Область техники
Настоящее изобретение относится к селективному и эффективному способу получения кардиомиоцитов из клеток ES и других полипотентных клеток, а также к клеткам для использования в регенеративной медицине, получаемых данным способом.
Предпосылки создания изобретения
Как правило, до рождения кардиомиоциты активно делятся, формируя орган, способный к автономной пульсации, однако после рождения они утрачивают способность к делению, и, поскольку существует мало недифференцированных клеток-предшественников, кардиомиоциты, погибшие вследствие воздействия разных видов стресса, в том числе инфаркта миокарда, миокардита и т.п., не восполняются. В результате живые кардиомиоциты пытаются поддержать функцию миокарда посредством компенсаторной гипертрофии и т.п., однако если воздействие стресса продолжается и превышает допустимый предел, то оно приводит к дополнительному истощению, гибели кардиомиоцитов и к последующему ухудшению функции миокарда (то есть к сердечной недостаточности).
Сердечная недостаточность и другие типы сердечных заболеваний являются второй основной причиной смерти в Японии и имеют очень плохой прогноз: 5-летний коэффициент выживаемости пациентов с заболеваниями сердца составляет приблизительно только 50%. Следовательно, можно надеяться, что разработка высокоэффективных способов лечения сердечной недостаточности приведет к значительному снижению расходов на медицинское обслуживание и к улучшению экономических показателей медицины. Традиционно для лечения сердечной недостаточности используют препараты наперстянки, которые усиливают сократительную активность миокарда, ксантиновые препараты и другие сердечные стимуляторы, однако известно, что длительное введение этих лекарственных средств приводит к ухудшению состояния из-за чрезмерного расходования энергии в миокарде. В последнее время предпочтение отдается бета-блокаторам и ингибиторам ACE, которые уменьшают чрезмерную нагрузку на сердце вследствие стимуляции симпатической нервной системы и ренин-ангиотензинной системы, однако данные методы только позволяют устранять непосредственные симптомы, но не восстанавливают поврежденную ткань сердца. Наоборот, трансплантация сердца является кардинальным способом лечения тяжелой сердечной недостаточности, однако ее применение обычно затруднено из-за таких причин, как дефицит доноров сердца, этические проблемы, большие физические нагрузки для пациентов, высокая стоимость и т.п.
Следовательно, способы трансплантации с целью замены ослабленных или утраченных кардиомиоцитов могут быть крайне полезными для лечения сердечной недостаточности. Фактически эксперименты на животных показали, что, если незрелые кардиомиоциты, полученные от зародыша, трансплантировать в ткань сердца взрослой особи, трансплантированные клетки эффективно функционируют (см. непатентный документ 1). Однако для такого способа трудно получить достаточное количество кардиомиоцитов, и, кроме того, применение такого способа в клинической медицине также затруднено из-за этических проблем.
Поэтому в последнее время повышенное внимание уделяется индукции дифференциации стволовых клеток в кардиомиоциты и применению данных клеток для трансплантации. В настоящее время пока невозможно однозначно идентифицировать популяцию клеток-предшественников или стволовых клеток, способных продуцировать кардиомиоциты во взрослой ткани сердца, поэтому предполагается, что для указанного способа можно использовать полипотентные стволовые клетки, которые являются менее дифференцированными и могут дифференцироваться в разные клетки.
Полипотентными стволовыми клетками называют клетки, способные к неограниченной или длительной клеточной пролиферации с сохранением недифференцированного состояния в клеточной культуре in vitro, которые сохраняют нормальный кариотип и способны дифференцироваться в клетки всех трех зародышевых слоев (эктодерм, мезодерм и эндодерм). В настоящее время известно три типа полипотентных стволовых клеток: эмбриональные стволовые клетки (клетки ES), получаемые из эмбрионов на ранней стадии развития, эмбриональные зародышевые клетки (клетки EG), получаемые из примордиальных зародышевых клеток на эмбриональной стадии, и полипотентные клетки-предшественники взрослых особей (МАРС), получаемые из костного мозга.
Давно известно, что in vitro можно индуцировать дифференциацию, в особенности клеток ES с получением кардиомиоцитов. Большую часть ранних исследований проводили с использованием мышиных клеток ES. Если клетки ES культивируют в суспензии в виде одиночных клеток (отдельных клеток, которые при диспергировании не прилипают друг к другу вследствие обработки ферментом или т.п.) в отсутствие фактора, ингибирующего дифференциацию, такого как фактор, ингибирующий лейкоз (LIF) и т.п., клетки ES прилипают друг к другу и агрегируют, образуя структуру, называемую эмбриоидные тельца (EB), подобную ранним эмбриональным структурам. Также известно, что при культивировании ES в суспензии или прилипшими на поверхности приспособлений для культивирования кардиомиоциты образуют структуры, способные к спонтанной пульсации.
Кардиомиоциты, полученные из клеток ES по описанному выше способу, имеют свойства, очень похожие на свойства незрелых кардиомиоцитов фетальных сердец (см. непатентные документы 2 и 3). Более того, на экспериментах с животными было показано, что трансплантация кардиомиоцитов, полученных из клеток ES, в ткани сердца взрослой особи дает эффективные результаты, подобные полученным при трансплантации фетального миокарда (см. патентный документ 1; непатентный документ 4).
В 1995 г. Thomson et al. впервые выделили клетки ES из приматов (см. патентный документ 2; непатентный документ 5), и регенерационная терапия с использованием кардиомиоцитов, полученных из полипотентных стволовых клеток, стала реальной. Затем они также успешно выделили клетки ES из ранних человеческих эмбрионов и получили соответствующие клеточные линии (см. непатентный документ 6). Кроме того, Gearhart et al. получили линии клеток ES из примордиальных зародышевых клеток человека (см. непатентный документ 7; патентный документ 3).
Kehat et al. (см. непатентный документ 8) и Chunhui et al. (см. патентный документ 4; непатентный документ 9) описали, что человеческие клетки ES, как и мышиные клетки ES, могут дифференцироваться в кардиомиоциты in vitro. В соответствии с указанными публикациями кардиомиоциты, полученные в результате индукции дифференциации человеческих клеток ES, не только обладают способностью к спонтанной пульсации, но также экспрессируют и продуцируют специфичные для миокарда белки, такие как тяжелые и легкие цепи миозина, альфа-актинин, тропонин I и атриальный натрийуретический пептид (ANP), а также специфичные для миокарда факторы транскрипции, такие как GATA-4, Nkx2.5, MEF-2c и т.п., а результаты микроанатомических наблюдений и электрофизиологического анализа показывают, что полученные клетки сохраняют свойства незрелых кардиомиоцитов на стадии зародыша и могут использоваться для регенерационной терапии.
Однако при использовании кардиомиоцитов, полученных из полипотентных стволовых клеток, в терапии с применением трансплантации клеток и для других целей остается одна серьезная проблема. Если EBS получают из клеток ES и EG традиционными методами, то образуются не только кардиомиоциты, но и другие типы дифференцированных клеток, такие как кровяные клетки, клетки сосудов, нервные клетки, клетки кишечника, клетки костей, хрящей и т.п. Более того, доля кардиомиоцитов в данной популяции дифференцированных клеток не так высока и составляет приблизительно только 5-20% от общего количества клеток.
Способы выделения кардиомиоцитов из смеси клеток разных видов включают в себя добавление к генам клеток ES искусственной модификации, придающей устойчивость к лекарственному средству или обеспечивающей эктопическую экспрессию, и сбор клеток, обладающих свойствами кардиомиоцитов или их предшественников. Например, путем введения генной кассеты, обеспечивающей экспрессию гена устойчивости к неомицину (G418) под контролем промотора тяжелой цепи α-миозина, Field и соавторы получили систему, в которой во время дифференциации в кардиомиоциты и экспрессии гена тяжелой цепи α-миозина выживать в среде могут только те клетки ES, в которые был добавлен G418 (см. патентный документ 1; непатентный документ 4). Было подтверждено, что 99% или больше G418-устойчивых клеток, отобранных с помощью данного способа, являются кардиомиоцитами. Однако, хотя чистота кардиомиоцитов, полученных с помощью указанного способа, является очень высокой, конечное число полученных кардиомиоцитов составляет только несколько процентов от общего числа клеток, что затрудняет получение достаточного для трансплантации количества кардиомиоцитов.
Недавно Chunhui et al. описали, что, если человеческие клетки ES обработать 5-азацитидином, процент тропонин I-положительных клеток (подходящих кардиомиоцитов) в EB увеличивается от 15% до 44% (см. непатентный документ 9), но даже в данном способе процент кардиомиоцитов в EB не превышает 50%. Более того, 5-азацитидин является деметилирующим агентом, который изменяет экспрессию генов путем удаления метильных групп, связанных с ДНК, и, поскольку он действует непосредственно на хромосомы, он не подходит для получения клеток, предназначенных для трансплантации.
Другие способы более эффективного получения кардиомиоцитов из клеток ES включают в себя, в случае мышиных клеток ES, добавление ретиноевой кислоты (см. непатентный документ 10), аскорбиновой кислоты (см. непатентный документ 11), TGF-бета, BMP-2 (см. непатентный документ 12), PDGF (см. непатентный документ 13) и динорфин В (см. непатентный документ 14), а также обработку средствами, увеличивающими количество реакционноспособных частиц кислорода (ROS) (см. непатентный документ 15) и Ca2+ (см. непатентный документ 16) в клетках, причем известно, что все они оказывают положительное воздействие на индукцию дифференциации кардиомиоцитов. Однако кардимиоцит-специфичная, селективная дифференциация не достигается при применении указанных способов.
Секреторные белки Noggin и Chordin вначале были идентифицированы как факторы индукции нервов в эмбрионах Xenopus (см. непатентные документы 17 и 18; патентные документы 5-8). В другом исследовании было показано, что Noggin и Chordin связываются с семейством молекул BMP (костный морфогенетический белок (bone morphogenic protein)), которые ухудшают передачу сигнала и вызывают индукцию и дифференциацию нейронов (см. непатентные документы 19-21). Фактически эксперименты, проводимые с использованием мышиных клеток ES, показали, что дифференциация нервных клеток индуцируется в клетках, постоянно экспрессирующих ген Noggin или Chordin (см. непатентный документ 22).
Если человеческие клетки ES культивируют в среде, в которую добавлен Noggin, эндогенная продукция BMP-2 снижается и состояние клеток ES в экстраэмбриональных эндодермальных клетках ухудшается так, что они остаются в недифференцированном состоянии. Кроме того, если затем клетки ES, обработанные Noggin, культивируют в условиях дифференциации нейронов, то индуцируется развитие нервных клеток (см. патентный документ 9).
В более ранних исследованиях, проводимых с использованием эмбриональных раковых клеток кур (см. непатентный документ 23), Xenopus (см. непатентный документ 24) и мышей (см. непатентный документ 25), также было показано, что семейство молекул BMP стимулирует развитие и/или дифференциацию кардиомиоцитов, а после блокады этого эффекта путем обработки Noggin развитие и/или дифференциация кардиомиоцитов подавляется.
До настоящего времени не предпринимались попытки стимулировать развитие и/или дифференциацию кардиомиоцитов с помощью Noggin, Chordin или других факторов семейства BMP, ингибирующих передачу сигнала.
