СВЯЗАННЫЕ ЗАЯВКИ
По данной патентной заявке испрашивается приоритет по условным патентным заявкам США с серийными номерами № 60/780905, поданной 8 марта 2006 года, и 60/848274, поданной 29 сентября 2006 года, все из которых включены в настоящий документ в качестве ссылок в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится, главным образом, к области нуклеиновых кислот и, более конкретно, к аптамерам, способным связываться с белками системы комплемента, пригодным в качестве лекарственных средств и диагностических средств при связанных с комплементом глазных, сердечных, воспалительных, астматических и аутоиммунных нарушениях, ишемическом реперфузионном повреждении и/или других заболеваниях или нарушениях, в которые, в частности, вовлечена опосредуемая C5 активация комплемента. В предпочтительных вариантах осуществления, настоящее изобретение, более конкретно, относится к способам и материалам для лечения и выявления глазных нарушений, включая, но не ограничиваясь этим, лечение и выявление опосредуемых C5 нарушений, таких как опосредуемые C5 глазные нарушения. Кроме того, настоящее изобретение относится к материалам и способам для введения аптамеров, способных связывать белки системы комплемента, включая белки C5.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Аптамер по определению представляет собой выделенную молекулу нуклеиновой кислоты, которая связывается с высокой специфичностью и аффинностью с определенной мишенью, такой как белок, посредством взаимодействий, отличных от образования пар по принципу Уотсона-Крика. Несмотря на то, что аптамеры представляют собой молекулы на основе нуклеиновых кислот, существует фундаментальное отличие между аптамерами и другими молекулами нуклеиновых кислот, такими как гены и мРНК. В последних структура нуклеиновой кислоты кодирует информацию через ее линейную последовательность оснований и, таким образом, эта последовательность важна для функции хранения информации. В противоположность этому, функция аптамера, которая основана на специфичном связывании молекулы-мишени, не зависит от консервативной линейной последовательности оснований, а зависит от конкретной вторичной/третичной структуры. Значит, аптамеры представляют собой некодирующие последовательности. Любой потенциал в отношении кодирования, которым может обладать аптамер, является полностью случайным и абсолютно не участвует в связывании аптамера с его собственной мишенью. Таким образом, несмотря на то, что возможно, чтобы аптамеры, которые связываются с одной мишенью и даже с одним участком на этой мишени, обладали сходной линейной последовательностью оснований, большинство из них не обладают ей.
Также аптамеры необходимо отличать от встречающихся в природе последовательностей нуклеиновых кислот, которые связываются с определенными белками. Эти последние последовательности представляют собой встречающиеся в природе последовательности, заключенные в геном организма, которые связываются со специализированной подгруппой белков, которые вовлечены в транскрипцию, трансляцию и транспорт встречающихся в природе нуклеиновых кислот, т.е. связывающих нуклеиновые кислоты белков. С другой стороны, аптамеры представляют собой короткие, выделенные, не встречающиеся в природе молекулы нуклеиновых кислот. Несмотря на то, что могут быть идентифицированы аптамеры, которые связывают связывающие нуклеиновые кислоты белки, в большинстве случаев такие аптамеры обладают небольшой идентичностью с последовательностями, распознаваемыми связывающими нуклеиновую кислоту белками в природе, или не обладают ей. Более важным является то, что аптамеры могут связываться практически с любым белком (не только со связывающими нуклеиновые кислоты белками), а также практически с любой представляющей интерес мишенью, включая низкомолекулярные соединения, углеводы, пептиды и т.д. Для большинства мишеней, даже белков, встречающаяся в природе последовательность нуклеиновой кислоты, с которой они связываются, не существует; для тех мишеней, которые имеют такую последовательность, т.е. связывающих нуклеиновую кислоту белков, такие последовательности будут отличаться от аптамеров в результате относительно низкой аффинности связывания, используемой в природе по сравнению с сильно связывающимися аптамерами.
Аптамеры, подобно пептидам, полученным посредством фагового дисплея, или антителам, способны специфично связывать выбранные мишени и модулировать активность или связывающие взаимодействия мишени, например, связывающие аптамеры могут блокировать способность их мишени к функционированию. Как и в случае антител, это функциональное свойство специфичного связывания с мишенью представляет собой присущее им свойство. Так же, как в случае антител, несмотря на то, что квалифицированный специалист может не знать, какими точно структурными свойствами будет обладать аптамер к мишени, квалифицированному специалисту известно, как идентифицировать, создавать и применять такую молекулу в отсутствие точной структурной определенности.
Также аптамеры аналогичны низкомолекулярным лекарственным средствам в том, что единичное структурное изменение, каким бы незначительным оно ни казалось, может значительно влиять (на несколько порядков величины) на связывание и/или другой вид активности (или виды активности) аптамера. С другой стороны, некоторые структурные изменения будут оказывать небольшой эффект или не оказать никакого эффекта. Это является следствием важности вторичной/третичной структуры аптамеров. Иными словами, аптамер представляет собой трехмерную структуру, находящуюся в фиксированной конформации, которая обеспечивает химические контакты для специфичного связывания данной его мишени. Следовательно: (1) некоторые области или конкретные последовательности являются существенными в качестве (a) специфичных точек контакта с мишенью, и/или в качестве (b) последовательностей, которые обеспечивают контакт молекул с мишенью; (2) некоторые области или конкретные последовательности обладают диапазоном вариабельности, например нуклеотид X должен быть пиримидином, или нуклеотид Y должен быть пурином, или нуклеотиды X и Y должны быть комплементарными; и (3) некоторые области или конкретные последовательности могут быть любыми, т.е. по существу они представляют собой промежуточные элементы, например, они могут представлять собой любую цепь нуклеотидов данной длины или даже ненуклеотидный спейсер, такой как молекула PEG.
Открытые посредством процесса селекции in vitro из пулов олигонуклеотидов со случайной последовательностью, аптамеры были получены свыше чем для 130 белков, включая факторы роста, транскрипционные факторы, ферменты, иммуноглобулины и рецепторы. Типичный аптамер обладает размером 10-15 кДа (20-45 нуклеотидов), связывает его мишень с от наномолярной до субнаномолярной аффинностью, и отличает высокосходные мишени (например, аптамеры, как правило, не связывают другие белки из того же семейства генов). В ряде структурных исследований было показано, что аптамеры способны использовать те же типы связывающих взаимодействий (например, образование водородных связей, электростатическую комплементарность, гидрофобные взаимодействия, пространственное вытеснение), которые определяют аффинность и специфичность в комплексах антитело-антиген.
Аптамеры обладают рядом свойств, желательных для применения в качестве средств для диагностики и лечения, включая высокую специфичность и аффинность, биологическую эффективность и превосходные фармакокинетические свойства. Кроме того, они обладают определенными преимуществами, обеспечивающими конкурентоспособность, над антителами и другими белковыми биологическими веществами, например:
1) Скорость и контроль. Аптамеры полностью получают в процессе in vitro, что обеспечивает быстрое получение исходных цепей, включая терапевтические цепи. Отбор in vitro обеспечивает строгий контроль специфичности и аффинности и обеспечивает возможность получения цепей, включающих цепи против как токсичных, так и неиммуногенных мишеней.
2) Токсичность и иммуногенность. Было показано, что аптамеры как класс обладают терапевтически приемлемой токсичностью и отсутствием иммуногенности. В то время как эффективность множества моноклональных антител может быть значительно ограничена иммунным ответом на сами антитела, крайне трудно вызвать образование антител к аптамерам, наиболее вероятно, вследствие того, что аптамеры не могут быть представлены T-клетками посредством MHC, и иммунный ответ, главным образом, не направлен на распознавание фрагментов нуклеиновых кислот.
3) Введение. В то время как большинство одобренных к настоящему времени лекарственных средств на основе антител вводят посредством внутривенной инфузии (как правило, в течение 2-4 час), аптамеры можно вводить подкожной инъекцией (биодоступность аптамеров при подкожном введении в исследованиях на обезьянах составляет >80% (Tucker et al, J. Chromatography B. 732: 203-212, 1999)). Это отличие, главным образом, является следствием сравнительно низкой растворимости и, таким образом, для большинства терапевтических mAb необходимы большие объемы. При хорошей растворимости (>150 мг/мл) и сравнительно низкой молекулярной массе (аптамер: 10-50 кДа антитела: 150 кДа), недельная доза аптамера может быть доставлена инъекцией объемом менее 0,5 мл. Кроме того, небольшой размер аптамеров позволяет им проникать в области конформационных сужений, в которые не могут проникать антитела или фрагменты антител, что представляет собой дополнительное преимущество средств для лечения и профилактики на основе аптамеров.
4) Возможность увеличения масштаба продукции и стоимость. Терапевтические аптамеры химически синтезируют, и, таким образом, масштаб синтеза может быть легко увеличен для удовлетворения производственных потребностей по мере необходимости. В то время как сложность масштабного изготовления в настоящее время ограничивает доступность некоторых биологических препаратов, и затраты капитала на продуцирующие белки растения являются очень большими, с помощью отдельного устройства для крупномасштабного синтеза олигонуклеотидов можно получать свыше 100 кг/год и для этого требуется относительно небольшое первоначальное капиталовложение. Текущая стоимость материалов для синтеза аптамеров на килограммовом уровне оценивается как $500/г, что сравнимо со стоимостью высокооптимизированных антител. Ожидается, что продолжающиеся усовершенствования в развитии производственного процесса снизят стоимость материалов до <$100/г в течение пяти лет.
