ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение в основном относится к антителам к DR5, включая агонистические антитела, и к способам применения таких антител к DR5.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
В данной области выявлены различные лиганды и рецепторы, принадлежащие к суперсемейству фактора некроза опухоли (TNF). В число таких лигандов включены фактор некроза опухоли-альфа ("TNF-альфа"), фактор некроза опухоли-бета ("TNF-бета или "лимфотоксин-альфа"), лимфотоксин-бета ("LT-бета"), лиганд CD30, лиганд CD27, лиганд CD40, лиганд OX-40, лиганд 4-1BB, LIGHT, лиганд Apo-1 (также обозначаемый как лиганд Fas или лиганд CD95), лиганд Apo-2 (также обозначаемый как Apo2L или TRAIL), лиганд Apo-3 (также обозначаемый как TWEAK), APRIL, лиганд OPG (также обозначаемый как лиганд RANK, ODF или TRANCE) и TALL-1 (также обозначаемый как BlyS, BAFF или THANK) (см., например, Ashkenazi, Nature Review. 2:420-430 (2002); Ashkenazi and Dixit, Science, 281:1305-1308 (1998); Ashkenazi and Dixit, Curr. Opin. Cell Biol., 11:255-260 (2000); Golstein, Curr. Biol., 7:750-753 (1997) Wallach, Cytokine Reference, Academic Press, 2000, pp.377-411; Locksley et al. Cell, 104:487-501 (2001); Grass and Dower, Blood, 85:3378-3404 (1995); Schmid et al., Proc. Natl. Acad. Sci., 83:1881 (1986); Dealtry et al., Eur. J. Immunol., 17:689 (1987); Pitti et al., J. Biol. Chem., 271:12687-12690 (1996); Wiley et al., Immunity, 3:673-682 (1995); Browning et al., Cell, 72:847-856 (1993); Arrnitage et al. Nature, 357:80-82 (1992), WO 97/01633, опубликованную 16 января 1997 года; WO 97/25428, опубликованную 17 июля 1997 года; Marsters et al., Curr. Biol., 8:525-528 (1998); Chicheportiche et al., Biol. Chem., 272:32401-32410 (1997); Hahne et al., J. Exp. Med., 188:1185-1190 (1998); WO 98/28426, опубликованную 2 июля 1998 года; WO 98/46751, опубликованную 22 октября 1998 года; WO 98/18921, опубликованную 7 мая 1998 года; Moore et al., Science, 285:260-263 (1999); Shu et al., J. Leukocyte Biol., 65:680 (1999); Schneider et al., J. Exp. Med., 189:1747-1756 (1999); Mukhopadhyay et al., J. Biol. Chem., 274:15978-15981 (1999)).
Как правило, индукцию различных видов клеточного ответа, опосредованного такими лигандами семейства TNF, вызывает их связывание со специфичными клеточными рецепторами. Некоторые, но не все, лиганды семейства TNF связываются с "рецепторами смерти" на поверхности клетки и индуцируют посредством их различные виды биологической активности, активируя каспазы или ферменты, приводящие к гибели клетки или осуществляющие каскад реакций апоптоза (Salvesen et al., Cell, 91:443-446 (1997)). В число выявленных к настоящему времени представителей суперсемейства рецепторов TNF входят TNFR1, TNFR2, TACI, GITR, CD27, OX-40, CD30, CD40, HVEM, Fas (также обозначаемый как Apo-1 или CD95), DR4 (также обозначаемый как TRAIL-R1), DR5 (также обозначаемый как Apo-2 или TRAIL-R2), DcR1, DcR2, остеопротегерин (OPG), RANK и Apo-3 (также обозначаемый как DR3 или TRAMP) (см., например, Ashkenazi, Nature Reviews, 2:420-430 (2002); Ashkenazi and Dixit, Science, 281:1305-1308 (1998); Ashkenazi and Dixit, Curr. Opin. Cell Biol., 11:255-260 (2000); Golstein, Curr. Biol., 7:750-753 (1997); Wallach, Cytokine Reference. Academic Press, 2000, pp.377-411; Locksley et al. Cell, 104:487-501 (2001); Grass and Dower, Blood, 85:3378-3404 (1995); Hohman et al., J. Biol. Chem., 264:14927-14934 (1989); Brockhaus et al., Proc. Natl. Acad. Sci., 87:3127-3131 (1990); EP 417563, опубликованную 20 марта 1991 года; Loetscher et al., Cell, 61:351 (1990); Schall et al., Cell, 61:361 (1990); Smith et al., Science, 248:1019-1023 (1990); Lewis et al., Proc. Natl. Acad. Sci., 88:2830-2834 (1991); Goodwin et al., Mol. Cell. Biol., 11:3020-3026 (1991); Stamenkovic et al., EMBO J., 8:1403-1410 (1989); Mallett et al., EMBO J., 9:1063-1068 (1990); Anderson et al., Nature, 390:175-179 (1997); Chicheportiche et al., J. Biol. Chem., 272:32401-32410 (1997); Pan et al., Science, 276:111-113 (1997); Pan et al., Science, 277:815-818 (1997); Sheridan et al., Science, 277:818-821 (1997); Degli-Esposti et al., J. Exp. Med., 186:1165-1170 (1997); Marsters et al. Curr. Biol., 7:1003-1006 (1997); Tsuda et al., BBRC, 234:137-142 (1997); Nocentini et al., Proc. Natl. Acad. Sci., 94:6216-6221 (1997); vonBulow et al., Science, 278:138-141 (1997)).
Большинство этих представителей семейства рецепторов TNF обладает обычной структурой рецепторов клеточной поверхности, включая внеклеточную, трансмембранную и внутриклеточную области, тогда как у других рецепторов, встречающихся в природе в виде растворимых белков, отсутствуют трансмембранный и внутриклеточный домены. Внеклеточный участок типичных TNFR, начиная с NH2-конца, содержит повторяющийся характерный участок аминокислотной последовательности из множества богатых цистеином доменов (CRD).
Несколько лет назад как представитель семейства цитокинов TNF был идентифицирован лиганд, обозначаемый как Apo-2L или TRAIL (см., например, Wiley et al., Immunity, 3:673-682 (1995); Pitti et al., J. Biol. Chem., 271:12697-12690 (1996); WO 97/01633; WO 97/25428; патент США №5763223, выданный 9 июня 1998 года; патент США №6284236, выданный 4 сентября 2001 года). Полноразмерная природная последовательность полипептида Apo2L/TRAIL человека представляет собой трансмембранный белок II типа, длиной в 281 аминокислоту. Некоторые клетки могут продуцировать природную растворимую форму полипептида посредством ферментативного отщепления внеклеточной области полипептида (Mariani et al., J. Cell. Biol., 137:221-229 (1997)). Кристаллографические исследования растворимых форм Apo2L/TRAIL выявили гомотримерную структуру, сходную со структурами TNF и других родственных белков (Hymowitz et al., Molec. Cell, 4:563-571 (1999); Cha et al., Immunity, 11:253-261 (1999); Mongkolsapaya et al., Nature Structural Biology, 6:1048 (1999); Hymowitz et al., Biochemistry, 39:633-644 (2000)). Однако было обнаружено, что в отличие от других представителей семейства TNF, Apo2L/TRAIL обладает уникальным структурным свойством в том, что три остатка цистеина (в положении 230 каждой субъединицы в гомотримере) вместе координируют атом цинка и что связывание цинка важно для стабильности и биологической активности тримера (Hymowitz et al., выше; Bodmer et al., J. Biol. Chem., 275:20632-20637 (2000)).
В литературе было описано, что Apo2L/TRAIL может играть роль в регуляции иммунной системы, в том числе при аутоиммунных заболеваниях, таких как ревматоидный артрит (см., например, Thomas et al., J. Immunol, 161:2195-2200 (1998); Johnsen et al., Cytokine, 11:664-672 (1999); Griffith et al., J. Exp. Med., 189:1343-1353 (1999); Song et al., J. Exp. Med., 191:1095-1103 (2000)).
Также было описано, что растворимые формы Apo2L/TRAIL индуцируют апоптоз во множестве раковых клеток, включая опухоли толстой кишки, легких, молочной железы, предстательной железы, мочевого пузыря, почек, яичников и головного мозга, а также меланому, лейкоз и множественную миелому (см., например, Wiley et al., выше; Pitti et al., выше; патент США № 6030945, выданный 29 февраля 2000 года; патент США № 6746668, выданный 8 июня 2004 года; Rieger et al., FEBS Letters, 427:124-128 (1998); Ashkenazi et al., J. Clin. Invest., 104:155-162 (1999); Walczak et al., Nature Med., 5:157-163 (1999); Keane et al., Cancer Research, 59:734-741 (1999); Mizutani et al., Clin. Cancer Res., 5:2605-2612 (1999); Gazitt, Leukemia, 13:1817-1824 (1999); Yu et al., Cancer Res., 60:2384-2389 (2000); Chinnaiyan et al., Proc. Natl. Acad. Sci., 97:1754-1759 (2000)). Исследования на моделях опухолей на мышах in vivo позволили предположить, что Apo2L/TRAIL, отдельно или в сочетании с химиотерапией или лучевой терапией, может оказывать значительные противоопухолевые эффекты (см., например, Ashkenazi et al., выше; Walzcak et al., выше; Gliniak et al., Cancer Res., 59:6153-6158 (1999); Chinnaiyan et al., выше; Roth et al., Biochem. Biophys. Res. Comm., 265:1999 (1999); заявка PCT США/00/15512; заявка PCT США/01/23691). В отличие от многих типов злокачественных клеток у большинства обычных типов клеток человека наблюдают устойчивость к индукции апоптоза определенными рекомбинантными формами Apo2L/TRAIL (Ashkenazi et al., выше; Walzcak et al., выше). Jo et al. описывали, что меченная полигистидином растворимая форма Apo2L/TRAIL индуцировала апоптоз in vitro в нормальных выделенных гепатоцитах человека, но не в не принадлежащих человеку гепатоцитах (Jo et al., Nature Med., 6:564-567 (2000); см. также Nagata, Nature Med., 6:502-503 (2000)). Полагают, что определенные препараты рекомбинантного Apo2L/TRAIL могут различаться по биохимическим свойствам и биологической активности в отношении патологических клеток по сравнению с нормальными, в зависимости, например, от наличия или отсутствия молекулы-метки, содержания цинка и % содержания тримера (см., Lawrence et al., Nature Med., Letter to the Editor, 7:383-385 (2001); Qin et al., Nature Med., Letter to the Editor, 7:385-386 (2001)).
Обнаружено, что Apo2L/TRAIL связывает, по меньшей мере, пять различных рецепторов. По меньшей мере, два из рецепторов, связывающихся с Apo2L/TRAIL, содержат функциональный цитоплазматический домен смерти. Один такой рецептор был обозначен как "DR4" (а альтернативно как TR4 или TRAIL-R1) (Pan et al., Science, 276:111-113 (1997); см. также WO 98/32856, опубликованную 30 июля 1998 года; WO 99/37684, опубликованную 29 июля 1999 года; WO 00/773349, опубликованную 7 декабря 2000 года; патент США №6433147, выданный 13 августа 2002 года; патент США №6461823, выданный 8 октября 2002 года, и патент США №6342383, выданный 29 января 2002 года).
Другой такой рецептор для Apo2L/TRAIL был обозначен как DR5 (также он был альтернативно обозначен как Apo-2; TRAIL-R или TRAIL-R2, TR6, Tango-63, hAPO8, TRICK2 или KILLER) (см., например, Sheridan et al., Science, 277:818-821 (1997), Pan et al., Science, 277:815-818 (1997), WO 98/51793, опубликованную 19 ноября 1998 года; WO 98/41629, опубликованную 24 сентября 1998 года; Screaton et al., Curr. Biol., 7:693-696 (1997); Walczak et al., EMBO J., 16:5386-5387 (1997); Wu et al., Nature Genetics, 17:141-143 (1997); WO 98/35986, опубликованную 20 августа 1998 года; EP870827, опубликованную 14 октября 1998 года; WO 98/46643, опубликованную 22 октября 1998 года; WO 99/02653, опубликованную 21 января 1999 года; WO 99/09165, опубликованную 25 февраля 1999 года; WO 99/11791, опубликованную 11 марта 1999 года; патент США №2002/0072091, выданный 13 августа 2002 года; патент США №2002/0098550, выданный 7 декабря 2001 года; патент США №6313269, выданный 6 декабря 2001 года; патент США №2001/0010924, выданный 2 августа 2001 года; патент США №2003/01255540, выданный 3 июля 2003 года; патент США №2002/0160446, выданный 31 октября 2002 года; патент США №2002/0048785, выданный 25 апреля 2002 года; патент США №6342369, выданный в феврале 2002 года; патент США №6569642, выданный 27 мая 2003 года; патент США №6072047, выданный 6 июня 2000 года; патент США №6642358, выданный 4 ноября 2003 года; патент IS 6743625, выданный 1 июня 2004 года). Описано, что, подобно DR4, DR5 содержит три богатых цистеином домена в своей внеклеточной части и один цитоплазматический домен смерти и способен передавать сигнал апоптоза при связывании лиганда (или при связывании молекулы, такой как антитело-агонист, воспроизводящей активность лиганда). Кристаллическая структура комплекса, формируемого Apo-2L/TRAIL и DR5, описана в Hymowitz et al., Molecular Cell, 4:563-571 (1999).
При связывании лиганда DR4 и DR5 способны запускать апоптоз независимо от вовлечения и активации инициатора апоптоза, каспазы-8, посредством содержащей домен смерти вспомогательной молекулы, которую обозначают как FADD/Mort1 (Kischkel et al., Immunity, 12:611-620 (2000); Sprick et al., Immunity, 12:599-609 (2000); Bodmer et al., Nature Cell Biol., 2:241-243 (2000)). В частности, DR5 передает сигнал апоптоза через "внешний по отношению к клетке" путь, независимый от опухолевого супрессорного гена p53 (Ashkenazi and Dixit, Science 281:1305-8 (1998); Ashkenazi, Nat. Rev. Cancer 2:420-30 (2002)). Активация этого пути включает в себя формирование воздействия индуцирующего смерть сигнального комплекса (DISC) на цитоплазматический домен смерти активированного рецептора. Сначала адаптерная молекула FADD связывается с DR5 посредством гомофильного взаимодействия доменов смерти (Kischkel et al., выше, Sprick et al., выше, Bodmer et al., выше). Затем FADD привлекает инициирующие апоптоз протеазы каспазу-8 и каспазу-10, опосредуя их активацию посредством индуцированного сближения. Каспаза-9 и каспаза-10 претерпевают самопроцессинг с высвобождением растворимых активных субъединиц каспаз в цитоплазму, где они собираются и расщепляют эффекторные каспазы, такие как каспаза-3 и каспаза-7. Расщепление приводит к активации эффекторных каспаз, осуществляющих клеточную программу апоптоза (Thornberry and Lazebnik, Science 281:1312-6 (1998)).
Было описано, что Apo2L/TRAIL также связывает те рецепторы, которые обозначены как DcR1, DcR2 и OPG, как предполагают, функционирующие, в качестве ингибиторов, а не передатчиков сигнала (например, см., DcR1 (также обозначаемый как TRID, LIT или TRAIL-R3) (Pan et al., Science, 276:111-113 (1997); Sheridan et al., Science, 277:818-821 (1997); McFarlane et al., J. Biol. Chem., 272:25417-25420 (1997); Schneider et al., FEBS Letters, 416:329-334 (1997); Degli-Esposti et al., J. Exp. Med., 186:1165-1170 (1997) и Mongkolsapaya et al., J. Immunol., 160:3-6 (1998)); DcR2 (также называемый TRUNDD или TRAIL-R4) (Marsters et al., Curr. Biol., 7:1003-1006 (1997); Pan et al., FEBS Letters, 424:41-45 (1998); Degli-Esposti et al., Immunity, 7:813-820 (1997)) [Simonet et al., выше], а также OPG. В отличие от DR4 и DR5, рецепторы DcR1 и DcR2 не передают сигнал апоптоза.
В литературе были описаны определенные антитела, связывающиеся с рецепторами DR4 и/или DR5. Например, антитела к DR4, направленные к рецептору DR4 и обладающие агонистической или апоптотической активностью в определенных клетках млекопитающих, описаны, например, в WO 99/37684, опубликованной 29 июля 1999 года; WO 00/73349, опубликованной 12 июля 2000 года; WO 03/066661, опубликованной 14 августа 2003 года. См. также, например, Griffith et al., J. Immunol., 162:2597-2605 (1999); Chuntharapai et al., J. Immunol., 166:4891-4898 (2001); WO 02/097033, опубликованную 2 декабря 2002 года; WO 03/042367, опубликованную 22 мая 2003 года; WO 03/038043, опубликованную 8 мая 2003 года; WO 03/037913, опубликованную 8 мая 2003 года. Также были описаны некоторые антитела против DR5, например, см. WO 98/51793, опубликованную 8 ноября 1998 года; Griffith et al., J. Immunol., 162:2597-2605 (1999); Ichikawa et al., Nature Med., 7:954-960 (2001); Hylander et al., "An Antibody to DR5 (TRAIL-Receptor 2) Suppresses the Growth of Patient Derived Gastrointestinal Tumors Grown in SCID mice", Abstract, 2d International Congress on Monoclonal Antibodies in Cancers, Aug. 29-Sept. 1, 2002, Banff, Alberta, Canada; WO 03/038043, опубликованную 8 мая 2003 года; WO 03/037913, опубликованную 8 мая 2003 года. Кроме того, были описаны некоторые антитела, обладающие перекрестной реактивностью в отношении обоих рецепторов DR4 и DR5 (см., например, патент США №6252050, выданный 26 июня 2001 года).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к антителам к DR5, которые способны к специфическому связыванию с DR5 человека и/или способны к регуляции биологической активности, ассоциированной с DR5 и/или его лигандом(ами), в частности апоптоза, и, таким образом, пригодны для лечения различных заболеваний и патологических состояний, включая злокачественную опухоль или связанные с иммунной системой заболевания.
В одном из аспектов изобретение относится к антителу к DR5, несущему, по меньшей мере, одну мутацию в тяжелой и/или легкой цепи полноразмерного антитела 16E2 (SEQ ID NO: 11 и 13, соответственно), или его фрагменту, где антитело или фрагмент антитела демонстрирует, по меньшей мере, такую же аффинность к DR5 и/или проявляет, по меньшей мере, такую же биологическую активность и/или эффективность, как антитело 16E2. В конкретном варианте осуществления антитело или фрагмент антитела связываются по существу с тем же эпитопом, что и полноразмерное антитело 16E2. В другом варианте осуществления антитело к DR5 проявляет большую аффинность к DR5, чем полноразмерное антитело 16E2, и/или демонстрирует увеличенную биологическую активность и/или увеличенную эффективность по отношению к полноразмерному антителу 16E2. В еще одном варианте осуществления антитела и фрагменты антител к DR5 по настоящему изобретению демонстрируют, по меньшей мере, такую же аффинность к DR5 и/или проявляют, по меньшей мере, такую же биологическую активность и/или эффективность, как и антитело к DR5 с одной цепью Fc 16E2, описанное в WO 98/51793.
В одном из вариантов осуществления антитело к DR5 содержит тяжелую и/или легкую цепь, по меньшей мере, с одной заменой из перечисленных в любой из таблиц с 1 по 7 и 9-12 или их фрагмент.
В другом варианте осуществления антитело к DR5 несет одну или несколько мутаций в каркасной области вариабельного домена тяжелой цепи антитела 16E2.
В еще одном варианте осуществления антитело к DR5 несет мутацию в каркасной области, выбранную из группы, состоящей из Q6E, V11L, E12V, R13Q и K105Q.
В дополнительном варианте осуществления антитело к DR5 несет все мутации в каркасной области Q6E, V11L, E12V, R13Q и K105Q.
В еще одном дополнительном варианте осуществления антитело к DR5 несет, по меньшей мере, одну мутацию в тяжелой цепи (SEQ ID NO: 11) полноразмерного антитела 16E2 или его фрагмента.
В другом варианте осуществления антитело к DR5 несет, по меньшей мере, одну мутацию в аминокислотной последовательности SEQ ID NO: 11 или ее фрагменте, выбранную из группы, состоящей из T28A, G33A, M34L, M34A, M34I, M34S, N53Q, N53Y и L102Y.
В другом варианте осуществления антитело к DR5 несет, по меньшей мере, одну из мутаций G99A и R100A в аминокислотной последовательности SEQ ID NO: 11 или ее фрагменте.
В еще одном варианте осуществления антитело к DR5 в аминокислотной последовательности SEQ ID NO: 11 или ее фрагменте несет набор мутаций, выбранный из группы, состоящей из (i) N53Q, L102Y; (ii) M34L, N53Q, L102Y; (iii) N53Y, L102Y; (iv) M34L, N53Y, L102Y; (v) G33A, N53Q, L102Y; (vi) M34L, N53Y, L102Y; (vii) G33A, N53Q, L102Y; (viii) G33A, N53Y, L102Y; (ix) T28A, N53Q, L102Y и (x) T28A, N53Y, L102Y.
В дополнительном варианте осуществления антитело к DR5 несет, по меньшей мере, одну мутацию в легкой цепи (SEQ ID NO: 13) полноразмерного антитела 16E2 или его фрагменте.
В конкретном варианте осуществления легкая цепь представляет собой лямбда-цепь.
В другом конкретном варианте осуществления мутация в легкой цепи находится в L1 CDR.
В дополнительном варианте осуществления мутация в легкой цепи в аминокислотной последовательности SEQ ID NO: 13 выбрана из группы, состоящей из Q24A, Q24S, G25A, D26E, S27A, L28A, R29A, S30A, Y31A, Y31K, Y32H, A33G, S34A и S34Y.
В еще одном дополнительном варианте осуществления мутация в легкой цепи в аминокислотной последовательности SEQ ID NO: 13 выбрана из группы, состоящей из (i) Q24S, D26E, Y31K, S34Y и (ii) D26E, Y31K.
В другом варианте осуществления мутация в легкой цепи находится в L2 CDR.
Например, таким образом, мутацию можно выбрать из группы, состоящей из G50A, G50K, G50S, K51D, N52A, N52S, N52L, N52Q, N53A, N53E, N53Q, N53S, P55A и S56A в аминокислотной последовательности SEQ ID NO: 13.
В другом варианте осуществления антитело может нести набор мутаций, выбранных из группы, состоящей из (i) G50K, K52S, N53E; (ii) G50S, K51D, N52S, N53E; (ii) N52S, N53E и (iv) N52Q, N53S в аминокислотной последовательности SEQ ID NO: 13.
В дополнительном варианте осуществления мутация в легкой цепи находится в L3 CDR.
В еще одном дополнительном варианте осуществления антитело несет, по меньшей мере, одну мутацию, выбранную из группы, состоящей из N89A, N89L, N89Q, R91A, S93A, N95aA, N95aT, N95aQ, H95bA, N95bY, V96A, V97A в SEQ ID NO: 13.
Альтернативно антитело к DR5 может нести набор мутаций, выбранных из группы, состоящей из (i) N89L, R91A, N95aT, H95bY и (ii) N95aT, H95bY в последовательности SEQ ID NO: 13.
В дополнительном варианте осуществления антитело к DR5 несет набор мутаций в легкой цепи, выбранный из группы, состоящей из (i) Q24S, G50K, K51D, H95bY; (ii) Q24S, K51A, D92S, S93Y и (iii) Q24S, K51A, R91A в аминокислотной последовательности SEQ ID NO: 13, и может дополнительно нести набор мутаций в тяжелой цепи, выбранный из группы, состоящей из (i) M34L, N53Q, L102Y; (ii) M34L, N53Y, L102Y; (iii) G33A, N53Q, L102Y; (iv) G33A, N53Y, L102Y; (v) M34L, N53Q, L102Y; (vi) M34L, N53Y, L103Y; (vii) G33A, N53Q, L102Y; (viii) G33A, N53Y, L102Y и (ix) T28A, N53Q, L102Y в аминокислотной последовательности SEQ ID NO: 11, и, необязательно, набор мутаций в каркасной области, перечисленных в таблице 5.
В конкретном варианте осуществления антитело к DR5 несет следующие мутации: G33A, N53Q, L102Y в последовательности SEQ ID NO: 11 и Q24S, K51A, R91A в последовательности SEQ ID NO: 13, и может дополнительно нести, по меньшей мере, одну мутацию в каркасной области, которая может представлять собой, например, по меньшей мере, один из остатков 6, 11, 12, 13 и 105 SEQ ID NO: 11.
В конкретном варианте осуществления антитело к DR5 выбрано из группы, состоящей из Apomab 1.1, 2.1, 3.1, 4.1, 5.1, 6.1, 7.1, 8.1, 9.1, 1.2, 2.2, 3.2, 4.2, 5.2, 6.2, 7.2, 8.2, 9.2, 1.3, 2.2, 3.3, 4.3, 5.3, 6.3, 7.3, 8.3 и 9.3.
В конкретном варианте осуществления антитело к DR5 выбрано из группы, состоящей из Apomab 5.2, 5.3, 6.2, 6.3, 7.2, 7.3, 8.3 и 25.3.
В еще одном конкретном варианте осуществления антитело к DR5 представляет собой Apomab 7.3 или Apomab 8.3, особенно Apomab 7.3.
В еще одном дополнительном варианте осуществления антитело к DR5 представляет собой фрагмент антитела, который можно выбрать из группы, состоящей из фрагментов Fab, Fab', F(ab')2 и Fv, димерных антител, одноцепочечных молекул антител и полиспецифических антител, сформированных из фрагментов антител.
В других вариантах осуществления антитело может представлять собой одноцепочечное антитело.
Например, антитела к DR5 могут обладать активностью в отношении злокачественных опухолей, например, такой, что они могут обладать способностью активировать или стимулировать апоптоз в злокачественных клетках.
Злокачественная опухоль включает в себя, например, карциному, лимфому, бластому, саркому и лейкоз.
Более конкретные примеры злокачественной опухоли включают плоскоклеточный рак, мелкоклеточный рак легких, немелкоклеточный рак легких (NSCLC), неходжкинскую лимфому, бластому, желудочно-кишечный рак, рак почек, рак яичников, рак печени, рак желудка, рак мочевого пузыря, гепатому, рак молочной железы, рак толстой кишки, колоректальный рак, рак поджелудочной железы, карциному эндометрия, карциному слюнной железы, рак почки, рак печени, рак предстательной железы, рак женских наружных половых органов, рак щитовидной железы, печеночную карциному и рак головы и шеи.
Конкретные группы злокачественных опухолей включают в себя рак легких (например, немелкоклеточную карциному легких - NSCLC) или аденокарциному, которая может представлять собой, например, колоректальную аденокарциному, аденокарциному поджелудочной железы или метастатическую аденокарциному. Также включены злокачественные опухоли гематологического происхождения.
В объем настоящего документа включены химерные, гуманизированные антитела и антитела человека, такие как антитела, опосредующие обусловленную антителами клеточную цитотоксичность (ADCC).
В предпочтительном варианте осуществления антитело к DR5 представляет собой Apomab 7.3 или Apomab 8.3 или его фрагмент.
Антитела могут находиться в димерной форме и/или в форме, перекрестно связанной, например, с Fc-областью IgG к антителам человека.
В других вариантах осуществления антитела к DR5 по настоящему документу слиты с последовательностью эпитопной метки.
В другом аспекте изобретение относится к химерной молекуле, содержащей антитело или фрагмент антитела к DR5 по настоящему документу, слитые с гетерологичной аминокислотной последовательностью, где гетерологичная аминокислотная последовательность, например, может содержать иммуноглобулиновую последовательность, такую как Fc-область IgG к антителам человека.
В еще одном аспекте изобретение относится к выделенным молекулам нуклеиновой кислоты, кодирующим антитела или фрагменты антител к DR5 по настоящему документу, векторам, несущим такие молекулы нуклеиновой кислоты, клеткам-хозяевам, несущим такие молекулы нуклеиновой кислоты, и способам получения антител и фрагментов антител по настоящему документу.
Изобретение дополнительно относится к композиции, содержащей антитело к DR5, как определено выше в настоящем документе, и носитель.
Носитель может представлять собой фармацевтически приемлемый носитель, а композиция может дополнительно содержать дополнительное противораковое средство и/или дополнительное антитело к DR5.
В дополнительном аспекте изобретение относится к способу индукции апоптоза, включающему в себя воздействие на злокачественные клетки млекопитающих антитела к DR5, как определено выше в настоящем документе.
В еще одном дополнительном аспекте изобретение относится к способу лечения злокачественной опухоли, включающему в себя введение млекопитающему эффективного количества антитела к DR5, как определено выше.
Во всех аспектах субъект может представлять собой пациента-человека, а злокачественная опухоль может представлять собой любую злокачественную опухоль, включая злокачественные опухоли, перечисленные выше.
В дополнительном аспекте изобретение относится к промышленному изделию, содержащему контейнер и композицию, содержащуюся в указанном контейнере, где композиция содержит антитело к DR5 по настоящему изобретению. Промышленное изделие может дополнительно содержать инструкции по применению антитела к DR5 in vitro или in vivo. В предпочтительном варианте осуществления инструкции относятся к лечению злокачественной опухоли.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 приведена нуклеотидная последовательность кДНК лиганда Apo-2 человека (SEQ ID NO: 2) и выведенная из нее аминокислотная последовательность (SEQ ID NO: 1). "N" в положении нуклеотида 447 (в SEQ ID №2) используют для указания того, что нуклеотидное основание может представлять собой "T" или "G".
На фиг.2A-2C приведена нуклеотидная последовательность кДНК (SEQ ID NO: 4) полноразмерного рецептора DR4 человека и полученная из нее аминокислотная последовательность (SEQ ID NO: 3). О соответствующих нуклеотидной и аминокислотной последовательностях рецептора DR4 человека также сообщалось Pan et al., Science, 276:111 (1997).
На фиг.3A-3C приведена последовательность DR5 человека из 411 аминокислот рецептора (SEQ ID NO: 5), как опубликовано в WO 98/51793 19 ноября 1998 года, и кодирующая нуклеотидная последовательность (SEQ ID NO: 6).
На фиг.4A-4C приведена последовательность DR5 человека из 440 аминокислот (SEQ ID NO: 7) и кодирующая нуклеотидная последовательность (SEQ ID NO: 8), как также опубликовано в WO 98/35986 20 августа 1998 года.
На фиг.5 приведена нуклеотидная последовательность одноцепочечного антитела к DR5 16E2 (scFv 16E2) (SEQ ID NO: 9).
На фиг.6 приведена аминокислотная последовательность одноцепочечного антитела к DR5 16E2 (scFv 16E2) (SEQ ID NO: 10), где показаны сигнальная последовательность и CDR тяжелой и легкой цепей.
На фиг.7 приведена аминокислотная последовательность тяжелой цепи полноразмерного антитела 16E2 (SEQ ID NO: 11).
На фиг.8 приведена нуклеотидная последовательность тяжелой цепи полноразмерного антитела 16E2 (SEQ ID NO: 12).
На фиг.9 приведена аминокислотная последовательность легкой цепи полноразмерного антитела 16E2 (SEQ ID NO: 13).
На фиг.10 приведена нуклеотидная последовательность легкой цепи полноразмерного антитела 16E2 (SEQ ID NO: 14).
На фиг.11A и B приведена последовательность плазмиды pDR1 (SEQ ID NO: 15, 5391 п.н.) для экспрессии легких цепей иммуноглобулинов. pDR1 содержит последовательности, кодирующие не относящееся к изобретению антитело, легкую цепь гуманизированного антитела к CD3 (Shalaby et al., J. Exp. Med. 175:217-225 (1992)), старт- и стоп-кодоны которого указаны жирным шрифтом и подчеркнуты.
На фиг.12A и B приведена последовательность плазмиды pDR2 (SEQ ID NO: 16) для экспрессии тяжелых цепей иммуноглобулинов. pDR2 содержит последовательности, кодирующие не относящееся к изобретению антитело, тяжелую цепь гуманизированного антитела к CD3 (Shalaby et al., выше), старт- и стоп-кодоны которого указаны жирным шрифтом и подчеркнуты.
На фиг.13 приведена нуклеотидная последовательность тяжелой цепи Apomab 7.3 (SEQ ID NO: 17).
На фиг.14 приведена аминокислотная последовательность тяжелой цепи Apomab 7.3 (SEQ ID NO: 18).
На фиг.15 приведена нуклеотидная последовательность легкой цепи Apomab 7.3 (SEQ ID NO: 19).
На фиг.16 приведена аминокислотная последовательность легкой цепи Apomab 7.3 (SEQ ID NO: 20).
На фиг.17A и B приведено выравнивание тяжелых цепей 16E2 и Apomab 7.3.
На фиг.18 приведено выравнивание легких цепей 16E2 и Apomab 7.3.
Фиг.19 представляет собой модель гомологии для тяжелой цепи антитела к DR5.
Фиг.20 представляет собой модель гомологии для легкой цепи антитела к DR5.
На фиг.21 показана противораковая активность одной интраперитонеальной (и/п) дозы Apomab 5.3, 6.3 и 8.3 по сравнению с полноразмерным антителом 16E2 (версия 1) в модели рака толстой кишки человека у бестимусной мыши "nude" с ксенотрансплантатом Colo205.
На фиг.22 показана противораковая активность одной и/п дозы Apomab 5.2, 6.2, 5.3, 7.2 и 7.3 по сравнению с полноразмерным антителом 16E2 (версия 1) в модели рака толстой кишки человека у бестимусной мыши "nude" с ксенотрансплантатом Colo205.
На фиг.23 показана противораковая активность одной и/п дозы Apomab 5.2, 7.3 и 8.3 по сравнению с полноразмерным антителом 16E2 (версия 1) в модели рака толстой кишки человека у бестимусной мыши "nude" с ксенотрансплантатом Colo205.
На фиг.24 показана противораковая активность Apomab 23.3 и 25.3 по сравнению с Apomab 7.3 в модели рака толстой кишки человека у бестимусной мыши "nude" с ксенотрансплантатом Colo205.
На фиг.25 показана противораковая активность Apomab 7.3, полученного из стабильной клеточной линии, по сравнению с транзиторной клеточной линией в модели рака толстой кишки человека у бестимусной мыши "nude" с ксенотрансплантатом Colo205.
На фиг.26 показана противораковая активность Apomab 7.3 отдельно и в сочетании с CPT-11 в модели рака легких на ксенотрансплантате HCT15.
На фиг.27 показана противораковая активность Apomab 7.3 отдельно и в сочетании с CPT-11 в модели саркомы человека на ксенотрансплантате LS180.
На фиг.28 показана противораковая активность Apomab 7.3 отдельно и в сочетании с RITUXAN® (ритуксимаб) в модели неходжкинской лимфомы у мышей SCID CB17 ICR с ксенотрансплантатом BJAB.
На фиг.29 показана противораковая активность Apomab 7.3 отдельно и в сочетании с гемцитабином в модели аденокарциномы поджелудочной железы человека у бестимусных мышей "nude" с ксенотрансплантатом BxPC3.
На фиг.30 показана противораковая активность Apomab 7.3 отдельно и в сочетании с карбоплатином и таксолом в модели рака легких человека на ксенотрансплантате H460.
На фиг.31 показана противораковая активность Apomab 7.3 отдельно и в сочетании с карбоплатином и таксолом в модели рака легких человека на ксенотрансплантате H2122.
На фиг.32 приведена кривая доза-ответ Apomab 7.3 в модели рака легких человека на ксенотрансплантате H2122.
На фиг.33 показана противораковая активность Apomab 23.3 и 25.3 по сравнению с Apomab 7.3 в модели рака толстой кишки человека на ксенотрансплантате Colo205.
На фиг.34 приведены средний рост опухоли и диаграмма Каплана-Майера для Apo2L.0 отдельно, Apomab 7.3 отдельно и различных сочетаний по отношению к ксенотрансплантатам карциномы толстой кишки Colo205 у мышей "nude".
На фиг.35 и 36 приведены средний рост опухоли и диаграмма Каплана-Майера для Apo2L.0 отдельно, Apomab 7.3 отдельно и различных сочетаний по отношению к модели немелкоклеточной карциномы легких (NSCLC) человека у бестимусных мышей "nude" с ксенотрансплантатом клеток SKMES-I.
На фиг.37 приведены средний рост опухоли и диаграмма Каплана-Майера для Apo2L.0 отдельно, Apomab 7.3 отдельно и различных сочетаний в модели карциномы толстой кишки человека на ксенотрансплантате Colo205.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
I. Определения
Термины "лиганд Apo-2", "Apo-2L", "Apo2L", "лиганд Apo-2/TRAIL" и "TRAIL" в настоящем документе применяют взаимозаменяемо для обозначения полипептидной последовательности, содержащей аминокислотные остатки 114-281, включительно, 95-281, включительно, остатки 92-281, включительно, остатки 91-281, включительно, остатки 41-281, включительно, остатки 39-281, включительно, остатки 15-281, включительно, или остатки 1-281, включительно, аминокислотной последовательности, приведенной на фиг.1 (SEQ ID NO: 1), а также биологически активных фрагментов, делеционных, инсерционных и/или замещенных вариантов указанных выше последовательностей. В одном из вариантов осуществления полипептидная последовательность содержит остатки 114-281 из фиг.1 (SEQ ID NO: 1). Необязательно полипептидная последовательность содержит остатки 92-281 или остатки 91-281 из фиг.1 (SEQ ID NO: 1). Полипептиды Apo-2L может кодировать природная нуклеотидная последовательность, приведенная на фиг.1 (SEQ ID NO: 2). Необязательно кодон, кодирующий остаток Pro119 (фиг.1; SEQ ID NO: 2), может представлять собой "CCT" или "CCG". Необязательно фрагменты или варианты являются биологически активными и обладают, по меньшей мере, приблизительно 80% идентичностью аминокислотных последовательностей, или, по меньшей мере, приблизительно 90% идентичностью последовательностей, или, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичностью последовательностей с любой из указанных выше последовательностей. Определение включает в себя заместительные варианты лиганда Apo-2, в которых, по меньшей мере, одна из его природных аминокислот замещена другой аминокислотой, такой как аланиновый остаток. Определение также включает в себя природную последовательность лиганда Apo-2, выделенную из источника лиганда Apo-2 или полученную рекомбинантными и/или синтетическими способами. Лиганд Apo-2 по изобретению включает в себя полипептиды, обозначаемые как лиганд Apo-2 или TRAIL, описанные в WO 97/01633, опубликованной 16 января 1997 года, WO 97/25428, опубликованной 17 июля 1997 года, WO 99/36535, опубликованной 22 июля 1999 года, WO 01/00832, опубликованной 4 января 2001 года, WO02/09755, опубликованной 7 Февраля 2002 года, WO 00/75191, опубликованной 14 декабря 2000 года, и в патенте США №6030945, выданном 29 февраля 2000 года. Термины, как правило, используют для обозначения форм лиганда Apo-2, которые включают в себя мономерные, димерные, тримерные, гексамерные или высоко олигомерные формы полипептида. Если не указано иначе, при нумерации аминокислотных остатков, указанных в последовательности Apo-2L, используют нумерацию в соответствии с фиг.1 (SEQ ID NO: 1).