Патентный документ 1: патент США № 6015671;
Патентный документ 2: патент США № 5843780;
Патентный документ 3: патент США № 6015671;
Патентный документ 4: описание международного патента 03/06950, памфлет;
Патентный документ 5: описание международного патента 94/05791, памфлет;
Патентный документ 6: патент США № 5679783;
Патентный документ 7: патент США № 5846770;
Патентный документ 8: патент США № 5986056;
Патентный документ 9: описание международного патента 01/98463, памфлет;
Непатентный документ 1: Soonpaa et al., Science 264:98, 1994;
Непатентный документ 2: Maltsev et al., Mech. Dev. 44:41, 1993;
Непатентный документ 3: Maltsev et al., Circ. Res. 75:233, 1994;
Непатентный документ 4: Klug et al., J.Clin. Invest. 98:216, 1996;
Непатентный документ 5: Thomson et al., Proc. Natl. Acad. Sci. USA 92:7844, 1995;
Непатентный документ 6: Thomson et al., Science 282:114, 1998;
Непатентный документ 7: Shamblott et al. Proc. Natl. Acad. Sci. USA 95:13726, 1998;
Непатентный документ 8: Kehat et al., J. Clin. Invest. 108:407, 2001;
Непатентный документ 9: Chunhui et al., Circ. Res. 91:508, 2002;
Непатентный документ 10: Wobus et al., J. Mol. Cell Cardiol. 29:1525, 1997;
Непатентный документ 11: Takahashi et al., Circulatuon 107:1912, 2003;
Непатентный документ 12: Behfar et al., FASEB J. 16:1558, 2002;
Непатентный документ 13: Sachinidis et al., Cardiovasc. Res. 58:278, 2003;
Непатентный документ 14: Ventura et al., Circ. Res. 92:623, 2003;
Непатентный документ 15: Sauer et al., FEBS Lett. 476:218, 2000;
Непатентный документ 16: Li et al., J. Cell Biol. 158:103, 2002;
Непатентный документ 17: Smith & Harland, Cell 70:829, 1992;
Непатентный документ 18: Sasai et al., Cell 79:779, 1994;
Непатентный документ 19: Re'em-Kalma et al., Proc. Natl. Acad. Sci. USA 92:12141, 1995;
Непатентный документ 20: Zimmerman et al., Cell 86:599, 1996;
Непатентный документ 21: Piccolo et al., Cell 86:589, 1996;
Непатентный документ 22: Gratsch & O'Shea, Dev. Biol. 245:83, 2002;
Непатентный документ 23: Schultheiss et al., Genes Dev., 11:451, 1997;
Непатентный документ 24: Sparrow et al., Mech. Dev. 71:151, 1998;
Непатентный документ 25: Monzen et al., Mol. Cell. Biol. 19:7096, 1999.
Описание изобретения
Проблемы, решаемые изобретением
Целью настоящего изобретения является предоставление эффективного и селективного способа индукции дифференциации стволовых клеток в кардиомиоциты, а также кардиомиоцитов, полученных с помощью данного способа, и способа применения данных клеток в трансплантации, инъекции и других способах лечения, направленных на заболевания сердца.
Способы решения проблем
Настоящее изобретение, в основном, включает в себя:
(1) Способ индукции кардиомиоцитарной дифференциации стволовых клеток, где стволовые клетки культивируют с целью индукции дифференциации в присутствии вещества, ингибирующего сигнальный путь, опосредованный BMP.
(2) Способ по приведенному выше п.1, где культивирование стволовых клеток с целью индукции дифференциации включает в себя стадию получения эмбриоидных телец путем культивирования в суспензии с агрегацией.
(3) Способ по приведенному выше п.1, где культивирование стволовых клеток с целью индукции дифференциации включает в себя стадию совместного культивирования с питающими клетками.
(4) Способ по приведенному выше п.1, где культивирование стволовых клеток с целью индукции дифференциации включает в себя стадию культивирования в чашках на культуральном контейнере.
(5) Способ по любому из приведенных выше п.п.(1)-(4), включающий в себя стадию обработки стволовых клеток веществом, которое ингибирует сигнальный путь BMP, в течение первых нескольких дней стадии индукции дифференциации.
(6) Способ по любому из приведенных выше п.п.(1)-(4), включающий в себя стадию обработки стволовых клеток веществом, которое ингибирует сигнальный путь BMP, предшествующую дифференциации.
(7) Способ по любому из приведенных выше п.п.(1)-(4), включающий в себя стадию обработки стволовых клеток веществом, которое ингибирует сигнальный путь BMP, до стадии дифференциации и стадию обработки стволовых клеток веществом, которое ингибирует сигнальный путь BMP, в течение первых нескольких дней стадии индукции дифференциации.
(8) Способ по любому из приведенных выше п.п.(1)-(7), где вещество, которое ингибирует сигнальный путь BMP, представляет собой антагонист BMP.
(9) Способ по приведенному выше п.8, где антагонист BMP представляет собой одно или несколько средств, выбранных из группы, включающей в себя Noggin, Chordin, фетуин, фоллистатин, склеростин, DAN, Cerberus, гремлин, Dante и родственные им белки.
(10) Способ по любому из приведенных выше п.п.(1)-(9), где стволовые клетки представляют собой клетки млекопитающих, обладающие способностью к дифференциации в кардиомиоциты in vitro.
(11) Способ по приведенному выше п.10, где клетки млекопитающих, обладающие способностью к дифференциации в кардиомиоциты, представляют собой полипотентные стволовые клетки или клетки, полученные из них.
(12) Способ по приведенному выше п.11, где полипотентные стволовые клетки представляют собой эмбриональные стволовые клетки, клетки, обладающие свойствами эмбриональных стволовых клеток, эмбриональные зародышевые клетки или полипотентные клетки-предшественники взрослых особей.
(13) Способ по приведенному выше п.12, где полипотентные стволовые клетки представляют собой эмбриональные стволовые клетки.
(14) Способ по любому из приведенных выше п.п.(1)-(13), где стволовые клетки представляют собой человеческие клетки.
(15) Способ получения кардиомиоцитов, включающий в себя способ по любому из приведенных выше п.п.(1)-(14).
(16) Кардиомиоциты, полученные по способу любого из приведенных выше п.п.(1)-(14).
(17) Способ лечения заболеваний сердца, возникающих в результате истощения, функциональной недостаточности или гибели кардиомиоцитов, включающий в себя введение (трансплантацию) кардиомиоцитов по приведенному выше п.(16) в участок истощения, функциональной недостаточности или гибели кардиомиоцитов или вблизи данного участка.
(18) Способ скрининга веществ, пригодных для лечения заболеваний сердца, возникающих в результате истощения, функциональной недостаточности или гибели кардиомиоцитов, который включает в себя приведение в контакт тестируемого вещества и кардиомиоцитов по приведенному выше п.(16) и измерение количественных или качественных изменений клеточных функций или эффективности дифференциации клеток в кардиомиоциты.
(19) Композиция лекарственного средства или подложка для лечения заболеваний сердца, возникающих в результате истощения, функциональной недостаточности или гибели кардиомиоцитов, содержащая в качестве активного компонента кардиомиоциты по приведенному выше п.(16).
В качестве источника стволовых клеток для получения кардиомиоцитов авторы данного изобретения использовали полипотентные стволовые клетки, чаще всего клетки ES, и в результате экстенсивного изучения условий индукции дифференциации в кардиомиоциты или клетки-предшественники они обнаружили, что если на определенной, ограниченной стадии культивирования в среду добавить вещество, ингибирующее сигнальный путь BMP (костного морфогенетического белка), то популяции клеток, обладающих способностью к пульсации, которые были идентифицированы как кардиомиоциты, развиваются гораздо более селективно и эффективно, чем полученные с помощью традиционных методов, что и составило основу изобретения, также было обнаружено, что добавление избыточного количества BMP приводит к сильному снижению индукции миокардиальной дифференциации под действием вещества, ингибирующего сигнальный путь BMP, это показывает, что ингибирование сигнального пути BMP вызывает дифференциацию полипотентных стволовых клеток в кардиомиоциты.
В настоящем изобретении вещество, ингибирующее сигнальный путь BMP, представляет собой вещество, которое оказывает блокирующее действие на сигнальный путь, опосредованный BMP, или является конкурентом BMP, примеры данного вещества включают в себя антагонисты BMP, семейство специфичных нейтрализующих антител против BMP, солюбилизированные молекулы рецептора BMP (относящиеся к типу рецепторов, не связанных с клеточной мембраной) и т.п. Другие примеры включают в себя генные векторы экспрессии, специфичные антисмысловые олигонуклеотиды, рибозимы, антисмысловую РНК для связывания РНК, низкомолекулярные соединения и т.п., которые либо подавляют, либо останавливают экспрессию молекул семейства BMP или функциональных продуктов генов их рецепторов, или, иначе, подавляют или останавливают экспрессию генов, которые кодируют компоненты, препятствующие активации сигнального пути BMP.
Способные к пульсации клетки, полученные из полипотентных стволовых клеток по способу настоящего изобретения, представляют собой клетки, обладающие характеристиками "кардиомиоцитов", например, показано, что данные клетки экспрессируют гены GATA-4, TEF-1, Tbx-5, MEF2, MLC-2v и другие, специфичные для миокарда факторы транскрипции, а также белки, такие как специфичные для миокарда маркеры саркомерный миозин, тропонин I, α-актинин, ANP и т.п.
Кроме традиционно используемого способа культивирования в суспензии с получением агрегатов для индукции дифференциации полипотентных стволовых клеток настоящего изобретения можно использовать способ культивирования в висячей капле или способ совместного культивирования с питающими клетками.
При определении моментов и периодов времени, подходящих для добавления вещества, ингибирующего сигнальный путь BMP, было обнаружено, что если полипотентные стволовые клетки культивируют в виде отдельных клеток или клеточных агрегатов, состоящих из небольшого числа клеток, а дифференциацию индуцируют с помощью такого способа, как агрегационное культивирование в суспензии или совместное культивирование с питающими клетками, то более эффективные результаты получают при обработке полипотентных стволовых клеток BMP либо до дифференциации, либо в течение первых нескольких дней после начала культивирования с индукцией дифференциации, либо при объединении двух указанных методов. Также было показано, что если антагонист BMP присутствует в течение всего времени культивирования в суспензии, совместного культивирования с питающими клетками или т.п., то эффективность дифференциации полипотентных стволовых клеток в кардиомиоциты сильно снижается.
То есть настоящее изобретение относится к способу индукции дифференциации полипотентных стволовых клеток в кардиомиоциты, где стволовые клетки культивируют с использованием временной обработки веществом, ингибирующим сигнальный путь BMP.
В настоящем изобретении стволовые клетки, обладающие способностью к миокардиальной дифференциации, представляют собой клетки, которые могут дифференцироваться в кардиомиоциты в культуре in vitro, их примеры включают в себя полипотентные стволовые клетки, мезинхимальные стволовые клетки, клетки CMG, клетки Spoc и т.п. Полипотентные стволовые клетки представляют собой клетки, способные к неограниченной или длительной клеточной пролиферации с сохранением недифференцированного состояния в культуре in vitro, которые имеют нормальный кариотип и способны дифференцироваться в клетки всех трех зародышевых слоев (эктодерм, мезодерм и эндодерм) в подходящих условиях, примеры данных клеток включают в себя клетки ES, ES-подобные клетки, клетки EG, MAPC и т.п.
Другое воплощение настоящего изобретения относится к клеткам, которые обладают морфологическими, физиологическими и/или иммунологическими характеристиками кардиомиоцитов и которые получают из полипотентных стволовых клеток. С точки зрения физиологических и/или иммунологических свойств клетки, полученные по способу настоящего изобретения, могут экспрессировать один или несколько маркеров, специфичных для кардиомиоцитов, по которым распознаются кардиомиоциты, но это не является ограничением.
Настоящее изобретение также относится к способу скрининга с использованием клеток, полученных по способу настоящего изобретения, с целью идентификации возможных химиотерапевтических лекарственных средств или новых факторов, которые стимулируют развитие, дифференциацию, регенерацию, выживание и другие процессы, связанные с кардиомиоцитами.
Настоящее изобретение также относится к набору для индукции дифференциации полипотентных стволовых клеток в клетки, обладающие морфологическими, физиологическими и/или иммунологическими свойствами миокардиальных клеток-предшественников или кардиомиоцитов. Данный набор используют для осуществления способа настоящего изобретения.
Другое воплощение настоящего изобретения относится к способу лечения заболеваний сердца с использованием клеток, полученных по способу настоящего изобретения, и к лекарственному средству для лечения заболеваний сердца, которое в качестве активного компонента содержит клетки, полученные по способу настоящего изобретения.
Указанные и другие преимущества и характеристики настоящего изобретения соответствующим образом отражены в нижеследующем подробном описании предпочтительных воплощений.
При любом осуществлении настоящего изобретения можно использовать стандартные ссылки, касающиеся традиционных методов культивирования клеток и исследований в области развития и цитологии полипотентных стволовых клеток. Данные ссылки включают в себя (Wasserman et al. Eds., Academic Press, 1993); Embryonic Stem Cell Differentiation in vitro (M.V.Wiles, Meth. Enzymol. 225:900, 1993); Manipulating the Mouse Embryo: A laboratory manual (Hogan et al. Eds., Cold Spring Harbor Laboratory Press, 1994); and Embryonic Stem Cells (Turksen Ed., Humana Press, 2002). Реагенты и наборы для культивирования клеток и исследований в области развития и цитологии, упомянутых в данном описании, можно получить из коммерческих источников, в том числе Invitrogen/GIBCO, Sigma и т.п.
