Область техники
Настоящее изобретение относится к аптамеру NGF и его применению.
Предшествующий уровень техники
Фактор роста нервов (NGF) является первым нейротрофином, идентифицированным в 1951 году, и он является важным секреторным белком, участвующим в развитии и выживании периферических и центральных нейронов. Он состоит из 118 аминокислот, имеет молекулярную массу 13 кДа и S-S-связи в 3 положениях молекулы.
В качестве рецепторов NGF известны рецептор тирозинкиназного типа TrkA с высокой аффинностью и p75 с низкой аффинностью, принадлежащий к суперсемейству рецепторов фактора некроза опухоли. Эти рецепторы действуют как гомодимер или гетеродимер и интенсивно участвуют в развитии и поддержании нервной системы. TrkA является однопроходным трансмембранным рецептором и имеет тирозинкиназную структуру во внутриклеточном домене. При связывании с NGF происходит фосфорилирование тирозина, сигнал передается дальше и происходит стимуляция дифференцировки и поддержание выживания клетки.
В семействе рецепторов TrkA известны TrkB и TrkC. TrkB связывается с BDNF и NT-4/5, а TrkC связывается с NT-3. По сравнению с TrkA p75 демонстрирует более низкую специфичность к лиганду и, помимо NGF, также связывается с BDNF, NT-3 и NT-4/5. Хотя p75 является однопроходным трансмембранным рецептором, он не имеет тирозинкиназного домена в цитоплазматической части. Подобно TrkA, он экспрессируется не только в нейронах, но также и не в нервных клетках. Известно, что этот рецептор участвует в стимуляции дифференцировки и поддержании выживания клетки, а также связан с индукцией апоптоза и миграции клеток. Результаты кристаллографического анализа указывают на то, что гомодимер NGF связывается с TrkA в соотношении 2:2 и с p75 в соотношении 2:1. Иногда гомодимер NGF связывается с гетеродимером TrkA и p75.
Хорошо известно, что NGF играет ключевую роль в нервной системе. Было выяснено, что NGF действует, поддерживая выживаемость холинергических нейронов, и считают, что он каким-то образом связан с болезнью Альцгеймера. Кроме того, т.к. интрацеребральное введение NGF снижает нарушения памяти у старых крыс, также ожидают, что он будет терапевтическим средством против сенильной деменции.
NGF также связан с воспалением, повышенную экспрессию NGF наблюдают у пациентов с воспалительными заболеваниями и в воспалительных моделях на животных. Их примерами являются системная красная волчанка, рассеянный склероз, псориаз, артрит, интерстициальный цистит, астма и т.п. Сообщают, что в синовиальной жидкости пациентов с ревматоидным артритом обнаруживали более высокую концентрацию NGF. Кроме того, сообщают о повышенной экспрессии NGF у крыс в модели ревматоидного артрита, его повышении в тучных клетках и повышенной экспрессии NGF в модели артрита мыши.
NGF интенсивно участвует в болевой реакции. При подкожном введении NGF человеку сильная боль, такая как мышечная боль, продолжается в течение нескольких дней, и в месте инъекции происходит гипералгезия. У мышей, нокаутных по NGF, и мышей, нокаутных по TrkA, отсутствуют немиелинизированные нервы, и они не чувствуют боли. При интраперитонеальном введении NGF зрелой крысе в дозе 1 мг/кг в ответ на температурный и механический стимулы развивается гипералгезия. Трансгенная по NGF мышь демонстрирует гипералгезию, сопровождающуюся воспалительными состояниями. Кроме того, известно, что у пациентов с врожденным отсутствием чувствительности к боли с ангидрозом (CIPA) ген TrkA является аномальным, и при аномальности гена NGF чувствительность к боли снижается.
Учитывая вышесказанное, следует понимать, что ингибитор NGF можно использовать в качестве терапевтического средства от боли, такой как ноцицептивная боль, воспалительная боль, нейропатическая боль, карциноматозная боль, боль при фибромиалгии и т.п.
В последние годы привлекает внимание применение аптамеров РНК в качестве лекарственных средств, диагностических средств и тестовых лекарственных средств; некоторые аптамеры РНК уже находятся на стадии клинических испытаний, или их применяют на практике. В декабре 2004 года первое в мире лекарственное средство на основе аптамера РНК, макуген, одобрено для применения в качестве терапевтического лекарственного средства против возрастной дегенерации желтого пятна в США. Аптамер РНК относится к РНК, специфически связывающейся с веществом-мишенью, таким как белок, и его можно получать с использованием способа SELEX (систематической эволюции лигандов экспоненциальным обогащением) (патентные ссылки 1-3). В SELEX РНК, специфически связывающуюся с веществом-мишенью, выбирают из совокупности РНК из приблизительно 1014 различных нуклеотидных последовательностей. Используемая структура РНК имеет случайную последовательность из приблизительно 40 остатков, фланкированную последовательностями праймеров. Этой совокупности РНК позволяют связываться с веществом-мишенью и собирают только РНК, связывающуюся с веществом-мишенью, с использованием фильтра и т.п. Собранную РНК амплифицируют с помощью RT-ПЦР и используют в качестве матрицы для следующего раунда. Повторяя это действие приблизительно 10 раз, можно получать аптамер РНК, специфически связывающийся с веществом-мишенью.
Лекарственные средства на основе аптамеров, подобно лекарственным средствам на основе антител, могут быть направлены на внеклеточные факторы. Учитывая многочисленные научные статьи и другие справочные материалы в публичном доступе, лекарственные средства на основе аптамеров считают потенциально превосходящими лекарственные средства на основе антител в некоторых аспектах. Например, аптамеры часто демонстрируют большую силу связывания и большую специфичность, чем антитела. Иммунный клиренс аптамеров маловероятен, и при использовании аптамеров не развиваются побочные реакции, характерные для антител, такие как антителозависимая клеточная цитотоксичность (ADCC) и комплементзависимая цитотоксичность (CDC). Что касается доставки, т.к. аптамеры составляют приблизительно 1/10 размера антител, доставка лекарственного средства в желаемое место является более легкой. Т.к. аптамеры получают химическим синтезом, легче осуществлять различные модификации, и возможно снижение затрат при массовом производстве. При этом время полужизни аптамеров в кровотоке, как правило, меньше, чем у антител; однако иногда это свойство является преимуществом с точки зрения токсичности. Эти факты приводят к выводу, что даже при направленности против одной и той же молекулы лекарственные средства на основе аптамеров потенциально превосходят лекарственные средства на основе антител.
Авторы настоящего изобретения получали аптамер, связывающийся с NGF и ингибирующий связывание NGF с рецептором NGF, и обнаруживали, что аптамер ингибирует активность NGF в отношении роста нейритов (патентный документ 4). В патентном документе 5 описывают аптамер против NGF, получаемый с помощью автоматизированной SELEX, а в патентном документе 6 описывают измененный продукт и модифицированный продукт аптамера, получаемого в патентном документе 4.
Перечень документов
Патентные документы
Патентный документ 1: WO 91/19813.
Патентный документ 2: WO 94/08050.
Патентный документ 3: WO 95/07364.
Патентный документ 4: WO 2010/035725.
Патентный документ 5: WO 02/077262.
Патентный документ 6: WO 03/070984.
Сущность изобретения
Задачи, подлежащие решению посредством изобретения
Цели настоящего изобретения относятся к аптамеру против NGF, способу его применения и т.п. В частности, цель настоящего изобретения относится к аптамеру против NGF, пригодному для использования в качестве фармацевтического препарата, а именно, аптамеру, имеющему короткую цепь, активность, ингибирующую высокую активность NGF (активность в отношении роста нейритов, активность в отношении пролиферации клеток TF-1), и высокую специфичность к NGF.
Средства решения задач
Авторы настоящего изобретения проводили интенсивные исследования, пытаясь решить указанные выше задачи, и добивались успеха в получении аптамера против NGF более высокого качества, демонстрирующего значение IC50 1 нМ или ниже в отношении ингибирования роста нейритов, и исключительно высокую ингибиторную активность в отношении роста нейритов по сравнению с общеизвестными аптамерами против NGF, что привело к осуществлению настоящего изобретения.
Таким образом, настоящее изобретение относится к следующему.
[1] Аптамеру, связывающемуся с NGF и способному образовывать предполагаемую вторичную структуру, представленную формулой (I):

где N является одним нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T,
N11-N13, N21-N23, N32-N38 и N42-N48 являются одинаковыми или разными, и каждый из них является связью или 1 или 2 нуклеотидами, выбранными из группы, состоящей из A, G, C, U и T,
N14, N24, N31, N41, N39 и N49 являются одинаковыми или разными, и каждый из них является одним нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T,
каждые из N14 и N24, N31 и N41, и N39 и N49 образуют уотсон-криковскую пару оснований,
N11-N12-N13-N14 и N21-N22-N23-N24 являются нуклеотидными последовательностями, в комбинации способными образовывать структуру стебля, и
N31-N32-N33-N34-N35-N36-N37-N38-N39 и N41-N42-N43-N44-N45-N46-N47-N48-N49 являются нуклеотидными последовательностями, в комбинации способными образовывать структуру стебля.
[2] Аптамер по предшествующему п.[1], где N11-N13, N21-N23, N32-N38 и N42-N48 являются одинаковыми или разными, и каждый из них является одним нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T.
[3] Аптамер по предшествующему п.[1] или [2], где N14 является U, N24 является A, N31 является G, N41 является C, N39 является G, и N49 является C.
[4] Аптамер по любому из предшествующих пп.[1]-[3], где между N32-N33-N34-N35-N36-N37-N38 и N42-N43-N44-N45-N46-N47-N48 образуется не менее чем 4 уотсон-криковских пар оснований.
[5] Аптамер по предшествующему п.[1], являющийся (a) или (b):
(a) нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO: 3, SEQ ID NO: 9-13, SEQ ID NO: 22-117 и SEQ ID NO: 152-168 (где урацил может являться тимином);
(b) нуклеиновой кислотой, связывающейся с NGF и состоящей из нуклеотидной последовательности по предшествующему п. (a), где от 1 до нескольких нуклеотидов подвергают замене, делеции, инсерции или добавлению.
[6] Аптамер по любому из предшествующих пп. [1]-[5], имеющий длину не более 50 оснований.
[7] Аптамер по любому из предшествующих пп. [1]-[6], где по меньшей мере один нуклеотид модифицируют.
[8] Аптамер по предшествующему п. [7], модифицированный с использованием инвертированного dT или полиэтиленгликоля.
[9] Аптамер по предшествующему п. [8], где инвертированный dT или полиэтиленгликоль присоединяют к 5'-концу или 3'-концу аптамера.
[10] Аптамер по любому из предшествующих пп. [7]-[9], где гидроксильные группы в 2’-положении рибозы соответствующих пиримидиновых нуклеотидов являются одинаковыми или разными, и их не замещают или замещают атомом или группой, выбранной из группы, состоящей из атома водорода, атома фтора и метоксигруппы.
[11] Аптамер по любому из предшествующих пп. [7]-[9], где гидроксильные группы в 2’-положении рибозы соответствующих пуриновых нуклеотидов являются одинаковыми или разными, и их не замещают или замещают атомом или группой, выбранной из группы, состоящей из атома водорода, атома фтора и метоксигруппы.
[12] Аптамер по любому из предшествующих пп. [1]-[11], ингибирующий активность NGF в отношении роста нейритов и/или пролиферации клеток.
[13] Фармацевтическая композиция, содержащая аптамер по любому из предшествующих пп. [1]-[12].
[14] Средство против боли, содержащее аптамер по любому из предшествующих пп. [1]-[12].
[15] Противовоспалительное средство, содержащее аптамер по любому из предшествующих пп. [1]-[12].
[16] Способ лечения или профилактики заболевания, сопровождающегося болью или воспалением, включающий введение аптамера по любому из предшествующих пп. [1]-[12] нуждающемуся в этом индивидууму.
[17] Аптамер по любому из предшествующих пп. [1]-[12] для профилактики или лечения заболевания, сопровождающегося болью или воспалением.
Эффект изобретения
Т.к. аптамер или нуклеиновая кислота по настоящему изобретению демонстрируют лучшую ингибирующую NGF активность, в частности, высокую ингибиторную активность в отношении роста нейритов, благодаря указанному выше строению он может являться применимым в качестве лекарственного средства против заболеваний, таких как алгия, воспалительное заболевание и т.п.
Краткое описание чертежей
Фиг. 1 является схематическим представлением прогнозируемой вторичной структуры аптамера против NGF, приведенного в ID NO: 3, где структура "стебель-петля" сверху слева соответствует консенсусной вторичной структуре 1.
Фиг. 2 является схематическим представлением консенсусной вторичной структуры 1, представленной нуклеотидной последовательностью аптамера против NGF, приведенной в SEQ ID NO: 12.
Фиг.3 является сенсограммой, на которой показано, что аптамер против NGF, приведенный в SEQ ID NO: 82(2) (модифицированная форма), связывается с NGF, NT-3 и NT-4, где RU по вертикальной оси представляет собой относительную единицу, Resp. Diff. представляет собой различия ответов, и по горизонтальной оси отложено время (секунды) (Время (сек)). Эти обозначения по вертикальной оси и по горизонтальной оси являются одинаковыми для следующих фиг. 4-5.
Фиг. 4 является сенсограммой, на которой показано, что аптамер против NGF, приведенный в SEQ ID NO: 82(2) (модифицированная форма), ингибирует связывание NGF и рецептора NGF TrkA.
Фиг. 5 является сенсограммой, на которой показано, что аптамер NGF, приведенный в SEQ ID NO: 82(2) (модифицированная форма), ингибирует связывание NGF и рецептора NGF p75.
Описание вариантов осуществления
Настоящее изобретение относится к аптамеру, связывающемуся с NGF и способному образовывать предполагаемую вторичную структуру, представленную формулой (I):
где N является одним нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T,
N11-N13, N21-N23, N32-N38 и N42-N48 являются одинаковыми или разными, и каждый из них является связью или 1 или 2 нуклеотидами, выбранными из группы, состоящей из A, G, C, U и T,
N14, N24, N31, N41, N39 и N49 являются одинаковыми или разными, и каждый из них является одним нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T,
каждые из N14 и N24, N31 и N41, и N39 и N49 образуют уотсон-криковскую пару оснований,
N11-N12-N13-N14 и N21-N22-N23-N24 являются нуклеотидными последовательностями, в комбинации способными образовывать структуру стебля, и
N31-N32-N33-N34-N35-N36-N37-N38-N39 и N41-N42-N43-N44-N45-N46-N47-N48-N49 являются нуклеотидными последовательностями, в комбинации способными образовывать структуру стебля (далее в настоящем описании описываемому как "аптамер по настоящему изобретению"). Указанную выше нуклеотидную последовательность, необязательно, модифицируют, как указано ниже.
Аптамер относится к молекуле нуклеиновой кислоты, обладающей связывающей активностью в отношении конкретной молекулы-мишени. Аптамер может ингибировать активность конкретной молекулы-мишени посредством связывания с конкретной молекулой-мишенью. Аптамер по настоящему изобретению является аптамером, обладающим связывающей активностью по отношению к NGF. По предпочтительному варианту осуществления аптамер по настоящему изобретению может ингибировать активность NGF посредством связывания с NGF и ингибирования связывания NGF и рецептора NGF.
Аптамер по настоящему изобретению может являться нуклеиновой кислотой, такой как РНК, ДНК, модифицированная нуклеиновая кислота или их смесь. Таким образом, аптамер по настоящему изобретению в дальнейшем можно обозначать как "нуклеиновая кислота по настоящему изобретению".
Одноцепочечная нуклеиновая кислота может иметь различные вторичные структуры. Термин "предполагаемая вторичная структура" означает вторичную структуру, которую конкретная одноцепочечная нуклеиновая кислота термодинамически может принимать с учетом ее первичной структуры. В частности, предполагаемая вторичная структура аптамера по настоящему изобретению является вторичной структурой, прогнозируемой с использованием программы MFOLD, описываемой в примере 5. Таким образом, даже нуклеиновую кислоту, в настоящий момент не имеющую вторичную структуру, представленную указанной выше формулой (I), включают в аптамер по настоящему изобретению, при условии, что она имеет первичную структуру, способную образовывать указанную вторичную структуру.
Таким образом, предпочтительно, аптамер по настоящему изобретению является молекулой нуклеиновой кислоты, способной иметь вторичную структуру, представленную указанной выше формулой (I), термодинамически стабильную с учетом ее первичной структуры. В этом смысле аптамер по настоящему изобретению является аптамером, способным образовывать предполагаемую вторичную структуру, представленную формулой (I).
Предполагаемая вторичная структура, представленная формулой (I), является тем, что называют "структурой "стебель-петля", в частности, являющейся структурой, имеющей структуру петли (обозначаемую в настоящем описании как "внутренняя петля 1") между структурой стебля, которую может образовывать комбинация N11-N12-N13-N14 и N21-N22-N23-N24, и структурой стебля, которую может образовывать комбинация N31-N32-N33-N34-N35-N36-N37-N38-N39 и N41-N42-N43-N44-N45-N46-N47-N48-N49, и, кроме того, структуру петли между N39 и N49 (обозначаемую в настоящем описании как "петля 2").
"Структура стебля" является структурой, где частичные нуклеотидные последовательности, имеющие комплементарность в молекуле нуклеиновой кислоты, образуют уотсон-криковские пары оснований (G-C или A-U/T). В настоящем описании для N11-N12-N13-N14 и N21-N22-N23-N24, и N31-N32-N33-N34-N35-N36-N37-N38-N39 и N41-N42-N43-N44-N45-N46-N47-N48-N49 не требуется полной комплементарности, и допускают несовпадения и/или вобблинг G-U/T. Т.е. при условии, что нуклеотиды на обоих концах частичной нуклеотидной последовательности, образующие структуру стебля, образуют уотсон-криковские пары оснований, для всех других нуклеотидов не требуется образования уотсон-криковских пар оснований.
В формуле (I) "N", находящийся в участке петли 2, является одним нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T. В предпочтительном варианте осуществления "N" может являться G.
В формуле (I) N11-N13, N21-N23, N32-N38 и N42-N48 являются одинаковыми или разными, и каждый из них является связью или 1 или 2 нуклеотидами, выбранными из группы, состоящей из A, G, C, U и T. Если Ni (i является целым числом, выбранным из 11-13, 21-23, 32-38, 42-48) представляет "два нуклеотида", указанные два нуклеотида могут являться одинаковыми или различными. Если Ni представляет "два нуклеотида" или "связь", он, предпочтительно, содержится в каждой частичной последовательности N11-N13, N21-N23, N32-N38 и N42-N48 в количестве не более 2, более предпочтительно - не более 1. Таким образом, каждая из N11-N14 и N21-N24, образующая одну структуру стебля, предпочтительно, имеет длину 2-6 нуклеотидов, более предпочтительно - 3-5, и каждая из N31-N39 и N41-N49, образующая другую структуру стебля, предпочтительно, имеет длину, предпочтительно, 7-11 нуклеотидов, более предпочтительно - 8-10.
Указанная выше "связь" означает одинарную связь, и если любой Ni в формуле (I) является "связью", это означает, что нуклеотиды, смежные с нуклеотидом, соединены друг с другом посредством фосфодиэфирной связи.
В частности, предпочтительно, N11-N13, N21-N23, N32-N38 и N42-N48 являются одинаковыми или разными, и каждый из них является одним нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T. Таким образом, структура стебля, включая оба конца части, образующей вторичную структуру, имеет длину стебля предпочтительно не более 4 нуклеотидов, и внутренняя структура стебля, находящаяся между двумя петлями, имеет длину стебля предпочтительно 9 нуклеотидов.
