ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к композициям и способам блокирования проникновения бактерий рода Shigella (шигелла) в клетки животных и, таким образом, к обеспечению защиты от инфекций, вызываемых бактериями рода Shigella, или снижению тяжести инфекций, вызываемых Shigella. Более конкретно, изобретение относится к применению белка IpaD, получаемого из природных источников и/или посредством синтеза или технологии рекомбинантных белков, а также конъюгатов данного белка, для получения нейтрализующих антител, обладающих защитным действием против некоторых серотипов бактерий рода Shigella, в частности S. flexneri. Композиции согласно изобретению могут быть полезны в предотвращении или лечении шигеллиоза, вызываемого бактериями рода Shigella.
УРОВЕНЬ ТЕХНИКИ
Многие грамотрицательные патогенные бактерии используют систему секреции третьего типа (T3S) для взаимодействия с клетками своих хозяев. Каждая система T3S состоит из аппарата секреции (T3SA), который пронизывает бактериальную оболочку и выступает на поверхности бактериальной клетки;
транслокаторов, которые проходят через T3SA и встраиваются в мембрану клетки хозяина, где формируют пору; эффекторов, которые, пройдя через T3SA и пору, образованную транслокаторами, попадают в цитоплазму клетки; определенных шаперонов, которые ассоциируют с транслокаторами и эффекторами в цитоплазме бактерии; и регуляторов транскрипции. Для сборки T3SA требуется около 15 различных белков.
Бактерии, относящиеся к роду Shigella, являются возбудителями бактериальной дизентерии у человека [1]. Гены, необходимые для проникновения бактерий в клетки эпителия и индуцирования апоптоза макрофагов, сгруппированы в области размером 30 т.п.н., называемой областью проникновения («entry region»), плазмиды вирулентности размером 220 т.п.н. Область проникновения содержит гены mxi и spa, кодирующие компоненты T3SA; гены ipaA, В, С и D, ipgB1, ipgD и icsB, кодирующие белки, проникающие через T3SA; гены ipgA, ipgC, ipgE и spa15, кодирующие шапероны; и гены virB и mxiE, кодирующие регуляторы транскрипции [2].
T3SA, который слабо активен в бактериях, растущих на жидкой среде, активируется при контакте бактерий с клетками эпителия [3]. Инактивация ipaB, ipaC или ipaD, так же, как и большинства генов mxi и spa, нарушает способность бактерий к проникновению в клетки эпителия, индуцированию апоптоза в макрофагах и проявлению контактной гемолитической активности. IpaB и IpaC содержат гидрофобные сегменты и остаются связанными с мембраной лизированных эритроцитов, что свидетельствует о том, что два этих белка являются компонентами транслокатора S. fexneri. Кроме того, существует предположение, что белки IpaB и IpaC обладают эффекторными функциями [4,5,6,7,8]. Инактивация ipaB и ipaD, но не ipaC, приводит к нарушению регуляции T3SA, т.е. к его конститутивной активности, что свидетельствует о роли IpaB и IpaD в поддержании T3SA в неактивном состоянии в отсутствие индукторов [9, 10]. Небольшая доля IpaD ассоциирована с бактериальной оболочкой [9, 11]. Picking et al. [12] сообщили, что роль IpaD в контроле активности T3SA возможно не связана с его участием в проникновении бактерий в клетки эпителия.
Для получения дальнейших данных о структуре игольчатого комплекса авторы настоящего изобретения провели иммуно-электронный микроскопический анализ бактерий, обработанных поперечносшивающим агентом BS3, в отношении целых бактерий и в некоторой степени очищенных игольчатых комплексов (NC). Авторами изобретения показано, что IpaD является компонентом NC, расположенным на кончике иглы, а также, что антитела, выработанные против IpaD, обладают ингибирующим действием в отношении процесса проникновения S. flexneri в клетки эпителия.
КРАТКОЕ ОПИСАНИЕ ФИГУР
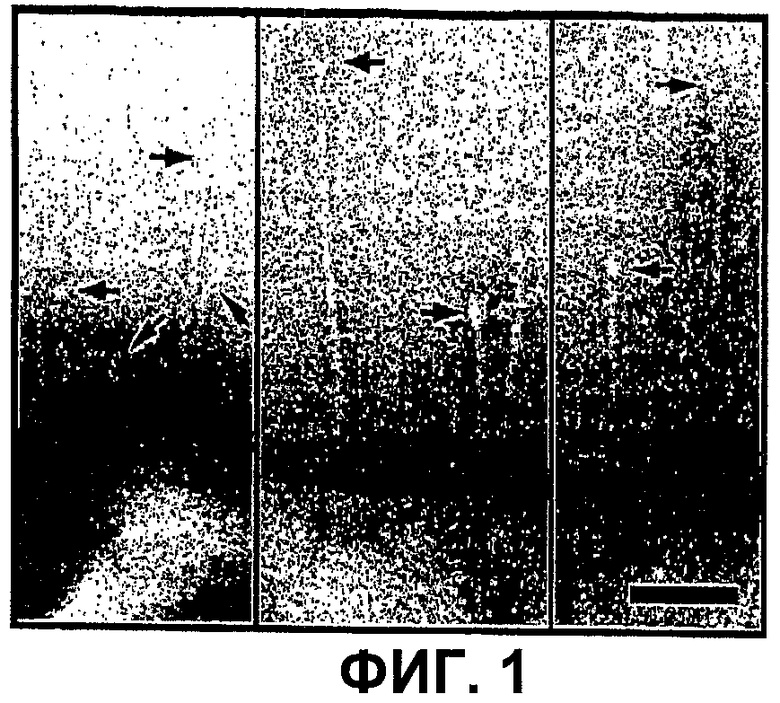
Фиг.1 - электронные микрофотографии отрицательно окрашенных бактерий, обработанных BS3. Стрелки указывают видимые уплотнения на кончиках игл, выступающих из поверхности бактериальной клетки. Масштабная линейка 100 нм.
