РОДСТВЕННЫЕ ЗАЯВКИ
По этой заявке испрашивается приоритет по предварительной заявке США номер 60/708677, поданной 16 августа 2005, и по предварительной заявке США номер 60/808076, поданной 24 мая 2006, содержания которых включены здесь в качестве ссылки.
ОБЛАСТЬ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Описанные здесь изобретения относятся к способам и анализам для выявления биомаркеров, прогнозирующих чувствительность клеток млекопитающих к Apo2L/TRAIL и/или антителам-агонистам рецепторов смерти. Более конкретно, представленные здесь изобретения относятся к способам и анализам, которые выявляют молекулы, связанные с семейством белков GalNac-T, которые прогнозируют чувствительность злокачественных клеток млекопитающих к Apo2L/TRAIL или антителам-агонистам рецепторов смерти, таким как антитела-агонисты DR4 или DR5.
ПРЕДПОСЫЛКИ К СОЗДАНИЮ ИЗОБРЕТЕНИЯ
В уровне техники были идентифицированы различные лиганды и рецепторы, принадлежащие к суперсемейству факторов некроза опухоли (TNF). В число таких лигандов включены фактор некроза опухоли альфа («TNF-альфа»), фактор некроза опухоли бета («TNF-бета» или «лимфотоксин-альфа»), лимфотоксин бета («LT-бета»), лиганд CD30, лиганд CD27, лиганд CD40, лиганд OX-40, лиганд 4-1BB, LIGHT, лиганд Apo-1 (также называемый Fas лигандом или лигандом CD95), лиганд Apo-2 (также называемый Apo2L или TRAIL), лиганд Apo-3 (также называемый TWEAK), APRIL, лиганд OPG (также называемый RANK лигандом, ODF или TRANCE), и TALL-1 (также называемый BlyS, BAFF или THANK) (См., например, Ashkenazi, Nature Review, 2:420-430 (2002); Ashkenazi and Dixit, Science, 281:1305-1308 (1998); Ashkenazi and Dixit, Curr. Opin. Cell Biol., 11:255-260 (2000); Golstein, Curr. Biol., 7:750-753 (1997) Wallach, Cytokine Reference, Academic Press, 2000, страницы 377-411; Locksley et al., Cell, 104:487-501 (2001); Gruss and Dower, Blood, 85:3378-3404 (1995); Schmid et al., Proc. Natl. Acad. Sci., 83:1881 (1986); Dealtry et al., Eur. J. Immunol., 17:689 (1987); Pitti et al., J. Biol. Chem., 271:12687-12690 (1996); Wiley et al., Immunity, 3:673-682 (1995); Browning et al., Cell, 72:847-856 (1993); Armitage et al. Nature, 357:80-82 (1992), WO 97/01633, опубликована 16 января 1997; WO 97/25428, опубликована 17 июля 1997; Marsters et al., Curr. Biol., 8:525-528 (1998); Chicheportiche et al., Biol. Chem., 272:32401-32410 (1997); Hahne et al., J. Exp. Med., 188:1185-1190 (1998); WO 98/28426, опубликована 2 июля 1998; WO 98/46751, опубликована 22 октября 1998; WO 98/18921, опубликована 7 мая 1998; Moore et al., Science, 285:260-263 (1999); Shu et al., J. Leukocyte Biol., 65:680 (1999); Schneider et al., J. Exp. Med., 189:1747-1756 (1999); Mukhopadhyay et al., J. Biol. Chem., 274:15978-15981 (1999)).
Индукция различных клеточных ответов, опосредованных такими лигандами семейства TNF, обычно начинается с их связывания со специфическими клеточными рецепторами. Некоторые, но не все, лиганды семейства TNF связываются с «рецепторами смерти» клеточной поверхности и посредством этого индуцируют различную биологическую активность, для активации каспаз, или ферментов, которые осуществляют процессы клеточной гибели или апоптоз (Salvesen et al., Cell, 91:443-446 (1997). В число представителей суперсемейства рецепторов TNF, идентифицированных к настоящему времени, включены TNFR1, TNFR2, TACI, GITR, CD27, OX-40, CD30, CD40, HVEM, Fas (также называемый Apo-1 или CD95), DR4 (также называемый TRAIL-R1), DR5 (также называемый Apo-2 или TRAIL-R2), DcR1, DcR2, остеопротегерин (OPG), RANK и Apo-3 (также называемый DR3 или TRAMP) (см., например, Ashkenazi, Nature Reviews, 2:420-430 (2002); Ashkenazi and Dixit, Science, 281:1305-1308 (1998); Ashkenazi and Dixit, Curr. Opin. Cell Biol., 11:255-260 (2000); Golstein, Curr. Biol., 7:750-753 (1997) Wallach, Cytokine Reference, Academic Press, 2000, страницы 377-411; Locksley et al., Cell, 104:487-501 (2001); Gruss and Dower, Blood, 85:3378-3404 (1995); Hohman et al., J. Biol. Chem., 264:14927-14934 (1989); Brockhaus et al., Proc. Natl. Acad. Sci., 87:3127-3131 (1990); EP 417563, опубликованную 20 марта 1991; Loetscher et al., Cell, 61:351 (1990); Schall et al., Cell, 61:361 (1990); Smith et al., Science, 248:1019-1023 (1990); Lewis et al., Proc. Natl. Acad. Sci., 88:2830-2834 (1991); Goodwin et al., Mol. Cell. Biol., 11:3020-3026 (1991); Stamenkovic et al., EMBO J., 8:1403-1410 (1989); Mallett et al., EMBO J., 9:1063-1068 (1990); Anderson et al., Nature, 390:175-179 (1997); Chicheportiche et al., J. Biol. Chem., 272:32401-32410 (1997); Pan et al., Science, 276:111-113 (1997); Pan et al., Science, 277:815-818 (1997); Sheridan et al., Science, 277:818-821 (1997); Degli-Esposti et al., J. Exp. Med., 186:1165-1170 (1997); Marsters et al., Curr. Biol., 7:1003-1006 (1997); Tsuda et al., BBRC, 234:137-142 (1997); Nocentini et al., Proc. Natl. Acad. Sci., 94:6216-6221 (1997); vonBulow et al., Science, 278:138-141 (1997)).
Большинство из этих представителей семейства рецепторов TNF имеют общую характерную структуру рецепторов клеточной поверхности, включая внеклеточные, трансмембранные и внутриклеточные области, тогда как другие встречаются в природе в виде растворимых белков, лишенных трансмембранного и внутриклеточного домена. Внеклеточная часть характерных TNFR содержит повторяющуюся структуру аминокислотной последовательности множественных доменов, богатых цистеином (CRD), начиная с NH2-конца.
Лиганд, называемый Apo-2L или TRAIL, был обнаружен несколько лет назад как представитель TNF семейства цитокинов (см., например, Wiley et al., Immunity, 3:673-682 (1995); Pitti et al., J. Biol. Chem., 271:12697-12690 (1996); WO 97/01633; WO 97/25428; патент США 5763223, выданный 9 июня 1998; патент США 6284236, выданный 4 сентября 2001). Полная длина нативной последовательности Apo2L/TRAIL полипептида человека составляет в длину 281 аминокислоту, II тип трансмембранного белка. Некоторые клетки могут продуцировать природную растворимую форму этого полипептида посредством ферментативного расщепления внеклеточной области этого полипептида (Mariani et al., J. Cell. Biol., 137:221-229 (1997)). Кристаллографические исследования растворимых форм Apo2L/TRAIL демонстрируют гомотримерную структуру, сходную со структурами TNF, и других родственных белков (Hymowitz et al., Molec. Cell, 4:563-571 (1999); Cha et al., Immunity, 11:253-261 (1999); Mongkolsapaya et al., Nature Structural Biology, 6:1048 (1999); Hymowitz et al., Biochemistry, 39:633-644 (2000)). Однако было обнаружено, что Apo2L/TRAIL, в отличие от других представителей семейства TNF, обладает уникальной структурной особенностью в том, что три остатка цистеина (в положении 230 каждой субъединицы в гомотримере) вместе координируют атом цинка и что связывание цинка является важным для стабильности триммера и биологической активности (Hymowitz et al., supra; Bodmer et al., J. Biol. Chem., 275:20632-20637 (2000)).
В литературных источниках сообщалось, что Apo2L/TRAIL может играть роль в модуляции иммунной системы, включая аутоиммунные заболевания, такие как ревматоидный артрит [см., например, Thomas et al., J. Immunol., 161:2195-2200 (1998); Johnsen et al., Cytokine, 11:664-672 (1999); Griffith et al., J. Exp. Med., 189:1343-1353 (1999); Song et al., J. Exp. Med., 191:1095-1103 (2000)].
Также сообщалось, что растворимые формы Apo2L/TRAIL индуцируют апоптоз в различных злокачественных клетках, в том числе в опухолях кишечника, легких, груди, предстательной железы, мочевого пузыря, почек, яичников и головного мозга, а также меланоме, при лейкозе и множественной миеломе (см., например, Wiley et al., supra; Pitti et al., supra; патент США 6030945, выданный 29 февраля 2000; патент США 6746668, выданный 8 июня 2004; Rieger et al., FEBS Letters, 427:124-128 (1998); Ashkenazi et al., J. Clin. Invest., 104:155-162 (1999); Walczak et al., Nature Med., 5:157-163 (1999); Keane et al., Cancer Research, 59:734-741 (1999); Mizutani et al., Clin. Cancer Res., 5:2605-2612 (1999); Gazitt, Leukemia, 13:1817-1824 (1999); Yu et al., Cancer Res., 60:2384-2389 (2000); Chinnaiyan et al., Proc. Natl. Acad. Sci., 97:1754-1759 (2000)). In vivo исследования на мышиных моделях опухолей дополнительно дают возможность предположить, что Apo2L/TRAIL, отдельно или в сочетании с химиотерапией или лучевой терапией, может оказывать значительные противоопухолевые воздействия (см., например, Ashkenazi et al., supra; Walzcak et al., supra; Gliniak et al., Cancer Res., 59:6153-6158 (1999); Chinnaiyan et al., supra; Roth et al., Biochem. Biophys. Res. Comm., 265:1999 (1999); заявку РСТ US/00/15512; заявку РСТ US/01/23691). В отличие от многих типов злокачественных клеток, у человека большинство нормальных клеточных типов являются резистентными к индукции апоптоза под действием определенных рекомбинантных форм Apo2L/TRAIL (Ashkenazi et al., supra; Walzcak et al., supra). Jo et al. сообщал о том, что растворимая форма Apo2L/TRAIL, меченая полигистидином, индуцировала апоптоз in vitro в нормальных выделенных гепатоцитах человека, но не в гепатоцитах, не принадлежащих человеку (Jo et al., Nature Med., 6:564-567 (2000); см. также Nagata, Nature Med., 6:502-503 (2000)). Считается, что некоторые рекомбинантные препараты Apo2L/TRAIL могут варьировать, исходя из биологических свойств и биологических активностей в отношении патологических клеток по сравнению с нормальными, в зависимости, например, от наличия или отсутствия таг-молекулы, содержания цинка и % содержания тримера (См., Lawrence et al., Nature Med., Letter to the Editor, 7:383-385 (2001); Qin et al., Nature Med., Letter to the Editor, 7:385-386 (2001)).
Было обнаружено, что Apo2L/TRAIL связывается по меньшей мере с пятью различными рецепторами. По меньшей мере два из рецепторов, которые связывают Apo2L/TRAIL, содержат функциональный, цитоплазматический домен смерти. Один такой рецептор был назван «DR4» (и, альтернативно, TR4 или TRAIL-R1) (Pan et al., Science, 276:111-113 (1997); см. также WO 98/32856, опубликованную 30 июля 1998; WO 99/37684, опубликованную 29 июля 1999; WO 00/73349, опубликованную 7 декабря 2000; US 2003/0036168, опубликованную 20 февраля 2003; US 6433147, выданный 13 августа 2002; US 6461823, выданный 8 октября 2002, и US 6342383, выданный 29 января 2002).
Другой рецептор для Apo2L/TRAIL был назван DR5 (альтернативно он также был назван Apo-2; TRAIL-R или TRAIL-R2, TR6, Tango-63, hAPO8, TRICK2 или KILLER) (см., например, Sheridan et al., Science, 277:818-821 (1997), Pan et al., Science, 277:815-818 (1997), WO 98/51793, опубликованную 19 ноября 1998; WO 98/41629, опубликованную 24 сентября 1998; Screaton et al., Curr. Biol., 7:693-696 (1997); Walczak et al., EMBO J., 16:5386-5387 (1997); Wu et al., Nature Genetics, 17:141-143 (1997); WO 98/35986, опубликованную 20 августа 1998; EP 870827, опубликованную 14 октября 1998; WO 98/46643, опубликованную 22 октября 1998; WO 99/02653, опубликованную 21 января 1999; WO 99/09165, опубликованную 25 февраля 1999; WO 99/11791, опубликованную 11 марта 1999; WO 03/042367, опубликованную 22 мая 2003; WO 02/097033, опубликованную 5 декабря 2002; WO 03/038043, опубликованную 8 мая 2003; US 2002/0072091, опубликованную 13 августа 2002; US 2002/0098550, опубликованную 7 декабря 2001; US 6313269, выданный 6 декабря 2001; US 2001/0010924, опубликованную 2 августа 2001; US 2003/01255540, опубликованную 3 июля 2003; US 2002/0160446, опубликованную 31 октября 2002, US 2002/0048785, опубликованную 25 апреля 2002; US 2004/0141952, опубликованную 22 июля 2004; US 2005/0129699, опубликованную 16 июня 2005; US 2005/0129616, опубликованную 16 июня 2005; US 6342369, выданный в феврале 2002; US 6569642, выданный 27 мая 2003, US 6072047, выданный 6 июня 2000, US 6642358, выданный 4 ноября 2003; US 6743625, выданный 1 июня 2004). Подобно DR4, сообщалось, что DR5 содержит цитоплазматический домен смерти и способен передавать сигнал апоптоза при связывании с лигандом (или при связывании с молекулой, такой как антитело-агонист, которое имитирует активность этого лиганда). Кристаллическая структура комплекса, образованного между Apo-2L/TRAIL и DR5, описана у Hymowitz et al., Molecular Cell, 4:563-571 (1999).
При связывании с лигандом как DR4, так и DR5 могут запускать апоптоз, независимо вовлекая и активируя инициатор апоптоза, каспазу-8 посредством молекулы-адаптора, содержащей домен смерти, названной FADD/Mort1 [Kischkel et al., Immunity, 12:611-620 (2000); Sprick et al., Immunity, 12:599-609 (2000); Bodmer et al., Nature Cell Biol., 2:241-243 (2000)].
Сообщалось, что Apo2L/TRAIL также связывается с рецепторами, называемыми DcR1, DcR2 и OPG, которые предположительно функционируют как ингибиторы, а не трансдукторы передачи сигнала (см., например, DCR1 (также называемый TRID, LIT или TRAIL-R3) [Pan et al., Science, 276:111-113 (1997); Sheridan et al., Science, 277:818-821 (1997); McFarlane et al., J. Biol. Chem., 272:25417-25420 (1997); Schneider et al., FEBS Letters, 416:329-334 (1997); Degli-Esposti et al., J. Exp. Med., 186:1165-1170 (1997); и Mongkolsapaya et al., J. Immunol., 160:3-6 (1998); DCR2 (также называемый TRUNDD или TRAIL-R4) [Marsters et al., Curr. Biol., 7:1003-1006 (1997); Pan et al., FEBS Letters, 424:41-45 (1998); Degli-Esposti et al., Immunity, 7:813-820 (1997)], и OPG [Simonet et al., supra]. В отличие от DR4 и DR5, рецепторы DcR1 и DcR2 не дают сигнала к апоптозу.
В литературных источниках сообщалось об определенных антителах, которые связываются с рецепторами DR4 и/или DR5. Например, анти-DR4 антитела к рецептору DR4 и обладающие агонистической или апоптотической активностью в некоторых клетках млекопитающих описаны, например, в WO 99/37684, опубликованной 29 июля 1999; WO 00/73349, опубликованной 12 июля 2000; WO 03/066661, опубликованной 14 августа 2003. Также, см., например, Griffith et al., J. Immunol., 162:2597-2605 (1999); Chuntharapai et al., J. Immunol., 166:4891-4898 (2001); WO 02/097033, опубликованную 2 декабря 2002; WO 03/042367, опубликованную 22 мая 2003; WO 03/038043, опубликованную 8 мая 2003; WO 03/037913, опубликованную 8 мая 2003; US 2003/0073187, опубликованную 17 апреля 2003; US 2003/0108516, опубликованную 12 июня 2003. Также были описаны некоторые анти-DR5 антитела, см., например, WO 98/51793, опубликованную 8 ноября 1998; Griffith et al., J. Immunol., 162:2597-2605 (1999); Ichikawa et al., Nature Med., 7:954-960 (2001); Hylander et al., “An Antibody to DR5 (TRAIL-Receptor 2) Suppresses the Growth of Patient Derived Gastrointestinal Tumors Grown in SCID mice”, Abstract, 2d International Congress on Monoclonal Antibodies in Cancers, Aug. 29-Sept. 1, 2002, Banff, Alberta, Canada; WO 03/038043, опубликованную 8 мая 2003; WO 03/037913, опубликованную 8 мая 2003; US 2003/0180296, опубликованную 25 сентября 2003. Кроме того, были описаны некоторые антитела, обладающие перекрестной реактивностью в отношении как DR4, так и DR5 рецепторов (см., например, патент США 6252050, выданный 26 июня 2001).
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Описанное здесь изобретение относится к способам и анализам для исследования экспрессии одного или нескольких биомаркеров в образце ткани млекопитающих или клеточном образце, в которых экспрессия одного или нескольких таких биомаркеров прогнозирует чувствительность образца ткани или клеточного образца к таким агентам, как Apo2L/TRAIL или антителам-агонистам к DR5. В различных вариантах осуществления этого изобретения способы и анализы исследования экспрессии молекул в GalNac-T семействе белков, в частности GalNAc-T14 или GalNAc-T3.
Как рассматривалось выше, у человека большинство нормальных клеточных типов проявляет резистентность к индукции апоптоза под действием некоторых рекомбинантных форм Apo2L/TRAIL (Ashkenazi et al., supra; Walzcak et al., supra). Также было отмечено, что некоторые популяции патологических типов клеток человека (например, некоторые популяции злокачественных клеток) являются резистентными к индукции апоптоза под действием некоторых рекомбинантных форм Apo2L/TRAIL (Ashkenazi et al., J. Clin. Invest., 1999, supra; Walczak et al., Nature Med., 1999, supra). Следовательно, исследуя образец ткани млекопитающего или клеточный образец на экспрессию выбранных биомаркеров путем анализа, можно удобно и эффективно получить информацию, полезную для оценки соответствующих или эффективных способов лечения пациентов. Например, информация, полученная в результате анализа, направленного на выявление экспрессии GalNac-T14 в образце ткани или клеточном образце млекопитающего, может предоставить лечащему врачу полезные сведения, которые могут быть использованы для определения оптимальной схемы лечения (с использованием Apo2L/TRAIL или антител-агонистов рецепторов смерти) для пациентов, страдающих таким заболеванием, как рак, или заболеванием, связанным с иммунной системой, таким как аутоиммунное заболевание.
Изобретение относится к способам прогнозирования чувствительности образца ткани или клеток млекопитающих (например, злокачественных клеток) к Apo2L/TRAIL или антителу-агонисту рецептора смерти. В некоторых вариантах осуществления способы включают в себя получение образца ткани или клеток млекопитающего и исследование ткани или клетки на экспрессию GalNac-T14. Способы можно проводить в различных форматах, включая анализы, определяющие экспрессию мРНК и/или белка, анализы ферментативной активности и другие, рассмотренные здесь способы. Определение экспрессии GalNac-T14 в указанных тканях или клетках будет прогнозировать то, что такие ткани или клетки будут чувствительными к апоптоз-индуцируемой активности Apo2L/TRAIL и/или антителу рецептора смерти. В необязательных вариантах осуществления ткани или клетки также могут быть исследованы на экспрессию рецепторов DR4, DR5, DcR1 или DcR2.
Дополнительные способы по изобретению включают в себя способы индукции апоптоза в образце ткани или клеток млекопитающего, включающие в себя стадии получения образца ткани или клеток млекопитающего, исследование ткани или клетки на экспрессию GalNac-T14, и при определении, что указанный образец ткани или клеток экспрессирует GalNac-T14, воздействие на указанный образец ткани или клеток эффективным количеством Apo2L/TRAIL или антителом-агонистом рецептора смерти. Стадии в способах исследования экспрессии GalNac-T14 можно проводить в различных аналитических форматах, включая исследования определения экспрессии мРНК и/или белка, ферментативной активности и другие, рассмотренные здесь способы. В необязательных вариантах осуществления способы также включают в себя исследование образца ткани или клеток на экспрессию рецепторов DR4, DR5, DcR1 или DcR2. Необязательно образец ткани или клеток содержит злокачественную ткань или клетки. Необязательно, образец ткани или клеток содержит клетки немелкоклеточного рака легких, клетки злокачественной опухоли поджелудочной железы, клетки рака груди или клетки неходжкинской лимфомы.
Другие способы по изобретению включают в себя способы лечения заболевания у млекопитающего, такого как заболевание, связанное с иммунной системой, или рак, предусматривающие стадии получения образца ткани или клеток этого млекопитающего, исследование ткани или клеток на экспрессию GalNac-T14, и при определении, что указанный образец ткани или клеток экспрессирует GalNac-T14, введение указанному млекопитающему эффективного количества Apo2L/TRAIL или антитела-агониста рецептора смерти. Стадии в этих способах исследования экспрессии одного или нескольких биомаркеров можно проводить в различных аналитических форматах, включая исследование определения экспрессии мРНК и/или белка, ферментативной активности и другие описанные здесь способы. В необязательных вариантах осуществления способы также включают в себя исследование образца ткани или клеток на экспрессию рецепторов DR4, DR5, DcR1, или DcR2. Необязательно способы включают в себя лечение злокачественного заболевания млекопитающего. Необязательно эти способы дополнительно к введению эффективного количества Apo2L/TRAIL и/или антитела-агониста рецептора смерти включают в себя введение указанному млекопитающему химиотерапевтических средств(а) или применение лучевой терапии.
В дополнительных вариантах осуществления упомянутые выше способы могут включать в себя исследование ткани или клеток млекопитающего на экспрессию других GalNac-T молекул, таких как GalNac-T3.
Другие варианты осуществления иллюстрируются в качестве примера следующими пунктами:
1. Способ прогнозирования чувствительности образца ткани или клеток млекопитающего к Apo2L/TRAIL, включающий в себя стадии:
получения образца ткани или клеток млекопитающего;
исследования образца ткани или клеток млекопитающего для выявления экспрессии GalNac-T14, где экспрессия указанной GalNac-T14 прогнозирует чувствительность указанного образца ткани или клеток к апоптоз-индуцирующей активности Apo2L/TRAIL.
2. Способ по пункту 1, в котором указанную экспрессию GalNac-T14 исследуют путем определения экспрессии мРНК GalNac-T14.
3. Способ по пункту 1, в котором указанную экспрессию GalNac-T14 исследуют с помощью иммуногистохимии.
4. Способ по пункту 1, дополнительно включающий в себя стадию исследования экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанном образце ткани или клеток.
5. Способ по пункту 1, в котором образец ткани или клеток содержит злокачественную ткань или клетки.
6. Способ по пункту 5, в котором указанные злокачественные клетки представляют собой клетки или ткань рака поджелудочной железы, лимфомы или немелкоклеточного рака легких.
7. Способ индукции апоптоза в образце ткани или клеток млекопитающих, включающий в себя стадии:
получения образца ткани или клеток млекопитающего;
исследования образца ткани или клеток для выявления экспрессии GalNac-T14, и
после выявления экспрессии указанной GalNac-T14 воздействие на указанный образец ткани или клеток эффективным количеством Apo2L/TRAIL.
8. Способ по пункту 7, в котором указанную экспрессию GalNac-T14 исследуют путем тестирования экспрессии мРНК GalNac-T14.
9. Способ по пункту 7, в котором указанную экспрессию GalNac-T14 исследуют с помощью иммуногистохимии.
10. Способ по пункту 7, дополнительно включающий стадию исследования экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанном образце ткани или клеток.
11. Способ по пункту 7, в котором указанный образец ткани или клеток содержит злокачественную ткань или клетки.
12. Способ по пункту 11, в котором указанные злокачественные клетки представляют собой злокачественные клетки или ткань рака поджелудочной железы, лимфомы или немелкоклеточного рака легких.
13. Способ по пункту 7, в котором на указанные клетки воздействуют эффективным количеством полипептида Apo2L/TRAIL, содержащего аминокислоты 114-281, представленные на Фиг.1.
14. Способ лечения заболевания у млекопитающего, например заболевания, связанного с иммунной системой, или злокачественного заболевания, включающий в себя стадии:
получения образца ткани или клеток указанного млекопитающего;
исследования образца ткани или клеток для определения экспрессии GalNac-T14, и
после определения экспрессии указанной GalNac-T14 введение указанному млекопитающему эффективного количества Apo2L/TRAIL.
15. Способ по пункту 14, в котором указанную экспрессию GalNac-T14 исследуют путем определения экспрессии мРНК GalNac-T14.
16. Способ по пункту 14, в котором указанную экспрессию GalNac-T14 исследуют с помощью иммуногистохимии.
17. Способ по пункту 14, дополнительно включающий в себя стадию исследования экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанной ткани или клетке.
18. Способ по пункту 14, в котором образец ткани или клеток содержит злокачественную ткань или клетки.
19. Способ по пункту 18, в котором указанные злокачественные клетки или ткань содержат клетки или ткань рака поджелудочной железы, лимфомы или немелкоклеточного рака легких.
20. Способ по пункту 14, в котором эффективное количество полипептида Apo2L/TRAIL, содержащего аминокислоты 114-281, представленные на Фиг.1, вводят указанному млекопитающему.
21. Способ по пункту 14, в котором указанному млекопитающему также вводят химиотерапевтическое средство (средства) или применяют лучевую терапию.
22. Способ по пункту 14, в котором указанному млекопитающему также вводят цитокин, цитотоксическое средство или ингибитор роста.
23. Способ по пункту 7, в котором указанный полипептид Apo2L/TRAIL связан с молекулой полиэтиленгликоля.
24. Способ по пункту 14, где указанный полипептид Apo2L/TRAIL связан с молекулой полиэтиленгликоля.
25. Способ прогнозирования чувствительности образца ткани или клеток млекопитающего к антителам рецептора смерти, включающий в себя стадии:
получения образца ткани или клеток млекопитающего;
исследования образца ткани или клеток для определения экспрессии GalNac-T14, где экспрессия указанной GalNac-T14 прогнозирует чувствительность указанного образца ткани или клеток к апоптоз-индуцирующей активности антител к рецептору смерти.
26. Способ по пункту 25, в котором указанную экспрессию GalNac-T14 исследуют путем определения экспрессии мРНК GalNac-T14.
27. Способ по пункту 25, в котором указанную экспрессию GalNac-T14 исследуют с помощью иммуногистохимии.
28. Способ по пункту 25, дополнительно включающий в себя стадию исследования экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанном образце ткани или клеток.
29. Способ по пункту 25, в котором образец ткани или клеток содержит ткань злокачественной опухоли или злокачественные клетки.
30. Способ по пункту 29, в котором указанные злокачественные клетки представляют собой злокачественные клетки или злокачественную ткань опухоли поджелудочной железы, лимфомы или немелкоклеточного рака легких.
31. Способ по пункту 25, в котором антитела к рецептору смерти представляют собой анти-DR4 или анти-DR5 антитела.
32. Способ индукции апоптоза в образце ткани или клеток млекопитающих, включающий в себя стадии:
получения образца ткани или клеток млекопитающего;
исследования образца ткани или клеток для выявления экспрессии GalNac-T14, и
после выявления экспрессии указанной GalNac-T14 воздействие на указанный образец ткани или клеток эффективным количеством антитела к рецептору смерти.
33. Способ по пункту 32, в котором указанную экспрессию GalNac-T14 исследуют путем тестирования экспрессии мРНК GalNac-T14.
34. Способ по пункту 32, в котором указанную экспрессию GalNac-T14 исследуют с помощью иммуногистохимии.
35. Способ по пункту 32, дополнительно включающий стадию исследования экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанном образце ткани или клеток.
36. Способ по пункту 32, в котором указанный образец ткани или клеток содержит злокачественную ткань или клетки.
37. Способ по пункту 36, в котором указанные злокачественные клетки представляют собой злокачественные клетки или ткань рака поджелудочной железы, лимфомы или немелкоклеточного рака легких.
38. Способ по пункту 32, в котором на указанные клетки воздействуют эффективным количеством антитела-агониста DR4 или DR5.
39. Способ по пункту 38, в котором на указанные клетки воздействуют эффективным количеством антитела-агониста DR5, которое связывает рецептор DR5, изображенный на Фиг.3A.
40. Способ лечения заболевания млекопитающего, например, заболевания, связанного с иммунной системой, или злокачественного заболевания, включающий в себя стадии:
получения образца ткани или клеток указанного млекопитающего;
исследования образца ткани или клеток для выявления экспрессии GalNac-T14, и
после определения экспрессии указанной GalNac-T14 введение указанному млекопитающему эффективного количества антитела к рецептору смерти.
41. Способ по пункту 40, в котором указанную экспрессию GalNac-T14 исследуют путем выявления экспрессии мРНК GalNac-T14.
42. Способ по пункту 40, в котором указанную экспрессию GalNac-T14 исследуют с помощью иммуногистохимии.
43. Способ по пункту 40, дополнительно включающий в себя стадию изучения экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанной ткани или клетке.
44. Способ по пункту 40, в котором образец ткани или клеток содержит злокачественную ткань или злокачественные клетки.
45. Способ по пункту 44, в котором указанные злокачественные клетки или злокачественная ткань содержит клетки или ткань злокачественной опухоли поджелудочной железы, лимфомы или немелкоклеточного рака легких.
46. Способ по пункту 40, где указанному млекопитающему вводят эффективное количество анти-DR4 или DR5 антитела.
47. Способ по пункту 40, где указанному млекопитающему также вводят химиотерапевтическое средство или применяют лучевую терапию.
48. Способ по пункту 40, в котором указанному млекопитающему также вводят цитокин, цитотоксическое средство или ингибитор фактора роста.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фиг.1 показана нуклеотидная последовательность кДНК лиганда Apo-2 человека (SEQ ID NO:2) и происходящая из нее аминокислотная последовательность (SEQ ID NO:1). «N» в 447 положении нуклеотидной последовательности используется для обозначения нуклеотидного основания, которое может быть «T» или «G».
На Фиг.2A и 2B показана нуклеотидная последовательность кДНК (SEQ ID NO:4) полной длины DR4 человека и происходящая из нее аминокислотная последовательность (SEQ ID NO:3). Соответствующие нуклеотидная и аминокислотная последовательности DR4 человека также описаны у Pan et al., Science, 276:111 (1997).
На Фиг.3A показана последовательность 411 аминокислот (SEQ ID NO:5) DR5 человека, опубликованная в WO 98/51793 19 ноября 1998. В уровне техники известен вариант транскрипционного сплайсинга DR5 человека. Этот вариант сплайсинга DR5 кодирует последовательность 440 аминокислот (SEQ ID NO:6) DR5 человека, показанную на Фиг.3B и 3C, опубликованную в WO 98/35986 20 августа 1998.
На Фиг.3D показаны нуклеотидные последовательности кДНК (SEQ ID NO:7) полной длины DcR1 человека и происходящая из них аминокислотная последовательность (SEQ ID NO:8). Соответствующие нуклеотидная и аминокислотная последовательности DcR1 человека (и его отдельные домены) также показаны и описаны в WO 98/58062.
На Фиг.3E показаны нуклеотидные последовательности кДНК (SEQ ID NO:9) полной длины DcR2 человека и происходящая из них аминокислотная последовательность (SEQ ID NO:10). Соответствующие нуклеотидная и аминокислотная последовательности DcR2 человека (и его отдельные домены) также представлены в WO 99/10484.
На Фиг.4A показана нуклеотидная последовательность GalNac-T14 человека (SEQ ID NO:11) и происходящая из нее аминокислотная последовательность (SEQ ID NO:12). Эти последовательности также описаны у Wang et al., BBRC, 300:738-744 (2003).
На Фиг.4B показана нуклеотидная последовательность GalNac-T3 человека (SEQ ID NO:13) и происходящая из нее аминокислотная последовательность (SEQ ID NO:14). Эти последовательности также описаны у Bennett et al., J. Biol. Chem., 271:17006-17012 (1996).
На Фиг.5 представлена суммарная таблица данных по IC50, полученных при анализе клеточных линий немелкоклеточного рака легких («NSCLC») на чувствительность или резистентность к апоптотической активности Apo2L (+ 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS) и моноклонального антитела к DR5 «DR5 ab», перекрестно-связанного «XL» или не перекрестно-связанного, + 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS), измеренную в MTT анализах цитотоксичности.