5) Стабильность. Терапевтические аптамеры являются химически устойчивыми. Они по своей природе адаптированы для восстановления активности после воздействия факторов, таких как нагревание и денатурирующие вещества, и в виде лиофилизированных порошков могут храниться в течение длительного периода времени (>1 года) при комнатной температуре. Напротив, антитела необходимо хранить замороженными.
Система комплемента. Система комплемента содержит набор по меньшей мере из 20-30 белков плазмы и мембранных белков, которые действуют совместно в регулируемой каскадной системе для атаки на внеклеточные формы патогенов (например, бактерии). Система комплемента включает три различных каскада активации ферментов: классический, лектиновый и альтернативный каскады (Фиг.1), которые сходятся на активации C5 и приводят к неферментативному каскаду, известному как мембраноатакующий каскад.
Первый ферментативно активируемый каскад, известный как классический каскад, включает несколько компонентов, C1, C4, C2, C3 и C5 (перечисленные по порядку в каскаде). Инициация классического каскада системы комплемента происходит после связывания и активации первого компонента комплемента (C1) как иммунными, так и неиммунными активаторами. C1 включает кальций-зависимый комплекс компонентов C1q, C1r и C1s и активируется посредством связывания компонента C1q. C1q содержит шесть идентичных субъединиц, и каждая субъединица содержит три цепи (цепи A, B и C). Каждая цепь обладает глобулярной областью головки, которая соединена с коллаген-подобным хвостом. Связывание и активация C1q комплексами антиген-антитело происходит через область головки C1q. Множество не являющихся антителами активаторов C1q, включая белки, липиды и нуклеиновые кислоты, связывают и активируют C1q через отдаленную область на коллаген-подобной области стебля. Молекулярное распознавание активаторов комплемента посредством C1q индуцирует изменение конформации, которое стимулирует аутоактивацию профермента C1r, который, в свою очередь, катализирует протеолитическую активацию C1s. Затем Cs катализирует активацию компонентов комплемента C4 и C2, образующих комплекс C4bC2a, который функционирует в качестве конвертазы C3.
Второй ферментативно активируемый каскад, известный как лектиновый каскад, является сходным с первым, за исключением того, что вместо комплекса C1 находится MBL/MASP-2. Маннан-связывающий лектин (MBL) прямо распознает содержащие маннозу полисахариды на поверхностях бактерий и является структурно и функционально гомологичным компоненту C1q из C1. Связывание MBL с активатором индуцирует активацию ассоциированной с MBL протеазы 2 (MASP-2). MASP-2, в свою очередь, катализирует активацию C4 и C2 способом, гомологичным функции C1s, что приводит к образованию конвертазы C3.
Третий ферментативно активный каскад, известный как альтернативный каскад, представляет собой быстрый, независимый от антител путь активации и усиления системы комплемента. Альтернативный каскад включает несколько компонентов: C3, фактор B и фактор D (приведенные по порядку в каскаде). Активация альтернативного каскада происходит, когда C3b, протеолитически расщепленная форма C3, связывается с активирующим поверхностным агентом, таким как бактерия. Затем с C3b связывается фактор B, и он расщепляется фактором D с образованием C3-конвертазы C3bBb. Усиление активности конвертазы C3 происходит по мере образования и накопления дополнительного C3b. Кроме того, усиленному ответу способствует связывание положительного регуляторного белка пропердина (P), который стабилизирует конвертазу против деградации, продлевая ее время полужизни от 1-2 минут до 18 минут.
Таким образом, все три каскада приводят к образованию конвертаз C3, которые расщепляют фактор C3 на C3a и C3b. В это время обе конвертазы C3 (классическая/лектиновая и альтернативная) далее собираются в конвертазы C5 (C4b2a3b и C3b3bBb). Эти комплексы впоследствии расщепляют компонент комплемента C5 на два компонента: полипептид C5a (9 кДа) и полипептид C5b (170 кДа). Полипептид C5a связывается с 7-кратно трансмембранным сопряженным с G-белком рецептором, который исходно ассоциировали с лейкоцитами и о котором в настоящее время известно, что он экспрессируется в различных тканях, включая гепатоциты и нейроны. Молекула C5a представляет собой первичный хемотактический компонент системы комплемента человека и может запускать различные биологические ответы, включая хемотаксис лейкоцитов, сокращение гладких мышц, активацию внутриклеточных каскадов передачи сигнала, нейтрофил-эндотелиальную адгезию, высвобождение цитокинов и липидных медиаторов и образование окисляющих веществ.
Более крупный фрагмент C5b последовательно связывается с последующими компонентами каскада комплемента, C6, C7, C8 и C9, с образованием мембраноатакующего комплекса ("MAC") C5b-9. MAC C5b-9 может непосредственно лизировать эритроциты, и в больших количествах он является литическим для лейкоцитов и повреждает ткани, такие как мышечные, эпителиальные и эндотелиальные клетки. В сублитических количествах MAC может стимулировать активацию молекул адгезии, повышение внутриклеточного содержания кальция и высвобождение цитокинов. Кроме того, MAC C5b-9 может стимулировать клетки, такие как эндотелиальные клетки и тромбоциты, не вызывая лизис клеток. Нелитические эффекты C5a и MAC C5b-9 иногда являются высокосходными.
Несмотря на то, что система комплемента играет важную роль в поддержании состояния здоровья, она может вызывать или участвовать в заболевании. Например, система комплемента вовлечена в побочные эффекты, связанные с хирургической операцией по аортокоронарному шунтированию ("CABG"), множеством почечных, ревматологических, неврологических, дерматологических, гематологических, сосудистых/легочных, аллергических, инфекционных и связанных с биологической совместимостью/шоком заболеваний и/или состояний. Система комплемента необязательно является единственной причиной болезненного состояния, однако она может представлять собой один из нескольких факторов, которые участвуют в патогенезе.
Недавно данные подтвердили, что комплемент также вовлечен в заболевания глаз. Таким образом, может быть полезным наличие новых ингибиторов системы комплемента для применения в качестве лекарственных и диагностических средств для лечения связанных с комплементом глазных нарушений.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На Фиг.1 представлена иллюстрация классического и альтернативного каскадов системы комплемента.
На Фиг.2 представлена схема процесса отбора аптамеров in vitro (SELEXTM) из наборов олигонуклеотидов со случайной последовательностью.
На Фиг.3A представлена иллюстрация, на которой изображена нуклеотидная последовательность и вторичная структура аптамера против C5 (SEQ ID NO:1), в котором подчеркнутые остатки представляют собой либо остатки 2'-H-пиримидина, либо остатки 2'-фторпиримидина, заключенные в рамку остатки представляют собой либо остатки 2'-фторпиримидина, либо остатки 2'-OMe-пиримидина, и остатки, указанные стрелкой (→), представляют собой остатки, которые должны сдержать модификацию 2'-фтор.
На Фиг.3B представлена иллюстрация, на которой изображена нуклеотидная последовательность и вторичная структура аптамера против C5 ARC330 (SEQ ID NO:2), в котором обведенные кружком остатки представляют собой 2'-H-остатки, остатки пиримидина являются 2'-фтор-замещенными, и большинство остатков пурина являются 2'-OMe-замещенными, за исключением остатков 2'-OH-пурина, показанных контурным изображением.
На Фиг.3C представлена иллюстрация, на которой изображена нуклеотидная последовательность и вторичная структура аптамера против C5 ARC186 (SEQ ID NO:4), в котором все 21 остатка пиримидина обладают 2'-фтор-модификациями, и большинство пуринов (14 остатков) обладают 2'-OMe-модификациями, за исключением трех остатков 2'-OH-пуринов, показанных контурным изображением.
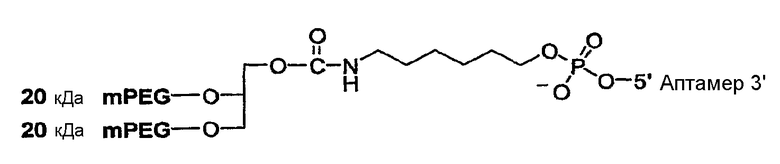
На Фиг.4 представлена иллюстрация разветвленного PEG массой 40 кДа (1,3-бис(mPEG-[20 кДа])-пропил-2-(4'-бутамид)).
На Фиг.5 представлена иллюстрация разветвленного PEG массой 40 кДа (1,3-бис(mPEG-[20 кДа])-пропил-2-(4'-бутамид), присоединенного к 5'-концу аптамера.
На Фиг.6 представлена иллюстрация, на которой изображены различные стратегии синтеза конъюгатов высокомолекулярный PEG-нуклеиновая кислота.
На Фиг.7A представлен график, на котором приведено сравнение зависимого от дозы ингибирования гемолиза посредством пегилированных аптамеров против C5 (ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5)), и непегилированного аптамера против C5 (ARC186 (SEQ ID NO:4)); на Фиг.7B представлена таблица значений IC50 для аптамеров, применяемых для анализа гемолиза, изображенного на Фиг.7A; На Фиг.7C представлен график для сравнения зависимого от дозы ингибирования гемолиза пегилированными аптамерами против С5 ARC187 (SEQ ID NO:5), ARC1537 (SEQ ID NO:65), ARC1730 (SEQ ID NO:66) и ARC1905 (SEQ ID NO:67); на Фиг.7D представлена таблица значений IC50 для аптамеров, применяемых для анализа гемолиза, представленного на Фиг.7C.