Эффекты данного изобретения
Миокардиальные клетки-предшественники и кардиомиоциты можно эффективно и селективно получать из стволовых клеток с помощью способа настоящего изобретения. Миокардиальные клетки (предшественники), полученные по данному способу, можно использовать для скрининга и разработки эффективных лекарственных средств для лечения заболеваний сердца, и потенциально могут использоваться для трансплантационной терапии миокарда при тяжелых сердечных заболеваниях.
Краткое описание чертежей
На фиг.1А показано влияние добавления белка Noggin (500 нг/мл) на частоту появления способных к пульсации EB, образующихся из клеток ES (EB3) при культивировании в суспензии.
На фиг.1В показано влияние добавления белка Noggin (500 нг/мл) на частоту появления способных к пульсации EB, образующихся из клеток ES (EB3) при культивировании методом висячей капли.
На фиг.2 приведена частота появления способных к пульсации EB в присутствии разных доз Noggin. EB получают из клеток ES (клеток EB3 и клеток R1) и количество способных к пульсации EB определяют на 10-й день культивирования в суспензии.
На фиг.3 показана экспрессия специфичных для миокарда маркерных генов в клетках ES, обработанных Noggin. EB (полученные из клеток EB3) собирают после 0, 5, 10 и 15 дней культивирования в суспензии и анализируют экспрессию каждого гена. TEF-1: фактор увеличения транскрипции-1, MEF-2c: повышающий мышечный фактор-2с, α-MHC: тяжелая цепь α-миозина, MLC-2v: легкая цепь миозина-2v, GAPDH: глицеральдегид-3-фосфатдегидрогеназа.
На фиг.4 приведены результаты иммунохимического окрашивания выделенных кардиомиоцитов группы EB Noggin (+) (полученных из клеток EB3) на 10 день культивирования в суспензии. a: саркомерный миозин, b: тропонин I, с: α-актинин, d: ANP.
На фиг.5 приведены результаты иммунохимического окрашивания кардиомиоцитов групп EB Noggin (+) и Noggin (-) (полученных из клеток EB3) на 10 день культивирования в суспензии. a,b: саркомерный миозин, c,d: тропонин I, e,f: α-актинин, g,h: ANP.
На фиг.6 показано ингибирующее действие BMP на индукцию кардиомиоцитов под действием Noggin.
На фиг.7А показано получение дифференцированных кардиомиоцитов методом совместного культивирования с питающими клетками. Клетки ES (R1) высевают на питающие клетки ST2 и миокардиальную дифференциацию анализируют методом иммуноцитохимического окрашивания с использованием антител против саркомерного миозина (MF20) на 8 день после высевания (n>5). При сравнении с группой Noggin (-) *p<0,01.
На фиг.7B показана частота появления миокардиальных колоний из клеток ES, обработанных Noggin, при использовании совместного культивирования с питающими клетками. Клетки ES (R1) высевают на питающие клетки ST2 и миокардиальную дифференциацию оценивают по частоте появления MF20-положительных колоний на 8 день после высевания (n>5). При сравнении с группой Noggin (-) *p<0,01.
На фиг.7С показана частота появления способных к пульсации колоний из клеток ES, обработанных Noggin, при использовании совместного культивирования с питающими клетками. Клетки ES (R1) высевают на питающие клетки ST2 и миокардиальную дифференциацию оценивают по частоте появления способных к пульсации колоний на 12 день после высевания (n>5). При сравнении с группой Noggin (-) *p<0,01.
На фиг.8 показано влияние добавления белка Chordin (500 нг/мл) на частоту появления способных к пульсации EB по сравнению с добавлением белка Noggin (500 нг/мл) при культивировании методом висячей капли. Дни культивирования считают от начала культивирования в висячей капле (n=8). При сравнении с необработанной группой (-) *p<0,01.
На фиг.9 показано влияние добавления белка Chordin (150 нг/мл) на частоту появления способных к пульсации EB из клеток ES (R1) с использованием чашки для культивирования в суспензии. С: Chordin, N: Noggin (150 нг/мл), (-): необработанные. В выражениях "А"→"В" А и В означают белок, добавляемый до и после образования EB соответственно.
На фиг.10 показано влияние добавления белков антагонистов BMP (150 нг/мл) на частоту образования способных к пульсации EB из клеток ES (R1) при использовании чашки для культивирования в суспензии.
Наилучший способ осуществления изобретения
В данном описании термин "кардиомиоциты" включает в себя предшественников клеток сердца, способные в будущем функционировать как кардиомиоциты, а также кардиомиоциты зародышей и взрослых особей на всех стадиях дифференциации; данный термин относится к клеткам, которые можно идентифицировать с помощью одного или, предпочтительно, нескольких из нижеследующих методов с использованием одного или, предпочтительно, нескольких маркеров или индексов.
Экспрессию различных маркеров, специфичных для кардиомиоцитов, детектируют с помощью традиционных биохимических или иммунохимических методов. Особых ограничений по данным методам не существует, однако предпочтительно используют иммунохимические методы, такие как иммунохимическое окрашивание или иммуноэлектрофорез. В данных способах можно использовать маркер-специфичные поликлональные антитела или моноклональные антитела, которые взаимодействуют с предшественниками клеток сердца или с кардиомиоцитами. Антитела против отдельных специфических маркеров коммерчески доступны и просты в использовании. Маркеры, специфичные для предшественников клеток сердца, включают в себя, например, тяжелую/легкую цепи миозина, α-актинин, тропонин I, ANP, GATA-4, Nkx2.5, MEF-2c и т.п.
Альтернативно, хотя используемые способы конкретно не ограничиваются, экспрессию генов маркеров, специфичных для предшественников клеток сердца или кардиомиоцитов, также можно подтвердить с помощью полимеразной цепной реакции с обратной транскриптазой (ОТ-ПЦР) или гибридизационного анализа, методов молекулярной биологии, которые в прошлом обычно использовались для амплификации, детекции и анализа мРНК, кодирующих маркерные белки. Последовательности нуклеиновых кислот, кодирующие маркеры, специфичные для предшественников клеток сердца или кардиомиоцитов (такие как тяжелая/легкая цепи миозина, α-актинин, тропонин I, ANP, GATA-4, Nkx2.5 и MEF-2c), уже известны и находятся в таких общедоступных базах данных, как GenBank, а маркер-специфичные последовательности, используемые в качестве праймеров или зондов, можно легко определить.
Для подтверждения дифференциации полипотентных клеток в кардиомиоциты также можно использовать физиологические показатели. Например, подходящие показатели включают в себя спонтанную пульсацию клеток, полученных из полипотентных клеток, экспрессию различных ионных каналов и способность взаимодействовать с электрофизиологическими стимулами.
Полипотентные стволовые клетки, которые можно использовать в настоящем изобретении, включают в себя клетки ES, MAPC и т.п., полученные из таких млекопитающих, как мыши, обезьяны и люди, данные клетки уже широко используются для культивирования. Конкретные примеры мышиных клеток ES включают в себя клетки EB3, клетки E14, клетки D3, клетки CCE, клетки R1, клетки 129SV, клетки J1 и т.п. Были разработаны стандартные методы получения, субкультивирования и хранения клеток ES, клеток EG и MAPC для получения таких полипотентных стволовых клеток, кроме указанных выше, можно использовать другие ссылки (Matsui et al., Cell 70:841, 1992; Shamblott et al., Proc. Natl. Acad. USA 95:13726, 1998; U.S. Patent № 6090622; Jiang et al., Nature 418:41, 2002; International Patent Disclosure 01/11011).
Клетки, которые можно использовать в настоящем изобретении, не ограничиваются тремя вышеуказанными типами и включают в себя все полипотентные стволовые клетки, полученные из эмбрионов и зародышей млекопитающих, пуповинной крови млекопитающих или из тканей, крови и т.п. взрослых млекопитающих, например из органов и костного мозга взрослых особей. Конкретные примеры включают в себя стволовые клетки, полученные путем обработки клеток эпителиального влагалища корня волоса или эпидермальных клеток таким средством, как 5-азацитидин (Sharda & Zahner, International Patent Disclosure 02/051980), стволовые клетки, полученные путем обработки моноцитов антителами CR3/43 (Abuljadayel, Curr. Med. Res. Opinion 19:355, 2003), и стволовые клетки, которые имеют характеристики, подобные характеристикам клеток ES, например стволовые клетки, полученные из клеток внутреннего уха взрослой особи (Li et al., Nature Med., Advance online publication). В данном случае характеристическое подобие клеткам ES определяется с точки зрения индивидуальных цитобиологических свойств клеток ES, таких как наличие поверхностных маркеров (антигенов), специфичных для клеток ES, экспрессия генов, характерных для клеток ES, или способность продуцировать тератомы или химерных мышей.
В способах настоящего изобретения можно использовать даже клетки, которые по характеристикам отличаются от клеток ES, или клетки, отличные от полипотентных стволовых клеток, если данные клетки способны дифференцироваться in vitro в клетки, обладающие признаками кардиомиоцитов. Примеры таких клеток включают в себя мезенхимальные стволовые клетки костного мозга (Bruder et al., патент США № 5736396; Pittenger et al., Science 284:143, 1999), клетки CMG (Makino et al., J. Clin. Invest. 103:697, 1999; описание международного патента 01/048151) и клетки Spoc, полученные из мышечной ткани (описание международного патента 031/035382).
Любой способ, подходящий для индукции дифференциации кардиомиоцитов, можно использовать как способ культивирования для получения кардиомиоцитов из полипотентных стволовых клеток в настоящем изобретении, в случае клеток ES примеры таких способов включают в себя культивирование в суспензии с получением агрегатов, культивирование в висячей капле, совместное культивирование с питающими клетками, культивирование с вращением, культивирование на мягком агаре, культивирование на микроносителе и т.п. Конкретным примером культивирования в суспензии с получением агрегатов является способ, включающий в себя суспендирование клеток ES в виде одиночных клеток (отдельные клетки, которые при диспергировании не прилипают друг к другу вследствие обработки ферментом или т.п.) в среде с получением плотности клеток, предпочтительно, от 10 клеток/мл до 1×107 клеток/мл или, более предпочтительно, от 100 клеток/мл до 1×106 клеток/мл, высевание их в чашку для культивирования и культивирование в течение 4-30 дней или, предпочтительно, 6-15 дней при 37°С в условиях продувания 5% СО2.
В другом воплощении при использовании способа совместного культивирования с питающими клетками вид данных клеток особо не ограничивается, однако, предпочтительно, они представляют собой клетки, обладающие характеристиками мезенхимальных клеток, и, более предпочтительно, клетки, обладающие свойствами, подобными свойствам клеток стромы костного мозга, такие как клетки ST2, клетки OP9, клетки PA6 и т.п. Данные питающие клетки культивируют до получения высокой плотности и формируют из них фидер, используя такие способы, как обработка митомицином С, облучение и т.п., затем клетки ES, суспендированные в виде отдельных клеток в среде с получением плотности, составляющей от 1 клетки/мл до 1×106 клеток/мл, предпочтительно, от 100 клеток/мл до 1×105 клеток/мл или, более предпочтительно, от 1×103 клеток/мл до 1×104 клеток/мл, высевают на полученный фидер и культивируют в течение 4-30 дней или, предпочтительно, 6-15 дней при 37°С в условиях продувания 5% СО2.
В настоящем изобретении вещество, ингибирующее сигнальный путь BMP, представляет собой вещество, которое блокирует или прерывает сигнальный путь, опосредованный BMP, примеры данного вещества включают в себя антагонисты BMP, семейство специфичных нейтрализующих антител против BMP, солюбилизированные молекулы рецептора BMP (относящиеся к типу рецепторов, не связанных с клеточной мембраной) и т.п. Другие примеры включают в себя генные векторы экспрессии, специфичные антисмысловые олигонуклеотиды, рибозимы, антисмысловую РНК для связывания РНК, низкомолекулярные соединения и т.п., которые либо подавляют, либо останавливают экспрессию молекул семейства BMP или функциональных продуктов генов их рецепторов, или, иначе, они подавляют или останавливают экспрессию генов, которые кодируют компоненты, препятствующие активации сигнального пути BMP.