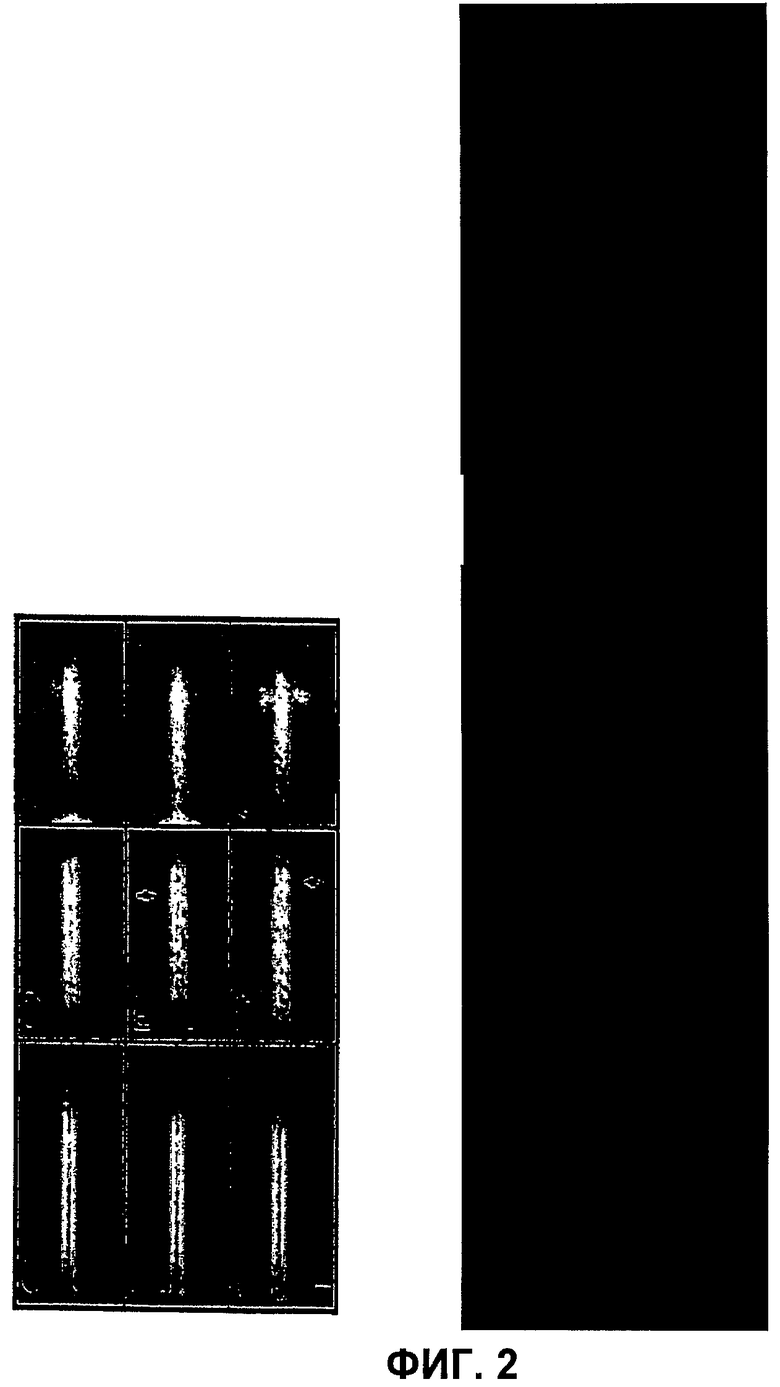
Фиг.2 - проекционные карты (суммация классов) на игольчатом участке NC, полученные методом одночастичного анализа. (А-В) карты NC, выделенных из бактерий «дикого типа», обработанных BS3 (А-С); из бактерий «дикого типа», не обработанных BS3 (D-F) и из бактерий мутантного штамма ipaD, обработанных BS3 (G-1). Стрелки указывают на участки остаточной плотности, ассоциированные с участками игл, приготовленными без воздействия BS3. Масштабная линейка 10 нм.
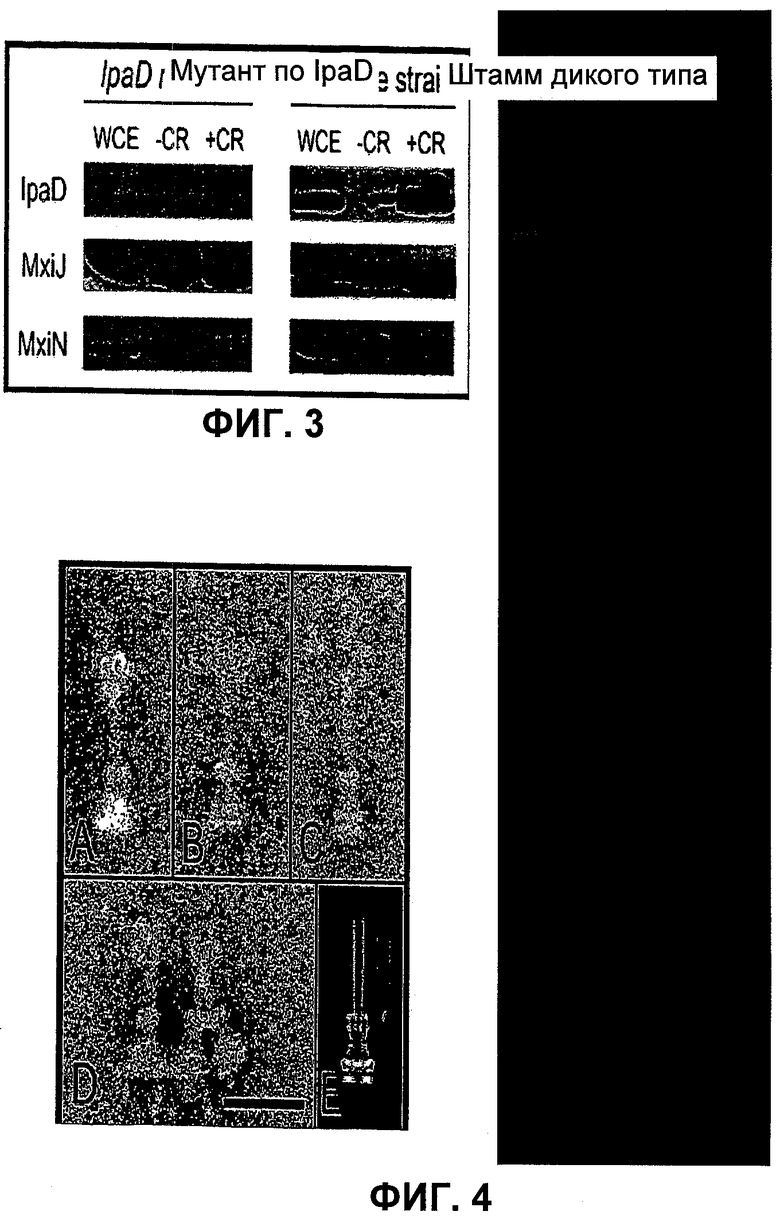
Фиг.3 - анализ очищенных NC методом иммуноблоттинга. Экстракты целых клеток (WCE) и поперечно сшитые (+CR) и NC без поперечной сшивки (-CR), выделенные из штаммов «дикого типа» и штаммов, дефектных по IpaD, исследовали методами электрофореза в ДСН-ПААГ (SDS-PAGE) и иммуноблоттинга с использованием антител, специфичных к MxiJ, MxiN, и IpaD. IpaD был обогащен до уровня WCE по NC, полученных из поперечносшитых бактерий дикого типа. MxiJ является положительным контролем для демонстрации интактного T3SA. MxiN, цитоплазматический компонент T3S, является положительным контролем для демонстрации загрязнения несвязанными цитоплазматическими белками.
Фиг.4 - локализация IpaD методом иммуно-электронной микроскопии. NC, очищенные из штаммов дикого типа, обработанных BS3, инкубировали в присутствии анти-IpaD антител и подвергали отрицательному окрашиванию (A-D). Усредненное изображение 250 NC, очищенных из бактерий, не обработанных BS3, показана на рисунке Е для сравнения. Масштабная линейка 10 нм.