"Рецептор лиганда Apo-2" включает в себя рецепторы, обозначаемые в данной области как "DR4" и "DR5", полинуклеотидные и полипептидные последовательности которых показаны на фиг.2A-2C (SEQ ID NO: 4 и 3) и 3A-3C (SEQ ID NO: 6 и 5), соответственно. У Pan et al. описан представитель семейства рецептора TNF, обозначаемый как "DR4" (Pan et al., Science, 276:111-113 (1997); также см. WO 98/32856, опубликованную 30 июля 1998 года; WO 99/37684, опубликованную 29 июля 1999 года; WO 00/73349, опубликованную 7 декабря 2000 года; патент США №6433147, выданный 13 августа 2002 года; патент США №6461823, выданный 8 октября 2002 года, и патент США №6342383, выданный 29 января 2002 года). У Sheridan et al., Science, 277:818-821 (1997), и Pan et al., Science, 277:815-818 (1997), описан другой рецептор для Apo2L/TRAIL (также см., WO 98/51793, опубликованную 19 ноября 1998 года; WO 98/41629, опубликованную 24 сентября 1998 года). Этот рецептор обозначают как DR5 (этот рецептор также альтернативно обозначают как Apo-2; TRAIL-R, TR6, Tango-63, hAPO8, TRICK2 или KILLER; Screaton et al., Curr. Biol., 7:693-696 (1997); Walczak et al., EMBO J., 16:5386-5387 (1997); Wu et al., Nature Genetics, 17:141-143 (1997); WO 98/35986, опубликованная 20 августа 1998 года; EP 870827, опубликованный 14 октября 1998 года; WO 98/46643, опубликованная 22 октября 1998 года; WO 99/02653, опубликованная 21 января 1999 года; WO 99/09165, опубликованная 25 февраля 1999 года; WO 99/11791, опубликованная 11 марта 1999 года; US 2002/0072091, опубликованная 13 августа 2002 года; US 2002/0098550, опубликованная 7 декабря 2001 года; патент США №6313269, выданный 6 декабря 2001 года; US 2001/0010924, опубликованная 2 августа 2001 года; US 2003/01255540, опубликованная 3 июля 2003 года; US 2002/0160446, опубликованная 31 октября 2002 года, US 2002/0048785, опубликованная 25 апреля 2002 года; патент США №6569642, выданный 27 мая 2003 года, патент США №6072047, выданный 6 июня 2000 года, патент США №6642358, выданный 4 ноября 2003 года). Как описано выше, другие рецепторы для Apo-2L включают в себя DcR1, DcR2 и OPG (см. Sheridan et al., выше; Marsters et al., выше, и Simonet et al., выше). Когда применяют в настоящем документе, термин "рецептор Apo-2L" относится к природной последовательности рецептора и вариантам рецептора. Эти термины относятся к рецептору Apo-2L, экспрессируемому у ряда млекопитающих, включая людей. Рецептор Apo-2L может быть эндогенно экспрессируемым, как происходит в природе в ряде тканевых линий человека, или может экспрессироваться рекомбинантным или синтетическим способами. "Рецептор Apo-2L с природной последовательностью" содержит полипептид с такой же аминокислотной последовательностью, что и рецептор Apo-2L, полученный в природных условиях. Таким образом, рецептор Apo-2L с природной последовательностью может обладать аминокислотной последовательностью встречающегося в природе рецептора Apo-2L любого млекопитающего, включая людей. Такой рецептор Apo-2L с природной последовательностью можно выделять в природных условиях или можно получать рекомбинантными или синтетическими способами. Термин "рецептор Apo-2L с природной последовательностью", в частности, относится к встречающимся в природе укороченным или секретируемым формам рецептора (например, растворимая форма, содержащая, например, последовательность внеклеточного домена), встречающимся в природе вариантным формам (например, формы альтернативного сплайсинга) и встречающимся в природе аллельным вариантам. Варианты рецептора могут включать в себя фрагменты и делеционные мутанты рецептора Apo-2L с природной последовательностью. На фиг.3A-3C показана последовательность DR5 человека из 411 аминокислот, как опубликовано в WO 98/51793 19 ноября 1998 года. В данной области известен вариант сплайсинга продукта транскрипции DR5 человека. Этот вариант сплайсинга DR5 кодирует последовательность DR5 человека из 440 аминокислот, как показано на фиг.4A-4C, вместе с ее нуклеотидной последовательностью (SEQ ID NO: 7 и 8), и как опубликовано в WO 98/35986 20 августа 1998 года.
"Антитело к рецептору смерти" применяют в настоящем документе в основном для обозначения антитела или антител к рецептору, входящему в суперсемейство рецептора фактора некроза опухоли и содержащему домен смерти, способный к передаче сигнала апоптоза, и такие антитела включают в себя антитело к DR5 и антитело к DR4.
"Антитело к рецептору DR5", "антитело к DR5" или "антитело против DR5" используют в широком смысле для обозначения антител, связывающихся, по меньшей мере, с одной формой рецептора DR5 или его внеклеточного домена. Необязательно антитело к DR5 слито или связано с гетерологичной последовательностью или молекулой. Предпочтительно гетерологичная последовательность позволяет антителу формировать комплексы более высокого порядка или олигомерные комплексы или способствует этому. Необязательно антитело к DR5 связывается с рецептором DR5, но не связывается или не вступает в перекрестную реакцию с любым дополнительным рецептором Apo-2L (например, DR4, DcR1 или DcR2). Необязательно антитело представляет собой агонист сигнальной активности DR5. Термин "антитело против DR5" и его грамматические эквиваленты, в частности, относится к антителам, описанным в примерах, и включает в себя в качестве неограничивающих примеров антитела "Apomab", перечисленные в таблицах 11 и 12, например, такие как Apomab 1.1, 2.1, 3.1, 4.1, 5.1, 6.1, 7.1, 8.1, 9.1, 1.2, 2.2, 3.2, 4.2, 5.2, 6.2, 7.2, 8.2, 9.2, 1.3, 2.2, 3.3, 4.3, 5.3, 6.3, 7.3, 8.3 и 9.3, предпочтительно Apomab 7.3.
Необязательно антитело к DR5 по изобретению связывается с рецептором DR5 в диапазоне концентраций от приблизительно 0,1 нМ до приблизительно 20 мМ, как измерено в анализе связывания BIAcore. Необязательно антитела к DR5 по изобретению демонстрируют значение Ic50 от приблизительно 0,6 нМ до приблизительно 18 мМ, как измерено в анализе связывания BIAcore.
"Антитело к рецептору DR4", "антитело к DR4" или "антитело против DR4" используют в широком смысле для обозначения антител, связывающихся, по меньшей мере, с одной формой рецептора DR4 или его внеклеточного домена. Необязательно антитело к DR4 слито или связано с гетерологичной последовательностью или молекулой. Предпочтительно гетерологичная последовательность позволяет антителу формировать комплексы более высокого порядка или олигомерные комплексы или способствует этому. Необязательно антитело к DR4 связывается с рецептором DR4, но не связывается или не вступает в перекрестную реакцию с любым дополнительным рецептором Apo-2L (например, DR5, DcR1 или DcR2). Необязательно антитело представляет собой агонист сигнальной активности DR4.
Необязательно антитело к DR4 по изобретению связывается с рецептором DR4 в диапазоне концентраций от приблизительно 0,1 нМ до приблизительно 20 мМ, как измерено в анализе связывания BIAcore. Необязательно антитела к DR4 по изобретению демонстрируют значение Ic 50 от приблизительно 0,6 нМ до приблизительно 18 мМ, как измерено в анализе связывания BIAcore.
Термин "агонист" используют в самом широком смысле, и он включает в себя любую молекулу, которая частично или полностью усиливает, стимулирует или активирует один или несколько видов биологической активности Apo2L/TRAIL, DR4 или DR5 in vitro, in situ или in vivo. Примеры таких видов биологической активности связывания Apo2L/TRAIL с DR4 или DR5 включают в себя апоптоз, а также виды биологической активности, описанные в литературе. Агонист может действовать прямым или непрямым образом. Например, агонист может действовать, частично или полностью усиливая, стимулируя или активируя один или несколько видов биологической активности DR4 или DR5 in vitro, in situ или in vivo, в результате его прямого связывания с DR4 или DR5, что вызывает активацию рецептора и передачу сигнала. Агонист также может действовать непрямым образом, частично или полностью усиливая, стимулируя или активируя один или несколько видов биологической активности DR4 или DR5 in vitro, in situ или in vivo, например, в результате стимуляции другой эффекторной молекулы, которая затем вызывает активацию DR4 или DR5 или передачу сигнала. Полагают, что агонист может действовать как энхансерная молекула, которая действует непрямым образом, усиливая или повышая активацию или активность DR4 или DR5. Например, агонист может усиливать активность эндогенного Apo-2L у млекопитающего. Это можно осуществлять, например, посредством предварительного образования комплексов DR4 или DR5 или посредством стабилизации комплексов соответствующего лиганда с рецептором DR4 или DR5 (такой как стабилизация природного комплекса, формируемого между Apo-2L и DR4 или DR5).
Термин "внеклеточный домен" или "ECD" относится к форме лиганда или рецептора, в которой по существу отсутствуют трансмембранный и цитоплазматический домены. Как правило, растворимый ECD содержит менее 1% таких трансмембранных и цитоплазматических доменов, а предпочтительно содержит менее 0,5% таких доменов.
Когда применяют в настоящем документе, термин "маркированный эпитопом" относится к химерному полипептиду, содержащему белок, такой как лиганд Apo-2 или рецептор DR5, или его часть или антитело, которое связывается с таким лигандом или рецептором, слитый с "маркерным полипептидом". Маркерный полипептид содержит достаточное количество остатков для образования эпитопа, к которому можно получать антитело, а также достаточно короток, так что он не препятствует активности лиганда или рецептора. Предпочтительно маркерный полипептид также в достаточной степени уникален, так что антитело по существу не вступает в перекрестные реакции с другими эпитопами. Подходящие маркерные полипептиды, как правило, состоят, по меньшей мере, из шести аминокислотных остатков, а обычно приблизительно из аминокислотных остатков в количестве от 8 до 50 (предпочтительно, приблизительно от 10 до 20 остатков).
"Выделенный", когда применяют для описания различных описанных в настоящем документе белков, означает белок, который идентифицирован и отделен от компонентов его природного окружения и/или выделен из них. Загрязняющими компонентами его природного окружения являются вещества, которые обычно могут мешать диагностическим или терапевтическим применениям белка, и они могут включать в себя ферменты, гормоны и другие белковые и небелковые растворы. В предпочтительных вариантах осуществления белок очищают (1) до степени, достаточной для получения, по меньшей мере, 15 остатков N-концевой или внутренней аминокислотной последовательности при применении секвенатора с вращающимся стаканом, или (2) до гомогенности при SDS-PAGE в невосстанавливающих или восстанавливающих условиях с применением красителя кумасси синего или, предпочтительно, серебра. Выделенный белок включает в себя белок в рекомбинантных клетках in situ, так как отсутствует, по меньшей мере, один компонент природного окружения лиганда Apo-2. Однако, как правило, выделенный белок получают посредством, по меньшей мере, одной стадии очистки.
"Процент (%) идентичности аминокислотной последовательности" по отношению к последовательностям, идентифицированным в настоящем документе, определен как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в сравниваемой последовательности лиганда, рецептора или антитела после выравнивания последовательностей и внесения, если необходимо, пропусков, для достижения максимального процента идентичности последовательностей, и не рассматривая никакие консервативные замены как часть идентичности последовательностей. Выравнивание с целью определения процента идентичности аминокислотных последовательностей можно получать различными способами, известными специалистам в данной области, которые могут определить подходящие параметры для оценки выравнивания, включающие в себя определенные алгоритмы, необходимые для достижения максимального выравнивания между сравниваемыми полноразмерными последовательностями. Для целей по настоящему документу значения процента идентичности аминокислот можно получать с применением компьютерной программы сравнения последовательностей ALIGN-2, созданной в Genentech, Inc. исходный код которой подан вместе с документацией для пользователя в U.S. Copyright Office, Washington D.C., 20559, зарегистрированный под U.S. Copyright Registration No. TXU510087. Программа ALIGN-2 общедоступна в Genentech, Inc., South San Francisco, California. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и не изменяются. Затем процент идентичности аминокислотных последовательностей подсчитывают относительно более длинной последовательности. Таким образом, даже если более короткая последовательность полностью содержится в более длинной последовательности, идентичность последовательностей будет составлять менее 100%.
Термин "контрольные последовательности" относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Контрольные последовательности, подходящие для прокариот, например, включают в себя промотор, необязательно, операторную последовательность и участок связывания рибосомы. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота "функционально связана", когда она находится в функциональной связи с другой последовательностью нуклеиновой кислоты. Например, ДНК предпоследовательности или секреторной лидирующей последовательности функционально связана с ДНК для полипептида, если он экспрессируется как пребелок, участвующий в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он воздействует на транскрипцию последовательности; или участок связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы облегчать трансляцию. Как правило, "функционально связанный" означает, что связанные последовательности ДНК являются смежными, а в случае секреторной лидирующей последовательности являются смежными и находятся в одной рамке считывания. Однако энхансеры не являются смежными. Связывание осуществляют лигированием по подходящим участкам рестрикции. Если такие участки не существуют, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с традиционной практикой.
Когда применяют в настоящем документе, термин "полиол" в широком смысле относится к многоатомным спиртовым соединениям. Полиолы могут представлять собой, например, любой водорастворимый поли(алкиленоксид)ный полимер и могут обладать линейной или разветвленной цепью. Предпочтительные полиолы включают в себя полиолы, замещенные в одном или нескольких гидроксильных положениях химическими группами, такими как алкильная группа с количеством атомов углерода от одного до четырех. Как правило, полиол представляет собой поли(алкиленгликоль), предпочтительно поли(этиленгликоль) (PEG). Однако специалистам в данной области понятно, что при использовании способов для конъюгации, описанных в настоящем документе для PEG, можно применять другие полиолы, например, такие как поли(пропиленгликоль) и сополимеры полиэтилен-полипропиленгликоля. Полиолы включают в себя хорошо известные в данной области и общедоступные полиолы, такие как полиолы из коммерчески доступных источников, таких как Nektar® Corporation.
Термин "конъюгат" в настоящем документы применяют в его наиболее широком определении для обозначения соединенного или связанного вместе. Молекулы "конъюгированы", когда они действуют или функционируют, как если они соединены.
"Строгость" реакций гибридизации легко определяется специалистом в данной области, и, как правило, она представляет собой эмпирический расчет, зависящий от длины зонда, температуры отмывки и концентрации солей. Как правило, для более длинных зондов для правильного отжига необходимы более высокие температуры, тогда как для коротких зондов необходимы более низкие температуры. Как правило, гибридизация зависит от способности денатурированной ДНК к повторному отжигу, когда комплементарные цепи находятся в среде с температурой ниже температуры плавления. Чем выше степень желаемой гомологии между зондом и гибридизуемой последовательностью, тем выше относительная температура, которую можно использовать. В результате, отсюда следует, что более высокие относительные температуры приводят к формированию более строгих условий реакции, тогда как более низкие температуры уменьшают строгость условий реакции. Для дополнительных подробностей и объяснения строгости реакций гибридизации см. Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers (1995).
Как определено в настоящем документе, "строгие условия" или "условия с высокой строгостью" определяют как условия, в которых: (1) применяют низкую ионную силу и высокую температуру для отмывки, например 0,015 M хлорид натрия/0,0015 M цитрат натрия/0,1% додецилсульфат натрия при 50°C; (2) в течение гибридизации применяют денатурирующее средство, такое как формамид, например 50% (об./об.) формамид с 0,1% бычьим сывороточным альбумином/0,1% фиколлом/0,1% поливинилпирролидоном/50 мМ натрий-фосфатным буфером при pH 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°C; или (3) применяют 50% формамид, 5×SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5× раствор Денхардта, разрушенную ультразвуком ДНК спермы лосося (50 мкг/мл), 0,1% SDS и 10% декстрансульфат при 42°C, с отмывкой при 42°C в 0,2×SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55°C с последующей отмывкой с высокой строгостью в растворе, состоящем из содержащей 0,1×SSC ЭДТА при 55°C.
"Умеренно строгие условия" можно определить, как описано в Sambrook et al., Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989, и они включают в себя применение отмывочного раствора и условий гибридизации (например, температура, ионная сила и % SDS), менее строгих, чем описанные выше условия. Пример умеренно строгих условий представляет собой инкубацию в течение ночи при 37°C в растворе, содержащем: 20% формамид, 5×SSC (150 мМ NaCl, 15 мМ трехосновный цитрат натрия), 50 мМ фосфат натрия (pH 7,6), 5× раствор Денхардта, 10% декстрансульфат и 20 мг/мл денатурированной расщепленной ДНК спермы лосося с последующей отмывкой фильтров в 1×SSC приблизительно при 37-50°C. Специалисты в данной области знают, как отрегулировать температуру, ионную силу и т.д., как необходимо для приведения в соответствие с такими факторами, как длина зонда и т.п.
Термины "аминокислота" и "аминокислоты" относятся ко всем встречающимся в природе L-альфа-аминокислотам. Это определение предназначено для включения норлейцина, орнитина и гомоцистеина. Аминокислоты идентифицируют посредством их однобуквенных или трехбуквенных обозначений:
На фигурах могут применяться некоторые другие однобуквенные и трехбуквенные обозначения для обозначения и идентификации двух или более аминокислот или нуклеотидов в данном положении последовательности.
Термин "антитело" применяют в самом широком смысле, и он конкретно относится к отдельным моноклональным антителам к DR5 (включающим в себя агонистические, антагонистические и нейтрализующие или блокирующие антитела) и композициям антител к DR5 с полиэпитопной специфичностью. Как применяют в настоящем документе, "антитело" включает в себя молекулы интактных иммуноглобулинов или антител, поликлональные антитела, полиспецифические антитела (т.е. биспецифические антитела, сформированные, по меньшей мере, двумя интактными антителами) и фрагменты иммуноглобулинов (такие как Fab, F(ab')2 или Fv), при условии, они проявляют желаемые агонистические или антагонистические свойства, описанные в настоящем документе.
Как правило, антитела представляют собой белки или полипептиды, проявляющие специфичность связывания с конкретным антигеном. Природные антитела, как правило, являются гетеротетрамерными гликопротеинами, состоящими из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Как правило, каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, тогда как количество дисульфидных связей у тяжелых цепей различных изотипов иммуноглобулинов варьирует. Каждая тяжелая и легкая цепь также обладает расположенными с равными интервалами дисульфидными мостиками внутри цепи. На одном из концов каждой тяжелой цепи находится вариабельный домен (VH), а за ним следует ряд константных доменов. У каждой легкой цепи на одном из концов находится вариабельный домен (VL), а на другом конце - константный домен; константный домен легкой цепи расположен параллельно первому константному домену тяжелой цепи, а вариабельный домен легкой цепи расположен параллельно вариабельному домену тяжелой цепи. Полагают, что область контакта между вариабельными доменами легкой цепи и тяжелой цепи формируют конкретные аминокислотные остатки [Chothia et al., J. Mol. Biol., 186:651-663 (1985); Novotny and Haber, Proc. Natl. Acad. Sci. USA, 82:4592-4596 (1985)]. Легкие цепи антител любого вида позвоночного на основе аминокислотных последовательностей их константных доменов можно отнести к одному из двух отчетливо различимых типов, называемых каппа и лямбда. В зависимости от аминокислотной последовательности константного домена их тяжелых цепей иммуноглобулины можно отнести к различным классам. Существует пять главных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, а некоторые из них можно дополнительно разделить на подклассы (изотипы), например IgG-1, IgG-2, IgG-3 и IgG-4; IgA-1 и IgA-2. Константные домены тяжелых цепей, соответствующие различным классам иммуноглобулинов, обозначают альфа, дельта, эпсилон, гамма и мю, соответственно.
"Фрагменты антител" содержат часть интактного антитела, как правило, антигенсвязывающую или вариабельную область интактного антитела. Примеры фрагментов антител включают в себя фрагменты Fab, Fab', F(ab')2 и Fv, димерные антитела, молекулы одноцепочечных антител и полиспецифические антитела, сформированные из фрагментов антител.
Термин "вариабельный" в настоящем документе используют для описания определенных частей вариабельных доменов, которые отличаются по последовательности у разных антител и используются для связывания каждого конкретного антитела с его конкретным антигеном и для формирования специфичности каждого конкретного антитела к его конкретному антигену. Однако вариабельность не всегда одинаково распределена в вариабельных доменах антител. Как правило, она концентрируется в трех участках вариабельных доменов легких цепей и тяжелых цепей, называемых определяющими комплементарность областями (CDR) или гипервариабельными областями. Более высококонсервативные части вариабельных доменов называют каркасом (FR). Каждый из вариабельных доменов природных тяжелых и легких цепей содержит четыре FR области, главным образом принимающие конфигурацию β-слоя, соединенные тремя гипервариабельными областями, формирующими петли, соединяющие β-слои, а в некоторых случаях, формирующие часть структуры β-слоя. CDR в каждой цепи удерживаются вместе в непосредственной близости с помощью FR областей и, совместно с CDR другой цепи, вносят вклад в формирование антигенсвязывающего участка антитела [см. Kabat, E.A. et al., Sequences of Proteins of Immunological Interest, National Institutes of Health, Bethesda, MD (1987)]. Константные домены непосредственно не участвуют в связывании антитела с антигеном, но осуществляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности.
Как применяют в настоящем документе, термин "моноклональное антитело" относится к антителу, полученному из совокупности по существу гомогенных антител, т.е. отдельные антитела, составляющие совокупность, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, являясь направленными к одному антигенному участку. Кроме того, в отличие от традиционных препаратов (поликлональных) антител, которые включают в себя различные антитела, направленные к различным детерминантам (эпитопам), каждое моноклональное антитело направлено к одной детерминанте на антигене.
Моноклональные антитела по настоящему документу включают в себя химерные, гибридные и рекомбинантные антитела, полученные посредством соединения вариабельного (включая гипервариабельный) домена антитела к DR5 с константным доменом (например, "гуманизированные" антитела), или легкой цепи с тяжелой цепью, или цепи одного вида с цепью другого вида, или посредством слияний с гетерологичными белками, вне зависимости от вида происхождения или обозначения класса или подкласса иммуноглобулина, а также фрагменты антител (например, Fab, F(ab')2 и Fv) при условии, что они проявляют желательную биологическую активность или свойства. См., например, патент США №4816567 и Mage et al., in Monoclonal Antibody Production Techniques and Applications, pp.79-97 (Marcel Defcker, Inc.: New York, 1987).
Таким образом, модификатор "моноклональное" указывает на характеристику антитела как полученного из по существу гомогенной совокупности антител, и его не следует истолковывать как требующего получение антител каким-либо конкретным способом. Например, моноклональные антитела для применения по настоящему изобретению можно получать гибридомным способом, впервые описанным в Kohler and Milstein, Nature, 256:495 (1975), или их можно получать способами рекомбинантных ДНК так, как описано в патенте США №4816567. "Моноклональные антитела" также можно выделять из фаговых библиотек, полученных с применением способов, описанных, например, в McCafferty et al., Nature, 348:552-554 (1990).
"Гуманизированные" формы не являющихся человеческими (например, мышиных) антител представляют собой специфические химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab' F(ab')2 или другие антигенсвязывающие подпоследовательности антитела), которые содержат минимальную последовательность, полученную из не являющегося человеческим иммуноглобулина. Преимущественно, гуманизированные антитела являются иммуноглобулинами человека (реципиентное антитело), в которых остатки из определяющей комплементарность области (CDR) реципиента заменены остатками из CDR не являющихся человеком видов (донорное антитело), таких как мышь, крыса или кролик, с желательной специфичностью, аффинностью и емкостью. В некоторых случаях соответствующими остатками, не принадлежащими человеку, заменены остатки каркасной области (FR) иммуноглобулина человека. Кроме того, гуманизированное антитело может содержать остатки, которых нет ни в реципиентном антителе, ни в импортируемых CDR или каркасных последовательностях. Эти модификации производят для дальнейшего улучшения и оптимизации характеристик антитела. Как правило, гуманизированное антитело содержит по существу все, по меньшей мере, из одного или, как правило, двух вариабельных доменов, в которых все или по существу все области CDR соответствуют областям CDR не принадлежащего человеку иммуноглобулина, а все или по существу все области FR представляют собой области FR консенсусной последовательности иммуноглобулина человека. Гуманизированное антитело также оптимально содержит, по меньшей мере, часть константной области или домена (Fc) иммуноглобулина, как правило, часть константной области или домена иммуноглобулина человека.
"Антитело человека" представляет собой антитело, которое обладает аминокислотной последовательностью, которая соответствует аминокислотной последовательности антитела, полученного у человека и/или полученного с применением любого из способов получения антител человека, известных в данной области, или как описано в настоящем документе. Это определение антитела человека включает в себя антитела, содержащие, по меньшей мере, один полипептид тяжелой цепи человека или, по меньшей мере, один полипептид легкой цепи человека, например антитело, содержащее полипептиды легкой цепи мыши и тяжелой цепи человека. Антитела человека можно получать с применением различных известных в данной области способов. В одном из вариантов осуществления антитело человека выбрано из фаговой библиотеки, где эта фаговая библиотека экспрессирует антитела человека (Vaughan et al. Nature Biotechnology, 14:309-314 (1996): Sheets et al. PNAS, (USA) 95:6157-6162 (1998)); Hoogenboom and Winter, J. Mol. Biol. 227:381(1991); Marks et al., J. Mol. Biol. 222:581 (1991)). Антитела человека также можно получать посредством введения локусов иммуноглобулина человека трансгенным животным, например мышам, у которых эндогенные гены иммуноглобулинов частично или полностью инактивированы. При антигенной стимуляции наблюдают продукцию антител человека, которая во всех аспектах, включая перестройку, сборку генов и репертуар антител, очень похожа на ту, которую наблюдают у людей. Этот подход описан, например, в патентах США №№5545807; 5545806; 5569825; 5625126; 5633425; 5661016 и в следующих научных публикациях: Marks et al., Bio/Technology, 10:779-783 (1992); Lonberg et al., Nature, 368:856-859 (1994); Morrison, Nature, 368:812-13 (1994); Fishwild et al., Nature Biotechnology, 14:845-51 (1996); Neuberger, Nature Biotechnology, 14:826 (1996); Lonberg and Huszar, Intern. Rev. Immunol. 13:65-93 (1995). Альтернативно антитело человека можно получать посредством иммортализации B-лимфоцитов человека, продуцирующих антитела, направленных к антигену-мишени (такие B-лимфоциты можно получать у индивидуума, или они могут быть иммунизированы in vitro). См., например, Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985); Boerner et al., J. Immunol. 147 (1):86-95 (1991), и патент США №5750373.
Термин "Fc-область" используют для определения C-концевой области тяжелой цепи иммуноглобулина, которую можно получить посредством расщепления интактного антитела папаином. Fc-область может представлять собой Fc-область с природной последовательностью или вариант Fc-области. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьировать, Fc-область тяжелой цепи IgG человека, как правило, определяют как располагающуюся от аминокислотного остатка в положении приблизительно Cys226 или приблизительно от положения Pro230 до C-конца Fc-области (применяя в настоящем документе систему нумерации по Kabat et al., выше). Fc-область иммуноглобулина, как правило, содержит два константных домена, домен CH2 и домен CH3, и необязательно содержит домен CH4.
Под "цепью Fc-области" в настоящем документе подразумевают одну из двух полипептидных цепей Fc-области.
"Домен CH2" Fc-области IgG человека (также обозначаемый как домен "Cγ2"), как правило, расположен от аминокислотного остатка в положении приблизительно 231 до аминокислотного остатка в положении приблизительно 340. Домен CH2 является уникальным в том, что он не образует тесной пары с другим доменом. Вместо этого, между двумя доменами CH2 интактной природной молекулы IgG расположены две N-связанные разветвленные углеводные цепи. Предполагают, что углевод может обеспечивать замену спариванию домен-домен и помогать стабилизации домена CH2. Burton, Molec. Immunol., 22:161-206 (1985). Домен CH2 по настоящему документу может представлять собой домен CH2 с природной последовательностью или вариант домена CH2.
"Домен CH3" содержит участок остатков в Fc-области с C-конца от домена CH2 (т.е. от аминокислотного остатка в положении приблизительно 341 до аминокислотного остатка в положении приблизительно 447 IgG). Область CH3 по настоящему документу может представлять собой домен CH3 с природной последовательностью или вариант домена CH3 (например, домен CH3 с введенным в одну его цепь "выступом" и соответствующей "полостью", введенной в другую его цепь; см. патент США №5821333). Такие вариантные домены CH3 можно использовать для получения полиспецифичных (например, биспецифичных) антител, как описано в настоящем документе.
Как правило, "шарнирная область" определена как участок приблизительно от Glu216 или приблизительно от Cys226 до приблизительно Pro230 IgG1 человека (Burton, Molec. Immunol. 22:161-206 (1985)). Шарнирные области других изотипов IgG можно выравнивать с последовательностью IgG1 посредством помещения предшествующих и последующих остатков цистеина, формирующих S-S-мостики между тяжелыми цепями в тех же положениях. Шарнирная область по настоящему документу может представлять собой шарнирную область с природной последовательностью или вариант шарнирной области. Две полипептидные цепи варианта шарнирной области, как правило, содержат, по меньшей мере, один цистеиновый остаток на полипептидную цепь так, чтобы две полипептидные цепи варианта шарнирной области могли формировать дисульфидную связь между двумя цепями. Предпочтительная шарнирная область по настоящему документу представляет собой шарнирную область с природной последовательностью человека, например шарнирную область с природной последовательностью IgG1 человека.
A "функциональная Fc-область" обладает, по меньшей мере, одной "эффекторной функцией" Fc-области с природной последовательностью. Примеры "эффекторной функции" включают в себя связывание C1q; обусловленную комплементом цитотоксичность (CDC); связывание Fc-рецептора; обусловленную антителами опосредованную клетками цитотоксичность (ADCC); фагоцитоз; снижение экспрессии рецепторов клеточной поверхности (например, B-клеточного рецептора; BCR) и т.д. Такие эффекторные функции, как правило, требуют, чтобы Fc-область была присоединена к домену связывания (например, вариабельному домену антитела), и их можно оценивать с применением известных в данной области различных анализов для оценки таких эффекторных функций антител.
"Fc-область с природной последовательностью" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, находящейся в природе. "Вариант Fc-области" содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности Fc-области с природной последовательностью благодаря, по меньшей мере, одной модификации аминокислоты. Предпочтительно, вариант Fc-области по сравнению с Fc-областью с природной последовательностью или с Fc-областью исходного полипептида несет, по меньшей мере, одну замену аминокислоты, например приблизительно от одной до приблизительно десяти замен аминокислот, а предпочтительно приблизительно от одной до приблизительно пяти замен аминокислот в Fc-области с природной последовательностью или в Fc-области исходного полипептида. Вариант Fc-области по настоящему документу предпочтительно обладает, по меньшей мере, приблизительно 80% идентичностью последовательности с Fc-областью с природной последовательностью и/или с Fc-областью исходного полипептида, а наиболее предпочтительно, по меньшей мере, приблизительно 90% идентичностью последовательности с ними, более предпочтительно, по меньшей мере, приблизительно 95% идентичностью последовательности с ними.
"Обусловленная антителами опосредованная клетками цитотоксичность" и "ADCC" относятся к опосредованной клетками реакции, в которой неспецифические цитотоксические клетки, экспрессирующие Fc-рецепторы (FcR) (например, натуральные киллерные (NK) клетки, нейтрофилы и макрофаги), распознают связанное на клетке-мишени антитело и впоследствии вызывают лизис клетки-мишени. Первичные клетки для осуществления ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках подытожена в таблице 3 на странице 464 Ravetch and Kinet, Annu. Rev. Immunol., 9:457-92 (1991). Для оценки активности ADCC представляющей интерес молекулы можно провести анализ ADCC in vitro, такой как анализ ADCC, описанный в патентах США №№5500362 или 5821337. Подходящие для таких анализов эффекторные клетки включают в себя мононуклеарные клетки периферической крови (PBMC) и натуральные киллерные (NK) клетки. Альтернативно или дополнительно активность ADCC представляющей интерес молекулы можно оценивать in vivo, например, в модели на животных, такой как модель на животных, описанная в Clynes et al. PNAS (USA), 95:652-656 (1998).
"Эффекторные клетки человека" представляют собой лейкоциты, экспрессирующие один или несколько FcR и выполняющие эффекторные функции. Предпочтительно клетки экспрессируют, по меньшей мере, FcγRIII и выполняют эффекторную функцию ADCC. Примеры лейкоцитов человека, опосредующих ADCC, включают в себя мононуклеарные клетки периферической крови (PBMC), натуральные киллерные (NK) клетки, моноциты, цитотоксические T-клетки и нейтрофилы; где предпочтительны клетки PBMC и NK. Эффекторные клетки можно выделять из их природного источника, например из крови или PBMC, как описано в настоящем документе.
Термины "Fc-рецептор" и "FcR" используют для описания рецептора, связывающегося с Fc-областью антитела. Предпочтительный FcR представляет собой природную последовательность FcR человека. Кроме того, предпочтительным FcR является FcR, который связывает антитело IgG (рецептор гамма) и включает в себя рецепторы подклассов FcγRI, FcγRII и FcγRIII, включающие в себя аллельные варианты и формы альтернативного сплайсинга этих рецепторов. Рецепторы FcγRII включают в себя FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые обладают сходными аминокислотными последовательностями, которые отличаются в основном своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит в своем цитоплазматическом домене активирующий мотив иммунорецептора на основе тирозина (ITAM). Ингибирующий рецептор FcγRIIB содержит в своем цитоплазматическом домене ингибирующий мотив иммунорецептора на основе тирозина (ITIM) (рассмотрено в Daëron, Annu. Rev. Immunol. 15:203-234 (1997)). FcR рассмотрены в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-492 (1991); Capel et al., Immunomethods 4:25-34 (1994), и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). В настоящем документе термин "FcR" относится к другим FcR, включающим в себя FcR, которые будут определены в будущем. Термин также относится к неонатальному рецептору, FcRn, ответственному за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117:587 (1976), и Kim et al., J. Immunol. 24:249 (1994)).
"Обусловленная комплементом цитотоксичность" и "CDC" относятся к лизису мишени в присутствии комплемента. Активация пути комплемента начинается при связывании первого компонента системы комплемента (C1q) с молекулой (например, антителом), образующей комплекс с распознаваемым ею антигеном. Для оценки активации комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996).
"Аффинно зрелое" антитело представляет собой антитело с одним или несколькими изменениями в его одной или нескольких CDR, приводящих к увеличению аффинности антитела к антигену по сравнению с исходным антителом, в котором нет этого (этих) изменения(ий). Предпочтительные аффинно зрелые антитела обладают наномолярными или даже пикомолярными аффинностями к антигену-мишени. Аффинно зрелые антитела получают известными в данной области способами. В Marks et al. Bio/Technology 10:779-783 (1992) описано созревание аффинности посредством перестановки доменов VH и VL. Случайный мутагенез CDR и/или каркасных остатков описан в: Barbas et al. Proc Nat. Acad. Sci, USA 91:3809-3813 (1994); Schier et al. Gene 169:147-155 (1995); Yelton et al. J. Immunol., 155:1994-2004 (1995); Jackson et al., J. Immunol., 154(7):3310-9 (1995), и Hawkins et al., J. Mol. Biol., 226:889-896 (1992).
Термин "иммуноспецифичный", как используют, например, в "иммуноспецифичном связывании антител" относится к специфичному для антигена связывающему взаимодействию, которое происходит между антигенсвязывающим участком антитела и конкретным антигеном, распознаваемым антителом.
"Биологически активный" и "желательная биологическая активность" для целей настоящего документа означает обладание способностью регулировать активность DR5 или активацию DR5, включая в качестве примера апоптоз (или агонистическим или стимулирующим образом или антагонистическим или блокирующим образом), по меньшей мере, в одном типе клеток млекопитающего in vivo или ex vivo, связывание с лигандом Apo-2 (TRAIL) или регуляцию активации одной или нескольких молекул во внутриклеточном пути передачи сигнала, таких как каспаза 3, каспаза 8, каспаза 10 или FADD. В данной области известны анализы для определения активации таких внутриклеточных молекул, например, см. Boldin et al., J. Biol. Chem., 270:7795-7798 (1995); Peter, Cell Death Differ., 7:759-760 (2000); Nagata, Cell, 88:355-365 (1998); Ashkenazi et al., Science, 281:1305-1308 (1999).
Термины "агонист" и "агонистический", когда применяют в настоящем документе, обозначают или описывают молекулу, способную, прямо или опосредованно, в значительной степени индуцировать, стимулировать или усиливать биологическую активность или активацию DR5. Необязательно, "агонистическое антитело к DR5" представляет собой антитело с активностью, сравнимой с лигандом DR5, известным как лиганд Apo-2 (TRAIL), или способное к активации рецептора DR5, что приводит к активации одного или нескольких внутриклеточных путей передачи сигнала, что может включать в себя активацию каспазы 3, каспазы 8, каспазы 10 или FADD.
Термины "антагонист" и "антагонистический", когда применяют в настоящем документе, обозначают или описывают молекулу, способную, прямо или опосредованно, в значительной степени нейтрализовать, снижать или ингибировать биологическую активность DR5 или активацию DR5. Необязательно антагонист представляет собой молекулу, нейтрализующую биологическую активность, являющуюся результатом активации DR5 или формирования комплекса между DR5 и ее лигандом, таким как лиганд Apo-2.
Термины "апоптоз" и "апоптотическая активность" применяются в широком смысле и обозначают запланированную или контролируемую форму клеточной гибели у млекопитающих, которая, как правило, сопровождается одним или несколькими характерными изменениями в клетке, включая конденсацию цитоплазмы, потерю микроворсинок плазматической мембраной, сегментацию ядра, деградацию хромосомной ДНК или снижение функционирования митохондрий. Эту активность можно определять и измерять, например, посредством анализов жизнеспособности клетки, анализов связывания аннексина V, анализов PARP, анализа FACS или электрофореза ДНК, где все они известны в данной области. Необязательно апоптотическую активность определяют посредством аннексина V или анализа PARP.