На Фиг.6 представлена суммарная таблица данных по IC50, полученных при анализе клеточных линий рака поджелудочной железы на чувствительность или резистентность к апоптотической активности Apo2L (+ 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS) и моноклонального антитела к DR5 «DR5 ab», перекрестно-связанного «XL» или не перекрестно-связанного, + 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS), измеренную в MTT анализах цитотоксичности.
На Фиг.7 представлена суммарная таблица данных по IC50, полученных при анализе клеточных линий неходжкинской лимфомы («NHL») на чувствительность или резистентность к апоптотической активности Apo2L (+ 10% эмбриональная сыворотка теленка «FBS») и моноклонального антитела к DR5 «DR5 ab», перекрестно-связанного «XL» или не перекрестно-связанного (+ 10% эмбриональная сыворотка теленка «FBS»), измеренную в MTT анализах цитотоксичности.
На Фиг.8 представлено сравнение чувствительности («sen») или резистентности («RES») выбранных клеточных линий NSCLC, рака поджелудочной железы и NHL к антителу DR5 и корреляция с экспрессией GalNac-T14, измеренной по экспрессии мРНК GalNac-T14.
На Фиг.9 представлена гистограмма различных клеточных линий NSCLC, поджелудочной железы и NHL, расположенных (в порядке убывания) по уровням экспрессии образцов мРНК GalNac-T14.
Фиг.10A-D иллюстрируют дифференциальную экспрессию специфических ферментов О-гликозилирования в Apo2L/TRAIL-чувствительных и резистентных линиях злокачественных клеток: (A) вариабельность клеток измеряли после инкубирования с различными дозами Apo2L/TRAIL. IC50 для каждой клеточной линии вычисляли как концентрацию Apo2L/TRAIL, которая дает 50% потерю жизнеспособности. Каждый эксперимент на жизнеспособность повторяли по меньшей мере три раза в присутствии низкой (0,5%) и высокой (10%) концентрации эмбриональной сыворотки теленка. Черные, серые или незаполненные символы обозначают клеточные линии, которые являются наиболее чувствительными, умеренно чувствительными или резистентными к Apo2L/TRAIL, соответственно. (B) уровни экспрессии мРНК ppGalNAcT-14 (набор зондов 219271_at) в клеточных линиях поджелудочной железы и злокачественной меланомы. Клеточные линии распределены по типу ткани и чувствительности к Apo2L/TRAIL. Черные, серые или незаштрихованные столбцы обозначают клеточные линии, как в А. (C) уровни экспрессии мРНК Fut-6 (верхняя панель, набор зондов 211885_x_at) и ppGalNAcT-3 (нижняя панель, набор зондов 203397_s_at) в клеточных линиях колоректального рака. Клеточные линии распределены как в B. Значения P на панелях B и С основаны на критерии Фишера корреляции между чувствительностью клеточной линии (включая высокую и умеренную) и экспрессией мРНК выше порогового значения. (D) Влияние Apo2L/TRAIL на рост прижившихся опухолевых ксенотрансплантатов. Бестимусным мышам «nude», несущим GalNAcT-3/Fut-6-позитивные (левая панель) или GalNAcT-3/Fut-6-негативные (правая панель) опухоли, вводили носитель или Apo2L/TRAIL (60 мг/кг/день внутрибрюшинно в дни 0-4) и наблюдали за размером опухоли (среднее±SE, N=10 мышей на группу).
Фиг.11 иллюстрирует модуляцию конкретных ферментов О-гликозилирования, изменяет чувствительность к Apo2L/TRAIL. (A) клетки Colo205 предварительно инкубировали с ингибитором фермента pan O-гликозилирования бензил-GalNAc (bGalNAc), обрабатывали Apo2L/TRAIL в течение 24 ч и определяли жизнеспособность клеток (DMSO=носитель-контроль). (B) Клетки PSN-1 (карцинома поджелудочной железы) и Hs294T (меланома) трансфектировали каспазой-8 или ppGalNAcT-14 siРНК в течение 48 ч, инкубировали с Apo2L/TRAIL в течение еще 24 ч и определяли жизнеспособность клеток. Дуплексы siРНК против нецелевой последовательности (Dharmacon) использовали в качестве контроля (NTC). (C) клетки DLD-1 колоректальной карциномы трансфектировали ppGalNAcT-3 или Fut-6 siРНК и тестировали, как в B. (D) Клетки HEK293 ко-трансфектировали плазмидами, кодирующими указанные гены в комбинации с ppGalNAcT-14 или векторным контролем. Апоптоз измеряли через 24 ч путем окрашивания Аннексином V (левая панель). Клетки H1569 меланомы трансдуцировали ретровирусом, направляющим экспрессию ppGalNAcT-14 или контрольным ретровирусом; полученные в результате пулы клеточных линий обрабатывали Apo2L/TRAIL в течение 24 ч и определяли жизнеспособность клеток (правая панель). Вестерн блот анализ с использованием анти-FLAG антител применяли для верификации экспрессии меченной эпитопом ppGalNAcT-14.
Фиг.12 иллюстрирует (A) анализ каспазного каскада, индуцированного действием Apo2L/TRAIL. Клетки PSN-1 и DLD-1 трансфектировали siРНК против ppGalNAcT-14 или Fut-6, соответственно, в течение 48 ч. Клетки обрабатывали Apo2L/TRAIL в течение 4 или 8 ч, и клеточные лизаты анализировали с помощью иммуноблоттинга с антителами, специфичными к каспазе-8, Bid, каспазе-9, каспазе-3, или актину в качестве контрольной нагрузки. (B) Клетки PSN-1 трансфектировали ppGalNAcT-14 siРНК как в A, обрабатывали Apo2L/TRAIL в течение 4 ч и определяли ферментативную активность каспаз-3/7 в клеточных лизатах. (C) Анализ Apo2L/TRAIL DISC. Клетки PSN-1 трансфектировали ppGalNAcT-14 siРНК как в A. Добавляли FLAG-Apo2L/TRAIL (1 мг/мл) в течение 0-60 мин, клетки лизировали и подвергали иммунопреципитации анти-FLAG антителом. DISC-ассоциированный FADD, каспазу-8, DR4, и выявляли иммуноблоттингом. (D) Клетки PSN-1 трансфектировали, обрабатывали и подвергали DISC иммунопреципитации как в C, и DISC-ассоциированную ферментативную активность каспазы-8 измеряли, как описано ранее (Sharp et al., J. Biol. Chem., 280:19401 (2005).
Фиг.13 иллюстрирует (A) моносахаридный анализ рекомбинантного DR5 человека (длинный вариант сплайсинга), продуцируемого в клетках CHO, выполненный с помощью HPAEC-PAD (высокоэффективная анионообменная хроматография с импульсным амперометрическим обнаружением). (B) Сравнение последовательностей Apo2L/TRAIL рецепторов человека (DR5 человека длиной 440 а.к. форма «hDR5L», DR5 человека короткая форма 411 а.к. «hDR5S» и hDR4), DR5 мыши или крысы (mDR5), Fas человека (hFas) TNFR1 человека (hTNFR1). Рамки указывают предполагаемые сайты О-гликозилирования. (C) Иммуноблоттинг суммарных клеточных лизатов, соответствующих D. DR5L-5T и DR5S-5T представляют собой конструкции, содержащие 5 замен треонин-на-аланин и DR5L-5T3S и DR5S-5T3S представляют собой конструкции, содержащие 5 замен треонин-на-аланин и три замены серин-на-аланин, соответственно, в остатках, которые являются возможными сайтами О-гликозилирования. (D) HEK293 клетки ко-трансфектировали указанными DR5 конструкциями вместе с вектором или плазмидой ppGalNAcT-14 в течение 48 ч и апоптоз измеряли окрашиванием Аннексином V. (E) уровни экспрессии мРНК для ppGalNAcT-14 (чип Affymetrix, набор зондов 219271_at) в образцах первичных опухолей человека, полученных из злокачественных опухолей кожи (SCC=сквамозно-клеточная карцинома), легких, поджелудочной железы (Panc), груди, яичников (Ov), эндометрия (Endo), мочевого пузыря (Bla, TCC= переходноклеточная карцинома) и NHL (FL=фолликулярная лимфома, DLBCL=диффузная крупноклеточная В-клеточная лимфома). Средние значения экспрессии в образцах указаны серой горизонтальной полосой для каждого класса. Показаны граничные значения 500 и 200 (меланома), соответствующие данным по клеточной линии с Фиг.10B.
Фиг.14 иллюстрирует (A) уменьшение экспрессии мРНК ppGalNAcT-14 или ppGalNAcT-3 в клетках PSN-1 или DLD-1 после 48 ч siРНК нокдауна с помощью Taqman анализа. (B) экспрессию GalNAcT-14 восстанавливают в клетках PSN-1 путем трансфекции пустой плазмиды (Empty), GalNAcT-14 дикого типа (GalNAcT-14) или GalNAcT-14, содержащих молчащие мутации siРНК (GalNAcT-14 si(1)Mut) после siGalNAcT-14 (1) опосредованных нокдауном ppGalNAcT-14. (C) Отрицательная регуляция ppGalNAcT-3 или Fut-6 под действием интерферирующих РНК ингибирует индуцированную Apo2L/TRAIL клеточную гибель в клетках C170 (колоректальный рак). Экспериментальная процедура как в 11C. (Таблица 1) A) Суммарная таблица фенотипов нокдауна под действием siРНК. Клеточные линии, в которых отрицательная регуляция GalNAcT-14 или ppGalNAcT-3 и Fut-6, в результате, приводит к защите от действия Apo2L/TRAIL, обозначены показывающими менее (+) или более 50% (++) защиту по меньшей мере с одним тестируемым олигонуклеотидом siРНК. (0) указывает на отсутствие защиты от Apo2L/TRAIL. (D), (E) После 48 ч нокдауна, указанными siРНК, клетки обрабатывали возрастающими дозами этопозида или стауроспорина (STS) в течение 24 ч и проводили анализ жизнеспособности клеток. (F) Ретровирусную ppGalNAcT-14, гиперэкспрессирующую PA-TU-8902 и пулы клеточных линий PL-45 подвергали анализу на жизнеспособность клеток после обработки Apo2L/TRAIL. Вестерн-блоттинг с использованием анти-FLAG антител показывает ретровирусную экспрессированную ppGalNAcT-14 в этих клетках.
Фиг.15 (A) Вестерн-блоттинг индуцированного Apo2L/TRAIL каскада активации каспазы в клеточных линиях колоректального рака, Colo205, чувствительных к Apo2L/TRAIL, и резистентных, RKO и SW1417. Клетки обрабатывали 1000 нг/мл Apo2L/TRAIL в течение 8 и 24 ч, и суммарные клеточные лизаты подвергали Вестерн-блоттингу с использованием антител, специфичных к каспазе-8, Bid, каспазе-9, каспазе-3 и актину в качестве контрольной нагрузки. (B) Нокдаун Fut-6 снижал обновление пула и активацию каспазы-8 в Apo2L/TRAIL DISC в клетках DLD-1. Экспериментальная процедура соответствует 12D. (C) экспрессию клеточной поверхности DR4 и DR5 измеряли с помощью FACS анализа в клетках, которые подвергали нокдауну siРНК в отношении указанных генов.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Способы и методики, описанные здесь или на которые дана ссылка, в основном хорошо понятны и широко применяются с использованием общепринятой методологии специалистами в данной области, например, такие как широкоиспользуемые методики молекулярного клонирования, описанные у Sambrook et al., Molecular Cloning: A Laboratory Manual 2nd. edition (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. Соответствующим образом, процедуры, включающие в себя использование коммерчески доступных наборов и реактивов, обычно выполняются в соответствии с протоколами, установленными производителями и/или параметрами, если не отмечено иное.
Перед тем, как будут описаны способы и анализы настоящего изобретения, должно быть понятно, что это изобретение не ограничивается конкретной методикой, протоколами, клеточными линиями, видами или родами животных, конструкциями, и реактивы, описанные как таковые, разумеется, могут меняться. Также должно быть понятно, что терминология, используемая в описании только с целью описания конкретных вариантов осуществления, не предназначена для ограничения объема настоящего изобретения, который будет ограничен только прилагаемой формулой изобретения.
Следует отметить, что используемые здесь и в прилагаемой формуле изобретения формы единственного числа включают в себя ссылки на формы множественного числа, если из контекста явным образом не следует иное. Таким образом, например, ссылка на «генетическую альтерацию» включает в себя множество таких альтераций, а ссылка на «зонд» включает в себя ссылку на один или несколько зондов и их эквиваленты, известные специалистам в данной области, и так далее.
Все публикации, упомянутые здесь, включены в настоящее описание в качестве ссылки для раскрытия и описания способов и/или веществ, в отношении которых эти публикации цитируются. Упомянутые здесь публикации цитируются в связи с их раскрытием до даты подачи настоящей заявки. Здесь ничто не должно интерпретироваться как признание того, что авторы изобретения не имеют права относить к более ранней дате публикации на основании более ранней даты приоритета или даты приоритета изобретения. В дальнейшем фактические даты публикаций могут отличаться от дат, представленных здесь, и требуют независимой проверки.
I. ОПРЕДЕЛЕНИЯ
Используемые здесь термины «Apo2L/TRAIL», «Apo-2L» и «TRAIL» относятся к полипептидной последовательности, которая включает в себя аминокислотные остатки 114-281, включительно, 95-281, включительно, остатки 92-281, включительно, остатки 91-281, включительно, остатки 41-281, включительно, остатки 15-281, включительно, или остатки 1-281, включительно, аминокислотной последовательности, показанной на Фиг.1, а также биологически активные фрагменты, делеционные варианты, варианты со вставками или заменами приведенных выше последовательностей. В одном варианте осуществления полипептидная последовательность включает в себя остатки 114-281 Фиг.1, и, необязательно, состоит из остатков 114-281 Фиг.1. Необязательно, полипептидная последовательность содержит остатки 92-281 или остатки 91-281 Фиг.1. Полипептиды Apo-2L могут кодироваться нативной нуклеотидной последовательностью, представленной на Фиг.1. Необязательно, кодоном, который кодирует остаток Pro119 (Фиг.1), может быть «CCT» или «CCG». В других вариантах осуществления фрагменты или варианты являются биологически активными и имеют по меньшей мере 80% идентичность аминокислотной последовательности, более предпочтительно, по меньшей мере 90% идентичность последовательности, и даже еще более предпочтительно, по меньшей мере 95%, 96%, 97%, 98% или 99% идентичность последовательности любой из упомянутых выше последовательностей Apo2L/TRAIL. Необязательно, полипептид Apo2L/TRAIL кодируется нуклеотидной последовательностью, которая гибридизируется в условиях строгости с кодирующей полинуклеотидной последовательностью, представленной на Фиг.1. Это определение охватывает варианты Apo2L/TRAIL с заменами, в которых по меньшей мере одна из его природных аминокислот заменена на остаток аланина. Конкретные варианты Apo2L/TRAIL с заменами включают в себя те, в которых по меньшей мере одна аминокислота заменена остатком аланина. Эти варианты с заменами включают в себя варианты, которые обозначены, например, как «D203A»; «D218A» и «D269A». Эта номенклатура используется для идентификации вариантов Apo2L/TRAIL, в которых остатки аспарагиновой кислоты в положениях 203, 218 и/или 269 (используя нумерацию, показанную на Фиг.1) замещены остатками аланина. Необязательно, варианты Apo2L могут содержать один или несколько замен аланином, которые упоминаются в Таблице I опубликованной заявки PCT WO 01/00832. Варианты с заменами включают в себя одну или несколько замен остатков, указанных в Таблице I заявки WO 01/00832, опубликованной 4 января 2001. Это определение также охватывает нативную последовательность Apo2L/TRAIL, выделенную из источника Apo2L/TRAIL или полученную рекомбинантными или синтетическими способами. Apo2L/TRAIL по изобретению включает в себя полипептиды, обозначаемые Apo2L/TRAIL или TRAIL, описанные в публикациях №№ WO 97/01633 и WO 97/25428. Термины «Apo2L/TRAIL» или «Apo2L» используются для обозначения главным образом форм Apo2L/TRAIL, которые включают в себя мономерные, димерные или тримерные формы этого полипептида. Во всей нумерации аминокислотных остатков, указанной в последовательности Apo2L, используется нумерация в соответствии с Фиг.1, если особым образом не указано иное. Например, «D203» или «Asp203» относится к остатку аспарагиновой кислоты в положении 203 в последовательности, представленной на Фиг.1.
Термин «внеклеточный домен Apo2L/TRAIL» или «Apo2L/TRAIL ECD» относится к форме Apo2L/TRAIL, которая, по существу, свободна от трансмембранного и цитоплазматического доменов. Как правило, ECD будет иметь менее 1% такого трансмембранного и цитоплазматического доменов, и, предпочтительно, будет иметь менее 0,5% таких доменов. Будет понятно, что любой трансмембранный домен(ы), идентифицированный для полипептидов настоящего изобретения, идентифицирован в соответствии с критериями, используемыми в данной области для идентификации этого типа гидрофобного домена. Точные границы трансмембранного домена могут изменяться, но наиболее вероятно, не более чем на 5 аминокислот с любого конца домена, установленного первоначально. В предпочтительных вариантах осуществления ECD будет состоять из последовательности полипептида растворимого внеклеточного домена, свободного от трансмембранного и цитоплазматического доменов (и не связанного с мембраной). Конкретные последовательности внеклеточного домена Apo-2L/TRAIL описаны в публикациях PCT №№ WO 97/01633 и WO 97/25428.
Термин «мономер Apo2L/TRAIL» или «мономер Apo2L» относится к ковалентной цепи последовательности внеклеточного домена Apo2L.
Термин «димер Apo2L/TRAIL» или «димер Apo2L» относится к двум мономерам Apo-2L, соединенных ковалентной связью посредством дисульфидной связи. Используемый здесь термин включает в себя автономные димеры Apo2L и димеры Apo2L, которые находятся в тримерных формах Apo2L (т.е. связаны с другим, третьим мономером Apo2L).
Термин «тример Apo2L/TRAIL» или «тример Apo2L» относится к трем мономерам Apo2L, которые не связаны нековалентно.
Термин «аггрегат Apo2L/TRAIL» используется для обозначения самоассоциированных олигомерных форм Apo2L/TRAIL более высокого порядка, таких как тримеры Apo2L/TRAIL, например, гексамерные и наномерные формы Apo2L/TRAIL. Определение присутствия и количества мономера, димера или тримера Apo2L/TRAIL (или других агрегированных форм) можно осуществить, используя способы и анализы, известные в уровне техники (и используя коммерчески доступные материалы), например, нативная эксклюзионная ВЭЖХ («SEC»), денатурирующая эксклюзионная с использованием додецилсульфата натрия («SDS-SEC»), ВЭЖХ с обращенной фазой и капиллярный электрофорез.
«Рецептор лиганда Apo-2» включает в себя рецепторы, называемые в уровне техники «DR4» и «DR5», полинуклеотидная и полипептидная последовательность которых показана на Фиг.2 и 3 соответственно. Pan et al. описали представителя семейства TNF рецепторов, названного «DR4» (Pan et al., Science, 276:111-113 (1997); см. также WO 98/32856, опубликованную 30 июля 1998; WO 99/37684, опубликованную 29 июля 1999; WO 00/73349, опубликованную 7 декабря 2000; US 6433147, выданный 13 августа 2002; US 6461823, выданный 8 октября 2002, и US 6342383, выданный 29 января 2002). Sheridan et al., Science, 277:818-821 (1997) и Pan et al., Science, 277:815-818 (1997) описали другой рецептор для Apo2L/TRAIL (см. также WO 98/51793, опубликованную 19 ноября 1998; WO 98/41629, опубликованную 24 сентября 1998). Этот рецептор назвали DR5 (этот рецептор альтернативно был назван Apo-2; TRAIL-R, TR6, Tango-63, hAPO8, TRICK2 или KILLER; Screaton et al., Curr. Biol., 7:693-696 (1997); Walczak et al., EMBO J., 16:5386-5387 (1997); Wu et al., Nature Genetics, 17:141-143 (1997); WO 98/35986, опубликованная 20 августа 1998; EP870827, опубликованная 14 октября 1998; WO 98/46643, опубликованная 22 октября 1998; WO 99/02653, опубликованная 21 января 1999; WO 99/09165, опубликованная 25 февраля 1999; WO 99/11791, опубликованная 11 марта 1999; US 2002/0072091, опубликованная 13 августа 2002; US 2002/0098550, опубликованная 7 декабря 2001; US 6313269, выданный 6 декабря 2001; US 2001/0010924, опубликованная 2 августа 2001; US 2003/01255540, опубликованная 3 июля 2003; US 2002/0160446, опубликованная 31 октября 2002, US 2002/0048785, опубликованная 25 апреля 2002; US 6569642, выданный 27 мая 2003, US 6072047, выданный 6 июня 2000, US 6642358, выданный 4 ноября 2003). Как описано выше, другие рецепторы для Apo-2L включают в себя DcR1, DcR2 и OPG (см. Sheridan et al., supra; Marsters et al., supra; и Simonet et al., supra). Термин «рецептор Apo-2L» при использовании здесь охватывает рецептор с нативной последовательностью и варианты рецептора. Эти термины охватывают рецептор Apo-2L, экспрессируемый у различных млекопитающих, в том числе у людей. Рецептор Apo-2L может экспрессироваться эндогенно, как происходит в природе в тканях различного происхождения у человека, или может быть экспрессирован рекомбинантными или синтетическими способами. «Рецептор Apo-2L с нативной последовательностью» содержит полипептид, имеющий такую же аминокислотную последовательность, что и рецептор Apo-2L природного происхождения. Следовательно, рецептор Apo-2L с нативной последовательностью может иметь аминокислотную последовательность природного рецептора Apo-2L любого млекопитающего. Такой рецептор Apo-2L с нативной последовательностью может быть выделен из природного источника или может быть получен рекомбинантным или синтетическим путем. Термин «рецептор Apo-2L с нативной последовательностью» особенно охватывает природные усеченные или секретированные формы этого рецептора (например, растворимая форма, содержащая, например, последовательность внеклеточного домена), варианты природных форм (например, формы в результате альтернативного сплайсинга) и природные аллельные варианты. Варианты рецептора могут включать в себя фрагменты или делеционные мутанты рецептора Apo-2L с нативной последовательностью. На Фиг.3A показана последовательность 411 аминокислот DR5 человека, как опубликовано в WO 98/51793 19 ноября 1998. Вариант транскрипционного сплайсинга DR5 человека известен в уровне техники. Этот вариант сплайсинга DR5 кодирует последовательность 440 аминокислот DR5 человека, показанную на Фиг.3B и 3C, как опубликовано в WO 98/35986 20 августа 1998.
«Антитело к рецептору смерти» в контексте настоящего изобретения относится главным образом к антителу, или антителам, направленным на рецептор суперсемейства рецепторов фактора некроза опухоли, и содержащий домен смерти, способный передавать сигнал апоптоза, и такие антитела включают в себя антитело к DR5 и антитело к DR4.
«Антитело к рецептору DR5» или «DR5 антитело» или «анти-DR5 антитело» используется в широком смысле для обозначения антител, которые связываются по меньшей мере с одной формой рецептора DR5, например, 1-411 последовательностью, показанной на Фиг.3A, или 1-440 последовательностью, показанной на Фиг.3B-3C, или его внеклеточным доменом. Необязательно антитело к DR5 слито или связано с гетерологичной последовательностью или молекулой. Предпочтительно гетерологичная последовательность дает возможность антителу образовать более высокий порядок или олигомерные комплексы, или способствует этому. Необязательно антитело к DR5 связывает рецептор DR5, но не связывает или перекрестно не реагирует с любым дополнительным рецептором к Apo-2L (например, DR4, DcR1 или DcR2). Необязательно антитело является агонистом сигнальной активности DR5.
Необязательно, антитело к DR5 по изобретению связывается с DR5 рецептором в диапазоне концентраций примерно от 0,1 нM примерно до 20 мM, измеренных в анализе связывания BIAcore. Необязательно антитела к DR5 по изобретению демонстрируют значение Ic 50 примерно от 0,6 нM примерно до 18 мM, измеренное в анализе связывания BIAcore.
«Антитело к рецептору DR4», «антитело к DR4» или «анти-DR4 антитело» используются в широком смысле для обозначения антител, которые связываются по меньшей мере с одной формой рецептора DR4 или его внеклеточного домена. Необязательно антитело к DR4 слито или связано с гетерологичной последовательностью или молекулой. Предпочтительно гетерологичная последовательность дает возможность антителу образовать более высокий порядок или олигомерные комплексы, или способствует этому. Необязательно антитело к DR4 связывается с рецептором DR4, но не связывается или перекрестно не реагирует с любым дополнительным рецептором к Apo-2L (например, DR5, DcR1 или DcR2). Необязательно антитело является агонистом сигнальной активности DR4.
Необязательно антитело к DR4 по изобретению связывается с рецептором DR4 в диапазоне концентраций примерно от 0,1 нM примерно до 20 мM, измеренных в анализе связывания BIAcore. Необязательно антитела к DR4 по изобретению демонстрируют значение Ic 50 примерно от 0,6 нM примерно до 18 мM, измеренное в анализе связывания BIAcore.
Термин «агонист» используется в самом широком смысле и включает в себя любую молекулу, которая частично или полностью усиливает, стимулирует или активирует одну или несколько биологических активностей Apo2L/TRAIL, DR4 или DR5, in vitro, in situ или in vivo. Примерами таких биологических активностей являются связывание Apo2L/TRAIL с DR4 или DR5, включая апоптоз, а также те, о которых дополнительно сообщается в литературе. Агонист может действовать прямо или опосредованно. Например, агонист может действовать для частичного или полного усиления, стимуляции или активации одной или нескольких биологических активностей DR4 или DR5, in vitro, in situ или in vivo как результат его непосредственного связывания с DR4 или DR5, что вызывает активацию рецептора или трансдукцию сигнала. Агонист также может действовать опосредованно для частичного или полного усиления, стимуляции или активации одной или нескольких биологических активностей DR4 или DR5, in vitro, in situ или in vivo как результат, например, стимуляции другой эффекторной молекулы, которая затем вызывает активацию DR4 или DR5 или передачу сигнала. Предполагается, что агонист может действовать как молекула энхансер, которая действует опосредованно для усиления или увеличения активации или активности DR4 или DR5. Например, агонист может усиливать активность эндогенного Apo-2L у млекопитающего. Это можно осуществить, например, путем предварительного комплексообразования DR4 или DR5 или путем стабилизации комплексов соответствующего лиганда с рецептором DR4 или DR5 (например, стабилизируя нативный комплекс, образованный между Apo-2L и DR4 или DR5).
Термин «биомаркер», используемый в настоящей заявке, относится главным образом к молекуле, в том числе к гену, белку, углеводной структуре или гликолипиду, экспрессия которой в ткани или на ткани млекопитающего, или в клетке, или на клетке можно определить стандартными способами (или способами, описанными здесь) и которая прогнозирует чувствительность клеток или тканей млекопитающих к Apo2L/TRAIL или антителу рецептора смерти. Такие биомаркеры, рассматриваемые в настоящем изобретении, включают в себя, но не ограничиваются ими, молекулы семейства белков GalNac-T. Были описаны представители семейства генов и белков N-ацетилгалактозаминилтрансферазы («GalNac-T») человека (см., например, Hang et al., “The chemistry and biology of mucin-type O-linked glycosylation initiated by the polypeptide N-acetylgalactosaminyltransferases”, Bioorganic & Medicinal Chemistry (доступно в мае 2005 на www.sciencedirect.com) и цитируемые там ссылки; Wang et al., BBRC, 300:738-744 (2003) и цитируемые там ссылки), и считается, что они функционируют для определения количества и положения О-связанных цепей сахаров в белках. Необязательно определяется, что экспрессия такого биомаркера выше экспрессии, наблюдаемой в образце контрольной ткани или клетки. Необязательно, например, экспрессия такого биомаркера будет определяться с использованием микрочипа для генной экспрессии, количественной ПЦР или иммуногистохимического (IHC) анализа. Необязательно, экспрессия биомаркера GalNac-T, такого как GalNac-T14 или GalNac-T3, будет определяться на уровне по меньшей мере 750, измеренном с помощью микрочипового исследования Affymetrix U133P, или в 500 раз, или предпочтительно, по меньшей мере в 1000 раз выше, в образце тестируемой ткани или клетке при выявлении экспрессии биомаркера с использованием количественной ПЦР.
«UDP-N-ацетил-D-галактозамин:полипептид N-ацетилгалактозаминилтрансфераза-T14», «pp-GalNac-T14», «GalNac-T14», «GALNT14» используется здесь для обозначения мембранного белка II типа, имеющего характерные признаки семейства молекул GalNac-T, содержащих N-концевой цитоплазматический домен, трансмембранный домен, стволовую область и каталитический домен. В дополнительном варианте осуществления молекула GalNac-T14 человека содержит 1659 пар оснований, кодирующих белок 552 аминокислот, как показано на Фиг.4A. кДНК человека полной длины была депонирована в GenBank с номером доступа № AB078144. Как описано у Wang et al., BBRC, 300:738-744 (2003), были идентифицированы сплайсированные изоформы GalNac-T14, которые включают в себя (или не включают) конкретные экзоны, такие как экзоны 2, 3 и/или 4. В настоящем изобретении рассматривается изучение экспрессии любой из таких разнообразных изоформ GalNac-T14, и что экспрессия любой из таких изоформ прогнозирует чувствительность образца ткани или клетки млекопитающего к Apo2L/TRAIL или антителу рецептора смерти.
«UDP-N-ацетил-D-галактозамин:полипептид N-ацетилгалактозаминилтрансфераза-T3», «pp-GalNac-T3», «GalNac-T3», «GALNT3» используются в настоящем описании для обозначения мембранного белка II типа, имеющего характерные признаки семейства молекул GalNac-T, содержащего N-концевой цитоплазматический домен, трансмембранный домен, стволовую область и каталитический домен. В дополнительном варианте осуществления GalNac-T3 полипептид человека содержит аминокислотную последовательность, показанную на Фиг.4B. GalNac-T3 дополнительно описана у Bennett et al., J. Biol. Chemistry, 271:17006-17012 (1996).
Под «индивидом» или «пациентом» подразумевается любой отдельный индивид, которому требуется лечение, включая людей. Также предназначен для включения в качестве индивида любых индивидов, участвующих в клинических испытаниях, не проявляющих каких-либо признаков заболевания, или индивиды, участвующие в эпидемиологических исследованиях, или индивиды, которые являются контрольными пациентами.
Используемый в описании термин «млекопитающее» относится к любому млекопитающему, относящемуся к млекопитающему, включая людей, коров, лошадей, собак и кошек. В предпочтительном варианте осуществления изобретения млекопитающим является человек.
Под «образцом ткани или клеток» подразумевается скопление однотипных клеток, полученных из ткани субъекта или пациента. Источником образца ткани или клеток может быть твердая ткань, как из только что полученного, замороженного и/или фиксированного органа или образца ткани, или биопсии, или аспирата; крови или любых компонентов крови; жидкостей тела, таких как спинномозговая жидкость, амниотическая жидкость, перитонеальная жидкость или интерстициальная жидкость; клетки с любого срока гестации или развития организма. Образец ткани также может представлять собой первичные или культивированные клетки или клеточные линии. Необязательно, образец ткани или клеток получают из первичной или метастазирующей опухоли. Образец ткани может содержать соединения, которые естественным образом не смешаны с тканью в природе, такие как консерванты, антикоагулянты, буферы, фиксаторы, питательные вещества, антибиотики и подобное.
Для целей настоящего изобретения «срез» образца ткани означает отдельную часть или фрагмент образца ткани, например тонкий слой ткани или клеток, срезанный с образца ткани. Понятно, что может быть сделано множество срезов образцов ткани и их можно подвергнуть исследованию в соответствии с настоящим изобретением, при условии, что подразумевается, что настоящее изобретение включает в себя способ, посредством которого один и тот же образец ткани анализируют как на морфологическом, так и на молекулярном уровнях, или его исследуют как в отношении белков, так и нуклеиновых кислот.
Под «коррелирует» или «коррелирующий» подразумевается сравнение, любым способом, проведение и/или результаты первого анализа или протокола с проведением, и/или результатами второго анализа или протокола. Например, один может использовать результаты первого анализа или протокола при выполнении других протоколов и/или может использовать результаты первого анализа или протокола для определения, следует ли осуществлять другой анализ или протокол. В отношении различных вариантов осуществления в контексте настоящего изобретения один может использовать результаты аналитического исследования, например, экспрессии мРНК или IHC для определения, следует ли проводить особую схему лечения с использованием Apo2L/TRAIL или антитела к рецептору смерти.