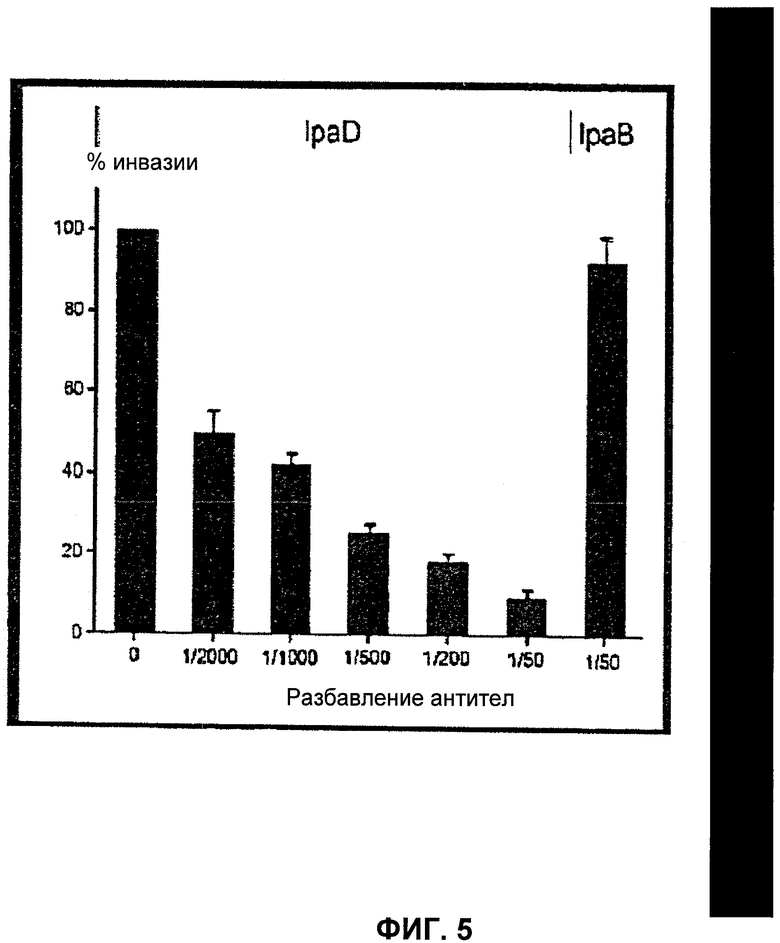
Фиг.5 - исследование заражения клеток эпителия S. flexneri дикого типа. Бактерии инкубировали в присутствии серии разведении анти-IpaD или анти-IpaB поликлональных антител, затем проводили подсчет гентамицин-устойчивых бактерий, находящихся внутри клеток путем высевания на чашку лизатов клеток. Эффективность проникновения в случае различных условий приведена относительно штамма дикого типа, обработанного ФБР (фосфатным буферным раствором). Приведены усредненные значения по меньшей мере по трем независимым экспериментам, в качестве ошибки указано стандартное отклонение.
Фиг.6 - известная последовательность аминокислот белка IpaD штамма Shigella (номер в базе GenBank AL391753).
Фиг.7 - известная последовательность нуклеотидов, кодирующая белок IpaD, представленный на Фиг.6 (номер в базе GenBank AL391753).
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы изобретения неожиданно обнаружили, что нейтрализующие антитела, специфичные к IpaD, могут блокировать проникновение Shigella в восприимчивые клетки, такие как клетки эпителия, и что этот нейтрализующий эффект наблюдается в отношении различных серотипов Shigella. Соответственно, настоящее изобретение более конкретно относится к применению белка IpaD, полинуклеотида, кодирующего указанный белок, или нейтрализующих анти-IpaD антител для получения композиций и разработки способов обеспечения перекрестной защиты от шигеллезных инфекций.
ОПРЕДЕЛЕНИЯ
Термины «животное» или «хозяин» относятся к любому животному, восприимчивому к какому-либо штамму Shigella, например S. flexneri, или к животному, о котором известно, что оно инфицировано каким-либо штаммом Shigella, например S. flexneri. Более конкретно, к животным относится человек. Термин «пермиссивная (восприимчивая) клетка» относится к клетке, которая может быть инфицирована штаммом Shigella. Например, такой клеткой может быть (не ограничиваясь этим) клетка эпителия или клетки иммунной системы, в частности клетки, являющиеся мишенями для проникновения Shigella в иммунной системе слизистой оболочки кишечника, например дендритные клетки и моноциты/макрофаги, а также В и Т-лимфоциты, в которые эффекторы Shigella могут попасть через систему секреции III типа, даже если это не приводит к их заражению.
Термин «лечение» относится к процессу, который обеспечивает снижение выраженности или устранение симптомов инфекции или заболевания, ассоциированных со штаммом Shigella. В настоящей заявке термин «предотвращение» относится к процессу, который обеспечивает подавление (блокирование) или задержку проявления симптомов инфекции или заболевания, ассоциированных со штаммом Shigella.
Термин «защитная реакция (ответ)» означает предотвращение начала заболевания, ассоциированного с Shigella, или инфекции, вызванной каким-либо видом, или снижение тяжести существующего заболевания у животного.
Выражение «приемлемый носитель» обозначает среду, которая может содержать элементы композиции, предусмотренной настоящим изобретением, который можно вводить животному-хозяину без побочных эффектов. Подходящими носителями, известными в данной области, являются (но не ограничиваются перечисленными) частицы золота, стерильная вода, физиологический раствор, глюкоза, декстроза или буферные растворы. Носители могут включать вспомогательные вещества, включая (но не ограничиваясь перечисленными) разбавители, стабилизаторы (например, сахара и аминокислоты), консерванты, смачивающие вещества, эмульгаторы, вещества, буферизующие рН, добавки, повышающие вязкость, красители и т.п.
«Функциональное производное» в настоящей заявке относится к последовательности белка/полипептида, которая обладает функциональной биологической активностью, которая, по существу, аналогична биологической активности полной последовательности белка/пептида IpaD. Другими словами, данное выражение предпочтительно относится к полипептиду или фрагментам полипептида, которые в значительной степени сохраняют способность вызывать продукцию нейтрализующих анти-IpaD антител против инфекционного штамма Shigella, при введении указанного функционального производного животному.
«Функциональный фрагмент» в настоящей заявке относится к последовательности нуклеиновой кислоты, которая определяет (кодирует) функциональную биологическую активность, по существу, аналогичную биологической активности полной последовательности нуклеиновой кислоты IpaD. Другими словами, в контексте настоящего изобретения данное выражение предпочтительно относится к нуклеиновой кислоте и фрагментам нуклеиновой кислоты, которые в значительной степени сохраняют способность кодировать полипептид/белок IpaD, который вызывает продукцию нейтрализующих анти-IpaD антител против инфекционного штамма Shigella, при введении животному.
Термин «серотип Shigella» относится к четырем группам Shigella, обозначаемым заглавными буквами от А до D: Shigella dysenteriae (A) Shigella flexneri (В), Shigella boydii (С) и Shigella sonnei (D). Они соответственно включают: группа А: 8 серотипов, группа В: 11 серотипов и суб-серотипов, группа С - 11 серотипов, группа D - 1 серотип. Например, серотип Shigella может представлять собой, без ограничения, Shigella flexneri 2a, 1b и 3а, Shigella dysenteriae 1 и Shigella sonnei.
Термин «нейтрализация», или «блокирование», относится к способности анти-IpaD антител, рассматриваемых в изобретении, специфичным образом связываться с белком IpaD и нарушать биологическую функцию белка IpaD, таким образом блокируя, например, способность бактерии доставлять эффекторы вирулентности в клетки-мишени. Другими словами, такие антитела преимущественно обезвреживают патоген и делают его одновременно неспособным причинять вред и эффективно противодействовать иммунной защите хозяина.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном из вариантов изобретение обеспечивает композицию для лечения и предотвращения инфекции Shigella (шигеллезной инфекции). Изобретение также обеспечивает композицию для блокирования проникновения по меньшей мере одного серотипа Shigella в пермиссивные клетки. Предусмотренные композиции включают по меньшей мере один из следующих элементов:
- полипептид IpaD или функциональное производное такогополипептида;
- полинуклеотид, кодирующий полипептид IpaD или функциональный фрагмент такого полинуклеотида;
- нейтрализующее анти-IpaD антитело.
Для специалиста в данной области понятно, что композиции согласно изобретению при введении хозяину преимущественно предоставляют перекрестную защиту против более чем одного серотипа Shigella. Другими словами, композиции согласно настоящему изобретению предотвращают или существенно снижают проникновение в пермиссивные клетки более одного серотипа Shigella.