Термины "злокачественная опухоль", "раковый" и "злокачественный" обозначают или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется нерегулируемым ростом клеток. Примеры злокачественной опухоли включают в себя в качестве неограничивающих примеров, карциному, включая аденокарциному, лимфому, бластому, меланому, глиому, саркому, миелому (такую как множественная миелома) и лейкоз. Более конкретные примеры таких злокачественных опухолей включают в себя плоскоклеточный рак, мелкоклеточный рак легких, немелкоклеточный рак легких, аденокарциному легких и сквамозную карциному легких, желудочно-кишечный рак, ходжкинскую и неходжкинскую лимфомы, рак поджелудочной железы, глиобластому, рак шейки матки, глиому, рак яичников, злокачественную опухоль печени, такую как печеночная карцинома и гепатома, рак мочевого пузыря, рак молочной железы, рак толстой кишки, колоректальный рак, карциному эндометрия или матки, карциному слюнных желез, рак почки, такой как почечноклеточный рак и опухоли Вильмса, базально-клеточный рак, меланому, рак предстательной железы, рак женских наружных половых органов, рак щитовидной железы, рак яичка, рак пищевода и различные типы злокачественных опухолей головы и шеи.
Термин "связанное с иммунной системой заболевание" означает заболевание, в котором какой-либо компонент иммунной системы млекопитающего вызывает, опосредует или другим образом вносит вклад в заболеваемость млекопитающего. Также включены заболевания, где стимуляция иммунного ответа или вмешательство в него оказывает благоприятный эффект на развитие заболевания. В этот термин включены аутоиммунные заболевания, опосредованные иммунной системой воспалительные заболевания, не опосредуемые иммунной системой воспалительные заболевания, инфекционные заболевания и иммунодефициты. Примеры связанных с иммунной системой заболеваний и воспалительных заболеваний, некоторые из которых являются опосредованными иммунными или T-клетками, которые можно подвергать лечению по изобретению, включают в себя системную красную волчанку, ревматоидный артрит, ювенильный ревматоидный артрит, спондилоартропатию, системный склероз (склеродермию), идиопатические воспалительные миопатии (дерматомиозит, полимиозит), синдром Шегрена, системный васкулит, саркоидоз, аутоиммунную гемолитическую анемию (иммунную панцитопению, пароксизмальную ночную гемоглобинурию), аутоиммунную тромбоцитопению (идиопатическую тромбоцитопеническую пурпуру, опосредованную иммунной системой тромбоцитопению), тиреоидит (болезнь Грэйва, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит, атрофический тиреоидит), сахарный диабет, опосредованное иммунной системой почечное заболевание (гломерулонефрит, тубулоинтерстициальный нефрит), демиелинизирующие заболевания центральной и периферической нервной систем, такие как рассеянный склероз, идиопатическая демиелинизирующая полиневропатия или синдром Гийена-Барре, и хроническую воспалительную демиелинизирующую полиневропатию, гепатобилиарные заболевания, такие как инфекционный гепатит (гепатит A, B, C, D, E и другие негепатотропные вирусы), аутоиммунный хронический активный гепатит, первичный биллиарный цирроз, гранулематозный гепатит, склерозирующий холангит, воспалительные и фиброзные легочные заболевания, такие как воспалительное заболевание кишечника (язвенный колит, болезнь Крона), глютензависимая энтеропатия и синдром Уипла, аутоиммунные или опосредованные иммунной системой заболевания кожи, включая буллезные заболевания кожи, полиморфную эритему и контактный дерматит, псориаз, аллергические заболевания, такие как астма, аллергический ринит, атопический дерматит, гиперчувствительность к пище и крапивница, иммунологические заболевания легких, такие как эозинофильные пневмонии, идиопатический фиброз легких и гиперчувствительный пневмонит, связанные с трансплантацией заболевания, включая отторжение трансплантата и реакцию "трансплантат против хозяина". Инфекционные заболевания включают в себя СПИД (инфекция ВИЧ), гепатит A, B, C, D и E, бактериальные инфекции, грибковые инфекции, протозойные инфекции и паразитарные инфекции.
"Аутоиммунное заболевание" в настоящем документе используют в широком, общем смысле для обозначения заболеваний или состояний у млекопитающих, у которых разрушение нормальных или здоровых тканей происходит вследствие гуморального или клеточного иммунного ответа конкретного млекопитающего на его или ее собственные тканевые компоненты. Примеры включают в себя в качестве неограничивающих примеров обыкновенную волчанку, тиреоидит, ревматоидный артрит, псориаз, рассеянный склероз, аутоиммунный диабет и воспалительное заболевание кишечника (IBD).
Когда применяют в настоящем документе, "ингибирующее рост средство" относится к соединению или композиции, которые ингибируют рост клетки in vitro и/или in vivo. Таким образом, ингибирующее рост средство может представлять собой средство, которое специфически уменьшает процент клеток в S-фазе. Примеры ингибирующих рост средств включают в себя средства, блокирующие прохождение клеточного цикла (в точке, отличной от S-фазы), такие как средства, индуцирующие арест в G1-фазе и арест в M-фазе. Классические блокаторы M-фазы включают в себя алкалоиды барвинка (винкристин и винбластин), ТАКСОЛ® и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Средства, осуществляющие арест в G1, также имеют побочным результатом арест в S-фазе, например алкилирующие ДНК средства, такие как тамоксифен, преднизон, дакарбазин, мехлоретамин, цисплатин, метотрексат, 5-фторурацил и ara-C. Дополнительную информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., глава 1, озаглавленная "Cell cycle regulation, oncogenes, and antineoplastic drugs" by Murakami et al. (WB Saunders: Philadelphia, 1995), особенно стр.13.
Как применяют в данной заявке, термин "пролекарство" относится к предшественнику или производной форме фармацевтически активного вещества, которые менее цитотоксичны для злокачественных клеток по сравнению с исходным лекарственным средством и способны к ферментативной активации или конверсии в более активную исходную форму. Например, см. Wilman, "Prodrugs in Cancer Chemotherapy" Biochemical Society Transactions, l4, pp.375-382, 615th Meeting Belfast (1986), и Stella et al.," Prodrugs: A Chemical Approach to Targeted Drug Delivery," Directed Drug Delivery, Borchardt et al., (ed.), pp.247-267, Humana Press (1985). Пролекарства по данному изобретению включают в себя в качестве неограничивающих примеров содержащие фосфаты пролекарства, содержащие тиофосфаты пролекарства, содержащие сульфаты пролекарства, содержащие пептиды пролекарства, модифицированные D-аминокислотами пролекарства, гликозилированные пролекарства, содержащие бета-лактамы пролекарства, содержащие необязательно замещенные феноксиацетамиды пролекарства или содержащие необязательно замещенные фенилацетамиды пролекарства, 5-фторцитозиновые и другие 5-фторуридиновые пролекарства, которые можно конвертировать в более активное цитотоксическое свободное лекарственное средство. Примеры цитотоксических лекарственных средств, которые можно дериватизировать в пролекарственную форму для применения по настоящему изобретению, включают в себя в качестве неограничивающих примеров те химиотерапевтические средства, которые описаны ниже.
Как применяют в настоящем документе, термин "цитотоксическое средство" относится к веществу, которое ингибирует или предотвращает функционирование клеток и/или вызывает разрушение клеток. Термин предназначен для включения в него радиоактивных изотопов (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивных изотопов Lu), химиотерапевтических средств и токсинов, таких как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты.
"Химиотерапевтическое средство" представляет собой химическое соединение, пригодное для лечения состояний, таких как злокачественная опухоль. Примеры химиотерапевтических средств включают в себя алкилирующие средства, такие как тиотепа и циклофосфамид (CYTOXAN™); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включающие в себя алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогенины (особенно буллатацин и буллатацинон); камптотецин (включающий в себя синтетический аналог топотекан); бриостатин; каллистатин; CC-1065 (включающий его адолезиновые, карзелезиновые и бизелезиновые синтетические аналоги); криптофицины (особенно криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая его синтетические аналоги, KW-2189 и CBI-TMI); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлоретамин, гидрохлорид оксида мехлоретамина, мелфалан, новэмбихин, фенестерин, преднимустин, трофосфамид, урамустин; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин, ранимустин; антибиотики, такие как ендииновые антибиотики (например, калихимицин, особенно калихимицин γ1 I и калихимицин ωI I (см., например, Agnew, Chem Intl. Ed. Engl., 33:183-186 (1994)); динемицин, включая динемицин A; эсперамицин; а также хромофор неокарциностатин и родственные хромопротеиновые ендииновые антибиотические хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (включая морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, циностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин, 5-FU; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; антиадренергические средства, такие как аминоглутетимид, митотан, трилостан; компенсаторы фолиевой кислоты, такие как фролиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновая кислота; амсакрин; бестрабуцил; бизантрен; эдатраксат; дефофамин; демеколцин; диазиквон; эльфорнитин; ацетат эллиптиния; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидамин; майтанзиноиды, такие как майтанзин и анзамитоцины; митогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамет; пирарубицин; подофилиновая кислота; 2-этилгидразид; прокарбазин; PSK®; разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновая кислота; триазиквон; 2,2',2''-трихлортриэтиламин; трихотецены (особенно токсин T-2, верракурин A, роридин A и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-C"); циклофосфамид; тиотепа; таксоиды, например паклитаксел (TAXOL®, Bristol-Myers Squibb Oncology, Princeton, NJ) и доцетаксел (TAXOTERE®, Rhône-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митомицин C; митоксантрон; винкристин; винорелбин; навелбин; новантрон; тенипозид; дауномицин; аминоптерин; кселода; ибандронат; CPT-11; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноевая кислота; капецитабин и фармацевтически приемлемые соли, кислоты или производные любого из указанного выше. Также в это определение включены антигормональные средства, регулирующие или ингибирующие действие гормонов на опухоли, такие как антиэстрогенные средства, например, включающие в себя тамоксифен, ралоксифен, ингибирующие ароматазу 4(5)-имидазолы, 4-гидрокситамоксифен, триоксифен, коксифен, LY117018, онапристон и торемифен (фарестон); и антиандрогенные средства, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; и фармацевтически приемлемые соли, кислоты или производные любого из указанного выше.
Термин "цитокин" представляет собой общий термин для белков, высвобождаемых одной группой клеток и действующих на другие клетки в качестве межклеточных медиаторов. Примерами таких цитокинов являются лимфокины, монокины и обычные полипептидные гормоны. Цитокины включают в себя гормон роста, такой как гормон роста человека, N-метионил гормон роста человека и бычий гормон роста; паратиреоидный гормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (FSH), тиреотропный гормон (TSH) и лютеинизирующий гормон (LH); фактор роста печени; фактор роста фибробластов; пролактин; плацентарный лактоген; факторы некроза опухоли-альфа и -бета; мюллерова ингибирующая субстанция; ассоциированный с гонадотропином пептид мыши; ингибин; активин; фактор роста эндотелия сосудов; интегрин; тромбопоетин (TPO); факторы роста нервов, такие как NGF-альфа; фактор роста тромбоцитов; трансформирующие факторы роста (TGF), такие как TGF-альфа и TGF-бета; инсулиноподобный фактор роста-I и -II; эритропоэтин (EPO); остеоиндуктивные факторы; интерфероны, такие как интерферон-альфа, -бета и -гамма; колониестимулирующие факторы (CSF), такие как макрофагальный CSF (M-CSF); гранулоцитарно-макрофагальный CSF (GM-CSF) и гранулоцитарный CSF (G-CSF); интерлейкины (IL), такие как IL-1, IL-1альфа, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12; фактор некроза опухоли, такой как TNF-альфа или TNF-бета, и другие полипептидные факторы, включая LIF и лиганд kit (KL). Как применяют в настоящем документе, термин цитокин включает в себя белки из природных источников или из культуры рекомбинантных клеток и биологически активные эквиваленты цитокинов с природной последовательностью.
Как применяют в настоящем документе, термины "лечение" и "терапия" относятся к лечебной терапии, профилактической терапии и превентивной терапии.
Термин "терапевтически эффективное количество" относится к количеству лекарственного средства, эффективному для лечения заболевания или нарушения у млекопитающего. В случае злокачественной опухоли терапевтически эффективное количество лекарственного средства может уменьшать количество злокачественных клеток; уменьшать размер опухоли; ингибировать (т.е. до некоторой степени замедлять, а предпочтительно останавливать) инфильтрацию злокачественных клеток в периферические органы; ингибировать (т.е. до некоторой степени замедлять, а предпочтительно останавливать) метастазирование опухоли; до некоторой степени ингибировать рост опухоли и/или до некоторой степени облегчать один или несколько симптомов, ассоциированных с нарушением. По той степени, в которой лекарственное средство может предотвращать рост существующих злокачественных клеток и/или уничтожать их, оно может быть цитостатическим и/или цитотоксическим. Для терапии злокачественных опухолей эффективность in vivo можно измерять, например, посредством оценки массы или объема опухоли, времени до прогрессирования заболевания (TTP) и/или определения доли ответивших (RR).
Как применяют в настоящем документе, термин "млекопитающее" относится к любому млекопитающему, классифицируемому как млекопитающее, включая людей, высших приматов, коров, лошадей, собак и кошек. В предпочтительном варианте осуществления изобретения млекопитающее представляет собой человека.
Термин "объективный ответ" определяют как полный или частичный ответ субъекта на лечение, как определяют, применяя критерий оценки ответа в солидных опухолях (Response Evaluation Criteria in Solid Tumors) (RECIST) (J. Nat. Cancer Inst. 92(3):205-216 (2000)).
Термин "длительность ответа" в настоящем документе используют для указания времени от начального полного или частичного ответа до момента прогрессирования заболевания или смерти.
Термин "продолжительность жизни без прогрессирования" в настоящем документе используют для указания времени от первых суток лечения до прогрессирования заболевания или гибели, смотря по тому, что произойдет сначала.
Термин "полная регрессия (CR)" используют для указания того, что объем опухоли в трех последовательных измерениях составляет ≤13,5 мм3.
Термин "частичная регрессия (PR)" указывает на то, что объем опухоли в трех последовательных измерениях составляет ≤50% от ее объема в первые сутки и ≥13,5 мм3 в одном или нескольких из этих измерений.
II. Композиции и способы по изобретению
A. Антитела к DR5
В одном из вариантов осуществления изобретения предоставлены антитела к DR5. Примеры антител включают в себя поликлональные, моноклональные, гуманизированные, биспецифические и гетероконъюгированные антитела. Эти антитела могут представлять собой агонистические, антагонистические или блокирующие антитела.
1. Поликлональные антитела
Антитела по изобретению могут включать в себя поликлональные антитела. Способы получения поликлональных антител известны специалистам в данной области. Поликлональные антитела можно индуцировать у млекопитающего, например, посредством одной или нескольких инъекций иммунизирующего вещества и, при желании, адъюванта. Как правило, иммунизирующее вещество и/или адъювант инъецируют животному посредством нескольких подкожных или интраперитонеальных инъекций. Иммунизирующее вещество может содержать полипептид DR5 (или ECD DR5) или его слитный белок. Может быть эффективным конъюгировать иммунизирующее вещество с белком, для которого известно, что он иммуногенен у иммунизируемого животного. Примеры таких иммуногенных белков включают в себя в качестве неограничивающих примеров гемоцианин морского блюдца, сывороточный альбумин, бычий тиреоглобулин и ингибитор трипсина сои. Примеры адъювантов, которые можно применять, включают в себя полный адъювант Фрейнда и адъювант MPL-TDM (монофосфорил липид A, синтетический дикориномиколат трегалозы). Специалист в данной области может выбрать протокол иммунизации без излишнего экспериментирования. Затем у млекопитающего можно взять кровь и проанализировать сыворотку на титр антитела к DR5. При желании млекопитающего можно повторно стимулировать до тех пор, пока титр антитела увеличивается или находится на плато.
2. Моноклональные антитела
Антитела по изобретению альтернативно могут представлять собой моноклональные антитела. Моноклональные антитела можно получить с применением гибридомных способов, таких как описанные в Kohler and Milstein, Nature, 256:495 (1975). В гибридомном способе мышь, хомяка или другого подходящего животного-хозяина, как правило, иммунизируют иммунизирующим веществом для стимуляции лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфически связываться с иммунизирующим веществом. Альтернативно лимфоциты можно иммунизировать in vitro.
Иммунизирующее вещество, как правило, включает в себя полипептид DR5 (или ECD DR5) или его слитный белок, такой как слитный белок ECD DR5-IgG. Иммунизирующее вещество альтернативно может содержать фрагмент или часть DR5 с одной или несколькими аминокислотами, принимающими участие в связывании Apo-2L с DR5. В предпочтительном варианте осуществления иммунизирующее вещество содержит последовательность внеклеточного домена DR5, слитую с последовательностью IgG.
Как правило, используют или лимфоциты периферической крови ("PBL"), если желательны клетки человеческого происхождения, или клетки селезенки или клетки лимфоузлов, если желательны источники из не относящихся к человеку млекопитающих. Затем лимфоциты сливают с иммортализованной клеточной линией с использованием подходящего вызывающего слияние клеток вещества, такого как полиэтиленгликоль, с получением гибридомной клетки [Goding, Monoclonal antibodies: Principles and Practice, Academic Press, (1986) pp.59-103]. Иммортализованные клеточные линии, как правило, представляют собой трансформированные клетки млекопитающих, в частности миеломные клетки, происходящие от грызунов, коров и человека. Как правило, применяют миеломные клеточные линии крыс или мышей. Гибридомные клетки можно культивировать в подходящей среде для культивирования, предпочтительно содержащей одно или несколько веществ, ингибирующих рост или выживание неслитых иммортализованных клеток. Например, если в родительских клетках отсутствует фермент гипоксантин-гуанин-фосфорибозилтрансфераза (HGPRT или HPRT), среда для культивирования гибридомы, как правило, будет содержать гипоксантин, аминоптерин и тимидин ("среда HAT"), которые предотвращают рост дефектных по HGPRT клеток.
Предпочтительными иммортализованными линиями клеток являются клетки, которые эффективно сливаются, поддерживают высокий уровень экспрессии антитела выбранными продуцирующими антитела клетками и являются чувствительными к такой среде, как среда HAT. Более предпочтительными иммортализованными линиями клеток являются линии миеломы мышей, которые можно получить, например, из Salk Institute Cell Distribution Center, San Diego, California и American Type Culture Collection, Manassas, Virginia. Примером такой мышиной линии миеломных клеток является P3X63Ag8U.1 (ATCC CRL 1580). Описано также получение моноклональных антител в линиях клеток миеломы человека и гетеромиеломы мыши-человека [Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp.51-63].
Затем среду для культивирования, в которой культивируют гибридомные клетки, можно анализировать на присутствие моноклональных антител, направленных против DR5. Предпочтительно специфичность связывания моноклональных антител, полученных посредством гибридомных клеток, определяют посредством иммунопреципитации или анализа связывания in vitro, такого как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Такие способы и анализы известны в данной области. Например, аффинность связывания моноклонального антитела можно определять анализом Скэтчарда по Munson and Pollard, Anal. Biochem., 107:220 (1980).
После идентификации желательных гибридомных клеток клоны можно субклонировать посредством серийных разведений и выращивать традиционными способами [Goding, выше]. Подходящие для данной цели среды для культивирования включают в себя, например, среду Игла, модифицированную Дульбекко и среду RPMI-1640. Альтернативно гибридомные клетки можно выращивать у млекопитающего in vivo в виде асцитов.
Секретируемые субклонами моноклональные антитела можно выделять или очищать из среды для культивирования или асцитной жидкости традиционными способами очистки иммуноглобулинов, например, такими как, очистка на белок А-сефарозе, хроматография на гидроксилапатите, гель-электрофорез, диализ или аффинная хроматография.
Моноклональные антитела также можно получить способами рекомбинантных ДНК, такими как описаны в патенте США №4816567. ДНК, кодирующие моноклональные антитела по изобретению, можно легко выделять и секвенировать с применением традиционных способов (например, с применением олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи моноклональных антител). Гибридомные клетки по изобретению служат в качестве предпочтительного источника такой ДНК. После выделения ДНК, для обеспечения синтеза моноклональных антител в рекомбинантных клетках-хозяевах, ее можно поместить в экспрессирующие векторы, которые затем трансфицируют в клетки-хозяева, такие как клетки E.coli, клетки COS обезьяны, клетки яичников китайского хомяка (CHO) или миеломные клетки, которые иначе не продуцируют белок иммуноглобулина. ДНК также можно модифицировать, например, посредством замены кодирующими последовательностями константных доменов тяжелых и легких цепей человека гомологичных последовательностей мыши, Morrison, et al., Proc. Nat. Acad. Sci. 81, 6851 (1984), или ковалентным присоединением к кодирующей последовательности иммуноглобулина всей или части кодирующей последовательности неиммуноглобулинового полипептида. Таким образом, получают "химерные" или "гибридные" антитела, которые обладают специфичностью связывания моноклонального антитела против DR5 по настоящему изобретению.
Как правило, такими неиммуноглобулиновыми полипептидами замещают константные домены антитела по изобретению или ими замещают вариабельные домены одного антигенсвязывающего участка антитела по изобретению с получением химерного бивалентного антитела, содержащего один антигенсвязывающий участок со специфичностью к DR5 и другой антигенсвязывающий участок со специфичностью к другому антигену.
Химерные или гибридные антитела также можно получать in vitro с применением известных способов химии синтеза белков, включая способы, включающие в себя использование сшивающих средств. Например, с применением реакции дисульфидного замещения или посредством формирования тиоэфирной связи можно конструировать иммунотоксины. Примеры подходящих для этой цели реагентов включают в себя иминотиолат и метил-4-меркаптобутиримидат.
Также можно получать одноцепочечные фрагменты Fv так, как описано в Iliades et al., FEBS Letters, 409:437-441 (1997). Связывание таких одноцепочечных фрагментов с применением различных линкеров описано в Kortt et al., Protein Engineering, 10:423-433 (1997). В данной области хорошо известен ряд способов рекомбинантных получения и обработки антител. Иллюстративные примеры таких способов, которые, как правило, применяют специалисты в данной области, более подробно описаны ниже.
(i) Гуманизированные антитела
Как правило, гуманизированное антитело несет один или несколько аминокислотных остатков, введенных в него из не принадлежащего человеку источника. Эти не принадлежащие человеку аминокислотные остатки часто обозначают как "импортируемые" остатки, которые, как правило, берут из "импортируемого" вариабельного домена. Гуманизирование по существу можно проводить способом Winter с коллегами [Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoeyen et al., Science, 239:1534-1536
(1988)], посредством замены CDR грызунов или последовательностей CDR на соответствующие последовательности антитела человека.
Таким образом, такие "гуманизированные" антитела представляют собой химерные антитела, где по существу соответствующими последовательностями из не являющихся человеком видов замещено менее одного исходного вариабельного домена человека. На практике гуманизированные антитела, как правило, представляют собой антитела человека, в которых некоторые остатки CDR и, возможно, некоторые остатки FR замещены на аналогичные участки из антител грызунов.
Важно, чтобы антитела гуманизировали с сохранением высокой аффинности к антигену и других благоприятных биологических свойств. Для достижения этой цели в соответствии с предпочтительным способом гуманизированные антитела получают способом анализа исходных последовательностей и различных воображаемых гуманизированных продуктов с применением трехмерных моделей исходной и гуманизированной последовательностей. Трехмерные модели иммуноглобулинов являются общедоступными и хорошо известны специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и отображают возможные трехмерные конформационные структуры выбранных возможных последовательностей иммуноглобулинов. Исследование этих изображений позволяет проводить анализ возможной роли остатков в функционировании возможной последовательности иммуноглобулина, т.е. анализ остатков, влияющих на способность иммуноглобулина-кандидата связывать его антиген. Таким образом, можно выбрать и объединить остатки FR из консенсусной и импортируемой последовательностей так, чтобы добиться желательной характеристики антитела, такой как увеличенная аффинность к антигену(ам)-мишени. Как правило, непосредственно и наиболее значительно в воздействие на связывание антигена вовлечены остатки CDR.
(ii) Антитела человека
Моноклональные антитела человека можно получать гибридомным способом. Для получения моноклональных тел человека описаны линии клеток миеломы человека и гетеромиеломы мыши-человека (Kozbor, J. Immunol., 133:3001 (1984), и Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987)).
В настоящее время можно получать трансгенных животных (например, мышей), которые после иммунизации способны к продукции спектра антител человека в отсутствие продукции эндогенных иммуноглобулинов. Например, описано, что гомозиготная делеция гена соединительной области (JH) тяжелой цепи антитела у химерных и мутантных по зародышевой линии мышей приводит к полному ингибированию продукции эндогенных антител. Перенос набора генов иммуноглобулинов зародышевой линии человека таким мутантным по зародышевой линии мышам приводит при стимуляции антигеном к продукции антител человека. Например, см. Jakobovits et al., Proc. Natl. Acad. Sci. USA. 90:2551-255 (1993); Jakobovits et al., Nature, 362:255-258 (1993).
Mendez et al. (Nature Genetics 15: 146-156 [1997]) дополнительно улучшили этот способ и получили линию трансгенных мышей, обозначенную как "Xenomouse II", которая при стимуляции антигеном продуцировала высокоаффинные полностью человеческие антитела. Этого достигли посредством интеграции в зародышевую линию мышей с делецией в эндогенном сегменте JH, как описано выше, локусов тяжелой цепи и легкой цепи размером в миллион пар нуклеотидов. Xenomouse II несет 1020 т.п.н. локуса тяжелой цепи человека, содержащего приблизительно 66 генов VH, целые области DH и JH и три другие константные области (µ, δ и χ), а также несет 800 т.п.н. локуса κ человека, содержащего 32 гена Vκ, сегменты Jκ и гены Cκ. Полученные у этих мышей антитела во всех аспектах, включая перестановку генов, сборку и спектр, имеют высокое сходство с антителами, наблюдаемыми у человека. Антитела человека экспрессируются предпочтительнее, чем эндогенные антитела, вследствие делеции в эндогенном сегменте JH, предотвращающей перестановку генов в локусе мыши.
Альтернативно, для получения антител и фрагментов антител человека in vitro из репертуара генов вариабельных (V) доменов иммуноглобулинов от неиммунизированных доноров можно применять технологию фагового дисплея (McCafferty et al., Nature, 348, 552-553 (1990)). В соответствии с этим способом гены V-доменов антител клонируют в рамку считывания гена главного или минорного белка оболочки нитевидного бактериофага, такого как M13 или fd, и представляют в виде функциональных фрагментов антител на поверхности фаговой частицы. Так как нитевидная частица содержит одноцепочечную копию ДНК фагового генома, отбор на основе функциональных свойств антитела также приводит к отбору гена, кодирующего антитело, проявляющее эти свойства. Таким образом, фаг имитирует некоторые из свойств B-клетки. Фаговый дисплей можно проводить множеством способов; для их обзора см., например, Johnson, Kevin S. and Chiswell, David J., Current Opinion in Structural Biology 3, 564-571 (1993). Для фагового дисплея можно использовать несколько источников сегментов V-генов. Clackson et al., Nature, 352, 624-628 (1991), выделили набор различных антител к оксазолину из малой библиотеки случайно комбинированных V-генов, полученных из селезенки иммунизированных мышей. Можно сконструировать репертуар V-генов от неиммунизированных доноров-людей, а антитела к набору разнообразных антигенов (включая собственные антигены) можно выделять по существу способом, описанным у Marks et al., J. Mol. Biol. 222, 581-597 (1991), или Griffith et al., EMBO J. 12, 725-734 (1993). При естественном иммунном ответе гены антител с высокой скоростью накапливают мутации (соматическое гипермутирование). Некоторые из вносимых изменений обеспечивают большую аффинность, а B-клетки, экспонирующие поверхностный иммуноглобулин с высокой аффинностью, реплицируются и дифференцируются при последующей стимуляции антигеном более предпочтительно. Этот природный процесс можно имитировать с применением способа, известного как "перестановка цепей" (Marks et al., Bio/Technol. 10, 779-783 [1992]). В данном способе аффинность "первичного" антитела человека, полученного посредством фагового дисплея, можно улучшить посредством последовательного замещения V-областями генов тяжелых и легких цепей со спектром встречающихся в природе вариантов (репертуаров) генов V-доменов, полученных от неиммунизированных доноров. Этот способ обеспечивает получение антител и фрагментов антител с аффинностями в нМ диапазоне. Стратегия получения очень больших репертуаров фаговых антител (также известная как "материнская библиотека") описана в Waterhouse et al., Nucl. Acids Res. 21, 2265-2266 (1993). Перестановку генов также можно использовать для получения антитела человека из антитела грызуна, где антитело человека обладает сходными аффинностями и специфичностями с исходным антителом грызуна. В соответствии с этим способом, который также обозначают как "импринтинг эпитопа", ген V-домена тяжелой или легкой цепи антитела грызуна, полученный способом фагового дисплея, заменяют спектром генов V-доменов человека, получая химеры грызуна-человека. Отбор на антигене приводит к выделению вариабельного домена человека, способного к восстановлению функционального антигенсвязывающего участка, т.е. эпитоп направляет (закрепляет) выбор партнера. Когда процесс повторяют для замены оставшегося V-домена грызуна, получают антитело человека (см. патентную заявку PCT WO 93/06213, опубликованную 1 апреля 1993 года). В отличие от традиционного гуманизирования антител грызунов посредством пересадки CDR, этот способ обеспечивает полностью человеческие антитела, в которых нет остатков каркаса или CDR, происходящих от грызуна.
Как подробно обсуждается ниже, антитела по изобретению необязательно могут включать в себя мономерные антитела, димерные антитела, а также поливалентные формы антител. Специалисты в данной области могут сконструировать такие димерные или поливалентные формы известными в данной области способами и с применением антител к DR5 по настоящему документу. В данной области также хорошо известны способы получения моновалентных антител. Например, один из способов включает в себя рекомбинантную экспрессию легкой цепи и модифицированной тяжелой цепи иммуноглобулина. Тяжелая цепь, как правило, укорочена в любой точке в Fc-области так, чтобы предотвратить перекрестное связывание тяжелых цепей. Альтернативно для предотвращения перекрестного связывания соответствующие остатки цистеина замещают другим аминокислотным остатком или удаляют.
(iii) Биспецифические антитела
Биспецифические антитела представляют собой моноклональные, предпочтительно человеческие или гуманизированные, антитела со специфичностями связывания, по меньшей мере, для двух различных антигенов. В настоящем случае одна из специфичностей связывания представляет собой специфичность для рецептора DR5, другая специфичность представляет собой специфичность для другого антигена, а предпочтительно для других рецептора или субъединицы рецептора. Например, в объем настоящего изобретения входят биспецифические антитела, специфически связывающие рецептор DR5 и другой рецептор апоптоза/передачи сигнала.
Способы получения биспецифических антител известны в данной области. Как правило, рекомбинантное получение биспецифических антител основано на совместной экспрессии двух пар тяжелых цепей-легких цепей иммуноглобулинов, где две тяжелые цепи обладают различными специфичностями (Millstein and Cuello, Nature 305, 537-539 (1983)). Вследствие случайной сортировки тяжелых и легких цепей иммуноглобулинов эти гибридомы (квадромы) продуцируют потенциальную смесь из 10 различных молекул антител, из которых только одна обладает корректной биспецифической структурой. Очистка правильной молекулы, которую обычно проводят посредством стадий аффинной хроматографии, является довольно трудоемкой, а выход продуктов является низким. Сходные способы описаны в публикации заявки PCT № WO 93/08829 (опубликованной 13 мая 1993 года) и в Traunecker et al., EMBO 10, 3655-3659 (1991).
В соответствии с другим и более предпочтительным подходом вариабельные домены антител с желательными специфичностями связывания (антигенсвязывающие участки антител) сливают с последовательностями константных доменов иммуноглобулинов. Предпочтительно слияние проводят с константным доменом тяжелой цепи иммуноглобулина, содержащим, по меньшей мере, часть областей шарнира, области CH2 и CH3. Предпочтительно, чтобы, по меньшей мере, на одном из слитных белков присутствовала первая константная область тяжелой цепи (CH1), содержащая участок, необходимый для связывания легкой цепи. ДНК, кодирующие слитные белки тяжелых цепей иммуноглобулинов и, при желании, легких цепей иммуноглобулина, вставляют в отдельные экспрессирующие векторы и котрансфицируют в подходящий организм-хозяин. Это предоставляет большую гибкость для регулировки соответствующих пропорций трех полипептидных фрагментов в вариантах осуществления, когда используемые при получении неравные соотношения трех полипептидных цепей обеспечивают оптимальный выход желательного биспецифического антитела. Однако когда экспрессия, по меньшей мере, двух полипептидных цепей в равных соотношениях приводит к высоким выходам или когда соотношения существенно не влияют на выход желательного сочетания цепей, кодирующие последовательности для двух или всех трех полипептидных цепей можно вставлять в один экспрессирующий вектор. В предпочтительном варианте осуществления этого подхода биспецифические антитела состоят из гибрида тяжелой цепи иммуноглобулина с первой специфичностью связывания на одном плече и гибрида из пары тяжелой цепи-легкой цепи иммуноглобулина (обеспечивающего вторую специфичность связывания) на другом плече. Обнаружено, что эта асимметрическая структура облегчает отделение желательного биспецифического соединения от нежелательных сочетаний иммуноглобулиновых цепей, так как наличие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий способ разделения. Этот подход описан в публикации PCT № WO 94/04690, опубликованной 3 марта 1994 года.
Более подробно о получении биспецифических антител см., например, Suresh et al., Methods in Enzymology 121, 210 (1986).
(iv) Гетероконъюгированные антитела
В объем настоящего изобретения также входят гетероконъюгированные антитела. Гетероконъюгированные антитела состоят из двух ковалентно связанных антител. Такие антитела предложены, например, для направления клеток иммунной системы к нежелательным клеткам (патент США №4676980) и для лечения ВИЧ-инфекции (публикации PCT №№ WO 91/00360, WO 92/200373 и EP 03089). Гетероконъюгированные антитела можно получать с применением подходящих способов перекрестного связывания. Подходящие сшивающие средства хорошо известны в данной области и описаны в патенте США №4676980, вместе с рядом способов перекрестного связывания.
(v) Фрагменты антител
В определенных вариантах осуществления антитело к DR5 (включая мышиные, человеческие и гуманизированные антитела и варианты антител) представляет собой фрагмент антитела. Для получения фрагментов антител разработаны различные способы. Как правило, эти фрагменты получают посредством протеолитического расщепления интактных антител (см., например, Morimoto et al., J. Biochem. Biophys. Methods 24:107-117 (1992), и Brennan et al., Science 229:81 (1985)). Однако эти фрагменты в настоящее время можно получать непосредственно из рекомбинантных клеток-хозяев. Например, фрагменты Fab'-SH можно непосредственно выделять из E.coli и химически связывать с формированием фрагментов F(ab')2 (Carter et al., Bio/Technology 10:163-167 (1992)). В другом варианте осуществления F(ab')2 формируют с применением для обеспечения сборки молекулы F(ab')2 лейциновой молнии GCN4. В соответствии с другим подходом фрагменты Fv, Fab или F(ab')2 можно выделять непосредственно из культуры рекомбинантных клеток-хозяев. Практикующим специалистам очевиден ряд способов получения фрагментов антител. Например, можно осуществлять расщепление с применением папаина. Примеры расщепления папаином описаны в WO 94/29348, опубликованной 12/22/94 и патенте США №4342566.
Расщепление антител папаином, как правило, дает два идентичных антигенсвязывающих фрагмента, называемых фрагментами Fab, каждый с одним антигенсвязывающим участком и остаточным фрагментом Fc. Обработка пепсином приводит к фрагменту F(ab')2 с двумя антигенсвязывающими участками, все еще способному к перекрестному связыванию антигена.
Полученные при расщеплении антитела фрагменты Fab также содержат константные домены легкой цепи и первый константный домен (CH1) тяжелой цепи. Фрагменты Fab' отличаются от фрагментов Fab добавлением небольшого количества остатков на С-конце домена CH1 тяжелой цепи, включая один или несколько остатков цистеина из шарнирной области антитела. Fab'-SH в настоящем документе представляет собой обозначение для Fab', в котором цистеиновый остаток(ки) константных доменов несут свободную тиольную группу. Фрагменты антител F(ab')2 исходно получали как пары фрагментов Fab' с остатками цистеинов шарнира между ними. Также известны другие способы химического связывания фрагментов антител.
(vi) Варианты аминокислотной последовательности антител
Варианты аминокислотной последовательности антител к DR5 получают введением в ДНК антитела к DR5 соответствующих изменений нуклеотидов или посредством пептидного синтеза. Такие варианты включают в себя, например, делеции, и/или инсерции, и/или замены остатков в пределах аминокислотных последовательностей антител к DR5 из примеров в настоящем документе. Для получения конечной конструкции можно осуществлять любое сочетание делеции, инсерции и замены при условии, что конечная конструкция обладает желательными характеристиками. Замены аминокислоты также могут изменять посттрансляционные процессы гуманизированных или вариантных антител к DR5, такие как изменение количества и положения участков гликозилирования.
Пригодный способ идентификации определенных остатков или областей антитела к DR5, которые представляют собой предпочтительные участки для мутагенеза, называется "мутагенез сканирования аланином", как описано в Cunningham and Wells, Science, 244:1081-1085 (1989). В статье определяют остаток или группу целевых остатков (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяют нейтральной или отрицательно заряженной аминокислотой (наиболее предпочтительно аланином или полиаланином) для воздействия на взаимодействие аминокислот с антигеном DR5. Затем те положения аминокислот, которые продемонстрировали функциональную чувствительность к заменам, уточняют посредством внесения дополнительных или других изменений в участках замены или для них. Таким образом, хотя участок для внесения изменения в аминокислотной последовательности предопределен, характер мутации по существу не нуждается в предопределении. Например, для анализа характеристики мутации в данном участке, в целевом кодоне или области проводят сканирование ala или случайный мутагенез и экспрессированные варианты антител против DR5 скринируют на желательную активность.
Вставки в аминокислотную последовательность включают в себя слияния на N- и/или C-конце в диапазоне длины от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутри последовательности одного или нескольких аминокислотных остатков. Примеры концевых вставок включают в себя антитело к DR5 с N-концевым остатком метионина или антитело, слитое с эпитопным маркером. Другие инсерционные варианты молекулы антитела к DR5 включают в себя слияние с N- или C-концом антитела к DR5 фермента, или полипептида, или полиола, увеличивающих время полужизни антитела в сыворотке.
Другим типом вариантов является вариант замены аминокислот. Эти варианты несут в молекуле антитела к DR5, по меньшей мере, один аминокислотный остаток, замещенный другим остатком. Участки наибольшего интереса для замещающего мутагенеза включают в себя гипервариабельные области, но также рассматривают изменения в FR. Консервативные замены представлены в таблице 1 под заголовком "предпочтительные замены". Если такие замены приводят к изменению биологической активности, тогда можно проводить более основательные изменения, обозначенные в таблице 1 как "допустимые замены", или как дополнительно описано ниже в отношении классов аминокислот, а продукты скринировать.