В формуле (I) N14, N24, N31, N41, N39 и N49 являются одинаковыми или разными, и каждый из них является одним нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T, и каждый из N14 и N24, N31 и N41, и N39 и N49 образует уотсон-криковскую пару оснований (G-C или A-U/T). Таким образом, структура стебля, содержащая оба конца части, образующей вторичную структуру, образует пару оснований, по меньшей мере, на конце участка внутренней петли 1, и внутренняя структура стебля, находящаяся между двумя петлями, образует пары оснований на обоих его концах. Более предпочтительно, N14 является U, N24 является A, N31 является G, N41 является C, N39 является G, и N49 является C.
С другой стороны, как определено выше, для N11-N12-N13 и N21-N22-N23, и N32-N33-N34-N35-N36-N37-N38 и N42-N43-N44-N45-N46-N47-N48 в "структуре стебля" не требуется полной комплементарности (образование уотсон-криковских пар оснований всеми из них является необязательным). Однако необходима комплементарность на уровне, делающем возможным образование структуры стебля (петля (пузырь) в стебле не образуется). Т.к. петля может образовываться, когда в каждой структуре стебля содержатся 3 непрерывных несовпадения или вобблинга G-U/T, для специфичности каждая структура стебля, желательно, не содержит 3 непрерывных несовпадения или вобблинга G-U/T. Также желательно, чтобы не менее чем 50%, предпочтительно - не менее чем 60%, более предпочтительно - не менее чем 70% каждого из N11-N12-N13 и N21-N22-N23, и N32-N33-N34-N35-N36-N37-N38 и N42-N43-N44-N45-N46-N47-N48 являлись нуклеотидами, образующими уотсон-криковские пары оснований.
Настоящее изобретение также относится к нуклеиновой кислоте, состоящей из нуклеотидной последовательности по следующим пп. (a) или (b):
(a) нуклеотидная последовательность, выбранная из SEQ ID NO: 3, SEQ ID NO: 9-13, SEQ ID NO: 22-117 и SEQ ID NO: 152-168 (где урацил может являться тимином);
(b) нуклеотидная последовательность по предшествующему п. (a) и связывающаяся с NGF, где от 1 до нескольких нуклеотидов подвергают замене, делеции, инсерции или добавлению.
Такие нуклеиновые кислоты могут образовывать предполагаемую вторичную структуру, представленную указанной выше формулой (I).
Т.к. любой урацил в любой последовательности можно заменять тимином, подлежащий замене урацил, предпочтительно, может являться урацилом в части, иной, чем участок внутренней петли 1 и участок петли 2 в указанной выше предполагаемой вторичной структуре, таким образом, что можно поддерживать активность аптамера по настоящему изобретению.
В настоящем описании последовательность, определяемая посредством "SEQ ID NO", означает нуклеотидную последовательность каждого аптамера или нуклеиновой кислоты, и, например, "нуклеиновая кислота, содержащая последовательность, приведенную в SEQ ID NO: 1" означает природную нуклеиновую кислоту или модифицированную нуклеиновую кислоту, содержащую последовательность, приведенную в SEQ ID NO: 1, или нуклеиновую кислоту, составленную из них обоих. Последовательность оснований SEQ ID NO каждого аптамера приводят в списке последовательностей.
В указанном выше (b) количество нуклеотидов, подвергнутых замене, делеции, инсерции или добавлению, составляет например, приблизительно 1-10, предпочтительно - 1-6, более предпочтительно 1-5, более предпочтительно - 1-3, наиболее предпочтительно - 1 или 2.
В указанном выше (b), хотя положение нуклеотида, подлежащего замене, делеции, инсерции или добавлению, в частности, не ограничено, предпочтительно, нуклеотид может находиться в части, иной, чем участок внутренней петли 1 и участок петли 2 в указанной выше предполагаемой вторичной структуре, таким образом, что можно поддерживать активность аптамера по настоящему изобретению.
Хотя длина аптамера или нуклеиновой кислоты по настоящему изобретению, в частности, не ограничена, она, как правило, составляет от 34 до приблизительно 200 нуклеотидов, предпочтительно - от 34 до приблизительно 100 нуклеотидов, более предпочтительно - 36-60 нуклеотидов, более предпочтительно - 38-44 нуклеотидов. Длина аптамера или нуклеиновой кислоты по настоящему изобретению, предпочтительно, составляет не более 50, более предпочтительно - не более 44. Химический синтез и массовое производство аптамера упрощаются при снижении общего количества нуклеотидов, попадающего в диапазон, делающий возможным образование предполагаемой вторичной структуры, представленной указанной выше формулой (I), а также существует основное преимущество в отношении затрат. Также считают, что такой аптамер делает возможной легкую химическую модификацию, высокую стабильность в организме и низкую токсичность.
Аптамер по настоящему изобретению также может являться конъюгатом, выбранным из группы, состоящей из конъюгата множества нуклеиновых кислот, состоящих из нуклеотидной последовательности по указанному выше п. (a), конъюгата множества нуклеиновых кислот, состоящих из нуклеотидной последовательности по указанному выше п. (b), и конъюгата множества нуклеиновых кислот, состоящих из нуклеотидной последовательности по указанному выше п. (a), и нуклеиновых кислот, состоящих из нуклеотидной последовательности по указанному выше п. (b).
Эти конъюгаты также могут связываться с NGF и/или ингибировать активность NGF (активность связывания рецептора NGF и т.д.).
В настоящем описании конъюгации можно достигать посредством тандемного связывания. В конъюгации можно использовать линкер. В качестве линкера можно упомянуть нуклеотидные цепи (например, от 1 до приблизительно 20 нуклеотидов) и ненуклеотидные цепи (например, линкер -(CH2)n-, линкер -(CH2CH2O)n-, гексаэтиленгликолевый линкер, линкер TEG, пептид-содержащий линкер, линкер, содержащий связь -S-S-, линкер, содержащий связь -CONH-, линкер, содержащий связь -OPO3-). Множество, как указано для описываемого выше конъюгата множества, в частности, не ограничено при условии, что оно составляет два или более, и множество может составлять, например, 2, 3 или 4.
Каждый нуклеотид, содержащийся в аптамере по настоящему изобретению, является одинаковым или разным и может являться нуклеотидом, содержащим гидроксильную группу в 2’-положении рибозы (например, рибозы пиримидинового нуклеотида, рибозы пуринового нуклеотида) (т.е. незамещенным нуклеотидом), или нуклеотидом, где гидроксильную группу замещают любым атомом или группой в 2ʹ-положении рибозы. В качестве примеров любого такого атома или группы можно упомянуть нуклеотид, замещенный атомом водорода, атомом фтора или -O-алкильной группой (например, группой -O-Me), -O-ацильной группой (например, группой -O-CHO) или аминогруппой (например, группой -NH2). В следующих случаях гидроксильную группу замещают атомом водорода, атомом фтора или группой -O-Me, соответственно, в 2’-положении рибозы.
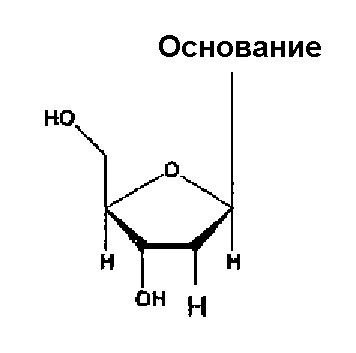
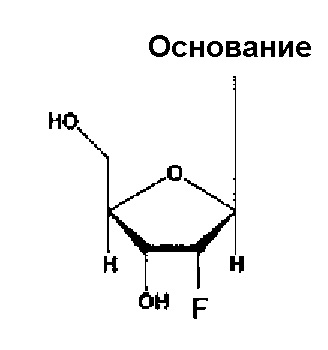
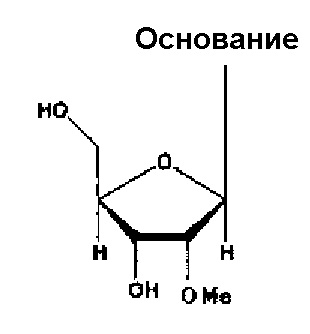
Аптамер по настоящему изобретению также может являться нуклеотидом, где, по меньшей мере, один тип (например, 1, 2, 3 или 4 типа) нуклеотидов содержит гидроксильную группу или описываемый выше любой атом или группу, например, по меньшей мере, два типа (например, 2, 3 или 4 типа) групп, выбранных из группы, состоящей из атома водорода, атома фтора, гидроксильной группы и группы -O-Me, в 2ʹ-положении рибозы.
Кроме того, в аптамере по настоящему изобретению все пиримидиновые нуклеотиды являются одинаковыми или разными, и каждый из них может являться нуклеотидом, замещенным атомом фтора, или нуклеотидом, замещенным любым указанным выше атомом или группой, предпочтительно, атомом или группой, выбранной из группы, состоящей из атома водорода, гидроксильной группы и метоксигруппы в 2ʹ-положении рибозы.
Кроме того, в аптамерах по настоящему изобретению все пуриновые нуклеотиды являются одинаковыми или разными, и каждый из них может являться нуклеотидом, замещенным гидроксильной группой, или нуклеотидом, замещенным любым указанным выше атомом или группой, предпочтительно, атомом или группой, выбранной из группы, состоящей из атома водорода, метоксигруппы и атома фтора в 2’-положении рибозы.
Кроме того, в аптамерах по настоящему изобретению все нуклеотиды могут содержать гидроксильную группу или любой указанный выше атом или группу, например, идентичную группу, выбранную из группы, состоящей из атома водорода, атома фтора, гидроксильной группы и группы -O-Me в 2ʹ-положении рибозы.
В настоящем описании при описании модификации групп сахаров в нуклеотиды нуклеотиды, из которых состоит аптамер, считают РНК (т.е. группы сахаров считают рибозой). Однако это не означает, что ДНК исключают из нуклеотидов, из которых состоит аптамер, и при необходимости модификацию РНК необходимо рассматривать как модификацию ДНК. Если нуклеотид, из которого состоит аптамер, является ДНК, например, замену гидроксильной группы в 2ʹ-положении рибозы на X необходимо рассматривать как замену одного атома водорода в 2’-положении дезоксирибозы на X.
Если в аптамере по настоящему изобретению урацил замещают тимином, можно повышать активность связывания NGF, активность ингибирования связывания NGF и рецептора NGF, ингибирования активности NGF в отношении роста нейритов, ингибирования активности NGF в отношении пролиферации клеток, стабильность, доставляемость лекарственного средства и стабильность аптамера в крови и т.п.
Аптамер по настоящему изобретению связывается с NGF, являющимся известным нейротрофином и важным секреторным белком, участвующим в развитии и выживании периферических и центральных нейронов. В частности, в настоящем изобретении NGF означает β-тип NGF. Аминокислотные последовательности β-NGF человека являются теми, которые представлены под регистрационными номерами NP002497, P01138, AAI26151, AAI26149 и CAB75625, которые также могут содержать мутацию, его функциональным доменом или пептидным фрагментом. Он может являться не только мономером, но также и димером или мультимером. Кроме того, он включает NGF, получаемым не являющихся человеком млекопитающих, например, приматов (например, обезьян), грызунов (например, мыши, крысы, морской свинки) и животных-компаньонов, домашних животных и рабочих животных (например, собаки, кошки, лошади, коровы, козы, овцы, свиньи).
Аптамер по настоящему изобретению ингибирует активность NGF посредством связывания с NGF и ингибирования связывания NGF и рецептора NGF. Аптамер по настоящему изобретению может связываться с любой частью NGF при условии, что можно ингибировать связывание NGF и рецептора NGF.
В настоящем описании "ингибиторная активность в отношении NGF" означает способность к ингибированию любой активности NGF. Например, это означает активность в отношении ингибирования связывания NGF с рецептором NGF, ингибирования передачи сигнала ниже рецептора NGF (через путь киназ Ras-MAP, путь киназы PI3), ингибирования повышения экспрессии TRPV1, SP, BDNF и т.п., активность ингибирования экспрессии HA, BK, PG, NGF и других цитокинов, высвобождаемых из тучных клеток и т.п., что является результатом связывания NGF с рецептором NGF, кроме того, можно отметить ингибирование дифференцировки, выживания, роста нейритов нейрона, индуцируемого NGF, повышения проницаемости кровеносных сосудов, повышения иммунного ответа T-клеток и B-клеток, дифференцировки лимфоцитов, роста и т.п. различных клеток, таких как тучные клетки, эритролейкозные клетки, злокачественные клетки и т.п., снижение боли, гипералгезии и т.п., индуцируемых NGF.
Предпочтительная "ингибиторная активность в отношении NGF", которой обладает аптамер по настоящему изобретению, является активность ингибирования связывания NGF с рецептором NGF, активность ингибирования роста нейритов, индуцируемого NGF, активность ингибирования пролиферации клеток, индуцируемой NGF и т.п.
Аптамер по настоящему изобретению связывается с NGF в физиологическом буфере (например, растворе A: см. пример 1). Аптамер по настоящему изобретению связывается, например, с NGF с интенсивностью, определяемой с помощью следующего теста.
Для измерения используют BIAcore2000, производимый BIAcore. Аптамер иммобилизуют на сенсорном чипе. Количество, подлежащее иммобилизации, устанавливают на 1000 RU. Для получения раствора NGF (0,5 мкМ) используют физиологический буфер, содержащий 0,3M NaCl (раствор A: см. пример 1). Этот раствор NGF (20 мкл) инъецируют и определяют связывание NGF с аптамером. Используя РНК, содержащую случайный нуклеотид, состоящий из 40 нуклеотидов, в качестве отрицательного контроля, когда NGF связывается с аптамером, значимо сильнее по сравнению с контрольной РНК, оценивают способность аптамера к связыванию с NGF.
В настоящем описании термин "рецептор NGF" означает белок поверхности клетки, с которой связывается NGF. TrkA и p75 известны как рецепторы NGF. Рецептор NGF, указанный в настоящем изобретении, может являться белком, содержащим природную аминокислотную последовательность или ее вариант. В настоящем описании термин "ее вариант" означает белок или пептид, где несколько аминокислот в аминокислотной последовательности "рецептора NGF" замещены, или его частичную аминокислотную последовательность, имеющую активность связывания с NGF и ингибирующую связывание NGF и рецептора NGF.
Аптамер по настоящему изобретению связывается с NGF и ингибирует связывание NGF и рецептора NGF. Ингибирует ли аптамер связывание NGF с рецептором NGF или нет, например, можно оценивать с помощью следующего теста.
Для измерения используют BIAcore2000, производимый BIAcore. На сенсорном чипе CM5 иммобилизуют слитный белок рецептора NGF и Fc (например, TrkA-Fc (175-TK, R&D systems) или p75-Fc (R&D systems)). Количество, подлежащее иммобилизации, составляет от 500 до 700 RU. NGF (0,1 мкМ) и аптамера (0,2 мкМ) смешивают в физиологическом буфере (раствор A: см. пример 1), и в течение 30 мин получают смесь, являющуюся образцом. Эту смесь инъецируют в BIAcore2000 и определяют связывание NGF с рецептором NGF.
В одном из вариантов осуществления аптамер по настоящему изобретению может ингибировать и связывание NGF и TrkA, и связывания NGF и p75.
Аптамер по настоящему изобретению является аптамером, ингибирующим активность NGF в отношении роста нейритов и/или активность NGF в отношении пролиферации клеток. Может ли аптамер по настоящему изобретению ингибировать активность NGF в отношении роста нейритов или нет, можно оценивать, например, с помощью теста, описываемого в примере 3. Кроме того, может ли аптамер по настоящему изобретению ингибировать активность NGF в отношении пролиферации клеток или нет, можно оценивать, например, с помощью теста, описываемого в примере 8.
Аптамер по настоящему изобретению характеризуют по концентрации, необходимой для ингибирования активности NGF в отношении роста нейритов на 50% (IC50 или 50%-ная ингибиторная концентрация), составляющей не более 1 нМ. Т.к. общеизвестный аптамер против NGF имеет значение IC50 приблизительно несколько нМ в случае активности NGF в отношении роста нейритов, аптамер по настоящему изобретению демонстрирует лучшее свойство в отношении активности ингибирования роста нейритов.
В предпочтительном варианте осуществления аптамер по настоящему изобретению также демонстрирует значение IC50 не более 1 нМ в случае активности NGF в отношении пролиферации клеток.
С другой стороны, имеет ли аптамер по настоящему изобретению активность ингибирования активности нейротрофина, иного, чем NGF, в частности, нейротрофического фактора мозга (BDNF), нейротрофина-3 (NT-3) и нейротрофина 4/5 (NT-4/5), в отношении пролиферации клеток, варьируется в зависимости от аптамера. В настоящем описании термины BDNF, NT-3 и NT-4/5, соответственно, означают BDNF, NT-3 и NT-4/5 всех видов млекопитающих, включая человека.
Уровень ингибиторной активности других нейротрофинов (BDNF, NT-3, NT-4/5) в отношении пролиферации клеток можно оценивать с помощью теста, описываемого в примере 16. Ингибиторная активность аптамера по настоящему изобретению в отношении пролиферации клеток, как описано в примере 16 и таблице 2, представлена значением IC50 не более 0,1 нМ в случае NGF и не менее 1000 нМ в случае BDNF, что означает, что аптамер по настоящему изобретению не ингибирует активность BDNF в отношении пролиферации клеток. Однако она составляет от 0,97 нМ до не менее 10 нМ в случае NT-3; и от не более 3 нМ до не менее 30 нМ в случае NT-4. Таким образом, ингибирование активности NT-3 и NT-4 в отношении пролиферации клеток варьируется в зависимости от аптамера.
Аптамер по настоящему изобретению может являться аптамером, где остаток сахара (например, рибозы) в каждом нуклеотиде модифицируют для повышения активности связывания с NGF, активности ингибирования связывания NGF и рецептора NGF, ингибирования активности NGF в отношении роста нейритов, ингибирования активности NGF в отношении пролиферации клеток, стабильности, доставляемости лекарственного средства и стабильности аптамера в крови и т.п. Примеры модификации остатка сахара включают замещение атома кислорода в 2’-положении, 3’-положении и/или 4’-положении остатка сахара другим атомом и т.п. В качестве типа модификации можно упомянуть фторирование, O-алкилирование (например, O-метилирование, O-этилирование), O-арилирование, S-алкилирование (например, S-метилирование, S-этилирование), S-арилирование и аминирование (например, -NH2). Кроме того, ее примеры включают 4’-SRNA, где кислород в 4’-положении замещают серой, ЗНК (замкнутую нуклеиновую кислоту), где 2’-положение и 4’-положение поперечно сшивают с помощью метилена, 3’-N-фосфоамидатную нуклеиновую кислоту, где гидроксильную группу в 3’-положении замещают аминогруппой и т.п. Такие изменения в остатке сахара можно осуществлять способом, известным per se (см., например, Sproat et al., (1991) Nucl. Acid. Res. 19, 733-738; Cotton et al., (1991) Nucl. Acid. Res. 19, 2629-2635; Hobbs et al., (1973) 12 Biochemistry 5138-5145).