Полипептид IpaD или функциональное производное полипептида IpaD, предусмотренные настоящим изобретением, имеют последовательность аминокислот, идентичную или в значительной степени идентичную последовательности аминокислот, имеющей номер AL391753 в базе данных GenBank и обозначенной здесь SEQ ID. NO:1 (см. фиг.6). В случае функционального производного полипептида IpaD можно использовать плазмиду pMal-IpaD, депонированную в CNCM 10 октября 2007 г. под номером 1-3839. Эта плазмида кодирует фрагмент белка IpaD, начинающийся с кодона 130 белка IpaD.
«В значительной степени идентичная» по отношению к аминокислотной последовательности будет означать, что полипептид, предусмотренный настоящим изобретением, имеет, например, последовательность аминокислот, обладающую по меньшей мере 75% идентичностью, или 85% идентичностью, или даже 95% идентичностью, с частью или полной последовательности, обозначенной SEQ ID NO:1. Полинуклеотид или его функциональный фрагмент, предусмотренный настоящим изобретением, кодирует полипептид IpaD или его функциональное производное, определенные выше. Например, такой полинуклеотид имеет последовательность нуклеотидов (последовательность нуклеиновой кислоты), идентичную или в значительной степени идентичную последовательности нуклеотидов, имеющей номер AL391753 в базе GenBank и обозначенной здесь SEQ ID NO:2 (см. фиг.7). В случае функционального фрагмента полинуклеотида IpaD можно использовать плазмиду pMal-IpaD, определенную выше.
«В значительной степени идентичная» по отношению к нуклеотидной или полинуклеотидной последовательности будет пониматься, что полинуклеотид, предусмотренный настоящим изобретением, имеет, например, последовательность нуклеиновой кислоты, обладающую по меньшей мере 65% идентичностью, или 80% идентичностью, или даже 95% идентичностью, с полной последовательностью, указанной в SEQ ID NO:2, или ее частью.
Методики определения степени идентичности для последовательностей нуклеотидов и аминокислот хорошо известны в данной области. Обычно эти методики включают определение последовательности нуклеотидов мРНК гена, последовательности самой ДНК и/или определение последовательности аминокислот, кодируемой ими, и сравнение этих последовательностей со второй последовательностью аминокислот или нуклеотидов. В общем смысле «идентичность» относится к точному совпадению соответствующих нуклеотидов или аминокислот в последовательностях двух полинуклеотидов или полипептидов, соответственно. Две или более последовательности (полинуклеотидные или аминокислотные) можно сравнивать путем определения их «процентной идентичности». Степень идентичности двух последовательностей - это число точных совпадений между двумя выровненными последовательностями, разделенное на длину более короткой последовательности и умноженное на 100. Приблизительное выравнивание для нуклеиновых кислот осуществляется по алгоритму локальной гомологии согласно Smith and Waterman, Advances in Applied Mathematics 2:482-489 (1981). Этот алгоритм можно применять также для последовательностей аминокислот при помощи матриц, разработанных Dayhoff, Atlas of Protein Sequences and Structure, M. O. Dayhoff ed., 5 suppl. 3:353-358, National Biomedical Research Foundation, Washington, D.C., USA, и нормализованных по Gribskov, Nucl. Acids Res. 14(6):6745-6763 (1986). Данный алгоритм для определения степени сходства последовательностей реализован, например, Genetics Computer Group (Madison, Wis.) в приложении "BestFit". Параметры, заданные по умолчанию для этого метода, описаны в Wisconsin Sequence Analysis Package Program Manual, Version 8 (1995) (доступен в Genetics Computer Group, Madison, Wis.). Предпочтительным способом установления степени идентичности в контексте настоящего изобретения является использование пакета программ MPSRCH, охраняемого авторским правом Университета Эдинбурга (University of Edinburgh), разработанного John F. Collins и Shane S. Sturrok и распространяемого IntelliGenetics, Inc. (Mountain View, Calif.). Из данного пакета программ можно использовать алгоритм Smith-Waterman с параметрами по умолчанию для таблиц сравнения (например, gap open penalty 12, gap extension penalty 1, и gap - 6). Среди генерируемых данных значение "Match" отражает «идентичность последовательности». Другие подходящие программы для расчета степени идентичности или подобия между последовательностями широко известны в данной области исследования, например другой программой для выравнивания является BLAST, используемая с параметрами, заданными по умолчанию. Например, BLASTN и BLASTP можно использовать со следующими параметрами по умолчанию: genetic code=standard; filter=none; strand=both; cutoff=60; expect=10; Matrix BLOSUM62; Descriptions=50 sequences; sort by=HIGH SCORE; Databases=non-redundant, GenBank+EMBL+DDBJ+PDB+GenBank CDS translations+Swiss protein+Spupdate+PIR.
Нейтрализующие антитела, рассматриваемые в изобретении, которые специфично связываются с белком IpaD, могут быть получены различными методами, известными специалистам в данной области. Например, можно вводить полипептид IpaD животным для стимуляции продукции поликлональных антител. В качестве альтернативы нейтрализующие анти-IpaD антитела для применения согласно настоящему описанию могут представлять собой моноклональные антитела, приготовленные с использованием гибридомных технологий (см. например Hammerling et al., In Monoclonal Antibodies and T-Cell Hybridomas, Elsevier, NY, 1981).
По отношению к антителам, предусмотренным настоящим изобретением, термин «специфично связывается с» относится к антителам, которые связываются с относительно высокой аффинностью с одним или более эпитопами полипептида IpaD, но при этом практически не распознают и не связывают молекулы, отличные от полипептида IpaD. В настоящей заявке термин «относительно высокая аффинность» обозначает аффинность связывания между антителом и полипептидом IpaD, достигающую по меньшей мере 106 М-1, и предпочтительно по меньшей мере 107 М-1, и даже более предпочтительно от 108 М-1 до 1010 М-1. Определение аффинности предпочтительно осуществляют путем иммунохимического анализа конкурентного связывания в стандартных условиях, которые известны специалистам в данной области.
При этом понимается, что специалист в данной области также способен отобрать и идентифицировать антитела, нейтрализующие проникновение Shigella в клетку.
В предпочтительном варианте реализации изобретения композиция согласно изобретению дополнительно включает вспомогательное вещество. В настоящей заявке термин «адъювант» обозначает вещество, добавляемое к композиции согласно настоящему изобретению для усиления иммуногенности композиции. Механизм действия адъювантов не выяснен полностью. Вероятно, некоторые адъюванты усиливают иммунный ответ (гуморальный и клеточный ответ) за счет медленного высвобождения антигена, в то время как другие адъюванты исходно являются сильными иммуногенами и действуют синергитически. Известные адъюванты включают в себя (но не ограничиваются перечисленными) масляные и водные эмульсии (например, полный адъювант Фрейнда (complete Freund's adjuvant) и неполный адъювант Фрейнда (incomplete Freund's adjuvant)), Corytzebactei-ium parvuin, Quil А, цитокины, такие как IL12, Emulsigen-Plus®, Bacillus Calmette Guerin, гидроксид алюминия, глюкан, сульфат декстрана, оксид железа, альгинат натрия, Bacto Adjuvant, некоторые синтетические полимеры, такие как полиаминокислоты и сополимеры аминокислот, сапонин, парафиновое масло и мурамил-дипептид. К адъювантам также относятся вспомогательные генетические вещества, такие как иммуномодулирующие молекулы, кодируемые одновременно вводимой ДНК, или олигонуклеотиды CpG. ДНК, вводимая совместно, может находиться в той же плазмидной конструкции, как плазмидный иммуноген, или в отдельном ДНК-векторе.