Значительные модификации биологических свойств антитела осуществляют посредством выбора замен, которые значительно отличаются по своему воздействию на поддержание (a) структуры полипептидного каркаса в области замены, например, такой как конформации слоя или спирали, (b) заряда или гидрофобности молекулы в намеченном участке или (c) объема боковой цепи. Встречающиеся в природе остатки разделяют на группы на основе общих свойств боковых цепей:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr;
(3) кислые: asp, glu;
(4) основные: asn, gln, his, lys, arg;
(5) остатки, влияющие на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены влекут за собой замену представителя одного из этих классов на представителя другого класса.
Также можно замещать любой цистеиновый остаток, не вовлеченный в поддержание правильной конформации гуманизированного или вариантного антитела к DR5, как правило, серином, для увеличения устойчивости молекулы к окислению и предотвращения аберрантного перекрестного связывания. С другой стороны, в антитело можно добавлять цистеиновую связь(и) для увеличения ее стабильности (особенно когда антитело представляет собой фрагмент антитела, такой как фрагмент Fv).
Особенно предпочтительный тип заместительных вариантов включает в себя замену одного или нескольких остатков гипервариабельной области исходного антитела (например, гуманизированного или человеческого антитела). Как правило, полученный вариант(ы), отбираемый для дальнейшего преобразования, обладает улучшенными биологическими свойствами относительно исходного антитела, из которого его получают. Удобный способ получения таких заместительных вариантов включает в себя созревание аффинности с применением фагового дисплея. В кратком изложении, несколько участков гипервариабельной области (например, 6-7 участков) подвергают мутированию с получением всех возможных аминокислотных замен в каждом из участков. Полученные таким образом варианты антител в моновалентном виде экспонируют на частицах нитевидного фага в виде слитных белков с продуктом гена III M13, упакованных в каждую частицу. Затем экспонированные на фагах варианты скринируют на их биологическую активность (например, связывающую активность), как описано в настоящем документе. Для идентификации участков гипервариабельных областей, являющихся кандидатами для модификации, можно провести мутагенез сканирования аланином для идентификации остатков гипервариабельной области, значительным образом участвующих в связывании антигена. Альтернативно или дополнительно может быть полезным анализировать кристаллическую структуру комплекса антиген-антитело для идентификации точек контакта между антителом и DR5 человека. Такие контактирующие остатки и расположенные рядом остатки являются кандидатами для замены в соответствии с разработанными по настоящему документу способами. После получения таких вариантов набор вариантов подвергают скринингу, как описано в настоящем документе, и антитела с наилучшими свойствами в одном или нескольких соответствующих анализах можно выбирать для дальнейшего усовершенствования.
(vii) Варианты гликозилирования антител
Антитела гликозилируются в консервативных положениях в их константных областях (Jefferis and Lund, Chem. Immunol. 65:111-128 [1997]; Wright and Morrison, TibTECH 15:26-32 [1997]). Олигосахаридные боковые цепи иммуноглобулинов влияют на функции белков (Boyd et al., Mol. Immunol. 32:1311-1318 [1996]; Wittwe and Howard, Biochem. 29:4175-4180 [1990]) и на внутримолекулярное взаимодействие между частями гликопротеина, что может влиять на конформацию и презентируемую трехмерную поверхность гликопротеина (Jefferis and Lund, выше; Wyss and Wagner, Current Opin. Biotech. 7:409-416 [1996]). Олигосахариды также могут служить для направления данного гликопротеина к определенным молекулам на основе специфических распознающих структур. Например, сообщалось, что в агалактозилированном IgG олигосахаридный остаток "перебрасывается" из внутреннего пространства CH2 и концевые остатки N-ацетилглюкозамина становятся доступными для связывания со связывающим маннозу белком (Malhotra et al., Nature Med. 1:237-243 [1995]). Удаление гликопептидазой олигосахаридов из CAMPATH-1H (рекомбинантное гуманизированное мышиное моноклональное антитело IgG1, распознающее антиген CDw52 лимфоцитов человека), продуцируемого в клетках яичника китайского хомяка (CHO), приводило к полному подавлению опосредованного комплементом лизиса (CMCL) (Boyd et al., Mol. Immunol. 32:1311-1318 [1996]), тогда как селективное удаление остатков сиаловой кислоты с применением нейраминидазы не приводило к утрате DMCL. Также сообщалось, что гликозилирование антител влияет на обусловленную антителами клеточную цитотоксичность (ADCC). В частности, сообщалось, что клетки CHO с регулируемой тетрациклином экспрессией β-(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII), гликозилтрансферазы, катализирующей формирование разветвленных GlcNAc, обладают улучшенной активностью ADCC (Umana et al., Mature Biotech. 17:176-180 [1999]).
Варианты гликозилирования антител представляют собой варианты, в которых изменен профиль гликозилирования антител. Под изменением подразумевают делецию одной или нескольких молекул углеводов, находящихся в антителе, добавление одной или нескольких молекул углеводов к антителу, изменения состава гликозилирования (профиль гликозилирования), степени гликозилирования и т.д. Например, варианты гликозилирования можно получать, удаляя, изменяя и/или добавляя в кодирующей антитело последовательности нуклеиновой кислоты один или несколько участков гликозилирования.
Гликозилирование антител, как правило, является или N-связанным, или O-связанным. N-связанное относится к присоединению молекулы углевода к боковой цепи остатка аспарагина. Распознаваемыми последовательностями для ферментативного присоединения молекулы углевода к боковой цепи аспарагина являются трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту за исключением пролина. Таким образом, присутствие в полипептиде любой из этих трипептидных последовательностей создает потенциальный участок гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров: N-ацетилгалактозамина, галактозы или ксилозы, к гидроксиаминокислоте, чаще всего серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление участков гликозилирования к антителу удобно проводить посредством изменения аминокислотной последовательности так, чтобы она содержала одну или несколько описанных выше трипептидных последовательностей (для N-связанных участков гликозилирования). Изменение также можно проводить посредством добавления к последовательности исходного антитела одного или нескольких остатков серина или треонина или замены в последовательности исходного антитела на один или несколько остатков серина или треонина (для O-связанных участков гликозилирования).
Молекулы нуклеиновой кислоты, кодирующие варианты аминокислотной последовательности антитела к DR5, получают рядом известных в данной области способов. Эти способы включают в себя в качестве неограничивающих примеров выделение из природного источника (в случае встречающихся в природе вариантов аминокислотной последовательности) или получение олигонуклеотид-опосредованным (или сайт-специфическим) мутагенезом, мутагенезом ПЦР и кассетным мутагенезом ранее полученных вариантов или безвариантной версии антитела к DR5.
Гликозилирование (включая профиль гликозилирования) антител также можно изменять без изменения лежащей в основе нуклеотидной последовательности. Гликозилирование в основном зависит от клетки-хозяина, используемой для экспрессии антитела. Так как тип клеток, используемых для экспрессии в качестве потенциальных терапевтических средств рекомбинантных гликопротеинов, например антител, крайне редко представляет собой природную клетку, можно ожидать значительных вариаций в профиле гликозилирования антител (см., например, Hse et al., J. Biol. Chem. 272:9062-9070 [1997]). Кроме выбора клеток-хозяев, факторы, влияющие на гликозилирование при рекомбинантной продукции антител, включают в себя способ роста, состав сред, плотность культуры, оксигенацию, pH, схемы очистки и т.п. Для изменения профиля гликозилирования, достигаемого в конкретном организме-хозяине, предложены различные способы, включая вставку или сверхэкспрессию определенных ферментов, вовлеченных в продукцию олигосахаридов (патенты США №№5047335; 5510261 и 5278299). Гликозилирование или определенные типы гликозилирования можно удалить из гликопротеина ферментативно, например, с применением эндогликозидазы H (Endo H). Кроме того, рекомбинантную клетку-хозяина можно сконструировать генетически, сделав ее дефектной по процессингу определенных типов полисахаридов. Эти и подобные способы хорошо известны в данной области.
Структуру гликозилирования антител можно легко анализировать общепринятыми способами анализа углеводов, включая хроматографию лектинов, ЯМР, масс-спектрометрию, ВЭЖХ, GPC, анализ состава моносахаридов, последовательное ферментативное расщепление и HPAEC-PAD, где для разделения олигосахаридов на основе заряда применяют анионообменную хроматографию с высоким pH. Способы выделения олигосахаридов для аналитических целей также известны и включают в себя в качестве неограничивающих примеров ферментативную обработку (как правило, проводимую с применением пептид-N-гликозидазы F/эндо-β-галактозидазы), элиминирование с применением жестких щелочных условий в основном для высвобождения O-связанных структур и химические способы с применением безводного гидразина для высвобождения N- и O-связанных олигосахаридов.
(viii) Иллюстративные антитела
Описанное в настоящем документе изобретение имеет ряд иллюстративных вариантов осуществления. Ниже описано множество типичных вариантов осуществления изобретения. Следующие ниже варианты осуществления предложены только в целях иллюстрации и никоим образом не предназначены для ограничения объема настоящего изобретения.
Как описано в примерах ниже, идентифицирован ряд моноклональных антител к DR5. В одном из вариантов осуществления антитела к DR5 по изобретению обладают теми же биологическими характеристиками, как любые антитела к DR5, в частности описанные в настоящем документе.
Термин "биологические характеристики" применяют для обозначения активности или свойств моноклональных антител in vitro и/или in vivo, таких как способность специфически связываться с DR5 или блокировать, индуцировать или усиливать активацию DR5 (или связанные с DR5 виды активности). Свойства и виды активности антител к DR5 дополнительно описаны в примерах ниже.
Необязательно моноклональные антитела по настоящему изобретению обладают такими же биологическими характеристиками, как антитело 16E2 или любое из антител, перечисленных в таблицах 11-13, и/или связываются с тем же эпитопом(ами), что и антитело 16E2 или любые из антител, перечисленных в таблицах 11-13, в частности Apomab 7.3 или Apomab 8.3. Это можно определить, проводя различные анализы так, как описано в настоящем документе и в примерах. Например, для определения того, обладает ли моноклональное антитело такой же специфичностью, что и антитела к DR5, в частности указанные в настоящем документе, можно сравнить их активность в анализах конкурентного связывания или анализах индукции апоптоза, таких как анализы, описанные в примерах ниже. Кроме того, эпитоп, с которым связывается конкретное антитело к DR5, можно определить посредством кристаллографического исследования комплекса между DR5 и рассматриваемым антителом.
Таким образом, кристаллографическое исследование комплекса между фрагментом Fab Apomab 7.3 и внеклеточным доменом DR5 с применением рентгенографии показало, что Apomab 7.3 связывается с эпитопом DR5, который перекрывается с участком связывания Apo2L/TRAIL на рецепторе DR5, все еще отличаясь от него.
Человеческие, химерные, гибридные или рекомбинантные антитела к DR5 (а также, например, димерные антитела или тримерные антитела, описанные в настоящем документе) могут образовывать антитело с полноразмерными тяжелыми и легкими цепями или их фрагментами, такими как фрагменты Fab, Fab', F(ab')2 или Fv, мономер или димер такой легкой или тяжелой цепи, одноцепочечный Fv, в котором такая тяжелая или легкая цепь(и) соединены линкерной молекулой, или с вариабельными доменами (или гипервариабельными доменами) такой легкой или тяжелой цепи(ей), связанными с другими типами доменов антител.
Как описано в настоящем документе, антитела к DR5 необязательно обладают одним или несколькими желательными видами биологической активности или свойствами. Такие антитела к DR5 могут включать в себя в качестве неограничивающих примеров химерные, гуманизированные, человеческие и аффинно зрелые антитела. Как описано выше, антитела к DR5 для достижения этих желательных видов активности или свойств можно синтезировать или конструировать с применением различных способов. В одном из вариантов осуществления антитело к DR5 обладает аффинностью связывания рецептора DR5 величиной, по меньшей мере, 105 M-1, предпочтительно, по меньшей мере, в диапазоне от 106 до 107 M-1, более предпочтительно, по меньшей мере, в диапазоне от 108 до 1012 M-1 и даже более предпочтительно, по меньшей мере, в диапазоне от 109 до 1012 M-1. Аффинность связывания антитела к DR5 можно без лишнего экспериментирования определять тестированием антитела к DR5 в соответствии с известными в данной области способами, включая анализ Скэтчарда (см. Munson et al., выше).
В другом варианте осуществления антитело к DR5 по изобретению может связываться с тем же эпитопом на DR5, с которым связывается Apo-2L, или связываться с эпитопом на DR5, который совпадает или перекрывается с эпитопом на DR5, с которым связывается Apo-2L, как, например, антитело, обозначенное Apomab 7.3. Антитело к DR5 также может взаимодействовать таким образом, чтобы создавать стерическую конформацию, предотвращающую связывание лиганда Apo-2 с DR5. Как указано выше, свойство антитела к DR5 по настоящему изобретению связывать эпитоп можно определять с применением известных в данной области способов. Например, для определения способности антитела к DR5 блокировать или ингибировать связывание Apo-2L с DR5 антителом к DR5 можно тестировать в анализе in vitro, таком как анализ конкурентного ингибирования. Необязательно антитело к DR5 можно тестировать в анализе конкурентного ингибирования для определения способности антитела к DR5 ингибировать связывание полипептида Apo-2L с конструкцией DR5-IgG или с клеткой, экспрессирующей DR5. Необязательно антитело к DR5 способно к блокированию или ингибированию связывания Apo-2L с DR5, по меньшей мере, на 50%, предпочтительно, по меньшей мере, на 75% и даже более предпочтительно, по меньшей мере, на 90%, что в качестве примера можно определить посредством анализа конкурентного ингибирования in vitro с применением растворимой формы лиганда Apo-2 (TRAIL) (такой как последовательность внеклеточного домена из аминокислот 114-281, описанная в Pitti et al., J. Biol. Chem., выше, также обозначаемой как Apo2L.0) и ECD DR5-IgG. Свойство антитела к DR5 связывать эпитоп также можно определить с применением анализов in vitro для тестирования способности антитела к DR5 блокировать индуцированный Apo-2L апоптоз или посредством кристаллографических исследований.
В дополнительном варианте осуществления антитело к DR5 является антителом-агонистом с активностью, сравнимой с лигандом Apo-2 (TRAIL). Предпочтительно, такое агонистическое антитело к DR5 индуцирует апоптоз, по меньшей мере, в одном типе злокачественной опухоли, или линии опухолевых клеток, или первичной опухоли. Апоптотическую активность агонистического антитела к DR5 можно определить с применением известных анализов in vitro или in vivo. Примеры ряда таких анализов in vitro и in vivo хорошо известны в данной области. In vitro апоптотическую активность можно определять с применением таких известных способов, как связывание аннексина V. In vivo апоптотическую активность можно определять, например, измерением уменьшения массы или объема опухоли.
Как указано выше, описанные в настоящем документе антитела обладают рядом свойств, включающих в себя способность регулировать некоторые физиологические взаимодействия и/или процессы. Как показано в примерах ниже, описанные в настоящем документе антитела способны индуцировать опосредованный DR5 апоптоз и демонстрируют мощные противоопухолевые свойства в различных моделях злокачественной опухоли на ксенотрансплантатах у мышей. В конкретном варианте осуществления изобретения агонистическую активность антитела увеличивают посредством связывания антитела с Fc IgG против антител человека. В предпочтительном варианте осуществления изобретения этот усиленный апоптоз сравним с апоптотической активностью Apo-2L.
Дополнительные варианты осуществления изобретения включают в себя описанное в настоящем документе антитело к рецептору DR5, которое связано с одним или несколькими небелковыми полимерами, выбранными из группы, состоящей из полиэтиленгликоля, полипропиленгликоля и полиоксиалкилена. В альтернативном варианте осуществления описанное в настоящем документе антитело к рецептору DR5 связано с цитотоксическим средством или ферментом. Еще в одном варианте осуществления описанное в настоящем документе антитело к рецептору DR5 связано с радиоактивным изотопом, флуоресцентным соединением или хемилюминесцентным соединением. Необязательно описанное в настоящем документе антитело к рецептору DR5 является гликозилированным или, альтернативно, дегликозилированным.
Как подробно обсуждается ниже, антитела по изобретению можно использовать в ряде способов регулирования физиологических процессов. Один такой вариант осуществления изобретения включает в себя способ индукции апоптоза в клетках млекопитающих, включающий в себя воздействие на клетки млекопитающих, экспрессирующие рецептор DR5, терапевтически эффективного количества выделенного моноклонального антитела к рецептору DR5, представляющего собой связывающееся с рецептором DR5 антитело, приведенное на фиг.3A-3C (411 аминокислот) или фиг.4A-4C (440 аминокислот), особенно с его внеклеточным доменом. В таких способах клетки млекопитающих, как правило, представляют собой злокачественные клетки. В предпочтительных вариантах осуществления используемое в этих способах антитело к рецептору DR5 представляет собой описанное в примерах ниже антитело Apomab, такое как антитело Apomab 7.3 или Apomab 8.3.
Еще один вариант осуществления изобретения представляет собой способ индукции апоптоза в клетках млекопитающих, включающий в себя воздействие на экспрессирующие рецептор DR5 клетки млекопитающих терапевтически эффективного количества выделенного моноклонального антитела к рецептору DR5, представляющего собой антитело, связывающееся с рецептором DR5, как определено выше в настоящем документе, или с его внеклеточным доменом.
3. Тримерные антитела
Также в объеме изобретения находятся тримерные антитела. Такие антитела описаны, например, в Iliades et al., выше, и Kortt et al., выше.
4. Другие модификации
В настоящем документе рассматривают другие модификации антител к DR5. Антитела по настоящему изобретению можно модифицировать посредством конъюгации антитела с цитотоксическим средством (таким как молекула токсина) или с активирующим пролекарство ферментом, который конвертирует пролекарство (например, пептидильное химиотерапевтическое средство, см. WO 81/01145) в активное противораковое лекарственное средство. См., например, WO 88/07378 и патент США №4975278. Эту технологию также обозначают как "антителозависимая опосредованная ферментами пролекарственная терапия" (ADEPT).
Ферментативный компонент иммуноконъюгата, пригодного для ADEPT, включает в себя фермент, способный действовать на пролекарство таким образом, что оно конвертируется в его более активную цитотоксическую форму. Ферменты, пригодные для этого способа по данному изобретению, включают в себя в качестве неограничивающих примеров щелочную фосфатазу, пригодную для преобразования в свободные лекарственные средства пролекарств, содержащих фосфаты; арилсульфатазу, пригодную для преобразования в свободные лекарственные средства пролекарств, содержащих сульфаты; цитозиндезаминазу, пригодную для преобразования в противораковое лекарственное средство, 5-фторурацил, нетоксического 5-фторцитозина; протеазы, такие как протеазы Serratia, термолизин, субтилизин, карбоксипептидазы и катепсины (такие как катепсины B и L), пригодные для преобразования в свободные лекарственные средства пролекарств, содержащих пептиды; каспазы, такие как каспаза-3; D-аланилкарбоксипептидазы, пригодные для преобразования пролекарств, содержащих заместители из D-аминокислот; расщепляющие углеводы ферменты, такие как β-галактозидаза и нейраминидаза, пригодные для преобразования в свободные лекарственные средства гликозилированных пролекарств; бета-лактамазу, пригодную для преобразования в свободные лекарственные средства лекарственных средств, дериватизированных β-лактамами; и пенициллинамидазы, такие как пенициллин V амидаза или пенициллин G амидаза, пригодные для преобразования в свободные лекарственные средства лекарственных средств, дериватизированных по их азотам аминов феноксиацетильными или фенилацетильными группами, соответственно. Альтернативно, для преобразования пролекарств в свободные лекарственные средства можно применять антитела с ферментативной активностью, также известные в данной области как "абзимы" (см., например, Massey, Nature 328:457-458 (1987)). Конъюгаты антитело-абзим можно получать, как описано в настоящем документе, для доставки абзима в популяцию опухолевых клеток.
Ферменты можно ковалентно связывать с антителами посредством хорошо известных в данной области способов, таких как применение гетеробифункциональных сшивающих средств. Альтернативно, с применением хорошо известных в данной области способов рекомбинантных ДНК, можно сконструировать слитные белки, содержащие, по меньшей мере, антигенсвязывающую область антитела по изобретению, связанную, по меньшей мере, с функционально активной частью фермента по изобретению (см., например, Neuberger et al., Nature 312:604-608 (1984)).
Рассмотрены дополнительные модификации антител. Например, антитело можно связывать с одним из множества небелковых полимеров, например полиэтиленгликолем, полипропиленгликолем, полиоксиалкиленами или сополимерами полиэтиленгликоля и полипропиленгликоля. Антитело также можно заключать в микрокапсулы, полученные, например, посредством способов коацервации или межфазной полимеризации (например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли(метилметацилатные) микрокапсулы, соответственно), в коллоидные системы доставки лекарственного средства (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences, 16th edition, Oslo, A., Ed., (1980). Для увеличения времени полужизни антитела в сыворотке в антитело можно вставлять эпитоп связывания рецептора спасения (особенно во фрагмент антитела), например, как описано в патенте США №5739277. Как применяют в настоящем документе, термин "эпитоп связывания рецептора спасения" относится к эпитопу Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), ответственному за увеличение времени полужизни молекулы IgG в сыворотке in vivo.
Описанные в настоящем документе антитела к DR5 также можно составлять в виде иммунолипосом. Содержащие антитело липосомы получают известными в данной области способами так, как описано в Epstein et al., Proc. Natl. Acad. Sci. USA, 82:3688 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA, 77:4030 (1980); патентах США №№4485045 и 4544545. В патенте США №5013556 описаны липосомы с увеличенным временем циркуляции. Особенно пригодные липосомы можно получать способом обращенно-фазового выпаривания липидной композиции, содержащей фосфатидилхолин, холестерин и дериватизированный PEG фосфатидилэтаноламин (PEG-PE). Липосомы экструдируют через фильтры с определенным размером пор с получением липосом с желательным диаметром. Фрагменты Fab' антитела по настоящему изобретению можно конъюгировать с липосомами, как описано в Martin et al., J. Biol. Chem. 257:286-288 (1982), посредством реакции дисульфидного обмена. В липосоме необязательно содержится химиотерапевтическое средство (такое как доксорубицин). См. Gabizon et al., J. National Cancer Inst. 81(19):1484 (1989).
Антитела по изобретению включают в себя "сшитые" антитела к DR5. Как применяют в настоящем документе, термин "сшитый" относится к связыванию, по меньшей мере, двух молекул IgG вместе с формированием одной (или отдельной) молекулы. Антитела к DR5 можно сшивать различными линкерными молекулами, предпочтительно антитела к DR5 сшивают с применением молекулы антитела к IgG, комплемента, химической модификации или молекулярной инженерии. Специалистам в данной области понятно, что комплемент обладает относительно высокой аффинностью к молекулам антител после связывания антител с клеточной поверхностной мембраной. Таким образом, полагают, что комплемент можно использовать как сшивающую молекулу для связывания двух или более антител к DR5, связанных с клеточной поверхностной мембраной.
5. Рекомбинантные способы
Изобретение также относится к выделенным нуклеиновым кислотам, кодирующим антитела к DR5, как описано в настоящем документе, векторам и клеткам-хозяевам, содержащим нуклеиновую кислоту, и рекомбинантным способам получения антител.
Для рекомбинантного получения антитела кодирующую его нуклеиновую кислоту выделяют и вставляют в реплицирующийся вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии. Кодирующую антитело ДНК легко выделять и секвенировать с применением общепринятых способов (например, с применением олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими антитело). Доступно множество векторов. Компоненты векторов, как правило, включают в себя в качестве неограничивающих примеров один или несколько из следующего: сигнальная последовательность, участок начала репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
Способы по настоящему документу включают в себя способы получения химерных или рекомбинантных антител к DR5, включающих в себя стадии получения вектора, содержащего последовательность ДНК, кодирующую легкую цепь или тяжелую цепь антитела к DR5 (или и легкую цепь, и тяжелую цепь), трансфекции или трансформации клетки-хозяина вектором и культивирования клетки(ок)-хозяина в условиях, достаточных для получения продукта в виде рекомбинантного антитела к DR5.
(i) Компонент сигнальной последовательности
Антитело к DR5 по данному изобретению можно получать рекомбинантным способом не только непосредственно, но и в виде слитного белка с гетерологичным полипептидом, которой предпочтительно представляет собой сигнальную последовательность или другой полипептид со специфическим участком расщепления на N-конце зрелого белка или полипептида. Выбираемая гетерологичная сигнальная последовательность предпочтительно представляет собой сигнальную последовательность, которая распознается и процессируется (т.е. расщепляется сигнальной пептидазой) клеткой-хозяином. Для прокариотических клеток-хозяев, которые не распознают и не процессируют природную сигнальную последовательность антитела, сигнальную последовательность заменяют прокариотической сигнальной последовательностью, например, выбранной из группы лидирующих последовательностей щелочной фосфатазы, пенициллиназы, lpp или термостабильного энтеротоксина II. Для секреции в дрожжах природную сигнальную последовательность можно заменять, например, на лидирующую последовательность дрожжевой инвертазы, лидирующую последовательность α-фактора (включая лидирующие последовательности α-фактора Saccharomyces и Kluyveromyces) или лидирующую последовательность кислой фосфатазы, лидирующую последовательность глюкоамилазы C.albicans или сигнальную последовательность, описанную в WO 90/13646. Для экспрессии в клетках млекопитающих доступны сигнальные последовательности млекопитающих, а также секреторные лидирующие последовательности вирусов, например сигнальная последовательность gD вируса простого герпеса.
ДНК для такой предшествующей области лигируют в рамку считывания кодирующей антитело ДНК.
(ii) Компонент участка начала репликации
И экспрессирующие, и клонирующие векторы содержат последовательность нуклеиновой кислоты, позволяющую векторам реплицироваться в одной или нескольких выбранных клетках-хозяевах. Как правило, в клонирующих векторах эта последовательность представляет собой последовательность, которая позволяет вектору реплицироваться независимо от хромосомной ДНК хозяина и включает в себя участки начала репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для множества бактерий, дрожжей и вирусов. Участок начала репликации из плазмиды pBR322 подходит для большинства грамотрицательных бактерий, точка начала репликации плазмиды 2μ подходит для дрожжей, а различные вирусные точки начала репликации (SV40, полиомы, аденовируса, VSV или BPV) подходят для клонирования векторов в клетках млекопитающих. Как правило, компонент участка начала репликации для экспрессирующих векторов млекопитающих не нужен (участок начала репликации SV40, как правило, можно использовать только потому, что он содержит ранний промотор).
(iii) Компонент селективного гена
Экспрессирующие и клонирующие векторы, как правило, содержат селективный ген, также называемый селективным маркером. Типичные селективные гены кодируют белки, которые (a) обеспечивают устойчивость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (b) дополняют ауксотрофный дефицит или (c) обеспечивают необходимые питательные вещества, недоступные из комплексных сред, например ген, кодирующий D-аланинрацемазу для Bacilli.
В одном из примеров схемы селекции используют лекарственное средство для ареста роста клетки-хозяина. Те клетки, которые успешно подверглись трансформации гетерологичным геном, продуцируют белок, придающий устойчивость к лекарственному циклу и, таким образом, выживают в режиме селекции. В примерах такой доминантной селекции используют лекарственные средства неомицин, микофеноловую кислоту и гигромицин.
Другим примером подходящих селективных маркеров для клеток млекопитающих являются селективные маркеры, обеспечивающие идентификацию клеток, способных захватывать кодирующую антитело нуклеиновую кислоту, такие как DHFR, тимидинкиназа, металлотионеин-I и -II, предпочтительно гены металлотионеина приматов, аденозиндезаминаза, орнитиндекарбоксилаза и т.д.
Например, клетки, трансформированные селективным геном DHFR, сначала идентифицируют посредством культивирования всех трансформантов в среде для культивирования, содержащей метотрексат (Mtx), конкурентный антагонист DHFR. Подходящей клеткой-хозяином при применении DHFR дикого типа является клеточная линия яичника китайского хомяка (CHO) с дефицитом активности DHFR.
Альтернативно клетки-хозяева (в особенности хозяева дикого типа, несущие эндогенную DHFR), трансформированные или котрансформированные последовательностями ДНК, кодирующими антитело к DR5, белок DHFR дикого типа и другой селективный маркер, такой как аминогликозид-3'-фосфотрансфераза (APH), можно отбирать по росту клеток в среде, содержащей селективное вещество для селективного маркера, такое как аминогликозидный антибиотик, например канамицин, неомицин или G418. См. патент США №4965199.
Подходящий селективный ген для применения в дрожжах представляет собой ген trp1, находящийся в дрожжевой плазмиде YRp7 (Stinchcomb et al., Nature, 282:39 (1979)). Ген trp1 предоставляет селективный маркер для мутантного штамма дрожжей с отсутствующей способностью расти на триптофане, например ATCC №44076 или PEP4-1. Jones, Genetics, 85:12 (1977). Таким образом, наличие повреждения trp1 в геноме дрожжевой клетки-хозяина обеспечивает эффективные условия для детекции трансформации посредством роста в отсутствие триптофана. Подобным образом, дефектные по Leu2 штаммы дрожжей (ATCC 20622 или 38626) комплементируют известными плазмидами, несущими ген Leu2.
Кроме того, для трансформации Kluyveromyces yeasts можно применять векторы, полученные из 1,6 мкм кольцевой плазмиды pKD1. Альтернативно для K.lactis представлена экспрессирующая система для крупномасштабного получения телячьего химозина. Van den Berg, Bio/Technology, 8:135 (1990). Также описаны стабильные мультикопийные экспрессирующие векторы для секреции зрелого рекомбинантного сывороточного альбумина промышленными штаммами Kluyveromyces. Fleer et al., Bio/Technology, 9:968-975 (1991).
(iv) Промоторный компонент
Экспрессирующие и клонирующие векторы, как правило, содержат промотор, распознаваемый организмом хозяина и функционально связанный с кодирующей антитело нуклеиновой кислотой. Промоторы, подходящие для применения в прокариотических хозяевах, включают в себя промотор phoA, промоторные системы β-лактамазы и лактозы, щелочной фосфатазы, промоторную систему триптофана (trp) и гибридные промоторы, такие как промотор tac. Однако подходят и другие известные бактериальные промоторы. Промоторы для применения в бактериальных системах также содержат последовательность Шайна-Дальгарно (S.D.), функционально связанную с кодирующей антитело к DR5 ДНК.
Известны промоторные последовательности для эукариот. Фактически все эукариотические гены имеют AT-богатую область, расположенную приблизительно от 25 до 30 основаниями выше участка инициации транскрипции. Другая последовательность, находящаяся на расстоянии от 70 до 80 оснований выше старта транскрипции множества генов, представляет собой область CNCAAT, где N может представлять собой любой нуклеотид. На 3'-конце большинства эукариотических генов находится последовательность AATAAA, которая может представлять собой сигнал добавления поли-A хвоста на 3'-конце кодирующей последовательности. Соответственно, все эти последовательности вставлены в эукариотические экспрессирующие векторы.
Примеры подходящих промоторных последовательностей для применения с дрожжевыми хозяевами включают в себя промоторы для 3-фосфоглицераткиназы или других гликолитических ферментов, таких как енолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
Другие дрожжевые промоторы, которые являются индуцибельными промоторами с дополнительным преимуществом транскрипции, контролируемой условиями роста, представляют собой промоторные области алкогольдегидрогеназы 2, изоцитохрома C, кислой фосфатазы, катаболических ферментов, связанных с метаболизмом азота, металлотионеина, глицеральдегид-3-фосфатдегидрогеназы и ферментов, отвечающих за утилизацию мальтозы и галактозы. Подходящие векторы и промоторы для применения при экспрессии в дрожжах дополнительно описаны в EP 73657. С дрожжевыми промоторами также успешно применяются дрожжевые энхансеры.
Транскрипция антитела к DR5 с векторов в клетках-хозяевах млекопитающих контролируется, например, промоторами, полученными из геномов вирусов, таких как вирус полиомы, вирус куриной оспы, аденовирус (такой как аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита B и вирус обезьян 40 (SV40), из гетерологичных промоторов млекопитающих, например промотора актина или промотора иммуноглобулина и из промоторов белков теплового шока, при условии, что такие промоторы совместимы с системами клетки-хозяина.
Ранний и поздний промоторы вируса SV40 удобно получать в виде рестрикционного фрагмента SV40, который также содержит участок начала репликации вируса SV40. Предранний промотор цитомегаловируса человека удобно получать в виде рестрикционного фрагмента E HindIII. Система для экспрессии ДНК в хозяевах-млекопитающих с применением вируса папилломы крупного рогатого скота описана в патенте США №4419446. Модификация этой системы описана в патенте США №4601978. Также см. Reyes et al., Nature 297:598-601 (1982), об экспрессии кДНК интерферона β человека в клетках мыши под контролем промотора тимидинкиназы из вируса простого герпеса. Альтернативно в качестве промотора можно использовать длинный концевой повтор вируса саркомы Рауса.
(v) Компонент энхансерного элемента
Транскрипцию высшими эукариотами кодирующей антитело к DR5 по данному изобретению ДНК часто усиливают посредством вставки в вектор энхансерной последовательности. В настоящее время известно множество энхансерных последовательностей из генов млекопитающих (глобин, эластаза, альбумин, α-фетопротеин и инсулин). Однако, как правило, используют энхансер из вируса эукариотических клеток. Примеры включают в себя энхансер SV40 в поздней области от точки начала репликации (100-270 п.н.), энхансер раннего промотора цитомегаловируса, энхансер полиомы в поздней области от точки начала репликации и энхансеры аденовирусов. Также об энхансерных элементах для активации эукариотических промоторов см. Yaniv, Nature 297:17-18 (1982). Энхансер можно вставлять в вектор в положении 5' или 3' от кодирующей последовательности антитела, но предпочтительно он расположен в положении 5' от промотора.
(vi) Компонент терминации транскрипции
Экспрессирующие векторы, используемые в эукариотических клетках-хозяевах (дрожжи, грибы, насекомые, растения, животные, человек или ядросодержащие клетки других многоклеточных организмов), также содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно доступны из 5' и иногда из 3' нетранслируемых областей эукариотической или вирусной ДНК или кДНК. Эти области содержат нуклеотидные сегменты, транскрибируемые в виде полиаденилированных фрагментов в нетранслируемой части мРНК, кодирующей поливалентное антитело. Один из пригодных компонентов терминации транскрипции представляет собой область полиаденилирования гормона роста крупного рогатого скота. См. WO 94/11026 и описанный в ней экспрессирующий вектор.
(vii) Отбор и трансформация клеток-хозяев
Подходящие для клонирования или экспрессии ДНК в векторах по настоящему документу клетки-хозяева представляют собой клетки прокариот, дрожжей или высших эукариот, описанные выше. Подходящие для этой цели прокариоты включают в себя эубактерии, такие как грамотрицательные или грамположительные организмы, например Enterobacteriaceae, такие как Escherichia, например E.coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans и Shigella, а также Bacilli, такие как B.subtilis и B.licheniformis (например, B.licheniformis 41P, описанная в DD 266710, опубликованной 12 апреля 1989 года), Pseudomonas, такие как P.aeruginosa, и Streptomyces. Один из предпочтительных для клонирования хозяев E.coli представляет собой E.coli 294 (ATCC 31446), хотя подходят и другие штаммы, такие как E.coli B, E.coli X1776 (ATCC 31537) и E.coli W3110 (ATCC 27325). Эти примеры являются иллюстративными, а не лимитирующими.
Кроме прокариот, подходящими хозяевами для клонирования или экспрессии кодирующих антитела к DR5 векторов являются эукариотические микроорганизмы, такие как нитевидные грибы или дрожжи. Наиболее часто используемым низшим эукариотическим микроорганизмом-хозяином являются Saccharomyces cerevisiae. Однако широкодоступным и подходящим по настоящему документу является ряд других родов, видов и штаммов, таких как Schizosaccharomyces pombe; хозяева Kluyveromyces, например, такие как K.lactis, K.fragilis (ATCC 12424), K.bulgaricus (ATCC 16045), K.wickeramii (ATCC 24178), K.waltii (ATCC 56500), K.drosophilarum (ATCC 36906), K.thermotolerans и K.marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis и нитевидные грибы, например, такие как Neurospora, Penicillium, Tolypocladium, и хозяева Aspergillus, такие как A.nidulans и A.niger.
Подходящие клетки-хозяева для экспрессии гликозилированных антител получают из многоклеточных организмов. Примеры клеток беспозвоночных включают в себя клетки растений и насекомых. Идентифицировано множество бакуловирусных штаммов и вариантов и соответствующие пермиссивные клетки-хозяева насекомых таких организмов, как Spodoptera frugiperda (пиявка), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Общедоступным является ряд вирусных штаммов для трансфекции, например вариант L-1 NPV Autographa californica и штамм Bm-5 NPV Bombyx mori, и такие вирусы по настоящему документу можно использовать в качестве вируса по настоящему изобретению, особенно для трансфекции клеток Spodoptera frugiperda.
В качестве хозяев также можно использовать культуры растительных клеток хлопка, кукурузы, картофеля, сои, петунии, томата и табака.
Однако наибольший интерес представляют клетки позвоночных, и культивирование клеток позвоночных в культуре (тканевая культура) стало обычным способом. Примеры подходящих хозяйских клеточных линий млекопитающих представляют собой линия CV1 почек обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); линия эмбриональной почки человека (293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol. 36:59 (1977)); клетки почки детеныша хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени буйволовой крысы (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мыши (MMT 060562, ATCC CCL 51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; клетки FS4 и линия гепатомы человека (Hep G2) и миеломные или лимфомные клетки (например, клетки Y0, J558L, P3 и NS0) (см. патент США №5807715).
Клетки-хозяева трансформируют описанными выше экспрессирующими или клонирующими векторами для продукции антител и культивируют в общепринятых питательных средах, соответствующим образом модифицированных для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих желательные последовательности.
(viii) Культивирование клеток-хозяев
Клетки-хозяева, используемые для получения антител по данному изобретению, можно культивировать в ряде сред. Для культивирования клеток-хозяев подходят коммерчески доступные среды, такие как Ham's F10 (Sigma), минимальная поддерживающая среда ((MEM), Sigma)), RPMI-1640 (Sigma) и модифицированная по Дульбекко среда Игла ((DMEM), Sigma)). Кроме того, в качестве сред для культивирования клеток-хозяев можно использовать любую из сред, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102:255 (1980), патентах США №№4767704; 4657866; 4927762; 4560655 или 5122469; WO 90/03430; WO 87/00195 или ссылке на патент США 30985. Любую из этих сред можно по мере необходимости дополнять гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальция, магния и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство GENTAMYCIN™), микроэлементами (определенными как неорганические соединения, как правило, присутствующие в конечных концентрациях в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Также можно добавлять любые другие необходимые добавки в подходящих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., представляют собой условия культивирования, используемые ранее для клеток-хозяев, отбираемых по экспрессии, и они, как правило, очевидны специалистам в данной области.
