[Область техники]
Настоящее изобретение относится к способу получения и способу очистки кардиомиоцитов. Более конкретно, настоящее изобретение относится к способу получения и способу очистки кардиомиоцитов с использованием ингибитора тирозинкиназы рецепторного типа.
[Уровень техники]
Хотя частота инфаркта миокарда в последние годы снижается, болезни сердца, включая инфаркт миокарда, по-прежнему остаются ведущей причиной смерти во всем мире. Трансплантация сердца в настоящее время является единственным методом лечения пациентов с тяжелой сердечной недостаточностью, но нехватка доноров создает проблему для трансплантации сердца. Поэтому клеточная терапия с использованием кардиомиоцитов привлекает внимание как терапевтический способ, улучшающий сердечные заболевания. Кроме того, внимание также привлекает создание теста для оценки эффективности лекарственного средства in vitro или теста на безопасность лекарственного средства с использованием кардиомиоцитов. Следовательно, требуется стабильное поступление однородных кардиомиоцитов, которые можно использовать для клеточной терапии и тестов in vitro.
Один из способов стабильного получения однородных кардиомиоцитов заключается в индукции дифференциации кардиомиоцитов из стволовых клеток или клеток-предшественников кардиомиоцитов, и были предприняты различные попытки разработать эффективный способ индукции дифференциации в кардиомиоциты. В качестве такого способа индукции дифференциации сообщали о способе стимулирования индукции дифференциации из плюрипотентных стволовых клеток в кардиомиоциты путем культивирования плюрипотентных стволовых клеток в среде, содержащей ингибитор EGFR (патентная литература 1), способе индукции дифференциации индуцированных плюрипотентных стволовых клеток в кардиомиоциты, и затем созревание кардиомиоцитов путем контакта кардиомиоцитов с антагонистом нейрегулина 1 или антагонистом ErbB (патентная литература 2), способе стимулирования индукции дифференциации недифференцированных клеток-предшественников, таких как миобласты, путем контакта недифференцированных клеток-предшественников с ингибитором деацетилазы (патентная литература 3), способе стимулирования индукции дифференциации взрослых клеток-предшественников, таких как клетки-предшественники миокарда и подобные, путем контакта клеток-предшественников с ингибитором гистондеацетилазы (HDAC) (патентная литература 4) и подобные. Кроме того, также сообщалось о способе, который не подвергается процессу индукции дифференциации из стволовых клеток. Например, в патентной литературе 5 описан способ получения клеток-предшественников миокарда или кардиомиоцитов из соматических клеток, таких как фибробласты и подобные, путем прямого перепрограммирования.
[Перечень ссылок]
[Патентная литература]
[PTL 1]
WO 2014/136519
[PTL 2]
US-A-2010/0183565
[PTL 3]
WO 2003/033678
[PTL 4]
WO 2009/073618
[PTL 5]
WO 2015/038704
[Сущность изобретения]
[Техническая проблема]
Целью настоящего изобретения является создание способа получения клеточной популяции, содержащей высокочистые кардиомиоциты, с помощью средств, отличных от упомянутых выше обычных способов. Другой целью настоящего изобретения является создание способа очистки кардиомиоцитов высокой чистоты из клеточной популяции, содержащей кардиомиоциты.
[Решение проблемы]
Авторы настоящего изобретения провели интенсивные исследования в попытке решить вышеупомянутые проблемы и пришли к выводу, что в клеточной популяции, уже содержащей кардиомиоциты, доля кардиомиоцитов в клеточной популяции может быть увеличена, другими словами, кардиомиоциты могут быть очищены путем подавления пролиферации клеток, отличных от кардиомиоцитов, или уменьшения количества клеток, отличных от кардиомиоцитов, с помощью ингибиторов, таргетирующих белки, высоко экспрессированные в клетках, отличных от кардиомиоцитов, вместо стимулирования индукции дифференциации из недифференцированных клеток в кардиомиоциты. Следовательно, сначала с использованием клеточной популяции, содержащей кардиомиоциты, индуцированные для дифференциации из iPS клеток, проводят анализ последовательности РНК одиночной клетки для кластеризации клеток в клеточной популяции. В результате кластеризации, было обнаружено, что экспрессия тирозинкиназ рецепторного типа различна между кардиомиоцитами и другими клетками. Таким образом, изменение доли кардиомиоцитов в клеточной популяции исследовали с помощью ингибиторов тирозинкиназы рецепторного типа и обнаружили, что кардиомиоциты можно успешно очищать с помощью ингибитора тирозинкиназы рецепторного типа. Авторы настоящего изобретения провели дополнительные исследования, основанные на этих открытиях, и завершили настоящее изобретение.
Соответственно, настоящее изобретение предлагает следующее.
[1] Способ получения клеточной популяции, содержащей кардиомиоциты, включающий
(1) стадию контакта ингибитора тирозинкиназы рецепторного типа (за исключением ингибиторов рецептора EGF) с клеточной популяцией, содержащей кардиомиоциты или сердечные клетки-предшественники, и другими клетками, где клеточную популяцию получают путем культивирования плюрипотентных стволовых клеток в среде для дифференциации кардиомиоцитов, и
(2) стадию культивирования клеточной популяции.
[2] Способ по [1], где клеточная популяция контактирует с ингибитором тирозинкиназы рецепторного типа на вышеупомянутой стадии (1) через 4 дня после начала индукции дифференциации плюрипотентных стволовых клеток.
[3] Способ по [1] или [2], где клеточная популяция контактирует с ингибитором тирозинкиназы рецепторного типа в течение не менее одного дня на вышеупомянутой стадии (1).
[4] Способ по любому из [1]-[3], где указанный выше ингибитор представляет собой, по меньшей мере, один ингибитор тирозинкиназы рецепторного типа, выбранный из группы, состоящей из рецептора VEGF, рецептора PDGF, рецептора HGF и рецептора FGF.
[5] Способ по любому из [1]-[4], где вышеупомянутый ингибитор представляет собой, по меньшей мере, один член, выбранный из группы, состоящей из N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамида, N-{4-[(6,7-диметоксихинолин-4-ил)окси]-3-фторфенил}-N'-(4-фторфенил)циклопропан-1,1-дикарбоксамида, AMG337, ASP5878, BGJ398, форетиниба, ZM323881, CP-673451, креноланиба и кризотиниба.
[6] Способ по любому из [1]-[5], где вышеупомянутая плюрипотентная стволовая клетка представляет собой индуцированную плюрипотентную стволовую клетку.
[7] Популяция клеток, содержащая кардиомиоциты, полученная способом по любому из [1]-[6].
[8] Агент для терапии на основе трансплантации клеток, включающий клеточную популяцию [7].
[9] Способ очистки кардиомиоцитов, включающий
(1) стадию контакта ингибитора тирозинкиназы рецепторного типа с клеточной популяцией, содержащей кардиомиоциты или сердечные клетки-предшественники, и другие клетки, где клеточную популяцию получают путем культивирования плюрипотентных стволовых клеток в среде для дифференциации кардиомиоцитов, и
(2) стадию культивирования клеточной популяции.
[Преимущества изобретения]
Согласно настоящему изобретению, предложена клеточная популяция, содержащая кардиомиоциты высокой чистоты. Такая клеточная популяция может быть предпочтительно использована для терапии сердечных заболеваний на основе трансплантации клеток. Кроме того, также предложен способ очистки кардиомиоцитов с высокой степенью чистоты из клеточной популяции, содержащей кардиомиоциты и сердечные клетки-предшественники.
[Краткое описание чертежей]
[Фиг. 1]
На фиг. 1 показан график t-SNE кластеризации и уровней экспрессии саркомерного-α-актинина и cTnT (сердечного тропонина T) каждого кластера, основанный на данных секвенирования одиночной РНК кардиомиоцитов, индуцированных для дифференциации из iPS клеток. Обведенные части на верхней панели показывают популяции, не относящиеся к кардиомиоцитам. Вертикальные оси центральной и нижней панелей показывают уровень экспрессии гена, и метка идентичности на горизонтальной оси показывает номер кластера на фиг. 1. Каждая точка показывает отдельную клетку, и часть в рамке показывает кластер не кардиомиоцитов.
[Фиг. 2-1]
На фиг. 2-1 показан график t-SNE результатов кластеризации кардиомиоцитов, полученных из iPS-клеток, и экспрессии генов VEGFR1 и VEGFR2 в каждой клетке на основе данных секвенирования одиночной РНК. На панелях, показывающих экспрессию гена, темно-серые точки показывают клетки с высоким уровнем экспрессии, и светло-серые точки показывают клетки с низким уровнем экспрессии. Популяции клеток с высокой экспрессией обведены кружком, и названия популяций указаны на рисунке (СМ: кардиомиоциты (далее СМ), SMC: клетки, подобные гладким мышцам (далее SMC), END: клетки эндодермальной линии (далее END), EC: клетки, подобные эндотелию).
[Фиг. 2-2]
На фиг. 2-2 показан график t-SNE результатов кластеризации кардиомиоцитов, полученных из iPS-клеток, и экспрессии генов VEGFR3, PDGFRA и PDGFRB в каждой клетке на основе данных секвенирования одиночной РНК. На панели, показывающей экспрессию гена, темно-серые точки показывают клетки с высоким уровнем экспрессии, и светло-серые точки показывают клетки с низким уровнем экспрессии. Популяции клеток с высокой экспрессией обведены кружком, и названия популяций указаны на рисунке (SMC: клетки, подобные гладким мышцам (далее SMC), ЕС: клетки, подобные эндотелию).
[Фиг. 2-3]
На фиг. 2-3 показан график t-SNE результатов кластеризации кардиомиоцитов, полученных из iPS-клеток, и экспрессии генов FGFR4, HGFR (c-Met) и EGFR1 в каждой клетке, основанный на данных секвенирования одиночной РНК. На панели, показывающей экспрессию гена, темно-серые точки показывают клетки с высоким уровнем экспрессии, и светло-серые точки показывают клетки с низким уровнем экспрессии. Популяции клеток с высокой экспрессией обведены кружком, и названия популяций указаны на рисунке (END: клетки линии эндодермы (далее END)).
[Фиг. 2-4]
На фиг. 2-4 показан график t-SNE результатов кластеризации кардиомиоцитов, полученных из iPS-клеток, и экспрессии гена EGFR3 в каждой клетке на основе данных секвенирования одиночной РНК. На панели, показывающей экспрессию гена, темно-серые точки показывают клетки с высоким уровнем экспрессии, и светло-серые точки показывают клетки с низким уровнем экспрессии. Популяция клеток с высокой экспрессией обведена кружком, и название популяции указано на рисунке (END: клетки линии эндодермы (далее END)).
[Фиг. 3]
На фиг. 3 показаны результаты разделения клеточной популяции кардиомиоцитов, происходящих из iPS-клеток, на основе экспрессии CD326, CD31 и CD49a. С помощью проточной цитометрии, кардиомиоциты, полученные из iPS-клеток-репортеров TNNI1, имеющие долю кардиомиоцитов (TNNI1-положительные показатели) 89,6% (левая крайняя панель), делят на CD326-положительные клетки (END, клетки в рамке в центре левой панели), CD326-отрицательные CD31-положительные клетки (EC, клетки в рамке в центре правой панели), CD326-отрицательные CD31-отрицательные CD49a-положительные клетки (SMC, клетки в рамке в центре справа) и CD326-отрицательные CD31-отрицательные CD49a-отрицательные клетки (трижды отрицательные: TN, клетки расположены в левом нижнем углу центральной правой панели). 99% или более TN являются клетками, положительными по маркеру сердечной мышцы TNNI1 (кардиомиоцитами) (правая крайняя панель). Доли EC и SMC показывают не клетки на панели, а числовые значения, также включающие CD326-отрицательные клетки.
[Фиг. 4]
На фиг.4 показана доля кардиомиоцитов после обработки соединением. *: не измерено из-за низкого количества клеток. Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю актинин-положительных клеток (долю кардиомиоцитов).
[Фиг. 5]
На фиг. 5 показана доля не кардиомиоцитов после обработки соединением (END: клетки эндодермальной линии, SMC: клетки, подобные гладкомышечным, EC: клетки, подобные эндотелию). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю не кардиомиоцитов.
[Фиг. 6]
На фиг. 6 показана средняя частота кардиомиоцитов после обработки соединением (стандартное отклонение) (n=4). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю кардиомиоцитов.
[Фиг. 7]
На фиг. 7 показана доля не кардиомиоцитов после обработки соединением (END: клетки эндодермальной линии, SMC: клетки, подобные гладким мышцам, EC: клетки, подобные эндотелию). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю не кардиомиоцитов.
[Фиг. 8]
На фиг. 8 показана средняя частота кардиомиоцитов после обработки соединением (стандартное отклонение) (n=3). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю кардиомиоцитов.
[Фиг. 9]
На фиг. 9 показано среднее относительное соотношение количества восстановленных клеток после обработки соединением, где количество восстановленных клеток без обработки соединением равно 1 (стандартное отклонение) (n=3). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает относительное соотношение количества восстановленных клеток.
[Фиг. 10]
На фиг. 10 показана средняя доля кардиомиоцитов после обработки соединением (стандартное отклонение) (n=4). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю кардиомиоцитов.
[Фиг. 11]
На фиг. 11 показано среднее относительное количество восстановленных клеток после обработки соединением, где количество восстановленных клеток без обработки соединением равно 1 (стандартное отклонение) (n=4). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает относительное соотношение количества восстановленных клеток.
[Фиг. 12]
На фиг.12 показана средняя доля кардиомиоцитов после обработки соединением (стандартное отклонение) (n=4). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю кардиомиоцитов.
[Фиг. 13]
На фиг. 13 показана средняя доля не кардиомиоцитов после обработки соединением (стандартное отклонение) (n=4) (END: клетки эндодермальной линии, SMC: клетки, подобные гладким мышцам, EC: клетки, подобные эндотелию). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю не кардиомиоцитов.
[Фиг. 14]
На фиг. 14 показано среднее относительное количество восстановленных клеток после обработки соединением, где количество восстановленных клеток без обработки соединением равно 1 (стандартное отклонение) (n=4). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает относительное соотношение количества восстановленных клеток.
[Фиг. 15]
На фиг. 15 показана доля кардиомиоцитов после обработки соединением (эксперимент номер 1 является средним значением n=2, n=1 в других). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю кардиомиоцитов.
[Фиг. 16]
На фиг. 16 показана доля не кардиомиоцитов после обработки соединением (END: клетки эндодермальной линии, SMC: клетки, подобные гладким мышцам, EC: клетки, подобные эндотелию). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю не кардиомиоцитов. *: не измерено из-за низкого количества клеток.
[Фиг. 17]
На фиг. 17 показана доля кардиомиоцитов после обработки соединением (эксперимент номер 1 является средним значением n=4, эксперименты номер 2, 3 являются средним значением n=3 (стандартное отклонение), n=1 в экспериментах номер 4, 5). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю кардиомиоцитов.
[Фиг. 18]
На фиг. 18 показана доля не кардиомиоцитов после обработки соединением (эксперимент номер 1 представляет собой среднее значение n=4, эксперимент номер 2 представляет собой среднее значение n=3 (стандартное отклонение), эксперимент номер 3 представляет собой среднее значение n=2, n=1 в экспериментах номер 4, 5) (END: клетки эндодермальной линии, SMC: клетки, подобные гладким мышцам, EC: клетки, подобные эндотелию). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю не кардиомиоцитов.
[Фиг. 19]
На фиг.19 показана средняя доля кардиомиоцитов после обработки соединением (стандартное отклонение) (n=4; n=3 в эксперименте номер 2). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю кардиомиоцитов.
[Фиг. 20]
На фиг. 20 показана средняя доля не кардиомиоцитов после обработки соединением (стандартное отклонение) (n=4) (END: клетки эндодермальной линии, SMC: клетки, подобные гладким мышцам, EC: клетки, подобные эндотелию). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает долю не кардиомиоцитов.
[Фиг. 21]
На фиг. 21 показано среднее относительное количество восстановленных клеток после обработки соединением, где количество восстановленных клеток без обработки соединением равно 1 (стандартное отклонение) (n=4). Вертикальная ось показывает номер эксперимента, и горизонтальная ось показывает относительное соотношение количества восстановленных клеток.
[Подробное описание изобретения]
1. Способ получения клеточной популяции, содержащей кардиомиоциты
Настоящее изобретение предлагает способ получения клеточной популяции, содержащей кардиомиоциты (далее также именуемый «способ получения по настоящему изобретению»). Способ получения по настоящему изобретению включает (1) стадию контакта ингибитора тирозинкиназы рецепторного типа с клеточной популяцией, содержащей кардиомиоциты или сердечные клетки-предшественники, и другие клетки, и (2) стадию культивирования клеточной популяции. В способе получения по настоящему изобретению, ингибиторы рецептора EGF исключены из «ингибитора тирозинкиназы рецепторного типа» в упомянутой выше (1).
В настоящем описании, «кардиомиоциты» означают клетки, положительные, по меньшей мере, по одному из саркомерного α-актинина, сердечного тропонина Т (cTnT) и тропонина I типа 1 (TNNI1), и, предпочтительно, представляют собой клетки, положительные по саркомерному α-актинину. Как правило, это клетка сердечной мышцы, обладающая способностью к самопульсации. «Сердечная клетка-предшественник» означает клетку-предшественник вышеупомянутых кардиомиоцитов, положительную, по меньшей мере, по одному из Nkx2.5, GATA4, MEF2C и MESP1. Вышеупомянутая «другая клетка» означает клетку, которая не является кардиомиоцитом или сердечной клеткой-предшественником (далее также именуемую «не кардиомиоцитом»), и примеры конкретной клетки включают гладкомышечную клетку, эндотелиальную клетку, стволовую клетку (например, плюрипотентную стволовую клетку) и подобные.