Антагонистом BMP называют вещество, которое связывается с молекулой семейства BMP (такой как BMP-2, BMP-4, BMP-7 или т.п.) и блокирует или ингибирует связывание молекулы BMP с рецептором BMP на поверхности клетки, или вещество, которое подавляет или прерывает сигнальный путь BMP путем связывания с рецептором BMP. Примеры предпочтительных антагонистов BMP в настоящем изобретении включают в себя Noggin, Chordin и т.п. Другие примеры включают в себя Noggin-родственные белки или Chordin-родственные белки, которые на 80% или более или, предпочтительно, на 90% или более гомологичны аминокислотной последовательности соответствующего белка и обладают активностью антагонистов BMP. Примеры антагонистов BMP также включают в себя Noggin-родственные белки или Chordin-родственные белки, кодируемые нуклеотидами, которые в жестких условиях (например, 2×SSC) гибридизуются с нуклеотидами, кодирующими Noggin или Chordin.
Настоящее изобретение также описывает культивирование полипотентных стволовых клеток в присутствии вещества, которое временно ингибирует сигнальный путь BMP, и, хотя способ с использованием такого вещества особо не ограничивается, предпочтительно использовать способ, включающий в себя добавление в питательную среду очищенного рекомбинантного белка. Однако можно использовать любой другой способ, который обеспечивает такие же эффекты, как и способ, включающий в себя добавление в культуру вещества, которое ингибирует сигнальный путь BMP, в виде очищенного рекомбинантного белка. Примеры включают в себя добавление Noggin или другого антагониста BMP, экстрагированного и выделенного из живых тканей, введение генного вектора экспрессии Noggin или другого антагониста BMP непосредственно в полипотентные стволовые клетки, введение генного вектора экспрессии Noggin или другого антагониста BMP в поддерживающие клетки и использование полученных трансфицированных клеток для совместного культивирования, а также применение культурального супернатанта или другого клеточного продукта данных трансфицированных клеток и т.п., все указанные способы включены в настоящее изобретение как часть воплощения, связанного с добавлением в среду антагониста BMP.
При осуществлении настоящего изобретения вещество, используемое для ингибирования сигнального пути BMP, в виде белка и его ген предпочтительно получают из животных тех же видов, которые используются для получения полипотентных стволовых клеток. Например, при осуществлении настоящего изобретения с использованием мышиных полипотентных стволовых клеток в качестве мышиного белка Noggin можно использовать простой в использовании и коммерчески доступный очищенный рекомбинантный белок (рекомбинантный белок мышиный Noggin/Fc-химера; R&D Systems, Genzyme Technology), который получают из мышиных клеток, содержащих и экспрессирующих гибридный ген, состоящий из кДНК мышиного Noggin, связанной с кДНК константного участка (Fc) иммуноглобулина. Однако можно также использовать вещества, полученные от животных других видов, например вместо упомянутого белка можно использовать очищенный рекомбинантный белок (PeproTech), полученный путем введения кДНК человеческого Noggin в E.coli и его экспрессии. Также можно использовать белки Noggin, полученные от свиней, овец, лошадей, птиц (таких как куры) или амфибий (таких как Xenopus). Подобные белки можно использовать в случае Chordin и других антагонистов BMP. Нуклеотидные последовательности генов, кодирующих данные факторы, имеются в общедоступных базах данных ДНК, таких как Национальный центр биотехнологии США (NCBI), а кДНК данных генов могут быть получены и использованы специалистами в данной области. Например, гены человеческих и мышиных Noggin и Chordin уже идентифицированы, нуклеотидные последовательности человеческого Noggin, мышиного Noggin, человеческого Chordin и мышиного Chordin находятся в базе данных NCBI под номерами NM#005450, NM#008711, NM#073411 и NM#009893 соответственно.
Антагонист BMP в настоящем изобретении может представлять собой вещество, которое связывается с молекулой BMP (такой как BMP-2, BMP-4, BMP-7 или т.п.) и блокирует или ингибирует связывание молекулы BMP с рецептором BMP на поверхности клетки, или вещество, которое подавляет или прерывает сигнальный путь BMP путем связывания с рецептором BMP, типичные примеры включают в себя Noggin (Re'em-Kalma et al., Proc. Natl. Acad. Sci. USA 92:12141, 1995; Zimmerman et al., Cell 86:599, 1996) и Chordinn (Piccolo et al., Cell 86:589, 1996; De Robertis et al., U.S. Patent № 5679783; LaVallie et al., U.S. Patent № 5846770; Lavallie et al., U.S. Patent № 5986056). Конкретные примеры других антагонистов BMP включают в себя фоллистатин, фетуин, склеростин и молекулы, принадлежащие к семейству DAN/Cerberus, такие как DAN, Cerberus, гремлин, Dante и т.п. (Balemans & Hul, Dev. Biol. 250:231, 2002). При осуществлении настоящего изобретения также можно использовать синтетические или рекомбинантные аналоги встречающихся в природе антагонистов BMP.
Способы обработки полипотентных стволовых клеток настоящего изобретения не ограничиваются способами, в которых применяются Noggin и другие антагонисты BMP, можно также использовать способы, обеспечивающие такие же эффекты, как и способы, в которых применяются антагонисты BMP, другими словами, способы, вызывающие ингибирование сигнального пути BMP. Конкретные примеры способов, которые вызывают ингибирование сигнального пути BMP, включают в себя способы, в которых используются специфичные нейтрализующие антитела против молекул семейства BMP, и способы, в которых используются солюбилизированные (не связанные с клеточной мембраной) молекулы рецепторов BMP. Другие примеры включают в себя способы введения в клетки генных векторов экспрессии, специфичных антисмысловых олигонуклеотидов, рибозимов, антисмысловых РНК для связывания РНК, низкомолекулярных соединений и т.п., которые либо подавляют, либо останавливают экспрессию молекул семейства BMP или функциональных продуктов генов их рецепторов, или, иначе, подавляют или останавливают экспрессию генов, которые кодируют компоненты, препятствующие активации сигнального пути BMP.
Если при осуществлении настоящего изобретения используются вещества, ингибирующие сигнальный путь BMP (такие как Noggin, Chordin или другие антагонисты BMP), и полипотентные стволовые клетки, применение данных факторов можно разделить на две стадии. Данные стадии относятся к периоду до и после распределения полипотентных стволовых клеток либо в виде отдельных клеток, либо в виде скоплений клеток, состоящих из небольшого количества клеток, и культивирования агрегатов в суспензии или совместного культивирования с поддерживающими клетками и т.п., причем первую стадию ниже называют стадией, предшествующей дифференциации, а последнюю стадию называют стадией индукции дифференциации.
На стадии, предшествующей дифференциации, вещество, ингибирующее сигнальный путь BMP в полипотентных стволовых клетках, применяют за 1-2 дня до или, предпочтительно, за 3 дня или более до образования EB, чтобы индуцировать дифференциацию.
Кроме того, на стадии, предшествующей дифференциации, клетки ES предпочтительно культивируют в обычных условиях, подходящих для данного вида животных и позволяющих поддерживать их в недифференцированном состоянии. То есть в случае мышиных клеток ES желательно добавить в среду фактор, ингибирующий лейкоз (LIF) в концентрации от 100 до 10000 ед/мл, или, предпочтительно, от 500 до 2000 ед/мл.
Вещество, ингибирующее сигнальный путь BMP, не следует применять в течение всего периода культивирования, например, если в качестве вещества, ингибирующего сигнальный путь BMP, используют антагонист BMP, такой как Noggin или Chordin, то культивирование в среде, содержащей антагонист BMP в активном состоянии, проводят в течение первых 5 дней или, более предпочтительно, в течение первых 3 дней периода индукции дифференциации. Однако время, в течение которого применяется вещество, ингибирующее сигнальный путь BMP, может варьировать соответствующим образом в зависимости от условий, таких как вид животного, от которого получены данные клетки, используемый штамм клеток, способ индукции дифференциации и тип вещества, используемого для ингибирования сигнального пути BMP.
Если при осуществлении настоящего изобретения в качестве вещества, ингибирующего сигнальный путь BMP, используют антагонист BMP (такой как белок Noggin или белок Chordin), старую среду удаляют в стерильных условиях и заменяют на среду, содержащую белок Noggin или белок Chordin в концентрации от 1 нг/мл до 2 мкг/мл, предпочтительно, от 5 нг/мл до 1000 нг/мл, более предпочтительно, от 10 нг/мл до 500 нг/мл, после чего культивирование продолжают предпочтительно в течение нескольких дней. Если для ингибирования сигнального пути BMP используется другое вещество, его концентрация может варьировать соответствующим образом в зависимости от типа вещества.
Затем кардиомиоциты, полученные из клеток ES или других полипотентных стволовых клеток с помощью вышеописанных способов, можно собрать, выделить и очистить, используя известные методы, с эффективным получением больших количеств кардиомиоцитов высокой степени чистоты. Полученные таким образом кардиомиоциты далее называют кардиомиоциты настоящего изобретения.
Для очистки кардиомиоцитов можно использовать любой известный способ выделения и очистки клеток, конкретные примеры которого включают в себя проточную цитометрию, магнитные шарики, пэннинг и другие методы, включающие в себя реакции антиген-антитело (см. Monoclonal Antibodies: principles and practice, Third Edition (Acad. Press, 1993); Antibody Engineering: A Practical; Approach (IRL Press at Oxford University Press, 1996), а также фракционирование клеток путем центрифугирования в градиенте плотности с использованием таких носителей, как сахароза, перколл и т.п. Другой способ отбора кардиомиоцитов включает в себя вначале введение генетической модификации в клетки ES или другие полипотентные стволовые клетки, придающей им устойчивость к лекарственному средству или способность к экспрессии эктопического белка, и затем сбор клеток, обладающих морфологией кардиомиоцитов. Например, путем введения в мышиные клетки ES генной кассеты, обеспечивающей экспрессию гена устойчивости к неомицину (G418) под контролем промотора тяжелой цепи α-миозина, Field и соавторы успешно сконструировали систему, в которой клетки ES дифференцируются в кардиомиоциты, причем выживать в среде, содержащей G418, могут только те клетки, которые экспрессируют ген тяжелой цепи α-миозина, и доказано, что 99% или более клеток, отобранных по данному способу как G418-устойчивые клетки, являются кардиомиоцитами (U.S. Patent № 6015671, Specifications; Klug et al., J. Clin. Invest. 98:216, 1996).
Кардиомиоциты настоящего изобретения используются для фармакологического анализа и оценки активности различных биологически активных веществ (например, лекарственных средств) и новых генных продуктов с неизвестными функциями. Например, их можно использовать для скрининга веществ и лекарственных средств, участвующих в регуляции дифференцировки кардиомиоцитов из клеток ES и других полипотентных стволовых клеток, веществ и лекарственных средств, участвующих в регуляции функционирования кардиомиоцитов, а также веществ и лекарственных средств, токсичных для кардиомиоцитов или ингибирующих их функции. В частности, в настоящее время существует очень мало способов скрининга с использованием человеческих кардиомиоцитов, а кардиомиоциты настоящего изобретения являются источником клеток, пригодным для осуществления таких способов скрининга. С другой стороны, для такого скрининга можно также использовать аналитический набор, содержащий кардиомиоциты настоящего изобретения.
Тестируемые вещества, которые можно подвергать скринингу, включают в себя любые вещества, которые можно добавлять в культуру, такие как низкомолекулярные соединения, органические соединения, неорганические соединения, белки, пептиды, гены, вирусы, клетки, культуральные жидкости, жидкости микробных культур и т.п. Эффективными способами введения генов в культуральные системы являются способы, включающие в себя добавление в культуральные системы ретровирусов, аденовирусов и других вирусных векторов, а также способы, включающие в себя добавление факторов после внедрения в липосомы и другие искусственные конструкты.
Тестируемое вещество можно анализировать путем оценки эффективности индукции дифференциации клеток ES и других полипотентных стволовых клеток в кардиомиоциты или количественных и качественных изменений клеточных функций миокарда. Например, эффективность тестируемого соединения при индукции миокардиальной дифференциации можно определить биохимическими и иммунохимическими методами путем детекции экспрессии различных специфичных для миокарда маркеров в полипотентных стволовых клетках после их культивирования по способу настоящего изобретения в течение 5-15 или, предпочтительно, 7-12 дней. Особого ограничения по биохимическим или иммунохимическим методам не существует, однако предпочтительно используют иммунохимические методы, такие как иммунохимическое окрашивание или иммуноэлектрофорез. В данных методах можно использовать маркер-специфичные поликлональные антитела или моноклональные антитела, которые связываются с кардиомиоцитами. Антитела против отдельных специфичных маркеров коммерчески доступны и просты в использовании. Примеры специфичных для кардиомиоцитов маркеров включают в себя тяжелую и легкую цепи миозина, α-актинин, тропонин I, ANP, GATA-4, Nkx2.5, MEF-2c и т.п.
