Область техники
Настоящее изобретение относится к аптамеру против мидкина, способу применения его и подобного ему.
Уровень техники
Мидкин (в дальнейшем сокращаемый как "МК") является фактором роста/дифференцировки, который был впервые обнаружен как продукт гена, транзиентно экспрессирующийся в процессе индукции дифференцировки эмбриональных раковых клеток (EC) ретиноевой кислотой и представляющий собой полипептид с молекулярной массой 13 кДа, богатый основными аминокислотами и цистеином (см., например, непатентный документ 1 и непатентный документ 2).
Стерическая структура МК была определена при помощи ЯМР и опубликована (см., например, непатентный документ 3). По структурным характеристикам МК состоит в основном из двух доменов. В частности, МК состоит из фрагмента в N-концевой части, состоящего из аминокислотных остатков 1-52 (в дальнейшем называемый “N-концевой фрагмент”), фрагмента в C-концевой части, состоящего из аминокислотных остатков 62-121 (в дальнейшем называемый “C-концевой фрагмент”) и области петли, которая соединяет эти фрагменты (аминокислотные остатки 53-61). Связанный с областью за пределами каждого домена находится хвост, который богат основными аминокислотами. В молекуле МК каждый из N-концевого фрагмента и С-концевого фрагмента имеет стерическую структуру, состоящую главным образом из трех структур обратных бета-листов (в дальнейшем называемые "доменами"; домен, состоящий из аминокислотных остатков 15-52 в N-концевом фрагменте, называемый “N-доменом”, домен, состоящий из аминокислотных остатков 62-104 в C-концевом фрагменте, называемый “C-доменом”), и свободно перемещающиеся структуры, не предполагающие определенной структуры (в дальнейшем называемые "хвостами"; хвост, состоящий из аминокислотных остатков 1-14 в N-концевом фрагменте, называемый “N-хвостом”, и хвост, состоящий из аминокислотных остатков 105-121 в C-концевом фрагменте, называемый “C-хвостом”).
Известные рецепторы МК включают белок рецепторного типа тирозиновую фосфатазу ζ (PTPζ), LRP (белок, сходный с рецепторами липопротеинов низкой плотности), ALK (киназа анапластической лейкемии), интегрин и синдекан и т.п. МК является высоко положительно заряженным белком, содержащим большое количество основных аминокислот, таких как лизин (K) и аргинин (R). Он имеет гепарин-связывающий участок в C-домене, и, как известно, сильно связывается с отрицательно заряженными молекулами, такими как гепарин и сульфат хондроитина E. В результате мутагенезного анализа и ЯМР анализа считается, что кластер I, в конфигурации с K79, R81 и K102, и кластер II, в конфигурации с K86, K87 и R89, важны для связывания с гепарином. В то же время имеется публикация, что только кластер I необходим для связывания с сульфатом хондроитина E. В случае, когда R81 кластера I заменен на A, активность связывания с гепарином уменьшается. В результате наблюдается уменьшение активности связывания с PTPζ и подавление вызываемой МК элонгации нейритов и движения нервных клеток.
Ряд факторов роста, таких как фактор роста фибробластов (bFGF) и фактор роста васкулярных эндотелиальных клеток (VEGF), имеют гепарин-связывающий сайт. Предполагается, что эти факторы роста связываются с протеогликан сульфатом гепарина, внеклеточным матриксом, находятся в соответствующих положениях и высвобождаются, когда это необходимо. Известно также, что те же факторы связываются с сульфатом гепарина, экспрессирующимся в нервных клетках и васкулярных эндотелиальных клетках, и способствуют удлинению нейритов и усилению фибринолитической активности. В случае, когда чашка Петри была покрыта МК и на нее высевались нервные клетки эмбриона мыши, происходило удлинение нейритов. В этой ситуации расщепление нервных клеток гепаритиназой подавляет удлинение нейритов. В то же время, когда культивируются васкулярные эндотелиальные клетки и к ним добавляется МК, активность активатора плазминогена в клетках растет. В этом случае также расщепление клеток гепаритиназой подавляет увеличение активности плазминогена.
Считается, что МК связывается с PTPζ по двум сайтам. Один сайт включает в себя высокоафинную связь с сульфатом хондроитина (Кд=0,58 нМ). Эта связь исчезает после расщепления хондроитиназой. Другой сайт включает в себя связь с белком, которая, будучи низкоафинной связью, сохраняется после расщепления с хондроитиназой (Кд=3 нМ). МК способствует миграции эмбриональных нервных клеток, экспрессирующих PTPζ; обработка нервных клеток хондроитиназой ABC препятствует миграции. Остеобласт-подобные клетки UMR106 экспрессируют PTPζ, и, как известно, обладают МК-зависимой миграцией, которая подавляется обработкой хондроитиназой ABC. МК-зависимая миграция макрофага также подавляется обработкой хондроитиназой ABC, хондроитиназой B или гепариназой. Поскольку считается, что макрофаг не экспрессирует PTPζ, предполагается, что в процессе участвует другой рецептор.
Все отрицательно заряженные частицы не связываются с гепарин-связывающим сайтом МК. В случае, когда МК был иммобилизован амино-связыванием и подвергался анализу поверхностного плазмонного резонанса, полученные результаты показали, что сульфат хондроитина E и гепарин сильно связываются с МК, тогда как сульфат хондроитина A, B, C и D с МК не связывается.
Известно, что МК обладает широким спектром биологической активности. Например, известно, что в человеческих раковых клетках экспрессия МК увеличена. Такая увеличенная экспрессия наблюдалась в многочисленных видах рака, включая рак пищевода, рак щитовидной железы, рак мочевого пузыря, рак прямой кишки, рак желудка, рак поджелудочной железы, рак груди, рак печени, рак легких, рак молочной железы, нейробластому, глиобластому, рак матки, рак яичников и опухоль Вильмса (см., например, патентный документ 1 и непатентный документ 4). Также предполагается, что МК способствует выживанию и перемещению раковых клеток и облегчает неоваскуляризацию, что приводит к прогрессированию рака.
Также известно, что МК является одной из молекул, которые играют центральную роль в процессе развития воспаления. Например, известно, что образование новой интимы после повреждения кровеносного сосуда и начало нефрита при ишемическом повреждении являются ослабленными в нокаутных мышах, не имеющих гена МК. Также известно, что в модели ревматизма послеоперационная адгезия также значительно подавлена в мышах с нокаутом МК (см., например, патентный документ 2, патентный документ 3 и патентный документ 4). Таким образом, известно, что МК участвует в воспалительных заболеваниях, таких как артрит, аутоиммунная болезнь, ревматические артриты (ревматический артрит (РА), остеоартрит), рассеянный склероз, постоперационная адгезия, воспалительный колит, псориаз, волчанка, астма и функциональные отклонения нейтрофилов. Кроме того, известно, что МК способствует движению (миграции) клеток, таких как макрофаги и нейтрофилы, при воспалении. Поскольку это движение необходимо для развития воспаления, считается, что при отсутствии мидкина болезни, основанные на воспалении, скорее всего не будут возникать. (См., например, патентный документ 5).
В связи с тем, что уровень МК увеличивается в перитонеальной жидкости женщин с распространенным эндометриозом, а также потому, что МК стимулирует пролиферацию эндометриальных интерстициальных клеток в культуре, считается, что МК участвует в возникновении и распространении эндометриоза (см., например, патентный документ 6).
Кроме того, в связи с действием на утолщение сосудистой интимы считается, что МК участвует в сосудистых обструктивных заболеваниях, таких как рестеноз после операции сосудистой реконструкции, обструктивное заболевание сердечных коронарных артериальных сосудов, обструктивное заболевание сосудов мозга, обструктивное заболевание сосудов почек, обструктивное заболевание периферических сосудов, артериосклероз и мозговой инфаркт (см., например, патентный документ 2).
Известно, что клеточная миграция важна для механизмов проникновения/метастаз раковых клеток, утолщения интимы в артериосклеротических очагах, при неоваскуляризации и т.д. Также известно, что миграция клеток при воспалении тесно связана с сердечно-сосудистыми заболеваниями, такими как стенокардия, инфаркт миокарда, мозговой инфаркт, мозговое кровоизлияние и гипертония.
Плейотропин (PTN или HB-GAM) является единственным белком в семействе МК, имеющим приблизительно 50% гомологии с МК. И МК, и плейотропин являются белками, содержащими большое количество цистеина и основных аминокислот. Все 10 цистеиновых аминокислот являются консервативными в МК и PTN, и структурно оба могут быть разделены на N-домен и C-домен. В результате ЯМР анализа известно, что эти две молекулы имеют очень похожие трехмерные структуры. Каждый домен состоит из трех бета листов, связанных через подвижную линкерную область. Аминокислоты K79, R81 и K102, которые предположительно важны для связывания с сульфатом хондроитина и гепарином, являются консервативными в этих двух белках. K79 и R81 находятся на одном и том же бета листе, в то время как K102 расположена на другом бета листе. Когда МК и PTN образуют пространственную структуру, эти основные аминокислоты располагаются вблизи поверхности белка.
В последние годы внимание привлекали применения РНК аптамеров в терапевтических лекарствах, диагностических реагентах и реактивах для анализа; некоторые РНК аптамеры уже находятся на стадии клинических испытаний или в практическом применении. В декабре 2004 г. первый в мире препарат РНК аптамера, Macugen, был одобрен в качестве терапевтического препарата для возрастной макулярной дегенерации в США. РНК аптамер обозначает РНК, которая специфично связывается с веществом-мишенью, таким как белок, и может быть получен с использованием способа SELEX (Системная Эволюция Лигандов Экспоненциальным Обогащением) (непатентные документы 5, 6). Способ SELEX является способом, при котором РНК, которая специфично связывается с веществом-мишенью, отбирается приблизительно из набора 1014 РНК, имеющих различные нуклеотидные последовательности. Используемая РНК имеет структуру, в которой случайная последовательность из приблизительно 40 остатков находится между последовательностями праймеров. Этому набору РНК давали возможность связаться с веществом-мишенью, и только та РНК, которая связалась с веществом-мишенью, выделялась с использованием фильтра и т.д. Полученная РНК амплифицировалась ПЦР с обратной транскрипцией, и полученный продукт использовался как матрица для следующего цикла. После повторения этой операции приблизительно 10 раз иногда можно получить РНК аптамер, который специфически связывается с веществом-мишенью.
[патентный документ 1] JP-A-6-172218
[патентный документ 2] WO2000/10608
[патентный документ 3] WO2004/078210
[патентный документ 4] WO2004/085642
[патентный документ 5] WO1999/03493
[патентный документ 6] WO2006/016571
[непатентный документ 1] Kadomatsu, K. et al., Biochem. Biophys. Res. Commun., 151:p.1312-1318
[непатентный документ 2] Tomokura, M. et al.: J.Biol. Chem, 265: p.10765-10770
[непатентный документ 3] Iwasaki, W. et al., (1997) EMBO J. 16, p.6936-6946
[непатентный документ 4] Muramatsu, T., (2002) J. Biochem. 132, p.359-371
[непатентный документ 5] Ellington et al., (1990) Nature, 346, 818-822
[непатентный документ 6] Tuerk et al., (1990) Science, 249, 505-510
Раскрытие изобретения
Задачи, решаемые изобретением
Настоящее изобретение направлено на обеспечение аптамера для мидкина и способа применения его и подобного ему.
Способы решения указанных задач
Авторам настоящего изобретения в результате активных исследований, направленных на решение описанных выше проблем, удалось получить аптамер хорошего качества для мидкина, что привело к разработке настоящего изобретения.
Соответственно, настоящее изобретение обеспечивает следующее:
(1) аптамер, обладающий ингибирующей активностью против мидкина,
(2) аптамер по п. (1), который не обладает ингибирующей активностью против плейотропина,
(3) аптамер по п. (1), обладающий активностью связывания с N-концевым фрагментом мидкина,
(4) аптамер по п. (1), обладающий активностью связывания с C-концевым фрагментом мидкина,
(5) аптамер по п. (2), обладающий активностью связывания с N-концевым фрагментом мидкина,
(6) аптамер по п. (2), обладающий активностью связывания с C-концевым фрагментом мидкина,
(7) аптамер, который проявляет ингибирующую активность против мидкина посредством ингибирования связывания мидкина и PTPζ,
(8) аптамер по п. (1), который соответствует либо (a), или (b) из описанного ниже:
(a) аптамер, включающий нуклеотидные последовательности, выбираемые из SEQ ID NO: 1-70 (с условием, что урацил может быть тимином), в которых нуклеотиды, содержащиеся в аптамере, такие, что
(i) в 2'-положениях пиримидиновых нуклеотидов, одинаковых или различных, находятся атомы фтора или замещены на атомы или группы, выбираемые из группы, состоящей из атомов водорода, гидроксильных групп и метоксильных групп, и
(ii) в 2'-положениях пуриновых нуклеотидов, одинаковых или различных, находятся гидроксильные группы или замещены на атомы или группы, выбираемые из группы, состоящей из атомов водорода, метоксильных групп и атомов фтора;
(b) аптамер, включающий в себя нуклеотидные последовательности, выбираемые из SEQ ID NO: 1-70 (с условием, что урацил может быть тимином), в которых один или несколько нуклеотидов замещены, удалены, вставлены или добавлены, в которых нуклеотиды, содержащиеся в аптамере, такие, что
(i) в 2'-положениях пиримидиновых нуклеотидов, одинаковых или различных, находятся атомы фтора или замещены на атомы или группы, выбираемые из группы, состоящей из атомов водорода, гидроксильных групп и метоксильных групп, и
(ii) в 2'-положениях пуриновых нуклеотидов, одинаковых или различных, находятся гидроксильные группы или замещены на атомы или группы, выбираемые из группы, состоящей из атомов водорода, метоксильных групп и атомов фтора;
(9) аптамер по любому из пп. (1)-(8), в которых нуклеотид, содержащийся в аптамере, модифицирован,
(10) комплекс, включающий аптамер по любому из пп. (1)-(9) и функциональное вещество,
(11) комплекс по п. (10), в котором функциональным веществом является вещество аффинности, вещество для мечения, фермент, носитель для доставки лекарственных средств или лекарственное средство,
(12) фармацевтический препарат, включающий аптамер по любому из пп. (1)-(9) или комплекс по п. (10) или (11),
(13) ингибитор миграции клеток, включающий аптамер по любому из пп. (1)-(9) или комплекс по п. (10) или (11),
(14) реактив для диагностики, включающий аптамер по любому из пп. (1)-(9) или комплекс по п. (10) или (11),
(15) агент для мечения, включающий аптамер по любому из пп. (1)-(9) или комплекс по п. (10) или (11), и
(16) способ детектирования аптамера по любому из пп. (1)-(9) или комплекс по п. (10) или (11).
Применение изобретения
Аптамер или комплекс настоящего изобретения может использоваться как фармацевтические препараты или реактивы, такие как диагностические реактивы для различных болезней, таких как аутоиммунная болезнь, рак, послеоперационная адгезия и эндометриоз. Аптамер или комплекс настоящего изобретения может также использоваться в очистке и концентрировании МК, и детектировании и количественном определении МК.
Краткое описание фигур
Фиг.1A показывает одну из двух вторичных структур РНК, представленных SEQ ID NO:1, предсказанную программой MFOLD.
Фиг.1B показывает вторую вторичную структуру РНК, представленную SEQ ID NO:1, предсказанную программой MFOLD.
Фиг.2A показывает одну из двух вторичных структур РНК, представленную SEQ ID NO:2, предсказанную программой MFOLD, в которой часть, заключенная в квадрат, показывает консенсусную область.
Фиг.2B показывает другую вторичную структуру РНК, представленную SEQ ID NO:2, предсказанную программой MFOLD, в которой часть, заключенная в квадрат, показывает консенсусную область.
Фиг.3A показывает одну из двух вторичных структур РНК, представленной SEQ ID NO:3, предсказанную программой MFOLD.
Фиг.3B показывает другую вторичную структуру РНК, представленной SEQ ID NO:3, предсказанную программой MFOLD, в которой часть, заключенная в квадрат, показывает консенсусную область.
Фиг.4 показывает вторичную структуру РНК, представленной SEQ ID NO:4, предсказанную программой MFOLD, в которой часть, заключенная в квадрат, показывает консенсусную область.
Фиг.5 показывает вторичную структуру РНК, представленной SEQ ID NO:5, предсказанную программой MFOLD.
Фиг.6 показывает взаимодействия между РНК, представленной SEQ ID NO:5 и мидкином, и между РНК и IgG1 человека (сенсограмма получена с использованием BIAcore2000).
Фиг.7 показывает взаимодействие между РНК, представленной SEQ ID NO:4 и мидкином (сенсограмма получена с использованием BIAcore2000).
Фиг.8 показывает вторичную структуру РНК, представленной SEQ ID NO:20, предсказанную программой MFOLD.
Фиг.9 показывает вторичную структуру РНК, представленной SEQ ID NO:61, предсказанную программой MFOLD.
Наилучший способ осуществления изобретения
Настоящее изобретение обеспечивает аптамер, обладающий активностью связывания с мидкином (МК). Аптамеры настоящего изобретения обладают активностью, ингибирующей МК.
Аптамер обозначает молекулу нуклеиновой кислоты, имеющую аффинность связывания со специфической молекулой-мишенью. Кроме того, аптамер может также ингибировать активность специфической молекулы-мишени, связываясь со специфической молекулой-мишенью. Аптамер настоящего изобретения может быть РНК, ДНК, модифицированной нуклеиновой кислотой или их смесью. Аптамер настоящего изобретения может также находиться в линейной или кольцевой форме.
Ингибирующая активность против МК означает ингибирование любой биологической активности МК. В качестве примера биологических активностей МК можно упомянуть миграционную активность клеток (например, макрофагов, нейтрофилов, эозинофилов, васкулярных гладкомышечных клеток, опухолевых клеток, остеобластов, нервных клеток и их прогениторных клеток) (Takada et al., 1997, J. Biochem. 122, 453-458, Horiba et al., 2000, J. Clin. Invest. 105, 489-495, Maeda et al., 1999, J. Biol. Chem. 274, 12474-12479, Qi et al., 2001, J. Biol. Chem. 276, 15868-15875), активности, способствующие пролиферации и дифференцировке клеток (например, опухолевых клеток, фибробластов, кератиноцитов, нервных клеток, хондроцитов и их прогениторных клеток) (Muramatsu и Muramatsu, 1991, Biochem. Biophys. Res. Commun. 177, 652-658, Muramatsu et al., 1993, Dev. Biol. 159, 392-402, Takei et al, 2001., Cancer Res. 61, 8486-8491), ингибиторные активности против пролиферации и функций регуляторных T клеток, активности, способствующие элонгации нейритов нервных клеток, ингибиторные активности против апоптоза клеток (например, опухолевых клеток, нервных клеток), активности, вызывающие нейроваскуляризацию (например, опухолевых клеток), активности, вызывающие формирование синапса для миобластов, активности, способствующие действиям фибринолитической системы для сосудистых эндотелиальных клеток, активности, вызывающие продукцию IL-8 для васкулярных гладкомышечных клеток и т.п. Таким образом, в качестве примеров ингибиторных активностей против МК можно упомянуть ингибиторные активности против этих активностей.
Аптамер настоящего изобретения может обладать ингибиторными активностями против МК, полученного из любых видов млекопитающих. В качестве примера таких млекопитающих можно упомянуть приматов (например, люди, обезьяны), грызунов (например, мыши, крысы, морские свинки), а также домашних животных, одомашненных животных и рабочих животных (например, собаки, кошки, лошади, быки, козы, овцы, свиньи).
Аптамеры настоящего изобретения не имеют особенных ограничений при условии, что они способны связываться с произвольно выбранной частью МК для ингибирования соответствующей активности; например, связываясь с N-концевым фрагментом или C-концевым фрагментом МК, аптамеры настоящего изобретения способны к ингибирующим активностям МК. Аминокислотная последовательность человеческого МК содержится в GenBank под номером BC011704, при этом секреторный белок образуется 121 аминокислотами от лизина 23 до аспарагиновой кислоты 143. Как правило, лизин 23 обозначается аминокислотой в положении 1. Человеческий МК состоит из N-концевого фрагмента, состоящего из аминокислот 1-52, C-концевого фрагмента, состоящего из аминокислот 62-121, и области петли, которая соединяет эти фрагменты, но граница N-концевого фрагмента и фрагмента C-концевого фрагмента может быть любой частью петли МК (53-61) и не может быть точно определена.
Длина аптамера настоящего изобретения не ограничена и обычно может быть приблизительно от 15 до 200 нуклеотидов, и может быть, например, не более чем приблизительно 100 нуклеотидов, предпочтительно не более чем приблизительно 80 нуклеотидов, более предпочтительно не более чем приблизительно 60 нуклеотидов, наиболее предпочтительно не более чем приблизительно 45 нуклеотидов. Длина аптамера настоящего изобретения может быть, например, не менее чем приблизительно 18, 20 или 25 нуклеотидов. Если общее количество нуклеотидов является меньшим, химический синтез и массовое производство будет легче, и это дает значительное преимущество с точки зрения стоимости. Также считается, что химическая модификация аптамера является легкой, его стабильность высокой и токсичность низкой.