(ix) Очистка
При применении рекомбинантных способов антитело может продуцироваться внутриклеточно, в периплазматическом пространстве или непосредственно секретироваться в среду. Если антитело продуцируется внутриклеточно, в качестве первой стадии удаляют дебрис из макрочастиц из клеток-хозяев или из фрагментов лизиза, например, центрифугированием или ультрацентрифугированием. В Carter et al., Bio/Technology 10:163-167 (1992), описан способ выделения антител, секретируемых в периплазматическое пространство E.coli. В кратком изложении, клеточную массу размораживают в присутствии ацетата натрия (pH 3,5), ЭДТА и фенилметилсульфонилфторида (PMSF) в течение приблизительно 30 мин. Клеточный дебрис можно удалять центрифугированием. Когда антитело секретируется в среду, супернатанты из таких систем экспрессии, как правило, вначале концентрируют с применением коммерчески доступного фильтра для концентрации белков, например устройства для ультрафильтрации Amicon или Millipore Pellicon. На любой из указанных выше стадий для ингибирования протеолиза можно добавлять ингибитор протеаз, такой как PMSF, а также можно добавлять антибиотики для предотвращения роста случайных загрязнителей.
Композиции антител, полученные из клеток, можно очищать с применением, например, хроматографии на гидроксиапатите, гель-электрофореза, диализа и аффинной хроматографии, где аффинная хроматография является предпочтительным способом очистки. Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа какого-либо домена Fc иммуноглобулина, который присутствует в антителе. Белок A можно использовать для очистки антител, основанных на тяжелых цепях γ1, γ2 или γ4 человека (Lindmark et al., J. Immunol. Meth. 62:1-13 (1983)). Белок G рекомендован для всех изотипов мышей и для γ3 человека (Guss et al., EMBO J. 5:1567-1575 (1986)). Матрикс, к которому прикреплен аффинный лиганд, чаще всего представляет собой агарозу, но также доступны другие матриксы. Механически устойчивые матриксы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокие скорости потока и более короткое время обработки, чем те, которые можно достичь с применением агарозы. Когда антитело содержит домен CH3, для очистки подходит смола Bakerbond ABX™ (J. T. Baker, Phillipsburg, NJ). В зависимости от антитела для восстановления также доступны другие способы очистки белков, такие как разделение на ионообменной колонке, осаждение этанолом, ВЭЖХ с обращенной фазой, хроматография на силикагеле, хроматография на SEPHAROSE™ с гепарином, хроматография на анионо- или катионообменной смоле (такой как колонка с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония.
B. Применения антител к DR5
Антитела к DR5 по изобретению имеют различные применения.
Известно, что DR5 опосредует передачу сигнала апоптоза. Хотя DR5 экспрессируют несколько типов нормальных клеток, по-видимому, передача сигнала апоптоза через этот рецептор в основном ограничена злокачественными клетками, которые становятся более чувствительными к опосредованному рецептором смерти апоптозу в случае их трансформации такими онкогенами, как MYC или RAS (Wang et al., Cancer Cell 5:501-12 (2004); Nesterov et al., Cancer Res. 64:3922-7 (2004)). DR5 часто экспрессируется линиями злокачественных клеток человека, а также первичными опухолями. Таким образом, антитела к DR5 находят применение при диагностике и лечении злокачественных опухолей. Например, агонистические антитела к DR5 можно использовать в способах лечения злокачественных опухолей у млекопитающих, включая людей. В этих способах антитело к DR5, предпочтительно агонистическое антитело, вводят млекопитающему отдельно или в сочетании с другими терапевтическими средствами или способами. Злокачественная опухоль может представлять собой экспрессирующую DR5 злокачественную опухоль любого типа, включая солидные опухоли, в частности развитые или метастатические солидные опухоли, прогрессировавшие до терапии или для которых не известно эффективной терапии. Отдельные типы злокачественных опухолей включают в себя в качестве неограничивающих примеров колоректальный рак, немелкоклеточный рак легких (NSCLC), рак поджелудочной железы, рак яичников, рак молочной железы, неходжкинскую лимфому (NHL), глиобластому или меланому, предпочтительно колоректальный рак, NSCLC или NHL.
Кроме того, антитела к DR5 пригодны в диагностике и лечении других ассоциированных с DR5 патологических состояний, таких как связанные с иммунной системой заболевания у млекопитающих, включая людей.
Диагноз различных патологических состояний млекопитающих, описанных в настоящем документе, может быть поставлен практикующими специалистами. В данной области доступны способы диагностики, позволяющие, например, диагностировать или выявлять злокачественную опухоль или связанное с иммунной системой заболевание у млекопитающего. Например, злокачественные опухоли можно определять способами, включающими в себя, в качестве неограничивающих примеров, пальпацию, анализ крови, рентгенограмму, ЯМР и т.п.
Связанные с иммунной системой заболевания также можно легко выявить.
При системной красной волчанке основным медиатором заболевания является продукция аутореактивных антител к собственным белкам/тканям и последующее формирование опосредованного иммунной системой воспаления. Клинически поражены несколько органов и систем, включая почки, легкие, костно-мышечную систему, кожно-слизистую систему, глаза, центральную нервную систему, сердечно-сосудистую систему, желудочно-кишечный тракт, костный мозг и кровь.
Ревматоидный артрит (RA) представляет собой хроническое системное аутоиммунное воспалительное заболевание, которое в основном затрагивает синовиальную мембрану нескольких суставов, с формирующимся в результате повреждением суставного хряща. Патогенез зависит от T-лимфоцитов и связан с образованием ревматоидных факторов, аутоантител, направленных против собственных IgG, с образующимися в результате иммунными комплексами, которые достигают высоких уровней в суставной жидкости и крови. Эти комплексы в суставе могут индуцировать заметный инфильтрат лимфоцитов и моноцитов в синовиальной оболочке и последующие заметные изменения в синовиальной оболочке; суставное пространство/жидкость инфильтрировано сходными клетками с добавлением многочисленных нейтрофилов. Поражаемые ткани в основном представляют собой сосуды, часто симметричным образом. Однако также происходит внесуставное поражение в виде двух основных форм. Одна из форм представляет собой развитие внесуставных очагов повреждения с постоянно прогрессирующим поражением сосудов и типичными очагами фиброза легких, васкулита и кожных язв. Вторая форма внесуставного поражения представляет собой так называемый синдром Фелти, который возникает на поздних стадиях течения RA, иногда после того как поражение суставов становится неактивным, и включает в себя наличие нейтропении, тромбоцитопении и спленомегалии. Это может сопровождаться васкулитом в нескольких органах с формированием инфарктов, язв кожи и гангрены. Часто у пациентов в подкожной ткани над пораженными суставами также развиваются ревматоидные узелки; узелки поздних стадий содержат некротические центры, окруженные инфильтратом разнородных воспалительных клеток. Другие проявления, которые могут происходить при RA, включают в себя перикардит, плеврит, коронарный артериит, интерстициальный пневмонит с фиброзом легких, сухой кератоконъюнктивит и ревматоидные узелки.
Ювенильный хронический артрит представляет собой хроническое идиопатическое воспалительное заболевание, часто начинающееся в возрасте менее 16 лет. Его фенотип имеет некоторые сходства с RA; некоторых пациентов, положительных по ревматоидному фактору, классифицируют как пациентов с ювенильным ревматоидным артритом. Заболевание подразделяют на три основных категории: малосуставное, полисуставное и системное. Артрит может быть тяжелым и, по меньшей мере, деструктивным и приводит к анкилозу суставов и задержке роста. Другие проявления могут включать хронический передний увеит и системный амилоидоз.
Спондилоартропатии представляют собой группу заболеваний с некоторыми общими клиническими признаками и общей связью с экспрессией продукта гена HLA-B27. Заболевания включают в себя анкилозирующий спондилит, синдром Рейтера (реактивный артрит), артрит, ассоциированный с воспалительным заболеванием кишечника, спондилит, ассоциированный с псориазом, спондилоартропатией с началом в юношеском возрасте и недифференцированной спондилоартропатией. Дифференциальные характеристики включают в себя сакроилеит со спондилитом или без; воспалительный ассиметричный артрит; ассоциацию с HLA-B27 (серологически определенный аллель локуса HLA-B MHC I класса); воспаление глаз и отсутствие аутоантител, ассоциированных с другим ревматоидным заболеванием. Клетка, наиболее вовлеченная в качестве ключа в индукцию заболевания, представляет собой CD8+ T-лимфоцит, клетку, направленную к антигену, представляемому молекулами MHC I класса. CD8+ T-клетки могут реагировать против аллеля HLA-B27 MHC I класса, как если бы он был чужеродным пептидом, экспрессированным молекулами MHC I класса. Выдвинуто предположение, что эпитоп HLA-B27 может имитировать бактериальный или другой микробный антигенный эпитоп и, таким образом, индуцировать ответ CD8+ T-клеток.
Этиология системного склероза (склеродермии) неизвестна. Отличительной чертой этого заболевания является уплотнение кожи; вероятно, это индуцировано активным воспалительным процессом. Склеродермия может быть локализованной или системной; частыми являются повреждения сосудов, и повреждение эндотелиальных клеток в микроциркуляторном русле является ранним и важным событием в развитии системного склероза; повреждение сосудов может быть обусловлено иммунной системой. Иммунологическое основание полагают вследствие присутствия в острых очагах повреждения инфильтратов мононуклеарных клеток и наличия у многих пациентов антител к содержимому ядер. Часто на поверхности фибробластов в очагах повреждения на коже повышена экспрессия ICAM-1, позволяя предположить, что в патогенезе заболевания может играть роль взаимодействие с этими клетками T-клеток. Другие вовлеченные органы включают в себя желудочно-кишечный тракт: атрофию гладкой мускулатуры и фиброз, приводящий к аномальной перестальтике/сократительной активности; почки: концентрическая эндотелиальная пролиферация интимы, влияющая на малые дуговые и междольковые артерии, с происходящим в результате снижением почечного коркового кровотока, приводящего к протеинурии, азотемии и гипертензии; скелетная мускулатура: атрофия, интерстициальный фиброз, воспаление; легкие: интерстициальный пневмонит и интерстициальный фиброз; и сердца, некроз сократительных волокон, образование рубцов/фиброз.
Идиопатические воспалительные миопатии, включающие в себя дерматомиозит, полимиозит и другие, представляют собой нарушения в виде хронического мышечного воспаления неизвестной этиологии, приводящие к мышечной слабости. Повреждение/воспаление мышц часто является симметричным и прогрессирующим. С большинством форм ассоциированы аутоантитела. Эти специфичные для миозита аутоантитела направлены против и ингибируют функционирование компонентов, белков и РНК, вовлеченных в синтез белков.
Синдром Шегрена возникает вследствие иммуноопосредованного воспаления и последующей функциональной деструкции слезных и слюнных желез. Заболевание может быть ассоциировано с воспалительными заболеваниями соединительной ткани и сопровождаться ими. Это заболевание ассоциировано с продукцией антител к антигенам Ro и La, которые оба представляют собой небольшие комплексы РНК-белок. Повреждение приводит к сухому кератоконъюнктивиту, ксеростомии с другими проявлениями или корреляциями, включая биллиарный цирроз, периферическую или сенсорную нейропатию и пальпируемую пурпуру.
Системный васкулит включает заболевания, в которых первичное повреждение представляет собой воспаление и последующее повреждение кровеносных сосудов, приводящее к ишемии/некрозу/дегенерации тканей, питаемых пораженными сосудами, а в некоторых случаях - к возможной дисфункции конечных органов. Васкулиты также могут возникать как вторичное повреждение или осложнение при других иммунных воспалительных заболеваниях, таких как ревматоидный артрит, системный склероз и т.д., особенно при заболеваниях, также ассоциированных с формированием иммунных комплексов. Заболевания в группе первичных системных васкулитов включают в себя системный некротизирующий васкулит: узелковый полиартериит, аллергический васкулит и гранулематоз, полиангиит; гранулематоз Вегенера; лимфоматоидный гранулематоз и гигантоклеточный артериит. Прочие васкулиты включают в себя слизисто-кожный лимфоузелковый синдром (MLNS или болезнь Кавасаки), изолированный васкулит ЦНС, болезнь Бехчета, облитерирующий тромбангиит (болезнь Бюргера) и кожный некротизирующий венулит. Полагают, что патогенетический механизм большинства типов перечисленных васкулитов, в основном, основан на отложении комплексов иммуноглобулинов в сосудистой стенке и последующей индукции противовоспалительного ответа посредством ADCC, активации комплемента или обоих.
Саркоидоз представляет собой состояние неизвестной этиологии, характеризующееся наличием эпителиоидных гранулем почти во всех тканях организма; наиболее часто вовлекаются легкие. Патогенез включает в себя персистенцию активированных макрофагов и лимфоидных клеток в очагах заболевания с последующими хроническими последствиями, приводящими к высвобождению локально и системно активных продуктов, высвобождаемых этими типами клеток.
Аутоиммунная гемолитическая анемия, включающая в себя аутоиммунную гемолитическую анемию, иммунную панцитопению и пароксизмальную ночную гемоглобинурию, является результатом продукции антител, реагирующих с антигенами, экспрессируемыми на поверхности красных кровяных клеток (а в некоторых случаях - других клеток крови, включая также и тромбоциты), и представляет собой отражение удаления этих покрытых антителами клеток вследствие опосредованного комплементом лизиса и/или опосредованных ADCC/Fc-рецептором механизмов.
При аутоиммунной тромбоцитопении, включающей в себя тромбоцитопеническую пурпуру и опосредованную иммунной системой тромбоцитопению в других клинических условиях, происходит разрушение/удаление тромбоцитов в результате прикрепления к тромбоцитами антител или комплемента и последующего удаления посредством лизиса комплементом, ADCC или опосредованными FC-рецептором механизмами.
Тиреоидит, включающий в себя болезнь Грейва, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит и атрофический тиреоидит, представляет собой результат аутоиммунного ответа на антигены щитовидной железы с продукцией антител, которые реагируют с белками, представленными на щитовидной железе и часто специфичными для нее. Существуют экспериментальные модели, включающие в себя модели с самопроизвольным началом заболевания: крысы (BUF и крысы BB) и куры (линия тучных кур); модели с индуцируемым началом заболевания: иммунизация животных или тиреоглобулином, или антигеном микросомальной фракции щитовидной железы (пероксидаза щитовидной железы).
Сахарный диабет I типа или инсулинозависимый диабет представляет собой аутоиммунное разрушение β-клеток островков поджелудочной железы; это разрушение опосредовано аутоантителами и аутореактивными T-клетками. Антитела к инсулину или инсулиновому рецептору также могут давать фенотип неотвечаемости на инсулин.
Опосредованные иммунной системой заболевания почек, включающие в себя гломерулонефрит и тубулоинтерстициальный нефрит, являются результатом опосредованного антителами или T-лимфоцитами повреждения ткани почек или непосредственно, как результат продукции аутореактивных антител или T-клеток против почечных антигенов, или опосредованно, как результат депонирования в почках антител и/или иммунных комплексов, реактивных в отношении других, непочечных антигенов. Таким образом, другие опосредованные иммунной системой заболевания, приводящие к формированию иммунных комплексов, также могут индуцировать опосредованное иммунной системой заболевание почек в виде непрямого осложнения. Прямые и непрямые иммунные механизмы приводят к воспалительному ответу, продуцирующему/индуцирующему развитие повреждения в почечной ткани с возникающей в результате функциональной недостаточностью, а в некоторых случаях - с прогрессированием до почечной недостаточности. В патогенез повреждения могут быть вовлечены и гуморальные, и клеточные иммунные механизмы.
Полагают, что демиелинизирующие заболевания центральной и периферической нервной системы, включающие в себя рассеянный склероз; идиопатическую демиелинизирующую полинейропатию или синдром Гийена-Барре и хроническую воспалительную демиелинизирующую полинейропатию, имеют аутоиммунное основание и приводят к демиелинизации нервов в результате повреждения, причиняемого олигодендроцитам или непосредственно миелину. При РС существуют основания полагать, что индукция и прогрессирование заболевания зависят от T-лимфоцитов. Рассеянный склероз представляет собой демиелинизирующее заболевание, являющееся T-лимфоцитзависимым, и обладает или ремитирующим курсом течения, или хронически прогрессирующим курсом течения. Этиология РС неизвестна; однако вклад вносят вирусные инфекции, генетическая предрасположенность, окружающая среда и аутоиммунность. Очаги повреждения содержат инфильтраты, преимущественно опосредованные T-лимфоцитами, микроглиальные клетки и инфильтрирующие макрофаги; преобладающим клеточным типом в очагах повреждения являются CD4+ T-лимфоциты. Механизм клеточной гибели олигодендроцитов и последующей демиелинизации неизвестен, но, вероятно, он управляется T-лимфоцитами.
Воспалительное и фиброзное заболевание легких, включающее в себя эозинофильные пневмонии; идиопатический легочный фиброз и гиперчувствительный пневмонит, могут включать в себя нерегулируемый иммунный воспалительный ответ. Ингибирование этого ответа может быть терапевтически полезным.
Аутоиммунное или опосредованное иммунной системой кожное заболевание, включающее в себя буллезные заболевания кожи, полиморфную эритему и контактный дерматит, опосредованы антителами, происхождение которых зависит от T-лимфоцитов.
Псориаз представляет собой опосредованное T-лимфоцитами воспалительное заболевание. Очаги повреждения содержат инфильтраты из T-лимфоцитов, макрофагов и процессирующих антиген клеток и некоторого количества нейтрофилов.
Аллергические заболевания, включающие в себя астму; аллергический ринит; атопический дерматит; гиперчувствительность к пище и крапивницу, зависят от T-лимфоцитов. Эти заболевания преимущественно опосредованы индуцированным T-лимфоцитами воспалением, опосредованным IgE воспалением или сочетанием обоих.
Связанные с трансплантацией заболевания, включающие в себя отторжение трансплантата и реакцию трансплантат-против-хозяина (GVHD), зависят от T-лимфоцитов; ингибирование функционирования T-лимфоцитов приводит к улучшению.
Другими заболеваниями, при которых вмешательство иммунного и/или воспалительного ответа может быть полезным, являются инфекционные заболевания, включающие в себя в качестве неограничивающих примеров вирусную инфекцию (включающую в себя в качестве неограничивающих примеров СПИД, гепатит A, B, C, D, E), бактериальную инфекцию, грибковые инфекции и протозойные и паразитические инфекции (молекулы (или дериваты/агонисты), которые стимулируют MLR, можно использовать терапевтически для усиления иммунного ответа на возбудители инфекции), иммунодефицитные состояния (молекулы/дериваты/агонисты), стимулирующие MLR, можно использовать терапевтически для усиления иммунного ответа на унаследованные, приобретенные, индуцированные инфекциями (как при ВИЧ-инфекции) или ятрогенные (т.е. в результате химиотерапии) состояния и неоплазию.
Антитело предпочтительно вводят млекопитающему в носителе; предпочтительно фармацевтически приемлемом носителе. Подходящие носители и их составы описаны в Remington's Pharmaceutical Sciences, 16th ed., 1980, Mack Publishing Co., под редакцией Oslo et al. Как правило, подходящее количество фармацевтически приемлемой соли применяют в составе для придания составу изотоничности. Примеры носителя включают в себя физиологический раствор, раствор Рингера и раствор декстрозы. pH раствора предпочтительно составляет от приблизительно 5 до приблизительно 8, а более предпочтительно от приблизительно 7 до приблизительно 7,5. Дополнительные носители включают в себя препараты с замедленным высвобождением, такие как полупроницаемые матриксы твердых гидрофобных полимеров, содержащие антитело, где эти матрицы находятся в форме профилированный изделий, например пленок, липосом или микрочастиц. Специалистам в данной области понятно, что определенные носители могут быть более предпочтительными, например, в зависимости от маршрута введения и концентрации вводимого антитела.
Антитело можно вводить млекопитающему посредством инъекции (например, внутривенной, интраперитонеальной, подкожной, внутримышечной, интрапортальной) или другими способами, такими как вливание, которые обеспечивают его доставку в кровоток в эффективной форме. Антитело также можно вводить способами изолированной перфузии, такими как изолированная тканевая перфузия, для оказания локального терапевтического действия. Предпочтительны локальная и внутривенная инъекции.
Эффективные дозировки и протоколы введения антитела можно определять эмпирически, и осуществление таких определений известно специалистам в данной области. Специалистам в данной области понятно, что дозировка антитела, которую необходимо вводить, варьирует в зависимости, например, от млекопитающего, получающего антитело, маршрута введения, конкретного типа используемого антитела и других лекарственных средств, вводимых млекопитающему. Руководство по выбору подходящих доз для антитела находится в литературе по терапевтическому применению антител, например Handbook of Monoclonal Antibodies, Ferrone et al., eds., Noges Publications, Park Ridge, NJ., (1985), ch. 22 and pp.303-357; Smith et al., Antibodies in Human Diagnosis and Therapy, Haber et al., eds., Raven Press, New York (1977), pp.365-389. Типичная суточная доза антитела, применяемого отдельно, может находиться в диапазоне приблизительно от 1 мкг/кг до 100 мг/кг массы тела или более в сутки, в зависимости от указанных выше факторов.
Антитело также можно вводить млекопитающему в сочетании с эффективными количествами одного или нескольких других терапевтических средств. При лечении злокачественной опухоли это особенно верно, так как многие опухоли приобретают устойчивость к химиотерапии или радиотерапии вследствие инактивации гена p53, супрессирующего опухоли. Так как DR5 стимулирует апоптоз независимо от p53, ожидают, что он будет клинически полезным не только в качестве отдельного средства, но также и в сочетании с другими типами противоракового лечения, такими как, например, химиотерапия (химиотерапевтические средства), лучевая терапия, иммуноадъюванты, ингибирующие рост средства, цитотоксические средства и/или цитокины. Также можно применять другие средства, для которых известно, что они индуцируют апоптоз в клетках млекопитающих, и такие средства включают в себя TNF-альфа, TNF-бета, лиганд CD30, лиганд 4-1BB и лиганд Apo-2, а также другие антитела, которые могут индуцировать апоптоз. Одно или несколько других лекарственных средств могут включать в себя терапевтические антитела (отличные от антитела к DR5), и такие антитела могут включать в себя антитела к рецептору Her (такие как Herceptin® (трастузумаб), Genentech, Inc.), антитела к VEGF, антитела к CD20 (такие как Rituxan® (ритуксимаб), Genentech, Inc.) и антитела к другим рецепторам лиганда Apo-2, такие как антитела к DR4, или антитела к другим представителям семейства рецептора TNF, такие как ENBREL® (этанерцепт) (Immunex).
Рассматриваемые в изобретении химиотерапевтические препараты включают в себя химические вещества или лекарственные средства, которые известны в данной области и коммерчески доступны, такие как доксорубицин, 5-фторурацил, этопозид, камптотецин, лейковорин, цитозинарабинозид, циклофосфамид, тиотепа, бусульфан, цитоксин, таксол, метотрексат, цисплатин, мелфалан, винбластин и карбоплатин. Состав и схемы дозирования для такой химиотерапии можно применять по инструкциям производителя или как определено эмпирически практикующими специалистами. Состав и схемы дозирования для такой химиотерапии также описаны в Chemotherapy Service Ed., M.C. Perry, Williams & Wilkins, Baltimore, MD (1992).
Химиотерапевтический препарат предпочтительно вводят в фармацевтически приемлемом носителе, таком как описанные выше носители. Способ введения млекопитающему химиотерапевтического препарата может быть таким же, как применяют для антитела к DR5, или его можно вводить другим способом. Например, млекопитающему антитело к DR5 можно инъецировать, тогда как химиотерапевтическое средство вводят перорально.
Радиационную терапию можно проводить млекопитающему согласно протоколам, обычно используемым в данной области и известным специалистам в данной области. Такая терапия может включать в себя радиоактивное излучение цезия, иридия, йода или кобальта. Радиационная терапия может представлять собой облучение всего тела, или ее можно применять локально к конкретному участку или ткани в организме или на теле. Как правило, радиационную терапию проводят импульсами в течение периода времени от приблизительно 1 до приблизительно 2 недель. Однако радиационную терапию можно проводить в течение более длительных периодов времени. Необязательно, радиационную терапию можно проводить в виде однократной дозы или в виде множества последовательных доз.
Антитело можно вводить последовательно или одновременно с одним или несколькими другими терапевтическими средствами. Количества антитела и терапевтического средства зависят, например, от типа применяемых лекарственных средств, подвергаемого лечению патологического состояния и схемы и путей введения, но, как правило, составляют менее, чем если их использовать по отдельности.
После введения антитела млекопитающему физиологическое состояние млекопитающего можно контролировать различными способами, хорошо известными практикующим специалистам.
Полагают, что в терапии также можно использовать антагонистические или блокирующие антитела к DR5. Например, антитело к DR5 можно вводить млекопитающему (так, как описано выше) для блокирования связывания рецептора DR5 с Apo-2L, таким образом, увеличивая биодоступность Apo-2L, вводимого при терапии Apo-2L, для индукции апоптоза у злокачественных клеток.
Терапевтические эффекты антител к DR5 по изобретению можно контролировать в анализах in vitro и с применением моделей на животных in vivo. Для дальнейшего понимания роли определенных в настоящем документе антител к DR5 в развитии и патогенезе, например, злокачественной опухоли или связанных с иммунной системой заболеваний и для тестирования эффективности терапевтических средств-кандидатов можно использовать ряд хорошо известных моделей на животных. In vivo характер таких моделей делает их особенно предсказательными в отношении ответа у пациентов, являющихся людьми.
Модели связанных с иммунной системой заболеваний на животных включают в себя нерекомбинантных и рекомбинантных (трансгенных) животных. Модели на нерекомбинантных животных включают в себя, например, грызунов, например модели на мышах. Такие модели можно получать посредством введения клеток сингенным мышам с применением стандартных способов, например подкожной инъекции, инъекции в хвостовую вену, имплантации в селезенку, интраперитонеальной имплантации и имплантации под почечную капсулу.
Модели на животных, например, для реакции трансплантат-против-хозяина известны. Реакция трансплантат-против-хозяина происходит, когда иммуносупрессированным или толерантным пациентам трансплантируют иммунокомпетентные клетки. Донорные клетки распознают антигены хозяина и отвечают на них. Ответ может варьировать от опасного для жизни тяжелого воспаления до мягкой формы диареи и потери массы. Модели реакции трансплантат-против-хозяина обеспечивают средства для оценки реактивности T-клеток в отношении антигенов MHC и второстепенных антигенов трансплантации. Подходящая процедура подробно описана в Current Protocols in Immunology, unit 4.3.
Модель на животном для отторжения кожного трансплантата представляет собой средство для тестирования T-клеток на способность опосредовать разрушение ткани in vivo, что является показателем и мерой их роли в противовирусном и противоопухолевом иммунитете. В самых распространенных и признанных моделях используют трансплантаты кожи хвоста мышей. Повторные эксперименты показали, что отторжение аллотрансплантатов кожи опосредуется T-клетками, хелперными T-клетками и киллерными-эффекторными T-клетками, а не антителами [Auchincloss, H. Jr. and Sachs, D. H., Fundamental Immunology, 2nd ed., W. E. Paul ed., Raven Press, NY, 1989, 889-992]. Подходящая процедура подробно описана в Current Protocols in Immunology, unit 4.4. Другие модели отторжения трансплантата, которые можно использовать для тестирования композиций по изобретению, представляют собой модели аллогенной трансплантации сердца, описанные в Tanabe, M. et al., Transplantation, (1994) 58:23, и Tinubu, S. A. et al., J. Immunol. (1994) 4330-4338.
Модели на животных для гиперчувствительности замедленного типа также обеспечивают анализ опосредованных клетками функций иммунной системы. Реакции гиперчувствительности замедленного типа представляют собой опосредованный T-клетками иммунный ответ in vivo, характеризующийся воспалением, которое не достигает пика до окончания промежутка времени, прошедшего после стимуляции антигеном. Эти реакции также происходят при тканеспецифических аутоиммунных заболеваниях, таких как рассеянный склероз (РС) и экспериментальный аутоиммунный энцефаломиелит (EAE, модель для РС). Подходящая процедура подробно описана в Current Protocols in Immunology, unit 4.5.
Модель на животных для артрита представляет собой индуцированный коллагеном артрит. Эта модель обладает клиническими, гистологическими и иммунологическими характеристиками аутоиммунного ревматоидного артрита человека и представляет собой приемлемую модель для аутоиммунного артрита человека. Мышиные и крысиные модели характеризуются синовиитом, эрозией хряща и подхрящевой кости. Антитела к DR5 по изобретению можно тестировать на активность против аутоиммунного артрита с применением протоколов, описанных в Current Protocols in Immunology, выше, units 15.5. Также см. модель с применением моноклонального антитела к интегринам CD18 и VLA-4, описанную в Issekutz, A. C. et al., Immunology, (1996) 88:569.
Описана модель для астмы, в которой индуцированную антигеном гиперреактивность бронхов, легочную эозинофилию и воспаление индуцируют посредством сенсибилизации животного овальбумином, а затем стимулируя животного тем же белком, вводимым посредством аэрозоля. Некоторые модели на животных (морская свинка, крыса, не являющийся человеком примат) демонстрируют симптомы, сходные с атопической астмой у людей при стимуляции аэрозольными антигенами. Модели на мышах обладают многими свойствами астмы человека. Подходящие процедуры для тестирования композиций по изобретению на активность и эффективность лечения астмы описаны в Wolyniec, W. W. et al., Am. J. Respir. Cell Mol. Biol. (1998) 18:777, и, цитируемых в ней ссылках.
Кроме того, антитела к DR5 по изобретению можно тестировать в моделях на животных для заболеваний, подобных псориазу. Антитела к DR5 по изобретению можно тестировать на мышиной модели scid/scid, описанной в Schon, M. P. et al., Nat. Med. (1997) 3:183, где мыши демонстрируют гистопатологические очаги повреждения кожи, сходные с псориазом. Другую подходящую модель представляет собой химерная мышь Scid/кожа человека, полученная, как описано у Nickoloff, B. J. et al., Am. J. Path. (1995) 146:580.
Для тестирования безопасности и противораковой активности терапевтической композиции-кандидата хорошо известны различные модели на животных. Они включают в себя ксенотрансплантацию опухоли человека бестимусным мышам nude или мышам scid/scid или генетические модели опухолей у мышей, такие как мыши с нокаутом p53.
Рекомбинантные (трансгенные) модели на животных можно конструировать посредством введения кодирующей части определяемых в настоящем документе молекул в геном представляющих интерес животных с применением стандартных способов получения трансгенных животных. Животные, которые могут служить в качестве мишени для трансгенных манипуляций, включают в себя в качестве неограничивающих примеров мышей, крыс, кроликов, морских свинок, овец, коз, свиней и не являющихся человеком приматов, например бабуинов, шимпанзе и мартышек, таких как яванская макака. Известные в данной области способы введения трансгена такому животному включают в себя микроинъекцию в пронуклеус (Hoppe and Wanger, патент США №4873191); опосредуемый ретровирусами перенос генов в зародышевые линии (например, Van der Putten et al., Proc. Natl. Acad. Sci. USA, 82, 6148-615 [1985]); воздействие на гены в эмбриональных стволовых клетках (Thompson et al., Cell, 56, 313-321 [1989]); электропорацию эмбрионов (Lo, Mol. Cel. Biol., 3, 1803-1814 [1983]); опосредованный спермием перенос гена (Lavitrano et al., Cell, 57, 717-73 [1989]). Для обзора см., например, патент США №4736866.
Для целей настоящего изобретения трансгенные животные включают в себя животных, которые несут трансген только в части своих клеток ("мозаичные животные"). Трансген может быть интегрирован в виде одного трансгена или в конкатемеры, например в тандемы "голова-к-голове" или "голова-к-хвосту". Селективное введение трансгена в конкретный тип клеток также возможно, например, способом Lasko et al., Proc. Natl. Acad. Sci. USA, 89, 6232-636 (1992).
Экспрессию трансгена у трансгенных животных можно контролировать стандартными способами. Например, для подтверждения интеграции трансгена можно использовать анализ "саузерн"-блот или амплификацию ПЦР. Затем можно анализировать уровень экспрессии мРНК с применением таких способов, как гибридизация in situ, анализ "нозерн"-блот, ПЦР или иммуногистохимия. Животных можно дополнительно проверять на патологические признаки иммунного заболевания, например, посредством гистологической проверки для определения инфильтрации иммунных клеток в конкретную ткань или на присутствие раковой или злокачественной ткани.
Альтернативно, можно получать животное с "нокаутом", которое несет дефектный или измененный ген, кодирующий полипептид, определяемый в настоящем документе, как результат гомологичной рекомбинации между эндогенным геном, кодирующим полипептид, и измененной геномной ДНК, кодирующей тот же пептид, введенный в эмбриональную клетку животного. Например, кодирующую конкретный полипептид кДНК можно применять для клонирования кодирующей этот полипептид геномной ДНК в соответствии с широко известными способами. Часть кодирующей конкретный полипептид геномной ДНК можно удалить или заменить другим геном, таким как ген, кодирующий селективный маркер, который можно применять для контроля интеграции. Как правило, в вектор включают несколько тысяч оснований неизмененной фланкирующей ДНК (с 5'- и 3'-концов) [для описания векторов для гомологичной рекомбинации см., например, Thomas and Capecchi, Cell, 51:503 (1987)]. Вектор вводят в линию эмбриональных стволовых клеток (например, посредством электропорации) и отбирают клетки, в которых введенная ДНК подверглась гомологичной рекомбинации с эндогенной ДНК [см. например, Li et al., Cell, 69:915 (1992)]. Затем отобранные клетки инъецируют в бластоцисту животного (например, мыши или крысы) с образованием агрегационных химер (см., например, Bradley, in Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson, ed. (IRL, Oxford, 1987), стр. 113-152]. Затем химерный эмбрион можно имплантировать в подходящую псевдобеременную приемную самку животного и доводить эмбрион до нормального срока окончания беременности с получением животного с "нокаутом". Потомство, несущее в своих зародышевых клетках ДНК, подвергшуюся гомологичной рекомбинации, можно идентифицировать стандартными способами и применять для выведения животных, у которых все клетки животного содержат гомологично рекомбинированную ДНК. Животных с нокаутом можно отличать по их способности противостоять определенным патологическим состояниям и по развитию у них патологических состояний вследствие отсутствия полипептида.
В другом варианте осуществления изобретения предоставлены способы для применения антитела в диагностических анализах. Например, антитела можно применять в диагностических анализах для детекции экспрессии или сверхэкспрессии DR5 в конкретных клетках и тканях. Можно использовать различные известные в данной области способы диагностического анализа, такие как анализ изображений in vivo, анализ конкурентного связывания in vitro, прямой и непрямой сэндвич-анализ и анализ иммунопреципитации, проводимый в гетерогенной или гомогенной фазах [Zola, Monoclonal Antibodies: A Manual of Techniques, CRC Press, Inc. (1987), pp.147-158]. Применяемые в диагностических анализах антитела можно метить детектируемой группой. Детектируемая группа может быть способной к прямой или непрямой продукции детектируемого сигнала. Например, детектируемая группа может представлять собой радиоактивный изотоп, такой как 3H, 14C, 32P, 35S или 125I, флуоресцентное или хемилюминесцентное соединение, такое как флуоресцеинизотиоцианат, родамин или люциферин, или фермент, такой как щелочная фосфатаза, бета-галактозидаза или пероксидаза хрена. Можно применять любой известный в данной области способ конъюгации антитела с детектируемой группой, включая способы, описанные в Hunter et al., Nature, 144:945 (1962); David et al., Biochemistry, 13:1014-1021 (1974); Pain et al., J. Immunol. Meth., 40:219-230 (1981), и Nygren, J. Histochem. and Cytochem., 30:407-412 (1982).
Антитела к DR5 также пригодны для аффинной очистки DR5 из культуры рекомбинантных клеток или природных источников. В этом способе антитела к DR5 иммобилизуют на подходящей подложке, такой как смола сефадекс или фильтровальная бумага, с применением хорошо известных в данной области способов. Затем иммобилизованное антитело приводят в контакт с образцом, содержащим DR5 для очистки, а затем подложку промывают подходящим растворителем, который по существу удаляет все вещество в образце, за исключением DR5, который связывается с иммобилизованным антителом. В конце подложку промывают другим подходящим растворителем, который высвобождает DR5 из антитела.
В дополнительном варианте осуществления изобретения предоставлены промышленные изделия и наборы, содержащие вещества, пригодные для лечения патологических состояний или детекции или очистки DR5. Промышленное изделие содержит контейнер с этикеткой. Подходящие контейнеры включают, например, пызырьки, флаконы и тестовые пробирки. Контейнеры можно производить из ряда материалов, таких как стекло или пластик. Контейнер содержит композицию с активным веществом, эффективным для лечения патологических состояний или для детекции или очистки DR5. Активное вещество в композиции представляет собой антитело к DR5 и предпочтительно содержит моноклональные антитела, специфичные к DR5. Этикетка на контейнере указывает, что композицию применяют для лечения патологических состояний или детекции или очистки DR5, и может также указывать способы применения in vivo или in vitro, такие как описанные выше способы.
Набор по изобретению содержит описанный выше контейнер и второй контейнер, содержащий буфер. Он может дополнительно содержать другие материалы, желательные с коммерческой и пользовательской точек зрения, включая другие буферы, разбавители, фильтры, иглы, шприцы и вкладыши в упаковку с инструкциями по применению.
Следующие примеры предложены только в целях иллюстрации и никаким образом не предназначены для ограничения объема настоящего изобретения.
Все патенты и литературные ссылки, цитируемые в настоящем описании, таким образом, включены в виде ссылки в полном объеме.
ПРИМЕРЫ
Если не указано иначе, указываемые в примерах коммерчески доступные реагенты применяли по инструкциям производителя. Источником клеток, определяемых в следующих ниже примерах и на всем протяжении спецификации посредством инвентарных номеров ATCC, является American Type Culture Collection, Manassas, Virginia. Ряд реагентов и протоколов, описанных в настоящем документе, дополнительно описаны в WO 99/37684, WO 00/73349, WO 98/32856, WO 98/51793 и WO 99/64461, содержание которых, таким образом, включено в виде ссылки в полном объеме.
Пример 1
Конструирование и тестирование вариантов антител к DR5
Антитело к DR5 16E2 получено в форме scFv из библиотеки фагового дисплея антител человека и описано в WO 98/51793, опубликованной 19 ноября 1998 года (см. пример 14). Нуклеотидная и аминокислотная последовательности 16E2 scFv показаны на фиг.5 (SEQ ID NO: 9) и фиг.6 (SEQ ID NO: 10), соответственно. На фиг.6 указана сигнальная последовательность и области CDR тяжелой и легкой цепи (области CDR1, CDR2 и CDR3 подчеркнуты).
