ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу избирательного и эффективного получения кардиомиоцитов из ЭС (ES) клеток и других плюрипотентных стволовых клеток.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
(1) Получение кардиомиоцитов с использованием плюрипотентных стволовых клеток
Как правило, кардиомиоциты претерпевают активное клеточное деление, автономно ритмически сокращаясь, до момента рождения, однако, сразу после рождения они теряют способность к делению, а поскольку у них имеется крайне мало недифференцированных стволовых клеток и клеток-предшественников, чьи способности к росту и дифференциации чрезвычайно низки, то когда кардиомиоциты гибнут, подвергаясь различным формам стресса, включая инфаркт миокарда, миокардит и тому подобное, погибшие кардиомиоциты не могут регенерировать. В результате функция миокарда поддерживается за счет сохранившихся кардиомиоцитов, которые подвергаются компенсаторной гипертрофии и тому подобному, однако, если стресс продолжается и его уровень превышает допустимое пороговое значение, это ведет к дальнейшему истощению и гибели кардиомиоцитов и последующему снижению функции миокарда (то есть сердечной недостаточности).
Сердечная недостаточность и другие типы сердечных заболеваний являются второй по значению причиной смерти в Японии, и прогноз является весьма неблагоприятным при 5-летней выживаемости всего лишь примерно 50% пациентов с сердечными заболеваниями. Поэтому есть надежда, что разработка высокоэффективных способов лечения сердечной недостаточности приведет к значительным успехам в области медицинского обслуживания и экономики здравоохранения. Общепринятые лекарственные средства для терапии сердечной недостаточности включают препараты дигиталиса, которые увеличивают силу сокращения миокарда, а также препараты ксантина и других сердечных стимуляторов, однако, как известно, долгосрочное введение данных лекарственных средств приводит к ухудшению состояния, так как расходуется слишком много энергии миокарда. Совсем недавно основной акцент в терапии сместился в сторону β-блокаторов и ингибиторов АПФ (ACE), которые снижают избыточную нагрузку на сердце за счет стимуляции симпатической нервной системы и системы «ренин-ангиотензин», однако, данные способы действуют только на непосредственные симптомы и не могут восстанавливать поврежденную сердечную ткань. Напротив, трансплантация сердца является фундаментальным лечением тяжелой сердечной недостаточности, однако, ее сложно применять повсеместно вследствие таких проблем, как нехватка доноров сердца, этических вопросов, физической и финансовой нагрузки на пациентов и тому подобного.
Вследствие этого, очевидно, что способы трансплантации для замены ослабленных или погибших кардиомиоцитов будут чрезвычайно полезны для лечения сердечной недостаточности. Действительно, из экспериментов с животными известно, что если незрелые кардиомиоциты, полученные из плода, трансплантировать во взрослую сердечную ткань, трансплантированные клетки функционируют эффективно (см. не патентный документ 1). Однако сложно получать достаточное количество кардиомиоцитов для данного способа и применение его в клинической медицине также осложнено с этической точки зрения.
Вследствие этого в последние годы внимание было сосредоточено на индукции дифференциации стволовых клеток в кардиомиоциты и использовании этих клеток для трансплантации. В настоящее время еще не представляется возможным точно идентифицировать популяцию клеток-предшественников или стволовых клеток, способных образовывать кардиомиоциты в сердечной ткани взрослых особей, так что плюрипотентные стволовые клетки, которые являются менее дифференцированными и способны дифференцироваться в различные клетки, считаются полезными для вышеуказанного способа.
Плюрипотентные стволовые клетки определяют как клетки, которые способны к бесконечной или долговременной клеточной пролиферации в культуре in vitro, оставаясь при этом в недифференцированном состоянии, которые сохраняют нормальные кариотипы и которые обладают способностью дифференцироваться во все три зародышевых слоя (эктодерму, мезодерму и эндодерму) при соответствующих условиях. Три хорошо известных типа плюрипотентных стволовых клеток представляют собой эмбриональные стволовые клетки (ЭС (ES) клетки), происходящие из эмбрионов на ранней стадии развития, эмбриональные зародышевые клетки (ЭЗ (EG) клетки), происходящие из примордиальных зародышевых клеток на эмбриональной стадии, и зародышевые стволовые клетки (ЗС (GS) клетки), происходящие из семенников непосредственно после рождения.
В частности, давно известно, что можно индуцировать дифференциацию ЭС клеток в кардиомиоциты in vitro. В большинстве ранних исследований использовали мышиные ЭС клетки. Если ЭС клетки культивируют в суспензионной культуре как одиночные клетки (отдельные клетки диспергированы без межклеточной адгезии в результате обработки ферментами или т.п.) в отсутствие фактора, ингибирующего дифференциацию, такого как фактор, ингибирующий лейкоз (LIF), или т.п., ЭС клетки слипаются друг с другом и агрегируют, образуя структуру, называемую эмбриоидные тельца (ЭТ) (EBs), которые похожи на ранние эмбриональные структуры. Также известно, что кардиомиоциты со способностью к спонтанному ритмическому сокращению возникают, когда данные ЭТ культивируют в суспензии или при прикреплении к поверхности устройств для культивирования.
Кардиомиоциты, происходящие из ЭС клеток, полученные как описано выше, обладают свойствами, очень схожими с таковыми у незрелых кардиомиоцитов из сердца плода (см. не патентные документы 2 и 3). Более того, эксперименты на животных подтвердили, что если кардиомиоциты, происходящие из ЭС клеток, трансплантировать во взрослую сердечную ткань, они являются высокоэффективными, демонстрируя результаты, сходные с теми, что получены при трансплантации миокарда плода (см. патентный документ 1, не патентный документ 4).
В 1995 г. Thomson et al. впервые получили линию ЭС клеток из приматов (см. патентный документ 2, не патентный документ 5), благодаря чему регенерационная терапия с использованием кардиомиоцитов, полученных из плюрипотентных стволовых клеток, стала реальностью. Впоследствии они также добились успеха в выделении и получении линий ЭС клеток человека из человеческих эмбрионов на ранней стадии развития (см. не патентный документ 6). Кроме того, Gearhart et al. получили линии ЭС клеток человека из человеческих примордиальных зародышевых клеток (см. не патентный документ 7, патентный документ 3).
Kehat et al. (см. не патентный документ 8) и Xu et al. (см. патентный документ 4, не патентный документ 9) сообщали, что человеческие ЭС клетки могут дифференцироваться в кардиомиоциты in vitro подобно мышиным ЭС клеткам. Согласно этим сообщениям кардиомиоциты, полученные из человеческих ЭС клеток, не только обладают способностью спонтанно ритмически сокращаться, но также экспрессируют и продуцируют специфические для миокарда белки, такие как тяжелые и легкие цепи миозина, α-актинин, тропонин I и предсердный натрийуретический пептид (ANP), а также специфические для миокарда факторы транскрипции, такие как GATA-4, Nkx2.5, MEF-2c и т.п., а из результатов гистологических исследований и электрофизиологического анализа следует, что они сохраняют свойства незрелых кардиомиоцитов на стадии плода и могут быть использованы для регенерационной терапии.
Однако, чтобы использовать кардиомиоциты, полученные из плюрипотентных стволовых клеток, для терапии методом клеточной трансплантации и других целей, следует решить одну серьезную проблему. Когда ЭТ образуются из ЭС клеток или ЭЗ клеток обычными способами, развиваются не только кардиомиоциты, но также и другие типы дифференцированных клеток, такие как клетки крови, клетки сосудов, нервные клетки, клетки кишечника, костные и хрящевые клетки и тому подобное. Кроме того, относительное содержание кардиомиоцитов в данной популяции дифференцированных клеток не слишком велико, всего лишь примерно от 5% до 20% от общего количества.
Способы выделения только кардиомиоцитов из смеси различных типов клеток включают способ введения искусственной модификации в гены ЭС клеток, придающей устойчивость к лекарственным средствам или эктопическую экспрессию, и отбор клеток, обладающих свойствами кардиомиоцитов или их клеток-предшественников. Например, путем введения генного кластера, способного экспрессировать ген устойчивости к неомицину (G418) под контролем промотора тяжелой цепи α-миозина, в мышиные ЭС клетки Field и его соавторы создали систему, в которой эти ЭС клетки могли выживать в среде, в которую добавлен G418, только когда они дифференцировались в кардиомиоциты и экспрессировали ген тяжелой цепи α-миозина (см. патентный документ 1, не патентный документ 4). Было подтверждено, что 99% или более G418-устойчивых клеток, отобранных с помощью данного способа, представляют собой кардиомиоциты. Однако, хотя в данном способе степень чистоты кардиомиоцитов является чрезвычайно высокой, количество кардиомиоцитов, полученных в итоге, составляет всего несколько процентов от общего количества клеток, что весьма затрудняет получение достаточных для трансплантации количеств кардиомиоцитов.
Xu et al. сообщали, что если человеческие ЭС клетки обработать 5-азацитидином, процентное содержание тропонин I-позитивных клеток (кандидатов в кардиомиоциты) в ЭТ возрастает с 15% до 44% (см. не патентный документ 9), но даже в данном способе процентное содержание кардиомиоцитов в ЭТ не превышает 50%. Кроме того, 5-азацитидин представляет собой деметилирующее вещество, которое изменяет экспрессию генов путем удаления метиловых групп, связанных с ДНК, и поскольку оно действует непосредственно на хромосомы, оно не является подходящим лекарственным средством для получения клеток, предназначенных для клеточной трансплантации.
Другие способы более эффективного получения кардиомиоцитов из ЭС клеток включают, в случае мышиных ЭС клеток, добавление ретиноевой кислоты (см. не патентный документ 10), аскорбиновой кислоты (см. не патентный документ 11), TGF β, BMP-2 (см. не патентный документ 12), PDGF (см. не патентный документ 13) и динорфина B (см. не патентный документ 14), а также обработку для увеличения содержания активных форм кислорода (ROS) (см. не патентный документ 15) и Ca2+ (см. не патентный документ 16) в клетках, все эти способы, как известно, положительно влияют на индукцию дифференциации кардиомиоцитов. Однако ни одним из данных способов не удается достичь специфической для кардиомиоцитов или избирательной дифференциации. Недавно исследовательская группа, включая авторов данного изобретения, продемонстрировала, что если ЭС клетки временно обработать антагонистом BMP, дифференциацию в кардиомиоциты можно индуцировать более эффективно и избирательно, чем обычными способами (патентный документ 5, не патентный документ 17).
(2) Функциональные роли белков Wnt в процессе дифференциации и развития кардиомиоцитов
Белки Wnt, которые являются секреторными белками, представляют собой членов семейной группы белков, которые широко представлены не только у позвоночных животных, но также у беспозвоночных животных, таких как нематоды и насекомые, и, как известно, их семейство генов имеет множество молекулярных видов (не патентные документы 18 и 19). Например, до настоящего времени были идентифицированы 19 генов Wnt у человека и мыши (Wnt-1, 2, 2b/13, 3, 3a, 4, 5a, 5b, 6, 7a, 7b, 8a, 8b, 9a, 9b, 10a, 10b, 11, 16). Белки Wnt, кодируемые данными генами Wnt, имеют различную тканевую специфичность, но являются сходными по структуре.
Если белки Wnt играют роль лигандов в межклеточных сигнальных системах, они связываются с расположенными на клеточной мембране рецепторами семейства Frizzled (далее сокращенное обозначение Fzd) с семью трансмембранными доменами. Существует несколько метаболических путей в нисходящем направлении от Fzd рецепторов и наиболее важный путь представляет собой ингибирование фосфорилирования β-катенина, опосредованное гликиген синтазой киназой (GSK)-3β. В отсутствие сигналов Wnt β-катенин захватывается вместе с GSK-3β аксином на белке Adenomatous polyposis coli (APC) и быстро фосфорилируется GSK-3β. Фосфорилированный β-катенин подвергается убиквитинированию и опосредованной протеасомами деградации.
С другой стороны, когда белки Wnt связываются с Fzd рецепторами, внутриклеточный фактор Dishevelled активируется для захвата GSK-3β, в результате чего β-катенин не фосфорилируется и остается в свободной форме в цитоплазме, а затем мигрирует в ядро. После миграции в ядро β-катенин связывается с лимфоидным энхансерным фактором-1/T-клеточным фактором (далее сокращенное обозначение LEF-1/TCF), присутствующими в ядре, с образованием транскрипционного активаторного комплекса, тем самым индуцируя транскрипцию целевого гена. Такой путь передачи сигнала, при котором происходит накопление и миграция в ядро β-катенина, называется «классическим» путем Wnt или каноническим сигнальным путем Wnt, а семейство видов молекул (например, Wnt-1, Wnt-3a, Wnt-8a), способных активировать данный путь, называется каноническими Wnt. Также известно, что активация канонического сигнального пути Wnt индуцируется обработкой различными ингибиторами GSK-3β.
Известно, что лиганды Wnt активируют не только β-катениновый путь, но также и другие сигнальные пути через Fzd рецепторы. Такие сигнальные пути включают путь планарной клеточной полярности (PCP), который активирует JNK (N-концевую киназу Jun), своего рода MAP-киназу, а также Ca2+-путь, который приводит к увеличению внутриклеточной концентрации Ca2+ и активирует протеинкиназу C путем активации тримерного G-белка и последующей активации фосфолипазы C (не патентные документы 19 и 20). Данные пути называют «неклассическими» путями Wnt или неканоническими сигнальными путями Wnt в отличие от канонического сигнального пути Wnt. Сообщают, что Wnt-4 и Wnt-11 представляют собой молекулы семейства Wnt, способные активировать такие пути, и эти лиганды Wnt действуют, ингибируя канонический сигнальный путь Wnt.
Следует отметить, что некоторые молекулярные виды белка Wnt обладают способностью активировать как канонический, так и неканонический пути в зависимости от типа клеток-мишеней и стадии их дифференциации, а также различий в Fzd рецепторах, экспрессированных в клетках-мишенях. Например, известно, что Wnt-5a действует как неканонический Wnt в обычно используемых системах анализа, таких как образование вторичной оси в эмбрионах Xenopus laevis и канцерогенез эпителиальных клеток молочной железы, в то время как сообщают также, что Wnt-5a индуцирует стабилизацию β-катенина и его транскрипционную активность в ЭС клетках, то есть активирует канонический сигнальный путь Wnt в ЭС клетках (не патентный документ 21).
Известно, что белки Wnt вовлечены в большое многообразие биологических функций на протяжении развития, роста и дифференциации различных клеток, тканей и злокачественных новообразований. Кардиомиоциты развиваются из части мезодермы боковой пластины на ранней стадии развития, а затем многократно делятся и растут, образуя сердце. Наличие или отсутствие сигналов Wnt играет важную роль в данном процессе, как продемонстрировано в разных случаях. В качестве примера, на ранней стадии развития птиц или Xenopus laevis, эктопическая и/или вынужденная экспрессия гена Wnt-3a или Wnt-8a, который активирует канонический сигнальный путь Wnt, существенно ингибирует формирование сердца (не патентные документы 22 и 23).
С другой стороны, так называемые антагонисты Wnt (например, Frzb, Dkk-1), которые связываются с Wnt-3a или Wnt-8a, ингибируя передачу ими сигнала, способствуют формированию сердца, из чего следует, что канонические сигналы Wnt ингибируют развитие миокарда.
Напротив, известно, что активация неканонических сигнальных путей Wnt, которые антагонистичны каноническим сигналам Wnt, способствует индукции развития и дифференциации кардиомиоцитов. Pandur et al. (не патентный документ 24) показали, что Wnt-11, который активирует неканонические пути без активирования канонического пути, является фактором, необходимым для развития сердца у Xenopus laevis. Впоследствии стимулирующее действие Wnt-11 было также подтверждено в системах индукции миокардиальной дифференциации для мышиных ЭС клеток (не патентный документ 25) и человеческих клеток-предшественников эндотелия сосудов (не патентный документ 26). Что касается активации неканонических сигнальных путей Wnt, то известно также, что можно индуцировать дифференциацию кардиомиоцитов из клеток ткани языка (патентный документ 6).
С другой стороны, в отличие от вышеизложенных случаев, известно, что активация канонического сигнального пути Wnt стимулирует миокардиальную дифференциацию из клеток эмбриональной карциномы (ЭК клетки). Клетки P19CL6, сублиния клеток P19, которые являются своего рода ЭК клетками, обладают свойством дифференцироваться в кардиомиоциты при стимуляции диметилсульфоксидом (DMSO). Когда в среду добавляли Wnt-3a или Wnt-8, клетки P19CL6 были стимулированы к дифференциации в кардиомиоциты поскольку β-катенин был стабилизирован (не патентный документ 27). В данной системе также показано, что период времени, достаточный для добавления белка Wnt, составляет 4 дня сразу же после индукции дифференциации (не патентный документ 28).
Клеточные линии P19 обладают характеристиками, частично схожими с таковыми у ЭС клеток в том, что их можно индуцировать для дифференциации в кардиомиоциты и нейроны. Однако клеточные линии P19 не обладают способностью дифференцироваться в различные клетки или способностью образовывать химеры в отличие от ЭС клеток. Кроме того, клеточные линии P19 сильно отличаются от ЭС клеток по маркерам клеточной поверхности, экспрессированным генам и так далее. То есть клеточные линии P19 можно использовать в качестве модельной системы для ЭС клеток в определенных экспериментах, но они не всегда обладают теми же характеристиками, что ЭС клетки. Таким образом, на основании научных данных было невозможно предсказать, можно ли результаты, полученные в данной экспериментальной системе, напрямую экстраполировать в системы индукции миокардиальной дифференциации для ЭС клеток и других плюрипотентных стволовых клеток.
Недавно сообщалось, что в экспериментальных системах с использованием мышиных ЭС клеток белок Wnt-3a, член канонического Wnt, стимулирует миокардиальную дифференциацию из ЭС клеток, будучи добавлен на 3 дня после начала индукции дифференциации (Naito A. et al., 28th Annual Meeting of the Molecular Biology Society of Japan, 2005.12.7 to 2005.12.10, Hakata, Japan; не патентный документ 30). Однако аналогичные исследования, проведенные авторами данного изобретения, свидетельствовали о том, что существенный стимулирующий эффект на дифференциацию отсутствовал (пример 2), и другие исследовательские группы также сообщали, что обработка мышиных ЭС клеток Wnt-3a не приводит к существенному эффекту на индукцию миокардиальной дифференциации (не патентный документ 25) или вызывает ингибиторный эффект на миокардиальную дифференциацию (не патентный документ 29). То есть, не ясно, как активированный канонический путь Wnt влияет на миокардиальную дифференциацию из ЭС клеток или других плюрипотентных стволовых клеток. При данных обстоятельствах не существует разработанного оптимального способа культивирования для индукции миокардиальной дифференциации.