Выживание миокардиальных клеток представляет собой одну из клеточных функций, которую можно использовать в качестве показателя для оценки тестируемого вещества. А именно, апоптоз (гибель клеток) можно индуцировать путем высевания кардиомиоцитов настоящего изобретения в культуральную чашку до получения подходящей плотности клеток и культивирования их в среде, не содержащей сыворотки, при этом в среду можно добавить соответствующее количество тестируемого вещества и определить степень выживания или гибели кардиомиоцитов. Степень выживания или гибели кардиомиоцитов можно определить путем макроскопического наблюдения по окрашиванию таким красителем, как триптановый синий, с помощью метода, включающего в себя использование дегидрогеназной активности (восстановительной активности) в качестве маркера, или с помощью метода, включающего в себя использование в качестве маркера экспрессии аннексина V или каспазной активности, специфичных для клеток в состоянии апоптоза. Наборы, использующиеся на основе данных механизмов, изготовляются многими производителями, в том числе Sigma, Clonetech и Promega, и просты в употреблении.
Поскольку вещество или лекарственное средство, полученное с помощью такого способа скрининга, индуцирует дифференциацию кардиомиоцитов и регулирует их функцию, его можно использовать, например, как средство для профилактики или лечения заболеваний сердца, включающих в себя инфаркт миокарда, ишемическую болезнь сердца, застойную сердечную недостаточность, гипертрофическую кардиомиопатию, расширяющую кардиомиопатию, миокардит, хроническую сердечную недостаточность и т.п. Данные соединения могут быть новыми соединениями или известными соединениями.
Кроме того, кардиомиоциты настоящего изобретения можно использовать в качестве средств для регенерации миокарда или для лечения заболеваний сердца. Примеры заболеваний сердца включают в себя инфаркт миокарда, ишемическую болезнь сердца, застойную сердечную недостаточность, гипертрофическую кардиомиопатию, расширяющую кардиомиопатию, миокардит, хроническую сердечную недостаточность и т.п. При использовании в качестве средств для регенерации миокарда или для лечения заболеваний сердца кардиомиоциты настоящего изобретения, при условии высокой степени чистоты, можно использовать в виде любой формы, такой как клетки, суспендированные в среде или другом водном носителе, клетки, заключенные в биодеградируемом носителе или другой подложке, или клетки, образующие однослойный или многослойный лист миокарда (Shimizu et al., Circ. Res. 90:40, 2002).
Хотя и не ограниченные особо указанными, способы транспортирования вышеуказанных терапевтических средств к участку повреждения включают в себя непосредственное введение в сердце через открытую грудную клетку или шприц, трансплантацию через хирургическое рассечение сердца и трансплантацию через кровеносные сосуды с использованием катетера Murry et al., Cold Spring Harb. Symp. Quant. Biol. 67:519, 2002; Menasche, Ann. Thorac. Surg. 75:S20, 2003; Dowell et al., Cardiovasc. Res. 58:336, 2003). Крайне благоприятные терапевтические эффекты были описаны для трансплантации кардиомиоцитов, полученных из фетального сердца, в пораженные сердца животных с использованием таких методов (Menasche, Ann. Thorac. Surg. 75:S20, 2003; Reffelmann et al., Heart Fail. Rev. 8:201, 2003). Кардиомиоциты, полученные из клеток ES, имеют характеристики, очень похожие на характеристики кардиомиоцитов, полученных их фетальных сердец (Maltsev et al., Mech. Dev. 44:41, 1993; Cicr. Res. 75:233, 1994). Более того, наблюдается большое сходство результатов, полученных при трансплантации фетального миокарда, и результатов, полученных в экспериментах на животных, в которых проводилась трансплантация кардиомиоцитов, полученных из клеток ES, в сердца взрослых особей (Klug et al., J. Clin. Invest. 98:216, 1996). Следовательно, ожидается, что восполнительная трансплантация кардиомиоцитов настоящего изобретения в ткань больного сердца будет способствовать улучшению функционирования сердца в случаях вышеупомянутых заболеваний сердца, развивающихся в результате повреждения или утраты клеток сердца.
Примеры
Настоящее изобретение разъясняется более подробно с помощью приведенных ниже примеров, однако данные примеры служат только для иллюстрации настоящего изобретения и не ограничивают его объем.
Пример 1
Влияние обработки Noggin на дифференциацию клеток ES в кардиомиоциты
Влияние добавления белка Noggin в культуральную среду на развитие кардиомиоцитов изучают на экспериментальной системе, в которой в результате культивирования клеток ES в суспензии с получением агрегатов (ниже в примерах данной заявки называемом просто культивирование в суспензии) образуются EB с последующей дифференциацией в кардиомиоциты.
В описанных ниже экспериментах в качестве клеток ES используют клетки EB3 (поставляемые Prof. Hitoshi Niwa of Riken, Japan), клетки R1 (поставляемые Andrew Nagy of Mount Sinai Hospital, Canada) и клетки 129SV (поставляемые Dainippon Pharmaceutical Co.), но, как правило, при использовании данных линий клеток ES различий не наблюдают. Указанные клетки ES пересевают и поддерживают в недифференцированном состоянии с помощью методов, описанных (Hogan et al. Eds., Cold Spring Harbor Laboratory Press, 1994), Embryonic Stem Cells: Methods and Protocols (Turksen Ed., Humana Press, 2002) и т.п., с использованием минимальной поддерживающей среды Глазго (GMEM, Sigma), содержащей 10% фетальной бычьей сыворотки (FBS), 0,1 мМ МЕМ не эссенциальных аминокислот, 2 мМ L-глутамин и 0,1 мМ 2-меркаптоэтанол, к которой добавлено 2000 ед/мл фактора, ингибирующего лейкоз (LIF) (ESGRO; Chemicon).
За 3 дня до начала культивирования в суспензии клетки ES, которые образуют колонии в описанных выше условиях, промывают дважды PBS, диспергируют в виде отдельных клеток путем обработки 0,25% раствором трипсина, содержащим 1 мМ EDTA, и получают суспензию клеток с плотностью 2,5×105 клеток/мл с использованием α-модифицированной минимальной поддерживающей среды (α-МЕМ, Sigma), содержащей 10% FBS, 0,1 мМ МЕМ не эссенциальных аминокислот, 2 мМ L-глутамин и 0,1 мМ 2-меркаптоэтанол (далее называемой "среда для дифференциации"), в присутствии 2000 ед/мл LIF. К данной суспензии добавляют или не добавляют 500 нг/мл рекомбинантного белка мышиный Noggin/Fc-химера (R&D Systems, далее называемого "белок Noggin"), после чего в случае клеток EB3, которые представляют собой клеточную линию ES, не зависимую от питателя, суспензию высевают на коммерчески доступные чашки для культивирования клеток, покрытые желатином (колба T75, Greiner). В случае не зависимых от питателя клеточных штаммов R1 и 129SV покрытые желатином чашки засевают фибробластами мышиных эмбрионов, обработанных митомицином (Dainippon Pharmaceutical), с получением питающих клеток, на которые затем высевают суспензию клеток.
EB получают из клеток ES путем культивирования в суспензии следующим способом. Клетки ES, которые культивировали в течение 3 дней в присутствии или в отсутствии Noggin, дважды промывают PBS, диспергируют в виде отдельных клеток путем обработки 0,25% раствором трипсина, содержащим 1 мМ EDTA, и помещают в коммерчески доступные чашки Петри с низким прилипанием клеток (диаметр 100 мм; Valmalk) в концентрации 1×102 клеток/мл или 2×105 клеток/мл. За три дня до начала культивирования в суспензии в среду некоторых групп добавляют 500 нг/мл белка Noggin. Затем клетки держат в условиях, предотвращающих прилипание к чашкам, и культивируют в суспензии в течение 4-14 дней. В данных экспериментальных условиях культивирования клетки ES начинают агрегировать и образовывать EB сразу после начала культивирования в суспензии, а автономная пульсация наблюдается в некоторых EB приблизительно на 7-8 день культивирования.
EB можно также получить из клеток ES путем культивирования в висячей капле, которое проводят следующим образом. Клетки ES, которые культивировали в течение 3 дней в среде для дифференциации в присутствии или в отсутствие Noggin, дважды промывают PBS и диспергируют в виде отдельных клеток путем обработки 0,25% раствором трипсина, содержащим 1 мМ EDTA. Затем получают капли, содержащие 500 клеток в 15 мкл среды для дифференциации, подвешенные на крышках культуральных чашек, и культивируют в течение 4 дней. В это время в среду групп клеток ES, предварительно обработанных Noggin в течение 3 дней, добавляют 500 нг/мл белка Noggin. После культивирования в висячей капле, EB, образовавшиеся в каплях, помещают на коммерчески доступные культуральные чашки (4-луночные Multidish; Nunc), заполненные средой для дифференциации, и культивируют в прилипшем состоянии, индуцируя дифференциацию в кардиомиоциты. После культивирования в прилипшем состоянии половину среды заменяют на свежую среду каждые 2 дня. В данных экспериментальных условиях наблюдают агрегацию клеток ES и образование EB сразу после начала культивирования в висячей капле, как и при культивировании в суспензии, а спонтанная пульсация наблюдается в некоторых колониях EB приблизительно на 3 день культивирования в прилипшем состоянии собранных EB (7-8 день после начала культивирования в висячей капле).
Периодически определяют процент EB, способных к спонтанной пульсации, как удобный показатель дифференциации и развития клеток ES в кардиомиоциты. Если клетки EB3 используют в качестве клеток ES и культивируют в суспензии с плотностью 1×102 клеток/мл, очень небольшое количество EB, полученных из клеток ES, не обработанных Noggin (группа Noggin(-)), способно к пульсации даже после 14 дней культивирования (фиг.1А). С другой стороны, пульсацию наблюдают у EB, полученных из клеток ES, обработанных Noggin (группа Noggin(+)), начиная с 7-го дня культивирования в суспензии, а на 14 день пульсацию наблюдают у 80 или более % EB. такую же тенденцию наблюдают у клеток, культивированных в суспензии в концентрации 2×105 клеток/мл, причем, в конечном счете, пульсация наблюдается менее чем у 10% EB в группе Noggin(-) и у 30% или более в группе Noggin(+) на 10 день и у 90% или более на 14 день. У EB Noggin(-) пульсация наблюдается в некоторых ограниченных участках, тогда как у EB Noggin(+) пульсация неожиданно наблюдается во всех участках поверхностного слоя.
Даже в условиях культивирования в висячей капле способность к миокардиальной дифференциации у клеток ES Noggin(+) гораздо выше, чем у клеток ES Noggin(-), причем пульсация наблюдается у 40% или более колоний EB на 7 день после образования EB (3 день культивирования с прилипанием), у 80% на 10 день и практически у всех на 14 день (фиг.1В). Как и в условиях культивирования в суспензии, в группе Noggin(-) пульсация клеток наблюдается в некоторых частях колоний, образовавшихся в результате прилипания EB к культуральным чашкам, тогда как в группе Noggin(+) пульсация клеток наблюдается практически во всех колониях EB.
Различные рекомбинантные цитокины и факторы роста, такие как IGF(инсулин-подобный фактор роста)-1, FGF(фактор роста фибробластов)-2, BMP-2 и т.п. добавляют в среду в тех же условиях, что и Noggin, однако эффекта индукции дифференциации, равного или превышающего эффект Noggin, не наблюдается.
Далее, авторы настоящего изобретения исследовали, как влияет концентрация белка Noggin на индукцию дифференциации клеток ES в кардиомиоциты. Клетки культивируют в суспензии в условиях, приведенных на фиг.1, за исключением того, что концентрация добавленного белка Noggin варьирует от 0,5 до 1500 нг/мл, как показано на фиг.2. Клетки EB3 и клетки R1 демонстрируют почти одинаковую зависимость от дозы, а процент способных к пульсации EB при добавлении белка Noggin в концентрации от 5 до 1500 нг/мл значительно выше, чем в группе Noggin(-). В частности, очень хорошая частота появления способных к пульсации EB наблюдается при добавлении белка Noggin в концентрации от 50 до 150 нг/мл.
