зеленые кубики, желтые цилиндры сходного размера, приблизительно 4 см). Регистрировали продолжительность и частоту, с которой мышь исследовала объекты. Во время фазы запоминания, 2,5 час спустя, мышей возвращали на площадку, теперь содержащую, в дополнение к известному объекту, неизвестный объект, случайно выбранный из других объектов. Распознавание нового объекта регистрировали как время, в течение которого мышь исследовала новый объект, по отношению к общему времени (исследование старого и нового объекта). «Показатель распознавания» выражает это отношение (время для нового объекта/общее время). Мышь, которая не запоминает известный объект, будет считать его таким же интересным, как новый объект, и тратить такое же количество времени на его исследование, иными словами, обладать показателем распознавания 50%. Мышь, которая запоминает известный объект, будет считать его неинтересным и, таким образом, обладать значительно более высоким показателем распознавания. Известно, что мыши APP/L обладают когнитивными нарушениями в возрасте 4,5 месяцев и обладают показателем распознавания при измерении на уровне случайного, т.е. 50%.
Результаты показаны на фигуре 3.
Тест распознавания объектов для мышей. Тест описывает распознавание известного объекта по сравнению с неизвестным объектом, измеренное в переводе на исследовательское поведение в течение 10 минутной фазы тестирования. Показатель распознавания определяют как процент времени, которое мышь тратит на исследование неизвестного объекта относительно времени, потраченного на исследование обоих объектов. Мышь исследовала известный объект в течение 10 минутной фазы обучения за три часа перед фазой тестирования. Сравнивали пять групп мышей (номер n дан под столбцами). Нормальные мыши C57BI/6 (дикий тип) обладали высоким R1, значимо отличающимся от случайного уровня (50%, т.е. равное время исследования как известного, так и неизвестного объекта) (*** = p < 0,001; t-тест Стьюдента). Четыре другие группы трансгенных мышей APP подвергали активной иммунизации тремя месяцами ранее. Используемые иммуногены представляли собой мономер Aβ(1-42), глобуломер Aβ(1-42) и глобуломер Aβ(20-42). Фосфатно-солевой буфер (PBS) использовали в качестве контроля. Значимые различия между PBS и другими группами указаны кругами: ° = p < 0,05; °° = p < 0,01 (вторичный t-тест после p < 0,05 в ANOVA).
Известно, что мыши APP/L обладают, в отличие от нетрансгенных мышей, когнитивными нарушениями в возрасте 4,5 месяцев, с оценкой результатов близко к случайному уровню (т.е. 50% показатель распознавания). Фактически, обработанные PBS мыши обладают случайным поведением, в отличие от нетрансгенных мышей (дикий тип). Иммунизация нативным глобуломером Aβ(1-42), так же как глобуломером Aβ(20-42), приводит к значительно улучшенному распознаванию объектов у мышей APP/L.
Так как оба препарата глобуломера (нативного и усеченного) приводят к улучшению памяти у трансгенных животных APP и даже превосходящему распознаванию у животных, обработанных глобуломером Aβ(20-42), является разумным заключить, что индукция антител против усеченного глобуломера Aβ(20-42) будет приводить к лучшим результатам и что пассивная иммунизация антителами, специфически реагирующими с этими видами, представляет собой оптимальную стратегию лечения.
Пример 5: Дот-блот анализ профиля антител для различных форм Aβ после активной иммунизации мышей APP/L Tg глобуломером Aβ(20-42).
После иммунизации мышей (сравни с примером 4) APP/L (Moechars et al., 1999, J. Biol. Chem. 274, 6483-6492) различными формами Aβ, в образцах плазмы оценивали антитела против Aβ. С этой целью получали серийные разведения выделенных форм Aβ(1-42) в диапазоне от 100 пмоль/мкл до 0,01 пмоль/мкл в PBS, дополненном 0,2 мг/мл BSA. По 1 мкл каждого образца наносили точками на нитроцеллюлозную мембрану. Для детекции использовали соответствующие образцы плазмы мыши (разведенные 1:400). Иммуноокрашивание выполняли с использованием щелочной фосфатазы, конъюгированной с антителами против IgG мыши и окрашивающего реагента NBT/BCIP.
Стандарты Aβ для дот-блоттинга:
1. глобуломер Aβ(1-42)
Получение глобуломера Aβ(1-42) описано в примере 1a.
2. Предварительно обработанный HFIP мономер Aβ(1-42) в Плюроник F68
3 мг Aβ(1-42), (Bachem Inc.; кат. no. H-1368) растворяли в 0,5 мл HFIP (суспензия 6 мг/мл) в 1,7 мл пробирке Eppendorff и встряхивали (Eppendorff Thermo mixer, 1400 об/мин) в течение 1,5 час при 37°C до получения прозрачного раствора. Образец сушили в концентраторе SpeedVac (1,5 час) и ресуспендировали в 13,2 мкл DMSO, встряхивали в течение 10 сек, с последующей обработкой ультразвуком (20 сек), и встряхивали (например, в Eppendorff Thermo mixer, 1400 об/мин) в течение 10 мин. Добавляли 6 мл 20 мМ NaH2PO4; 140 мМ NaCl; 0,1% Плюроник F68; pH 7,4 и перемешивали в течение 1 час при комнатной температуре. Образец центрифугировали в течение 20 мин при 3000 g. Супернатант удаляли и преципитат растворяли в 0,6 мл 20 мМ NaH2PO4; 140 мМ NaCl; 1 % Плюроник F68, pH 7,4. Добавляли 3,4 мл H2O и перемешивали в течение 1 час при комнатной температуре с последующим центрифугированием в течение 20 мин при 3000g. Восемь аликвот из каждых 0,5 мл супернатанта сохраняли при -20°C для дальнейшего использования.
3. Глобуломер Aβ(20-42)
Получение глобуломера Aβ(20-42) описано в примере 1c.
4. Глобуломер Aβ(12-42)
Получение глобуломера Aβ(12-42) описано в примере 1d.
5. Предварительно обработанный HFIP мономер Aβ(1-40), 5 мМ в DMSO
1 мг Aβ(1-40), (Bachem Inc, кат. № H-1194) суспендировали в 0,25 мл HFIP (суспензия 4 мг/мл) в пробирке Eppendorff. Пробирку встряхивали (например, в Eppendorff Thermo mixer, 1400 об/мин) в течение 1,5 ч при 37°C для получения прозрачного раствора и затем сушили в концентраторе speed vac (1,5 ч). Образец повторно растворяли в 46 мкл DMSO (21,7 мг/мл раствора = 5 мМ), встряхивали в течение 10 сек и затем обрабатывали ультразвуком в течение 20 сек. После 10 мин встряхивания (например, в Eppendorff Thermo mixer, 1400 об/мин) Образец сохраняли при -20°C для дальнейшего использования.
6. Мономер Aβ(1-42), 0,1% NH4OH
1 мг Aβ(1-42) (Bachem Inc., кат. № H-1368) растворяли в 0,5 мл 0,1% NH4OH в H2O (свежеприготовленном) (= 2 мг/мл) и немедленно встряхивали в течение 30 сек при комнатной температуре для получения прозрачного раствора. Образец сохраняли при -20°C для дальнейшего использования.
7. Фибриллы Aβ(1-42)
1 мг Aβ(1-42) (Bachem Inc. каталожный № H-1368) растворяли в 500 мкл водного 0,1% NH4OH (пробирка Eppendorff), и образец перемешивали в течение 1 мин при комнатной температуре. 100 мкл этого свежеприготовленного раствора Aβ(1-42) нейтрализовали 300 мкл 20 мМ NaH2PO4; 140 мМ NaCl, pH 7,4. pH доводили до pH 7,4 с помощью 1% HCl. Образец инкубировали в течение 24 ч при 37°C и центрифугировали (10 мин при 10000g). Супернатант удаляли, и осадок фибрилл ресуспендировали в 400 мкл 20 мМ NaH2PO4; 140 мМ NaCl, pH 7,4 посредством встряхивания в течение 1 мин.
8. sAPPα
Поставлен из Sigma (кат. № S9564; 25 мкг в 20 мМ NaH2PO4; 140 мМ NaCl; pH 7,4). sAPPα разводили в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, 0,2 мг/мл BSA до 0,1 мг/мл (= 1 пмоль/мкл).
Материалы для дот-блоттинга
Стандарты Aβ:
Серийное разведение антигенов Aβ в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4+ 0,2 мг/мл BSA
1) 100 пмоль/мкл;
2) 10 пмоль/мкл;
3) 1 пмоль/мкл;
4) 0,1 пмоль/мкл;
5) 0,01 пмоль/мкл.
Нитроцеллюлоза:
Среда для переноса транс-блоттингом, чистая нитроцеллюлозная мембрана (0,45 мкм); BIO-RAD
Анти-мышь-AP:
AQ330A (Chemicon)
Реагент для детекции:
Таблетки NBT/BCIP (Roche)
Бычий сывороточный альбумин, (BSA):
A-7888 (SIGMA)
Блокирующий реагент:
5% молоко с низкой жирностью в TBS
Буферные растворы:
TBS
25 мМ Трис/HCl-буфер pH 7,5
+ 150 мМ NaCl
TTBS
25 мМ Трис/HCl-буфер pH 7,5
+ 150 мМ NaCl
+ 0,05 % Tween 20
PBS + 0,2 мг/мл BSA
20 мМ буфер NaH2PO4 pH 7,4
+ 140 мМ NaCl
+ 0,2 мг/мл BSA
Раствор антитела I:
Образцы плазмы мыши из исследования активной иммунизации глобуломером Aβ(20-42) (разведение 1:400 в 20 мл 1% молока с низкой жирностью в TBS)
Раствор антитела II:
разведение 1:5000
Анти-мышь-AP в 1% молоке с низкой жирностью в TBS
Способ дот-блоттинга:
1) 1 мкл каждого из различных стандартов Aβ (в 5 их серийных разведениях) наносили точками на нитроцеллюлозную мембрану на расстоянии приблизительно 1 см друг от друга.
2) Точкам стандартов Aβ позволяли высохнуть на нитроцеллюлозной мембране на воздухе в течение по меньшей мере 10 мин при комнатной температуре (RT) (= дот-блот)
3) Блокирование:
Дот-блот инкубировали с 30 мл 5% молока с низкой жирностью в TBS в течение 1,5 час при RT.
4) Промывка:
Блокирующий раствор сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT.
5) Раствор антитела I:
Буфер для промывки сливали, и дот-блот инкубировали с раствором антитела I в течение ночи при RT.
6) Промывка:
Раствор антитела I сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TBS в течение 10 мин при RT.
7) Раствор антитела II:
Буфер для промывки сливали, и дот-блот инкубировали с раствором антитела II в течение 1 ч при RT
8) Промывка:
Раствор антитела II сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали, и дот-блот инкубировали при встряхивании с 20 мл TBS в течение 10 мин при RT.
9) Проявка:
Раствор для промывки сливали. 1 таблетку NBT/BCIP растворяли в 20 мл H2O, и дот-блот инкубировали в течение 5 мин с этим раствором. Проявку останавливали интенсивной промывкой H2O.
Результаты показаны на фигуре 4.
Анализ дот-блоттингом антител против Aβ, полученных после активной иммунизации мышей глобуломером Aβ(20-42) для оценки их специфичности по отношению к различным формам Aβ. Выделенные формы Aβ наносили точками в серийных разведениях и инкубировали с соответствующей плазмой мыши, содержащей антитела против Aβ, полученные в ходе иммунной реакции. Отдельные дот-блоты соответствуют различным индивидуумам иммунизированных мышей.
1. Глобуломер Aβ(1-42).
2. Мономер Aβ(1-42), предварительно обработанный HFIP, в 0,1% Плюроник F68.
3. Глобуломер Aβ(20-42).
4. Глобуломер Aβ(12-42).
5. Мономер Aβ(1-40), предварительно обработанный HFIP, 5 мМ в DMSO.
6. Мономер Aβ(1-42), 0,1% NH4OH.
7. Препарат фибрилл Aβ(1-42).
8. sAPPα (Sigma); (первая точка: 1 пмоль).
В исследовании активной иммунизации мышей APP/L Tg показали, что иммунизация глобуломером Aβ(20-42) приводит к лучшим результатам для облегчения когнитивных нарушений у этих мышей по сравнению с обработкой PBS. Для образцов плазмы от мышей APP/L Tg после активной иммунизации глобуломеров Aβ(20-42) показали профиль антител (предпочтительное распознавание глобуломера Aβ(20-42) и глобуломера Aβ(12-42)), напоминающий профиль mAb против глобуломера Aβ(20-42), заявленных здесь.
Пример 6: Концентрация растворимого и нерастворимого пептида Aβ(1-42) и Aβ(1-40) в экстрактах мозга мышей, активно иммунизированных APP/PS1 Tg мономером Aβ(1-42), глобуломером Aβ(1-42), глобуломером Aβ(20-42) или носителем в качестве контроля.