Патентный документ 1: патент США № 6015671
Патентный документ 2: патент США № 5843780
Патентный документ 3: патент США № 6090622
Патентный документ 4: WO03/06950
Патентный документ 5: WO2005/033298
Патентный документ 6: JP 2005-224155 A
Не патентный документ 1: Soonpaa MH et al., Science, 264:98, 1994
Не патентный документ 2: Maltsev VA et al., Mechanism of Development, 44:41, 1993
Не патентный документ 3: Maltsev VA et al., Circulation Research, 75:233, 1994
Не патентный документ 4: Klug MG et al., Journal of Clinical Investigation, 98:216, 1996
Не патентный документ 5: Thomson JA et al., Proceedings of the National Academy of Sciences of the United States of America, 92:7844, 1995
Не патентный документ 6: Thomson JA et al., Science, 282:1145, 1998
Не патентный документ 7: Shamblott MJ et al., Proceedings of the National Academy of Sciences of the United States of America, 95:13726, 1998
Не патентный документ 8: Kehat I et al., Journal of Clinical Investigation, 108:407, 2001
Не патентный документ 9: Xu C et al., Circulation Research, 91:501, 2002
Не патентный документ 10: Wobus AM et al., Journal of Molecular and cellular Cardiology, 29:1525, 1997
Не патентный документ 11: Takahashi T et al., Circulation, 107:1912, 2003
Не патентный документ 12: Behfar A et al., FASEB Journal, 16:1558, 2002
Не патентный документ 13: Sachinidis et al., Cardiovascular Research, 58:278, 2003
Не патентный документ 14: Ventura C et al., Circulation Research, 92:623, 2003
Не патентный документ 15: Sauer H et al., FEBS Letters, 476:218, 2000
Не патентный документ 16: Li J et al., Journal of Cell Biology, 158:103, 2002
Не патентный документ 17: Yuasa S et al., Nature Biotechnology, 23:607, 2005
Не патентный документ 18: Nusse R, Cell Research, 15:28, 2005
Не патентный документ 19: Widelitz R, Growth Factors, 23:111, 2005
Не патентный документ 20: Kühl M et al., Trends in Genetics, 16:279, 2000
Не патентный документ 21: Hao J et al., Developmental Biology, 290:81, 2006
Не патентный документ 22: Schneider VA & Mercola M, Genes and development, 15:304, 2001
Не патентный документ 23: Marvin MJ et al., Genes and Development, 15:316, 2001
Не патентный документ 24: Pandur P et al., Nature, 418:636, 2002
Не патентный документ 25: Terami H et al., Biochemical and Biophysical Research Communication, 325:968, 2004
Не патентный документ 26: Koyanagi M et al., Journal of Biological Chemistry, 280:16838, 2005
Не патентный документ 27: Nakamura T et al., Proceedings of the National Academy of Sciences of the United States of America, 100:5834, 2003
Не патентный документ 28: Naito AT et al., Circulation Research, 97:144, 2005
Не патентный документ 29: Yamashita JK et al., FASEB Journal, 19:1534, 2002
Не патентный документ 30: Naito AT et al., Proceedings of the National Academy of Sciences of the United States of America, 103:19812, 2006.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
ПРОБЛЕМЫ, РЕШАЕМЫЕ С ПОМОЩЬЮ ДАННОГО ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу эффективной и избирательной индукции дифференциации недифференцированных плюрипотентных стволовых клеток в кардиомиоциты посредством активирования канонического сигнального пути Wnt, а также к кардиомиоцитам, полученным с помощью данного способа, и к способу использования данных клеток для клеточной трансплантации и инъекции, а также других видов терапии, направленной на заболевания сердца.
СПОСОБЫ РЕШЕНИЯ ПРОБЛЕМ
В качестве источника стволовых клеток для получения кардиомиоцитов авторы данного изобретения использовали плюрипотентные стволовые клетки, особенно ЭС клетки, которые наиболее часто используют, и в результате интенсивных исследований условий для индукции дифференциации в кардиомиоциты или их клетки-предшественники они совершили настоящее изобретение, когда обнаружили, что если вещество, которое стимулирует активацию канонического сигнального пути Wnt (далее называемое «активатор передачи сигнала Wnt»), добавляли к среде во время определенной ограниченной стадии клеточной культуры, популяции клеток, обладающих способностью ритмично сокращаться, которые были идентифицированы как кардиомиоциты, развивались гораздо более избирательно и эффективно, чем при обычно используемых способах.
Плюрипотентные стволовые клетки, которые можно использовать по настоящему изобретению, включают ЭС клетки, ЭЗ клетки и ЗС клетки, полученные от млекопитающих, таких как мыши, обезьяны и люди, а также все плюрипотентные стволовые клетки, которые сходны по характеристикам с ЭС клетками. В данном случае сходство по характеристикам с ЭС клетками определяют в контексте клеточно-биологических свойств, уникальных для ЭС клеток, таких как присутствие специфических для ЭС клеток поверхностных маркеров (антигенов), экспрессия специфических для ЭС клеток генов или способность образовывать тератомы или мышей-химер.
По настоящему изобретению конкретные примеры вещества, которое стимулирует активацию канонического сигнального пути Wnt, включают различные канонические белки Wnt, ингибиторы GSK-3β и другие низкомолекулярные соединения, способные активировать канонический сигнальный путь Wnt. Можно также использовать гены, способные активировать канонический сигнальный путь Wnt, например различные гены канонических Wnt, а также ген β-катенина или их активные мутанты, которые модифицированы для удаления N-конца или для замены сайтов фосфорилирования GSK-3β нефосфорилированными аминокислотами.
По настоящему изобретению канонические белки Wnt являются членами группы белков семейства Wnt и их определяют как вещества, которые связываются с рецепторами семейства Fzd и ингибируют опосредованное GSK-3β фосфорилирование β-катенина, тем самым стимулируя стабилизацию β-катенина и его способность активировать транскрипцию. Предпочтительные канонические белки Wnt по настоящему изобретению включают, например, Wnt-1, Wnt-3a, Wnt-5a и Wnt-8a, а также те, которые имеют гомологию аминокислотной последовательности по меньшей мере 80%, более предпочтительно, по меньшей мере 90% с данными белками и обладают способностью активировать β-катенин.
Одной из характерных особенностей настоящего изобретения является то, что ЭС клетки или другие плюрипотентные стволовые клетки временно стимулируют активатором передачи сигнала Wnt, и хотя способ стимулирования особо не ограничен, предпочтительным является способ культивирования клеток в среде, содержащей канонический белок Wnt, например рекомбинантный вариант белка (далее называемый «рекомбинантный белок Wnt»), полученный при экспрессии очищенного гена канонического Wnt. Канонический белок Wnt для использования и ген, кодирующий его, предпочтительно получают от животных тех же видов, которые использовали для получения плюрипотентных стволовых клеток, однако можно использовать и те, что получены от животных других видов. В случае использования рекомбинантного белка Wnt культуральную среду стерильно удаляют и заменяют свежей средой, содержащей рекомбинантный белок Wnt в концентрации от 0,1 нг/мл до 500 нг/мл, предпочтительно от 1 нг/мл до 200 нг/мл, более предпочтительно от 10 нг/мл до 100 нг/мл, и продолжают культивирование.
В соответствии с настоящим изобретением ингибиторы GSK-3β определяют как вещества, которые ингибируют киназную активность белка GSK-3β (например, способность фосфорилировать β-катенин), и к настоящему времени известны более нескольких десятков ингибиторов. Конкретные примеры включают производное индирубина BIO (также называемое ингибитор IX GSK-3β, 6-броминдирубин-3'-оксим), производное малеимида SB216763 (3-(2,4-дихлорфенил)-4-(1-метил-1H-индол-3-ил)-1H-пиррол-2,5-дион), фенил-α-бромметилкетоновое соединение ингибитор VII GSK-3β (4-дибромацетофенон), а также проникающий в клетку фосфорилированный пептид L803-mts (также называемый пептидный ингибитор GSK-3β, Myr-N-GKEAPPAPPQSpP-NH2). Данные соединения коммерчески доступны от Calbiochem или Biomol и легки в применении, однако список ими не ограничивается.
В случае использования этих ингибиторов GSK-3β, их оптимальная концентрация будет сильно варьировать в зависимости от различий в свойствах соединений. По этой причине необходимо определять оптимальную концентрацию каждого соединения, которое будет использовано. Например, в случае BIO или SB216763, для замены среды используют среду, содержащую ингибитор GSK-3β предпочтительно в концентрации от 10 нмоль/л до 1 мкмоль/л, более предпочтительно от 50 нмоль/л до 200 нмоль/л, и продолжают культивирование. При добавлении ингибитора VII GSK-3β концентрация предпочтительно составляет от 2 мкмоль/л до 100 мкмоль/л и более предпочтительно от 5 мкмоль/л до 20 мкмоль/л. При добавлении L803-mts концентрация предпочтительно составляет от 5 мкмоль/л до 500 мкмоль/л, более предпочтительно от 20 мкмоль/л до 200 мкмоль/л, и еще более предпочтительно от 25 мкмоль/л до 200 мкмоль/л.
Кроме ингибиторов GSK-3β, лекарственные средства, используемые для осуществления настоящего изобретения, могут представлять собой низкомолекулярные соединения, которые стимулируют активацию канонического сигнального пути Wnt (далее называемые «агонисты Wnt»). Предпочтительные примеры включают производное аминопиримидина (2-амино-4-[3,4-(метилендиокси)бензиламино]-6-(3-метоксифенил)пиримидин, Calbiochem) (Liu et al., Angew. Chem. Int. Ed. Engl. 44:1987, 2005). В случае использования такого агониста Wnt, для замены среды используют среду, содержащую агонист Wnt в концентрации от 1 нмоль/л до 1000 нмоль/л, предпочтительно от 10 нмоль/л до 500 нмоль/л, более предпочтительно от 50 нмоль/л до 200 нмоль/л, и продолжают культивирование.
Срок для обработки активатором передачи сигнала Wnt можно определить на основании схем экспрессии различных генов канонических Wnt в процессе дифференциации в плюрипотентных стволовых клетках, используемых для осуществления настоящего изобретения. Более конкретно, индуцируют дифференциацию плюрипотентных стволовых клеток обычным образом и из периодически отобранных образцов выделяют мРНК, чтобы анализировать уровни экспрессии различных генов канонических Wnt стандартными методами, такими как ОТ-ПЦР (RT-PCR). Момент времени, в который уровни экспрессии генов канонических Wnt существенно повышены после индукции дифференциации по сравнению с недифференцированными плюрипотентными столовыми клетками, определяют как «период повышенной экспрессии генов Wnt». Хотя для анализа можно использовать один ген канонического Wnt, предпочтительно использовать два или больше, более предпочтительно три или больше генов.
При осуществлении настоящего изобретения плюрипотентные стволовые клетки культивируют в среде, не содержащей активатора передачи сигнала Wnt на протяжении периода времени между моментом сразу после инициации культуры для индукции миокардиальной дифференциации и 24 часами до периода повышенной экспрессии генов Wnt, определенного как описано выше. Далее плюрипотентные стволовые клетки культивируют в среде, содержащей активатор передачи сигнала Wnt, предпочтительно в течение 24-96 часов, более предпочтительно в течение 48-72 часов, начиная с момента времени, составляющего от 24 до 0 часов, предпочтительно 24 часа до периода повышенной экспрессии генов Wnt, определенного как описано выше. Следует отметить, что период времени, в течение которого клетки обрабатывают активатором передачи сигнала Wnt, можно изменять для достижения оптимального периода (часы) в зависимости от различий в условиях, таких как виды животных, от которых получены клетки, которые будут использованы, тип используемой клеточной линии и/или тип используемого активатора передачи сигнала Wnt.
Кардиомиоциты, полученные из ЭС клеток или других плюрипотентных стволовых клеток вышеуказанным способом, затем можно собрать, выделить и очистить известными способами, чтобы эффективно получить большие количества высокоочищенных кардиомиоцитов. Полученные таким образом кардиомиоциты далее называются кардиомиоцитами, полученными в соответствии с настоящим изобретением.
Кардиомиоциты, полученные в соответствии с настоящим изобретением, представляют собой клетки, которые обладают морфологическими, физиологическими и/или иммуноцитологическими характеристиками кардиомиоцитов. В контексте физиологических и/или иммуноцитологических характеристик клетки, полученные в соответствии с настоящим изобретением, могут экспрессировать один или более маркеров, специфических для кардиомиоцитов, которые распознаются как кардиомиоциты, однако этим не ограничивается.
Кроме того, кардиомиоциты, полученные в соответствии с настоящим изобретением, можно использовать в методах скрининга, направленного на выявление потенциальных химиотерапевтических лекарственных средств или новых факторов, которые стимулируют развитие, дифференциацию, регенерацию, выживание, и тому подобное, кардиомиоцитов.
Кроме того, кардиомиоциты, полученные в соответствии с настоящим изобретением, можно использовать в способах лечения сердца, пораженного сердечными заболеваниями.
То есть настоящее изобретение относится, но без ограничения, к следующему.
(1) Способ индукции дифференциации кардиомиоцитов из плюрипотентных стволовых клеток, который включает:
i) культивирование плюрипотентных стволовых клеток в культуральной среде, не содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, на протяжении периода времени между началом индукции дифференциации и 24 часами перед периодом повышенной экспрессии генов канонических Wnt, а затем
ii) культивирование плюрипотентных стволовых клеток в культуральной среде, содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, на протяжении периода времени от 24 до 96 часов, начиная с 24 до 0 часов перед периодом повышенной экспрессии генов канонических Wnt.
(2) Способ по п. (1) выше, где плюрипотентные стволовые клетки культивируют в культуральной среде, содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, начиная с 24 часов перед периодом повышенной экспрессии генов канонических Wnt.
(3) Способ по п. (1) или (2) выше, где плюрипотентные стволовые клетки культивируют в культуральной среде, содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, на протяжении периода времени от 48 до 72 часов.
(4) Способ по любому из п.п. (1)-(3) выше, где вещество, которое стимулирует активацию канонического сигнального пути Wnt, представляет собой вещество, выбранное из группы, состоящей из канонического белка Wnt, ингибитора GSK3β и агониста Wnt.
(5) Способ по п. (4) выше, где вещество, которое стимулирует активацию канонического сигнального пути Wnt, представляет собой канонический белок Wnt.
(6) Способ по п. (5) выше, где канонический белок Wnt представляет собой, по меньшей мере, один белок Wnt, выбранный из группы, состоящей из Wnt-1, Wnt-3a и Wnt-5a.
(7) Способ по п. (5) или (6) выше, где концентрация канонического белка Wnt в культуральной среде составляет от 0,1 нг/мл до 500 нг/мл.
(8) Способ по п. (4) выше, где вещество, которое стимулирует активацию канонического сигнального пути Wnt, представляет собой ингибитор GSK3β.
(9) Способ по п. (8) выше, где ингибитор GSK3β представляет собой, по меньшей мере, один ингибитор, выбранный из группы, состоящей из ингибитора VII GSK3β, L803-mts, SB216763 и ингибитора IX GSK3β (BIO).
(10) Способ по п. (8) или (9) выше, где концентрация ингибитора GSK3β в культуральной среде составляет от 2 мкмоль/л до 100 мкмоль/л для ингибитора VII GSK3β, от 5 мкмоль/л до 500 мкмоль/л для L803-mts, от 10 нмоль/л до 1 мкмоль/л для SB216763 или от 10 нмоль/л до 1 мкмоль/л для ингибитора IX GSK3β (BIO).
(11) Способ по п. (4) выше, где вещество, которое стимулирует активацию канонического сигнального пути Wnt, представляет собой агонист Wnt.
(12) Способ по п. (11) выше, где агонист Wnt представляет собой производное аминопиримидина.
(13) Способ по п. (11) или (12) выше, где концентрация агониста Wnt в культуральной среде составляет от 1 нмоль/л до 1000 нмоль/л.
(14) Способ по любому из п.п. (1)-(13) выше, где плюрипотентные стволовые клетки представляют собой эмбриональные стволовые клетки, эмбриональные зародышевые клетки или зародышевые стволовые клетки.
(15) Способ по п. (14) выше, где плюрипотентные стволовые клетки представляют собой эмбриональные стволовые клетки.
(16) Способ по п. (14) или (15) выше, где плюрипотентные стволовые клетки человеческого происхождения.
ПРЕИМУЩЕСТВА ИЗОБРЕТЕНИЯ
Используя способ настоящего изобретения, можно очень эффективно и избирательно получать миокардиальные клетки-предшественники и кардиомиоциты из ЭС клеток и других плюрипотентных стволовых клеток. Клетки кардиомиоциты (предшественники), полученные способом настоящего изобретения, можно использовать для поиска и разработки эффективных лекарственных средств для лечения заболеваний сердца и потенциально можно применять для трансплантационной терапии миокарда в случае тяжелого сердечного заболевания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[Фиг. 1A] На фиг. 1A представлены изменения экспрессии гена Wnt в процессе индукции дифференциации в ЭС клетках. Символы на фигуре означают следующее. Белые кружки: необработанная группа, заштрихованные квадраты: группа, обработанная хордином, заштрихованные треугольники: группа, обработанная DAN. Вертикальные оси представляют собой сравнительное отношение уровней экспресии гена Wnt и гена GAPDH, использованного в качестве внутреннего стандарта. Аналогично, звездочкой (*) отмечен момент времени, в который уровень экспрессии гена Wnt был значительно повышен по сравнению с недифференцированными ЭС клетками.