В соответствии с предпочтительным вариантом реализации изобретения, композиции могут далее иметь дополнительно полиозидный (polyosidic) антиген, например, описанный в WO 2005/003995. Например, предусмотренный полиозидный антиген может представлять собой (но не ограничиваясь перечисленными) синтетические полисахариды, соответствующие O-боковым цепям (определяющим серотип) представляющего интерес штамма, в частности, тех серотипов, которые считаются предпочтительными благодаря высокой распространенности: Shigella flexneri 2a, 1b и 3а, Shigella dysenteriae 1 и Shigella sonnei. Рассматриваемые полиозидные антигены также могут представлять собой детоксифицированные липополисахариды (LPS), выделенные из бактериальных культур близких серотипов.
СПОСОБЫ ЛЕЧЕНИЯ И КОМПОЗИЦИИ
Полипептид IpaD или его функциональные производные, полинуклеотиды или функциональные фрагменты, кодирующие полипептид IpaD или его функциональные производные, и антитела согласно настоящему изобретению можно применять множеством способов для лечения и/или предотвращения инфекции, вызываемой Shigella, или для блокирования проникновения штаммов Shigella в пермиссивные клетки.
Например, и в соответствии с одним из способов реализации изобретения, полипептид IpaD можно применять в качестве иммуногена для получения специфичных нейтрализующих анти-IpaD антител. Как отмечалось ранее, подходящие нейтрализующие анти-IpaD антитела можно определять с помощью известных соответствующих методов скрининга, например, путем измерения способности конкретного антитела нейтрализовать или блокировать проникновение штамма Shigella в клетку.
В соответствии с другим вариантом реализации изобретения, полинуклеотиды, кодирующие полипептид IpaD или его производные, можно применять в способе ДНК-иммунизации для выработки нейтрализующих анти-IpaD антител. Это означает, что их можно встраивать в вектор, способный к репликации или экспрессии после введения, что обеспечивает выработку антигенного полипептида in vivo. Например, полинуклеотиды могут быть встроены в плазмиду-вектор под контролем промотора CMV, который способен функционировать в эукариотических клетках. Предпочтительно, вектор вводят путем внутримышечной инъекции. Применение полинуклеотидов согласно настоящему изобретению для генетической иммунизации предпочтительно включает подходящий способ доставки или систему, такую как прямая инъекция плазмидной ДНК в мышцы [Wolf et al. H M G (1992) 1: 363, Turnes et al., Vaccine (1999), 17: 2089, Le et al., Vaccine (2000) 18: 1893, Alves et al., Vaccine (2001)19: 788], инъекция плазмидной ДНК с и без вспомогательных веществ [Ulmer et al., Vaccine (1999) 18: 18, MacLaughlin et al., J. Control Release (1998) 56: 259, Hartikka et al., Gene Then (2000) 7: 1171-82, Benvenisty and Reshef, PNAS USA (1986) 83: 9551, Singh et al., PNAS USA (2000) 97: 811], мечение клеток путем доставки ДНК, связанной со специфичными носителями [Wa et al., J Biol Chem (1989) 264: 16985, Chaplin et al., Infect. Immun. (1999) 67:6434], инъекция плазмиды, связанной или инкапсулированной в различных формах липосом [Ishii et al., AIDS Research and Human Retroviruses (1997) 13: 142, Perrie et al., Vaccine (2001) 19:3301], нацеливание ДНК путем различных методов бомбардировки [Tang et al., Nature (1992) 356: 152, Eisenbraun et al., DNA Cell Biol (1993) 12: 791, Chen et al., Vaccine (2001) 19:2908], нацеливание ДНК с использованием живых векторов [Tubulekas et al., Gene (1997) 190: 191, Pushko et al., Virology (1997) 239: 389, Spreng et al. FEMS (2000) 27: 299, Dietrich et al., Vaccine (2001) 19: 2506].
Дальнейшим вариантом реализации изобретения является применение специфичных нейтрализующих анти-IpaD антител для пассивной иммунизации.
Еще одним вариантом реализации изобретения является обеспечение способа лечения и/или предотвращения инфекции Shigella у животных в соответствии с изобретением.
Дальнейшим вариантом реализации изобретения является обеспечение способа блокирования проникновения по меньшей мере одного серотипа Shigella в пермиссивную клетку, включающего этап, обеспечивающий возможность образования иммунного комплекса путем контакта нейтрализующего анти-IpaD антитела со штаммом Shigella, способным инфицировать указанную пермиссивную клетку; причем упомянутый комплекс предотвращает или существенно снижает проникновение упомянутого штамма Shigella в пермиссивную клетку.
Количество компонентов или элементов композиций согласно настоящему изобретению предпочтительно представляет собой терапевтически эффективное количество. Терапевтически эффективное количество рассматриваемого компонента - это количество компонента, необходимое для реазизации его иммунологической функции (т.е. выработки нейтрализующих анти-IpaD антител), без стимуляции излишних негативных эффектов у хозяина, которому вводят композицию. Точное количество компонентов для применения и композиций для введения варьирует в зависимости от таких факторов, как тип состояния, которое лечат, тип и возраст животного, которое лечат, способ введения, а также другие ингредиенты композиции.
Композиции согласно изобретению можно давать животному посредством различных путей введения. Например, композиции можно вводить в форме стерильных препаратов для инъекций, таких как стерильные водные или масляные суспензии. Данные суспензии могут быть изготовлены в соответствии с технологиями, известными специалисту в данной области, с использованием подходящих диспергирующих или смачивающих агентов, и суспендирующих агентов. Стерильные препараты для инъекций могут также представлять собой стерильные растворы для инъекций или суспензии в нетоксичных растворителях, подходящих для парентерального введения. Их можно вводить парентерально, например внутривенно, внутримышечно или подкожно, путем инъекции, вливания (инфузии) или перорально. Подходящие дозировки будут варьировать в зависимости от таких факторов, как, например, количество компонентов в композиции, желаемый эффект (кратковременный или продолжительный), пути введения, возраста и веса животного, которое лечат. Для введения композиций согласно настоящему изобретению можно использовать любые другие способы, хорошо известные в данной области.
Следующий вариант реализации изобретения обеспечивает наборы для применения в любом из способов согласно настоящему изобретению. Набор может включать два или более компонентов, необходимых для проведения определенного теста. Компоненты могут представлять собой соединения, реагенты, контейнеры и/или оборудование. Например, один контейнер из набора может содержать моноклональное антитело или фрагмент моноклонального антитела, или поликлональные антитела, которые специфичным образом нейтрализуют белок IpaD. Один или более дополнительных контейнеров могут содержать элементы, такие как реагенты или буферы, для использования в тесте. Другие наборы, предусмотренные настоящим изобретением, могут содержать по меньшей мере один полипептид IpaD или полинуклеотид, кодирующий такой полипептид, описанные выше, для стимуляции нейтрализующего иммунного ответа против белка IpaD шигеллы.
Настоящее изобретение станет более понятно из приведенных ниже примеров. Данные примеры иллюстрируют широкий диапазон применения настоящего изобретения и ни в коем случае не ограничивают объем изобретения. В настоящее изобретение можно вносить модификации и вариации, не выходящие за пределы сущности и объема изобретения. Хотя на практике для тестирования настоящего изобретения можно использовать любые методы и материалы, подобные или эквивалентные описанным в настоящей заявке, предпочтительные методы и материалы описаны ниже.
ПРИМЕРЫ
Системы секреции III типа (T3S) используются многими грамотрицательными патогенными бактериями для введения белков вирулентности в клетки-хозяева животных и растений. Основа аппарата T3S, известная как игольчатый комплекс, состоит из базального тела, проходящего через обе бактериальные мембраны, и иглы, выступающей над поверхностью бактерии. У Shigella flexneri белок IpaD необходим для ингибирования активности аппарата T3S перед контактом бактерии с хозяином; предполагают, что IpaD способствует переносу (транслокации) бактериальных белков в клетку хозяина. Авторы настоящего изобретения изучили локализацию IpaD путем электронной микроскопии поперечносшитых бактерий и в некоторой степени очищенных игольчатых комплексов. Данный анализ выявил существование отчетливо видимых электронноплотных образований на кончике иглы. Комбинирование методов одночастичного анализа, иммуномечения и биохимического анализа позволило показать, что IpaD образует часть указанной структуры на кончике иглы. Было показано, что анти-IpaD антитела блокируют проникновение бактерий в клетки эпителия.