40 самок мышей из дважды трансгенной модели болезни Альцгеймера на мышах (мыши APP/PS1 Tg) на фоне FVBxC57BI/6J в возрасте 4 месяцев использовали для этого исследования. Мыши APP/PS1 Tg обладают 695 аминокислотной формой APP человека с мутацией V717I (положение по отношению к наиболее длинной изоформе APP) и, кроме того, ген пенесилина 1 человека с мутацией A264E. Оба гена находятся под контролем промотора Thy1. Мышей получили и охарактеризовали в одной из основных лабораторий reMYND, Experimental Genetics Group, Campus Gasthuisberg, Catholic University Leuven, by Prof. Fred Van Leuven et al.
Всех мышей генотипировали посредством полимеразной цепной реакции (ПЦР) в возрасте 3 недель и присваивали уникальный идентификационный номер, как только становились известными результаты ПЦР.
Мыши обладали свободным доступом к предварительно фильтрованной и стерильной воде (УФ-лампа) и стандартному корму для мышей. Корм хранили в сухих и холодных условиях в хорошо вентилированном хранилище. Количество воды и корма проверяли ежесуточно, добавляли по необходимости и по умолчанию обновляли дважды в неделю.
Мышей содержали при обращенном ритме день-ночь: 14 часов света/10 часов темноты начиная с 19-00, в стандартных металлических клетках типа RVS T2 (площадь 540 см2). Клетки оборудованы твердыми полами и слоем соломенной подстилки. Число мышей на клетку ограничивали в соответствии с законодательством о благосостоянии животных. За пять суток до начала теста поведения мышей помещали в другие клетки макролон типа 2 и транспортировали в лабораторию для адаптации к лабораторному окружению при подготовке к тесту поведения.
Мышам вводили 100 мкг мономера Aβ(1-42) (0,1% NH4OH), глобуломера Aβ(1-42) или глобуломера Aβ(20-42) в фосфатно-солевом буфере (PBS), смешанного с равным количеством полного адъюванта Фрейнда внутрибрюшинно, с последующими бустер-инъекциями таким же количеством антигена в неполном адъюванте Фрейнда каждую третью неделю в течение четырех месяцев.
Биохимия
Aβ(1-40) и Aβ(1-42) в растворимой фракции 1 полушария головного мозга определяли посредством ELISA. Кроме того, Aβ(1-40) и Aβ(1-42) в нерастворимой мембранной фракции 1 полушария головного мозга определяли посредством ELISA.
Мышей анестезировали смесью 2:1:1 Кеталара (кетамина), Ромпуна (ксилазина 2%) и атропина и проводили транскардиальную промывку физиологической сывороткой при 4°C. Это проводили для удаления крови из сосудов мозга, процедуры, не влияющей на целостность органов. Цереброспинальную жидкость (CSF) собирали посредством надреза мышц, между черепом и первым шейным позвонком. Выполняли пункцию в мостомозжечковую цистерну иглой размера 26 и 10-20 мкл, CSF собирали тонкой стеклянной пипеткой.
Кровь собирали посредством пункции сердца, и 1 мл отбирали шприцом в покрытые гепарином пробирки Eppendorf. Кровь центрифугировали при 14000 об/мин при 4°C в течение 5 минут. Сыворотку хранили при -70°C.
Мышам проводили транскардиальную промывку физиологической сывороткой при 4°C.
Мозг удаляли из черепа, задний мозг и передний мозг разделяли разрезом в коронарной/фронтальной плоскости. Мозжечок удаляли. Передний мозг разделяли на левую и правую полусферу с использованием разреза по срединной линии.
Одну полусферу немедленно погружали в жидкий азот и хранили при -70°C до биохимического анализа.
Гомогенизация и фракционирование полусферы мозга
Мозг гомогенизировали с использованием гомогенизатора Поттера, стеклянной пробирки (свободной от детергента, 2 см3) и механического гомогенизатора (650 об/мин). Объем 6,5 × ½ массы мозга свежеприготовленного 20 мМ буфера Трис/HCl (pH 8,5) с ингибиторами протеаз (1 таблетка на 50 мл буфера Трис/HCl, Complete™, Roche, Mannheim, Germany) использовали в качестве буфера для гомогенизации.
Образцы переносили с -70°C в держатель для образцов с жидким азотом, и каждый отдельный образец предварительно нагревали посредством инкубации на столе в течение нескольких секунд перед гомогенизацией. Гомогенаты собирали в пробирки для центрифугирования Beckman TLX и хранили во льду перед центрифугированием. Между двумя образцами гомогенизатор Поттера и стеклянную пробирку тщательно промывали дистиллированной водой (AD) без детергентов и высушивали абсорбирующей бумагой.
Образцы центрифугировали в предварительно охлажденной ультрацентрифуге (Beckman, Mannheim, Germany) в течение 1 часа и 20 минут при 48000 об/мин (135000 × g) при 4°C. Из-за ограниченного числа держателей в центрифуге (N=8) образцы сортировали по массе мозга (для уравновешивания центрифуги) и распределяли случайным образом, чтобы разделить различные группы обработки по различным сеансам центрифугирования.
Супернатант (растворимую фракцию, содержащую секретированный APP и пептиды амилоида) отделяли от осадка (мембранной фракции, содержащей связанные с мембраной фрагменты APP и связанные с бляшками пептиды амилоида в случае старых мышей). Супернатант разделяли на две пробирки, из которых одну сохраняли при -20°C в качестве резерва, а другую далее обрабатывали хроматографией на колонке для концентрации пептидов амилоида.
Массы мозга, объем использованного буфера Трис/HCl, сеансы центрифугирования (помеченные цветом) и объем растворимой фракции, использованной для хроматографии на колонке, приведены в качестве примера в следующей таблице.
(мкл)
(мкл)
Небольшие колонки с обращенной фазой (C18-Sep-Pack Vac 3cc cartridges, Waters, Massachusetts, MA) устанавливали в вакуумной системе и промывали 80% ацетонитрилом в 0,1% трифторуксусной кислоте (A-TFA) с последующими двумя промывками 0,1% TFA. Затем образцы наносили на колонки, и колонки последовательно промывали 5% и 25% A-TFA. Пептиды амилоида элюировали 75% A-TFA, и элюаты собирали в 2 мл пробирки во льду. Элюаты лиофилизировали в концентраторе SpeedVac (Savant, Farmingdale, NY) в течение ночи и повторно растворяли в 330 мкл разбавителя для образцов, поставляемого с наборами для ELISA.
Осадки далее фракционировали на различные мембранные фракции: мембранную фракцию A (MFA), мембранную фракцию B (MFB), содержащую полноразмерный APP и мембранную фракцию C (MFC), содержащую связанный с бляшками амилоид. Таким образом образцы растворяли в буфере TBS с ингибиторами протеаз (1 таблетка на 50 мл буфера TBS, Complete™, Roche, Mannheim, Germany), и MFA разделяли на две пробирки, из которых одну сохраняли при -20°C в качестве резерва. 60% MFA дополнительно обрабатывали добавлением NP40 (2% от конечного объема) и Triton X-100 (2% от конечного объема) в TBS с ингибиторами протеазы и центрифугировали в течение одного часа при 27000 об/мин (98000 × g) в ультрацентрифуге Beckman при 4°C с использованием качающегося ротора (SW60). Супернатант (MFB) отделяли от осадка (MFC), и оба хранили при -20°C.
Массы мозга, 60% от массы мозга, объемы использованного буфера TBS + PI + NP40 + Triton X-100 и сеансы центрифугирования (помеченные цветом) приведены в качестве примера в следующей таблице.
(мг)
ELISA Aβ человека в растворимой фракции одной полусферы
Для оценки количества Aβ(1-40) человека и Aβ(1-42) человека в растворимой фракции гомогенатов мозга и/или в цереброспинальной жидкости (CSF) использовали коммерчески доступные наборы для твердофазного иммуноферментного анализа (ELISA) (высокочувствительный ELISA для амилоида h β40 или β42, The Genetics Company, Zurich, Switzerland). ELISA проводили согласно протоколу производителя. Коротко, стандарты (разведения синтетического Aβ(1-40) или Aβ(1-42)) и образцы подготавливали в 96-луночном полипропиленовом планшете без способности связывания белка (Greiner bio-one, Frickenhausen, Germany). Стандартные разведения с конечными концентрациями 1000, 500, 250, 125, 62,5, 31,3 и 15,6 пг/мл и образцы получали в разбавителе для образцов, поставленном с набором для ELISA, в конечном объеме 60 мкл. Поскольку уровни амилоида увеличиваются с возрастом мыши и поскольку для истинной оценки необходимо чтобы считывания образцов находились внутри линейной части стандартной кривой, образцы для анализов Aβ(1-40) разводили 1:3, образцы для анализов Aβ(1-42) разводили 1:6.
Образцы, стандарты и пустые контроли (50 мкл) наносили в покрытый анти-Aβ полистироловый планшет (с фиксированным антителом, избирательно узнающим C-конец антигена) в дополнение к избирательному конъюгату антитела против Aβ (биотинилированное антитело для детекции) и инкубировали в течение ночи при 4°C, чтобы позволить формирование комплекса антитело-амилоид-антитело. На следующие сутки добавляли конъюгат стрептавидин-пероксидаза с последующим добавлением через 30 минут смеси TMB/пероксид, что приводит к превращению субстрата в окрашенный продукт. Эту реакцию останавливали добавлением серной кислоты (1 M), и интенсивность окраски измеряли посредством фотометрии с помощью считывателя для ELISA с фильтром 450 нм. Количественную оценку содержания Aβ в образцах получали посредством сравнения поглощения со стандартной кривой, полученной с синтетическим Aβ(1-40) или Aβ(1-42).
ELISA Aβ человека в нерастворимой фракции одной полусферы
Для оценки количества Aβ(1-40) человека и Aβ(1-42) человека в нерастворимой мембранной фракции гомогенатов мозга образцы MFC дополнительно обрабатывали и растворяли в 8M гуанидине в 80 мМ Трис/HCl. Затем образцы инкубировали в течение 3 часов в термовстряхивателе при 25°C и пипетировали вверх и вниз с помощью 100 мкл пипеткой каждый час для растворения осадка MFC в гуанидиновом буфере. Наконец образцы центрифугировали в течение только 1 минуты при 4000 об/мин для удаления дебриса.
Масса мозга, масса осадка MFC и объем 8 М гуанидинового буфера приведены в качестве примера в следующей таблице.
Для оценки количества Aβ(1-40) человека и Aβ(1-42) человека в конечных образцах, использовали коммерчески доступные наборы для твердофазного иммуноферментного анализа (ELISA) (высокочувствительный ELISA для амилоида h β40 или β42 ELISA, The Genetics Company, Zurich, Switzerland). ELISA проводили согласно протоколу производителя, за исключением получения стандартов (разведения синтетического Aβ(1-40) или Aβ(1-42)). Образцы получали в разбавителе для образцов, поставленном с набором для ELISA, в конечном объеме 60 мкл. Поскольку гуанидин влияет на значения OD стандартной кривой, получали стандартные разведения с конечные концентрациями 1000, 500, 250, 125, 62,5, 31,3 и 15,6 пг/мл получали в разбавителе для образцов с той же самой концентрацией гуанидина, как и в образцах. Это выполняли в 96-луночном полипропиленовом планшете без способности связывания белка (Greiner bio-one, Frickenhausen, Germany).
Поскольку уровни амилоида увеличиваются с возрастом мыши и поскольку для истинной оценки необходимо, чтобы считывания образцов находились внутри линейной части стандартной кривой, образцы для анализов Aβ(1-40) разводили 1:3, образцы для анализа нерастворимого Aβ(1-40) и нерастворимого Aβ(1-42) разводили 1:500.
Образцы, стандарты и пустые контроли (50 мкл) наносили в покрытый анти-Aβ полистироловый планшет (с фиксированным антителом, избирательно узнающим C-конец антигена) в дополнение к избирательному конъюгату антитела против Aβ (биотинилированное антитело для детекции) и инкубировали в течение ночи при 4°C, чтобы позволить формирование комплекса антитело-амилоид-антитело. На следующие сутки добавляли конъюгат стрептавидин-пероксидаза с последующим добавлением через 30 минут смеси TMB/пероксид, что приводит к превращению субстрата в окрашенный продукт. Эту реакцию останавливали добавлением серной кислоты (1 M), и интенсивность окраски измеряли посредством фотометрии с помощью считывателя для ELISA с фильтром 450 нм. Количественную оценку содержания Aβ в образцах получали посредством сравнения поглощения со стандартной кривой, полученной с синтетическим Aβ(1-40) или Aβ(1-42).
Результаты показаны на фигуре 5
Концентрация растворимого и нерастворимого пептида Aβ(1-42) и пептида Aβ(1-40) в экстрактах мозга активно иммунизированных мышей APP/PS1 Tg мономером Aβ(1-42) (0,1% NH4OH), глобуломером Aβ(1-42), глобуломером Aβ(20-42) или носителем в качестве контроля.