Аптамер по настоящему изобретению также может содержать изменения (например, химическое замещение) оснований нуклеиновой кислоты (например, пурина или пиримидина) для повышения активности связывания NGF, активности ингибирования связывания NGF и рецептора NGF, активности ингибирования NGF в отношении роста нейритов, активности ингибирования NGF в отношении пролиферации клеток, стабильности, доставляемости лекарственного средства и стабильности аптамера в крови и т.п. В качестве примеров таких изменений можно упомянуть изменение пиримидина в 5-положении, изменение пурина в 6- и/или 8-положениях (O-метильную модификацию и т.п.), изменение с использованием экстрациклического амина, замещение 4-тиоуридином, замещение 5-бромо- или 5-иодоурацилом, модификацию 5-аминокислоты и модификацию боковой цепи 5-триптофана. Фосфатную группу, содержащуюся в аптамере по настоящему изобретению, можно изменять для придания устойчивости к действию нуклеаз и гидролизу. Например, фосфатную область аптамера можно замещать P(O)S (тиоатом), P(S)S (дитиоатом), P(O)NR2 (амидатом), P(O)R, P(O)OR’, CO или CH2 (формацеталем), P(O)BH3 (боранофосфатом) или 3’-амином (-NH-CH2-CH2-) [где каждая единица R или R’ независимо является H или замещенным или незамещенным алкилом (например, метилом, этилом)].
Соединительная группа является, например, -O-, -N- или -S-, и нуклеотиды можно соединять с соседним нуклеотидом через эти соединительные группы.
Изменения также могут включать изменения, такие как кэпирование на 3’- и 5’-конце.
Изменение дополнительно можно осуществлять, добавляя с конца полиэтиленгликоль (далее в настоящем описании иногда описываемый как "PEG"), аминокислоту, пептид, инвертированный dT, миристоил, литохолевый олеил, докозанил, лауроил, стеароил, пальмитоил, олеоил, линолеоил, другие липиды, стероиды, холестерол, кофеин, витамины, пигменты, флуоресцентные вещества, противораковое средство, токсин, ферменты, радиоактивные вещества, биотин и т.п. Такие изменения, см., например, в патентах США №№ 5660985 и 5756703.
В частности, если изменение осуществляют добавлением PEG с конца, молекулярная масса PEG, в частности, не ограничена и составляет, предпочтительно, 1000-100000, более предпочтительно - 30000-90000. PEG может являться линейным или разветвленным с двумя или более цепями (многолучевой PEG).
Такой PEG, в частности, не ограничен, и специалисты в этой области соответствующим образом могут выбирать и использовать коммерчески доступный или известный PEG (например, http://www.peg-drug.com/peg_product/branched.html). Конкретные предпочтительные примеры PEG для использования в аптамере по настоящему изобретению включают 2-разветвленный PEG типа GS, имеющий молекулярную массу 40000 (SUNBRIGHT GL2-400GS2, производимый NOF CORPORATION), 2-разветвленный PEG типа TS, имеющий молекулярную массу 40000 (SUNBRIGHT GL2-400TS, производимый NOF CORPORATION), 4-разветвленный PEG типа TS, имеющий молекулярную массу 40000 (SUNBRIGHT GL4-400TS, производимый NOF CORPORATION), 2-разветвленный PEG типа TS, имеющий молекулярную массу 80000 (SUNBRIGHT GL2-800TS, производимый NOF CORPORATION), 4-разветвленный PEG типа TS, имеющий молекулярную массу 80000 (SUNBRIGHT GL4-800TS, производимый NOF CORPORATION) и т.п.
В этом случае в аптамере по настоящему изобретению PEG можно добавлять непосредственно с конца. Более предпочтительно добавлять линкер, имеющий группу, связывающуюся с PEG и т.п., на его конец, и добавлять PEG к аптамеру по настоящему изобретению через линкер.
Линкер для PEG и аптамера по настоящему изобретению, в частности, не ограничен, и количество углеродных цепей, функциональные группы и т.п. можно соответствующим образом выбирать, учитывая участок связывания, тип PEG и т.п. Примеры такого линкера включают линкер, имеющий аминогруппу. В частности, в случае добавления с 5'-конца можно упомянуть ssH Linker (SAFC) или DMS(O)MT-AMINO-MODIFIER (GLEN RESEARCH), и в случае добавления с 3'-конца можно упомянуть TFA Amino C-6 lcaa CPG (ChemGenes) и т.п. При выборе этого линкера, например, активную группу N-гидроксисукцинимида добавляют к PEG и проводят реакцию с аминогруппой со стороны линкера, посредством чего аптамер по настоящему изобретению можно соединять с PEG через линкер.
В качестве PEG и линкера предпочтительно использовать коммерчески доступные продукты. Специалисты в этой области могут соответствующим образом определять условия реакции и т.п. для связывания PEG, линкера и аптамера по настоящему изобретению.
Аптамер по настоящему изобретению можно синтезировать химически, как представлено в настоящем описании, и с помощью способа, известного per se в этой области. Аптамер связывается с веществом-мишенью множеством способов связывания, например, с помощью ионных связей с учетом отрицательного заряда фосфатной группы, гидрофобных связей и водородных связей, обусловленных рибозой, и водородных связей и стэкинг-взаимодействий, обусловленных основаниями нуклеиновых кислот. В частности, ионные связи, обусловленные отрицательным зарядом фосфатной группы, присутствующие в том же количестве, что и образующие их нуклеотиды, являются сильными и образуются благодаря положительному заряду лизина и аргинина, присутствующих на поверхности белка. По этой причине можно замещать основания нуклеиновой кислоты, не участвующие в непосредственном связывании с веществом-мишенью. В частности, т.к. участок структуры стебля уже образовал пары оснований и обращен к внутренней части двойной винтовой структуры, основания нуклеиновой кислоты, вероятно, не связываются напрямую с веществом-мишенью. Таким образом, даже если пару оснований замещают другой парой оснований, активность аптамера часто не снижается. В структурах, где не образуются пары оснований, такие как структуры петли, при условии, что основание нуклеиновой кислоты не участвует в прямом связывании с молекулой-мишенью, возможна замена оснований. Что касается модификаций 2’-положения рибозы, функциональная группа в 2’-положении рибозы редко взаимодействует напрямую с молекулой-мишенью, но во многих случаях это не имеет значения, и ее можно замещать другой модифицированной молекулой. Таким образом, если функциональную группу, участвующую в прямом связывании с молекулой-мишенью не подвергают замене или делеции, аптамер часто сохраняет свою активность. Также важно, что общая трехмерная структура не изменяется значительно.
Аптамер можно получать с использованием способа SELEX или его улучшенной версии (например, Ellington et al., (1990) Nature, 346, 818-822; Tuerk et al., (1990) Science, 249, 505-510). В способе SELEX при повышении количества раундов или использовании конкурентного вещества концентрируют и выбирают аптамер, проявляющий больший потенциал связывания для молекулы-мишени. Таким образом, корректируя количество раундов SELEX и/или изменяя условия конкуренции, в некоторых случаях можно получать аптамеры с различной силой связывания, аптамеры с различными способами связывания и аптамеры с одинаковой силой связывания или способом связывания, но разными последовательностями оснований. Способ SELEX включает амплификацию с помощью ПЦР; вызывая мутацию с использованием ионов магния и т.п., можно осуществлять SELEX с более высоким разнообразием.
Аптамеры, получаемые с помощью SELEX, являются нуклеиновыми кислотами, проявляющими высокую аффинность для вещества-мишени, но это не означает связывание с активным центром вещества-мишени. Таким образом, аптамеры, получаемые с помощью SELEX, необязательно действуют на функцию вещества-мишени. NGF является основным белком, и считают, что он, вероятно, позволяет нуклеиновым кислотам связываться с ним неспецифически. Аптамер, не связывающийся с активным центром, не влияет на активность вещества-мишени.
Таким образом, учитывая выбранный активный аптамер, можно осуществлять SELEX, основываясь на последовательности полученного аптамера для получения аптамера, обладающего большей активностью. В частности, после получения матрицы, где аптамер с определенной последовательностью частично рандомизирован, или матрицы дополнены приблизительно от 10 до 30% случайных последовательностей, снова осуществляют SELEX.
Аптамер, получаемый с помощью SELEX, имеет длину приблизительно 80 нуклеотидов, и его трудно получать в качестве лекарственного средства в таком виде. Таким образом, необходимо повторять попытки методом проб и ошибок для укорачивания аптамера до длины приблизительно 60 нуклеотидов или менее, делающей возможным легкий химический синтез, более предпочтительно - приблизительно 50 нуклеотидов или менее, наиболее предпочтительно - 45 нуклеотидов или менее. В зависимости от дизайна праймеров для аптамера, получаемого с помощью SELEX, упрощается последующая минимизация операций. Если праймер сконструирован неудачно, последующая разработка будет невозможна, даже если аптамер с активностью выбран с помощью SELEX.
Аптамеры легко изменяют, т.к. они делают возможным химический синтез. В случае аптамеров, прогнозируя вторичную структуру с использованием программы MFOLD или прогнозируя стерическую структуру посредством рентгеноструктурного анализа или анализа ЯМР, можно до некоторой степени прогнозировать, какой нуклеотид можно подвергать замещению или делеции и куда встраивать новый нуклеотид. Прогнозируемый аптамер с новой последовательностью можно легко синтезировать химически, и с использованием существующей аналитической системы можно определять, сохранит ли аптамер активность или нет.
Если область, важную для связывания получаемого аптамера с веществом-мишенью, идентифицируют с помощью повторных попыток методом проб и ошибок, как описано выше, активность во многих случаях остается неизмененной, даже если новую последовательность добавляют с обоих концов последовательности. В частности, такая длина новой последовательности не ограничена.
Модификации, как и последовательности, обеспечивают широту дизайна или изменений.
Как указано выше, аптамеры делают возможным широту дизайна или изменений. Настоящее изобретение также относится к способу получения аптамера, делающего возможным широту дизайна или изменения аптамера, содержащего конкретную последовательность (например, последовательность, соответствующую части, выбранной из участка стебля, участка внутренней петли, участка шпильки петли и одноцепочечного участка: далее в настоящем описании при необходимости сокращаемую как фиксированная последовательность).
Например, способ получения такого аптамера включает получение аптамера, содержащего фиксированную последовательность, с использованием одного типа молекулы нуклеиновой кислоты, состоящей из нуклеотидной последовательности, представленной следующим образом:
Последовательность праймера (i)-(N)a-фиксированная последовательность-(N)b-последовательность праймера (ii),
где (N)a представляет собой цепь нуклеотидов, состоящую из "a" единиц N; (N)b представляет собой цепь нуклеотидов, состоящую из "b" единиц N; каждая из единиц N, являющаяся идентичной или различной, является нуклеотидом, выбранным из группы, состоящей из A, G, C, U и T (предпочтительно, A, G, C и U). Каждый из "a" и "b", являющийся идентичной или различной, может являться любым числом и может составлять, например, от 1 до приблизительно 100, предпочтительно - от 1 до приблизительно 50, более предпочтительно - от 1 до приблизительно 30, еще более предпочтительно - от 1 до приблизительно 20 или от 1 до приблизительно 10], или многих типов молекул нуклеиновой кислоты (например, библиотеки молекул нуклеиновой кислоты, различающихся по числу a, b и т.д.) и пар праймеров, соответствующих последовательностям праймеров (i) и (ii), соответственно.
Аптамер по настоящему изобретению, предпочтительно, является аптамером, связывающимся с NGF, как правило, содержит последовательность, приведенную в SEQ ID NO: 82, и имеет длину не более 50 нуклеотидов.
Последовательность, приведенная в SEQ ID NO: 82, является областью, важной для аптамера по настоящему изобретению для его функционирования в качестве аптамера по настоящему изобретению, такого как связывание с NGF и ингибирование активности NGF, в частности, активности в отношении роста нейритов, активности в отношении пролиферации клеток и т.п. Даже если новую последовательность добавляют с обоих концов последовательности, функционирование аптамера по настоящему изобретению не нарушается. Последовательность можно подвергать модификации указанного выше остатка сахара, изменению основания и фосфатной группы нуклеиновой кислоты и т.п.
Таким образом, конкретные предпочтительные примеры аптамера по настоящему изобретению включают
аптамеры, содержащие последовательность, приведенную в SEQ ID NO: 82, имеющие длину не более 50 нуклеотидов, и связывающиеся с NGF, являющиеся
(i) аптамером, содержащим, по меньшей мере, один тип нуклеотидов, где гидроксильную группу замещают атомом водорода, атомом фтора, -O-алкильной группой, -O-ацильной группой или аминогруппой в 2’-положении рибозы;
(ii) аптамером, где с конца добавляют PEG, аминокислоту, пептид, инвертированный dT, миристоил, литохолевый олеил, докозанил, лауроил, стеароил, пальмитоил, олеоил, линолеоил, другой липид, стероид, холестерол, кофеин, витамин, краситель, флуоресцентное вещество, противоопухолевое средство, токсин, фермент, радиоактивное вещество или биотин;
(iii) аптамером, удовлетворяющим условиям (i) и (ii);
и т.п.
Настоящее изобретение также относится к комплексу, содержащему аптамер по настоящему изобретению и связанное с ним функциональное вещество. Связь между аптамером и функциональным веществом в комплексе по настоящему изобретению может являться ковалентной связью или нековалентной связью. Комплекс по настоящему изобретению может являться комплексом, в котором соединены аптамер по настоящему изобретению и одно или несколько (например, 2 или 3) функциональных веществ одного типа или различных типов. В частности, функциональное вещество не ограничено до той степени, пока оно придает аптамеру по настоящему изобретению конкретную новую функцию или способно изменять (например, улучшать) конкретное характерное свойство, которым аптамер по настоящему изобретению может обладать. В качестве примеров функционального вещества можно упомянуть белки, пептиды, аминокислоты, липиды, сахара, моносахариды, полинуклеотиды и нуклеотиды. В качестве примеров функционального вещества можно упомянуть вещества с высоким сродством (например, биотин, стрептавидин, полинуклеотиды, обладающие аффинностью для комплементарной последовательности-мишени, антитела, глутатион-сефарозу, гистидин), вещества для мечения (например, флуоресцентные вещества, люминесцентные вещества, радиоактивные изотопы), ферменты (например, пероксидазу хрена, щелочную фосфатазу), средства доставки лекарственных веществ (например, липосому, микросферы, пептиды, полиэтиленгликоли), лекарственные средства (например, используемые в реактивной терапии, такие как калихимицин и дуокармицин; аналоги азотистого иприта, такие как циклофосфамид, мелфалан, ифосфамид или трофосфамид; этиленимины, такие как тиотепа; нитрозомочевину, такую как кармустин; алкилирующие средства, такие как темозоломид или дакарбазин; антиметаболитов-антагонистов фолиевой кислоты, такие как метотрексат или ралтитрексед; аналоги пурина, такие как тиогуанин, кладрибин или флударабин; аналоги пиримидина, такие как фторурацил, тегафур или гемцитабин; алкалоиды барвинка, такие как винбластин, винкристин или винорелбин и их аналоги; производные подофиллотоксина, такие как этопозид, таксаны, доцетаксел или паклитаксел; антрациклины, такие как доксорубицин, эпирубицин, идарубицин и митоксантрон и их аналоги; другие цитотоксические антибиотики, такие как блеомицин и митомицин; соединения платины, такие как цисплатин, карбоплатин и оксалиплатин; пентостатин, милтефозин, эстрамустин, топотекан, иринотекан и бикалутамид) и токсины (например, рицин, лиатоксин и веротоксин). В некоторых случаях эти функциональные молекулы, в конечном итоге, удаляют. Кроме того, молекулы могут являться пептидами, которые могут распознаваться и расщепляться ферментами, такими как тромбин, матричная металлопротеиназа (MMP) и фактор X, и могут являться полинуклеотидами, которые могут расщепляться нуклеазами или эндонуклеазами рестрикции.
Аптамер или комплекс по настоящему изобретению можно использовать, например, в качестве лекарственного средства или средства для диагностики, тестового лекарственного средства, реагента, добавки для питьевой воды и пищи, энхансера и ограничителя.
Аптамер и комплекс по настоящему изобретению может иметь активность ингибирования функции NGF посредством связывания с NGF и ингибирования связывания NGF и рецептора NGF. Как указано выше, NGF в значительной степени участвует в развитии болевой реакции и воспаления. Таким образом, аптамер и комплекс по настоящему изобретению применимы в качестве лекарственных средств для профилактики или лечения заболеваний, сопровождающихся болью или воспалением (средства против боли, противовоспалительного средства и т.д.).
В настоящем описании примеры боли включают ноцицептивную боль (мышечную боль, позвоночную боль, боль в верхней конечности, хлыстовую травму, артралгию, остеоартрит, подагру, ревматоидный артрит, головную боль, мигрень, кататоническую головную боль, кластерную головную боль, вторичную головную боль, орофациальную боль, зубную боль, каузалгию после удаления зуба, фантомную зубную боль, боль в органе, боль в сердце, боль в области живота, боль при овуляции, дисменорею, родовые схватки, нефралгию, боль в мочеточнике, боль в костях и т.п.), воспалительную боль, нейропатическую боль (диабетическую невропатию, токсическую нейропатию, послеоперационную боль, фантомную боль в конечностях, боль при ампутации, симпатическую рефлекторную дистрофию, каузалгию, постгерпетическую боль, невралгию тройничного нерва, центральную боль), карциноматозную боль (боль в результате опухолевой инфильтрации внутреннего органа, боль, вызываемая обструкцией кровеносного сосуда в результате инфильтрации кровеносного сосуда опухолевой ткани, боль от метастазов в костях, боль, ассоциированная с интрацеребральными метастазами, боль, вызываемая инфильтрацией периферических нервов опухолевой ткани), фибромиалгию и т.п.
Хотя в настоящем описании заболевание, ассоциированное с воспалением, в частности, не ограничено, можно упомянуть системную красную волчанку, рассеянный склероз, псориаз, остеоартрит, ревматоидный артрит, интерстициальный цистит, астму и т.п.
Хотя указанное выше злокачественное новообразование, в частности, не ограничено, можно упомянуть рак пищевода, рак щитовидной железы, рак мочевого пузыря, колоректальный рак, рак желудка, рак поджелудочной железы, злокачественное новообразование в грудной клетке, рак печени, рак легких, немелкоклеточный рак легких, рак молочной железы, нейробластому, глиобластому, рак матки, рак шейки матки, рак яичников, опухоль Вильмса, рак предстательной железы и т.п.
При связывании NGF с его рецептором TrkA он активирует фосфорилирование тирозина TrkA и Ras-MAPK, PLC-γ, PI3K и т.п. ниже TrkA, и проявляет свою физиологическую функцию, такую как выживание и дифференцировка нейронов. С другой стороны, он индуцирует гибель клеток через сигнальный путь рецептора p75. Таким образом, аптамер и комплекс по настоящему изобретению можно использовать в качестве лекарственных средств, средств для диагностики, тестовых лекарственных средств или реагентов для заболевания, имеющего отношение к активации этих путей передачи сигнала. Примеры заболеваний, имеющих отношение к активации этих путей передачи сигнала, включают указанную выше алгию, воспалительное заболевание и злокачественные новообразования.
Если аптамер и комплекс по настоящему изобретению используют в качестве лекарственных средств, средств для диагностики, тестовых лекарственных средств, реагентов и т.п., субъект, которому их вводят, в частности, не ограничен, и можно упомянуть, например, приматов (например, человека, обезьян), грызунов (например, мышь, крысу, морскую свинку), и животных-компаньонов, домашних животных и рабочих животных (например, собаку, кошку, лошадь, крупный рогатый скот, козу, овцу, свинью).