ОБЩИЕ МАТЕРИАЛЫ И МЕТОДЫ
Штаммы бактерий и среды для выращивания
Штаммы, которые использовали в настоящем исследовании, представляли собой штамм дикого типа S. flexneri 5 M90T-Sm [13] и полученный из него дефектный по ipaD штамм SF622 [14]. Бактерии выращивали в трипсин-казеин-соевом бульоне (TSB) (Sigma) при 37°С.
Очистка игольчатого комплекса (NC)
NC очищали, как описано в [15]. Бактерии, находящиеся в экспоненциальной фазе роста в 1 литре среды TSB при 37°С, собирали путем центрифугирования, ресуспендировали в 25 мл фосфатного буфера и инкубировали в присутствии 1 мМ Bis (Sulfosuccinimidyl)suberate (BS3) в течение 30 минут при 37°С. К смеси добавляли 100 мМ Tris-HCl и инкубировали в течение 15 минут при 37°С. Культуры, обработанные BS3, собирали и ресуспендировали в ледяном лизирующем буфере (0.5 М сахарозы, 20 мМ Tris-HCl [рН 7.5], 2 мМ ЭДТА, 0.5 мг/мл лизозима) с добавлением 1 мМ фенилметилсульфонил-фторида и инкубировали в течение 45 минут при 4°С и в течение 15 минут при 37°С. Полученные сферопласты инкубировали в присутствии 0.01% Triton Х-100 в течение 30 минут и обрабатывали 4 мМ MgCl2 и 80 мкг/мл ДНКазы (Sigma) в течение 20 минут при 30°С.Остатки клеток удаляли путем центрифугирования (20,000 g в течение 20 минут при 4°С), мембранную фракцию также осаждали путем центрифугирования (110,000 g в течение 30 минут при 4°С) и ресуспендировали в буфере ТЕТ (20 мМ Tris-HCl рН 7.5, 1 мМ ЭДТА, 0.01% Triton Х-100). Анализ методом иммуноблоттинга осуществляли с антителами, выработанными против MxiJ, MxiN и IpaD по описанной методике [16].
Электронная микроскопия и анализ изображений
Целые клетки и образцы очищенных NC негативно окрашивали 2% уранил-ацетатом на медных сетках, покрытых углеродом при тлеющем разряде. Электронную микроскопию осуществляли на Philips CM120FEG, снабженном эмиссионной пушкой, при 120 кВ. Изображения фиксировали на медленно сканирующую CCD камеру 4000 SP 4K при увеличении 80,000 х, при размере пиксела (после загрузки изображения) 3.75 А на уровне образца, с использованием программного обеспечения "GRACE" для полуавтоматического отбора образцов и сбора данных [17]. Одночастичный анализ, включая мультиреференсные и нереференсные процедуры, мультивариантный статистический анализ и классификацию, проводили, как описано в [15]. Для иммуномечения очищенные NC инкубировали с аффинно очищенными поликлональными антителами к IpaD (pAbs) в конечной концентрации 0.132 нг/мкл в течение 1 часа при 20°С. Образцы окрашивали 2% уранил-ацетатом и изучали, как указано выше.
Анализ инвазии
Два миллилитра культуры штаммов дикого типа и mxiD в экспоненциальной фазе роста (OD600nm=0.4) инкубировали в присутствии антител анти-IpaD (разведения от 1/2000 до 1/50) и анти-IpaB (1/50) в течение 1 часа при 37°С, затем бактерии центрифугировали на чашках, содержащих 2·105 клеток Hela в течение 10 минут при 2000 g. После 1 часа инкубирования при 37°С клетки промывали 3 раза 2 миллилитрами EBSS и инкубировали в течение 1 часа в присутствии 2 мл среды MEM, содержащей 50 µg/ml гентамицина. После трех промывок 2 миллилитрами EBSS (сбалансированный солевой раствор Эрла) чашки инкубировали в 0.5% растворе деоксохолата в течение 15 минут при 20°С, лизаты клеток растворяли и помещали на чашки с агаром для подсчета числа колоний.
Пример 1: Характерная структура на кончике иглы T3SA
Процедуры очистки белков обычно приводят к отбору наиболее стабильных комплексов, которые возможно не содержат слабо ассоциированные субъединицы. Авторами настоящего изобретения недавно было показано, что низкая концентрация детергента Triton-Х100 (0.01%) эффективна для индукции высвобождения NC из мембран (Sani et al., 2006). Для выявления потенциально лабильных субъединиц, присоединенных к игле, исследователи осуществляли этап перекрестных сшивок бактерий с использованием BS3 перед каждой очисткой. Анализ методом электронной микроскопии показал, что после действия BS3 у большинства бактерий присутствовали игольчатые выступы с повышенной плотностью вблизи кончика (фиг.1).
NC очищали из бактерий, обработанных BS3 после солюбилизации мембран детергентами, как описано в [18]. Препараты содержали достаточное для проведения структурного анализа количество NC с повышенной плотностью на кончике иглы. Изображения, полученные с помощью электронного микроскопа, анализировали путем одночастичного анализа с расчетом двухмерных проекционных карт. Авторы изобретения отобрали несколько сотен изображений NC с относительно прямыми и короткими игольчатыми выступами с длиной, близкой к 45 нм. Усредненные NC ясно демонстрировали присутствие электронной плотности в районе кончика иглы, а также в верхней части базального тела (фиг.2А и 2В, см. также фиг.4Е для общего вида NC). Тем не менее оказалось, что изображения NC после усреднения получаются несколько смазанными вследствие различий в длине игл. Более четкие подробности кончиков игл были получены после выравнивания проекций и классификации их после маскировки базальной части (фиг.2C). Характерной особенностью усредненной карты перекрестно-сшитых частиц является присутствие плотностей на любой стороне кончика иглы. В отличие от этого усредненные карты частиц, созданные на основе анализ штамма дикого типа без перекрестных сшивок, демонстрировали иглы, в которых большинство этих участков плотности отсутствовали (фиг.2D-F). На этих образцах видны участки с низкой плотностью, расположенные в тех же местах, где присутствует высокая плотность в препаратах, приготовленных с перекрестными сшивками (стрелки, фиг.Фиг.2Е и 2F). Данные результаты свидетельствуют о том, что в отсутствие перекрестных сшивок большинство очищенных NC теряют дополнительные молекулы, формирующие плотность, наблюдаемую после процедуры перекрестного сшивания. Для идентфикации молекул, образующих плотность на кончике T3SA, авторы изобретения провели аналогичные эксперименты с мутантом по ipaD, не экспрессирующим IpaD. IpaD, вместе с IpaB, необходим для поддержания T3SA в неактивном состоянии в отсутствие индукторов, и небольшая часть IpaD связана с мембраной [9]. NC, очищенные из мутанта ipaD, обработанного BS3, не демонстрировали наличия плотности ни на одной стороне кончика иглы (фиг.2G-I), что свидетельствует о том, что IpaD является частью данного структурного элемента или необходим для его сборки.