Под «нуклеиновой кислотой» подразумевается включение любой ДНК или РНК. Например, хромосомной, митохондриальной, вирусной и/или бактериальной нуклеиновой кислоты, присутствующей в образце ткани. Термин «нуклеиновая кислота» охватывает любую или обе спирали двуспиральной молекулы нуклеиновой кислоты и включает в себя любой фрагмент или часть интактной молекулы нуклеиновой кислоты.
Под «геном» подразумевается любая последовательность нуклеиновой кислоты или ее часть с функциональной ролью в кодировании или транскрипции белка, или регуляцией экспрессии других генов. Ген может состоять из всех нуклеиновых кислот, отвечающих за кодирование функционального белка, или только части нуклеиновых кислот, ответственных за кодирование или экспрессию белка. Последовательность нуклеиновой кислоты может содержать генетическую аномалию в экзонах, интронах, областях инициации или терминации, промоторных последовательностях, других регуляторных последовательностях или уникальных участках, смежных с этим геном.
При использовании в описании слово «метка» относится к соединению или композиции, конъюгированной или слитой прямо или опосредованно с реагентом, таким как зонд нуклеиновой кислоты или антитело, и облегчает выявление этого реагента, с которым она конъюгирована или конденсирована. Метка сама по себе может быть детектируемой (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментной метки, может катализировать химическое изменение детектируемого соединения-субстрата или композиции.
Термин «антитело» используется в описании в самом широком смысле и, главным образом, относится к интактным моноклональным антителам, поликлональным антителам, мультиспецифическим антителам (например, биспецифическим антителам), образованным по меньшей мере из двух интактных антител или фрагментов антител, при условии, что они проявляют желаемую биологическую активность.
«Фрагменты антител» включают в себя часть интактного антитела, предпочтительно содержащую его антигенсвязывающую или вариабельную область. Примеры фрагментов антител включают в себя Fab, Fab', F(ab')2 и Fv фрагменты; диатела; линейные антитела; одноцепочечные молекулы антител; и мультиспецифические антитела, образованные из фрагментов антител.
«Нативные антитела» обычно представляют собой гетеротетрамерные гликопротеины примерно 150000 дальтон, состоящие из двух идентичных легких цепей (L) и двух идентичных тяжелых цепей (H). Каждая легкая цепь соединена с тяжелой цепью одной ковалентной дисульфидной связью, тогда как количество дисульфидных связей варьирует среди тяжелых цепей различных изотипов иммуноглобулинов. Каждая тяжелая и легкая цепь также имеет правильно расположенные внутрицепочечные дисульфидные мостики. Каждая тяжелая цепь на одном конце имеет вариабельный домен (VH), за которым следует ряд константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен - на другом конце; константный домен легкой цепи совпадает с первым константным доменом тяжелой цепи, а вариабельный домен легкой цепи совпадает с вариабельным доменом тяжелой цепи. Считается, что определенные аминокислотные остатки образуют границу раздела между вариабельными доменами легкой и тяжелой цепей.
Термин «вариабельный» относится к тому, что последовательности некоторых участков вариабельных доменов сильно отличаются среди антител и используются для связывания и специфичности каждого конкретного антитела с его конкретным антигеном. Однако вариабельность не является равномерно распределенной по всем вариабельным доменам антител. Она сконцентрирована в трех сегментах, называемых гипервариабельными или областями, определяющими комплементарность, как в легкой цепи, так и в тяжелой цепи вариабельных доменов. Более высококонсервативные участки вариабельных доменов называются каркасными областями (FR). Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре FR, в основном имеющих β-складчатую конфигурацию, соединенных тремя гипервариабельными участками, которые образуют петли, соединяющие и в некоторых случаях образующие часть β-складчатой структуры. Гипервариабельные участки в каждой цепи удерживаются вместе в непосредственной близости с FR и с гипервариабельными участками другой цепи, участвуя в образовании антигенсвязывающего сайта антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Константные домены непосредственно не участвуют в связывании антитела с антигеном, но демонстрируют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности (ADCC).
Расщепление антител папаином дает два идентичных антигенсвязывающих фрагмента, называемых «Fab»-фрагментами, каждый с одним антигенсвязывающим сайтом, и остаточный «Fc»-фрагмент, название которого отражает его способность к легкой кристаллизации. Обработка пепсином дает F(ab')2 фрагмент, который имеет два антигенсвязывающих сайта и по-прежнему способен перекрестно связываться с антигеном.
«Fv» представляет минимальный фрагмент антитела, который содержит полный антигенраспознающий и антигенсвязывающий сайт. Этот участок состоит из димера вариабельного домена одной тяжелой цепи и одной легкой цепи в близкой, нековалентной связи. Он находится в такой конфигурации, что три гипервариабельных участка каждого вариабельного домена взаимодействуют с образованием антигенсвязывающего сайта на поверхности димера VH-VL. Совместно шесть гипервариабельных участков придают антителу антигенсвязывающую специфичность. Однако даже один вариабельный домен (или половина Fv, включающая только три гипервариабельных участка, специфичных к антигену) обладает способностью распознавать и связывать антиген, хотя с меньшей аффинностью, чем связывающий сайт целиком.
Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на карбоксильном конце CH1 домена тяжелой цепи, включая один или несколько цистеинов из шарнирной области антитела. В данном случае Fab'-SH является обозначением Fab', в котором цистеиновый остаток(и) константных доменов несет по меньшей мере одну свободную тиольную группу. F(ab')2 фрагменты антитела первоначально были получены в виде пары Fab'-фрагментов, которые имеют между собой шарнирные цистеины. Также известны другие химические соединения фрагментов антитела.
«Легкие цепи» антител (иммуноглобулинов) позвоночного любого вида можно отнести к одному из двух четко различаемых типов, названных каппа (κ) и лямбда (λ), на основе аминокислотных последовательностей их константных доменов.
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей антитела можно отнести к различным классам. Существует пять основных классов интактных антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них дополнительно можно разделить на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA и IgA2. Константные домены тяжелых цепей, которые соответствуют различным классам антител, называются α, δ, ε, γ и μ соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны.
«Одноцепочечные Fv» или «scFv» фрагменты антитела содержат VH и VL домены антитела, где эти домены находятся в одной полипептидной цепи. Предпочтительно, чтобы полипептид Fv дополнительно содержал полипептидный линкер между VH и VL доменами, который дает возможность scFv образовывать желаемую структуру для связывания антигена. Обзор по scFv см. у Plűckthun в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994).
Термин «диатела» относится к небольшим фрагментам антитела с двумя антигенсвязывающими сайтами, эти фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом (VL) легкой цепи в одной полипептидной цепи (VH - VL). Используя линкер, который является слишком коротким для образования пары между двумя доменами в одной цепи, домены вынуждены образовывать с комплементарными доменами другой цепи и создавать два антигенсвязывающих сайта. Более полно диатела описаны, например, в EP 404097; WO 93/11161; и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993).
Используемый здесь термин «моноклональное антитело» относится к антителу, полученному из популяции, по существу, гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных естественных мутаций, которые могут присутствовать в небольших количествах. Моноклональные антитела являются высокоспецифичными, являясь направленными на один антигенный сайт. Кроме того, в отличие от препаратов обычных (поликлональных) антител, которые обычно включают в себя различные антитела, направленные на различные детерминанты (эпитопы), каждое моноклональное антитело направлено на одну антигенную детерминанту. Помимо их специфичности, моноклональные антитела имеют преимущество в том, что они синтезируются гибридомной культурой, не загрязненной другими иммуноглобулинами. Определение «моноклональное» указывает на характер антитела, как полученного, по существу, из гомогенной популяции антител, и не рассматривается требующим продукции антитела каким-либо конкретным способом. Например, моноклональные антитела, используемые в соответствии с настоящим изоберетением, могут быть получены гибридомным способом, впервые описанным Kohler et al., Nature, 256:495 (1975), или могут быть получены способами рекомбинатной ДНК (см., например, U.S. Patent No. 4816567). «Моноклональные антитела» также можно выделить из фаговой библиотеки антител, используя методики, описанные, например, у Clackson et al., Nature, 352:624-628 (1991) and Marks et al., J. Mol. Biol., 222:581-597 (1991).
В контексте этого изобретения моноклональные антитела особенно включают в себя «химерные» антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям антител, полученных от конкретных видов, или принадлежащих конкретному классу или подклассу антител, тогда как остаток цепи (цепей) идентичен или гомологичен соответствующим последовательностям антител, полученных из других видов, или принадлежит другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют желаемую биологическую активность (патент США № 4816567; Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). В контексте изобретения представляющие интерес химерные антитела включают в себя «приматизированные» антитела, содержащие последовательности антигенсвязывающего вариабельного домена, полученные от нечеловекообразных приматов (например, Обезьяны Старого Света, такие как павианы, макаки-резус или яванские макаки), и последовательности константных областей, принадлежащих человеку (патент США № 5693780).
«Гуманизированные» формы антител, отличных от антител человека (например, мышиных), представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, не относящегося к иммуноглобулину человека. В основном, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельного участка реципиента замещены остатками из гипервариабельного участка, видов, не относящихся к человеку (донорное антитело), таких как мышь, крыса, кролик и нечеловекообразный примат, с желаемой специфичностью, аффинностью и активностью. В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека замещаются соответствующими остатками, не относящимися к иммуноглобулину человека. Кроме того, гуманизированные антитела могут содержать остатки, которые не обнаружены в реципиентном антителе или в донорном антителе. Эти модификации проводят для дополнительного совершенствования функциональной активности антитела. В основном, гуманизированное антитело будет содержать, по существу, все по меньшей мере из одного и, обычно, двух вариабельных доменов, в которых все, или по существу все, из гипервариабельных петлей соответствуют таковым иммуноглобулина, не относящегося к человеку, и все, или по существу все, FR являются таковыми с последовательностью иммуноглобулина человека. Гуманизированное антитело также необязательно будет содержать по меньшей мере часть константной области (Fc) иммуноглобулина, обычно иммуноглобулина человека. Более подробно см. у Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992).
При использовании в этом изобретении термин «гипервариабельный участок» относится к аминокислотным остаткам антитела, которые ответственны за связывание антигена. Гипервариабельный участок включает в себя аминокислотные остатки из «области, определяющей комплементарность» или «CDR» (например, остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)) и/или остатки из «гипервариабельной петли» (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (H1), 53-55 (H2) и 96-101 (H3) в тяжелой цепи вариабельного домена; Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). «Каркасными» или «FR» остатками являются остатки вариабельного домена, отличные от остатков гипервариабельного участка, определенного в настоящем описании.
Антитело, «которое связывает» представляющий интерес антиген, представляет собой антитело, способное связывать этот антиген с достаточной аффинностью и/или авидностью таким образом, что это антитело применимо в качестве терапевтического или диагностического средства для нацеливания на клетку, экспрессирующую этот антиген.
В целях настоящего изобретения «иммунотерапия» будет относиться к способу лечения млекопитающего (предпочтительно пациента человека) антителом, где антитело может быть неконъюгированным или «обнаженным» антителом, или антитело может быть конъюгировано или слито с гетерологичной молекулой (молекулами) или агентом (агентами), например, одним или несколькими цитотоксическими агентами, тем самым воспроизводя «иммуноконъюгат».
«Выделенное антитело» представляет собой антитело, которое было идентифицировано и отделено и/или восстановлено из компонента его природного окружения. Загрязняющими компонентами его природного окружения являются вещества, которые бы препятствовали его диагностическому или терапевтическому использованию этого антитела, и могут включать в себя ферменты, гормоны и другие белковоподобные и небелковые растворенные вещества. В предпочтительных вариантах осуществления антитело будет очищено (1) более чем до 95% по массе антитела, как определено методом Лоури, и наиболее предпочтительно более 99% по массе (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся стаканом, или (3) до гомогенности с помощью SDS-PAGE в редуцирующих или нередуцирующих условиях с использованием Кумасси голубого или, предпочтительно, окрашивания серебром. Выделенное антитело включает в себя антитело in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент природного окружения антитела не будет присутствовать. Как правило, однако, выделенное антитело будет получено по меньшей мере за одну стадию очистки.
Выражение «эффективное количество» относится к количеству средства (например, Apo2L/TRAIL, анти-DR4 или DR5 антитело и т.д.), которое является эффективным для профилактики, облегчения или лечения рассматриваемого заболевания или состояния.
Используемые здесь термины «лечение», «лечебное воздействие» и «терапия» относятся к лечебной терапии, профилактической терапии и превентивной терапии. Последовательное лечебное воздействие или введение относится по меньшей мере к лечебному воздействию на ежедневной основе без перерыва в лечении на один или несколько дней. Дробное лечебное воздействие или введение, или лечебное воздействие или введение в периодическом режиме относится к лечению, которое является не последовательным, а скорее периодическим по своей природе.
Термин «цитокин» является родовым термином для белков, секретируемых одной клеточной популяцией, которые действуют на другие клетки как межклеточные медиаторы. Примерами таких цитокинов являются лимфокины, монокины и обычные полипептидные гормоны. В число цитокинов входит гормон роста, например гормон роста человека; N-метионил гормона роста человека и гормон роста быка; паратиреоидный гормон; тироксин, инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (ФСГ), тиреотропный гормон (ТТГ) и лютеинизирующий гормон (ЛГ); печеночный фактор роста; фактор роста фибробластов; пролактин; плацентарный лактоген; фактор некроза опухоли -α и -β; мюллерова ингибирующая субстанция; мышиный пептид, связанный с гонадотропином; ингибин; активин; сосудистый эндотелиальный фактор роста; интегрин; тромбопоэтин (ТПО); факторы роста нервов; тромбоцитарный фактор роста; трансформирующие факторы роста (TGF), такие как TGF-α и TGF-β; инсулиноподобный фактор роста -I и -II; эритропоэтин (ЭПО); остеоиндуктивные факторы; интерфероны, такие как интерферон-α, -β и -γ; колониестимулирующие факторы (CSF), такие как макрофагальный CSF (M-CSF); гранулоцитомакрофагальный-CSF (GM-CSF) и гранулоцитарный-CSF (G-CSF); интерлейкины (IL), такие как IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-11, IL-12, IL-13, IL-17; и другие полипептидные факторы, включая LIF и кит-лиганд (KL). Использованный в описании термин цитокин включает в себя белки из природных источников или из рекомбинантной клеточной культуры и биологически активные эквиваленты нативных последовательностей цитокинов.
Используемый здесь термин «цитотоксическое средство» относится к веществу, которое ингибирует или препятствует функции клеток и/или вызывает деструкцию клеток. Этот термин включает в себя радиоактивные изотопы (например, I131, I125, Y90 и Re186), химиотерапевтические средства и токсины, такие как ферментативно активные токсины бактерий, грибов, растений или токсины животного происхождения, или их фрагменты.
«Химиотерапевтическое средство» представляет химическое соединение, пригодное для лечения опухолей. Примеры химиотерапевтических средств включают алкилирующие агенты, такие как тиотепа и циклофосфамид (CYTOXAN™); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквуон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогенины (главным образом, буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекан); бриостатин; каллистатин; CC-1065 (включая его адозелезиновые, карзелезиновые и бизелезиновые синтетические аналоги); криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги, KW-2189 и CBI-TMI); элеутеробин; панкратистатин; саркодистин; спонгистатин; азотистого иприта, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, меклоретамин, гидрохлорид оксида меклоретамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урамустин; нитрозомочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин, ранимустин; антибиотики такие, как энединовые антибиотики (например, каликеамицин, главным образом, каликеамицин гамма1I и каликеамицин phiI1, см., например, Agnew, Chem Intl. Ed. Engl., 33:183-186 (1994); динемицин, в том числе динемицин A; бисфосфонаты, такие как клодронат; эсперамицин; а также неокарзиностатин хромофор и родственные хромопротеин энединовым антибиотикам хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (AdriamycinTM) (в том числе морфолино-доксорубицин, цианоморфолино-доксорубицин, 2-пирролино-доксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин C, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-ФУ); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пуринов, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидинов, такие как анцитабин, азацитидин, 6-азауридин, карморфур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолон пропионат, эпитиостанол, мепитиостан, тестолактон; антагонисты гормонов надпочечников, такие как аминоглутетимид, митотан, трилостан; метаболит фолиевой кислоты, такой как фолиновая кислота; ацеглатон; альдофосфамидгликозид; аминолевулиновая кислота; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазихинон; элфорнитин; эллиптиния ацетат; эпотилон; этоглуцид; галлия нитрат; оксимочевина; лентинан; лонидамин; майтанзиноиды, такие как майтанзин и анзамитоцины; митогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновую кислоту; 2-этилгидразид; прокарбазин; PSK®; разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновая кислота; триазиквон; 2, 2',2''-трихлортриэтиламин; трихотецены (особенно T-2 токсин, верракурин A, роридин A и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ара-C»); циклофосфамид; тиотепа; таксоиды, например, паклитаксел (TAXOL®, Bristol-Myers Squibb Oncology, Princeton, NJ) и доксетаксел (TAXOTERE®, Rhône-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин (GemzarTM); 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин (NavelbineTM); новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселода; ибандронат; CPT-11; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; и фармацевтически приемлемые соли или производные любого из указанного выше. В это определение также включены антигормональные средства, которые действуют для регуляции или подавления действия гормонов на опухоли, такие как антиэстрогены и селективные модуляторы эстрогеновых рецепторов (SERM), в том числе, например, тамоксифен (в том числе NolvadexTM), ралоксифен, дропоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен (FarestonTM); ингибиторы ароматазы, которые ингибируют фермент ароматазу, который регулирует синтез эстрогена в надпочечниках, например, такие как 4(5)-имидазолы, аминоглутетимиды, мегестрола ацетат (MegaceTM), экземестан, форместан, фадрозол, ворозол (RivisorTM), летрозол (FemaraTM), и анастрозол (ArimidexTM); и анти-андрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; и фармацевтически приемлемые соли, кислоты и производные любого из изложенного выше.
«Ингибитор роста» при использовании в контексте настоящего изобретения относится к соединению или композиции, которая ингибирует рост клетки, главным образом, злокачественной клетки, гиперэкспрессирующей любой из указанных здесь генов, либо in vitro, либо in vivo. Таким образом, ингибитор роста представляет собой средство, которое существенно снижает процентное содержание клеток, гиперэкспрессирующих такие гены в S фазе. Примеры ингибиторов роста включают в себя средства, которые блокируют развитие клеточного цикла (не в S фазе), такие как средства, индуцирующие задержку G1 и M-фазы. Классические блокаторы M-фазы включают в себя винкас (винкристин и винбластин), таксол и ингибиторы топо II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Средства, которые задерживают G1, также переходят в задержку S-фазы, например, средства, алкилирующие ДНК, такие как тамоксифен, преднизон, дакарбазин, меклоретамин, цисплатин, метотрексат, 5-фторурацил и ара-С. Дополнительную информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Глава 1, под названием "Cell cycle regulation, oncogens, and antineoplastic drugs" авторов Murakami et al. (WB Saunders: Philadelphia, 1995), в основном, стр. 13.
Термины «апоптоз» и «апоптотическая активность» используются в широком смысле и относятся к упорядоченной или контролируемой форме клеточной гибели у млекопитающих, то есть обычно сопровождаемой одной или несколькими характерными клеточными изменениями, включая конденсацию цитоплазмы, утрату микроворсинок цитоплазматической мембраной, сегментацию ядра, деградацию хромосомной ДНК или утрату митохондриальной функции. Эту активность можно определить и измерить, например, исследованиями жизнеспособности клетки (таких как исследования с Аламаром синим или MTT), FACS анализом, активации каспазы, фрагментации ДНК (см., например, Nicoletti et al., J. Immunol. Methods, 139:271-279 (1991) и полимеразы поли-АДФ рибозы, «PARP», анализами расщепления, известными в уровне техники.
Используемый в описании термин «нарушение» в основном относится к любому состоянию, которое бы улучшилось в результате лечения описанными здесь композициями, в том числе любое заболевание или нарушение, которое можно лечить эффективными количествами Apo2L/TRAIL или анти-DR4 антитела, и/или анти-DR5 антитела. Он включает в себя хронические и острые нарушения, а также патофизиологические состояния, которые предрасполагают млекопитающего к рассматриваемому нарушению. Неограничивающие примеры нарушений, которые лечат в настоящем изобретении, включают в себя доброкачественные и злокачественные опухоли; воспалительные, ангиогенные и иммунологические нарушения, аутоиммунные нарушения, артрит (включая ревматоидный артрит), рассеянный склероз и ВИЧ/СПИД.
Термины «рак», «раковый» или «злокачественный» относятся к физиологическому состоянию млекопитающих, которое обычно характеризуется нерегулируемым клеточным ростом, или описывают такое состояние. Примеры злокачественных опухолей включают в себя, но не только, карциному, лимфому, лейкоз, бластому и саркому. Более конкретные примеры таких злокачественных опухолей включают в себя сквамозноклеточную карциному, миелому, мелкоклеточный рак легких, глиому, Ходжкинскую лимфому, нехождкинскую лимфому, рак желудочно-кишечного тракта, рак почек, рак яичников, рак печени, лимфобластный лейкоз, лимфоцитарный лейкоз, колоректальный рак, рак эндометрия, рак почек, рак предстательной железы, рак щитовидной железы, меланому, хондросаркому, нейробластому, рак поджелудочной железы, полиморфную глиобластому, рак шейки матки, рак головного мозга, рак желудка, рак мочевого пузыря, гепатому, рак груди, карциному кишечника и рак тканей головы и шеи.
Термин «заболевание, связанное с иммунной системой», означает заболевание, в котором компонент иммунной системы млекопитающего вызывает, опосредует или иным путем вносит вклад в заболеваемость млекопитающего. Также включены заболевания, в которых стимуляция или вмешательство иммунного ответа оказывает благоприятное воздействие на течение заболевания. В этот термин включены аутоиммунные заболевания, иммуно-опосредованные воспалительные заболевания, воспалительные заболевания, не опосредованные иммунной системой, инфекционные заболевания и иммунодефицитные заболевания. Примеры заболеваний, связанных с иммунной системой, и воспалительных заболеваний, некоторые из которых являются иммунологически опосредованными или опосредованными Т-клетками, которые можно лечить в соответствии с этим изобретением, включают в себя системную красную волчанку, ревматоидный артрит, хронический ювенильный артрит, спондилоартропации, системный склероз (склеродермия), идиопатические воспалительные миопатии (дерматомиозит, полимиозит), синдром Шегрена, системный васкулит, саркоидоз, аутоиммунную гемолитическую анемию, (иммунная панцитопения, пароксизмальная ночная гемоглобулинурия), аутоиммунную тромбоцитопению (идиопатическая тромбоцитопеническая пурпура, иммунологически опосредованная тромбоцитопения), тироидит (болезнь Грейвса, тироидит Хашимото, ювенильный лимфоцитарный тироидит, атрофический тироидит), сахарный диабет, иммунологически опосредованное заболевание почек (гломерулонефрит, тубулоинтерстициальный нефрит), демиелинизирующие заболевания центральной и периферической нервной системы, такие как рассеянный склероз, идиопатическая демиелинизирующая полинейропатия или синдром Жиллиана-Барра и хроническая демилиенизирующая полинейропатия, гепатобиллиарные заболевания, такие как инфекционный гепатит (гепатиты A, B, C, D, E и другие негепатотропные вирусы), аутоиммунный хронический активный гепатит, первичный биллиарный цирроз, грануломатозный гепатит и склерозирующий холангит, воспалительные и фиброзные заболевания легких, такие как воспалительное заболевание кишечника (язвенный колит: болезнь Крона), глютеновая энтеропатия и болезнь Уипла, аутоиммунные или иммунологически опосредованные заболевания кожи, включая буллезные заболевания кожи, полиморфную эритему и контактный дерматит, псориаз, аллергические заболевания, такие как астма, аллергический ринит, атопический дерматит, пищевая сенсибилизация и крапивница, иммунологические заболевания легких, такие как эозинофильная пневмония, идиопатический легочный фиброз и гиперчувствительный пневмонит, заболевания, связанные с трансплантацией, в том числе отторжение трансплантата и заболевание трансплантат-против-хозяина. Инфекционные заболевания включают в себя СПИД (ВИЧ инфекцию), гепатит A, B, C, D и E, бактериальные инфекции, грибковые инфекции, протозойные инфекции и паразитические инфекции.
«Аутоиммунное заболевание» используется в описании в широком общем смысле для обозначения нарушений или состояний у млекопитающих, при котором разрушение нормальной или здоровой ткани происходит в результате гуморальной или клеточной иммунной реакции отдельного млекопитающего на компоненты его или ее собственных тканей. Примеры включают в себя, но не только, системную красную волчанку, тироидит, ревматоидный артрит, псориаз, рассеянный склероз, аутоиммунный диабет и воспалительное заболевание кишечника (IBD).
Термин «меченый» при использовании в настоящем изобретении относится к химерной молекуле, содержащей антитело или полипептид, конденсированный с «таг-полипептидом». Таг-полипептид имеет достаточно остатков для предоставления эпитопа, в отношении которого может быть получено антитело, или для обеспечения некоторых других функций, таких как способность к олигомеризации (например, как происходит с пептидами, имеющими домены лейциновых застежек), тем не менее достаточно короткий, так что в основном не препятствует активности антитела или полипептида. Таг-полипептид предпочтительно также является в известной степени уникальным, так что таг-специфическое антитело, по существу, перекрестно не реагирует с другими эпитопами. Подходящие таг-полипептиды в основном имеют по меньшей мере шесть аминокислотных остатков и обычно примерно от 8 примерно до 50 аминокислотных остатков (предпочтительно, примерно от 10 примерно до 20 остатков).
Термин «двухвалентный ион металла» относится к иону металла, имеющему два положительных заряда. Примеры двухвалентных ионов металла включают в себя, но не только, цинк, кобальт, никель, кадмий, магний и марганец. Особые формы таких металлов, которые могут быть использованы, включают в себя солевые формы (например, фармацевтически приемлемые солевые формы), такие как формы хлорида, ацетата, карбоната, цитрата и сульфата вышеупомянутых двухвалентных ионов металла. Необязательно, двухвалентный ион металла для использования в настоящем изобретении представляет собой цинк и, предпочтительно, солевую форму, сульфат цинка или хлорид цинка.
«Выделенный» при использовании для описания различных пептидов или белков, раскрытых в этом изобретении, означает пептид или белок, который был идентифицирован или отделен и/или извлечен из компонент его природного окружения. Загрязняющие компоненты его природного окружения представляют собой вещества, которые обычно препятствовали бы его диагностическим или терапевтическим применениям этого пептида или белка, и могут включать в себя ферменты, гормоны и другие белковоподобные или небелковые растворенные вещества. В предпочтительных вариантах осуществления этот пептид или белок будет очищен (1) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся стаканом, или (2) до гомогенности с помощью SDS-PAGE в редуцирующих или нередуцирующих условиях с использованием Кумасси голубого или, предпочтительно, окрашивания серебром, или (3) до гомогенности с помощью масс-спектроскопических способов или методик пептидного картирования. Выделенное вещество включает в себя пептид или белок in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент из природного окружения не будет присутствовать. Как правило, однако, выделенный пептид или белок будет получен по меньшей мере за одну стадию очистки.
«Процент (%) идентичности аминокислотных последовательностей» в отношении последовательностей, идентифицированных в настоящем изобретении, определяется как процентное содержание аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам в эталонной последовательности, после выравнивания последовательностей и, при необходимости, введения разрывов для достижения максимальной процентной идентичности последовательностей, и не принимая во внимание какие-либо консервативные замены как часть идентичности последовательностей. Выравнивания с целью определения процентной аминокислотной идентичности можно достичь различными способами, которые относятся к специальным знаниям в этой области, можно определить соответствующие параметры для определения выравнивания, включая задание алгоритмов, необходимых для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Для целей настоящего изобретения процентные значения аминокислотной идентичности могут быть получены с использованием компьютерной программы для сравнения последовательностей, ALIGN-2, которая разработана Genentech, Inc. и исходная программа которой была заявлена пользовательской документацией в Ведомстве по охране авторских прав США, Вашингтон, DC, 20559, зарегистрированной в соответствии с регистрацией авторских прав США № TXU510087. Программа ALIGN-2 общедоступна через Genentech, Inc., South San Francisco, CA. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не изменяются.
«Строгость» реакций гибридизации легко определяется специалистом в данной области и в основном представляет собой эмпирический расчет в зависимости от длины зонда, температуры промывки и концентрации соли. Обычно более длинные зонды требуют более высоких температур для надлежащего отжига, тогда как для более коротких зондов нужны более низкие температуры. Гибридизация в основном зависит от способности денатурированной ДНК к повторному отжигу, когда комплементарныхе цепи находятся в среде ниже их температуры плавления. Чем выше степень желаемой идентичности между зондом и гибридируемой последовательностью, тем выше относительная температура, которая может быть использована. Как результат, следует, что более высокие относительные температуры имели бы тенденцию делать условия реакции более строгими, тогда как более низкие температуры - менее. Дополнительные подробности и объяснения строгости условий гибридизации см. у Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers (1995).
«Условия высокой строгости», определенные в настоящем изобретении, определяются тем, что: (1) используется низкая ионная сила и высокая температура для промывки; 0,015 M хлорид натрия/0,0015, M цитрат натрия/0,1%, додецил сульфат натрия при 50oC; (2) во время гибридизации используется денатурирующий агент; 50% (об./об.) формамид с 0,1% бычьим сывороточным альбумином/0,1%, Фиколл/0,1% поливинилпирролидон/50 мM, фосфатно-натриевый буфер при pH 6,5 с 750 мM хлорида натрия, 75 мM цитрата натрия при 42oC; или (3) используется 50% формамид, 5 × SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мM фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5 × раствор Денхардта, ДНК спермы лосося, подвергнутая действию ультразвука (50 мкг/мл), 0,1% SDS и 10% сульфат декстрана при 42oC с промывками при 42oC в 0,2 × SSC (хлорид натрия/цитрат натрия) и 50% формамид при 55oC с последующей промывкой высокой строгости, состоящей из 0,1 × SSC, содержащих EDTA при 55oC.
«Условия умеренной строгости» могут быть определены, как описано у Sambrook et al., Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989, и включают в себя инкубацию в течение ночи при 37oC в растворе, содержащем: 20% формамид, 5 × SSC (150 мM NaCl, 15 мM тринатриевый цитрат), 50 мM фосфат натрия (pH 7,6), 5 × раствор Денхардта, 10% сульфат декстрана и 20 мг/мл денатурированной ДНК спермы лосося с последующей промывкой фильтров в 1 × SSC примерно при 37-50oC. Специалисту в данной области будет понятно, как установить температуру, ионную силу и т.д., необходимые для приведения в соответствие таких факторов, как длина зонда и подобное.
Термин «праймер» или «праймеры» относится к олигонуклеотидным последовательностям, которые гибридизируются с комплементарным целевым полинуклеотидом РНК или ДНК и служат в качестве отправных точек для поэтапного синтеза полинуклеотида из мононуклеотидов под действием нуклеотидилтрансферазы, например, как происходит в полимеразно-цепной реакции.
Термин «контрольные последовательности» относится к последовательностям ДНК, необходимым для экспрессии оперативно связанной кодирующей последовательности в конкретном организме-хозяине. Контрольные последовательности, подходящие для прокариот, например, включают в себя промотор, необязательно операторную последовательность и сайт связывания с рибосомой. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является «оперативно связанной», когда она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК для предпоследовательности или секреторного лидера является оперативно связанной с ДНК для полипептида, если она экспрессируется в виде препробелка, который участвует в секреции полипептида; промотор или энхансер является оперативно связанным с кодирующей последовательностью, если он влияет на транскрипцию этой последовательности; или сайт связывания с рибосомой является оперативно связанным с кодирующей последовательностью, если он расположен так, что облегчает трансляцию. В основном, «оперативно связанная» означает, что последовательности ДНК, будучи связанными, являются непрерывными и в случае секреторного лидера непрерывными и на стадии считывания. Однако энхансеры не должны быть непрерывными. Связывание осуществляется лигированием в подходящих сайтах рестрикции. Если таких сайтов нет, используют синтетические олигонуклеотидные адапторы или линкеры, в соответствии с общепринятой практикой.
«Антитело-зависимая опосредованная клетками цитотоксичность» или «ADCC» относится к клеточно-опосредованной реакции, в которой неспецифические цитотоксические клетки, которые экспрессируют Fc рецепторы (FcR) (например, естественные киллеры (NK), нейтрофилы и макрофаги), распознают связанное антитело на клетке-мишени и впоследствии вызывают лизис клетки-мишени. Первичные клетки для опосредования ADCC, NK клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках в суммированном виде предствляет собой Таблицу 3 на странице 464 публикации Ravetch и Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Для оценки ADCC активности, представляющей интерес молекулы, можно провести анализ ADCC in vitro, например, описанный в патенте США № 5500362 или 5821337. Подходящие эффекторные клетки для таких исследований включают в себя мононуклеарные клетки периферической крови (PBMC) и клетки естественные киллеры (NK). Альтернативно или дополнительно ADCC активность молекулы, представляющей интерес, можно оценить activity in vivo, например, в модели на животных, например, описанной у Clynes et al. PNAS (USA) 95:652-656 (1998).