[Фиг. 1B] На фиг. 1B представлены изменения экспрессии гена Wnt в процессе индукции дифференциации в ЭС клетках. Символы на фигуре означают следующее. Белые кружки: необработанная группа, заштрихованные квадраты: группа, обработанная хордином, заштрихованные треугольники: группа, обработанная DAN. Вертикальные оси представляют собой сравнительное отношение уровней экспресии гена Wnt и гена GAPDH, использованного в качестве внутреннего стандарта. Аналогично, звездочкой (*) отмечен момент времени, в который уровень экспрессии гена Wnt был значительно повышен по сравнению с недифференцированными ЭС клетками.
[Фиг. 1C] На фиг. 1C представлены изменения экспрессии гена Wnt в процессе индукции дифференциации в ЭС клетках. Символы на фигуре означают следующее. Белые кружки: необработанная группа, заштрихованные квадраты: группа, обработанная хордином, заштрихованные треугольники: группа, обработанная DAN. Вертикальные оси представляют собой сравнительное отношение уровней экспресии гена Wnt и гена GAPDH, использованного в качестве внутреннего стандарта. Аналогично, звездочкой (*) отмечен момент времени, в который уровень экспрессии гена Wnt был значительно повышен по сравнению с недифференцированными ЭС клетками.
[Фиг. 2A] На фиг. 2A показано влияние на появление ритмически сокращающихся ЭТ, вызванное различиями в сроках добавления рекомбинантного белка Wnt к культуральной среде.
[Фиг. 2B] На фиг. 2B показано влияние на появление ритмически сокращающихся ЭТ, вызванное различиями в сроках добавления рекомбинантного белка Wnt к культуральной среде.
[Фиг. 3A] На фиг. 3A представлена экспрессия маркерного гена, специфического для кардиомиоцитов, в ритмически сокращающихся ЭТ, которые появляются после индукции дифференциации в ЭС клетках. Вертикальная ось представляет собой коэффициент относительно уровня генной экспрессии в необработанной группе (ничего), который принят за 1.
[Фиг. 3B] На фиг. 3B представлена экспрессия маркерного гена, специфического для кардиомиоцитов, в ритмически сокращающихся ЭТ, которые появляются после индукции дифференциации в ЭС клетках. Вертикальная ось представляет собой коэффициент относительно уровня генной экспрессии в необработанной группе (ничего), который принят за 1.
[Фиг. 3C] На фиг. 3C представлена экспрессия маркерного гена, специфического для кардиомиоцитов, в ритмически сокращающихся ЭТ, которые появляются после индукции дифференциации в ЭС клетках. Вертикальная ось представляет собой коэффициент относительно уровня генной экспрессии в необработанной группе (ничего), который принят за 1.
[Фиг. 3D] На фиг. 3D представлена экспрессия маркерного гена, специфического для кардиомиоцитов, в ритмически сокращающихся ЭТ, которые появляются после индукции дифференциации в ЭС клетках. Вертикальная ось представляет собой коэффициент относительно уровня генной экспрессии в необработанной группе (ничего), который принят за 1.
[Фиг. 4] На фиг. 4 представлено иммуногистохимическое окрашивание маркерных белков, специфических для кардиомиоцитов, в ритмически сокращающихся ЭТ, которые появляются после индукции дифференциации в ЭС клетках.
[Фиг. 5A] На фиг. 5A показано влияние ингибитора GSK3β на появление ритмически сокращающихся ЭТ.
[Фиг. 5B] На фиг. 5B показано влияние ингибитора GSK3β на появление ритмически сокращающихся ЭТ.
[Фиг. 5C] На фиг. 5C показано влияние ингибитора GSK3β на появление ритмически сокращающихся ЭТ.
[Фиг. 5D] На фиг. 5D показано влияние ингибитора GSK3β на появление ритмически сокращающихся ЭТ.
[Фиг. 5E] На фиг. 5E показано влияние ингибитора GSK3β на появление ритмически сокращающихся ЭТ.
[Фиг. 6] На фиг. 6 показано изменение в экспрессии гена Wnt-3 в процессе индукции дифференциации в ЭС клетках обыкновенной игрунки (оиЭС) (обезьяна).
[Фиг. 7] На фиг. 7 представлена экспрессия маркерного гена, специфического для кардиомиоцитов, в ритмически сокращающихся ЭТ, которые появляются после индукции дифференциации в оиЭС клетках.
[Фиг. 8] На фиг. 8 представлено иммуногистохимическое окрашивание маркерных белков, специфических для кардиомиоцитов, в ритмически сокращающихся ЭТ, которые появляются после индукции дифференциации в оиЭС клетках.
СПОСОБЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже приведены способы осуществления изобретения, включая вышеуказанные эффекты настоящего изобретения, а также другие преимущества и характеристики.
При осуществлении настоящего изобретения каждый может обратиться к стандартным литературным источникам касательно методов генетической инженерии, таких как методики молекулярной биологии и рекомбинантной ДНК, а также обычных методов клеточной биологии и известного уровня техники, если не указано особо. Такие литературные источники включают, например, «Molecular Cloning: A Laboratory Manual, Third Edition» (Sambrook & Russell, Cold Spring Harbor Laboratory Press, 2001), «Current Protocols in Molecular biology» (Ausubel et al., eds., John Wiley & Sons, 1987), «Methods in Enzymology series» (Academic Press), «PCR Protocols: Methods in Molecular Biology» (Bartlett & Striling, eds., Humana Press, 2003), «Animal Cell Culture: A Practical Approach, Third Edition» (Masters, ed., Oxford University Press, 2000) и «Antibodies: A Laboratory Manual» (Harlow et al. & Lane, eds., Cold Spring Harbor Laboratory Press, 1987). Реагенты и наборы для культивирования клеток и клеточных биологических исследований, приведенные в данном описании, можно получить из коммерческих источников, включая компании Sigma, Aldrich, Invitrogen/GIBCO, Clontech, Stratagene и тому подобные.
Аналогичным образом, при осуществлении настоящего изобретения каждый может обратиться к стандартным литературным источникам касательно обычных методов культивирования клеток, а также относящихся к развитию и клеткам биологических исследований с использованием плюрипотентных стволовых клеток. Они включают «Guide to Techniques in Mouse Development» (Wasserman et al., eds., Academic Press, 1993), «Embryonic Stem Cell Differentiation in vitro» (M.V. Wiles, Meth. Enzymol. 225:900, 1993), «Manipulating the Mouse Embryo: A laboratory manual» (Hogan et al., eds., Cold Spring Harbor Laboratory Press, 1994) и «Embryonic Stem Cells» (Turksen ed., Humana Press, 2002). Реагенты и наборы для культивирования клеток, а также для относящихся к развитию и клеткам биологических исследований, приведенных в данном описании, можно получить из коммерческих источников, включая компании Invitrogen/GIBCO, Sigma и тому подобные.
Также разработаны стандартные протоколы для получения, субкультивирования и сохранения мышиных и человеческих плюрипотентных стволовых клеток, и в дополнение к вышеприведенным литературным источникам исполнитель может при использовании таких плюрипотентных стволовых клеток обращаться за информацией к различным другим литературным источникам. Такие литературные источники включают Matsui et al., Cell 70:841, 1992; Thomson et al., патент США № 5843780; Thomson et al., Science 282:114, 1998; Shamblott et al., Proc. Natl. Acad. Sci. USA 95:13726, 1998; Shamblott et al., патент США № 6090622; Reubinoff et al., Nat. Biotech. 18:399, 2000 и международная публикация № WO00/27995. Также известны способы создания культур ЭС клеток или ЭС-подобных клеток для других видов животных, например обезьян (Thomson et al., патент США № 5843780; Proc. Natl. Acad. Sci. USA, 92, 7844, 1996), крыс (Iannaccone et al., Dev. Biol. 163:288, 1994; Loring et al., международная публикация № WO99/27076), птиц (Pain et al., Development 122:2339, 1996; патент США № 5340740; патент США № 5656479) и свиней (Wheeler et al., Reprod. Fertil. Dev. 6:563, 1994; Shim et al., Biol. Reprod. 57:1089, 1997). В соответствии с данными описанными методами можно получать и применять ЭС клетки по настоящему изобретению.
В данном описании «кардиомиоциты» включают сердечные клетки-предшественники, обладающие способностью становиться в будущем функциональными кардиомиоцитами, а также фетальные или взрослые кардиомиоциты на всех стадиях дифференциации, и определяются как клетки, которые можно идентифицировать одним или, предпочтительно, более чем одним из следующих способов, используя один или, предпочтительно, более чем один маркер или показатель.
Экспрессию различных маркеров, специфических для кардиомиоцитов, выявляют общепринятыми биохимическими или иммунохимическими методами. Не существует определенных ограничений по методу, однако, предпочтительно использовать иммунохимический метод, такой как иммуногистохимическое окрашивание или иммуноэлектрофорез. В данных методах можно использовать специфические для маркера поликлональные антитела или моноклональные антитела, которые реагируют с сердечными клетками-предшественниками или кардиомиоцитами. Антитела для отдельных специфических маркеров коммерчески доступны и могут легко быть использованы. Маркеры, специфические для сердечных клеток-предшественников или кардиомиоцитов, включают, например, тяжелую и легкую цепи миозина, α-актинин, тропонин I, ANP, GATA-4, Nkx2.5, MEF-2c и т.п.
Альтернативно, хотя не существует определенных ограничений по методу, экспрессию маркерных генов, специфических для сердечных клеток-предшественников или специфических для кардиомиоцитов, можно также подтверждать полимеразной цепной реакцией с обратной транскрипцией (ОТ-ПЦР) или гибридизационным анализом, методами молекулярной биологии, которые в прошлом обычно использовали, чтобы амплифицировать, выявлять и анализировать мРНК, кодирующую любые маркерные белки. Последовательности нуклеиновых кислот, кодирующие маркерные белки, специфические для сердечных клеток-предшественников и кардиомиоцитов (такие как тяжелая и легкая цепи миозина, α-актинин, тропонин I, ANP, GATA-4, Nkx2.5 и MEF-2c) уже известны и доступны через публичные базы данных, такие как GenBank Национального центра биотехнологической информации (National Center for Biotechnology Information (NCBI)), и специфические для маркеров последовательности, необходимые для использования в качестве праймеров или зондов, могут быть легко определены.
Кроме того, можно также использовать физиологические показатели, чтобы подтвердить дифференциацию плюрипотентных клеток в кардиомиоциты. Например, полезные маркеры включают спонтанные ритмические сокращения клеток, полученных из плюрипотентных клеток, экспрессию различных ионных каналов и способность реагировать на электрофизиологический стимул.
Способ по настоящему изобретению можно применять для плюрипотентных стволовых клеток из любого вида млекопитающих. Например, способ по настоящему изобретению можно применять для плюрипотентных стволовых клеток, полученных от мышей, коров, коз, собак, кошек, игрунок, макак-резусов или людей, однако список не ограничен плюрипотентными стволовыми клетками, полученными из этих видов животных. Примеры плюрипотентных стволовых клеток, доступных для использования по настоящему изобретению, включают ЭС клетки, полученные от млекопитающих, таких как мыши, обезьяны и люди, которые уже широко используют в качестве клеточных культур.
Конкретные примеры ЭС клеток, полученных от мышей, включают клетки EB3, клетки E14, клетки D3, клетки CCE, клетки R1, клетки 129SV, клетки J1 и т.п. Мышиные ЭС клетки для использования по настоящему изобретению можно получить, например, из Американской коллекции типовых культур (American Type Culture Collection (ATCC)), Chemicon или Cell & Molecular Technologies.
Сообщалось, что культуры обезьяньих ЭС клеток были получены от макак-резусов (Macaca mulatta) (Thomson et al., Proc. Natl. Acad. Sci. USA 92:7844, 1995), яванских макак (Macaca fascicularis) (Suemori et al., Dev. Dyn. 222:273, 2001) и обыкновенных игрунок (Callithrix jacchus) (Sasaki et al., Stem Cells. 23:1304, 2005), и их можно использовать. Например, ЭС клетки игрунок можно также получить из Central Institute for Experimental Animals, Япония.
В настоящее время во всем мире было получено более нескольких десятков человеческих линий ЭС клеток. Например, многие клеточные линии внесены в список US National Institutes of Health (http://stemcells.nih.gov /registry/ index.asp) и могут быть использованы, в то время как данные клеточные линии можно также получить, например, от Cellartis, ES Cell International или Wisconsin Alumni Research Foundation. В Японии человеческие линии ЭС клеток можно также получить из Stem Cell Research Center, Institute for Frontier Medical Sciences, Kyoto University (Suemori et al., Biochem. Biophys. Res. Commun., 345:926, 2006).
Кроме того, сообщали о создании культуры ЭС клеток коров (Mitalipova et al., Cloning 3:59, 2001), птиц (Petitte et al., Mech. Dev. 121:1159, 2004) и данио (Fishman, Science 294:1290, 2001).
Хотя культуру ЭС клеток, как правило, получают культивируя эмбрионы на ранней стадии развития, ЭС клетки можно также получать из эмбрионов на ранней стадии развития, которые были модифицированы трансплантацией ядер, чтобы иметь ядро соматических клеток (Munsie et al., Curr. Biol. 10:989, 2000; Wakayama et al., Science 292:740, 2001; Hwang et al., Science 303:1669, 2004). Кроме того, сообщают о попытке получения ЭС клеток из партеногенетических эмбрионов, которые развились до стадии, эквивалентной стадии бластоцисты (патентная публикация США № 02/168763; Vrana K et al., Proc. Natl. Acad. Sci. USA 100:11911-6), а также о способе получения ЭС клеток, обладающих генетической информацией ядра соматической клетки в результате слияния между ЭС клетками и соматическими клетками (международная публикация № WO00/49137; Tada et al., Curr. Biol. 11:1553, 2001). ЭС клетки для использования по настоящему изобретению также включают ЭС клетки, полученные таким образом, или ЭС клетки, хромосомные гены которых модифицированы методами генетической инженерии.
Плюрипотентные стволовые клетки, применимые в способе по настоящему изобретению, не ограничиваются ЭС клетками и также включают все плюрипотентные стволовые клетки, сходные по характеристикам с ЭС клетками, которые получены, например, из клеток взрослого органа или ткани млекопитающего, клеток костного мозга и клеток крови, а также из клеток эмбриона или плода млекопитающего. В данном случае сходство по характеристикам с ЭС клетками определяют в контексте цитобиологических свойств, уникальных для ЭС клеток, таких как присутствие специфических для ЭС клеток поверхностных маркеров (антигенов), экспрессия специфических для ЭС клеток генов или способность образовывать тератомы или мышей-химер. Конкретные примеры включают ЭЗ клетки, полученные из примордиальных зародышевых клеток, ЗС клетки, полученные из тестикулярных зародышевых клеток и индуцированные плюрипотентные стволовые клетки (иПС (iPS) клетки), полученные из соматических клеток, таких как фибробласты, путем особых генетических процедур.
Любой способ, пригодный для индукции дифференциации кардиомиоцитов, можно использовать в качестве способа культивирования для получения кардиомиоцитов из ЭС клеток или других плюрипотентных стволовых клеток по настоящему изобретению, и примеры включают суспензионную культуру, культуру в висячей капле, совместную культуру с поддерживающими клетками, ротационную культуру, культуру в мягком агаре, культуру микроносителей и тому подобное. Конкретным примером является способ суспендирования ЭС клеток в виде одиночных клеток (отдельные клетки диспергированы в жидкой фазе без межклеточной адгезии в результате обработки ферментами или тому подобного) в среде до плотности клеток, равной 1×103-1×105 клеток/мл и осаждение капли суспензии объемом 10-100 мкл на внутренней стороне крышки культурального планшета для создания эффекта культуры в висячей капле. Альтернативно, вышеуказанную клеточную суспензию можно высевать в коммерчески доступный планшет, такой как 96-луночный культуральный планшет для образования сфероида (например, Sumilon Celltight Spheroid; Sumitomo Bakelite Co., Ltd., Japan), культуральный планшет без клеточной адгезии (например, Coaster ultra-low-attachment plate; Corning) или необработанный полистирольный планшет. Суспензию, содержащую ЭС клетки, затем культивируют при 37°C в атмосфере CO2 с аэрацией 5% двуокисью углерода, в результате чего ЭС образуются и индуцируются для дифференциации в кардиомиоциты или другие клетки.
В настоящем изобретении активация канонического сигнального пути Wnt означает состояние, при котором β-катенин не фосфорилирован с помощью GSK-3β и стабилизирован в цитоплазме и/или ядре, и/или состояние, при котором β-катенин связывается с LEF-1/TCF в ядре с образованием транскрипционного активаторного комплекса и, вследствие этого, обладает способностью индуцировать транскрипцию целевого гена. Для того чтобы определить, активирован ли канонический сигнальный путь Wnt, можно использовать любой способ, включая, но не ограничиваясь им, способ измерения количества цитоплазматического и/или ядерного β-катенина, например, путем иммуногистологического окрашивания с помощью антител, специфических для β-катенина, или анализа вестерн-блоттингом. Кроме того, моноклональные антитела, специфически распознающие нефосфорилированный β-катенин, то есть активный β-катенин, также коммерчески доступны и крайне полезны. Помимо этого, эффективны также анализы с использованием репортеров, в которых репортерный ген присоединен ниже LEF-1/TCF-связывающей последовательности, а способность производить продукт репортерного гена применяется в анализе в качестве маркера. Плазмиду, содержащую LEF-1/TCF-связывающую последовательность и ген-репортер, для использования в таких анализах можно приобрести у Upstate под торговым названием TOPflash.
Конкретные примеры активаторов передачи сигнала Wnt включают различные канонические белки Wnt, ингибиторы GSK-3β и агонисты Wnt. Также можно использовать гены, способные активировать канонический сигнальный путь Wnt, например различные гены канонических Wnt, а также ген β-катенина или их активные мутанты, которые модифицированы для удаления N-конца или для замены сайтов фосфорилирования GSK-3β нефосфорилированными аминокислотами. Альтернативно, экспрессию таких генов, как Axin или APC, которые подавляют канонический сигнальный путь Wnt, можно супрессировать или блокировать с помощью специфических антисмысловых ДНК или рибозимов, антисмысловых РНК для РНК-интерференции, низкомолекулярных соединений и так далее. Необходимо отметить, что нуклеотидные последовательности генов, кодирующих эти молекулы, доступны в таких публичных базах данных по ДНК, как принадлежащая NCBI, и любые специалисты в данной области смогут получить, создать и использовать кДНК, миРНК (siRNAs) и/или антисмысловые ДНК этих генов.