Каждый из нуклеотидов, содержащихся в аптамере настоящего изобретения, одинаковый или различный, может быть нуклеотидом, включающим гидроксильную группу в 2'-положении рибозы (например, рибозы нуклеотида пиримидина) (то есть незамещенного нуклеотида) или нуклеотидом, имеющим гидроксильную группу, замещенную произвольно выбранным атомом или группой в 2'-положении рибозы. В качестве примера такого произвольно выбранного атома или группы может быть упомянут нуклеотид с замещением на атом водорода, атом фтора или алкил группу (например, -O-Me группу), алкил группу (например, -O-CHO группу), или аминогруппу (например, -NH2 группу). Аптамер настоящего изобретения может также быть аптамером, в котором, по меньшей мере один тип (например, 1, 2, 3 или 4 типа) нуклеотида включает нуклеотид, содержащий в 2'-положениях рибозы гидроксильную группу, или вышеописанные произвольно выбранные атом или группу, например, по меньшей мере два вида (например, 2, 3 или 4 вида) групп, выбираемых из группы, состоящей из атома водорода, атома фтора, гидроксильной группы и -O-Me группы. В аптамерах настоящего изобретения все нуклеотиды могут быть нуклеотидами, содержащими во 2'-положении рибозы гидроксильную группу или произвольно выбранный атом или группу, описанную выше, например, группу, выбираемую из группы, состоящей из атома водорода, атома фтора, гидроксильной группы и -O-Me группы.
Пример аптамера настоящего изобретения может иметь потенциальную вторичную структуру, включающую одну или более частей, выбираемых из группы, состоящей из одноцепочечных частей (например, gggagaggaac), первых стволовых частей (например, gacg и их комплементарных цепей), внутренних областей петли (например, aggagua и gg), вторых стволовых частей (например, gcc и их комплементарных цепей) и внутренних областей петли (например, ggaaagaa). Другой пример аптамера настоящего изобретения может иметь потенциальную вторичную структуру, включающую одну или более частей, выбираемых из группы, состоящей из одноцепочечных частей (например, gggaaggaggaa), первых стволовых частей (например, gugcac и их комплементарных цепей), внутренних областей петли (например, ag и gg), вторых стволовых частей (например, gg и их комплементарных цепей) и внутренних областей петли (например, guuggug).
Использующийся здесь термин “потенциальная вторичная структура” относится к вторичной структуре, способной к образованию при физиологических условиях; например, присутствует ли или нет потенциальная вторичная структура, может быть определено с использованием программ предсказания структуры, описанных в Примерах. Стволовая область относится к части, в которой двойная цепь образуется парой оснований в двух или более последовательных нуклеотидах (например, G-C, A-U, A-T). Часть внутренней петли обозначает нестволовую область, образованную между двумя различными стволовыми областями. Область шпильки петли относится к частичной структуре, образованной одной стволовой областью, которая является областью петли, образованной на противоположной стороне к 5' концу и 3' концу цепи аптамера. Одноцепочечная область относится к конечной части полинуклеотидной цепи и является областью, которая не соответствует вышеупомянутой стволовой области, внутренней области петли или области шпильки петли.
Аптамер настоящего изобретения может также обладать способностью связывания с N-концевым фрагментом и/или C-концевым фрагментом МК. Аптамер, представленный SEQ ID NO:39, и его измененные формы, как гепарин и сульфат E хондроитина, проявляют высокую активность связывания с C-концевым фрагментом. Предполагается, что гепарин связывается с C-концевым фрагментом в областях кластера I и кластера II. Предполагается, что сульфат хондроитина E связывается с C-концевым фрагментом в области кластера I. Известно, что МК взаимодействует с PTPς, который включает сульфат хондроитина как составляющую молекулу. PTPς экспрессируется в эмбриональных нервных клетках и клетках остеобластной структуры, и в присутствии МК происходит стимуляция миграции этих клеток. В настоящем изобретении описываются аптамер, способный к связыванию с C-концевым фрагментом и ингибирующий миграцию клеток, и другой аптамер, который связывается главным образом с N-концевым фрагментом и ингибирует миграцию клеток.
Аптамеры настоящего изобретения также способны к ингибированию активности МК (например, активности МК в отношении миграции клеток) и могут обладать свойством неспособности ингибировать активность PTN (например, активность PTN в отношении клеточной миграции). PTN является единственным белком в семействе, имеющим гомологию с МК 50%, они имеют очень схожие трехмерные структуры и аминокислоты, которые важны для связывания с сульфатом хондроитина и гепарина, являются консервативными.
Аптамер настоящего изобретения может также быть (a) аптамером, содержащим нуклеотидную последовательность, выбираемую из одного из SEQ ID NO:1-70 (но где урацил может быть тимином), (b) аптамером, содержащим нуклеотидную последовательность, выбираемую из одного из SEQ ID NO:1-70 (но где урацил может быть тимином), имеющим один или несколько нуклеотидов замещенных, удаленных, вставленных или добавленных, или (c) конъюгатом, выбираемым из группы, состоящей из конъюгатов из множества вышеупомянутых единиц (a), конъюгатом из множества вышеупомянутых единиц (b) и конъюгатов из множества вышеупомянутых единиц (a) и (b). В вышеупомянутом (b) количество замещенных, удаленных, вставленных или добавленных нуклеотидов не ограничено специально при условии, что это ограничивается несколькими нуклеотидами, и число нуклеотидов может быть, например, не более чем приблизительно 30, предпочтительно не более чем приблизительно 20, более предпочтительно не более чем приблизительно 10, более предпочтительно не более чем 5, наиболее предпочтительно 4, 3, 2 или 1. В вышеупомянутом (c) конъюгирование может быть достигнуто тандемным связыванием. В этом конъюгировании может использоваться линкер. В качестве линкера можно отметить нуклеотидные цепи (например, от 1 до приблизительно 20 нуклеотидов) и ненуклеотидные цепи (например, линкер -(CH2)n-, линкер -(CH2CH2O)n-, линкер гексаэтиленгликоля, TEG линкер, пептид-содержащий линкер, линкер, содержащий -S-S- связи, линкер, содержащий -CONH- связи, линкер, содержащий -OPO3- связи. Множество, упомянутое в вышеописанном множестве конъюгатов, не является специально ограниченным при условии, что их два или больше, и множество может быть, например, 2, 3 или 4. Каждый из нуклеотидов в упомянутых выше (a)-(c), одинаковый или различный, может быть нуклеотидом, включающим гидроксильную группу в 2'-положении рибозы, или нуклеотидом, имеющим замещенную гидроксильную группу на произвольно выбранную группу (например, атом водорода, атом фтора или группу -O-Me) в 2'-положении рибозы (например, рибозы нуклеотида пиримидина).
В одном конкретном аспекте аптамеры настоящего изобретения можно классифицировать приблизительно на три типа в соответствии своим структурам. Первый тип аптамера - это аптамер, состоящий из нуклеотидной последовательности, представленной SEQ ID NO:61, или его мутированная форма. Аптамер, состоящий из нуклеотидной последовательности, представленной SEQ ID NO:61, когда его вторичная структура предсказана программой MFOLD, имеет потенциальную вторичную структуру, показанную на фиг.9, формируемую одноцепочечной частью, первой стволовой частью, внутренней областью петли, второй стволовой частью и областью шпильки петли. В этом аптамере замена, удаление, вставка и/или добавление нескольких нуклеотидов являются приемлемыми в одноцепочечной части, первой стволовой части, внутренней области петли, второй стволовой части и области шпильки петли. Например, в этом аптамере допускаются вставка нескольких нуклеотидов в одноцепочечную часть, вставка нескольких нуклеотидов в первую стволовую часть и добавление нескольких нуклеотидов к 3' концу одноцепочечной части (например, SEQ ID NO:5). Такой аптамер связывается сильнее с N-концевым фрагментом, чем с C-концевым фрагментом МК.
Вторым видом аптамера является аптамер, состоящий из нуклеотидной последовательности, представленный SEQ ID NO:20, или его мутантная форма. Аптамер, состоящий из нуклеотидной последовательности, представленной SEQ ID NO:20, когда его вторичная структура предсказана программой MFOLD, имеет потенциальную вторичную структуру, показанную на Фиг.8, формируемую одноцепочечной частью, первой стволовой частью, внутренней областью петли, второй стволовой частью и областью шпильки петли. В этом аптамере замена, удаление, вставка и/или добавление нескольких нуклеотидов являются приемлемыми в одноцепочечной части, первой стволовой части, внутренней области петли, второй стволовой части и области шпильки петли. Например, в этом аптамере допускаются вставка нескольких нуклеотидов в одноцепочечную часть, вставка нескольких нуклеотидов в первую стволовую часть и добавление нескольких нуклеотидов к 3' концу одноцепочечной части (например, SEQ ID NO:4). Такой аптамер почти не проявляет связывания к N-концевому фрагменту МК и связывается сильно с C-концевым фрагментом.
Третьим видом аптамера является аптамер, состоящий из нуклеотидной последовательности, представленной SEQ ID NO:1, или его мутированная форма.
Аптамер настоящего изобретения может быть аптамером, в котором остаток сахара (например, рибоза) каждого нуклеотида является модифицированным для увеличения активности связывания, стабильности, доставки лекарственных препаратов МК и т.п. В качестве примеров модифицированного участка в остатке сахара можно отметить участок, имеющий атом кислорода во 2'-положении, 3'-положении и/или 4'-положении остатка сахара, замещенные на другие атомы. В качестве примеров модификации можно отметить фторирование, O-алкилирование (например, O-метилирование, O-этилирование), O-арилирование, S-алкилирование (например, S-метилирование, S-этилирование), S-арилирование и аминирование (например, -NH2). Такие изменения в остатке сахара могут выполняться известным способом (см., например, Sproat et al., (1991) Nucle. Acid. Res. 19, 733-738; Cotton et al., (1991) Nucl. Acid. Res. 19, 2629-2635; Hobbs et al., (1973) Biochemistry 12, 5138-5145).
Аптамер настоящего изобретения может также иметь измененную (например, с помощью химического замещения) основу нуклеиновой кислоты (например, пурин или пиримидин) для увеличения активности связывания МК и т.п. В качестве примеров таких изменений можно упомянуть изменение пиримидина в 5 положении, изменение пурина в 6 и/или 8 положениях, изменение с экстрациклическим амином, замещение на 4-тиуридин, замена на 5-бромо или 5-иодоурацил. Фосфатная группа, содержащаяся в аптамере настоящего изобретения, может быть изменена для придания устойчивости к нуклеазам и гидролизу. Например, P(O)O группа может быть замещена на P(O)S (тиоат), P(S)S (дитиоат), P(O)NR2 (амидат), P(O)R, R(O)OR', CO или CH2 (формацетал) или 3'-амин (-NH-CH2-CH2-) (в котором каждая группа R или R' независимо является H или замещенным или незамещенным алкилом (например, метилом, этилом)).
Группой связывания может, например, быть -O-,-N- или -S-, и нуклеотиды могут связываться с соседним нуклеотидом посредством этих групп связывания.
Изменения могут также включать такие изменения, как кэппинг в 3' и 5'-положениях.
Изменение может также осуществляться добавлением к концу полиэтиленгликоля, аминокислоты, пептида, инвертированного dT, нуклеиновой кислоты, нуклеозидов, миристоила, литоколик-олеила, докозанила, лауроила, стеароила, пальмитоила, олеоила, линолеоила, других липидов, стероидов, холестерина, кофеина, витаминов, пигментов, флуоресцентных веществ, противораковых агентов, токсина, ферментов, радиоактивных веществ, биотина и т.п. Для таких изменений см., например, патенты США 5660985 и 5756703.
Аптамер настоящего изобретения может быть химически синтезирован по описаниям, представленным здесь, и по способу, известному специалисту. Аптамер связывается с веществом-мишенью большим разнообразием способов связывания, таких как ионные связи, основанные на отрицательном заряде фосфатных групп, гидрофобные связи и водородные связи, основанные на рибозе, и водородные связи и связи сжатия, основанные на основаниях нуклеиновой кислоты. В частности, ионные связи, основанные на отрицательном заряде фосфатных групп, которые присутствуют в таком же количестве, как и количество составляющих нуклеотидов, являются сильными связями и связываются с лизином и аргинином, присутствующими на поверхности положительного заряда белка. Поэтому основания нуклеиновой кислоты, не участвующие в прямом связывании с веществом-мишенью, могут быть заменены. В частности, в связи с тем, что в области стволовой структуры пары оснований уже сформированы и направлены внутрь двойной спирали, маловероятно, что основания нуклеиновой кислоты непосредственно связываются с веществом-мишенью. Поэтому часто, даже когда одна пара оснований замещается на другую пару оснований, активность аптамера не уменьшается. В структурах, в которых пары оснований не сформированы (таких как структуры петли), замена оснований возможна при условии, что основания нуклеиновой кислоты не участвуют в прямом связывании с молекулой-мишенью. Что касается модификаций 2'-положений рибозы, функциональная группа во 2'-положении рибозы иногда непосредственно взаимодействует с молекулой-мишенью, но во многих случаях это не является важным, и она может быть замещена другой модифицированной молекулой. Таким образом, аптамер часто сохраняет свою активность при условии, что не изменена или не удалена функциональная группа, участвующая в прямом связывании с молекулой-мишенью. Также важно, чтобы полная стерическая структура не сильно менялась.
Аптамер может быть получен с использованием способа SELEX или его улучшенной модификации (например, Ellington et al., (1990) Nature, 346, 818-822; Tuerk et al., (1990) Nature, 249, 505-510). В способе SELEX при увеличении число раундов или использовании конкурирующего вещества концентрируется и отбирается аптамер, проявляющий более сильные силы связывания к веществу-мишени. Следовательно, регулируя число раундов SELEX и/или изменяя условия конкуренции, в ряде случаев могут быть получены аптамеры с различными силами связывания, аптамеры с различными способами связывания и аптамеры с одинаковыми силами связывания и способами связывания, но различными последовательностями оснований. Способ SELEX включает процесс ПЦР амплификации; посредством осуществления мутации ионами марганца и т.п. возможно осуществить SELEX с более высоким многообразием.
Аптамеры, полученные SELEX, являются нуклеиновыми кислотами, которые проявляют высокую аффинность к веществу-мишени, и это не означает его связывания с активным участком вещества-мишени. Поэтому аптамеры, полученные SELEX, не всегда действуют на функцию вещества-мишени. У МК имеется богатая лизином область в области хвоста каждого из N-концов и C-концов; предполагается, что нуклеиновая кислота связывается с ними неспецифически. Предполагается, что эта часть хвоста не является важной для связывания с гепарином или хондроитином сульфатом. Не легко получить аптамер, который эффективно ингибирует активность МК в таких условиях. Фактически в настоящем изобретении были исследованы активности, ингибирующие миграцию клеток, 23 типов аптамеров и только 4 типа аптамеров сохраняли не менее 50% своей активности.
Отобранные таким образом активные аптамеры могут быть наделены еще большей активностью с помощью оптимизированного способа SELEX. Оптимизированный способ SELEX относится к способу, в котором SELEX выполняется еще раз после получения матрицы, в которой аптамер с определенной фиксированной последовательностью частично изменен с тем, чтобы включать случайные последовательности или матрицу с включениями, содержащими приблизительно 10-30% случайных последовательностей.
Аптамер, полученный способом SELEX, имеет приблизительно 80 нуклеотидов в длину, и это создает трудности при его получении в качестве фармацевтического продукта. Следовательно, необходимо повторить действия «попытки-и-ошибки» с тем, чтобы уменьшить аптамер приблизительно до 50 нуклеотидов в длину или меньше для облегчения химического синтеза.
В зависимости от дизайна праймеров для аптамера, полученного способом SELEX, изменяется легкость последующих операций минимизации. Если дизайн праймеров не проведен успешно, последующее совершенствование становится невозможным, даже если активный аптамер будет отобран способом SELEX.
Аптамеры легко изменяемы, потому что они допускают химический синтез. Для аптамеров возможно с некоторой точностью предсказать, какой нуклеотид можно заменить или удалить, а также где вставить новый нуклеотид, с помощью программы MFOLD предсказания вторичной структуры или с помощью предсказания стерической структуры рентгеновским анализом или ЯМР анализом. Аптамер с предсказанной новой последовательностью может легко быть химически синтезирован, а также с использованием существующей системы анализа может быть определено, сохраняет ли аптамер свою активность.
Если область, важная для связывания полученного аптамера с веществом-мишенью, определена несколькими последовательными действиями «попытки-и-ошибки», как описано выше, активность остается неизменной во многих случаях, даже когда к обоим концам последовательности добавляется новая последовательность. Длина новой последовательности не является специально ограниченной.
Модификации, как и последовательности, дают широкие возможности дизайна или изменений.
Как указано выше, аптамеры дают широкие возможности дизайна или изменений. Настоящее изобретение также обеспечивает способ производства аптамера, который позволяет широкие возможности дизайна или изменения аптамера, включая конкретную последовательность (например, последовательность, соответствующую части, выбираемой из числа стволовых частей, внутренних областей петли, областей шпильки петли и одноцепочечных частей; в дальнейшем установленные последовательности обозначаются сокращениями, как требуется).
Например, способ полученя такого аптамера включает производство аптамера, содержащего фиксированную последовательность с использованием единственного вида молекулы нуклеиновой кислоты или множества видов молекул нуклеиновой кислоты (например, библиотеки молекул нуклеиновой кислоты с различным количеством для “a” или “b”), состоящих из нуклеотидной последовательности, представленной формулой:
Сиквенс праймера (i) - (N)a - фиксированная последовательность - (N)b - Сиквенс праймера (ii),
где (N)a обозначает нуклеотидную цепь, состоящую из “a” единиц N; (N)b обозначает нуклеотидную цепь, состоящую из “b” единиц N; каждая из единиц N, одинаковая или различная, является нуклеотидом, выбираемым из группы, состоящей из A, G, C, U и T (предпочтительно, A, G, C и U). Каждая единица из “a” и “b”, одинаковая или различная, может быть произвольно выбранным числом и может быть, например, от 1 до приблизительно 100, предпочтительно от 1 до приблизительно 50, более предпочтительно от 1 до приблизительно 30, еще более предпочтительно от 1 до приблизительно 20 или от 1 до приблизительно 10), и пары праймеров соответствуют сиквенсам праймеров (i) и (ii) соответственно.
Настоящее изобретение также обеспечивает комплекс, включающий аптамер изобретения и функциональное вещество, связанное с ним. Связи между аптамером и функциональным веществом в комплексе настоящего изобретения могут быть ковалентными связями или нековалентными связями. Комплекс настоящего изобретения может быть комплексом, в котором аптамер изобретения и один или более (например, 2 или 3) функциональных веществ одинакового или различных видов связаны между собой. Функциональное вещество не ограничено специально при условии, что оно придает новую определенную функцию аптамеру настоящего изобретения или способно изменить (например, улучшить) определенную особенность, которой может обладать аптамер настоящего изобретения. В качестве примеров функционального вещества можно упомянуть белки, пептиды, аминокислоты, липиды, сахара, моносахариды, полинуклеотиды и нуклеотиды. В качестве примеров функционального вещества можно упомянуть вещества с аффинностью (например, биотин, стрептавидин, полинуклеотиды, обладающие аффинностью к комплементарным последовательностям-мишеням, антитела, глутатион Сефарозу, гистидин), вещества для мечения (например, флуоресцентные вещества, люминесцентные вещества, радиоизотопы), ферменты (например, пероксидазу хрена, щелочную фосфатазу), наполнители для доставки лекарственных средств (например, липосомы, микросферы, пептиды, полиэтиленгликоли), лекарственные средства (например, средства, используемые в направленной терапии, такие как каликеамицин и дуокармицин; аналоги изотиациана азота, такие как циклофосфамид, мелфалан, ифосфамид или трофосфамид; этиленимины, такие как тиотепа; нитрозомочевины, такие как курмустин; алкилирующие агенты, такие как темозоломид или декарбазин; метаболические антагонисты, подобные фолату, такие как метотрексат или ралтитрексед; аналоги пурина, такие как тиогуанин, кладрибин или флударабин; аналоги пиримидина, такие как фтороурацил, тегафур или гемцитабин; алкалоиды барвинка, такие как винбластин, винкристин или винорелбин или их аналоги; производные подофиллотоксина, такие как этопозид, таксаны, доцетаксел или паклитаксел; антрациклины, такие как доксорубицин, эпирубицин, идарубицин и митоксантрон и их аналоги; другие цитостатические антибиотики, такие как блеомицин и митомицин; платиновые составы, такие как цисплатин, карбоплатин и оксалиплатин; пентастатин, милтефозин, эстрамустин, топотекан, иринотекан и бикалутамид) и токсины (например, токсин рицина, лиатоксин и токсин Веро). В ряде случаев эти функциональные молекулы удаляются в конце. Кроме того, молекулы могут быть пептидами, которые могут узнаваться и отщепляться ферментами, такими как тромбин, матричная металлопротеаза (MMP) и фактор X, полинуклеотидами, которые могут расщепляться нуклеазами или эндонуклеазами рестрикции.
