Настоящее изобретение относится к слитому белку для применения в лечении и/или профилактике инфекции вирусом гепатита В (HBV).
Гепатит B представляет собой заболевание печени, вызываемое вирусами гепатита В. Болезнь поражает миллионы людей в год по всему миру. HBV присутствует в крови и биологических жидкостях инфицированных людей и, следовательно, может распространяться при контакте этих жидкостей с жидкостями здоровых людей.
HBV, в первую очередь, нарушает функции печени вследствие репликации в клетках печени. Во время инфекции HBV иммунный ответ хозяина вызывает как гепатоцеллюлярное повреждение, так и клиренс вируса.
Острую инфекцию HBV, как правило, не лечат, поскольку большинство людей способны спонтанно избавляться от инфекции. Однако хроническую инфекцию HBV необходимо лечить, чтобы снизить риск возникновения цирроза и рака печени. Противовирусные препараты, в настоящее время используемые при лечении инфекции HBV, включают ламивудин, адефовир, тенофовир, телбивудин и энтекавир. Кроме того, при лечении также может использоваться интерферон альфа-2а, действующий в качестве модулятора иммунной системы. Однако ни один из этих препаратов не способен избавить организм от инфекции HBV. Эти препараты могут только остановить репликацию HBV, таким образом, минимизируя повреждение печени.
Задача настоящего изобретения заключается в создании новых средств лечения и/или профилактики инфекции HBV, которые преодолевают недостатки существующих методов лечения HBV. Особенно важной задачей является клиренс вируса HBV у хронически инфицированных пациентов путем восстановления эффективного гуморального и клеточного иммунного ответа.
Эти задачи выполняются с помощью гибридного белка для применения в лечении и/или профилактике инфекции вирусом гепатита В, содержащего по меньшей мере один полипептид PreS вируса гепатита B (HBV PreS) или его фрагмент, слитый по меньшей мере с одним пептидом, состоящим из аминокислотной последовательности, имеющей по меньшей мере 80% идентичности с последовательностью, выбранный из группы, состоящей из SEQ ID Nо. 1, SEQ ID Nо. 2, SEQ ID Nо. 3 и SEQ ID No. 4.
Неожиданно оказалось, что слитый белок, содержащий полипептид PreS вируса гепатита B или его фрагмент и по меньшей мере один пептид, состоящий из аминокислотной последовательности, имеющей по меньшей мере 80% идентичности с последовательностью, выбранной из группы, состоящей из SEQ ID Nо. 1, SEQ ID Nо. 2, SEQ ID NO. 3 и SEQ ID NO. 4, вызывает образование специфичных к PreS антител у индивидуума в гораздо более высокой степени по сравнению только с одним PreS или по сравнению с другими слитыми белками, содержащими пептиды, отличные от SEQ ID Nо. 1, SEQ ID Nо. 2, SEQ ID NO. 3 и SEQ ID NO. 4. Кроме того, антитела, вырабатываемые в ответ на введение слитого белка по настоящему изобретению, имеют превосходные нейтрализующие эффекты в отношении вируса гепатита В и способны ингибировать заражение вирусом гепатита В. Впервые введение слитого белка, содержащего PreS, можно успешно использовать для лечения и/или профилактики инфекции вирусом гепатита В у человека.
Как показано на фиг. 2B, введение слитого белка по настоящему изобретению приводит к образованию антител, которые специфичны к первым 30 (пептиду P1) и 50 (пептиду Р2) аминокислотным остаткам HBV PreS, в более низкой степени - к С-концевой области (пептиды P6-P8), и в незначительной степени - к центральной части HBV PreS (пептидам P4 и P5). Поскольку N-концевая часть HBV PreS, как известно, играет важную роль в прикреплении HBV к клеткам печени и инфекции HBV, антитела, направленные на эту часть PreS-полипептида, особенно полезны для лечения и/или профилактики инфекции HBV. В отличие от этого, одиночное введение HBV PreS не вызывает таких эффектов. Антитела, полученные таким образом, способны связываться с практически любой частью HBV PreS (см. фиг. 2А). Это показывает, что иммунный ответ, индуцированный слитыми белками по настоящему изобретению, более сфокусирован на тех частях полипептида HBV PreS, которые участвуют в инфекции HBV.
Слитый белок по настоящему изобретению может содержать один или несколько полипептидов PreS вируса гепатита B или их один или несколько фрагментов. Наличие более одного полипептида PreS вируса гепатита B или их фрагментов в слитом белке имеет то преимущество, что иммунной системе представляется несколько антигенов, что позволяет образование еще большего количества антител, направленных на PreS. В особенно предпочтительном варианте осуществления настоящего изобретения слитый белок содержит один, два, три, четыре, пять, шесть, семь, восемь, девять или десять полипептидов PreS вируса гепатита В или их фрагментов. Полипептиды HBV PreS, а также их фрагменты, описанные в данном документе, являясь частью слитого белка по настоящему изобретению, могут быть получены из одного генотипа HBV или из различных генотипов. Например, слитый белок по настоящему изобретению может содержать полипептид PreS или его фрагмент только из генотипа А HBV или может быть объединен с дополнительным полипептидом PreS или его фрагментом, полученным из генотипа B, C, D, E, F, G или H HBV.
В особенно предпочтительном варианте осуществления настоящего изобретения слитый белок содержит по меньшей мере один пептид, состоящий из аминокислотной последовательности, имеющей по меньшей мере 80% идентичности с SEQ ID Nо. 1, по меньшей мере один пептид, состоящий из аминокислотной последовательности, имеющей по меньшей мере 80% идентичности с SEQ ID Nо. 2, по меньшей мере один пептид, состоящий из аминокислотной последовательности, имеющей по меньшей мере 80% идентичность с SEQ ID № 3, и по меньшей мере один пептид, состоящий из аминокислотной последовательности, имеющей по меньшей мере 80% идентичности с SEQ ID Nо. 4. В альтернативном варианте слитый белок по настоящему изобретению может содержать один, два, три, четыре, пять шесть, семь, восемь, девять или десять этих пептидов в любой возможной комбинации или даже только один конкретный пептид в таком же количестве.
Термины «слитый с» или «слитый белок», используемые в данном описании, относятся к белку, содержащему полипептид PreS вируса гепатита B или его фрагмент, который экспрессируется и получается в виде одной рекомбинантной полипептидной цепи.
Способы получения слитых белков хорошо известны в данной области и могут быть найдены в стандартных источниках по молекулярной биологии, таких как Sambrook et al. (Molecular Cloning, 2nd ed., Cold Spring Harbor Laboratory Press, 1989) и Ausubel et al. (Current protocols in molecular biology, 3rd ed.; Wiley and Sons, 1995). В общем, слитый белок получают путем сначала создания конструкции слитого гена, которая вставляется в подходящий экспрессионный вектор, который, в свою очередь, используется для трансфекции подходящей клетки-хозяина. В общем, рекомбинантные слитые конструкции получают рядом расщеплений рестрикционными ферментами и реакций лигирования, которые приводят к включению целевых последовательностей в плазмиду. Если подходящие сайты рестрикции недоступны, то могут быть использованы синтетические олигонуклеотидные адаптеры или линкеры, известные специалистам в данной области техники и описанные в приведенных выше ссылках. Полинуклеотидные последовательности, кодирующие аллергены и нативные белки, могут быть собраны до вставки в подходящий вектор, или же последовательность, кодирующая аллерген, может быть вставлена рядом с уже присутствующей в векторе последовательностью, кодирующей нативную последовательность. Вставка последовательности в вектор должна осуществляться с сохранением рамки считывания, так что последовательность может транскрибироваться в белок. Среднему специалисту в данной области будет очевидно, что точные требуемые рестрикционные ферменты, линкеры и/или адаптеры, а также точные условия реакции, будут меняться в зависимости от используемых последовательностей и клонирующих векторов. Создание конструкций ДНК, тем не менее, является обычным в данной области техники и может быть легко осуществлено специалистом в данной области техники.
Фрагмент полипептида PreS вируса гепатита B состоит, предпочтительно, по меньшей мере из 30, предпочтительно, по меньшей мере из 40, более предпочтительно, по меньшей мере из 50 последовательных аминокислотных остатков и может содержать PreS1 и/или PreS2 из полипептида PreS вируса гепатита В. В особенно предпочтительном варианте осуществления настоящего изобретения фрагмент полипептида PreS вируса гепатита В может включать аминокислотные остатки от 1 до 70, предпочтительно, аминокислотные остатки от 1 до 65, более предпочтительно, аминокислотные остатки от 1 до 60, более предпочтительно, аминокислотные остатки от 1 до 55, более предпочтительно, аминокислотные остатки от 1 до 50, более предпочтительно, от 1 до 45, более предпочтительно, аминокислотные остатки от 1 до 40, более предпочтительно, аминокислотные остатки от 1 до 35, более предпочтительно, аминокислотные остатки от 5 до 70, более предпочтительно, аминокислотные остатки от 5 до 65, более предпочтительно, аминокислотные остатки от 5 до 60, более предпочтительно, аминокислотные остатки от 5 до 55, более предпочтительно, аминокислотные остатки от 5 до 50, более предпочтительно, от 5 до 45, более предпочтительно, аминокислотные остатки от 5 до 40, более предпочтительно, аминокислотные остатки от 5 до 35, более предпочтительно, аминокислотные остатки от 10 до 70, более предпочтительно, аминокислотные остатки от 10 до 65, более предпочтительно, аминокислотные остатки от 10 до 60, более предпочтительно, аминокислотные остатки от 10 до 55, более предпочтительно, аминокислотные остатки от 10 до 50, более предпочтительно, от 10 до 45, более предпочтительно, аминокислотные остатки от 10 до 40, более предпочтительно, аминокислотные остатки от 10 до 35, более предпочтительно, аминокислотные остатки от 15 до 70, более предпочтительно, аминокислотные остатки от 15 до 65, более предпочтительно, аминокислотные остатки от 15 до 60, более предпочтительно, аминокислотные остатки от 15 до 55, более предпочтительно, аминокислотные остатки от 15 до 50, более предпочтительно, от 15 до 45, более предпочтительно, аминокислотные остатки от 15 до 40, более предпочтительно, аминокислотные остатки от 15 до 35 полипептида PreS вируса гепатита B, предпочтительно, полипептидов PreS HBV, состоящих из последовательностей SEQ ID No. 5, 7, 8, 9, 10, 11, 12, 13 или 14, причем SEQ ID Nо. 8-14 принадлежат генотипам В-Н HBV, соответственно.
По меньшей мере один пептид, подлежащий слиянию по меньшей мере с одним полипептидом PreS вируса гепатита B или его фрагментом, имеет идентичность по меньшей мере на 80%, предпочтительно, по меньшей мере на 85%, более предпочтительно, по меньшей мере на 90%, более предпочтительно, по меньшей мере на 92%, более предпочтительно, по меньшей мере на 94%, более предпочтительно, по меньшей мере на 96%, более предпочтительно, по меньшей мере на 98%, более предпочтительно, по меньшей мере на 99%, в частности, на 100% с SEQ ID Nо. 1, SEQ ID Nо. 2, SEQ ID Nо. 3 и SEQ ID Nо. 4. Степень идентичности первой аминокислотной последовательности и второй аминокислотной последовательности может быть определена путем прямого сравнения двух аминокислотных последовательностей с использованием некоторых алгоритмов. Идентичность последовательностей предпочтительно определять BLAST-выравниванием (http://blast.ncbi.nlm.nih.gov/; Altschul SF et al., J. Mol. Biol. 215 (1990): 403-410) с использованием матрицы BLOSUM62, штрафом за разрыв 11 и штрафом за удлинение разрыва 1.
В соответствии с предпочтительным вариантом осуществления настоящего изобретения аминокислотная последовательность полипептида PreS по меньшей мере на 80% идентична SEQ ID Nо. 5, SEQ ID Nо. 7, SEQ ID Nо. 8, SEQ ID Nо. 9, SEQ ID Nо. 10, SEQ ID Nо. 11, SEQ ID Nо. 12 или SEQ ID Nо. 13, наиболее предпочтительно, SEQ ID Nо. 5.
Полипептид PreS вируса гепатита B, слитый по меньшей мере с одним из пептидов, описанных выше, имеет идентичность по меньшей мере 80%, предпочтительно, по меньшей мере 85%, более предпочтительно. по меньшей мере 90%, более предпочтительно, по меньшей мере 92%, более предпочтительно, по меньшей мере 94%, более предпочтительно, по меньшей мере 96%, более предпочтительно, по меньшей мере 98%, более предпочтительно, по меньшей мере 99%, в частности, 100% с SEQ ID Nо. 5, SEQ ID Nо. 7, SEQ ID Nо. 8, SEQ ID Nо. 9, SEQ ID Nо. 10, SEQ ID Nо. 11, SEQ ID Nо. 12 или SEQ ID Nо. 13.