В растворимой и нерастворимой фракции экстракта мозга мышей APP/PS1 Tg, активно иммунизированных глобуломером Aβ(20-42), уровень пептидов Aβ(1-40) и Aβ(1-42) значимо не отличался от контроля-носителя. В отличие от этого иммунизация глобуломером Aβ(1-42) и мономером Aβ(1-42) приводит к уменьшению в мозге уровней Aβ(1-40) и Aβ(1-42). Это указывает на то, что способ прямой иммунизации глобуломером Aβ(20-42) значительно не изменяет общие уровни Aβ в мозге, однако, тем не менее, является эффективным для облегчения связанных с пептидом Aβ когнитивных нарушений (см. пример 4).
Пример 7: Анализ когнитивного поведения посредством теста распознавания объектов для трансгенных мышей APP/L после пассивной иммунизации антителами против глобуломера Aβ(20-42).
В этих экспериментах использовали мышей со сверэкспрессией APP человека с точечной мутацией. Точечная мутация относится к аминокислоте 717 (замене валина на изолейцин) и обнаружена в семье из Лондона, где она приводила к дебюту AD до начала шестого десятилетия жизни (Mullan et al., Nature Genetics 2 (1992) 340-342). Трансгенных мышей, обозначенных здесь как APP/L, получили и впервые описали в Leuven (Moechars et al., J. Biol. Chem. 274 (1999) 6483-6492). Самок мышей APP/L подвергали пассивной иммунизации в возрасте 3 месяцев. Мышам вводили 250 мкг любого из моноклональных антител мыши 5F7, 10F11 или 7C6 в 100 мкл фосфатно-солевого буфера (PBS). В течение периода времени эксперимента животных содержали в стандартных условиях с обращенным циклом день/ночь (14 часов света, начиная с 19-00/10 часов темноты). Они хорошо переносили пассивную иммунизацию, без каких-либо признаков неблагоприятных эффектов.
После третьей инъекции (15 сутки эксперимента) когнитивную способность мышей тестировали посредством теста распознавания объектов, как описано в данной области (Dewachter et al. Journal of Neuroscience 22 (2002) 3445-3453). С этой целью мышей приучали к площадке и затем в течение 10 минут подвергали фазе обучения, во время которой их по отдельности помещали на площадку, теперь содержащую два одинаковых элемента (зеленые кубики или оранжевые цилиндры сходного размера, приблизительно 4 см). Регистрировали продолжительность и частоту, с которой мышь исследовала объекты. Во время фазы запоминания, 2,5 час спустя, мышей возвращали на площадку, теперь содержащую, в дополнение к известному объекту, другой объект. Распознавание нового объекта регистрировали как время, в течение которого мышь исследовала новый объект, по отношению к общему времени (исследование старого и нового объекта). «Показатель распознавания» выражает это отношение (время для нового объекта/общее время). Мышь, которая не запоминает известный объект, будет считать его таким же интересным, как новый объект, и тратить такое же количество времени на его исследование, иными словами, обладать показателем распознавания 50%. Мышь, которая запоминает известный объект, будет считать его неинтересным и, таким образом, обладать значительно более высоким показателем распознавания. Известно, что мыши APP/L обладают когнитивными нарушениями в возрасте 4,5 месяцев и обладают показателем распознавания при измерении на уровне случайного, т.е. 50%.
Результаты показаны на фигуре 6.
Тест распознавания объектов для мышей. Тест описывает распознавание известного объекта по сравнению с неизвестным объектом, измеренное в переводе на исследовательское поведение в течение 10-минутной фазы тестирования. Показатель распознавания определяют как процент времени, которое мышь тратит на исследование неизвестного объекта относительно времени, потраченного на исследование обоих объектов. Мышь исследовала известный объект в течение 10-минутной фазы обучения за 2,5 часа перед фазой тестирования.
a) Трансгенных мышей APP иммунизировали один раз в неделю в течение трех недель посредством внутрибрюшинной инъекции 250 мкг антитела 5F7 (n=9), антитела 10F11 (n=11) или антитела 7C6 (n=11); контрольным животным вводили PBS (n=6). Значимые отличия от случайного уровня (50%, т.е. равное время исследования как известного, так и неизвестного объекта) отмечены звездочками. * = p < 0,05 (t-тест)
b) Сравнение всех мышей, обработанных антителами (5F7, 10F11 и 7C6; (n=31)) и мышей, обработанных фосфатно-солевым буфером (PBS; n=6). RI обработанной антителами группы значимо отличался от случайного уровня (** = P<0,01; t-тест).
Известно, что мыши APP/L обладают, когнитивными нарушениями в возрасте 4,5 месяцев и обладают показателем распознавания в шкале значений случайного уровня, т.е. 50%.
Действительно, для обработанных PBS мышей показали случайное поведение. Пассивная иммунизация всеми тремя антителами (5F7, 10F11 и 7C6) приводила к заметно увеличенному показателю распознавания. При сравнении пулированной группы по сравнению с контролями, показатель распознавания является значительно увеличенным. Этот благоприятный эффект на показатели памяти мышей APP/L после введения всех трех антител позволяет предполагать, что антитело против усеченного глобуломера Aβ(20-42) является достаточным для достижения когнитивного улучшения.
Пример 8: Дот-блот анализ профиля избирательности антител против глобуломера Aβ(20-42).
Чтобы охарактеризовать избирательность моноклональных антител против глобуломера Aβ(20-42), их использовали в качестве зондов для распознавания различных форм Aβ. С этой целью получали серийные разведения выделенных форм Aβ(1-42) в диапазоне от 100 пмоль/мкл до 0,01 пмоль/мкл в PBS, дополненном 0,2 мг/мл BSA. По 1 мкл каждого образца наносили точками на нитроцеллюлозную мембрану. Для детекции использовали соответствующее антитело (0,2 мкг/мл). Иммуноокрашивание выполняли с использованием конъюгированного с пероксидазой антитела против IgG мыши и окрашивающего реагента субстрата BM Blue POD (Roche).
Стандарты Aβ для дот-блоттинга:
1. Мономер Aβ(1-42), 0,1 % NH4OH
1 мг Aβ(1-42) (Bachem Inc., кат. № H-1368) растворяли в 0,5 мл 0,1% NH4OH в H2O (свежеприготовленного) (= 2 мг/мл) и немедленно встряхивали в течение 30 сек при комнатной температуре для получения прозрачного раствора. Образец хранили при -20°C для дальнейшего использования.
2. Мономер Aβ(1-40), 0,1% NH4OH
1 мг Aβ(1-40) (Bachem Inc., cat. № H-1368) растворяли в 0,5 мл 0,1% NH4OH в H2O (свежеприготовленного) (= 2 мг/мл) и немедленно встряхивали в течение 30 сек при комнатной температуре для получения прозрачного раствора. Образец хранили при -20°C для дальнейшего использования.
3. Мономер Aβ(1-42), 0,1% NaOH
2,5 мг Aβ(1-42) (Bachem Inc., кат. № H-1368) растворяли в 0,5 мл 0,1% NaOH в H2O (свежеприготовленного) (= 5 мг/мл) и немедленно встряхивали в течение 30 сек при комнатной температуре для получения прозрачного раствора. Образец хранили при -20°C для дальнейшего использования.
4. Мономер Aβ(1-40), 0,1% NaOH
2,5 мг Aβ(1-40) (Bachem Inc., кат. № H-1368) растворяли в 0,5 мл 0,1% NaOH в H2O (свежеприготовленного) (= 5 мг/мл) и немедленно встряхивали в течение 30 сек при комнатной температуре для получения прозрачного раствора. Образец хранили при -20°C для дальнейшего использования.
5. Глобуломер Aβ(1-42)
Получение глобуломера Aβ(1-42) описано в примере 1a.
6. Глобуломер Aβ(12-42)
Получение глобуломера Aβ(12-42) описано в примере 1d.
7. Глобуломер Aβ(20-42)
Получение глобуломера Aβ(20-42) описано в примере 1c.
8. Фибриллы Aβ(1-42)
1 мг Aβ(1-42) (Bachem Inc. кат. № H-1368) растворяли в 500 мкл водного 0,1% NH4OH (пробирка Eppendorff) и образец перемешивали в течение 1 мин при комнатной температуре. 100 мкл этого свежеприготовленного раствора Aβ(1-42) нейтрализовали 300 мкл 20 мМ NaH2PO4; 140 мМ aCl, pH 7,4. pH доводили до pH 7,4 с помощью 1% HCl. Образец инкубировали в течение 24 час при 37°C и центрифугировали (10 мин при 10000 g). Супернатант удаляли и осадок фибрилл ресуспендировали в 400 мкл 20 мМ NaH2PO4; 140 мМ NaCl, pH 7,4 посредством встряхивания в течение 1 мин.
9. sAPPα
Поставлен из Sigma (кат. № S9564; 25 мкг в 20 мМ NaH2PO4; 140 мМ NaCl; pH 7,4). sAPPα разводили до 0,1 мг/мл (= 1 пмоль/мкл) 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, 0,2 мг/мл BSA.
Материалы для дот-блоттинга:
Стандарты Aβ:
Серийное разведение антигенов Aβ в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 + 0,2 мг/мл BSA
1) 100 пмоль/мкл
2) 10 пмоль/мкл
3) 1 пмоль/мкл
4) 0,1 пмоль/мкл
5) 0,01 пмоль/мкл
Нитроцеллюлоза:
Среда для переноса транс-блоттингом, чистая нитроцеллюлозная мембрана (0,45 мкМ); BIO-RAD
Анти-мышь-POD:
Кат № 715-035-150 (Jackson lmmuno Research)
Реагент для детекции: Субстрат BM Blue POD, преципитирующий (Roche)
Бычий сывороточный альбумин, (BSA):
Кат. № A-7888 (SIGMA)
Блокирующий реагент:
5 % молока с низкой жирностью в TBS
Буферные растворы:
TBS 25 мМ Трис/HCl-буфер pH 7,5
+ 150 мМ NaCl
TTBS
25 мМ Трис/HCl-буфер pH 7,5 + 150 мМ NaCl + 0,05 % Tween 20
PBS + 0,2 мг/мл BSA
20 мМ буфер NaH2PO4 pH 7,4
+ 140 мМ NaCl
+ 0,2 мг/мл BSA
Раствор антитела I:
0,2 мкг/мл антитело, разведенное в 20 мл 1 % молока с низкой жирностью в TBS
Раствор антитела II:
разведение 1:5000
Анти-мышь-POD в 1% молоке с низкой жирностью в TBS
Способ дот-блоттинга:
1) 1 мкл каждого из различных стандартов Aβ (в 5 их серийных разведениях) наносили точками на нитроцеллюлозную мембрану на расстоянии приблизительно 1 см друг от друга.
2) Точкам стандартов Aβ позволяли высохнуть на нитроцеллюлозной мембране на воздухе в течение по меньшей мере 10 мин при комнатной температуре (RT) (=дот-блот).
3) Блокирование:
Дот-блот инкубировали с 30 мл 5% молока с низкой жирностью в TBS в течение 1,5 час при RT.
4) Промывка:
Блокирующий раствор сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT.
5) Раствор антитела I:
Буфер для промывки сливали и дот-блот инкубировали с раствором антитела I в течение 2 час при RT
6) Промывка:
Раствор антитела I сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TBS в течение 10 мин при RT.
7) Раствор антитела II:
Буфер для промывки сливали и дот-блот инкубировали с раствором антитела II в течение ночи при RT
8) Промывка:
Раствор антитела II сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TBS в течение 10 мин при RT.
9) Проявка:
Раствор для промывки сливали. Дот-блот проявляли с помощью 10 мл субстрата BM Blue POD в течение 10 мин. Проявку останавливали интенсивной промывкой дот-блота H2O. Количественную оценку выполняли с использованием денситометрического анализа (денситометр GS800 (BioRad) и пакета программного обеспечения Quantity one, версии 4.5.0 (BioRad)) по интенсивности точек. Оценивали только точки, обладающие относительной плотностью большей, чем 20% от относительной плотности последней однозначно определенной точки глобуломера Aβ(20-42). Это значение порога независимо определяли для каждого дот-блота независимо. Рассчитанное значение указывает на соотношение между узнаванием глобуломера Aβ(20-42) и соответствующей формой Aβ для данного антитела.
Результаты показаны на фигуре 7.
Анализ дот-блоттингом специфичности различных антител против Aβ (6E10, 5F7, 4B7, 10F11, 6A2, 4D10, 2F2; 3B10, 7C6, 7E5, 10C1) по отношению к различным формам Aβ. Тестированные моноклональные антитела получены (за исключением 6E10) посредством активной иммунизации мышей глобуломером Aβ(20-42) с последующим отбором слитых клеток гибридомы. Отдельные формы Aβ наносили в серийных разведениях и инкубировали с соответствующими антителами для иммунной реакции.