Аптамер или комплекс настоящего изобретения может использоваться как, например, фармацевтическое вещество или реактив (например, диагностические реактивы, реактивы для анализов (включая экспериментальные реактивы)). Например, аптамеры или комплекс настоящего изобретения могут использоваться как ингибиторы клеточной миграции, ускорители регуляции пролиферации T клеток, ускорители супрессивной функции регуляторных Т клеток, супрессанты ингибирования апоптоза, ингибиторы клеточной пролиферации, ингибиторы клеточной дифференцировки, агенты доставки лекарственных средств, зонды для изображений ин-виво, зонды для измерения концентрации МК в крови, зонды для окрашивания ткани, зонды для ELISA и лиганды для отделения и очистки МК.
Аптамеры или комплекс настоящего изобретения могут также использоваться для предотвращения или лечения различных болезней, таких как аутоиммунные заболевания (например, рассеянный склероз, системная эритематозная волчанка, болезнь Шегрена, полимиозит, дерматомиозит, ревматический артрит (ревматоидный артрит, остеоартрит), воспалительный энтерит (болезнь Крона и т.п.), прогрессивный системный склероз, нодозный панартериит (PN), болезни щитовидной железы (Базедова болезнь и т.п.), синдром Гийена-Барре, первичный желчный цирроз печени (PBC), идиопатическая тромбоцитопеническая пурпура, аутоиммунная гемолитическая анемия, миастения гравис (MG), амиотрофический боковой склероз (ALS), диабет I типа, псориаз, астма, функциональные отклонения нейтрофилов), раковые заболевания (например, эзофагальный рак, рак щитовидной железы, рак мочевого пузыря, рак ободочной и прямой кишки, рак желудка, рак поджелудочной железы, рак груди, рак печени, рак легкого, рак молочной железы, нейробластома, глиобластома, рак матки, рак яичников, опухоль Уилмса, рак простаты), постоперационная адгезия, эндометриоз, отторжения при трансплантации, аллергии, рестенозис после сосудистой хирургической реконструкции, сердечная коронарная артериальная сосудистая обструктивная болезнь, мозговая сосудистая обструктивная болезнь, почечная сосудистая обструктивная болезнь, периферийная обструктивная болезнь сосудов, артериосклероз и мозговой инфаркт. В частности, аптамеры настоящего изобретения ингибируют клеточную миграционную активность МК и поэтому важны для предотвращения или лечения рассеянного склероза, послеоперационной адгезии, эндометриоза, ревматического артрита и сосудистого стеноза.
Фармацевтические препараты настоящего изобретения могут приготовляться с фармацевтически допустимыми носителями. В качестве примеров фармацевтически допустимых носителей могут быть упомянуты (без ограничений) такие эксципиенты, как сахароза, крахмал, маннит, сорбит, лактоза, глюкоза, целлюлоза, тальк, фосфат кальция и карбонат кальция; связывающие вещества, такие как целлюлоза, метилцеллюлоза, гидроксилпропилцеллюлоза, полипропилпирралидон, желатин, гуммиарабик, гликоль полиэтилена, сахароза и крахмал; дизентеграторы, такие как крахмал, карбоксиметилцеллюлоза, гидроксилпропилкрахмал, крахмал гликоля-натрия, карбонат водорода натрия, фосфат кальция и цитрат кальция; лубриканты, такие как стеарат магния, Аэрозил, тальк и лаурил сульфат натрия; ароматические агенты, такие как лимонная кислота, ментол, соль аммония глицирризина, глицин и апельсиновый порошок; консерванты, такие как бензоат натрия, сульфит водорода натрия, метилпарабен и пропилпарабен; стабилизаторы, такие как лимонная кислота, цитрат натрия и уксусная кислота; суспендирующие агенты, такие как метилцеллюлоза, поливинилпирролидон, алуминум стеарат; диспергирующие агенты, такие как сурфактанты; растворители, такие как вода, физиологический раствор и апельсиновый сок; основные воски, такие как масло какао, полиэтиленгликоль и керосин; и т.п.
Препараты, подходящие для орального введения, являются жидкими препаратами, полученными растворением эффективного количества лиганда в растворителе, таком как вода, физиологический раствор или апельсиновый сок; капсулами, пакетиками или таблетками, содержащими эффективное количество лиганда в твердой или гранулированной форме; суспензией, полученной суспендированием эффективного количества активного компонента в соответствующем дисперсанте; эмульсией, полученной дисперсией и эмульсификацией раствора эффективного количества активного компонента в соответствующем дисперсанте и т.п.
Фармацевтические препараты настоящего изобретения могут быть покрыты по известным способам для целей маскировки вкуса, энтеральной растворимости, замедленного высвобождения и т.п., как требуется. В качестве примеров покрывающих веществ, используемых для покрытия, используются гидроксипропилметилцеллюлоза, этилцеллюлоза, гидроксиметилцеллюлоза, гидроксипропилцеллюлоза, полиоксиэтиленгликоль, Твин 80, Плуроник F68, ацетат фталат целлюлозы, фталат гидроксипропилметилцеллюлозы, ацетат сукцинат гидроксиметилцеллюлозы, Эудрагит (произведенный компанией Ром (Rohm), Германия, сополимер метакриловой кислоты/акриловый кислоты) и т.п., пигменты (например, красный оксид железа, диоксид титана и т.п.) и т.д. Фармацевтические препараты могут быть препаратами быстрого высвобождения или препаратами медленного высвобождения. В качестве примеров материалов для медленного высвобождения можно отметить липосомы, атероколлаген, желатин, гидроксиапатита, PLGA и т.п.
В качестве препаратов, подходящих для парентерального введения (например, внутривенного введения, подкожного введения, внутримышечного введения, локального введения, интраперитонеального введения, интраназального введения, легочного введения и т.п.), доступны водные и неводные изотонические стерильные жидкости для инъекций, которые могут включить в себя антиоксидант, буферный раствор, бактериостатический агент, изотонирующий агент и т.п. Могут также быть упомянуты водные и неводные стерильные суспензии, которые могут содержать суспендирующий агент, растворитель, загуститель, стабилизатор, антисептик и т.п. Препарат может быть заключен в сосуд, такой как ампула или пузырек в единице дозирующего объема или в нескольких разделенных дозах. Активный компонент и фармацевтически допустимый носитель могут также быть высушены сублимацией и сохраняться в таком состоянии, которое может быть растворено или суспендировано в соответствующем стерильном наполнителе непосредственно перед использованием. Кроме того, в дополнение к жидкостям для инъекций, ингаляции и мази также возможны. В случае ингаляций активный компонент в высушенном сублимацией состоянии измельчается и вводится ингаляцией с использованием соответствующего устройства ингаляции. Ингаляция может быть соответствующим образом составлена с традиционно используемым сурфактантом, маслом, добавками, циклодекстрином или их производными и т.п., как это требуется.
Здесь, как примеры сурфактанта, могут быть упомянуты олеиновая кислота, лецитин, диэтиленгликоль диолеат, тетрагидрофтофурил олеат, этил олеат, изопропил миристат, глицерил триолеат, глицерил монолаурат, глицерил монолеат, глицерил моностеарат, глицерил монолизиноат, цетиловый спирт, стеариловый спирт, полиэтиленгликоль 400, цетилпиридин хлорид, сорбитан триолеат (торговая марка Span 85), сорбитан монолеат (торговая марка Span 80), сорбитан монолаурат (торговая марка Span 20), усиленное полиоксиэтиленом касторовое масло (торговая марка HCO-60), полиоксиэтилен (20) сорбитан монолаурат (торговая марка Твин 20), полиоксиэтилен (20) сорбитан монолеат (торговая марка Твин 80), лецитин природного происхождения (торговая марка EPICLON), эфир олеилполиоксиэтилена (2) (торговая марка Brij 92), эфир стеарилполиоксиэтилена (2) (торговая марка Brij 72), эфир лаурилполиоксиэтилена (4) (торговая марка Brij 30), эфир олеилполиоксиэтилена (2) (торговая марка Genapol 0-020), блоковый сополимер оксиэтилена и оксипропилена (торговая марка Synperonic) и т.п. В качестве примеров масел могут быть упомнуты кукурузное масло, оливковое масло, хлопковое масло, подсолнечное масло и т.п. В случае мази соответствующая фармацевтически приемлемая основа (желтый парафин, белый парафин, керосин, пластибейз, силикон, белая мазь, воск, сало, растительные масла, гидрофильньная мазь, гидрофильный парафин, очищенный ланолин, гидролизированный ланолин, водопоглощающая мазь, гидрофильный пластибейз, мазь на основе макрогола и т.п.) смешивается с активным компонентом и используется в качестве препарата.
Средство для ингаляции может быть получено стандартным способом. В частности, средство для ингаляции может быть получено превращением вышеописанного аптамера или комплекса настоящего изобретения в порошок или в жидкость, смешивая их с ингаляционным пропеллентом и/или носителем и заполняя соответствующий сосуд ингаляции. В случае, когда вышеописанный аптамер или комплекс настоящего изобретения является порошком, может использоваться обычный механический порошковый ингалятор; в случае жидкости может использоваться такой вид ингалятора, как распылитель. В этом случае в качестве пропеллента могут широко использоваться хорошо известные вещества; можно упомянуть составы хлорфторуглеродного ряда, такие как хлорфторуглерод-11, хлорфторуглерод-12, хлорфторуглерод-21, хлорфторуглерод-22, хлорфторуглерод-113, хлорфторуглерод-114, хлорфторуглерод-123, хлорфторуглерод-142c, хлорфторуглерод-134a, хлорфторуглерод-227, хлорфторуглерод-C318 и 1,1,1,2-тетрафторэтан, углеводороды, такие как пропан, изобутан и n-бутан, эфиры, такие как диэтиловый эфир, сжатые газы, такие как газообразный азот и газообразный углекислый газ и т.п.
Дозировка фармацевтического препарата настоящего изобретения изменяется в зависимости от вида и активности активного компонента, серьезности заболевания, вида животных, являющихся объектом введения, переносимости препарата, массы тела, возраста и т.п.; обычная дозировка, основанная на количестве активного компонента в день для взрослого, может быть приблизительно от 0,0001 до 100 мг/кг, например, от приблизительно 0,0001 до 10 мг/кг, предпочтительно от приблизительно 0,005 до 1 мг/кг.
Настоящее изобретение также обеспечивает твердофазный носитель с иммобилизованным аптамером и/или комплексом настоящего изобретения. В качестве примеров твердофазного носителя могут быть упомянуты субстрат, смола, культуральная чашка (например, мультилуночный планшет), фильтр, картридж, колонка и пористый материал. Субстратом может быть субстрат, используемый в ДНК чипах, белковых чипах и т.п.; например, никель-PTFE (политетрафторэтилен) субстраты, субстраты стекла, субстраты апатита, субстраты кремния, субстраты алюминия и т.п., а также субстраты, полученные покрытием этих субстратов полимером и т.п. В качестве примеров смол могут быть упомянуты частицы агарозы, частицы кварца, сополимер акриламида и метиленбисакриламида, частицы дивинилбензола, сшитые с полистиролом, частицы декстрана, сшитые с эпихлорогидрином, волокна целлюлозы, сшитые полимеры арилдекстрана и метиленбисакриламида, монодисперсные синтетические полимеры, монодисперсные гидрофильньные полимеры, сефароза, Toyopearl и т.п., а также смолы, полученные связыванием различных функциональных групп с этими смолами. Твердофазный носитель настоящего изобретения может использоваться, например, для получения, детектирования и количественного определения МК.
Аптамер и/или комплекс настоящего изобретения могут быть иммобилизованы на твердофазный носитель известными способами. Например, можно упомянуть способ, при котором вводят аффинное вещество (например, описанное выше) или заранее определенную функциональную группу в аптамер и/или комплекс настоящего изобретения, и затем иммобилизуют аптамер или комплекс на твердофазный носитель посредством аффинного вещества или предварительно заданной функциональной группы. Изобретение также обеспечивает такие способы. Предварительно заданная функциональная группа может быть функциональной группой, которая может подвергаться реакции связывания; например, можно упомянуть амино группу, тиольную группу, гидроксильную группу и карбоксильную группу. Настоящее изобретение также обеспечивает аптамер с такой функциональной группой, присоединенной к нему.
Настоящее изобретение также обеспечивает способ получения и концентрирования МК. Способ получения и концентрирования настоящего изобретения может включать адсорбирующийся МК на твердофазный носитель настоящего изобретения и элюцию адсорбированного МК с элюентом. Адсорбция МК на твердофазный носитель настоящего изобретения может осуществляться известным способом. Например, образец, содержащий МК (например, бактериальная или клеточная культура или культуральный супернатант, кровь), вводится в твердофазный носитель настоящего изобретения или состав, содержащий то же самое. Элюция МК может производиться с использованием элюента, такого как нейтральный раствор. Нейтральный элюент не имеет специальных ограничений и может иметь значение pH, например, приблизительно от 6 до 9, предпочтительно приблизительно от 6,5 до 8,5 и более предпочтительно приблизительно от 7 до 8. Нейтральным раствором может также быть раствор, содержащий, например, соль калия (например, NaCl, KCl), соль магния (например, MgCl2), сурфактант (например, Твин 20, Тритон, NP40) или глицерин. Способ получения и концентрирования настоящего изобретения может также включать промывку твердофазного носителя с использованием промывочного раствора после адсорбции МК. В качестве примеров промывочного раствора можно упомянуть растворы, которые содержат мочевину, хелатный агент (например, ЭДТА), Трис, кислоту, щелочь и т.п. Способ получения и концентрирования настоящего изобретения может также включать нагревание твердофазного носителя. Эта стадия позволяет осуществлять регенерацию и стерилизацию твердофазного носителя.
Настоящее изобретение также обеспечивает способ детектирования и количественного определения МК. Способ детектирования и количественного определения настоящего изобретения может включать измерение МК с использованием аптамера изобретения (например, при помощи комплекса и твердофазного носителя настоящего изобретения). Способ детектирования и количественного определения МК может осуществляться подобно иммунологическому способу за исключением того, что вместо антитела используется аптамер изобретения. Поэтому при использовании аптамера настоящего изобретения вместо антитела для детектирования и количественного определения могут также использоваться такие способы, как ферментный имунноанализ (EIA) (например, прямой конкурирующий ELISA, обратный конкурирующий ELISA, сандвич-ELISA), радиоиммуноанализ (RIA), флуоресцентный иммуноланализ (FIA), Вестерн Блот (например, при использовании вместо вторичного антитела в Вестерн блоте), способ иммуногистохимического окрашивания и способ сортировки клеток. Эти способы могут применяться, например, для измерения состава МК в живом организме или биологическом образце и диагностике связанных с МК болезней.
Раскрытия, описанные во всех публикациях, упомянутых здесь, включая патенты и спецификации патентных заявок, включены в настоящее изобретение посредством ссылки до известной степени.
Настоящее изобретение в дальнейшем описано более подробно посредством следующих Примеров, которые, однако, не ограничивают объем изобретения.
Пример 1
Получение нуклеиновых кислот, которые специфически связываются с мидкином 1
Нуклеиновые кислоты, которые специфически связываются с мидкином, были получены с использованием способа SELEX. SELEX осуществлялся по способу, описанному Ellington et al. (Ellington and Szostak, Nature 346, 818-822, 1990) и способу, описанному Tuerk et al. (Tuerk and Gold, Science 249, 505-510, 1990) с усовершенствованиями. В качестве вещества-мишени мидкин человека получали с использованием дрожжей по способу Murasugi et al. (Murasugi and Tohma-Aiba, Protein expression and purification 27, 244-252, 2003). В дальнейшем, если не определено иначе, мидкин означает мидкин человека. Мидкин иммобилизовывали на смоле агарозы (NHS-активизированная Сефароза, произведенная Amersham Bioscience) аминосвязыванием. Аминосвязывание осуществляли так, как рекомендовано в описаниях Amersham Bioscience. Количество иммобилизованного мидкина подтверждалось исследованием раствора мидкина непосредственно перед иммобилизацией и супернатанта сразу после иммобилизации в полиакриламидном геле. В результате этого исследования в полиакриламидном геле полоса мидкина в супернатанте не была обнаружена; было подтверждено, что почти весь используемый мидкин был связан. Это означает, что приблизительно 175 микрограммов мидкина было иммобилизовано на приблизительно 70 микролитрах смолы.
РНК, используемая в первом раунде (40N-РНК), была получена с помощью транскрипции химически синтезируемой ДНК, используя набор для транскрипции DuraScribeTM T7 (произведенный Epicentre). РНК, полученная таким способом, имеет фторозамещенный нуклеотид пиримидина во 2'-положении рибозы. В качестве матрицы ДНК использовалась ДНК длиной 94 нуклеотида, приведенная ниже, имеющая последовательность праймеров со случайным 40-нуклеотидным сиквенсом на каждом конце. Матрица ДНК и праймеры были получены химическим синтезом (произведенный Operon).
Матрица ДНК: 5 '-tcctcattcctgtcctcta-40N-ttcctcttctcctctccc-3' (SEQ ID NO:71)
Праймер прямой: 5 '-taatacgactcactatagggagaggagaagaggaa-3' (SEQ ID NO:72)
Праймер обратный: 5 '-tcctcattcctgtcctcta-3' (SEQ ID NO:73)
N обозначает любой из A, G, C и T. Прямой праймер включает последовательность промотора T7 РНК полимеразы. Вариации набора РНК, используемого в первом раунде, теоретически составляли 1014.
Пул РНК добавляли к мидкин-иммобилизованной смоле и оставляли при комнатной температуре в течение 30 минут. После 30 минут для удаления РНК, не связавшейся с мидкином, смола промывалась раствором A. Раствор A являлся смесью раствора 145 мМ хлорида натрия, 5,4 мМ хлорида калия, 1,8 мМ хлорида кальция, 0,8 мМ хлорида магния и 20 мМ Трис (pH 7,6). Мидкин-связанная РНК извлекалась путем нагревания до 95°С в течение 10 минут с добавлением элюента. В качестве элюента использовалась смесь раствора 7М мочевины, 3 мМ ЭДТА и 100 мМ Трис, со значением pH, доведенным до 6,6. Полученная РНК амплифицировалась ПЦР с обратной транскрипцией и транскрибировалась с использованием набора для транскрипции DuraScribeTM T7, полученный продукт использовался в качестве пула для следующего раунда. Эта операция повторялась 7 раундов так же, как на первом раунде. После завершения SELEX ПЦР продукт был заклонирован в вектор pGEM-T Easy (произведенный Promega), который был трансформирован в штамм DH5α Escherichia coli (произведенный Toyobo). После получения плазмид из одиночных колоний последовательности оснований 48 клонов были определены с использованием ДНК сиквернатора (ABI PRISM 3100, произведены ABI).
После 7 раундов SELEX были исследованы полученные последовательности: они проявляли конвергенцию. Среди них было двадцать копий последовательности, представленной SEQ ID NO:1, и одна копия ее формы с заменой 2 нуклеотидов. Также присутствовали две копии последовательности, представленной SEQ ID NO:2. Присутствовала одна копия каждой из последовательностей, представленных SEQ ID NO:3-5. Вторичные структуры РНК, представленных SEQ ID NO:1-5, оценивались с использованием программы MFOLD (M. Zuker, Nucleic Acids Res. 31 (13), 3406-3415, 2003). Таким образом, были замечены внутренние структуры петля-ствол-шпилька, морфологически подобные РНК, представленной SEQ ID NO:2, 3 и 4 (фиг.1-5). Все петли шпилек были составлены из восьми нуклеотидов; 2 и 3 были одно-нуклеотидными замещенными формами по сравнению с SEQ ID NO:4. Что касается стволов, SEQ ID NO:2 формировалась с помощью двух пар оснований, а SEQ ID NO:3 и 4 формировались с помощью трех пар оснований.
Каждая нуклеотидная последовательность показана ниже. Круглые скобки в каждом нуклеотиде обозначают модификации в 2'-положении, и F является атомом фтора (те же обозначения в дальнейшем).