Канонические белки Wnt, пригодные для использования по настоящему изобретению, являются членами группы белков семейства Wnt и их определяют как вещества, которые связываются с рецепторами семейства Fzd и ингибируют опосредованное GSK-3β фосфорилирование β-катенина, тем самым способствуя стабилизации β-катенина и его способности активировать транскрипцию. Предпочтительные канонические белки Wnt по настоящему изобретению включают, например, Wnt-1 (SEQ ID №: 1), Wnt-3a (SEQ ID №: 2), Wnt-5a (SEQ ID №: 3) и Wnt-8a (SEQ ID №: 4), а также те, которые имеют гомологию аминокислотной последовательности по меньшей мере 80%, более предпочтительно, по меньшей мере 90% с данными белками и обладают способностью активировать β-катенин.
Одной из характерных особенностей настоящего изобретения является то, что ЭС клетки или другие плюрипотентные стволовые клетки временно стимулируют активатором передачи сигнала Wnt, и несмотря на то, что способ стимулирования особо не ограничен, предпочтительным является способ культивирования клеток в среде, содержащей канонический белок Wnt, например рекомбинантный белок Wnt. Однако можно использовать любой другой способ, оказывающий такое же действие. Примеры включают способ культивирования клеток в присутствии канонического белка Wnt, который экстрагирован и очищен из живых тканей, способ введения экспрессирующего вектора, несущего ген, кодирующий канонический белок Wnt, в сами плюрипотентные стволовые клетки, способ введения такого экспрессирующего вектора в поддерживающие клетки и использования таких трансфецированных клеток в качестве совместно культивируемых клеток и способ, использующий культуральный супернатант, или другой клеточный продукт таких трансфецированных клеток и тому подобное, которые все включены как часть варианта осуществления для добавления канонического белка Wnt в среду в способе по настоящему изобретению.
При осуществлении настоящего изобретения канонический белок Wnt для использования и ген, кодирующий его, предпочтительно получают от животных тех же видов, которые использовали для получения плюрипотентных стволовых клеток, однако можно использовать и те, что получены от животных других видов. Например, если в настоящем изобретении используют ЭС клетки мыши или ЭС клетки обезьяны, то можно использовать человеческий белок WNT-1. В качестве рекомбинантных белков Wnt мышиные Wnt-3a и Wnt-5a, а также человеческий WNT-7A коммерчески доступны от компании R&D Systems, а человеческий WNT-1 коммерчески доступен от компании Peprotech. Такие рекомбинантные белки Wnt легко использовать. В случае использования таких рекомбинантных белков культуральную среду стерильно удаляют и заменяют новой средой, содержащей рекомбинантный белок Wnt в концентрации от 0,1 нг/мл до 500 нг/мл, предпочтительно от 1 нг/мл до 200 нг/мл, более предпочтительно от 10 нг/мл до 100 нг/мл, и продолжают культивирование.
В случае, если желательный белок Wnt получен самостоятельно, необходимо осуществить введение и экспрессию экспрессирующего вектора, несущего интересующий ген, в животных клетках (например, L-клетках) и очистить рекомбинантный белок, секретируемый в культуральный супернатант, поскольку известно, что белки Wnt не проявляют своей биологической активности, не будучи модифицированы пальмитиновой кислотой. Подробные методики для этой цели уже известны (Willert et al., Nature 423:448, 2003; Kishida et al., Mol. Cell. Biol. 24:4487; http://www.stanford.edu/~rnusse/ wntwindow.html).
Необходимо отметить, что нуклеотидные последовательности генов, кодирующих эти факторы, доступны в таких публичных базах данных по ДНК, как принадлежащая NCBI, и любые специалисты в данной области смогут получить и использовать кДНК этих генов. Например, гены Wnt-3a и Wnt-8a уже идентифицированы у людей и мышей, и нуклеотидные последовательности человеческого WNT-3A (SEQ ID №: 5), мышиного Wnt-3a (SEQ ID №: 2), человеческого WNT-8A (SEQ ID №: 6) и мышиного Wnt-8a (SEQ ID №: 4) зарегистрированы под входящими номерами NM_033131, NM_009522, NM_031933 и NM_009290 соответственно.
Ингибиторы GSK-3β согласно настоящему изобретению определяются как вещества, ингибирующие киназную активность белка GSK-3β (например, способность фосфорилировать β-катенин), и более чем несколько десятков ингибиторов уже известны (Martinez et al., Med. Res. Rev. 22:373, 2002; Meijer L et al., Trends Pharmacol. Sci. 25:471, 2004). Конкретные примеры включают литий, вальпроевую кислоту, членов семейства бензазепинонов кенполлон (9-бром-7,12-дигидроиндол[3,2-d][1]бензазепин-6(5H)он) и альстерполлон (9-нитро-7,12-дигидроиндол[3,2-d][1]бензазепин-6(5H)он), производные индирубина 5-хлориндирубин, индирубин-3'-моноксим и BIO (также называемый ингибитором IX GSK-3β, 6-броминдирубин-3'-оксим), производные малеимида SB216763 (3-(2,4-дихлорфенил)-4-(1-метил-1H-индол-3-ил)-1H-пиррол-2,5-дион) и SB415286 (3-[(3-хлор-4-гидроксифенил)амино]-4-(2-нитрофенил)-1H-пиррол-2,5-дион), аналоги тиадиазолидинона (TDZD) TDZD-8 (также называемый ингибитором I GSK-3β, 4-бензил-2-метил-1,2,4-тиадиазолидин-3,5-дион) и OTDZT (также называемый ингибитором III GSK-3β; 2,4-дибензил-5-оксотиадиазолидин-3-тион), фенил-α-бромметилкетоновое соединение ингибитор VII GSK-3β (4-дибромацетофенон) и проникающий в клетку фосфорилированый пептид L803-mts (также называемый пептидным ингибитором GSK-3β; Myr-N-GKEAPPAPPQSpP-NH2). Такие соединения коммерчески доступны от Calbiochem или Biomol и легки в применении, однако список ими не ограничивается.
В случае использования этих ингибиторов GSK-3β, их оптимальная концентрация будет сильно варьировать в зависимости от различий в свойствах соединений. По этой причине необходимо определять оптимальную концентрацию каждого соединения, которое будет использовано, и для культивирования использовать среду, содержащую ингибитор GSK-3β в нужной концентрации.
Например, в случае BIO или SB216763, для культивирования используют среду, содержащую ингибитор при концентрации предпочтительно от 10 нмоль/л до 1 мкмоль/л, более предпочтительно от 50 нмоль/л до 200 нмоль/л. В случае ингибитора VII GSK-3β, его концентрация предпочтительно составляет от 2 мкмоль/л до 100 мкмоль/л, и более предпочтительно от 5 мкмоль/л до 20 мкмоль/л. Аналогичным образом, в случае L803-mts его концентрация предпочтительно составляет от 5 мкмоль/л до 500 мкмоль/л, более предпочтительно от 20 мкмоль/л до 200 мкмоль/л, и еще более предпочтительно от 25 мкмоль/л до 200 мкмоль/л.
Кроме ингибиторов GSK-3β, лекарственные средства, используемые для осуществления настоящего изобретения, могут представлять собой низкомолекулярные соединения, которые способствуют активации канонического сигнального пути Wnt (агонисты Wnt), включая органические или неорганические соединения и пептидные фрагменты. Предпочтительные примеры включают производное аминопиримидина (2-амино-4-[3,4-(метилендиокси)бензиламино]-6-(3-метоксифенил)пиримидин, (Calbiochem) (Liu et al., Angew. Chem. Int. Ed. Engl. 44:1987, 2005). В случае использования такого агониста Wnt для культивирования применяли среду, содержащую агонист Wnt при концентрациях от 1 нмоль/л до 1000 нмоль/л, предпочтительно от 10 нмоль/л до 500 нмоль/л, более предпочтительно от 50 нмоль/л до 200 нмоль/л.
Определение срока, во время которого плюрипотентные стволовые клетки обрабатывают активатором передачи сигнала Wnt, является очень важным требованием для осуществления настоящего изобретения. А именно, при неправильно выбранном сроке активатор передачи сигнала Wnt не оказывает стимулирующего действия или даже может иметь ингибирующий эффект на способность плюрипотентных стволовых клеток дифференцироваться в миокард. В качестве примера, если плюрипотентные стволовые клетки культивируют в течение примерно 1 недели в культуральной среде с добавлением активатора передачи сигнала Wnt, начиная сразу после индукции дифференциации, способность к миокардиальной дифференциации может быть ниже, чем наблюдаемая в группе (необработанная группа), где используют культуральную среду, не содержащую дополнительный ингредиент.
Срок для обработки активатором передачи сигнала Wnt можно определить на основании схем экспрессии различных генов канонических Wnt в процессе индукции дифференциации в плюрипотентных стволовых клетках, используемых для осуществления настоящего изобретения. Более конкретно, можно индуцировать дифференциацию плюрипотентных стволовых клеток обычным образом и из периодически отобранных образцов можно выделять мРНК, чтобы анализировать уровни экспрессии различных генов канонических Wnt стандартными методами, такими как ОТ-ПЦР (RT-PCR). Образцы отбирают предпочтительно каждые 24 часа, более предпочтительно каждые 12 часов, в течение периода времени между началом культивирования для индукции дифференциации и появлением (ритмически сокращающихся) кардиомиоцитов, например, приблизительно от 6 до 14 дней для мышиных, обезьяньих и человеческих ЭС клеток. Хотя для анализа можно использовать один ген канонического Wnt, предпочтительно использовать два или больше, более предпочтительно три или больше генов.
В ЭС клетках и других плюрипотентных стволовых клетках экспрессия различных генов канонических Wnt обычно низка как при недифференцированном состоянии, так и сразу после индукции дифференциации, однако их экспрессия быстро возрастает в течение нескольких дней после индукции дифференциации (пример 1). Следовательно, момент времени, в который уровни экспрессии генов канонических Wnt существенно повышены после индукции дифференциации по сравнению с недифференцированными плюрипотентными стволовыми клетками, определяют как «период повышенной экспрессии генов Wnt». Значительное возрастание экспрессии генов можно определять с помощью обычных статистических тестов, таких как t-тест Стьюдента (уровень достоверности 5%). Уровень достоверности, используемый в данном случае в качестве критерия, предпочтительно составляет 5%, более предпочтительно 1%. Альтернативно, когда измеряемая экспрессия генов канонических Wnt быстро возрастает в течение нескольких дней после индукции дифференциации, а затем исчезает в течение нескольких дней, то есть когда гены канонических Wnt демонстрируют повышенную экспрессию только в течение короткого периода, момент времени, в который они достигают максимальных уровней экспрессии, может быть определен как период повышенной экспрессии генов Wnt.
Если плюрипотентные стволовые клетки культивируют в среде, содержащей антагонист BMP, начиная за 2 или 3 дня до индукции дифференциации и/или начиная сразу после индукции дифференциации, известно, что их способность к миокардиальной дифференциации значительно возрастает (WO2005/033298; Yuasa et al., Nat. Biotechnol. 23:607, 2005). В этом случае было установлено, что вышеуказанные различные гены канонических Wnt демонстрируют повышенную экспрессию во время культивирования. Этот факт полезен при определении периода повышенной экспрессии генов канонических Wnt по настоящему изобретению и желательно использовать среду, содержащую антагонист BMP для культивирования при определении периода повышенной экспрессии. Антагонистом BMP называется вещество, которое связывает молекулу BMP (например, BMP-2, BMP-4, BMP-7), чтобы ингибировать сигнализацию BMP, и примеры включают ноггин, хордин и DAN. Эти вещества, которые могут быть добавлены к среде, можно приобрести, например, у R&D systems.
В настоящем изобретении плюрипотентные стволовые клетки культивируют в среде, не содержащей активатора передачи сигнала Wnt на протяжении периода времени между моментом сразу после инициации культуры для индукции миокардиальной дифференциации и 24 часами до периода повышенной экспрессии генов Wnt, определенного, как описано выше. Затем клетки культивируют дальше в среде, содержащей активатор передачи сигнала Wnt, в течение 24-96 часов, предпочтительно в течение 48-72 часов, начиная с момента времени, составляющего от 24 до 0 часов, предпочтительно 24 часа до периода повышенной экспрессии генов Wnt, определенного, как описано выше. Например, в одном случае ЭС клеток мыши, культивируемых для индукции миокардиальной дифференциации, экспрессия типичных генов канонических Wnt Wnt-3, Wnt-3a и Wnt-8a является крайне низкой, как в недифференцированном состоянии, так и сразу после индукции дифференциации, однако эти гены демонстрируют сильную экспрессию между 72 и 96 часами после индукции дифференциации (пример 1). По этой причине, в случае, если ЭС клетки мыши используют в способе по настоящему изобретению, период повышенной экспрессии генов канонических Wnt определен как 72 часа после индукции дифференциации и с этого времени клетки культивируют в среде, не содержащей активатор передачи сигнала Wnt, вплоть до 48 часов после начала индукции дифференциации. Затем клетки культивируют дальше в среде, содержащей активатор передачи сигнала Wnt, в течение 24-96 часов, предпочтительно в течение 48-72 часов, начиная от 48 часов после начала индукции дифференциации. Необходимо отметить, что период времени (часы), в течение которого клетки обрабатывают активатором передачи сигнала Wnt, может быть доведен при необходимости до оптимального периода (часов), в зависимости от различий в таких условиях, как видовая принадлежность животных, из которых происходят используемые клетки, тип используемой клеточной линии и/или тип используемого активатора передачи сигнала Wnt, и такой период (часы) можно определять на основе периода повышенной экспрессии генов канонических Wnt, полученного указанным выше способом определения сроков обработки активатором передачи сигнала Wnt. Например, в случае ЭС клеток обезьяны (обыкновенная игрунка), ген Wnt-3 демонстрирует значительную экспрессию между 72 и 120 час после индукции дифференциации (пример 5). Аналогичным образом, в случае человеческих ЭС клеток ген Wnt-3а демонстрирует экспрессию с пиком около 72 часов после индукции дифференциации (Beqqali et al., Stem Cells 24:1956, 2006).
Кардиомиоциты, полученные из ЭС клеток или других плюрипотентных стволовых клеток с помощью вышеуказанного способа, могут в дальнейшем быть собраны, выделены и очищены с помощью известных методов для того, чтобы эффективно получить большие количества высокоочищенных кардиомиоцитов (кардиомиоциты, полученные в соответствии с настоящим изобретением).
В качестве способа очистки кардиомиоцитов можно использовать любой известный способ выделения и очистки клеток, и конкретные примеры включают проточную цитометрию, магнитные гранулы, пэннинг и другие способы, основанные на взаимодействии антиген-антитело (см. «Monoclonal Antibodies: principles and practice, Third Edition» (Acad. Press, 1993); «Antibody Engineering: A Practical Approach» (IRL Press at Oxford University Press, 1996), а также клеточное фракционирование с помощью центрифугирования в градиенте плотности с использованием таких носителей, как сахароза, перколл или тому подобное. Другой способ селекции кардиомиоцитов заключается в том, чтобы сначала искусственно ввести модификации в гены ЭС клеток или других плюрипотентных стволовых клеток, делая их устойчивыми к лекарственным средствам или способными к эктопической экспрессии белка, и собрать клетки, обладающие морфологией кардиомиоцитов. Например, путем введения генного кластера, способного экспрессировать ген устойчивости к неомицину (G418) под контролем промотора тяжелой цепи α-миозина, в мышиные ЭС клетки Field и его соавторы смогли создать систему, в которой ЭС клетки дифференцировались в кардиомиоциты, и только те клетки, которые экспрессировали ген тяжелой цепи α-миозина, могли выживать в среде, в которую был добавлен G418, и было подтверждено, что 99% или более клеток, отобранных этим способом как клетки, устойчивые к G418, являлись кардиомиоцитами (патент США № 6015671; Klug et al., J. Clin. Invest. 98: 216, 1996). В качестве другого примера эффективным является также способ, основанный на более высоком содержании митохондрий в кардиомиоцитах по сравнению с прочими клетками, в котором богатую митохондриями популяцию клеток, а именно кардиомиоциты, специфически собирали, используя избирательный для митохондрий флуоресцентный краситель, или реагент, чувствительный к мембранному потенциалу митохондрий (WO2006/022377). В качестве еще одного примера также предпочтительным является способ, основанный на специфических метаболических свойствах кардиомиоцитов, в котором кардиомиоциты специфически очищают в условиях пониженного содержания глюкозы путем добавления молочной кислоты или аминокислоты, такой как аспарагиновая кислота (японская патентная заявка № 2006-23770).
Кардиомиоциты, полученные в соответствии с настоящим изобретением, полезны для фармакологической оценки и оценки активности различных биологически активных веществ (например, лекарственных средств) и новых генных продуктов с неизвестной функцией. Например, их можно использовать для скрининга веществ и лекарственных средств, вовлеченных в контроль дифференциации кардиомиоцитов из ЭС клеток и других плюрипотентных стволовых клеток, веществ и лекарственных средств, вовлеченных в регуляцию функции кардиомиоцитов, а также веществ и лекарственных средств, которые токсичны или являются ингибиторами для кардиомиоцитов. В частности, в настоящее время существует крайне мало способов скрининга с использованием человеческих кардиомиоцитов, и кардиомиоциты, полученные в соответствии с настоящим изобретением, являются полезным источником клеток для осуществления таких способов скрининга. В другом варианте для такого скрининга также полезен испытательный набор, содержащий кардиомиоциты, полученные в соответствии с настоящим изобретением.
Тестируемые вещества для скрининга могут включать любые, которые можно добавлять к культуре, такие как низкомолекулярные соединения, высокомолекулярные соединения, органические соединения, неорганические соединения, белки, пептиды, гены, вирусы, клетки, жидкости клеточных культур, жидкости культур микроорганизмов и тому подобное. Эффективные способы введения генов в культуральные системы включают способы добавления в культуральные системы с использованием ретровирусов, аденовирусов и других вирусных векторов, а также способы добавления после встраивания в липосомы и другие искусственные конструкции.