Аптамер и комплекс по настоящему изобретению способны специфически связываться с NGF. Таким образом, аптамер и комплекс по настоящему изобретению применимы в качестве зондов для определения NGF. Зонды применимы в визуализации NGF in vivo, измерениях концентраций в крови, окрашивании тканей, ELISA и т.п. Зонды также применимы в качестве средств для диагностики, тестовых лекарственных средств, реагентов и т.п. для заболеваний, ассоциированных с NGF (заболеваний, сопровождающихся болью или воспалением и т.п.).
Учитывая их специфическое связывание с NGF, аптамер и комплекс по настоящему изобретению можно использовать в качестве лигандов для разделения и очистки NGF.
Кроме того, аптамер и комплекс по настоящему изобретению можно использовать в качестве тестовых лекарственных средств для исследования психического состояния при влюбленности и т.п. или лекарственных средств, регуляторов, энхансеров или ограничитель для контроля психического состояния.
Аптамер и комплекс по настоящему изобретению можно использовать в качестве средств для доставки лекарственных средств.
Лекарственное средство по настоящему изобретению может являться лекарственным средством, составляемым с фармацевтически приемлемым носителем. В качестве примеров фармацевтически приемлемого носителя можно упомянуть эксципиенты, такие как сахароза, крахмал, маннит, сорбит, лактоза, глюкоза, целлюлоза, тальк, фосфат кальция и карбонат кальция; связывающие средства, такие как целлюлоза, метилцеллюлоза, гидроксилпропилцеллюлоза, полипропилпирролидон, желатин, гуммиарабик, полиэтиленгликоль, сахароза и крахмал; разрыхлители, такие как крахмал, карбоксиметилцеллюлоза, гидроксилпропилкрахмал, натрий-гликоль-крахмал, бикарбонат натрия, фосфат кальция и цитрат кальция; смазочные средства, такие как стеарат магния, аэросил, тальк и лаурилсульфат натрия; ароматизаторы, такие как лимонная кислота, ментол, глицирризининат аммония, глицин и порошкообразный ароматизатор апельсина; консерванты, такие как бензоат натрия, гидросульфит натрия, метилпарабен и пропилпарабен; стабилизаторы, такие как лимонная кислота, цитрат натрия и уксусная кислота; суспендирующие средства, такие как метилцеллюлоза, поливинилпирролидон и стеарат алюминия; диспергирующие средства, такие как поверхностно-активные вещества; разбавители, такие как вода, физиологический раствор и апельсиновый сок; основные воски, такие как какао-масло, полиэтиленгликоль и керосин и т.п., но эти примеры являются неограничивающими.
Препаратами, подходящими для перорального введения, являются раствор, получаемый растворением эффективного количества лиганда в разбавителе, таком как вода, физиологический раствор или апельсиновый сок; капсулы, саше или таблетки, содержащие эффективное количество лиганда в твердой или гранулированный форме; суспензия, получаемая суспендированием эффективного количества активного ингредиента в подходящем дисперсанте; эмульсия, получаемая диспергированием и эмульгированием раствора эффективного количества активного ингредиента в подходящем дисперсанте, и т.п.
Лекарственное средство по настоящему изобретению можно покрывать способом, известным per se, в целях маскировки вкуса, растворения в желудочно-кишечном тракте, замедленного высвобождения и т.п. по мере необходимости. В качестве примеров покрывающих средств, используемых для покрывания, используют гидроксипропилметилцеллюлозу, этилцеллюлозу, гидроксиметилцеллюлозу, гидроксипропилцеллюлозу, полиоксиэтиленгликоль, Tween 80, плюроник F68, ацетатфталат целлюлозы, фталат гидроксипропилметилцеллюлозы, ацетатсукцинат гидроксиметилцеллюлозы, эудражит (производимый Rohm, Germany, coполимер метакриловой кислоты/акриловой кислоты), пигменты (например, оксид железа (III), диоксид титана и т.п.) и т.п. Лекарственное средство может являться препаратом с быстрым высвобождением или препаратом с замедленным высвобождением. Примеры основ с замедленным высвобождением включают липосому, ателоколлаген, желатин, гидроксиапатит, PLGA и т.п.
В качестве препаратов, подходящих для парентерального введения (например, внутривенного введения, подкожного введения, внутримышечного введения, местного введения, интраперитонеального введения, интраназального введения, ингаляционного введения и т.п.), доступны водные и неводные изотонические стерильные инъецируемые жидкости, которые могут содержать антиоксидант, буферный раствор, бактериостатическое средство, средство придания изотоничности и т.п. Также можно упомянуть водные и неводные стерильные суспензии, которые могут содержать суспендирующее средство, солюбилизатор, загуститель, стабилизатор, антисептик и т.п. Препарат можно заключать в контейнер, такой как ампула или сосуд в объеме единицы дозирования или в нескольких отдельных дозах. Активный ингредиент и фармацевтически приемлемый носитель также можно лиофилизировать и хранить в таком состоянии, что его можно растворять или суспендировать в подходящем стерильном средстве непосредственно перед использованием. Препараты с замедленным высвобождением также являются подходящими препаратами. Препараты с замедленным высвобождением включают замедленное высвобождение из носителей или контейнеров, введенных в организм, таких как искусственные кости, биодеградируемые или деградируемые губки, мешки, помпы с лекарственными средствами, осмотические помпы и т.п. Устройства для непрерывной или периодической, системной или местной доставки извне организма также включены в объем препаратов с замедленным высвобождением. Биодеградируемые основы включают липосому, катионную липосому, сополимер молочной и гликолевой кислот (PLGA), ателоколлаген, желатин, гидроксиапатит, полисахарид сизофиран. В дополнение к инъецируемым жидкостям и к препарату с замедленным высвобождением также подходят ингаляционные средства и мази. В случае ингаляционного средства активный ингредиент в лиофилизированном состоянии тонко измельчают и вводят посредством ингаляции с использованием подходящего устройства для ингаляции. Ингаляционное средство при необходимости можно составлять с общеупотребительным поверхностно-активным веществом, маслом, вкусовой добавкой, циклодекстрином или его производным и т.п.
В настоящем описании в качестве примеров поверхностно-активного вещества можно упомянуть олеиновую кислоту, лецитин, диэтиленгликоль диолеат, тетрагидрофурфурилолеат, этилолеат, изопропилмиристат, глицерилтриолеат, глицерилмонолаурат, глицерилмоноолеат, глицерилмоностеарат, глицерилмонолизиноат, цетиловый спирт, стеариловый спирт, полиэтиленгликоль 400, хлорид цетилпиридиния, сорбитантриолеат (торговое название Span 85), сорбитанмоноолеат (торговое название Span 80), сорбитанмонолаурат (торговое название Span 20), касторовое масло, гидрогенизированное полиоксиэтиленом (торговое название HCO-60), полиоксиэтилен (20) сорбитанмонолаурат (торговое название Tween 20), полиоксиэтилен (20) сорбитан моноолеат (торговое название Tween 80), лецитин природного происхождения (торговое название EPICLON), олеиловый простой эфир полиоксиэтилена (2) (торговое название Brij 92), стеариловый простой эфир полиоксиэтилена (2) (торговое название Brij 72), лауриловый простой эфир полиоксиэтилена (4) (торговое название Brij 30), олеиловый простой эфир полиоксиэтилена (2) (торговое название Genapol 0-020), блок-сополимер оксиэтилена и оксипропилена (торговое название Synperonic) и т.п. В качестве примеров масла можно упомянуть кукурузное масло, оливковое масло, хлопковое масло, подсолнечное масло и т.п. В случае мази подходящую фармацевтически приемлемую основу (желтый вазелин, белый вазелин, парафин, пластибейз, силикон, вазелиновая мазь, пчелиный воск, лярд, растительные масла, гидрофильную основу для мази, гидрофильный петролатум, очищенный ланолин, гидролизованный ланолин, влагопоглощающую основу для мази, гидрофильный пластибейз, макроголовую основу для мази и т.п.) смешивают с активным ингредиентом и используют в качестве препарата.
Ингаляционное средство можно получать общепринятым способом. В частности, ингаляционное средство можно получать измельчением в порошок или разжижением описываемого выше аптамера и комплекса по настоящему изобретению, смешиванием его в пропелленте и/или носителе для ингаляции и наполнением им подходящего сосуда для ингаляции. Если описываемый выше аптамер и комплекс по настоящему изобретению является порошком, можно использовать общепринятый механический ингалятор для порошка; в случае жидкости можно использовать ингалятор, такой как небулайзер. В настоящем описании в качестве пропеллента можно широко использовать общеизвестный пропеллент; можно упомянуть хлорфторуглеродные соединения, такие как хлорфторуглерод-11, хлорфторуглерод-12, хлорфторуглерод-21, хлорфторуглерод-22, хлорфторуглерод-113, хлорфторуглерод-114, хлорфторуглерод-123, хлорфторуглерод-142c, хлорфторуглерод-134a, хлорфторуглерод-227, хлорфторуглерод-C318 и 1,1,1,2-тетрафторэтан, углеводороды, такие как пропан, изобутан и n-бутан, простые эфиры, такие как диэтиловый простой эфир, сжатые газы, такие как газообразный азот и газообразный диоксид углерода и т.п.
Дозировка лекарственного средства по настоящему изобретению варьируется в зависимости от типа и активности активного ингредиента, тяжести заболевания, вида животного, являющегося субъектом для введения, переносимости лекарственного средства субъектом для введения, массы тела, возраста и т.п., и типичная дозировка с учетом количества активного ингредиента в сутки для взрослого может составлять от приблизительно 0,0001 до приблизительно 100 мг/кг, например, от приблизительно 0,0001 до приблизительно 10 мг/кг, предпочтительно - от приблизительно 0,005 до приблизительно 1 мг/кг.
Настоящее изобретение также относится к твердофазному носителю, содержащему иммобилизованный на нем аптамер и комплекс по настоящему изобретению. В качестве примеров твердофазного носителя можно упомянуть подложку, смолу, планшет (например, многолуночный планшет), фильтр, картридж, колонку и пористый материал. Подложка может являться подложкой, используемой в ДНК-чипах, белковых чипах и т.п.; например, можно упомянуть подложки никель-PTFE (политетрафторэтилен), стеклянные подложки, апатитовые подложки, силиконовые подложки, подложки из оксида алюминия и т.п., и подложки, получаемые покрыванием этих подложек полимером и т.п. В качестве примеров смолы можно упомянуть частицы агарозы, частицы диоксида кремния, coполимер акриламида и N,N’-метиленбисакриламида, частицы полистирола, поперечно сшитого с дивинилбензолом, частицы декстрана, поперечно сшитого с эпихлоргидрином, целлюлозное волокно, сшитые полимеры арилдекстрана и N,N’-метиленбисакриламида, монодисперсные синтетические полимеры, монодисперсные гидрофильные полимеры, сефарозу, Toyopearl и т.п., а также смолы, получаемые связыванием различных функциональных групп с этими смолами. Твердофазный носитель по настоящему изобретению можно использовать, например, в очистке, определении и количественном анализе NGF.
Аптамер и комплекс по настоящему изобретению можно иммобилизовать на твердофазном носителе способом, известным per se. Например, можно упомянуть способ, при котором в аптамер или комплекс по настоящему изобретению включают вещество для аффинной хроматографии (например, описываемое выше) или заранее определенную функциональную группу, а затем иммобилизуют аптамер и комплекс на твердофазном носителе через вещество для аффинной хроматографии или заранее определенную функциональную группу.Настоящее изобретение также относится к таким способам. Заранее определенная функциональная группа может являться функциональной группой, которую можно подвергать реакции присоединения; например, можно упомянуть аминогруппу, тиоловую группу, гидроксильную группу и карбоксильную группу. Настоящее изобретение также относится к аптамеру, содержащему такую встроенную в него функциональную группу.
Настоящее изобретение также относится к способу очистки и концентрирования NGF. В частности, настоящее изобретение делает возможным отделение NGF от белков из других семейств белков. Способ очистки и концентрирования по настоящему изобретению может включать адсорбцию NGF на твердофазном носителе по настоящему изобретению и элюцию адсорбированного NGF с элюентом. Адсорбцию NGF на твердофазном носителе по настоящему изобретению можно осуществлять способом, известным per se. Например, содержащий NGF образец (например, бактериальную или клеточную культуру или супернатант культуры, кровь) наносят на твердофазный носитель по настоящему изобретению или содержащую его композицию. NGF модно элюировать с использованием элюента, такого как нейтральный раствор. Не существует ограничений в отношении нейтрального элюента, который может иметь pH, например, от приблизительно 6 до приблизительно 9, предпочтительно - от приблизительно 6,5 до приблизительно 8,5, и более предпочтительно - от приблизительно 7 до приблизительно 8. Нейтральный раствор также может содержать, например, мочевину, хелатирующее средство (например, ЭДТА), соль калия (например, KCl), соль магния (например, MgCl2), поверхностно-активное вещество (например, Tween 20, Triton, NP40) и глицерин. Способ очистки и концентрирования по настоящему изобретению дополнительно может включать промывку твердофазного носителя после адсорбции NGF с использованием промывочного раствора. Примеры промывочного раствора включают растворы, содержащие мочевину, хелатирующее средство (например, ЭДТА), Трис, кислоту, щелочь, транспортную РНК, ДНК, поверхностно-активные вещества, такие как Tween 20, соли, такие как NaCl и т.п. Способ очистки и концентрирования по настоящему изобретению дополнительно может включать нагревание твердофазного носителя. Этот этап делает возможным восстановление и стерилизацию твердофазного носителя.
Настоящее изобретение также относится к способу определения и количественного анализа NGF. В частности, настоящее изобретение делает возможным определение и количественный анализ NGF отдельно от белков из других семейств белков. Способ определения и количественного анализа по настоящему изобретению может включать измерение NGF с использованием аптамера по настоящему изобретению (например, с использованием комплекса и твердофазного носителя по настоящему изобретению). Способ определения и количественного анализа NGF можно осуществлять таким же образом, что и иммунологический способ анализа, за исключением того, что вместо антитела используют аптамер по настоящему изобретению. Таким образом, при использовании аптамера по настоящему изобретению в качестве зонда вместо антитела определение и количественный анализ можно осуществлять таким же образом, как иммуноферментный анализ (EIA) (например, прямой конкурентный ELISA, непрямой конкурентный ELISA, сэндвич-ELISA), радиоиммунологический анализ (RIA), флуоресцентный иммуноанализ (FIA), вестерн-блоттинг, способ иммуногистохимического окрашивания и способ сортировки клеток. Аптамер по настоящему изобретению также можно использовать в качестве молекулярного зонда для PET и т.п. Эти способы можно использовать, например, в измерении содержания NGF в живых организмах или биологических образцах и диагностике заболевания, ассоциированного с NGF.
Описания всех публикаций, упомянутых в настоящем описании, включая описания патентов и патентных заявок, включены в настоящее описание в качестве ссылки в такой степени, как если бы они были указаны конкретно.
Далее в настоящем описании более подробно описывают настоящее изобретение с помощью следующих примеров, которые, однако, не в коей мере не ограничивают объем изобретения.
ПРИМЕРЫ
Пример 1: Получение аптамера против NGF - 1
Аптамеры РНК, специфически связывающиеся с NGF, получали с использованием способа SELEX. SELEX осуществляли способом Ellington et al. (Ellington и Szostak, Nature 346, 818-822, 1990) и способом Tuerk et al. (Tuerk и Gold, Science 249, 505-510, 1990). В качестве вещества-мишени использовали NGF человека (производимый R&D Systems).
РНК, используемую в первом раунде (40N), получали транскрибированием химически синтезированной ДНК с использованием набора для транскрипции DuraScribeтм T7 (производимого Epicentre). Из NTP, содержащихся в наборе, 2’-OH АТФ заменяли 2’-дезоксиаденозин-5’-трифосфатом (2’-H АТФ или dATP, производимым GE Healthcare) и использовали другие субстраты, содержащиеся в наборе. РНК, получаемая этим способом, содержит фторированное 2’-положение рибозы пиримидинового нуклеотида и G (пуриновый нуклеотид) РНК-типа и A ДНК-типа. Представленную ниже ДНК из 83 нуклеотидов, содержащую последовательность праймера на каждом конце 40-нуклеотидной случайной последовательности, использовали в качестве ДНК-матрицы. ДНК-матрицу и праймеры получали химическим синтезом.
ДНК матрица 1:
5’-gaggatccatgtatgcgcacata-40n-cttctggtcgaagttctccc-3’ (SEQ ID NO: 118)
Праймер Fwd1:
5’-cggaattctaatacgactcactatagggagaacttcgaccagaag-3’ (SEQ ID NO: 119)
Праймер Rev1:
5’-gaggatccatgtatgcgcacata-3’ (SEQ ID NO: 120)
В указанной выше последовательности n представляет собой любой из a, g, c и t. Праймер Fwd1 содержит промоторную последовательность РНК-полимеразы T7. Разброс совокупности РНК, используемой в первом раунде, теоретически составляет 1014.
После 10 раундов SELEX продукт ПЦР клонировали в вектор pGEM-T Easy (производимый Promega), используемый для трансформации штамма DH5α Escherichia coli (производимого Toyobo). Плазмиду выделяли из одной колонии и определяли последовательности оснований с помощью ДНК-секвенатора (ABI PRISM3100, производимого ABI). Исследовали 48 клонов и секвенировали 45 последовательностей. Среди них находились 7 типов одних и тех же 2 последовательностей, и остальные 31 последовательность являлись единичными последовательностями. Дополнительно проводили 3 раунда SELEX и исследовали последовательность. В результате, наблюдали еще большую конвергенцию.
Последовательности, демонстрирующие конвергенцию в 10 и 13 раундах и несколько единичных последовательностей, оценивали на активность связывания с NGF с помощью способа поверхностного плазмонного резонанса.
В качестве устройства для измерения использовали BIAcore2000, производимый BIAcore, а в качестве сенсорного чипа использовали CM5, реагирующий на аминогруппу. NGF человека растворяли в растворе для иммобилизации (10 мМ ацетат натрия, pH 6) при концентрации 25-40 мкг/мл. Для реакции аминогруппы белка и карбоксильной группы чипа использовали этил-3-карбодиимид гидрохлорид и N-гидроксисукцинимид. После реакции осуществляли блокирование этаноламином-HCl. Иммобилизованное количество NGF устанавливали на 3000-4000 RU. Получали аптамер для аналита в концентрации 0,15 мкМ - 0,5 мкМ. В качестве подвижного буфера использовали раствор A. В настоящем описании раствор A является смешанным раствором 145 мМ хлорида натрия, 5,4 мМ хлорида калия, 1,8 мМ хлорида кальция, 0,8 мМ хлорида магния, 20 мМ Трис (pH 7,6), 0,05% Tween 20. В качестве раствора для восстановления использовали смешанный раствор 1M NaCl и 50 мМ NaOH. NGF иммобилизовывали на FC2 и вычитали результаты для FC1 для получения конечной сенсограммы.
Измеряли активность связывания 34 последовательностей и обнаруживали, что все РНК сильнее связывались с NGF, чем контрольная 40N. В настоящем описании 40N относится к совокупности РНК, используемой для первого раунда, содержащей 40-нуклеотидную случайную последовательность. Учитывая приведенное выше, показано, что эти РНК являются аптамерами, связывающимися с NGF.
Пример 2: Аптамер, ингибирующий связывание NGF и рецептора NGF
С использованием способа поверхностного плазмонного резонанса определяли, ингибируют ли аптамеры, полученные в примере 1, связывание NGF и рецептора NGF (TrkA и p75) или нет.