1. Мономер Aβ(1-42), 0,1 % NH4OH
2. Мономер Aβ(1-40), 0,1% NH4OH
3. Мономер Aβ(1-42), 0,1%NaOH
4. Мономер Aβ(1-40), 0,1% NaOH
5. Глобуломер Aβ(1-42)
6. Глобуломер Aβ(12-42)
7. Глобуломер Aβ(20-42)
8. Препарат фибрилл Aβ(1-42)
9. sAPPα (Sigma); (первая точка: 1 пмоль)
Избирательные mAb против глобуломера Aβ(20-42) можно разделить на 3 класса по отношению к дискриминации глобуломера Aβ(1-42) и глобуломера Aβ(12-42). Первый класс, включающий в себя антитела 6A2, 5F7 и 2F2, узнает предпочтительно глобуломер Aβ(20-42) и, в некоторой степени, глобуломер Aβ(1-42) (и также глобуломер Aβ(12-42)). Второй класс, включающий в себя антитела 10F11, 4D10 и 3B10, предпочтительно узнает глобуломер Aβ(20-42) и также узнает глобуломер Aβ(12-42), но, в меньшей степени, и не узнает в значительной степени глобуломер Aβ(1-42). Третий класс, включающий в себя антитела 7C6, 4B7, 7E5 и 10C1 узнает глобуломер Aβ(20-42), но для него не показали значительное узнавание. Все три класса не узнают в значительной степени мономерный Aβ(1-42), мономерный Aβ(1-40), фибриллы Aβ(1-42) или sAPPα.
По профилю избирательности антител против глобуломера Aβ(20-42) показали, что значительно увеличенный показатель распознавания при пассивной иммунизации (на фигуре 6) в основном должен быть обусловлен избирательным распознаванием усеченного глобуломера Aβ(20-42) и глобуломера Aβ(12-42) и в меньшей степени глобуломера Aβ(1-42) и не мономерного Aβ(1-42), мономерного Aβ(1-40), фибрилл Aβ(1-42) или sAPPα.
Пример 9: Анализ in situ специфической реакции антител, избирательных для Aβ(20-42), с фибриллярным пептидом Aβ в форме бляшек Aβ у старых мышей TG2576 и с амилоидом Aβ сосудов оболочек мозга.
Для этих экспериментов использовали материал мозга мышей TG2576 в возрасте 19 месяцев (Hsiao et al., 1996, Science; 274(5284), 99-102) или мышей APP/LxPS1 в возрасте 9 месяцев (описание, как выше; ReMYND, Leuven, Belgium) или материал аутопсии двух пациентов с болезнью Альцгеймера (RZ16 и RZ55; получены из BrainNet, Munich). У мышей сверхэкспрессирован APP человека с так называемой шведской мутацией (K670N/M671L; Tg2576) или так называемой лондонской мутацией (V717I), помимо гена пресенилина 1 человека с мутацией A264E (APP/LxPS1) и формируются отложения β-амилоида в паренхиме мозга в возрасте приблизительно 7-11 месяцев и отложения β-амилоида в более крупных сосудах головного мозга в возрасте приблизительно 18 месяцев (Tg2576). Животных подвергали глубокой анестезии и транскардиальной перфузии 0,1 M фосфатно-солевым буфером (PBS) для отмывки крови. Затем мозг удаляли из черепа и разделяли в продольном направлении. Одну полусферу мозга быстро замораживали, другие фиксировали погружением в 4% параформальдегид. Фиксированные погружением полусферы подвергали криопротекции смачиванием в 30% сахарозе в PBS и монтировали на замороженном микротоме. Целый передний мозг разрезали на срезы 40 мкМ, которые собирали в PBS и использовали для последующего способа окрашивания. Материал мозга человека представлял собой глубоко замороженный брусок из неокортекса приблизительно 1 см3. Небольшую часть бруска фиксировали погружением в 4% параформальдегид и далее обрабатывали как материал мозга мыши.
Отдельные срезы окрашивали Конго красным с использованием следующего способа:
Материал:
- набор с красителем для амилоида Конго красным (Sigma-Aldrich; HT-60), состоящий из спиртового раствора NaCl, раствора NaOH и раствора Конго красного;
- кюветы для окрашивания;
- предметные стекла для микроскопа SuperfrostPlus и покровные стекла;
- этанол, ксилол, среда для заливки.
Реагенты:
- разведением NaOH 1:100 раствором NaCl получают щелочной физиологический раствор;
- разведением щелочного физиологического раствора 1:100 раствором Конго красного получают щелочной раствор Конго красного (полученный не более, чем за 15 мин перед использованием, фильтрат);
- монтируют срезы на предметном стекле и позволяют им высохнуть;
- инкубируют предметное стекло в кювете для окрашивания, сначала в течение 30-40 минут в щелочном физиологическом растворе, затем в течение 30-40 минут в щелочном растворе Конго красного;
- промывают три раза свежим этанолом и заливают ксилолом.
Окрашивание сначала фотографировали с использованием микроскопа Zeiss Axioplan и оценивали количественно. Красная окраска указывала на отложения амилоида как в форме бляшек, так и в более крупных сосудах оболочек мозга. Эти результаты показаны на фиг. 8A. В дальнейшем оценку окрашивания антителом фокусировали на этих структурах.
Окрашивание антителом проводили посредством инкубации срезов с раствором, содержащим 0,07-7,0 мкг/мл соответствующего антитела в соответствии со следующим способом:
Материалы:
- раствор для промывки TBST (забуференный Трисом физиологический раствор с Tween 20; 10× концентрат; DakoCytomation; S3306 1:10 в бидистилированной воде)
- 0,3% H2O2 в метаноле;
- сыворотка осла (Serotec), 5% в TBST;
- моноклональное антитело мыши против глобуломера, разведенное в TBST;
- вторичное антитело: биотинилированное антитело осла против антител мыши (Jackson Immuno; 715-065- 150; разведенные 1:500 в TBST);
- StreptABComplex (DakoCytomation; K 0377);
- набор с субстратом для пероксидазы диаминобензидином (=DAB; Vector Laboratories; SK-4100);
- предметные и покровные стекла для микроскопа SuperFrost Plus;
- свободная от ксилола среда для заливки (Medite; X-tra Kitt).
Способ:
- затопляют срезы ледяной 0,3% H2O2 и инкубируют в течение 30 мин;
- промывают в течение 5 мин в буфере TBST;
- инкубируют с сывороткой осла/TBST в течение 20 минут;
- инкубируют с первичным антителом в течение 24 часов при комнатной температуре;
- промывают в буфере TBST в течение 5 минут;
- инкубируют с блокирующей сывороткой из набора с пероксидазой Vectastain Elite ABC в течение 20 минут;
- промывают в буфере TBST в течение 5 минут;
- инкубируют с вторичным антителом в течение 60 минут при температуре окружающей среды;
- промывают в буфере TBST в течение 5 минут;
- инкубируют с StreptABComplex в течение 60 минут при температуре окружающей среды;
- промывают в буфере TBST в течение 5 минут;
- инкубируют с DAB из набора с пероксидазой Vectastain Elite ABC в течение 20 минут;
- монтируют срезы на предметных стеклах, сушат их на воздухе, дегидратируют их спиртом и фиксируют.
Помимо визуальной проверки окрашивания, окрашивание бляшек, кроме того, оценивали количественно посредством графического вырезания 10 случайно выбранных бляшек из гистологических изображений с использованием системы анализа изображений ImagePro 5.0 и определения среднего значения для них в шкале серого. Значения оптической плотности рассчитывали из значений в шкале серого посредством вычитания средней фоновой плотности окрашенного материала из плотности бляшек амилоида (0% - отсутствие окрашивания бляшки выше окружающего фона, 100% - отсутствие перехода/максимальное окрашивание), и тестировали статистическую значимость между контролем и антителами, а также между 6G1 и антителами, избирательными для Aβ(20-42), соответственно, с помощью ANOVA.
Результаты окрашивания для мышей Tg2576 и APP/LxPS1 показаны на фигуре 8B-D и H.
Связывание различных антител в концентрации 0,7 мкг/мл с поперечными срезами неокортикса трансгенных мышей TG 2576 в возрасте 19 месяцев:
C) Паренхимальные отложения Aβ (амилоидные бляшки) являлись окрашенными только 6G1 и 6E10, но не избирательными для глобуломеров антителами (т.е. 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1).
D) для всех избирательных для глобуломеров антител (т.е. 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1) показали значительно меньшее окрашивание паренхимальных бляшек по сравнению с коммерчески доступными антителами 6E10 и 4G8.
Связывание различных антител в концентрации 0,07-7,0 мкг/мл с поперечными срезами неокортикса трансгенных мышей APP/LxPS1 в возрасте 11 месяцев:
E) Паренхимальные отложения Aβ (амилоидные бляшки) являлись значительно сильнее окрашенными и при более низкой концентрации с помощью 6G1, 6E10 и 4G8, чем с помощью избирательных для глобуломеров антител (т.е. 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1).
Все отложения амилоида верифицировали посредством предварительного конгофильного окрашивания (Конго красный; см. Фиг. 8A). шкала = 100 мкм.
По оценке коричневых отложений DAB показали, что не избирательные для Aβ антитела 6G1 и 6E10 окрашивают бляшки и сосуды оболочек мозга, в то время как избирательные для глобуломеров Aβ(20-42) антитела 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1 не окрашивают. Эти результаты показали отсутствие связывания или заметно меньшее связывание этих антител с фибриллами Aβ или другими видами Aβ, присутствующими в амилоидных структурах in vivo. Предполагают, что это уменьшенное связывание уменьшает опасность побочных эффектов, индуцированных слишком быстрым растворением бляшек и последующим увеличением уровня растворимого Aβ или нейровоспаления из-за взаимодействия связанных с бляшками антител с микроглией.
Результаты окрашивания для мозга человека при болезни Альцгеймера показаны на фигуре 8B, F-H.
Связывание различных антител в концентрации 0,7 мкг/мл с поперечными срезами неокортекса пациента RZ55:
B) Паренхимальные отложения Aβ (амилоидные бляшки) являлись окрашенными только 6G1 и 6E10, но не избирательными для глобуломеров антителами (т.е. 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1).
F) Для всех избирательных для глобуломеров антител (т.е. 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1) показали значительно меньшее окрашивание по сравнению с коммерчески доступными антителами 6E10 и 4G8.
H) Отложения Aβ в сосудах (стрелки) являлись окрашенными только 6G1 и 6E10, но не избирательными для глобуломеров антителами (т.е. 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1).
Связывание различных антител в концентрации 0,07-7,0 мкг/мл с поперечными срезами неокортексов трансгенных мышей APP/LxPS1 в возрасте 11 месяцев:
G) Паренхимальные отложения Aβ (амилоидные бляшки) являлись значительно сильнее окрашенными и при более низкой концентрации с помощью 6G1, 6E10 и 4G8, чем с помощью избирательных для глобуломеров антител (т.е. 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1).
Все отложения амилоида верифицировали посредством предварительного конгофильного окрашивания (Конго красный; см. Фиг. 8A).
По оценке коричневых отложений DAB показали, что не избирательные для Aβ антитела 6G1 и 6E10 окрашивают бляшки и сосуды оболочек мозга, в то время как избирательные для глобуломеров Aβ(20-42) антитела 5F7, 2F2, 6A2, 4D10, 10F11, 3B10, 7C6, 7E5 и 10C1 не окрашивают. Для коммерчески доступных антител 6E10 и 4G8 показали более сильное окрашивание по сравнению с избирательными для глобуломеров антител, но меньшее окрашивание, чем для 6G1. Эти результаты подтверждают характер окрашивания для трансгенных мышей, где отсутствовало связывание или присутствовало заметно меньшее связывание избирательных для глобуломеров антител с фибриллами Aβ или другими видами Aβ, присутствующими в амилоидных структурах in vivo. Предполагают, что это уменьшенное связывание уменьшает опасность побочных эффектов, индуцированных слишком быстрым растворением бляшек и последующим увеличением уровня растворимого Aβ или нейровоспаления из-за взаимодействия связанных с бляшками антител с микроглией.
Пример 10: титр антитело против Aβ и профиль избирательности при дот-блоттинге в плазме мышей TG2576 спустя приблизительно один год после активной иммунизации.
Спустя приблизительно один год после последней иммунизации (глобуломером Aβ(20-42), глобуломером Aβ(12-42), мономером Aβ(1-42) и носителем) мышей Tg 2576 (из примера 9) в образцах плазмы оценивали, что антитела против Aβ продуцированы и еще присутствуют. С этой целью получали серийные разведения различных форм Aβ(1-42) в диапазоне концентраций от 100 пмоль/мкл до 0,01 пмоль/мкл в PBS + 0,2 мг/мл BSA. Из каждого образца 1 мкл наносили на нитроцеллюлозную мембрану. Детекцию проводили с помощью подходящих образцов плазмы мыши (разведенными 1:400). Окрашивание проводили с помощью конъюгированной с антителами против IgG щелочной фосфатазы и добавления окрашивающего реагента NBT/BCIP.