Тестируемое вещество можно оценивать, определяя эффективность индукции дифференциации из ЭС клеток или других плюрипотентных стволовых клеток в кардиомиоциты, либо качественные или количественные изменения функций клеток миокарда. Например, эффективность тестируемого вещества для индукции миокардиальной дифференциации можно определять, используя биохимические или иммунохимические средства для выявления экспрессии различных специфических для кардиомиоцитов маркеров в плюрипотентных стволовых клетках, культивируемых с помощью способа по настоящему изобретению, после того, как их культивировали в течение 5-15, или предпочтительно, 7-12 дней. Не существует особых ограничений в отношении биохимических или иммунохимических средств, но предпочтительно можно использовать такой иммунохимический метод, как иммуногистохимическое окрашивание или иммуноэлектрофорез. В данных методах можно использовать специфические к маркеру поликлональные антитела или моноклональные антитела, которые связываются с кардиомиоцитами. Антитела, направленные на отдельные специфические маркеры, коммерчески доступны и могут легко быть использованы. Примеры специфических для кардиомиоцитов маркеров включают тяжелые и легкие цепи миозина, α-актинин, тропонин I, ANP, GATA-4, Nkx2.5, MEF-2c и тому подобное.
Выживаемость клеток миокарда представляет собой один из примеров функций клеток миокарда, которая может быть использована в качестве маркера для оценки тестируемого вещества. Конкретно, гибель клеток (апоптоз) можно вызвать посевом кардиомиоцитов, полученных способом по настоящему изобретению, на культуральную чашку до подходящей плотности клеток и культивированием их в бессывороточной среде, и в этом случае соответствующее количество тестируемого вещества можно добавлять к среде и определять коэффициент выживаемости или коэффициент гибели кардиомиоцитов. Коэффициент выживаемости или коэффициент гибели кардиомиоцитов можно определять макроскопическим наблюдением, используя в качестве маркера внедрение такого красителя, как трипановый синий, способом, в котором в качестве маркера используют дегидрогеназную активность (восстановительную активность), или способом, в котором в качестве маркера используют экспрессию аннексина V или каспазную активность, которые специфичны для апоптозных клеток. Наборы, основанные на таких механизмах, доступны от многих производителей, включая Sigma, Clonetech и Promega, и просты в применении.
Поскольку вещество или лекарственное средство, полученное таким методом скрининга, действует, индуцируя дифференциацию кардиомиоцитов и регулируя их функции, оно может быть использовано, например, в качестве профилактического или терапевтического лекарственного средства при болезнях сердца, включая инфаркт миокарда, ишемическую болезнь сердца, застойную сердечную недостаточность, гипертрофическую кардиомиопатию, расширяющуюся кардиомиопатию, миокардит, хроническую сердечную недостаточность и т.п. Такие соединения могут быть новыми соединениями или известными соединениями.
Кроме того, кардиомиоциты, полученные в соответствии с настоящим изобретением, могут быть использованы в качестве регенерирующих лекарственных средств для миокарда или лекарственных средств для лечения сердечных заболеваний. Примеры сердечных заболеваний включают инфаркт миокарда, ишемическую болезнь сердца, застойную сердечную недостаточность, гипертрофическую кардиомиопатию, расширяющуюся кардиомиопатию, миокардит, хроническую сердечную недостаточность и тому подобное. Будучи использованы в качестве регенерирующих лекарственных средств для миокарда или лекарственных средств для лечения сердечных заболеваний, кардиомиоциты, полученные в соответствии с настоящим изобретением, можно вводить в любой форме при условии высокой степени их очистки, такой как клетки, суспендированные в среде или другом водном носителе, клетки, заключенные в биодеградируемый субстрат или другую основу, либо клетки, полученные в виде однослойного или многослойного миокардиального листа (Shimizu et al., Circ. Res. 90:e40, 2002).
Способы доставки вышеуказанного терапевтического лекарственного средства к поврежденному участку включают, хотя и не ограничиваются ими, прямую инъекцию в сердце посредством вскрытия грудной клетки, либо шприцем, способы трансплантации посредством хирургического разреза на сердце, и способы трансплантации через кровеносные сосуды с помощью катетера (Murry et al., Cold Spring Harb. Symp. Quant. Biol. 67:519, 2002; Menasche, Ann. Thorac. Surg. 75:S20, 2003; Dowell et al., Cardiovasc. Res. 58:336, 2003). Сообщали, что очень хорошие терапевтические эффекты были получены, когда кардиомиоциты, собранные из сердца плода, трансплантировали такими способами в сердца животных с сердечными нарушениями (Menasche, Ann. Thorac. Surg. 75:S20, 2003; Reffelmann et al., Heart Fail. Rev. 8:201, 2003). Кардиомиоциты, полученные из ЭС клеток, обладают характеристиками, в высшей степени сходными с таковыми для кардиомиоцитов, полученных из сердца плода (Maltsev et al., Mech. Dev. 44:41, 1993; Circ. Res. 75:233, 1994; Doevendans et al., J. Mol. Cell. Cardiol. 32:839, 2000). Кроме того, крайне высокая степень приживаемости, эквивалентная той, что достигается при трансплантации миокарда плода, была подтверждена в экспериментах на животных, во время которых кардиомиоциты, полученные из ЭС клеток, фактически трансплантировали во взрослые сердца (Klug et al., J. Clin. Invest. 98:216, 1996; Laflamme et al., Am. J. Pathol. 167:663). Вследствие этого ожидается, что восполнительная трансплантация кардиомиоцитов, полученных в соответствии с настоящим изобретением, в пораженные сердечные ткани будет стимулировать улучшение сердечных функций в случаях вышеупомянутых заболеваний сердца, обусловленных разрушениями или утратой клеток сердца.
ПРИМЕРЫ
Настоящее изобретение более детально поясняется ниже с помощью примеров.
Пример 1: Изучение схем экспрессии различных генов Wnt в процессе индукции дифференциации в ЭС клетках (1)
Экспрессию различных генов Wnt изучали в процессе дифференциации в ЭС клетках мыши. Для использования в экспериментах ЭС клетки мыши пассировали и поддерживали в недифференцированном состоянии в соответствии с методами, описанными в «Manipulating the Mouse Embryo: A Laboratory Manual» (Hogan et al., eds., Cold Spring Harbor Laboratory Press, 1994) и «Embryonic Stem Cells: Methods and Protocols» (Turksen ed., Humana Press, 2002), используя среду Knockout-DMEM (Invitrogen), содержащую 20% фетальной бычьей сыворотки, 2 ммоль/л L-глутамин и 0,1 ммоль/л 2-меркаптоэтанол (далее называемую ESM), к которой добавляли 1000 ед/мл LIF (ESGRO; Chemicon). ЭС клетки, пассированные в таких условиях, далее называются «ЭС клетки, пассированные в обычных условиях культивирования». ЭС клетки мыши, использованные в следующих экспериментах, представляли собой клетки D3, клетки R1 и клетки 129SV (приобретенные у Dainippon Pharmaceutical Co., Ltd., Japan), но, как правило, не существовало различий в результатах экспериментов между этими линиями ЭС клеток. Если не указано иначе, ниже представлены экспериментальные данные, полученные на клеточной линии D3. Следует отметить, что в экспериментах примеров 1-4 использовали ЭС клетки мыши.
ЭС клетки, пассированные в обычных условиях культивирования, промывали дважды фосфатно-солевым буфером (далее называемом PBS) и обрабатывали 0,25% раствором трипсина, содержащим 1 ммоль/л EDTA, чтобы получить отдельные клетки, которые затем суспендировали в ESM. Если не указано иначе, применяли одинаковые условия при снятии ЭС клеток с чашек для использования в экспериментах по индукции дифференциации и других экспериментах.
Культивирование для индукции дифференциации ЭС клеток в кардиомиоциты или нейроны осуществляли обычным способом, как описано далее. ЭС клетки суспендировали в не содержащей LIF среде и полученную суспензию высевали при 500 клеток/50 мкл в лунку коммерчески доступного 96-луночного культурального планшета для образования сфероидов (Sumilon Celltight Spheroid; Sumitomo Bakelite Co., Ltd., Japan). В этих экспериментальных условиях ЭС клетки начинали агрегировать и образовывать ЭТ сразу после культивирования в суспензии и некоторые ЭТ начинали демонстрировать спонтанные ритмические сокращения примерно через 7 или 8 дней после культивирования в неприкрепленном агрегированном состоянии (индукция дифференциации), свидетельствуя о том, что как минимум часть ЭТ дифференцировались в кардиомиоциты.
В этом эксперименте некоторые из экспериментальных групп получали в качестве добавки в среду коммерчески доступные рекомбинантный хордин или белок DAN (15 нг/мл; оба приобретены у R&D systems) за три дня до или сразу после индукции дифференциации. Известно, что при такой непродолжительной обработке антагонистом BMP возрастает способность ЭС клеток к миокардиальной дифференциации (WO2005-033298; Yuasa et al., Nat. Biotechnol. 23:607, 2005). Обработка ЭС клеток в среде с добавкой антагониста BMP, такого как белок хордин или белок DAN, далее называется «обработка антагонистом BMP».
Полученные таким образом ЭТ периодически собирали и с помощью набора RNeasy mini kit (Qiagen) получали суммарную РНК с последующей обработкой ДНКазой. Из обработанной ДНКазой суммарной РНК (1 мкг) синтезировали кДНК с помощью SuperScriptTM First-Strand Synthesis System for RT-PCR (Invitrogen). Анализ экспрессии генов проводили с помощью ABI PRISM 7700 (PE Applied Biosystems), применяя систему количественной оценки полимеразной цепной реакции (ПЦР) в реальном времени с Lux праймерами, чтобы изучить уровень экспрессии каждого гена. Количественную оценку ПЦР в реальном времени проводили с помощью вышеуказанной кДНК в качестве матрицы и используя Platinum Quantitative PCR SuperMix-UDG (Invitrogen) согласно приложенным инструкциям.
Для выявления различных генов Wnt были разработаны Lux праймеры с помощью программного обеспечения для разработки праймеров (D-LUXTM Designer; Invitrogen) на основе информации по нуклеотидным последовательностям генов. Нуклеотидные последовательности Lux праймеров, использованных для выявления различных генных транскриптов Wnt, представлены ниже.
Wnt-3
(Прямая) 5'-CAACAGTAGCAAGGAGCATGGACTGTTG-3' (SEQ ID №: 7)
(Обратная) 5'-GGCTGGGTCCAGGTCGTTTA-3' (SEQ ID №: 8)
Wnt-3a
(Прямая) 5'-GACAAACCGGGAGTCAGCCTTTGTC-3' (SEQ ID №: 9)
(Обратная) 5'-TGCTGCACCCACAGATAGCA-3' (SEQ ID №: 10)
Wnt-8a
(Обратная) 5'-GTACATGCGCTCTGCTGCCATCATGTAC-3' (SEQ ID №: 11)
(Прямая) 5'-GACTCGTCACAGCCGCAGTT-3' (SEQ ID №: 12)
На фиг. 1 представлен один из примеров экспериментов, проведенных так, как описано выше. Экспрессию генов Wnt изучали между 24 часами (1 день) и 168 часами (7 дней) после индукции дифференциации в ЭС клетках и установили, что экспрессия генов Wnt-3, Wnt-3a и Wnt-8a значительно повышена. Каждый из этих генов Wnt демонстрировал пик интенсивной экспрессии между 72 и 96 часами после индукции дифференциации и затем их экспрессия значительно снижалась, начиная со 120 часов после индукции дифференциации. Таким образом, можно определить, что в данных ЭС клетках период повышенной экспрессии генов Wnt приходится на 72 часа после индукции дифференциации.
Группы, обработанные антагонистом BMP, таким как белок хордин или белок DAN, демонстрировали сильное повышение экспрессии генов Wnt через 72 часа после индукции дифференциации, как в случае необработанной группы, и было обнаружено, что уровни экспрессии генов Wnt были значительно выше, чем у необработанной группы. Эти результаты свидетельствуют о том, что обработка антагонистом BMP является способом, способным более точно определять период повышенной экспрессии генов Wnt в процессе дифференциации ЭС клеток.
Пример 2: Усиливающий эффект обработки рекомбинантным белком Wnt на появление кардиомиоцитов, происходящих из ЭС клеток (1)
На ранней стадии дифференциации в ЭС клетках наблюдались временные увеличения экспрессии различных генов Wnt перед появлением кардиомиоцитов. Затем ЭС клетки на этой стадии обрабатывали рекомбинантными белками Wnt, чтобы изучить индуцирующий миокардиальную дифференциацию эффект этих белков. Индукцию дифференциации ЭС клеток осуществляли таким же образом, что и в примере 1, за исключением того, что некоторые из экспериментальных групп культивировали в среде, содержащей коммерчески доступные рекомбинантные белки WNT-1 (Peprotech), Wnt-3a (R&D systems) или Wnt-5a (R&D systems). Обработка ЭС клеток в среде, содержащей рекомбинантный канонический белок Wnt, такой как WNT-1, далее называется «обработка Wnt».
Частоту появления ЭТ, демонстрирующих спонтанные ритмические сокращения, исследовали периодически как один из полезных показателей дифференциации и развития кардиомиоцитов из ЭС клеток. В необработанной группе частота появления ритмически сокращающихся ЭТ составляла примерно 20% на 13 день после культивирования в суспензии, тогда как в группе, подвергнутой обработке Wnt в течение 48 часов (2 дня) между 48 и 96 часами после индукции дифференциации (Wnt48~96ч), а также в группе, подвергнутой обработке Wnt в течение 72 часов (3 дня) между 48 и 120 часами после индукции дифференциации (Wnt48~120ч), ритмические сокращения наблюдали у значительно большего процента ЭТ (фиг.2A и 2B). Эффект обработки Wnt был настолько высок, что был сравним с обработкой антагонистом BMP (на чертежах - «хордин»).
Напротив, в группе, подвергнутой обработке Wnt в течение первых 48, 72, 96 или 120 часов (2, 3, 4 или 5 дней) после индукции дифференциации (Wnt~48ч, Wnt~72ч, Wnt~96ч или Wnt~120ч соответственно), а также в группе, подвергнутой обработке Wnt от 120 или 144 часов (5 или 6 дней) после индукции дифференциации (Wnt120ч~ или Wnt144ч~), ЭТ со способностью к ритмическим сокращениям появлялись на том же уровне, что и у необработанной группы. Более того, у ЭТ из необработанной группы и других групп с малой долей ритмически сокращающихся ЭТ сокращения были ограничены определенными узкими областями ЭТ, в то время как было обнаружено, что ЭТ из групп Wnt48~96ч и Wnt48~120ч демонстрируют ритмические сокращения на протяжении практически всех областей своего поверхностного слоя, как в случае обработанных хордином ЭТ. То есть, даже когда ЭС клетки культивировали в среде, содержащей белок Wnt, на протяжении периода времени между моментом сразу после индукции дифференциации и моментом за 24 часа до периода повышенной экспрессии генов Wnt (то есть спустя 72 часа после начала индукции дифференциации) отсутствовал значительный эффект на индукцию миокардиальной дифференциации.
Напротив, когда ЭС клетки культивировали в среде, содержащей рекомбинантный белок Wnt, в течение 48 часов (2 дня) или 72 часов (3 дня), начиная с момента за 24 часа до периода повышенной экспрессии генов Wnt (то есть спустя 72 часа после начала индукции дифференциации), отмечался значительный эффект усиления на способность к миокардиальной дифференциации.
Данные результаты свидетельствовали о том, что хотя обработка Wnt значительно индуцировала миокардиальную дифференциацию из ЭС клеток, ее эффект наблюдался только в течение очень ограниченного периода времени в процессе индукции дифференциации. В следующих экспериментах термин «обработка Wnt» применяется для обозначения обработки Wnt в течение 48 часов (2 дней) между 48 и 96 часами после начала индукции дифференциации или в течение 72 часов (3 дней) между 48 и 120 часами после начала индукции дифференциации, если только не указано иное.
Были проведены дальнейшие исследования по «обработке Wnt», чтобы изучить, как различия в концентрациях добавленных рекомбинантных белков Wnt затрагивают способность к миокардиальной дифференциации. В качестве примера, в случае использования Wnt-3a, Wnt-5a и WNT-1, они демонстрировали почти одинаковую концентрационную зависимость, и частота появления ритмически сокращающихся ЭТ была значительно выше, чем в необработанной группе, когда рекомбинантные белки добавляли в концентрациях от 1 нг/мл до 100 нг/мл. В частности, очень хороший результат с точки зрения ритмически сокращающихся ЭТ был получен при добавлении белков Wnt в концентрациях от 10 нг/мл до 50 нг/мл.
Пример 3: Свойства кардиомиоцитов, происходящих от обработанных Wnt ЭС клеток.
Как показано в примере 2, ритмические сокращения ЭТ, полученных из ЭС клеток, значительно возрастают в результате обработки Wnt, и чтобы подтвердить, что ритмически сокращающиеся клетки в таких ЭТ являются кардиомиоцитами, были проведены дальнейшие исследования по изучению генной экспрессии и белковой продукции различных специфических для миокарда маркерных молекул. Таким же способом, как и в примере 2, ЭС клетки индуцировали для дифференциации и образовавшиеся ЭТ собирали через 10 дней после индукции дифференциации для получения кДНК. Проводили количественную оценку ПЦР в реальном времени методом TaqMan-зондирования, то есть используя вышеуказанную кДНК (1 мкл) в качестве матрицы и применяя TaqMan Universal PCR Master Mix (PE Applied Biosystems) согласно приложенной инструкции. Были разработаны TaqMan-зонды для детекции различных генов с помощью программного обеспечения для разработки праймеров (ABI PRISM Primer Express) на основании информации по нуклеотидным последовательностям генов. Нуклеотидные последовательности праймеров и TaqMan-зондов, использованных для детекции транскриптов GATA-4, Nkx-2.5, MLC-2a, MLC-2v и GAPDH, представлены ниже.