последовательность петли шпильки
SEQ ID NO:2 SEQ-ggaaugaa-
SEQ ID NO:3 SEQ-ggaaagca-
SEQ ID NO:4 SEQ-ggaaagaa-
Пример 2
Получение нуклеиновых кислот, которые специфически связываются с мидкином 2
Для получения аптамеров, которые связываются с мидкином, но не связывают с плейотропином, белком из семейства мидкинов, выполнялся SELEX с предварительной субтракцией с использованием плейотропина. На первом этапе, как и с мидкином, плейотропин иммобилизовали со смолой агарозы с помощью аминосвязывания. Затем к плейотропин-связанной смоле добавляли пул РНК и оставляли при комнатной температуре в течение 30 минут. После этого собирали только супернатант. Теоретически этот супернатант не должен содержать РНК, которая связывается с плейотропином. Этот супернатант добавляли к мидкин-связанной смоле и осуществляли SELEX таким же способом, как и в Примере 1. Используемый плейотропин был экспрессирован в дрожжах по способу Murasugi et al. (Murasugi, Kido, Kumai и Asami, Biosci. Biotech. Biochem. 67 (10), 2288-2290, 2003). Матричная ДНК и используемые праймеры были такими же, как и используемые в Примере 1.
После завершения 7 раундов были исследованы последовательности 48 клонов; в последовательностях наблюдалась конвергенция. Среди них присутствовали десять копий той же самой последовательности, как и SEQ ID NO:3, полученной в Примере 1, и 1-нуклеотидно замещенная форма. Также было шесть копий такой же последовательности, как и SEQ ID NO:2, и две копии 1-нуклеотидно замещенной формы. Кроме того, присутствовала одна копия такой же последовательности, как и SEQ ID NO:5.
Пример 3
Получение нуклеиновых кислот, которые специфически связываются с мидкином 3
В случае, когда мидкин иммобилизован с помощью аминосвязывания, важные его части могут разрушаться в зависимости от участка аминосвязывания. Поэтому был осуществлен фильтр-связывающий SELEX с использованием нитроцеллюлозной мембраны, который не включает иммобилизацию на носитель. Это делалось с целью отделить нуклеиновые кислоты, которые связываются с белком-мишенью от нуклеиновых кислот, которые с ним не связываются, основываясь на том факте, что белки, скорее всего, связываются с нитроцеллюлозными мембранами, в то время как нуклеиновые кислоты, скорее всего, с ними не связываются. Пул РНК и мидкин смешивались, выдерживались в течение 30 минут при комнатной температуре, а затем смесь фильтровалась с использованием нитроцеллюлозной мембраны. После того как нитроцеллюлозная мембрана была полностью промыта раствором A, ее помещали в элюент B и нагревали до 90°C в течение 5 минут. После этого таким же способом, как и в Примере 1, получали РНК осаждением этанолом, которая затем была амплифицирована ПЦР с обратной транскрипцией и транскрибирована в пул РНК для использования на следующем раунде. Матричная ДНК и используемые праймеры были такими же, как и в Примере 1. Элюент B является жидкой смесью 50%-ого фенола и 6 М мочевины.
После завершения шести раундов были проверены последовательности 48 клонов; существенной конвергенции не было получено. Следовательно, SELEX был выполнен еще три раза; после завершения девяти раундов были проверены последовательности 48 клонов; наблюдалась достаточная конвергенция. Среди последовательностей присутствовали 21 копия такой же последовательности, как SEQ ID NO:2 и четыре копии 1-нуклеотидно замещенной формы. Присутствовало десять копий такой же последовательности, как SEQ ID NO:4. Были обнаружены три новых последовательности, ни одна из которых не проявляла конвергенции.
Пример 4
Оценка связывающих активностей с помощью способа поверхностного плазмонного резонанса
Активности связывания РНК, представленных SEQ ID NO:1-5 для мидкина, исследовались способом поверхностного плазмонного резонанса. Измерения осуществлялись с использованием BIAcore2000, произведенного BIAcore. Используемый сенсорный чип был SA чипом, который несет иммобилизованный стрептавидин. С последним было связано около 1000 RU 16-нуклеотидного poly dT с биотином на 5' конце. РНК, являющаяся лигандом, имела 16 нуклеотидный Poly A, добавленный к 3' концу и была иммобилизована к SA чипу посредством связи между dT и A. Иммобилизованного количества было приблизительно 1000 RU. Вводилось 70 микролитров мидкина для аналита, приготовленного в концентрации 0,5 мкМ. Проточным буфером, используемым для BIAcore, был раствор A. В результате измерений было обнаружено, что все РНК, представленные SEQ ID NO:1-5, связываются с мидкином (фиг.6). В качестве отрицательного контроля такое же измерение выполнялось с использованием иммобилизованной 40N-РНК, которая содержала случайную последовательность из 40 нуклеотидов. В результате было найдено, что 40N-РНК также обладала аффинностью к мидкину. Степень аффинности была высокой, подобно аффинности РНК, представленной SEQ ID NO:1-5. Поскольку мидкин содержит большое количество основных аминокислот, таких как лизин, он, как предполагается, связывается неспецифически с отрицательно заряженными нуклеиновыми кислотами.
Поэтому измерение осуществлялось с использованием в качестве проточного буфера для BIAcore буфер с высокой концентрацией соли (раствор B), приготовленный с помощью изменения концентрации хлорида натрия в растворе А до 500 мМ. Предполагалось, что при использовании буфера с высокой концентрацией хлорида натрия неспецифическая адсорбция ионных связей может уменьшиться. В результате измерения было показано, что 40N-РНК почти не связывается с мидкином. В то же время РНК, представленные SEQ ID NO:2-5, связываются с мидкином в более высокой степени, чем с 40N-РНК (фиг.7). Факт связывания при высокой концентрации соли означает, что связь, скорее всего, является гидрофобной связью. Это говорит о том, что эти РНК не связываются неспецифически с лизиновой частью, а специфически узнает мидкин.
Далее, был проведен эксперимент, в котором мидкин был иммобилизован на CM4 сенсорный чип с помощью аминосвязывания, и РНК, представленная SEQ ID NO:4 или 5, была введена в качестве аналита, посредством чего была проверена аффинность РНК и мидкина. Иммобилизация мидкина достигалась с использованием гидроксисукцинимида (NHS, 11 мг/л) и этилдиметиламинопропил карбодиимил гидрохлорида (EDC, 75 мг/л) в соответствии с рекомендациями BIAcore. Мидкин растворяли в буфере HBS-EP (произведенный BIAcore) и использовали при концентрации 20 мг/мл. Для блокирования использовался 1 М гидрохлорид этаноламина (pH 8,5). МК (1-59, MK-N, произведенный Peptide Institute, Inc.) иммобилизовали к проточной ячейке 2 одного сенсорного чипа, МК (60-121, MK-C, произведенный Peptide Institute, Inc.) иммобилизовали к проточной ячейке 3 и полноразмерный мидкин (MK-NC) иммобилизовали к проточной ячейке 4. Проточная ячейка 1 использовалась как контрольная ячейка. При иммобилизации 3 видов мидкина и фрагмента мидкина к одному сенсорному чипу так, как описано выше, одновременно могут быть измерены аффинность для 3 видов лиганда. В результате этого измерения было найдено, что РНК, представленная SEQ ID NO:4, связывается с полноразмерным мидкином (в дальнейшем, обозначаемым МК-NC) и C-концевым доменом мидкина (в дальнейшем, обозначаемым МК-C), но не связывается с N-концевым доменом мидкина (в дальнейшем, обозначаемым МК-N) (Таблица 1). В то же время РНК, представленная SEQ ID NO:5, связывается со всеми МК-NC, МК-N и МК-C, но аффинность для МК-N является более высокой, чем для МК-C.
Аффинность мидкина и различных аналитов
Измеренный способом поверхностного плазмонного резонанса, мидкин был иммобилизован к сенсорного чипу CM4, и были введены различные аналиты. Аффинность в порядке убывания представлена следующим образом +++, ++, + и -.
Подобные эксперименты выполнялись с использованием в качестве аналита, вместо РНК, гепарина (Гепарин, натриевая соль, интестинальная слизистая свиньи, Низкомолекулярная, Мол Вес: 5000, произведенный Calbiochem), хондроитин сульфат E (из хряща кальмара, произведенный Seikagaku), хондроитин сульфат C (из хряща акулы, Мол Вес: 40,000-80,000, произведенный Seikagaku) или тРНК (произведенный Sigma). В результате было обнаружено, что все эти аналиты имели низкую аффинность к МК-N и связывались главным образом с МК-C (Таблица 1).
Из вышеуказанного следует, что было найдено, что РНК, представленная SEQ ID NO:4, как гепарин и ему подобное, связывается с C-концевым доменом мидкина. В то же время было найдено, что РНК, представленная SEQ ID NO:5, имеет низкую аффинность к C-концевому домену и связывается сильнее с N-концевым доменом. Это иллюстрирует, что РНК, представленные SEQ ID NO:4 и 5, связываются с различными участками мидкина. Известно, что у мидкина в C-концевом домене имеется активный участок, являющийся гепарин-связывающим центром (Muramatsu H et al., Biochem Biophys Res Commun. 1994 Sep 15; 203 (2):1131-9, 106; Iwasaki W et al., EMBO J. 1997 1 Dec; 16 (23):6936-46).
Способом поверхностного плазмонного резонанса, как описано выше, определялось, обладала ли РНК, представленная SEQ ID NO:5, связывающей активностью для белка плейотропина из семейства мидкинов. РНК была иммобилизована к сенсорному чипу SA, и на него вводилось 0,5 микроМ плейотропина. Для уменьшения неспецифической адсорбции к раствору плейотропина добавлялось 0,4 мг/мл тРНК. В результате измерения было обнаружено, что РНК, представленная SEQ ID NO:5, обладает связывающей активностью для плейотропина, но степень связывания для плейотрпина ниже, чем степень связывания для мидкина. Подобное измерение было также произведено с использованием 40N-РНК в качестве лиганда. И мидкин и плейотропин связываются с 40N-РНК, но степень связывания была ниже, чем степень связывания для РНК, представленной SEQ ID NO:5. 40N-РНК проявляла более высокую аффинность к мидкину, чем к плейотропину. Из вышеизложенного следует, что было найдено, что плейотропин, как и мидкин, обладает склонностью к связыванию с нуклеиновыми кислотами. Было также найдено, что РНК, представленная SEQ ID NO:5, обладает более высокой аффинностью к мидкину, чем к плейотропину.
Далее, было определено, обладали ли РНК, представленные SEQ ID NO:4 и 5, аффинностью к другим белкам. В качестве белков использовался IgG1 человека (произведенный Calbiochem) и человеческий альбумин (произведенный Sigma). Каждая РНК была иммобилизована с использованием сенсорного чипа SA, как описано выше, и каждый белок вводился в качестве аналита. В результате IgG1 человека и человеческий альбумин не связывался ни с одним из РНК, представленных SEQ ID NO:4 и 5. Из вышеуказанного следует, что было найдено, что РНК, представленные SEQ ID NO:4 и 5, не связывались с человеческим альбумином и IgG человека, которые присутствуют в большом количестве в крови.
Были измерены связывающие активности РНК, представленные SEQ ID NO:2 SEQ 7, 31, 32, 36, 40, 40-1 и 40-2 с МК. Как описано выше, измерение выполнялось с МК, иммобилизованным к сенсорному чипу CM4. В результате было найдено, что все эти РНК обладают аффинностью к МК.
Пример 5
Оценка аптамеров РНК экспериментом ингибирования клеточной миграции
Известно, что мидкин обладает действием инфильтрации прогениторной клетки остеобластов (Qi et al., J. Biol. Chem. 276 (19), 15868-15875, 2001). Следовательно, было исследовано, проявляют ли полученные РНК аптамеры ингибирующую активность в отношении активности мидкина на миграцию клеток, используя клетки UMR106 клеточной линии прогенеторных остеобластов (ATCC No CRL1661). 30 микролитров 1,5 микроM мидкина наносились на внешнюю поверхность мембраны ячейки Chemotaxicell (мембранный диаметр пор 8 микрометров, произведенные Kurabo) для иммобилизации мидкина на внешней поверхности мембраны. Мидкин-иммобилизованная ячейка Chemotaxicell помещалась в 24-луночную культуральную планшету, содержащую 500 микролитров среды (с добавлением 0,3% бычьего сывороточного альбумина в среду ДМЕМ) с добавлением каждого РНК аптамера в концентрации 100 нМ. 200 микролитров клеток UMR106 помещалось на внутренний слой ячейки Chemotaxicell с плотностью 1×106 клеток/мл и культивировалось при 37°C в течение 4 часов. Клетки, остающиеся во внутреннем слое ячейки Chemotaxicell, удалялись, а клетки, которые инфильтровывались и оставались на мидкин-покрытой поверхности, фиксировались метанолом. Ячейка Chemotaxicell погружалась в 1%-ный водный раствор кристаллического фиолетового в течение 30 минут для окрашивания клеток. После того как ячейка Chemotaxicell промывалась дистиллированной водой и высушивалась, пигмент экстрагировался с использованием 200 микролитров смеси растворов 1%-ного SDS и 1%-ного тритона X100. 150 микролитров экстракта переносилось в 96-луночный планшет и определялось его поглощение при 590 нМ.
В результате измерения было найдено, что РНК, представленные SEQ ID NO:1, 2, 4 и 5, обладают значительной активностью, ингибирующей клеточную миграцию. Результаты представлены в Таблице 2. Аптамер, представленный SEQ ID NO:5, проявлял самую высокую ингибирующую активность: среднее значение для 14 измерений составляет 76%. 40N-РНК, используемая как отрицательный контроль, почти не проявляла ингибирующую активность.
Ингибиторные активности клеточной миграции полученных аптамеров в отношении мидкина и плейотропина
Концентрация РНК: 100 нм
Здесь % ингибиторной активности является величиной, полученной вычитанием числа клеток, перемещающихся после добавления аптамера, из числа клеток, перемещающихся без добавления аптамера (поглощение окрашенного клеточного экстракта), взятых за 100. В таблице каждая величина % является средним значением указанных измерений.
После этого было измерено, обладали ли аптамеры, представленные SEQ ID NO:4 и 5, активностью, ингибирующей клеточную миграцию в отношении плейотропина. Эксперимент выполнялся так, как описано выше, за исключением того, что плейотропин использовался вместо мидкина. В результате эксперимента было обнаружено, что эти аптамеры не проявляют заметной ингибиторной активности в отношении плейотропина (Таблица 2).
Затем это было измерено, ингибируют ли гепарин сульфат E хондроитина, сульфат C хондроитина клеточную миграцию в отношении мидкина и плейотропина. Эксперимент выполнялся так, как описано выше, за исключением того, что аптамеры были заменены на гепарин, сульфат E хондроитина, сульфат C хондроитина. Источником используемого гепарина был продукт, произведенный Nacalai Tesque. Источниками используемых сульфата E и C хондроитина были те же, как и в Примере 4. Концентрации гепарина и сульфата E хондроитина были 0,1, 1, 10 и 100 микрограмм/мл. В результате эксперимента гепарин в концентрации 0,1 микрограмм/мл ингибировал ативность клеточной миграции в отношении мидкина и плейотропина. При концентрации 1 микрограмм/мл гепарин ингибировал активность клеточной миграции в отношении мидкина и плейотропина не менее чем на 80%. В то же время сульфат E хондроитина при концентрации 10 микрограмм/мл ингибировал мидкин на 49%, а плейотропин на 69%. Принимая молекулярную массу сульфата C хондроитина за 40,000, эксперимент выполнялся при 500 нМ (20 микрограмм/мл). В результате, в случае, когда ингибиторная активность аптамера, представленного SEQ ID NO:4 (500 нм), была взята за 100, ингибиторная активность сульфата C хондроитина была равна 44.
Исходя из вышесказанного, было обнаружено, что аптамеры, представленные SEQ ID NO:1, 2, 4 и 5, специфически связываются с мидкином, ингибируя активность клеточной миграции. 40N-РНК неспецифически адсорбируется к мидкину электростатически, но не ингибирует активность клеточной миграции. Это показывает, что РНК, полученные способом SELEX, свойства которых нельзя объяснить неспецифической адсорбцией, связываются с важным участком, связанным с активностью клеточной миграции. Гепарин и сульфат E хондроитина одинаково ингибировали активность клеточной миграции, не делая различий между мидкином и плейотропином. В то же время аптамеры, представленные SEQ ID NO:4 и 5, ингибировали только активность мидкина. Поскольку мидкин и плейотропин имеют гомологию 50% и также в связи с тем, что гепарин, связывающий участок, является высококонсервативным, становится понятной высокая специфичность аптамеров.
Пример 6
Миниатюризация и стабилизация аптамера, представленного SEQ ID NO:4
Аптамер, представленный SEQ ID NO:4, имеет 77 нуклеотидов в длину с фторозамещенными нуклеотидами пиримидина в 2'-положениях рибозы. С целью химического синтеза, уменьшения токсичности и улучшения стабильности в крови были выполнены миниатюризация и стабилизация этого аптамера. Операции по миниатюризации и стабилизации были выполнены на основе вторичной структуры, оцененной программой MFOLD, и активность оценивалась в эксперименте ингибирования клеточной миграции. В эксперименте ингибирования клеточной миграции концентрация РНК составляла 100 нм или 500 нм. Так как в экспериментальных результатах встречаются некоторые ошибки в зависимости от состояния клеток, в каждый эксперимент включался ранее проанализированный образец в качестве положительного контроля. Ингибиторные активности, полученные в случае, когда концентрация РНК составляла 500 нм, представлены в Таблице 3 (Таблицы 3-1 и 3-2). Ингибиторные активности выражены как относительные значения по отношению к активности аптамера, представленного SEQ ID NO:4, взятого за 100, для объяснения различий в активностях среди измененных форм. % ингибиторной активности аптамера, представленного SEQ ID NO:4 (значение, полученное вычитанием числа клеток, перемещающихся после добавления аптамера от значения 100, которое является числом клеток, перемещающихся без добавления аптамера), составлял 73% в случае, когда концентрация РНК составляла 500 нм. Это значение является средним значением для 4 измерений. Средним значением % ингибирующей активности для 6 измерений было 63% в случае, когда концентрация РНК составляла 100 нм.
Ингибиторные активности клеточной миграции измененных форм РНК, представленных SEQ ID NO:4, в отношении мидкина
Концентрация РНК составляла 500 нм. Активности выражены в относительных значениях относительно активности РНК, представленной SEQ ID NO:4, взятой за 100. % ингибирующей активности РНК, представленной SEQ ID NO:4, составлял 73%. Эта величина является средним значением для 4 измерений. (мышь) указывает на то, что значение относится к мидкину мыши. Хонд-C обозначает сульфат хондроитина С.
Измененные части в измененных формах (SEQ ID NO:6-29) объяснены ниже.
SEQ ID NO:6: 10 нуклеотидов были удалены из одноцепочечной части на 3' концах РНК, представленной SEQ ID NO:4.
gggagaggagaagaggaac(F)gu(F)gc(F)u(F)c(F)u(F)gu(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggu(F)gu(F)gu(F)agaggac(F)a
SEQ ID NO:7: 14 нуклеотидов были удалены из одноцепочечной части на 5' концах РНК, представленной SEQ ID NO:4, и один G был добавлен для транскрипции.
gggaac(F)gu(F)gc(F)u(F)c(F)u(F)gu(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggu(F)gu(F)gu(F)agaggac(F)aggaau(F)gagga
SEQ ID NO:8: 4 пары оснований были удалены из ствола в концевой части РНК, представленной SEQ ID NO:4.
gggagaggagaagaggaac(F)gc(F)u(F)gu(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggu(F)gu(F)gu(F)agc(F)aggaau(F)gagga
SEQ ID NO:9: 8 нуклеотидов из внутренней петли и CGG на противоположной ее части были удалены из РНК, представленной SEQ ID NO:4.
gggagaggagaagaggaac(F)gu(F)gc(F)u(F)c(F)u(F)gu(F)ac(F)gc(F)c(F)ggaaagaaggu(F)gu(F)gu(F)agaggac(F)aggaau(F)gagga
SEQ ID NO:10: часть петли была заменена на GAAA тетрапетлю в РНК, представленной SEQ ID NO:4.
gggagaggagaagaggaac(F)gu(F)gc(F)u(F)c(F)u(F)gu(F)ac(F)gaggagu(F)agc(F)c(F)gaaaggc(F)ggu(F)gu(F)gu(F)agaggac(F)aggaau(F)gagga
SEQ ID NO:11: 3 пары оснований G-U были заменены парами оснований G-C в стволе в концевой части РНК, представленной SEQ ID NO:4.
gggagaggagaagaggaac(F)gu(F)gc(F)u(F)c(F)u(F)gc(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)gc(F)agaggac(F)aggaau(F)gagga
SEQ ID NO:12: три пары оснований G-U были заменены парами оснований G-C в стволе в концевой части, и 11 нуклеотидов были удалены из одноцепочечной части на 3' конце РНК, представленной SEQ ID NO:8.
gggagaggagaagaggaac(F)gc(F)u(F)gc(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)gc(F)agc(F)
SEQ ID NO:13: 11 нуклеотидов были удалены из одноцепочечной части на 5' конце РНК, представленной SEQ ID NO:12, и GGG был добавлен к ней для транскрипции.
gggagaggaac(F)gc(F)u(F)gc(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)gc(F)agc(F)
SEQ ID NO:14: одна пара оснований G-C и одна пара оснований C-G были удалены из ствола в концевой части РНК, представленной SEQ ID NO:12.
gggagaggagaagaggaac(F)gc(F)u(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)agc(F)
SEQ ID NO:15: A36 и A37 были удалены из части петли РНК, показанной SEQ ID NO:12.
gggagaggagaagaggaac(F)gc(F)u(F)gc(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagggc(F)ggc(F)gu(F)gc(F)agc(F)
SEQ ID NO:16: A23 был удален из внутренней части петли РНК, представленной SEQ ID NO:12.
gggagaggagaagaggaac(F)gc(F)u(F)gc(F)ac(F)gagggu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)gc(F)agc(F)
SEQ ID NO:17: одна пара оснований G-C и одна пара оснований C-G были удалены из ствола в концевой части РНК, представленной SEQ ID NO:13.
gggagaggaac(F)gc(F)u(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)agc(F)
SEQ ID NO:18: 11 нуклеотидов были удалены из одноцепочечной части в 5' концевой части РНК, представленной SEQ ID NO:17, и GG было добавлено к ней для транскрипции.
gggc(F)u(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)agc(F)
SEQ ID NO:19: одна пара оснований C-G была удалена из части ствола в концевой части РНК, представленной SEQ ID NO:17.
gggagaggaac(F)gu(F)ac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)ac(F)
SEQ ID NO:20: одна пара оснований C-G и одна пара оснований U-A были удалены из части ствола в концевой части РНК, представленной SEQ ID NO:17.
gggagaggaac(F)gac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)c(F)
SEQ ID NO:20-1: одноцепочечная часть РНК, представленная SEQ ID NO:20, была полностью модифицирована с OMe.
g(M)g(M)g(M)a(M)g(M)a(M)g(M)g(M)a(M)a(M)c(F)gac(F)gaggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)gu(F)c(F)
SEQ ID NO:20-2: первый ствол в РНК, представленной SEQ ID NO:20, был модифицирован с OMe.
gggagaggaac(F)g(M)a(M)c(F)g(M)aggagu(F)agc(F)c(F)ggaaagaaggc(F)ggc(F)g(M)u(F)c(F)
SEQ ID NO:20-3: второй ствол в РНК, представленной SEQ ID NO:20, был модифицирован с OMe.
gggagaggaac(F)gac(F)gaggagu(F)ag(M)c(F)c(F)ggaaagaag(M)g(M)c(F)ggc(F)gu(F)c(F)
SEQ ID NO:20-4: G в части петли РНК, представленной SEQ ID NO:20, был модифицирован с ОМе.