«Эффекторные клетки человека» представляют собой лейкоциты, которые экспрессируют один или несколько FcR и выполняют эффекторные функции. Предпочтительно, клетки, экспрессирующие по меньшей мере FcγRIII и выполняющие эффекторную функцию ADCC. Примеры лейкоцитов человека, которые опосредуют ADCC, включают в себя мононуклеарные клетки периферической крови (PBMC), клетки естественные киллеры (NK), моноциты, цитотоксические Т клетки и нейтрофилы; с наиболее предпочтительными PBMC и NK клетками.
Термины «Fc рецептор» или «FcR» используют для описания рецептора, который связывается с Fc участком антитела. Предпочтительный FcR представляет собой нативную последовательность FcR человека. Более того, предпочтительным FcR является рецептор, который связывается с IgG антителом (гамма рецептор) и включает в себя рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и формы альтернативного сплайсинга этих рецептров. FcγRII рецепторы включают в себя FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые имеют сходные аминокислотные последовательности, которые отличаются главным образом своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит мотив, активирующий иммунорецептор, основанный на тирозине (ITAM) в своем цитоплазматическом домене. Ингибирующий рецептор FcγRIIB в своем цитоплазматическом домене содержит мотив, ингибирующий иммунорецептор, основанный на тирозине (ITIM). (См. Daëron, Annu. Rev. Immunol. 15:203-234 (1997)). Обзор по FcR представлен у Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). Другие FcR, в том числе те, которые будут установлены в будущем, здесь охватываются термином «FcR». Этот термин также включает в себя неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG к плоду (Guyer et al., J. Immunol. 117:587 (1976) and Kim et al., J. Immunol. 24:249 (1994)). В настоящем изобретении FcR включают в себя полиморфизмы, такие как генетический диморфизм в гене, который кодирует FcγRIIIa, приводящий, в результате, либо к фенилаланину (F), либо валину (V) в аминокислотном положении 158, расположенном в той области рецептора, которая связывается с IgG1. Было показано, что гомозиготные FcγRIIIa с валином (FcγRIIIa-158V) имеют более высокую аффинность к IgG1 человека и опосредуют повышенную ADCC in vitro по сравнению с гомозиготными FcγRIIIa с фенилаланином (FcγRIIIa-158F) или гетерозиготными (FcγRIIIa-158F/V) рецепторами.
«Комплемент-зависимая цитотоксичность» или «CDC» относится к способности молекулы лизировать мишень в присутствии комплемента. Путь активации комплемента запускается связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом), образовавшей комплекс с родственным антигеном. Для оценки активации комплемента можно провести анализ CDC, например, как описано у Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996).
II. СПОСОБЫ И МАТЕРИАЛЫ ИЗОБРЕТЕНИЯ, ПРИВОДИМЫЕ В КАЧЕСТВЕ ПРИМЕРА
Способы и исследования, описанные в настоящем описании, направлены на изучение экспрессии одного или нескольких биомаркеров в образце ткани млекопитающего или образце клеток, где определение экспрессии одного или нескольких таких биомаркеров прогнозирует или указывает, будет ли образец ткани или клеток чувствительным к таким агентам, как Apo2L/TRAIL и/или антителам рецепторов смерти, таким как антитела-агонисты к DR5 или антитела-агонисты к DR4. Эти способы и исследования включают в себя способы и исследования, которые изучают экспрессию представителей семейства молекул GalNac-T, в том числе GalNac-T14 и GalNac-T3.
Как рассматривалось выше, существует несколько популяций патологических типов клеток человека (например, определенные популяции злокачественных клеток), которые резистентны к действиям Apo2L/TRAIL, индуцирующим клеточную гибель, в том числе к действиям антител рецепторов смерти. Следовательно, считается, что описанные способы и исследования могут обеспечить подходящие, эффективные и потенциально рентабельные средства получения данных и информации, используемой для оценки адекватных или эффективных схем для лечения пациентов. Например, у пациента, у которого диагностировано злокачественное заболевание или патологическое состояние, связанное с иммунной системой, можно было бы взять биопсию для получения образца ткани или клеток, и этот образец можно было бы проверить с помощью различных анализов in vitro для определения, будут ли клетки пациента чувствительны к терапевтическому средству, такому как Apo2L/TRAIL или антитело рецептора смерти.
Изобретение относится к способам прогноза чувствительности образца ткани или клеток млекопитающего (например, злокачественных клеток) к Apo2L/TRAIL или антителу-агонисту рецептора смерти. Необязательно, получают образец ткани или клеток млекопитающих и исследуют на экспрессию GalNac-T14. Эти способы можно проводить в различных форматах исследований, включая исследования, определяющие экспрессию мРНК, экспрессию белка (например, иммуногистохимические исследования) и биохимические анализы, выявляющие ферментативную активность UDP-N-ацетил-D-галактозамин:полипептид N-ацетилгалактозаминилтрансферазы. Выявление экспрессии таких GalNac-T14 биомаркеров в (или на) указанных тканях или клетках, будет прогнозом того, что такие ткани или клетки будут чувствительными к биологическим эффектам Apo2L/TRAIL и/или антитела рецептора смерти. Заявители неожиданно обнаружили, что экспрессия GalNac-T14 коррелирует с чувствительностью таких тканей и клеток к Apo2L/TRAIL и антителам-агонистам рецепторов смерти.
Как рассмотрено ниже, экспрессия различных биомаркеров, таких как GalNac-T14 в образце, можно проанализировать с помощью ряда методик, многие из которых известны в уровне техники и понятны специалистам, в том числе, но не только, иммуногистохимический и/или Вестерн анализ, количественные анализы крови (как, например, Serum ELISA) (например, для исследования уровней экспрессии белка), биохимические исследования ферментативной активности, гибридизация in situ, Нозерн анализ и/или ПЦР анализ мРНК и геномный Саузерн анализ (например, для исследования делеции или амплификации гена), а также любым из разнообразных способов анализа, которые можно проводить с помощью матричного анализа генов и/или тканей. Стандартные протоколы для оценки статуса генов или генных продуктов находятся, например, в публикации Ausubel et al. eds., 1995, Current Protocols In Molecular Biology, Units 2 (Нозерн блоттинг), 4 (Саузерн блоттинг), 15 (Иммуноблоттинг) и 18 (ПЦР анализ).
Приведенные ниже протоколы, относящиеся к выявлению GalNac-T14 в образце, представлены ниже с целью иллюстрации.
*************************************
Необязательные способы по изобретению включают в себя протоколы, которыми проверяют или тестируют наличие GalNac-T14 в образце ткани или клеток млекопитающих. Для выявления GalNac-T14 могут быть использованы разнообразные способы, и они включают в себя, например, иммуногистохимический анализ, иммунопреципитацию, Вестерн блот анализ, исследования молекулярного связывания, ELISA, ELIFA, активируемую флуоресценцией сортировку клеток (FACS) и иммунопреципитацию с последующим MS, анализом моносахаридов. Например, необязательный способ выявления экспрессии GalNac-T14 в ткани или образце включает в себя контактирование образца с анти-GalNac-T14 антителом, а затем выявление связывания антитела с GalNac-T14 в образце.
В конкретных вариантах осуществления изобретения экспрессию GalNac-T14 в образце исследуют с использованием иммуногистохимии протоколов окрашивания. Было показано, что иммуногистохимическое окрашивание срезов ткани является надежным способом оценки или выявления наличия белков в образце. В иммуногистохимических методиках («IHC») используют антитела для зондирования и визуализации клеточных антигенов in situ, главным образом с помощью хромогенных или флуоресцентных способов.
Для получения образца может быть использован образец ткани или клеток млекопитающего (обычно пациента человека). Примеры образцов включают в себя, но не только, злокачественные клетки, такие как злокачественные клетки кишечника, груди, предстательной железы, яичника, легких, желудка, поджелудочной железы, лимфомы и лейкозные клетки. Необязательно образцы включают в себя злокачественные клетки немелкоклеточного рака легких, злокачественные клетки поджелудочной железы или злокачественные клетки неходжкинской лимфомы. Образец может быть получен с помощью ряда методик, известных в уровне техники, в том числе, но не только, хирургическим путем, аспирацией или биопсией. Ткань может быть свежей или замороженной. В одном варианте осуществления образец фиксируют и заключают в парафин, или подобное.
Образец ткани может быть фиксирован (т.е. консервирован) с помощью общепринятой методики (см., например, “Manual of Histological Staining Method of the Armed Forces Institute of Pathology,” 3rd edition (1960) Lee G. Luna, HT (ASCP) Editor, The Blakston Division McGraw-Hill Book Company, New York; The Armed Forces Institute of Pathology Advanced Laboratory Methods in Histology and Pathology (1994) Ulreka V. Mikel, Editor, Armed Forces Institute of Pathology, American Registry of Pathology, Washington, D.C.). Специалисту в данной области будет понятно, что выбор фиксатора зависит от цели гистологического окрашивания или иного исследования этого образца. Специалисту в данной области также будет понятно, что длительность фиксации зависит от размера образца ткани и используемого фиксатора. В качестве примера, для фиксации образца может быть использован нейтральный забуференный формалин, фиксатор Буэна или параформальдегид.
В основном, сначала образец фиксируют, а затем дегидрируют посредством возрастающих серий спиртов, пропитывают и заключают в парафин или другую среду для изготовления срезов таким образом, чтобы из образца ткани можно было получить срез. Альтернативно, можно сделать срез ткани и фиксировать полученные срезы. В качестве примера образец ткани может быть заключен и обработан в парафине с помощью общепринятой методики (см., например, “Manual of Histological Staining Method of the Armed Forces Institute of Pathology", supra). Примеры парафина, который может быть использован, включают в себя, но не ограничиваются ими, Paraplast, Broloid и Tissuemay. Как только образец ткани заключен в парафин, можно делать срезы этого образца с помощью микротома или подобного (см., например, “Manual of Histological Staining Method of the Armed Forces Institute of Pathology”, supra). В качестве примера этой методики, толщина срезов может варьировать примерно от трех микрон примерно до пяти микрон. После получения срезы можно присоединить к предметным стеклам несколькими стандартными способами. Примеры адгезивов для предметных стекол включают в себя, но не только, силан, желатин, поли-L-лизин и подобное. В качестве примера срезы, заключенные в парафин, можно присоединить к положительно заряженным предметным стеклам и/или предметным стеклам, покрытым поли-L-лизином.
При использовании парафина в качестве вещества для заключения препарата срезы тканей, как правило, освобождают от парафина и регидрируют в воде. Образцы ткани можно освободить от парафина с использованием нескольких общепринятых стандартных методик. Например, могут быть использованы ксилены и градиентно снижающиеся серии спиртов (см., например, “Manual of Histological Staining Method of the Armed Forces Institute of Pathology”, supra). Альтернативно, можно использовать коммерчески доступные депарафинизирующие неорганические агенты, такие как Hemo-De7 (CMS, Houston, Texas).
Необязательно, после приготовления образца, срез ткани можно анализировать с помощью IHC. IHC можно проводить в сочетании с дополнительными методиками, такими как морфологическое окрашивание и/или флуоресцентная гибридизация in situ. Существует два основных способа IHC; прямой и непрямой анализ. В соответствии с первым анализом, связывание антитела с целевым антигеном (например, GalNac-T14) определяют напрямую. В этом прямом анализе используется меченый реактив, такой как флуоресцентная метка, или меченное ферментом первичное антитело, который можно визуализировать без дальнейшего взаимодействия антитела. В стандартном непрямом анализе неконъюгированное первичное антитело связывается с антигеном, а затем меченое вторичное антитело связывается с первичным антителом. Там, где вторичное антитело конъюгировано с ферментной меткой, добавляют хромогенный или флуорогенный субстрат для обеспечения визуализации антигена. Происходит амплификация сигнала, поскольку несколько вторичных антител могут взаимодействовать с различными эпитопами на первичном антителе.
Первичное и/или вторичное антитело, используемое в иммуногистохимии, обычно будет мечено детектируемой молекулой. Существуют многочисленные метки, которые в основном можно объединить в следующие категории:
(a) Радиоизотопы, такие как 35S, 14C, 125I, 3H и 131I. Антитело может быть мечено радиоизотопом с использованием методик, описанных в Current Protocols in Immunology, Volumes 1 and 2, Coligen et al., Ed. Wiley-Interscience, New York, New York, Pubs. (1991), например, и радиоактивность можно измерить с помощью сцинтилляционного счета.
(b) Частицы коллоидного золота.
(c) Флуоресцентные метки, включающие в себя, но не только, хелаты редкоземельных металлов (хелаты европия), техасский красный, родамин, флуоресцеин, дансил, лиссамин, умбеллиферон, фикоэритерин, фикоцианин или коммерчески доступные флуорофоры, такие как SPECTRUM ORANGE7 и SPECTRUM GREEN7 и/или производные любого одного или нескольких, указанного выше. Флуоресцентные метки могут быть конъюгированы с антителом с помощью методик, описанных, например, в Current Protocols in Immunology, supra. Флуоресценцию можно измерить количественно с использованием флуориметра.
(d) Существуют различные фермент-субстратные метки, и в патенте США № 4275149 представлен обзор некоторых из них. Фермент, главным образом, катализирует химическое изменение хромогенного субстрата, которое может быть измерено с использованием различных методик. Например, фермент может катализировать изменение цвета в субстрате, которое можно количественно измерить спектрофотометрически. Альтернативно, фермент может изменять флуоресценцию или хемилюминесценцию субстрата. Методики количественного определения изменения флуоресценции описаны выше. Происходит электронное возбуждение хемилюминесцентного субстрата под действием химической реакции, а затем может излучать свет, который можно определить количественно (например, используя хемилюминометр) или передает энергию акцептору флуоресценции. Примеры ферментных меток включают в себя люциферазы (например, люциферазы светлячков и бактериальная люцифераза; патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназа, уреаза, пероксидаза, такая как пероксидаза хрена (HRPO), щелочная фосфатаза, β-галактозидаза, глюкоамилаза, лизозим, оксидазы сахаридов (например, глюкозоксидаза, галактозоксидаза и глюкозо-6-фосфат дегидрогеназа), гетероциклические оксидазы (такие как уриказа и ксантиноксидаза), лактопероксидаза, микропероксидаза, и подобное. Методики конъюгации ферментов с антителами описаны у O'Sullivan et al., Methods for the Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay, in Methods in Enzym. (ed. J. Langone & H. Van Vunakis), Academic press, New York, 73:147-166 (1981).
Примеры фермент-субстратных сочетаний включают в себя, например:
(i) пероксидазу хрена (HRPO) с перекисью водорода в качестве субстрата, где перекись водорода окисляет окрашивающий предшественник (например, ортофенилендиамин (OPD) или 3,3',5,5'-тетраметилбензидингидрохлорид (TMB));
(ii) щелочную фосфатазу (AP) с пара-нитрофенилфосфатом в качестве хромогенного субстрата; и
(iii) β-D-галактозидазу (β-D-Gal) с хромогенным субстратом (например, п-нитрофенил-β-D-галактозидазой) или флуорогенным субстратом (например, 4-метилумбеллиферил-β-D-галактозидазой).
Другие многочисленные фермент-субстратные сочетания доступны специалистам в данной области. Для общего обзора которых см. патенты США №№ 4275149 и 4318980. Иногда метка опосредованно конъюгирована с антителом. Различные методики для достижения этого находятся в компетенции специалиста. Например, антитело может быть конъюгировано с биотином, и любая из четырех широких категорий меток, упомянутых выше, может быть конъюгирована с авидином, или vice versa. Биотин специфически связывается с авидином, и, следовательно, эта метка может быть конъюгирована с антителом таким непрямым способом. Альтернативно, для достижения непрямой конъюгации метки с антителом, антитело конъюгируют с небольшим гаптеном, а одну из различных типов меток, упомянутых выше, конъюгируют с антителом к гаптену. Таким образом, можно достичь непрямой конъюгации метки с антителом.
Помимо методик получения образца, описанных выше, может быть целесообразна дополнительная обработка среза ткани до, во время или после IHC, например, методы воспроизведения эпитопов, например, можно провести нагревание образца ткани в цитратном буфере (см., например, Leong et al. Appl. Immunohistochem. 4(3):201 (1996)).
После необязательной стадии блокировки срез ткани подвергают действию первичного антитела в течение достаточного периода времени и в подходящих условиях так, чтобы первичное антитело связалось с целевым белковым антигеном в образце ткани. Соответствующие условия для достижения этого можно определить проведением стандартных экспериментов. Степень связывания антитела с образцом определяется использованием любой из детектируемых меток, рассмотренных выше. Предпочтительно, метка представляет собой ферментную метку (например, HRPO), которая катализирует химическое изменение хромогенного субстрата, такого как 3,3'-диаминобензидинхромоген. Предпочтительно, ферментную метку конъюгируют с антителом, которое специфически связывается с первичным антителом (например, первичное антитело представляет собой кроличье поликлональное антитело, а вторичное антитело представляет собой козье антикроличье антитело).
Необязательно, антитела, используемые в IHC анализе для выявления экспрессии GalNac-T14, представляют собой анти-GalNac-T14 антитела. Альтернативно, могут быть использованы антитела к другим GalNac-T антигенам, которые перекрестно реагируют с GalNac-T14. Необязательно, анти-GalNac-T14 антитело представляет собой моноклональное антитело.
Таким образом приготовленные образцы могут быть заключены в среду и заклеены покровными стеклами. Затем можно провести оценку гистологического препарата, например, используя микроскоп, и критерии интенсивности окрашивания, обычно используемые в данной области. Критерии интенсивности окрашивания можно оценить следующим образом:
Обычно считается, что балл окрашивания образца примерно 2+ или выше в таком IHC исследовании является прогнозом или показателем чувствительности клетки млекопитающего (например, злокачественной клетки млекопитающего) к Apo2L/TRAIL антителу-агонисту рецептора смерти.
В альтернативных способах образец может контактировать с антителом, специфичным к указанному биомаркеру, в условиях, достаточных для образования комплекса антитело-биомаркер, а затем выявление указанного комплекса. Наличие биомаркера можно определить рядом способов, например Вестерн-блоттингом (с иммунопреципитацией или без нее) и ELISA для оценки большого разнообразия тканей и образцов, включая плазму или сыворотку. Существует большое количество способов иммунологического анализа с использованием такого формата исследования, см., например, патенты США №№ 4016043, 4424279 и 4018653. Они включают в себя как односайтовый, так и двухсайтовый или «сэндвич» анализ неконкурентных типов, а также традиционные конкурентные анализы связывания. Эти исследования также включают в себя прямое связывание меченого антитела с целевым биомаркером.
Среди наиболее подходящих и обычно используемых находятся сэндвич-анализы. Существует ряд вариантов методики проведения сэндвич-анализа, и все они охватываются настоящим изобретением. Кратко, в обычном прямом анализе немеченое антитело иммобилизируют на твердом субстрате, а тестируемый образец приводят в контакт со связанной молекулой. После подходящего периода инкубации в течение периода времени, достаточного для образования комплекса антитело-антиген, затем добавляют второе антитело, специфичное к этому антигену, меченное репортерной молекулой, способной давать детектируемый сигнал, и инкубируют, предоставляя время, достаточное для образования другого комплекса антитело-антиген-меченое антитело. Любое непрореагировавшее вещество смывают, а наличие антигена определяют путем наблюдения сигнала, генерируемого репортерной молекулой. Результаты могут быть либо качественными, просто наблюдением видимого сигнала, либо могут быть количественными, путем сравнения с контрольным образцом, содержащим известные количества биомаркера.
Варианты прямого анализа включают в себя одновременный анализ, в котором и образец, и меченное антитело добавляют одновременно к связанному антителу. Эти методики хорошо известны специалистам в этой области, включая любые минимальные изменения, которые будут совершенно очевидны. В стандартном прямом сэндвич-анализе первое антитело, имеющее специфичность к биомаркеру, является либо ковалентно, либо пассивно связанным с твердой поверхностью. Твердая поверхность обычно представляет собой стекло или полимер, полимерами, используемыми чаще всего, являются целлюлоза, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен. Твердые подложки могут быть в форме пробирок, бус, дисков или микропланшетов, или любой другой поверхности, подходящей для проведения иммуноанализа. Процессы связывания хорошо известны в уровне техники и в основном состоят из перекрестного ковалентного связывания или физической адсорбции, комплекс полимер-антитело промывают в препарате для тестирования образца. Затем добавляют аликвоту тестируемого образца к твердофазному комплексу и инкубируют в течение периода времени, достаточного (например, 2-40 минут или в течение ночи, если это удобнее), и в соответствующих условиях (например, от комнатной температуры до 40oC, например, от 25oC до 32oC включительно) для обеспечения связывания любой субъединицы, присутствующей в антителе. После периода инкубации субъединицу антитела в твердой фазе промывают и сушат, и инкубируют со вторым антителом, специфичным к части биомаркера. Второе антитело связывается с репортерной молекулой, которую используют для индикации связывания второго антитела с молекулярным маркером.
Альтернативный способ включает в себя иммобилизацию целевых биомаркеров в образце, а затем воздействие на иммобилизованную мишень специфического антитела, которое может быть мечено или может не быть мечено репортерной молекулой. В зависимости от количества мишени и силы сигнала репортерной молекулы связанную мишень можно выявить непосредственным мечением антителом. Альтернативно на второе меченое антитело, специфичное к первому антителу, действует комплекс мишень-первое антитело с образованием третичного комплекса мишень-первое антитело-второе антитело. Этот комплекс выявляют с помощью сигнала, испускаемого репортерной молекулой. Под «репортерной молекулой» в настоящем изобретении подразумеваются молекулы, которые своей химической природой обеспечивают аналитически идентифицируемый сигнал, который позволяет осуществлять выявление антиген-связанное антитело. Наиболее часто используемыми репортерными молекулами в этом типе исследования являются либо ферменты, флуорофоры, либо молекулы, содержащие радионуклеиды (т.е. радиоизотопы) и хемилюминесцентные молекулы.
В случае иммуноферментного анализа фермент конъюгируют со вторым антителом, обычно с помощью глютаральдегида или периодата. Однако без труда будет понятно, что существует широкое разнообразие различных методик конъюгации, которые легко доступны специалистам. Обычно используемые ферменты среди прочих включают в себя пероксидазу хрена, глюкозоксидазу, галактозидазу и щелочную фосфатазу. Субстраты, используемые со специфическими ферментами, главным образом, выбирают для получения при гидролизе под действием соответствующего фермента детектируемого изменения цвета. Примеры подходящих ферментов включают в себя щелочную фосфатазу и пероксидазу. Также можно использовать флуорогенные субстраты, которые дают флуоресцентный продукт, а не хромогенные субстраты, отмеченные выше. Во всех случаях, антитело, меченное ферментом, добавляют к первому комплексу антитело-молекулярный маркер, дают возможность для связывания, а затем смывают избыток реактива. Затем добавляют раствор, содержащий соответствующий субстрат, к комплексу антитело-антиген-антитело. Субстрат будет взаимодействовать с ферментом, связанным со вторым антителом, давая качественный визуальный сигнал, который может быть в дальнейшем определен количественно, обычно спектрофотометрически, для получения указания на количество биомаркера, который присутствовал в образце. Альтернативно, флуоресцентные соединения, такие как флуоресцеин и родамин, могут быть химически соединены с антителами без изменения их связывающей способности. При активации освещением светом определенной длины волны, антитело, меченное флуорохромом, поглощает световую энергию, вызывая в молекуле состояние возбуждения, с последующей эмиссией света характерного цвета, визуально детектируемого в световой микроскоп. Как в EIA, флуоресцентно меченому антителу обеспечивают связывание с первым комплексом антитело-молекулярный маркер. После отмывания несвязанного реагента оставшийся третичный комплекс затем подвергают воздействию света соответствующей длины волны, наблюдаемая флуоресценция указывает на присутствие молекулярного маркера, представляющего интерес. Как методики иммунофлуоресценции, так и EIA являются общепринятыми в этой области. Однако другие репортерные молекулы, такие как радиоизотопы, хемилюминесцентные или биолюминесцентные молекулы также могут быть использованы.
Предполагается, что описанные выше методики также могут быть использованы для выявления экспрессии GalNac-T14.
Способы по изобретению дополнительно включают в себя протоколы, которые исследуют наличие и/или экспрессию мРНК GalNac-T14 в образце ткани или клеток. Способы оценки мРНК в клетках хорошо известны и включают в себя, например, гибридизационные анализы с использованием комплементарных ДНК зондов (например, in situ гибридизация с использованием меченых рибопроб GalNac-T14, Нозерн блот и родственные методики) и различные исследования амплификации нуклеиновых кислот (например, ОТ-ПЦР с использованием комплементарных праймеров, специфичных к GalNac-T14, и другие способы детекции амплификационного типа, например, такие как разветвленная ДНК, SISBA, TMA и подобное).
Образцы ткани или клеток млекопитающих можно удобно исследовать, например, на мРНК GalNac-T14 с использованием Нозерн, дот блот или ПЦР анализов. Например, ОТ-ПЦР анализ, такой как количественный ПЦР анализ, хорошо известен в уровне техники. В иллюстративном варианте осуществления этого изобретения способ детекции мРНК GalNac-T14 в биологическом образце включает в себя получение кДНК из образца посредством обратной транскрипции с использованием по меньшей мере одного праймера; амплификацию кДНК, полученную таким образом, используя полинуклеотид GalNac-T14 в качестве смыслового и антисмыслового праймеров для амплификации кДНК GalNac-T14; и выявление наличия амплифицированной кДНК GalNac-T14. Кроме того, такие способы могут включать в себя один или несколько стадий, которые дают возможность определить уровни мРНК GalNac-T14 в биологическом образце (например, путем одновременного исследования уровней сравнительной контрольной последовательности мРНК «облигатных» генов, например, представителя семейства актина). Необязательно, можно определить последовательность амплифицированной кДНК GalNac-T14.
Практические варианты осуществления этого аспекта изобретения включают GalNac-T14 праймеры и пары праймеров, которые дают возможность специфической амплификации полинуклеотидов по изобретению или любых их специфических частей, и зонды, которые селективно или специфически гибридизируются с молекулами нуклеиновой кислоты по изобретению или с любой их частью. Зонды могут быть мечены детектируемым маркером, например, таким как радиоизотоп, флуоресцентное соединение, биолюминесцентное соединение, хемилюминесцентное соединение, хелатор металла или фермент. Такие зонды и праймеры могут быть использованы для выявления наличия полинуклеотидов GalNac-T14 в образце, и как средства выявления клеток, экспрессирующих GalNac-T14 белки. Специалисту в данной области будет понятно, что очень большое число различных праймеров и зондов может быть получено на основе представленных здесь последовательностей и эффективно используемых для амплификации, клонирования и/или определения наличия и/или уровней мРНК GalNac-T14.
Необязательные способы по изобретению включают в себя протоколы для изучения или выявления мРНК, таких как мРНК GalNac-T14, в образце ткани или клеток с помощью технологий микрочипов. Используя микрочипы нуклеиновой кислоты, исследуемый и контрольный образец мРНК из тестируемых и контрольных образцов ткани обратно транскрибируют и метят для получения кДНК зондов. Зонды затем гибридизируют на матрицу нуклеиновых кислот, иммобилизованных на твердой подложке. Конфигурацию матрицы создают таким образом, что последовательность и положение каждого члена матрицы известны. Например, набор генов, которые могут экспрессироваться при определенных патологических состояниях, может быть выстроен в определенном порядке на твердой подложке. Гибридизация меченого зонда с конкретным членом матрицы указывает на то, что образец, из которого получен этот зонд, экспрессирует этот ген. Анализ дифференциальной генной экспрессии патологической ткани может дать ценную информацию. В технологии микрочипов используются методики гибридизации нуклеиновой кислоты и вычислительная техника для оценки профиля экспрессии мРНК тысяч генов за один эксперимент (см., например, WO 01/75166, опубликованную 11 октября 2001; (См., например, US 5700637, патент США 5445934 и патент США 5807522, Lockart, Nature Biotechnology, 14:1675-1680 (1996); Cheung, V.G. et al., Nature Genetics 21(Suppl):15-19 (1999) для рассмотрения изготовления матриц). Микрочипы ДНК представляют собой миниатюрные матрицы, содержащие фрагменты генов, которые либо синтезируют непосредственно или на стекле, или других субстратах, либо наносят пятнами. Тысячи генов обычно представлены в одной матрице. Стандартный эксперимент с использованием микрочипа включает в себя следующие стадии: 1) получение флуоресцентно меченой мишени из РНК, выделенной из образца, 2) гибридизация меченой мишени с микрочипом, 3) промывка, окрашивание и сканирование матрицы, 4) анализ сканированного изображения и 5) создание профилей экспрессии генов. В настоящее время используют два основных типа ДНК микрочипов: олигонуклеотидные (обычно от 25 до 70 мономерных звеньев) матрицы и матрицы генной экспрессии, содержащие продукты ПЦР, полученные из кДНК. В первой матрице олигонуклеотиды могут быть либо изготовлены заранее и нанесены на поверхность в виде пятен, либо синтезированы непосредственно на этой поверхности (in situ).
Система Affymetrix GeneChip® представляет собой коммерчески доступную систему микрочипов, которая содержит матрицы, изготовленные прямым синтезом олигонуклеотидов на стеклянной поверхности. Матрицы зонд/ген: олигонуклеотиды, обычно 25 мономерных звеньев, непосредственно синтезируют на стеклянной пластинке путем объединения технологий фотолитографии на полупроводниковой основе и твердофазного химического синтеза. Каждая матрица содержит до 400000 различных олигомеров, и каждый олигомер представлен в миллионах копий. Поскольку олигонуклеотидные зонды синтезируют в известных положениях на матрице, особенности гибридизации и интенсивности сигналов можно интерпретировать в понятиях генной идентичности и относительных уровней экспрессии с помощью программного обеспечения Affymetrix Microarray Suite. Каждый ген представлен на матрице серией различных олигонуклеотидных зондов. Каждая пара зондов состоит из комплементарного олигонуклеотида и некомплементарного олигонуклеотида. Комплементарный зонд имеет последовательность полностью комплементарную конкретному гену и, таким образом, определяет экспрессию гена. Некомплементарный зонд отличается от комплементарного зонда однонуклеотидной заменой в центральном положении основания, нарушая связывание целевого транскрипта гена. Это помогает определить фоновую и неспецифическую гибридизацию, которая вносит свой вклад в сигнал, определяемый в отношении комплементарного олигонуклеотида. В программном обеспечении Microarray Suite интенсивности гибридизации некомплементарных зондов вычитаются из интенсивностей гибридизации комплементарных зондов для определения величины абсолютной или специфичной интенсивности для каждого набора зондов. Зонды выбирают, основываясь на текущей информации из Genbank и других нуклеотидных баз данных. Считается, что последовательности распознают уникальные участки на 3' конце гена. Гибридизационную печь GeneChip (печь «гриль») используют для проведения гибридизации вплоть до 64 матриц за один раз. Струйная установка выполняет промывание и окрашивание матриц зондов. Она полностью автоматизирована и содержит четыре модуля, каждый модуль удерживает одну матрицу зондов. Каждый модуль контролируется независимо посредством программного обеспечения Microarray Suite с использованием заранее запрограммированных протоколов для струйных установок. Сканер представляет собой конфокальный лазерный флуоресцентный сканер, который измеряет интенсивность флуоресценции, испускаемой меченой кРНК, связанной с матрицами зондов. Компьютерная рабочая станция с программным обеспечением Microarray Suite контролирует струйную установку и сканер. Программное обеспечение Microarray Suite может контролировать до восьми струйных установок, использующих протоколы для заранее запрограммированной гибридизации, промывки и окрашивания для каждой матрицы зондов. Это программное обеспечение также получает информацию и преобразует данные по интенсивности гибридизации в сигнал присутствие/отсутствие для каждого гена, используя соответствующие алгоритмы. Наконец, это программное обеспечение детектирует изменения генной экспрессии между экспериментами путем сравнения анализа и форматирует выходные данные в .txt файлы, которые можно использовать с другими программами для дальнейшего анализа данных.
Флуоресцентную гибридизацию in-situ (FISH) также можно использовать для выявления экспрессии мРНК биомаркера, используя меченые зонды. Такие способы известны в уровне техники (см., например, Kallioniemi et al., 1992; патент США 6358682).
Экспрессию выбранного биомаркера также можно оценить, изучая делецию гена или амплификацию гена. Делецию гена или амплификацию можно определить любым из широкого разнообразия протоколов, известных в уровне техники, например стандартным Саузерн блоттингом, Нозерн блоттингом для количественного определения транскрипции мРНК (Thomas, Proc. Natl. Acad. Sci. USA, 77:5201-5205 (1980)), дот-блоттингом (ДНК анализ) или гибридизацией in situ (например, FISH), используя соответственно меченый зонд, цитогенетическими способами или сравнительной геномной гибридизации (CGH), используя соответственно меченый зонд. Для примера эти способы можно использовать для выявления делеции или амплификации генов GalNac-T14.