В настоящем описании, «положительный» означает, что белок или ген экспрессируется в количестве, определяемом, по меньшей мере, любым способом, известным в данной области техники. Белок можно обнаружить с помощью иммунологического анализа с использованием антитела, такого как ELISA, иммуноокрашивания или проточной цитометрии. В случае белка, который экспрессируется внутриклеточно и не появляется на клеточной поверхности (например, фактора транскрипции или его субъединицы и подобных), репортерный белок экспрессируется вместе с белком, и белок-мишень может быть обнаружен с помощью обнаружения репортерного белка. Ген можно обнаружить, например, способом амплификации нуклеиновой кислоты и/или способом обнаружения нуклеиновой кислоты, таким как ОТ-ПЦР, биочип (например, микрочип), секвенирование РНК или подобные. Экспрессию белка или гена можно определить обычным способом. Например, когда используется проточная цитометрия, и уровень экспрессии относительно высок по сравнению с уровнем экспрессии контрольной группы, демонстрирующей отрицательную экспрессию белка, можно определить, что белок обнаруживаемо экспрессируется.
В настоящем описании, «отрицательный» означает, что уровень экспрессии белка или гена меньше нижнего предела обнаружения всеми или любым из вышеупомянутых известных способов. Нижний предел обнаружения экспрессии белка или гена может варьироваться в зависимости от каждого способа, но может быть определен общим способом.
В настоящем описании, «клеточная популяция» означает популяцию, состоящую из двух или нескольких клеток одного типа или разных типов. «Клеточная популяция» также означает массу клеток одного или разных типов. Популяцию клеток, содержащую кардиомиоциты или сердечные клетки-предшественники, а также не кардиомиоциты для использования на вышеупомянутой стадии (1), можно получить путем культивирования плюрипотентных стволовых клеток в среде для дифференциации кардиомиоцитов. Следовательно, способ получения по настоящему изобретению может включать, перед вышеупомянутой стадией (1), стадию (0) индукции дифференциации кардиомиоцитов или сердечных клеток-предшественников из плюрипотентных стволовых клеток.
Примеры плюрипотентной стволовой клетки для использования в настоящем изобретении включают индуцированную плюрипотентную стволовую клетку (iPS клетку), эмбриональную стволовую клетку (ES клетку), эмбриональную стволовую клетку, полученную из клонированного эмбриона, полученного трансплантацией ядра (эмбриональную стволовую клетка ядерного переноса: ntES клетку), мультипотентную стволовую клетку зародышевой линии (mGS клетку), эмбриональную зародышевую клетку (EG клетку), Muse клетку (мультилинейно-дифференцирующуюся устойчивую к стрессу клетку). iPS клетка является предпочтительной (iPS клетка человека является более предпочтительной). Когда вышеупомянутая плюрипотентная стволовая клетка представляет собой ES клетку или любую клетку, полученную из эмбриона человека, клетка может быть клеткой, полученной путем разрушения эмбриона, или клеткой, полученной без разрушения эмбриона. Предпочтительно, она представляет собой клетку, полученную без разрушения эмбриона. Вышеупомянутая плюрипотентная стволовая клетка предпочтительно происходит от млекопитающего (например, мыши, крысы, хомяка, морской свинки, собаки, обезьяны, орангутанга, шимпанзе, человека), более предпочтительно, человека. Следовательно, плюрипотентная стволовая клетка, используемая в настоящем изобретении, наиболее предпочтительно представляет собой iPS клетку человека.
«Индуцированная плюрипотентная стволовая клетка (iPS клетка)» относится к клетке, полученной путем перепрограммирования путем введения специфического фактора (фактора ядерного перепрограммирования) в соматическую клетку млекопитающего или недифференцированную стволовую клетку. В настоящее время, существуют различные типы «индуцированных плюрипотентных стволовых клеток (iPS клеток)». В дополнение к iPS клеткам, установленным Yamanaka et al. путем введения четырех факторов Oct3/4, Sox2, Klf4 и c-Myc в фибробласты мыши (Takahashi K, Yamanaka S., Cell, (2006) 126: 663-676), iPS-клетки, полученные из клеток человека, получают путем введения тех же четырех факторов в фибробласты человека (Takahashi K, Yamanaka S., et al. Cell, (2007) 131: 861-872.), клетки Nanog-iPS, полученные путем выбора экспрессии Nanog в качестве показателя после введения вышеупомянутых четырех факторов (Okita, K., Ichisaka, T., and Yamanaka, S. (2007). Nature 448, 313-317.), iPS клетки, полученные способом, не включающим c-Myc (Nakagawa M, Yamanaka S., et al. Nature Biotechnology, (2008) 26, 101-106), и iPS-клетки, полученные путем введения шести факторов безвирусным методом (Okita K et al. Nat. Methods 2011 May; 8(5):409-12, Okita K et al., Stem Cells, 31(3):458-66.). Кроме того, индуцированные плюрипотентные стволовые клетки, полученные путем введения четырех факторов OCT3/4, SOX2, NANOG и LIN28, полученные Thomson et al. (Yu J., Thomson JA. et al., Science (2007) 318: 1917-1920.), индуцированные плюрипотентные стволовые клетки, полученные Daley et al. (Park IH, Daley GQ. et al., Nature (2007) 451: 141-146), индуцированные плюрипотентные стволовые клетки, полученные Sakurada et al. (JP-A-2008-307007) и подобные.
Кроме того, можно использовать любые индуцированные плюрипотентные стволовые клетки, известные в данной области техники, которые описаны во всех опубликованных статьях (например, Shi Y., Ding S., et al., Cell Stem Cell, (2008) Vol3, Issue 5, 568-574; Kim JB., Scholer HR, et al., Nature, (2008) 454, 646-650; Huangfu D., Melton, DA, et al., Nature Biotechnology, (2008) 26, № 7, 795-797) или патентах (например, JP-A-2008-307007, JP-A-2008-283972, US2008-2336610, US2009-047263, WO2007-069666, WO2008-118220, WO2008-124133, WO2008-151058, WO2009-006930, WO2009-006997, WO2009-007852). В качестве линии индуцированных плюрипотентных стволовых клеток можно использовать различные линии iPS клеток, созданные NIH, RIKEN, Киотским университетом и подобными. Например, в случае линии клеток iPS человека, может быть упомянут штамм HiPS-RIKEN-1A, штамм HiPS-RIKEN-2A, штамм HiPS-RIKEN-12A и штамм Nips-B2 RIKEN, а также штамм 253G1, штамм 201B7, штамм 409B2, штамм 454E2, штамм 606A1, штамм 610B1, штамм 648A1 и запас клеток iPS для регенеративной медицины Киотского университета, и подобные.
«Соматическая клетка» в настоящем описании означает любую клетку животного, за исключением клеток зародышевой линии и дифференцированных тотипотентных клеток, таких как яйцеклетка, ооцит, ES клетки и подобные (предпочтительно, клетки млекопитающих, включая человека). Соматическая клетка конкретно не ограничена и охватывает любые соматические клетки плодов, соматические клетки новорожденных и зрелые здоровые или патогенные соматические клетки, и любые первично культивированные клетки, пассированные клетки и установленные линии клеток. Конкретные примеры соматических клеток включают (1) стволовые клетки тканей (соматические стволовые клетки), такие как нейральные стволовые клетки, гемопоэтические стволовые клетки, мезенхимальные стволовые клетки, стволовые клетки пульпы зуба и подобные, (2) клетки-предшественники тканей, (3) дифференцированные клетки, такие как лимфоциты, эпителиальные клетки, эндотелиальные клетки, миоциты, фибробласты (клетки кожи и т. д.), волосковые клетки, гепатоциты, клетки слизистой оболочки желудка, энтероциты, спленоциты, клетки поджелудочной железы (экзокринные клетки поджелудочной железы и т. д.), клетки мозга, клетки легкого, клетки почки, адипоциты и подобные, и подобные.
ES клетка представляет собой плюрипотентную и самореплицирующуюся стволовую клетку, полученную из внутренней клеточной массы раннего эмбриона (например, бластоцисты) млекопитающего, такого как человек, мышь или подобное. ES клетки были обнаружены у мышей в 1981 году (M.J. Evans and M.H. Kaufman (1981), Nature 292:154-156), после чего линия ES клеток была также установлена у приматов, таких как человек, обезьяна и подобные (J.A. Thomson et al. (1998), Science 282:1145-1147, J.A. Thomson et al. (1995), Proc. Natl. Acad. Sci. USA, 92:7844-7848, J.A. Thomson et al. (1996), Biol. Reprod., 55:254-259, J.A. Thomson and V.S. Marshall (1998), Curr. Top. Dev. Biol., 38:133-165). ES клетка может быть получена путем удаления внутренней клеточной массы из бластоцисты оплодотворенной яйцеклетки животного-мишени и культивирования внутренней клеточной массы на питателе фибробластов. Способы создания и поддержания ES клеток человека и обезьяны описаны, например, в USP 5,843,780; Thomson JA, et al. (1995), Proc Natl. Acad. Sci. U S A. 92:7844-7848; Thomson JA, et al. (1998), Science. 282:1145-1147; Suemori H. et al. (2006), Biochem. Biophys. Res. Commun., 345:926-932; Ueno M. et al. (2006), Proc. Natl. Acad. Sci. USA, 103:9554-9559; Suemori H. et al. (2001), Dev. Dyn., 222:273-279; Kawasaki H. et al. (2002), Proc. Natl. Acad. Sci. USA, 99:1580-1585; Klimanskaya I. et al. (2006), Nature. 444:481-485 и подобные. Альтернативно, ES клетка может быть получена с использованием только одного бластомера эмбриона на стадии расщепления перед стадией бластоцисты (Chung Y. et al. (2008), Cell Stem Cell 2: 113-117) или также может быть получена с использованием эмбриона, который перестал развиваться (Zhang X. et al. (2006), Stem Cells 24: 2669-2676). Что касается «ES клетки», различные линии ES клеток мыши, полученные таргетной лабораторией inGenious, RIKEN, и подобные можно использовать для ES клеток мыши. Для ES клеток человека можно использовать различные линии ES клеток человека, полученные Университетом Висконсина, NIH, RIKEN, Киотским университетом, Национальным центром детского здоровья и развития, Cellartis и подобными. Например, в качестве ES клеточных линий человека, можно использовать штаммы CHB-1 - CHB-12, штамм RUES1, штамм RUES2, штаммы HUES1 - HUES28 и подобные, распространяемые ESI Bio, и штамм H1, штамм H9 и подобные, распространяемых WiCell Research, и штамм KhES-1, штамм KhES-2, штамм KhES-3, штамм KhES-4, штамм KhES-5, штамм SSES1, штамм SSES2, штамм SSES3 и подобные, распространяемые RIKEN.
nt ES клетка (ES клетка ядерного переноса) представляет собой ES клетку, полученную из клонированного эмбриона, полученного методом ядерной трансплантации, и обладает почти теми же свойствами, что и ES клетка, полученная из оплодотворенной яйцеклетки (Wakayama T. et al. (2001), Science, 292: 740-743; S. Wakayama et al. (2005), Biol. Reprod., 72: 932-936; J. Byrne et al. (2007), Nature, 450: 497-502). То есть, ES клетка, созданная из внутренней клеточной массы бластоцисты, полученной из клонированного эмбриона, полученного путем замещения ядра неоплодотворенной яйцеклетки ядром соматической клетки, представляет собой nt ES (ES ядерного переноса) клетку. Для получения nt ES клетки используют комбинацию методики ядерной трансплантации (Cibelli J.B. et al. (1998), Nature Biotechnol., 16: 642-646) и методики получения ES клеток (упомянутой выше) (Kiyoka Wakayama et al., (2008), Experimental Medicine, Vol.26, No.5 (Suppl.), pp. 47-52). При ядерной трансплантации, перепрограммирование может быть выполнено путем инъекции ядра соматической клетки в неоплодотворенную яйцеклетку млекопитающего без ядра и культивирования в течение нескольких часов.
mGS клетка представляет собой плюрипотентную стволовую клетку, полученную из яичка, которая становится источником сперматогенеза. Эта клетка может быть индуцирована дифференциацией в различные линии клеток, такие как ES клетки, и проявляет свойства, например, образования химерной мыши путем трансплантации в бластоцисту мыши и подобные (Kanatsu-Shinohara M. et al. (2003) Biol. Reprod., 69: 612-616; Shinohara K. et al. (2004), Cell, 119: 1001-1012). Она является самообновляемой в культуральной среде, содержащей нейротрофический фактор глиальной клеточной линии (GDNF), может продуцировать зародышевые стволовые клетки путем повторения пассажей в условиях культивирования, сходных с условиями культивирования ES клеток (Masanori Takehashi et al., (2008), Experimental Medicine, Vol. 26, No. 5 (Suppl.), pp. 41 to 46, YODOSHA (Tokyo, Japan)).
EG клетка представляет собой клетку, имеющую плюрипотентность, аналогичную таковой у ES клеток, которую получают из первоначальной зародышевой клетки в пренатальный период. Ее можно получить путем культивирования первичных зародышевых клеток в присутствии таких веществ, как LIF, bFGF, фактор стволовых клеток или подобные (Matsui Y. et al. (1992), Cell, 70: 841-847; J.L. Resnick et al. (1992), Nature, 359: 550-551).
Muse клетка представляет собой in vivo эндогенную неопухолевую плюрипотентную стволовую клетку и может быть получена, например, способом, описанным в WO 2011/007900. Более подробно, Muse клетка представляет собой клетку, обладающую плюрипотентностью, которую получают путем обработки фибробласта или стромальной клетки костного мозга трипсином в течение длительного времени, предпочтительно в течение 8 или 16 ч, и последующим культивированием клеток в суспендированном состоянии, и которая является положительной по SSEA-3 и CD105.
Вышеупомянутая стадия (0) конкретно не ограничена, поскольку плюрипотентные стволовые клетки могут быть индуцированы для дифференциации в кардиомиоциты или сердечные клетки-предшественники. Например, дифференциацию в кардиомиоциты или сердечные клетки-предшественники можно индуцировать путем культивирования плюрипотентных стволовых клеток в среде для дифференциации кардиомиоцитов. В одном варианте осуществления настоящего изобретения, вышеупомянутая стадия (0) может включать (0-1) стадию индукции дифференциации плюрипотентных стволовых клеток в мезодермальные клетки и (0-2) стадию индукции дифференциации мезодермальных клеток в кардиомиоциты или сердечные клетки-предшественники. В настоящем описании, «среда для дифференциации кардиомиоцитов» означает среду, содержащую фактор, способствующий индукции дифференциации кардиомиоцитов, такой как цитокин и подобные (в дальнейшем иногда называемый «фактором, индуцирующим дифференциацию кардиомиоцитов»), и минимальная среда. Вышеупомянутый фактор, который способствует индукции дифференциации в кардиомиоциты, охватывает фактор, необходимый для индукции дифференциации в промежуточные клетки (например, мезодермальные клетки и подобные) в процессе индукции дифференциации из плюрипотентных стволовых клеток в кардиомиоциты или сердечные клетки-предшественники.
Примеры минимальной среды, используемой в настоящем изобретении, включают StemFit (например, StemFit AK03N, StemFit AK02N) (Ajinomoto Co., Inc.), StemPro-34 (Thermo Fisher Scientific), PECM (среда для ES клеток приматов), GMEM (минимальная поддерживающая среда Глазго), IMDM (модифицированная по Искову среда Дульбекко), среда 199, минимальная поддерживающая среда Игла (EMEM), αMEM, модифицированная по Дульбекко среда Игла (DMEM), среда Хэма F12, среда RPMI 1640, среда Фишера, и их комбинированная среда, и подобные.
Можно надлежащим образом добавить ингибиторы ROCK (например, Y-27632, Fasudil/HA1077, SR3677, GSK269962, H-1152, Wf-536 и т. д.), сыворотки (например, сыворотку эмбриона крупного рогатого скота (FBS), сыворотку человека, сыворотку лошади и т.д.) или заменители сыворотки, инсулины, различные витамины (например, витамин С (например, аскорбиновую кислоту)), различные аминокислоты, такие как L-глутамин, заменимая аминокислота и подобные, 2-меркаптоэтанол, тиоглицерин (например, α-монотиоглицерин (MTG)), различные цитокины, факторы стволовых клеток (SCF (фактор стволовых клеток)), активин и т. д.), различные гормоны, различные факторы роста (ингибирующий лейкоз фактор (LIF), основной фактор роста фибробластов (bFGF), TGF-β и т.д.), различные внеклеточные матриксы, различные молекулы клеточной адгезии, антибиотики, такие как пенициллин/стрептомицин, пуромицин и подобные, индикаторы рН, такие как феноловый красный и подобные, и подобные к минимальной среде. Примеры заменителя сыворотки включают альбумин, трансферрин, жирную кислоту, инсулин, предшественник коллагена, микроэлемент, заменитель нокаутирующей сыворотки (KSR), ITS-добавку и их смеси, и подобные.