SEQ ID NO:20-5: А в выпуклой части РНК, представленной SEQ ID NO:20-1, был модифицирован с ОМе.
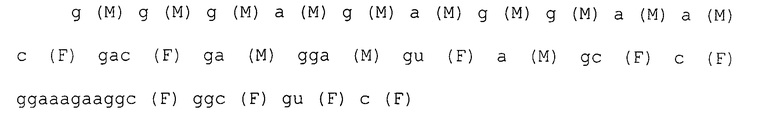
SEQ SEQ ID NO:20-6: G в выпуклой части РНК, представленной SEQ ID NO:20-1, был модифицирован с ОМе.
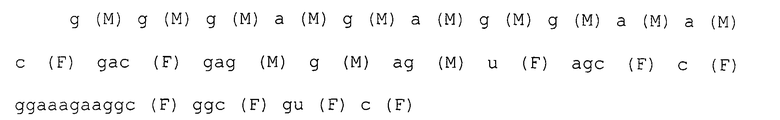
SEQ ID NO:20-7: А в части петли РНК, представленной SEQ ID NO: 20-1, была модифицирована с ОМе.

SEQ ID NO:20-8: А в части петли РНК, представленной SEQ ID N0:20-5 SEQ, была модифицирована с ОМе.
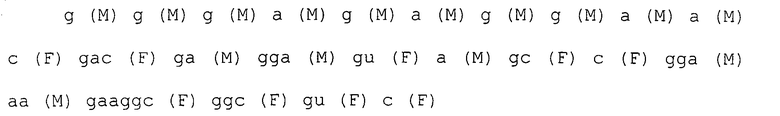
SEQ ID NO:20-9: первый ствол в РНК, представленной SEQ ID NO:20-5, был модифицирован с ОМе.
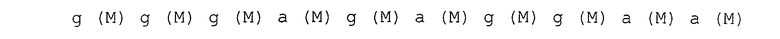