В соответствии с дополнительным предпочтительным вариантом осуществления настоящего изобретения по меньшей мере один пептид слит с N- и/или С-концом полипептида PreS.
«Слитый с N- и/или С-концом» в контексте настоящего изобретения означает, что по меньшей мере один пептид слит с N- и/или С-концом полипептида PreS или его фрагмента. Слитый белок по настоящему изобретению может содержать один или несколько пептидов, слитых с N-концом полипептида PreS или его фрагмента, либо с их С-концом.
В соответствии с предпочтительным вариантом осуществления настоящего изобретения слитый белок содержит аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID Nо. 6.
Слитый белок по настоящему изобретению имеет идентичность по меньшей мере 80%, предпочтительно, по меньшей мере 85%, более предпочтительно, по меньшей мере 90%, более предпочтительно, по меньшей мере 92%, более предпочтительно, по меньшей мере 94%, более предпочтительно, по меньшей мере 96%, более предпочтительно, по меньшей мере 98%, более предпочтительно, по меньшей мере 99%, в частности, 100% с SEQ ID Nо. 6.
В соответствии с еще одним предпочтительным вариантом осуществления настоящего изобретения инфекция вирусом гепатита В вызвана генотипом А, В, С, D, Е, F, G, Н вируса гепатита В или их подтипом. Предпочтительно использовать полипептид PreS HBV или его фрагмент такого же генотипа для лечения и/или профилактики инфекции HBV, вызванной данным генотипом HBV (например, PreS HBV с генотипом A используется для лечения/профилактики инфекции генотипом A HBV или одним из его подтипов). Благодаря консервативным аминокислотным последовательностям в этих частях полипептида PreS, которые, как известно, участвуют в инфекции HBV, конечно, также можно использовать полипептид PreS HBV или его фрагмент одного генотипа для лечения/профилактики инфекции другим генотипом HBV (например, PreS HBV с генотипом A используется для лечения/предупреждения инфекции HBV с генотипом В, С, D, Е, F, G и/или Н или их подтипом).
Слитый белок по настоящему изобретению может быть использован для лечения и/или профилактики инфекции HBV с различными генотипами и их подтипами. Подтипы вирусов гепатита B, включают A1, A2, A3, A4, A5, B1, B2, B3, B4, B5, C1, C2, C3, C4, C5, D1, D2, D3, D4, D5, F1, F2, F3 и F4, описанные у Schaefer и др. (World J Gastroenterol 13 (2007): 14-21).
В соответствии с особенно предпочтительным вариантом осуществления настоящего изобретения слитый белок вводят индивидууму по меньшей мере один раз в количестве от 0,01 мкг/кг массы тела до 5 мг/кг массы тела, предпочтительно, от 0,1 мкг/кг массы тела до 2 мг/кг массы тела. В соответствии с еще одним предпочтительным вариантом осуществления настоящего изобретения слитый белок вводят пациенту в количестве от 5 до 50 мкг, предпочтительно, от 10 до 40 мкг, более предпочтительно, от 15 до 30 мкг, либо независимо от массы тела (т.е. доза может включать 15, 20, 25 или 30 мкг), либо в пересчете на кг массы тела.
Количество слитого белка, которое может быть объединено с вспомогательными веществами для получения единичной лекарственной формы, будет меняться в зависимости от получающего лечение пациента и конкретного способа введения. Доза слитого белка может варьировать в зависимости от таких факторов, как состояние заболевания, возраст, пол и вес индивидуума, а также от способности вызывать желаемую выработку антител у индивидуума. Схема введения может регулироваться для обеспечения оптимального терапевтического ответа. Например, несколько дробных доз можно вводить ежедневно, или доза может быть пропорционально снижена в зависимости от конкретной терапевтической ситуации. Доза вакцины также может быть изменена, чтобы обеспечить оптимальный ответ на профилактическую дозу в зависимости от обстоятельств. Например, полипептиды и вакцины по настоящему изобретению могут быть введены индивидууму с интервалами в несколько дней, от одной до двух недель или даже месяцев, всегда в зависимости от уровня индукции IgG специфичных к PreS вируса гепатита B.
В предпочтительном варианте осуществления настоящего изобретения слитый белок по настоящему изобретению вводится от 2 до 10 раз, предпочтительно, от 2 до 7 раз, еще более предпочтительно, до 5 раз и, наиболее предпочтительно, до 3 раз. В особенно предпочтительном варианте осуществления временной интервал между соседними вакцинациями выбирается так, чтобы он составлял от 2 недель до 5 лет, предпочтительно, от 1 месяца до 3 лет, более предпочтительно, от 2 месяцев до 1,5 лет. Повторное введение слитого белка по настоящему изобретению может максимизировать конечный эффект лечения.
В соответствии с дополнительным предпочтительным вариантом осуществления настоящего изобретения слитый белок вводят по меньшей мере совместно с одним адъювантом и/или фармацевтически приемлемым вспомогательным веществом.
Слитый белок по настоящему изобретению можно вводить подкожно, внутримышечно, внутривенно, через слизистые и т.д. В зависимости от лекарственной формы и способа введения слитый белок по настоящему изобретению может быть объединен со вспомогательными веществами, разбавителями, адъювантами и/или носителями. Предпочтительным адъювантом являются квасцы. Подходящие протоколы для получения вакцинных препаратов известны специалистам в данной области и могут быть найдены, например, в ʺVaccine protocolsʺ (A. Robinson, M. P. Cranage, M. Hudson; Humana Press Inc., U. S.; 2nd edition 2003).
Слитый белок по настоящему изобретению может быть введен в состав также с другими адъювантами, регулярно используемыми в вакцинах. Например, подходящие адъювантами могут являться MF59, фосфат алюминия, фосфат кальция, цитокины (например, IL-2, IL-12, GM-CSF), сапонины (например, QS21), производные MDP, CpG-олигонуклеотиды, LPS, MPL, полифосфазены, эмульсии (например, Фрейнда, SAF), липосомы, виросомы, ISCOM, кохлеаты, PLG-микрочастицы, частицы полоксамеров, вирусоподобные частицы, термолабильный энтеротоксин (LT), холерный токсин (СТ), мутантные токсины (например, LTK63 и LTR72), микрочастицы и/или полимеризованные липосомы. Подходящие адъюванты являются коммерчески доступными, как, например, AS01B (MPL и QS21 в липосомном составе), AS02A, AS15, AS-2, АS-03 и их производные (GlaxoSmithKline, США); CWS (скелет клеточной стенки), TDM (трегалоза-6,6'-димиколят), LeIF (фактор инициации элонгации лейшманий), соли алюминия, такие как гелевый гидроксид алюминия (квасцы) или фосфат алюминия; соли кальция, железа или цинка; нерастворимая суспензия ацилированного тирозина; ацилированные сахара; катионно или анионно-дериватизированные полисахариды; полифосфазены; биоразлагаемые микросферы; монофосфориллипид А и Квил-А. Цитокины, такие как GM-CSF или интерлейкин-2, -7 или -12, также могут быть использованы в качестве адъювантов. Предпочтительными адъюванты для индукции преимущественно ответа Th1-типа включают, например, комбинацию монофосфориллипида А, предпочтительно, 3-О-деацилированного монофосфориллипида А (3D-MPL), необязательно, с солью алюминия. Водные составы, содержащие монофосфориллипид А и поверхностно-активное вещество, описаны в WO 98/43670.
Другим предпочтительным адъювантом является сапонин, либо миметики или производные сапонина, предпочтительно, QS21 (Aquila Biopharmaceuticals Inc.), которые могут использоваться отдельно или в комбинации с другими адъювантами. Например, улучшенная система включает комбинацию монофосфориллипида А и производного сапонина, такую как комбинация QS21 и 3D-MPL. Другие предпочтительные композиции содержат эмульсию типа масло-в-воде и токоферол. Особенно сильный адъювантный состав представляет собой QS21, 3D-MPL и токоферол в эмульсии масло-в-воде. Дополнительные сапониновые адъюванты для использования в настоящем изобретении, включают QS7 (описанный в WO 96/33739 и WO 96/11711) и QS17 (описанный в патенте США № 5057540 и европейском патенте № 0362279 B1).
Последовательности SEQ ID No., раскрытые в настоящем документе, имеют следующие аминокислотные последовательности с номерам доступа в безе данных Genbank:
ANSNNPDWDFNPIKDHWPAANQVGVGAFGPGLTP
PHGGILGWSPQAQGILTTVSTIPPPASTNRQSGR
QPTPISPPLRDSHPQAMQWNSTAFHQALQDPRVR
GLYFPAGGSSSGTVNPAPNIASHISSISARTGDP
VTN
VKVIPAGELQVIEKVDAAFKVAATAANAAPANDK
GGWSSKPRKGMGTNLSVPNPLGFFPDHQLDPAFG
ANSNNPDWDFNPIKDHWPAANQVGVGAFGPGLTP
PHGGILGWSPQAQGILTTVSTIPPPASTNRQSGR
QPTPISPPLRDSHPQAMQWNSTAFHQALQDPRVR
GLYFPAGGSSSGTVNPAPNIASHISSISARTGDP
VTNADLGYGPATPAAPAAGYTPATPAAPAEAAPA
GKATTEEQKLIEKINAGFKAALAAAAGVQPADKY
R
ANSDNPDWDLNPHKDNWPDSNKVGVGAFGLGFTP
PHGGLLGWSPQAQGILTTVPAAPPPASTNRQVAR
PPTPLSPPLRDTHPQAMQWNSTTFHQTLQDPRVR
ALYFPAGGSSSGTVNPVQNTASSISSILSKTGDP
VPN
ANSNNPDWDFNPNKDPWPEAWQVGVGAFGPGFTP
PHGGLLGWSPQAQGILTPVPAAPPPASTNRQSGR
QPTPISPPLRDSHPQAMQWNSTTFHQALLDPRVR
GLYFPAGGSSSGTTNPVPTTASPISSIFSRTGDP
APN
PNKDTWPDANKVGAGAFGLGFTPPHGGLLGWSPQ
AQGILQTLPTNPPPASTNRQSGRQPTPLSPPLRN
THPQAMQWNSTTFHQTLQDPRVRGLYFPAGGSSS
GTVNPVPTTVSHISSIFSRIGDPALN
NTRNPDWDHNPNKDHWTAANKVGVGAFGPGFTPP
HGGLLGWSPQAQGMLKTLPADPPPASTNRQSGRQ
PTPITPPLRDTHPQAMQWNSTTFHQALQDPRVRG
LYFPAGGSSSGTVNPVPTTASLIССIFSRIGDPA
PN
ANSSSPDWDFNKSKDNWPMANKVGVGGYGPGFTP
PHGGLLGWSPQAQGVLTTLPADPPPASTNRRSGR
KPTPVSPPLRDTHPQAMQWNSTQFHQALLDPRVR
ALYFPAGGSSSETQNPAPTIASLTSSIFLKTGGP
AMN
NTNNPDWDFNPKKDPWPEANKVGVGAYGPGFTPP
HGGLLGWSPQSQGTLTTLPADPPPASTNRQSGRQ
PTPISPPLRDSHPQAMQWNSTAFHQALQNPKVRG
LYFPAGGSSSGIVNPVPTIASHISSIFSRIGDPA
PN
ANSSSPDWDFNTNKDNWPMANKVGVGGFGPGFTP
PHGGLLGWSPQAQGILTTSPPDPPPASTNRRSGR
KPTPVSPPLRDTHPQAMQWNSTQFHQALLDPRVR
GLYFPAGGSSSETQNPAPTIASLTSSIFSKTGDP
AMN
Настоящее изобретение далее проиллюстрировано нижеследующими фигурами и примерами, однако не ограничено ими.