Стандарты Aβ для дот-блоттинга:
1. Глобуломер Aβ(1-42)
Получение глобуломера Aβ(1-42) описано в примере 1a.
2. Предварительно обработанный HFIP мономер Aβ(1-42) в Плюроник F68
3 мг Aβ(1-42), (Bachem Inc.; кат. № H-1368) растворяли в 0,5 мл HFIP (суспензия 6 мг/мл) в 1,7 мл пробирке Eppendorff и встряхивали (Eppendorff Thermo mixer, 1400 об/мин) в течение 1,5 час при 37°C до получения прозрачного раствора. Образец сушили в концентраторе SpeedVac (1,5 час) и ресуспендировали в 13,2 мкл DMSO, встряхивали в течение 10 сек, с последующей обработкой ультразвуком (20 сек), и встряхивали (например, в Eppendorff Thermo mixer, 1400 об/мин) в течение 10 мин. Добавляли 6 мл 20 мМ NaH2PO4; 140 мМ NaCl; 0,1% Плюроник F68; pH 7,4 и перемешивали в течение 1 час при комнатной температуре. Образец центрифугировали в течение 20 мин при 3000g. Супернатант удаляли, и преципитат растворяли в 0,6 мл 20 мМ NaH2PO4; 140 мМ NaCl; 1% Плюроник F68, pH 7,4. 3,4 мл H2O добавляли и перемешивали в течение 1 час при комнатной температуре с последующим центрифугированием в течение 20 мин при 3000g. Восемь аликвот из каждых 0,5 мл супернатанта сохраняли при -20°C для дальнейшего использования.
3. Глобуломер Aβ(20-42)
Получение глобуломера Aβ(20-42) описано в примере 1c.
4. Глобуломер Aβ(12-42)
Получение глобуломера Aβ(1-42) описано в примере 1d.
5. Предварительно обработанный HFIP мономер Aβ(1-40), 5 мМ в DMSO
1 мг Aβ(1-40), (Bachem Inc, кат. № H-1194) суспендировали в 0,25 мл HFIP (суспензия 4 мг/мл) в пробирке Eppendorff. Пробирку встряхивали (например, в Eppendorff Thermo mixer, 1400 об/мин) в течение 1,5 час при 37°C для получения прозрачного раствора и затем сушили в концентраторе SpeedVac (в течение 1,5 час). Образец повторно растворяли в 46 мкл DMSO (21,7 мг/мл раствора = 5 мМ), встряхивали в течение 10 сек и затем обрабатывали ультразвуком в течение 20 сек. После встряхивания (например, в Eppendorff Thermo mixer, 1400 об/мин) в течение 10 мин образец сохраняли при -20°C для дальнейшего использования.
6. Мономер Aβ(1-42), 0,1% NH4OH
1 мг Aβ(1-42) (Bachem Inc., кат. № H-1368) растворяли в 0,5 мл 0,1% NH4OH в H2O (свежеприготовленном) (= 2 мг/мл) и немедленно встряхивали в течение 30 сек при комнатной температуре для получения прозрачного раствора. Образец сохраняли при -20 °C для дальнейшего использования.
7. Фибриллы Aβ(1-42)
1 мг Aβ(1-42) (Bachem Inc. каталожный Nr.: H-1368) растворяли в 500 мкл водного 0,1% NH4OH (пробирка Eppendorff) и образец перемешивали в течение 1 мин при комнатной температуре. 100 мкл этого свежеприготовленного раствора Aβ(1-42) нейтрализовали 300 мкл 20 мМ NaH2PO4; 140 мМ NaCl, pH 7,4. pH доводили до pH 7,4 с помощью 1% HCl. Образец инкубировали в течение 24 час при 37°C и центрифугировали (10 мин при 10000 g). Супернатант удаляли и осадок фибрилл ресуспендировали в 400 мкл 20 мМ NaH2PO4; 140 мМ NaCl, pH 7,4 посредством встряхивания в течение 1 мин.
8. sAPPα
Поставлен из Sigma (кат. № S9564; 25 мкг in 20 мМ NaH2PO4; 140 мМ NaCl; pH 7,4). sAPPα разводили в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, 0,2 мг/мл BSA до 0,1 мг/мл (= 1 пмоль/мкл).
Материалы для дот-блоттинга
Стандарты Aβ:
Серийное разведение антигенов Aβ в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4 + 0,2 мг/мл BSA
1) 100 пмоль/мкл
2) 10 пмоль/мкл
3) 1 пмоль/мкл
4) 0,1 пмоль/мкл
5) 0,01 пмоль/мкл
Нитроцеллюлоза:
Среда для переноса транс-блоттингом, чистая нитроцеллюлозная мембрана (0,45 мкм); BIO-RAD
Анти-мышь-AP:
AQ330A (Chemicon)
Реагент для детекции:
Таблетки NBT/BCIP (Roche)
Бычий сывороточный альбумин, (BSA):
A-7888 (Fa. SIGMA)
Блокирующий реагент:
5% молоко с низкой жирностью в TBS
Буферные растворы:
TBS
25 мМ Трис/HCl - буфер pH 7,5
+ 150 мМ NaCl
TTBS
25 мМ Трис/HCl - буфер pH 7,5
+ 150 мМ NaCl
+ 0,05% Tween 20
PBS + 0,2 мг/мл BSA
20 мМ N буфер NaH2PO4 pH 7,4
+ 140 мМ NaCl
+ 0,2 мг/мл BSA
Раствор антитела I:
Плазма активно иммунизированных мышей TG2576, разведенная 1/400 в 20 мл 1 % молока с низкой жирностью в TBS
Раствор антитела II:
разведение 1:5000
Анти-мышь-AP в 1% молоке с низкой жирностью в TBS
Способ дот-блоттинга:
1) 1 мкл каждого из различных стандартов Aβ (в 5 их серийных разведениях) наносили точками на нитроцеллюлозную мембрану на расстоянии приблизительно 1 см друг от друга.
2) Точкам стандартов Aβ позволяли высохнуть на нитроцеллюлозной мембране на воздухе в течение по меньшей мере 10 мин при комнатной температуре (RT) (= дот-блот)
3) Блокирование:
Дот-блот инкубировали с 30 мл 5% молока с низкой жирностью в TBS в течение 1,5 час при RT.
4) Промывка:
Блокирующий раствор сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT.
5) Раствор антитела I:
Буфер для промывки сливали, и дот-блот инкубировали с раствором антитела I в течение ночи при RT.
6) Промывка:
Раствор антитела I сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TBS в течение 10 мин при RT.
7) Раствор антитела II:
Буфер для промывки сливали, и дот-блот инкубировали с раствором антитела II в течение 1 час при RT
8) Промывка:
Раствор антитела II сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TTBS в течение 10 мин при RT. Раствор для промывки сливали и дот-блот инкубировали при встряхивании с 20 мл TBS в течение 10 мин при RT.
9) Проявка:
Раствор для промывки сливали. 1 таблетку NBT/BCIP растворяли в 20 мл H2O и дот-блот инкубировали в течение 5 мин с этим раствором. Проявку останавливали интенсивной промывкой H2O.
Результаты показаны на фигуре 9.
Сыворотку из различных групп иммунизации: a) глобуломерами Aβ(20-42); b) глобуломерами Aβ(12-42); c) мономером Aβ(1-42), 0,1% NH4OH; d) контролем-носителем тестировали против различных форм Aβ в анализе дот-блоттингом различных профилей антител.
1. Глобуломер Aβ(1-42).
2. Мономер Aβ(1-42), предварительно обработанный HFIP, в 0,1% Плюроник F68.
3. Глобуломер Aβ(20-42).
4. Глобуломер Aβ(12-42).
5. Мономер Aβ(1-40), предварительно обработанный HFIP, 5 мМ в DMSO.
6. Мономер Aβ(1-42), растворенный в 0,1% NH4OH.
7. Препарат фибрилл Aβ(1-42).
8. sAPPα (Sigma); (первая точка: 1 пмоль).
В отличие от активной иммунизации носителем в качестве контроля или мономером Aβ(1-42) для иммунизации глобуломером Aβ(20-42) или глобуломером Aβ(12-42) показали высокий титр антител даже спустя приблизительно один год после последней иммунизации. Эти антитела являются избирательными для глобуломера Aβ(20-42) в случае иммунизации глобуломером Aβ(20-42) или избирательными для глобуломера Aβ(20-42) и глобуломера Aβ(12-42) в случае иммунизации глобуломером Aβ(12-42). Это указывает на то, что усеченный глобуломер Aβ(20-42) и глобуломер Aβ(12-42) представляют собой очень хорошие антигены и что антитела, направленные против них, обладают очень долгой персистенцией in vivo.
(Следует заметить, что на некоторых дот-блотах наблюдали неспецифический сигнал окрашивания, что, наиболее вероятно, представляет собой перекрестную реакцию антител мышей на BSA, используемый для серийных разведений пептидов Aβ.)
Пример 11: Уровни эпитопов глобуломера Aβ(20-42) в мозге пациентов при болезни Альцгеймера.
Экстракт мозга с SDS-DTT:
Образцы мозга с AD: RZ 16; RZ 52 и RZ 55 (получены из Brain Net, Munich)
Контрольный образец: RZ 92 (получен из Brain Net, Munich)
Одну таблетку полного ингибитора протеаз (Roche, кат. No. 1697 498) растворяли в 1 мл H2O (= раствор ингибитора протеаз). 100 мг образца мозга с AD гомогенизировали в 2,5 мл NaH2PO4, 140 мМ NaCl, 0,05% Tween 20, 0,5% BSA (дополненном 25 мкл раствор ингибитора протеаз) с помощью 20 ударов в стеклянном гомогенизаторе Поттера. Суспензию обрабатывали ультразвуком в течение 30 сек во льду, затем инкубировали при 37°C в течение 16 час. Суспензию центрифугировали при 100000g и 8°C в течение одного часа, затем собирали супернатант. Осадок растворяли в 5 мМ NaH2PO4, 35 мМ NaCl, pH 7,4 и гомогенизировали с помощью 10 ударов в стеклянном гомогенизаторе Поттера. Добавляли 75 мкл 10% SDS и 125 мкл 0,16 мг/мл DTT и перемешивали в течение 20 минут при температуре окружающей среды. Образец центрифугировали в течение 10 минут при 10000g, и супернатант хранили в течение ночи при -20°C. Перед использованием супернатант оттаивали и центрифугировали еще 10 мин при 10000g. Супернатант (= экстракт мозга SDS/DTT) использовали для ELISA.
a) Сэндвич-ELISA для эпитопов глобуломера Aβ(20-42)
Список реагентов:
1. Иммунологический планшет F96 Cert. Maxisorp NUNC- (кат. № 439454)
2. Связывающее антитело: 5F7, 7C6, 10F11
3. Буфер для связывания:
100 мМ гидрокарбонат натрия, pH9,6
4. Блокирующий реагент для ELISA (Roche Diagnostics GmbH кат. № 1112589)
5. Буфер PBST:
20 мМ NaH2PO4, 140 мМ NaCl, 0,05% Tween 20, pH 7,4
6. Стандарт Aβ(20-42) для калибровки
7. Первичное антитело: анти-Aβ pRAb BA199; аффинно очищенное (посредством глобуломер Aβ(1-42)-сефарозы), раствор IgG в PBS; конц.: 0,22 мг/мл
8. Вторичное антитело: конъюгат анти-кролик-POD; (Jackson ImmunoResearch, кат. № 111-036-045)
9. Проявка:
- TMB; (Roche Diagnostics GmbH Cat. № 92817060), 42 мМ в DMSO
- 3% H2O2 в H2O
- 100 мМ ацетат натрия, pH 4,9
- Останавливающий раствор: 2 М серная кислота
Получение реагентов:
1. Связывающее антитело:
Отдельные связывающие антитела 5F7, 7C6 и 10F11 разводили до конечной концентрации 0,7 мкг/мл в буфере для связывания.
2. Блокирующий реагент:
Для получения исходного блокирующего раствора блокирующий реагент растворяли в 100 мл H2O и хранили при -20°C в аликвотах по 10 мл каждая.
3 мл исходного блокирующего раствора разводили 27 мл H2O для блокирования одного планшета для ELISA.