В настоящем изобретении, «витамин Cs» означает L-аскорбиновую кислоту и ее производные, и «производное L-аскорбиновой кислоты» означает производные, которые превращаются в витамин C в результате ферментативной реакции в живом организме. Примеры производных L-аскорбиновой кислоты для использования в настоящем изобретении включают фосфат витамина С (например, 2-фосфат аскорбиновой кислоты), глюкозид аскорбиновой кислоты, аскорбил этил, сложный эфир витамина С, аскорбил тетрагексилдеканоат, аскорбил стеарат и аскорбил 2-фосфат 6-пальмитат. Предпочтительным является фосфат витамина С (например, 2-фосфат аскорбиновой кислоты). Примеры фосфата витамина С включают соли фосфата L-аскорбиновой кислоты, такие как фосфат L-аскорбиновой кислоты Na и фосфат L-аскорбиновой кислоты Mg.
Индуцированные плюрипотентные стволовые клетки или эмбриоидные тельца можно культивировать с помощью адгезивной культуры или суспензионной культуры. В случае адгезивной культуры, культивирование можно проводить в сосуде для культивирования, покрытом компонентом внеклеточного матрикса, и/или можно культивировать совместно с питающими клетками. Хотя питающая клетка особо не ограничена, можно упомянуть, например, фибробласт (фибробласт эмбриона мыши (MEF), фибробласт мыши (STO) и подобные). Питающие клетки предпочтительно инактивируют известным способом, например, облучением (гамма-лучами и подобными), обработкой противораковым агентом (митомицином С и подобными) или подобными. В качестве компонента внеклеточного матрикса могут быть упомянуты волокнистые белки, такие как матригель (Niwa A, et al., PLoS One.6(7): e22261, 2011), желатин, коллаген, эластин и подобные, глюкозаминогликан и протеогликан, такие как гиалуроновая кислота, хондроитинсульфат и подобные, белки клеточной адгезии, такие как фибронектин, витронектин, ламинин и подобные, и подобные.
Температурные условия культивирования особо не ограничены. Температура составляет, например, от примерно 37°С до 42°С, предпочтительно, от примерно 37°С до 39°С. Культивирование можно проводить в условиях с низким содержанием кислорода, и условия с низким содержанием кислорода в настоящем изобретении означают, например, концентрацию кислорода 15%, 10%, 9%, 8%, 7%, 6%, 5% или ниже этих значений.
Культивирование в суспензии означает культивирование клеток в состоянии, когда они не прилипают к культуральному контейнеру, и конкретно не ограничена. Оно может быть выполнено с использованием культурального контейнера, не подвергнутого искусственной обработке (например, обработке покрытием с внеклеточным матриксом или подобным) для улучшения адгезии к клеткам, или культурального контейнера, искусственно обработанного для подавления адгезии (например, обработкой покрытием с поли-гидроксиэтилметакриловой кислотой (поли-НЕМА) или неионогенным полиолом с поверхностно-активной активностью (плюроник F-127 и др.)). Например, культивирование в суспензии можно проводить с использованием инкубатора, снабженного крыльчаткой, такого как одноразовый биореактор (Biott Corporation), одноразовый биореактор (Thermo Fisher), одноразовый биореактор (Sartorius Stedim), одноразовый биореактор ( GE Healthcare Life Sciences) или подобные. Тип и скорость перемешивания используемого инкубатора могут быть соответствующим образом выбраны специалистами в данной области техники в соответствии с типом культивируемых клеток. Например, скорость перемешивания может составлять, но не ограничена ими, от 0 до 100 об/мин, от 20 до 80 об/мин или от 45 до 65 об/мин.
Во время культивирования в суспензии предпочтительно сформировать эмбриоидное тельце (ЕВ) и культивировать его. Следовательно, вышеупомянутая стадия (0) может включать стадию формирования эмбриоидного тельца из плюрипотентных стволовых клеток. На таком стадии предпочтительно диссоциировать колониеобразующие плюрипотентные стволовые клетки на отдельные клетки, и затем сформировать эмбриоидное тельце. На стадии диссоциации плюрипотентных стволовых клеток, клетки, образующие популяцию путем прилипания друг к другу, диссоциируют (разделяют) на отдельные клетки. Способ диссоциации плюрипотентных стволовых клеток включает, например, способ механической диссоциации и способ диссоциации с использованием раствора для диссоциации, обладающего протеазной и коллагеназной активностью (например, Accutase™, Accumax™ и подобные), или раствора для диссоциации, обладающего только коллагеназной активностью. Предпочтительно, используют способ диссоциации плюрипотентных стволовых клеток с использованием раствора для диссоциации, обладающего протеазной активностью и коллагеназной активностью (особенно предпочтительно, Accumax™). Среда, используемая на вышеуказанной стадии, предпочтительно содержит тиоглицерин, L-глутамин и/или аскорбиновую кислоту.
Факторы, индуцирующие дифференциацию кардиомиоцитов, используемые на вышеупомянутой стадии (0-1), включают, например, вещество, активирующее сигнал Wnt, активин A, BMP4 и bFGF, и их можно использовать отдельно или в комбинации. В одном варианте осуществления настоящего изобретения, активин А, BMP4 и bFGF используют в комбинации. Кроме того, среда, используемая на вышеупомянутой стадии (0-1), предпочтительно содержит тиоглицерин, L-глутамин и/или аскорбиновую кислоту.
В настоящем описании «активатор сигнала Wnt» означает вещество, которое активирует сигнальный путь Wnt. Примеры активатора сигнала Wnt включают белок Wnt, ингибитор GSK3β (например, BIO, CHIR99021 и т.д.) и подобные. Их можно использовать по отдельности или в комбинации. Когда используется активатор сигнала Wnt, его концентрация в среде особо не ограничена. Когда BIO или CHIR99021 используют в качестве активатора сигнала Wnt, его предпочтительно использовать в конечной концентрации от 100 нМ до 100 мкМ, предпочтительно, от 1 мкМ до 10 мкМ в среде.
При использовании активина А, его концентрация в среде предпочтительно составляет от 1 нг/мл до 100 нг/мл, и составляет, например, 1 нг/мл, 2 нг/мл, 3 нг/мл, 4 нг/мл, 5 нг/мл, 6 нг/мл, 7 нг/мл, 8 нг/мл, 9 нг/мл, 10 нг/мл, 11 нг/мл, 12 нг/мл, 13 нг/мл, 14 нг/мл, 15 нг/мл, 16 нг/мл, 17 нг/мл, 18 нг/мл, 19 нг/мл, 20 нг/мл, 30 нг/мл, 40 нг/мл, 50 нг/мл, 60 нг /мл, 70 нг/мл, 80 нг/мл, 90 нг/мл или 100 нг/мл.
При использовании BMP4, его концентрация в среде предпочтительно составляет от 1 нг/мл до 1 мкг/мл, и составляет, например, 1 нг/мл, 2 нг/мл, 3 нг/мл, 4 нг/мл, 5 нг/мл, 6 нг/мл, 7 нг/мл, 8 нг/мл, 9 нг/мл, 10 нг/мл, 11 нг/мл, 12 нг/мл, 13 нг/мл, 14 нг/мл, 15 нг/мл, 16 нг/мл, 17 нг/мл, 18 нг/мл, 19 нг/мл, 20 нг/мл, 30 нг/мл, 40 нг/мл, 50 нг/мл, 60 нг/мл, 70 нг/мл, 80 нг/мл, 90 нг/мл, 100 нг/мл, 200 нг/мл, 300 нг/мл, 400 нг/мл, 500 нг/мл, 600 нг/мл, 700 нг/мл, 800 нг/мл, 900 нг/мл или 1 мкг/мл.
При использовании bFGF, его концентрация в среде предпочтительно составляет от 1 нг/мл до 100 нг/мл, и составляет, например, 1 нг/мл, 2 нг/мл, 3 нг/мл, 4 нг/мл, 5 нг/мл, 6 нг/мл, 7 нг/мл, 8 нг/мл, 9 нг/мл, 10 нг/мл, 11 нг/мл, 12 нг/мл, 13 нг/мл, 14 нг/мл, 15 нг/мл, 16 нг/мл, 17 нг/мл, 18 нг/мл, 19 нг/мл, 20 нг/мл, 30 нг/мл, 40 нг/мл, 50 нг/мл, 60 нг/мл, 70 нг/мл, 80 нг/мл, 90 нг/мл или 100 нг/мл.
Период вышеупомянутой стадии (0-1) конкретно не ограничен, пока получают мезодермальные клетки. Предпочтительно, он составляет не менее 12 часов (например, 1 день, 2 дня или больше) или может составлять не более 6 дней (например, 5 дней, 4 дня, 3 дня или меньше). Кроме того, можно контролировать, получены или нет мезодермальные клетки, и в этом случае это можно определить по экспрессии гена мезодермального маркера. Примеры гена маркера мезодермы включают T, MIXL1, NODAL и подобные.
Примеры фактора, индуцирующего дифференциацию кардиомиоцитов, используемого на вышеупомянутой стадии (0-2), включают ингибитор Wnt и VEGF. Их можно использовать по отдельности или в комбинации. Среда, используемая на вышеупомянутой стадии (0-2), предпочтительно содержит тиоглицерин, L-глутамин и/или аскорбиновую кислоту.
В настоящем описании «ингибитор Wnt» означает вещество, которое ингибирует трансдукцию сигнала от связывания Wnt с рецепторами до накопления β-катенина. Это может быть вещество, которое ингибирует связывание с рецепторами семейства Frizzled, или вещество, которое способствует деградации β-катенина. Примеры ингибитора Wnt включают белок DKK1 (например, в случае человека, номер доступа NCBI: NM_012242), склеростин (например, в случае человека, номер доступа NCBI: NM_025237), IWR-1 (Merck Millipore), IWP-2 (Sigma-Aldrich), IWP-3 (Sigma-Aldrich), IWP-4 (Sigma-Aldrich), PNU-74654 (Sigma-Aldrich), XAV939 (Sigma-Aldrich), их производные и подобные. Среди них предпочтительными являются IWP-3 или IWP-4. Можно использовать только один тип ингибитора Wnt или можно использовать несколько их видов в комбинации.
При использовании ингибитора Wnt его концентрация в среде предпочтительно составляет от 1 нМ до 50 мкМ и составляет, например, 1 нМ, 10 нМ, 50 нМ, 100 нМ, 500 нМ, 750 нМ, 1 мкМ, 2 мкМ, 3 мкМ, 4 мкМ, 5 мкМ, 6 мкМ, 7 мкМ, 8 мкМ, 9 мкМ, 10 мкМ, 15 мкМ, 20 мкМ, 25 мкМ, 30 мкМ, 40 мкМ или 50 мкМ, но не ограничена ими. Более предпочтительно, 1 мкМ.
При использовании VEGF, его концентрация в среде предпочтительно составляет от 1 до 100 нг/мл, и составляет, например, 1 нг/мл, 2 нг/мл, 3 нг/мл, 4 нг/мл, 5 нг/мл, 6 нг/мл, 7 нг/мл, 8 нг/мл, 9 нг/мл, 10 нг/мл, 11 нг/мл, 12 нг/мл, 13 нг/мл, 14 нг/мл, 15 нг/мл, 16 нг/мл, 17 нг/мл, 18 нг/мл, 19 нг/мл, 20 нг/мл, 30 нг/мл, 40 нг/мл, 50 нг/мл, 60 нг/мл, 70 нг/мл, 80 нг/мл, 90 нг/мл или 100 нг/мл.
На вышеупомянутой стадии (0-2), ингибитор BMP и/или ингибитор TGFβ можно дополнительно добавить в минимальную среду в качестве фактора, индуцирующего дифференциацию кардиомиоцитов. В настоящем описании, примеры «ингибитора BMP» включают белковые ингибиторы, такие как хордин, ноггин, фоллистатин и подобные, дорсоморфин (6-[4-(2-пиперидин-1-ил-этокси)фенил]-3-пиридин-4-ил-пиразоло[1,5-а]пиримидин) и его производное (P. B. Yu et al. (2007), Circulation, 116: II_60; P. B. Yu et al. (2008), Nat. Chem. Biol., 4: 33-41; J. Hao et al. (2008), PLoS ONE, 3(8): e2904), LDN-193189 (4-(6-(4-(пиперазин-1-ил)фенил)пиразоло[1,5-а]пиримидин-3-ил)хинолин) и подобные. Среди них предпочтительным является дорсоморфин. Можно использовать только один тип ингибитора BMP или ингибитора TGFβ или можно использовать несколько их видов в комбинации.
При использовании ингибитора BMP, его концентрация в среде предпочтительно составляет от 1 нМ до 50 мкМ, и составляет, например, 1 нМ, 10 нМ, 50 нМ, 100 нМ, 500 нМ, 600 нМ, 700 нМ, 800 нМ, 900 нМ, 1 мкм, 2 мкм, 3 мкм, 4 мкм, 5 мкм, 6 мкм, 7 мкм, 8 мкм, 9 мкм, 10 мкм, 15 мкм, 20 мкм, 25 мкм, 30 мкм, 40 мкм, или 50 мкМ, но не ограничена ими.
В настоящем описании, ингибитор TGFβ означает вещество, которое ингибирует трансдукцию сигнала от связывания TGFβ с рецептором, что приводит к SMAD. Это может быть вещество, которое ингибирует связывание с рецепторами семейства ALK, или вещество, которое ингибирует фосфорилирование SMAD семейством ALK. Примеры ингибитора TGFβ включают Lefty-1 (например, мышь: NM_010094, человек: NM_020997 в номере доступа NCBI), SB431542, SB202190 (все R.K. Lindemann et al., Mol. Cancer, 2003, 2: 20), SB505124. (GlaxoSmithKline), NPC30345, SD093, SD908, SD208 (Scios), LY2109761, LY364947, LY580276 (Lilly Research Laboratories), A-83-01 (WO 2009146408), их производные и подобные. Среди них предпочтительным является SB431542.
При использовании ингибитора TGFβ, его концентрация в среде предпочтительно составляет от 1 нМ до 50 мкМ, и составляет, например, 1 нМ, 10 нМ, 50 нМ, 100 нМ, 500 нМ, 750 нМ, 1 мкМ, 2 мкм, 3 мкм, 4 мкм, 5 мкм, 5,2 мкм, 5,4 мкм, 5,6 мкм, 5,8 мкм, 6 мкм, 7 мкм, 8 мкм, 9 мкм, 10 мкм, 15 мкм, 20 мкм, 25 мкм, 30 мкм, 40 мкм или 50 мкм, но не ограничена ими.
Период вышеупомянутой стадии (0-2) конкретно не ограничен, пока получают кардиомиоциты или сердечные клетки-предшественники. Он составляет, например, не менее 1 дня (например, 1 день, 2 дня, 3 дня, 4 дня, 5 дней, 6 дней, 7 дней или дольше). Поскольку длительное культивирование не влияет на формирование кардиомиоцитов или сердечных клеток-предшественников, верхний предел конкретно не установлен, но обычно он не превышает 40 дней. Кроме того, можно контролировать, получены ли кардиомиоциты или сердечные клетки-предшественники. В этом случае это может быть подтверждено количеством сокращающихся кардиомиоцитов, экспрессией маркеров кардиомиоцитов или сердечных клеток-предшественников, экспрессией ионных каналов, реакцией на электрическую физиологическую стимуляцию и подобным.
Вышеупомянутая стадия (0) может дополнительно включать стадию (0-3) культивирования кардиомиоцитов или сердечных клеток-предшественников, полученных на стадии (0-2), в присутствии или в отсутствие VEGF и/или bFGF. Среда, используемая на этой стадии, предпочтительно содержит тиоглицерин, L-глутамин и/или аскорбиновую кислоту. Кроме того, среда, используемая на этой стадии, может также содержать соединение для созревания миокарда (например, N-(1,1-диоксо-2,3-дигидро-1H-1-бензотиофен-5-ил)-2-(4-{5-[1-оксо-5-(пиперидин-1-ил)-1,3-дигидро-2H-изоиндол-2-ил]-1H-бензоимидазол-2-ил}фенокси)ацетамид) и/или ингибитор мультикиназы (например, 2-(4-{3-[3-(2-амино-2-фенилэтокси)-4-цианофенил]пиразоло[1,5-а]пиримидин-6-ил}-1H-пиразол-1-ил)-N-(2-метоксиэтил)ацетамид).
Когда VEGF используют на стадии (0-3), его концентрация в среде предпочтительно составляет от 1 до 100 нг/мл, и составляет, например, 1 нг/мл, 2 нг/мл, 3 нг/мл, 4 нг/мл, 5 нг/мл, 6 нг/мл, 7 нг/мл, 8 нг/мл, 9 нг/мл, 10 нг/мл, 11 нг/мл, 12 нг/мл, 13 нг/мл, 14 нг/мл, 15 нг/мл, 16 нг/мл, 17 нг/мл, 18 нг/мл, 19 нг/мл, 20 нг/мл, 30 нг/мл, 40 нг/мл, 50 нг/мл, 60 нг/мл, 70 нг/мл, 80 нг/мл, 90 нг/мл или 100 нг/мл.
Когда bFGF используется на стадии (0-3), его концентрация в среде, предпочтительно, составляет от 1 до 100 нг/мл, и составляет, например, 1 нг/мл, 2 нг/мл, 3 нг/мл, 4 нг/мл, 5 нг/мл, 6 нг/мл, 7 нг/мл, 8 нг/мл, 9 нг/мл, 10 нг/мл, 11 нг/мл, 12 нг/мл, 13 нг/мл, 14 нг/мл, 15 нг/мл, 16 нг/мл, 17 нг/мл, 18 нг/мл, 19 нг/мл, 20 нг/мл, 30 нг/мл, 40 нг/мл, 50 нг/мл, 60 нг/мл, 70 нг/мл, 80 нг/мл, 90 нг/мл или 100 нг/мл. Более предпочтительно, 5 нг/мл.