Как указано в протоколе BIAcore, протеин A (21181, PIERCE) иммобилизовывали на сенсорном чипе CM5. На нем иммобилизовывали приблизительно от 700 до 1200 RU TrkA человека, подвергнутых слиянию с Fc-областью IgG (175-TK, R&D systems), или P75 человека (367-NR, R&D systems). После отстаивания в течение 30 минут смесь NGF (0,1 мкМ) и каждого аптамера (0,3 мкМ) инъецировали в качестве аналита. Если аптамер ингибирует связывание NGF и TrkA или p75, ожидают, что сигнал на сенсограмме не будет возрастать; если аптамер не ингибирует связывание, будет образовываться тройной комплекс, и ожидают, что сигнал будет возрастать. Если NGF сильнее связывается с рецептором, чем с аптамером, аптамер можно удалять, и NGF может связываться с рецептором. Перед началом эксперимента по ингибированию подтверждали связывание TrkA или p75 и NGF.
Измеряли ингибиторную активность 34 последовательностей и обнаруживали, что все аптамеры ингибируют связывание NGF и TrkA или p75. В частности, аптамеры, приведенные в SEQ ID NO: 1, 2, 3, 4, 5, 7, демонстрировали сильную ингибиторную активность. Учитывая приведенное выше, показано, что эти РНК являются аптамерами, ингибирующими связывание NGF и рецептора NGF.
Пример 3: Ингибиторная активность аптамера в отношении роста нейритов
Ингибиторную активность аптамера, полученного в примере 1, в отношении роста нейритов оценивали с использованием клетки Neuroscreen-1, являющейся субклоном клеток PC-12.
Клетки (2500 клеток на лунку) культивировали в течение одного дня в среде RPMI-1640, содержащей 2,5% лошадиной сыворотки и 1,25% эмбриональной телячьей сыворотки, в 96-луночном плоскодонном планшете, покрытом коллагеном типа IV. Добавляли смешанный раствор NGF человека (конечная концентрация 0,38 нМ или 1,14 нМ) и аптамера (конечная концентрация 500 - 0,01 нМ), предварительно прореагировавших в бессывороточной среде RPMI-1640 при комнатной температуре или 37°C в течение периода от 30 мин до 1 ч. Через два дня цитоплазму и ядра окрашивали с использованием набора Cellomics Neurite Outgrowth (производимого Thermo Scientific) и измеряли длину нейрита на клетку с использованием Cellomics ArrayScan VTI (производимого Thermo Scientific). Используя длину нейрита на клетку, полученную при добавлении только NGF, в качестве ингибиторной активности 0%, и длину нейрита на клетку, полученную в культуре без NGF в течение 2 дней, в качестве ингибиторной активности 100%, из длины нейрита на клетку, полученной при культивировании с добавлением NGF и аптамера в смесь, вычисляли ингибиторную активность аптамера.
Измеряли ингибиторную активность 34 типов аптамеров, полученных в примере 1, и обнаруживали, что аптамеры, приведенные в SEQ ID NO: 1-8, сильно ингибируют рост нейритов при добавлении их в концентрации 10 нМ. Другие аптамеры не продемонстрировали значительного ингибирования при концентрации 10 нМ.
Фактически, полученные нуклеотидные последовательности, соответствующие каждому SEQ ID NO, представлены ниже. Прописными буквами показана РНК, строчными буквами показана ДНК, нуклеотиды в круглых скобках представляют собой модификацию в 2’-положении, и F является атомом фтора (так же далее в настоящем описании).
SEQ ID NO: 1:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GaC(F)GaC(F)C(F)aaC(F)U(F)C(F)GU(F)C(F)U(F)C(F)U(F)U(F)aU(F)GGaU(F)U(F)U(F)aC(F)GU(F)GaaC(F)C(F)C(F)GU(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 2:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)C(F)C(F)aaaC(F)GGGaC(F)U(F)U(F)U(F)aU(F)aC(F)C(F)U(F)C(F)U(F)GaGU(F)C(F)GC(F)C(F)U(F)aC(F)GC(F)U(F)C(F)C(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 3:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 4:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)aC(F)GU(F)U(F)aGU(F)aC(F)GU(F)U(F)U(F)GC(F)aU(F)aU(F)GU(F)aC(F)aaC(F)C(F)U(F)U(F)GC(F)aU(F)aC(F)GaU(F)aC(F)GU(F)aGaU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 5:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)aGaaGaGGaC(F)U(F)aGU(F)U(F)GC(F)U(F)aaU(F)GC(F)C(F)C(F)U(F)GGU(F)U(F)C(F)GU(F)C(F)GC(F)U(F)aU(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 6:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)GC(F)aaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 7:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGaC(F)GC(F)aC(F)C(F)U(F)C(F)U(F)U(F)aU(F)C(F)aC(F)aC(F)aU(F)GC(F)GU(F)C(F)aGC(F)C(F)U(F)U(F)GU(F)GaU(F)aC(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 8:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGaU(F)C(F)C(F)aC(F)U(F)GGU(F)aC(F)U(F)aC(F)GU(F)GaC(F)C(F)C(F)C(F)GC(F)aU(F)aGGC(F)aaU(F)C(F)C(F)U(F)GC(F)U(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
Пример 4: Укорачивание аптамеров
Аптамеры, приведенные в SEQ ID NO: 3 и 6, подвергали укорачиванию. Вторичную структуру РНК прогнозировали с использованием программы MFOLD (Zuker, Nucleic Acids Res. 31, 3406-3415, 2003) и укорачивали цепь, учитывая ее структуру. Укороченную форму получали химическим синтезом ДНК целевой последовательности и ее транскрипцией с использованием набора для транскрипции DuraScribe T7 (производимого Epicentre). Продукт транскрипции обрабатывали ДНКазой, белок удаляли обработкой фенол-хлороформом и собирали РНК осаждением этанолом. Чистоту выделенной РНК подтверждали электрофорезом в полиакриламидном геле, а количество подтверждали способом измерения поглощения. Фактически, полученные последовательности в укороченной форме представлены ниже.
SEQ ID NO: 9: 73-нуклеотидная РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 3
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)
SEQ ID NO: 10: 68-нуклеотидная РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 3
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)G
SEQ ID NO: 11: 46-нуклеотидная РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 3
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 12: 40-нуклеотидная РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 3
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 13: 42-нуклеотидная РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 3
GGGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 121: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из стебля 2 удаляют 1 пару оснований
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 122: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из стебля 2 удаляют 1 пару оснований
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 123: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из стебля 2 удаляют 1 пару оснований
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GU(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 124: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из стебля 2 удаляют 1 пару оснований
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GGU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)U(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 125: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из стебля 2 удаляют 1 пару оснований
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)U(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 126: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из стебля 2 удаляют 1 пару оснований
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)GU(F)U(F)U(F)C(F)C(F)GU(F)C(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 127: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из стебля 2 удаляют 1 пару оснований
GGGGU(F)C(F)C(F)U(F)C(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGU(F)GU(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 128: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из стебля 2 удаляют 1 пару оснований
GGGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aU(F)U(F)U(F)C(F)C(F)GU(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 129: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из стебля 1 удаляют 1 пару оснований
GGGGC(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)C(F)C(F)C(F)C(F)
SEQ ID NO: 130: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из петли 2 удаляют один U
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 131: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из петли 2 удаляют два C’
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 132: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из петли 2 удаляют один G
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)U(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 133: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из петли 2 удаляют один U
GGGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GC(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 134: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 11, где из внутренней петли 1 удаляют один U
GGGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)aU(F)GU(F)GC(F)
SEQ ID NO: 135: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из внутренней петли 1 удаляют один C
GGGGU(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 136: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из внутренней петли 1 удаляют один U
GGGGU(F)C(F)C(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 137: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из внутренней петли 1 удаляют один C и один U
GGGGU(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 138: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из внутренней петли 1 удаляют один C и один G
GGGGU(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)U(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 139: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из внутренней петли 1 удаляют два U
GGGGU(F)C(F)C(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)GU(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 140: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из внутренней петли 1 удаляют один U и один G
GGGGU(F)C(F)C(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)U(F)U(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 141: РНК, являющаяся измененной формой аптамера, приведенного в SEQ ID NO: 13, где из внутренней петли 1 удаляют два U
GGGGU(F)C(F)C(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)aC(F)C(F)C(F)C(F)
SEQ ID NO: 14: 78-нуклеотидный аптамер, являющийся измененной формой аптамера, приведенного в SEQ ID NO: 6
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)GC(F)aaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGa
SEQ ID NO: 15: 73-нуклеотидный аптамер, являющийся измененной формой аптамера, приведенного в SEQ ID NO: 6
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)GC(F)aaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)
SEQ ID NO: 16: 63-нуклеотидный аптамер, являющийся измененной формой аптамера, приведенного в SEQ ID NO: 6
C(F)GaC(F)C(F)aGaaGU(F)GC(F)aaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)
SEQ ID NO: 17: 58-нуклеотидный аптамер, являющийся измененной формой аптамера, приведенного в SEQ ID NO: 6
aGaaGU(F)GC(F)aaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)
SEQ ID NO: 18: 48-нуклеотидный аптамер, являющийся измененной формой аптамера, приведенного в SEQ ID NO: 6
U(F)GC(F)aaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)GC(F)G
SEQ ID NO: 19: 46-нуклеотидный аптамер, являющийся измененной формой аптамера, приведенного в SEQ ID NO: 6
GC(F)aaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)GC(F)
SEQ ID NO: 20: 50-нуклеотидный аптамер, являющийся измененной формой аптамера, приведенного в SEQ ID NO: 6
GU(F)GC(F)aaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)GC(F)GC(F)
SEQ ID NO: 21: 48-нуклеотидный аптамер, являющийся измененной формой аптамера, приведенного в SEQ ID NO: 6
GGGaaU(F)aC(F)U(F)U(F)U(F)C(F)GC(F)GGC(F)aU(F)aU(F)GU(F)GC(F)aaaC(F)C(F)U(F)U(F)GC(F)C(F)aC(F)GaC(F)U(F)aU(F)GU(F)C(F)C(F)C(F)
Активность связывания этих аптамеров с NGF оценивали с помощью способа поверхностного плазмонного резонанса, что представлено в примере 1. В результате, обнаруживали, что РНК, приведенная в SEQ ID NO: 9-21, связывается с NGF сильнее, чем контрольная 40N. С другой стороны, степень связывания РНК, приведенной в SEQ ID NO: 121-141, значительно снижено по сравнению с аптамером, приведенным в SEQ ID NO: 11 или 13.
Ингибирование связывания NGF с его рецепторами (TrkA и p75) оценивали с помощью способа поверхностного плазмонного резонанса аналогично представленному в примере 2 и обнаруживали, что аптамеры, приведенные в SEQ ID NO: 9-16, обладают высокой ингибиторной активностью.
Активность ингибирования роста нейритов исследовали способом, аналогичным представленному в примере 3, и обнаруживали, что аптамеры, приведенные в SEQ ID NO: 9-21, обладают высокой ингибиторной активностью при концентрации 10 нМ. С другой стороны, аптамеры, приведенные в SEQ ID NO: 127, 128, 131, 133, 135, 141, не продемонстрировали значительной ингибиторной активности при концентрации 10 нМ.
Пример 5: Прогнозирование вторичной структуры аптамера, приведенного в SEQ ID NO: 3, и его укороченной формы
Вторичную структуру РНК, приведенной в SEQ ID NO: 3, 9-13 и SEQ ID NO: 121-141, прогнозировали с использованием программы MFOLD. Все аптамеры, обладающие активностью, содержали вторичную структуру аптамера, приведенного в SEQ ID NO: 12 (фиг. 1). Вторичную структуру охарактеризовывали с использованием 4 структур петли 1, внутренней петли 1, стебля 2 и петли 2, начиная с 5'-конца (фиг. 2). Стебель 1 состоит из 4 пар оснований, внутренняя петля 1 состоит из 3 нуклеотидов и 4 нуклеотидов, стебель 2 состоит из 9 пар оснований, и петля 2 состоит из 7 нуклеотидов.
Все активности РНК, приведенной в SEQ ID NO: 121-128, где из стебля 2 удаляют 1 пару оснований, значительно снижались. Таким образом, предполагали, что для стебля 2 необходимо 9 пар оснований.
Аптамеры, приведенные в SEQ ID NO: 12 и 13, являются измененными формами аптамера, приведенного в SEQ ID NO: 11, где стебель 1 замещают парой G-C. Аптамеры, приведенные в SEQ ID NO: 12 и 13, демонстрировали активность ингибирования роста нейритов, равную таковой для SEQ ID NO: 11. Таким образом, предполагали, что пара оснований не влияет значительно на активность при условии, что стебель 1 является структурой стебля. С другой стороны, значительно снижалась активность РНК, приведенной в SEQ ID NO: 129, являющейся аптамером, приведенным в SEQ ID NO: 13, где пару оснований U-a удаляют из стебля 1. Таким образом, обнаруживали, что 4-ая пара оснований стебля 1 должна представлять собой U-a.
Все РНК, приведенные в SEQ ID NO: 130-133, где из петли 2 удаляют один нуклеотид, демонстрировали значительно сниженную активность. Таким образом, предполагали, что петля 2 должна состоять из 7 нуклеотидов.
Все РНК, приведенные в SEQ ID NO: 134-141, где из внутренней петли 1 удаляли 1 или 2 нуклеотида, демонстрировали значительно сниженную активность. Таким образом, предполагали, что внутренняя петля 1 должна содержать всего 7 нуклеотидов.
Структуру, представленную на фиг.2, далее в настоящем описании обозначают как консенсусную вторичную структуру 1.
Пример 6: Получение аптамера против NGF - 2
SELEX осуществляли с использованием праймера, отличающегося от праймера в примере 1, и определяли, можно ли получать аптамер, имеющий консенсусную вторичную структуру 1. Используемая ДНК-матрица и последовательности праймеров представлены ниже.
ДНК-матрица 2:
5’-ccagttgttggtgacaatgcnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnngcagctccacaggcttccc (SEQ ID NO: 142)
Праймер Fwd2:
5’-taatacgactcactatagggaagcctgtggagctgc (SEQ ID NO: 143)
Праймер Rev2:
5’-gcattgtcaccaacaactgg (SEQ ID NO: 144)
В указанных выше последовательностях n представляет любой из a, g, c и t. Праймер Fwd2 содержит промоторную последовательность РНК-полимеразы T7. Разброс совокупности РНК, используемой в первом раунде, теоретически составлял 1014.
SELEX осуществляли аналогично примеру 1. После 10 раундов SELEX исследовали 48 клонов и секвенировали 46 последовательностей. Среди них была 1 последовательность, где 5 клонов являлись одинаковыми, 4 последовательности, где 3 клона являлись одинаковыми, и 3 последовательности, где 2 клона являлись одинаковыми, и всего 8 последовательностей демонстрировали конвергенцию. Остальные 23 последовательности являлись единичными последовательностями.
Выбирали 8 последовательностей, демонстрирующих конвергенцию, и оценивали активность связывания с NGF с помощью способа поверхностного плазмонного резонанса. Способ измерения являлся аналогичным способу в примере 1. В результате, все последовательности связывались с NGF лишь незначительно.
Вторичную структуру всех последовательностей, включая единичные последовательности, прогнозировали с использованием программы MFOLD и не обнаруживали последовательность, содержащую консенсусную вторичную структуру 1.
Пример 7: Получение аптамера против NGF - 3
SELEX осуществляли с использованием совокупности РНК, содержащей последовательность, приведенную в SEQ ID NO: 12, дополненную 15% случайной последовательности и последовательностями праймеров, аналогичных представленным в примере 1, с обоих ее концов. SELEX осуществляли почти аналогично представленному в примере 1. Последовательности матрицы представлены ниже.
матрица 3:
5’-gaggatccatgtatgcgcacata-acagccacggagacggaaactacgcagcaggatgtgccaa-cttctggtcgaagttctccc-3’ (SEQ ID NO: 145)
Последовательность оснований, подчеркнутая в последовательности, представлена ниже.
a: a(85%), g(5%), c(5%), t(5%)
g: a(5%), g(85%), c(5%), t(5%)
c: a(5%), g(5%), c(85%), t(5%)
t: a(5%), g(5%), c(5%), t(85%)
После завершения 4 раундов подтверждали последовательности 48 клонов и обнаруживали, что приблизительно половина является последовательностью, содержащей SEQ ID NO: 12, а остальная часть является последовательностью с мутациями в нескольких участках. Для удаления той же последовательности, что и SEQ ID NO: 12, к совокупности РНК добавляли антисмысловой олигонуклеотид SEQ ID NO: 12, и осуществляли SELEX в течение еще 3 раундов. Последовательность антисмыслового нуклеотида представлена ниже.
5’-agacggaaactacgcagcagga-3’- (SEQ ID NO: 146)
Антисмысловой олигонуклеотид добавляли в 10-кратном количестве относительно совокупности РНК. Подтверждали последовательность полученной РНК и обнаруживали, что приблизительно половина являлась последовательностью, приведенной в SEQ ID NO: 12 с мутациями в нескольких участках, а остальная часть являлась последовательностью, полностью отличающейся от последовательности, приведенной в SEQ ID NO: 12.
Из 4 и 7 раундов выбирали всего 16 последовательностей, приведенных в SEQ ID NO: 22-37, и исследовали активность связывания с NGF и активность ингибирования связывания NGF и рецептора NGF. Измерение являлось аналогичным представленному в примерах 1 и 2, и использовали способ поверхностного плазмонного резонанса. В результате измерения обнаруживали, что все последовательности связываются с NGF сильнее, чем контрольная 40N, и ингибируют связывание NGF и рецептора NGF. Кроме того, активность ингибирования роста нейритов измеряли способом, аналогичным представленному в примере 3. В результате, все последовательности демонстрировали высокую ингибиторную активность при концентрации 10 нМ. Фактически, полученные нуклеотидные последовательности, соответствующие каждому SEQ ID NO, представлены ниже.
SEQ ID NO: 22:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 23:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 24:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GaaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 25:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GC(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 26:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)C(F)GC(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 27:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GC(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 28:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)aU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 29:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)C(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GaaGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 30:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GaaGGU(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)aC(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 31:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GC(F)aGU(F)U(F)U(F)C(F)C(F)C(F)U(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 32:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GGaGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 33:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)C(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)aaU(F)C(F)U(F)U(F)GGU(F)GGC(F)GU(F)GU(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 34:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)C(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 35:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GU(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 36:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGGU(F)aC(F)GU(F)U(F)aGU(F)aC(F)GU(F)U(F)U(F)GC(F)aU(F)aU(F)GU(F)aC(F)aaC(F)C(F)U(F)U(F)GC(F)aU(F)aC(F)GaU(F)aC(F)GU(F)aGGU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 37:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)aC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
Вторичную структуру аптамеров, приведенных в SEQ ID NO: 22-37, прогнозировали с использованием программы MFOLD и обнаруживали, что все аптамеры, иные, чем аптамеры, приведенные в SEQ ID NO: 30, 33, 36, содержат консенсусную вторичную структуру 1. 5’-сторона всех последовательностей внутренних петель 1 представлял собой CCU, и 3’-сторона представляла собой UGUU (фиг. 2). Кроме того, петля 2 содержала консенсусную последовательность, представленную 5’-UUUCCXU-3’. В этом случае X является любым из A, G, C и U. Все конечные пары оснований стебля 1 представляли собой U-a. 1-ая, 5-ая, 6-ая, 8-ая и 9-ая пары оснований стебля 2 представляли собой G-C, C-G, G-C, a-U и G-C, соответственно. 2-ая - 4-ая и 7-ая содержали некоторые другие пары оснований.