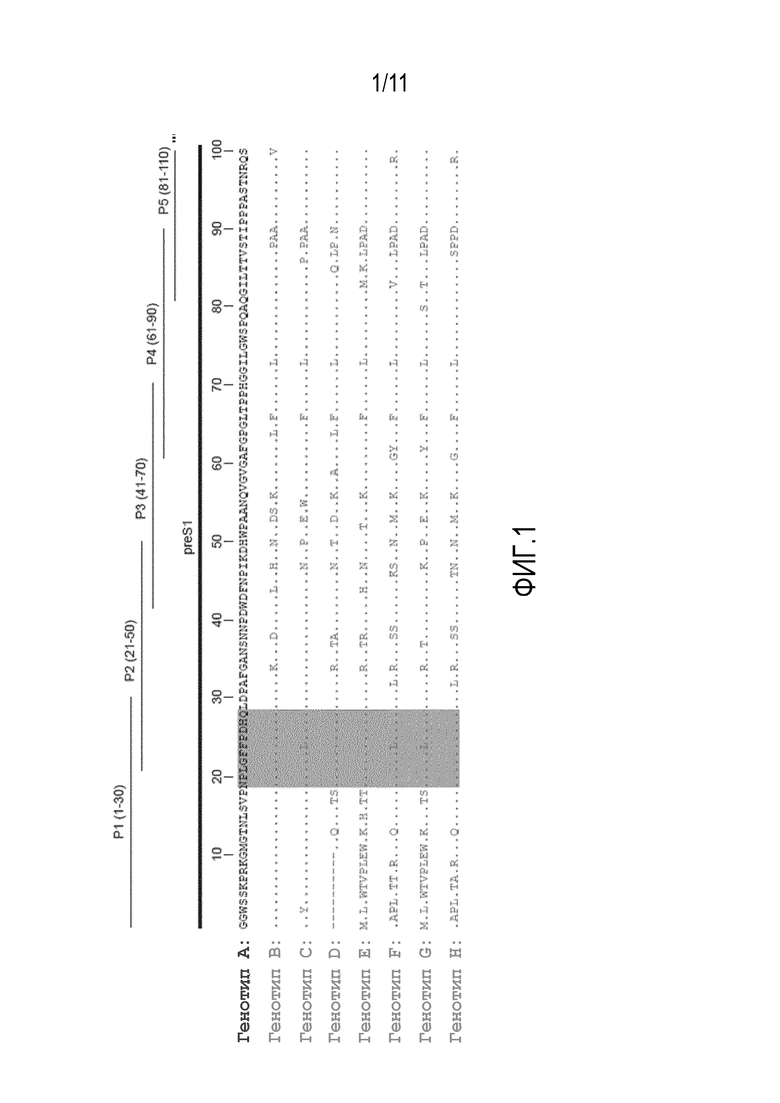
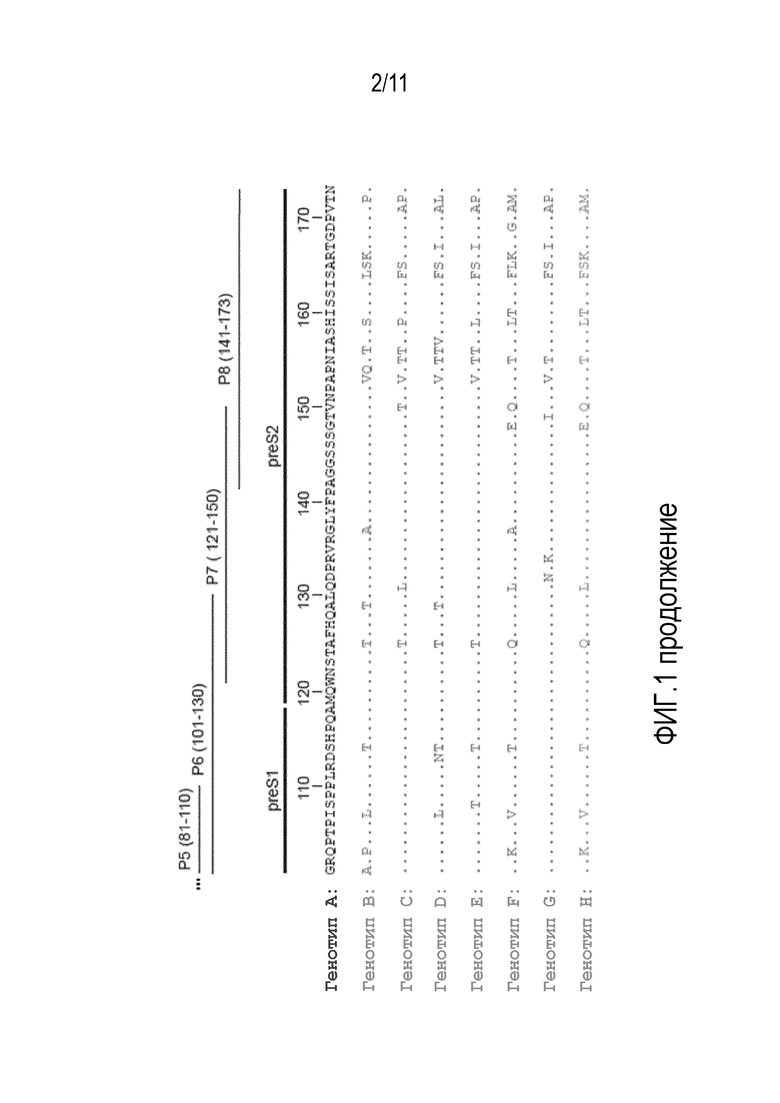
На фиг. 1 показано распределение пептидов PreS на выравненных последовательностях PreS из различных генотипов. Идентичные аминокислоты обозначены точками, домен PreS1 включает аминокислотные остатки 1-118, и домен PreS2 - аминокислотные остатки 119-173 (см. также SEQ ID Nо. 5), а аминокислотные остатки 19-28 (в сером прямоугольнике) играют ключевую роль в прикреплении к клеткам печени и инфекции HBV.
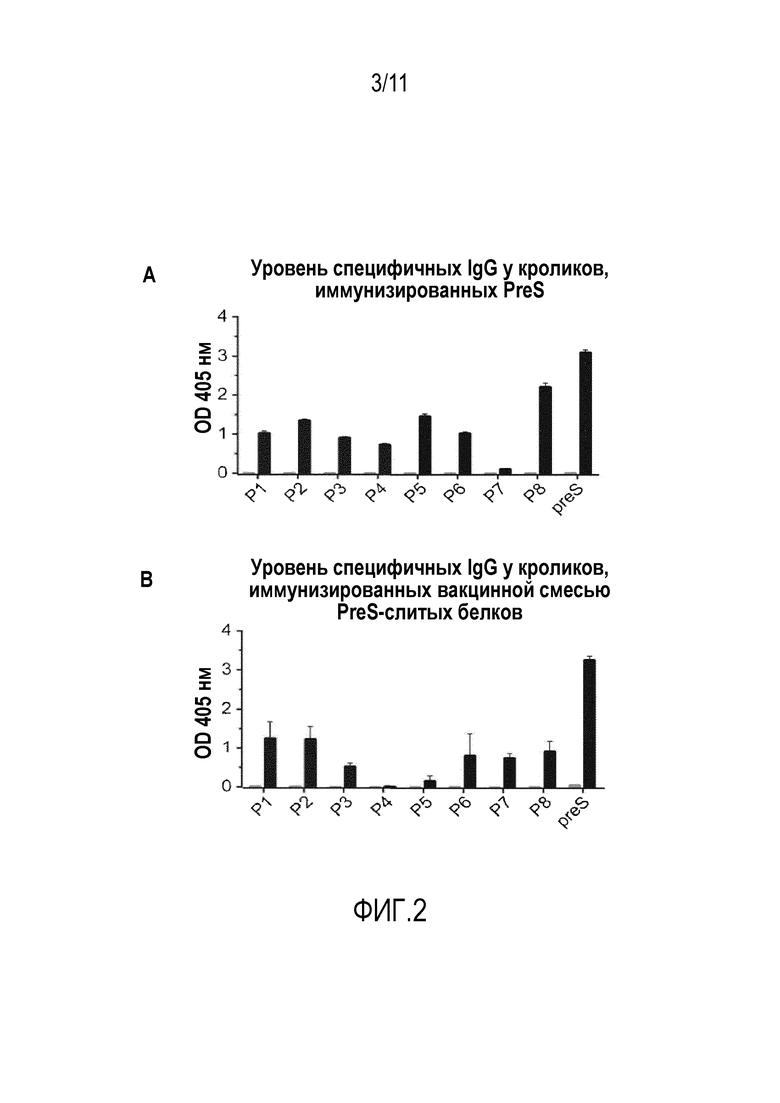
На фиг. 2 показана выработка IgG у кроликов, в ответ на иммунизацию PreS (n=1;. фиг.2А) или 20 мкг вакцинной смеси PreS-слитых белков (n= 2;. фиг.2В) до (левые столбики серого цвета) и после (правые столбики черного цвета) иммунизации. Значения оптической плотности (ось y: значения OD при 405нм) соответствуют уровням IgG к PreS и синтетическим перекрывающимся пептидам-производным PreS Р1-Р8 (ось x). Результаты представляют собой средние значения с SD (среднеквадратичным отклонением) для трех измерений.
На фиг. 3А-3С показана выработка IgG к PreS (фиг. 3А) и синтетическим перекрывающимся пептидам-производным PreS Р1-Р8 (фиг. 3B и 3C) у субъектов в ответ на вакцинацию вакцинной смесью PreS-слитых белков или плацебо. Показаны значения оптической плотности (ось y: значения OD, среднее из трех определений), соответствующие уровням IgG к PreS и пептидам Р1-Р8, измеренные у пациентов с или без предварительной вакцинации против гепатита В, которые были иммунизированы вакцинной смесью PreS-слитых белков (n=22) или плацебо (n=8) до (точка V5) и в различные моменты времени после иммунизации (точки V8 и V15 на оси x). Результаты представлены в виде средних значений с SD, и достоверные различия (у всех индивидуумов, вакцинированных вакцинной смесью PreS-слитых белков, в точках V5, V8 и V15) указаны как *р<0,05, **р<0,01, ***р<0,001.
На фиг. 4 показана выработка специфичных к PreS антител у субъектов в ответ на вакцинацию вакцинной смесью PreS-слитых белков (PreS-FVM) или плацебо и показаны антитела, присутствующие у инфицированных гепатитом В индивидуумов. Показаны значения оптической плотности (ось y: значения OD), соответствующие уровням IgA, IgE, IgM, IgG и подклассам IgG (IgG1-IgG4), специфичным к PreS, у субъектов, иммунизированных плацебо (n=8), 20 мкг (n=10) или 40 мкг (n=12) вакцинной смеси PreS-слитых белков, а также у инфицированных гепатитом B индивидуумов (n=19) (ось x). На графиках показаны средние значения с SD. Достоверные различия указаны как ***р<0,001.
На фиг. 5 показана выработка IgG, специфичных к PreS-пептидам P1-P8, у субъектов в ответ на вакцинацию Вакцинной смесью PreS-слитых белков (PreS-FVM) или плацебо, а также показаны IgG, присутствующие у инфицированных гепатитом В индивидуумов. Показаны значения оптической плотности (ось y: значения OD), которые соответствуют уровням IgG, специфичным к пептидам-производным PreS (Р1-Р8), у субъектов, иммунизированных плацебо (n=8), 20 мкг (n=10) или 40 мкг вакцинной смеси PreS-слитых белков (n=12), в точке V15, а также у инфицированных гепатитом В индивидуумов (n=19) (ось x). Результаты представлены в виде средних значений с SD.
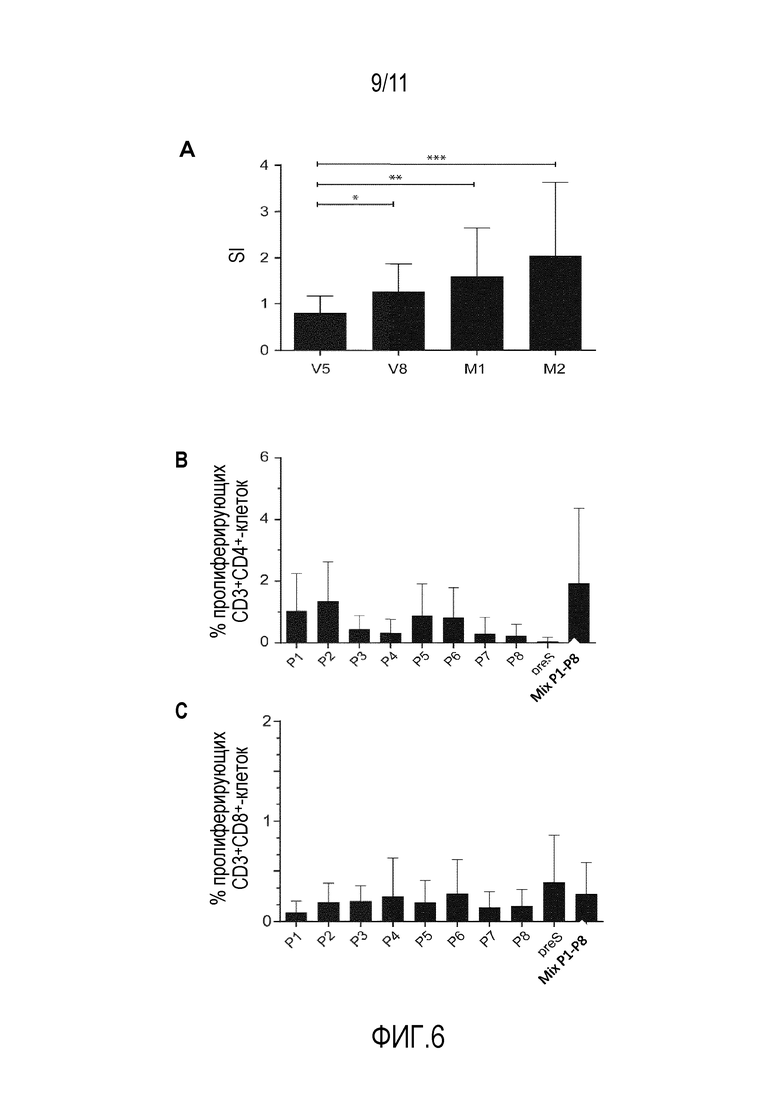
На фиг. 6 показаны специфичные к PreS и пептидам Т-клеточные ответы. Фиг.6А: пролиферацию PreS-специфичных РВМС (ось y: индекс стимуляции, SI) оценивали с помощью включения [3H]тимидина у субъектов, иммунизированных вакцинной смесью PreS-слитых белков (n=19) в различные моменты времени (ось х). Средние значения с SD и достоверные различия указаны как *р<0,05, **р<0,01, ***р<0,001. Фиг.6В и фиг.6С: процент пролиферировавших CD4 (В) и CD8 (С) Т-клеток (ось y) после стимуляции PreS-пептидами (Р1-Р8), PreS или эквимолярной пептидной смесью (ось x) в образцах крови субъектов, иммунизированных вакцинной смесью PreS-слитых белков (n=11) в момент времени М2. Результаты представлены в виде средних значений с SD.