Кроме того, можно исследовать статус метилирования биомаркера, такого как ген GalNac-T14, в образце ткани или клетки. Аберрантное деметилирование и/или гиперметилирование CpG-островков в 5' регуляторных областях гена часто происходит в иммортализованных и трансформированных клетках, и в результате может приводить к измененной экспрессии различных генов. В уровне техники известен ряд анализов для исследования статуса метилирования гена. Например, можно использовать, в подходах Саузерн гибридизации, ферменты рестрикции, чувствительные к метилированию, которые не могут расщеплять последовательности, которые содержат метилированные CpG сайты для оценки статуса метилирования CpG-островков. Кроме того, с помощью MSP (ПЦР, специфичная к метилированию) можно быстро изобразить профиль статуса метилирования всех CpG сайтов, присутствующих в CpG-островке заданного гена. Эта методика включает в себя начальную модификацию ДНК под действием бисульфита натрия (которая будет превращать весь неметилированный цитозин в урацил) с последующей амплификацией с использованием праймеров, специфичных к метилированной ДНК, в отличие от неметилированной ДНК. Протоколы, включающие в себя метилирующую интерференцию, также можно найти, например, в Current Protocols In Molecular Biology, Unit 12, Frederick M. Ausubel et al. eds., 1995; De Marzo et al., Am. J. Pathol. 155(6): 1985-1992 (1999); Brooks et al, Cancer Epidemiol. Biomarkers Prev., 1998, 7:531-536); и Lethe et al., Int. J. Cancer 76(6): 903-908 (1998).
Экспрессию GalNac-T14 в образце ткани или клеток также можно исследовать с помощью анализов функции или активности. Например, можно провести исследования, известные в уровне техники, для определения или выявления наличия заданной ферментативной активности N-ацетилгалактозаминилтрансферазы в образце ткани или клеток (см., например, Bennett et al., J. Biol. Chem., 271:17006-17012 (1996); Wang et al., BBRC, 300:738-744 (2003); Hang et al., supra, доступные с мая 2005 на www.sciencedirect.com.
В способах по настоящему изобретению предполагается, что образец ткани или клетки также можно исследовать на экспрессию Apo2L/TRAIL или рецепторы в образце, которые связывают Apo2L/TRAIL или антитело к рецептору смерти. Как описано выше и в уровне техники, в настоящее время считается, что Apo2L/TRAIL связывает по меньшей мере пять различных рецепторов: DR4, DR5, DcR1, DcR2 и OPG. Используя способы, известные в уровне техники, в том числе описанные в этом изобретении, экспрессию Apo2L/TRAIL, DR4, DR5, DcR1, DcR2 и/или OPG можно определить по уровню мРНК и по уровню белка. Для примера, методики IHC, описанные выше, можно использовать для выявления присутствия одной или нескольких таких молекул в образце. Предполагается, что в способах, в которых ткань или образец исследуют не только на наличие маркера GalNac-T14, а также на наличие, например, DR4, DR5 или DcR1, отдельные гистологические препараты могут быть получены из одной и той же ткани или образца, и каждый гистологический препарат тестируют с реактивом, специфичным для каждого конкретного биомаркера или рецептора. Альтернативно, отдельный гистологический препарат может быть получен из образца ткани или клеток, а антитела, направленные на каждый биомаркер или рецептор, могут быть использованы применительно к многоцветному протоколу окрашивания для возможности визуализации и детекции соответствующих биомаркеров или рецепторов.
После определения, что в образце ткани или клеток экспрессируется GalNac-T14, указывая на то, что этот образец ткани или клеток будет чувствительным к Apo2L/TRAIL или антителу к рецептору смерти, предполагается, что эффективное количество Apo2L/TRAIL или антитела к рецептору смерти можно вводить млекопитающему для лечения такого нарушения, как злокачественное заболевание или заболевание, связанное с иммунной системой, которое поражает млекопитающее. Диагностику у млекопитающих различных патологических состояний, описанных здесь, может провести опытный практикующий врач. В уровне техники существуют способы диагностики, которые позволяют, например, осуществлять диагностику или выявление у млекопитающего злокачественного заболевания или заболевания, связанного с иммунной системой. Например, злокачественные заболевания можно идентифицировать посредством методик, включающих в себя, но не только, пальпацию, анализ крови, рентгенологическое исследование, ЯМР и подобные. Заболевания, связанные с иммунной системой, также могут быть легко идентифицированы.
Apo2L/TRAIL или антитело к рецептору смерти можно вводить в соответствии с известными способами, такими как внутривенное введение в виде болюса или путем непрерывной инфузии на протяжении периода времени, внутримышечным, внутрибрюшинным, интрацереброспинальным, подкожным путем, внутрисуставным, интрасиновиальным, в полость позвоночного канала, пероральным, местным или ингаляционным путем. Необязательно, введение можно осуществлять посредством инфузии мини-насосом, используя различные коммерчески доступные устройства.
Эффективные дозы и схемы введения Apo2L/TRAIL или антитела к рецептору смерти можно определить эмпирически, и такие определения находятся в компетенции специалиста в данной области. Можно использовать однократные или многократные дозы. В настоящее время считается, что эффективная доза или количество Apo2L/TRAIL, используемого отдельно, может варьировать примерно от 1 мкг/кг примерно до 100 мг/кг массы тела или более в день. Межвидовой перенос доз может быть выполнен способом, известным в уровне техники, например, как описано у Mordenti et al., Pharmaceut. Res., 8:1351 (1991).
При использовании in vivo введения Apo2L/TRAIL стандартная величина дозы может варьировать примерно от 10 нг/кг до 100 мг/кг массы тела млекопитающего или более в день, предпочтительно примерно от 1 мкг/кг/день до 10 мг/кг/день в зависимости от пути введения. Руководство по конкретным дозам и способам доставки представлено в литературе; см., например, патенты США №№ 4657760; 5206344; или 5225212. Ожидается, что различные композиции будут эффективны для различных лекарственных соединений и различных заболеваний, что для введения, нацеленного на один орган или ткань, например, может быть необходима доставка, отличная от таковой для другого органа или ткани.
Предполагается, что еще и дополнительные лечебные воздействия могут быть использованы в этих способах. Один или несколько других лечебных воздействий могут включать в себя, но не только, применение лучевой терапии, цитокина (цитокинов), ингибитора (ингибиторов) роста, химиотерапевтических средств(а), цитотоксических средств(а), ингибиторов тирозинкиназы, ингибиторов фарнезилтрансферазы ras, ингибиторов ангиогенеза и ингибиторов циклинзависимых киназ, которые известны в уровне техники и дополнительно характеризуемые указанной выше особенностью. Предполагается, что другие подобные лечебные воздействия могут быть использованы в качестве средства, отдельного от Apo2L/TRAIL или антитела к рецептору смерти. Кроме того, лечебные воздействия, основанные на терапевтических антителах, которые нацелены на опухолевые антигены, такие как РитуксанTM или ГерцептинTM, а также антиангиогенные антитела, такие как анти-VEGF.
Препарат и схемы введения химиотерапевтических средств могут быть использованы в соответствии с инструкциями производителя или получены эмпирически опытным практикующим врачом. Препарат и схемы введения для такой химиотерапии также описаны в Chemotherapy Service Ed., M.C. Perry, Williams & Wilkins, Baltimore, MD (1992). Химиотерапевтическое средство может предшествовать введению или следовать за введением Apo2L/TRAIL, или антитела к рецептору смерти, или его можно вводить одновременно с ними.
Может быть желательным также вводить антитела к другим антигенам, например антитела, которые связываются с CD20, CD11a, CD18, CD40, ErbB2, EGFR, ErbB3, ErbB4, сосудистым эндотелиальным фактором (VEGF), или другими членами семейства TNFR (такими как OPG, TNFR1, TNFR2, GITR, Apo-3, TACI, BCMA, BR3). Альтернативно, или дополнительно, два или более антител, связывающих один и тот же или два или более различных антигенов, описанных здесь, можно вводить пациенту совместно. Иногда может быть эффективным также введение пациенту одного или нескольких цитокинов. После введения можно анализировать обработанные клетки in vitro. При воздействии in vivo млекопитающее, которому проводили лечение, можно подвергнуть мониторингу различными способами, хорошо известными опытному практикующему врачу. Например, опухолевые клетки можно исследовать в патологических анализах для оценки некроза или можно исследовать сыворотку на ответы иммунной системы.
Для использования по описанным и предложенным выше назначениям изобретение также относится к наборам или изделиям. Такие наборы могут содержать средства доставки, компартментализированные для получения строгой изоляции одного или нескольких контейнеров, таких как флаконы, пробирки и подобное, каждый из контейнеров содержит один из отдельных элементов, используемых в этом способе. Например, один из контейнеров может содержать зонд, который является или может быть детектируемо мечен. Такой зонд может представлять антитело или полинуклеотид, специфичный в отношении белка GalNac-T14 или GalNac-T14 гена или единицы генетического кода, соответственно. В тех случаях, когда в наборе используется гибридизация нуклеиновой кислоты для выявления целевой нуклеиновой кислоты, в наборе также могут находиться контейнеры, содержащие нуклеотид(ы) для амплификации целевой последовательности нуклеиновой кислоты, в набор также могут входить контейнеры, содержащие нуклеотид(ы) для амплификации целевой последовательности нуклеиновой кислоты и/или контейнер, содержащий репортеры, биотин-связывающий белок, такой как авидин или стрептавидин, связанный с репортерной молекулой, такой как ферментная, флуоресцентная или радиоизотопная метка.
Набор по изобретению обычно будет содержать контейнер, описанный выше, и один или несколько других контейнеров, содержащих материалы, необходимые с коммерческой и пользовательской точек зрения, включая буферы, разбавители, фильтры, иглы, шприцы и вкладыши с инструкциями по применению. Этикетка может находиться в контейнере для указания того, что композицию используют для специфической терапии или нетерапевтического применения, и также может указывать способ для применения либо in vivo, либо in vitro, как описано выше.
Наборы по изобретению имеют ряд вариантов осуществления. Стандартным вариантом осуществления является набор, содержащий контейнер, этикетку на указанном контейнере и композицию, заключенную в указанном контейнере; где композиция включает в себя первичное антитело, которое связывается с полипептидной последовательностью GalNac-T14, этикетка на указанном контейнере указывает на то, что композицию можно использовать для оценки наличия белков GalNac-T14 по меньшей мере в одном типе клеток млекопитающих и инструкции по применению антитела к GalNac-T14 для оценки наличия белков по меньшей мере в одном типе клеток млекопитающих. Дополнительно набор может содержать комплект инструкций и материалов для получения образца ткани и нанесения антитела и зонда на вышеупомянутый срез образца ткани. Набор может включать в себя как первичное, так и вторичное антитело, где вторичное антитело конъюгировано с меткой, например ферментной меткой.
Другой вариант осуществления представляет собой набор, содержащий контейнер, этикетку на указанном контейнере и композицию, заключенную в этом контейнере; где композиция включает в себя полинуклеотид, который гибридизируется с комплементом полинуклеотида GalNac-T14 в строгих условиях, этикетка на этом контейнере указывает, что композицию можно использовать для оценки наличия GalNac-T14 по меньшей мере в одном типе клеток млекопитающих и инструкции для использования полинуклеотида GalNac-T14 для оценки наличия РНК или ДНК GalNac-T14 по меньшей мере в одном типе клеток млекопитающих.
Другие необязательные компоненты в наборе включают в себя один или несколько буферов (например, блокировочный буфер, промывочный буфер, субстратный буфер и т.д.), другие реактивы, такие как субстрат (например, хромоген), который химически изменяется ферментной меткой, раствор для восстановления эпитопов, контрольные образцы (положительный и/или отрицательный контроли), контрольный гистологический препарат(ы) и т.д.
ПРИМЕРЫ
Различные аспекты изобретения дополнительно описаны и проиллюстрированы следующими примерами, ни один из которых не предназначен для ограничения объема изобретения.
СПОСОБЫ И МАТЕРИАЛЫ:
Клеточная культура и клеточные линии
Следующие линии клеток человека: линии немелкоклеточного рака легких (NSCLC): H2122, A427, H647, SK-MES-1, H838, H358, H2126, H460, H1703, H2405, H650, H1568, H1666, H322T, SW1573, H292, H1650, H522, EKVX, H661, H23, LXFL 529, H226, A549, H1781, H1299, HOP 62, H2009, HOP 92, H1793, H1975, H1651, calu-1, H1435, HOP 18, H520, H441, H2030, H1155, H1838, H596, HLFa; линии рака поджелудочной железы: Panc 05.04, BxPC3, HPAC, SU.86.86, HuP-T3, PSN1, Panc 08.13, MiaPaCa-2, PA-TU-8988T, Panc 03.27, Capan-1, SW 1990, CFPAC-1, PA-TU-8902, Panc 02.03, Panc 04.03, PL45, Aspc-1, Hs766T, Panc 10.05, Panc1, Capan-2, HPAF-II и NHL: JEKO-1, SU-DHL-4, OCI-LY-19, SR, Farage, DOHH-2, Toledo, WSU-NHL, KARPAS-422, GRANTA-519, Pfeiffer, HT, SC-1, DB. Эти линии были получены из депозитария ATCC (Manassas, Virginia), DSMZ (Немецкая коллекция микроорганизмов и клеточных культур), JCRB (Японский банк клеточных ресурсов) или ECACC (Европейская коллекция клеточных культур), и их культивировали в среде RPMI-1640, дополненной 10% инактивированной нагреванием эмбриональной сывороткой теленка, 2 мM L-глутамина и 10 мM HEPES.
Исследования цитотоксичности
MTT анализ (CellTiter96® нерадиоактивный анализ клеточной пролиферации от Promega), который представляет собой колориметрический анализ, основанный на способности жизнеспособных клеток восстанавливать растворимую соль тетразолия желтого (MTT) в голубые кристаллы формазана), использовали для определения количества жизнеспособных клеток после обработки Apo2L/TRAIL или антителом к DR5. MTT анализ проводили путем добавления предварительно смешанного оптимизированного раствора красителя в культуральные лунки 96-луночного планшета, содержащего различные концентрации (от 0 до 1000 нг/мл) Apo2L/TRAIL или антитела к DR5. Во время 4-часовой инкубации живые клетки превращают тетразолиевый компонент раствора красителя в формазановый продукт. Затем к культуральным лункам добавляли солюбилизация/стоп раствор для растворения формазанового продукта и регистрировали поглощение при 570 нм, используя 96-луночный планшетный ридер (SpectraMax). Показание поглощения при 570 нм прямопропорционально количеству клеток, обычно используемых в анализах пролиферации. Хотя максимум поглощения для формазанового продукта составляет 570 нм, и чистый растор выглядит голубым, цвет в конце исследования может не быть голубым и зависит от количества присутствующего формазана относительно других компонентов (включая сыворотку, окисленный феноловый красный и невосстановленный MTT) в культуральной среде.
Количество клеток было оптимизировано проведением тирования клеток для получения аналитического сигнала, близкого к верхней границе линейной области этого исследования. Поскольку различные типы клеток имеют различные уровни метаболической активности, для каждой клеточной линии это делали отдельно. Для большинства исследуемых опухолевых клеток использовали от 5000 клеток на лунку до 20000 клеток на лунку.
Следующее представляет постадийное описание используемых исследований:
1. Клетки, используемые для биологического анализа, были из чистых культур.
2. Определение количества клеток и жизнеспособности по трепановому синему и суспендирование клеток до конечного количества от 5000 до 20000 клеток на лунку.
3. Распределение по 50 мкл клеточной суспензии в 96-луночный планшет.
4. Инкубирование планшетов при 37°C в увлажненной атмосфере 5% CO2 в течение ночи.
5. Добавление 50 мкл культуральной среды, содержащей различные концентрации в диапазоне от 0 до 1000 мг/мл Apo2L/TRAIL или антитела к DR5 к образцам в 96-луночном планшете. Контролями были 50 мкл культуральной среды (без Apo2L/TRAIL или антитела к DR5) и 100 мкл культуральной среды (без клеток).
Каждый эксперимент проводили в тройной серии лунок и в три отдельных дня. Общий объем лунок составил 100 мкл/лунку.
6. Инкубирование планшетов при 37°C в течение 72 часов в увлажненной атмосфере 5% CO2.
7. Добавление 15 мкл раствора красителя в каждую лунку.
8. Инкубирование планшетов при 37°C в течение периода времени до 4 часов в увлажненной атмосфере 5% CO2.
9. Добавление 100 мкл солюбилизирующего/стоп раствора в каждую лунку.
10. Инкубирование планшетов в течение ночи при 37°C.
11. Регистрация поглощения при длине волны 570 нм, используя 96-луночный планшетный ридер. Эталонную длину волны 750 нм использовали для уменьшения фона, который вносил клеточный дебрис, отпечатки пальцев и другое неспецифическое поглощение.
12. Средние значения поглощения для отрицательного контроля использовали в качестве пустого значения и вычитали из всех других показаний. Средние значения поглощения для каждой концентрации Apo2L/TRAIL или антитела к DR5 делили на средние значения поглощения положительного контроля (100% жизнеспособные клетки - необработанные) для вычисления количества жизнеспособных клеток (в %).
13. Процент жизнеспособных клеток (ось Y) в зависимости от концентрации Apo2L/TRAIL или антитела к DR5 (ось X, логарифмическая шкала) отмечали на графике, и значения IC50 определяли расположением значения по оси Х (нг/мл), соответствующего 50% жизнеспособных клеток.
Протокол введения метки от Affymetrix
Для всех образцов брали показатели OD260/280, и образцы прогоняли через BioAnalyzer. Использовали 5 мкг общей РНК высокого качества.
A. Синтез первой цепи кДНК:
1. Гибридизация праймера
DEPC-H2O x мкл Перемешивание встряхиванием. Быстро осадить центрифугированием.
РНК (5 мкг) y мкл Инкубирование при 70°C в течение 10 минут.
Контрольный раствор (1:4 разведение исходного раствора для 5 мкг) 1 мкл Высокие обороты и помещение на лед
T7-(dT)24 праймер 1 мкл
Объем 12 мкл
2. Установка температуры
5X-буфер для 1-й цепи кДНК 4 мкл
Добавление 7 мкл (к смеси, указанной слева) к каждому образцу.
0,1 M DTT 2 мкл Перемешивание встряхиванием. Быстрое вращение.
10 мM смесь dNTP 1 мкл Инкубирование при 42°C в течение 2 минут.
объем 7 мкл
3. Синтез первой цепи
Добавление 1 мкл SSII RT к каждому образцу.
SSII RT 1 мкл Перемешивание пипетированием вверх и вниз -ИЛИ- легкое перемешивание.
Быстрое вращение.
Общий объем 20 мкл Инкубирование при 42°C в течение 1 часа.
B. Синтез второй цепи кДНК
1. Поместить реакционные смеси для первой цепи на лед. Короткое центрифугирование, чтобы снизить конденсацию по бокам пробирки.
2. Получение следующей рабочей смеси для второй цепи.
DEPC-обработанная H2O 91 мкл
5X Реакционный буфер для второй цепи 30 мкл
10 мM смеси dNTP 3 мкл
10 Ед/мкл ДНК лигазы 1 мкл
10 Ед/мкл ДНК полимеразы I 4 мкл
2 Ед/мкл РНКазы H 1 мкл
Общий объем 130 мкл
3. Добавить 130 мкл рабочей смеси для второй цепи к 20 мкл первой цепи кДНК. (Конечный объем = 150 мкл)
4. Перемешать пипетированием вверх и вниз -ИЛИ- легким встряхиванием. Быстрое вращение.
5. Инкубирование при 16°C в течение 2 часов на охлаждающей водяной бане.
6. Добавить 2 мкл [10 Ед] T4 ДНК полимеразы. Перемешать пипетированием вверх и вниз -ИЛИ- легким встряхиванием. Быстрое вращение.
7. Инкубировать в течение 5 минут при 16°C.
8. Добавить 10 мкл 0,5 M EDTA. Слабое встряхивание. Быстрое вращение.
9. Перейти к процедуре очистки кДНК -ИЛИ- хранить при -20°C для последующего использования.
Очистка двухспиральной кДНК (Модуль для очистки образцов GeneChip)
1. Добавить 600 мкл кДНК связывающего буфера к 162 мкл конечному препарату синтеза двухспиральной кДНК.
Перемешивание встряхиванием в течение 3 секунд.
2. Проверить, что цвет смеси желтый (сходный с кДНК связывающим буфером w/o синтез кДНК rxn.)
Если цвет смеси оранжевый или фиолетовый, добавить 10 мкл 3 М ацетата натрия, pH 5,0 и перемешать.
Цвет смеси перейдет в желтый.
3. Нанести 500 мкл образца на колонку для хроматографии центрифугированием для очистки кДНК, находящуюся в 2 мл пробирке для сбора, и центрифугировать в течение 1 минуты при ≥8000 x g (≥10000 об/мин). Утилизировать проскок как *вредные отходы.
4. Перезагрузить спиновую колонку оставшейся смесью (262 мкл) и центрифугировать, как указано выше.
Утилизировать проскок и пробирку для сбора как *вредные отходы.
5. Перенести спиновую колонку в новую 2-мл коллекторную трубку (входит в комплект). Перенести пипеткой 750 мкл промывочного буфера кДНК на спиновую колонку. Центрифугировать в течение 1 минуты при ≥8000 x g (≥10000 об/мин).
Отбросить непрерывный поток.
6. Открыть крышку спиновой колонки и центрифугировать в течение 5 минут с максимальной скоростью (≤ 25000 x g). Расположить колонки в центрифуге, используя каждое второе гнездо ротора, расположить крышки пробирок над соседними гнездами ротора так, чтобы они были ориентированы в направлении, противоположном вращению, т.е. если вращение по часовой стрелке, крышки ориентируют в направлении против часовой стрелки. Это позволяет избежать повреждения крышек.
Отбросить непрерывный поток и снять коллекторную трубку.
7. Перенести спиновую колонку в 1,5-мл коллекторную трубку. Перенести пипеткой 10 мкл буфера для элюирования кДНК непосредственно на мембрану спиновой колонки. Убедиться, что буфер для элюирования кДНК распределен непосредственно на мембране.
Инкубировать в течение 1 минуты при комнатной температуре и центрифугировать 1 минуту с максимальной скоростью (≤ 25000 x g) для элюции.
Подготовка и проведение реакции IVT
Enzo: Набор для мечения РНК транскрипта Bioarray HighYield (Партия № 900182)
1. Использовать 10 мкл очищенной двухспиральной кДНК.
2. Приготовить следующую рабочую смесь для IVT:
Дистиллированная или деионизированная H2O 12 мкл
10X HY реакционного буфера 4 мкл
10x рибонуклеотидов, меченных биотином 4 мкл
10X DTT 4 мкл
10X смесь, ингибирующая РНазы 4 мкл
20X T7 РНК полимеразы 2 мкл
Общий объем: 30 мкл
3. Добавить 30 мкл рабочей смеси IVT к 10 мкл двухспиральной кДНК (Общий объем = 40 мкл).
4. Перемешать пипетированием вверх и вниз -ИЛИ- легким встряхиванием. Быстрое вращение.
5. Сразу же поместить пробирку на водяную баню 37°C. Инкубировать в течение 5 часов.
6. Хранить при -20°C, если не производить немедленную очистку РНК.
Очистка кРНК, меченной биотином (GeneChip Sample Cleanup Module).
1. Добавить 60 мкл H2O к реакции IVT и перемешать встряхиванием в течение 3 секунд.
2. Добавить 350 мкл связывающего буфера IVT кРНК к образцу, перемешать встряхиванием в течение 3 секунд.
3. Добавить 250 мкл этанола (96-100%) к этому лизату и тщательно перемешать пипетированием. Не центрифугировать.
4. Нанести образец (700 мкл) на очищающую спиновую колонку для IVT кРНК, установленную в 2 мл коллекторной трубке.
Центрифугировать 15 секунд при ≥8000 x g (≥10000 об/мин).
5. Пропустить элюат через колонку еще раз.
Центрифугировать в течение 15 секунд при ≥8000 x g (≥10000 об/мин).
Отбросить непрерывный поток как **вредные отходы и снять коллекторную трубку.
6. Перенести спиновую колонку в новую 2-мл коллекторную трубку (входит в комплект).
7. Добавить 500 мкл промывочного буфера для IVT кРНК и центрифугировать в течение 15 секунд при ≥8000 x g (≥10000 об/мин) для промывания.
Отбросить непрерывный поток.
8. Нанести пипеткой 500 мкл 80% (об./об.) этанола на спиновую колонку и центрифугировать в течение 15 секунд при ≥8000 x g (≥10000 об/мин). Отбросить непрерывный поток.
9. Открыть крышку спиновой колонки и центрифугировать в течение 5 минут с максимальной скоростью (≤25000 x g).
Отбросить непрерывный поток и снять коллекторную трубку.
10. Перенести спиновую колонку в новую 1,5-мл коллекторную трубку.
11. Нанести пипеткой 11 мкл воды без РНКазы непосредственно на мембрану спиновой колонки. Оставить на 1 минуту.
Центрифугировать в течение 1 минуты с максимальной скоростью (≤25000 x g) для элюирования.
12. Нанести пипеткой 10 мкл воды без РНКазы непосредственно на мембрану спиновой колонки. Оставить на 1 минуту.
Центрифугировать в течение 1 минуты с максимальной скоростью (≤25000 x g) для элюирования.
Определение количества кРНК (продукт IVT).
Для определения выхода РНК используют спектрофотометрический анализ. Принять условие, что 1 OD при 260 нм равняется 40 мкг/мл РНК.
Проверить OD при 260 нм и 280 нм для определения концентрации образца и чистоты.
Поддерживать соотношение A260/A280, близкое к 2,0, для чистой РНК (приемлемы соотношения между от 1,9 до 2,1).
Для количественного определения кРНК при использовании общей РНК в качестве исходного материала скорректированный выход кРНК следует вычислить для выражения остатка немеченой общей РНК. С использованием оценки 100% переноса использовать приведенную ниже формулу для определения скорректированного выхода кРНК:
Скорректированный выход кРНК = РНКm - (общая РНКi)(y)
РНКm = количество кРНК, измеренное после IVT (мкг)
общая РНКi = исходное количество общей РНК (мкг)
y = фракция кДНК реакции, используемой в IVT
Фрагментация кРНК для получения мишени
Для фрагментации использовать скорректированную концентрацию кРНК.
1. Добавить 2 мкл 5x буфера для фрагментации для каждых 8 мкл РНК плюс H2O.
20 мкг кРНК 1 к 32 мкл
5X буфера для фрагментации 8 мкл
Вода без РНазы до 40 мкл
Общий объем: 40 мкл
2. Инкубировать при 94°C в течение 30 минут. После инкубации сразу же поместить на лед.
Получение мишени для гибридизации
1. Нагревать 20X гибридизационные контроли для эукариот и олиго B2 в течение 5 минут при 65°C.
Набор для гибридизационного контроля эукариот Affymetrix GeneChip, Партия #900362 (для 150 реакций)
2. Легкое встряхивание, замедленное вращение.
3. Рабочая смесь (предполагая, что концентрация фрагментированной кРНК составляет 0,5 мкг/мкл):
Стандартная матрица (мкл) Конечная концентрация
Фрагментированная кРНК 15 мкг 30 0,05 мкг/мкл
Олиго B2 (3 нM) 5 50 пM
20x контрольный Spike 15 1,5, 5, 25, 100 пM
(Bio B, C, D, Cre)
ДНК спермы сельди 3 0,1 мг/мл
Ацетилированный BSA 3 0,5 мг/мл
Hu cot-1 ДНК (1 мг/мл) 30 0,1 мг/мл
2X MES гиб. буфер 150 1X
H2O 64
Конечный объем 300
4. Разлить аликвоту рабочей смеси 270 мкл в пробирки и в каждую добавить 30 мкл фрагментированной кРНК. Это - гибридизационная смесь.
5. Непосредственно перед использованием матрицы зондов привести в равновесие с комнатной температурой.
6. Наполнить матрицу зондов 1x MES гибридизационным буфером и инкубировать в печи-гриль в течение 10 минут при 45°C, 60 об/мин.
7. Нагреть гибридизационную смесь на водяной бане 99°C в течение 5 минут.
8. Перенести гибридизационную смесь на водяную баню 45°C на 5 минут.
9. Центрифугировать гибридизационную смесь в течение 5 минут с максимальной скоростью.
10. Удалить 1x MES гибридизационного буфера из матрицы зондов.
11. Наполнить матрицу зондов до краев 200 мкл гибридизационной смеси.
12. Герметизировать ячейки с помощью Tough-Spots.
13. Гибридизировать матрицу зондов при 45°C, 60 RPM в течение 19 часов.
14. Промыть, окрасить и сканировать матрицу зондов в соответствии с протоколами Affymetrix.
Материалы Affymetrix
Наименование Фирма-поставщик № каталога
T7-(dT)24 праймер Biosearch Technolgies на заказ
Контрольные spikes собственного производства -
Superscript II/5X Буфер
для синтеза первой цепи/0,1 M DTT Invitrogen 18064-014
5X буфер для синтеза второй цепи Invitrogen 10812-014
10 мM dNTP Invitrogen 18427-088
10 Ед/мкл ДНК лигазы E. coli Invitrogen 18052-019
10 Ед/мкл ДНК полимеразы I E. coli Invitrogen 18010-025
2 Ед/мкл РНазы H Invitrogen 18021-071
10 Ед/мкл ДНК полимеразы T4 Invitrogen 18005-025
0,5 M EDTA Sigma E-7889
ENZO High Yield RNA Transcript
labeling kit Affymetrix или ENZO 900182 (ENZO)
GeneChip Sample Cleanup Module Affymetrix 900371
Ацетилированный бычий
сывороточный альбумин Invitrogen 15561-020
Козьи IgG - химически чистые Sigma I-5256
Антитела к стрептавидину (козьи),
биотинилированные Vector Labs BA-0500
R-рикоэритрин стрептавидин Molecular Probes S-866
20X SSPE BioWhittaker 51214
Eukaryotic Control Kit Affymetrix 900362
Вода, Molecular Biology Grade Ambion 9934
ДНК человека Cot-1 Roche 1-581-074
5 M NaCl без РНазы, без ДНазы Ambion 9760
Противовспенивающее вещество 0-30 Sigma A-8082
10% Твин-20 Pierce Chemical 28320
Моногидрат свободной кислоты MES Sigma M5287
Натриевая соль MES Sigma M3885
Динатриевая соль EDTA, 0,5 M раствор Sigma E7889
Tough Spots, Label Dots USA Scientific 9902
GeneChip гибридизационная печь 640 Affymetrix 800139
GeneChip сканер 3000 w/рабочая станция Affymetrix 00-0074
Струйная установка Affymetrix 00-0081
Autoloader w/External Barcode Reader Affymetrix 00-0129
Количественная ПЦР
Синтез кДНК:
Условия инкубации:
25° в течение 10 минут
37° в течение 2 часов
TaqMan реакция с использованием ABI Prism 7700 Sequencing Detector:
(Assays-on-Demand™)
Термические условия циклов:
95° по 10 минут
40 циклов: 95° по 15 секунд
60° по 1 минуте
TaqMan зонды: Assays-on-Demand™ (TaqMan® MGB зонды, FAM™ меченные красителем)
Амплификацию внутреннего контроля, GAPDH (концентрация зонда 100 нM, концентрации прямого и обратного праймеров 200 нM) проводили для стандартизации количества РНК образца (кДНК), добавляемого в каждой реакции.
Сравнительный количественный анализ проводили с использованием калибровочной кривой. Для количественного анализа, нормированного к внутреннему контролю, калибровочные кривые строили как для мишени, так и для внутреннего стандарта. Для каждого экспериментального образца количество мишени и внутреннего стандарта определяют по соответствующей калибровочной кривой. Затем количество мишени делят на количество внутреннего контроля для получения нормализованной искомой величины. Один из экспериментальных образцов служит калибратором или 1x образцом. Каждую из нормализованных искомых величин затем делили на нормализованную искомую величину для калибратора для получения относительных уровней экспрессии.
Экспериментальные результаты.
Эксперименты проводили с использованием способов и материалов, описанных выше. Результаты этих экспериментов показаны на Фиг. 5-9, рассматриваемых ниже.
На Фиг.5 представлена сводная таблица данных по IC50, полученных в анализе чувствительности или резистентности клеточных линий немелкоклеточного рака легких («NSCLC») к апоптотической активности Apo2L (+ 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS) или моноклонального антитела DR5 «mab», поперечно-связанного «XL» или поперечно не связанного, + 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS), как определено в MTT анализах цитотоксичности.