SEQ ID NO:20-10: некоторые части РНК, представленной SEQ ID NO:20-5, были модифицированы с ОМе.

SEQ ID NO:20-11: некоторые части РНК, представленной SEQ ID NO:20-5, были модифицированы с ОМе.

SEQ ID NO:20-12: некоторые части РНК, представленной SEQ ID NO:20-5, были модифицированы с ОМе.

SEQ ID NO:20-13 SEQ: некоторые части РНК, представленной SEQ ID NO:20-5, были модифицированы с ОМе.

SEQ ID NO:21: одна пара оснований G-C была удалена из части ствола в части петли РНК, представленной SEQ ID NO:17.

SEQ ID NO:22: 11 нуклеотидов были удалены из одноцепочечной части в 5' конце РНК, представленной SEQ ID NO:20, и два G были добавлены для транскрипции.
gggac (F) gaggagu (F) agc (F) c (F) ggaaagaaggc (F) ggc (F) gu (F) c (F)
SEQ ID NO:23: часть петли была заменена на GAAA тетрапетлю в РНК, представленной SEQ ID NO:20.
gggagaggaac (F) gac (F) gaggagu (F) agc (F) c (F) gaaaggc (F) ggc (F) gu (F) c (F)
SEQ ID NO:24: часть петли была заменена на UUCG тетрапетлю в РНК, представленной SEQ ID NO:20.
gggagaggaac (F) gac (F) gaggagu (F) agc (F) c (F) u (F) u (F) c (F) gggc (F) ggc (F) gu (F) c (F)
SEQ ID NO:25: внутренняя часть петли в РНК, представленной SEQ ID NO:20, была заменена внутренней петлей аптамера, представленного SEQ ID NO:2.
gggagaggaac (F) gac (F) gagaac (F) agc (F) c (F) ggaaagaaggc (F) ggc (F) gu (F) c (F)
SEQ ID NO:26: G18 внутренней петли в РНК, представленной SEQ ID NO:20, был удален.
gggagaggaac (F) gac (F) gagagu (F) agc (F) c (F) ggaaagaaggc (F) ggc (F) gu (F) c (F)
SEQ ID NO:27: A19 внутренней петли в РНК, представленной SEQ ID NO:20, был удален.
gggagaggaac (F) gac (F) gagggu (F) agc (F) c (F) ggaaagaaggc (F) ggc (F) gu (F) c (F)
SEQ ID NO:28: Два g были удалены от 5' концов и второй пары оснований, A-U был изменен на G-C в РНК, представленной SEQ ID NO:22.
ggc (F) gaggagu (F) agc (F) c (F) ggaaagaaggc (F) ggc (F) gc (F) c (F)
SEQ ID NO:29: Два g были удалены от 5' концов РНК, представленной SEQ ID NO:22.
gac (F) gaggagu (F) agc (F) c (F) ggaaagaaggc (F) ggc (F) gu (F) c (F)
Была исследована ингибиторная активность клеточной миграции аптамера, представленного SEQ ID NO:4, против мидкина мыши. Экспериментальный способ был таким же, как способ для вышеописанного эксперимента с мидкином человека. В результате эксперимента % ингибиторной активности этого аптамера составлял 50%. Это эквивалентно приблизительно 57 относительно ингибиторной активности этого аптамера против человеческого мидкина, взятого за 100; активность явно уменьшилась по сравнению с ингибиторной активностью против человеческого мидкина. Таким образом, было обнаружено, что этот аптамер является аптамером, который проявляет более высокую ингибиторную активность против человеческого мидкина.
Как показано в Таблице 3, аптамер, представленный SEQ ID NO:4, который первоначально имел 77 нуклеотидов в длину, может быть уменьшен до 31 нуклеотида без существенного снижения активности (SEQ ID NO:28-29). Часть петли шпильки этого полученного аптамера не обязательно должна быть GGAAAGAA; аптамер сохраняет активность, даже когда часть петли шпильки является GAAA или тетра-петлей UUCG (SEQ ID NO:23, 24). Даже когда внутренняя часть петли была заменена внутренней частью петли аптамера, представленного SEQ ID NO:2, активность сохранялась (SEQ ID NO:25). Даже когда A19 был удален, активность сохранялась, но когда G18 был удален, активность значительно уменьшалась (SEQ ID NO:26, 27). В то же время было найдено, что, когда пара оснований C-G ствола на стороне петли была удалена, вторичная структура существенно изменялась, и ингибиторная активность исчезала (SEQ ID NO:21). Исходя из вышесказанного, было найдено, что аптамер с 42 нуклеотидами, представленный SEQ ID NO:20 (фиг.8) сохраняет активность даже после замены или удаления некоторых нуклеотидов другими нуклеотидами при условии, что его основная структура изменялась несущественно.
Пример 7
Миниатюризация и стабилизация аптамера, представленного SEQ ID NO:5
Аптамер, представленный SEQ ID NO:5, имеет 77 нуклеотидов в длину и фторозамещенные нуклеотиды пиримидина в 2'-положениях рибозы. С целью обеспечения химического синтеза, уменьшения токсичности и улучшения стабильности в крови осуществлялись миниатюризация и стабилизация этого аптамера. Операции миниатюризации и стабилизации были выполнены на основе вторичной структуры, оцененной программой MFOLD, и активность оценивалась в эксперименте ингибирования клеточной миграции. Концентрация РНК в эксперименте ингибирования клеточной миграции составляла 100 нм или 500 нм. Так как в экспериментальных результатах встречаются некоторые ошибки из-за состояния клеток, в каждое измерение включался ранее проанализированный образец в качестве положительного контроля. Ингибиторные активности, полученные, когда концентрация РНК составляла 100 нм, показаны в Таблице 4-1. Ингибиторные активности выражены как относительные значения по отношению к активности аптамера, представленного SEQ ID NO:5, взятого за 100 для объяснения различия в активностях среди измененных форм. % ингибиторной активности аптамера, представленного SEQ ID NO:5 (значение, полученное вычитанием число клеток, перемещающихся после добавления аптамера от значения 100, который является числом клеток, перемещающихся без добавления аптамера), составлял 76% в случае, когда концентрация РНК составляла 100 нм. Это является средним значением для 14 измерений. Подобный эксперимент был выполнен при концентрации РНК, измененной до 500 нм. Результаты представлены в Таблице 4-2 (Таблица 4-2-1, Таблица 4-2-2). Активности выражены как относительные значения по отношению к активности аптамера, представленного SEQ ID NO:40, взятым за 100. % ингибиторной активности аптамера, представленного SEQ ID NO:40, составлял 82%. Это является средним значением для 4 измерений.
Концентрация РНК составляла 100 нм. Активности выражены как относительные значения по отношению к ингибиторной активности РНК, представленной SEQ ID NO:5 против мидкина, взятого за 100. % ингибиторной активности РНК, представленной SEQ ID NO:5 против мидкина, составлял 76%. Это значение является средним для 14 измерений.
*: приблизительно полученное значение
Измененные части в измененных формах (SEQ ID NOs: 30-70) объяснены ниже.
SEQ ID NO:30: 6 нуклеотидов были удалены из одноцепочечной части в 5' концевой части РНК, представленной SEQ ID NO:5, и один G был добавлен для транскрипции.
ggagaagaggaagu (F) gu (F) gc (F) ac (F) aggggu (F) u (F) ggu (F) gu (F) c (F) gggu (F) gc (F) au (F) ac (F) agu (F) au (F) aagau (F) agaggac (F) aggaau (F) gagga
SEQ ID NO:31: 10 нуклеотидов были удалены из одноцепочечной части в 3' концевой части РНК, представленной SEQ ID NO:5.
gggagaggagaagaggaagu (F) gu (F) gc (F) ac (F) aggggu (F) u (F) ggu (F) gu (F) c (F) gggu (F) gc (F) au (F) ac (F) agu (F) au (F) aagau (F) agaggac (F) a
SEQ ID NO:32: 20 нуклеотидов были удалены из одноцепочечной части в 3' сторонах конца РНК, показанной SEQ ID NO:5.
gggagaggagaagaggaagu (F) gu (F) gc (F) ac (F) aggggu (F) u 
SEQ ID NO:33: 6 нуклеотидов были удалены из одноцепочечной части в 5' концевой части, и 10 нуклеотидов были удалены из одноцепочечной части в 3' концевой части РНК, представленной SEQ ID NO:5.

SEQ ID NO:34: 12 нуклеотидов были удалены из одноцепочечной части в 5' концевой части, и 20 нуклеотидов были удалены из одноцепочечной части в 3' концевой части РНК, представленной SEQ ID NO:5.

SEQ ID NO:35: 6 нуклеотидов были удалены из одноцепочечной части в 3' концевой части РНК, представленной SEQ ID NO:32.

SEQ ID NO:36: две пары оснований были удалены из ствола в концевой части РНК, представленной SEQ ID NO:32.
SEQ ID NO:36-1: одноцепочечная часть в 5' концевой области РНК, представленной SEQ ID N0:36, была полностью модифицирована с ОМе.

SEQ ID NO:37: четыре пары оснований были удалены из ствола в концевой области РНК, представленной SEQ ID NO:32.

SEQ ID NO:38: одноцепочечная часть в 5' концевой области РНК, представленной SEQ ID NO:32, была изменена на poly U, где U обозначает рибозу с фтором в 2'-положении.

SEQ ID NO:39: 6 нуклеотидов были удалены из одноцепочечной части в 3' концевой области, и одна пара оснований была удалена из ствола в концевой области РНК, представленной SEQ ID NO:36.
SEQ ID NO:40: 8 нуклеотидов были удалены из одноцепочечной части в 3' концевой области РНК, представленной SEQ ID NO:36.
SEQ ID NO:40-1: полиэтиленгликоль с молекулярной массой 2000 был добавлен к 5' концу посредством линкера С12, и idT был добавлен к 3' концу РНК, представленной SEQ ID NO:40.
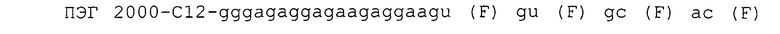
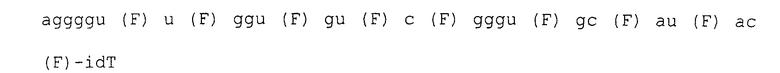
SEQ ID NO:40-2: Все G в одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:40, были модифицированы с ОМе.

SEQ ID NO:40-3: Все А в одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:40, были полностью модифицированы с ОМе.

SEQ ID NO:41: G5 был удален из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:36.
SEQ ID NO:42: A11 был удален из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:36.

SEQ ID NO:43: A17 был удален из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO: 36.

SEQ ID NO:44: одна пара оснований была удалена из ствола в концевой части РНК, представленной SEQ ID NO:40.

SEQ ID NO:45: две пары оснований были удалены из ствола в концевой части РНК, представленной SEQ ID NO:40.

SEQ ID NO:45-1: полиэтиленгликоль с молекулярной массой 2000 был добавлен к 5' концу посредством линкера С12, и idT был добавлен к 3' концу РНК, представленной SEQ ID NO:45.

SEQ ID NO:45-2: все А в одноцепочечной части и все G в части петли в 5' концевой области РНК, представленной SEQ ID NO:45, были полностью модифицированы с ОМе.

SEQ ID NO:45-3: все А в одноцепочечной части и все А и G во внутренней части петли в 5' концевой области РНК, представленной SEQ ID NO:45, были полностью модифицированы с ОМе.

SEQ ID NO:45-4: все А в одноцепочечной части в 5' концевой области и все А и G в части ствола в концевой части РНК, представленной SEQ ID NO:45, были полностью модифицированы с ОМе.

SEQ ID NO:45-4-1: все G в части петли РНК, представленной SEQ ID NO:45-4, были полностью модифицированы с ОМе.

SEQ ID NO:45-4-1-1: С24 РНК, представленной SEQ ID NO:45-4-1, был изменен на нуклеотид РНК.

SEQ ID NO:46: пара оснований A-U была заменена парой оснований G-C в стволе в концевой части РНК, представленной SEQ ID NO:40.

SEQ ID NO:47: U32 был удален из петли РНК, представленной SEQ ID NO:40.

SEQ ID NO:48: G34 был удален из петли РНК, представленной SEQ ID NO:40.

SEQ ID NO:49: U36 был удален из петли РНК, представленной SEQ ID NO:40.

SEQ ID NO:50: G4 и G10 были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:45.
SEQ ID NO:51: G5 и A11 были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:36.

SEQ ID NO:52: G1 и G5 были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:36.

SEQ ID NO:53: G5 и G10 были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:36.

SEQ ID NO:54: все G были изменены с F, и все А были модифицированы с ОМе в одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:45.
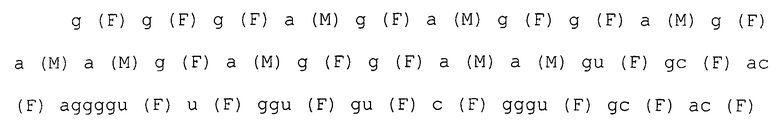
SEQ ID NO:55: A11 и A12 были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:45.
gggagaggaggaggaagu (F) gc (F) ac (F) aggggu (F) u (F) ggu (F) gu (F) c (F) gggu (F) gc (F) ac (F)
SEQ ID NO:56: G13 и A14 были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:45.
gggagaggagaaggaagu (F) gc (F) ac (F) aggggu (F) u (F) ggu (F) gu (F) c (F) gggu (F) gc (F) ac (F)
SEQ ID NO:57: G15 и G16 были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:45.
gggagaggagaagaaagu (F) gc (F) ac (F) aggggu (F) u (F) ggu (F) gu (F) c (F) gggu (F) gc (F) ac (F)
SEQ ID NO:58: A17 и A18 были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:45.
gggagaggagaagagggu (F) gc (F) ac (F) aggggu (F) u (F) ggu (F) gu (F) c (F) gggu (F) gc (F) ac (F)
SEQ ID NO:59: 18 нуклеотидов были удалены из одноцепочечной части в 5' концевой области РНК, представленной SEQ ID NO:45, и два G были добавлены для транскрипции.
gggu (F) gc (F) ac (F) aggggu (F) u (F) ggu (F) gu (F) c (F) gggu (F) gc (F) ac (F)
SEQ ID NO:60: одноцепочечная часть в 5' концевой области РНК, представленной SEQ ID NO:45, была изменена на GGGAAGGA.
gggaaggagu (F) gc (F) ac (F) aggggu (F) u (F) ggu (F) gu (F) c (F) gggu (F) gc (F) ac (F)
SEQ ID NO:61: G5, G10, A11, A12, G13 и A14 были удалены из одноцепочечной части в 5' концевой части РНК, представленной SEQ ID NO:45.

SEQ ID NO:61-1: G был изменен на нуклеотид ДНК, и А был модифицирован с ОМе в одноцепочечной части, и G в части петли был модифицирован с ОМе в 5' концевой области РНК, представленной SEQ ID NO:61.

SEQ ID NO:61-2: G и А в одноцепочечной части и G в части петли в 5' концевой части РНК, представленной SEQ ID N:61, были модифицированы с ОМе.

SEQ ID NO:61-3: часть РНК, представленной SEQ ID NO:61, была модифицирована с F и ОМе.

SEQ ID NO:61-4: часть РНК, представленной SEQ ID NO:61, была модифицирована с ОМе.

SEQ ID NO:61-5: разветвленная цепь полиэтиленгликоля 40 кДа была добавлена к 5' концу, и idT был добавлен к 3' концу РНК, представленной SEQ ID NO:61-4.

SEQ ID NO:61-6: цепь полиэтиленгликоля 30 кДа была добавлена к обоим концам РНК, представленной SEQ ID NO:61-5.

SEQ ID NO:61-7: часть РНК, показанной SEQ ID NO:61 SEQ, была модифицирована с ОМе.

SEQ ID NO:61-8: часть РНК, представленной SEQ ID NO:61, была модифицирована с ОМе.

SEQ ID NO:61-9: часть РНК, представленной SEQ ID NO:61, была изменена с ОМе, и полиэтиленгликоль 2 kDa был добавлен к 5' концу.

SEQ ID NO:62: G13, A14, G15, G16, A17 и A18 были удалены из одноцепочечной части на 5' конце РНК, представленной SEQ ID NO:45.

SEQ ID NO:63: A25 и G26 в РНК, представленной SEQ ID NO:45, были изменены на С, чтобы сделать внутреннюю петлю стволом.

SEQ ID NO:64: U-A был удален из ствола в 5' концевой области РНК, представленной SEQ ID NO:61.

SEQ ID NO:65: A19 и G20 во внутренней петле РНК, представленной SEQ ID NO:61, были заменены на С.

SEQ ID NO:66: некоторая часть РНК, представленной SEQ ID NO:61, была модифицирована с ОМе, и G был модифицирован с F.