На фиг. 7 показано индуцированное антителами ингибирование вирусной инфекции гепатитом В в анализе нейтрализации вируса in vitro, который основан на in vitro культивируемых клетках печени. Процент ингибирования инфекции гепатитом В культивированных HepG2-hNTCP (ось х) получают путем предварительной инкубации вируса с анти-сывороткой, содержащей нейтрализующие вирус антитела. Фиг. 7А: ингибирование вирусной инфекции Ма 18/7 (положительным контролем), сывороткой от получавшего плацебо человека, сывороткой от людей после иммунизации вакцинной смесью PreS-слитых белков (n=7), причем все субъекты не имели предварительной вакцинации от гепатита В. Фиг. 7В: ингибирование вирусной инфекции сывороткой кроликов, иммунизированных коммерческой вакциной против гепатита В (Engerix) или вакцинной смесью PreS-слитых белков.
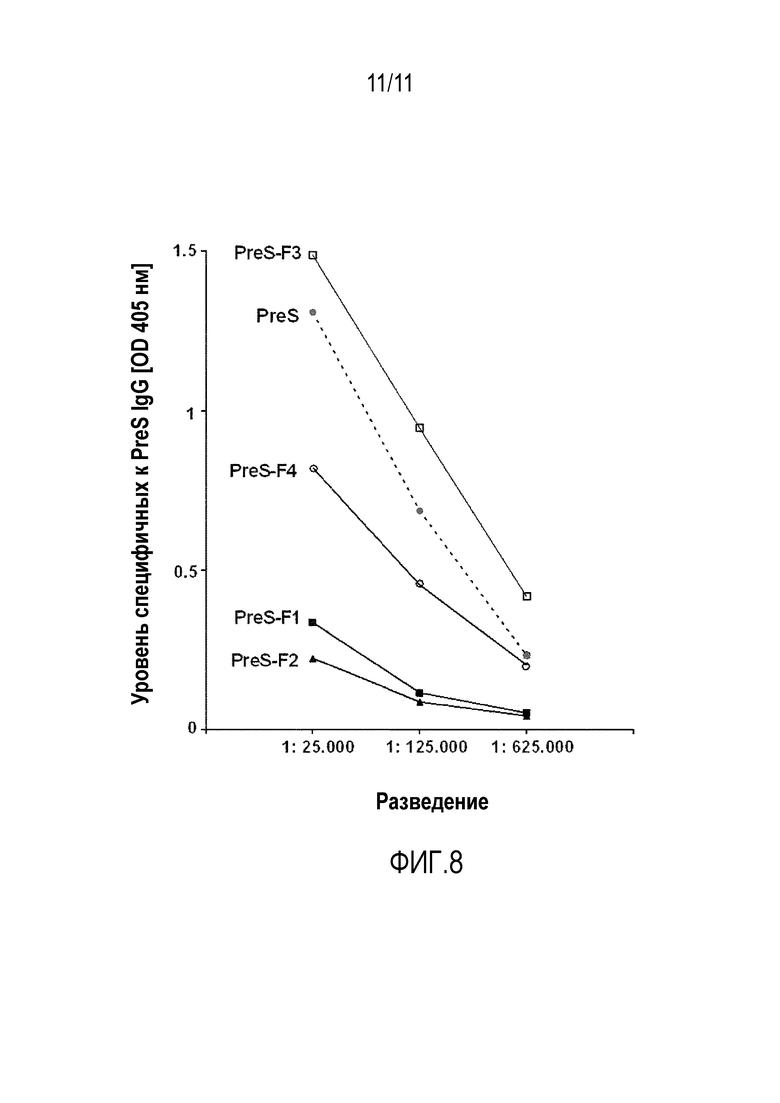
На фиг.8 показано сравнение общего сывороточного IgG к PreS в сыворотке новозеландских белых кроликов (NZW), иммунизированных либо рекомбинантным PreS, либо PreS-слитыми белками (PreS-F1 - PreS-F4), в виде эмульсии в полном адъюванте Фрейнда. На оси x показано разведение сывороток, а на оси y показаны значения оптической плотности, измеренные при 405 нм. Эксперимент проводили в дупликатах.
ПРИМЕРЫ:
Пример 1: Экспрессия и очистка рекомбинантного PreS, синтез перекрывающихся пептидов PreS, выравнивание последовательностей
Экспрессию и очистку меченого гексагистидиновой меткой рекомбинантного белка PreS (PreS1+PreS2 (SEQ ID Nо. 5; генотип A; подтип adw2, получен из GenBank: AAT28735.1) в Escherichia coli BL21 (DE3, Stratagene, США) выполняли, как описано у Niespodziana K. и др. (J Allergy Clin Immunol 127 (2011): 1562-70).
Восемь пептидов длиной приблизительно 30 аминокислот и перекрывающихся на 10 аминокислот, охватывающих полную последовательность PreS (генотип A, подтип adw2; таблица А; фиг. 1), были синтезированы с помощью Fmoc (9-флуоренилметоксикарбонил)-стратегии с активацией HBTU [2-(1H-бензотриазол-1-ил)-1,1,3,3-тетраметилуронийгексафторфосфатом] (СЕМ-Liberty, Matthews, NC; Applied Biosystems, Life Technologies, USA).
Таблица A:
(подчеркнуты перекрывающиеся области пептидов)
Пептиды очищали с помощью препаративной ВЭЖХ и их идентичность подтверждали масс-спектрометрически (Microflex MALDI-TOF, Bruker, USA).
Выравнивание последовательности PreS генотипа А серотипа adw2 и ее пептидных последовательностей с генотипами В-Н HBV было осуществлено с помощью CLUSTAL W с использованием эталонных последовательностей HBV из базы данных (HBVdb: https://hbvdb.ibcp/HBVdb/HBVdbIndex) (Hayer J et al. Nucleic Acids Res 2012; gksl 022) (см. фиг. 1).
Пример 2: Иммунизация кроликов
Кроличьи антитела, специфичные к рекомбинантному PreS, были получены иммунизацией белого новозеландского кролика очищенным PreS (200 мкг на инъекцию) с использованием полного адъювант Фрейнда (CFA) для первой и неполного адъюванта Фрейнда (IFA) для второй и третьей инъекций (Charles River, Germany). Кроме того, белых новозеландских кроликов иммунизировали три раза смесью, содержащей 20 мкг (n=2) или 40 мкг (n=2), каждого из четырех PreS-компонентов вакцинной смеси (PreS-вакцинная смесь-20/PreS-вакцинная смесь-40) с использованием Al(OH)3 в качестве адъюванта. Четыре компонента PreS-вакцинной смеси включают PreS-слитые белки PreSF1, PreSF2, PreSF3 и PreSF4, имеющие следующие аминокислотные последовательности:
PreSF1 (SEQ ID Nо. 22)
MVRYTTEGGTKTEAEDVIPEGWKADTSYESKVRYTTEGGTKTEAEDVIPEGWKADT SYESKGGWSSKPRKGMGTNLSVPNPLGFFPDHQLDPAFGANSNNPDWDFNPIKDHW PAANQVGVGAFGPGLTPPHGGILGWSPQAQGILTTVSTIPPPASTNRQSGRQPTPI SPPLRDSHPQAMQWNSTAFHQALQDPRVRGLYFPAGGSSSGTVNPAPNIASHISSI SARTGDPVTNVRYTTEGGTKTEAEDVIPEGWKADTSYESKVRYTTEGGTKTEAEDV IPEGWKADTSYESK
PreSF2 (SEQ ID Nо. 23):
MFRFLTEKGMKNVFDDVVPEKYTIGATYAPEEFRFLTEKGMKNVFDDVVPEKYTIG ATYAPEEGGWSSKPRKGMGTNLSVPNPLGFFPDHQLDPAFGANSNNPDWDFNPIKD HWPAANQVGVGAFGPGLTPPHGGILGWSPQAQGILTTVSTIPPPASTNRQSGRQPT PISPPLRDSHPQAMQWNSTAFHQALQDPRVRGLYFPAGGSSSGTVNPAPNIASHIS SISARTGDPVTNFRFLTEKGMKNVFDDVVPEKYTIGATYAPEEFRFLTEKGMKNVF DDVVPEKYTIGATYAPEE
PreSF3 (SEQ ID Nо. 6):
MEAAFNDAIKASTGGAYESYKFIPALEAAVKAEEVKVIPAGELQVIEKVDAAFKVA ATAANAAPANDKGGWSSKPRKGMGTNLSVPNPLGFFPDHQLDPAFGANSNNPDWDF NPIKDHWPAANQVGVGAFGPGLTPPHGGILGWSPQAQGILTTVSTIPPPASTNRQS GRQPTPISPPLRDSHPQAMQWNSTAFHQALQDPRVRGLYFPAGGSSSGTVNPAPNI ASHISSISARTGDPVTNADLGYGPATPAAPAAGYTPATPAAPAEAAPAGKATTEEQ KLIEKINAGFKAALAAAAGVQPADKYR
PreSF4 (SEQ ID Nо. 24):
MGKATTEEQKLIEDVNASFRAAMATTANVPPADKGKATTEEQKLIEDVNASFRAAM ATTANVPPADKGGWSSKPRKGMGTNLSVPNPLGFFPDHQLDPAFGANSNNPDWDFN PIKDHWPAANQVGVGAFGPGLTPPHGGILGWSPQAQGILTTVSTIPPPASTNRQSG RQPTPISPPLRDSHPQAMQWNSTAFHQALQDPRVRGLYFPAGGSSSGTVNPAPNIA SHISSISARTGDPVTNGKATTEEQKLIEDVNASFRAAMATTANVPPADKGKATTEE QKLIEDVNASFRAAMATTANVPPADK
Кроме того, специфичные к зарегистрированной вакцине против гепатита B (ENGERIX-B) кроличьи антитела были получены трехкратной иммунизацией новозеландских белых кроликов (n=2) с использованием коммерчески доступных и готовых к использованию предварительно заполненных шприцев с интервалом в один месяц.
Образцы сыворотки отбирали перед иммунизацией и приблизительно через четыре недели после третьей иммунизации и хранили при -20°С до проведения анализа.
Иммунизация PreS-вакцинной смесью показала индукцию образования IgG-антител со специфичностью к последовательным эпитопам в PreS. На фиг. 2 показано сравнение выработки IgG (IgG-ответа) против PreS и синтетических пептидов-производных PreS, индуцируемых у кроликов при использовании PreS в CFA или сорбированной на гидроксиде алюминия PreS-вакцинной смеси (фиг. 2В). Кроличьи антитела, индуцированные с использованием PreS в CFA, распознавали PreS и каждый из пептидов-производных PreS, за исключением Р7 (фиг. 2А). Сорбированная на гидроксиде алюминия PreS-вакцинная смесь в дозировке 20 мкг вызывала образование PreS-специфичных IgG-антител и IgG-антител, направленных главным образом на N-концевые пептиды P1, P2, пептид P6 и на С-концевой пептид P8 (фиг. 2В). Никаких PreS или пептид-специфичных IgG-антител не было найдено у кроликов до иммунизации (фиг. 2, левые столбики).