Период вышеупомянутой стадии (0-3) конкретно не ограничен. Например, он составляет не менее одного дня (например, 1 день, 2 дня, 3 дня, 4 дня, 5 дней, 6 дней, 7 дней, 8 дней, 9 дней, 10 дней, 11 дней, 12 дней, 13 дней, 14 дней, 15 дней, 16 дней, 17 дней, 18 дней, 19 дней, 20 дней, 21 день, 22 дня, 23 дня, 24 дня, 25 дней, 26 дней, 27 дней, 28 дней и более). Поскольку длительное культивирование не влияет на формирование кардиомиоцитов или сердечных клеток-предшественников, верхний предел конкретно не установлен, но обычно он составляет не более 60 дней. Вышеупомянутая стадия (0-3) может повысить эффективность дифференциации в кардиомиоциты или сердечные клетки-предшественники.
Кроме того, эмбриоидные тельца могут быть диссоциированы способом, аналогичным описанному выше, перед вышеупомянутой стадией (0-2) или стадией (0-3).
Способ, конкретно описанный выше, является просто примером, и способ не ограничен вышеупомянутым способом. Например, могут быть упомянуты способ совместного культивирования клеток END2, которые являются поддерживающими клетками, происходящими от мыши, и плюрипотентных стволовых клеток (Mummery, C., et al., Differentiation of human embryonic stem cells to cardiomyocytes: role of coculture with visceral endoderm-like cells. Circulation. 107(21), 2733-40 (2003)), способ индукции кардиомиоцитов путем культивирования эмбриоидных телец с BMP4, FGF2, инсулином и сывороткой (Paul, W B., et al., A Universal System for Highly Efficient Cardiac Differentiation of Human Induced Pluripotent Stem Cells That Eliminates Interline Variability. PLoSone. 6(4), e18293 (2011)) и подобные. Кроме того, также могут быть использованы способ индукции дифференциации кардиомиоцитов без использования цитокина в адгезионной культуре (Lian X, et al., Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling., Proc Natl Acad Sci USA., 2012 July 3; 109(27): E1848-57), способ индукции дифференциации кардиомиоцитов с помощью адгезионной культуры и суспензионной культуры в комбинации без использования цитокина (Minami I, et al., A small molecule that promotes cardiac differentiation of human pluripotent stem cells under defined, cytokine-and xeno-free conditions., Cell Rep., 2012 Nov 29; 2(5): 1448-60) и подобные.
Популяцию клеток с высокой чистотой кардиомиоцитов по сравнению с клеточной популяцией до контакта с ингибитором тирозинкиназы рецепторного типа можно получить путем контакта клеточной популяции, содержащей кардиомиоциты или сердечные клетки-предшественники, полученные, как указано выше, с ингибитором тирозинкиназы рецепторного типа, с последующим культивированием клеточной популяции. То есть, кардиомиоциты могут быть очищены с использованием ингибитора тирозинкиназы рецепторного типа. Следовательно, в другом варианте осуществления настоящего изобретения, предложен способ очистки кардиомиоцитов, включающий (1') стадию контакта ингибитора тирозинкиназы рецепторного типа с клеточной популяцией, содержащей кардиомиоциты или сердечные клетки-предшественники, и не кардиомиоциты, где клеточную популяцию получают путем культивирования плюрипотентных стволовых клеток в среде для дифференциации кардиомиоцитов, и (2') стадию культивирования клеточной популяции (далее также именуемый «способом очистки по настоящему изобретению»).
В настоящем описании, очистка кардиомиоцитов означает, что доля кардиомиоцитов в клеточной популяции (количество кардиомиоцитов в клеточной популяции/общее количество клеток в клеточной популяции) увеличивается из-за того, что скорость снижения числа клеток, не являющихся кардиомиоцитами («количество клеток» означает количество жизнеспособных клеток, в дальнейшем то же самое) превышает скорость снижения кардиомиоцитов, или ингибируется пролиферация не кардиомиоцитов и, таким образом, превышает скорость пролиферации кардиомиоцитов, все из-за ингибитора тирозинкиназы рецепторного типа. Поэтому его отличают от увеличения доли кардиомиоцитов из-за стимулирования индукции дифференциации из сердечных клеток-предшественников в кардиомиоциты или ингибирования индукции дифференциации из сердечных клеток-предшественников в клетки, отличные от кардиомиоцитов. Не желая быть связанными какой-либо теорией, предполагают, что уменьшение количества клеток из-за ингибитора тирозинкиназы рецепторного типа является результатом индукции клеточного апоптоза ингибитором тирозинкиназы рецепторного типа.
На вышеупомянутых стадиях (1) и (1'), период контакта клеточной популяции, содержащей кардиомиоциты или сердечные клетки-предшественники, и ингибитора тирозинкиназы рецепторного типа конкретно не ограничен. Например, он составляет, предпочтительно, не менее 1 часа (например, 2 часов, 3 часов, 5 часов, 12 часов, 1 дня, 2 дней, 3 дней или дольше). Поскольку долговременное культивирование не влияет на формирование кардиомиоцитов или сердечных клеток-предшественников, верхний предел конкретно не устанавливается, но обычно он составляет не более 60 дней (например, 50 дней, 40 дней, 30 дней, 20 дней, 14 дней, 13 дней, 12 дней, 11 дней или меньше). Контакт между клеточной популяцией, содержащей кардиомиоциты или сердечные клетки-предшественники, и ингибитором тирозинкиназы рецепторного типа может быть осуществлен путем добавления ингибитора тирозинкиназы рецепторного типа в среду, содержащую клеточную популяцию, или путем посева клеточной популяции в среду, в которую предварительно добавлен ингибитор тирозинкиназы рецепторного типа. Время контакта с ингибитором тирозинкиназы рецепторного типа конкретно не ограничено, пока клеточная популяция содержит кардиомиоциты или сердечные клетки-предшественники. Например, предпочтительным является контакт на вышеупомянутой стадии (0-2) или (0-3). Основываясь на дате начала индукции дифференциации плюрипотентных стволовых клеток, предпочтительным является контакт через 4 дня или позже (например, через 5 дней, 6 дней, 7 дней или позже) от начала индукции дифференциации.
Примеры тирозинкиназы рецепторного типа, которые ингибируются ингибитором тирозинкиназы рецепторного типа, используемым в настоящем изобретении, включают рецепторы EGF (также обозначаемые как ErbB или HER) (например, ErbB1 (EGFR), ErbB2 (HER2), ErbB3 (HER3), ErbB4 (HER4)), рецепторы инсулина (например, IR-A, IR-B), рецептор инсулиноподобного фактора роста 1, рецепторы VEGF (например, VEGFR-1 (Flt-1), VEGFR-2 (KDR/Flk-1), VEGFR-3 (Flt-4)), рецепторы PDGF (например, PDGFRα, PDGFRβ), рецепторы HGF (также называемые c-Met), рецепторы FGF (например, FGFR1, FGFR2, FGFR3, FGFR4), CCK, рецепторы NGF (также называемые рецептором Trk) (например, TrkA, TrkB, TrkC), рецепторы Eph (эфрина) (например, EPHA1, EPHA2, EPHA3, EPHA4, EPHA5, EPHA6, EPHA7, EPHA8, EPHA9, EPHA10, EPHB1, EPHB2, EPHB3, EPHB4, EPHB5, EPHB6), AXL (также называемый ТАМ рецептором), TIE (например, TIE-1, TIE-2), RYK, DDR (например, DDR1), RET, ROS, LTK, ROR (например, ROR1, ROR2), MuSK и LMR. Среди них, рецепторы VEGF, рецепторы PDGF, рецепторы HGF и рецепторы FGF являются предпочтительными в качестве мишеней для ингибитора тирозинкиназы рецепторного типа. Ингибиторы рецептора EGF исключаются из ингибитора тирозинкиназы рецепторного типа, используемого на вышеупомянутой стадии (1); однако ингибиторы рецептора EGF можно использовать в качестве ингибитора тирозинкиназы рецепторного типа, используемого на вышеупомянутой стадии (1').
В настоящем описании, ингибитор тирозинкиназы рецепторного типа может обладать другими активностями, такими как ингибирующая активность в отношении других тирозинкиназ рецепторного типа, или может обладать ингибирующей активностью, специфичной в отношении одного типа тирозинкиназы рецепторного типа, при условии, что он обладает ингибирующей активностью, по меньшей мере, в отношении одной из вышеупомянутых тирозинкиназ рецепторного типа. Кроме того, ингибитор тирозинкиназы рецепторного типа, отличный от ингибиторов рецептора EGF, не исключает ингибиторы, обладающие ингибирующей активностью в отношении тирозинкиназ рецепторного типа, отличных от рецепторов EGF, и обладающих ингибирующей активностью в отношении рецепторов EGF. В настоящем описании, «ингибитор рецептора EGF» относится к веществу, обладающему ингибирующей активностью в отношении рецептора EGF, превосходящей другие виды активности, и, в частности, относится к веществу, которое ингибирует, по меньшей мере, 90% рецептора EGF или демонстрирует 50% ингибирующую концентрацию рецептора EGF не более 10 мкМ.
Примеры ингибитора рецептора EGF, используемого в настоящем изобретении, включают цетуксимаб, эрлотиниб HCl (OSI-744), гефитиниб (ZD1839), дитозилат лапатиниба (GW-572016), афатиниб (BIBW2992), канертиниб (CI-1033), TAS6417, PD153035, MTX-211, HS-10296, телиатиниб (HMPL-309), лапатиниб, AG-490 (тирфостин B42), CP-724714, дакомитиниб (PF-00299804), WZ4002, сапитиниб (AZD8931), CUDC-101, AG-1478 (тирфостин AG-1478), PD153035 HCl, пелитиниб (EKB-569), AC480 (BMS-599626), AEE788 (NVP-AEE788), AP26113-аналог (ALK-IN-1), OSI-420, WZ3146, HER2-ингибитор-1, WZ8040, AST-1306, роцилетиниб (CO-1686), генистеин, варлитиниб, икотиниб, TAK-285, WHI-P154, дафнетин, PD168393, тирфостин 9, CNX-2006, AG-18, AZ5104, лазертиниб, AZD9291, CL-387785 (EKI-785), олмутиниб (BI 1482694), (-)-эпигаллокатехин галлат, эрлотиниб, гидрохлорид гефитиниба, афатиниб (BIBW2992) дималеат, AZD3759, позиотиниб (HM781-36B), бригатиниб (AP26113), осимертиниба мезилат, накотиниб (ASP8273), хризофановую кислоту, назартиниб (EGF816), норкантаридин, лифирафениб (BGB-283), гидрохлорид лидокаина, бутеин, EAI045, циастерон, авитиниб (AC0010) и подобные.
Примеры ингибитора инсулинового рецептора или рецептора инсулиноподобного фактора роста 1 включают HNMPA, GSK1904529A, довитиниб (TKI-258) димолочную кислоту, довитиниб (TKI258) лактат, NVP-AEW541, довитиниб (TKI-258), NVP-ADW742, линситиниб (OSI-906), GSK1904529A, BMS-754807, TAE226 (NVP-TAE226), церитиниб (LDK378) и подобные.
Примеры ингибитора PDGFRα включают N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1Н-пиразол-5-карбоксамид (соединение формулы (I))
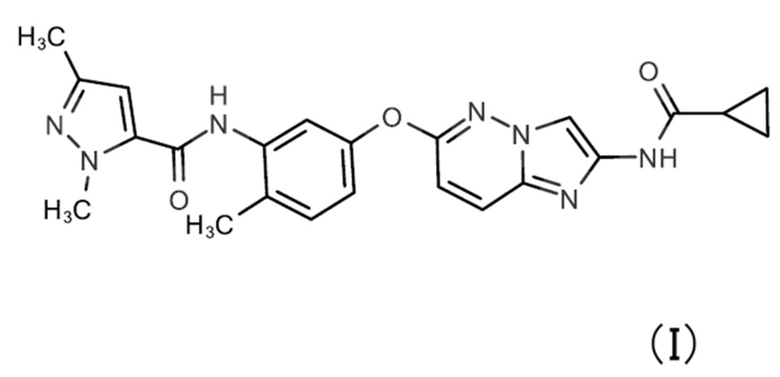 ,
,
понатиниб (АР24534), телатиниб, амуватиниб (МР-470), Ki8751, KRN 633, креноланиб (СР-868596), акситиниб, СР-673451, тивозаниб (AV-951), нинтеданиб (BIBF 1120), довитиниб (TKI-258, CHIR-258), довитиниб (TKI258) лактат, довитиниб (TKI-258) димолочную кислоту, маситиниб (AB1010) и подобные. Примеры ингибитора PDGFRβ включают N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамид, CP-673451, сунитиниб малат, сунитиниб, TSU-68 (SU6668, орантиниб), MK-2461, сорафениба тозилат, линифаниб (ABT-869), акситиниб, креноланиб (CP-868596), довитиниб (TKI-258) димолочную кислоту, довитиниб (TKI-258, CHIR-258), довитиниб (TKI258) лактат, тивозаниб (AV-951), нинтеданиб (BIBF 1120), маситиниб (AB1010), KRN 633 и подобные.
Примеры ингибитора VEGFR-1 включают N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамид, ZM 306416, акситиниб, мотесаниб дифосфат (AMG-706), линифаниб (ABT-869), MGCD-265, цедираниб (AZD2171), форетиниб (GSK1363089), OSI-930, довитиниб (TKI-258, CHIR-258), пазопаниб HCl (GW786034 HCl), пазопаниб, довитиниб (TKI-258) димолочную кислоту, довитиниб (TKI258) лактат, кабозантиниб (XL184, BMS-907351), кабозантиниб малат (XL184), регорафениб (BAY 73-4506), ленватиниб (E7080), тивозаниб (AV-951), нинтеданиб (BIBF 1120), AEE788 (NVP-AEE788), ваталаниб (PTK787) 2HCl, KRN 633, бриваниб (BMS-540215), бриваниба аланинат (BMS-582664) и подобные. Примеры ингибитора VEGFR-2 включают N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамид, N-{4-[(6,7-диметоксихинолин-4-ил)окси]-3-фторфенил}-N'-(4-фторфенил)циклопропан-1,1-дикарбоксамид (соединение формулы (II))

кабозантиниб (XL184, BMS-907351), кабозантиниба малат (XL184), Ki8751, апатиниб, понатиниб (AP24534), ZM323881, LY2874455, сорафениба тозилат, BMS-794833, голватиниб (E7050), RAF265 (CHIR-265), SKLB1002, вандетаниб (ZD6474), CYC116, сунитиниб малат, сунитиниб, PD173074, семаксаниб (SU5416), TSU-68 (SU6668, орантиниб), акситиниб, цедираниб (AZD2171), форетиниб (GSK1363089), MGCD-265, мотесаниб дифосфат (AMG-706), линифаниб (ABT-869), ленватиниб (E7080), регорафениб (BAY 73-4506), телатиниб, тивозаниб (AV-951), OSI-930, довитиниб (TKI-258), димолочную кислоту, нинтеданиб (BIBF 1120), довитиниб (TKI258) лактат, довитиниб (TKI-258, CHIR-258), бриваниба аланинат (BMS-582664), бриваниб (BMS-540215), пазопаниб, пазопаниб HCl (GW786034 HCl), ваталаниб (PTK787) 2HCl, ENMD-2076 L-(+)-винную кислоту, ENMD-2076 L-(+)-винную кислоту, AEE788 (NVP-AEE788), KRN 633 и подобные. Примеры ингибитора VEGFR-3 включают N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамид, SAR131675, акситиниб, форетиниб (GSK1363089), цедираниб (AZD2171), телатиниб, MGCD-265, ленватиниб (E7080), кабозантиниб (XL184, BMS-907351), кабозантиниба малат (XL184), мотесаниба дифосфат (AMG-706), довитиниб (TKI-258, CHIR-258), довитиниб (TKI-258) димолочную кислоту, довитиниб (TKI258) лактат, нинтеданиб (BIBF 1120), тивозаниб (AV-951), ENMD-2076 L-(+)-винную кислоту, ENMD-2076, регорафениб (BAY 73-4506), пазопаниб HCl (GW786034 HCl), пазопаниб, KRN 633, линифаниб (ABT-869), AEE788 (NVP-AEE788), ваталаниб (PTK787)2HCl и подобные.
Примеры ингибитора рецептора HGF включают N-{4-[(6,7-диметоксихинолин-4-ил)окси]-3-фторфенил}-N'-(4-фторфенил)циклопропан-1,1-дикарбоксамид, кризотиниб (PF-02341066), кабозантиниб (BMS-907351), форетиниб (GSK1363089), PHA-665752, SU11274, JNJ-38877618 (OMO-1), глуметиниб (SCC244), альтиратиниб, SAR125844, SGX-523, BMS-777607, тивантиниб (ARQ 197), JNJ-38877605, PF-04217903, аналог MGCD-265, капматиниб (INCB28060), BMS-754807, BMS-794833, AMG-208, MK-2461, голватиниб (E7050), AMG-458, NVP-BVU972, AMG 337, мерестиниб (LY2801653), S49076, пульсатилла сапонин D, норкантаридин, NPS-1034, саволитиниб (AZD6094) и подобные.