Материалы и методы
Конструирование полноразмерного антитела к DR5 16E2
Для описанных ниже экспериментов конструировали полноразмерные IgG. Таким образом, вариабельные домены 16E2 клонировали в ранее описанные векторы pRK, подходящие для экспрессии полноразмерных антител IgG1 в клетках млекопитающих (Gorman et al., DNA Prot. Eng. Tech. 2:3-10 (1990)). Сравнение аминокислотной последовательности вариабельного домена 16E2 с базой данных Kabat (Kabat et al., Sequences of Proteins of Immunological Interest, U.S. Dept. of Health and Human Services, NIH, 5th edition) показало, что вариабельная область легкой цепи (VL) 16E2 получена из семейства генов легкой цепи лямбда человека. Таким образом, вариабельный домен 16E2 первый субклонировали в вектор, содержащий константный домен лямбда. Сконструировали праймеры ПЦР для добавления участков распознавания рестрикционных ферментов SfiI и MscI, а затем амплифицированный вариабельный домен расщепили этими двумя ферментами. Этот фрагмент вставили в подобным образом расщепленный вектор, содержащий константный домен лямбда. Так как этот вектор конструировали для экспрессии Fab в E.coli, для экспрессии IgG область, кодирующую целую легкую цепь, снова подвергали амплификации ПЦР с применением праймеров для добавления участка рестрикции AgeI на 5'-конце кодирующей области и HindIII на 3'-конце. Затем этот фрагмент от AgeI до HindIII вставляли в подобным образом расщепленный вектор, pDR1 (Clontech). Полная последовательность плазмиды pDR1 показана на фиг.11 (SEQ ID NO: 15).
Для тяжелой цепи версии 1 вариабельный домен (VH) тяжелой цепи scFv 16E2 подвергали амплификации ПЦР с применением праймеров, сконструированных для добавления участка PvuII на 5'-конце и участка ApaI на 3'-конце домена. Этот фрагмент клонировали по участкам PvuII/ApaI вектора pDR2 (Clontech) для экспрессии целой тяжелой цепи (домены VH-CH1-CH2-CH3). Полная последовательность плазмиды pDR2 показана на фиг.12 (SEQ ID NO: 16).
Нуклеотидная и аминокислотная последовательности тяжелой и легкой цепей полноразмерного антитела 16E2 показаны на фиг.7-10 (SEQ ID NO: 11-14), соответственно, в частности, на фиг.7 и 8 (SEQ ID NO: 11 и 12) показаны аминокислотная и нуклеотидная последовательности полноразмерной тяжелой цепи 16E2, а на фиг.9 и 10 (SEQ ID NO: 13 и 14) показаны аминокислотная и нуклеотидная последовательности полноразмерной легкой цепи 16E2. Тяжелая и легкая цепи полноразмерного антитела 16E2 далее в настоящем документе также обозначают как "Версия 1."
Конструирование вариантов IgG. Варианты конструировали из легкой и тяжелой цепей отдельно с применением сайт-специфического мутагенеза (Kunkel et al., Proc. Natl. Acad. Sci USA 82:488-492 (1985)). Плазмиду pDR1, кодирующую легкую цепь версии 1, или pDR2, кодирующую тяжелую цепь версии 1, трансформировали в штамм E.coli CJ236 (BioRad, Joyce and Grindley, J. Bacteriol. 158:636-643 (1984)) для получения дезоксирибонуклеотидсодержащих одноцепочечных матриц ДНК. Аликвоты реакционных смесей мутагенеза трансформировали в штамм E.coli XL-I Blue (Stratagene, San Diego, CA) для очистки двухцепочечных ДНК. Для каждого варианта ДНК, кодирующие легкую или тяжелую цепь, полностью секвенировали с применением автоматического секвенатора ДНК ABI377x1 или ABI3730x1 (Perkin-Elmer Corp.).
Для каждого варианта IgG проводили транзиторную трансфекцию посредством котрансфекции экспрессирующей легкую цепь плазмиды с экспрессирующей тяжелую цепь плазмидой в трансформированную аденовирусом клеточную линию 293 эмбриональной почки человека (Graham et al., J. Gen. Virol, 36:59-74, (1977)). В кратком изложении, клетки 293 за день до трансфекции разделяли и высевали в содержащую сыворотку среду. На следующий день из двухцепочечной ДНК легкой или тяжелой цепей получали кальций-фосфатный преципитат вместе с pAdVantage™DNA (Promega, Madison, WI) и капельно добавляли в планшеты. Клетки инкубировали в течение ночи при 37°C, затем отмывали PBS и культивировали в среде без сыворотки в течение 4 суток, после чего собирали кондиционированную среду. Антитела очищали из супернатантов культуры с применением белка A-сефарозы CL-4B, затем буфер заменяли на 10 мМ сукцинат натрия, 140 мМ NaCl, pH 6,0, и концентрировали с применением Centricon-10 (Amicon). Концентрации белков определяли посредством измерения поглощения при 280 нм или посредством количественного аминокислотного анализа.
Электрохемилюминесцентный анализ связывания DR5. Относительное связывание антител к DR5 определяли в фазе раствора в формате конкуретного ELISA. Слитный белок DR5-Fc биотинилировали с применением биотина-X-NHS (Research Organics, Cleveland, OH), а стандартное антитело (версии 1 или Apomab 7.3) метили сложным эфиром ORI-TAG NHS (IGEN International, Gaithersburg, MD) по указаниям производителя. Для проведения анализа связывания образцы тестируемого антитела серийно разводили в буфере для анализа (PBS, pH 7,4, содержащий 0,5% BSA и 0,5% Tween-20). Равные объемы (каждый по 25 мкл) образца антитела (концентрации в диапазоне от 50000 до 0,85 нг/мл), стандартного антитела с ORI-TAG (150 нг/мл) и биотинилированного DR5 человека-Fc (15 нг/мл) добавляли в 96-луночные полипропиленовые планшеты и инкубировали в течение 1,5 ч при комнатной температуре со слабым перемешиванием. Затем добавляли магнитные бусы со стрептавидином (IGEN International) (25 мкл/лунку), а затем планшеты инкубировали, как указано выше, в течение дополнительных 30 минут. Добавляли буфер для анализа для доведения конечного объема до 250 мкл на лунку и планшеты считывали с применением устройства ORIGEN M384 (IGEN International). Подсчитывали значения IC50 с применением четырех параметров, соответствующих выборочным кривым.
Биологический анализ: ингибирование роста/уничтожение опухолевых клеток. Истинную активность каждого варианта антитела определяли в анализе уничтожения опухолевых клеток in vitro. Клеточную линию карциномы толстой кишки человека Colo205 культивировали в среде RPMI, содержащей 10% эмбриональной телячьей сыворотки. В 96-луночных планшетах для тканевых культур, содержащих среду со сшивающим антителом (аффинно очищенные козьи F(ab')2 к Fc человека) в концентрации 10 микрограммов/мл или без него, проводили двукратные серийные разведения стандарта (или версия 1, или Apomab 7.3) и образцов. Затем в планшеты добавляли клетки (20000/лунка). Планшеты инкубировали при 37°C всего в течение 48 ч. В течение последних 3 часов инкубации к лункам добавляли AlamarBlue. Показатели флуоресценции считывали с применением флуорометра с возбуждением при 530 нм и испусканием 590 нм. Данные анализировали с применением программы обработки кривых с четырьмя параметрами.
Результаты
Общей целью данной работы являлось сконструировать антитела к DR5 с улучшенными биологическими свойствами и увеличенной эффективностью без ухудшения безопасности.
Для достижения желательных улучшений использовали некоторые подходы, включая замены аминокислот в тяжелой и легкой цепях DR5 для увеличения химической и термической стабильности; фолдинга; сканирование аланином остатков CDR для определения того, какие остатки могут быть важными для связывания или фолдинга и, таким образом, могут быть заменены для увеличения аффинности; и применение библиотек фагового дисплея CDR для идентификации клонов с увеличенной активностью. Также изучали изменения в каркасных областях на их положительное влияние на иммуногенность и биологическую активность.
Модели гомологии (фиг.19 и 20) использовали в качестве помощи для селекции множества из этих изменений.
Варианты тяжелой цепи
Ряд 1
Версия 2 тяжелой цепи несет 5 замен в версии 1. Эти изменения (Q6E, V11L, E12V, R13Q и K105Q) находятся в каркасной области вариабельного домена и добавлены для того, чтобы сделать каркас ближе к консенсусной последовательности VHIII человека. В таблице 1 показаны первые варианты, сконструированные с заменами в CDR тяжелой цепи. Матрицей оцДНК для этих мутантов была версия 2. Замену Leu на Tyr в положении 102 проводили для улучшения укладки и, таким образом, стабильности. В сочетании с этой заменой Asn53 заменяли на Gln или Tyr для удаления потенциального участка дезамидирования. Met34 заменяли на Leu для удаления потенциального участка окисления. Эти варианты тяжелой цепи экспрессировали с исходной легкой цепью с получением вариантов 20-23 (таблица 1). Тяжелая цепь, содержащая три мутации M34L, N53Q и L102Y, и замены в каркасе, как в версии 2, обозначается в дальнейшем как "тяжелая цепь с тройной заменой один" или TH1, тогда как тяжелая цепь с тремя мутациями M34L, N53Y и L102Y и каркасом из версии 2 подобным образом обозначается TH2.
b Анализ конкурентного связывания DR5 человека Origen®
c Анализ ингибирования опухолевых клеток
d Анализ ингибирования опухолевых клеток
Так как при проведении мутации M34L (т.е. v21 по сравнению с v20, v23 по сравнению с v22, таблица 1) происходила потеря связывания и клеточной киллерной активности in vitro, в CDR1 тяжелой цепи с применением в качестве матрицы тяжелой цепи версии 20 проведен ряд дополнительных мутаций, замещающих или аланин, или другие остатки, предположение о которых сделано при сканировании базы данных Kabat. Эти тяжелые цепи экспрессировали с легкой цепью версии 1. Подвергавшиеся мутации аминокислоты, получаемое в результате связывание относительно v1 и данные биологического анализа для этих версий приведены в таблице 2. Замена Gly33 на Ala приводила к усилению связывания, а также к увеличенной эффективности. Эту тяжелую цепь, содержащую три мутации, G33A, N53Q и L102Y, обозначили TH3. Подобным образом сконструирована TH4, содержащая G33A, N53Y и L102Y. Замена Thr28 на Ala также приводила к увеличенной активности по сравнению с v1, и тяжелую цепь, содержащую T28A, N53Q, L102Y, обозначили TH9. Замены в CDR в TH1, TH2, TH3, TH4 и TH9 суммированы в таблице 5. Эти пять тяжелых цепей изучали далее после коэкспрессии их с сочетаниями легких цепей (см. ниже).
b Анализ конкурентного связывания DR5 человека Origen®
c Анализ ингибирования опухолевых клеток
d Анализ ингибирования опухолевых клеток
Ala-сканирование CDRH2 и CDRH3. Для дальнейшего выяснения вклада каждой аминокислоты из CDR2 и CDR3 тяжелой цепи получали мутанты с заменой каждого из этих остатков аланином. С применением тяжелой цепи версии 1 в качестве матрицы проводили сайт-специфический мутагенез с синтетическими олигонуклеотидами, кодирующими замену аланином. Эти тяжелые цепи экспрессировали с применением легкой цепи версии 1. Изменения наблюдаемой аффинности и эффективности полученных в результате антител приведены в таблицах 3 и 4. Каждая из Gly99Ala и Arg100Ala приводила к увеличенной активности, и, следовательно, эти замены можно применять для конструирования дополнительных комбинаторных мутантов тяжелой цепи.
b Анализ конкурентного связывания DR5 человека Origen®
c Анализ ингибирования опухолевых клеток
d Анализ ингибирования опухолевых клеток
b Анализ конкурентного связывания DR5 человека Origen®
c Анализ ингибирования опухолевых клеток
d Анализ ингибирования опухолевых клеток
Ряд 2
С применением тех же CDR, что и в версиях TH1, TH2, TH3 и TH4, в которых каркасные остатки E6, L11, V12, Q13 и Q105 заменяли на аминокислоты, присутствующие в версии 1, т.е. Q6, V1l, E12, R13 и K105, конструировали второй ряд тяжелых цепей. Эти тяжелые цепи обозначили как TH5, TH6, TH7 и TH8 (см. таблицу 5).
Варианты легких цепей
Сканирование аланином CDR легких цепей
Для лучшего понимания вклада остатков CDR легкой цепи в связывание и биологическую активность каждую аминокислоту с применением сайт-специфического мутагенеза заменяли на аланин. Каждый из вариантов легкой цепи объединяли с тяжелой цепью v1 для транзиторной экспрессии IgG, как описано выше. Полученные при сканировании CDR легких цепей аланином результаты подытожены в таблице 6. Представляет интерес, что в отличие от многих других антител, создается впечатление, что CDR L1 играет важную роль в связывании антигена. Эта легкая цепь представляет собой цепь лямбда, и модель, представленная на фиг.20, позволяет предположить, что CDR1 может формировать альфа-спираль. Замены в L2 и L3 на аланин более допустимы, за исключением G50A в CDR2, разрушающей связывание. Напротив, некоторые замены на ala, особенно R91A и K51A, усиливали связывание и биологическую активность.
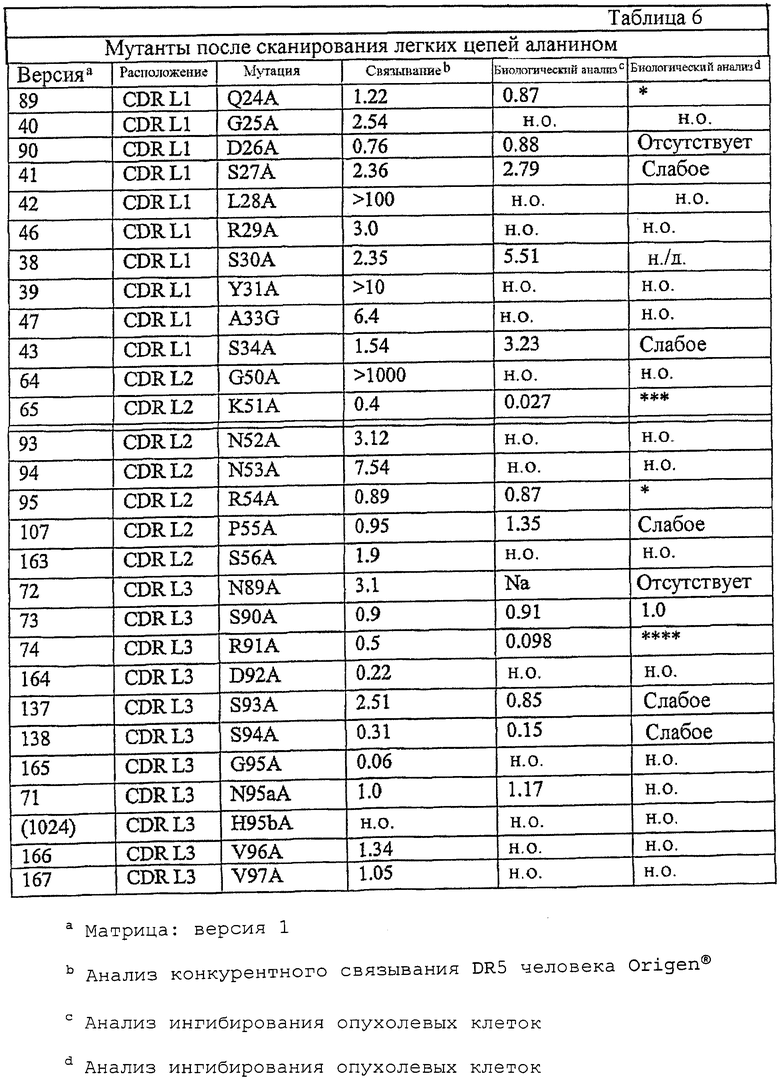
Замены остатков из семейства генов
Второй тип прямых мутантов использовали для исследования легкой цепи. При этом подходе остатки CDR, которые в модели находились с одной из сторон петель и, таким образом, могут играть поддерживающую роль, а не быть напрямую вовлеченными в связывание антигена, заменяли на остатки в соответствующих положениях в других близкородственных семействах лямбда-генов. Эти мутанты также экспрессировали с использованием тяжелой цепи v1, а результаты суммированы в таблице 7. В CDR L2 сочетания G50K, K51D приводили к значимому увеличению связывания и биологической активности, а сочетание, содержащее четыре мутации G50K, K51D, N52S, N53E, также являлось улучшенным по сравнению с v1. Другие более консервативные замены в этой же области, включающие только одну замену основания, были недопустимы.
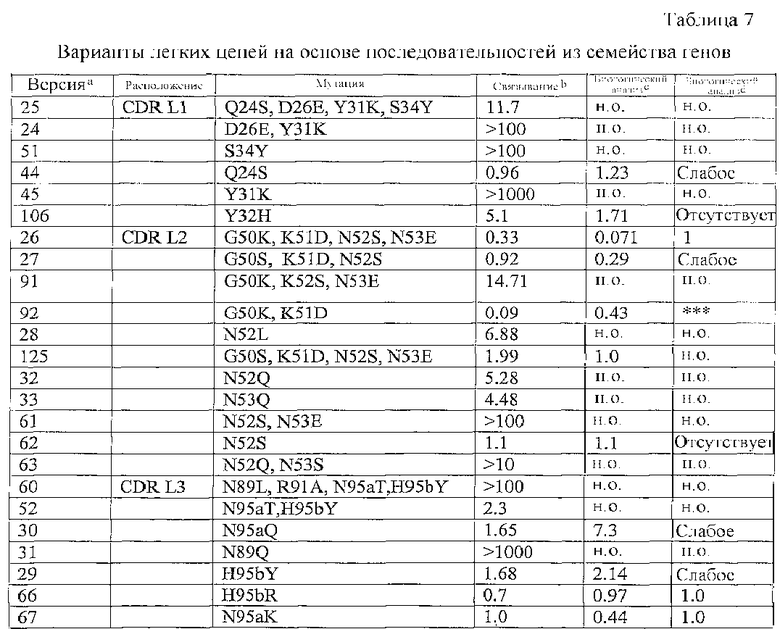
a Матрица: версия 1
b Анализ конкурентного связывания DR5 человека Origen®
c Анализ ингибирования опухолевых клеток
d Анализ ингибирования опухолевых клеток
Аффинный отбор в фаговых библиотеках антител
Для каждой из CDR L1, L2 и L3 отдельно сконструированы библиотеки фагового дисплея и проводился отбор на клоны с увеличенной аффинностью к DR5-Ig. Исследование модели (фиг.20) указало, какие остатки CDR, вероятно, были экспонированы, и эти остатки выбирали для рандомизации. Целый домен легкой цепи лямбда и домен VH версии 1 клонировали в вектор фагового дисплея pS1602, указанный в Vajdos et al., J. Mol. Biol. 320:415-428 (2002), и дополнительно описанный в Sidhu et al., Curr. Opin. Biotechnol. 11:610-616 (2002). Для конструирования библиотек использовали мутагенез по Kunkel. Для получения одноцепочечных ДНК фагмиду v1 трансформировали в штамм E.coli CJ236 и для получения библиотечных матриц использовали олигонуклеотиды, содержащие кодоны TAA в каждом участке, выбранном для рандомизации. Затем для получения библиотек использовали олигонуклеотиды с вырожденным кодоном NNS (где N представляет собой смесь одинакового количества G, A, T и C, а S представляет собой смесь одинакового количества G и C). CDRL1 подвергали мутированию с использованием олигонуклеотида стоп-матрицы CA945 и библиотечного олигонуклеотида CA946. CDRL2 подвергали мутированию с использованием олигонуклеотида стоп-матрицы CA947 и библиотечного олигонуклеотида CA948. CDRL3 подвергали мутированию с использованием олигонуклеотида стоп-матрицы CA949 и библиотечного олигонуклеотида CA950. Конструирование библиотеки суммировано в таблице 8.
Продукты реакций случайного мутагенеза посредством электропорации вводили в клетки E.coli XLI-Blue (Stratagene) и амплифицировали посредством роста в течение 14-16 часов с фагом-помощником M13K07. Размер библиотеки оценивали посредством серийного разведения и посева на чашках, если исходную трансформацию проводили на чашках с карбенициллином, и он составлял от 1,9×109 до 2,2×109 клонов.
Библиотеки подвергали пэннингу в течение 4 циклов в растворе с применением биотинилированных DR5-Ig (Genentech). Приблизительно 1011 фагов блокировали в одном мл раствора 3% обезжиренного сухого молока, 0,2% Tween в PBS (блокирующий фаг раствор) на карусельном столе в течение 1 часа при RT. Для уменьшения неспецифического связывания к блокирующему раствору добавляли DR4-IG и CD4-IG в концентрации 1 мкМ. Затем в концентрации 100 нМ для первого цикла добавляли биотинилированный антиген и связыванию позволяли происходить в течение 2 часов. В последующих циклах пэннинга концентрацию антигена снижали до 10, 5 и 1 нМ.
Для захвата антигенсвязывающего фага покрытые стрептавидином магнитные гранулы (Dynal) сначала три раза отмывали блокирующим фаг раствором, а затем блокировали в одном мл блокирующего фаг раствора в течение 1 часа при КТ. Гранулы концентрировали с применением магнита и добавляли в раствор антиген-фаг в течение 15 минут. Затем магнит использовали для извлечения комплексов гранулы-антиген-фаг из раствора. Затем эти частицы 3 раза отмывали в блокирующем фаг растворе, 3 раза в PBS-Tween (0,02% Tween) и 1 раз в PBS. Фаги элюировали с гранул 100 микролитрами 0,1 M HCl в течение 10 мин и нейтрализовали с применением NaOH. Элюированные фаги использовали для инфицирования E.coli XL1Blue и выращивали, как указано выше, для последующих циклов. В каждом цикле увеличивали строгость.
Клоны из каждой библиотеки секвенировали. Для библиотек L1 получали последовательности только дикого типа, подтверждая идею о том, что эта CDR важна для связывания антигена или конформации антитела и допустимо только небольшое количество мутаций в потенциальной спирали. Для библиотек в L2 консенсусных последовательностей выявлено не было, что позволяет предположить, что для связывания с DR5 приемлемы несколько последовательностей CDRL2. Для библиотек L3 некоторые последовательности встречались несколько раз, и эти последовательности вставили в вектор с полноразмерной легкой цепью с применением сайт-специфического мутагенеза. Экспрессию IgG снова проводили в клетках 293 с применением для котрансфекции тяжелой цепи версии 1. В таблице 9 описаны последовательности L3, экспрессируемые в качестве полноразмерных антител, и результаты связывания и биологических анализов. Две из тестируемых последовательностей, вставленные в версии 69 и 70, приводили к антителам с улучшенными по сравнению с версией 1 связыванием и биологической активностью.
CDR L3


b Анализ конкурентного связывания DR5 человека Origen®
c Анализ ингибирования опухолевых клеток
d Анализ ингибирования опухолевых клеток
Комбинаторные легкие цепи
Как описано выше, в каждой CDR легкой цепи идентифицированы мутации, которые по отдельности увеличивают связывание и гибель опухолевых клеток in vitro. Затем эти мутации комбинировали с получением нескольких легких цепей с улучшениями в каждой из CDR. Их обозначили TL1, TL2 и TL3, т.е. "легкая цепь с тройной заменой 1" и т.д., и они описаны в таблице 10. Таким образом, TL1 содержит мутантную L1, идентифицированную в версии 44, в сочетании с мутантной L2, идентифицированной в версии 26, и мутантной L3 из версии 29. Подобным образом, TL2 содержит мутантную L1 из версии 44 с мутантной L2 из v65 и мутантной L3 из v69, тогда как TL3 сочетает L1 из v44 с L2 из v65 и L3 из v74.
CDR L1
CDR L2
CDR L3
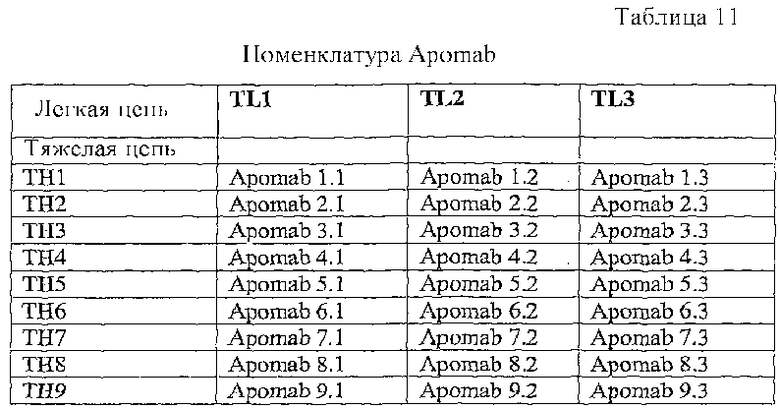
Кроме того, антитела Apomab приведены в таблице 12.
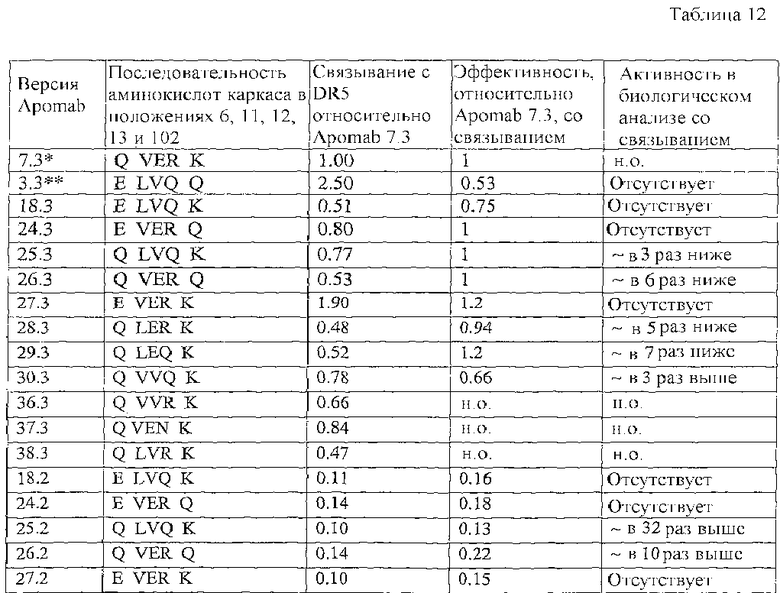
*такой же каркас, как в версии 1
**такой же каркас, как в консенсусной VH-III человека
Экспрессия Apomab
После получения тяжелых цепей с тройной заменой и легких цепей с тройной заменой получили сеть 9×3 сочетанных антител посредством котрансфекции каждой из трех легких цепей с каждой из девяти тяжелых цепей в клетки 293 и очистки полученных антител, как описано выше. Исследования in vitro этих Apomab показали, что определенные версии были достаточно эффективны в биологических анализах. Таким образом, получили материал, который подходил для исследований в моделях опухоли на мышах in vivo.
Восстановление и очистка Apomab
Описан способ восстановления и очистки антител Apomab из жидкости, собранной из клеточной культуры (HCCF), как указано ниже.
Хроматография на Prosep Protein A - жидкость, собранную из клеточной культуры (HCCF), полученную из клеток яичника китайского хомяка (CHO), доводили до pH 7,0 с применением основания 1,5 M Tris, а затем наносили на колонку Prosep Protein A (Millipore, U.S.A.), уравновешенную 25 мМ NaCl/25 мМ Tris/5 мМ EDTA, pH 7,5. Несвязавшиеся белки проходили через колонку и удалялись отмывкой уравновешивающим буфером с последующей второй стадией отмывки с применением 0,5 M TMAC в уравновешивающем буфере и третьей отмывкой с применением уравновешивающего буфера. Антитело Apomab элюируют из колонки с белком A с применением этапа элюции 0,1 M уксусной кислотой. Элюент колонки контролируют при A280. Пик антитела Apomab объединяют.
Хроматография с применением SP-Sepharose Fast Flow - объединенный объем антитела Apomab со стадии Prosep Protein A доводят до pH 5,5 с применением основания 1,5 M Tris, а затем помещают на колонку с SP-Sepharose Fast Flow (Amersham Pharmacia, Sweden), уравновешенную 25 мМ MOPS, pH 7,1. После сбора образцов колонку промывают уравновешивающим буфером до исходного уровня при A280. Антитело Apomab элюируют из колонки с применением линейного градиента от 0 до 0,2 M хлорида натрия в уравновешивающем буфере, pH 8, объемом 12 колонок. Элюент колонки контролируют при A280. Фракции собирают, и те из них, которые содержат правильно уложенное антитело Apomab, как определяют посредством анализа SEC-HPLC, объединяют.
Хроматография с применением Q-Sephrose Fast Flow - затем объединенный объем фракций после SP-Sepharose наносят на колонку с Q-Sepharose FF (Amersham pharmacia, Sweden), уравновешенную 50 мМ хлорида натрия/25 мМ буфером Tris, pH 8. Колонку промывают уравновешивающим буфером, а антитело Apomab собирают в стоке колонки.
Состав UF/DF - объединенный объем после Q-Sepharose концентрируют ультрафильтрацией на мембране с отделением молекулярной массы приблизительно от 10000 дальтон. Затем концентрированный объединенный объем после Q-Sepharose FF подвергают диафильтрации 10 объемами 10 мМ гистидина/8% сахарозы, pH 6. Подвергнутый диафильтрации объединенный объем после Q-Sepharose кондиционируют 10% полисорбатом 20 до достижения конечной концентрации полисорбата 20 0,02%. Полученный объем фильтруют через стерильный 0,22 мкм фильтр и хранят при 2-8°C или -70°C. Конечную чистоту антител Apomab определяют посредством SDS-PAGE, SEC-HPLC и анализа аминокислотной последовательности.
Секвенирование проводят на устройствах для определения последовательности ABI3700 или ABI3730 Applied Biosystems. Хроматограммы секвенирования анализируют посредством программного обеспечения по анализу последовательностей Sequencer (GeneCodes, Ann Arbor, MI).
Тестирование активности Apomab in vitro
Перед началом исследований in vivo с применением Apomab, каждую серию тестировали на активность in vitro, как описано выше. Эти результаты приведены в таблице 13.
ПРИМЕР 2
Оценка противоопухолевой активности Apomab в модели карциномы толстой кишки человека на ксенотрансплантате Colo205 и других моделях колоректального рака на ксенотрансплантатах
Часто используемые в этом и последующих примерах сокращения представляют собой следующее:
Логарифм уничтожения клеток=log10
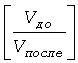 разница между логарифмом фактического объема опухоли на момент лечения, log10(V
до) и log10(V
после) (Chenevert et al., Clin. Cancer Res. 3:1457-1466 (1997)
разница между логарифмом фактического объема опухоли на момент лечения, log10(V
до) и log10(V
после) (Chenevert et al., Clin. Cancer Res. 3:1457-1466 (1997)
Бестимусным мышам "nude" в возрасте 6-8 недель (Charles River Laboratories) инокулировали 5 миллионов клеток Colo205 на мышь в объеме 0,2 мл на мышь, подкожно, сзади в область правого бока. Всем мышам сделали ушную метку для идентификации. После того как объем опухоли достиг приблизительно 100-200 мм3, несущих опухоль Colo205 мышей случайным образом группировали и проводили лечение.
Схема лечения представляла собой одну дозу интраперитонеально с дозами носителя для контроля или стандартных и тестовых антител, при 3 мг/кг/мышь или 10 мг/кг/мышь. В некоторых случаях 3 или 4 мышей из групп с носителем и 10 мг/кг подвергали эвтаназии и собирали сыворотку и образцы опухоли через 5 минут, 24 часа или 48 часов после лечения для исследования концентрации лекарственного средства в сыворотке и гистологии опухоли. У оставшихся мышей измерения опухоли проводили дважды в неделю в первые 2 недели, затем один раз в неделю в течение следующих 4 недель.
Полученные результаты для модели на бестимусных мышах "nude" с ксенотрансплантатом Colo205 приведены на фиг.21-25.
На фиг.21 показано, что каждый из Apomab 5.3, 6.3 и 8.3 был высокоэффективен для уменьшения среднего объема опухоли при всех тестируемых дозировках, и их эффективность была по существу такой же, как эффективность антитела 16E2 версии 1.
Эффективность единичных интраперитонеальных доз Apomab 5.2, 6.2, 5.3, 7.2 и 7.3 тестировали на модели на бестимусных мышах "nude" с ксенотрансплантатом Colo205, а результаты показаны на фиг.22. Все тестируемые Apomab были высокоэффективны в отношении уменьшения объема опухоли, и их эффективность была по существу такой же, как эффективность антитела 16E2 версии 1.
Подобным образом, результаты, приведенные на фиг.23, показывают, что Apomab 5.2, 7.3 и 8.3 были эффективны в отношении уменьшения объема опухоли в этой модели колоректального рака. В этом эксперименте Apomab и 16E2 вводили в дозах 1 мг/кг и 3 мг/кг, но в остальном лечили, как описано выше. Эффективность Apomab 7.3 и 8.3 особенно необычна и не демонстрировала какой-либо реверсии в течение 20-суточного тестового периода.
Как показано на фиг.24, противораковая активность Apomab 7.3 значительно превосходит активность Apomab 23.3 и 25.3 в модели с ксенотрансплантатом Colo205.
На фиг.25 проиллюстрированы результаты анализа сравнения противоопухолевой активности Apomab 7.3, полученного из стабильных и транзиторных клеточных линий в модели на мышах с ксенотрансплантатом Colo205. В кратком изложении, самкам бестимусных мышей "nude" в возрасте 6-8 недель инокулировали 5 миллионов клеток Colo205 на мышь в объеме 0,2 мл на мышь, подкожно сзади в область правого бока, как описано выше. Дозы приведены на фигуре. Данные на фиг.25 показывают, что Apomab 7.3, полученный из стабильных и транзиторных клеточных линий, соответственно, равным образом эффективен в модели на мышах с ксенотрансплантатом Colo205.
На фиг.26 приведены результаты эксперимента тестирования противораковой активности Apomab 7.3 (в дозировке 10 мг/кг) в качестве монотерапии или в качестве комбинированного лечения с 80 мг/кг CPT-11 (иринотекан, известное лекарственное средство для лечения колоректального рака) в модели колоректального рака на ксенотрансплантате HCT15. Как продемонстрировано приведенными результатами, хотя и Apomab 7.3, и CPT-11 были эффективны при самостоятельном введении, комбинация из данных двух лекарственных средств продемонстрировала превосходный эффект, превышающий активность Apomab 7.3 и CPT-11, вводимых в качестве монотерапии.
Результаты репрезентативного эксперимента у несущих ксенотрансплантаты LS180 мышей "nude", обработанных Apomab 7.3 в сочетании с CPT-11 (иринотекан), показаны на фиг.27. Снова обнаружено, что комбинированное лечение было лучше относительно введения Apomab 7.3 или CPT-11 в качестве единственного средства, и различия являлись статистически значимыми (вторая панель). Для всех сравниваемых пар по тесту Тьюки-Крамера P<0,05.
ПРИМЕР 3
Оценка противораковой активности Apomab 7.3 в модели неходжкинской лимфомы на ксенотрансплантате BJAB
Противораковую активность Apomab 7.3 (10 мг/кг 1 р/неделя) отдельно и в сочетании с Rituxan® (ритуксимаб, Genentech. Inc.) (4 мг/кг, 1 р/неделя) оценивали на модели неходжкинской лимфомы на ксенотрансплантате BJAB. Так как известно, что клетки лимфомы лучше растут у мышей SCID, для данного исследования использовали мышей SCID (Charles River Laboratory) в возрасте 6-8 недель. Параметры лечения и результаты показаны на фиг.28. Apomab 7.3 и Rituxan® при введении в виде комбинации продемонстрировали синергическую активность.
ПРИМЕР 4
Оценка противораковой активности Apomab 7.3 на модели аденокарциномы поджелудочной железы человека на ксенотрансплантате BxPC3
Противораковую активность Apomab 7.3 (10 мг/кг, в/в) отдельно и в сочетании с гемцитабином (160 мг/кг, в/в) исследовали на модели аденокарциномы поджелудочной железы человека на ксенотрансплантате BxPC3 у самок бестимусных мышей "nude" (Charles River Laboratory). Параметры лечения и результаты показаны на фиг.29. Данные показывают, что противораковая активность Apomab 7.3, вводимого в качестве монотерапии, была значительно лучше относительно эффективности гемцитабина. Комбинированное введение Apomab 7.3 и гемцитабина приводило к дополнительному увеличению эффективности.
ПРИМЕР 5
Оценка противораковой активности Apomab 7.3 отдельно и в сочетании с карбоплатином и таксолом на модели рака легких человека на ксенотрансплантате H460
Противораковую активность Apomab 7.3 (10 мг/кг, 1 р/неделя, в/в) отдельно и в сочетании с карбоплатином и таксолом оценивали относительно носителя в качестве контроля и комбинации из карбоплатина и таксола. Шестидесяти самкам бестимусных мышей "nude" (Charles River Laboratory) инокулировали 5 миллионов клеток H460 на мышь в объеме 0,2 мл на мышь, подкожно сзади в область правого бока. Всем мышам сделали ушную метку для идентификации. Опухолям позволяли достичь среднего размера опухоли 100-200 мм3 и обрабатывали, как показано на фиг.30. В кратком изложении, мышей делили на четыре группы. Группа 1 представляла собой контрольную группу, обрабатываемую носителем (10 мМ гистидин, 8% сахароза и 0,02% Tween 20 (pH 6)). Группу 2 обрабатывали Apomab 7.3 в дозе 10 мг/кг/мышь, в/в, 1 р/неделю в течение 2 недель. Группе 3 вводили карбоплатин (100 мг/кг/мышь, в/в, одну дозу на сутки 0) + таксол (6,25 мг/кг/мышь, п/к, ежедневно в течение 5 последовательных суток в течение 2 недель). Группа 4 получала Apomab 7.3 (дозу 10 мг/кг/мышь, в/в, 1 р/неделю в течение 2 недель) + карбоплатин (100 мг/кг/мышь, в/в, одну дозу на сутки 0) + таксол (6,25 мг/кг/мышь, п/к, ежедневно в течение 5 последовательных суток в течение 2 недель). Противораковая активность Apomab 7.3, вводимого в качестве монотерапии, была сравнима с противораковой активностью сочетания карбоплатин + таксол. Обнаружено, что комбинированное введение Apomab 7.3 + карбоплатин + таксол является лучшим по сравнению с другими формами лечения.