Пример 3: Оценка специфичных к PreS и PreS-пептидам гуморальных иммунных ответов
Образцы сыворотки были получены от пациентов, которые получили три инъекции сорбированной на Al(OH)3 PreS-вакцинной смеси (т.е. смеси из 10, 20 или 40 мкг каждого компонента PreS-вакцинной смеси или плацебо, т.е. Al(OH)3). Сыворотку собирали перед иммунизацией и через четыре недели после третьей иммунизации и хранили при -20°С до использования. Второй набор образцов сыворотки получали от пациентов, которых лечили в течение двух лет 7 подкожными инъекциями сорбированной на Al(OH)3 PreS-вакцинной смеси (т.е. смеси из 20 или 40 мкг каждого компонента PreS-вакцинной смеси или плацебо, т.е. Al(OH)3). Кроме того, образцы сыворотки были получены от пациентов, страдающих гепатитом В, которым был поставлен диагноз на основе клинических данных, тестирования функции печени и сывороточных маркеров HBV.
Все сыворотки были проанализированы, скринированы на серологические маркеры HBV (а именно, поверхностный антиген гепатита В [HBsAg], а также антитела к ядерному антигену гепатита В [анти-НВс]).
ИФА-планшеты (Nunc MaxiSorp®, Denmark) покрывали антигенами (рекомбинантным PreS, синтетическими перекрывающимися пептидами PreS: P1-P8) или сывороточным альбумином человека (отрицательный контроль) (Behring, USA). Инкубацию проводили с кроличьей сывороткой в разведении 1:10000 (CFA) или 1:500 (PreS-вакцинная смесь-20/PreS-вакциная смесь-40), с мышиной сывороткой в разведении 1:1000 и с сывороткой человека, разбавленной по-разному для изотипов и подклассов IgG. Для обнаружения общего IgG человека, сыворотку разводили 1:100, для IgA, IgG1, IgG2, IgG3, IgG4, а также для IgM - сыворотку разводили 1:20, а для обнаружения IgE-антител - сыворотку разводили в соотношении 1:10.
Кроличьи IgG детектировали конъюгированными с пероксидазой хрена ослиными антителами к IgG кролика в разведении 1:2500 (GE Healthcare, Buckinghamshire, Great Britain). Связанные мышиные IgG1 детектировали моноклональными крысиными антителами к мышиным IgG1 (BD Pharmingen, USA) в разведении 1:1000, а затем конъюгированными с пероксидазой хрена козьими антителами к крысиным IgG (Amersham Bioscience, Sweden) в разведении 1:2500.
IgG человека детектировали кроличьими антителами, специфичными к Fc области IgG человека (Jackson-Dianova, Germany), разведенным 1:10000, а затем конъюгированными с пероксидазой ослиными антителами к кроличьим IgG (GE Healthcare) в разведении 1:2500. IgA, IgG-подклассы IgG1, IgG2 и IgG4, а также IgM человека, детектировали очищенными мышиными антителами к антителам IgA1/IgA2, IgG1, IgG2, IgG4 и IgM человека (BD Pharmingen), разведенными 1:1000, соответственно, с последующей детекцией связанными с пероксидазой овечьими антителами к мышиным IgG (GE Healthcare) в разведении 1:2500. Моноклональные антитела к IgG3 человека (Sigma Aldrich, USA) разводили 1:5000. IgE человека детектировали конъюгированными с пероксидазой хрена козьими антителами к IgE человека (KPL, USA).
Пример 4: на выработку PreS-специфичных антител у иммунизированных PreS-вакцинными смесями субъектов не влиял предшествующий иммунитет к гепатиту В
Образцы сыворотки от людей, получавших иммунотерапию PreS-вакцинной смесью или плацебо, были протестированы на присутствие IgG к PreS и синтетическим пептидам PreS (фиг. 3а-3с). Эти пациенты (n=30) были обследованы на специфичные сывороточные маркеры гепатита B (HBsAg, анти-HBs и анти-НВс антитела) до лечения и получены отрицательные результаты по HBsAg и анти-НВс антителам. После предшествующей вакцинации вакциной против гепатита В, двадцать два субъекта имели анти-HBs антитела (фиг. 3а-3с). Было обнаружено, что у каждого из пациентов, получавших иммунотерапию PreS-вакцинной смесью (вне зависимости от того, были ли они вакцинированы от гепатита В перед этим или нет), но не пациенты, получавшие плацебо, индуцировалась выработка PreS-специфичных IgG при тестировании сыворотки после третьей (точка V8, через три месяца после первой инъекции), а также после седьмой инъекции (точка V15, через 15 месяцев после первой инъекции) (фиг.3а-3с). Выработка PreS-специфичных IgG достоверно возросла по сравнению с базовым уровнем до иммунотерапии (то есть, в точке V8 относительно V5) и дополнительно значительно увеличилась в интервале между V8 и V15 (то есть, после седьмой инъекции) (фиг. 3а-3с). PreS-специфичные IgG у этих пациентов были направлены, главным образом, на N-концевые пептиды P1, P2 и P3, и аналогичным образок количество P1- и P2-специфичных IgG значительно увеличилось относительно базового уровня (от V5 до V8 и от V8 до V15) (фиг. 3а 3с). Кроме того, повышение уровней IgG против других пептидов-производных PreS Р4, Р5, Р6, Р7 и Р8 было обнаружено в сыворотке крови у пациентов, получавших иммунотерапию PreS-вакцинной смесью, но не у пациентов, получавших плацебо (фиг. 3а-3с).
Пример 5: вырабатываемые PreS-специфичные антитела у иммунизированных PreS-вакцинной смесью субъектов направлены против нейтрализующих эпитопов и отличаются от таковых у инфицированных гепатитом B индивидуумов
На фиг. 4 показано сравнение изотипов и IgG-подклассов PreS-специфичных антител у пациентов в ответ на иммунотерапию с использованием PreS-вакцинной смеси или плацебо с таковыми у инфицированных гепатитом В индивидуумов. Иммунотерапия обеими дозами PreS-вакцинной смеси индуцировала сильный PreS-специфичный IgG-ответ у каждого из пациентов, который был значительно выше, чем IgG-ответ у инфицированных гепатитом В индивидуумов (фиг. 4). Никакого значимого присутствия PreS-специфичных IgA, IgE или IgM не было обнаружено в сыворотках пациентов, которых лечили PreS-вакцинной смесью или плацебо, а также у инфицированных гепатитом В индивидуумов (фиг. 4). Подкласс PreS-специфичных IgG отличался у иммунизированных PreS-вакцинной смесью субъектов и у инфицированных гепатитом В индивидуумов. Получавшие PreS-вакцинную смесь субъекты показывали преимущественно IgG1- и IgG4-ответы на PreS, тогда как у инфицированных гепатитом В индивидуумов вырабатывались антитела IgG1 и IgG2 (фиг. 4).
Также были обнаружены значительные различия в отношении эпитопной специфичности PreS-специфичных антител у иммунизированных PreS-вакцинной смесью пациентов относительно инфицированных гепатитом B индивидуумов (фиг. 5). Иммунизированные PreS-вакцинной смесью пациенты, но не инфицированные гепатитом B индивидуумы, давали сильные IgG-ответы на Р1 и Р3 (фиг. 5). Это открытие оказалось неожиданным, поскольку область, занимаемая P1, соответствует мотиву в PreS1 (см. также фиг. 1), который, как сообщалось, содержит важные остатки для ингибирования инфекции гепатитом B. Кроме того, Р7 распознавался только у иммунизированных PreS-вакцинной смесью субъектов, но не у инфицированных гепатитом В индивидуумов, тогда как IgG к P2 и P6 были также найдены у инфицированных гепатитом В индивидуумов (фиг. 5).
Пример 6: оценка Т-клеточных ответов
Мононуклеарные клетки периферической крови (РВМС) получали из гепаринизированных образцов крови центрифугированием в градиенте плотности, получаемом с помощью фиколла (Ficoll, Amersham Biosciences, Sweden). После получения образцов крови определяли PreS-специфичную пролиферацию РВМС у иммунизированных PreS-вакцинной смесью субъектов (n=19) в точках V5, V8, M1 (5 месяцев после первой вакцинации) и М2 (через 17 месяцев после первой вакцинации) по включению [3Н]-тимидина.
Для некоторых иммунизированных PreS-вакцинной смесью пациентов (n=11) можно было оценить CD4+ и CD8+ Т-клеточные ответы в М2 путем мечения сукцинимидильным эфиром карбоксифлуоресцеина (CFSE).
Меченые флуоресцентным красителем клетки высевали по 200000 клеток/лунку в бессывороточной среде Ultra culture™ (Lonza, Belgium) с добавлением 2 ммоль/л L-глутамина (Sigma Aldrich, США), 50 ммоль/л β-меркаптоэтанола (Sigma Aldrich) и 0,02 мг гентамицина на миллилитр (Sigma Aldrich) в общем объеме 200 мкл в 96-луночных микропланшетах с U-образным дном (Termo Fisher, USA). Клетки либо были оставлены нестимулированными (отрицательный контроль), либо были стимулированы Dynabeads® Human T-Activator CD3/CD28 (3 мкг/лунку (Invitrogen, USA)) в качестве положительного контроля, PreS (0,15 мкг/лунку), эквимолярными количествами PreS-перекрывающихся пептидов (0,03 мкг/лунку) или смесью PreS-перекрывающихся пептидов, содержащей по 0,03 мкг/лунку каждого пептида, и далее клетки культивировали при 37°С в 5% CO2 в течение 7 дней перед проведением окрашивания антителами и FACS-анализом.
Для проточной цитометрии были использованы следующие реактивы: PerCP/Cy5.5-антитело против СD3 человека (клон HIT3a), Brilliant Violet 421™-антитело против CD4 человека (клон RPA-Т4), APC-антитело против CD8a человека (клон HIT8a), а также контроли изотипа, а именно PerCP/Cy5.5-мышиные IgG2а, Brilliant Violet 421™-мышиные IgG1, APC-мышиные IgG1 (BioLegend, USA) и подходящий для фиксирования клеток краситель жизнеспособности клеток eFluor® 780 (eBioscience, USA).
Проточную цитометрию проводили на BD FACS Canto II (Becton, Dickinson and Company, USA). Для каждого образца регистрировали двадцать тысяч событий, и анализ проводили с помощью программного обеспечения FlowJo, Version 10. Лимфоциты отбирали в соответствии с морфологическими критериями на точечном графике прямого и бокового рассеяния, мертвые клетки исключали окрашиванием красителем жизнеспособности клеток, и последующий отбор был сфокусирован на CD3CD4 и CD3CD8-положительных Т-клетках. Те клетки, которые пролиферировали в ответ на стимуляцию антигеном были идентифицированы по снижению интенсивности флуоресценции CFSE в них. Результаты представляют собой среднее из трипликатов культур, и стимуляция CD3+CD4+ и CD3+CD8+ на 235 медианных процента выше фона была показана для различных антигенов и анализируемых пациентов.
На фиг. 6 показано развитие PreS-специфичных Т-клеточных ответов у больных, получавших иммунотерапию со PreS-вакцинной смесью. Был обнаружен постепенно увеличивающийся PreS-специфичный Т-клеточный ответ, который был значительно выше в точках V8, M1 и М2 по сравнению с исходным уровнем в точке V5 (фиг. 6А). При анализе эпитопной специфичности PreS-специфичных CD4-клеточных ответов с помощью CFSE-окрашивания авторы обнаружили, что P1, Р2, Р5 и Р6 вызывали самую сильную пролиферация клеток CD4, хотя ответы CD4 в отношении P3, P4 и Р7 также были обнаружены (фиг. 6В). Интересно, что пептиды и смесь пептидов индуцировали пролиферацию CD4-клеток сильнее, чем белок PreS (фиг. 6В). Хотя и с низкой частотой, но был обнаружен некоторый PreS- и PreS-пептид-специфичный CD8-клеточный ответ, который в основном был направлен на Р2, Р3, Р6 и Р8 и полноразмерный PreS (фиг. 6В).
Пример 7: анализы нейтрализации вируса гепатита B
Инокулят HBV для заражения получали из супернатантов клеток HepAd38 с использованием гепариновой колонки (GE Healthcare, Great Brtain) для выделения вирусных частиц. Клетки HepG2-hNTCP высевали с плотностью 3×105 клеток/лунку в 24-луночный планшет. На второй день после посева в инфекционную среду (DMEM, Invitrogen, USA) добавляли 2,5% DMSO (Merck, Germany), и на третий день клетки инфицировали HBV. Для нейтрализации частиц вируса гепатита В, сыворотку от пациентов (10 мкл) предварительно инкубировали с HBV-инокулятом (6,9×107 геномных эквивалентов (GE)/лунку) в течение 30 минут при 37°С с последующей совместной инкубацией клеток с сывороткой пациентов и вирусом в присутствии 4%-го полиэтиленгликоля 800 (Sigma Aldrich, USA) в течение 16 часов при 37°С. Нейтрализующие моноклональные антитела Ma18/721 были использованы в качестве положительного контроля.