Примеры ингибитора FGFR1 включают ASP5878, понатиниб (AP24534), PD173074, данусертиб (PHA-739358), бриваниба аланинат (BMS-582664), бриваниб (BMS-540215), TSU-68 (SU6668, орантиниб), SSR128129E, AZD4547, BGJ398 (NVP-BGJ398), LY2874455, довитиниб (TKI-258, CHIR-258), довитиниб (TKI258) лактат, довитиниб (TKI-258) димолочную кислоту, CH5183284 (Debio-1347), MK-2461, нинтеданиб (BIBF 1120) и подобные. Примеры ингибитора FGFR2 включают ASP5878, BGJ398 (NVP-BGJ398), AZD4547, LY2874455, CH5183284 (Debio-1347), нинтеданиб (BIBF 1120), MK-2461 и подобные. Примеры ингибитора FGFR3 включают ASP5878, BGJ398 (NVP-BGJ398), AZD4547, LY2874455, довитиниб (TKI-258) димолочную кислоту, довитиниб (TKI258) лактат, довитиниб (TKI-258, CHIR-258), CH5183284 (Debio-1347), MK-2461, нинтеданиб (BIBF 1120) и подобные. Примеры ингибитора FGFR4 включают ASP5878, LY2874455, AZD4547, CH5183284 (Debio-1347), нинтеданиб (BIBF 1120) и подобные.
Примеры ингибитора рецептора NGF включают BMS-754807, GW441756, DS-6051b, GNF-5837, CH7057288, альтиратиниб, селитректиниб (LOXO-195), BMS-935177, энтректиниб (RXDX-101), ситраватиниб (MGCD516), PF-06273340, белизатиниб (TSR-011), ларотректиниб (LOXO-101) сульфат и подобные.
Примеры ингибитора Eph включают NVP-BHG712, ситраватиниб (MGCD516) и подобные.
Примеры ингибитора AXL включают BMS-777607, бемцентиниб (R428), кабозантиниба малат (XL184), UNC2250, дуберматиниб (TP-0903), UNC-2025, LDC1267, UNC2881, RXDX-106 (CEP-40783), S49076, ситраватиниб (MGCD516), 2-D08, гилтеритиниб (ASP2215), NPS-1034 и подобные.
Примеры ингибитора TIE включают аналог MGCD-265, ингибитор киназы Tie2, альтиратиниб, пексметиниб (ARRY-614) и подобные.
Кроме того, могут быть надлежащим образом выбраны другие ингибиторы тирозинкиназы рецепторного типа. В качестве ингибитора тирозинкиназы рецепторного типа предпочтительными являются N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамид, N-{4-[(6,7-диметоксихинолин-4-ил)окси]-3-фторфенил}-N'-(4-фторфенил)циклопропан-1,1-дикарбоксамид, AMG337, ASP5878, BGJ398, форетиниб, ZM323881, CP-673451, креноланиб или кризотиниб. Можно использовать только один тип ингибитора тирозинкиназы рецепторного типа или можно использовать несколько видов в комбинации. Для получения кардиомиоцитов с более высокой степенью чистоты, другие лекарственные средства (ингибитор гистондеацетилазы и т. д.) также могут быть использованы одновременно с ингибитором тирозинкиназы рецепторного типа или на стадии до или после.
Вышеуказанный ингибитор может содержать один вид вышеупомянутого соединения или его соли или может содержать два или несколько их видов.
Вышеупомянутое соединение или его соль могут быть получены в соответствии со способом, известным per se.
Когда вышеупомянутое соединение находится в форме соли, соль предпочтительно представляет собой фармакологически приемлемую соль, и примеры такой соли включают соль с неорганическим основанием, соль с органическим основанием, соль с неорганической кислотой, соль с органической кислотой, соль основной или кислой аминокислоты и подобные.
Предпочтительные примеры соли с неорганическим основанием включают соли щелочных металлов, такие как соль натрия, соль калия и подобные, соли щелочноземельного металла, такие как соль кальция, соль магния и подобные, соль алюминия, соль аммония и подобные.
Предпочтительные примеры соли с органическим основанием включают соли с триметиламином, триэтиламином, пиридином, пиколином, этаноламином, диэтаноламином, триэтаноламином, трометамином [трис(гидроксиметил)метиламином], трет-бутиламином, циклогексиламином, бензиламином, дициклогексиламином, N,N-дибензилэтилендиамином или подобным.
Предпочтительные примеры соли с неорганической кислотой включают соль с хлористоводородной кислотой, бромистоводородной кислотой, азотной кислотой, серной кислотой, фосфорной кислотой и подобными.
Предпочтительные примеры соли органической кислоты включают соли муравьиной кислоты, уксусной кислоты, трифторуксусной кислоты, фталевой кислоты, фумаровой кислоты, щавелевой кислоты, винной кислоты, малеиновой кислоты, лимонной кислоты, янтарной кислоты, яблочной кислоты, метансульфоновой кислоты, бензолсульфоновой кислоты, п-толуолсульфоновой кислоты и подобные.
Предпочтительные примеры соли с основной аминокислотой включают соль с аргинином, лизином, орнитином и подобными.
Предпочтительные примеры соли с кислой аминокислотой включают соль с аспарагиновой кислотой, глутаминовой кислотой и подобными.
Вышеупомянутые соединения могут быть гидратами, не гидратами, сольватами или не сольватами.
Кроме того, вышеупомянутые соединения могут быть помечены изотопом или замещены им (например, 2H, 3H, 11C, 14C, 18F, 35S, 125I, и подобные) или подобные.
Формы превращения дейтерия, в которых 1H превращается в 2H (D), также охватываются вышеупомянутыми соединениями.
Таутомеры также охватываются вышеупомянутыми соединениями.
Вышеупомянутые соединения могут представлять собой фармацевтически приемлемые сокристаллы или соли сокристаллов. Сокристалл или соль сокристалла означает кристаллическое вещество, состоящее из двух или нескольких особых твердых веществ при комнатной температуре, каждое из которых имеет различные физические свойства (например, структуру, точку плавления, теплоту плавления, гигроскопичность, растворимость и стабильность). Сокристалл или соль сокристалла может быть получена способом сокристаллизации, известным per se.
Специалист в данной области техники может соответствующим образом выбрать концентрацию ингибитора тирозинкиназы рецепторного типа в среде. Например, предпочтительно она составляет от 1 нМ до 10 мкМ, особенно, более предпочтительно, от 10 нМ до 3 мкМ, и конкретно составляет 1 нМ, 2 нМ, 3 нМ, 5 нМ, 10 нМ, 20 нМ, 30 нМ, 40 нМ, 50 нМ, 0,1 мкМ, 0,2 мкМ, 0,3 мкМ, 0,4 мкМ, 0,5 мкМ, 1,0 мкМ, 1,5 мкМ, 2 мкМ, 3 мкМ, 4 мкМ, 5 мкМ, 6 мкМ, 7 мкМ, 8 мкМ, 9 мкМ или 10 мкМ. Концентрация также может быть изменена в соответствии с типом соединения. Например, когда используют N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамид, она составляет, предпочтительно, от 20 нМ до 10 мкМ, когда используют N-{4-[(6,7-диметоксихинолин-4-ил)окси]-3-фторфенил}-N'-(4-фторфенил)циклопропан-1,1-дикарбоксамид, она составляет, предпочтительно, от 0,2 мкМ до 10 мкМ, когда используют AMG337, она составляет, предпочтительно, от 0,2 мкМ до 10 мкМ, когда используют ASP5878, она составляет, предпочтительно, от 1 нМ до 1 мкМ, когда используют BGJ398, она составляет, предпочтительно от 5 нМ до 5 мкМ, когда используют форетиниб, она составляет, предпочтительно, от 5 нМ до 5 мкМ, когда используют ZM323881, она составляет, предпочтительно, от 0,1 мкМ до 10 мкМ, когда используют CP-673451, она составляет, предпочтительно, от 0,1 мкМ до 10 мкМ, когда используют креноланиб, она составляет, предпочтительно от 0,1 мкМ до 10 мкМ, и когда используют кризотиниб, она составляет, предпочтительно от 0,1 мкМ до 10 мкМ. Концентрация не ограничивается этими концентрациями.
Способ культивирования клеточной популяции на вышеупомянутых стадиях (2) и (2') является таким же, как и на вышеупомянутых (0-2) или (0-3). Период культивирования такой же, и культивирование можно продолжать, по меньшей мере, пока клеточная популяция находится в контакте с ингибитором тирозинкиназы рецепторного типа.
2. Популяция клеток, содержащая кардиомиоциты.
Настоящее изобретение также относится к клеточной популяции, содержащей кардиомиоциты (далее также называемой «клеточной популяцией по настоящему изобретению»), полученной способом получения или очистки по настоящему изобретению. Как описано выше, клеточная популяция по настоящему изобретению содержит кардиомиоциты высокой чистоты. Высокая чистота означает, что соотношение кардиомиоцитов в клеточной популяции (количество кардиомиоцитов в клеточной популяции/общее количество клеток в клеточной популяции) конкретно составляет не менее 80% (например, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или выше). Когда вышеупомянутая клеточная популяция и другие клетки или клеточные популяции, такие как мезенхимальные стволовые клетки и подобные, смешивают и используют, соотношение вышеупомянутых кардиомиоцитов, естественно, такое же, как до смешивания с другими клетками или клеточными популяциями. В предпочтительном варианте осуществления, клеточная популяция по настоящему изобретению содержит кардиомиоциты в более высокой доле, чем клеточная популяция, полученная обычным способом индукции кардиомиоцитов из плюрипотентных стволовых клеток. Такие клеточные популяции могут быть дополнительно очищены путем сортировки клеток и подобным, и такая очищенная клеточная популяция также охватывается «клеточной популяцией по настоящему изобретению».
3. Агент для терапии на основе трансплантации клеток
Настоящее изобретение также относится к агенту для терапии на основе трансплантации клеток, содержащему клеточную популяцию по настоящему изобретению (далее также упоминаемому как «терапевтический агент для клеточной трансплантации по настоящему изобретению»). Терапевтический агент для клеточной трансплантации по настоящему изобретению может быть использован для аутологичной трансплантации или аллотрансплантации. Кроме того, его можно использовать в сочетании с другими лекарствами, такими как иммунодепрессанты. Как упоминалось выше, поскольку клеточная популяция по настоящему изобретению содержит кардиомиоциты высокой чистоты, ее целесообразно использовать в качестве исходного материала для агента для терапии на основе трансплантации клеток, и клеточная популяция по настоящему изобретению и агент для терапии на основе трансплантации клеток по изобретению полезны для лечения или профилактики сердечных заболеваний. Таким образом, способ лечения или профилактики сердечных заболеваний, включающий введение или трансплантацию эффективного количества клеточной популяции или агента для терапии на основе трансплантации клеток по настоящему изобретению млекопитающему (например, человеку, мыши, крысе, обезьяне, быку, лошади, свиньи, собаки и т.д.) в качестве мишени лечения или профилактики также включены в настоящее изобретение. Примеры сердечного заболевания, которое является мишенью лечения или профилактики, включают, но не ограничены ими, такие заболевания, как сердечная недостаточность, ишемическая болезнь сердца, инфаркт миокарда, кардиомиопатия, миокардит, гипертрофическая кардиомиопатия, дилатационная фаза гипертрофической кардиомиопатии, дилатационная кардиомиопатия, и подобные, дефицит из-за нарушения, и подобные.
С точки зрения того аспекта, что отторжения не происходит, когда клеточная популяция по настоящему изобретению используется в качестве агента для терапии на основе трансплантации клеток, желательно использовать клеточную популяцию, содержащую клетки, полученные из iPS клеток, полученных из соматических клеток, которые имеют тот же или практически тот же генотип HLA, что и у индивидуума, которому проводится трансплантация. Используемый здесь термин «по существу одинаковый» означает, что генотипы HLA совпадают до такой степени, что иммунодепрессант может подавлять иммунный ответ на трансплантированные клетки. Например, это относится к соматической клетке с типом HLA, в котором совпадают 3 локуса HLA-A, HLA-B и HLA-DR или 4 локуса, включая HLA-C. Кроме того, клетки также можно поместить в капсулу, такую как полиэтиленгликоль или силикон, или в пористый контейнер и т. д., и трансплантировать клетки, избегая отторжения.
Популяцию клеток по настоящему изобретению можно получить в виде парентерального состава, такого как инъекция, суспензия, капельное вливание или подобные, в смеси с фармацевтически приемлемым носителем, обычными способами. Примеры фармацевтически приемлемого носителя, который может содержаться в парентеральном препарате, включают водные жидкости для инъекций, такие как изотонический раствор, содержащий физиологический раствор, глюкозу и другие вспомогательные лекарственные средства (например, D-сорбит, D-маннит, хлорид натрия и подобные). Агент для терапии на основе трансплантации клеток по настоящему изобретению может быть составлен, например, с буферным агентом (например, буферным раствором фосфата, буферным раствором ацетата натрия), смягчающим агентом (например, хлоридом бензалкония, гидрохлоридом прокаина и подобными), стабилизатором (например, сывороточным альбумином человека, полиэтиленгликолем и подобными), консервантом, антиоксидантом и подобными. Когда агент для терапии на основе трансплантации по настоящему изобретению составлен в виде водной суспензии, клеточная популяция, содержащая кардиомиоциты, может быть суспендирована в указанном выше водном растворе в концентрации от примерно 1×106 до примерно 1×108 клеток/мл. Его также можно вводить с трехмерной подложкой-носителем, который способствует приживлению. Примеры трехмерной подложки-носителя включают, но не ограничены ими, компоненты биологического происхождения, такие как коллаген и подобные, и синтетические полимеры, такие как полимолочная кислота и подобные, которые заменяют компоненты.
Альтернативно, сердечные заболевания можно лечить путем приклеивания пластины, сформированной из полученных кардиомиоцитов, к сердцу пациентов. Когда вводят пластину сердечной мышцы, пластину располагают таким образом, чтобы покрыть желаемую область. В данном случае, нанесение покрытия на желаемую область может быть выполнено с использованием методики, хорошо известной в соответствующей области техники. Во время размещения, когда требуемая площадь велика, пластину также можно поместить так, чтобы она окружала ткань. Для достижения желаемого эффекта, введение может включать несколько раз нанесения на одну и ту же область. Когда нанесение проводят несколько раз, желательно обеспечить достаточное время для того, чтобы нужные клетки прижились в ткани, чтобы обеспечить ангиогенез. Механизмом такого лечения сердечных заболеваний может быть эффект, возникающий в результате приживления пластины сердечной мышцы, или косвенное действие, не основанное на приживлении клеток (например, эффект за счет рекрутирования клеток реципиента в поврежденный участок, вызванный секрецией аттрактанта). Когда пластина сердечной мышцы используется для лечения сердечных заболеваний, она может содержать трехмерную подложку-носитель для клеток (каркас), такой как коллаген, фибронектин, ламинин или подобные, в дополнение к кардиомиоцитам. Альтернативно, он может содержать клетки любого типа (возможно, во множестве) в дополнение к кардиомиоцитам. Количество кардиомиоцитов, используемых для лечения сердечных заболеваний, конкретно не ограничивается, пока подлежащий введению сердечный мышечный слой проявляет эффект при лечении сердечных заболеваний, и может быть соответствующим образом увеличен или уменьшен для корректировки, в зависимости от размера пораженной части и размера тела.
В еще одном варианте осуществления, клеточная популяция по настоящему изобретению также может быть использована для скрининга лекарственных средств или оценки кардиотоксичности лекарств для лечения сердечного заболевания. Например, путем введения тестируемого лекарственного средства в клеточную популяцию по настоящему изобретению и последующего измерения реакции кардиомиоцитов, можно оценить эффект и токсичность тестируемого лекарственного средства.
Хотя настоящее изобретение дополнительно поясняется конкретно в следующих примерах, объем настоящего изобретения не ограничивается такими примерами.
[Пример]
Пример 1. Анализ секвенирования одноклеточной РНК клеток, индуцированных к дифференциации в кардиомиоциты из iPS клеток.
Используют штамм для оценки клинической линии iPS клеток, произведенный Центром исследований и применения iPS клеток Киотского университета (CiRA). Поддерживающую культуру линии iPS клеток проводят в соответствии с общепринятым способом (Okita K, et al. Stem Cells. 2012 Nov 29. doi: 10.1002/stem.1293). Индукцию дифференциации в кардиомиоциты проводят по способу, описанному в статье (Miki et al., Cell Stem Cell. 2015 Jun 4; 16. doi: 10.1016/j.stem. 2015.04.005.). Кардиомиоциты через 22 дня после начала индукции дифференциации подвергают анализу последовательности РНК одиночных клеток с использованием Chromium производства 10X Genomics. Программное обеспечение (Cellranger, Seurat, Loupe Cell Browser) используют для кластеризации клеток и анализа каждого кластера. Было обнаружено, что клеточные популяции, индуцированные к дифференциации из iPS клеток в кардиомиоциты, содержат, помимо популяции кардиомиоцитов, экспрессирующих саркомерный-α-актинин и сердечный тропонин Т, три популяции не кардиомиоцитов, которые не экспрессируют саркомерный-α-актинин или сердечный тропонин T (считаются клетками, подобными гладким мышцам (SMC), клетками, подобными эндотелию (EC), и клетками эндодермальной линии (END) из экспрессии генов) (фиг. 1).