На Фиг.6 представлена сводная таблица данных по IC50, полученных в анализе чувствительности или резистентности линий злокачественных клеток поджелудочной железы к апоптотической активности Apo2L (+ 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS) или моноклонального антитела DR5 «mab», поперечно-связанного «XL» или поперечно не связанного, + 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS), как определено в MTT анализах цитотоксичности.
На Фиг.7 представлена сводная таблица данных по IC50, полученных в анализе чувствительности или резистентности линий злокачественных клеток неходжкинской лимфомы («NHL») к апоптотической активности Apo2L (+ 10% эмбриональная сыворотка теленка «FBS») или моноклонального антитела DR5 «mab», поперечно-связанного «XL» или поперечно не связанного, + 0,5% эмбриональная сыворотка теленка «FBS» или 10% FBS), как определено в MTT анализах цитотоксичности.
На Фиг.8 приведено сравнение чувствительности («sen») или резистентности («RES») выбранных линий злокачественных клеток NSCLC, поджелудочной железы и NHL к антителу к DR5 и корреляция с экспрессией GalNac-T14, измеренной посредством экспрессии мРНК GalNac-T14.
На Фиг.9 представлена гистограмма различных клеточных линий NSCLC, поджелудочной железы и NHL cell, расположенных (в порядке убывания) по уровням экспрессии мРНК.
*********************************************************
Программа апоптотической гибели клетки играет важную роль в развитии и гомеостазе многоклеточных организмов (Danial et al., Cell, 116:205 (2004)). Внутриклеточные стимулы могут запускать апоптоз через эндогенный клеточный путь, который обусловлен представителями суперсемейства генов Bcl-2 для активации апоптотического каспазного механизма (Cory et al., Nat. Rev. Cancer, 2:647 (2002)). Определенные цитокины, которые принадлежат суперсемейству фактора некроза опухоли (TNF), могут активировать апоптоз через внешний клеточный путь путем взаимодействия с некоторыми рецепторами, которые содержат функциональный индуцирующий апоптоз «домен смерти» (DD) (Ashkenazi et al., Science, 281:1305 (1998)). Fas лиганд (FasL) стимулирует апоптоз через Fas (Apo1/CD95), тогда как Apo-2 лиганд/TNF-связанный апоптоз-индуцирующий лиганд (Apo2L/TRAIL) запускает апоптоз через DR4 (TRAIL-R1) и/или DR5 (TRAIL-R2) (LeBlanc et al., Cell Death Differ., 10:66 (2003)). При связывании родственного лиганда эти рецепторы связывают адаптерную молекулу FADD (связанный с Fas домен смерти), которая привлекает инициатор апоптоза каспазу-8 к образованию индуцирующего гибель сигнального комплекса (DISC) (см., например, Kischkel et al., EMBO J., 14:5579 (1995); Kischkel et al., Immunity, 12:611 (2000)). DISC-ассоциация стимулирует каспазу-8, которая, в свою очередь, активирует эффекторыне протеазы, такие как каспаза-3, 6 и 7, для выполнения апоптотической программы смерти. Во многих клеточных типах взаимное влияние на внутренний клеточный путь может дополнительно амплифицировать внешний клеточный сигнал смерти (Scaffidi et al., J. Biol. Chem., 274:1541 (1999)). Apo2L/TRAIL индуцирует апоптоз в разнообразных типах опухолевых клеток с небольшим воздействием или без воздействия на нормальные ткани, указывая на то, что Apo2L/TRAIL можно применять для лечения злокачественных заболеваний (см., например, Ashkenazi, Nat. Rev. Cancer, 2:420 (2002); Kelley et al., Curr. Opin. Pharmacol., 4:333 (2004)). Изменения в различных компонентах путей апоптоза могут снизить чувствительность к Apo2L/TRAIL в определенных линиях злокачественных клеток (Igney et al., Nat. Rev. Cancer, 2:277 (2002)).
Проводили различные эксперименты в соответствии со способами и протоколами, изложенными ниже, и данные представлены на Фиг.10-15.
Для изучения чувствительности к активации рецептора тестировали выживаемость клетки в зависимости от концентрации Apo2L/TRAIL в группе линий злокачественных клеток, в том числе 23 аденокарциномы поджелудочной железы, 18 злокачественных меланом и 36 колоректальных аденокарцином (Фиг.10A и данные, которые не показаны). В результате этого анализа 29/77 (38%) клеточных линий были классифицированы как высоко или умеренно чувствительные к Apo2L/TRAIL. Концентрация Apo2L/TRAIL, необходимая для достижения 50% клеточной гибели в этих 29 клеточных типах, находилась в диапазоне от 3 до 800 нг/мл со средним значением 250 нг/мл.
Группу клеточных линий также исследовали на профиль генной экспрессии, используя микрочип с наборами 54613 геннных зондов. Хоть и с некоторыми исключениями, клеточные линии рака поджелудочной железы и меланомы, которые демонстрировали сильную или среднюю чувствительность к Apo2L/TRAIL, экспрессировали значительно более высокие уровни мРНК фермента O-гликозилирования ppGalNAcT-14, чем соответствующие резистентные клеточные линии (p=0,5×10-4 по точному критерию Фишера, предельная величина определена многократно при 750 для карциномы поджелудочной железы и 300 для меланомы) (Фиг.10B). Большинство Apo2L/TRAIL-чувствительных клеточных линий колоректального рака демонстрировало высокую экспрессию мРНК родственного O-гликозилированного фермента ppGalNAcT-3, хотя несколько резистентных клеточных линий также экспрессировали этот ген, приводя, в результате, к слабому, все же значимому отличию (p=0,026, пороговая величина определена при 2000) (Фиг.10C, внизу). Исключениями из всей группы являлись: (a) 5/29 (17%) клеточных линий, которые были чувствительными, все же экспрессировали ppGalNAcT-14 или ppGalNAcT-3 ниже пороговых уровней; (b) 16/48 (33%) резистентных клеточных линий, которые, тем не менее, имели уровни ppGalNAcT-14 или ppGalNAcT-3 выше пороговых значений. Исследуя экспрессию мРНК других ферментов О-гликозилирования в колоректальных клеточных линиях, более высокие уровни Fut-6 были выявлены в 10/12 (83%) чувствительных по сравнению с 6/24 (25%) резистентных клеточных линий (p=0,013; пороговая величина определена при 200) (Фиг.10C, вверху). Комбинированная экспрессия ppGalNAcT-14 при раке поджелудочной железы и меланоме с Fut-6 в клеточных линиях колоректального рака очень сильно коррелировала с чувствительностью к Apo2L/TRAIL (p = 1,83×10-7, N=77). Этот генный набор верно прогнозировал чувствительность или резистентность для 23/32 (72%) маркер-позитивных и 39/45 (87%) маркер-негативных клеточных линий соответственно.
Apo2L/TRAIL чувствительность также исследовали in vivo, используя опухолевые ксенотрансплантаты. 5-дневная обработка Apo2L/TRAIL мышей, с опухолями, происходящими из Fut-6-позитивных клеточных линий колоректального рака Colo205 и DLD-1 вызывала регрессию опухоли вслед за значительной задержкой опухолевой прогрессии (Фиг.10D). Напротив, опухоли, происходящие из Fut-6-негативных клеточных линий колоректального рака Colo320 и RKO, не отвечали на эту обработку.
Предварительное инкубирование ppGalNAcT-3/Fut-6-позитивных клеточных линий Colo205 с ингибитором pan O-гликозилтрансферазы бензил-GalNAc (Delannoy et al., Glycoconj., 13:717 (1996)) заметно снижало чувствительность к Apo2L/TRAIL (Фиг.11A), указывая на функциональную связь между О-гликозилированием и сигнальным путем Apo2L/TRAIL. Для дополнительного исследования специфические малые интерферирующие (si)РНК олигонуклеотиды использовали таким образом, чтобы они были нацелены на мРНК ppGalNacT-14, ppGalNacT-3 или Fut-6. Для исключения нецелевых эффектов синтезировали многочисленные неперекрывающие siРНК для каждого гена и проверяли их способность уменьшать целевую экспрессию с помощью количественной ОТ-ПЦР (Фиг.14A). Специфичность siРНК дополнительно подтверждали с помощью мутантной плазмиды ppGalNacT-14, содержащей 6 «молчащих» нуклеотидных изменений в пределах целевого участка для siРНК (Editorial, Nat. Cell. Biol., 5:489 (2003)) (Фиг.14B). Трансфекция клеточных линий ppGalNAcT-14-позитивной PSN-1 карциномы поджелудочной железы и меланомы Hs294T с использованием ppGalNAcT-14 siРНК существенно снижала чувствительность к Apo2L/TRAIL, тогда как siРНК каспазы-8, как ожидалось, обеспечивала, по существу, полную защиту (Фиг.11B). Подобным образом, трансфекция DLD-1 или C170 клеток колоректального рака с использованием GalNAcT-3 или Fut-6 siРНК существенно снижала чувствительность к Apo2L/TRAIL (Фиг.11C и Фиг.14C). В сумме, GalNAcT-14 siРНК снижала чувствительность к Apo2L/TRAIL в 4/5 клеточных линиях рака поджелудочной железы и 2/2 клеточных линиях меланомы, тогда как ppGalNAcT-3 или Fut-6 siРНК - каждая уменьшала чувствительность в 2/3 клеточных линиях колоректального рака. В отличие от этого, трансфекция PSN-1 или Hs294T клеток с использованием GalNAcT-14 siРНК не изменяла чувствительности к ингибитору топоизомеразы II этопозиду (Фиг.14D). Подобным образом, трансфекция PSN-1 или C170 клетки с использованием GalNAcT-14 или GalNAcT-3 siРНК не влияла на чувствительность к ингибитору протеинкиназ широкого спектра действия стауроспорину (Фиг.14E). Поскольку и этопозид, и стауроспорин стимулируют апоптоз посредством внутреннего клеточного пути (Wei et al., Science, 292:727 (2001), эти исследования указывают на то, что ферменты O-гликозилирования могут модулировать передачу апоптотического сигнала через внешний клеточный путь.
Трансфекция клеток HEK293 с использованием ppGalNAcT-14 обнаруживала клеточную смерть при совместной трансфекции с DR4 или DR5, но не родственными рецепторами Fas и TNFR1 или агонистом Bax внутреннего клеточного пути (Фиг.11D). Более того, ppGalNAcT-14 трансфекция повышала чувствительность к Apo2L/TRAIL резистентных клеточных линий H1568 меланомы (Фиг.11E) и PA-TU-8902 и PL-45 карциномы поджелудочной железы (Фиг.14F), но не изменяла чувствительности к этопозиду (данные не показаны). В итоге, гиперэкспрессия GalNAcT-14 сенсибилизировала 4/7 клеточных линий к действию Apo2L/TRAIL.
Эффект siРНК нокдауна ppGalNacT-14 или Fut-6 исследовали на Apo2L/TRAIL-индуцированном процессинге каспазы. В клетках PSN-1 и DLD-1 клетки трансфектировали контрольной siРНК, Apo2L/TRAIL индуцировал, по существу, полный процессинг каспазы-8, ведущий к расщеплению Bid, каспазы-9 и каспазы-3 (Фиг.12A). Трансфекция с использованием siРНК каспазы-8 препятствовала этим событиям. Нокдаун ppGalNAcT-14 в клетках PSN-1 или Fut-6 в клетках DLD-1 также заметно ослаблял Apo2L/TRAIL-индуцированный процессинг каспазы-8, Bid, каспазы-9 и каспазы-3 (Фиг.12A) и стимуляцию активности каспазы-3/7 (Фиг.12B). Apo2L/TRAIL-резистентные клеточные линии колоректального рака RKO и SW1417, которые экспрессируют низкие уровни ppGalNAcT-3 и Fut-6, демонстрировали аналогичную блокаду на уровне процессинга каспазы-8 (Фиг.15A). Таким образом, ферменты O-гликозилирования могут модулировать события, предшествующие Apo2L/TRAIL каскаду, которые ведут к активации каспазы-8.
Для активации каспазы-8 требуется сборка DISC (Ashkenazi et al., Science, 281:1305 (1998)). Анализ Apo2L/TRAIL DISC (Kischkel et al., Immunity, 12:611 (2000)) в клетках PSN-1 и DLD-1 указывал на то, что нокдаун ppGalNAcT-14 или Fut-6 снижал рекрутинг FADD каспазы-8 для DISC, процессинг DISC-связанной каспазы-8 и стимуляцию связанной с DISC ферментативной активности каспазы-8 (Sharp et al., J. Biol. Chem., 280:19401 (2005)) (Фиг.12C, 12D и Фиг.15B). Ни ppGalNacT-14, ни Fut-6 siРНК, по существу, не изменяли количества DR4 и DR5 в DISC или дозозависимого связывания Apo2L/TRAIL с клетками PSN-1 или DLD-1, которые экспрессируют как DR4, так и DR5 (Фиг.12C, Фиг.15B и данные, которые не показаны). Таким образом, ppGalNAcT-14 и Fut-6, по-видимому, не модулируют апоптоз путем влияния на уровни рецепторов клеточной поверхности или Apo2L/TRAIL связывание. В соответствии с этим, Apo2L/TRAIL чувствительность в группе 77 клеточных линий не показала значительной корреляции с экспрессией на клеточной поверхности распознаваемых сигнальных рецепторов DR4 и DR5 или ложных рецепторов DcR1 и DcR2 (данные не показаны). Более того, большинство siРНК против ppGalNAcT-14, ppGalNacT-3 или Fut-6 не изменяли уровней DR4 и DR5 на клетках PSN-1, C170 или DLD-1 (Фиг.15C). Две siРНК не вызывали выявляемого уменьшения DR4 и DR5 в определенных клеточных линиях (Фиг.15C). Однако другие siРНК против тех же ферментов ингибировали Apo2L/TRAIL-индуцированный апоптоз, не влияя на уровни рецепторов.
Внеклеточный домен (ECD) DR5 человека экспрессировали в ооцитах китайского хомячка, секретированный белок очищали, подвергали кислотному гидролизу и анализировали соответствующие моносахариды (Фиг.13A). В соответствии с отсутствием ожидаемых сайтов N-гликозилирования в ECD DR5, авторы не обнаружили N-связанных гликанов. Однако в двух образцах 2 независимых экспериментов было идентифицировано 3 моль GalNAc и 3 моль Gal на моль ECD DR5 (Фиг.13A), указывая на O-связанную модификацию трех сайтов на DR5 с коровым гликаном GalNAc-Gal.
O-гликозилирование белка модифицирует серины или треонины. Используя ранее созданный инструмент биоинформатики для предопределения возможных сайтов O-гликозилирования (http://www.cbs.dtu.dk/services/NetOGlyc) (Julenius et al., Glycobiology, 15:153 (2005)), авторы идентифицировали две такие области в общей последовательности ECD длинного (DR5-L) и короткого (DR5-S) вариантов сплайсинга DR5 человека, а третий сайт - в пределах области альтернативного сплайсинга (Фиг.13B). Первый аминокислотный сегмент (74-77) содержит 3 серина; второй (130-144) имеет 5 треонинов, тогда как третий имеет 4 треонина и 3 серина. DR5 мыши имеет последовательности, сходные с первыми 2 сегментами с 2 серинами и 4 треонинами соответственно, тогда как DR4 человека также имеет 2 сходные последовательности, содержащие 1 серин и 5 треонинов. Для тестирования, могут ли эти сайты быть важными для пост-трансляционной модификации DR5, была получена серия DR5L и DR5S мутантов с заменой на аланины либо 5 треонинов сегмента 130-144 (DR5L-5T, DR5S-5T), либо этих же 5 треонинов, а также 3 серинов сегмента 74-77 (DR5L-5T3S, DR5S-5T3S). DR5 антительный иммуноблоттинг лизатов клеток HEK293, трансфектированных DR5L или DR5S, выявил наличие ожидаемых полос DR5L или DR5S (Фиг.13C). Это антитело также выявило полосы DR5 более высокой молекулярной массы (ММ), которая становится более распространенной при ко-трансфекции DR5L или DR5S с использованием ppGalNAcT-14 по сравнению с контролем (Фиг.13C, звездочки). Распространение этих полос более высокой ММ и их нарастание под действием ppGalNAcT-14 значительно снижалось с DR5L-5T или DR5S-5T и почти исчезало с DR5L-5T3S или DR5S-5T3S по сравнению с конструкциями дикого типа. Эти результаты указывают на то, что полосы более высокой ММ представляют О-гликозилированные формы DR5: ppGalNAcT-14 способствует их образованию, а постепенное удаление предполагаемых сайтов О-гликозилирования посредством замены аланином последовательно изменяет этот эффект. Трансфекция клеток HEK293 с использованием DR5 мыши или DR4 человека, DR5L или DR5S демонстрировала гибель клетки (Фиг.13D); каждый мутант DR5 демонстрировал меньшую активность, чем соответствующая ему конструкция дикого типа, с DR5S-5T3S (у которого отсутствуют все три сайта), имеющим наиболее слабую активность. Ко-трансфекция с ppGalNAcT-14 заметно усиливала гибель клетки под действием всех конструкций DR4 и DR5, за исключением DR5S-5T3S, которая показала значительно меньшую активность.
Большинство образцов нормальной ткани и образцов опухоли рака кожи, легких, поджелудочной железы, груди, яичников, эндометрия и мочевого пузыря или неходжкинской лимфомы демонстрировали экспрессию мРНК ppGalNAcT-14 ниже пороговых значений (определенных при 500 для большинства злокачественных опухолей и при 200 для рака кожи, Фиг.13E). Однако существенная подгруппа образцов опухоли в интервале от 10% при лобулярном раке груди до 30% при раке легких и диффузной крупноклеточной В-клеточной лимфоме демонстрировала гиперэкспрессию ppGalNAcT-14. Уровни экспрессии мРНК некоторых образцов злокачественных клеток были более чем 1000 раз выше значений в соответствующих нормальных тканях. Динамичная экспрессия ppGalNAcT-14 при злокачественных опухолях указывает на то, что этот ген и, возможно, другие родственные ферменты могут предоставить полезные биомаркеры для идентификации опухолей с большей чувствительностью к Apo2L/TRAIL.
O-связанные гликаны демонстрируют большое структурное разнообразие, и они модулируют различные аспекты биологии белков плазматических мембран, в том числе конформацию, агрегацию, направленную миграцию, период полужизни, а также клеточную адгезию и активность путей передачи сигнала (Hang et al., Bioorg. Med. Chem., 13:5021 (2005); Hanisch, Biol. Chem., 382:143 (2001)). В злокачественных клетках зачастую проявляются поразительные изменения профилей О-гликанов, создающие уникальные ассоциированные с опухолью углеводные антигены (Brockhausen, Biochim. Biophys. Acta, 1473:67 (1999); Dube et al., Nat. Rev. Drug Discovery, 4:477 (2005); Fuster et al., Nat. Rev. Cancer, 5:526 (2005)). O-гликозилирование также играет важную роль в возвращении опухолевых клеток к специфичным участкам метастаз (Fuster et al., Cancer Res., 63:2775 (2003); Ohyama et al., EMBO J., 18:1516 (1999); Takada et al., Cancer Res., 53:354 (1993)). Значительная подгруппа образцов первичных опухолей из различных злокачественных опухолей человека демонстрирует повышенную экспрессию фермента О-гликозилирования ppGalNAcT-14, в том числе образцы клеток толстой кишки и ободочной и прямой кишки, образцы клеток меланомы и образцы клеток хондросаркомы.
СПОСОБЫ
Материалы
Реактивы для клеточных культур покупали на фирме Gibco (Invitrogen/Gibco, Carlsbad, CA), немеченый растворимый Apo2L/TRAIL готовили, как описано ранее (Lawrence et al., Nat. Med., 7:383 (2001)), ингибитор O-связанного гликозилирования бензил-a-GalNAc покупали у Calbiochem, а все остальные химические препараты (в том числе этопозид и стауроспорин) были от Sigma Aldrich (St. Louis, MO).
Клеточная культура и клеточные линии
Все 119 клеточных линий карциномы человека (наименования и номера каталогов см. в дополнительных сведениях) получали из ATCC или DSMZ (Braunschweig, Germany) и культивировали при 37°C и 5% CO2 в RPMI1640, дополненной 10% инактивированной нагреванием эмбриональной сывороткой теленка, 2 мM L-глутамина и 10 мM HEPES без антибиотиков, подобных пенициллину/стрептомицину. 293 эмбриональные почечные клетки человека (номер каталога CRL-1573) также получали из ATCC и культивировали в 100% модифицированной по Дульбекко среде Игла, дополненной 10% FBS. На O-гликозилированную мутантную клеточную линию CHO, ldlD CHO было получено разрешение от Dr. Monty Kreiger, MIT (Boston MA).
Анализы жизнеспособности клеток и анализы апоптоза
Для определения IC50 для Apo2L/TRAIL клетки высевали в трех повторах в 96-луночные планшеты, оставляли для адгезии на 24 часа, а затем обрабатывали рекомбинантным Apo2L/TRAIL человека в возрастающих концентрациях, вплоть до 1000 нг/мл. После инкубации в течение 72 ч их затем подвергали исследованию на жизнеспособность - МТТ анализу (Pierce) или люминесцентному анализу жизнеспособности клеток CellTiter-Glo (Promega) - в соответствии с протоколом производителя. Каждый эксперимент на жизнеспособность клеток повторяли по меньшей мере три раза при низкой (0,5%) и высокой (10% FBS) концентрации сыворотки, и клеточные линии с переходной чувствительностью характеризовались колебаниями между значениями IC50 независимых экспериментов или между сывороткой низкой и высокой концентрации. Клеточная линия характеризовалась как чувствительная, исходя из индукции апоптоза по меньшей мере 50% клеток при концентрации Apo2L/TRAIL 1 мгл/мл и как с переходной чувствительностью на основе изменения степени апоптоза, индуцированного в независимых экспериментах или в присуствии сыворотки низкой концентрации (0,5%) по сравнению с сывороткой высокой (10%) концентрации. Апоптоз количественно оценивали с помощью проточной цитометрии среднего процентного содержания собранных клеток (адгезированных + плавающих в среде) окрашенных аннексином V (BD Pharmingen).
Гибридизация на микрочипе и анализ данных
Общую клеточную РНК получали из необработанных клеток (3 x 106), используя набор RNeasy Kit (Quiagen). Меченую кРНК получали и гибридизировали с нуклеотидными микрочипами (U133P GeneChip; Affymetrix Incorporated, Santa Clara, CA), как описано ранее (Hoffman et al., Nat. Rev. Genetics, 5:229 (2004); Yauch et al., Clin. Cancer Res., 11:8686 (2005)). Файлы сканированных изображений анализировали с использованием GENECHIP 3.1 (Affymetrix), Spotfire, GenePattern и Cluster/TreeView. Для идентификации наиболее различно экспрессированных генов между чувствительными и резистентными клеточными линиями авторы подвергли значения генной экспрессии выборке изменений, в которой исключили гены с минимальным изменением по всем анализируемым образцам, тестируя на кратность изменения и абсолютное изменение по всем образцам, сравнивая соотношение максимума и минимума (макс/мин) и различие между максимумом и минимумом min (макс-мин) с предварительно определенными значениями и исключая гены, не удовлетворяющие обоим условиям.
Экспрессионные конструкции и ретровирусная трансдукция
Фрагмент ДНК, кодирующий ppGalNAcT-14, клонировали из кДНК, объединенных в пул из Apo2L/TRAIL чувствительных клеточных линий, и вставляли в экспрессионную плазмиду pcDNA3.1 (Invitrogen) с N-концевой Flag меткой. Эту конструкцию затем подвергали сайт-направленному мутагенезу (Quikchange Mutagenesis kit, Stratagene) для генерации siРНК молчащих мутантов, которые имели 4-6 “wobble” изменения пар оснований в последовательности, гомологичной siРНК, без изменения последовательности белка. Эти мутации перекрывали область 10 нуклеотидных пар в центре связывающей последовательности siРНК длиной 19 н.п. Последовательности ДНК для DR5Long и DR5Short, DR4, TRAIL рецептора мыши, DR4, Fas (вариант 1), TNFR1 и Bax (вариант бета) клонировали из пулов кДНК и вставляли в вектор экспрессии pRK (Genentech). O-гликозилированные мутанты DR5L и DR5S получали сайт-специфичной мутацией четырех треонинов на остатки аланина, Mut4xTA (T130, T131, T132, T135) или пяти треонинов на остатки аланина Mut5xTA (T130, T131, T132, T135, T143). Транзиторную трансфекцию в клетки HEK293 экспрессионными конструкциями проапоптотических молекул осуществляли в 6-луночных планшетах с концентрацией 0,5 мкг/лунку проапоптотической молекулы и 2,0 мкг ppGalNAcT-14 или векторного контроля. Клетки трансфектировали, используя липофектамин 2000 в соответствии с протоколом производителя. После 48 часовой инкубации клетки подвергали анализу на апоптоз.
Для получения ретровирусных конструкций ppGalNAcT-14 и мутанты клонировали в ретровирусный вектор pQCXIP (Clontech). Высокий титр ретровирусных супернатантов получали с использованием линии хелперных клеток ΦNX-Ampho. Упаковывающие клетки трансфектировали с использованием фосфата кальция (Invitrogen). Супернатанты выделяли через 48 часов после трансфекции и добавляли к клеткам-мишеням наряду с 10 мкг/мл полибрена, с последующей стадией 1 ч центрифугирования при 2700 об/мин для усиления инфицирования. После трансдукции клетки подвергали селекции 2 мкг/мл пуромицина.
Конструкция siРНК и протоколы трансфекции
siРНК против ppGalNAcT-14, ppGalNAcT-3, каспазы-8 и DR5 были сконструированы Dharmacon (Lafayette, CO) с использованием их собственного критерия отбора. Выбранные последовательности представляли собой:
siGalNAcT-14 (1): 5' GACCATCCGCAGTGTATTA-dTdT 3' (=14-4) (SEQ ID NO:15)
siGalNAcT-14 (2): 5' ATACAGATATGTTCGGTGA-dTdT 3' (=14-6) (SEQ ID NO:16)
siGalNAcT-3 (1): 5' CCATAGATCTGAACACGTT-dTdT 3' (=3-2) (SEQ ID NO:17)
siGalNAcT-3 (2): 5' GCAAGGATATTATACAGCA-dTdT 3' (=3-7) (SEQ ID NO:18)
siFut-6 (1) 5' GUACCAGACACGCGGCAUA-dTdT 3' (=6-1) (SEQ ID NO:19)
siFut-6 (2) 5' ACCGAGAGGUCAUGUACAA-dTdT 3' (=6-2) (SEQ ID NO:20)
siКаспаза-8: 5' GGACAAAGTTTACCAAATG-dTdT 3' (SEQ ID NO:21)
siРНК приобретали в виде двухспиральных РНК олигонуклеотидов и трансфектировали в соответствующие клеточные линии с конечной концентрацией 25 нM для каждой siРНК. Дуплексы siРНК против нецелевой последовательности (Dharmacon) использовали в качестве контроля. Клетки трансфектировали с использованием липофектамина 2000 (Invitrogen) путем обратной трасфекции, где клетки добавлены в суспензию к предварительно нанесенным комплексам липид-siРНК. Концентрации липофектамина 2000 соответствовали протоколу производителя. После инкубации 48 ч клетки собирали для анализа мРНК или инкубировали с рекомбинантным Apo2L/TRAIL человека, этопозидом или стауроспорином в течение еще 24-72 часов для исследований жизнеспособности или в течение 4, 8 или 24 ч для Вестерн блоттинга.
Ингибирование O-гликозилирования под действием бензил-a-GalNAc.
Клетки Colo205 выращивали в присутствии бензил-a-GalNAc (2 мM или 4 мM) в течение 72 часов. В этот момент их пересаживали в 96-луночные планшеты, оставляя для адгезии на 24 часа, все еще в присутствии ингибитора. Затем их стимулировали возрастающими концентрациями Apo2L/TRAIL, как указано, и подвергали исследованию на жизнеспособность.
Количественная ПЦР
Уровни экспрессии транскриптов GalNacT-14 и GalNacT-3 оценивали с помощью количественной ОТ-ПЦР, используя стандартные методики Taqman. Уровни транскриптов нормировали к облигатному гену, GAPDH, и результаты выражали в виде нормированных значений экспрессии (=2-DCt). Наборы праймер/зонд для GalNacT-14 (cat#: Hs00226180_m1_GT14), GalNacT-3 (cat#:Hs00237084_m1_GT3) и GAPDH (cat#: 402869) приобретали у Applied Biosystems (Foster City, CA).
Иммунопреципитация, Вестерн блоттинг и антитела
Иммунопреципитация: анти-Apo2L (2E11; ATCC регистрационный номер HB-12256), анти-DR4 (3G1 и 4G7, ATCC регистрационный номер PTA-99) и анти-DR5 (3H3, ATCC регистрационный номер 12534 и 5C7) моноклональные антитела получали в Genentech, Inc., используя в качестве антигенов слитые белки рецептор-Fc. Анти-DR4 (4G7) и анти-DR5 (5C7) моноклональные антитела, используемые для иммунопреципитации Apo2L/TRAIL DISC, конъюгировали с агарозой, используя набор ImmunoPure Protein G IgG Plus orientation kit (каталожный номер 44990) от Pierce. Анти-DR4 (3G1) и анти-DR5 (3H3) моноклональные антитела, используемые для иммунодетекции DR4/5 в иммунопреципитатах DISC, биотинилировали, используя набор для биотинилирования EZ-link Sulfo-NHS-LC (каталожный номер 21217) от Pierce. Получали Apo2L/TRAIL, меченные FLAG, и перекрестно сшивали с анти-FLAG антителом M2 (Sigma), как описано (Kischkel, Immunity, 12:611 (2000)). Эти эксперименты проводили, как описано ранее для Apo2L/TRAIL-FLAG + анти-FLAG DISC анализа (Kischkel, supra). Эксперименты по DR4/5 DISC иммунопреципитации проводили, как описано, за исключением того, что анти-DR4 (4G7) и анти-DR5 (5C7) моноклональные антитела непосредственно конъюгировали с агарозой для иммунопреципитации (Sharp et al., J. Biol. Chem., 280:19401 (2005)).
Вестерн блоттинг: 5×105 клеток на лунку высевали в 6-луночные планшеты. Для экспериментов по РНКi нокдауну клетки обрабатывали различными siРНК в течение 48 часов, затем Apo2L/TRAIL в течение 4, 8 или 24 ч. После указанных периодов времени клетки промывали в ледяном PBS и лизировали в 1% Тритоне X-100, содержащем гипотонический лизирующий буфер (20 мM HEPES pH 7,5, 10 мM KCL, 1,5 мM MgCl2, 1 мM EDTA и 1 мM DTT). Для каждого образца 40 мкг белка разделяли в редуцирующих условиях на 10% или 10-20% градиентных SDS-полиакриламидных гелях. После переноса на нитроцеллюлозные мембраны (Schleicher и Schuell) мембраны инкубировали в течение 1 ч в 10% обезжиренном молочном порошке с последующей 1 ч инкубацией со следующими первичными антителами: козье антитело к каспазе-3 (1:1000, R&D), кроличье антитело к каспазе-8 (1:1000, Pharmingen), мышиное антитело к каспазе-9 5B4 (1:1000, MBL), кроличье антитело к Bid (1:1000, Pharmingen), кроличье антитело к DR5 (1:500, Cayman) или козье антитело к актину (1:200, Santa Cruz Biotechnology). Мембраны промывали пять раз TBS/0,05% Твином, а затем инкубировали с соотвествующим конъюгированным с пероксидазой аффинно очищенным вторичным антителом (1:5000, Biorad) в течение 30 мин. Мембраны снова промывали пять раз и проявляли, используя усиленную химилюминесценцию (ECL, Amersham) и пленки Kodak Biomax.
Проточная цитометрия/FACS анализ.
Поверхностную экспрессию рецепторов семейства TNF DR4 и DR5 определяли с помощью активированного флуоресценцией клеточного сортинга (FACS), используя проточный цитометр FACS Calibur (Becton Dickinson Immunocytometry System, San Jose, CA). Клетки C170 и PSN-1, трансфектированные указанными siРНК в течение 48 ч, окрашивали 10 мкг/мл первичным антителом, 4G7 (анти-DR4) или 3H3 (анти-DR5), или контрольным антителом мышиным IgG в течение 1 ч при 4°C. Затем клетки промывали PBS, а затем инкубировали со вторичным козьим антимышиным антителом, конъюгированным с флуоресцеином (FITC) (Jackson Laboratories) в течение 30 мин при 4°C. Затем клетки анализировали проточной цитометрией, используя проточный цитометр FACS Calibur.
Исследования каспазы
Активности каспазы-3/-7 исследовали при 37°C в 40 мкл каспазного буфера (50 мM HEPES pH 7,4, 100 мM NaCl, 10 % сахарозы, 1 мM EDTA, 0,1% CHAPS и 10 мM DTT), содержащего 100 мкM флуорогенного пептида Ac-DEVD-AFC. Активность измеряли непрерывно на протяжении указанного времени по высвобождению AFC из DEVD-AFC, используя молекулярные устройства флуориметра в динамическом режиме с парой фильтров 405-510. Для оценки каспазной активности 20 мкг общего клеточного белка (экстракты Тритоном X-100) использовали в 40 мкл каспазного буфера (содержащего 100 мкM DEVD-AFC).