ПРИМЕР 6
Оценка противораковой активности Apomab 7.3 отдельно и в сочетании с карбоплатином и таксолом на модели рака легких человека на ксенотрансплантате H2122
В данном исследовании самкам бестимусных мышей "nude" (Charles River Laboratory, 10 мышей/группу) инокулировали 5 миллионов клеток H2122 на мышь в объеме 0,2 мл на мышь, подкожно сзади в область правого бока. Всем мышам сделали ушную метку для идентификации. Опухолям позволяли достичь среднего размера опухоли 100-200 мм3, мышей случайным образом группировали в четыре группы (6 мышей/группу) и обрабатывали, как показано на фиг.31. Группа 1 представляла собой контрольную группу, обрабатываемую носителем (10 мМ гистидин, 8% сахароза и 0,02% Tween 20 (pH 6)). Группе 2 вводили карбоплатин (100 мг/кг/мышь, в/в, одну дозу на сутки 0) + таксол (6,25 мг/кг/мышь, п/к, ежедневно в течение 5 последовательных суток в течение 2 недель). Группа 3 получала Apomab 7.3 (в дозе 10 мг/кг/мышь, в/в, 1 р/неделю в течение 2 недель). Группа 4 получала Apomab 7.3 (дозу 10 мг/кг/мышь, в/в, 1р/неделю в течение 2 недель) + карбоплатин (100 мг/кг/мышь, в/в, одну дозу на сутки 0) + таксол (6,25 мг/кг/мышь, п/к, ежедневно в течение 5 последовательных суток в течение 2 недель). Как показано на фиг.31, комбинация карбоплатин + таксол не продемонстрировала значимой противораковой активности относительно обрабатываемой носителем контрольной группы. Напротив, Apomab 7.3, вводимый в качестве монотерапии, продемонстрировал заметную противоопухолевую активность. Все шесть мышей, обработанных Apomab 7.3, продемонстрировали полный ответ (CR) без какой-либо реверсии в течение 70-суточного периода лечения. Подобную активность выявили в группе, обрабатываемой сочетанием Apomab 7.3 + карбоплатин + таксол.
Для определения максимума эффективной дозы Apomab 7.3 в этой модели самкам бестимусных мышей "nude" (Charles River Laboratory) в возрасте 6-8 недель инокулировали 5 миллионов клеток H2122 на мышь в объеме 0,2 мл на мышь, подкожно сзади в область правого бока. Всем мышам сделали ушную метку для идентификации. Опухолям позволяли достичь среднего размера опухоли 100-200 мм3, мышей случайным образом группировали (13 мышей/группу) и обрабатывали, как описано ниже. Каждую мышь, исключаемую из групп обработки, подвергали эвтаназии.
Группа A представляла собой контрольную группу, обрабатываемую носителем (0,5 M аргининсукцинат, 20 мМ Tris, 0,02% Tween 20, pH 7,2). Носитель вводили 5 р/неделю, в/в в течение одной недели. Группе B вводили Apomab 7.3 в одной в/в дозе 1 мг/кг/мышь. Группе C вводили Apomab 7.3 в одной в/в дозе 3 мг/кг/мышь. Группе D вводили Apomab 7.3 в одной в/в дозе 10 мг/кг/мышь. Через 48 часов после первой обработки трех мышей из каждой из групп B, C и D подвергали эвтаназии и их опухоли собирали, как указано ниже. Одну из опухолей фиксировали в 10% формалине для гистологического исследования. Одну из опухолей замораживали в жидком азоте для исследований РНК. Одну из опухолей замораживали в жидком азоте для "вестерн"-блоттинга. Измерения опухолей проводили 2 р/неделю в течение первых 2 недель, а затем 1 р/неделю в течение следующих 4 недель. По окончании 6 недель или после того, как опухоль достигнет объема приблизительно 800-1000 мм3, всех оставшихся мышей подвергали эвтаназии. Кривые доза-ответ, показанные на фиг.32, первая панель, указывают на то, что Apomab 7.3 эффективен при всех тестируемых дозах, но дозы 3 мг/кг и 10 мг/кг продемонстрировали явное (хотя и статистически незначимое) улучшение относительно дозы 1 мг/кг. Для всех сравниваемых пар по тесту Тьюки-Крамера P<0,05 (см., фиг.32, вторая панель).
ПРИМЕР 7
Оценка противораковой активности Apomab 23.3, 25.3 и 7.3 на модели колоректального рака человека на ксенотрансплантате Colo205
В данном исследовании самкам бестимусных мышей "nude" (Charles River Laboratory) инокулировали 5 миллионов клеток Colo205 на мышь в объеме 0,2 мл на мышь, подкожно сзади в область правого бока. Всем мышам сделали ушную метку для идентификации. Опухолям позволяли достичь среднего размера опухоли 100-200 мм3, мышей случайным образом группировали на семь групп (10 мышей/группу) и обрабатывали, как показано на фиг.33. Группа 1 представляла собой контрольную группу, обрабатываемую носителем (10 мМ гистидин, 8% сахароза и 0,02% Tween 20 (pH 6)). Группе 2 вводили одну дозу 3 мг/кг/мышь, в/в, Apomab 7.3. Группе 3 вводили одну дозу 10 мг/кг/мышь, в/в, Apomab 7.3. Группе 4 вводили одну дозу 3 мг/кг/мышь, в/в, Apomab 23.3. Группе 5 вводили одну дозу 10 мг/кг/мышь, в/в, Apomab 23.3. Группе 6 вводили одну дозу 3 мг/кг/мышь, в/в, Apomab 25.3. Группе 7 вводили одну дозу 10 мг/кг/мышь, в/в, Apomab 25.3. Через 24 часа после обработки мышей взвешивали. Опухоли измеряли 2р/неделю в течение первых 2 недель, а затем каждую неделю в течение следующих 4 недель. Через 6 недель или когда опухоль достигала размера >1000 мм3 мышей умерщвляли.
Результаты показаны на фиг.33. Как проиллюстрировано данными через 25 суток и Apomab 7.3, и Apomab 25.3 продемонстрировали в этой модели значимую противораковую активность.
ПРИМЕР 8
Оценка противораковой активности моноклонального антитела Apomab 7.3 в отношении ксенотрансплантатов карциномы толстой кишки человека Colo205 мышам "nude"
Животные
Самки бестимусных мышей "nude" (nu/nu, Harlan) на сутки 1 исследования находились в возрасте 11 или 12 недель. Животных снабжали ad libitum водой и NIH 31 Modified and Irradiated Lab Diet®, состоящей из 18,0% общего белка, 5,0% общего жира и 5,0% сырой клетчатки. Животных содержали на облученных ALPHA-Dri® bed-o'cobs® Laboratory Animal Bedding в статических микроизоляторах при 12-часовом световом цикле при 21-22°C и 40-60% влажности.
Имплантация опухоли
Ксенотрансплантаты происходили из культивируемых клеток карциномы толстой кишки человека Colo205. Опухолевые клетки растили до фазы середины логарифмического роста в RPMI-1640, дополненной 10% инактивированной нагреванием эмбриональной телячьей сывороткой, 100 единиц/мл пенициллином G натрия, 100 мкг/мл стрептомицинсульфатом, 0,25 мкг/мл амфотерицином B и 25 мкг/мл гентамицином, 2 мМ глутамином, 1 мМ пируватом натрия, 10 мМ HEPES и 0,075% бикарбонатом натрия. Клеточные культуры содержали во флаконах для тканевых культур во влажном инкубаторе при 37°C, в атмосфере 5% CO2 и 95% воздуха. В сутки имплантации опухолевых клеток клетки Colo205 собирали и ресуспендировали в 50% матриксе Matrigel (BD Biosciences) в PBS при концентрации 5×106 клеток/мл. Каждая мышь в тесте получала 1×106 клеток Colo205, имплантированных подкожно в правый бок, и контролировали рост опухоли, когда средний размер достигнет от 100 до 300 мм3. Через тринадцать суток, обозначенных как сутки 1 исследования, индивидуальные объемы опухолей находились в диапазоне от 126 до 288 мм3, и животных сортировали на шесть групп, где каждая состояла из десяти мышей со средним объемом опухоли 188 мм3.
Тестируемые вещества и лечение
Тестируемые вещества в течение дозирования хранили на льду, а дозируемые растворы впоследствии хранили при 4°C. В контрольной группе с носителем (группа 1) мыши получали носитель, вводимый интраперитонеально раз в сутки в течение пяти суток с последующими двумя сутками отдыха, затем один раз в сутки в течение дополнительных пяти суток. В тестовых группах (группы 2-4) животных обрабатывали лигандом Apo2L.0 (60 мг/мг и/п по схеме 5/2/5), Apomab 7.3 (3 мг/кг в/в на сутки 1, 8) и мышиным моноклональным антителом к VEGF B20-4.1 (10 мг/кг и/п на сутки 1 и 8). Группы 5 и 6 получали сочетание B20-4.1 с Apo2L.0 и B20-4.1 с Apomab 7.3, соответственно. Каждую дозу вводили в объеме 0,2 мл на 20 г массы тела (10 мл/кг), в соответствии с массой животного.
Конечная точка
Опухоли измеряли дважды в неделю с применением циркулей. Каждое животное подвергали эвтаназии, когда размер его опухоли достигал объема 2000 мм3 или после конечной точки исследования на сутки 68, в зависимости от того, что произойдет первым. Конечный объем опухоли 1000 мм3 выбрали для анализа задержки опухолевого роста вследствие того, что ряд опухолей не достигал размера 2000 мм3. Время до конечной точки (TTE) для каждой мыши рассчитывали на основании следующего уравнения:
TTE (сутки)=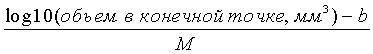
где b представляет собой пересечение, а M представляет собой угловой коэффициент линии, получаемой посредством линейной регрессии множества данных опухолевого роста. Множество данных содержало первое наблюдение, превышающее конечный объем исследования, и три последовательных наблюдения, которые непосредственно предшествовали достижению конечного объема. У животных, не достигавших конечной точки, значение TTE приравнивали последней дате исследования. У животных, погибших вследствие связанной с лечением гибели или не связанной с лечением гибели вследствие метастазов, значение TTE приравнивали суткам гибели. Животных, погибших вследствие не связанной с лечением гибели или гибели с неизвестной причиной, исключали из расчетов TTE.
Исход лечения оценивали по задержке опухолевого роста (TGD), которую определяли как увеличение среднего времени до конечной точки (TTE) в группе, получавшей лечение, по сравнению с контрольной группой:
TGD=T-C,
выраженной в сутках, или как процент средней TTE контрольной группы
%TGD=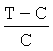 ×100,
×100,
где T=среднее TTE для группы, получавшей лечение,
C=среднее TTE для контрольной группы.
Лечение может вызывать частичную регрессию (PR) или полную регрессию (CR) опухоли у животного. При PR-ответе объем опухоли составляет 50% или менее от ее объема на сутки 1 для трех последовательных измерений в течение исследования и равен или больше 13,5 мм3 для одного или нескольких из этих трех измерений. При CR-ответе объем опухоли составляет менее 13,5 мм3 для одного или нескольких из этих трех измерений в течение исследования. Животное с CR-ответом при завершении исследования дополнительно классифицируют как выжившее без опухоли (TFS). Ответы с регрессией контролировали и записывали.
Отбор образцов
У животных из каждой группы получали образцы опухолей в конечной точке. Этих животных подвергали эвтаназии посредством смещения шейных позвонков точно перед отбором образцов. Опухоли трех животных из группы собирали, разрезали и фиксировали в 10% формалине в нейтральном буфере в течение срока от 12 до 24 часов при комнатной температуре.
Статистический и графический анализы
Для анализа значимости различий между значениями TTE подвергаемых лечению и контрольной групп использовали логарифмический ранговый тест. Проводили двусторонний статистический анализ на уровне значимости P=0,05, с результатами, считающимися значимыми при 0,01≤P≤0,05 и высоко значимыми при P<0,01.
Кривые среднего опухолевого роста представляют групповые средние объемы опухоли как функцию времени. Когда животное выходило из исследования из-за размера опухоли, конечный объем опухоли, записываемый для животного, включали в данные, используемые для расчета группового среднего объема опухоли в последующие временные точки. Для демонстрации доли животных, остающихся в исследовании, как функции времени строили графики Каплана-Мейера. Для этих графиков использовали то же множество данных, что и для логарифмического рангового теста.
Результаты
На фиг.34 приведены кривые группового среднего опухолевого роста (верхняя панель) и графики Каплана-Мейера (нижняя панель) для каждой группы в этом исследовании. Среднее TTE контрольных мышей, обрабатываемых носителем, составляло 10,0 суток, с одной опухолью (из десяти), не достигшей 1000 мм3 конечного объема опухоли. B20-4.1, вводимое при 10 мг/кг и/п на сутки 1 и 8, приводило к умеренной TGD в размере 19,9 суток (113%), которая было статистически незначимой. Монотерапия Apo2L.0 и Apomab 7.3 были эффективными против Colo205, приводя к задержке опухолевого роста в размере 28,4 суток (190%) и 53,0 суток (355%), соответственно. Добавление B20-4.1 к Apo2L.0 или Apomab 7.3 не увеличивало эффективности лечения в отношении TGD или ответов с регрессией.
ПРИМЕР 9
Оценка противораковой активности моноклонального антитела Apomab 7.3 в качестве монотерапии и в сочетании с карбоплатином и паклитакселом против SKMES-1 NSCLC человека
Тестировали противоопухолевую активность лиганда Apo2L.0 и Apomab 7.3 в качестве монотерапии и в сочетании с карбоплатином и паклитакселом против SKMES-1 NSCLC человека.
Животные
Самки бестимусных мышей "nude" (nu/nu, Harlan) на сутки 1 исследования находились в возрасте от 9 до 10 недель и обладали массой тела от 17,4 до 25,4 г. Животных снабжали ad libitum водой и NIH 31 Modified and Irradiated Lab Diet®, состоящей из 18,0% общего белка, 5,0% общего жира и 5,0% сырой клетчатки. Животных содержали на облученных ALPHA-Dri® bed-o'cobs® Laboratory Animal Bedding в статических микроизоляторах при 12-часовом световом цикле при 21-22°C и 40-60% влажности.
Имплантация опухоли
Ксенотрансплантаты происходили из опухолей легких SKMES-1, поддерживаемых серийными трансплантациями при PRC. Каждая мышь в тесте получала фрагмент опухоли SKMES-1 объемом 1 мм3, имплантируемый подкожно в правый бок, и рост опухоли контролировали. Через тринадцать суток, обозначенных как сутки 1 исследования, индивидуальные объемы опухолей находились в диапазоне от 63 до 144 мм3, и животных сортировали на четыре группы, где каждая состояла из десяти мышей, и две группы, состоящие из девяти мышей. Средний объем опухолей составлял от 93 до 95 мм3.
Тестируемые вещества и лечение
Все тестируемые вещества предоставляли готовыми к дозированию при 0,1 мл на 20 г массы тела (5 мл/кг) и после получения хранили при -80°C. Тестируемые вещества оттаивали на первые сутки дозирования, хранили на льду в течение дозирования, затем хранили при 4°C. Карбоплатин (инъекция Paraplatin®, Bristol Myers Squibb) разводили 5% декстрозой в воде (D5W) с получением желаемой дозы в объеме 0,2 мл на 10 г массы тела (10 мл/кг). Паклитаксел (Natural Pharmaceuticals, Inc.) разбавляли D5W на каждые сутки дозирования из 10× исходного раствора с получением носителя, содержащего 5% этанола и 5% кремофора EL в 90% D5W (5% носитель EC) так, что желаемую дозу доставляли в 0,1 мл на 20 г массы тела.
Группа 1 (носитель для контроля) мышей получала носитель, вводимый интраперитонеально один раз в сутки в течение пяти суток (и/п, qd×5), и служила в качестве контролей опухолевого роста. Мышам в группах 2 и 3 проводили монотерапию Apo2L.0 (60 мг/кг, п/к qd×5) и Apomab 7.3 (10 мг/кг, в/в qd×1), соответственно. Мышам в группе 4 вводили комбинацию карбоплатина (100 мг/кг, и/п qd×1) и паклитаксела (6,25 мг/кг, п/к qd×5). Мышам в группе 5 и 6 вводили карбоплатин и паклитаксел в сочетании с Apo2L или Apomab 7.3, соответственно. Каждую дозу вводили в объеме, указанном в предыдущем разделе, и приводили в соответствие с массой тела животного.
Отбор образцов и статистический анализ в конечной точке
Конечную точку определяли и отбор образцов и статистический анализ проводили, как описано в предыдущем примере 8.
Результаты
На фиг.35 и 36 показаны кривые группового среднего опухолевого роста и графики Каплана-Мейера для групп, обрабатываемых Apo2L.0 и Apomab 7.3, соответственно.
Опухоли всех обработанных носителем контрольных мышей росли до конечного объема 1500 мм3 со средним TTE 18,9 суток. Таким образом, максимальный TGD, достижимый в этом 45-суточном исследовании, составлял 26,1 суток (138%). Кривая среднего опухолевого роста и график Каплана-Мейера для контрольной группы включены в верхние и нижние панели на фиг.35 и 36.
Для обработанной Apo2L.0 группы (группа 2), среднее TTE составляло 22,9 суток и соответствовало TGD в размере 4,0 суток (21%) и статистически незначимой активности. На фиг.28 (верхняя панель) показано, что средние объемы опухолей в группе 2 сокращаются в течение периода лечения, затем восстанавливается быстрый рост опухоли.
Среднее TTE в группе 3 составляло 2,0 суток и соответствовало TGD в размере 7,1 суток (38%) и статистически высокозначимой активности (P=0,005). Не зарегистрировано ни одного регрессивного ответа, и все опухоли достигали конечного объема 1500 мм3. Кривая среднего опухолевого роста группы 3 показывает, что существует исходная задержка опухолевого роста относительно контролей.
Среднее TTE группы мышей 4, обработанных карбоплатином и паклитакселом, составляло 26,5 суток и соответствовало TGD в размере 7,6 суток (40%) и статистически значимой активности (P=0,01). Все опухоли в группе 4 росли до конечного объема 1500 мм3, и не зарегистрировано ни одного регрессивного ответа. Кривая среднего опухолевого роста для группы мышей 4 указывает на умеренную задержку роста опухоли относительно контрольных мышей (см. фиг.35 и 36, верхние панели).
Обработка тройным сочетанием Apo2L.0, карбоплатина и паклитаксела приводила к среднему TTE 32,7 суток, что соответствовало TGD 13,9 суток (73%) и высокозначимой активности относительно группы 1 (P=0,002). Среднее TTE в размере 32,7 суток при применении этого тройного сочетания было длиннее, чем среднее TTE в размере 22,9 суток группы с монотерапией Apo2L.0 или среднее TTE в размере 16,5 суток при контрольной химиотерапевтической обработке, но различия при логарифмическом ранговом тесте не достигали статистической значимости. Несмотря на отсутствие статистической значимости кривые среднего опухолевого роста указывали большую активность с сочетанием из группы 5 по сравнению с монотерапией Apo2L.0 или химиотерапией карбоплатин и паклитаксел (см. фиг.28, верхняя панель).