Пример 2. Анализ не кардиомиоцитов, присутствующих в клеточной популяции, индуцированной для дифференциации в миокард из iPS клеток
Экспрессию генов рецепторного типа тирозинкиназы анализируют с помощью анализа Loupe Cell Browser данных секвенирования одноклеточной РНК. В результате было установлено, что экспрессия генов тирозинкиназы рецепторного типа, таких как PDGFRA, PDGFRB, VEGFR1, VEGFR2, c-Met (HGFR), FGFR4 и подобные, является высокой в популяциях не кардиомиоцитов, но невысокой в популяциях кардиомиоцитов (фиг. 2).
Пример 3. Окрашивание не кардиомиоцитов маркерами клеточной поверхности
Линию iPS клеток человека с двойным нокином получают путем вставки последовательности репортерного белка EmGFP (SEQ ID NO: 1) в локус гена TNNI1, и mCherry (SEQ ID NO: 2) в локус гена TNNI3. Линию iPS клеток человека получают с использованием PBMC (LP_167, ID образца: 20130318), приобретенных у CTL, и эписомального вектора (загруженный ген; OCT3/4, KLF4, SOX2, L-MYC, LIN28, p53DD мыши) (справочный документ; Okita K, et al. Stem Cells. 2012 Nov 29. doi: 10.1002/stem.1293).
Поддерживающую культуру линии iPS клеток получают в соответствии с общепринятым способом (Okita K, et al. Stem Cells. 2012 Nov 29. doi: 10.1002/stem.1293).
Индукцию дифференциации в кардиомиоциты проводят по способу, описанному в статье (Miki et al, Cell Stem Cell. 2015 Jun 4; 16. doi: 10.1016/j.stem. 2015.04.005.), и с использованием 6-луночного планшета. Для краткого пояснения, индукцию дифференциации в кардиомиоциты проводят путем обработки линии репортерных iPS клеток в течение 4-5 минут с помощью TrypLE select (Life Technologies), разведенного до 1/2 0,5 мМ ЭДТК/PBS, с отделением клеток клеточным скребком (IWAKI) и диссоциацией клеток на отдельные клетки с помощью пипетирования. Среду удаляют центрифугированием при 1000 об/мин в течение 5 мин, полученные клетки высевают в количестве 2×106 клеток на лунку 6-луночного планшета и культивируют в среде StemPro34 с добавлением 1% L-глутамина, трансферрина 150 мкг/мл, аскорбиновой кислоты 50 мкг/мл (сигма), монотиоглицерина 4×10-4 М, 10 мкМ Y-27632, 2 нг/мл BMP4 (R&D) и 0,5% фактора роста восстановленного матригеля, 1,5 мл на лунку, в условиях 37°C, 5% кислорода для образования эмбриоидных телец (день 0).
На следующий день (день 1) 1,5 мл среды StemPro34 с добавлением 1% L-глутамина, трансферрина 150 мкг/мл, аскорбиновой кислоты 50 мкг/мл (сигма), монотиоглицерина 4×10-4 М, 2 нг/мл BMP4 (R&D) активина А 12 нг/мл, bFGF 5 нг/мл и ВМР4 18 нг/мл добавляют в каждую лунку, и клетки культивируют при 37°C, 5% кислороде в течение еще двух дней.
Затем (3 день) 6-луночный планшет выдерживают под наклоном, чтобы дать возможность эмбриоидным тельцам образовать осадок, удаляют от 80 до 90% среды и в каждую лунку добавляют IMDM (1,5 мл). Планшет с лунками выдерживают, снова под наклоном, чтобы эмбриоидные тельца образовали осадок, удаляют 80-90% среды и клетки культивируют в среде StemPro34 с добавлением 1% L-глутамина, трансферрина 150 мкг/мл, аскорбиновой кислоты 50 мкг/мл (сигма), монотиоглицерина 4×10-4 М, 10 нг/мл VEGF, 1 мкМ IWP-3, 0,6 мкМ дорсоморфина и 5,4 мкМ SB431542, в условиях 37°С, 5% кислорода в течение 3 дней.
Затем (6 день) 6-луночный планшет выдерживают под наклоном, чтобы позволить эмбриоидным тельцам образовать осадок, удаляют от 80 до 90% среды и добавляют среду StemPro34 с добавлением 1% L-глутамина, трансферрина 150 мкг/мл, аскорбиновой кислоты 50 мкг/мл (сигма), монотиоглицерина 4×10-4 М и 10 нг/мл VEGF. Клетки культивируют в течение 8 дней в условиях 37°С, 5% кислорода. В течение этого периода, среду заменяют средой в тех же условиях один раз в 2-3 дня.
На 15 день индуцирования дифференциации, культуральную среду, содержащую эмбриоидные тельца, центрифугируют при 200 g в течение 1 мин и удаляют супернатант аспиратором. Добавляют PBS, смесь центрифугируют при 200×g в течение 1 мин, и надосадочную жидкость удаляют аспиратором. В каждую пробирку добавляют раствор (3 мл), полученный добавлением 10 мкг/мл ДНКазы и 100 мкг/мл либеразы к IMDM (модифицированная по Искову среда Дульбекко), и смесь выдерживают при 37°C в обычных кислородных условиях в течение 1 часа. Через 1 ч, пробирку центрифугируют при 400 g в течение 5 мин и удаляют супернатант, избегая всасывания эмбриоидных телец. В каждую пробирку добавляют раствор (2 мл), полученный добавлением ДНКазы 10 мкг/мл к аккутазе (Thermo), и смесь выдерживают при 37°С в обычных кислородных условиях в течение 10 мин. После отстаивания, отдельные клетки получают пипетированием, в каждую пробирку добавляют среду (2 мл), полученную добавлением ДНКазы 10 мкг/мл к IMDM, и смесь перемешивают переворачиванием с получением суспензии отдельных клеток.
Вышеупомянутую суспензию отдельных клеток центрифугируют при 200xg в течение 5 минут, удаляют супернатант и клетки подвергают криоконсервации при -80°C.
Криоконсервированные клетки размораживают в теплой ванне при 37°С, центрифугируют при 400xg в течение 5 мин и удаляют супернатант.
После постукивания клеточного дебриса, добавляют 1 мл PBS, содержащего 1% BSA, и смесь центрифугируют при 300×g в течение 3 мин. Супернатант удаляют. Клетки окрашивают меченным APCFire750 анти-CD326 антителом, меченным PE анти-CD49a антителом, меченым BV605 анти-CD31 антителом и DAPI в PBS, содержащем 1% BSA. DAPI-положительные мертвые клетки удаляют, и разделение клеточной популяции и соотношение каждой клеточной популяции измеряют путем измерения количества сигнала каждого флуоресцентного красителя в ядросодержащих клетках.
Кардиомиоциты, полученные путем индукции дифференциации вышеупомянутых репортерных iPS клеток в сердечную мышцу, разделяют на 4 основные клеточные популяции: CD326-положительные клетки, CD326-отрицательные CD31-положительные клетки, CD326-отрицательные CD31-отрицательные CD49a-положительные клетки и CD326-отрицательные CD31-отрицательные CD49a-отрицательные клетки из-за различий в экспрессии этих трех поверхностных маркеров. Четыре популяции клеток состоят из трех популяций, в основном состоящих из не кардиомиоцитов, экспрессирующих один или несколько из этих трех поверхностных маркеров (популяций, содержащих много клеток, не экспрессирующих репортерные гены сердечной мышцы, и популяции кардиомиоцитов, отрицательная по всем трем маркерам (популяции клеток, экспрессирующей репортерные гены сердечной мышцы). Не менее 99% клеточной популяции, отрицательной по всем трем маркерам, составляют кардиомиоциты (клетки, экспрессирующие репортерный ген сердечной мышцы) (фиг. 3).
Затем, принимая во внимание тип рецепторной тирозинкиназы, обнаруженной в примере 2, то могут ли или нет кардиомиоциты с относительно низкой экспрессией тирозинкиназы рецепторного типа, быть очищены с использованием ингибитора тирозинкиназы рецепторного типа, проверяют в различных условиях (экспериментальные примеры 1-9).
В следующих экспериментальных примерах 1-9, соединение А представляет собой N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамид, и соединение B представляет собой N-{4-[(6,7-диметоксихинолин-4-ил)окси]-3-фторфенил}-N'-(4-фторфенил)циклопропан-1,1-дикарбоксамид.
Экспериментальный пример 1. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
В качестве линии клеток iPS человека используют клеточную линию (исходная клеточная линия репортерных клеток, используемая для окрашивания не кардиомиоцитов маркером клеточной поверхности), полученную с использованием РВМС (LP_167, ID образца: 20130318), приобретенных у CTL, и эписомального вектора (нагруженный ген; OCT3/4, KLF4, SOX2, L-MYC, LIN28, p53DD мыши) (справочный документ; Okita K, et al. Stem Cells. 2012 Nov 29. doi: 10.1002/stem.1293).
Поддерживающую культуру линии iPS клеток получают в соответствии с общепринятым способом (Okita K, et al. Stem Cells. 2012 Nov 29. doi: 10.1002/stem.1293). Индукцию дифференциации в кардиомиоциты проводят по способу, описанному в статье (Miki et al., Cell Stem Cell. 2015 Jun 4; 16. doi: 10.1016/j.stem. 2015.04.005.), и с использованием 6-луночного планшета. VEGF не добавляют после 8 дня.
На 8 день индукции дифференциации, эмбриоидные тельца собирают в одну центрифужную пробирку, оставляют на несколько минут при комнатной температуре, чтобы вызвать осаждение эмбриоидных телец. Супернатант удаляют аспиратором, и вместо среды добавляют среду. Суспензию эмбриоидных телец переносят в 6-луночный планшет и добавляют 1/100 количества оцениваемого соединения (соединения и концентрации экспериментов №№ 2-8 показаны в таблице 1 ниже) в концентрации, в 100 раз превышающей конечную концентрацию в каждой лунке. Планшет перемешивают для получения культуры клеток.
[Таблица 1]
На 10 день индукции дифференциации, 6-луночный планшет выдерживают в течение нескольких минут под наклоном, чтобы позволить эмбриоидным тельцам образовать осадок. Супернатант удаляют аспиратором и добавляют среду для замены среды в каждую лунку. Как и в день 8, добавляют 1/100 количества исследуемого соединения в концентрации, в 100 раз превышающей конечную концентрацию, посредством чего добавляют соединение. На 13 день индукции дифференциации, 6-луночный планшет наклоняют, чтобы позволить эмбриоидным тельцам образовать осадок, и эмбриоидные тельца переносят в 1,5 мл пробирку с помощью пипетки, снабженной чипом для 1 мл пипетки с широким диаметром. Диссоциацию на отдельные клетки проводят в соответствии с окрашиванием не кардиомиоцитов маркером клеточной поверхности, и суспензию отдельных клеток используют для измерения доли клеток, положительных по маркеру кардиомиоцитов, и измерения доли клеток, положительных по маркеру не кардиомиоцитов.
Измерение скорости клеток с положительным маркером сердечной мышцы (доля кардиомиоцитов)
Вышеупомянутую суспензию отдельных клеток помещают в пробирку объемом 1,5 мл, центрифугируют при 400 g в течение 3 мин и удаляют надосадочную жидкость. Осадки клеток ресуспендируют в растворе Cytofix/Cytoperm Fixation/Permeabilization Solution (BD) и фиксируют, оставляя при комнатной температуре на 15 минут. В каждую пробирку добавляют 1xPerm/промывочный буфер (1 мл), смесь центрифугируют при 1500 g в течение 3 мин и супернатант удаляют для промывания клеток. После постукивания клеточных дебрисов добавляют 1xPerm/промывочный буфер (1 мл) и аналогичную операцию выполняют для промывания клеток. После постукивания клеточных дебрисов, их окрашивают флуоресцентным красителем Alexa647 с использованием анти-саркомерного α-актининового антитела в 1xPerm/промывочном буфере, содержащем 1% BSA, а затем проводят окрашивание ДНК с использованием DAPI (дигидрохлорида 4',6-диамидино-2-фенилиндола). Долю положительных по саркомерносу α-актинину клеток анализируют путем измерения количества флуоресцентного сигнала Alexa647 в клеточной популяции, исключая мертвые клетки (клетки с количеством ДНК Sub-G1), с использованием проточного цитометра. В результате, доля кардиомиоцитов (доля актинин-положительных клеток) увеличивается при обработке соединением А (N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамидом), AMG337, ASP5878 или BGJ398 по сравнению с отсутствием обработки соединением (фиг. 4).
Измерение доли клеток, положительных по маркеру не кардиомиоцитов (доли не кардиомиоцитов)
Используя незамороженную суспензию отдельных клеток, окрашивание, измерение и анализ проводят в соответствии с окрашиванием не кардиомиоцитов маркером клеточной поверхности. В результате, доля не кардиомиоцитов (количество клеток, положительных по любому из CD326, CD31, CD49a) снижается при обработке соединением A, AMG337, ASP5878 или BGJ398 по сравнению с отсутствием обработки соединением (фиг. 5).
Экспериментальный пример 2. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
В качестве iPS клеток человека используют ту же клеточную линию, что и в экспериментальном примере 1, и индукцию дифференциации в кардиомиоциты также проводят аналогичным способом. Среду меняют на 8, 10 и 13 день индукции дифференциации, и это осуществляют способом, аналогичным способу в экспериментальном примере 1, на 8 и 10 день индукции дифференциации, и проводят способом, аналогичным 10 дню в экспериментальном примере 1, на 13 день индукции дифференциации. На 8, 10 и 13 день индукции дифференциации добавляют оцениваемые соединения (соединения и концентрации экспериментов №№ 2, 3 показаны в таблице 2 ниже) способом, аналогичным способу, описанному в экспериментальном примере 1. ASP5878 в эксперименте номер 3 добавляют только на 8 и 10 день индукции дифференциации. Соответствующие эксперименты проводят с использованием 4 лунок каждый.
[Таблица 2]
На 16 день индукции дифференциации проводят диссоциацию на отдельные клетки, фиксацию и измерение доли кардиомиоцитов (доли положительных по саркомерному α-актинину клеток) способами, аналогичными тем, что описаны в экспериментальном примере 1. В результате, доля кардиомиоцитов увеличивается при обработке форетинибом или ASP5878 по сравнению с отсутствием обработки соединением (фиг. 6).
Экспериментальный пример 3. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
В качестве линии клеток iPS человека используют линию клеток, полученную с использованием PBMC (LP_140, идентификатор образца: 20120808), приобретенных у CTL, и эписомального вектора (нагруженный ген; OCT3/4, KLF4, SOX2, L-MYC, LIN28, p53DD мыши) (ссылочный документ; Okita K, et al. Stem Cells. 2012 Nov 29. doi: 10.1002/stem.1293).
Поддерживающую культуру линии iPS-клеток получают в соответствии с общепринятым способом (Okita K, et al. Stem Cells. 2012 Nov 29. doi: 10.1002/stem.1293). Индукцию дифференциации в кардиомиоциты проводят по методике, аналогичной методике в примере 1. VEGF не добавляют после 10 дня.
Среду меняют на 8, 10 и 13 день индукции дифференциации, и это осуществляют способом, аналогичным способу в экспериментальном примере 1, на 8 и 10 день индукции дифференциации, и проводят способом, аналогичным 10 дню в экспериментальном примере 1 на 13 день индукции дифференциации. На 8, 10 и 13 день индукции дифференциации, оценочные соединения (соединения и концентрации экспериментов №№ 2-6, показаны в таблице 3 ниже) добавляют способом, аналогичным способу, описанному в экспериментальном примере 1.
[Таблица 3]
На 17 день индукции дифференциации проводят измерение диссоциации на отдельные клетки и доли не кардиомиоцитов (количества клеток, положительных по любому из CD326, CD31, CD49a) способами, аналогичными тем, что описаны в экспериментальном примере 1. В результате доля не-кардиомиоцитов снижается при обработке соединением А, форетинибом, ZM323881, креноланибом или кризотинибом по сравнению с отсутствием обработки соединением (фиг. 7).
Экспериментальный пример 4. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
В качестве iPS клеток человека используют ту же клеточную линию, что и в экспериментальном примере 3, и индукцию дифференциации в кардиомиоциты также проводят аналогичным способом.
Среду меняют на 8, 10 и 13 день индукции дифференциации, и это осуществляют способом, аналогичным способу в экспериментальном примере 1, на 8 и 10 день индукции дифференциации, и проводят способом, аналогичным 10 дню в экспериментальном примере 1 на 13 день индукции дифференциации. На 8, 10 и 13 день индукции дифференциации добавляют оцениваемые соединения (соединения и концентрации экспериментов №№ 2, 3, показаны в таблице 4 ниже) способом, аналогичным способу, описанному в экспериментальном примере 1. Соответствующие эксперименты проводят с использованием 4 лунок каждый.
[Таблица 4]
На 17 день индукции дифференциации проводят диссоциацию на отдельные клетки, измерение количества клеток и измерение доли кардиомиоцитов. Измерение фиксации и доли кардиомиоцитов (доли положительных по саркомерному α-актинину клеток) проводят способами, аналогичными тем, которые описаны в экспериментальном примере 1. В результате, доля кардиомиоцитов повышается при обработке соединением А и форетинибом по сравнению с отсутствием обработки (фиг. 8).