SEQ ID NO:67: модификация РНК, представленной SEQ ID NO:66, была изменена.
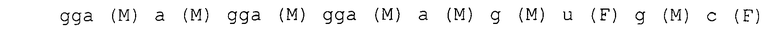

SEQ ID NO:68: часть РНК, представленной SEQ ID NO:61, была модифицирована с ОМе, и U28 был заменен (М).

SEQ ID NO:69: часть РНК, представленной SEQ ID NO:61, была модифицирована с ОМе, и U25 был заменен (М).

SEQ ID NO:70: часть РНК, представленной SEQ ID NO:61, была модифицирована с ОМе, и U24 был заменен (М).

Здесь n (М) представляет рибозу, модифицированную с ОМе в 2'-положениях, n (F) представляет рибозу, модифицированную с F в 2'-положениях, n (Н) представляет дезоксирибозу, ПЭГ 2000 представляет полиэтиленгликоль 2000 Да, ПЭГ 40к представляет разветвленный полиэтиленгликоль 40 кДа, ПЭГ 30к представляет полиэтиленгликоль 30кДа, С12 представляет линкер С12, и idT представляет инвертированный dT.
Был выполнен эксперимент по ингибированию миграции клетки для плейотропина с помощью аптамера, представленного SEQ ID NO:5, и его измененных форм. Экспериментальный способ был таким же, как описан выше, за исключением того, что вместо мидкина использовался плейотропин. Концентрация аптамера составляла 100 нм, и ингибиторная активность аптамера, представленного SEQ ID NO:5 против мидкина, была взята за 100. В результате эксперимента ингибиторная активность против плейотропина составляла 13 (Таблица 4-1). Это является средним значением для 6 измерений. В измененных формах никакой значительной ингибиторной активности против плейотропина не наблюдалось.
Была исследована ингибиторная активность клеточной миграции аптамера, представленного SEQ ID NO:35, против мышиного мидкина. Эксперимент был таким же, как вышеописанный экспериментальный способ с использованием мидкина человека. В результате эксперимента % ингибиторной активности этого аптамера составлял 84%. Таким образом, было показано, что этот аптамер обладал активностью против мышиного мидкина, эквивалентной ингибиторной активности против мидкина человека.
Был выполнен эксперимент ингибирования клеточной миграции, как описано выше, с использованием вместо аптамеров тРНК (произведенной Sigma), Тромбина и ВИЧ-S, которые, скорее всего, не связываются специфически с мидкином. Здесь Тромбин является ДНК аптамером t'ggttggtgtggttgg'taaaaaaaaaaaaaaaa (SEQ ID NO:74), а ВИЧ-S является ДНК аптамером g'tggtgggtgggtggg't (SEQ ID NO:75). Каждый знак "'" представляет фосфотиоатную связь. Фосфотиоатные связи были добавлены для увеличения устойчивости нуклеазы. Эти РНК использовались в концентрации 500 нм. Когда ингибиторная активность аптамера, представленного SEQ ID NO:45 против мидкина человека, была взята за 100, было определено, что активность тРНК равна 28, активность Тромбина равна 0 и ВИЧ-S - 48. Таким образом, было предположено, что аптамеры, полученные в настоящем исследовании, специфически связываются с важными участками, связанными с активностью мидкина на клеточную миграцию.
Аптамер, представленный SEQ ID NO:5, который имел 77 нуклеотидов в длину, мог быть уменьшен до 39 нуклеотидов без существенного снижения активности (SEQ ID NO:61). Одноцепочечная часть на 5' конце не может быть полностью удалена; постулируется, что эта одноцепочечная часть вовлечена в формирование стерической структуры аптамера. В то время как G этой одноцепочечной части может быть нуклеотидом с F модификацией, было обнаружено, что активность уменьшалась в случае OMe-модифицированных нуклеотидов (SEQ ID NO:40-2, 54, 61-2). В то же время, даже когда A был OMe-модифицированным нуклеотидом, активность оставалась неизменной (SEQ ID NO:40-3). Даже когда некоторые пары оснований A-U были заменены парами оснований G-C в стволе в 5' концевой области, это не сильно влияло на активность (SEQ ID NO:46). Даже когда A и G этой части ствола были заменены OMe-измененными нуклеотидами, активность сохранялась (SEQ ID NO:45-4). Даже когда внутренняя часть петли была заменена структурой ствола G-C с помощью нуклеотидной замены, активность не изменялась (SEQ ID NO:63); однако когда одноцепочечная часть была уменьшена, активность уменьшалась (SEQ ID NO:59). Когда G и А внутренней петли были заменены OMe-измененными нуклеотидами, активность уменьшалась (SEQ ID NO:45-3). Когда из части петли был убран 1 нуклеотид, активность уменьшалась (SEQ ID NO:47-49). Даже когда G части петли был заменен OMe-измененным нуклеотидом, активность сохранялась (SEQ ID NO:45-2).
Из вышеизложенного следует, что было найдено, что активность этого полученного аптамера не зависила даже от замен некоторых оснований или изменения модификаций. Было также найдено, что этот аптамер специфически связывается с мидкином и ингибирует активность клеточной миграции. В то же время было обнаружено, что этот аптамер также связывается с семейством белков плейотропина, но не обладает значительной ингибиторной активностью клеточной миграции (Пример 8).
Ингибиторный эффект аптамера на адгезию органа, используя мышиную модель формирования постоперационной адгезии
Открывалась брюшная полость нормальной мыши, и брюшина рассекалась хирургическим ножом и т.п., после чего внутренние органы подсушивались, и затем лапаротомизованная часть зашивалась; в течение 5 дней после этого наблюдается адгезия органа (Am J Obstet Gynecol 179, 438-443, 1998). Имеется сообщение, что когда при этом способе индукции постоперационной адгезии органов использовались мыши, нокаутные по мидкину, постоперационная адгезия органа не происходила (Biochemical and Biophysical Research Communications, 317, 108-113, 2004). Следовательно, был исследован эффект аптамера, представленного SEQ ID NO:76, на предотвращение постоперационной адгезии органа с использованием модели формирования мышиной постоперационной адгезии. Мышь C57BL/6 в возрасте 8 недель (самка) была лапаротамизирована под анестезией, после чего брюшина была протерта впитывающей влагу салфеткой. После этого в брюшине был сделан разрез около 2 см в пяти положениях с использованием ножниц. После гемостаза с использованием впитывающей влагу салфетки в течение 10 минут рана зашивалась с использованием иглы и нити. После выхода их наркоза внутрибрюшинно вводился аптамер, представленный SEQ ID NO:76, в дозе 1 мг/25 мл/кг. В контрольной группе внутрибрюшинно вводился физиологический раствор, содержащий 1 мМ MgCl2 в дозе 25 мл/кг тем же способом. Введение производилось один раз в день в общей сложности три раза в послеоперационные день 0, день 1 и день 2; после этого животное было лапаратомизировано под анестезией в день 3, и степень адгезии органа к ране оценивалась, используя критерии, представленные ниже.
0: нет адгезии
1: с адгезией, низкой (низкой) адгезией
2: с адгезией, умеренной (умеренной) адгезией
3: с адгезией, серьезной адгезией, которая не может быть отделена даже при натяжении органа в части адгезии.
Результаты представлены как средние значения и стандартные ошибки множества степеней адгезии для 9-10 животных в каждой группе (Таблица 5). В результате показатель 3 был для всех животных в группе, получающей физиологический раствор, в то время как средний показатель группы, получающей аптамер, представленный SEQ ID NO:76, равнялся 2,4. В группе, получающей аптамер, представленный SEQ ID NO:76, по сравнению с группой, получающей физиологический раствор, наблюдалось статистически значимое различие (p<0,05). Для статистической обработки использовался Mann-Whitney U тест. Из вышеизложенных результатов следует, что было продемонстрировано, что аптамер, представленный SEQ ID NO:76, обладал активностью, препятствующей постоперационной адгезии органа.
Аптамер, представленный SEQ ID NO:76, является следующим.
SEQ ID NO:76 SEQ: РНК, представленная SEQ ID NO:40, в которой все "А" 5' концевой одноцепочечной части модифицированы с ОМе, холестерин (Chol) был добавлен к 5' концу через линкер насыщенной углеводородной цепи (С12), имеющий 12 атомов углерода, и инвертированный dT (idT) был добавлен к 3' концу.

Эта заявка основана на патентной заявке Номер 2006-308482, зарегистрированной в Японии (дата регистрации: 14 ноября 2006 г.), содержание которой включено здесь полностью посредством этой ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| АПТАМЕР ПРОТИВ NGF И ЕГО ПРИМЕНЕНИЕ | 2012 |
|
RU2633510C2 |
| АПТАМЕР ДЛЯ FGF2 И ЕГО ПРИМЕНЕНИЕ | 2015 |
|
RU2686992C2 |
| АПТАМЕР К ХИМАЗЕ И ЕГО ПРИМЕНЕНИЕ | 2010 |
|
RU2579667C2 |
| АПТАМЕРЫ НА ОСНОВЕ ДНК ДЛЯ КАТЕПСИНА G ЧЕЛОВЕКА | 2004 |
|
RU2360000C2 |
| МОДУЛЯТОРЫ ГЛИКОПРОТЕИНА VI, ПРЕДСТАВЛЯЮЩИЕ СОБОЙ НУКЛЕИНОВУЮ КИСЛОТУ | 2010 |
|
RU2571660C2 |
| СОЕДИНЕНИЯ НУКЛЕИНОВОЙ КИСЛОТЫ ДЛЯ СВЯЗЫВАНИЯ РОСТОВОГО ФАКТОРА ДИФФЕРЕНЦИРОВКИ 11 | 2015 |
|
RU2708170C2 |
| Аптамер, проходящий через гематоэнцефалический барьер головного мозга мыши | 2016 |
|
RU2711912C2 |
| СВЯЗЫВАЮЩИЕ КОМПЛЕМЕНТ АПТАМЕРЫ И СРЕДСТВА ПРОТИВ С5, ПРИГОДНЫЕ ДЛЯ ЛЕЧЕНИЯ ГЛАЗНЫХ НАРУШЕНИЙ | 2007 |
|
RU2477137C2 |
| АПТАМЕРНЫЕ ТЕРАПЕВТИЧЕСКИЕ СРЕДСТВА, ПРИМЕНИМЫЕ ДЛЯ ЛЕЧЕНИЯ СВЯЗАННЫХ С КОМПЛЕМЕНТОМ РАССТРОЙСТВ | 2006 |
|
RU2406507C2 |
| СПОСОБЫ РАЗДЕЛЕНИЯ СПЕРМАТОЗОИДОВ И ПРИМЕНЯЕМЫЕ ДЛЯ ЭТОГО КОМПОЗИЦИИ, СОДЕРЖАЩИЕ АПТАМЕРЫ ИЛИ ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВЫХ КИСЛОТ | 2007 |
|
RU2475539C2 |



