Через 16 часов после инокуляции, клетки хорошо промывали PBS и добавляли свежую среду для дифференцировки с добавлением 2,5% DMSO (Invitrogen). Дополнительную смену среды проводили на третий и пятый день после заражения.
Количественную оценку инфекции HBV проводили путем измерения секретируемого антигена «е» гепатита В (HBeAg) в супернатанте клеток на 5-7 день после заражения. HBeAg определяли с помощью автоматизированной хемилюминесцентной системы ADVIA Centaur XPT (Siemens, Germany). Образцы считались положительными при сигнале выше 1-го индекса.
Экспрессию ядерного белка HBV детектировали специфичной иммунофлуоресценцией. Супернатант удаляли и клетки промывали PBS перед фиксацией 4%-м параформальдегидом (Sigma Aldrich) в течение 30 минут при комнатной температуре (RT). Затем клетки промывали PBS после чего пермеабилизовали 0,25%-м Triton X 100 (AppliChem GmbH, Germany) в PBS в течение 30 минут при комнатной температуре. Затем клетки инкубировали в течение ночи при 4°С с первичными антителами (кроличьими поликлональными антителами к ядерному белку HBV, DAKO Deutschland GmbH, Hamburg, Germany), разведенными в 2%-м (вес/объем) BSA в PBS. На следующий день клетки промывали PBS и в конце инкубировали со вторичными антителами (конъюгированными с Alexa 488 козьими антителами против кроличьих антител; Invitrogen, Carlsbad, CA) и с 4',6-диамидин-2-фенилиндо/Хекст 33342 (Roche Applied Science, Germany) в темноте. Для детекции ядерного белка HBV вторичное антитело инкубировали в течение 2 часов при комнатной температуре в защищенном от света месте. Клетки исследовали под флуоресцентным микроскопом, используя возбуждение при 488 нм для меченых Alexa 488 вторичных антител (Invitrogen, Carlsbad, CA) и при 360 нм для окрашивания ядер.
В первом типе анализа экспрессию ядерного антигена гепатита В (HBсAg) после инфицирования клеток детектируют с помощью специфичной иммунофлуоресценции. HBсAg не был обнаружен в незараженных клетках, кроме зараженных и необработанных клеток, и эта экспрессия может быть предотвращена предварительной инкубацией вируса с нейтрализующим моноклональным антителом Ma18/721, который направлен против PreS1-домена большого поверхностного белка вируса гепатита В. Аналогичным образом было установлено, что предварительная инкубация вируса гепатита В с кроличьими антителами, индуцированными коммерческой вакциной Engerix-B, или с кроличьими антителами, индуцированными PreS-вакцинной смесью (в дозировке 20 мкг), ингибирует инфицирование клеток HepG2-hNTCP. Аналогичный набор экспериментов проводили с сыворотками от получавших PreS-вакцинную смесь или плацебо пациентов. Сыворотка, полученная от пациента перед и после иммунизации плацебо, не ингибировала инфекцию клеток HepG2-hNTCP, тогда как сыворотка, полученная от пациента после иммунизации 20 мкг или от пациента после иммунизации 40 мкг PreS-вакцинной смеси, ингибировала инфекцию клеток HepG2-hNTCP.
В дополнении к окрашиванию HBcAg использовали анализ на основе измерения секретируемого антигена е вируса гепатита В (HBeAg) клетками HepG2-hNTCP через семь дней после инфицирования HBV качестве еще одного суррогатного маркера для количественного определения ингибирования HBV-инфекции. Было обнаружено, что сыворотка после иммунизации PreS-вакцинной смесью ингибировала инфекцию HBV на 50-99% (фиг. 7А). Никакой значимой зависимости от дозы и числа инъекций PreS-вакцинной смеси не было обнаружено, поскольку схожее ингибирование наблюдали для сывороток от пациентов, которые получили по три инъекции (фиг. 7а), а также для сывороток от пациентов, которые получили по семь инъекций (фиг. 7A, черный цвет). Кроме того, не было явных отличий в отношении степени ингибирования между пациентами, которые получили дозы 20 мкг или 40 мкг PreS-вакцинной смеси (фиг. 7А). Никакого ингибирования не наблюдалось для сыворотки от получавшего плацебо пациента, и более чем 90%-е ингибирование наблюдалось для моноклонального антитела Ма 18/7 (фиг. 7А). Кроличьи антитела после иммунизации Engerix-B и PreS-вакцинной смесью вызвали более чем 99%-е ингибирование инфекции HBV (фиг. 7B).
Пример 8:
Рекомбинантным PreS и человеческим сывороточным альбумином (Behring, USA) в качестве отрицательного контроля покрывали микропланшеты Nunc Maxisorb (Thermo-Fisher Scientific, USA) в концентрации 2 мкг/мл в 100 мМ натрий-фосфатном буфере, рН 9,6, в течение ночи при 4°С. Промывочный буфер состоял из PBS, 0,05%-го (объем/объем) Tween 20 (PBS/Т), и процедуру блокирования неспецифичных сайтов связывания проводили с использованием 2%-го (вес/объем) BSA в PBS/T в течение 2 часов при температуре 37°С. Все последующие разведения сыворотки и реагентов проводили в 0,5%-м (вес/объем) BSA в PBS/T.
Для определения гуморального иммунного ответа у кроликов, прошедших полный курс иммунизации рекомбинантным PreS или PreS-слитыми белками в виде эмульсии в полном адъюванте Фрейнда (CFA), использовали сыворотки в различных разведениях (4°С, в течение ночи), и связавшиеся общие IgG кролика детектировали конъюгированными с пероксидазой хрена ослиными антителами против кроличьих IgG (GE Healthcare, Great Britain), разведенными 1:2000. Цветную реакцию индуцировали ABTS [2,2'-азино-бис(3-этилбензотиазолин-6-сульфоновой кислотой] и детекцию поглощения, соответствующего уровню антиген-специфичных антител, проводили при 405 нм и 490 нм с использованием микропланшетного спектрофотометра (Molecular Devices, USA). Все измерения были выполнены в трех повторах.
Удивительным оказалось, что только слитый белок, содержащий один или несколько пептидов, имеющих аминокислотные последовательности SEQ ID Nо. 1, SEQ ID Nо. 2, SEQ ID NO. 3 и/или SEQ ID NO. 4, и PreS (PreS-F3), был способен вызвать образование PreS-специфичных IgG в гораздо большей степени по сравнению с одним только PreS или другими слитыми белками, также содержащими PreS, слитый с различными пептидами (PreS-F1, PreS-F2, PreS-F4), как показано на фиг. 8.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Biomay AG
<120> Слитый белок
<130> 47640
<150> EP 15183983.4
<151> 2015-09-05
<160> 24
<170> PatentIn version 3.5
<210> 1
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственный пептид
<400> 1
Glu Ala Ala Phe Asn Asp Ala Ile Lys Ala Ser Thr Gly Gly Ala Tyr
1 5 10 15
Glu Ser Tyr Lys Phe Ile Pro Ala Leu Glu Ala Ala Val Lys
20 25 30
<210> 2
<211> 37
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственный пептид
<400> 2
Ala Glu Glu Val Lys Val Ile Pro Ala Gly Glu Leu Gln Val Ile Glu
1 5 10 15
Lys Val Asp Ala Ala Phe Lys Val Ala Ala Thr Ala Ala Asn Ala Ala
20 25 30
Pro Ala Asn Asp Lys
35
<210> 3
<211> 33
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственный пептид
<400> 3
Ala Asp Leu Gly Tyr Gly Pro Ala Thr Pro Ala Ala Pro Ala Ala Gly
1 5 10 15
Tyr Thr Pro Ala Thr Pro Ala Ala Pro Ala Glu Ala Ala Pro Ala Gly
20 25 30
Lys
<210> 4
<211> 33
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственный пептид
<400> 4
Ala Thr Thr Glu Glu Gln Lys Leu Ile Glu Lys Ile Asn Ala Gly Phe
1 5 10 15
Lys Ala Ala Leu Ala Ala Ala Ala Gly Val Gln Pro Ala Asp Lys Tyr
20 25 30
Arg
<210> 5
<211> 173
<212> Белок
<213> Вирус гепатита В
<400> 5
Gly Gly Trp Ser Ser Lys Pro Arg Lys Gly Met Gly Thr Asn Leu Ser
1 5 10 15
Val Pro Asn Pro Leu Gly Phe Phe Pro Asp His Gln Leu Asp Pro Ala
20 25 30
Phe Gly Ala Asn Ser Asn Asn Pro Asp Trp Asp Phe Asn Pro Ile Lys
35 40 45
Asp His Trp Pro Ala Ala Asn Gln Val Gly Val Gly Ala Phe Gly Pro
50 55 60
Gly Leu Thr Pro Pro His Gly Gly Ile Leu Gly Trp Ser Pro Gln Ala
65 70 75 80
Gln Gly Ile Leu Thr Thr Val Ser Thr Ile Pro Pro Pro Ala Ser Thr
85 90 95
Asn Arg Gln Ser Gly Arg Gln Pro Thr Pro Ile Ser Pro Pro Leu Arg
100 105 110
Asp Ser His Pro Gln Ala Met Gln Trp Asn Ser Thr Ala Phe His Gln
115 120 125
Ala Leu Gln Asp Pro Arg Val Arg Gly Leu Tyr Phe Pro Ala Gly Gly
130 135 140
Ser Ser Ser Gly Thr Val Asn Pro Ala Pro Asn Ile Ala Ser His Ile
145 150 155 160
Ser Ser Ile Ser Ala Arg Thr Gly Asp Pro Val Thr Asn
165 170
<210> 6
<211> 307
<212> Белок
<213> Искусственная последовательность
<220>
<223> PreSF3
<400> 6
Met Glu Ala Ala Phe Asn Asp Ala Ile Lys Ala Ser Thr Gly Gly Ala
1 5 10 15
Tyr Glu Ser Tyr Lys Phe Ile Pro Ala Leu Glu Ala Ala Val Lys Ala
20 25 30
Glu Glu Val Lys Val Ile Pro Ala Gly Glu Leu Gln Val Ile Glu Lys
35 40 45
Val Asp Ala Ala Phe Lys Val Ala Ala Thr Ala Ala Asn Ala Ala Pro
50 55 60
Ala Asn Asp Lys Gly Gly Trp Ser Ser Lys Pro Arg Lys Gly Met Gly
65 70 75 80