Вышеупомянутую суспензию отдельных клеток разводят в 5 раз добавлением к среде IMDM, приготовленной в 1,5 мл пробирке, и перемешивают переворачиванием, и количество клеток измеряют с помощью счетчика клеток NC-200 (Chemometec). В результате, примерно 70% клеток восстановлено при обработке форетинибом и почти такое же количество клеток восстановлено при обработке соединением А по сравнению с отсутствием обработки (фиг. 9).
Экспериментальный пример 5. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
В качестве iPS клеток человека используют ту же клеточную линию, что и в экспериментальном примере 3, и индукцию дифференциации в кардиомиоциты также проводят аналогичным способом.
Среду меняют на 8, 10 и 13 день индукции дифференциации, и это осуществляют способом, аналогичным способу в экспериментальном примере 1, на 8 и 10 день индукции дифференциации, и проводят способом, аналогичным 10 дню в экспериментальном примере 1 на 13 день индукции дифференциации. На 8, 10 и 13 день индукции дифференциации добавляют оцениваемые соединения (соединения и концентрации экспериментов №№ 2-4 показаны в таблице 5 ниже) способом, аналогичным способу, описанному в экспериментальном примере 1. ASP5878 в эксперименте номер 4 добавляют только на 8 и 10 день индукции дифференциации. Соответствующие эксперименты проводят с использованием 4 лунок в каждом.
[Таблица 5]
На 16 день индукции дифференциации проводят диссоциацию на отдельные клетки, фиксацию и долю кардиомиоцитов (долю положительных по саркомерному α-актинину клеток) способами, аналогичными тем, что описаны в экспериментальном примере 1. В результате, доля кардиомиоцитов (долю положительных по саркомерному α- актинину клеток) увеличивается при обработке креноланибом, соединением B (N-{4-[(6,7-диметоксихинолин-4-ил)окси]-3-фторфенил}-N'-(4-фторфенил)циклопропан-1,1-дикарбоксамид), или ASP5878, по сравнению с отсутствием обработки соединением (фиг. 10).
Используя вышеупомянутую суспензию одиночных клеток, количество клеток измеряют способом, описанным в экспериментальном примере 4. В результате, количество восстановленных клеток не сильно меняется в зависимости от присутствия или отсутствия обработки соединением (фиг. 11).
Экспериментальный пример 6. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
Используют клиническую линию iPS клеток, произведенную CiRA. Поддерживающую культуру линии iPS клеток проводят в соответствии с общепринятым способом (Okita K, et al. Stem Cells. 2012 Nov 29. doi: 10.1002/stem.1293). Индукцию дифференциации в кардиомиоциты проводят по способу, описанному в статье (Miki et al, Cell Stem Cell. 2015 Jun 4; 16. doi: 10.1016/j.stem. 2015.04.005.).
Среду заменяют на 8, 10, 14, 17 и 21 день индукции дифференциации, и это осуществляют способом, аналогичным способу, описанному в экспериментальном примере 1, на 8 и 10 день индукции дифференциации, и проводят с помощью способа, аналогичного тому, который используют для 10 дня в экспериментальном примере 1, на 14 день и после индукции дифференциации.
На 14, 17 и 21 день индукции дифференциации добавляют соединение А, которое является одним из оцениваемых соединений, способом, подобным способу, описанному в экспериментальном примере 1. Соответствующие эксперименты проводят с использованием 4 лунок в каждом.
[Таблица 6]
Диссоциацию на отдельные клетки, фиксацию, измерение скорости кардиомиоцитов и измерение скорости не кардиомиоцитов проводят способами, аналогичными тем, что описаны в экспериментальном примере 1, и измерение количества клеток проводят способом, аналогичным тому, который используют в экспериментальном примере 4. В результате, доля кардиомиоцитов увеличивается (фиг. 12), и доля не кардиомиоцитов снижается (фиг. 13) при обработке соединением А по сравнению с отсутствием обработки соединением. Количество восстановленных клеток не сильно меняется в зависимости от наличия или отсутствия обработки соединением (фиг. 14).
Экспериментальный пример 7. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
В качестве iPS клеток человека используют клиническую линию iPS клеток, продуцируемую CiRA. С использованием той же клеточной линии, что и в экспериментальном примере 6, индукцию дифференциации в кардиомиоциты также проводят аналогичным способом.
Среду меняют на 8, 10 и 13 день индукции дифференциации, и это осуществляют способом, аналогичным способу в экспериментальном примере 1, на 8 и 10 день индукции дифференциации, и проводят способом, аналогичным 10 дню в экспериментальном примере 1 на 13 день индукции дифференциации. На 8, 10 и 13 день индукции дифференциации, оценочные соединения (соединения и концентрации экспериментов №№ 2-9 показаны в таблице 7 ниже) добавляют способом, подобным способу, описанному в экспериментальном примере 1 (две лунки используют для эксперимента №1, и одну лунку используют для других).
[Таблица 7]
На 17 день индукции дифференциации проводят диссоциацию на отдельные клетки, фиксацию, измерение доли кардиомиоцитов (доли положительных по саркомерному α-актинину клеток) и измерение доли не кардиомиоцитов способами, аналогичными тем, что описаны в экспериментальном примере 1. Доля кардиомиоцитов увеличивается (фиг. 15) и доля не кардиомиоцитов уменьшается (фиг. 16) при обработке соединением А, форетинибом, ZM323881 или CP-67351, фризотинибом, по сравнению с отсутствием обработки соединением (фиг. 16).
Экспериментальный пример 8. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
В качестве iPS-клеток человека используют ту же клеточную линию, что и в экспериментальном примере 6, и индукцию дифференциации в кардиомиоциты также проводят аналогичным способом.
Среду меняют на 8, 10, 13, 17 и 20 день индукции дифференциации, и это осуществляют способом, подобным тому, что применяют на 8 день в экспериментальном примере 1 на 8 день индукции дифференциации, и проводят с помощью способа, аналогичного тому, который применяют для 10 дня в экспериментальном примере 1, на 10 день и после индукции дифференциации. На 8, 10, 13, 17 и 20 дни индукции дифференциации добавляют исследуемое соединение креноланиб в соответствии с условиями, описанными в таблице 8, и способом, аналогичным описанному в экспериментальном примере 1. Соединения добавляют на 8 день или позже индукции дифференциации в эксперименте номер 4, на 10 день или после индукции дифференциации в экспериментах номер 2 и 3 и на 13 день или после индукции дифференциации в эксперименте номер 5. Для эксперимента номер 1 используют четыре лунки, три лунки используют для экспериментов номер 2 и 3, и одну лунку используют для экспериментов номер 4 и 5.
[Таблица 8]
На 23 день индукции дифференциации проводят диссоциацию на отдельные клетки, фиксацию, измерение доли кардиомиоцитов (доли положительных по саркомерному α-актинину клеток) и измерение доли не кардиомиоцитов способами, аналогичными тем, что описаны в экспериментальном примере 1. Доля кардиомиоцитов увеличивается (фиг. 17), и количество не кардиомиоцитов уменьшается (фиг. 18) при обработке креноланибом.
Экспериментальный пример 9. Верификация действия очистки кардиомиоцита ингибитором тирозинкиназы рецепторного типа
Индукция дифференциации в кардиомиоциты
В качестве iPS клеток человека используют ту же клеточную линию, что и в экспериментальном примере 6, и индукцию дифференциации в кардиомиоциты также проводят аналогичным способом.