Пример 2: IpaD присутствует на кончике иглы T3SA
Чтобы проверить, действительно ли наблюдаемая плотность образована IpaD, NC, очищенные из бактерий дикого типа, обработанных BS3, исследовали методами электрофореза (SDS-PAGE) и иммуноблоттинга (фиг.3). Содержание IpaD было выше в NC, выделенных из перекрестно-сшитых бактерий дикого типа, по сравнению с NC, полученными из бактерий, не подвергнутых перекрестному сшиванию (фиг.3, правая панель, правая дорожка), хотя небольшие количества проявляются при выделении из бактерий, не подвергнутых перекрестному сшиванию, что подтверждает наличие участков слабой плотности на кончике игл у типичных NC, выделенных из бактерий, не подвергнутых перекрестному сшиванию. MxiJ, который является основным компонентом NC, присутствует в близких количествах во всех препаратах (фиг.3). Контрольные эксперименты с использованием антител, распознающих цитоплазматические компоненты системы T3S, например, MxiN, не выявили загрязнения NC внутриклеточными компонентами (фиг.3), что свидетельствует о том, что присутствие IpaD в препаратах не обусловлено загрязнением этих препаратов цитоплазматическими белками.
Для подтверждения того, что плотные участки, выявленные на кончиках NC, содержат IpaD, авторы изобретения провели иммунохимическое окрашивание с использованием анти-IpaD сыворотки. Антитела специфичным образом связывались с кончиком иглы в NC, выделенных из бактерий дикого типа, обработанных BS3 (фиг.4 A-D). Наблюдали, что некоторые иглы также ассоциированы друг с другом в концевых участках, что, вероятно, является результатом взаимодействия двухвалентного антитела с двумя иглами (нижний левый кадр, фиг.4D). В контрольных экспериментах не было обнаружено связывания антител с иглами NC, выделенными из мутанта по ipaD (данные не представлены).
Пример 3: Антитела анти-IpaD блокируют проникновение бактерий в клетки эпителия.
Поскольку IpaD необходим для проникновения бактерий в клетки эпителия [14], а также поскольку, как показано в настоящем изобретении, он расположен на кончике T3SA, авторы настоящего изобретения исследовали вопрос, могут ли анти-IpaD антитела препятствовать проникновению бактерий в клетки линии HeLa. Для инфицирования клеток HeLa использовали бактерии, инкубированные в присутствии различных концентраций анти-IpaD сыворотки или анти-IpaB сыворотки в качестве контроля. Воздействие анти-IpaD сыворотки на бактерии, в отличие от анти-IpaB, ингибировало проникновение бактерий дозозависимым образом (фиг.5). Обработка антителами анти-IpaD также ингибировало проникновение штамма 2а S. flexneri (данные не представлены).
Общее обсуждение примеров 1-3.
T3SA S. flexneri активируется при контакте бактерий с клетками эпителия, и его регуляция нарушается из-за инактивации ipaB или ipaD. Предполагают, что эти белки необходимы для формирования комплекса, блокирующего T3SA. Авторами настоящей заявки представлены свидетельства того, что IpaD присутствует на кончике иглы. Трансмиссионные электронные микрофотографии игл, экспонированных на поверхности перекрестно сшитых бактерий, имеют характерную особенную структуру, присутствующую на кончике иглы, и иммуноблоттинг в некоторой степени очищенных NC показывает, что очистка приводит к совместному выделению IpaD и поперечно сшитых NC.
Расчетные средние по NC, выделенным из поперечно сшитых бактерий дикого типа, продемонстрировали присутствие характерных электронно-плотных участков на обеих сторонах кончика иглы. Эта особенность не была отмечена для NC, выделенных из штамма дикого типа, не обработанного перекрестно-сшивающим агентом, и из штамма, мутантного по ipaD, обработанного поперечно-сшивающим агентом. Результаты иммунно-электронной микроскопии указывают на то, что наблюдаемая на кончике иглы плотность содержит молекулы IpaD. Однако точную конфигурацию повышенной плотности нельзя определить по картам двумерной проекции. Две дополнительные структуры имеют размеры приблизительно 7×7 нм. Поскольку размер IpaD равен 37 кДа, вероятно, в этих структурах присутствуют несколько копий IpaD. Действительно, считают, что IpaD формирует олигомеры [19]. IpaD демонстрирует некоторую функциональную аналогию с LcrV из Yersinia enterocolitica, поскольку оба эти белка имеют сходные размеры, и оба необходимы для встраивания предполагаемых транслокаторов:
IpaB и IpaC в случае S. flexneri и YopB и UopD в случае Y. enterocolitica, в мембрану клеток-хозяев [9, 20, 21]. Недавно полученные данные свидетельствуют о том, что LcrV локализуется на кончике иглы T3SA [20]. Оказалось, что указанная структура у Yersinia слегка отличается от структуры Shigella, поскольку имеет меньшие выступающие уплотнения и другой кончик.
Идентификация структурного элемента, содержащего IpaD на кончике иглы T3SA, способствует дальнейшему пониманию устройства и структуры T3SA S. flexneri. Совсем недавно с использованием биохимических методов было также показано, что IpaD локализуется на кончике иглы T3SA, где выполняет функции контроля секреции и дальнейшего встраивания транслокаторов в мембраны клеток-хозяев [22]. Настоящий одночастичный анализ тем не менее напрямую указывает на расположение IpaD на кончике иглы и дополнительно поддерживает гипотезу о том, что IpaD действует как блок (заглушка) в T3SA перед контактом бактерии с клетками. Также как в случая LcrV у Yersinia, предполагается, что IpaD может также способствовать встраиванию компонентов транслокаторов в клеточную мембрану. Ингибирование проникновения бактерий в клетки HeLa путем обработки клеток нейтрализующими анти-IpaD антителами указывает на то, что связывание антител с IpaD нарушает функционирование белка. В отношении LcrV также было показано, что он является защитным антигеном в отношении чумы при исследованиях на животных [23, 24]. Соответственно, IpaD представляет собой интересную мишень для приготовления вакцин, которые могут быть эффективны против нескольких серотипов Shigella.
Пример 4. Защитное действие анти-IpaD антител in vivo
Петли кишечника кролика инокулировали суспензией Shigella flexneri серотипа 5а в количестве 109 КОЕ или инкубировали в присутствии различных разведений поликлональной сыворотки кролика, специфичной к IpaD. Поликлональная анти-IpaD сыворотка была получена после иммунизации с использованием функционального производного белка IpaD, экспрессируемого в экспрессионном векторе pMal-IpaD, депонированном в базе CNCM 10 октября 2007 года под номером 1-3839. Данная модель объединяет ряд поражений, наблюдаемых при шигеллиозе у человека вследствие прободения, инвазии и воспалительного разрушения эпителия кишечника данной энтероинвазивной бактерией. Эти поражения проявляются как комбинация морфологических изменений кишечных ворсинок и отека, в комбинации с воспалительным клеточным фильтратом, в частности полиядерных нейтрофилов, и абсцессами, которые, в конечном итоге, изъязвляются в просвете кишечника. В итоге это приводит к слизистогнойным выделениям в просвете кишечника, которые часто являются инвазивными.
В результате сравнения поражения тканей кишечных петель, инокулированных только бактериями и бактериями в присутствии поликлональной анти-IpaD сыворотки, было обнаружено защитное действие, зависящее от концентрации анти-IpaD антител. Действительно, при использовании не разбавленной сыворотки не наблюдали никаких поражений. Однако при использовании сыворотки, разбавленной 1/10, появляются отдельные поражения, которые становятся более выраженными при использовании разведения 1/100, оставаясь при этом менее серьезными, чем поражения, наблюдаемые при введении бактерий без сыворотки. При использовании контрольной поликлональной сыворотки, полученной к другому, нерелевантному белку, защитного действия не наблюдали. Таким образом, наблюдаемое защитное действие специфично связано с присутствием антител анти-IpaD.