3. Стандарт Aβ(20-42) для калибровки (CS1)
Получение глобуломера Aβ(1-42) описано в примере 1a. Определили концентрацию белка для глобуломера Aβ(20-42) (6,81 мг/мл) по Брэдфорд (BioRad). 14,68 мкл глобуломера Aβ(20-42) (6,81 мг/мл) разводили в 10 мл 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% Tween 20, pH 7,4, 0,5% BSA (= 10 мкг/мл). 10 мкл раствора 10 мкг/мл далее разводили в 10 мл 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% Tween20, pH 7,4, 0,5% BSA (= 10 нг/мл = CS1)
Стандарты для калибровки для Aβ(20-42):
экстракты мозга с SDS/DTT:
экстракты мозга с SDS/DTT = E#
(# обозначает 4 образца мозга человека (1) RZ 16; (2) RZ 52; (3) RZ 55; (4) RZ 92)
4. Первичное антитело:
Исходный раствор анти-Aβ pRAb разводили до 0,05 мкг/мл в PBST + 0,5% BSA. Раствор антитела использовали немедленно.
5. Вторичное антитело:
Лиофилизированный конъюгат анти-кролик-POD растворяли в 0,5 мл H2O и смешивали с 500 мкл глицерина. Затем концентрат антитела хранили при -20°C в аликвотах по 100 мкл. Концентрат разводили 1:10000 в буфере PBST. Раствор антитела использовали немедленно.
6. Раствор TMB:
20 мл 100 мМ ацетата натрия, pH 4,9, смешивали с 200 мкл раствора TMB и 29,5 мкл 3% пероксида водорода. Этот раствор использовали немедленно.
Планшет для ELISA для Aβ(20-42):
Стандарты для калибровки (CS1.1-CS1.8) и экстракты мозга в SDS/DTT из 4 образцов мозга человека (1) RZ 16; (2) RZ 52; (3) RZ 55; (4) RZ 92 (= E1-E4 в 4 их серийных разведениях E#.1-E#.4) определяли в двух параллелях:
Этот ELISA проводили с каждым из связывающихся моноклональных антител 5F7, 7C6, 10F11.
Способ
1. Добавить 100 мкл раствора mAb на лунку. Инкубировать планшет для ELISA в течение ночи при +6°C (холодильник).
2. Слить раствор антитела и промыть лунки три раза по 250 мкл буфера PBST каждый.
3. Добавить 250 мкл/лунку блокирующего раствора. Инкубировать в течение 2 часов при температуре окружающей среды.
4. Слить блокирующий раствор и промыть лунки три раза по 250 мкл буфера PBST каждый.
5. Добавить 100 мкл/лунку каждого из стандартов для калибровки и экстрактов мозга с SDS/DTT. Инкубировать планшет в течение 2 часов при температуре окружающей среды, затем в течение ночи при 6°C.
6. Слить стандарты для калибровки и растворы экстрактов мозга с SDS/DTT и промыть лунки три раза по 250 мкл буфера PBST каждый.
7. Добавить 200 мкл/лунку раствора первичного антитела и инкубировать в течение 1 часа при температуре окружающей среды.
8. Слить раствор первичного антитела и промыть лунки три раза по 250 мкл буфера PBST каждый.
9. Добавить 200 мкл/лунку раствора вторичного антитела и инкубировать в течение 1 часа при температуре окружающей среды.
10. Слить раствор вторичного антитела и промыть лунки три раза по 250 мкл буфера PBST каждый.
11. Добавить 100 мкл/лунку раствора TMB.
12. Контролировать окраску планшета во время развития (5-15 мин при температуре окружающей среды) и остановить реакцию добавлением 50 мкл/лунку останавливающего раствора при развитии подходящей окраски.
13. Измерить поглощение при 450 нм.
14. Рассчитать результаты с использованием калибровки.
15. Оценка: Если поглощение образцов выходит из линейного диапазона калибровки, развести их опять и повторить.
Результаты показаны на фигуре 10.
Уровни эпитопов глобуломера Aβ(20-42) в мозге по экстрактам мозга от пациентов с AD и контрольных субъектов
Сэндвич-ELISA использовали для оценки содержания усеченного эпитопа глобуломера Aβ(20-42) в экстрактах мозга. ELISA с соответствующими антителами против глобуломера Aβ(20-42) использовали для калибровки.
Экстракцией ткани мозга при болезни Альцгеймера показали, что содержание эпитопа глобуломера Aβ(20-42) значительно повышено по сравнению с контрольным пациентом. Это указывает на то, что эпитоп глобуломера Aβ(20-42) действительно является релевантным видом Aβ в мозге человека при болезни Альцгеймера, а не только релевантным для моделей болезни Альцгеймера на животных. Антитела, направленные против эпитопа глобуломера Aβ(20-42), таким образом, являются очень желательными для лечения болезни Альцгеймера.