На Фиг.8 представлен график процентного ингибирования гемолиза аптамером против C5, ARC658 (SEQ ID NO:62), комплемента сыворотки яванских макак против комплемента сыворотки человека.
На Фиг.9 представлен график, на котором изображено связывание ARC186 (SEQ ID NO:4) с очищенным белком C5 как при 37°С, так и при комнатной температуре (23°С) после инкубации в течение 15 минут.
На Фиг.10 представлен другой график, на котором изображено связывание ARC186 (SEQ ID NO:4) с очищенным белком C5 как при 37°С, так и при комнатной температуре (23°С) после инкубации в течение 4 часов.
На Фиг.11 представлен график, на котором показан ход диссоциации комплекса C5-ARC186 при 23°С.
На Фиг.12 представлен график, на котором представлен ход уравновешивания образования комплекса C5-ARC186 комплекс при 23°С.
На Фиг.13 представлен график, на котором изображено связывание ARC186 (SEQ ID NO:4) с белком C5 относительно предшествующих и последующих белковых компонентов каскада комплемента.
На Фиг.14 представлен график, на котором изображено процентное количество меченного радиоактивно меткой ARC186 (SEQ ID NO:4), который связывает C5 в присутствии немеченого конкурента ARC186 (SEQ ID NO:4), ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) или ARC187 (SEQ TD NO:5).
На Фиг.15 представлен график, на котором изображено количество белка комплемента C5b, образованного в образцах крови, инкубированных в течение 5 часов при 25°С и 37°С в присутствии варьирующих концентраций аптамера ARC186 (SEQ ID NO:4).
На Фиг.16 представлен график, на котором изображено процентное ингибирование комплемента посредством ARC187 (SEQ ID NO:5) в присутствии зимозана в неразбавленной сыворотке человека, цитратной крови человека или в сыворотке яванской макаки.
На Фиг.17 представлен график, на котором показано, что ARC658 (SEQ ID NO:62) полностью ингибирует активацию комплемента (C5a) в модели с петлей трубки, описанной в примере 1D.
На Фиг.18 представлен график, на котором изображены константы диссоциации пулов C5 для 10 раунда селекции. Константы диссоциации (Kd) оценивали подстановкой данных в уравнение: фракция связанной РНК = амплитуда*Kd/(Kd + [C5]). "ARC520" (SEQ ID NO:70) относится к интактному не подвергнутому селекции пулу dRmY и "+" указывает на наличие конкурента (0,1 мг/мл тРНК, 0,1 мг/мл ДНК спермы лосося).
На Фиг.19 представлен график, на котором изображены кривые констант диссоциации клонов C5. Константы диссоциации (Kd) оценивали подстановкой данных в уравнение: фракция связанной РНК = амплитуда*Kd/(Kd + [C5]).
На Фиг.20 представлен график, на котором изображена кривая IC50, которая иллюстрирует ингибиторный эффект гемолитической активности различных концентраций аптамера против C5 отдельно ARC913 (SEQ ID NO:75) по сравнению с ARC186 (SEQ ID NO:4).
На Фиг.21 представлена иллюстрация структуры ARC187 (SEQ ID NO:5).
На Фиг.22 представлена иллюстрация структуры ARC1905 (SEQ ID NO:67).
На Фиг.23 представлена таблица, в которой приведена схема эксперимента первого исследования с изолированным перфузируемым сердцем.
На Фиг.24 представлен график для сравнения кривой давления для внутрижелудочкого давления в левом желудочке (LV) изолированного сердца, подвергнутого воздействию плазмы человека (A), с кривой давления LVP изолированного сердца, подвергнутого воздействию контрольного раствора аптамера (B).
На Фиг.25 представлен график для сравнения кривых давления для внутрижелудочкого давления в левом желудочке (LV) изолированных сердец, подвергнутых воздействию молярно эквивалентного, 10X и 50X растворов аптамер/C5 (где в нормальной неразбавленной плазме допустима концентрация C5 приблизительно 500 нМ).
На Фиг.26 представлен график для сравнения изменений частоты сердечных сокращений в ударах в минуту (bpm) в изолированных мышиных сердцах после воздействия плазмы человека и различных растворов плазма/аптамер.
На Фиг.27 представлен график для сравнения изменений массы сердца изолированных мышиных сердец до и после воздействия плазмы человека, содержащей ARC186 (SEQ ID NO:4) с 0-1X молярной концентрацией (остановившиеся сердца), или с 10-50X молярной концентрацией (сердца, защищенные аптамером против C5).
На Фиг.28 представлен график для сравнения относительной продукции C5a в плазме человека, содержащей различные концентрации аптамера, после перфузии через изолированные мышиные сердца. Относительные концентрации C5a нанесены на график в качестве единиц поглощения (Abs), где более высокие данные отражают наличие более высоких уровней C5a.
На Фиг.29 представлен график для сравнения относительной продукции растворимого C5b-9 в плазме человека, содержащей различные концентрации аптамера, после перфузии изолированных мышиных сердец.
На Фиг.30 представлен график, на котором показан эффект ARC186 (SEQ ID NO:4) на расщепление C3 в вытекающей из мышиных сердец жидкости.
На Фиг.31 представлена таблица, на которой показаны результаты иммуногистохимического окрашивания в исследовании с изолированным перфузируемым сердцем мыши.
На Фиг.32 представлена таблица, на которой показана молярная концентрация ARC658 (SEQ ID NO:62), необходимая для защиты сердца от опосредуемого C5b повреждения в сыворотке человека или примата.
На Фиг.33 представлено изображение, на котором показан логарифмически-линейный график остающегося процентного количества полноразмерного ARC186 в зависимости от времени инкубации в плазме как у крыс, так и яванских макак.
На Фиг.34 представлена таблица, в которой показана схема эксперимента для фармакологического исследования, проведенного на крысах Sprague-Dawley, как описано в примере 5.
На Фиг.35 представлена таблица, в которой показана средняя концентрация ARC657 (SEQ ED NO:61), ARC658 (SEQ ED NO:62) или ARC187 (SEQ ID NO:5) в плазме в зависимости от периода времени у крыс Sprague-Dawley.
На Фиг.36 представлен график, на котором изображена средняя концентрация в плазме ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) и ARC187 (SEQ ED NO:5) с течением времени после внутривенного введения аптамера у крыс.
На Фиг.37 представлена таблица, на которой показан некомпартментный анализ концентрации против временных данных, представленных на Фиг.35 и 36.
На Фиг.38A представлена таблица, на которой показана схема фармакокинетического исследования ARC187 (SEQ ED NO:5) и ARC1905 (SEQ ED NO:67) у мышей; на Фиг.38B представлен график, на котором изображен фармакокинетический профиль ARC187 (SEQ ID NO:5) и ARC1905 (SEQ ID NO:67) у мышей CD-1 после однократного внутривенного болюсного введения; на Фиг.38C представлена таблица, в которой показан некомпартментный анализ концентрации против временных данных, представленных на Фиг.38B.
На Фиг.39 представлена таблица, в которой показана детекция приведенных аптамеров в ткани сердца мыши после внутривенного введения.
На Фиг.40 представлена таблица, в которой показана экспериментальная схема исследования 1 на животных, описанного в примере 5E.
На Фиг.41 представлена таблица, в которой показана концентрация аптамера в плазме относительно времени после внутривенного болюсного введения аптамера яванским макакам.
На Фиг.42 представлена таблица, в которой перечислены фармакокинетические параметры ARC657 (SEQ ED NO:61), ARC658 (SEQ ID NO:62) и ARC187 (SEQ ED NO:5), вводимых внутривенно яванской макаке в исследовании 1.
На Фиг.43(a) и 43(c) представлены графики, на которых изображены концентрации sC5b-9 и C5a в плазме с течением времени после внутривенного введения аптамеров против C5 ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) или ARC187 (SEQ ID NO:5) яванским макакам; На Фиг.43(b) и 43(d) представлены графики, на которых изображены концентрации sC5b-9 и C5a в плазме против концентрации аптамеров против C5, ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) или ARC187 (SEQ ID NO:5).
На Фиг.44 представлена таблица, в которой показана экспериментальная схема исследования 2, описанного в примере 5F.
На Фиг.45 представлен график, в котором показана средняя концентрация аптамера в плазме в различные моменты времени после внутривенного введения ARC658 (SEQ ID NO:62) или ARC187 (SEQ ID NO:5) яванским макакам.
На Фиг.46 представлена таблица, в которой показан двухкомпартментный анализ концентрации относительно временных данных после внутривенного болюсного введения аптамера яванской макаке.
На Фиг.47 представлен график, на котором изображена концентрация C5b-9 относительно концентрации ARC187 (SEQ ID NO:5) или ARC658 (SEQ ID NO:62) в присутствии зимозана в плазме яванских макак.
На Фиг.48 представлен график, на котором изображена концентрация C5a относительно концентрации ARC187 (SEQ ID NO:5) или ARC658 (SEQ ID NO:62) в присутствии зимозана в плазме яванских макак.
На Фиг.49 представлена таблица, обобщающая исследование PK-PD ARC187 (SEQ ID NO:5) в процессе и после введения внутривенной болюсной инфузией яванским макакам.