Thr Asn Leu Ser Val Pro Asn Pro Leu Gly Phe Phe Pro Asp His Gln
85 90 95
Leu Asp Pro Ala Phe Gly Ala Asn Ser Asn Asn Pro Asp Trp Asp Phe
100 105 110
Asn Pro Ile Lys Asp His Trp Pro Ala Ala Asn Gln Val Gly Val Gly
115 120 125
Ala Phe Gly Pro Gly Leu Thr Pro Pro His Gly Gly Ile Leu Gly Trp
130 135 140
Ser Pro Gln Ala Gln Gly Ile Leu Thr Thr Val Ser Thr Ile Pro Pro
145 150 155 160
Pro Ala Ser Thr Asn Arg Gln Ser Gly Arg Gln Pro Thr Pro Ile Ser
165 170 175
Pro Pro Leu Arg Asp Ser His Pro Gln Ala Met Gln Trp Asn Ser Thr
180 185 190
Ala Phe His Gln Ala Leu Gln Asp Pro Arg Val Arg Gly Leu Tyr Phe
195 200 205
Pro Ala Gly Gly Ser Ser Ser Gly Thr Val Asn Pro Ala Pro Asn Ile
210 215 220
Ala Ser His Ile Ser Ser Ile Ser Ala Arg Thr Gly Asp Pro Val Thr
225 230 235 240
Asn Ala Asp Leu Gly Tyr Gly Pro Ala Thr Pro Ala Ala Pro Ala Ala
245 250 255
Gly Tyr Thr Pro Ala Thr Pro Ala Ala Pro Ala Glu Ala Ala Pro Ala
260 265 270
Gly Lys Ala Thr Thr Glu Glu Gln Lys Leu Ile Glu Lys Ile Asn Ala
275 280 285
Gly Phe Lys Ala Ala Leu Ala Ala Ala Ala Gly Val Gln Pro Ala Asp
290 295 300
Lys Tyr Arg
305
<210> 7
<211> 173
<212> Белок
<213> Вирус гепатита В
<400> 7
Gly Gly Trp Ser Ser Lys Pro Arg Lys Gly Met Gly Thr Asn Leu Ser
1 5 10 15
Val Pro Asn Pro Leu Gly Phe Phe Pro Asp His Gln Leu Asp Pro Ala
20 25 30
Phe Lys Ala Asn Ser Asp Asn Pro Asp Trp Asp Leu Asn Pro His Lys
35 40 45
Asp Asn Trp Pro Asp Ser Asn Lys Val Gly Val Gly Ala Phe Gly Leu
50 55 60
Gly Phe Thr Pro Pro His Gly Gly Leu Leu Gly Trp Ser Pro Gln Ala
65 70 75 80
Gln Gly Ile Leu Thr Thr Val Pro Ala Ala Pro Pro Pro Ala Ser Thr
85 90 95
Asn Arg Gln Val Ala Arg Pro Pro Thr Pro Leu Ser Pro Pro Leu Arg
100 105 110
Asp Thr His Pro Gln Ala Met Gln Trp Asn Ser Thr Thr Phe His Gln
115 120 125
Thr Leu Gln Asp Pro Arg Val Arg Ala Leu Tyr Phe Pro Ala Gly Gly
130 135 140
Ser Ser Ser Gly Thr Val Asn Pro Val Gln Asn Thr Ala Ser Ser Ile
145 150 155 160
Ser Ser Ile Leu Ser Lys Thr Gly Asp Pro Val Pro Asn
165 170
<210> 8
<211> 173
<212> Белок
<213> Вирус гепатита В
<400> 8
Gly Gly Tyr Ser Ser Lys Pro Arg Lys Gly Met Gly Thr Asn Leu Ser
1 5 10 15
Val Pro Asn Pro Leu Gly Phe Leu Pro Asp His Gln Leu Asp Pro Ala
20 25 30
Phe Gly Ala Asn Ser Asn Asn Pro Asp Trp Asp Phe Asn Pro Asn Lys
35 40 45
Asp Pro Trp Pro Glu Ala Trp Gln Val Gly Val Gly Ala Phe Gly Pro
50 55 60
Gly Phe Thr Pro Pro His Gly Gly Leu Leu Gly Trp Ser Pro Gln Ala
65 70 75 80
Gln Gly Ile Leu Thr Pro Val Pro Ala Ala Pro Pro Pro Ala Ser Thr
85 90 95
Asn Arg Gln Ser Gly Arg Gln Pro Thr Pro Ile Ser Pro Pro Leu Arg
100 105 110
Asp Ser His Pro Gln Ala Met Gln Trp Asn Ser Thr Thr Phe His Gln
115 120 125
Ala Leu Leu Asp Pro Arg Val Arg Gly Leu Tyr Phe Pro Ala Gly Gly
130 135 140
Ser Ser Ser Gly Thr Thr Asn Pro Val Pro Thr Thr Ala Ser Pro Ile
145 150 155 160
Ser Ser Ile Phe Ser Arg Thr Gly Asp Pro Ala Pro Asn
165 170
<210> 9
<211> 162
<212> Белок
<213> Вирус гепатита В
<400> 9
Gly Gln Asn Leu Ser Thr Ser Asn Pro Leu Gly Phe Phe Pro Asp His
1 5 10 15
Gln Leu Asp Pro Ala Phe Arg Ala Asn Thr Ala Asn Pro Asp Trp Asp
20 25 30
Phe Asn Pro Asn Lys Asp Thr Trp Pro Asp Ala Asn Lys Val Gly Ala
35 40 45
Gly Ala Phe Gly Leu Gly Phe Thr Pro Pro His Gly Gly Leu Leu Gly
50 55 60
Trp Ser Pro Gln Ala Gln Gly Ile Leu Gln Thr Leu Pro Thr Asn Pro
65 70 75 80
Pro Pro Ala Ser Thr Asn Arg Gln Ser Gly Arg Gln Pro Thr Pro Leu
85 90 95
Ser Pro Pro Leu Arg Asn Thr His Pro Gln Ala Met Gln Trp Asn Ser
100 105 110
Thr Thr Phe His Gln Thr Leu Gln Asp Pro Arg Val Arg Gly Leu Tyr
115 120 125
Phe Pro Ala Gly Gly Ser Ser Ser Gly Thr Val Asn Pro Val Pro Thr
130 135 140
Thr Val Ser His Ile Ser Ser Ile Phe Ser Arg Ile Gly Asp Pro Ala
145 150 155 160
Leu Asn
<210> 10
<211> 172
<212> Белок
<213> Вирус гепатита В
<400> 10
Gly Leu Ser Trp Thr Val Pro Leu Glu Trp Gly Lys Asn His Ser Thr
1 5 10 15
Thr Asn Pro Leu Gly Phe Phe Pro Asp His Gln Leu Asp Pro Ala Phe
20 25 30
Arg Ala Asn Thr Arg Asn Pro Asp Trp Asp His Asn Pro Asn Lys Asp
35 40 45
His Trp Thr Ala Ala Asn Lys Val Gly Val Gly Ala Phe Gly Pro Gly
50 55 60
Phe Thr Pro Pro His Gly Gly Leu Leu Gly Trp Ser Pro Gln Ala Gln
65 70 75 80
Gly Met Leu Lys Thr Leu Pro Ala Asp Pro Pro Pro Ala Ser Thr Asn
85 90 95
Arg Gln Ser Gly Arg Gln Pro Thr Pro Ile Thr Pro Pro Leu Arg Asp
100 105 110
Thr His Pro Gln Ala Met Gln Trp Asn Ser Thr Thr Phe His Gln Ala
115 120 125
Leu Gln Asp Pro Arg Val Arg Gly Leu Tyr Phe Pro Ala Gly Gly Ser
130 135 140
Ser Ser Gly Thr Val Asn Pro Val Pro Thr Thr Ala Ser Leu Ile Ser
145 150 155 160
Ser Ile Phe Ser Arg Ile Gly Asp Pro Ala Pro Asn
165 170
<210> 11
<211> 173
<212> Белок
<213> Вирус гепатита В
<400> 11
Gly Ala Pro Leu Ser Thr Thr Arg Arg Gly Met Gly Gln Asn Leu Ser
1 5 10 15
Val Pro Asn Pro Leu Gly Phe Leu Pro Asp His Gln Leu Asp Pro Leu
20 25 30
Phe Arg Ala Asn Ser Ser Ser Pro Asp Trp Asp Phe Asn Lys Ser Lys
35 40 45
Asp Asn Trp Pro Met Ala Asn Lys Val Gly Val Gly Gly Tyr Gly Pro
50 55 60
Gly Phe Thr Pro Pro His Gly Gly Leu Leu Gly Trp Ser Pro Gln Ala
65 70 75 80
Gln Gly Val Leu Thr Thr Leu Pro Ala Asp Pro Pro Pro Ala Ser Thr
85 90 95
Asn Arg Arg Ser Gly Arg Lys Pro Thr Pro Val Ser Pro Pro Leu Arg
100 105 110
Asp Thr His Pro Gln Ala Met Gln Trp Asn Ser Thr Gln Phe His Gln
115 120 125
Ala Leu Leu Asp Pro Arg Val Arg Ala Leu Tyr Phe Pro Ala Gly Gly
130 135 140
Ser Ser Ser Glu Thr Gln Asn Pro Ala Pro Thr Ile Ala Ser Leu Thr
145 150 155 160
Ser Ser Ile Phe Leu Lys Thr Gly Gly Pro Ala Met Asn
165 170
<210> 12
<211> 172
<212> Белок
<213> Вирус гепатита В
<400> 12
Gly Leu Ser Trp Thr Val Pro Leu Glu Trp Gly Lys Asn Leu Ser Thr
1 5 10 15
Ser Asn Pro Leu Gly Phe Leu Pro Asp His Gln Leu Asp Pro Ala Phe
20 25 30
Arg Ala Asn Thr Asn Asn Pro Asp Trp Asp Phe Asn Pro Lys Lys Asp
35 40 45
Pro Trp Pro Glu Ala Asn Lys Val Gly Val Gly Ala Tyr Gly Pro Gly
50 55 60
Phe Thr Pro Pro His Gly Gly Leu Leu Gly Trp Ser Pro Gln Ser Gln
65 70 75 80
Gly Thr Leu Thr Thr Leu Pro Ala Asp Pro Pro Pro Ala Ser Thr Asn
85 90 95
Arg Gln Ser Gly Arg Gln Pro Thr Pro Ile Ser Pro Pro Leu Arg Asp
100 105 110
Ser His Pro Gln Ala Met Gln Trp Asn Ser Thr Ala Phe His Gln Ala
115 120 125
Leu Gln Asn Pro Lys Val Arg Gly Leu Tyr Phe Pro Ala Gly Gly Ser
130 135 140
Ser Ser Gly Ile Val Asn Pro Val Pro Thr Ile Ala Ser His Ile Ser
145 150 155 160
Ser Ile Phe Ser Arg Ile Gly Asp Pro Ala Pro Asn
165 170
<210> 13
<211> 173
<212> Белок
<213> Вирус гепатита В
<400> 13
Gly Ala Pro Leu Ser Thr Ala Arg Arg Gly Met Gly Gln Asn Leu Ser
1 5 10 15
Val Pro Asn Pro Leu Gly Phe Phe Pro Asp His Gln Leu Asp Pro Leu
20 25 30
Phe Arg Ala Asn Ser Ser Ser Pro Asp Trp Asp Phe Asn Thr Asn Lys
35 40 45
Asp Asn Trp Pro Met Ala Asn Lys Val Gly Val Gly Gly Phe Gly Pro
50 55 60
Gly Phe Thr Pro Pro His Gly Gly Leu Leu Gly Trp Ser Pro Gln Ala
65 70 75 80
Gln Gly Ile Leu Thr Thr Ser Pro Pro Asp Pro Pro Pro Ala Ser Thr
85 90 95
Asn Arg Arg Ser Gly Arg Lys Pro Thr Pro Val Ser Pro Pro Leu Arg
100 105 110
Asp Thr His Pro Gln Ala Met Gln Trp Asn Ser Thr Gln Phe His Gln
115 120 125
Ala Leu Leu Asp Pro Arg Val Arg Gly Leu Tyr Phe Pro Ala Gly Gly
130 135 140
Ser Ser Ser Glu Thr Gln Asn Pro Ala Pro Thr Ile Ala Ser Leu Thr
145 150 155 160
Ser Ser Ile Phe Ser Lys Thr Gly Asp Pro Ala Met Asn
165 170
<210> 14
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> фрагмент preS
<400> 14
Gly Gly Trp Ser Ser Lys Pro Arg Lys Gly Met Gly Thr Asn Leu Ser
1 5 10 15
Val Pro Asn Pro Leu Gly Phe Phe Pro Asp His Gln Leu Asp
20 25 30
<210> 15
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> фрагмент preS
<400> 15
Leu Gly Phe Phe Pro Asp His Gln Leu Asp Pro Ala Phe Gly Ala Asn
1 5 10 15
Ser Asn Asn Pro Asp Trp Asp Phe Asn Pro Ile Lys Asp His
20 25 30
<210> 16
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> фрагмент preS
<400> 16
Asp Trp Asp Phe Asn Pro Ile Lys Asp His Trp Pro Ala Ala Asn Gln
1 5 10 15
Val Gly Val Gly Ala Phe Gly Pro Gly Leu Thr Pro Pro His
20 25 30
<210> 17
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> фрагмент preS
<400> 17
Ala Phe Gly Pro Gly Leu Thr Pro Pro His Gly Gly Ile Leu Gly Trp
1 5 10 15
Ser Pro Gln Ala Gln Gly Ile Leu Thr Thr Val Ser Thr Ile
20 25 30
<210> 18
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> фрагмент preS
<400> 18
Gln Gly Ile Leu Thr Thr Val Ser Thr Ile Pro Pro Pro Ala Ser Thr
1 5 10 15
Asn Arg Gln Ser Gly Arg Gln Pro Thr Pro Ile Ser Pro Pro
20 25 30
<210> 19
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> фрагмент preS
<400> 19
Gly Arg Gln Pro Thr Pro Ile Ser Pro Pro Leu Arg Asp Ser His Pro