Пример 2
Свойства кардиомиоцитов, полученных из клеток ES, обработанных Noggin
Как показано в примере 1, количество способных к пульсации EB, полученных из клеток ES, значительно возрастает при обработке Noggin, и, чтобы подтвердить, что способные к пульсации клетки среди данных EB являются кардиомиоцитами, авторы исследовали экспрессию различных маркерных генов и белков, специфичных для миокарда.
Экспрессия различных, специфичных для клеток миокарда, маркерных генов в группах Noggin(+) и Noggin(-) показана на фиг.3. EB, полученные путем культивирования в суспензии в условиях, приведенных на фиг.1, периодически собирают и выделяют общую РНК с использованием набора RNeasy (Qiagen). Затем получают кДНК путем обратной транскрипции традиционными методами с использованием Superscript II (Invitrogen), после чего специфичные для кардиомиоцитов гены детектируют с помощью полимеразной цепной реакции (ПЦР). Для детекции транскриптов GATA-4, TEF-1, Tbx-5, MEF-2c, αMHC, MLC-2v, α-кардиальный актин и GAPDH используют следующие праймеры.
GATA-4 (прямой) 5'-CTGTCATCTC ACTATGGGCA-3' (SEQ ID NO:1)
GATA-4 (обратный) 5'-CCAAGTCCGA GCAGGAATTT-3' (SEQ ID NO:2)
TEF-1 (прямой) 5'-AAGACGTCAA GCCCTTTGTG-3' (SEQ ID NO:3)
TEF-1 (обратный) 5'-AAAGGAGCAC ACTTTGGTGG-3' (SEQ ID NO:4)
Tbx-5 (прямой) 5'-GGAGCCTGAT TCCAAAGACA-3' (SEQ ID NO:5)
Tbx-5 (обратный) 5'-TTCAGCCACA GТТСАСGTТС-3' (SEQ ID NO:6)
МЕF-2c (прямой) 5'-AGCAAGAATA CGATGCCATC-3' (SEQ ID NO:7)
MEF-2c (обратный) 5'-GAAGGGGTGG TGGTACGGTC-3' (SEQ ID NO:8)
αMHC (прямой) 5'-GGAAGAGTGA GCGGCCATCA AGG-3' (SEQ ID NO:9)
αMНС (обратный) 5'-CTGCTGGAGA GGTTATTCCT CG-3' (SEQ ID NO:10)
MLC-2v (прямой) 5'-GCCAAGAAGC GGATAGAAGG-3' (SEQ ID NO:11)
MLC-2v (обратный) 5'-CTGTGCTTCA GGGCTCAGTC-3' (SEQ ID NO:12)
α-кардиальный актин (прямой) 5'-CTGAGATGTC ТСТСТСТСТС TТАG-3' (SEQ ID NO:13)
α-кардиальный актин (обратный) 5'-ACAATGACTG ATGAGAGATG-3' (SEQ ID NO:14)
GAPDH (прямой) 5'-TTCAACGGCA CAGTCAAGG-3' (SEQ ID NO:15)
GAPDH (обратный) 5'-CATGGACTGT GGTCATGAG-3' (SEQ ID NO:16)
ПЦР проводят, используя GeneAmp PCR System 9600 (Perkin-Elmer) и TaKaRa Taq (Takara) в качестве устойчивой к нагреванию ДНК-полимеразы. Вначале реакционный раствор ПЦР, содержащий кДНК, нагревают в течение 3 минут при 94°С, затем его подвергают 30 циклам, включающим в себя 1 минуту при 94°С, 1 минуту при 55°С и 1 минуту при 72°С, и в конце нагревают в течение 5 минут при 72°С и охлаждают до 4°С. Продукт ПЦР подвергают электрофорезу на 3% полиакриламидном геле, окрашивают SYBR Green I (Takara) и детектируют с помощью Molecular Imager FX (Bio-Rad).
В результате в группе EB Noggin(-) в период от 5 до 10 дня культивирования в суспензии наблюдается относительно слабая экспрессия GATA-4, TEF-1, Tbx-5, MLC-2v и других генов, тогда как в группе EB Noggin(+) экспрессия наблюдается с первого дня (день 0) культивирования, и затем в период от 5 до 15 дня наблюдается высокий уровень экспрессии. Экспрессия MEF-2c и специфичного для миокарда α-актина наблюдается, начиная с 5 дня культивирования, и уровень экспрессии значительно выше в группе EB Noggin(+). Высокий уровень экспрессии гена α-МНС (тяжелой цепи миозина) наблюдается на 10 и 15 дни культивирования в группе EB Noggin(+), но не является достоверным в группе EB Noggin(-). Полученные результаты показывают, что обработка Noggin быстро и эффективно стимулирует дифференциацию EB в кардиомиоциты.
Экспрессия специфичных для кардиомиоцитов маркерных белков в способных к пульсации клетках, образовавшихся в группе EB Noggin(+), также подтверждается иммунохимическим окрашиванием. EB группы Noggin(+), полученные по методу, приведенному на фиг.1, собирают на 12-й день культивирования и обрабатывают 0,25% раствором трипсина, содержащим 1 мМ EDTA, чтобы диспергировать клетки. Диспергированные клетки с низкой плотностью высевают на покрытые желатином покровные стекла так, чтобы клетки не прилипали друг к другу и культивируют в коммерчески доступных культуральных чашках, заполненных средой для дифференциации. На следующий день клетки на покровных стеклах фиксируют 4% раствором формальдегида, затем подвергают взаимодействию с антителами против саркомерного миозина (MF20; Американская коллекция типовых культур), с антителами против тропонина (#sc-8120; Santa Cruz Biotechnology), с антителами против α-актинина (#sc-15335; Santa Cruz) и с антителами против ANP (#AB5490; Chemicon) в качестве первичных антител и с Alexa488-меченными вторичными антителами (Molecular Probes), после чего наблюдают под флюоресцентным микроскопом.
В результате обнаружено, что большое количество клеток являются положительными по отношению к специфичным для миокарда маркерным белкам саркомерному миозину, тропонину I, α-актинину и ANP (фиг.4), это свидетельствует о том, что способные к пульсации клетки, полученные из клеток ES, представляют собой кардиомиоциты.
Затем с помощью описанного выше метода иммуноцитохимического окрашивания подтверждают распределение и долю кардиомиоцитов в EB. EB, собранные на 12 день культивирования в суспензии, сразу помещают в приспособление для получения замороженных срезов (OCT Compound, Sakura Finetek USA Inc.) и затем замораживают жидким азотом. Из замороженных образцов получают срезы толщиной 5 мкм с помощью криостата (Leica CM3050-S) и помещают на предметные стекла. Полученные замороженные срезы подвергают взаимодействию с антителами против вышеупомянутых маркеров, специфичных для кардиомиоцитов, и клетки, положительные по отношению к маркерным белкам, определяют описанными выше методами.
Результаты приведены на фиг.5. В то время как клетки, положительные по отношению к маркерным белкам, специфичным для кардиомиоцитов, наблюдаются в очень ограниченных участках группы EB Noggin(-), в группе EB Noggin(+) миокардиальный маркер-положительные клетки обнаружены почти во всех участках поверхностного слоя EB. Данный факт хорошо согласуется с результатами наблюдений, полученными в примере 1, которые показывают, что, в то время как в группе EB Noggin(-) способный к пульсации участок ограничен только частью EB, в группе EB Noggin(+) способность к пульсации наблюдается почти во всех участках EB.
Пример 3
Влияние разницы во времени и периоде обработки Noggin на дифференциацию клеток ES в кардиомиоциты
Чтобы определить оптимальное время и период обработки Noggin, варьируют время добавления белка Noggin в описанную выше систему культивирования в суспензии и регистрируют влияние на индукцию миокардиальной дифференциации.
Результаты приведены в таблице 1.
В таблице 1 приведена частота появления способных к пульсации EB (полученных из клеток EB3) в разных условиях культивирования, приведенные данные показывают, что период добавления Noggin (до или после образования EB) имеет большое значение. В таблице 1 "стадия, предшествующая дифференциации", относится к периоду от 3 дней до начала культивирования в суспензии, тогда как "стадия индукции дифференциации" относится к периоду образования EB. В таблице 1 "+" означает, что Noggin (150 нг/мл) добавляют в среду, а "-" означает, что Noggin не добавляют.
Вначале исследовали необходимость обработки Noggin до и после дифференциации при использовании способа культивирования в суспензии. Способные к пульсации EB появляются с высокой частотой, если клетки ES культивируют в среде, содержащей белок Noggin (150 нг/мл) в течение 3 дней до образования EB, и затем белок Noggin (150 нг/мл) также добавляют в начале культивирования в суспензии (день 0), как в примерах 1 и 2, но частота появления способных к пульсации EB гораздо ниже, если клетки ES предварительно культивируют в среде, не содержащей белка Noggin, даже если белок Noggin затем добавляют на стадии образования EB. Также имеет значение присутствие LIF, который добавляют в среду, чтобы поддержать клетки ES в недифференцированном состоянии, в культуре мышиных клеток ES перед культивированием в суспензии, и, если клетки ES на стадии, предшествующей дифференциации, культивируют, не добавляя LIF (2000 ед/мл), частота появления способных к пульсации EB значительно меньше даже при обработке Noggin.
Однако появление способных к пульсации кардиомиоцитов значительно подавляется, если белок Noggin (150 нг/мл) добавляют в культуру позже 5 дня. Более того, если он постоянно присутствует в течение 7 или более дней при культивировании в суспензии, способные к пульсации кардиомиоциты практически отсутствуют и наблюдается индукция дифференциации в нейрон-подобные клетки, как было описано ранее (Gratsch & O'Shea, Dev. Biol. 245:83, 2002).
Полученные результаты показывают, что обработка Noggin важна для индукции дифференциации клеток ES в кардиомиоциты как до, так и сразу после образования ЕВ и что постоянное присутствие высокой концентрации белка Noggin после индукции, наоборот, препятствует развитию кардиомиоцитов.
Пример 4
Ингибиторное влияние BMP на индукцию миокардиальной дифференциации под действием Noggin
Чтобы подтвердить, что индукция миокардиальной дифференциации путем обработки Noggin клеток ES происходит в результате активности Noggin как антагониста BMP, авторы исследовали эффекты, оказываемые BMP-2 при добавлении в среду одновременно с Noggin.
Культивирование в суспензии проводят в условиях, описанных в примерах 1 и 2, с добавлением разных концентраций BMP-2 одновременно с белком Noggin (150 нг/мл), результаты приведены на фиг.6. BMP-2 в зависимости от концентрации ингибирует индукцию миокардиальной дифференциации в результате обработки Noggin клеток ES, а при добавлении 5 нг/мл или более BMP-2 появление способных к пульсации EB практически не наблюдается.
Пример 5
Влияние обработки Noggin на систему индукции миокардиальной дифференциации с использованием питающих клеток
Было описано, что дифференциацию кардиомиоцитов можно индуцировать, не используя трехмерную систему культивирования, такую как культивирование в суспензии или культивирование в висячей капле, путем совместного культивирования клеток ES с клетками ST2, OP9 или другими стромальными клетками, высеянными заранее на культуральные чашки в качестве поддерживающих клеток (питающих клеток) (Yamane et al., Methods Mol. Biol. 184:261, 2002; Schroeder et al., Proc. Natl. Acad. Sci. USA 100:4018, 2003). Авторы исследовали влияние обработки Noggin в системе индукции дифференциации с использованием клеток ST2 (полученных из банка клеток Riken) в качестве питающих клеток. Клетки ST2 высевают на коммерчески доступные 6-луночные планшеты для культивирования клеток (Corning), культивируют до слияния, используя среду, содержащую MEM Дульбекко (Invitrogen) и 10% FBS (Invitrogen), и используют их в качестве питающих клеток. Суспензию клеток ES в состоянии отдельных клеток получают, как описано выше, фидер промывают дважды PBS и высевают с плотностью 2000 клеток/2 мл среды/лунку. Затем каждый день под микроскопом наблюдают появление способных к пульсации кардиомиоцитов и через 8 дней некоторые клетки фиксируют 70% раствором этанола, подвергают взаимодействию с антителами против саркомерного миозина (MF20; Американская коллекция типовых культур) в качестве первичных антител и затем с вторичными антителами, конъюгированными с пероксидазой хрена (Histofine Simple Stain PO(M); Nichirei), после чего проводят цветную реакцию с использованием субстратной жидкости (Nichirei) ACE (3-амино-9-этилкарбазол) и наблюдают с помощью оптического микроскопа.