Пример 8: Ингибиторная активность аптамера в отношении пролиферации клеток (анализ TF-1)
Ингибиторные активности аптамеров, приведенных в SEQ NO: 22-37, оценивали с помощью анализа ингибирования пролиферации с использованием клеток TF-1.
Гены двух рецепторов NGF (TrkA человека и p75 человека) встраивали в клетки TF-1 (регистрационный номер ATCC: CRL-2003), являющиеся линией эритролейкозных клеток человека, с использованием ретровирусного вектора для получения клеток, одновременно и стабильно высоко экспрессирующих два рецептора. Клетки суспендировали в среде RPMI-1640, содержащей 20% эмбриональную телячью сыворотку, и высевали в белый 96-луночный плоскодонный планшет в концентрации 1000 клеток (50 мкл) на лунку. Туда добавляли смешанный раствор 50 мкл NGF человека (конечная концентрация 0,076 нМ) и аптамера (конечная концентрация 30-0,01 нМ), предварительно прореагировавших при комнатной температуре в течение 30 мин в бессывороточной среде RPMI-1640, через 3 дня в каждую лунку добавляли 100 мкл реагента CellTiter-Glo для анализа CellTiter-Glo Luminescent Cell Viability Assay (производимый Promega), измеряли хемилюминесценцию с использованием микроспектрофотометра для чтения планшетов и оценивали рост клеток TF-1 при стимуляции NGF. Используя интенсивность люминесценции на лунку, полученную при добавлении только NGF, и культивировании клеток в течение 3 дней, в качестве ингибиторной активности 0%, и интенсивность люминесценции на лунку, полученную при культивировании без NGF в течение 3 дней, в качестве ингибиторной активности 100%, из интенсивности люминесценции на лунку, полученной при культивировании с добавлением NGF и аптамера в смеси, вычисляли ингибиторную активность аптамера. В результате, обнаруживали, что все эти аптамеры демонстрировали высокую ингибиторную активность при концентрации 10 нМ.
Для сравнения исследовали ингибиторную активность аптамеров, приведенных в SEQ ID NO: 62 и 68, описываемых в WO 2010/035725 A1. В результате, IC50 составляла 6,1 и 7,5 нМ, соответственно.
Пример 9: Укорачивание аптамера - 2
Аптамеры, приведенные в SEQ ID NO: 23, 25, 26, 27, 28, 29, 31, 32, 34 и 35, демонстрировавшие высокую ингибиторную активность, подвергали укорачиванию с учетом консенсусной вторичной структуры 1. Активность связывания с NGF измеряли с помощью способа поверхностного плазмонного резонанса, аналогично приведенному в примере 1, и обнаруживали, что все укороченные формы сильно связываются с NGF. Кроме того, ингибиторную активность в отношении роста нейритов и ингибиторную активность в отношении пролиферации клеток TF-1 измеряли способом, аналогичным представленному в примерах 3 и 8, и обнаруживали сильную ингибиторную активность в концентрации 10 нМ. Фактически, полученные нуклеотидные последовательности представлены ниже.
SEQ ID NO: 38: (укороченная форма SEQ ID NO: 26)
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)C(F)GC(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 39: (укороченная форма SEQ ID NO: 27)
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GC(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 40: (укороченная форма SEQ ID NO: 28)
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)aU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 41:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GaaGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 42:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GC(F)aGU(F)U(F)U(F)C(F)C(F)C(F)U(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 43:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GGaGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 44:
GGGU(F)С(F)С(F)U(F)GC(F)С(F)GC(F)GU(F)aGU(F)U(F)U(F)С(F)С(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 45:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 46: (укороченная форма SEQ ID NO: 25)
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GC(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 47: (укороченная форма SEQ ID NO: 26)
GGGU(F)С(F)С(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)С(F)С(F)GU(F)C(F)U(F)U(F)C(F)GC(F)GGC(F)U(F)GU(F)U(F)aC(F)С(F)С(F)
Вторичную структуру этих аптамеров прогнозировали с использованием программы MFOLD и обнаруживали, что все из них обладают структурой, представленной консенсусной вторичной структурой 1.
Пример 10: Получение аптамера против NGF - 4
SELEX осуществляли с использованием совокупности РНК, содержащей последовательность, приведенную в SEQ ID NO: 12, дополненную 21% случайной последовательности и последовательностями праймеров, аналогичным представленным в примере 1, с обоих ее концов. SELEX осуществляли аналогично представленному в примере 1. Последовательность матрицы представлена ниже.
Матрица 3:
5'-gaggatccatgtatgcgcacata-acagccacggagacggaaactacgcagcaggatgtgccaa-cttctggtcgaagttctccc-3'
(SEQ ID NO: 145)
Последовательность оснований, подчеркнутую в последовательности, описывают ниже.
a: a(79%), g(7%), c(7%), t(7%)
g: a(7%), g(79%), c(7%), t(7%)
c: a(7%), g(7%), c(79%), t(7%)
t: a(7%), g(7%), c(7%), t(79%)
После завершения 4 раундов подтверждали последовательности 48 клонов, но не наблюдали конвергенцию последовательностей. Таким образом, осуществляли еще 3 раунда. В 5, 6, 7 раундах подтверждали последовательности 48 клонов, чтобы обнаружить конвергенцию последовательностей в последующих раундах. В 7-м раунде почти все последовательности демонстрировали конвергенцию.
Из 5, 6, 7 раундов выбирали всего 14 последовательностей, активность связывания с NGF измеряли с помощью способа поверхностного плазмонного резонанса. Способ измерения представлен в примере 1. В результате измерения все последовательности связывались с NGF сильнее, чем контрольная 40N. Фактически, полученные нуклеотидные последовательности представлены ниже.
SEQ ID NO: 48:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GGU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 49:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)U(F)GC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 50:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGC(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GU(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GaC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 51:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aC(F)C(F)C(F)U(F)GC(F)U(F)aC(F)GGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GGU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 52:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)aU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)U(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 53:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)U(F)GC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GC(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 54:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GaU(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GU(F)C(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 55:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GU(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 56:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GU(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)aU(F)GGU(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 57:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)aGGC(F)aC(F)GU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)aU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GU(F)GU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 58:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GaC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGU(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 59:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)U(F)GC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)GC(F)GU(F)GGU(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 60:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)GGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)aGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GGU(F)GGC(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 61:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)U(F)aGC(F)aC(F)aU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)C(F)GC(F)GGU(F)U(F)GU(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
Вторичную структуру этих аптамеров прогнозировали с использованием программы MFOLD и обнаруживали, что все из них обладают структурой, представленной консенсусной вторичной структурой 1. 5’-сторона всех последовательностей внутренних петель 1 представляла собой CCU, а 3’-сторона представляла собой UGUU. Кроме того, петля 2 содержала консенсусную последовательность, представленную 5’-UUUCCXU-3’. В этом случае X является G или U. 1-ая, 3-я, 8-ая и 9-ая пары оснований стебля 2 представляли собой G-C, U-G, a-U и G-C, соответственно. 2-ая и 4-ая - 7-ая содержали некоторые другие пары оснований.
Аналогично примеру 4, эти аптамеры подвергали укорачиванию. В результате, во всех аптамерах цепь можно укорачивать до структуры, аналогичной консенсусной вторичной структуре 1, при сохранении активности связывания.
Аналогично примерам 3 и 8, оценивали ингибиторную активность в отношении роста нейритов и ингибиторную активность в отношении пролиферации клеток TF-1. В результате, все эти аптамеры демонстрировали высокую ингибиторную активность при концентрации 10 нМ. Фактически, полученные нуклеотидные последовательности представлены ниже.
SEQ ID NO: 62:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GGU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 63:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 64:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 65:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)aC(F)GGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GGU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 66:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)aU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)U(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 67:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GC(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 68:
GGGU(F)C(F)C(F)U(F)GaU(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 69:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 70:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GU(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)aU(F)GGU(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 71:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)aU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GU(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 72:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGU(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 73:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)GC(F)GU(F)GGU(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 74:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)aGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GGU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 75:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)C(F)GC(F)GGU(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
Пример 11: Получение аптамера против NGF - 5
SELEX осуществляли аналогично примеру 1, используя следующую матрицу, где стебель 2 в консенсусной вторичной структуре 1 рандомизирован.
Матрица 4:
5’-gaggatccatgtatgcgcacataacagnnnnnnngacggaaacnnnnnnncaggatgtgccaacttctggtcgaagttctccc-3’ (SEQ ID NO: 147)
После завершения 7 раундов подтверждали последовательности 48 клонов, но не наблюдали конвергенцию последовательностей. Таким образом, осуществляли еще 3 раунда. После завершения 10 раундов подтверждали последовательности 48 клонов и обнаруживали конвергенцию приблизительно в половине последовательностей. Оставшаяся половина последовательностей представляла собой единичные последовательности. Из них выбирали 17 последовательностей и измеряли активность связывания с NGF с помощью способа поверхностного плазмонного резонанса. Способ измерения являлся аналогичным представленному в примере 1. В результате измерения обнаруживали, что все последовательности связываются с NGF сильнее, чем контрольная 40N.
Аналогично примеру 4, указанные выше аптамеры подвергали укорачиванию. В результате, во всех аптамерах цепь можно укорачивать до структуры, аналогичной консенсусной вторичной структуре 1, при сохранении активности связывания. Фактически полученные нуклеотидные последовательности представлены ниже.
SEQ ID NO: 76:
GGGU(F)C(F)C(F)U(F)GaU(F)U(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aaU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 77:
GGGU(F)C(F)C(F)U(F)GaC(F)U(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aGU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 78:
GGGU(F)C(F)C(F)U(F)GGC(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GGU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 79:
GGGU(F)C(F)C(F)U(F)GGU(F)U(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aaC(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 80:
GGGU(F)C(F)C(F)U(F)GGC(F)U(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aGU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 81:
GGGU(F)C(F)C(F)U(F)GGU(F)GaU(F)aaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)aU(F)C(F)aC(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 82:
GGGU(F)C(F)C(F)U(F)GGaU(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 83:
GGGU(F)C(F)C(F)U(F)GU(F)aC(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GU(F)aC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 84:
GGGU(F)C(F)C(F)U(F)GU(F)C(F)GC(F)U(F)aaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)U(F)GC(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 85:
GGGU(F)C(F)C(F)U(F)GaGU(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)aU(F)aC(F)U(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 86:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)GC(F)GU(F)GGU(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 87:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GGGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 88:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GaaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 89:
GGGU(F)C(F)C(F)U(F)GU(F)GGaaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)C(F)C(F)aC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 90:
GGGU(F)C(F)C(F)U(F)GC(F)GU(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aU(F)GC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 91:
GGGU(F)C(F)C(F)U(F)GC(F)C(F)U(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aGGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 92:
GGGU(F)C(F)C(F)U(F)GaC(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GGU(F)C(F)U(F)GU(F)aC(F)C(F)C(F)
Используя длину нейрита на клетку, полученную при добавлении только NGF, в качестве ингибиторной активности 0% и длину нейрита на клетку, полученную при культивировании без NGF в течение 2 дней, в качестве ингибиторной активности 100%, из длины нейрита на клетку, полученную при культивировании с добавлением NGF и аптамера в смеси, вычисляли ингибиторные активности этих аптамеров в отношении роста нейритов. 50%-ную ингибиторную концентрацию (IC50) определяли из концентраций в двух точках выше и ниже 50%-ной ингибиторной активности. Результаты экспериментов представлены в таблице 1. В таблице 1 значение IC50, указанное как "<X", означает, что ингибиторная активность составляла не менее 50%, если указанная концентрация X являлась минимальной измеряемой концентрацией. Все тестируемые аптамеры демонстрировали сильную ингибиторную активность. Их значения IC50 частично представлены в таблице 1.
Что касается ингибиторной активности в отношении пролиферации клеток TF-1, используя интенсивность люминесценции на лунку, полученную при добавлении только NGF и культивировании клеток в течение 3 дней, в качестве ингибиторной активности 0%, и интенсивность люминесценции на лунку, полученную при культивировании без NGF в течение 3 дней, в качестве ингибиторной активности 100%, из интенсивности люминесценции на лунку, полученной при культивировании с добавлением NGF и аптамера в смеси, вычисляли ингибиторную активность аптамера. 50%-ную ингибиторную концентрацию (IC50) определяли из концентраций в двух точках выше и ниже 50%-ной ингибиторной активности. Результаты представлены в таблице 1. Значение IC50, указанное как "<X", означает, что ингибиторная активность составляла не менее 50%, если указанная концентрация X являлась минимальной измеряемой концентрацией. В результате эксперимента обнаруживали, что значение IC50 аптамеров, иных, чем SEQ ID NO: 81, 84, 86, 89, составляло не более 1 нМ.
5’-сторона всех последовательностей внутренних петель 1 в этих аптамерах представляла собой CCU, а 3’-сторона представляла собой UGUU. Кроме того, петля 2 содержала консенсусную последовательность, представленную 5’-UUUCCXU-3’. В этом случае X являлся G или U. Конечная пара оснований стебля 1 всегда являлась U-a. 1-ая, 8-ая и 9-ая пары оснований стебля 2 представляли собой G-C, a-U и G-C, соответственно. 2-ая - 7-ая содержали некоторые другие пары оснований.
Пример 12: Получение аптамера против NGF - 6
Для получения аптамера, ингибирующего NGF, но не ингибирующего NT-3 и NT-4, осуществляли новый SELEX. В качестве первой совокупности смешивали в соотношении 1:1 и использовали совокупности РНК, впервые используемые в примерах 9 и 10. Перед селекцией совокупность РНК смешивали с NT-3 (производимым R&D Systems, 294 пмоль), NT-4 (производимым R&D Systems, 179 пмоль) и BDNF (производимым R&D Systems, 148 пмоль) и добавляли смесь к бусам с иммобилизированным на них NGF (380 пмоль).
После завершения 4 раундов подтверждали последовательности 48 клонов и не обнаруживали конвергенцию последовательностей. Некоторые единичные последовательности имели те же последовательности, что и SEQ ID NO: 27, 28, 34, 64, 72. Выбирали 14 новых последовательностей, прогнозировали вторичную структуру с использованием программы MFOLD и обнаруживали, что все содержали консенсусную вторичную структуру 1. Таким образом, эти аптамеры подвергали укорачиванию до 40 нуклеотидов аналогично консенсусной вторичной структуре 1. Активность связывания этих укороченных форм с NGF и NT-3 измеряли с помощью способа поверхностного плазмонного резонанса. Для способа измерения использовали BIAcore2000, производимый BIAcore, и в качестве сенсорного чипа использовали CM5, реагирующий на аминогруппу, как представлено в примере 1. Белок иммобилизовывали аналогично примеру 1 с использованием этил-3-карбодиимида гидрохлорида и N-гидроксисукцинимида. NGF или NT-3 человека растворяли в растворе для иммобилизации (10 мМ ацетат натрия, pH 6) и использовали при концентрации 25-40 мкг/мл. После иммобилизации белка осуществляли блокирование этаноламином-HCl. Количество иммобилизованных NGF и NT-3 составляло 3000-4000 RU и 3000-5000 RU, соответственно. Аптамер для аналита получали в концентрации 0,15-0,5 мкМ. Подвижный буфер и раствор для восстановления являлись теми же, что и в примере 1. NT-3 иммобилизовывали на FC2, а NGF на FC3, и получали конечную сенсограмму, вычитая результаты для FC1. Таким образом, обнаруживали 7 последовательностей, сильно связывающихся с NGF. С другой стороны, почти все последовательности не связывались с NT-3.
Аналогично примеру 3 измеряли ингибиторную активность в отношении роста нейритов и обнаруживали, что аптамер, приведенный в SEQ ID NO: 93-98, демонстрировал сильную ингибиторную активность при концентрации 10 нМ. Фактически полученные нуклеотидные последовательности, соответствующие каждому SEQ ID NO, представлены ниже.
SEQ ID NO: 93:
GGGU(F)C(F)C(F)U(F)GC(F)C(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)C(F)GC(F)GGU(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 94:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GGaGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 95:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)aU(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 96:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aU(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 97:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)aC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 98:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)aC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)aC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
5’-сторона всех последовательностей внутренних петель 1 в этих аптамерах представляла собой CCU, а 3’-сторона представляла UGUU. Кроме того, петля 2 содержала консенсусную последовательность, представленную 5’-UUUCCXU-3’. В этом случае X являлся G или a. Конечная пара оснований стебля 1 всегда являлась U-a. 2-ая, 5-ая, 8-ая и 9-ая пары оснований стебля 2 представляли собой C-G, C-G, a-U и G-C, соответственно. В других находились некоторые другие пары оснований.
Осуществляли до 7 раундов SELEX, подтверждали последовательности 48 клонов и обнаружили конвергенцию большинства последовательностей в один тип последовательности. Из остального выбирали 13 единичных последовательностей и исследовали активность связывания с NGF и NT-3 с помощью способа поверхностного плазмонного резонанса и аналогично представленному выше. Таким образом, обнаруживали 7 последовательностей, сильно связывающихся с NGF. С другой стороны, почти все последовательности не связывались с NT-3.
Вторичную структуру этих аптамеров прогнозировали с использованием программы MFOLD и обнаруживали, что ни одна из них не содержит консенсусную вторичную структуру 1. Аналогично примеру 3 измеряли ингибиторную активность в отношении роста нейритов и обнаруживали, что аптамеры, приведенные в SEQ ID NO: 99 и 100, демонстрировали сильную ингибиторную активность при концентрации 10 нМ. С другой стороны, аналогично примеру 8 измеряли ингибиторную активность в отношении пролиферации клеток TF-1 и не обнаруживали ингибиторную активность. Фактически полученные нуклеотидные последовательности, соответствующие каждому SEQ ID NO, представлены ниже.
SEQ ID NO: 99:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGU(F)C(F)C(F)aaaC(F)GGGaC(F)U(F)U(F)U(F)aU(F)aC(F)C(F)U(F)C(F)U(F)GaGU(F)C(F)GC(F)C(F)U(F)U(F)U(F)GC(F)U(F)C(F)C(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
SEQ ID NO: 100:
GGGaGaaC(F)U(F)U(F)C(F)GaC(F)C(F)aGaaGaC(F)C(F)aaaC(F)GGaC(F)U(F)U(F)U(F)aU(F)aC(F)C(F)U(F)C(F)U(F)GaGU(F)C(F)GC(F)C(F)U(F)aU(F)GC(F)U(F)C(F)C(F)U(F)aU(F)GU(F)GC(F)GC(F)aU(F)aC(F)aU(F)GGaU(F)C(F)C(F)U(F)C(F)
Пример 13: Получение аптамера против NGF - 7
Новый SELEX осуществляли аналогично примеру 12 для получения аптамера, ингибирующего NGF, но не ингибирующего NT-3 и NT-4. Матрица и праймеры, впервые используемые для совокупности РНК, описаны ниже.
Матрица 5:
5’-gaggatccatgtatgcgcacatnnnnggatacgagnnnnnnnctcttatccnnnatgtgccaacttctggtcgaagttctccc-3’ (SEQ ID NO: 148)
Подчеркнутая последовательность оснований в последовательности описана ниже.
a: a(70%), g(10%), c(10%), t(10%)
g: a(10%), g(70%), c(10%), t(10%)
c: a(10%), g(10%), c(70%), t(10%)
t: a(10%), g(10%), c(10%), t(70%)
Праймер Fwd3:
5’-taatacgactcactatagggagaacttcgaccagaagttggcaca (SEQ ID NO: 149)
Праймер Rev3:
5’-gaggatccatgtatgcgcaca (SEQ ID NO: 150)
Последовательность матрицы основана на последовательности, приведенной в SEQ ID NO: 82, стебель 2 консенсусной вторичной структуры 1 дополнен 30% случайной последовательности, и участки внутренней петли 1 и петли 2 полностью рандомизированы (n).