GATA-4
(Прямая) 5'-ACGGAAGCCCAAGAACCTGA-3' (SEQ ID №: 13),
(Обратная) 5'-CATTGCTGGAGTTACCGCTG-3' (SEQ ID №: 14),
(TaqMan-зонд) 5'-TAAATCTAA GACGCCAGCAGGTCCTGCTG-3' (SEQ ID №: 15);
Nkx-2.5
(Прямая) 5'-TGACCCAGCCAAAGACCCT-3' (SEQ ID №: 16),
(Обратная) 5'-CCATCCGTCTCGGCTTTGT-3' (SEQ ID №: 17),
(TaqMan-зонд) 5'-CGGATAAAAAAGA GCTGTGCGCGC-3' (SEQ ID №: 18);
MLC-2a
(Прямая) 5'-CCAGGCAGACAAGTTCTCTCCT-3' (SEQ ID №: 19),
(Обратная) 5'-CTTGTAGTCAATGTTGCCGGC-3' (SEQ ID №: 20),
(TaqMan-зонд) 5'-CAACTGTTTGCGCTGACACCCATGGA-3' (SEQ ID №: 21);
MLC-2v
(Прямая) 5'-GCAGAGAGGTTCTCCAAAGAGG-3' (SEQ ID №: 22),
(Обратная) 5'-AAGATTGCCGGTAACGTCAGG-3' (SEQ ID №: 23),
(TaqMan-зонд) 5'-ATCGACCAGATGTTCGCAGCCTTTCC-3' (SEQ ID №: 24)
GAPDH
(Прямая) 5'-TGCACCACCAACTGCTTAG-3' (SEQ ID №: 25),
(Обратная) 5'-GGATGCAGGGATGATGTTC-3' (SEQ ID №: 26),
(TaqMan-зонд) 5'-CAGAAGACTGTG GATGGCCCCTC-3' (SEQ ID №: 27)
По сравнению с необработанной группой, ЭТ в обработанной Wnt группе (группа Wnt48~120ч) на 10 день после индукции дифференциации демонстрировали значительно более интенсивную экспрессию генов GATA-4, Nkx-2.5, MLC-2a и MLC-2v (Фиг. 3), а также генов αMHC и βMHC, каждый из которых известен как типичный маркер кардиомиоцитов.
Напротив, в группах Wnt~48ч, Wnt~120ч и Wnt~144ч, отличающихся низкой частотой появления ритмически сокращающихся ЭТ, уровни экспрессии маркерных генов были почти такими же, как у необработанной группы, и в значительной степени такие же тенденции наблюдали для частоты появления ритмически сокращающихся ЭТ и уровней экспрессии различных миокардиальных маркерных генов.
Далее проводили иммуногистохимическое окрашивание для подтверждения того, что ритмически сокращающиеся клетки, возникшие в ЭТ из обработанных Wnt групп, продуцируют специфические для кардиомиоцитов маркерные белки. ЭТ в обработанной Wnt группе (группа Wnt48~120ч) на 10 день после индукции дифференциации сразу погружали в состав для приготовления замороженных срезов (OCT Compound, Sakura Finetek USA Inc.) и затем замораживали жидким азотом. Замороженные образцы нарезали на срезы толщиной 6 мкм и помещали на предметные стекла. Такие замороженные срезы подвергали реакции с антителом против саркомерного миозина (MF20, American Type Culture Collection), антителом против GATA-4 (C-20, Santa Cruz) или антителом против Nkx-2.5 (N-19, Santa Cruz) в качестве первичного антитела, а затем реакции со вторичным антителом, меченым пероксидазой хрена (Bio-RAD), и в конце подвергали цветной реакции с субстратным раствором ACE (3-амино-9-этилкарбазол) (Nichirei Corporation, Japan) и рассматривали под оптическим микроскопом.
Полученные результаты представлены на фиг. 4. В необработанной группе клетки, позитивные по специфическим для кардиомиоцитов маркерным белкам саркомерному миозину (на фиг. 4 - «MHC»), Nkx-2.5 и GATA-4, наблюдали у крайне ограниченного числа ЭТ. Напротив, было обнаружено, что в обработанной Wnt группе значительное большинство клеток, составляющих ЭТ, являются позитивными по маркерным белкам, и было подтверждено, что они образуют кардиосферы, как в случае группы, обработанной антагонистом BMP (DAN). Эти результаты подтвердили, что ритмически сокращающиеся клетки, происходящие из обработанных Wnt ЭС клеток, являлись кардиомиоцитами, и также свидетельствовали, что данный способ в значительной степени стимулирует дифференциацию и развитие кардиомиоцитов в ЭТ.
Пример 4: Усиливающий эффект активатора β-катенина на появление кардиомиоцитов, происходящих из ЭС клеток.
Известно, что описанная выше обработка каноническими белками Wnt ингибирует действие GSK-3β внутри клеток и тем самым стимулирует стабилизацию β-катенина и его способность активировать транскрипцию. Затем были проведены дальнейшие исследования, чтобы подтвердить, что обработка различными лекарственными средствами, способными стимулировать стабилизацию β-катенина и его способность активировать транскрипцию, производит индуцирующий миокардиальную дифференциацию эффект на ЭС клетки, как в случае обработки Wnt. В качестве лекарственных средств, способных стимулировать стабилизацию β-катенина и его способность активировать транскрипцию, были использованы следующие пять коммерчески доступных соединений: ингибиторы GSK3β BIO (Calbiochem), ингибитор VII GSK3β (Calbiochem), проникающий в клетку пептидный ингибитор GSK3β (L803-mts, Calbiochem) и SB216763 (Biomol), а также агонист Wnt (Calbiochem), который стимулирует способность к активации транскрипции β-катенина без ингибирования GSK3β. ЭС клетки индуцировали для дифференциации таким же образом, что и в приведенных выше примерах, и культивировали в среде, содержащей вышеуказанные соединения, в течение периода времени между 48 и 120 часами (3 и 5 днями) после индукции дифференциации, как в случае рекомбинантных белков Wnt.
Как показано на фиг. 5, результаты подтверждают, что группы, обработанные вышеуказанными соединениями, при своих оптимальных концентрациях обладали сильным индуцирующим миокардиальную дифференциацию эффектом, сравнимым или даже превышающим таковой у групп, обработанных рекомбинантными белками Wnt. Тем же способом, что и в примере 3, обработанные такими соединениями ЭТ исследовали на их экспрессию миокардиальных маркерных генов и миокардиальных маркерных белков, убеждаясь в том, что они демонстрировали значительно больший эффект повышения генной и белковой экспрессии, чем необработанная группа, как в случае групп, обработанных рекомбинантными белками Wnt.
Пример 5: Исследование схем экспрессии генов Wnt в процессе индукции дифференциации в ЭС клетках (2)
ЭС клетки, полученные от обыкновенной игрунки вида обезьяны (далее называемые «оиЭС клетки»), использовали для изучения экспрессии генов Wnt в процессе их дифференциации. Для использования в экспериментах оиЭС клетки пассировали и поддерживали в недифференцированном состоянии на обработанных митомицином первичных эмбриональных фибробластах мыши, которые были высеяны в качестве питающих клеток, применяя среду Knockout-DMEM (Invitrogen), содержащую 20% нокаутный заменитель сыворотки (Invitrogen), 0,1 ммоль/л раствор второстепенных аминокислот MEM, 1 ммоль/л L-глутамин и 0,1 ммоль/л 2-меркаптоэтанол (далее называемую «оиЭС среда»), с добавками 10 нг/мл рекомбинантного LIF (Alomone Labs) и 10 нг/мл рекомбинантного основного фактора роста фибробластов (Invitrogen).
Культивирование для индукции дифференциации в оиЭС клетках выполняли обычным способом, как указано далее. Клетки оиЭС промывали PBS и затем обрабатывали коммерчески доступным раствором клеточной диссоциации для ЭС клеток приматов (ReproCELL) при 37°C в течение 5 минут, чтобы собрать клеточную суспензию, содержащую агрегаты оиЭС клеток. Затем, чтобы отделить оиЭС клетки от питающих клеток, клеточную суспензию пропускали через сито с размером пор 100 мкм и прошедшую через него клеточную фракцию затем пропускали через сито с размером пор 40 мкм, чтобы отобрать не прошедшую фракцию, оставшуюся на сите. Такую не прошедшую фракцию, содержащую агрегаты оиЭС клеток, в дальнейшем высевали на коммерчески доступные культуральные планшеты с высокой клеточной адгезией (Primaria; Becton Dickinson) и культивировали в течение 30 минут, после чего собирали клеточные агрегаты, свободно плавающие в среде без прикрепления к планшету. Полученные таким способом агрегаты оиЭС клеток индуцировали для дифференциации в ЭТ путем культивирования в коммерчески доступных культуральных планшетах без клеточной адгезии (HydroCell; CellSeed), заполненных оиЭС средой, в то же время предохраняя клеточные агрегаты от контактов и слипания между собой.
Полученные таким образом ЭТ периодически собирали и получали суммарную РНК с помощью набора RNeasy mini kit (Qiagen). Затем с помощью обратной транскриптазы синтезировали кДНК и анализировали ПЦР, чтобы детектировать экспрессию гена Wnt-3 (оиWnt-3) и β-актина (оиβактин) обыкновенной игрунки, служащих внутренним контролем. Использованные для детекции праймеры представлены ниже.
оиWnt-3
(Прямая) 5'-GAGGTGAAGACCTGCTGGTGGGC-3' (SEQ ID №: 28)
(Обратная) 5'-GTTGGGCTCACAAAAGTTGG-3' (SEQ ID №: 29)
оиβактин
(Прямая) 5'-TCCTGACCCTGAAGTACCCC-3' (SEQ ID №: 30)
(Обратная) 5'-GTGGTGGTGAAGCTGTAGCC-3' (SEQ ID №: 31)
На фиг. 6 представлен один из примеров экспериментов, проведенных, как описано выше. Экспрессию генов изучали между 24 часами (1 день) и 168 часами (7 дней) после индукции дифференциации в оиЭС клетках, установив, что ген Wnt-3 демонстрировал пик интенсивной экспрессии между 72 и 120 часами после индукции дифференциации, а затем его экспрессия исчезала (Фиг. 6). Таким образом, можно определить, что у данных ЭС клеток период повышенной экспрессии гена Wnt-3 приходится на 72 часа после индукции дифференциации, тем самым получив почти такие же результаты, как для ЭС клеток мыши.
Пример 6: Усиливающий эффект обработки рекомбинантными белками Wnt на появление кардиомиоцитов, происходящих из ЭС клеток (2).
Для изучения эффекта обработки рекомбинантными белками Wnt использовали оиЭС клетки. Таким же способом, что и в примере 5, оиЭС клетки культивировали и индуцировали для дифференциации. В данном случае некоторые экспериментальные группы культивировали в течение 72 часов (3 дня) между 48 и 120 часами после индукции дифференциации в среде, содержащей коммерчески доступные рекомбинантные белки WNT-1 (PeproTech), Wnt-3a (R&D systems) или WNT-7A (R&D systems).
Чтобы подтвердить дифференциацию и развитие кардиомиоцитов из оиЭС клеток, полученные в результате ЭТ изучали с точки зрения их способности к спонтанным ритмическим сокращениям, а также исследовали экспрессию генов и белков различных специфических для миокарда маркерных молекул. В необработанной группе 10% или менее от всех ЭТ демонстрировали частичные ритмические сокращения примерно через 2 недели после индукции дифференциации. Напротив, в группах, обработанных Wnt, спонтанные ритмические сокращения начиналась примерно через 10 дней после индукции дифференциации и почти половина ЭТ демонстрировала ритмические сокращения через 16 дней после индукции дифференциации.
Кроме того, для анализа экспрессируемых генов ЭТ собирали через 10 дней после индукции дифференциации для детекции экспрессии различных маркерных генов таким же образом, что и в примере 5. Праймеры, использованные для детекции транскриптов нестина, ANP, MLC-2a и MLC-2v обыкновенной игрунки (далее называемые оиНестин, оиANP, оиMLC-2a и оиMLC-2v соответственно), представлены ниже.
оиНестин
(Прямая) 5'-GCCCTGACCACTCCAGTTTA-3' (SEQ ID №: 32),
(Обратная) 5'-GGAGTCCTGGATTTCCTTCC-3' (SEQ ID №: 33)
оиANP
(Прямая) 5'-GAACCAGAGGGGAGAGACAGA-3' (SEQ ID №: 34),
(Обратная) 5'-CCCTCAGCTTGCTTTTTAGGAG-3' (SEQ ID №: 35)
оиMLC-2a
(Прямая) 5'-GAGGAGAATGGCCAGCAGGAA-3' (SEQ ID №: 36),
(Обратная) 5'-GCGAACATCTGCTCCACCTCA-3' (SEQ ID №: 37)
оиMLC-2v
(Прямая) 5'-AGGAGGCCTTCACTATCATGG-3' (SEQ ID №: 38),
(Обратная) 5'-GTGATGATGTGCACCAGGTTC-3' (SEQ ID №: 39)
При сравнении с необработанной группой ЭТ в группе, обработанной Wnt-3a, через 10 дней после индукции дифференциации демонстрировали значительно более интенсивную экспрессию типичных маркерных генов оиANP, оиMLC-2a и оиMLC-2v кардиомиоцитов (Фиг. 7). Схожие результаты были также получены для ЭТ в группе, обработанной WNT-1.
Напротив, экспрессия оиНестина, известного как нейронный маркер, была значительно снижена в группах, обработанных Wnt.
Далее проводили иммуногистохимическое окрашивание таким же образом, как и в примере 2, для подтверждения того, что ритмически сокращающиеся клетки, возникшие в ЭТ из обработанных Wnt групп, являлись кардиомиоцитами, продуцирующими специфические маркерные белки. Из ЭТ в обработанных Wnt группах (Wnt-3a, WNT-1) через 16 дней после индукции дифференциации получали замороженные срезы и проводили реакцию с антителом против саркомерного миозина, антителом против GATA-4 или антителом против Nkx-2.5 в качестве первичного антитела. После цветной реакции такие срезы рассматривали под оптическим микроскопом.
В результате, в необработанной группе клетки, позитивные по специфическим для кардиомиоцитов маркерным белкам саркомерному миозину и GATA-4, наблюдались в крайне ограниченном числе ЭТ, и было немного клеток, позитивных по Nkx-2.5. Напротив, в группах, обработанных Wnt-3a или WNT-1, было обнаружено, что значительное большинство клеток, составляющих ЭТ, являлись позитивными по таким маркерным белкам (Фиг. 8). Схожие результаты были получены для ЭТ в группе, обработанной WNT-7A.