Группа изобретений относится к области медицины и касается IpaD - белка Shigella и его применения в качестве вакцины против инфекций, вызываемых Shigella. Сущность изобретений включает композиции и способы блокирования проникновения бактерий Shigella в клетки млекопитающих и, таким образом, к обеспечению защиты от инфекций, вызываемых бактериями рода Shigella, или снижению тяжести инфекций, вызываемых Shigella, включающих полипептид IpaD, способный вызывать получение нейтрализующих антител, обладающих защитным действием против некоторых серотипов бактерий Shigella, в частности S.flexneri. Преимущество изобретений заключается в создании композиций и способов их применения, полезных для лечения дизентерии, вызываемой Shigella. 7 н. и 15 з.п. ф-лы, 4 пр., 7 ил.
1. Композиция для блокирования проникновения по меньшей мере одного серотипа Shigella в пермиссивную клетку, включающая по меньшей мере один из следующих компонентов:
- полипептид IpaD или его фрагмент, который сохраняет способность вызывать продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному, или функциональное полипептидное производное указанного полипептида;
- полинуклеотид, кодирующий полипептид IpaD, или его фрагмент, который сохраняет способность кодировать полипептид IpaD, который вызывает продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному;
и/или
- нейтрализующее анти-lpaD антитело.
2. Полипептид IpaD или его фрагмент, который сохраняет способность вызывать продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному, или функциональное полипептидное производное указанного полипептида; или
- полинуклеотид, кодирующий полипептид IpaD, или его функциональный фрагмент, который сохраняет способность кодировать полипептид IpaD, который вызывает продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному; и/или
- нейтрализующее анти-lpaD антитело
для применения в лечении и/или предотвращении инфекции, вызываемой Shigella.
3. Композиция по п.1, отличающаяся тем, что указанный полипептид IpaD имеет последовательность аминокислот, идентичную SEQ ID NO:1, или последовательность аминокислот, обладающую по меньшей мере 75% идентичностью, или 85% идентичностью, или 95% идентичностью с SEQ ID NO:1.
4. Композиция по п.1, отличающаяся тем, что указанный полинуклеотид имеет последовательность нуклеотидов, идентичную SEQ ID NO:2, или последовательность нуклеотидов, обладающую по меньшей мере 65% идентичностью, или по меньшей мере 80% идентичностью, или по меньшей мере 95% идентичностью с SEQ ID NO:2.
5. Композиция по п.1, отличающаяся тем, что нейтрализующее анти-lpaD антитело является поликлональным.
6. Композиция по п.1, отличающаяся тем, что нейтрализующее анти-lpaD антитело является моноклональным.
7. Композиция по п.1, включающая дополнительно полиозидный антиген.
8. Применение по меньшей мере одного из следующих компонентов для блокирования проникновения по меньшей мере одного серотипа Shigella в пермиссивную клетку, где указанные компоненты представляют собой:
- полипептид IpaD, или функциональное полипептидное производное указанного полипептида, или его фрагмент, который сохраняет способность вызывать продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному;
- полинуклеотид, кодирующий полипептид IpaD, или его фрагмент, который сохраняет способность кодировать полипетид IpaD, который вызывает продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному;
и/или
- нейтрализующее анти-lpaD антитело.
9. Применение по меньшей мере одного из следующих компонентов для изготовления композиции для лечения и/или предотвращения инфекции, вызываемой Shigella, где указанные компоненты представляют собой:
- полипептид IpaD, или функциональное полипептидное производное указанного полипептида, или его фрагмент, который сохраняет
способность вызывать продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному;
- полинуклеотид, кодирующий полипептид IpaD, или его фрагмент, который сохраняет способность кодировать полипетид IpaD, который вызывает продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному;
и/или
- нейтрализующее анти-lpaD антитело.
10. Применение по любому из пп.8 или 9, отличающееся тем, что указанный полипептид IpaD имеет последовательность аминокислот, идентичную SEQ ID NO:1, или последовательность аминокислот, обладающую по меньшей мере 75% идентичностью, или по меньшей мере 85% идентичностью, или по меньшей мере 95% идентичностью с SEQ ID NO:1.
11. Применение по любому из пп.8 или 9, отличающееся тем, что указанный полинуклеотид имеет последовательность нуклеотидов, идентичную SEQ ID NO:2, или последовательность нуклеотидов, обладающую по меньшей мере 65% идентичностью, или по меньшей мере 80% идентичностью, или по меньшей мере 95% идентичностью с SEQ ID NO:2.
12. Применение по любому из любому из пп.8 или 9, отличающееся тем, что нейтрализующее анти-lpaD антитело является поликлональным.
13. Применение по любому из пп.8 или 9, отличающееся тем, что нейтрализующее анти-lpaD антитело является моноклональным.
14. Применение по любому из пп.8 или 9, включающее дополнительно полиозидный антиген.
15. Набор для блокирования проникновения по меньшей мере одного серотипа Shigella в пермиссивную клетку, включающий:
- полипептид IpaD или функциональное производное указанного полипептида или его фрагмент, который сохраняет способность вызывать
продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному;
- полинуклеотид, кодирующий полипептид IpaD, или его фрагмент, который сохраняет способность кодировать полипетид IpaD, который вызывает продукцию нейтрализующих анти-lpaD антител против штамма Shigella при введении животному; и/или
- нейтрализующее анти-lpaD антитело.
16. Способ лечения и/или предотвращения инфекции, вызываемой Shigella, у животных, включающий этап введения указанному животному композиции по любому из из пп.1, 3-7.
17. Способ блокирования проникновения по меньшей мере одного серотипа Shigella в пермиссивную клетку, включающий этап обеспечения возможности образования иммунного комплекса путем обеспечения контакта нейтрализующего анти-lpaD антитела со штаммом Shigella, способным к инфицированию указанной пермиссивной клетки, причем указанный иммунный комплекс предотвращает или существенно снижает проникновение указанного штамма Shigella в пермиссивную клетку.
18. Способ по п.17, представляющий собой способ in vivo.
19. Способ по п.17 или 18, в котором нейтрализующее анти-lpaD антитело продуцировано хозяином, обладающим указанными пермиссивными клетками, после введения указанному хозяину полипептида IpaD или функционального производного указанного полипептида и/или полинуклеотида или функционального фрагмента указанного полинуклеотида, кодирующего полипептид IpaD.
20. Способ по п.19, отличающийся тем, что указанный полипептид IpaD имеет последовательность аминокислот, идентичную SEQ ID NO:1, или последовательность аминокислот, обладающую по меньшей мере 75% идентичностью, или по меньшей мере 85% идентичностью, или по меньшей мере 95% идентичностью с SEQ ID NO:1.
21. Способ по п.19, отличающийся тем, что указанный полинуклеотид имеет последовательность нуклеотидов, идентичную или по существу аналогичную SEQ ID NO:2, или последовательность нуклеотидов, обладающую по меньшей мере 65% идентичностью, или по меньшей мере 80% идентичностью, или по меньшей мере 95% идентичностью с SEQ ID NO:2.
22. Способ по п.17, отличающийся тем, что нейтрализующее анти-lpaD антитело вводят хозяину, обладающему указанными пассивными клетками путем пассивной иммунизации указанного хозяина.
| WO 00/18355 A2, 06.04.2000 | |||
| ROSS K | |||
| et al | |||
| Isolation and Characterization of a Shigella Flexnery invasion complex subunit vaccine, Infect Immun., 2000, v.68, №12, pp.6624-6632 | |||
| MENARD R | |||
| et al | |||
| The secreted Ipa complex Shigella Flexnery promotes entry into mammalian cells, Pros | |||
| Natl., Acad | |||
| Sci USA, 1996, v.93, pp.1254-1258. |