1 5 10 15
Gln Ala Met Gln Trp Asn Ser Thr Ala Phe His Gln Ala Leu
20 25 30
<210> 20
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> фрагмент preS
<400> 20
Trp Asn Ser Thr Ala Phe His Gln Ala Leu Gln Asp Pro Arg Val Arg
1 5 10 15
Gly Leu Tyr Phe Pro Ala Gly Gly Ser Ser Ser Gly Thr Val
20 25 30
<210> 21
<211> 33
<212> Белок
<213> Искусственная последовательность
<220>
<223> фрагмент preS
<400> 21
Pro Ala Gly Gly Ser Ser Ser Gly Thr Val Asn Pro Ala Pro Asn Ile
1 5 10 15
Ala Ser His Ile Ser Ser Ile Ser Ala Arg Thr Gly Asp Pro Val Thr
20 25 30
Asn
<210> 22
<211> 294
<212> Белок
<213> Искусственная последовательность
<220>
<223> PreSF1
<400> 22
Met Val Arg Tyr Thr Thr Glu Gly Gly Thr Lys Thr Glu Ala Glu Asp
1 5 10 15
Val Ile Pro Glu Gly Trp Lys Ala Asp Thr Ser Tyr Glu Ser Lys Val
20 25 30
Arg Tyr Thr Thr Glu Gly Gly Thr Lys Thr Glu Ala Glu Asp Val Ile
35 40 45
Pro Glu Gly Trp Lys Ala Asp Thr Ser Tyr Glu Ser Lys Gly Gly Trp
50 55 60
Ser Ser Lys Pro Arg Lys Gly Met Gly Thr Asn Leu Ser Val Pro Asn
65 70 75 80
Pro Leu Gly Phe Phe Pro Asp His Gln Leu Asp Pro Ala Phe Gly Ala
85 90 95
Asn Ser Asn Asn Pro Asp Trp Asp Phe Asn Pro Ile Lys Asp His Trp
100 105 110
Pro Ala Ala Asn Gln Val Gly Val Gly Ala Phe Gly Pro Gly Leu Thr
115 120 125
Pro Pro His Gly Gly Ile Leu Gly Trp Ser Pro Gln Ala Gln Gly Ile
130 135 140
Leu Thr Thr Val Ser Thr Ile Pro Pro Pro Ala Ser Thr Asn Arg Gln
145 150 155 160
Ser Gly Arg Gln Pro Thr Pro Ile Ser Pro Pro Leu Arg Asp Ser His
165 170 175
Pro Gln Ala Met Gln Trp Asn Ser Thr Ala Phe His Gln Ala Leu Gln
180 185 190
Asp Pro Arg Val Arg Gly Leu Tyr Phe Pro Ala Gly Gly Ser Ser Ser
195 200 205
Gly Thr Val Asn Pro Ala Pro Asn Ile Ala Ser His Ile Ser Ser Ile
210 215 220
Ser Ala Arg Thr Gly Asp Pro Val Thr Asn Val Arg Tyr Thr Thr Glu
225 230 235 240
Gly Gly Thr Lys Thr Glu Ala Glu Asp Val Ile Pro Glu Gly Trp Lys
245 250 255
Ala Asp Thr Ser Tyr Glu Ser Lys Val Arg Tyr Thr Thr Glu Gly Gly
260 265 270
Thr Lys Thr Glu Ala Glu Asp Val Ile Pro Glu Gly Trp Lys Ala Asp
275 280 285
Thr Ser Tyr Glu Ser Lys
290
<210> 23
<211> 298
<212> Белок
<213> Искусственная последовательность
<220>
<223> PreSF2
<400> 23
Met Phe Arg Phe Leu Thr Glu Lys Gly Met Lys Asn Val Phe Asp Asp
1 5 10 15
Val Val Pro Glu Lys Tyr Thr Ile Gly Ala Thr Tyr Ala Pro Glu Glu
20 25 30
Phe Arg Phe Leu Thr Glu Lys Gly Met Lys Asn Val Phe Asp Asp Val
35 40 45
Val Pro Glu Lys Tyr Thr Ile Gly Ala Thr Tyr Ala Pro Glu Glu Gly
50 55 60
Gly Trp Ser Ser Lys Pro Arg Lys Gly Met Gly Thr Asn Leu Ser Val
65 70 75 80
Pro Asn Pro Leu Gly Phe Phe Pro Asp His Gln Leu Asp Pro Ala Phe
85 90 95
Gly Ala Asn Ser Asn Asn Pro Asp Trp Asp Phe Asn Pro Ile Lys Asp
100 105 110
His Trp Pro Ala Ala Asn Gln Val Gly Val Gly Ala Phe Gly Pro Gly
115 120 125
Leu Thr Pro Pro His Gly Gly Ile Leu Gly Trp Ser Pro Gln Ala Gln
130 135 140
Gly Ile Leu Thr Thr Val Ser Thr Ile Pro Pro Pro Ala Ser Thr Asn
145 150 155 160
Arg Gln Ser Gly Arg Gln Pro Thr Pro Ile Ser Pro Pro Leu Arg Asp
165 170 175
Ser His Pro Gln Ala Met Gln Trp Asn Ser Thr Ala Phe His Gln Ala
180 185 190
Leu Gln Asp Pro Arg Val Arg Gly Leu Tyr Phe Pro Ala Gly Gly Ser
195 200 205
Ser Ser Gly Thr Val Asn Pro Ala Pro Asn Ile Ala Ser His Ile Ser
210 215 220
Ser Ile Ser Ala Arg Thr Gly Asp Pro Val Thr Asn Phe Arg Phe Leu
225 230 235 240
Thr Glu Lys Gly Met Lys Asn Val Phe Asp Asp Val Val Pro Glu Lys
245 250 255
Tyr Thr Ile Gly Ala Thr Tyr Ala Pro Glu Glu Phe Arg Phe Leu Thr
260 265 270
Glu Lys Gly Met Lys Asn Val Phe Asp Asp Val Val Pro Glu Lys Tyr
275 280 285
Thr Ile Gly Ala Thr Tyr Ala Pro Glu Glu
290 295
<210> 24
<211> 306
<212> Белок
<213> Искусственная последовательность
<220>
<223> PreSF4
<400> 24
Met Gly Lys Ala Thr Thr Glu Glu Gln Lys Leu Ile Glu Asp Val Asn
1 5 10 15
Ala Ser Phe Arg Ala Ala Met Ala Thr Thr Ala Asn Val Pro Pro Ala
20 25 30
Asp Lys Gly Lys Ala Thr Thr Glu Glu Gln Lys Leu Ile Glu Asp Val
35 40 45
Asn Ala Ser Phe Arg Ala Ala Met Ala Thr Thr Ala Asn Val Pro Pro
50 55 60
Ala Asp Lys Gly Gly Trp Ser Ser Lys Pro Arg Lys Gly Met Gly Thr
65 70 75 80
Asn Leu Ser Val Pro Asn Pro Leu Gly Phe Phe Pro Asp His Gln Leu
85 90 95
Asp Pro Ala Phe Gly Ala Asn Ser Asn Asn Pro Asp Trp Asp Phe Asn
100 105 110
Pro Ile Lys Asp His Trp Pro Ala Ala Asn Gln Val Gly Val Gly Ala
115 120 125
Phe Gly Pro Gly Leu Thr Pro Pro His Gly Gly Ile Leu Gly Trp Ser
130 135 140
Pro Gln Ala Gln Gly Ile Leu Thr Thr Val Ser Thr Ile Pro Pro Pro
145 150 155 160
Ala Ser Thr Asn Arg Gln Ser Gly Arg Gln Pro Thr Pro Ile Ser Pro
165 170 175
Pro Leu Arg Asp Ser His Pro Gln Ala Met Gln Trp Asn Ser Thr Ala
180 185 190
Phe His Gln Ala Leu Gln Asp Pro Arg Val Arg Gly Leu Tyr Phe Pro
195 200 205
Ala Gly Gly Ser Ser Ser Gly Thr Val Asn Pro Ala Pro Asn Ile Ala
210 215 220
Ser His Ile Ser Ser Ile Ser Ala Arg Thr Gly Asp Pro Val Thr Asn
225 230 235 240
Gly Lys Ala Thr Thr Glu Glu Gln Lys Leu Ile Glu Asp Val Asn Ala
245 250 255
Ser Phe Arg Ala Ala Met Ala Thr Thr Ala Asn Val Pro Pro Ala Asp
260 265 270
Lys Gly Lys Ala Thr Thr Glu Glu Gln Lys Leu Ile Glu Asp Val Asn
275 280 285
Ala Ser Phe Arg Ala Ala Met Ala Thr Thr Ala Asn Val Pro Pro Ala
290 295 300
Asp Lys
305
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ И ПРОФИЛАКТИКА АЛЛЕРГИИ НА КЛЕЩЕЙ ДОМАШНЕЙ ПЫЛИ | 2019 |
|
RU2815386C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ И ПРЕДОТВРАЩЕНИЯ ГЕПАТИТА В И D | 2021 |
|
RU2812764C1 |
| НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЮЩАЯ СЛИТЫЙ БЕЛОК, СОСТОЯЩИЙ ИЗ РАСТВОРИМОГО ВНЕКЛЕТОЧНОГО ФРАГМЕНТА ЧЕЛОВЕЧЕСКОГО Dll4 И КОНСТАНТНОЙ ЧАСТИ ТЯЖЕЛОЙ ЦЕПИ ЧЕЛОВЕЧЕСКОГО IgG4 | 2021 |
|
RU2787060C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРЕДОТВРАЩЕНИЯ ИЛИ ЛЕЧЕНИЯ ГЕПАТИТА, ФИБРОЗА ПЕЧЕНИ И ЦИРРОЗА ПЕЧЕНИ, ВКЛЮЧАЮЩАЯ СЛИТЫЕ БЕЛКИ | 2017 |
|
RU2795548C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ИНФЕКЦИИ, ВЫЗВАННОЙ STREPTOCOCCUS SUIS | 2015 |
|
RU2735101C2 |
| МУТАНТНЫЙ БЕЛОК F RSV И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2807742C1 |
| АНТИ-PRE-S1 HBV АНТИТЕЛА | 2016 |
|
RU2739955C2 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| ПРОМОТОРЫ ДЛЯ УСИЛЕНИЯ ЭКСПРЕССИИ В ПОКСВИРУСАХ | 2016 |
|
RU2753884C2 |
| ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ С ВЫСОКОПЛОТНЫМ ПОКРЫТИЕМ ДЛЯ ИНДУКЦИИ ЭКСПРЕССИИ АНТИТЕЛ | 2017 |
|
RU2813282C2 |






Изобретение относится к медицине и касается применения слитого белка в лечении и/или профилактике инфекции вирусом гепатита B, где указанный слитый белок содержит полипептид PreS вируса гепатита В, слитый с пептидом, состоящим из аминокислотной последовательности SEQ ID Nо. 1, SEQ ID Nо. 2, SEQ ID NO. 3 или SEQ ID NO. 4. Изобретение обеспечивает образование PreS-специфичных IgG в гораздо большей степени по сравнению с одним только PreS или другими слитыми белками, также содержащими PreS, слитый с различными пептидами, в частности с PreS-F1, PreS-F2 или PreS-F4. 6 з.п. ф-лы, 8 пр., 8 ил., 2 табл.
1. Применение слитого белка в лечении и/или профилактике инфекции вирусом гепатита B, где указанный слитый белок содержит полипептид PreS вируса гепатита В, слитый с пептидом, состоящим из аминокислотной последовательности SEQ ID Nо. 1, SEQ ID Nо. 2, SEQ ID NO. 3 или SEQ ID NO. 4.
2. Применение по п.1, где аминокислотная последовательность полипептида PreS состоит из SEQ ID Nо. 5.
3. Применение по п.1 или 2, где указанные пептиды, состоящие из SEQ ID Nо. 1, SEQ ID Nо. 2, SEQ ID NO. 3 и SEQ ID NO. 4, слиты с N- и/или С-концом полипептида PreS.
4. Применение по п.1 или 2, где слитый белок состоит из аминокислотной последовательности SEQ ID Nо. 6.
5. Применение по п.1 или 2, где инфекция вирусом гепатита В вызвана вирусом гепатита В с генотипом А, В, С, D, Е, F, G, Н или его подтипом.
6. Применение по п.1 или 2, где слитый белок вводят индивидууму по меньшей мере один раз в количестве от 0,01 мкг/кг массы тела до 5 мг/кг массы тела, предпочтительно от 0,1 мкг/кг массы тела до 2 мг/кг массы тела.
7. Применение по п.1 или 2, где слитый белок вводят вместе по меньшей мере с одним адъювантом и/или фармацевтически приемлемым вспомогательным веществом.
| WO 2012168487 A1, 13.12.2012 | |||
| GODON O | |||
| et al | |||
| Recognition of core-derived epitopes from a novel HBV-targeted immunotherapeutic by T-cells from patients infected by different viral genotypes.Vaccine | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| FOCKE-TEJKL M | |||
| et al | |||
| Development and characterization of a | |||