В данных условиях культивирования клетки ES, высеянные на клетки ST2, образуют колонии, размер которых позволяет видеть их при макроскопическом наблюдении через несколько дней после помещения в чашки, клетки (колонии) положительные по отношению к миокардиальному клеточному маркеру, саркомерному миозину (здесь: MF20-положительные) обнаруживают после 8 дней культивирования, тогда как способные к пульсации кардиомиоциты появляются приблизительно на 12 день культивирования. Клетки ES, которые культивировали в присутствии белка Noggin (150 нг/мл) в течение 3 дней перед помещением в чашки, высевают на клетки ST2 и культивируют вместе с ними в среде, содержащей Noggin (150 нг/мл) в течение 2 дней. Результаты приведены на фиг.7. В то время как более 60% колоний, образовавшихся из клеток ES на фидере, являются MF20-положительными в группе Noggin(-), в группе Noggin(+) положительными являются 90% или более колоний, а процент MF20-положительных клеток в каждой колонии гораздо выше в группе Noggin(+), чем в группе Noggin(-). Более того, способные к пульсации кардиомиоциты появляются с более высокой частотой в группе Noggin(+): в то время, как в группе Noggin(-) приблизительно 30% колоний, образовавшихся из клеток ES на питающих клетках, способны к пульсации, в группе Noggin(+) наблюдается приблизительно 70% колоний, способных к пульсации.
Пример 6
Влияние обработки Chordin на дифференциацию клеток ES в кардиомиоциты
Изучают влияние Chordin, который, как и Noggin, действует как антагонист BMP, на индукцию миокардиальной дифференциации в сравнении с Noggin. В качестве белка Chordin используют коммерчески доступный рекомбинантный химерный белок Chordin-Fc (рекомбинантный мышиный Chordin/Fc химера, R&D Systems: здесь называется просто "белок Chordin"). Культивирование в висячей капле проводят по способу примера 1 с использованием клеток ES, которые культивировали 3 дня в среде, в которую был добавлен Noggin (500 нг/мл) или Chordin (500 нг/мл), или в среде, не содержащей факторов. На 4-й день культивирования в висячей капле EB собирают и высевают на культуральных чашках, после чего индуцируют дифференциацию в кардиомиоциты путем культивирования с прилипанием. В результате способные к пульсации кардиомиоциты появляются среди клеток ES, обработанных Chordin, приблизительно с такой же частотой, как и среди клеток ES, обработанных Noggin (фиг.8), подтверждая, что, как и Noggin, Chordin оказывает индуцирующее действие на миокардиальную дифференциацию клеток ES.
Эффекты Chordin также исследовали, используя разные методы получения EB/индукции миокардиальной дифференциации. Клетки ES, которые культивировали в течение 3 дней в среде, содержащей Noggin (150 нг/мл) или Chordin (150 нг/мл) или не содержащей никаких факторов, высевают на коммерчески доступные сфероидные культуральные планшеты (96-луночные планшеты, Sumitomo Bakelite) с получением EB и индукцией дифференциации в кардиомиоциты. В данном случае устанавливается группа, получающая Chordin (150 нг/мл), и группа, не получающая Chordin.
Как показано на фиг.9, способные к пульсации кардиомиоциты наблюдаются среди клеток ES, обработанных Chordin (С→С на фигуре), как и среди клеток, обработанных Noggin (N→N на фигуре), снова подтверждая, что Chordin так же индуцирует миокардиальную дифференциацию клеток ES, как и Noggin. Стимуляция миокардиальной дифференциации существенно выше в группе, предварительно обработанной Chordin в течение 3 дней перед индукцией и не обработанной после образования ЕВ (С→(-) на фигуре), а в группе, не обработанной Chordin перед образованием EB, но с добавлением Chordin после образования EB ((-)→С на фигуре) стимуляция имеет такой же уровень, как и в необработанной группе ((-)→(-) на фигуре). Такой же эффект наблюдается и при использовании Noggin вместо Chordin, причем гораздо более сильный эффект стимуляции миокардиальной дифференциации наблюдается в группе, обработанной Noggin в течение 3 дней до образования ЕВ, но в которой Noggin не добавляют после образования EB, и в группе, которую не обрабатывают Noggin перед образованием EB, но в которой Noggin добавляют после образования EB, чем в необработанной группе.
Далее, для исследования эффективности обработки Noggin и Chordin человеческих клеток используют эмбриональные раковые клетки (называемые здесь клетки EC), которые подобны клеткам ES. Клетки NCCIT (полученные от Американской коллекции типовых культур), клеточная линия человеческих EC, культивируют в течение 3 дней таким же способом, как и вышеупомянутые мышиные клетки ES, в среде, содержащей Noggin (15 нг/мл) или Chordin (15 нг/мл) или не содержащей никаких факторов. Затем их высевают на коммерчески доступные чашки для культивирования в суспензии с получением EB, которые культивируют в течение 14 дней. Экспрессию генов или продукцию белков кардиомиоцит-специфичных маркеров анализируют методом ПЦР или иммуноцитохимического окрашивания, используя способы, описанные в примере 2. Результаты показывают, что в клетках NCCIT, обработанных Noggin или Chordin, наблюдается экспрессия/продукция миокардиального маркера миозина.
Пример 7
Влияние других антагонистов BMP на дифференциацию клеток ES в кардиомиоциты
Далее, исследуют влияние 4 антагонистов BMP, фоллистатина, DAN, Caronte (куриный Cerberus-подобный фактор) и гремлина (рекомбинантные белки типа Fc-химера; R&D System). Клетки ES, которые культивировали в течение 3 дней в среде, содержащей (150 нг/мл) или не содержащей вышеупомянутые антагонисты BMP, высевают на коммерчески доступные сфероидные культуральные планшеты (96-луночные планшеты, Sumitomo Bakelite) с получением EB и индукцией дифференциации в кардиомиоциты.
Результаты показывают, что данные факторы индуцируют миокардиальную дифференциацию с такой же или более высокой эффективностью, как и Noggin или Chordin (фиг.10). Более того, подобные эффекты получают при добавлении рекомбинантного белка (рекомбинант мышиный BMPR-1A/Fc-химера; R&D System), который соответствует внеклеточному домену рецептора BMP 1A и конкурентно ингибирует связывание молекул семейства BMP с эндогенными рецепторами.
Пример 8
Эффект обработки BMP4-siPHK на дифференциацию кардиомиоцитов из клеток ES
ВМР4 представляет собой молекулу семейства BMP, которая экспрессируется до стадии дифференциации и на ранних стадиях дифференциации. В этом примере было подтверждено, что введение и экспрессия siPHK, специфичной в отношении ВМР4, эффективно индуцирует дифференциацию кардиомициотов из клеток ES.
Аналогично способу, описанному в примере 1 настоящего изобретения, клетки ES мыши пересевали и поддерживали в недифференцируемом состоянии. За 3 дня до индукции дифференциации клетки ES диспергировали до единичных клеток, обрабатывая трипсином. Затем в клетки вводили siPHK, специфичную в отношении гена ВМР4 (BMP4-siPHK; Ambion), используя Nucleofector (Amaxa), и экспрессировали. Последовательности BMP4-siPHK были следующими.
(прямая) 5'-CCACAGCGGUCCAGGAAGATT-3'
(обратная) 5'-UCUUCCUGGACCGCUGUGGTT-3'
Экспрессия введенной BMP4-siPHK в клетках продолжалась в течение 72 часов и более, и ген, экспрессирующий ВМР4, полностью ингибировался. После культивирования клеток в течение 3 дней до дифференциации формировали ЕВ аналогичными способами, описанными в примере 6, используя сфероидные культуральные планшеты (мультилуночные планшеты SUMILONCELLTIGHT, Sumitomo Bakelite) для индукции дифференциации.
Как показано на фиг.11, лишь у нескольких ЕВ в необработанной группе ("без добавления") отмечали пульсацию даже через 14 дней культивирования. С другой стороны, в группах, обработанных антагонистами BMP, такими как Chordin и DAN, пульсация подтверждалось приблизительно у 50-60% ЕВ на 10-й день дифференциации. Способность к автономной пульсации также была подтверждена приблизительно у 40% ЕВ в группе, обработанной BMP4-siPHK. Более того, в необработанной группе пульсация была ограничена лишь определенными областями ЕВ, тогда как в группах, обработанных антагонистами BMP и BMP4-siPHK, пульсация отмечалась практически на протяжении всей поверхности ЕВ.
Кроме того, была исследована экспрессия маркерных генов, специфичных для кардиомиоцитов, используя те же способы, описанные в примере 2 настоящего изобретения. Как показано на фиг.12, высокие уровни экспрессии GATA4, Nkx2.5, αМНС, βМНС и других генов была подтверждена 14-й день дифференциации в группах, обработанных Chordin, DAN или BMP4-siPHK, по сравнению с необработанной группой.
С помощью иммуноцитохимического окрашивания была исследована продукция специфичных маркерных белков кардиомиоцитов, используя способы, аналогичные способам, описанным в примере 2 настоящего изобретения. Результаты представлены на фиг.13. Несмотря на то что лишь ограниченное количество необработанных ЕВ было положительно на маркеры, специфичные для кардиомиоцитов, большинство клеток в ЕВ в группах, обработанных BMP4-siPHK, а также в группах, обработанных антагонистами BMP (Chordin и DAN), были положительны и имели сильный сигнал.
Эти результаты демонстрируют, что пульсирующие кардиомиоциты возникали из клеток ES, в которых была ингибирована экспрессия гена ВМР4 за счет введения и экспрессии BMP4-siPHK, а также из клеток ES, обработанных антагонистами BMP.