После завершения 4 раундов подтверждали последовательности 48 клонов и обнаруживали, что 21 клон являлся идентичным последовательности, приведенной в SEQ ID NO: 22. Из остальных последовательностей 2 клона являлись одинаковыми, а другие являлись единичными последовательностями. Из них выбирали 13 последовательностей, прогнозировали вторичную структуру с использованием программы MFOLD и обнаруживали, что они имели консенсусную вторичную структуру 1. Затем аптамеры подвергали укорачиванию до 40 нуклеотидов аналогично консенсусной вторичной структуре 1. Активность связывания укороченных аптамеров подтверждали с помощью способа поверхностного плазмонного резонанса аналогично примеру 12. NT-4 измеряли аналогично NT-3. В результате, обнаруживали, что укороченные формы сильно связываются с NGF. С другой стороны, связывание с NT-3 и NT-4 являлось слабым.
Аналогично примеру 3 измеряли ингибиторную активность этих укороченных форм в отношении роста нейритов и обнаруживали, что аптамеры, приведенные в SEQ ID NO: 101 и 102, демонстрировали сильную ингибиторную активность, о чем свидетельствовало значение IC50 1 нМ или ниже (таблица 1). С другой стороны, ингибиторную активность в отношении пролиферации клеток TF-1 измеряли аналогично примеру 8 и обнаруживали, что значение IC50 тех же двух аптамеров составляло не менее 1 нМ. Фактически полученные нуклеотидные последовательности, соответствующие каждому SEQ ID NO, представлены ниже.
SEQ ID NO: 101:
GGGU(F)C(F)C(F)U(F)GaC(F)GU(F)aU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GU(F)aU(F)GU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 102:
GGGU(F)C(F)C(F)U(F)GaGC(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)aU(F)GC(F)U(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
Пример 14: Анализ последовательности с использованием высокоскоростного секвенатора
Для получения аптамера, имеющего консенсусную вторичную структуру 1, ингибирующую NGF, но не ингибирующую другие нейротрофины, анализировали последовательности с использованием высокоскоростного секвенатора GS FLX (производимый Roche). Хотя анализ последовательности 48 клонов в примере 1 осуществляли с помощью секвенирования по Сенгеру, использование высокоскоростного секвенатора делает возможным анализ десятков тысяч последовательностей. Измерение и анализ данных осуществляли с помощью Operon, и получение образцов осуществляли по протоколу Operon. Измеряемой целевой ДНК являлась эквимолярная смесь совокупностей ДНК, полученных после завершения 7, 9 и 10 раундов SELEX в примере 8, полученных после завершения 4 и 5 раундов SELEX в примере 12 и полученных после завершения 3 и 4 раундов, полученный SELEX в примере 13.
Общее количество полученных последовательностей составляло 69249. Среди них 40077 последовательностей содержали полностью идентичную последовательность праймера FLX или последовательность праймера FLX, где одно основание являлось замещенным, и частичную последовательность N40 с длиной 40 нуклеотидов. Вторичную структуру этих последовательностей прогнозировали с использованием программы RNAfold и обнаруживали 22453 последовательностей, содержащих ту же структуру, что и консенсусная вторичная структура 1. При сравнении с последовательностями, полученными в примерах 10, 12, 13 с помощью секвенирования по Сенгеру, 99% являлись новыми последовательностями. Среди новых последовательностей 1615 типов последовательностей имели конвергенцию, а 4168 последовательностей являлись единичными последовательностями. Из них выбирали 52 новые последовательности, встречавшиеся очень часто, и подвергали их укорачиванию до 40 нуклеотидов для получения формы консенсусной вторичной структуры 1. Кроме того, снова выбирали 10 единичных последовательностей, полученных в примере 13 с помощью секвенирования по Сенгеру, и аналогично подвергали укорачиванию до 40 нуклеотидов.
С помощью способа поверхностного плазмонного резонанса измеряли связывание укороченных последовательностей с NGF, NT-3, NT-4. Измерение осуществляли аналогично примеру 13. В результате, все последовательности связывались с NGF, и, в частности, следующие 15 последовательностей демонстрировали сильное связывание. С другой стороны, они почти не связывались с NT-3 и NT-4.
Таким образом, ингибиторную активность 15 последовательностей в отношении роста нейритов измеряли способом, аналогичным представленному в примере 3. В результате, обнаруживали, что все аптамеры имели значение IC50 не более 1 нМ (таблица 1). Кроме того, ингибиторную активность в отношении пролиферации клеток TF-1 измеряли способом, аналогичным представленному в примере 8, и обнаруживали, что аптамеры, приведенные в SEQ ID NO: 111, 112, 114-117, имели значение IC50 не более 1 нМ (таблица 1).
Фактически, полученные нуклеотидные последовательности, соответствующие каждому SEQ ID NO, представлены ниже. SEQ ID NO: 111 является последовательностью, полученной в примере 13, с помощью секвенирования по Сенгеру.
SEQ ID NO: 103:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGU(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 104:
GGGU(F)C(F)C(F)U(F)GaU(F)GU(F)C(F)aaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)GaU(F)GU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 105:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)GC(F)U(F)aaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)aGU(F)GaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 106:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)aGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 107:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GaaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)U(F)C(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 108:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 109:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)U(F)U(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 110:
GGGU(F)C(F)C(F)U(F)GC(F)C(F)GC(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)GU(F)GGC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 111:
GGGU(F)C(F)C(F)U(F)GU(F)C(F)U(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aGaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 112:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GGaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 113:
GGGU(F)C(F)C(F)U(F)GC(F)U(F)GU(F)GU(F)aGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)GC(F)aU(F)GGU(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 114:
GGGU(F)C(F)C(F)U(F)GaC(F)aaaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)U(F)GU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 115:
GGGU(F)C(F)C(F)U(F)GU(F)C(F)U(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)aGaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 116:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GaaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
SEQ ID NO: 117:
GGGU(F)C(F)C(F)U(F)GU(F)U(F)GaaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)C(F)aaC(F)U(F)GU(F)U(F)aC(F)C(F)C(F)
5’-сторона всех последовательностей внутренних петель 1 в этих аптамерах представляла собой CCU, а 3’-сторона представляла собой UGUU. Кроме того, петля 2 содержала консенсусную последовательность, представляющую собой 5’-UUUCCXU-3’. В этом случае X является G или U. Конечная пара оснований стебля 1 всегда представляла собой U-a. 8-ая и 9-ая пара оснований стебля 2 представляла собой a-U и G-C, соответственно. 1-ая - 7-ая содержали некоторые другие пары оснований.
Пример 15: Модификация укороченных аптамеров
Для повышения стабильности аптамера в крови получали варианты, где замещали модификацию в 2’-положении рибозы.
Последовательности модифицированных форм представлены ниже. Нуклеотиды в круглых скобках представляют собой модификацию в 2’-положении, F является атомом фтора, M является O-метильной группой, и L является замкнутой нуклеиновой кислотой (ЗНК). Прописными буквами указана РНК, строчными буквами указана ДНК, и idT означает инвертированный dT. Линкер, используемый для 5'-конца, являлся линкером ssH (SAFC) или DMS(O)MT-AMINO-MODIFIFIER C6 (GLEN RESEARCH), и линкер, используемый для 3'-конца, являлся TFA Amino C-6 lcaa CPG (ChemGenes). PEG40GS2 являлся 2-разветвленным PEG типа GS, имеющим молекулярную массу 40000 (SUNBRIGHT GL2-400GS2, производимый NOF CORPORATION), PEG40TS2 являлся 2-разветвленным PEG типа TS, имеющим молекулярную массу 40000 (SUNBRIGHT GL2-400TS, производимый NOF CORPORATION), PEG40TS4 являлся 4-разветвленным PEG типа TS, имеющим молекулярную массу 40000 (SUNBRIGHT GL4-400TS, производимый NOF CORPORATION), PEG80TS2 является 2-разветвленным PEG типа TS, имеющим молекулярную массу 80000 (SUNBRIGHT GL2-800TS, производимый NOF CORPORATION), и PEG80TS4 является 4-разветвленным PEG типа TS, имеющим молекулярную массу 80000 (SUNBRIGHT GL4-800TS, производимый NOF CORPORATION).
SEQ ID NO: 38(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)G(M)U(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)U(F)C(F)G(M)C(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 151:
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)G(M)U(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)GC(F)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 152:
idT-
G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)G(M)U(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)G(M)C(F)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 153:
PEG40GS2-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)G(M)U(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)G(M)C(F)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 154:
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)G(M)U(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)G(M)C(F)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-PEG40GS2
SEQ ID NO: 155:
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)G(M)C(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)aC(F)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 62(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)G(M)G(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 66(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)aU(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)U(F)U(F)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 68(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GaU(F)G(M)C(F)G(M)U(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)G(M)C(F)G(M)U(F)G(M)U(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 71(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)aU(F)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)G(M)U(F)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 74(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)G(M)C(F)aG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)G(M)U(F)G(M)G(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 76(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GaU(F)U(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aaU(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 77(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GaC(F)U(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aG(M)U(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 78(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)C(F)C(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)G(M)G(M)U(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 78(2):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)C(F)C(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)G(M)G(M)U(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 78(3):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)C(M)C(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)G(M)G(M)U(F)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 78(4):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)C(M)C(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)G(M)G(M)U(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 79(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)U(F)U(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aaC(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 80(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)C(F)U(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aG(M)U(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(2):
idT-G(M)G(M)G(M)U(M)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(3):
idT-G(M)G(M)G(M)U(F)C(F)C(M)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(4):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(5):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(M)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(6):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(7):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(M)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(8):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(M)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(9):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(M)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(10):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(M)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(11):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(M)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(12):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(M)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(13):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(M)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(14):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(M)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(15):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(M)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(16):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(M)C(F)C(F)-idT
SEQ ID NO: 82(17):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(M)C(F)-idT
SEQ ID NO: 82(18):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(M)-idT
SEQ ID NO: 82(19):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(M)C(M)G(M)U(F)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(20):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(F)aU(M)C(F)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(21):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(F)G(M)U(M)aU(M)C(F)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(22):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(F)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(23):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(F)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(24):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(F)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(25):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(26):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(M)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(27):
idT-G(M)G(M)G(M)U(F)cC(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 156:
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)tU(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 157:
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)tU(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(30):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)cC(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(31):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)cU(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 158:
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)tGU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 159:
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)taC(F)C(F)C(F)-idT
SEQ ID NO: 160:
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)tU(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 161:
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)tGU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 162:
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)tU(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)tGU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(37):
idT-G(M)G(M)G(M)U(M)C(F)C(M)uGG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(38):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)uU(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(39):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)uC(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(40):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)uC(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(41):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)uaC(M)C(M)C(M)-idT
SEQ ID NO: 82(42):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(43):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aA(M)G(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(44):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)A(M)aG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(45):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(46):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(47):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(48):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 163:
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)tU(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)tGU(M)uaC(M)C(M)C(M)-idT
SEQ ID NO: 164:
idT-G(M)G(M)G(M)U(M)C(F)C(M)uGG(M)aU(M)aaG(M)aG(M)tU(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)tGU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 165:
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)tU(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)tGU(M)uA(M)C(M)C(M)C(M)-idT
SEQ ID NO: 166:
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)aaG(M)A(M)G(M)tU(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)tGU(M)uA(M)C(M)C(M)C(M)-idT
SEQ ID NO: 167:
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)aaG(M)A(M)G(M)tU(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)tGU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 168:
idT-G(M)G(M)G(M)U(M)C(F)C(M)uGG(M)A(M)U(M)aaG(M)A(M)G(M)tU(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)tGU(M)uA(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(55):
PEG80TS4-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 82(56):
PEG40GS2-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GG(M)aU(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(F)G(M)U(F)C(F)U(F)C(F)G(M)U(F)aU(F)C(F)C(F)U(F)GU(F)U(F)aC(F)C(F)C(F)-idT
SEQ ID NO: 82(57):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)sGG(M)A(M)U(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(58):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GsG(M)A(M)U(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(59):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)saaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(60):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)asaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(61):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)aasG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(62):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)sGU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(63):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)A(M)U(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)U(M)C(M)C(M)U(F)GsU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 82(64):
PEG40TS2-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GG(M)aU(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)aU(M)C(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 87(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GC(F)U(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GGGC(F)U(F)GU(F)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 87(2):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GC(F)U(F)C(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)GGGC(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 87(3):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GC(F)U(F)C(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)G(M)G(M)G(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 87(4):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GC(M)U(M)C(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)G(M)G(M)G(M)C(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 87(5):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GC(M)U(M)C(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)G(M)G(M)G(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 88(1):
idT-GGGU(F)C(F)C(F)U(F)GU(F)U(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GaaC(F)U(F)GU(F)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 88(2):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GU(F)U(F)C(F)aaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)GaaC(F)U(F)GU(F)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 88(3):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GU(F)U(F)C(F)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)GaaC(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 88(4):
idT-G(M)G(M)G(M)U(M)C(F)C(M)U(F)GU(M)U(M)C(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)G(M)aaC(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 89(1):
idT-G(M)G(M)G(M)U(F)C(F)C(F)U(F)GU(F)GGaaGaGU(F)U(F)U(F)C(F)C(F)GU(F)C(F)U(F)C(F)GU(F)C(F)C(F)aC(F)U(F)GU(F)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 89(2):
idT-G(M)G(M)G(M)U(F)C(F)C(M)U(F)GU(F)GGaaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)C(F)C(F)aC(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 89(3):
idT-G(M)G(M)G(M)U(F)C(F)C(M)U(F)GU(F)G(M)G(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)C(F)C(F)aC(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 89(4):
idT-G(M)G(M)G(M)U(F)C(F)C(M)U(F)GU(M)G(M)G(M)aaG(M)aG(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)C(M)C(M)aC(M)U(F)GU(M)U(F)aC(M)C(M)C(M)-idT
SEQ ID NO: 89(5):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GU(M)G(M)G(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)C(M)C(M)A(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
SEQ ID NO: 111(1):
G(M)G(M)G(M)U(M)C(F)C(M)U(F)GU(M)C(M)U(M)aaG(M)A(M)G(M)U(F)U(F)U(F)C(F)C(M)G(M)U(M)C(F)U(F)C(M)G(M)U(M)A(M)G(M)A(M)C(M)U(F)GU(M)U(F)A(M)C(M)C(M)C(M)-idT
Активность связывания РНК, приведенной в SEQ ID NO: 82(2), с NGF, NT-3, NT-4 измеряли с помощью способа поверхностного плазмонного резонанса аналогично примеру 13. В результате, обнаруживали, что РНК имела активность связывания с любым белком (фиг.3). Кроме того, ингибирует ли аптамер, приведенный в SEQ ID NO: 82(2), связывание NGF и его рецептора (TrkA или p75), исследовали аналогично примеру 2 с помощью способа поверхностного плазмонного резонанса. В результате, обнаруживали, что аптамер сильно ингибирует связывание NGF и обоих рецепторов (фиг.4 и 5).
Ингибиторную активность всех указанных выше модифицированных форм в отношении роста нейритов измеряли способом, аналогичным представленному в примере 3. В результате, значение IC50 всех модифицированных форм, иных, чем SEQ ID NO: 62(1), составляло не более 1 нМ (таблица 1). В частности, значение IC50 аптамера, приведенного в SEQ ID NO: 162, составляло 0,033 нМ. Кроме того, ингибиторную активность в отношении пролиферации клеток TF-1 измеряли способом, аналогичным представленному в примере 8, и обнаруживали, что почти все аптамеры имели значение IC50 не более 1 нМ (Таблица 1). В частности, значение IC50 аптамера, приведенного в SEQ ID NO: 162, составляло 0,014 нМ. Значение IC50 аптамера, приведенного в SEQ ID NO: 62(1), составляло 0,49 нМ.
Из указанного выше очевидно, что аптамеры могут сохранять ингибиторную активность даже при изменении модификации в 2’-положении рибозы.
Пример 16: Подтверждение перекрестной реактивности с другими нейротрофинами с помощью анализа ингибирования пролиферации клеток TF-1
Используя клетки TF-1, исследовали, ингибирует ли аптамер против NGF BDNF, NT-3, NT-4. Гены рецепторов человека (TrkB, TrkC, p75) для соответствующих нейротрофических факторов встраивали в клетки TF-1 (регистрационный номер ATCC: CRL-2003), являющиеся линией эритролейкозных клеток человека, с использованием ретровирусного вектора для получения клеток, стабильно высокоэкспрессирующих эти рецепторы. Клетки TF-1 со встроенными TrkB и p75 использовали для оценки ингибиторной активности против BDNF, клетки TF-1 со встроенными TrkC и p75 использовали для оценки ингибиторной активности против NT-3 и клетки TF-1 только со встроенным TrkB использовали для оценки ингибиторной активности против NT-4. Эти клетки суспендировали в среде RPMI-1640, содержащей 20% эмбриональную телячью сыворотку и высевали в белый 96-луночный плоскодонный планшет в концентрации 1000 клеток (50 мкл) на лунку. Туда добавляли смешанный раствор 50 мкл BDNF (конечная концентрация 0,074 нМ) или NT-3 (конечная концентрация 0,074 нМ) или NT-4 (конечная концентрация 0,071 нМ) человека и аптамер (конечная концентрация 1 мкМ-0,01 нМ), предварительно прореагировавших при комнатной температуре в течение 30 мин в бессывороточной среде RPMI-1640, через 3 дня в каждую лунку добавляли 100 мкл реагента CellTiter-Glo для анализа CellTiter-Glo Luminescent Cell Viability (производимого Promega), измеряли хемилюминесценцию с помощью микроспектрофотометра для чтения планшетов. Используя интенсивность люминесценции на лунку, полученную при добавлении только BDNF или NT-3, или NT-4, и культивировании клеток в течение 3 дней, в качестве ингибиторной активности 0%, и интенсивность люминесценции на лунку, полученную при культивировании в течение 3 дней без добавления BDNF или NT-3, или NT-4, в качестве ингибиторной активности 100%, из интенсивности люминесценции на лунку, полученной при культивировании с добавлением BDNF или NT-3, или NT-4 и аптамера в смеси, вычисляли ингибиторную активность аптамера. Если ингибиторная активность составляла 0 или менее, указывали "0%". 50%-ную ингибиторную концентрацию (IC50) определяли из концентраций в двух точках, выше и ниже, 50%-ной ингибиторной активности. Результаты экспериментов представлены в таблице 2. Значение IC50, указанное как ">X", означает, что ингибиторная активность составляла не более 50%, если указанная концентрация X являлась максимальной измеряемой концентрацией. "N.D." означает отсутствие измерений.
Все тестируемые аптамеры демонстрировали сильную ингибиторную активность. Их значения IC50 частично представлены в таблице 2. Как представлено в таблице 2, в которой описывают ингибиторную активность аптамеров по настоящему изобретению, значение IC50 для NGF составляло не более 0,1 нМ, в то время как для BDNF оно составляло не менее 1000 нМ. Значение IC50 для NT-3 варьировалось от 0,97 нМ до не менее чем 10 нМ в зависимости от аптамера. Значение IC50 для NT-4 в зависимости от аптамера варьировалось от не более 3 нМ до не менее 30 нМ.