Анализ углеводов DR5, происходящего из CHO
Композицию моносахаридов DR5 из клеток CHO получали после гидролиза с 4 н. TFA. Анализ извлеченных моносахаридов проводили на системе ВЭЖХ Dionex BioLC, используя анионообменную хроматографию высокого разрешения, соединенную с импульсным амперметром.
Животные и изучение подкожного ксенотрансплантата
Самок бестимусных мышей «nude» (The Jackson Laboratory, Bar Harbor, ME, USA) приучали к условиям содержания животных Genentech минимум в течение 1 недели до постановки на экспериментальное исследование. Все экспериментальные процедуры были одобрены комитетом по биоэтике Genentech (IACAUC). Мышей заражали подкожно 5 × 106 клеток/мышь Colo205, DLD-1 и RKO или 20 × 106 клеток/мышь клетками карциномы толстой кишки человека Colo320HSR (Американская коллекция типовых культур). Размеры опухоли получали с помощью цифрового штангенциркуля, и объемы опухоли вычисляли, используя формулу □~□ (A= длина) (B=ширина)2. При достижении опухолями объема ~ 150-200 мм3, мышей случайным образом группировали и применяли внутрибрюшинную обработку (и.п.) с носителем или Apo2L/TRAIL (60 мг/кг/день) в дни 0-4.






















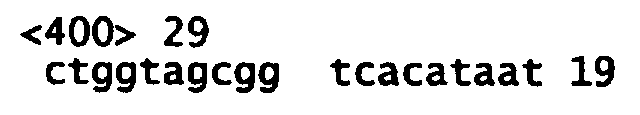


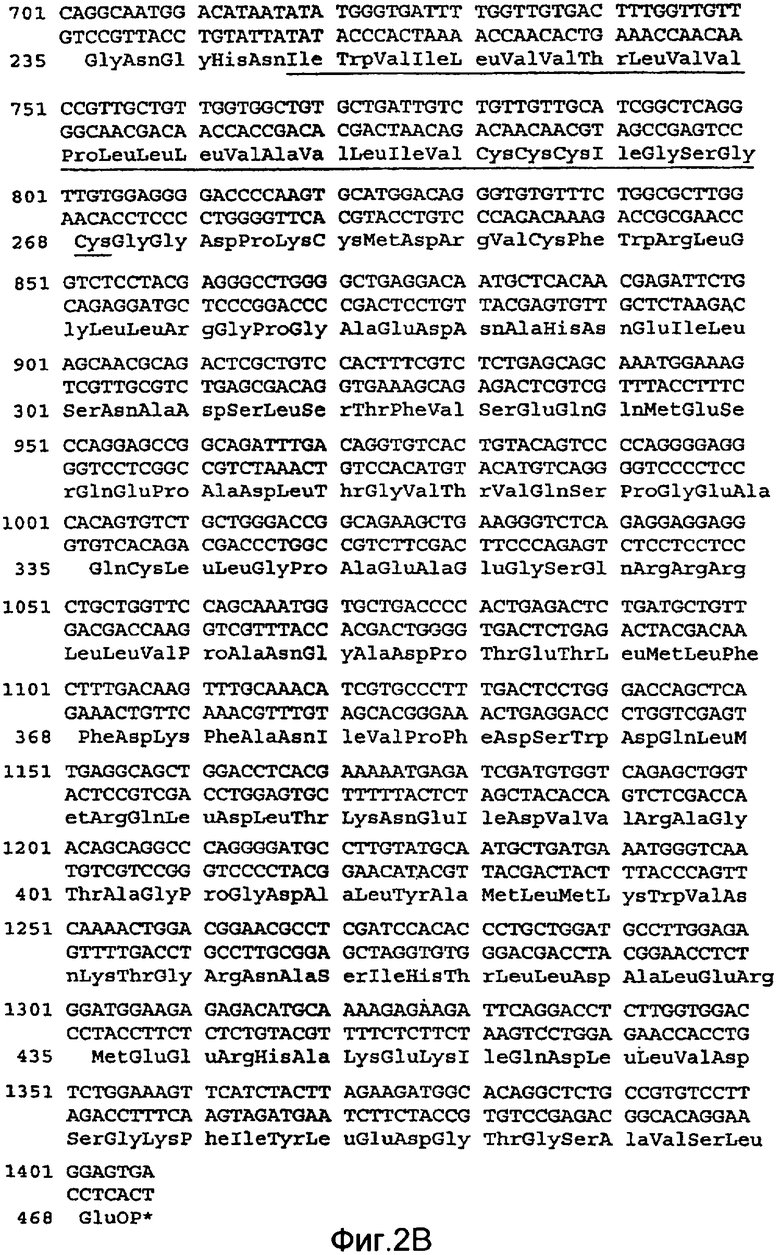






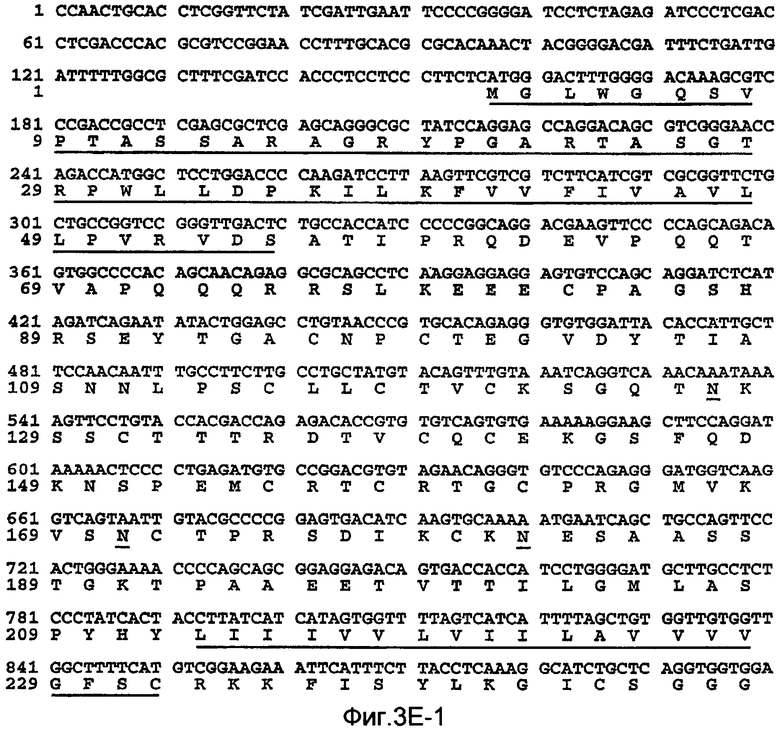
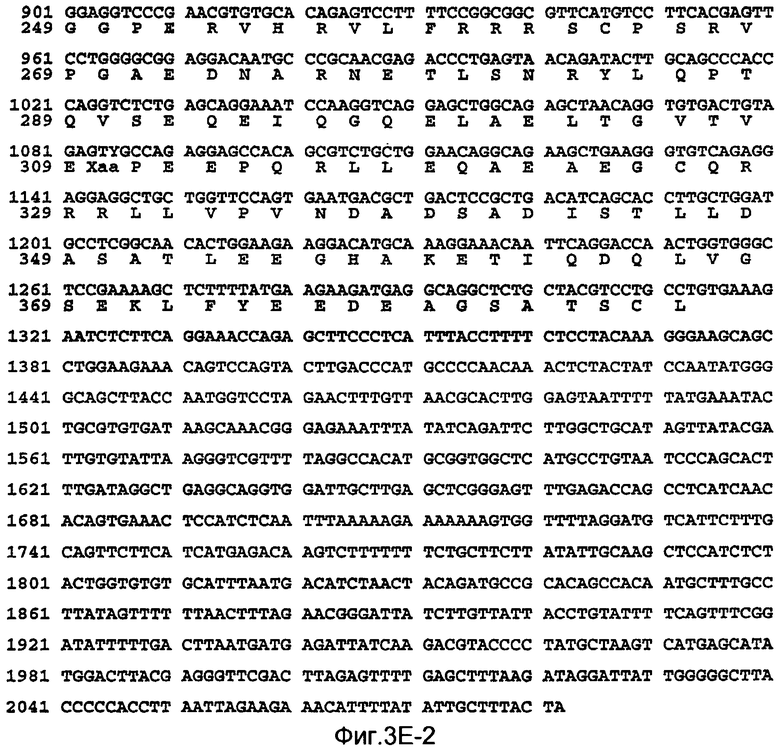
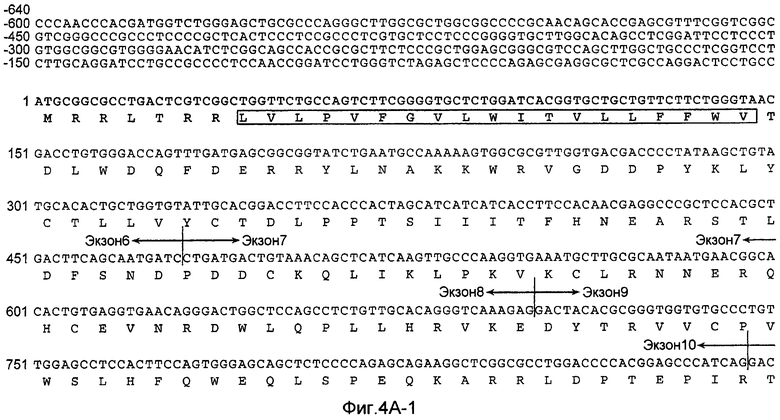
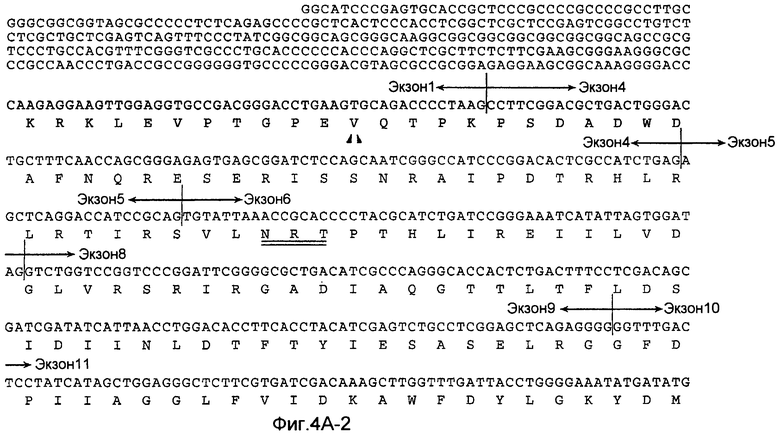
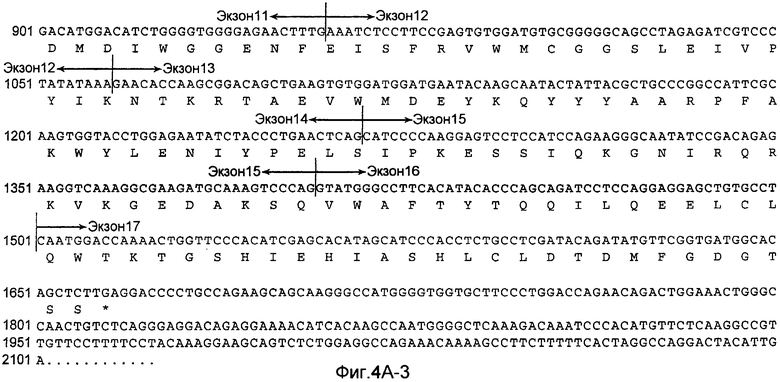
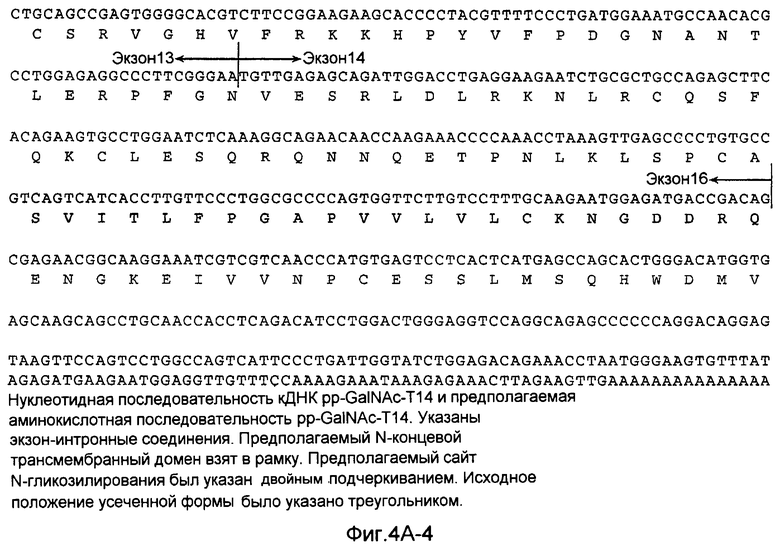
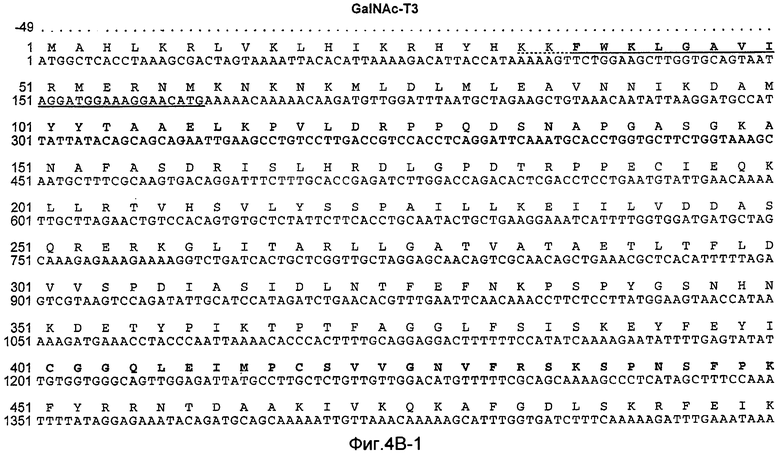
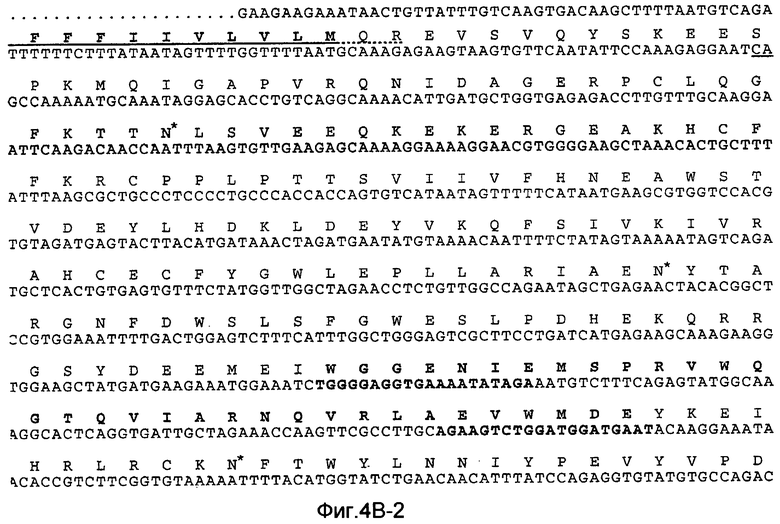
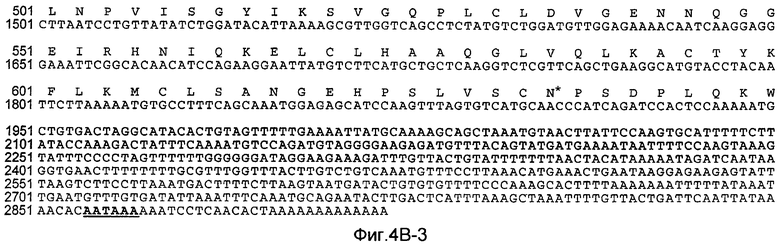

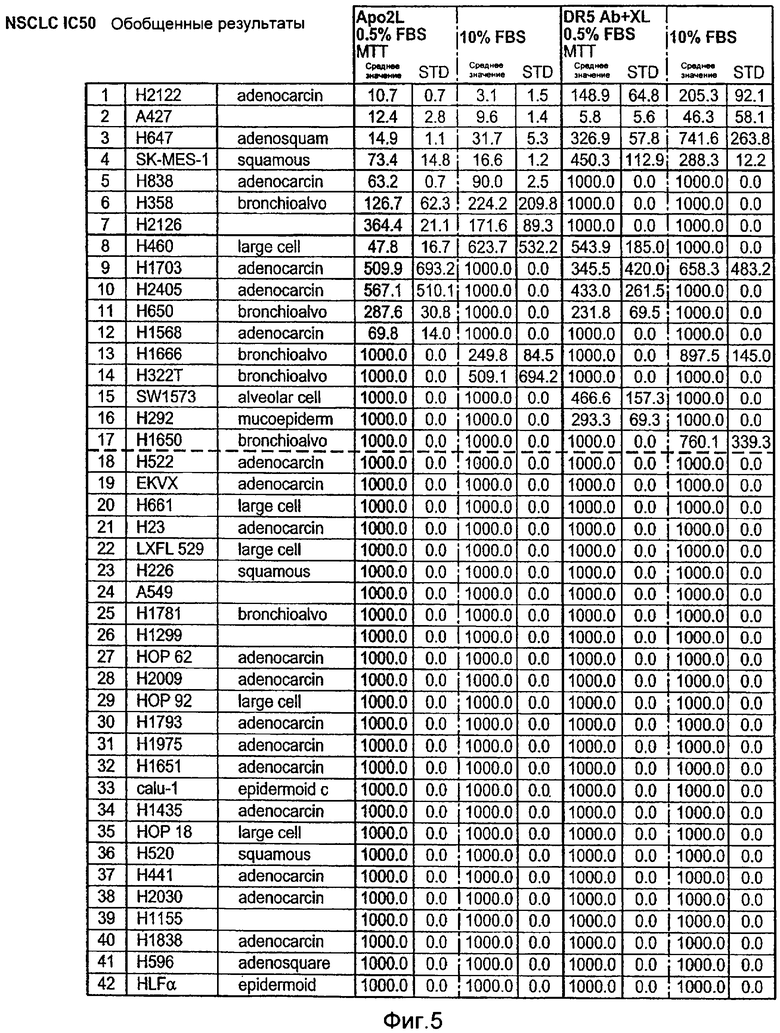
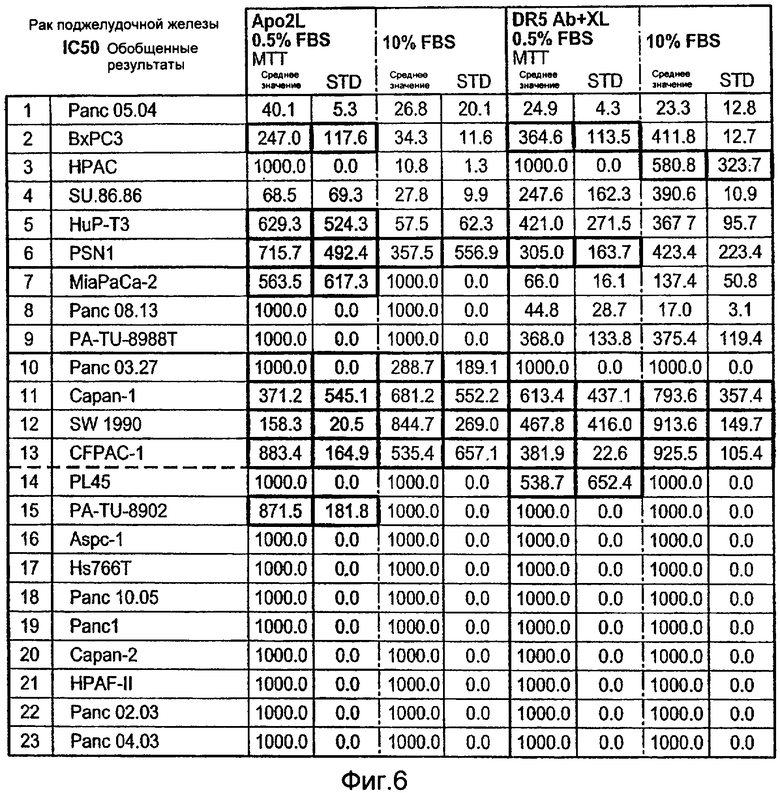
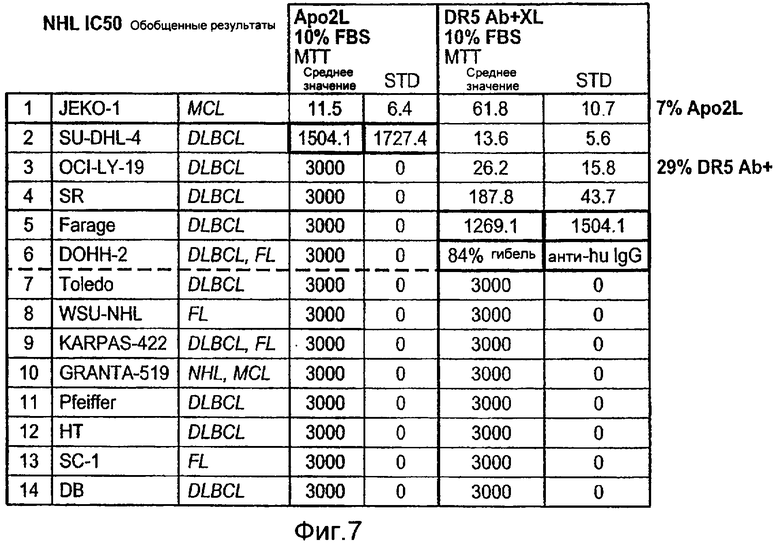
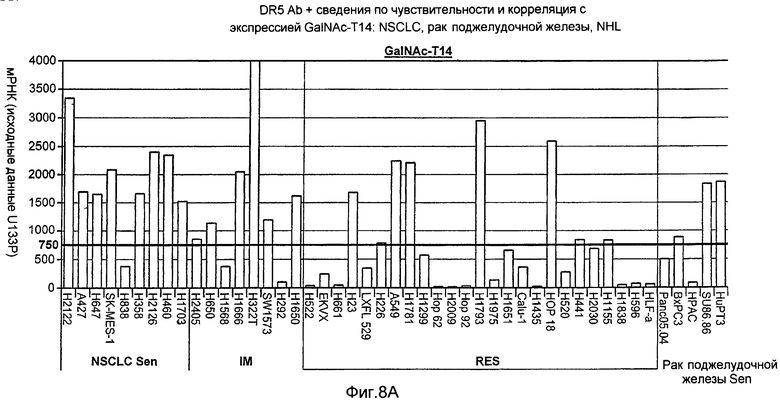
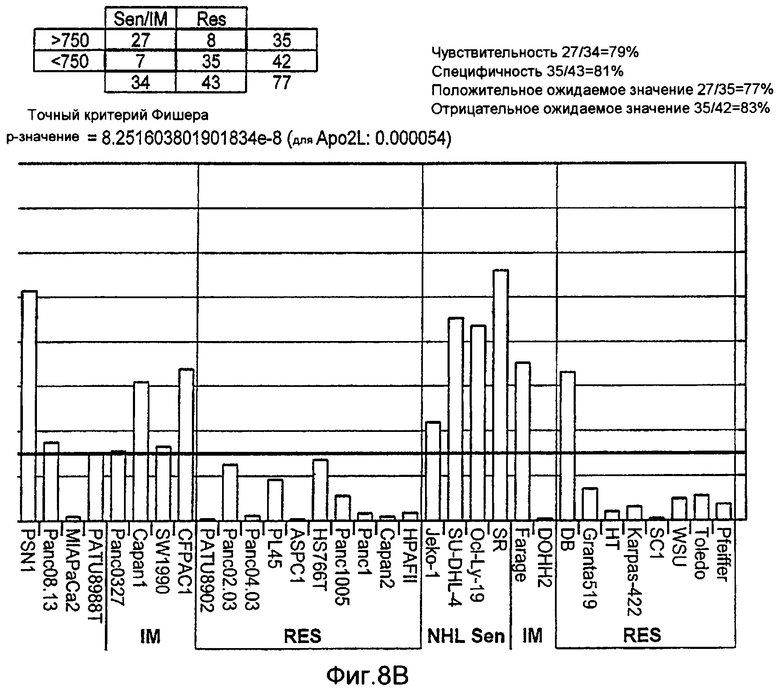

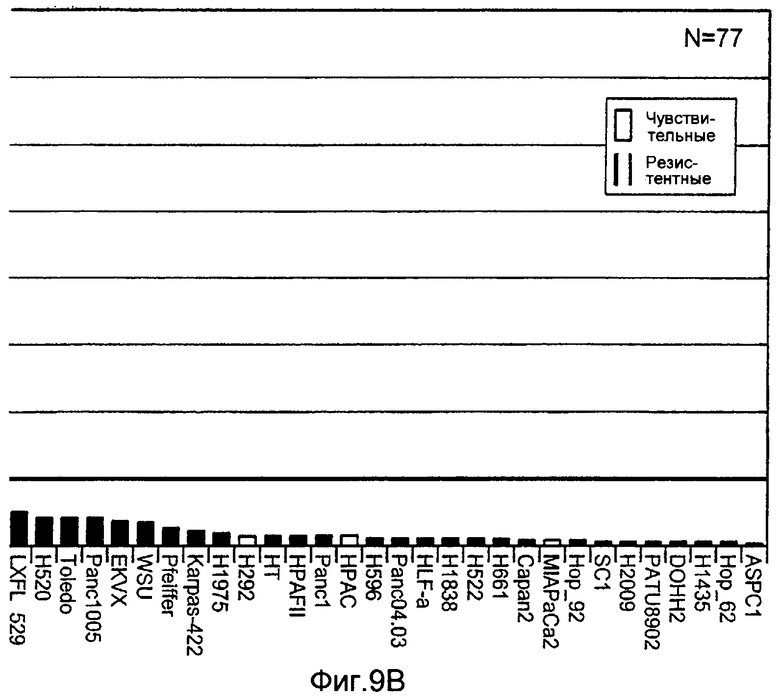
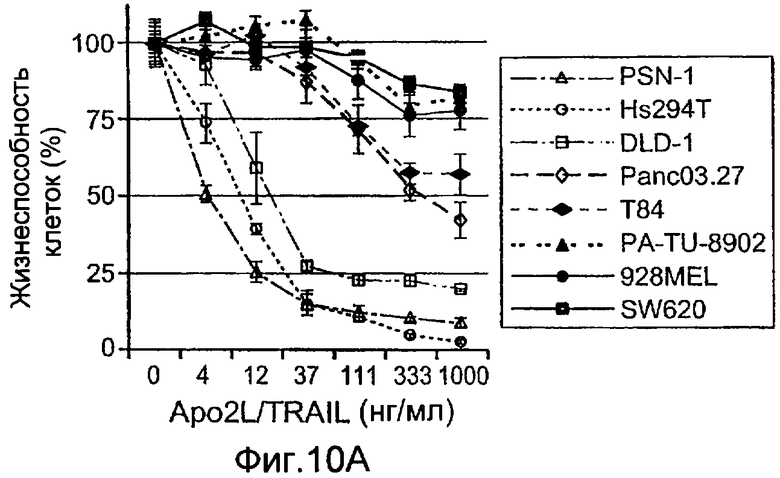
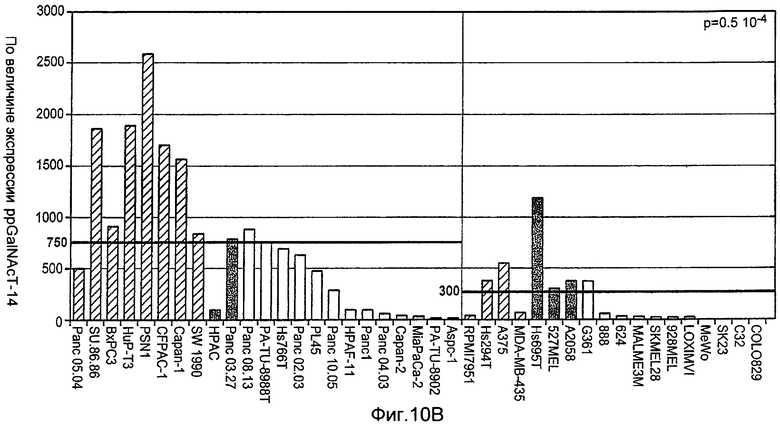
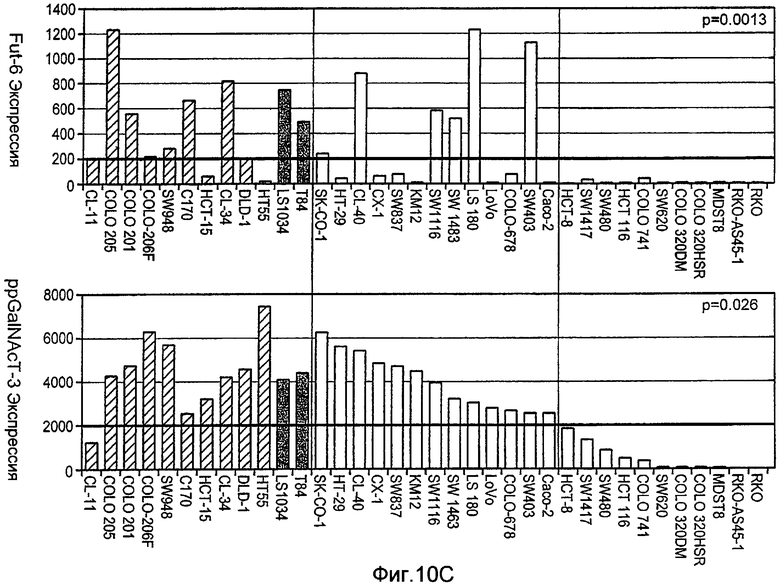
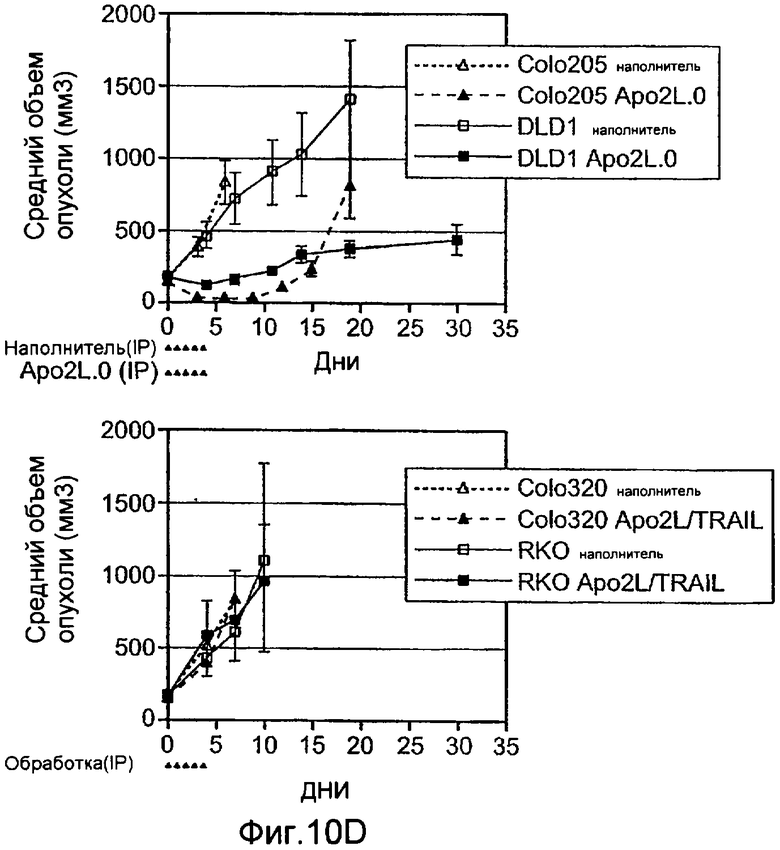
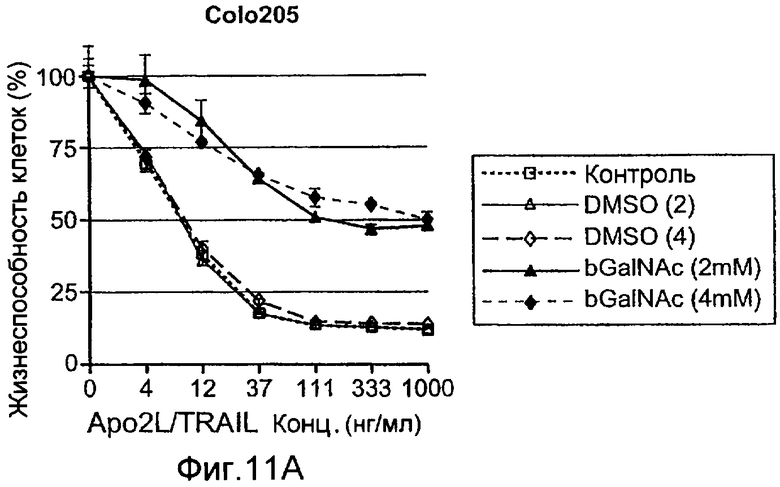

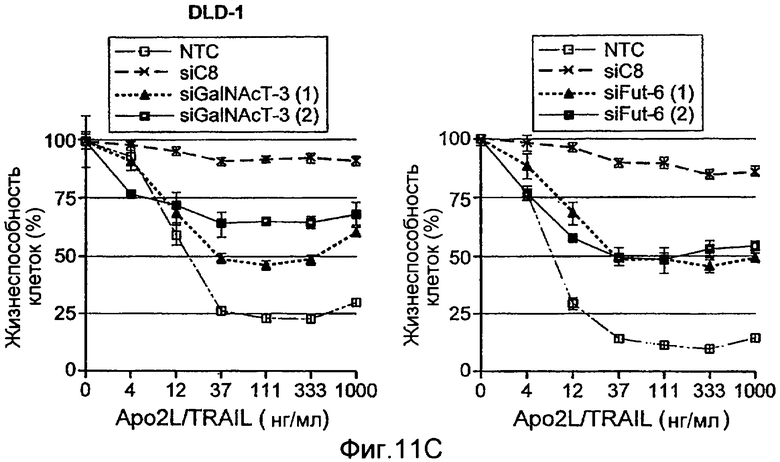

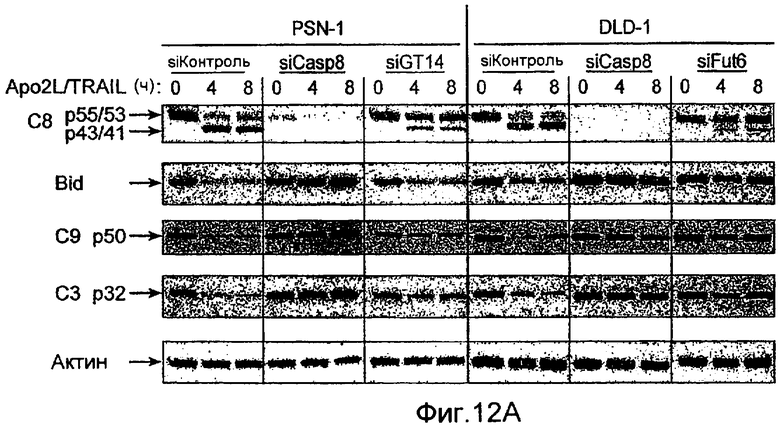
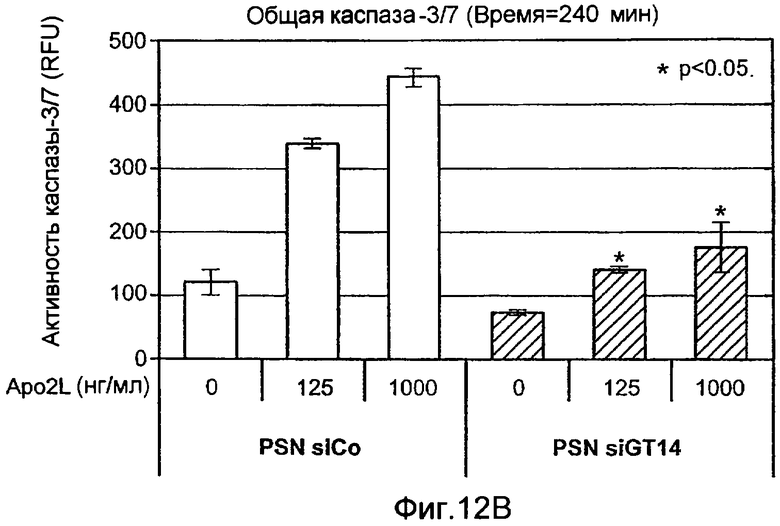
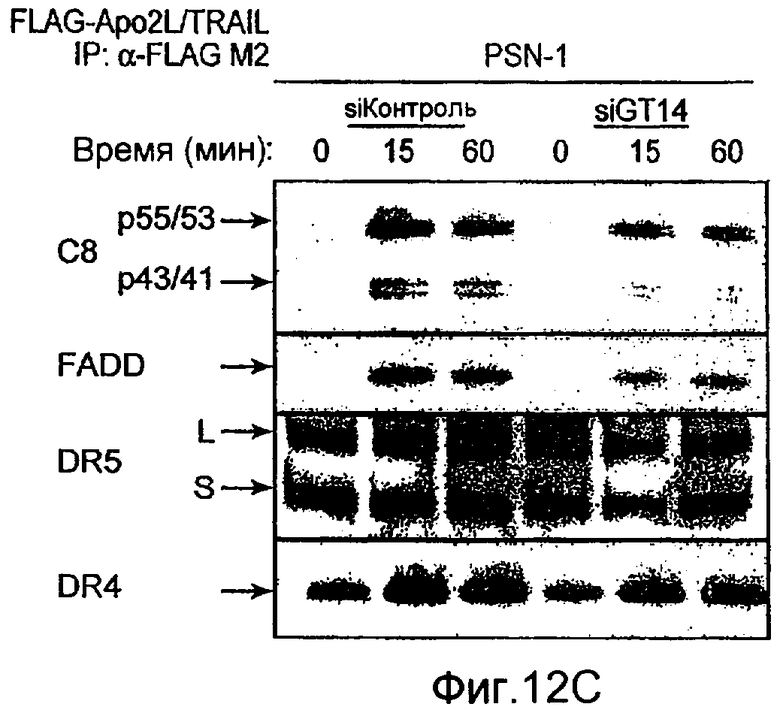
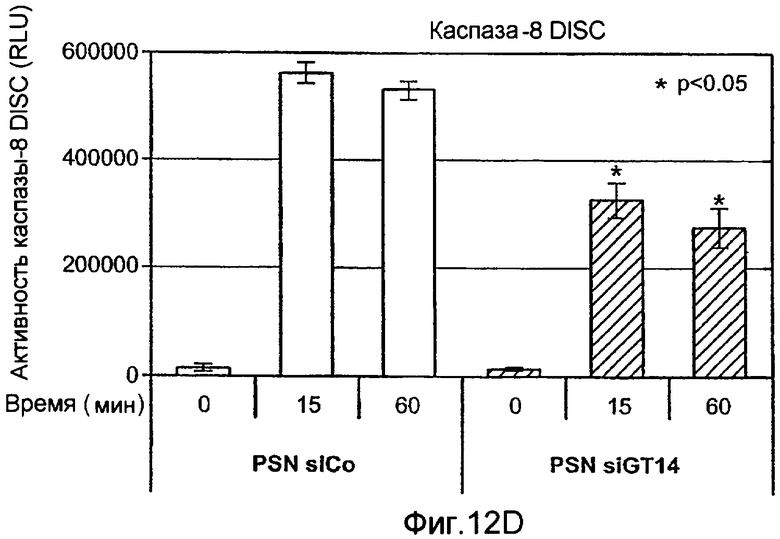

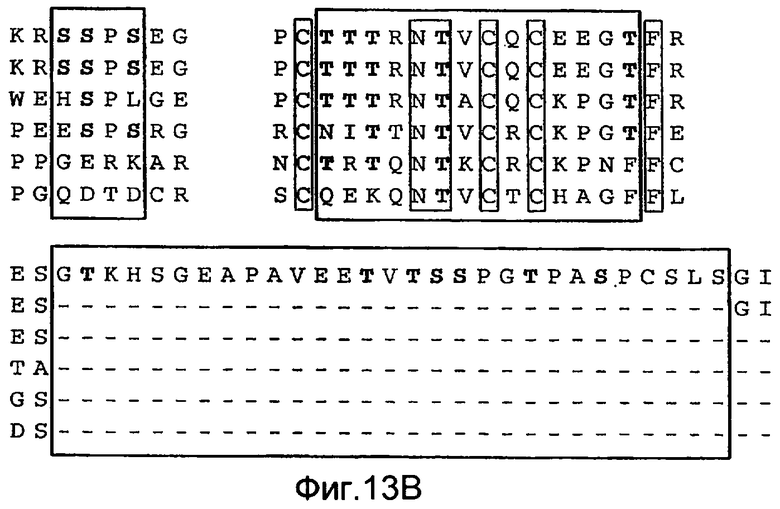
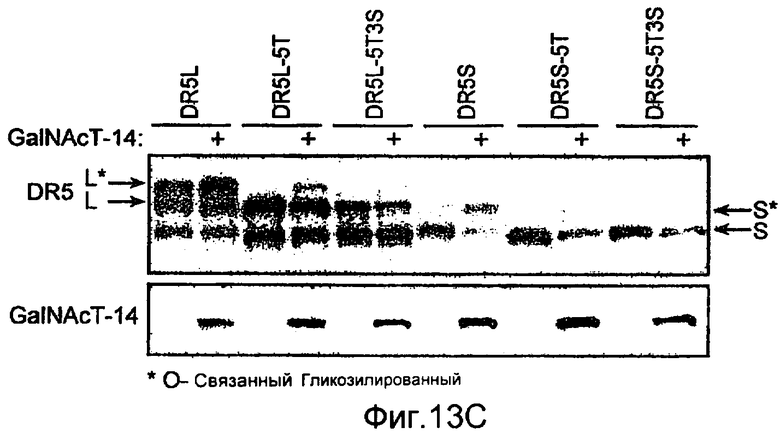
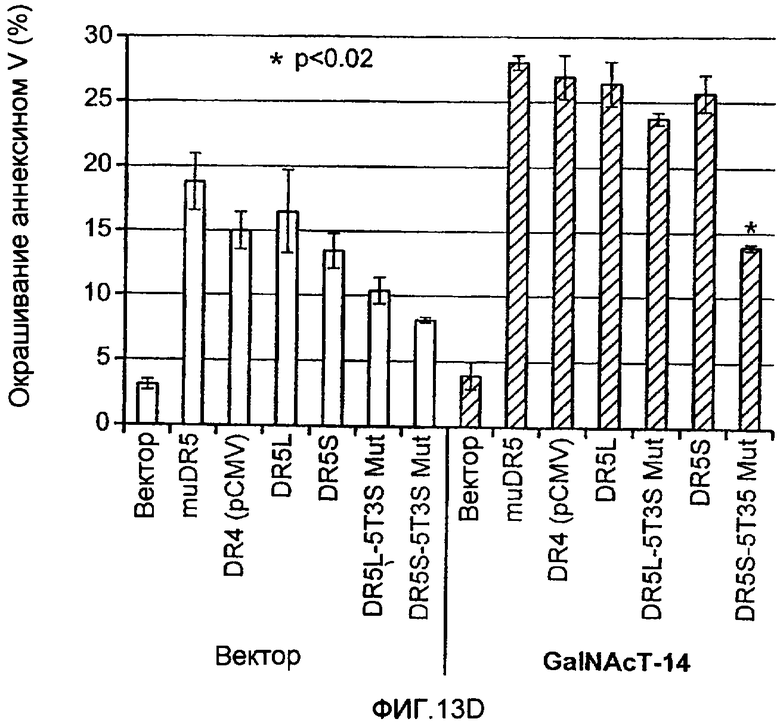
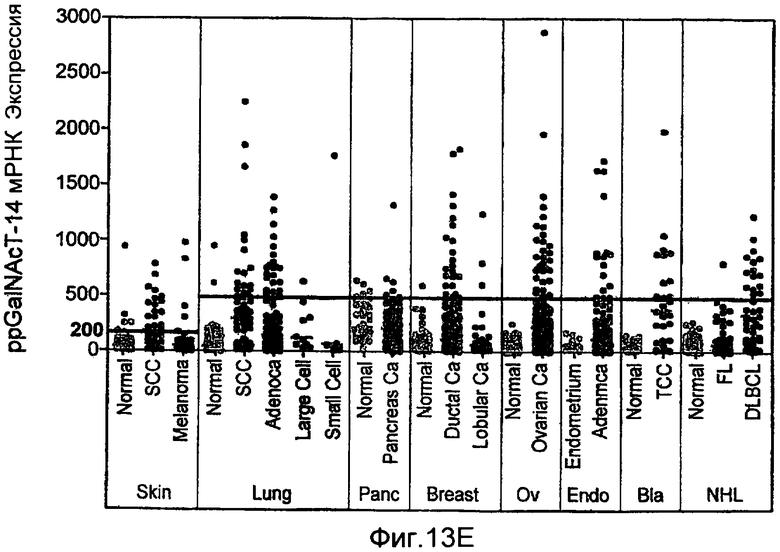
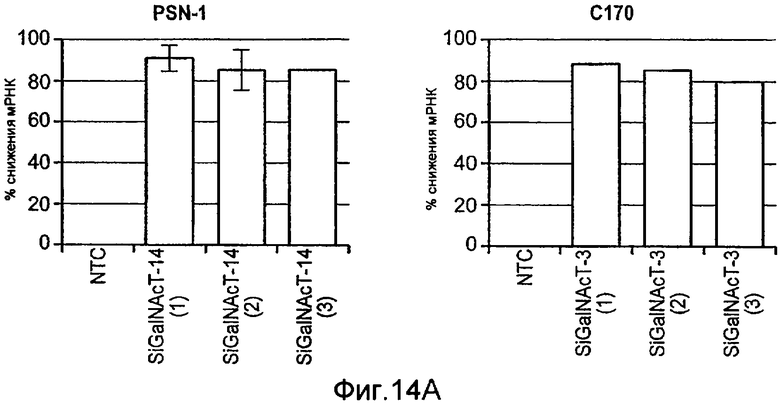

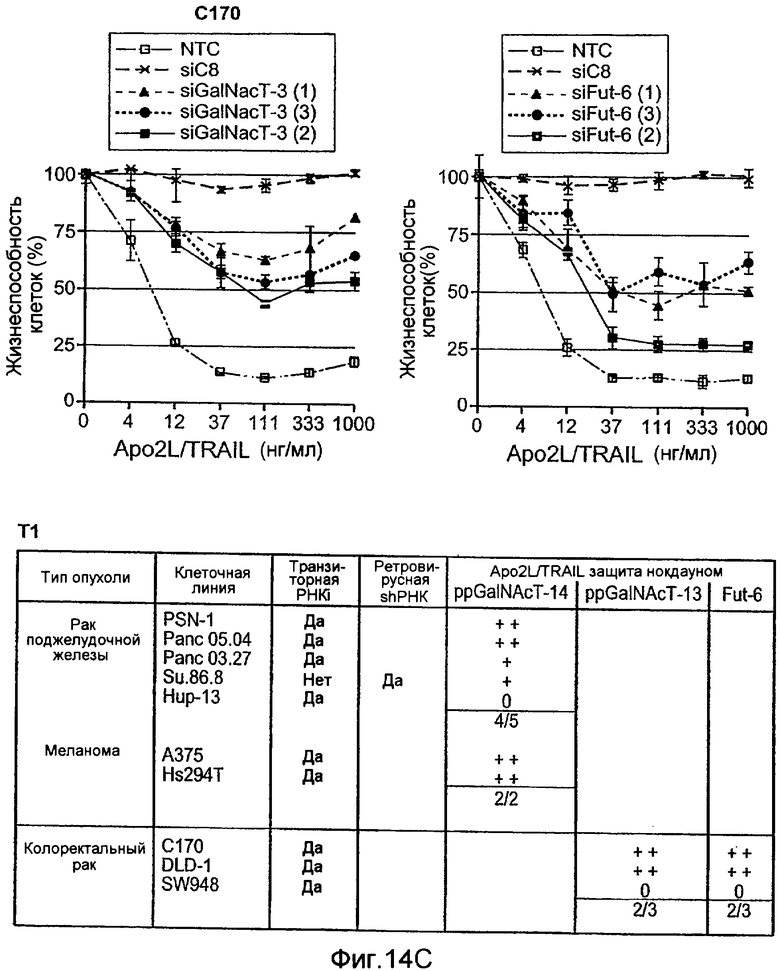
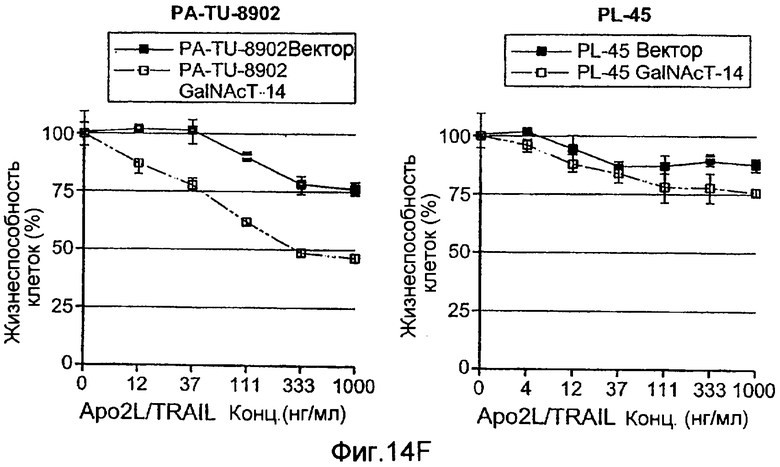
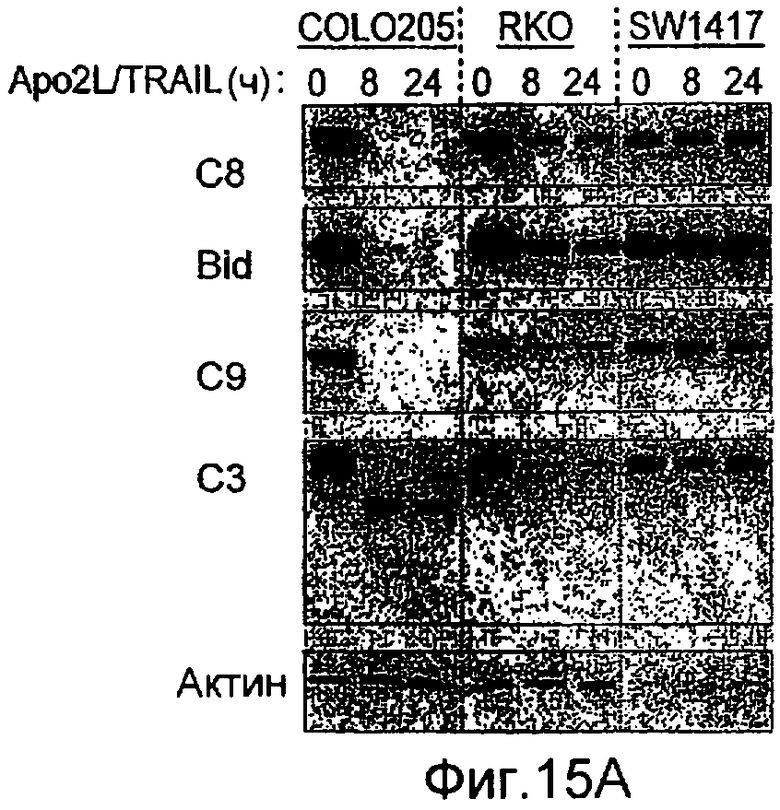
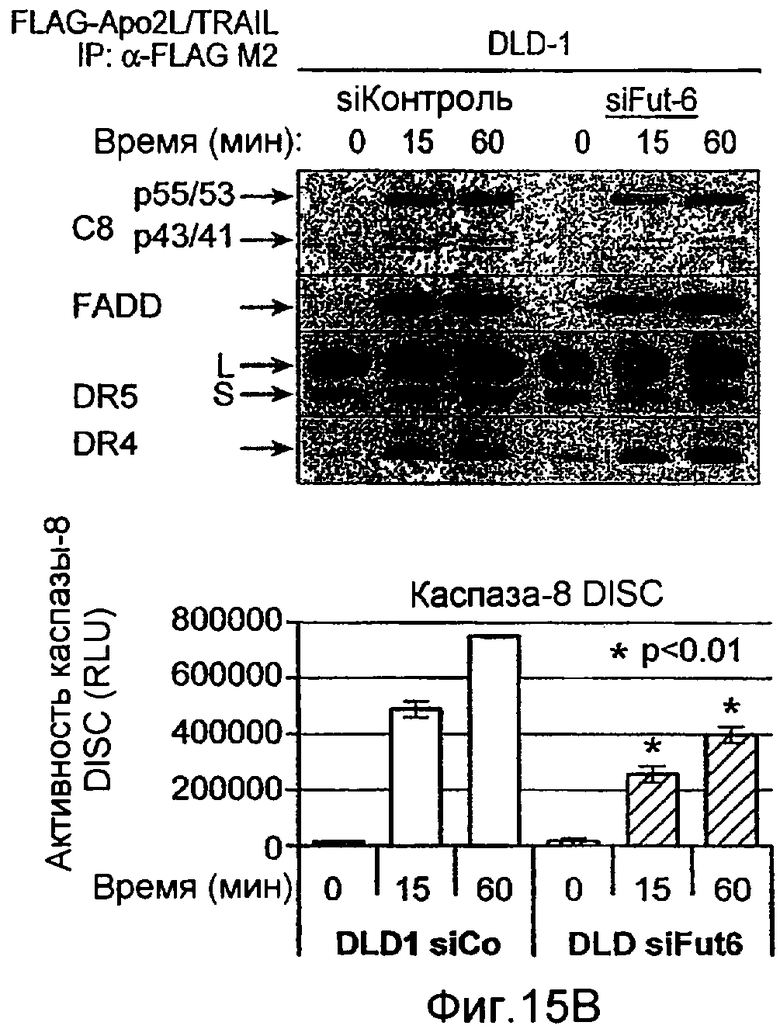

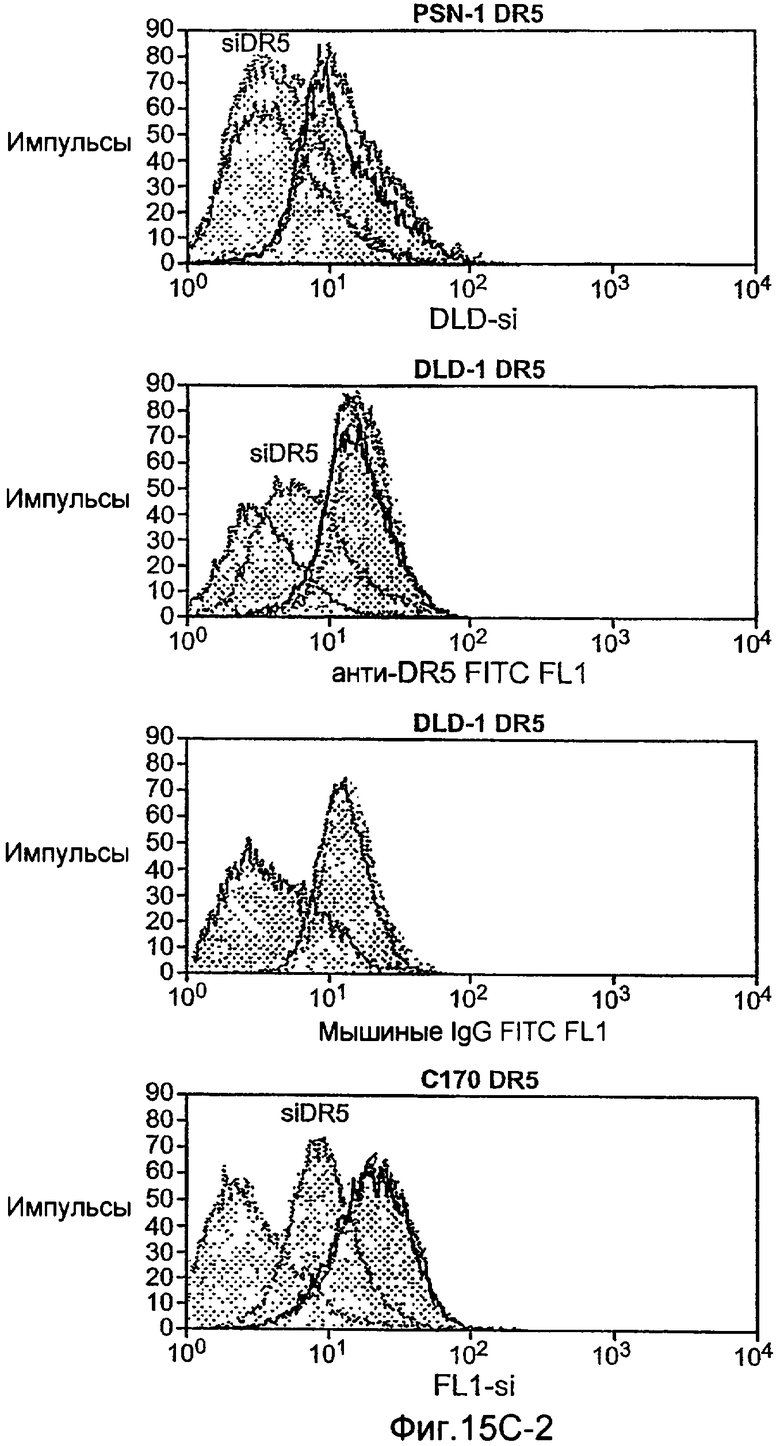
Изобретения касаются способов и анализов для исследования экспрессии одного или нескольких биомаркеров в образце ткани или клеток млекопитающего, а также представлены наборы и изделия для исследований. Выявление экспрессии молекул GalNac-T14 прогнозирует чувствительность или указывает на то, что образец ткани или клеток будет чувствительным к средствам, индуцирующим апоптоз, таким как Apo2L/TRAIL. Информация, полученная в результате анализа, направленного на выявление экспрессии GalNac-T14 в образце ткани или клеточном образце млекопитающего, может предоставить лечащему врачу сведения, которые могут быть использованы для определения оптимальной схемы лечения для пациентов, страдающих такими заболеваниями, как рак поджелудочной железы, лимфома, немелкоклеточный рак легких, рак толстой кишки, рак прямой кишки, меланома или хондросаркома. 5 н. и 17 з.п. ф-лы, 15 ил., 3 табл.
1. Способ прогнозирования чувствительности образца ткани или клеток злокачественной опухоли млекопитающего к полипептиду Apo2L/TRAIL, включающий стадию исследования образца ткани или клеток в иммуногистохимическом анализе для выявления экспрессии GalNac-T14, где указанный иммуногистохимический анализ проводят с помощью антител против GalNac-T14, которые связывают полипептид GalNac-T14 человека, содержащий аминокислоты 1-552 или аминокислоты 39-552, указанные на фигуре 4А (SEQ ID NO:12), и где экспрессия указанного GalNac-T14 является прогностическим признаком чувствительности указанного образца ткани или клеток к апоптоз-индуцирующей активности полипептида Apo2L/TRAIL.
2. Способ по п.1, в котором указанное антитело против GalNac-T14 представляет собой моноклональное антитело, которое связывает полипептид GalNac-T14 человека, содержащий аминокислоты 1-552, указанные на фигуре 4А (SEQ ID NO:12).
3. Способ по п.1, дополнительно включающий стадию исследования экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанном образце ткани или клеток.
4. Способ по п.1, в котором указанные клетки злокачественной опухоли представляют собой клетки или ткань рака поджелудочной железы, лимфомы, немелкоклеточного рака легких, рака толстой кишки, рака прямой кишки, меланомы или хондросаркомы.
5. Способ индукции апоптоза в образце ткани или клеток злокачественной опухоли млекопитающих, включающий последовательные стадии:
стадию исследования образца ткани или клеток в иммуногистохимическом анализе для выявления экспрессии полипептида GalNac-T14, где указанный иммуногистохимический анализ проводят с помощью антител против GalNac-T14, которые связывают полипептид GalNac-T14 человека, содержащий аминокислоты 1-552 или аминокислоты 39-552, указанные на фигуре 4А (SEQ ID NO:12), и где экспрессия указанного полипептида GalNac-T14 является прогностическим признаком чувствительности указанного образца ткани или клеток к апоптоз-индуцирующей активности полипептида Apo2L/TRAIL;
и, после обнаружения экспрессии указанного полипептида GalNac-T14, стадию воздействия на указанный образец ткани или клеток эффективным количеством полипептида Apo2L/TRAIL.
6. Способ по п.5, дополнительно включающий стадию исследования экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанном образце ткани или клеток.
7. Способ по п.5, в котором указанные клетки злокачественной опухоли представляют собой клетки или ткань рака поджелудочной железы, лимфомы, немелкоклеточного рака легких, рака толстой кишки, рака прямой кишки, меланомы или хондросаркомы.
8. Способ по п.5, в котором указанные клетки обрабатывают эффективным количеством полипептида Apo2L/TRAIL, содержащего аминокислоты 114-281, указанные на фиг.1 (SEQ ID NO:1).
9. Способ по п.5, в котором указанные клетки также подвергают воздействию химиотерапевтических(ого) средств(а) или лучевой терапии.
10. Способ по п.5, в котором указанные клетки также обрабатывают цитокином, цитотоксическим средством или ингибитором роста.
11. Способ по п.5, в котором указанный полипептид Apo2L/TRAIL связан с молекулой полиэтиленгликоля.
12. Способ по п.5, в котором указанный полипептид Apo2L/TRAIL содержит аминокислоты 41-281, указанные на фигуре 1 (SEQ ID NO:1), или биологически активный фрагмент.
13. Способ по п.5, в котором указанные клетки злокачественной опухоли являются клетками немелкоклеточного рака легких.
14. Способ по п.12, в котором указанный полипептид Apo2L/TRIAL состоит из аминокислот 114-281, указанных на фиг.1 (SEQ ID NO:1).
15. Применение антитела или полинуклеотида для выявления экспрессии белка GalNac-T14 или мРНК в образце ткани или клеток злокачественной опухоли млекопитающего для получения набора или изделия для прогнозирования чувствительности образца ткани или клеток млекопитающего к Apo2L/TRAIL, где экспрессия указанного GalNac-T14 является прогностическим признаком чувствительности указанного образца ткани или клеток к апоптоз-индуцирующей активности Apo2L/TRAIL.
16. Применение по п.15, при котором указанную экспрессию GalNac-T14 оценивают по экспрессии мРНК GalNac-T14.
17. Применение по п.15, при котором указанную экспрессию GalNac-T14 исследуют в иммуногистохимическом анализе.
18. Применение по п.15, при котором дополнительно проводят стадию исследования экспрессии рецепторов DR4, DR5, DcR1 или DcR2 в указанном образце ткани или клеток.
19. Применение по п.15, при котором указанные клетки злокачественной опухоли представляют собой клетки или ткань рака поджелудочной железы, лимфомы, немелкоклеточного рака легких, рака толстой кишки, рака прямой кишки, меланомы или хондросаркомы.
20. Применение набора или изделия в способе прогнозирования чувствительности образца ткани или клеток млекопитающего к полипептиду Apo2L/TRAIL, где набор или изделие содержит контейнер с этикеткой на указанном контейнере, причем в указанном контейнере находится композиция, которая включает первичное антитело, связывающееся с GalNac-T14, а на этикетке указанного контейнера указано, что композиция может использоваться для выявления GalNac-T14 по меньшей мере в одном типе клеток млекопитающих, и инструкции по применению антитела GalNac-T14 для оценки наличия GalNac-T14 по меньшей мере в одном типе клеток млекопитающих.
21. Применение набора или изделия по п.20, где указанное антитело является моноклональным антителом GalNac-T14.
22. Применение набора в способе прогнозирования чувствительности образца ткани или клеток млекопитающего к апоптозу, индуцированному полипептидом Apo2L/TRAIL, где набор содержит контейнер с этикеткой на указанном контейнере, причем в указанном контейнере находится композиция, которая включает полинуклеотид, который гибридизуется с комплементарным полинуклеотидом GalNac-T14 в жестких условиях, а на этикетке указанного контейнера указано, что композиция может использоваться для выявления GalNac-T14 по меньшей мере в одном типе клеток млекопитающих, и инструкции по применению полинуклеотида для оценки присутствия РНК или ДНК GalNac-T14 по меньшей мере в одном типе клеток млекопитающих.
| LeBLANC H.N | |||
| and ASHKENAZI A., Apo/TRAIL and its death and decoy receptors | |||
| Rev // Cell Death and Differentiation, 2003, 10, P.66-75 | |||
| CHEN Q | |||
| et al., Role ofApo2LTRAIL and Bcl-2-family proteins in apoptosis of multiple myeloma // Leuk | |||
| Lymphoma., 2003 44 (7), P.1209-1214 | |||
| FISHER U and SCHULZE-OSTHOFF K., Apoptosis-based therapies and drug targets | |||
| Rev | |||
| // Cell Death and Differentiation, 2005, 12, P.942-961. |