На Фиг.50 представлена таблица, в которой обобщены фармакокинетические параметры для ARC187 (SEQ ID NO:5) у яванских макак после внутривенного болюсного введения.
На Фиг.51 представлен график, на котором изображены вычисленные и истинные фармакокинетические профили ARC187 (SEQ ID NO:5) в процессе и после введения яванским макакам посредством внутривенного болюса плюс инфузия.
На Фиг.52 представлен график, на котором показаны уровни активного ARC187 (SEQ ID NO:5) в плазме, остающиеся постоянными в процессе и после введения яванским макакам посредством внутривенного болюса плюс инфузия.
На Фиг.53 представлена таблица, в которой показано предсказанное необходимое дозирование аптамеров против C5 при хирургической операции CABG.
На Фиг.54 представлен график, на котором изображено, что ARC187 (SEQ ID NO:5) относительно не оказывает эффекта in vitro на коагуляцию, измеренную посредством протромбинового времени (PT) и активированного частичного тромбопластинового времени (APTT).
На Фиг.55 представлена таблица, в которой обобщены эффекты in vitro ARC187 (SEQ ID NO:5) на противосвертывающую активность гепарина и прокоагулянтную активность протамина.
На Фиг.56 представлен график, на котором показано, что ARC187 (SEQ ID NO:5) не влияет на реверсию антикоагуляции посредством гепарина in vivo.
На Фиг.57 представлен график, на котором показано, что как гепарин, так и протамин не оказывают эффекта на направленную против комплемента функцию ARC187 (SEQ ID NO:5), измеренную по ингибированию активации комплементом зимозана.
На Фиг.58 представлен график, на котором изображено процентное ингибирование гемолиза эритроцитов овцы в присутствии сыворотки человека в зависимости от концентрации аптамеров против C5 ARC1905 (SEQ ID NO:67) или ARC672 (SEQ ID NO:63).
На Фиг.59A представлен график, на котором изображено процентное ингибирование гемолиза в присутствии сыворотки человека, яванской макаки и крысы посредством ARC1905 (SEQ ID NO:67); На Фиг.59B представлена таблица, в которой обобщены средние значения IC50 для ингибирования активации комплемента в сыворотке человека, яванской макаки и крысы посредством ARC1905, аптамера против C5 или ARC127, постороннего аптамера, который не связывает C5 (отрицательный контроль).
На Фиг.60 представлен график, на котором изображено значение IC50 для ингибирования меченного радиоактивной меткой ARC186 (SEQ ID NO:4) (вертикальная ось) в зависимости от концентрации немеченого конкурента ARC1905 (SEQ ID NO:67) или ARC672 (SEQ ID NO:63) (горизонтальная ось), в конкурентном анализе связывания.
На Фиг.61 представлен график, на котором изображено значение IC50 для ингибирования меченного радиоактивной меткой ARC186 (SEQ ID NO:4) (вертикальная ось) в зависимости от концентрации немеченого конкурента ARC1905 (SEQ ID NO:67) (горизонтальная ось) при 37°C и 25°C в конкурентном анализе связывания.
На Фиг.62 представлен график, на котором изображены стандартные кривые для C5a (hC5a) человека и C5a яванской макаки (hC5a экв.).
На Фиг.63 представлена таблица, в которой обобщены значения IC50, IC90 и IC99 для ингибирования активации C5 в сыворотке человека и яванской макаки посредством ARC1905 (SEQ ID NO:67), при измерении в анализе индуцируемой зимозаном активации комплемента.
На Фиг.64 представлен график, на котором изображено процентное ингибирование образования C5a в зависимости от концентрации ARC1905 (SEQ ID NO:67) в сыворотках человека и яванской макаки, измеренное в анализе индуцируемой зимозаном активации комплемента.
На Фиг.65 представлен график, на котором изображен эффект ARC1905 (SEQ ID NO:67) на образование C3a в сыворотке человека или яванской макаки, при измерении в анализе индуцируемой зимозаном активации комплемента.
На Фиг.66 представлена таблица, в которой обобщены средние значения IC50, IC90 и IC99 для ингибирования посредством ARC1905 активации комплемента (SEQ ID NO:67) в сыворотке человека от 5 доноров, при измерении в модели активации комплемента с помощью петли трубки.
На Фиг.67 представлен график, на котором изображено процентное ингибирование образования C5a и C3a в зависимости от концентрации ARC1905, аптамера против C5, или ARC127, постороннего аптамера, который не связывает C5 (отрицательный контроль) в модели активации комплемента с помощью петли трубки.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к материалам и способам для лечения, профилактики и/или стабилизации связанного с комплементом заболевания глаз (также называемого в настоящем документе глазными нарушениями).
В некоторых вариантах осуществления этого изобретения, аптамер против комплемента модулирует функцию компонента комплемента или его варианта. В особенно предпочтительных вариантах осуществления, аптамер против комплемента ингибирует или снижает функцию компонента комплемента или его варианта, предпочтительно in vivo, предпочтительно у позвоночного, предпочтительно млекопитающего, более предпочтительно in vivo у человека. В некоторых вариантах осуществления этого изобретения, например, где компонент комплемента-мишень представляет собой C2, C3, C4, C5 и/или фактор B, функция, модулируемая, предпочтительно ингибируемая, аптамером, представляет собой расщепление белка комплемента. В некоторых вариантах осуществления этого изобретения, например, где компонент комплемента-мишень представляет собой C2b, C5b, C6, C7, C8, C9, фактор B и/или пропердин, функция, модулируемая, предпочтительно ингибируемая аптамером, представляет собой сборку активного комплекса компонентов комплемента, такого как конвертазный или мембраноатакующий комплекс. В некоторых вариантах осуществления этого изобретения, например, где C3b, фактор D, C1 (включая C1r и/или C1s) и/или ассоциированную с маннозой сериновую протеазу ("MASP") представляет собой компонент комплемента-мишень, функция, модулируемая, предпочтительно ингибируемая аптамером, представляет собой ферментативную активность. В некоторых вариантах осуществления этого изобретения, например, где C3a, C5a, рецептор для C3a или рецептор для C5a представляет собой компонент комплемента-мишень, функция, модулируемая, предпочтительно ингибируемая, аптамером, представляет собой связывание лиганд/рецептор.
В одном варианте осуществления, предусмотрен способ стабилизации, лечения и/или профилактики опосредуемого C5, C5a и/или C5b-9 глазного нарушения, при этом способ включает стадию введения средства против C5 субъекту, нуждающемуся в этом, в количестве, достаточном для стабилизации, лечения и/или профилактики глазного нарушения. В некоторых вариантах осуществления, глазное нарушение, подлежащее стабилизации, лечению и/или профилактике, представляет глазное нарушение, связанное с неоваскуляризацией. В некоторых вариантах осуществления, глазное нарушение, подлежащее стабилизации, лечению и/или профилактике, представляет собой диабетическую ретинопатию или дегенерацию желтого пятна, в частности, связанную со старением дегенерацию желтого пятна ("AMD). В некоторых вариантах осуществления, AMD, подлежащая стабилизации, лечению и/или профилактике, представляет собой AMD экссудативного типа. В некоторых вариантах осуществления, AMD, подлежащая стабилизации, лечению и/или профилактике, является неэкссудативной.
В некоторых вариантах осуществления, предусмотрен способ стабилизации, лечения и/или профилактики опосредуемого комплементом глазного нарушения, при этом способ включает стадию введения терапевтически эффективного количества аптамера против комплемента субъекту, нуждающемуся в этом. В некоторых вариантах осуществления, терапевтически эффективное количество аптамера против комплемента представляет собой количество, достаточное для стабилизации, лечения и/или профилактики глазного нарушения. В некоторых вариантах осуществления этого изобретения, субъект является позвоночным, в некоторых вариантах осуществления млекопитающим и, в некоторых вариантах осуществления, человеком. В некоторых вариантах осуществления, опосредуемое комплементом глазное нарушение, подлежащее стабилизации, лечению и/или профилактике, представляет собой острое или хроническое воспалительное и/или опосредуемое иммунной системой глазное нарушение. В некоторых вариантах осуществления, опосредуемое комплементом глазное нарушение, подлежащее стабилизации, лечению и/или профилактике, выбрано из группы, состоящей из: воспалительного конъюнктивита, включая аллергический и гигантский папиллярный конъюнктивит, отека желтого пятна, увеита, эндофтальмита, склерита, язв роговицы, сухого кератита, глаукомы, ишемического заболевания сетчатки, отторжения трансплантата роговицы, осложнений, связанных с внутриглазной хирургической операцией, такой как внутриглазная имплантация искусственной линзы, и воспаления, обусловленного хирургической операцией по поводу катаракты, болезни Бехчета, болезни Штаргардта, иммунокомплексного васкулита, болезни Фуха, болезни Фогта-Каянаги-Харада, субретинального фиброза, кератита, витреоретинального воспаления, паразитического заражения/миграции глаз, пигментного ретинита, цитомегаловирусного ретинита и хориоидального воспаления. В некоторых вариантах осуществления, глазное нарушение, подлежащее стабилизации, лечению и/или профилактике, представляет собой дегенерацию желтого пятна, в частности, связанную со старением дегенерацию желтого пятна ("AMD"). В некоторых вариантах осуществления, глазное нарушение, подлежащее стабилизации, лечению и/или профилактике, представляет собой AMD неэкссудативного ("сухого" и/или "атрофического") типа. В некоторых вариантах осуществления, глазное нарушение, подлежащее стабилизации, лечению и/или профилактике, представляет глазное нарушение, связанное с неоваскуляризацией, такое как диабетическая ретинопатия или AMD экссудативного ("влажного") типа.