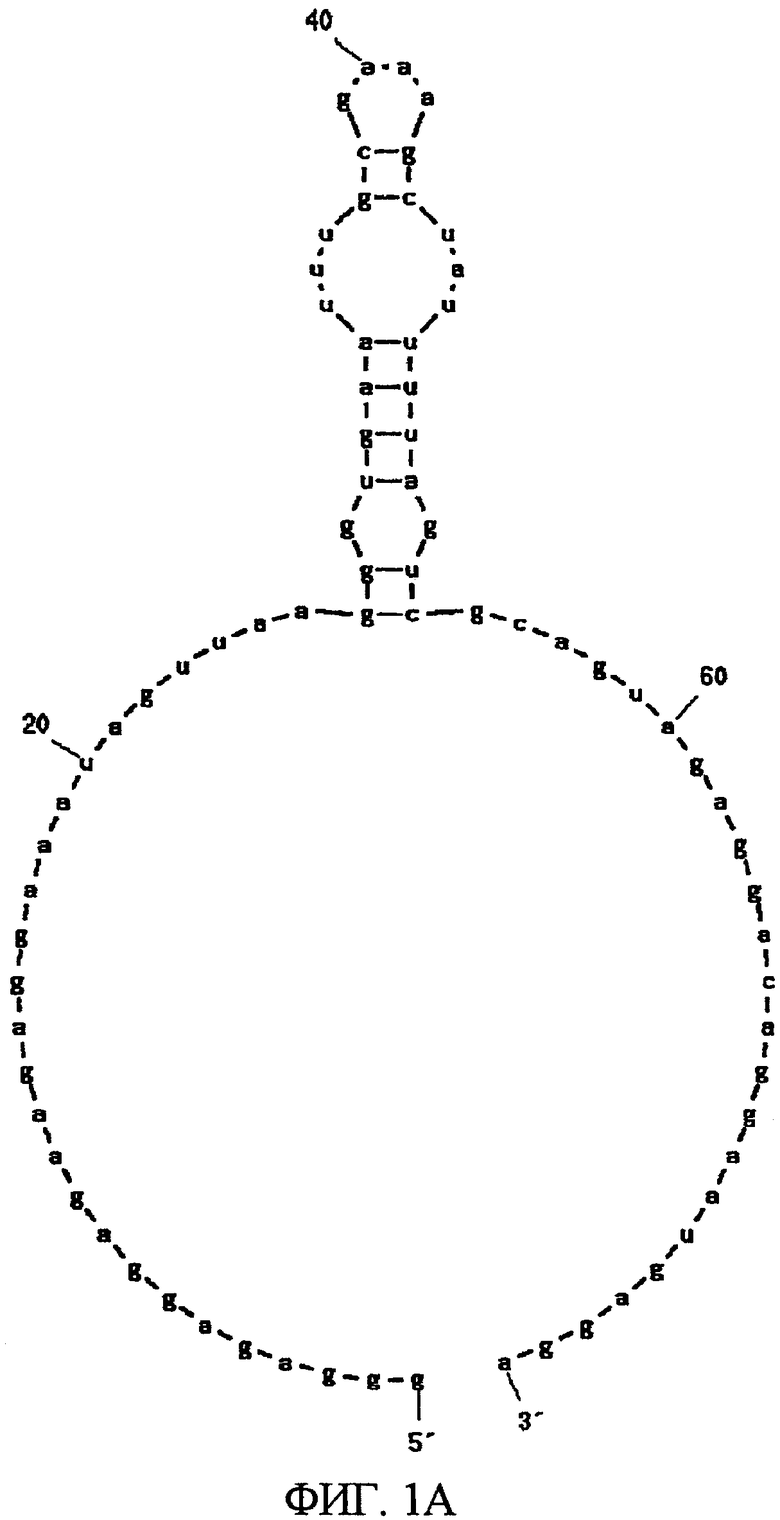


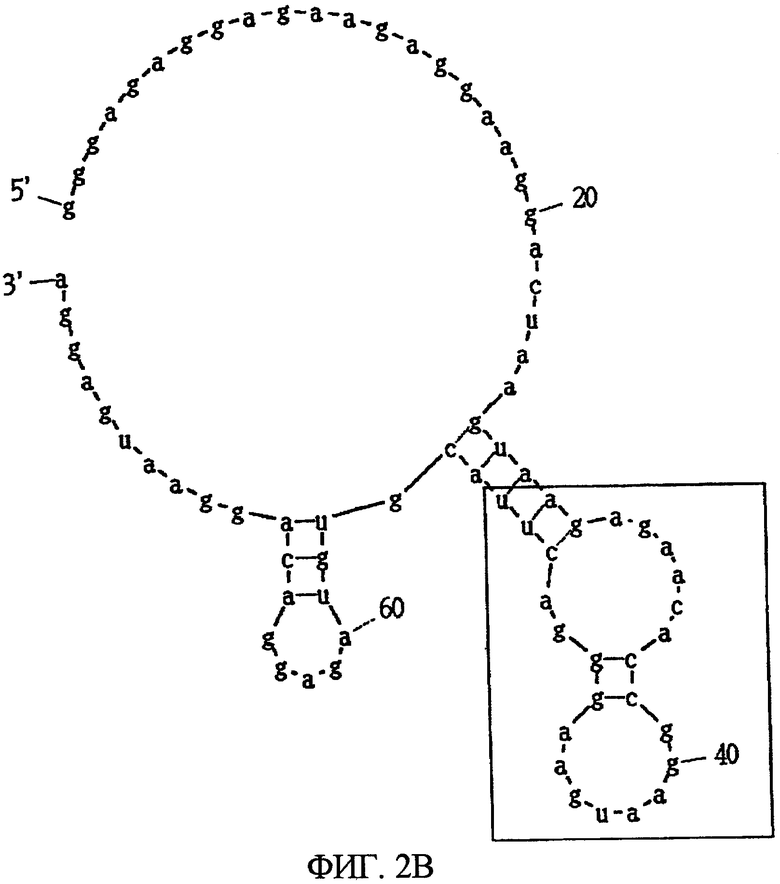

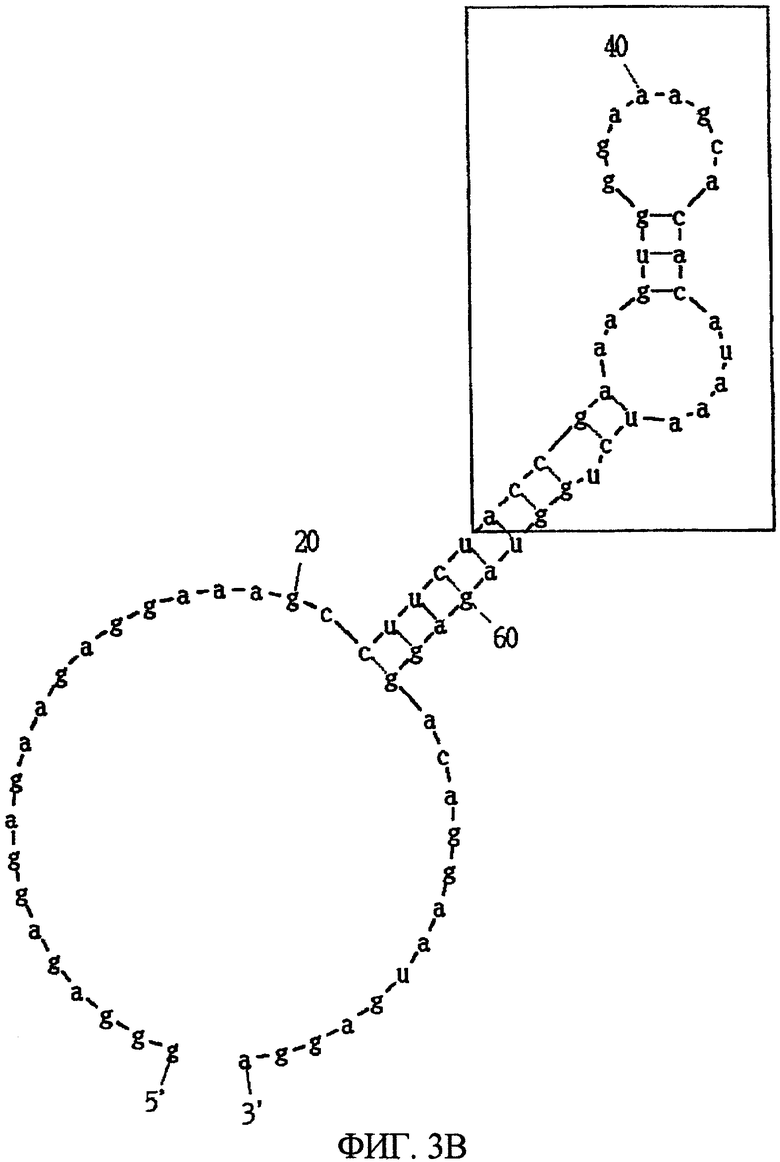
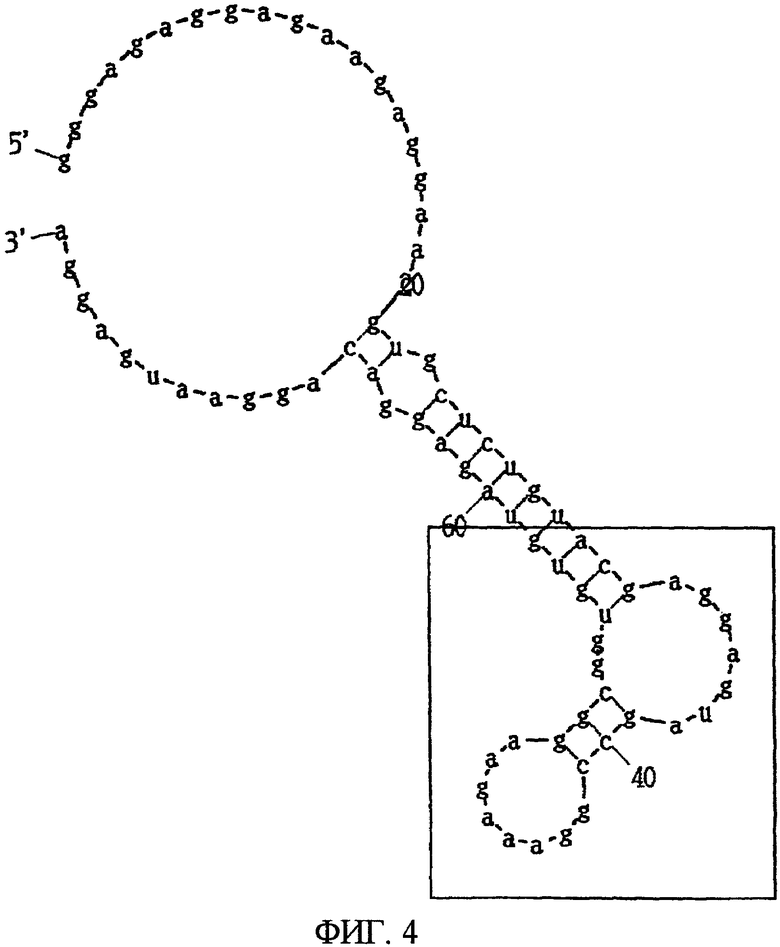
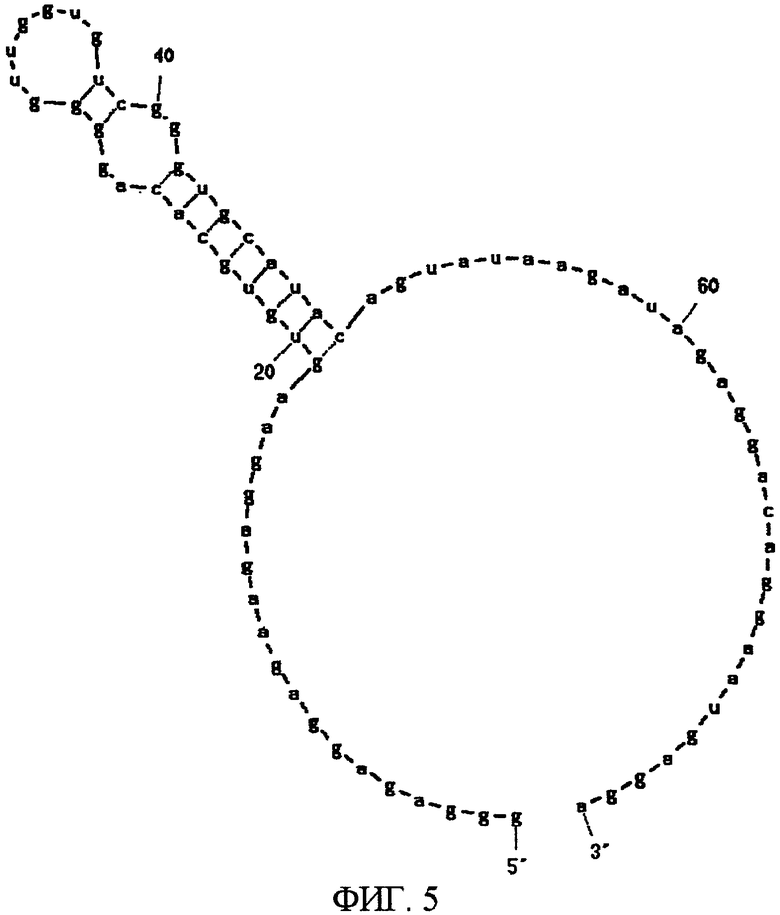

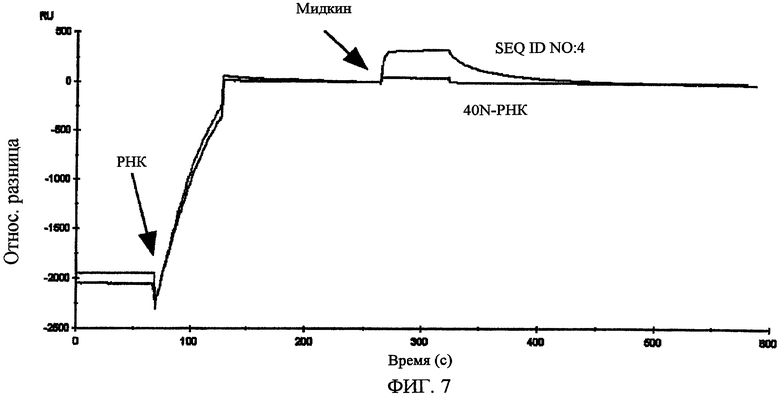
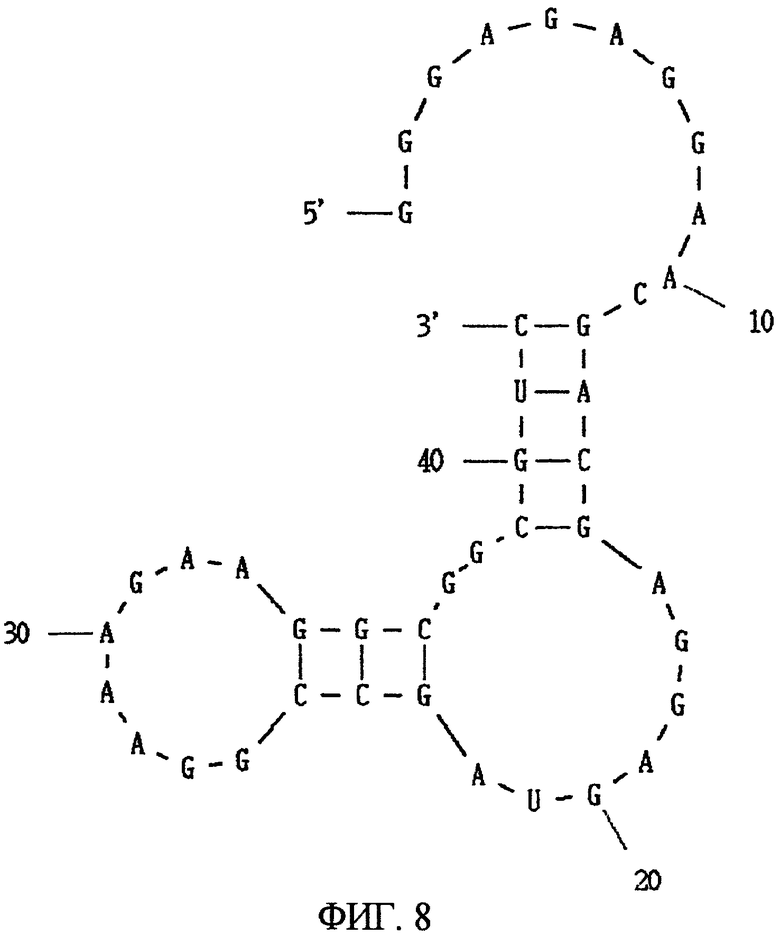
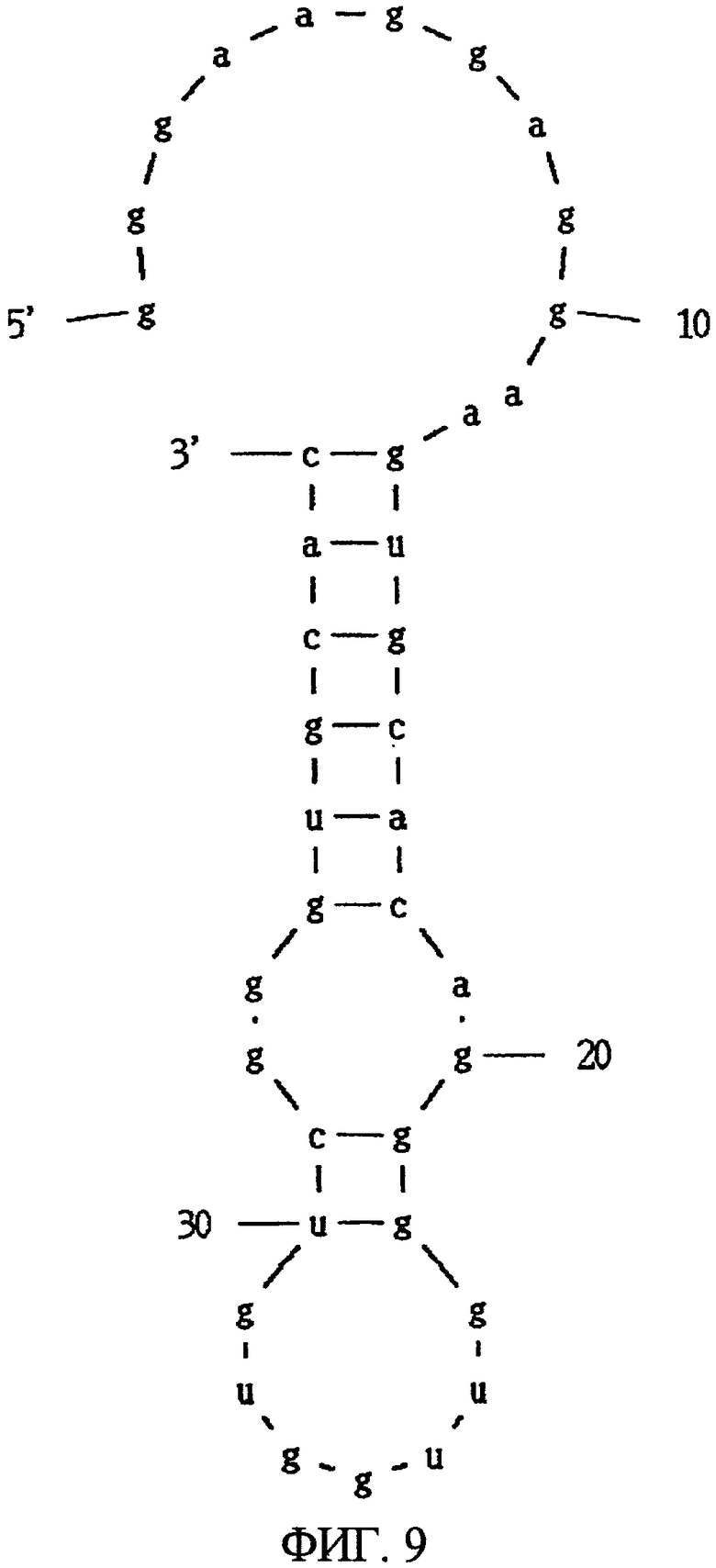
Изобретение относится к области биотехнологии. Изобретение включает высокоэффективный аптамер против мидкина и содержащее его фармацевтическое средство. Такой аптамер позволяет ингибировать активность мидкина и лечить связанные с ним заболевания. 3 н. и 7 з.п. ф-лы, 12 ил., 5 табл., 8 пр.
1. Аптамер, обладающий ингибирующей активностью против мидкина, где аптамер имеет длину не более 80 нуклеотидов, и имеет структуру (а) или (b) из указанного ниже;
(a) аптамер, включающий нуклеотидную последовательность, выбираемую из SEQ ID NO: 1, 2, 4-20, 22-33, 35, 36, 39-64, 66-70 (с условием, что урацил может быть тимином), причем нуклеотиды, содержавшиеся в аптамере, такие, что
(i) в 2'-положениях нуклеотидов пиримидина, одинаковых или различных, расположены атомы фтора или замещены атомами или группами, выбираемыми из группы, состоящей из атомов водорода, гидроксильных групп и метоксильных групп, и
(ii) в 2'-положениях нуклеотидов пурина, одинаковых или различных, расположены гидроксильные группы или замещены атомами или группами, выбираемыми из группы, состоящей из атомов водорода, метоксильных групп и атомов фтора;
(b) аптамер, включающий нуклеотидную последовательность, выбираемую из SEQ ID NO: 1, 2, 4-20, 22-33, 35, 36, 39-64, 66-70 (с условием, что урацил может быть тимином), причем от одного до трех нуклеотидов, за исключением нуклеотидов в стволовой части у петли и внутренней области петли, замещены, удалены, вставлены или добавлены, а нуклеотиды, содержавшиеся в аптамере, такие, что
(i) в 2'-положениях нуклеотидов пиримидина, одинаковых или различных, расположены атомы фтора или замещены атомами или группами, выбираемыми из группы, состоящей из атомов водорода, гидроксильных групп и метоксильных групп, и
(ii) в 2'-положениях нуклеотидов пурина, одинаковых или различных, расположены гидроксильные группы или замещены атомами или группами, выбираемыми из группы, состоящей из атомов водорода, метоксильных групп и атомов фтора.
2. Аптамер по п.1, в котором аптамер не обладает ингибирующей активностью против плейотропина.
3. Аптамер по п.1, обладающий активностью связывания с N-концевым фрагментом мидкина.
4. Аптамер по п.1, обладающий активностью связывания с С-концевым фрагментом мидкина.
5. Аптамер по п.2, обладающий активностью связывания с N-концевым фрагментом мидкина.
6. Аптамер по п.2, обладающий активностью связывания с С-концевым фрагментом мидкина.
7. Аптамер по п.1, который проявляет ингибирующую активность против мидкина, ингибируя связывание мидкина и PTPξ.
8. Аптамер по любому из пп.1-7, где один нуклеотид, содержащийся в аптамере, изменен.
9. Фармацевтический препарат, включающий аптамер по любому из пп.1-8, где препарат предназначен для предотвращения или лечения аутоиммунной болезни, рака, послеоперационной адгезии, эндометриоза, отторжения при трансплантации, аллергии, рестеноза после сосудистой хирургической реконструкции, сердечной коронарной артериально-сосудистой обструктивной болезни, мозговой сосудистой обструктивной болезни, почечной сосудистой обструктивной болезни, периферийной обструктивной болезни сосудов, артериосклероза и мозгового инфаркта.
10. Применение аптамера по любому из пп.1-8 в качестве агента связывания для детектирования мидкина.
| Ng Eugene W.M | |||
| et al | |||
| Pegaptanib, a targeted anti-VEGF aptamer for ocular vascular disease | |||
| Nat Rev Drug Discov | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Burmeister PE et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Chem Biol | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Inoh К et al | |||
| Doxorubicin-Conjugated Anti-Midkine Monoclonal Antibody as a Potential Anti-Tumor Drug | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| RU 2004112533, 16.09.2002. | |||