Данные результаты свидетельствуют, что обработка Wnt имела значительный стимулирующий эффект на индукцию миокардиальной дифференциации не только в ЭС клетках грызунов, но также и в ЭС клетках приматов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СТИМУЛЯЦИЯ ПУТИ Wnt ПРИ ПЕРЕПРОГРАММИРОВАНИИ СОМАТИЧЕСКИХ КЛЕТОК | 2008 |
|
RU2492232C2 |
| СПОСОБ ОЧИСТКИ КАРДИОМИОЦИТОВ | 2021 |
|
RU2813532C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕНАЛЬНЫХ КЛЕТОК-ПРЕДШЕСТВЕННИКОВ И СОДЕРЖАЩЕЕ ИХ ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2014 |
|
RU2696315C2 |
| СПОСОБ ДИФФЕРЕНЦИАЦИИ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК В КЛЕТКИ СОСУДИСТОГО РУСЛА | 2012 |
|
RU2618871C2 |
| СПОСОБ ИНДУКЦИИ ДИФФЕРЕНЦИАЦИИ СТВОЛОВЫХ КЛЕТОК В МИОКАРДИАЛЬНЫЕ | 2004 |
|
RU2392315C2 |
| ДИФФЕРЕНЦИРОВКА ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2008 |
|
RU2465323C2 |
| СПОСОБ ВЫРАЩИВАНИЯ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК | 2005 |
|
RU2375448C2 |
| ИНСУЛИНПРОДУЦИРУЮЩИЕ КЛЕТКИ | 2019 |
|
RU2829220C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛЕТОК, ПРОДУЦИРУЮЩИХ ПАНКРЕАТИЧЕСКИЕ ГОРМОНЫ | 2011 |
|
RU2576000C2 |
| ДИФФЕРЕНЦИРОВКА ЧЕЛОВЕЧЕСКИХ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК | 2008 |
|
RU2473685C2 |
















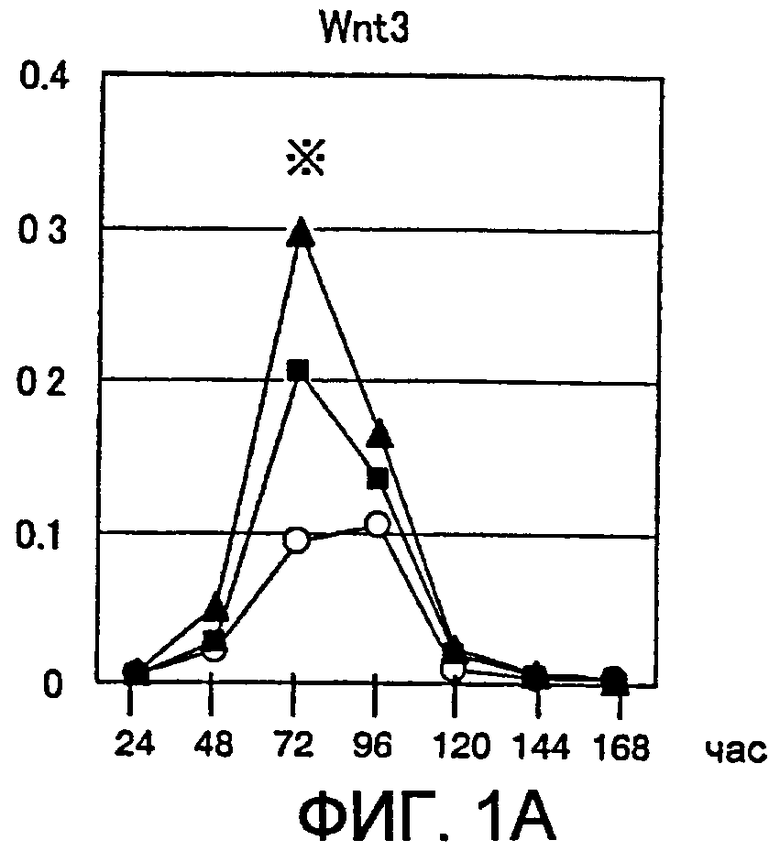
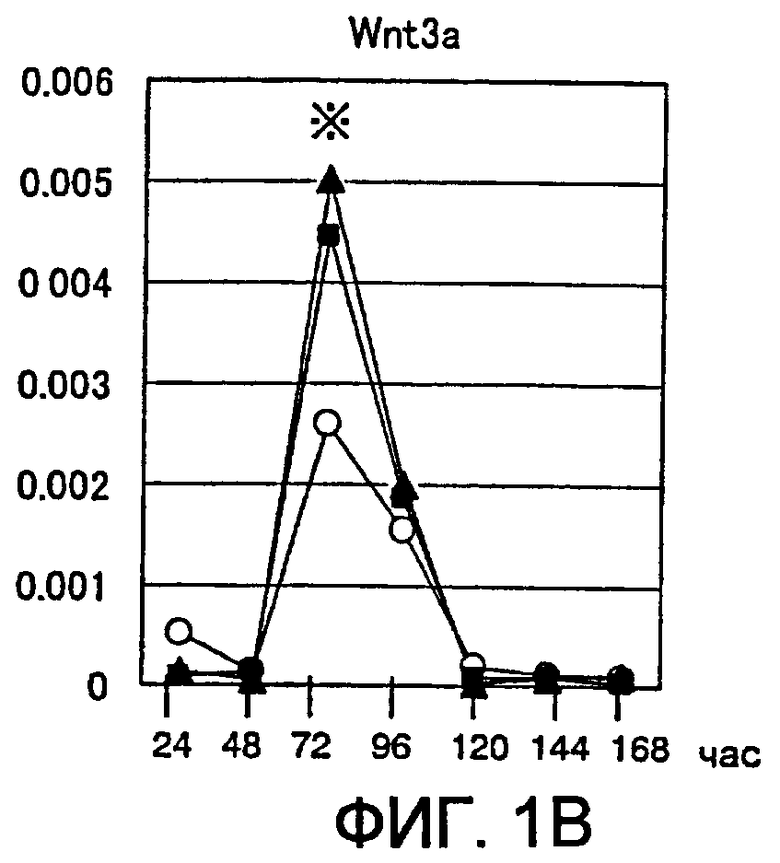
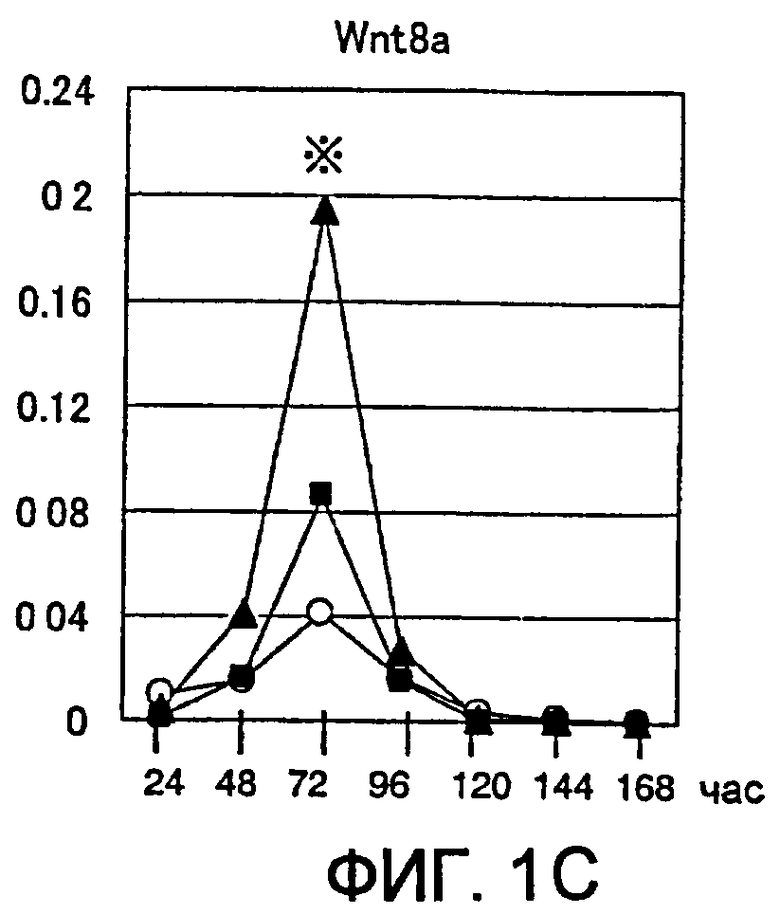
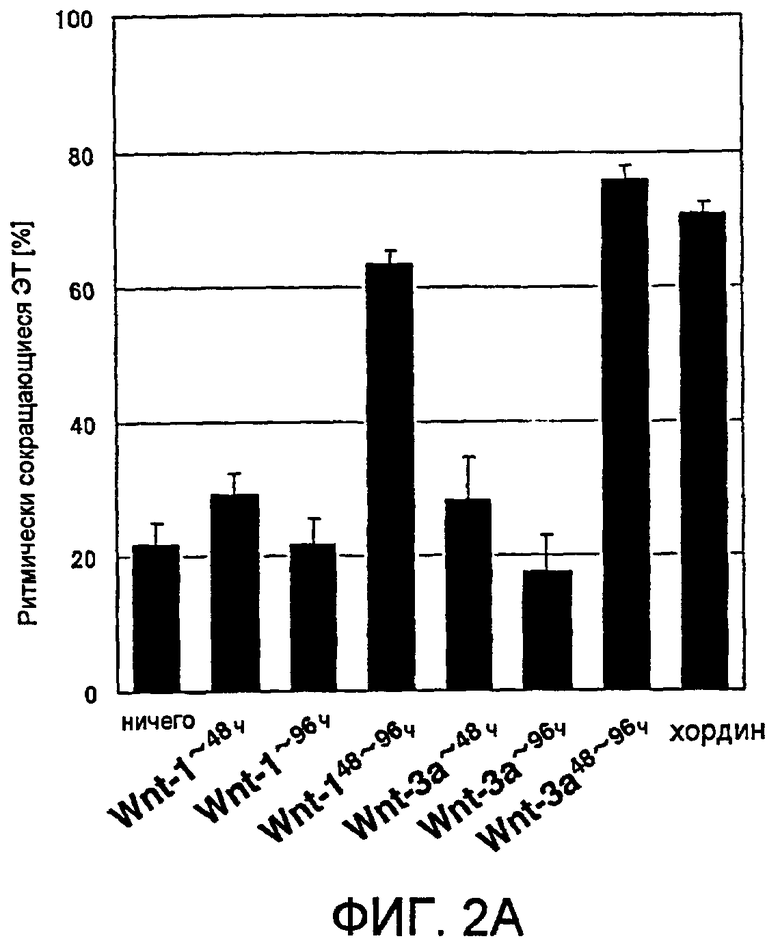
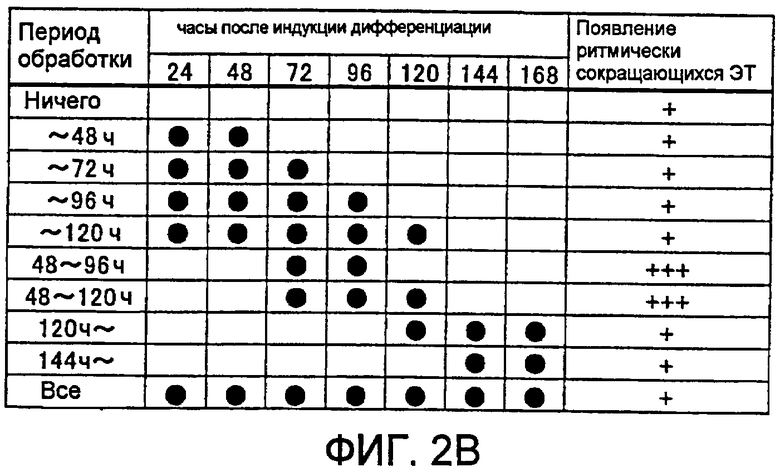
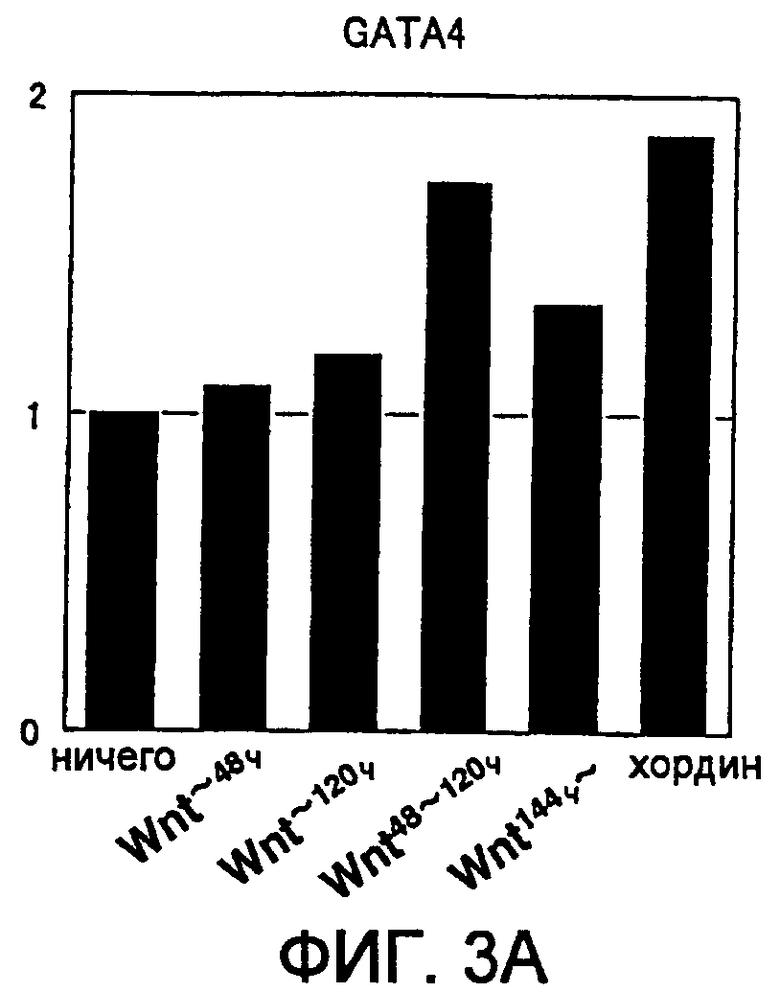
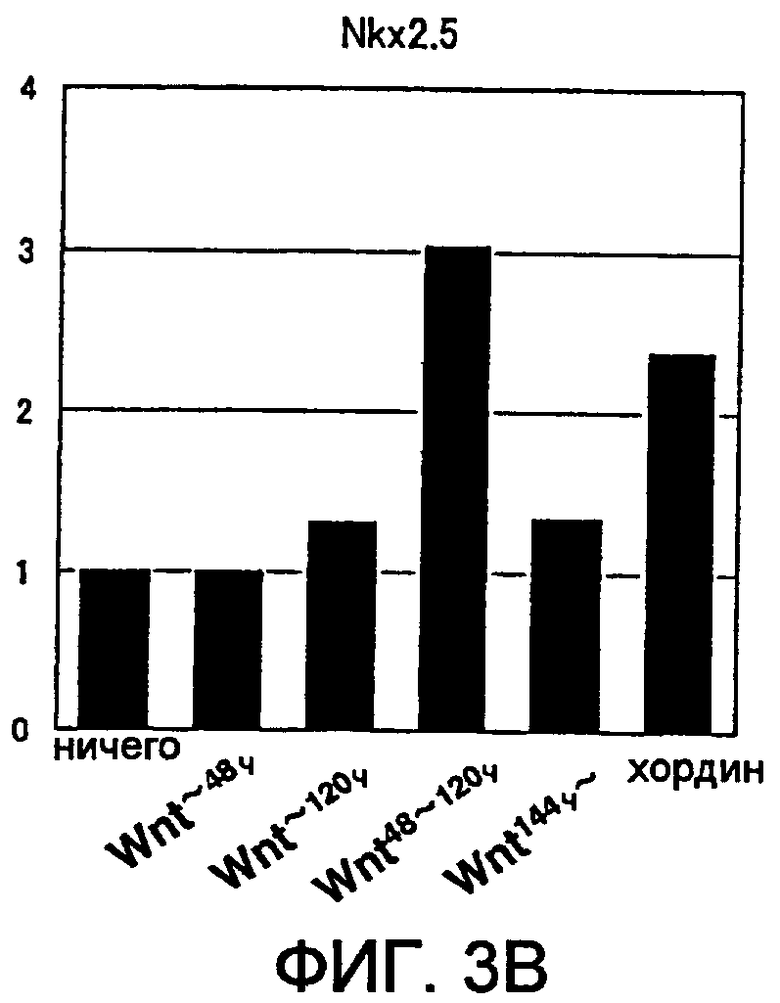
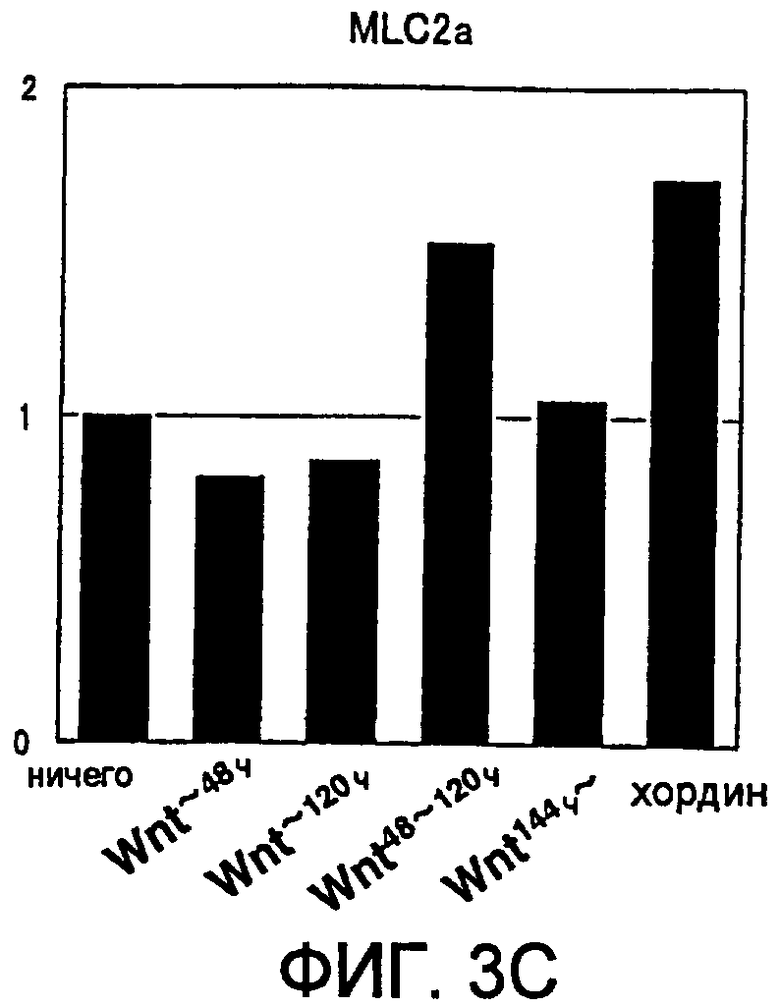
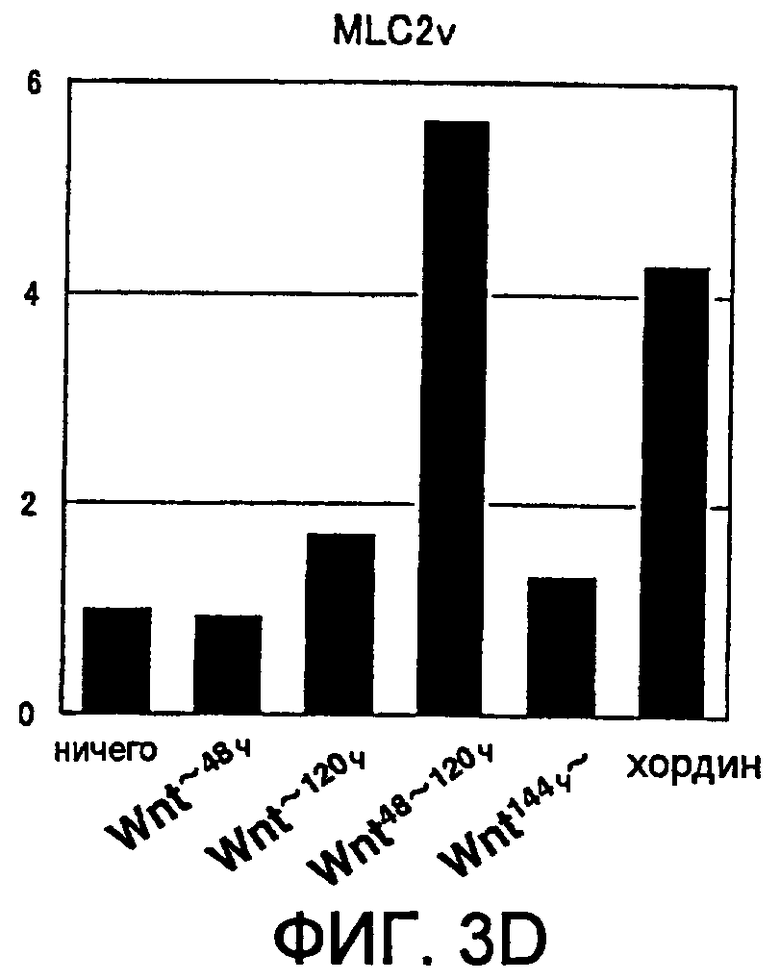
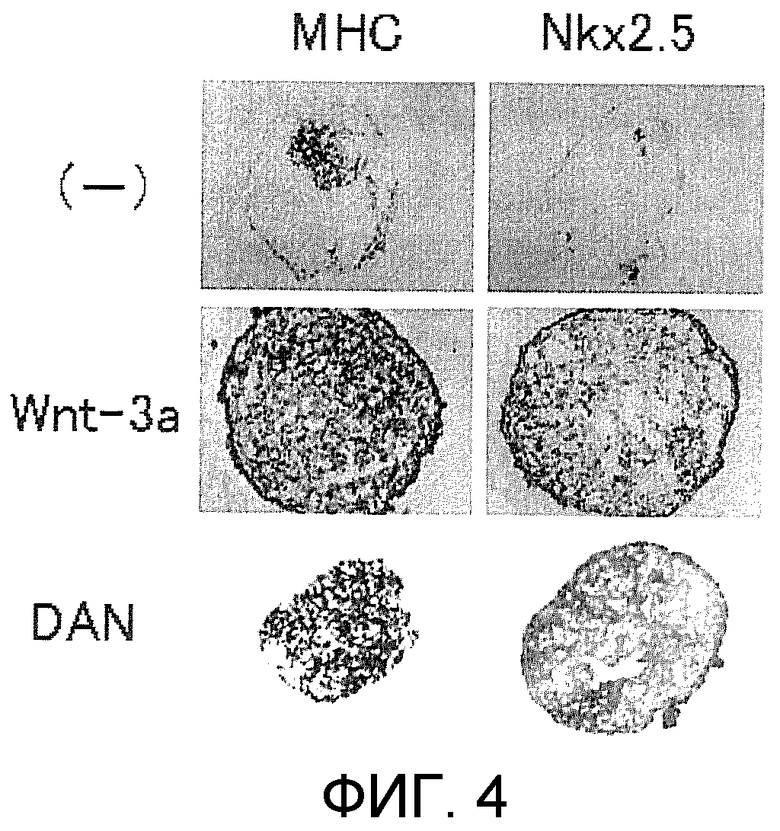
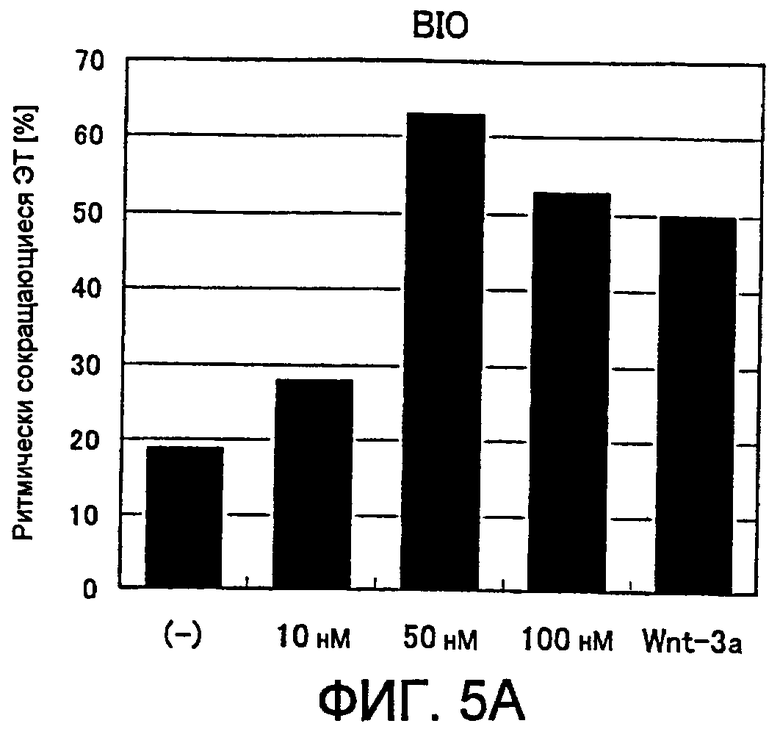
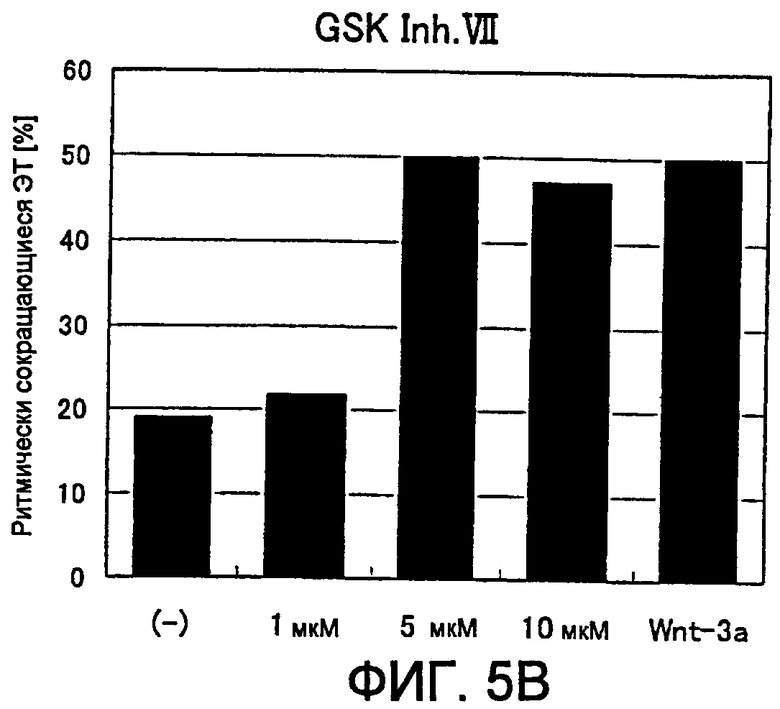
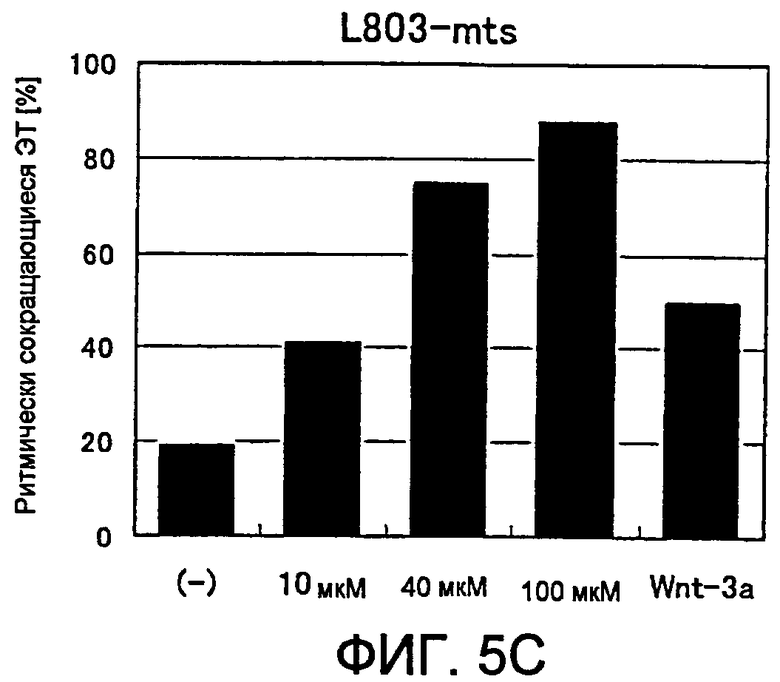
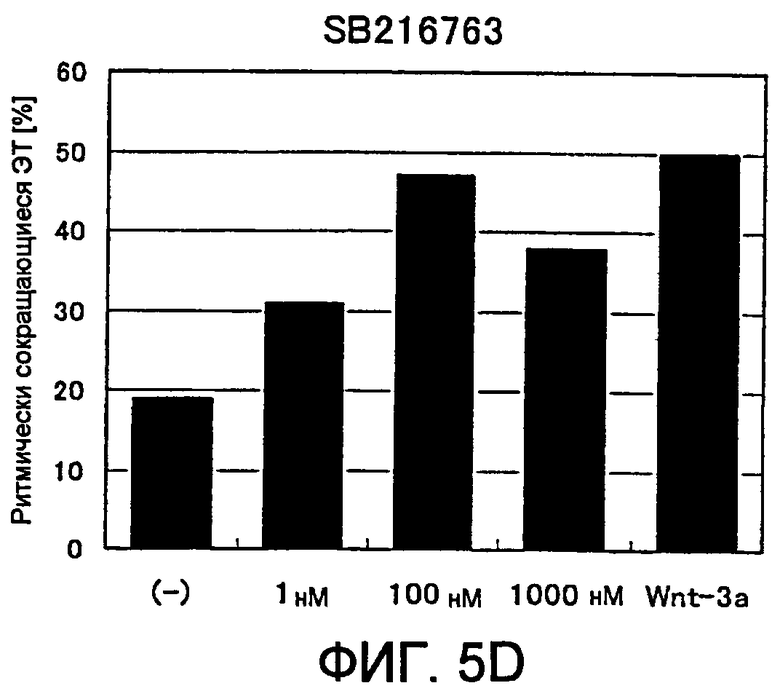
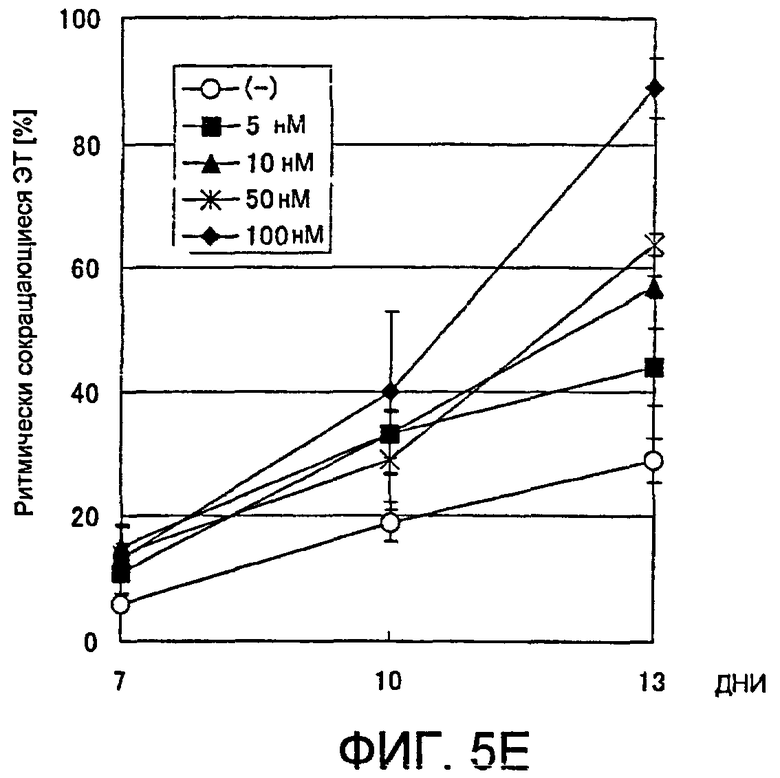
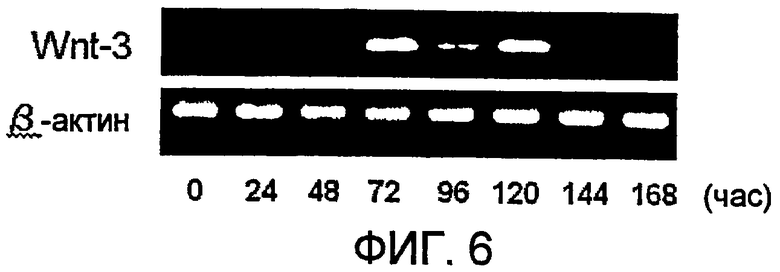
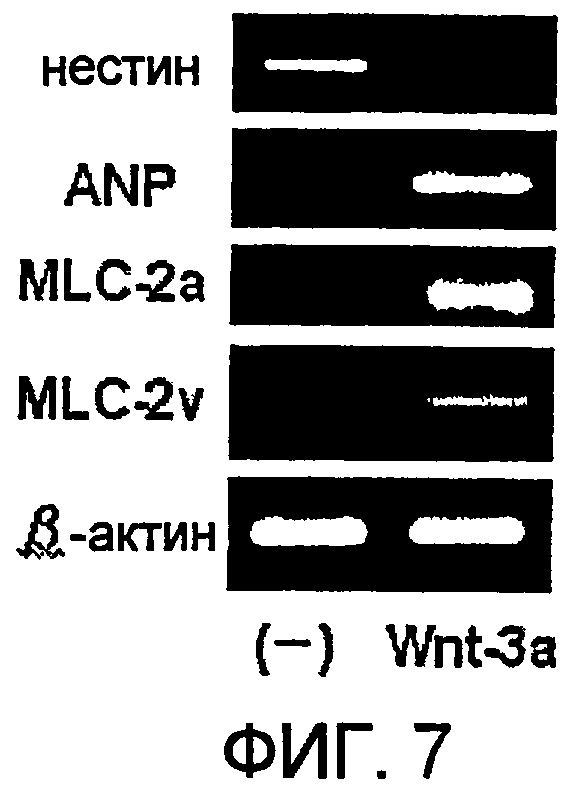
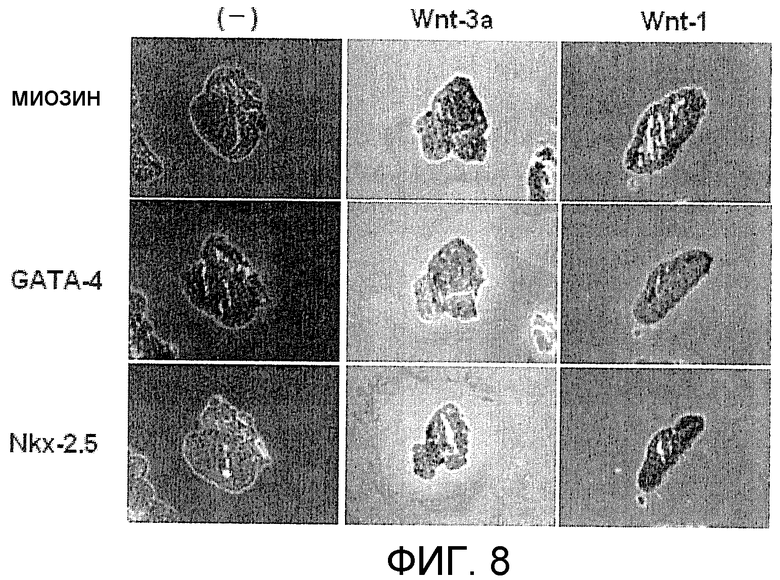
Изобретение относится к области биотехнологии, а именно к способу индукции дифференциации кардиомиоцитов из стволовых клеток. Способ предусматривает культивирование плюрипотентных стволовых клеток в культуральной среде, не содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, на протяжении периода времени между началом индукции дифференциации и 24 часами перед периодом повышенной экспрессии генов канонических Wnt. Затем проводят культивирование плюрипотентных стволовых клеток в культуральной среде, содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, на протяжении периода времени от 24 до 96 часов, начиная с 24 до 0 часов перед периодом повышенной экспрессии генов канонических Wnt. Способ позволяет эффективно и избирательно индуцировать дифференциацию кардиомиоцитов. Изобретение может быть использовано в медицине. 13 з.п. ф-лы, 18 ил.
1. Способ индукции дифференциации кардиомиоцитов из плюрипотентных стволовых клеток, который включает:
i) культивирование плюрипотентных стволовых клеток в культуральной среде, не содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, на протяжении периода, начинающегося от начала индукции дифференциации и заканчивающегося за 24 ч до периода повышенной экспрессии генов канонических Wnt, а затем