В некоторых вариантах осуществления, предусмотрен способ стабилизации опосредуемого C5, C5a и/или C5b-9 глазного нарушения, связанного с неоваскуляризацией, в частности AMD экссудативного типа или диабетической ретинопатии, включающий введение средства против C5 субъекту, нуждающемуся в этом, в количестве, достаточном для стабилизации опосредуемого C5, C5a и/или C5b-9 глазного нарушения, связанного с неоваскуляризацией. В некоторых вариантах осуществления, средство против C5 вводят в количестве, достаточном для поддержания по меньшей мере того же уровня остроты зрения у субъекта, что и уровень остроты зрения субъекта при введении средства против C5. В некоторых вариантах осуществления, средство против C5 вводят в количестве, достаточном для поддержания приблизительно того же уровня плотности сосудов сетчатки субъекта, что и уровень у субъекта при введении средства против C5.
В некоторых вариантах осуществления, предусмотрен способ стабилизации опосредуемого комплементом глазного нарушения, связанного с неоваскуляризацией, в частности AMD экссудативного типа или диабетической ретинопатии, включающий введение аптамера против комплемента субъекту, нуждающемуся в этом, в количестве, достаточном для стабилизации опосредуемого комплементом глазного нарушения, связанного с неоваскуляризацией. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для поддержания по меньшей мере того же уровня остроты зрения субъекта, что и уровень остроты зрения субъекта при введении аптамера против комплемента. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для поддержания приблизительно того же уровня плотности сосудов сетчатки субъекта, что и уровень у субъекта при введении аптамера. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для стабилизации или поддержания уровня обусловленного неоваскуляризацией кровотечения, скопления жидкости, отслоения сетчатки и/или образования рубцов у субъекта относительно уровня у субъекта обусловленного неоваскуляризацией кровотечения, скопления жидкости, отслоения сетчатки и/или образования рубцов при введении аптамера против комплемента.
В некоторых вариантах осуществления, предусмотрен способ лечения опосредуемого C5, C5a и/или C5b-9 глазного нарушения, связанного с неоваскуляризацией, в частности, AMD экссудативного типа или диабетической ретинопатии, включающий введение средства против C5 субъекту, нуждающемуся в этом, в количестве, достаточном для снижения симптома опосредуемого C5, C5a и/или C5b-9 глазного нарушения, связанного с неоваскуляризацией. В некоторых вариантах осуществления средство против C5 вводят в количестве, достаточном для повышения уровня остроты зрения у субъекта относительно уровня остроты зрения у субъекта при введении средства против C5. В некоторых вариантах осуществления, средство против C5 вводят в количестве, достаточном для снижения уровня плотности сосудов сетчатки у субъекта относительно уровня плотности сосудов сетчатки у субъекта при введении средства против C5.
В некоторых вариантах осуществления, предусмотрен способ лечения опосредуемого комплементом глазного нарушения, связанного с неоваскуляризацией, в частности AMD экссудативного типа или диабетической ретинопатии, включающий введение аптамера против комплемента субъекту, нуждающемуся в этом, в количестве, достаточном для снижения симптома опосредуемого комплементом глазного нарушения, связанного с неоваскуляризацией. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для повышения уровня остроты зрения у субъекта относительно уровня остроты зрения у субъекта при введении аптамера против комплемента. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для снижения уровня плотности сосудов сетчатки у субъекта относительно уровня плотности сосудов сетчатки у субъекта при введении аптамера против комплемента. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для снижения уровня обусловленного неоваскуляризацией кровотечения, скопления жидкости, отслоения сетчатки и/или образования рубцов у субъекта относительно уровня у субъекта обусловленного неоваскуляризацией кровотечения, скопления жидкости, отслоения сетчатки и/или образования рубцов при введении аптамера против комплемента.
В некоторых вариантах осуществления, предусмотрен способ профилактики клинического опосредуемого комплементом глазного нарушения, связанного с неоваскуляризацией, в частности AMD экссудативного типа или диабетической ретинопатии, у субъекта, при этом способ включает стадию введения аптамера против комплемента субъекту в количестве, достаточном для профилактики клинического симптома опосредуемого комплементом глазного нарушения, связанного с неоваскуляризацией. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для профилактики клинического снижения остроты зрения у субъекта. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для профилактики уровня плотности сосудов сетчатки у субъекта, соответствующего клиническому глазному связанному с неоваскуляризацией заболеванию. В некоторых вариантах осуществления, субъект обладает риском развития глазного нарушения, связанного с неоваскуляризацией. В некоторых вариантах осуществления, способ дополнительно включает выявление субъекта с риском развития опосредуемого комплементом глазного нарушения, связанного с неоваскуляризацией, перед введением аптамера против комплемента. В некоторых вариантах осуществления, стадия выявления включает детекцию наличия нарушений пигментации друз и/или сетчатки у субъекта и детекцию неклинического снижения остроты зрения. В некоторых вариантах осуществления, стадия выявления включает детекцию изменения фактора H комплемента у субъекта относительно фактора H дикого типа. Аминокислотная последовательность фактора H дикого типа описана в Ripoche et al. (1988) The complete amino acid sequence of human complement factor H. Biochem. J. 249, 593-602. В некоторых вариантах осуществления, где предусмотрен способ стабилизации, лечения и/или профилактики опосредуемого комплементом связанного с неоваскуляризацией глазного нарушения у субъекта, в частности диабетической ретинопатии, способ введения аптамера против комплемента представляет собой глазное или окологлазное введение.
В некоторых вариантах осуществления, предусмотрен способ профилактики клинического опосредуемого C5, C5a и/или C5b-9 глазного нарушения, связанного с неоваскуляризацией, в частности AMD экссудативного типа или диабетической ретинопатии, у субъекта, включающий введение средства против C5 субъекту, при этом способ включает стадию введения субъекту средства против C5 в количестве, достаточном для профилактики клинического симптома опосредуемого C5, C5a и/или C5b-9 глазного нарушения, связанного с неоваскуляризацией. В некоторых вариантах осуществления, средство против C5 вводят в количестве, достаточном для профилактики клинического снижения остроты зрения у субъекта. В некоторых вариантах осуществления, средство против C5 вводят в количестве, достаточном для профилактики уровня плотности сосудов сетчатки у субъекта, соответствующего клиническому глазному связанному с неоваскуляризацией заболеванию. В некоторых вариантах осуществления, субъект обладает риском развития глазного нарушения, связанного с неоваскуляризацией. В некоторых вариантах осуществления, способ дополнительно включает выявление субъекта с риском развития опосредуемого C5, C5a и/или C5b-9 глазного нарушения, связанного с неоваскуляризацией, перед введением средства против C5. В некоторых вариантах осуществления, стадия выявления включает детекцию наличия друз у субъекта и детекцию неклинического снижения остроты зрения. В некоторых вариантах осуществления, стадия выявления включает детекцию изменения фактора H комплемента у субъекта.
В некоторых вариантах осуществления описанных выше способов, способ дополнительно включает стадию введения субъекту средства против VEGF, в частности средства против VEGF, выбранного из группы, состоящей из: молекулы нуклеиновой кислоты, аптамера, антисмысловой молекулы, молекулы RNAi, белка, пептида, циклического пептида, антитела или фрагмента антитела, сахара, полимера и низкомолекулярного соединения.
В некоторых вариантах осуществления описанных выше способов, способ дополнительно включает стадию введения субъекту средства против PDGF, в частности средство против PDGF выбрано из группы, состоящей из: молекулы нуклеиновой кислоты, аптамера, антисмысловой молекулы, молекулы RNAi, белка, пептида, циклического пептида, антитела или фрагмента антитела, сахара, полимера и низкомолекулярного соединения.
В некоторых вариантах осуществления описанных выше способов, способ дополнительно включает введение субъекту средства против образования сосудов. В некоторых вариантах осуществления, средство против образования сосудов представляет собой производное порфирина. В некоторых вариантах осуществления производное порфирина представляет собой вертепорфин для инъекции (Visudyne®, Novartis Pharmaceuticals Corporation, East Hanover, NJ). В некоторых вариантах осуществления, способ дополнительно включает стадию активации производного порфирина лазерным излучением.