Пример 12: Разработка линий клеток гибридомы против глобуломера Aβ(20-42)
Моноклональные антитела можно получать с использованием широкого множества способов, известных в данной области, включая использование гибридомы, рекомбинантного способа и способа фагового дисплея или их сочетания. Например, моноклональные антитела можно получить с использованием способов гибридомы, включая известные в данной области и объясняемые, например, в Harlow et al., Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling, et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N. Y., 1981) (полное содержание указанных ссылок приведено здесь в полном объеме). Термин «моноклональное антитело», как применяют здесь, не является ограниченным антителами, полученными способом гибридомы. Термин «моноклональное антитело» относится к антителу, полученному из одного клона, включая любой эукариотический, прокариотический или фаговый клон, а не к способу, которым оно получено.
Конкретный способ, использованный для получения антител, описанных здесь, представляет собой следующий:
Иммунизация мышей: мышей Balb/c и A/J (в возрасте 6-8 недель) иммунизировали подкожно 50 мкг антигена в CFA. Животным вводили бустер-инъекции: каждые три недели 50 мкг антигена в Immuneasy™ (Qiagen) всего в трех бустер-инъекциях. За четверо суток перед слиянием мышам вводили бустер-инъекции 10 мкг антигена внутривенно.
Слияние клеток и скрининг гибридомы: клетки селезенки от иммунизированных животных сливали с клетками миеломы SP2/0-Ag14 в соотношении 5:1 с использованием общепринятых способов. От семи до десяти суток после слияния, когда наблюдали макроскопические колонии, SN тестировали посредством ELISA для антитела против глобуломер Aβ(20-42). Клетки из положительных по ELISA лунок размножали и клонировали посредством лимитирующих разведений.
Определение изотипа антитела: изотипы mAb против глобуломера Aβ(20-42) определяли с использованием набора для изотипирования Zymed EIA.
Получение в больших количествах и очистка моноклональных антител: гибридомы размножали в среде, содержащей 5% эмбриональную бычью сыворотку с низким содержанием IgG (Hyclone). Супернатант собирали и концентрировали. mAb очищали с использованием хроматографии с белком A и диализовали в PBS.
Титры в сыворотке: Десять мышей иммунизировали глобуломером Aβ(20-42). Для всех мышей наблюдали сероконверсию с титрами в ELISA (1/2 макс OD 450 нм) 1:5000-10000.
ОБОЗНАЧЕНИЯ ГИБРИДОМ, ПРОДУЦИРУЮЩИХ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА
Внутренние обозначения Abbott Laboratories, используемые для депонирования.
Депонированные линии клеток:
1) ML13-7C6.1 D4.4A9.5 G8 (обозначенная здесь также «7C6»);
2) ML15-5F7.5B10 (обозначенная здесь также «5F7»);
3) ML15-10F11.3D9 (обозначенная здесь также «10F11»);
4) ML15-4B7.3A6 (обозначенная здесь также «4B7»);
5) ML15-2F2.3E12 (обозначенная здесь также «2F2»);
6) ML15-6A2.4B10 (обозначенная здесь также «6A2»);
7) ML13-4D10.3F3 (обозначенная здесь также «4D10»);
8) ML15-7E5.5E12 (обозначенная здесь также «7E5»);
9) ML15-10C1.5C6.3H4 (обозначенная здесь также «10C1»);
10) ML15-3B10.2D5.3F1 (обозначенная здесь также «3B10»).
| название | год | авторы | номер документа |
|---|---|---|---|
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2432362C2 |
| ИММУНОГЕННЫЕ ПРОДУКТЫ, ПОЛУЧЕННЫЕ НА ОСНОВЕ АМИНОКИСЛОТНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ МУТЕИНОВОГО АМИЛОИДА β (Aβ), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2750268C2 |
| БЕЗОПАСНЫЕ И ФУНКЦИОНАЛЬНЫЕ ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА | 2011 |