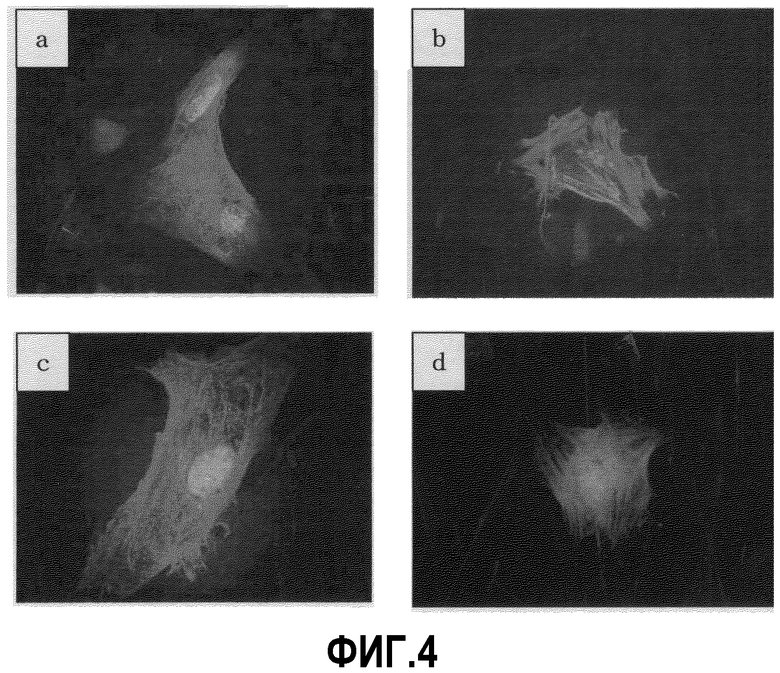
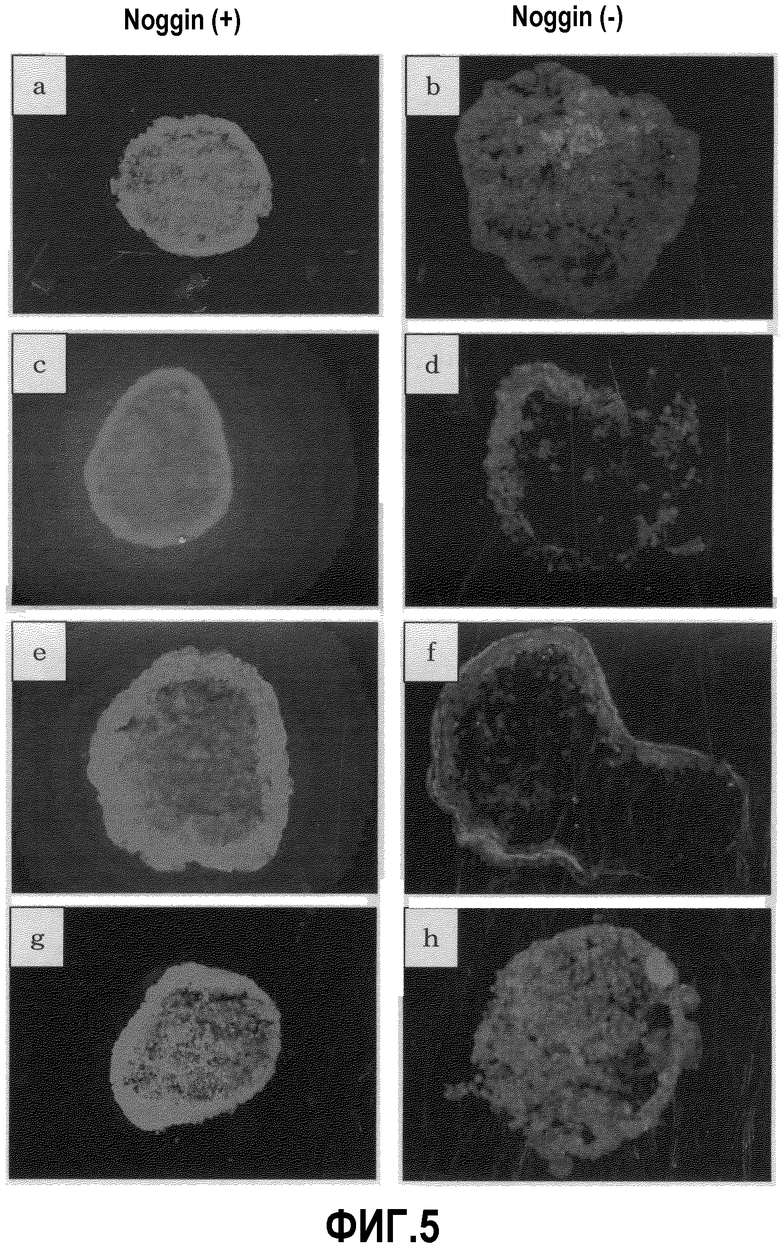
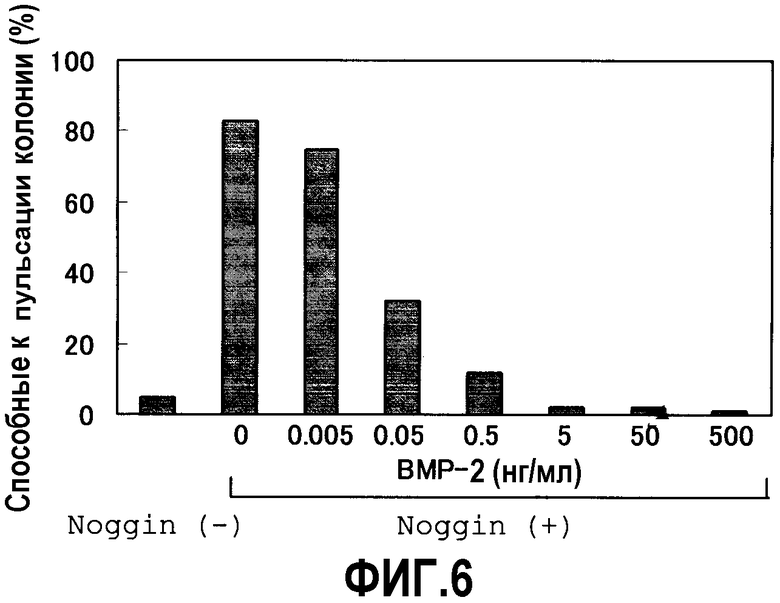

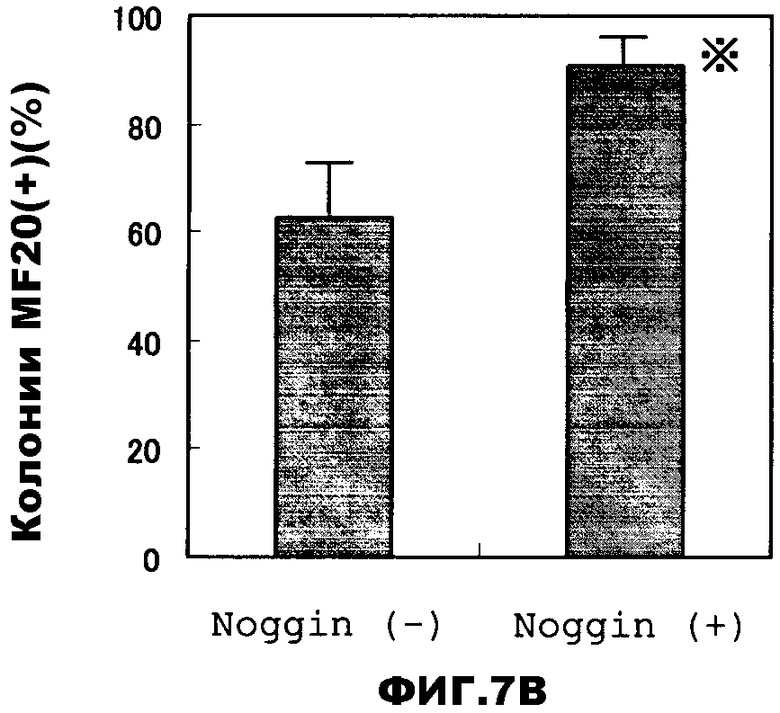



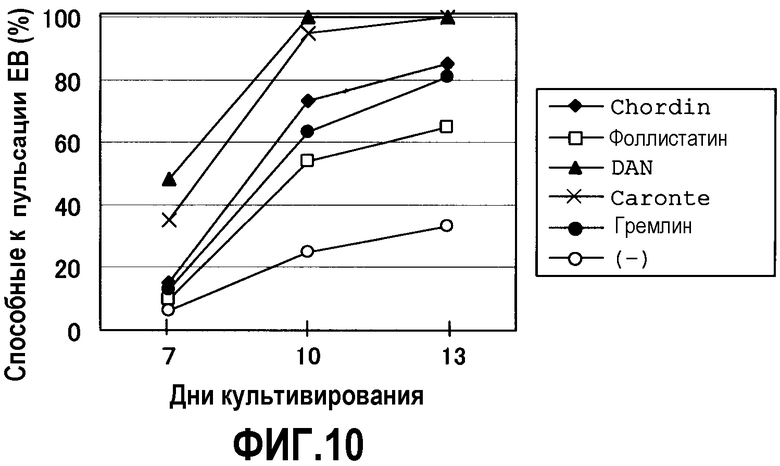

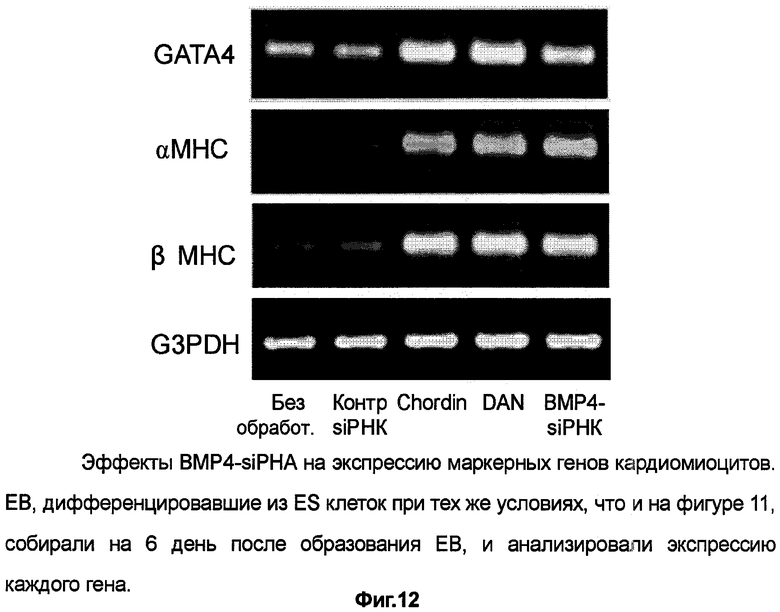

Изобретение относится к области клеточной биологии, конкретно к индукции дифференциации стволовых клеток в миокардиальные клетки-предшественники, которые могут быть использованы для эффективных методов лечения сердечной недостаточности. Культивируют полипотентные стволовые клетки с целью индукции дифференциации в присутствии вещества, ингибирующего сигнальный путь, опосредованный BMP, с помощью агрегационного культивирования в суспензии или совместного культивирования с питающими клетками. Вещество, ингибирующее сигнальный путь, опосредованный BMP, добавляют либо до дифференциации, либо в течение первых нескольких стадий культивирования с индукцией дифференциации, либо при объединении двух указанных методов. Изобретение позволяет увеличить эффективность дифференциации кардиомиоцитов из стволовых клеток и получить более чистую культуру дифференцированных клеток по сравнению с известным способом. 9 з.п. ф-лы, 13 ил., 1 табл.
1. Способ индукции дифференциации кардиомиоцитов из стволовых клеток, где
стволовые клетки культивируют с целью индукции дифференциации в присутствии вещества, ингибирующего сигнальный путь, опосредованный BMP, и включающий стадию обработки стволовых клеток веществом, которое ингибирует сигнальный путь BMP
(a) в течение стадии до дифференциации; и/или
(b) в течение первых нескольких дней стадии индукции дифференциации, и
где культивирование стволовых клеток с целью индукции дифференциации включает по меньшей мере одну из стадий, выбранных из группы, состоящей из:
a) стадию получения эмбриоидных телец путем культивирования в суспензии с агрегацией;
b) стадию совместного культивирования с питающими клетками; и
c) стадию культивирования в чашках на культуральном контейнере, где вещество, ингибирующее сигнальный путь, опосредованный BMP, представляет собой по меньшей мере одно из веществ, выбранных из группы, состоящей из: антагонистов BMP, специфически нейтрализующего антитела против молекул семейства BMP, солюбилизированной (не связанной с клеточной мембранной) молекулы рецептора BMP, генного вектора экспрессии, специфического антисмыслового олигонуклеотида, рибозима, антисмысловой РНК для РНК-интерференции и низкомолекулярного соединения.
2. Способ по п.1, где вещество, ингибирующее сигнальный путь, опосредованный BMP, представляет собой антагонист BMP и/или солюбилизированную (не связанную с клеточной мембранной) молекулу рецептора BMP.
3. Способ по п.1 или 2, где веществом, которое ингибирует сигнальный путь, опосредованный BMP, является антагонист BMP.
4. Способ по п.3, где стволовые клетки являются клетками млекопитающих, способными к дифференциации в кардиомиоциты in vitro.
5. Способ по п.3, где антагонист BMP представляет собой одно или несколько средств, выбранных из группы, включающей в себя Noggin, Chordin, фетуин, фоллистатин, склеростин, DAN, Cerberus, гремлин, Dante и родственные им белки.
6. Способ по п.5, где стволовые клетки являются клетками млекопитающих, способными к дифференциации в кардиомиоциты in vitro.
7. Способ по п.1, где стволовые клетки представляют собой клетки млекопитающих, обладающие способностью к дифференциации в кардиомиоциты in vitro.
8. Способ по п.7, где клетки млекопитающих, обладающие способностью к дифференциации в кардиомиоциты, представляет собой полипотентные стволовые клетки или клетки, полученные из них.
9. Способ по п.8, где полипотентные стволовые клетки представляют собой эмбриональные стволовые клетки, клетки, обладающие морфологией, подобной морфологии эмбриональных стволовых клеток, эмбриональные зародышевые клетки или полипотентные клетки-предшественники взрослых особей.
10. Способ по п.9, где полипотентные стволовые клетки представляют собой эмбриональные стволовые клетки.
| MONZEN К | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| MONZEN К | |||
| et al | |||
| Smads, TAK1, and their common target ATF-2 play a critical role in | |||