В одном варианте осуществления, предусмотрен способ стабилизации, лечения и/или профилактики опосредуемой C5, C5a и/или C5b-9 AMD неэкссудативного типа, включающий введение средства против C5 субъекту, нуждающемуся в этом, в количестве, достаточном для стабилизации, лечения и/или профилактики AMD неэкссудативного типа. В одном варианте осуществления, где AMD неэкссудативного типа подлежит стабилизации, средство против C5 вводят в количестве, достаточном для поддержания приблизительно того же уровня друз по сравнению с уровнем друз у субъекта при введении средства против C5. В одном варианте осуществления, где AMD неэкссудативного типа подлежит стабилизации, средство против C5 вводят в количестве, достаточном для поддержания приблизительно того же уровня остроты зрения у субъекта по сравнению с остротой зрения у субъекта при введении средства против C5. В одном варианте осуществления, где AMD неэкссудативного типа подлежит лечению, средство против C5 вводят в количестве, достаточном для снижения уровня друз по сравнению с уровнем друз у субъекта при введении средства против C5. В одном варианте осуществления, где AMD неэкссудативного типа подлежит лечению, средство против C5 вводят в количестве, достаточном для повышения остроты зрения у субъекта по сравнению с остротой зрения у субъекта при введении средства против C5. В одном варианте осуществления, где AMD неэкссудативного типа подлежит профилактике, способ включает введение средства против C5 субъекту, нуждающемуся в этом, в количестве, достаточном для профилактики клинического симптома опосредуемой C5, C5a и/или C5b-9 неэкссудативной AMD. В некоторых вариантах осуществления, средство против C5 вводят в количестве, достаточном для профилактики клинического снижения остроты зрения у субъекта. В некоторых вариантах осуществления, средство против C5 вводят в количестве, достаточном для профилактики повышения клинического уровня друз. В некоторых вариантах осуществления, субъект обладает риском развития неэкссудативной AMD. В некоторых вариантах осуществления, способ дополнительно включает выявление субъекта с риском развития опосредуемой C5, C5a и/или C5b-9 неэкссудативной AMD перед введением средства против C5. В некоторых вариантах осуществления, стадия выявления включает детекцию наличия друз у субъекта и детекцию неклинического снижения остроты зрения. В некоторых вариантах осуществления, стадия выявления включает детекцию у субъекта изменения фактора H комплемента.
В некоторых вариантах осуществления описанных выше способов, средство против C5 выбрано из группы, состоящей из: молекулы нуклеиновой кислоты, аптамера, антисмысловой молекулы, молекулы RNAi, белка, пептида, циклического пептида, антитела или фрагмента антитела, сахара, полимера и низкомолекулярного соединения. В конкретном варианте осуществления, средство против C5 представляет собой специфичный к C5 аптамер, более конкретно, специфичный к C5 аптамер, выбранный из группы, состоящей из SEQ ID NO:1-67, 75-81 и 88-98. В предпочтительном варианте осуществления, специфичный к C5 аптамер для применения в описанных выше способах выбран из группы, состоящей из ARC187 (SEQ ID NO:5) и ARC1905 (SEQ ID NO:67).
В некоторых вариантах осуществления описанных выше способов, средство против C5 доставляют внутриглазным введением, в частности, введением в стекловидное тело. В некоторых вариантах осуществления описанных выше способов средство против VEGF, средство против PDGF и/или средство против образования сосудов доставляют внутриглазным введением. В некоторых вариантах осуществления описанных выше способов, средство против C5, средство против VEGF, средство против PDGF и/или средство против образования сосудов, подлежащие введению, представляют собой пролекарства. В некоторых вариантах осуществления описанных выше способов, субъектом является человек.
Термин "при введении средства против C5", используемый в описанных выше способах, охватывает время клинического определения представляющего интерес симптома перед введением средства против C5.
В одном варианте осуществления, предусмотрен способ стабилизации, лечения и/или профилактики опосредуемой комплементом AMD неэкссудативного типа, включающий введение терапевтически эффективного количества аптамера против комплемента субъекту, нуждающемуся в этом. В одном варианте осуществления, где AMD неэкссудативного подлежит стабилизации, аптамер против комплемента вводят в количестве, достаточном для поддержания приблизительно того же уровня друз (например, размера, количества, площади и/или морфологии) по сравнению с уровнем друз у субъекта при введении аптамера против комплемента. В одном варианте осуществления, где AMD неэкссудативного типа подлежит стабилизации, аптамер против комплемента вводят в количестве, достаточном для стабилизации прогрессирования географической атрофии, включая атрофию пигментного эпителия сетчатки, фоторецепторов и/или хориоидальных капилляров, для поддержания приблизительно того же уровня географической атрофии по сравнению с уровнем у субъекта при введении аптамера против комплемента. В одном варианте осуществления, где AMD неэкссудативного типа подлежит стабилизации, аптамер против комплемента вводят в количестве, достаточном для поддержания приблизительно того же уровня остроты зрения у субъекта по сравнению с остротой зрения у субъекта при введении аптамера против комплемента.
В одном варианте осуществления, где AMD неэкссудативного типа подлежит лечению, аптамер против комплемента вводят в количестве, достаточном для снижения уровня друз, в частности больших мягких друз, по сравнению с уровнем друз у субъекта при введении аптамера против комплемента. В одном варианте осуществления, где AMD неэкссудативного типа подлежит лечению, аптамер против комплемента вводят в количестве, достаточном для повышения остроты зрения у субъекта по сравнению с остротой зрения у субъекта при введении аптамера против комплемента.
В одном варианте осуществления, где AMD неэкссудативного типа подлежит профилактике, способ включает введение аптамера против комплемента субъекту, нуждающемуся в этом, в количестве, достаточном для профилактики клинического симптома опосредуемой комплементом неэкссудативной AMD. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для профилактики клинического снижения остроты зрения у субъекта. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для профилактики повышения клинического уровня друз, в частности больших мягких друз. В некоторых вариантах осуществления, аптамер против комплемента вводят в количестве, достаточном для профилактики клинического уровня географической атрофии. В некоторых вариантах осуществления, аптамер против комплемента вводят субъекту, имеющему AMD неэкссудативного типа в количестве, достаточном для профилактики прогрессирования экссудативной AMD у субъекта. В некоторых вариантах осуществления, аптамер против комплемента вводят субъекту, имеющему связанную с возрастом макулопатию (характеризующуюся наличием друз, изменением пигментации сетчатки и/или небольшими областями атрофии) в количестве, достаточном для профилактики прогрессирования экссудативной AMD или клинического уровня географической атрофии у субъекта.
В некоторых вариантах осуществления, субъект обладает риском развития неэкссудативной AMD. В некоторых вариантах осуществления, способ дополнительно включает выявление субъекта с риском развития опосредуемой комплементом неэкссудативной AMD перед введением аптамера против комплемента. В некоторых вариантах осуществления, стадия выявления включает детекцию наличия друз, в частности больших мягких друз, изменений пигментации сетчатки и/или областей атрофии у субъекта и детекцию неклинической потери остроты зрения. В некоторых вариантах осуществления, стадия выявления включает детекцию изменения фактора H комплемента у субъекта по сравнению с диким типом.
В некоторых вариантах осуществления описанных выше способов, аптамер против комплемента ингибирует компонент комлемента-мишень, выбранный из группы, состоящей из: компонента классического каскада комплемента, компонента альтернативного каскада комплемента и компонента лектинового каскада. В некоторых вариантах осуществления, аптамер против комплемента ингибирует компонент комлемента-мишень в мембраноатакующем каскаде. В некоторых вариантах осуществления, аптамер против комплемента ингибирует компонент комлемента-мишень, выбранный из группы, состоящей из: C1, C1q, C1r, C1s, C2, C3, C3a, рецептора для C3a, C4, C5, C5a, рецептора для C5a, C5b, C6, C7, C8, C9, фактора B, фактора D, пропердина, маннан-связывающего лектина (в дальнейшем в настоящем документе "MBL"), ассоциированной с MBL сериновой протеазы 1 ("MASP 1") и ассоциированной с MBL сериновой протеазы 2 ("MASP 2"). В некоторых вариантах осуществления, аптамер против комплемента не является аптамером с аффинностью и высокой специфичностью к компоненту комплемента-мишени, выбранному из группы, состоящей из: C3a, рецептора для C3a, C5a и рецептора для C5a. В некоторых вариантах осуществления, аптамер против комплемента не является аптамером с аффинностью и высокой специфичностью к компоненту комплемента-мишени, выбранному из группы, состоящей из: фактора B и фактора D.
В некоторых вариантах осуществления описанных выше способов, аптамер против комплемента доставляют субъекту посредством внутриглазного введения, в частности, посредством введения в стекловидное тело или окологлазного введения. В некоторых вариантах осуществления, аптамер против комплемента, подлежащий введению субъекту, содержится в составе с замедленным высвобождением.
Как используют в настоящем документе, термин "при введении аптамера против комплемента" охватывает время определения или оценки представляющего интерес симптома, где определение и оценку проводят во время, варьирующееся от времени до введения аптамера против комплемента вплоть до и включая определение через короткий промежуток времени после введения аптамера против комплемента, например, вплоть до 12 часов после, 24 часов после или 48 часов после введения.
В некоторых вариантах осуществления, предусмотрена глазная фармацевтическая композиция, содержащая терапевтически эффективное количество аптамера против комплемента, например, количество, достаточное для стабилизации, лечения и/или профилактики опосредуемого комплементом глазного нарушения. Фармацевтическая композиция по этому изобретению может содержать фармацевтически приемлемый носитель или разбавитель. В этом аспекте, настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество аптамера, который ингибирует глазную функцию компонента комплемента-мишени in vivo, в частности, у субъекта-человека, или его соли, и фармацевтически приемлемый носитель или разбавитель. В некоторых вариантах осуществления, глазная фармацевтическая композиция содержит состав с замедленным высвобождением.