|
RU2607368C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ КОНЕЧНЫХ ПРОДУКТОВ ГЛУБОКОГО ГЛИКИРОВАНИЯ (RAGE) И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2518351C2 |
| УЛУЧШЕННЫЕ СЕЛЕКТИВНЫЕ В ОТНОШЕНИИ ПРОТОФИБРИЛЛ АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2429244C2 |
| АНТИТЕЛА К Aβ-ПЕПТИДУ И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2341533C2 |
| АНТИТЕЛА К БЕТА-АМИЛОИДУ | 2013 |
|
RU2689674C2 |
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2007 |
|
RU2472807C2 |
| ГУМАНИЗИРОВАННОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ Aβ и ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2783528C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С ПРОТОФИБРИЛЛАМИ, И ИХ ПРИМЕНЕНИЕ В МЕТОДАХ ТЕРАПИИ И ДИАГНОСТИКИ БОЛЕЗНИ ПАРКИНСОНА, ДЕМЕНЦИИ, АССОЦИИРОВАННОЙ С ОБРАЗОВАНИЕМ ДИФФУЗНЫХ ТЕЛЕЦ ЛЕВИ, И ДРУГИХ АЛЬФА-СИНУКЛЕИНОПАТИЙ | 2011 |
|
RU2555526C2 |
























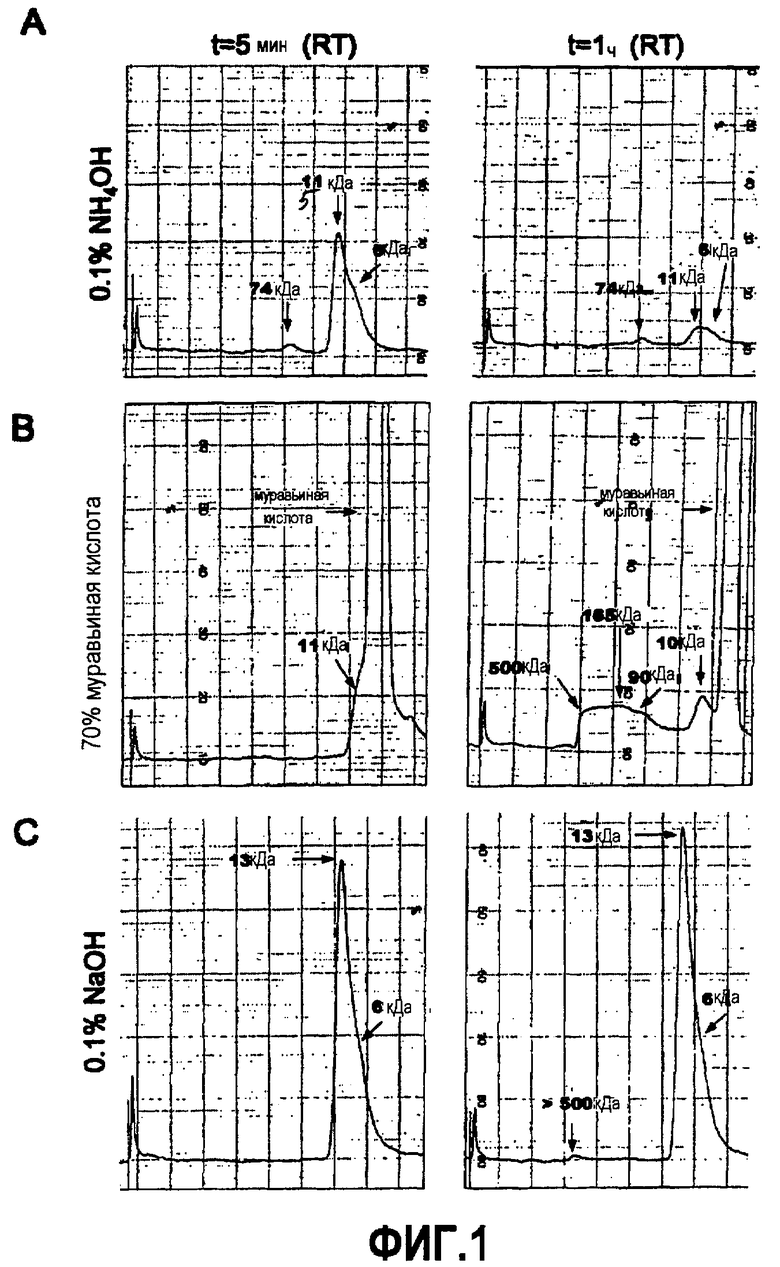
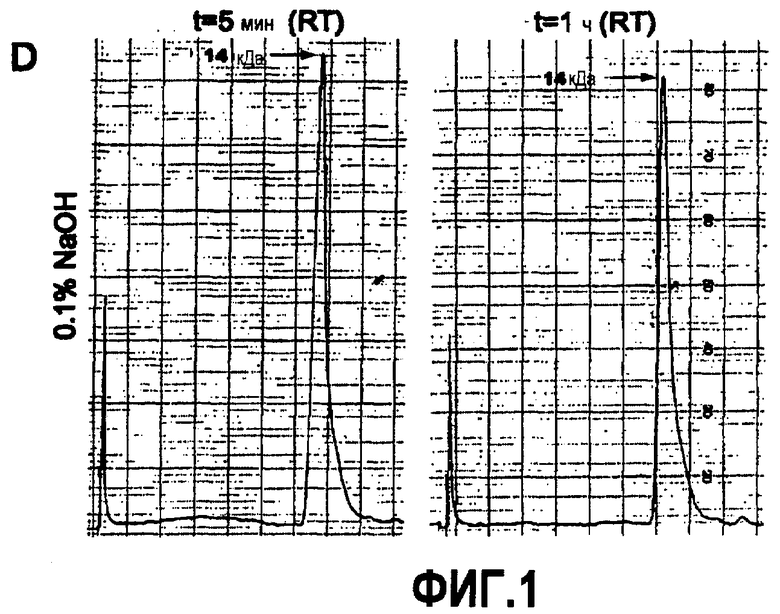
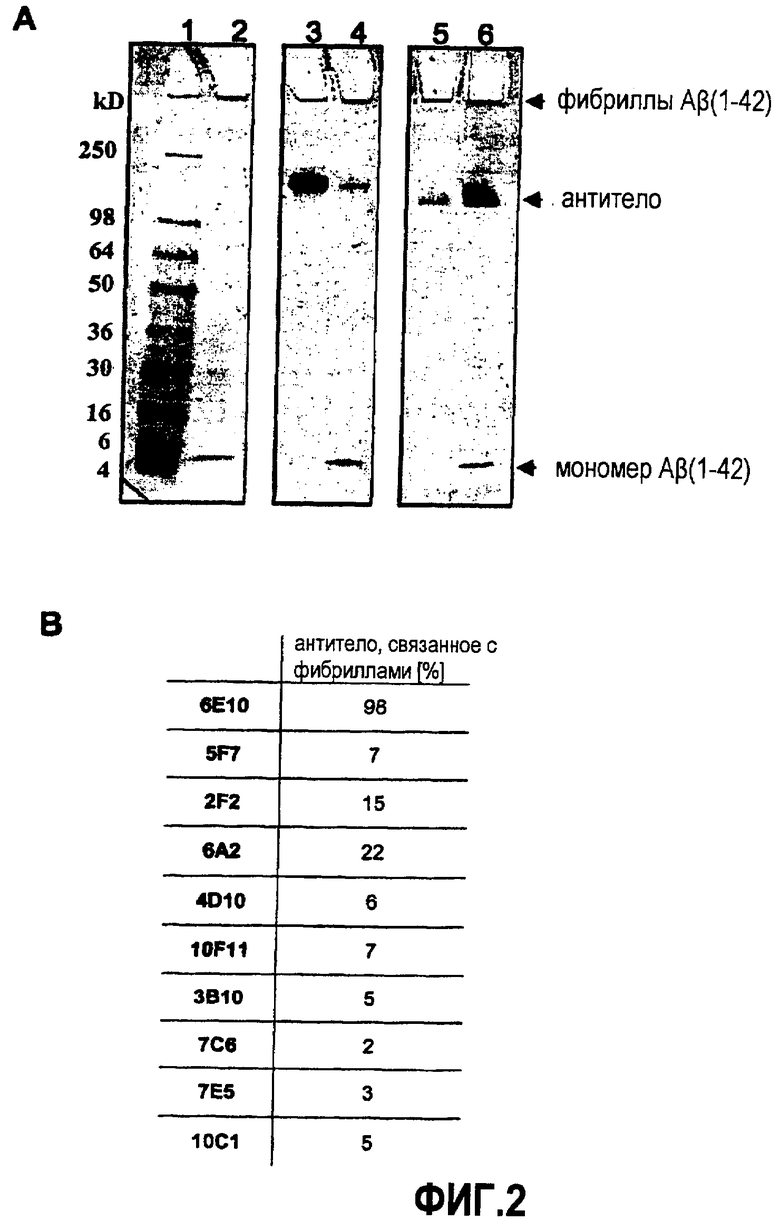
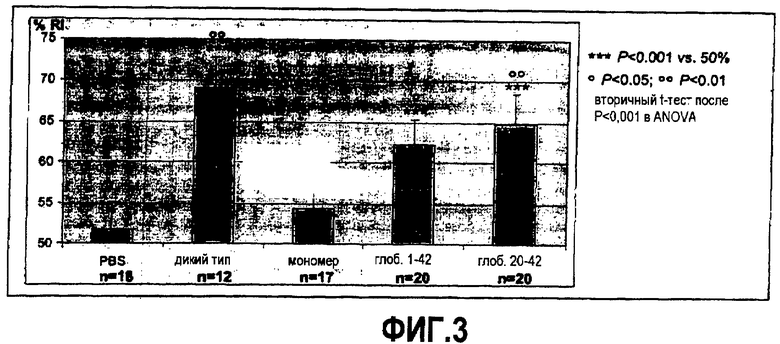
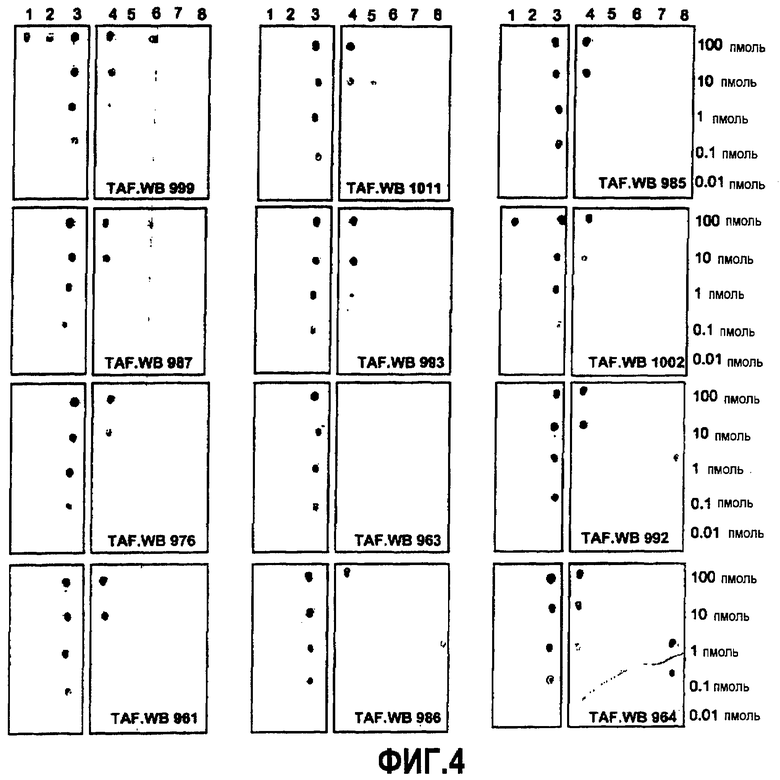
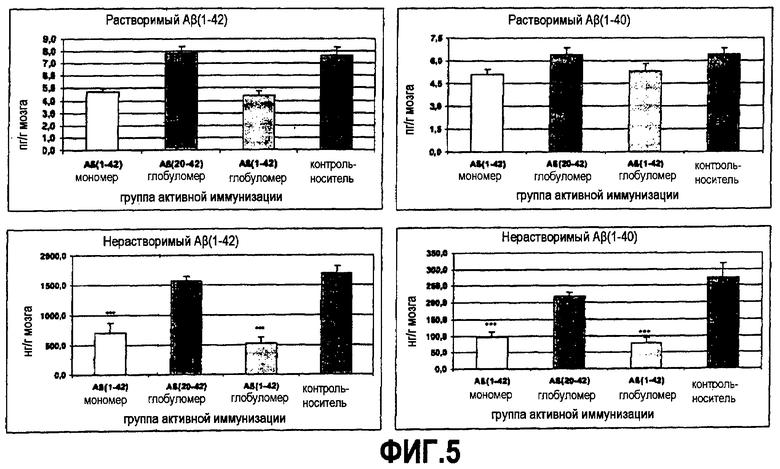
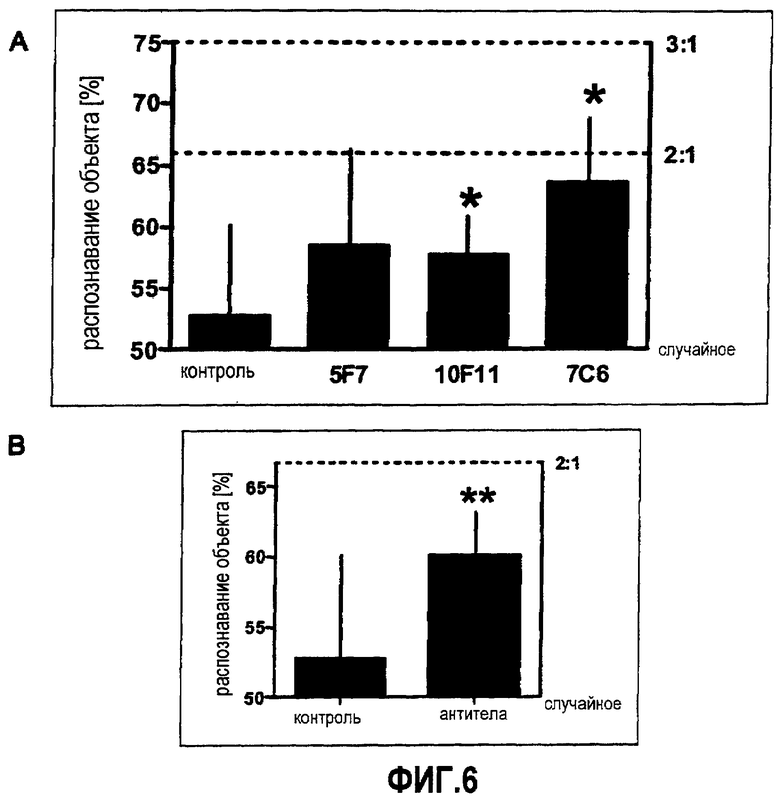
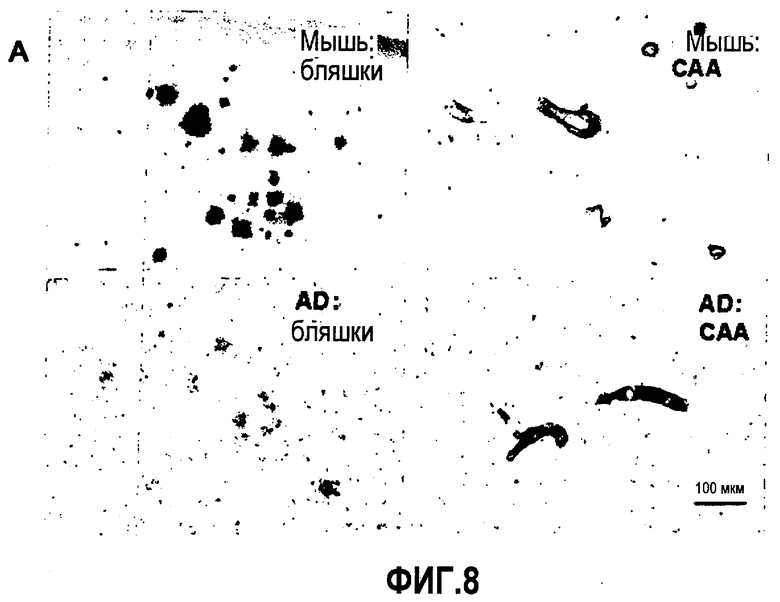

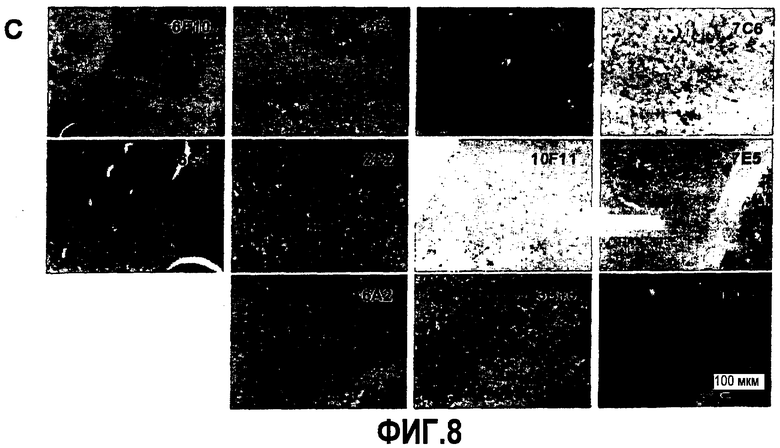
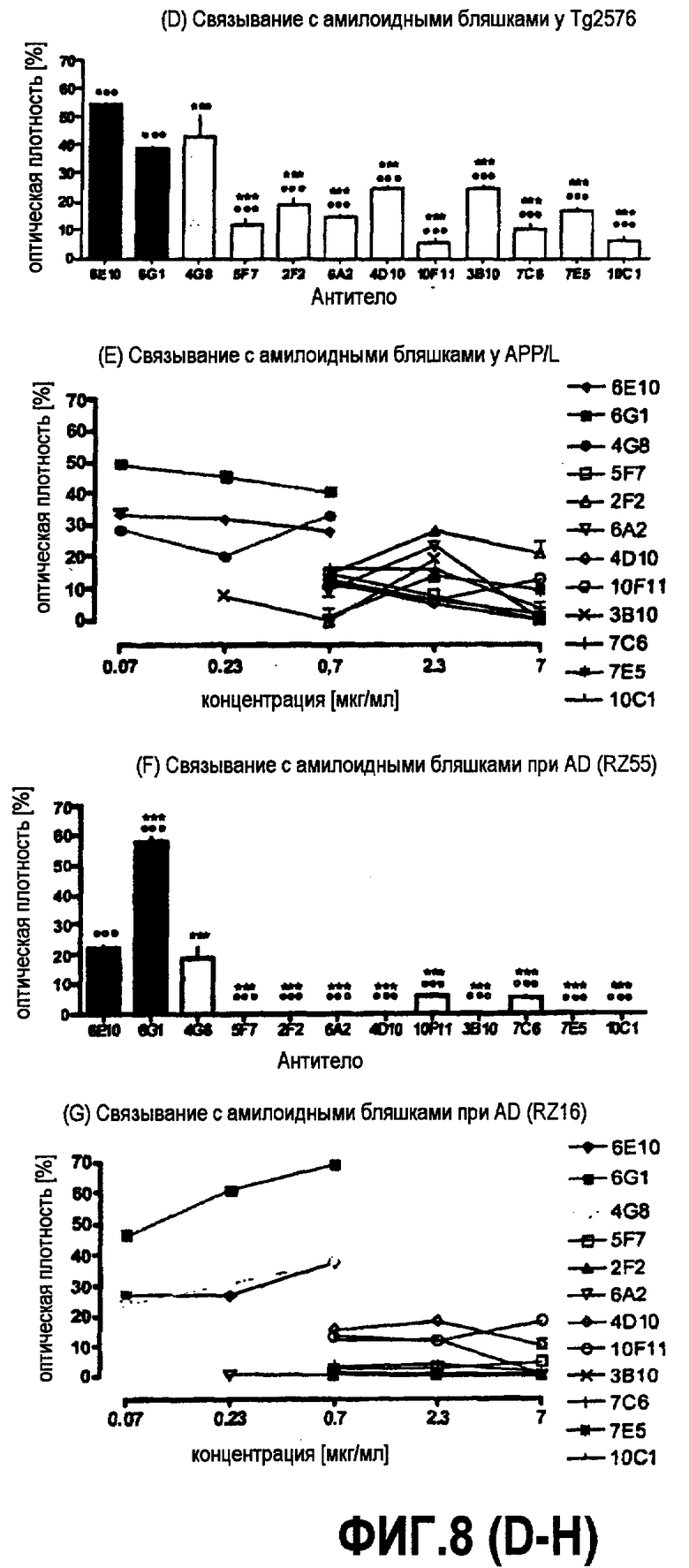
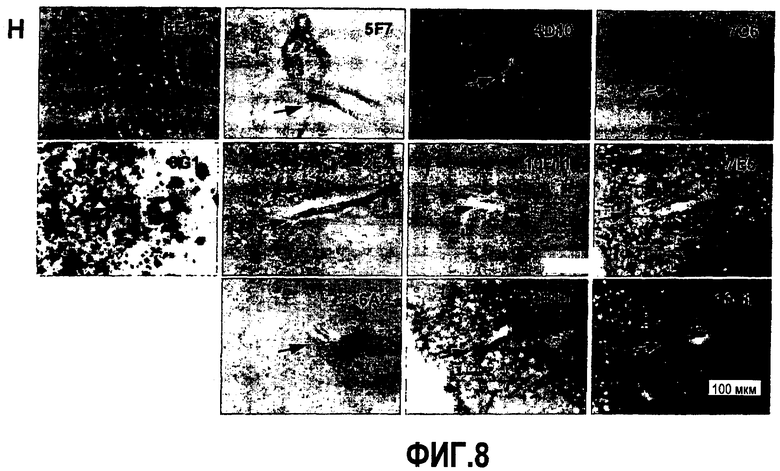

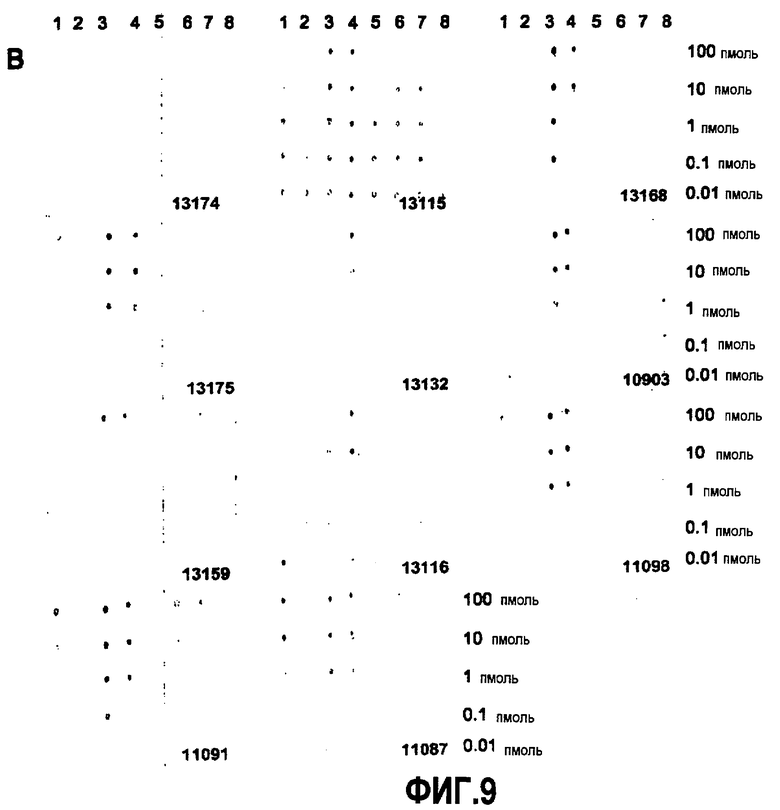
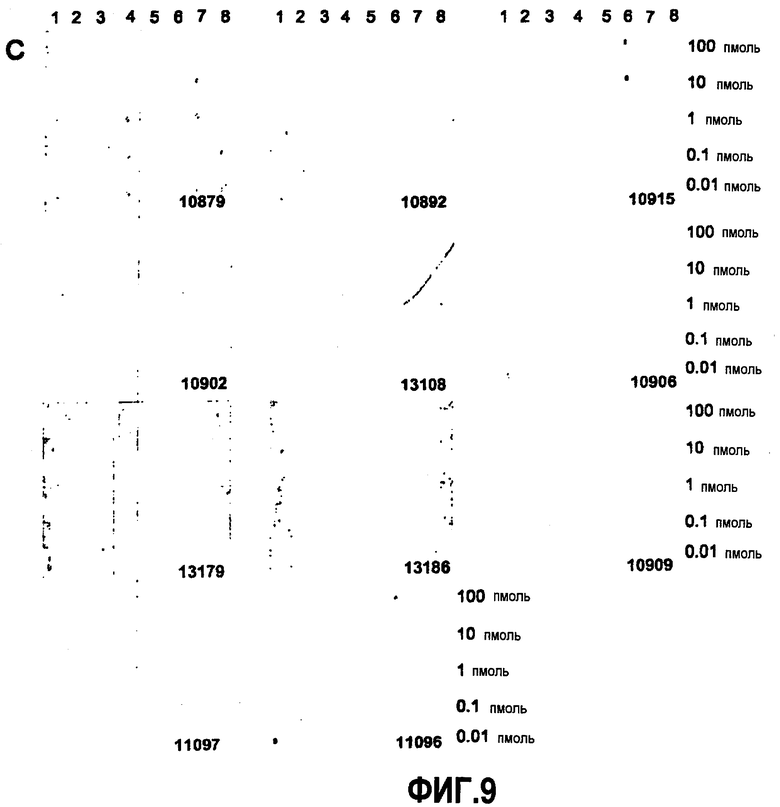
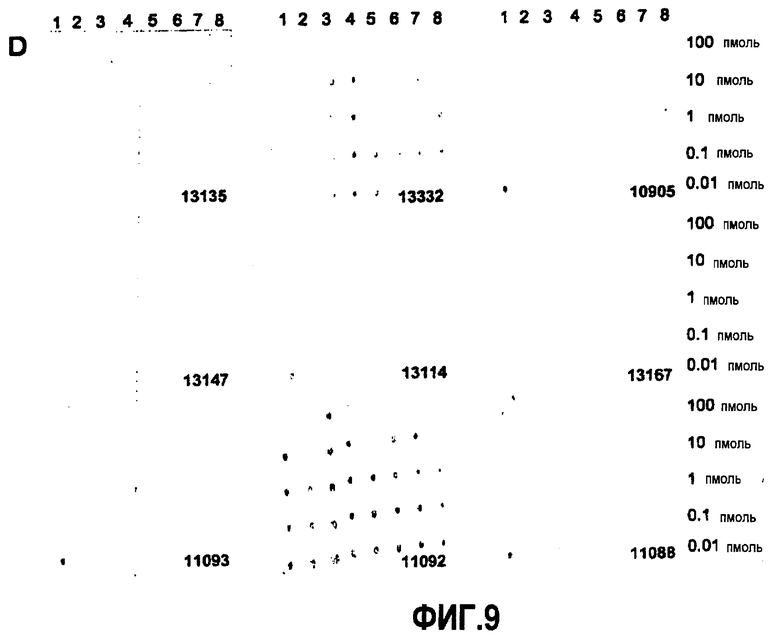











Настоящее изобретение относится к иммунологии и биотехнологии. Предложены варианты антитела, специфичного в отношении, по меньшей мере, одного глобуломера Аβ (20-42). Каждый из вариантов характеризуется тем, что включает VH и VL области, каждая из которых содержит по три соответствующих CDR. Раскрыта антигенсвязывающая область антитела. Описаны: кодирующая нуклеиновая кислота, а также вектор, ее содержащий, и клетка-хозяин, несущая вектор, - используемые для получения антитела. Раскрыт способ получения антитела с использованием клетки. Предложенные изобретения могут найти применение в терапии и диагностике болезни Альцгеймера и других амилоидозах. 6 н. и 4 з.п. ф-лы, 11 ил., 9 табл.
1. Антитело, специфичное в отношении, по меньшей мере, одного глобуломера Аβ (20-42), содержащее вариабельные области тяжелой (VH) и легкой (VL) цепей, где домены CDR вариабельных областей тяжелой и легкой цепей содержат аминокислотные последовательности, выбранные из группы, состоящей из следующих наборов CDR:
2. Антитело по п.1, где антитело является гуманизированным.
3. Антитело по п.1 или 2, где антитело содержит человеческую константную область.
4. Антитело по п.1 или 2, где антитело имеет характер гликозилирования человека.
5. Антигенсвязывающая часть антитела, как оно определено в любом из пп.1-4.
6. Антигенсвязывающая часть по п.5, где часть представляет собой Fab-фрагмент, F(ab')2-фрагмент или одноцепочечный Fv-фрагмент антитела.
7. Выделенная нуклеиновая кислота, кодирующая аминокислотную последовательность антитела по любому из пп.1-6.
8. Вектор для получения антитела, где вектор содержит выделенную нуклеиновую кислоту по п.7.
9. Клетка-хозяин для получения антитела, где клетка-хозяин содержит вектор по п.8.
10. Способ получения антитела, включающий культивирование клетки-хозяина по п.9 в культуральной среде в условиях, подходящих для продукции антитела.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| SERGEANT et al., "Truncated beta-amyloid peptide species in pre-clinical Alzheimer's disease as new targets for the vaccination approach", Journal of Neurochemistry, 2003, 85, 1581-1591 | |||
| СПОСОБ ЛЕЧЕНИЯ БОЛЕЗНИ АЛЬЦГЕЙМЕРА И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ КЛИОХИНОЛА ДЛЯ ЕЕ ЛЕЧЕНИЯ | 1997 |
|
RU2193406C2 |