Среду меняют способом, аналогичным таковому на 8 день в экспериментальном примере 1, на 8 день индукции дифференциации и проводят по методике, аналогичной таковой на 10 день в экспериментальном примере 1, на 10 и 13 день индукции дифференциации. На 8, 10 и 13 дни индукции дифференциации добавляют оцениваемые соединения (соединения и концентрации экспериментов №№ 2, 3 показаны в таблице 9 ниже) способом, аналогичным способу, описанному в экспериментальном примере 1. Соответствующие эксперименты проводят с использованием 4 лунок каждый.
[Таблица 9]
На 17 день индукции дифференциации проводят диссоциацию на отдельные клетки, фиксацию, измерение доли кардиомиоцитов (доли положительных по саркомерному α-актинину клеток) и измерение доли не кардиомиоцитов способами, аналогичными тем, что описаны в экспериментальном примере 1, и измерение количества клеток проводят методом, подобным тому, что описан в экспериментальном примере 4. Доля кардиомиоцитов увеличивается (фиг. 19) и доля не кардиомиоцитов снижается (фиг. 20) при обработке форетинибом или кризотинибом по сравнению с отсутствием обработки соединением. Около 80% количества клеток восстановлено с помощью кризотиниба, и почти такое же количество клеток восстановлено при обработке форетинибом по сравнению с отсутствием обработки соединением (фиг. 21).
Из вышеизложенного, показано, что кардиомиоциты в эмбриоидных тельцах могут быть очищены в различных клеточных линиях с помощью удобной обработки, заключающейся только в добавлении в среду различных видов ингибиторов тирозинкиназы рецепторного типа.
Тирозинкиназы рецепторного типа, которые должны быть мишенями для соединений, используемых в вышеупомянутых экспериментальных примерах 1-9, показаны в таблице 10.
[Таблица 10]
PDGFRα,β
c-Met
(HGFR)
[Промышленная применимость]
Согласно настоящему изобретению, предложена клеточная популяция, содержащая кардиомиоциты высокой чистоты. Такая клеточная популяция полезна, поскольку ее можно предпочтительно использовать для трансплантационной терапии сердечных заболеваний и скрининга терапевтических агентов для сердечных заболеваний.
Эта заявка основана на патентной заявке № 2020-050268, поданной в Японии (дата подачи: 19 марта 2020 г.), содержание которой полностью включено в настоящий документ.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИНДУКЦИИ ДИФФЕРЕНЦИАЦИИ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК В КАРДИОМИОЦИТЫ | 2007 |
|
RU2433174C2 |
| ИНСУЛИНПРОДУЦИРУЮЩИЕ КЛЕТКИ | 2019 |
|
RU2829220C2 |
| КОМПОЗИЦИИ КЛЕТОК, ПОЛУЧЕННЫЕ ИЗ ДЕДИФФЕРЕНЦИРОВАННЫХ ПЕРЕПРОГРАММИРОВАННЫХ КЛЕТОК | 2014 |
|
RU2675928C2 |
| СПОСОБ ИНДУКЦИИ ГИБЕЛИ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК И ДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК, ОТЛИЧНЫХ ОТ КАРДИОМИОЦИТОВ | 2010 |
|
RU2551778C2 |
| КОМПОЗИЦИИ КЛЕТОК, ПОЛУЧЕННЫЕ ИЗ ДЕДИФФЕРЕНЦИРОВАННЫХ ПЕРЕПРОГРАММИРОВАННЫХ КЛЕТОК | 2014 |
|
RU2754089C2 |
| СПОСОБ ПОЛУЧЕНИЯ БИОЛОГИЧЕСКОЙ ТКАНЕПОДОБНОЙ СТРУКТУРЫ | 2020 |
|
RU2815583C2 |
| СПОСОБ КОНСТРУИРОВАНИЯ МАССЫ МИОКАРДИАЛЬНЫХ КЛЕТОК И ПРИМЕНЕНИЕ МАССЫ МИОКАРДИАЛЬНЫХ КЛЕТОК | 2008 |
|
RU2467066C2 |
| СПОСОБ ДИФФЕРЕНЦИАЦИИ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК В КЛЕТКИ СОСУДИСТОГО РУСЛА | 2012 |
|
RU2618871C2 |
| СПОСОБ ПОЛУЧЕНИЯ ДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК ИЗ СТВОЛОВЫХ КЛЕТОК | 2009 |
|
RU2576003C2 |
| СПОСОБ ИНДУКЦИИ ДИФФЕРЕНЦИАЦИИ СТВОЛОВЫХ КЛЕТОК В МИОКАРДИАЛЬНЫЕ | 2004 |
|
RU2392315C2 |
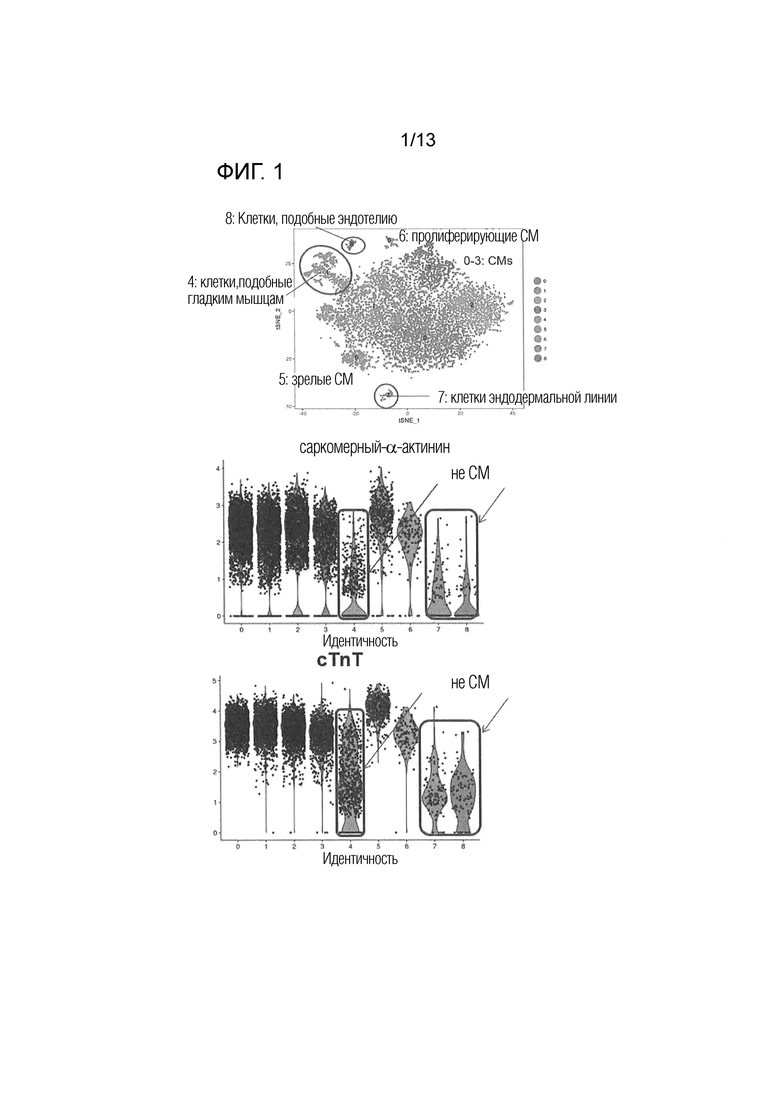



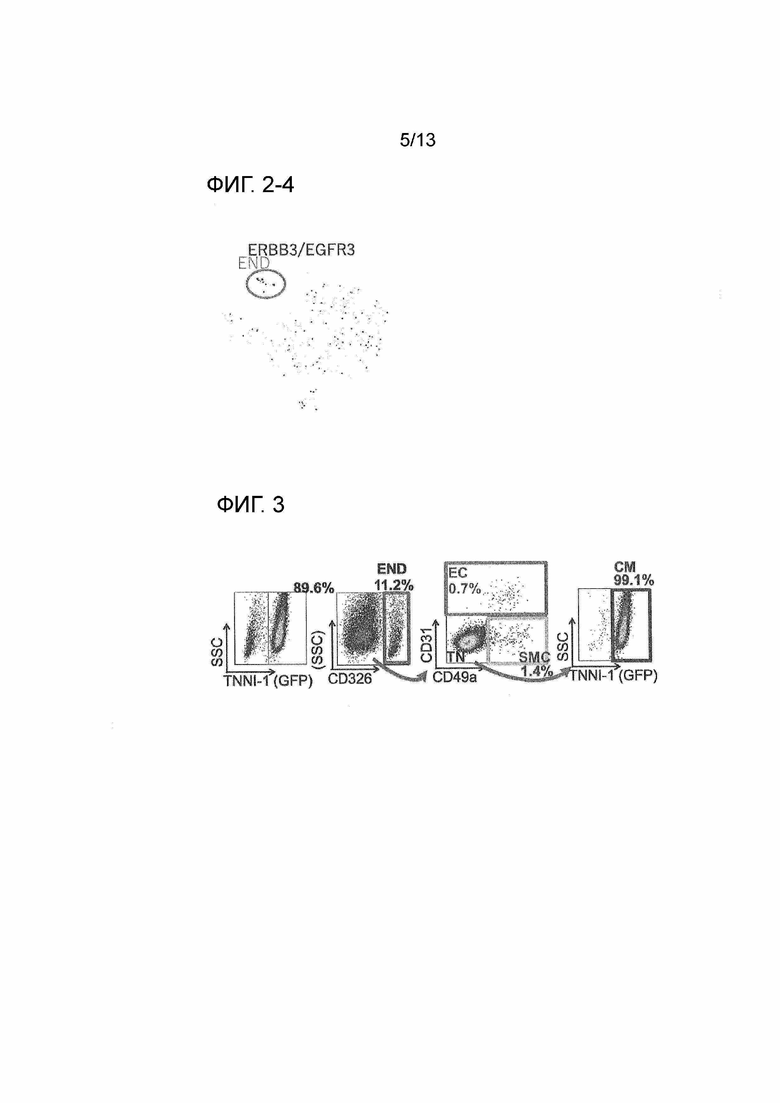
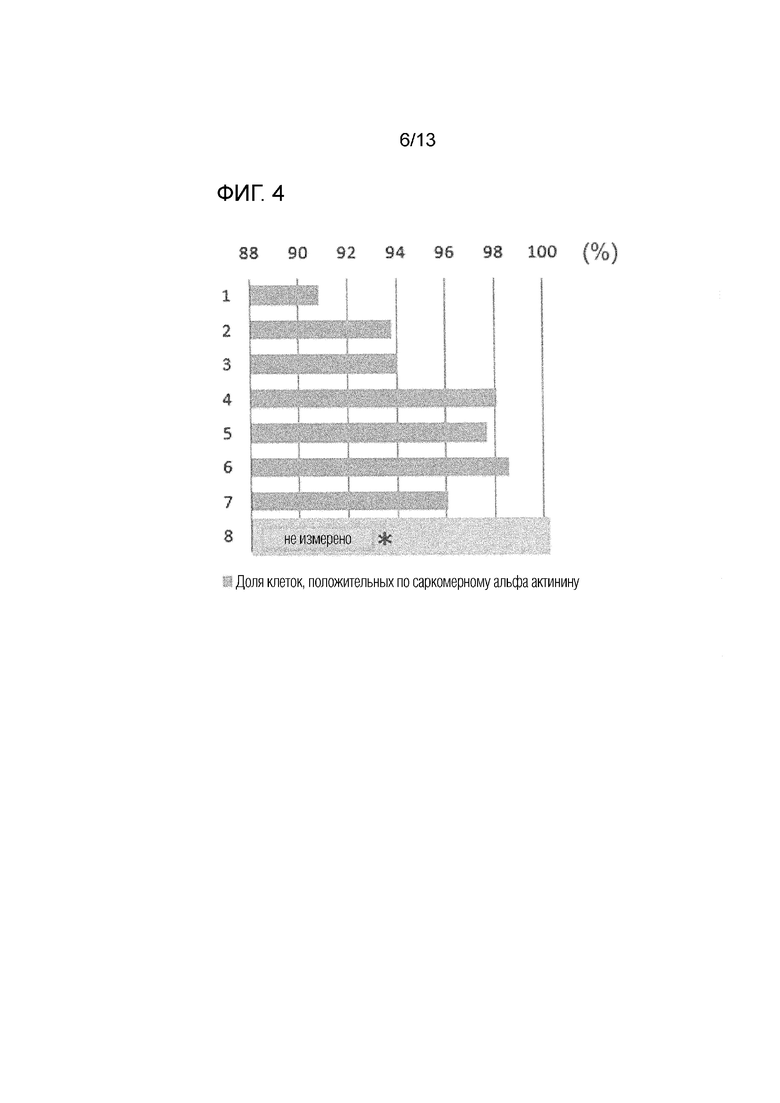
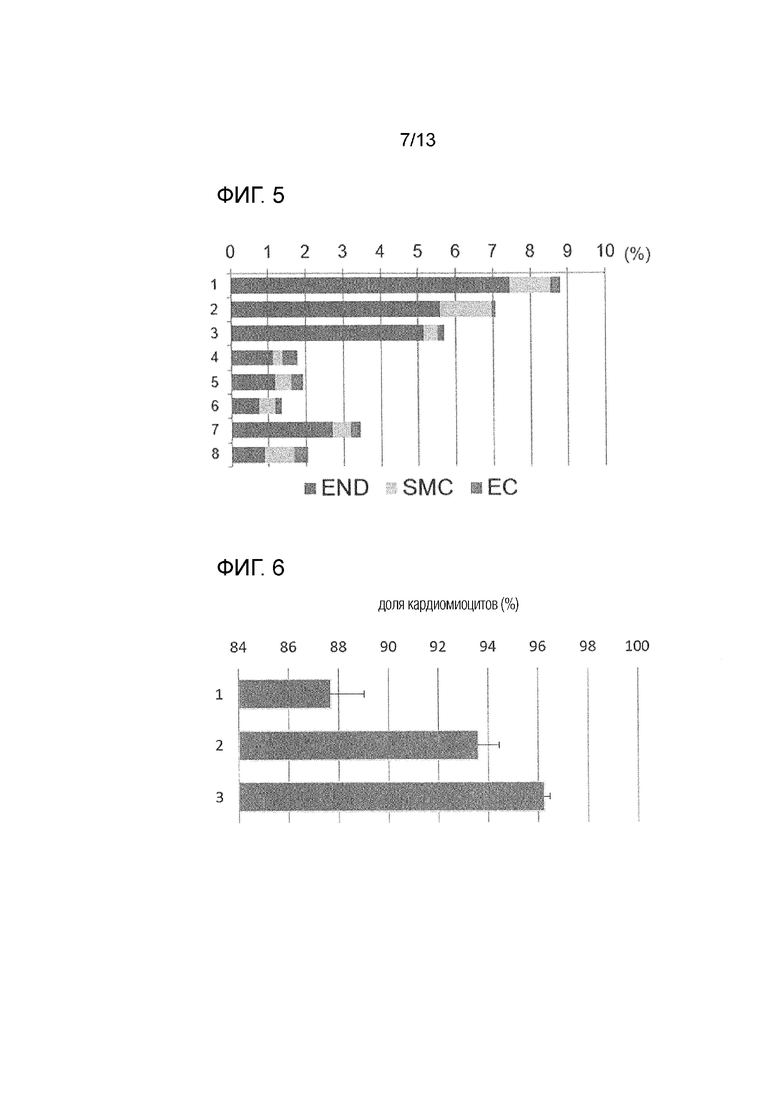

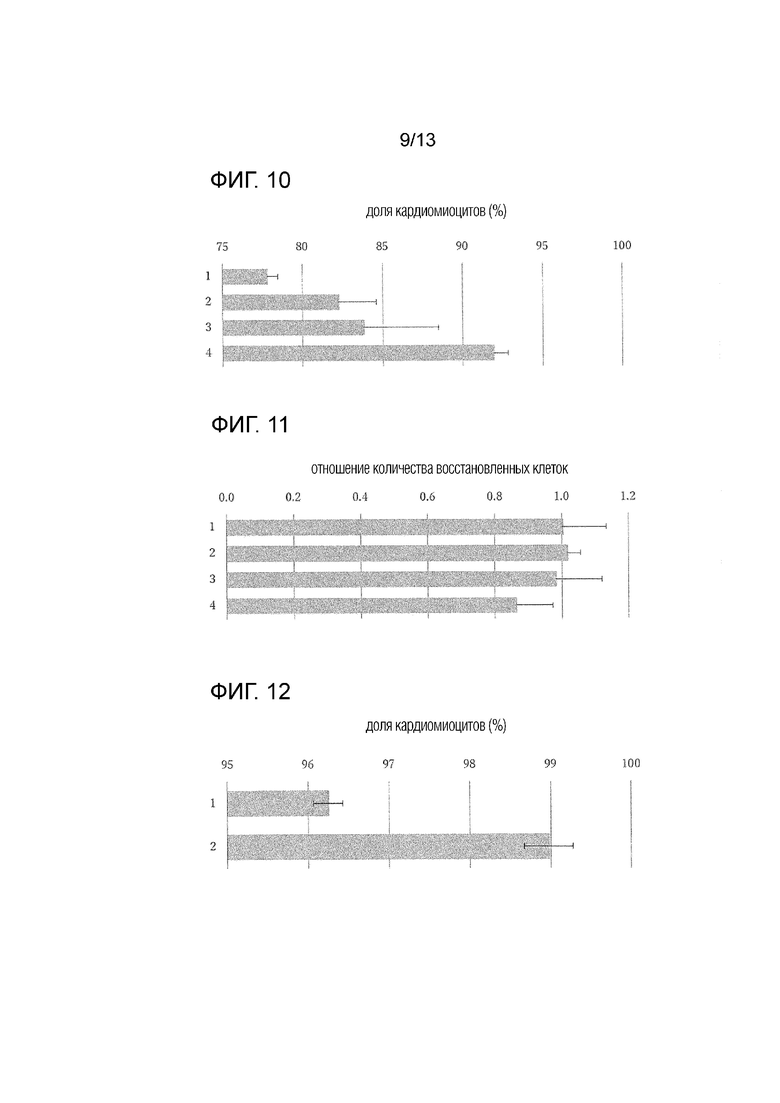

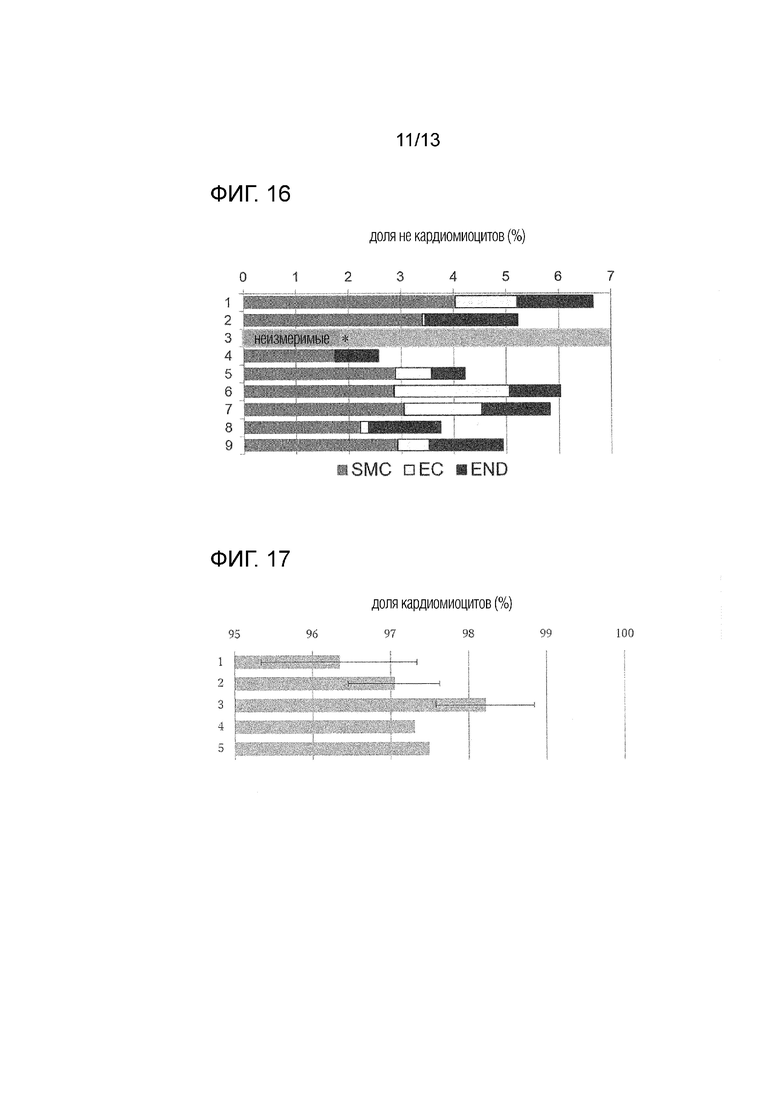
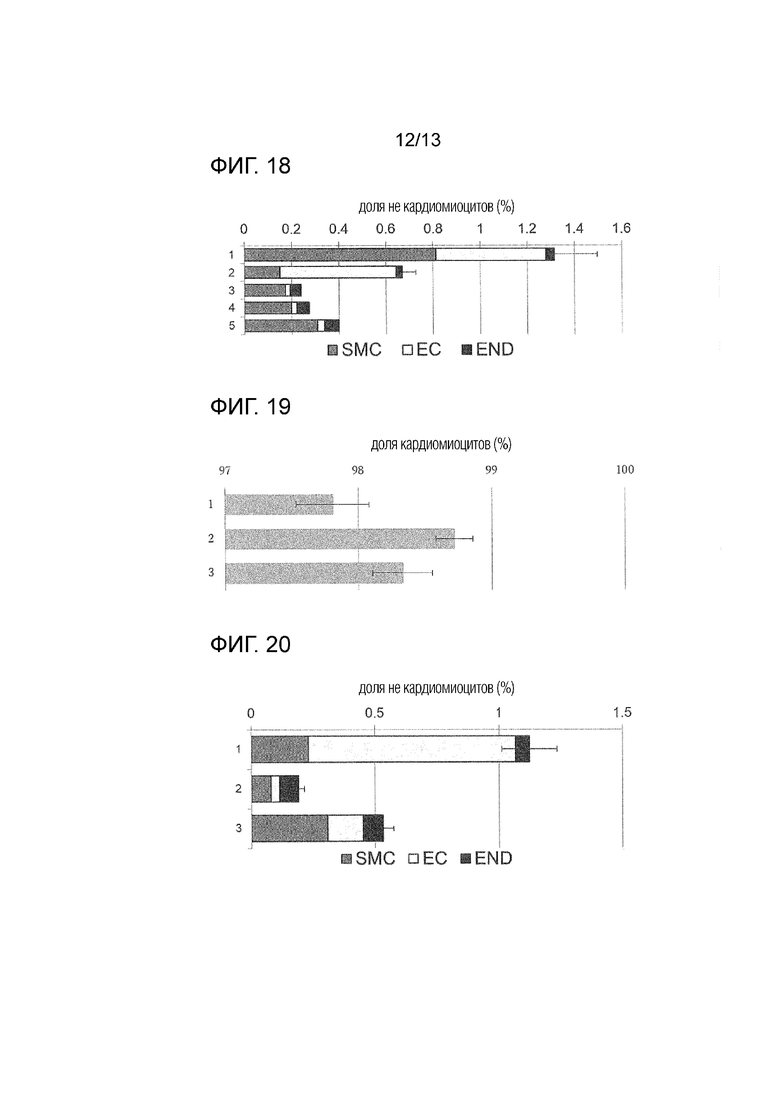

Изобретение относится к биотехнологии, в частности к способу получения клеточной популяции, содержащей кардиомиоциты, включающему (1) стадию контакта ингибитора тирозинкиназы рецепторного типа (за исключением ингибиторов рецептора EGF) с клеточной популяцией, содержащей кардиомиоциты или сердечные клетки-предшественники, и другие клетки, где клеточная популяция получена путем культивирования плюрипотентных стволовых клеток в среде для дифференциации кардиомиоцитов, и (2) стадию культивирования клеточной популяции. Изобретение позволяет получить популяцию клеток, подходящую для трансплантационной терапии сердечных заболеваний и скрининга терапевтических агентов для сердечных заболеваний. 2 н. и 5 з.п. ф-лы, 21 ил., 10 табл., 9 пр.
1. Способ получения клеточной популяции, содержащей кардиомиоциты, включающий
(1) стадию контакта ингибитора тирозинкиназы рецепторного типа, за исключением ингибиторов рецептора EGF, с клеточной популяцией, содержащей кардиомиоциты или сердечные клетки-предшественники и другие клетки, где клеточную популяцию получают путем культивирования плюрипотентных стволовых клеток в среде для дифференциации кардиомиоцитов, и
(2) стадию культивирования клеточной популяции в среде для дифференциации или поддержания кардиомиоцитов.
2. Способ по п. 1, отличающийся тем, что клеточная популяция контактирует с ингибитором тирозинкиназы рецепторного типа на стадии (1) через 4 дня после начала индукции дифференциации плюрипотентных стволовых клеток.
3. Способ по п. 1 или 2, отличающийся тем, что клеточная популяция контактирует с ингибитором тирозинкиназы рецепторного типа в течение не менее одного дня на стадии (1).
4. Способ по любому из пп. 1-3, отличающийся тем, что ингибитор представляет собой по меньшей мере один ингибитор тирозинкиназы рецепторного типа, выбранный из группы, состоящей из рецептора VEGF, рецептора PDGF, рецептора HGF и рецептора FGF.
5. Способ по любому из пп. 1-4, отличающийся тем, что ингибитор представляет собой по меньшей мере один член, выбранный из группы, состоящей из N-[5-({2-[(циклопропанкарбонил)амино]имидазо[1,2-b]пиридазин-6-ил}окси)-2-метилфенил]-1,3-диметил-1H-пиразол-5-карбоксамида, N-{4-[(6,7-диметоксихинолин-4-ил)окси]-3-фторфенил}-N'-(4-фторфенил)циклопропан-1,1-дикарбоксамида, AMG337, ASP5878, BGJ398, форетиниба, ZM323881, CP-673451, креноланиба и кризотиниба.
6. Способ по любому из пп. 1-5, отличающийся тем, что плюрипотентная стволовая клетка представляет собой индуцированную плюрипотентную стволовую клетку.
7. Способ очистки кардиомиоцитов, включающий
(1) стадию контакта ингибитора тирозинкиназы рецепторного типа с клеточной популяцией, содержащей кардиомиоциты или сердечные клетки-предшественники и другие клетки, где клеточную популяцию получают путем культивирования плюрипотентных стволовых клеток в среде для дифференциации кардиомиоцитов, и
(2) стадию культивирования клеточной популяции в среде для дифференциации или поддержания кардиомиоцитов.
| JP 2018510649 А, 19.04.2018 | |||
| JP 2017108705 A, 22.06.2017 | |||
| MIKI | |||
| et al., Efficient Detection and Purification of Cell Populations Using Synthetic MicroRNA Switches, Cell Stem Cell., 2015, 16, doi: 10.1016/j.stem | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| ABI-GERGES N | |||
| et al., Preservation of cardiomyocytes from the adult heart, J Mol Cell Cardiol, 2013, vol | |||
| Нефтяной конвертер | 1922 |
|
SU64A1 |
| Приспособление для останова мюля Dobson аnd Barlow при отработке съема | 1919 |
|
SU108A1 |