В одном варианте осуществления, аптамер против комплемента для применения в указанных выше способах представляет собой аптамер, который ингибирует C5 in vivo, предпочтительно C5 человека. В конкретном варианте осуществления, предусмотрен аптамер против C5 в соответствии с ARC186 (SEQ ID NO 4) или аптамер, содержащий нуклеотидную последовательность в соответствии с ARC186 (SEQ ID NO:4), конъюгированную с группой PEG для применения в указанных выше способах. В конкретных вариантах осуществления, этот конъюгат аптамер ARC186/PEG обладает по существу такой же аффинностью связывания в отношении белка комплемента C5, что и аптамер, состоящий из последовательности в соответствии с SEQ ID NO:4, но лишенный группы PEG. Как используют в настоящем документе, по существу такая же аффинность связывания означает отличие не более чем приблизительно в от 2 до десяти раз, предпочтительно отличие не более чем в от 2 до пяти раз в константах диссоциации, измеряемых дот-блот анализом. В некоторых вариантах осуществления константы диссоциации определяют конкурентным дот-блот анализом, описанным в примере 1A, ниже. В некоторых вариантах осуществления, группа полиэтиленгликоля обладает молекулярной массой более 10 кДа, в частности, молекулярной массой 20 кДа, более конкретно 30 кДа и более конкретно 40 кДа. В некоторых вариантах осуществления, группа PEG конъюгирована с 5'-концом ARC186 (SEQ ID NO:4). В некоторых вариантах осуществления конъюгат аптамер/PEG обладает временем полужизни, предпочтительно терминальным временем полужизни в двухкомпартментной модели, как определяют способом, описанным в примере 5E, ниже, составляющим по меньшей мере 15 часов, предпочтительно по меньшей мере 24 часа, более предпочтительно по меньшей мере 48 часов, у примата. В некоторых вариантах осуществления конъюгат аптамер/PEG обладает временем полужизни, предпочтительно терминальным временем полужизни в двухкомпартментной модели, составляющим по меньшей мере 10, предпочтительно по меньшей мере 15 часов, у крысы. В некоторых вариантах осуществления, PEG, конъюгированный с 5'-концом ARC186 (SEQ ID NO:4), представляет собой PEG массой 40 кДа. В конкретных вариантах осуществления PEG массой 40 кДа представляет собой разветвленный PEG. В некоторых вариантах осуществления разветвленный PEG массой 40 кДа представляет собой 1,3-бис(mPEG-[20 кДа])-пропил-2-(4'-бутамид). В других вариантах осуществления разветвленный PEG массой 40 кДа представляет собой 2,3-бис(mPEG-[20 кДа])-пропил-1-карбамоил.
В вариантах осуществления, где разветвленный PEG массой 40 кДа представляет собой 1,3-бис(mPEG-[20 кДа])-пропил-2-(4'-бутамид), предусмотрен аптамер, обладающей структурой, указанной ниже:
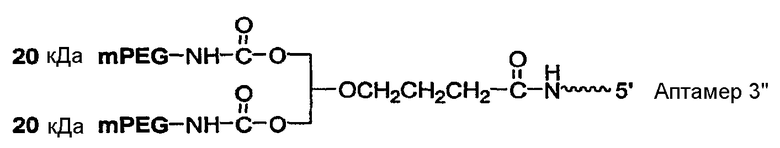
где
 указывает на линкер,
указывает на линкер,
Аптамер=
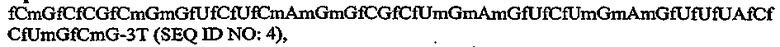
где fC и fU = 2'-фторнуклеотиды, и mG и mA = 2'-OMe-нуклеотиды, и все другие нуклеотиды являются 2'-OH, и 3T указывает на инвертированный дезокситимидин.
В вариантах осуществления, где разветвленный PEG массой 40 кДа представляет собой 2,3-бис(mPEG-[20 кДа])-пропил-1-карбамоил, предусмотрен аптамер, имеющий структуру, указанную ниже:
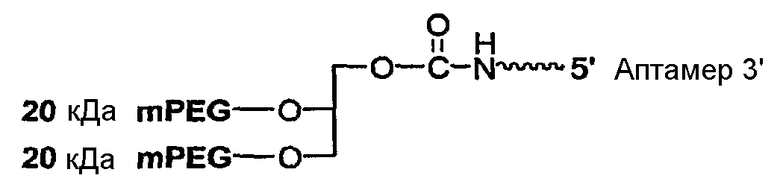
где
указывает на линкер,
Аптамер=
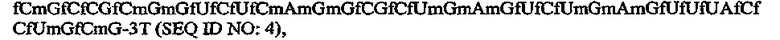
где fC и fU = 2'-фторнуклеотиды, и mG и mA = 2'-OMe-нуклеотиды, и все другие нуклеотиды являются 2'-OH, и 3T указывает на инвертированный дезокситимидин.
В некоторых вариантах осуществления данного аспекта этого изобретения линкер представляет собой алкильный линкер. В конкретных вариантах осуществления, алкильный линкер содержит от 2 до 18 последовательно расположенных групп CH2. В предпочтительных вариантах осуществления, алкильный линкер содержит от 2 до 12 последовательно расположенных групп CH2. В особенно предпочтительных вариантах осуществления алкильный линкер содержит от 3 до 6 последовательно расположенных групп CH2.
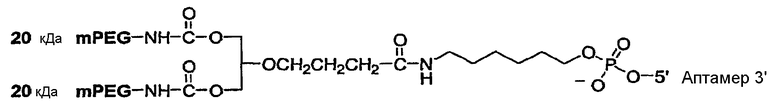
В конкретном варианте осуществления, предусмотрен аптамер, ARC187 (SEQ ID NO:5), имеющий структуру, указанную ниже:
где
Аптамер=
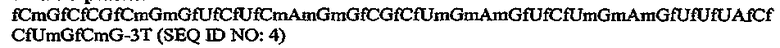
где fC и fU = 2'-фторнуклеотиды, и mG и mA = 2'-OMe-нуклеотиды, и все другие нуклеотиды являются 2'-OH, и где 3T указывает на инвертированный дезокситимидин.
В другом варианте осуществления, предусмотрен аптамер, ARC1905 (SEQ ID NO:67), имеющий структуру, указанную ниже:
где
Аптамер=
где fC и fU = 2'-фторнуклеотиды, и mG и mA = 2'-OMe нуклеотиды, и все другие нуклеотиды являются 2'-OH, и где 3T указывает на инвертированный дезокситимидин.
В одном варианте осуществления, предусмотрена глазная фармацевтическая композиция, содержащая количество ARC186 (SEQ ID NO 4), ARC187 (SEQ ED NO:5), или ARC1905 (SEQ ID NO:67), или их соли, эффективное для лечения, стабилизации и/или профилактики опосредуемого комплементом глазного нарушения у субъекта. Фармацевтическая композиция по этому изобретению может содержать фармацевтически приемлемый носитель или разбавитель. В этом аспекте, настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество аптамера, который ингибирует расщепление белка комплемента C5 in vivo, или его соли и фармацевтически приемлемый носитель или разбавитель. В данном аспекте этого изобретения предусмотрена фармацевтическая композиция ARC186 (SEQ ID NO 4), ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67) для применения в целях лечения, стабилизации и/или профилактики заболевания глаз in vivo. Также в данном аспекте этого изобретения, предусмотрены ARC186 (SEQ ID NO 4), ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67) для применения в целях изготовления фармацевтической композиции для лечения, стабилизации и/или профилактики опосредуемого комплементом заболевания глаз у субъекта.
В другом варианте осуществления, глазная фармацевтическая композиция по этому изобретению содержит терапевтически эффективное количество аптамера против C5, содержащего нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO: с 1 по 69, 75, 76, 81, 91, 95 и 96 для применения при изготовлении фармацевтической композиции в целях применения в способах лечения опосредуемых комплементом глазных заболеваний по этому изобретению. В этом аспекте, настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество аптамера, который ингибирует расщепление белка комплемента C5 in vivo, или его соли и фармацевтически приемлемый носитель или разбавитель.
В другом аспекте настоящее изобретение относится к фармацевтическим композициям. В одном варианте осуществления, предусмотрена фармацевтическая композиция, содержащая терапевтически эффективное количество ARC187 (SEQ ID NO:5), или ARC1905 (SEQ ID NO:67), или их соли. Фармацевтическая композиция по этому изобретению может содержать фармацевтически приемлемый носитель или разбавитель. В этом аспекте, настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество аптамера, который ингибирует расщепление белка комплемента C5 in vivo, или его соли и фармацевтически приемлемый носитель или разбавитель. В данном аспекте этого изобретения, предусмотрена фармацевтическая композиция ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67) для применения в целях лечения, профилактики или смягчения течения заболевания in vivo. Также, в данном аспекте этого изобретения предусмотрены ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67) для применения при изготовлении фармацевтической композиции.
В другом аспекте этого изобретения, предусмотрены способы лечения. В одном варианте осуществления, способ по этому изобретению включает лечение, профилактику или смягчение течения заболевания, опосредуемого белком комплемента C5, и/или его производными C5a и C5b-9, при этом способ включает введение фармацевтической композиции, содержащей ARC187 (SEQ ID NO:5), или ARC1905 (SEQ ID NO:67), или их соль, позвоночному. В некоторых вариантах осуществления, способ включает введение фармацевтической композиции по этому изобретению млекопитающему. В некоторых вариантах осуществления, млекопитающим является человек.