ii) культивирование плюрипотентных стволовых клеток в культуральной среде, содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, на протяжении 24-96 ч, начиная с 24-0 ч до периода повышенной экспрессии генов канонических Wnt, где вещество, которое стимулирует активацию канонического сигнального пути Wnt, представляет собой вещество, выбранное из группы, состоящей из канонического белка Wnt, ингибитора GSK3β и агониста Wnt,
при этом анализ экспрессии генов Wnt-3, Wnt-3a, Wnt-8a проводили в процессе дифференциации плюрипотентных стволовых клеток с применением системы количественной оценки полимеразной цепной реакции.
2. Способ по п.1, где плюрипотентные стволовые клетки культивируют в культуральной среде, содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, за 24 ч до периода повышенной экспрессии генов канонических Wnt.
3. Способ по п.1 или 2, где плюрипотентные стволовые клетки культивируют в культуральной среде, содержащей вещество, которое стимулирует активацию канонического сигнального пути Wnt, на протяжении периода времени 48-72 ч.
4. Способ по п.1, где вещество, которое стимулирует активацию канонического сигнального пути Wnt, представляет собой канонический белок Wnt.
5. Способ по п.4, где канонический белок Wnt представляет собой, по меньшей мере, один белок Wnt, выбранный из группы, состоящей из Wnt-1, Wnt-3а и Wnt-5a.
6. Способ по п.4 или 5, где концентрация канонического белка Wnt в культуральной среде составляет от 0,1 нг/мл до 500 нг/мл.
7. Способ по п.1, где вещество, которое стимулирует активацию канонического сигнального пути Wnt, представляет собой ингибитор GSK3β.
8. Способ по п.7, где ингибитор GSK3β представляет собой по меньшей мере один ингибитор, выбранный из группы, состоящей из ингибитора VII GSK3β, L803-mts, SB216763 и ингибитора IX GSK3β (ВIO).
9. Способ по п.7 или 8, где концентрация ингибитора GSK3β в культуральной среде составляет от 2 мкмоль/л до 100 мкмоль/л для ингибитора VII GSK3β от 5 мкмоль/л до 500 мкмоль/л для L803-mts, от 10 нмоль/л до 1 мкмоль/л для SB216763 или от 10 нмоль/л до 1 мкмоль/л для ингибитора IX GSK3β (ВIO).
10. Способ по п.1, где вещество, которое стимулирует активацию канонического сигнального пути Wnt, представляет собой агонист Wnt.
11. Способ по п.10, где агонист Wnt представляет собой производное аминопиримидина.
12. Способ по п.10 или 11, где концентрация агониста Wnt в культуральной среде составляет от 1 нмоль/л до 1000 нмоль/л.
13. Способ по п.1, где плюрипотентные стволовые клетки представляют собой эмбриональные стволовые клетки, эмбриональные зародышевые клетки или зародышевые стволовые клетки, исключая эмбриональные стволовые клетки, полученные путем разрушения эмбрионов человека.
14. Способ по п.13, где плюрипотентные стволовые клетки представляют собой эмбриональные стволовые клетки.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ORLANDO, FL, US, vol.325, no.3, 17 December 2004 (2004-12-17), pages 968-975 | |||
| KOYANAGI M ET AL: "Non-canonical Wnt signaling enhances differentiation of human circulating | |||