Пример 17: Анальгезирующее действие аптамера против NGF
Для исследования анальгезирующего действия аптамера против NGF в отношении индуцируемой NGF боли использовали модель термической гипералгезии, индуцируемой подкожным введением NGF в заднюю конечность крысы. Для эксперимента использовали крыс Jcl:SD (возрастом 6 недель). В качестве показателя термической гипералгезии использовали время ответа в виде реакции избегания на инфракрасное облучение подошвы устройством измерения термической стимуляции подошвы (производимым Ugo Basile). В предыдущий день тестирования осуществляли акклимацию к системе оценивания. В день тестирования перед введением измеряли время ответа в виде реакции избегания и использовали животных, демонстрировавших не менее чем 10 сек и менее чем 20 сек. β-NGF человека (R&D Systems, конечная концентрация 50 мкг/мл) и тестируемое вещество смешивали с наполнителем (20 мМ Tris-HCl (pH 7,6), 145 мМ NaCl, 5,4 мМ KCl, 0,8 мМ MgCl2, 1,8 мМ CaCl2, 0,1% BSA), инкубировали при комнатной температуре в течение 30 мин и подкожно вводили в подошву левой задней конечности в концентрации 10 мкл. Через 5 часов измеряли время ответа в виде реакции избегания. Аптамер, приведенный в SEQ ID NO: 153, вводили в конечной концентрации 50 мг/мл (молярное соотношение с NGF: 1000-кратное). В качестве контроля аналогично вводили наполнитель или смесь наполнителя и NGF. Результаты представлены в таблице 3 (среднее ± SEM, n=9).
Через 5 часов после введения группа NGF демонстрировала значимо низкое время ответа в виде реакции избегания по сравнению с группой наполнителя (p<0,01). Через 5 часов после введения время ответа в виде реакции избегания в группе, которой вводили аптамер, являлось высоким (p<0,01) по сравнению с группой, которой вводили только NGF. Учитывая представленные выше результаты, обнаруживали, что этот аптамер можно использовать в качестве лекарственного средства против индуцируемой NGF боли.
Пример 18: Анальгезирующее действие аптамера против NGF в модели послеоперационной боли
Для исследования эффективности терапии аптамером против NGF использовали модель послеоперационной боли, индуцируемой термической гипералгезией. Для эксперимента использовали крыс Crl:CD (SD) (возрастом 5 недель). Наконечник катетера вводили в бедренную вену, другой конец оставляли на спине крысы. Через одну неделю крысе устанавливали инфузионную систему Quick connect infusion system (производимую Strategic applications incorporated), через одну неделю оценивали термическую гипералгезию. В качестве показателя термической гипералгезии использовали время ответа в виде реакции избегания на инфракрасное излучение подошвы устройством измерения термической стимуляции подошвы (производимым Ugo Basile). Акклимацию к системе оценки проводили за 3 дня до начала тестирования. В день тестирования измеряли время ответа в виде реакции избегания и использовали животных, демонстрирующих время не менее 10 сек и менее 20 сек. Аптамер против NGF, растворенный в физиологическом растворе, вводили внутривенно с помощью шприцевого насоса (производимого TERUMO CORPORATION) на постоянной основе. В качестве аптамера против NGF использовали аптамер, приведенный в SEQ ID NO: 82(56) (вводимый в концентрации 21,2 мг/240 мл/кг/96 часов), и аптамер, приведенный в SEQ ID NO: 82(55) (вводимый в концентрации 10,08 мг/240 мл/кг/96 часов). В качестве контроля аналогичным образом вводили наполнитель. Через 1 ч после начала введения рассекали кожу и фасцию правой задней конечности, мышцу-сгибатель вертикально разделяли пополам и сшивали кожу. Измеряли время ответа в виде реакции избегания после операции рассечения через 1, 2, 3, 4 дня. Результаты представлены в таблице 4.
Группа, которой вводили наполнитель, демонстрировала значимо низкое (p<0,01) время ответа в виде реакции избегания через 1, 2, 3, 4 дня после введения-операции рассечения по сравнению с данными до введения-операции рассечения. Через 1, 2, 3, 4 дня после введения-операции рассечения время ответа в виде реакции избегания в любой группе, которой вводили аптамер, являлось значимо высоким (p<0,01) по сравнению с группой, которой вводили наполнитель. Результаты эксперимента представлены в таблице 4 и таблице 5 (среднее ± SEM, n=8-9). Это показало, что аптамер против NGF оказывает анальгезирующее действие в модели послеоперационной боли.
Таблица 5
Промышленная применимость
Аптамер по настоящему изобретению может являться применимым в качестве лекарственных средств, средств для диагностики или реагентов для заболеваний, таких как алгия, воспалительное заболевание и т.п. Аптамер и комплекс по настоящему изобретению также могут являться применимыми для очистки и концентрирования NGF, а также определения и количественного анализа NGF.
Эта заявка основана на патентной заявке № 2011-213585, зарегистрированной в Японии (дата регистрации: 28 сентября 2011года), содержание которой включено в настоящее описание в полном объеме.




































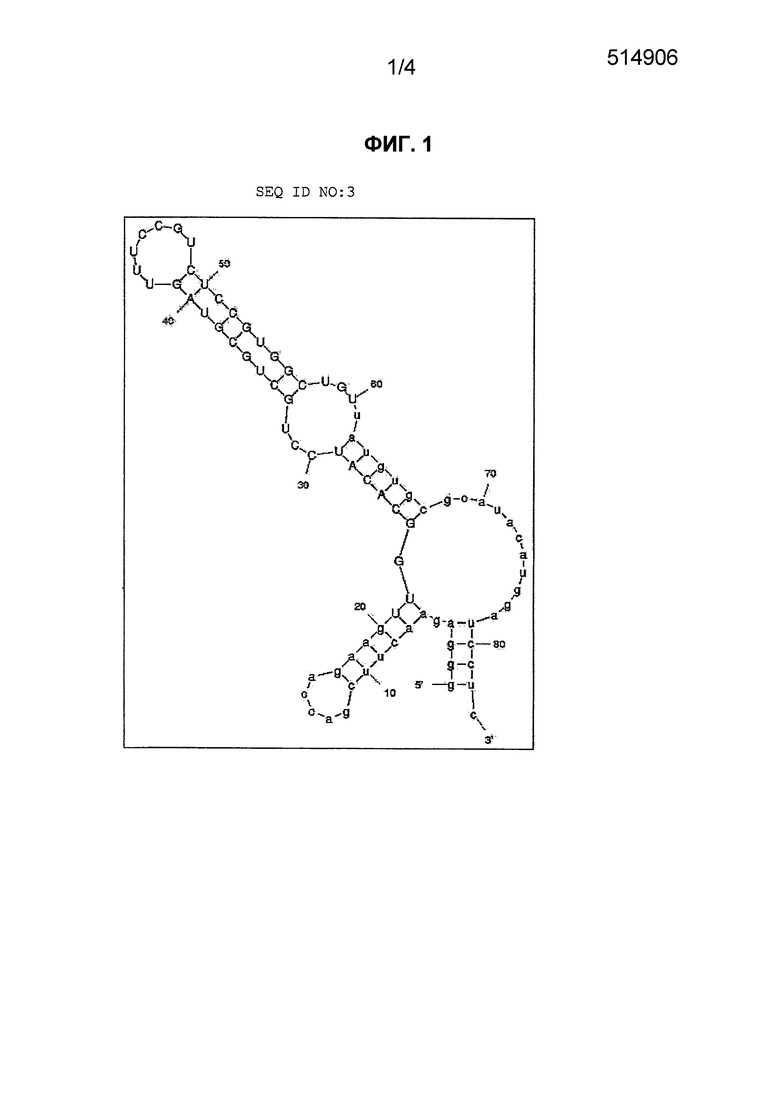
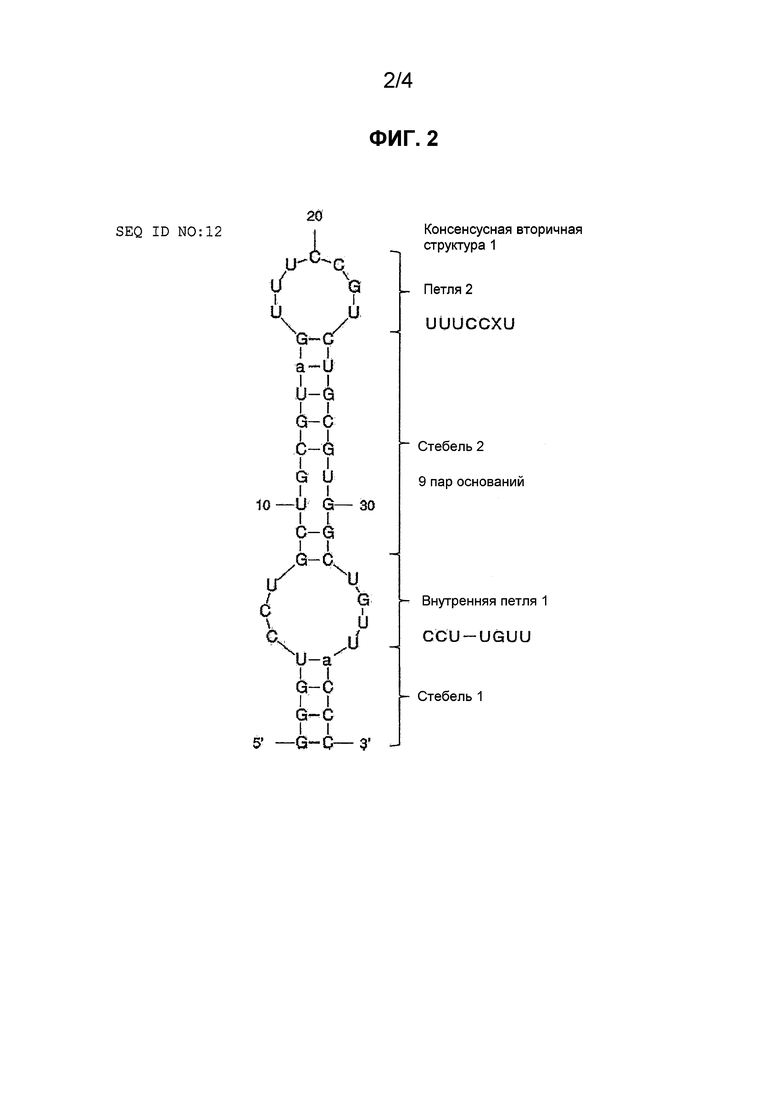
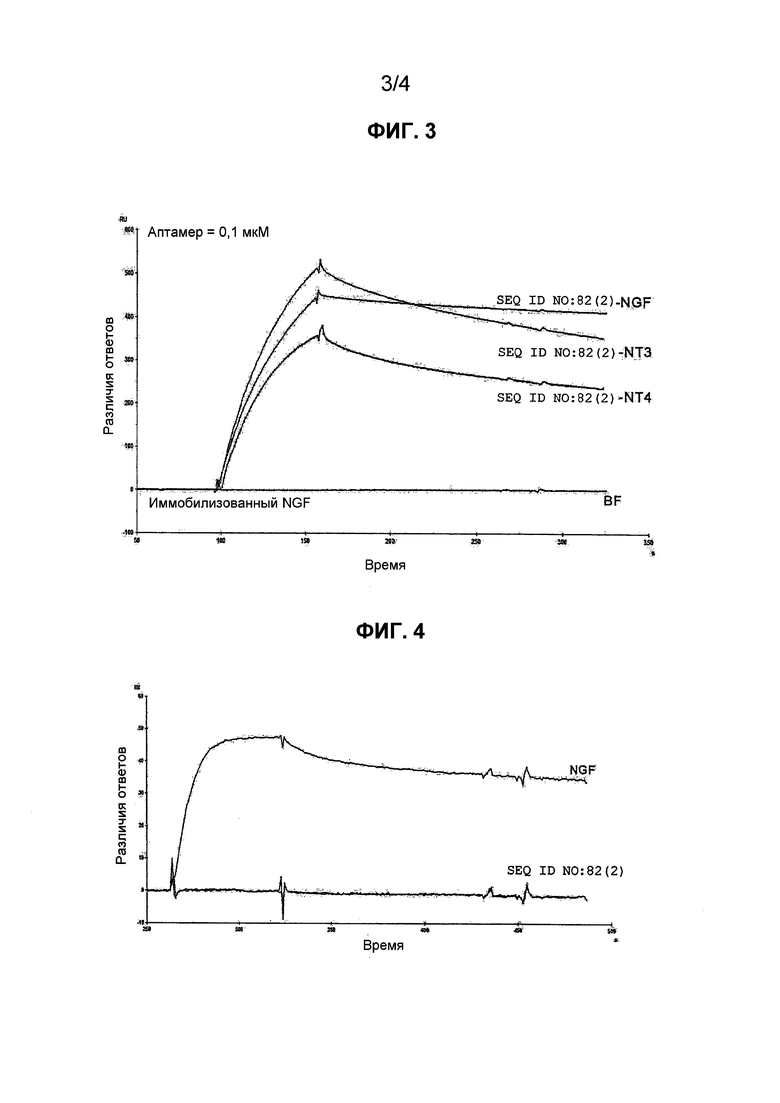
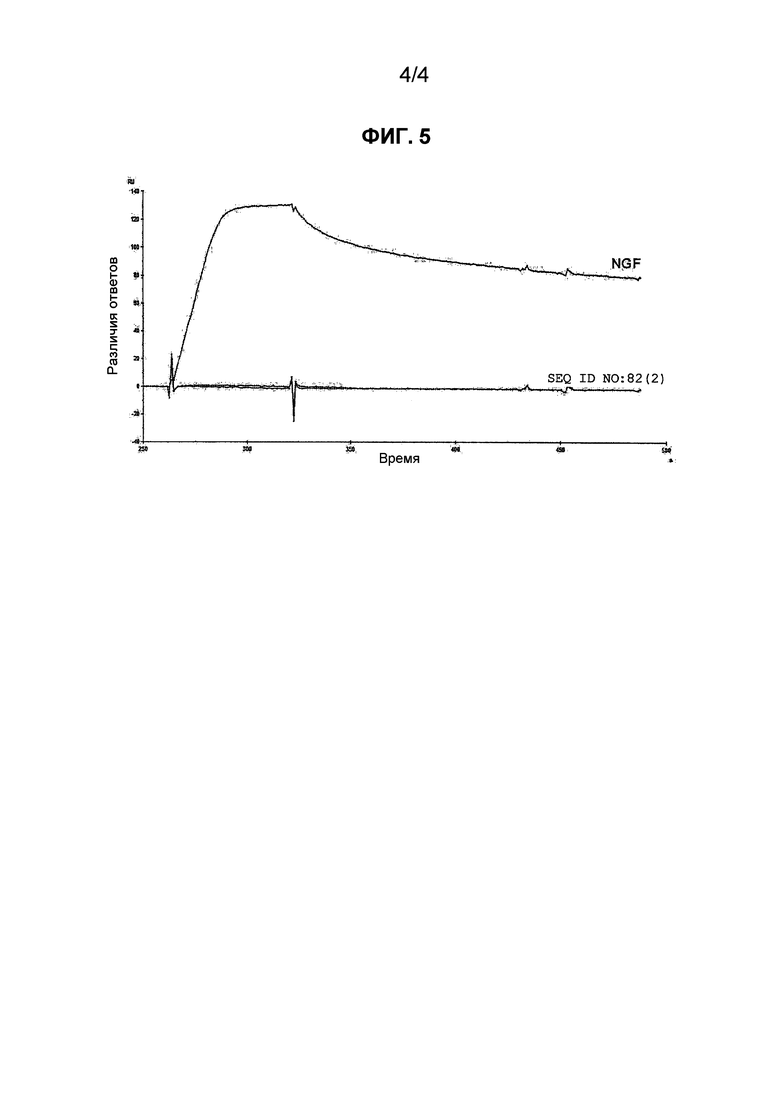
Изобретение относится к области биотехнологии, конкретно к аптамеру, специфически связывающемуся с фактором роста нервов (NGF), и может быть использовано в медицине как противовоспалительное или обезболивающее средство. Изобретение позволяет получить аптамер, способный образовывать вторичную структуру, представленную формулой (I)
 ,
,
где N является нуклеотидом, выбранным из группы, состоящей из A, G, C и U, притом, что каждый из N14 и N24, N31 и N41, и N39 и N49 образует уотсон-криковскую пару оснований, а остальные нуклеотиды формулы (I) способны образовывать структуру стебля. Изобретение позволяет получить аптамер, способный ингибировать активность NGF при IC50 1 нМ или менее. 3 н. и 2 з.п. ф-лы, 5 ил., 5 табл., 18 пр.
1. Аптамер, который связывается с фактором роста нервов (NGF) и ингибирует активность NGF при IC50 1 нМ или менее, где аптамер способен образовывать вторичную структуру, представленную формулой (I)

где N является одним нуклеотидом, выбранным из группы, состоящей из A, G, С и U,
N11, N12, N13, N21, N22, N23, N32, N33, N34, N35, N36, N37, N38, N42, N43, N44, N45, N46, N47 и N48 являются одинаковыми или разными, и каждый из них является связью или 1, или 2 нуклеотидами, выбранными из группы, состоящей из A, G, С и U,
N14, N24, N31, N41, N39 и N49 являются одинаковыми или разными, и каждый из них является одним нуклеотидом, выбранным из группы, состоящей из A, G, С, U и Т,
каждый N14 и N24, N31 и N41, и N39 и N49 образуют уотсон-криковскую пару оснований,
N11-N12-N13-N14 и N21-N22-N23-N24 являются нуклеотидными последовательностями, в комбинации способными образовывать структуру стебля, и
N31-N32-N33-N34-N35-N36-N37-N38-N39 и N41-N42-N43-N44-N45-N46-N47-N48-N49 являются нуклеотидными последовательностями, в комбинации способными образовывать структуру стебля, и
где аптамер имеет:
(а) нуклеотидную последовательность, выбранную из последовательностей, представленных SEQ ID NO: 38, SEQ ID NO: 66, SEQ ID NO: 68, SEQ ID NO: 71, SEQ ID NO: 74, SEQ ID NO: 76-85, SEQ ID NO: 87-89, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 104-112, SEQ ID NO: 114-117 и SEQ ID NO: 151-155, или
(b) нуклеотидную последовательность по приведенному выше (а), где от 1 до 4 нуклеотидов, отличных от нуклеотидов внутренней петли 1 и петли 2 (N исключен), подвергают замене, делеции, инсерции или добавлению, и
где аптамер может необязательно включать модифицированный нуклеотидный остаток(остатки), такой как инвертированный dT, или концевой нуклеотидный остаток(остатки), модифицированный полиэтиленгликолем, и/или нуклеотид(нуклеотиды), в котором модифицирован сахарный остаток в 2'-положении.
2. Аптамер по п. 1, который имеет длину не более 50 оснований.
3. Аптамер по п. 1, который дополнительно ингибирует активность NGF в отношении пролиферации клеток.
4. Средство против боли, содержащее аптамер по любому из пп. 1-3, где боль связана с активацией путей передачи сигналов через рецептор TrkA или рецептор р75.
5. Противовоспалительное средство, содержащее аптамер по любому из пп. 1-3, где воспаление связано с активацией путей передачи сигналов через рецептор TrkA или рецептор р75.
| WO 2010035725 A1, 01.04.2010 | |||
| WO 2010143714 A1, 16.12.2010 | |||
| RU 2009122476 A, 20.12.2010. |