Лечение тройным сочетанием Apomab 7.3 (группа 6), карбоплатина и паклитаксела приводило к 3/10 TR смерти, таким образом, эту группу не оценивали на TGD. Однако кривые среднего опухолевого роста указывали на большую активность с сочетанием из группы 6 по сравнению с монотерапией Apomab 7.3 или химиотерапией карбоплатин и паклитаксел (см. фиг.37, верхняя панель).
Вывод
Несмотря на относительно высокую смертность в этом эксперименте данные показывают, что Apo2L.0 и Apomab 7.3 могут добавлять противоопухолевое воздействие при лечении карбоплатином и паклитакселом.
ПРИМЕР 10
Оценка противораковой активности моноклонального антитела Apomab 7.3 в качестве монотерапии и в сочетании с антителом к VEGF в модели ксенотрансплантата карциномы Colo205
Животные
Самки бестимусных мышей "nude" (nu/nu, Harlan) на сутки 1 исследования находились в возрасте 7-8 недель. Животных снабжали ad libitum водой и NIH 31 Modified and Irradiated Lab Diet®, состоящей из 18,0% общего белка, 5,0% общего жира и 5,0% сырой клетчатки. Животных содержали на облученных ALPHA-Dri® bed-o'cobs® Laboratory Animal Bedding в статических микроизоляторах при 12-часовом световом цикле при 21-22°C и 40-60% влажности.
Культура опухолевых клеток
Клетки карциномы толстой кишки человека Colo205 культивировали в среде RPMI-1640, содержащей 100 единиц/мл пенициллина G натрия, 100 мкг/мл стрептомицинсульфата, 0,25 мкг/мл амфотерицина B и 25 мкг/мл гентамицина. Среду дополняли 10% инактивированной нагреванием эмбриональной телячьей сывороткой, 2 мМ глутамином и 1 мМ бикарбонатом натрия. Опухолевые клетки культивировали во флаконах для тканевых культур во влажном инкубаторе при 37°C, в атмосфере 5% CO2 и 95% воздуха.
Имплантация in vivo
Клетки карциномы человека Colo205, применяемые для имплантации, собирали в течение логарифмической фазы роста и ресуспендировали в 50% matrigel в концентрации 5×106 клеток/мл. Каждой мыши п/к в правый бок инъецировали 1×106 клеток (0,2 мл клеточная суспензия). Опухоли проверяли дважды в неделю, а затем ежедневно, когда их объем приближался к 100-300 мм3. На сутки 1 исследования животных сортировали на группы обработки с размерами опухолей 108,0-220,5 мм3 и групповым средним размером опухоли 149,8 мм. Массу опухоли рассчитывали в предположении, что 1 мг эквивалентен 1 мм3 объема опухоли.
Тестируемые вещества и лечение
Тестируемые вещества предоставляли готовыми для дозирования. Дозируемые растворы хранили при 4°C.
Мышей сортировали на шесть групп с десятью мышами на группу. Все средства для лечения вводили интраперитонеально (и/п).
Apo2L.0 и его носитель вводили один раз в сутки на сутки 1-5 (qd×5). Apomab 7.3 и мышиное антитело к VEGF к G6 вводили один раз на сутки 1 (qd×1). Мышей контрольной группы 1 обрабатывали носителем для Apo2L.0. Группе 2 проводили монотерапию Apo2L.0 при 60 мг/кг. Группе 3 проводили монотерапию Apomab 7.3 при 3 мг/кг. Группе 4 проводили монотерапию BY4 при 5 мг/кг. Группам 5 и 6 вводили Apo2L.0 при 60 мг/кг и Apomab 7.3 при 3 мг/кг, соответственно, каждое в сочетании с антителом к G6 при 5 мг/кг. Во всех группах объем дозирования 0,2 мл/20 г массы мыши приводили в соответствие с массой тела каждого животного.
Отбор образцов и статистические анализы в конечной точке
Конечную точку определяли и отбор образцов и статистические анализы проводили, как описано в предыдущем примере 8.
Результаты
Результаты показаны на фиг.37, где кривые на верхней панели демонстрируют групповые средние объемы опухолей в зависимости от времени, а график Каплана-Мейера на нижней панели демонстрирует долю оставшихся в каждой группе поддающихся оценке животных в зависимости от времени.
Мыши контрольной группы 1 получали носитель Apo2L.0 и служили в качестве контроля для всех групп, получавших лечение. Опухоли у всех десяти мышей росли до конечного объема 1500 мм3 со средним TTE 20,8 суток. Таким образом, максимально возможное TGD в этом 61-суточном исследовании составляло 193%.
Группе 2 проводили монотерапию Apo2L.0 при 60 мг/кг. Эта обработка приводила к высокозначимой противоопухолевой активности относительно контрольной группы, получавшей носитель (P<0,001), и среднему TTE в размере 53,6 суток. Это среднее TTE соответствовало T-C в размере 32,8 суток и TGD 158%. Средний объем опухоли на сутки 61 для пяти мышей составлял 1,210 мм3. Зафиксирован один LTTFS.
Группе 3 проводили монотерапию Apomab 7.3 при 3 мг/кг. Эта обработка приводила к высокозначимой активности (P<0,001) с максимально возможным TGD в размере 193% и MTV (6) 776 мм3. Зафиксирован один LTTFS, один транзиторный CR-ответ и три FR-ответа.
Группе 4 проводили монотерапию антителом к G6 при 5 мг/кг. Эта обработка приводила к высокозначимой активности (P<0,01), с 86% TGF и MTV(3) 1,224 мм3. Регрессивных ответов не зафиксировано.
Группа 5 получала Apo2L.0 при 60 мг/кг в сочетании с антителом к G6 при 5 мг/кг. Комбинированное лечение приводило к 138% TGD. Противоопухолевая активность была высокозначимой относительно обработки носителем (P<0,001), но незначимой относительно обоих видов монотерапии. В группе 5 MTV(3) составлял 1,080 мм3, и зафиксирован один PR-ответ.
Группе 6 проводили комбинированное лечение с применением Apomab 7.3 при 3 мг/кг и антитела к G6 при 5 мг/кг. Это лечение приводило к максимально возможным 193% TGD. Противоопухолевая активность была высокозначимой относительно обработки носителем (P<0,001), значимой относительно монотерапии антителом к G6, но незначимой относительно монотерапии Apomab 7.3. В группе 6 MTV(8) составлял 208 мм3, и зафиксировано пять PR-ответов.
Выводы
Опухоли хорошо отвечали на монотерапию Apomab 7.3 в концентрации 3 мг/кг qd×1 (группа 3). Это лечение приводило к высокозначимой активности относительно контрольной группы с применением носителя и максимально возможным 193% TGD. Из шести мышей, которые выжили до суток 61 с MTV 776 мм3, у пяти животных наблюдали регрессию опухоли. Эта монотерапия приводила к одному LTTFS, одному транзиторному CR-ответу и трем PR-ответам. Средний объем опухоли не увеличивался до окончания суток 15 (фиг.37).
Комбинированное лечение с применением Apomab 7.3 и мышиного антитела к VEGF к G6 приводило к более сильной активности, чем наблюдалась с применением Apomab 7.3 или антитела к G6 отдельно. Это комбинированное лечение приводило к восьми выжившим в течение 61 суток и позволяло получить наименьший MTV в исследовании, 208 мм3. Кривая среднего опухолевого роста демонстрирует уменьшение опухоли или состояние покоя до суток 33 с последующим очень медленным ростом опухоли (фиг.37). Сочетание приводило к значимо большей активности, чем при монотерапии антителом к G6, но значимо не отличалось от результатов монотерапии Apomab 7.3, кроме того, сочетание приводило к пяти PR-ответам, тогда как пять регрессивных ответов, полученных с применением монотерапии Apomab 7.3, включали в себя один CR и один LTTFS.
Все лекарственные средства хорошо переносились. В исследовании не наблюдали потери массы или другой явной токсичности.
Таким образом, комбинированное лечение Apomab 7.3/антитело к G6 приводило к большему числу выживших в течение 61 суток и меньшему MTV, чем лечение с применением любых соответствующих способов монотерапии. Однако сочетание Apomab 7.3/антитело к G6 не приводило к лечебной активности, которую наблюдали при применении монотерапии Apomab 7.3.



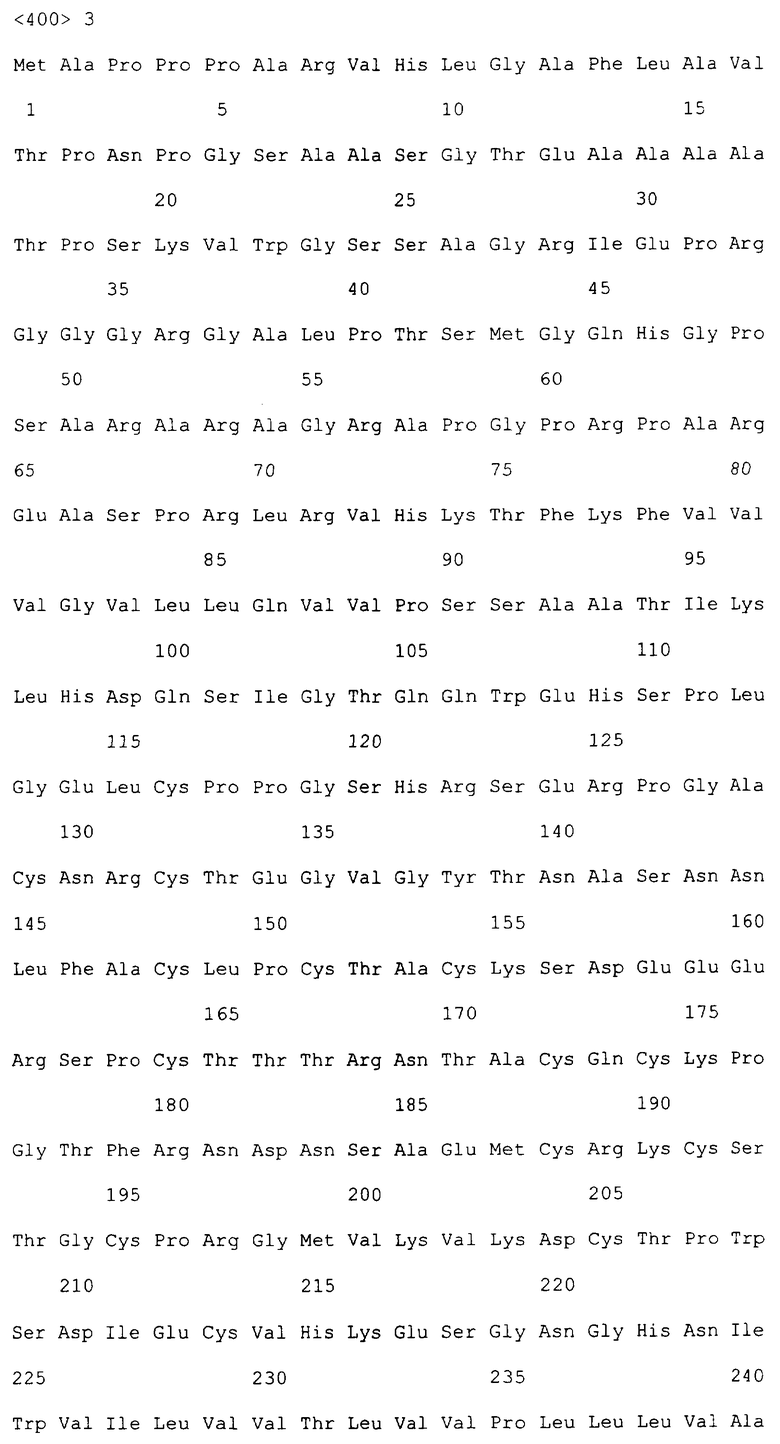
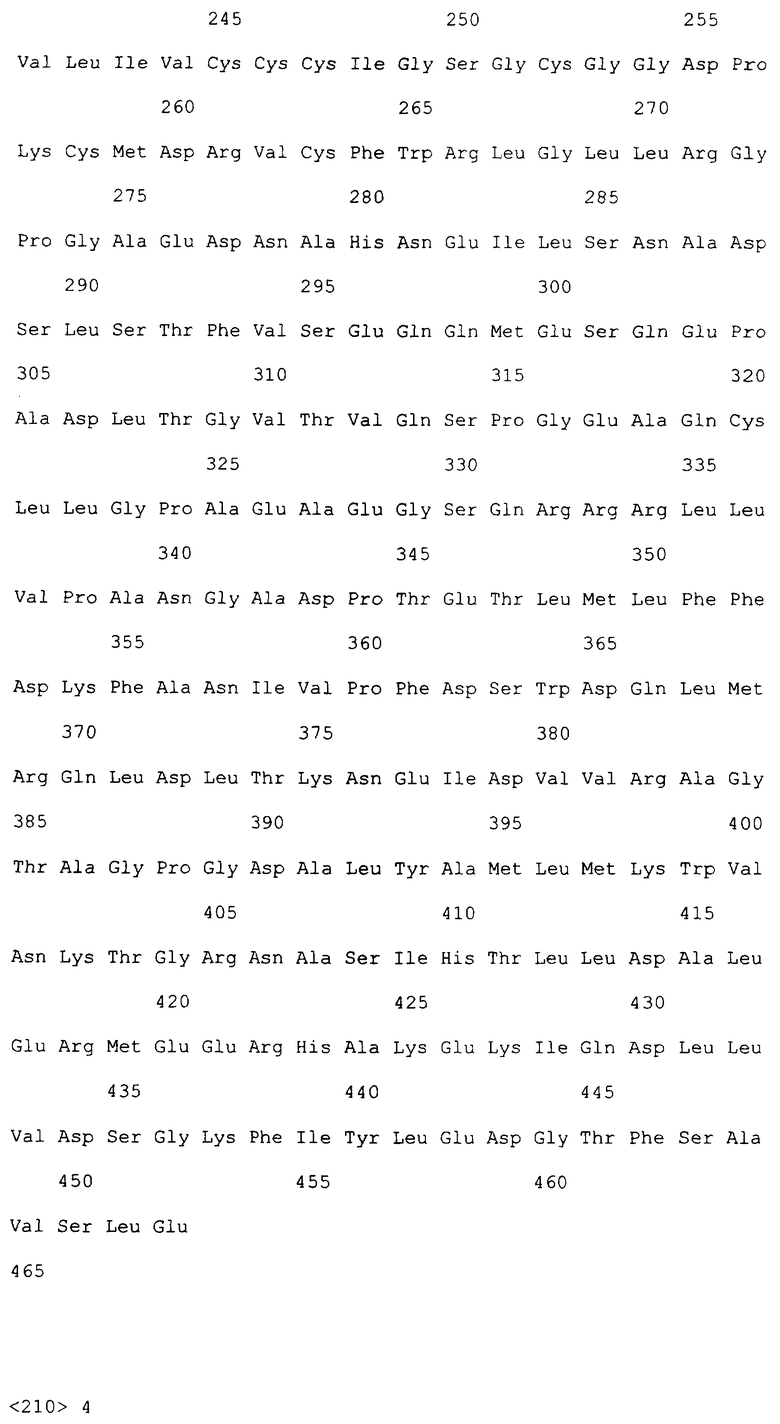





















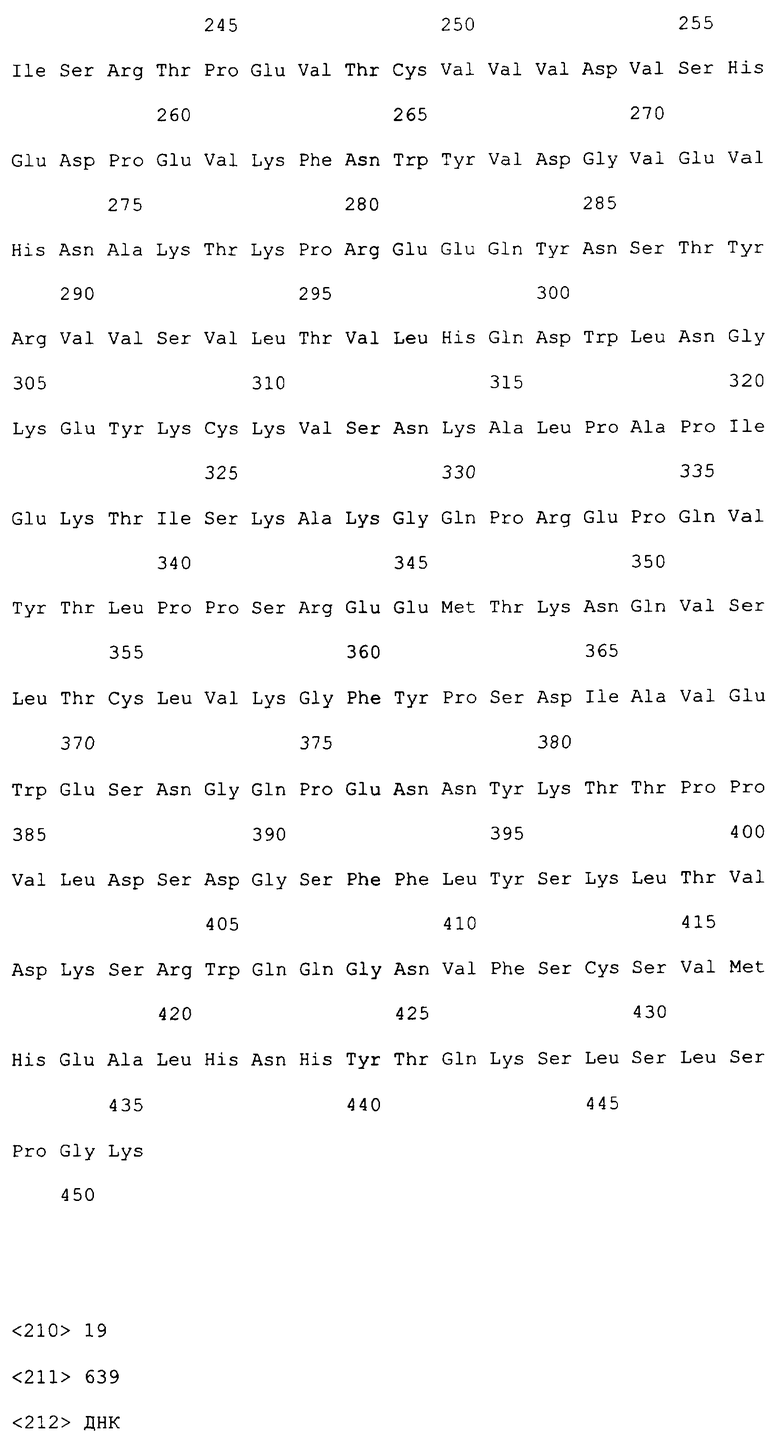







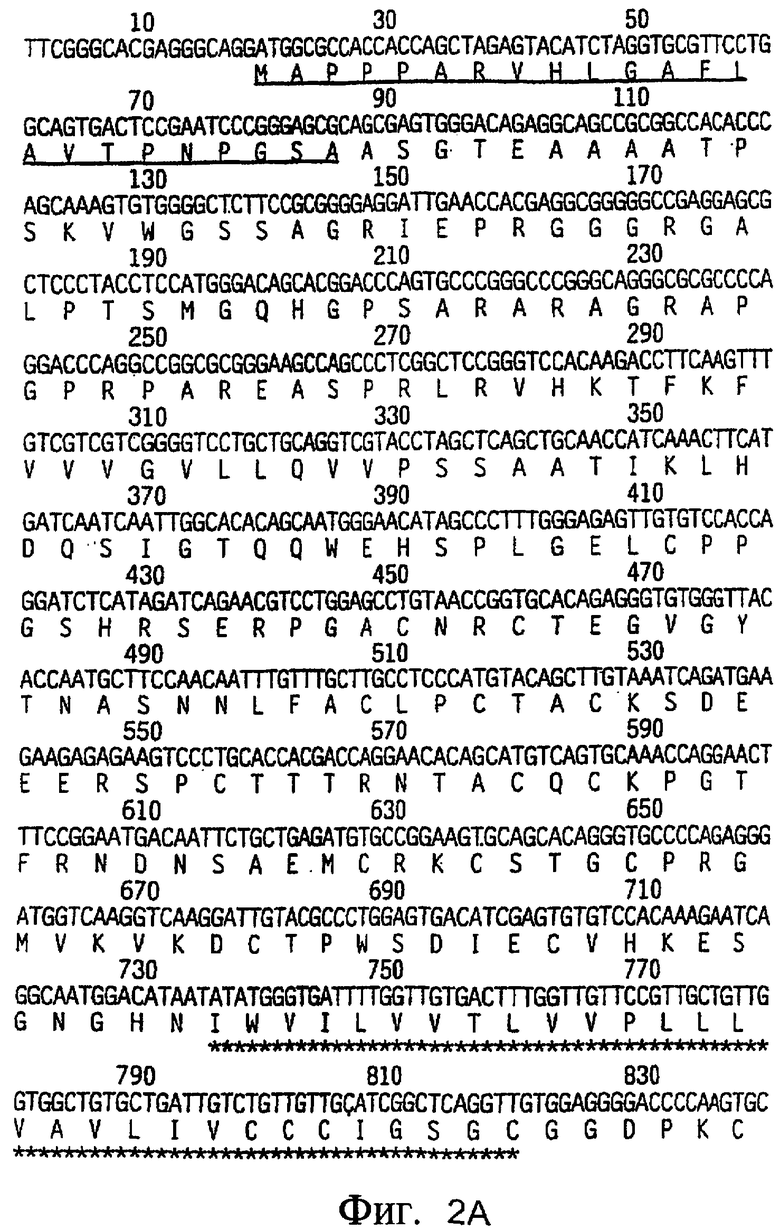
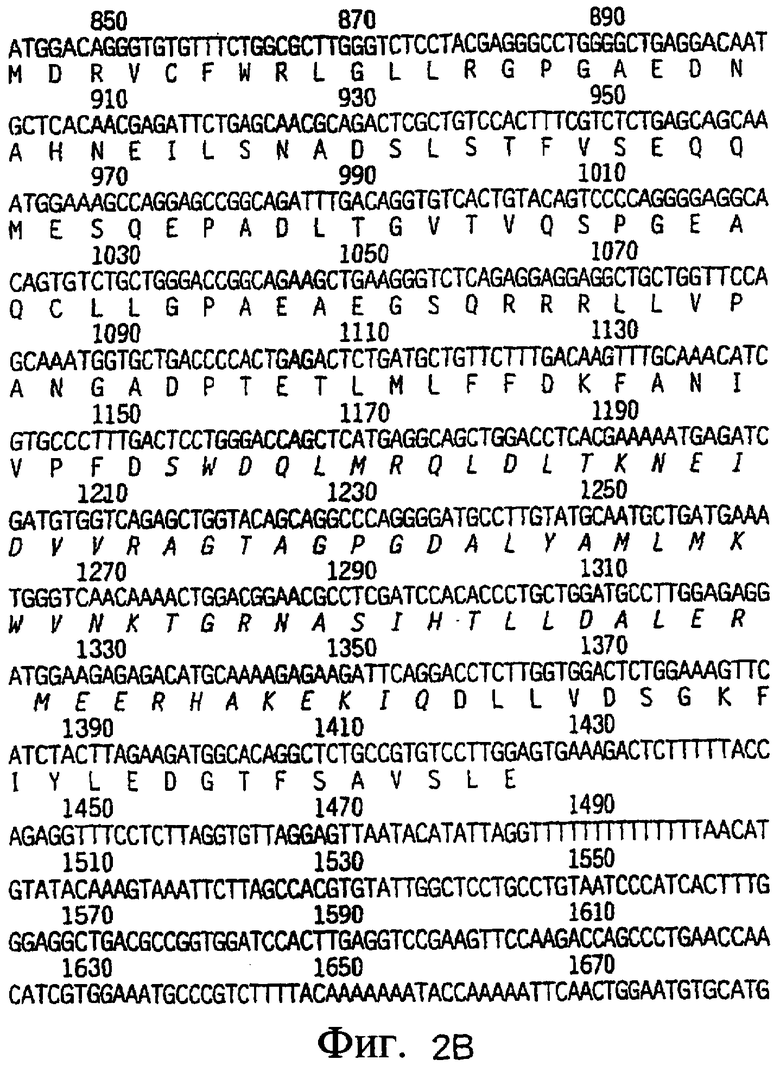












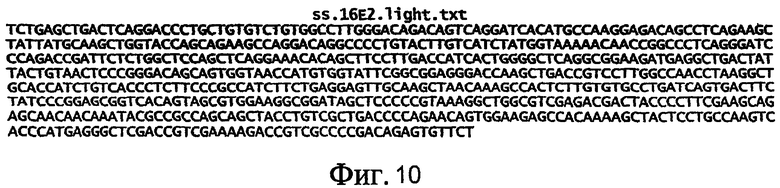














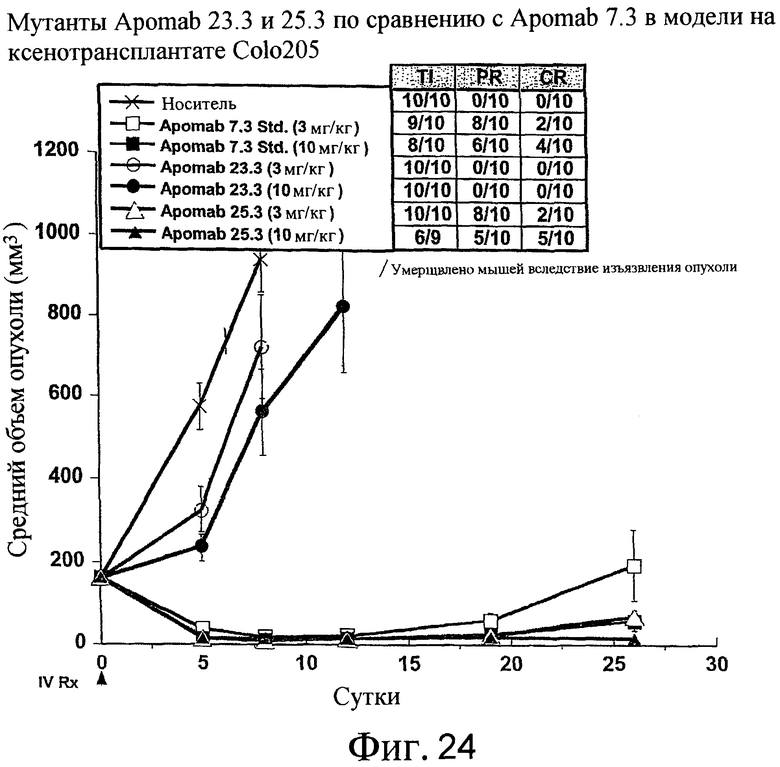
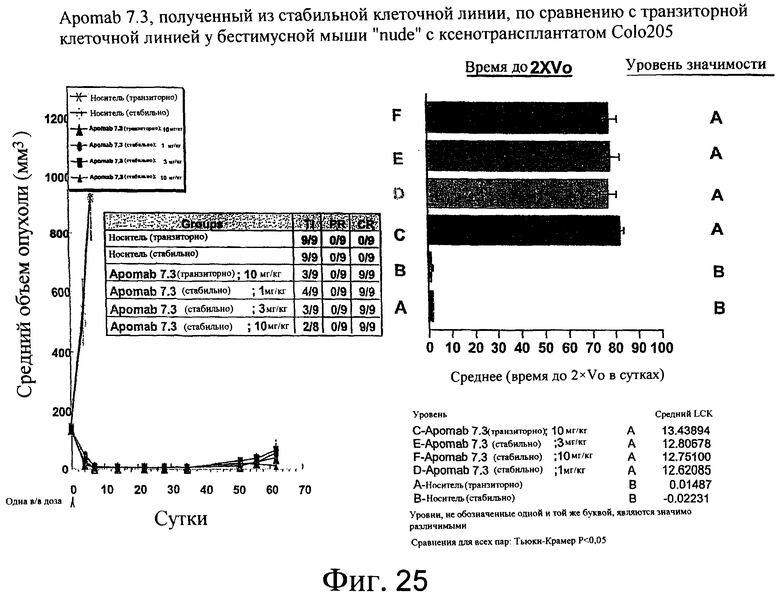
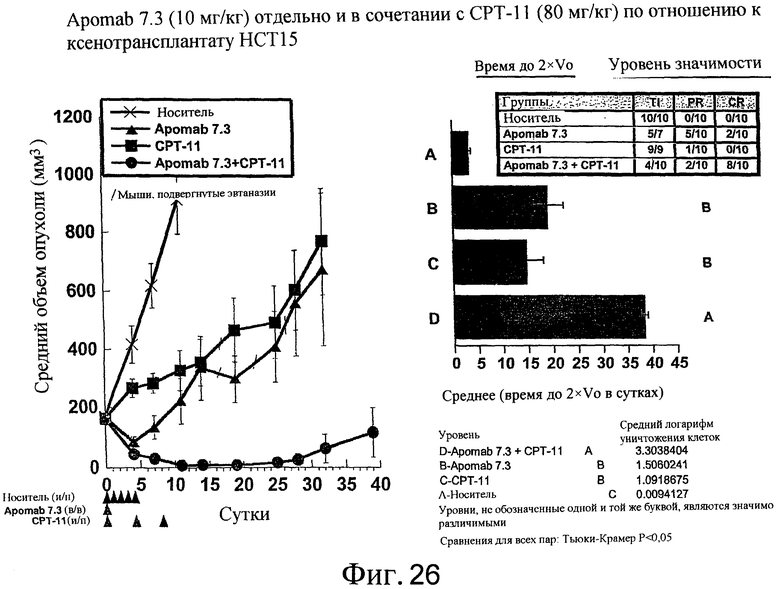
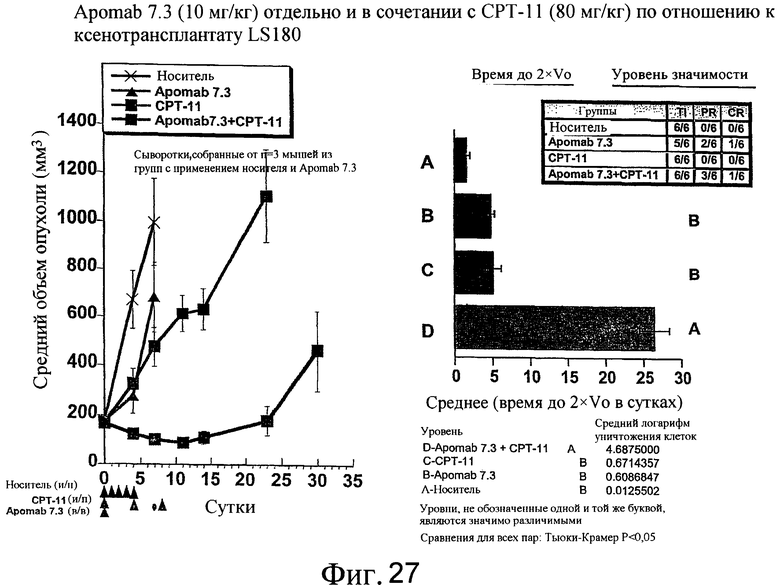
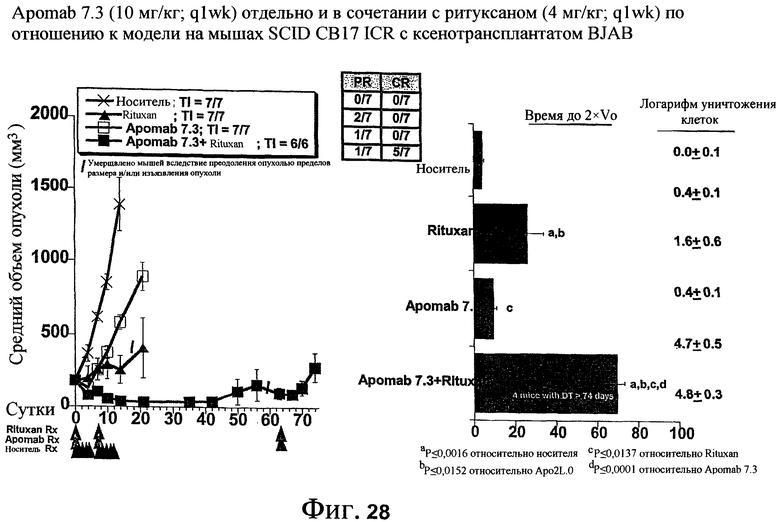
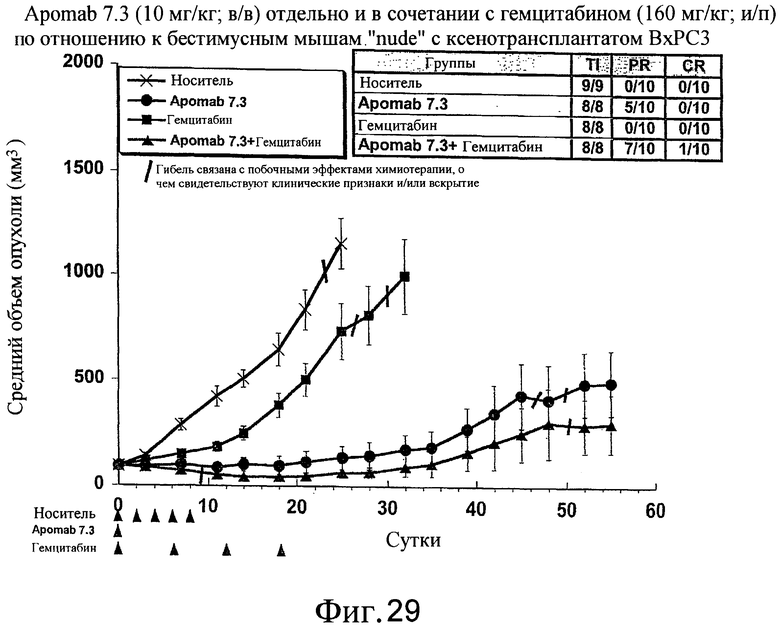
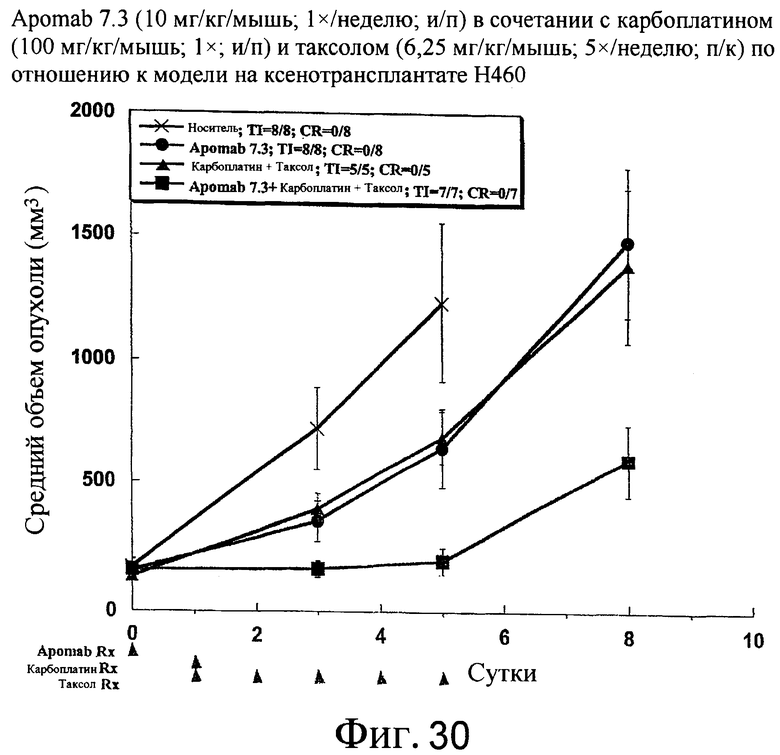

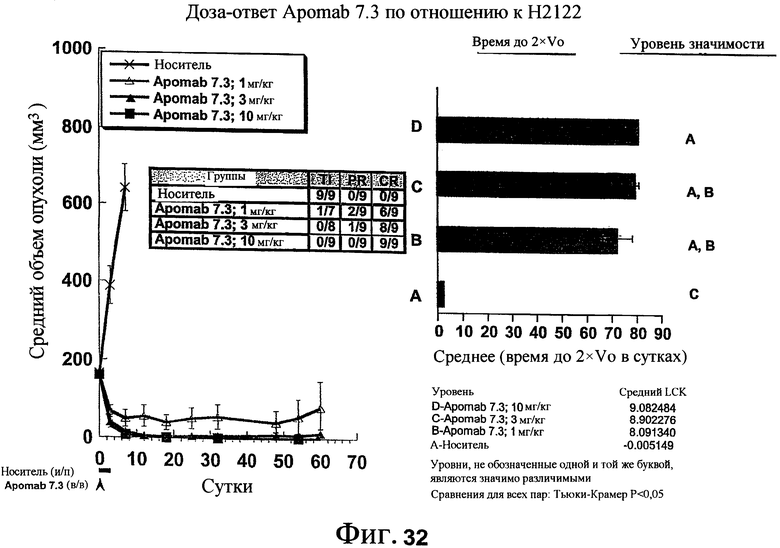
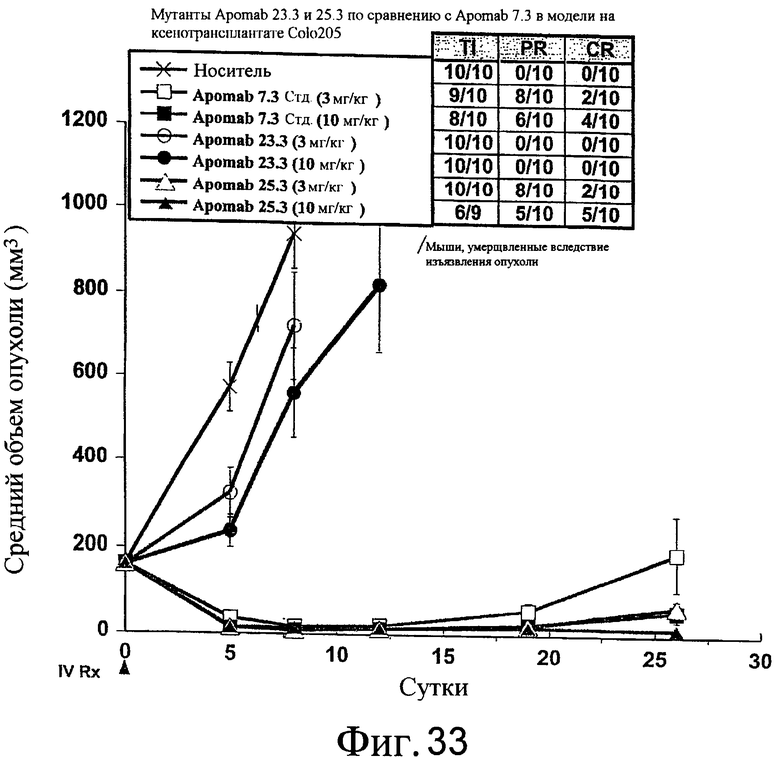
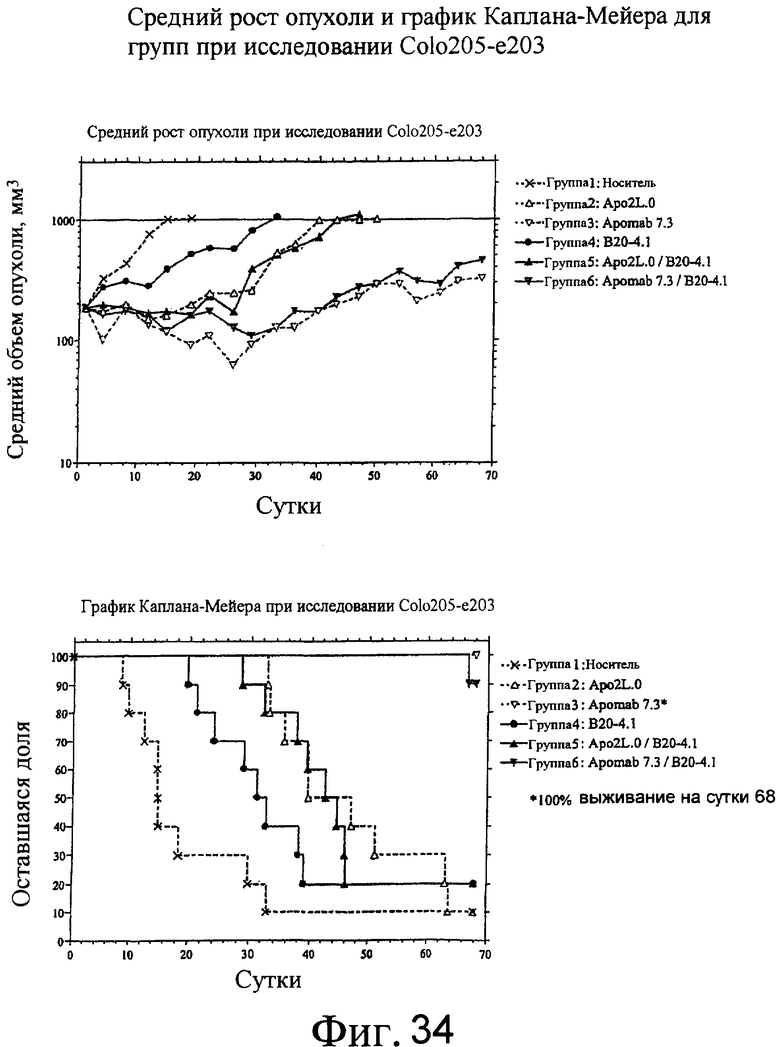
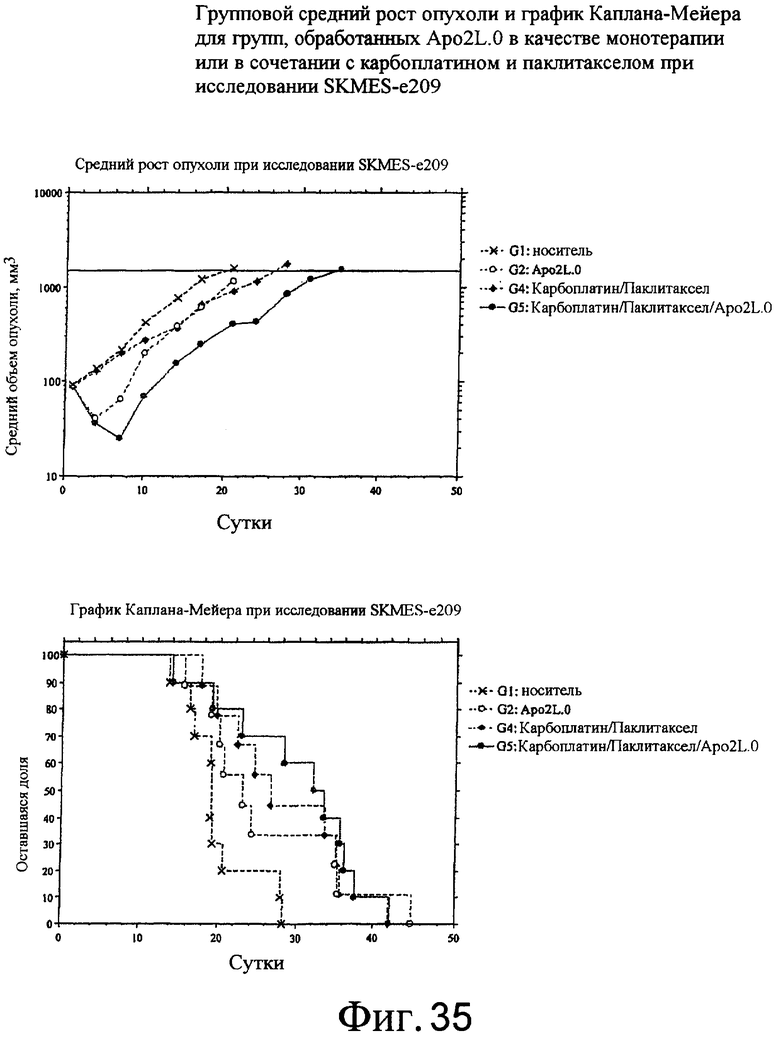
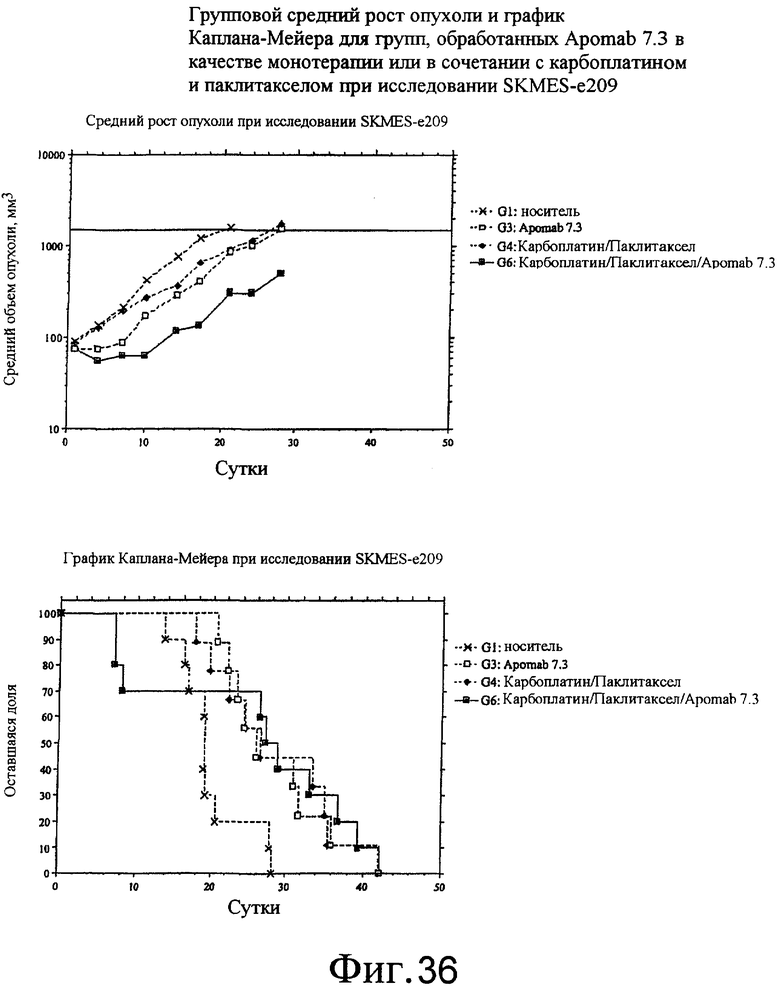
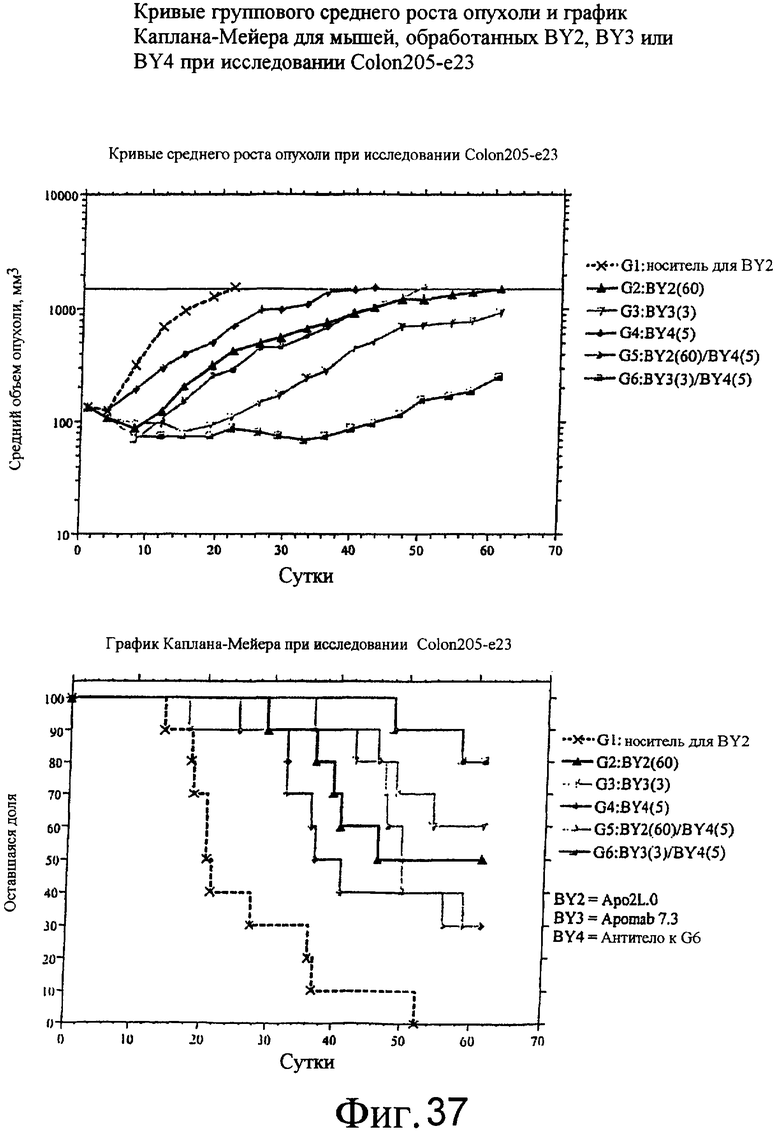
Изобретение относится к биотехнологии. Описано антитело к DR5 или его антигенсвязывающий фрагмент, включающее тяжелую и легкую цепи, где указанное антитело содержит по меньшей мере одну замену в последовательности полноразмерного антитела 16Е2. Представлена фармацевтическая композиция для лечения рака, содержащая описанное антитело. Предложен способ индукции апоптоза, включающий в себя воздействие на злокачественные клетки млекопитающих описанного антитела или его антигенсвязывающего фрагмента. Предложен способ лечения злокачественной опухоли, включающий в себя введение млекопитающему эффективного количества описанного антитела или его антигенсвязывающего фрагмента. Изобретение расширяет арсенал средств, направленных против рака. 11 н. и 34 з.п. ф-лы, 37 ил., 13 табл., 10 пр.
1. Антитело к DR5, включающее тяжелую и легкую цепи, где указанное антитело содержит по меньшей мере одну замену в последовательности полноразмерного антитела 16Е2 (SEQ ID NO: 11 и 13 соответственно), где замена в тяжелой цепи выбрана из группы, состоящей из:
(a) M34L, N53Q, L102Y;
(b) M34L, N53Y, L102Y;
(c) G33A, N53Q, L1-2Y;
(d) G33A, N53Y, L102Y;
(e) M34L, N53Q, L102Y;
(f) M34L, N53Y, L102Y;
(g) G33A, N53Q, L102Y;
(f) G33A, N53Y, L102Y;
(g) T28A, N53Q, L102Y; и
замена в легкой цепи выбрана из группы, состоящей из:
(h) Q24S, G50K, K51D, N52S, N53, H95bY;
(i) Q24S, K51A, D92S, S93Y;
(j) Q24S, K51A, R91A,
или его антигенсвязывающий фрагмент,
при этом расположение аминокислотных замен указана по нумерации Kabat.
2. Антитело к DR5 по п.1, обладающее большей аффинностью к DR5, чем указанное полноразмерное антитело 16Е2, и/или проявляет большую биологическую активность и/или эффективность, чем указанное полноразмерное антитело 16Е2 или его антигенсвязывающий фрагмент.
3. Антитело к DR5 по п.1 или 2, где указанное антитело, по существу, связывается с тем же эпитопом, что и полноразмерное антитело 16Е2 или его антигенсвязывающий фрагмент.
4. Антитело к DR5 по п.1, несущее один или несколько наборов мутаций в каркасной области вариабельного домена тяжелой цепи полноразмерного антитела 16Е2 (SEQ ID NO: 11), где мутация в каркасной области выбрана из группы, состоящей из
(i) Q6E, V11L, E12V, R13Q, K105Q,
(ii) Q6E, V11L, E12V, R13Q,
(iii) Q6E, K105Q;
(iv) V11L, E12V, R13Q;
(v) K105Q;
(vi) Q6E;
(vii) VHL;
(viii) R13Q;
(ix) E12V, R13Q;
(x) E12V;
(xi) E12N;
(xii) VHL, R12V,
или его антигенсвязывающий фрагмент,
где расположение мутаций в каркасной области указана по нумерации Kabat.
5. Антитело к DR5 по п.1, где антитело выбрано из группы, состоящей из 5.2, 5.3, 6.2, 7.2, 7.3, 8.3, 23.3, 2.3, или его антигенсвязывающий фрагмент.
6. Антитело к DR5 по п.5, где антитело представляет собой антитело 7.3 или его антигенсвязывающий фрагмент.
7. Антитело к DR5 по п.1, 5 или 6, где указанный антигенсвязывающий фрагмент выбран из группы, состоящей из фрагментов Fab, Fab', F(ab')2, и Fv, димерных антител, молекул одноцепочечных антител и полиспецифических антител, формирующихся из фрагментов антител.
8. Антитело к DR5 по п.7, где указанное антитело представляет собой одноцепочечное антитело.
9. Антитело к DR5 по п.7, где указанный антигенсвязывающий фрагмент представляет собой фрагмент Fv.
10. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.2, где указанная биологическая активность представляет собой противораковую активность.
11. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.2, где указанная биологическая активность представляет собой активацию или стимуляцию апоптоза в злокачественных клетках.
12. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.10 или 11, где злокачественная опухоль выбрана из группы, состоящей из карциномы, лимфомы, бластомы, саркомы и лейкоза.
13. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.10 или 11, где указанная злокачественная опухоль выбрана из группы, состоящей из плоскоклеточного рака, мелкоклеточного рака легких, немелкоклеточного рака легких (NSCLC), неходжкинской лимфомы, бластомы, желудочно-кишечного рака, рака почек, рака яичников, рака печени, рака желудка, рака мочевого пузыря, гепатомы, рака молочной железы, рака толстой кишки, колоректального рака, рака поджелудочной железы, карциномы эндометрия, карциномы слюнной железы, рака почки, рака печени, рака предстательной железы, рака женских наружных половых органов, рака щитовидной железы, печеночной карциномы и рака головы и шеи.
14. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.13, где указанная злокачественная опухоль представляет собой NSCLC, неходжкинскую лимфому, колоректальный рак или рак поджелудочной железы.
15. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.12, где указанная злокачественная опухоль представляет собой аденокарциному.
16. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.15, где указанная аденокарцинома представляет собой колоректальную аденокарциному, аденокарциному поджелудочной железы или метастазирующую аденокарциному.
17. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.2, где указанную эффективность определяют в анализе уничтожения опухоли in vitro.
18. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.1, которые представляют собой химерное, гуманизированное или человеческое антитело.
19. Антитело или антигенсвязывающий фрагмент антитела к DR5 по п.1, опосредующие обусловленную антителами клеточную цитотоксичность (ADCC).
20. Выделенная нуклеиновая кислота, кодирующая тяжелую цепь антитела к DR5 по п.1, 5 или 6, или антигенсвязывающий фрагмент указанной тяжелой цепи.
21. Выделенная нуклеиновая кислота, кодирующая легкую цепь антитела к DR5 по п.1, 5 или 6, или антигенсвязывающий фрагмент указанной легкой цепи.
22. Вектор экспрессии, содержащий нуклеиновую кислоту по п.20.
23. Вектор экспрессии, содержащий нуклеиновую кислоту по п.21.
24. Клетка-хозяин, содержащая нуклеиновую кислоту по п.20 и 21, и способная к их экспрессии.
25. Способ получения антитела к DR5 или его антигенсвязывающего фрагмента, включающий в себя культивирование клетки-хозяина по п.24, в условиях, в которых экспрессируются нуклеиновые кислоты.
26. Фармацевтическая композиция для лечения рака, содержащая антитело к DR5 по п.1, 5 или 6, или его антигенсвязывающий фрагмент, и фармацевтически приемлемый носитель.
27. Композиция по п.26, содержащая, кроме того, дополнительное противораковое средство.
28. Композиция по п.27, где указанное дополнительное противораковое средство представляет собой антитело.
29. Композиция по п.28, где указанное антитело выбрано из группы, состоящей из дополнительного антитела к DR5, ритуксимаба и антитела к VEGF.
30. Композиция по п.27, где указанное дополнительное противораковое средство представляет собой химиотерапевтическое средство.
31. Композиция по п.30, где указанное химиотерапевтическое средство выбрано из группы, состоящей из СРТ-11 (иринотекана), гемцитабина, карбоплатина, таксола и паклитаксела.
32. Композиция по п.27, где указанное дополнительное противораковое средство представляет собой лиганд Apo2L, содержащий аминокислоты 114-281 согласно фиг.1 (SEQ ID NO: 1).
33. Способ индукции апоптоза, включающий в себя воздействие на злокачественные клетки млекопитающих антитела к DR5 по п.1, 5 или 6, или его антигенсвязывающего фрагмента.
34. Способ по п.33, где указанное антитело представляет собой химерное, гуманизированное или человеческое антитело.
35. Способ по п.33, где указанные злокачественные клетки млекопитающих подвергают воздействию средства, активирующего DR5.
36. Способ лечения злокачественной опухоли, включающий в себя введение млекопитающему эффективного количества антитела к DR5 по п.1, 5 или 6, или его антигенсвязывающего фрагмента.
37. Способ по п.36, где указанное млекопитающее представляет собой человека.
38. Способ по п.37, где указанная злокачественная опухоль выбрана из группы, состоящей из плоскоклеточного рака, мелкоклеточного рака легких, немелкоклеточного рака легких (NSCLC), неходжкинской лимфомы, бластомы, желудочно-кишечного рака, рака почек, рака яичников, рака печени, рака желудка, рака мочевого пузыря, гепатомы, рака молочной железы, рака толстой кишки, колоректального рака, рака поджелудочной железы, карциномы эндометрия, карциномы слюнной железы, рака почки, рака печени, рака предстательной железы, рака женских наружных половых органов, рака щитовидной железы, печеночной карциномы и рака головы и шеи.
39. Способ по п.38, где указанная злокачественная опухоль представляет собой NSCLC, колоректальный рак, неходжкинскую лимфому или рак поджелудочной железы.
40. Способ по п.36, где указанная злокачественная опухоль представляет собой аденокарциному.
41. Способ по п.40, где указанная аденокарцинома представляет собой колоректальную аденокарциному, аденокарциному поджелудочной железы или метастазирующую аденокарциному.
42. Способ по любому из пп.36-41, дополнительно включающий в себя введение дополнительного противоракового средства.
43. Набор для лечения рака, включающий в себя контейнер и композиции, содержащиеся в указанном контейнере, где композиция включает антитело к DR5 по п.1, 5 или 6, или его антигенсвязывающий фрагмент.
44. Набор по п.43, дополнительно содержащее инструкции по применению антитела к DR5 in vitro или in vivo.
45. Набор по п.44, где указанные инструкции относятся к лечению злокачественной опухоли.
| US 6113898, 05.09.2000 | |||
| РЫБАЛЬСКИЙ Н.Г., СЕРОВА М.А, ИГНАТЬЕВА Г.А, СТАРЧЕУС А.П | |||
| Моноклональные антитела и гибридомы | |||
| - М.: ВАСХНИЛ, 1989. |