В некоторых вариантах осуществления, опосредуемое белком комплемента C5, C5a и/или C5b-9 заболевание, подлежащее лечению, представляет собой острые ишемические заболевания (инфаркт миокарда, инсульт, повреждение при ишемии/реперфузии); острые воспалительные заболевания (инфекционное заболевание, септицемия, шок, острое/сверхострое отторжение трансплантата); хронические воспалительные и/или опосредуемые иммунной системой заболевания, включая диабетическую ретинопатию, дегенерацию желтого пятна, включая экссудативную и неэкссудативную формы AMD, и также включая аллергию, астму, ревматоидный артрит и другие ревматологические заболевания, рассеянный склероз и другие неврологические заболевания, псориаз и другие дерматологические заболевания, миастению, системную красную волчанку (SLE)); и подострое/хроническое воспалительное и/или опосредуемое иммунной системой заболевание (включая отторжение трансплантата, гломерулонефрит и другие заболевания почек) и глазные заболевания. В некоторых вариантах осуществления, опосредуемые белком комплемента C5, C5a и/или C5b-9 заболевания, подлежащие лечению, включают активацию комплемента, обусловленную диализом или ситуациями, когда кровь проходит над и/или через синтетические трубки и/или другой материал. В некоторых вариантах осуществления, опосредуемое белком комплемента C5, C5a и/или C5b-9 заболевание, подлежащее лечению, выбрано из группы, состоящей из повреждения миокарда, связанного с хирургической операцией CABG, повреждения миокарда, связанного с баллонной ангиопластикой, и повреждения миокарда, связанного с рестенозом. В некоторых вариантах осуществления, опосредуемое белком комплемента C5, C5a и/или C5b-9 нарушение, подлежащее лечению, выбрано из группы, состоящей из: повреждения миокарда, связанного с хирургической операцией CABG, повреждения миокарда, связанного с баллонной ангиопластикой, повреждения миокарда, связанного с рестенозом, опосредуемых белком комплемента осложнений, связанных с хирургической операцией CABG, опосредуемых белком комплемента осложнений, связанных с чрескожным вмешательством на коронарных сосудах, пароксизмальной ночной гемоглобинурии, острого отторжения трансплантата, сверхострого отторжения трансплантата, подострого отторжения трансплантата и хронического отторжения трансплантата. В некоторых вариантах осуществления опосредуемое белком комплемента C5, C5a и/или C5b-9 заболевание, подлежащее лечению, представляет собой осложнения, связанные с хирургической операцией CABG. В конкретном варианте осуществления, заболевание, подлежащее лечению, представляет собой повреждение миокарда, связанное с хирургической операцией CABG. В конкретном варианте осуществления способа лечения по этому изобретению, заболевание, симптом которого подлежит уменьшению, стабилизации и/или профилактике, представляет собой глазное нарушение, в частности, диабетическую ретинопатию, экссудативную и/или неэкссудативную AMD.
В некоторых вариантах осуществления, способ по этому изобретению включает введение фармацевтической композиции, содержащей ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67), для достижения концентрации аптамера в плазме, которая изменена приблизительно в от 0,5 до 10 раз по сравнению с концентрацией эндогенного белка комплемента C5. В некоторых вариантах осуществления, фармацевтические композиции аптамеров ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67) вводят для достижения концентрации аптамера в плазме, которая изменена в от приблизительно 0,75 до приблизительно 5 раз, в от 0,75 до приблизительно 3 раз, и в от 1,5 до приблизительно 2 раз по сравнению с концентрацией эндогенного белка комплемента C5, в то время как в других вариантах осуществления композицию аптамера вводят для достижения концентрации, эквивалентной концентрации эндогенного белка комплемента. В некоторых вариантах осуществления, фармацевтическую композицию по этому изобретению, содержащую ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67), вводят для достижения концентрации аптамера в плазме приблизительно 5 мкМ, приблизительно 4 мкМ, приблизительно 3 мкМ, приблизительно 2 мкМ, приблизительно 1,5 мкМ, приблизительно 1 мкМ или приблизительно 500 нМ.
Можно применять любое сочетание способов, длительности и скорости введения, которое является достаточным для достижения концентраций аптамера в плазме по этому изобретению. В некоторых вариантах осуществления фармацевтическую композицию вводят внутривенно. В некоторых вариантах осуществления, фармацевтическую композицию вводят в виде болюса и/или посредством непрерывной инфузии.
В конкретных вариантах осуществления лечения, профилактики и/или смягчения осложнений, связанных с хирургической операцией CABG, в частности, повреждения миокарда, связанного с хирургической операцией CABG, способ по этому изобретению включает введение фармацевтической композиции перед хирургической операцией и продолжение введения по меньшей мере в течение 24 часов, в некоторых вариантах осуществления в течение приблизительно 48 часов или в некоторых вариантах осуществления, в течение приблизительно 72 часов. В конкретном варианте осуществления данного аспекта этого изобретения, концентрации аптамера в плазме, приблизительно в два раза превышающей концентрацию эндогенного белка комплемента, достигают посредством введения внутривенного болюса приблизительно из от 0,75 до 1,25, предпочтительно приблизительно из 1 мг аптамера на кг массы пациента, подлежащего лечению, до, одновременно или после внутривенной инфузии более низкой дозы аптамера, где количество мг не включает массу конъюгированного PEG. В некоторых вариантах осуществления инфузию более низкой дозы будут проводить со скоростью, выбранной из диапазона от 0,001 до 0,005 мг/кг/мин, где количество мг не включает массу конъюгированного PEG. В конкретном варианте осуществления, инфузию более низкой дозы будут проводить со скоростью приблизительно 0,0013 мг/кг/мин. В других вариантах осуществления данного аспекта этого изобретения, где аптамер/конъюгат обладает достаточно длительным временем полужизни, фармацевтическую композицию аптамера можно вводить один раз или два раза в сутки в качестве внутривенной болюсной дозы.
В другом аспекте этого изобретения, предусмотрены способы диагностики. В одном варианте осуществления, способ диагностики включает контактирование ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67) с композицией, предположительно содержащей белок комплемента C5 или его вариант, и детекцию наличия или отсутствия белка комплемента C5 или его варианта. В некоторых вариантах осуществления белок комплемента или вариант представляют собой белок или вариант позвоночного, в частности, млекопитающего, и более конкретно человека. Настоящее изобретение относится к композиции ARC187 (SEQ ID NO:5) или ARC1905 (SEQ ID NO:67) для применения в качестве диагностического средства in vitro или in vivo.
В другом аспекте этого изобретения, предусмотрен аптамер, содержащий нуклеотидную последовательность, выбранную из группы, состоящей из: ARC330 (SEQ ID NO:2), ARC188-189, ARC250, ARC296-297, ARC331-334, ARC411-440, ARC457-459, ARC473, ARC522-525, ARC532, ARC543-544, ARC550-554, ARC657-658, ARC672, ARC706, ARC1537 и ARC1730 (с SEQ ID NO:6 по SEQ ID NO:66). В другом варианте осуществления предусмотрен любой из ARC330 (SEQ ID NO:2) и ARC188-189, ARC250, ARC296-297, ARC331-334, ARC411-440, ARC457-459, ARC473, ARC522-525, ARC532, ARC543-544, ARC550-554, ARC657-658, ARC672, ARC706, ARC1537 и ARC1730 (с SEQ ID NO:6 по SEQ ID NO:66) для применения при получении фармацевтической композиции. В этом аспекте, настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество аптамера, который ингибирует расщепление белка комплемента C5 in vivo, или его соли и фармацевтически приемлемый носитель или разбавитель.
В конкретном варианте осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность в соответствии с SEQ ID NO:1. В конкретном варианте осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62 и с SEQ ID NO:64 по SEQ ID NO:66. В некоторых вариантах осуществления, где аптамер содержит нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62 и с SEQ ID NO:64 по SEQ ID NO:66, аптамер обладает по существу такой же аффинностью связывания в отношении белка комплемента C5, что и аптамер, состоящий из последовательности в соответствии с SEQ ID NO:4, но лишенный группы PEG.
В некоторых вариантах осуществления, где аптамер содержит нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62 и с SEQ ID NO:64 по SEQ ID NO:66, аптамер обладает временем полужизни, предпочтительно терминальным временем полужизни в двухкомпартментной модели, как определено в примере 5E, ниже, составляющим по меньшей мере 15, предпочтительно по меньшей мере 30 часов, у примата. В некоторых вариантах осуществления, где аптамер содержит нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62 и с SEQ ID NO:64 по SEQ ID NO:66, аптамер обладает временем полужизни, предпочтительно терминальным временем полужизни, в двухкомпартментной модели, составляющим по меньшей мере полтора часа, предпочтительно по меньшей мере семь часов, у крысы.
В некоторых вариантах осуществления данного аспекта этого изобретения, где аптамер содержит нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62 и с SEQ ID NO:64 по SEQ ID NO:66, аптамер синтезирован с 5'-линкером следующим образом: H2N 5'Аптамер3', где указывает на линкер. В некоторых вариантах осуществления линкер представляет собой следующий алкильный линкер: H2N-(CH2)n-5'Аптамер3', где n= от 2 до 18, предпочтительно n=2-12, более предпочтительно n= от 3 до 6, более предпочтительно n=6, и где Аптамер=
5'Аптамер3', где указывает на линкер. В некоторых вариантах осуществления линкер представляет собой следующий алкильный линкер: H2N-(CH2)n-5'Аптамер3', где n= от 2 до 18, предпочтительно n=2-12, более предпочтительно n= от 3 до 6, более предпочтительно n=6, и где Аптамер=
где fC и fU = 2'-фторнуклеотиды, и mG и mA = 2'-OMe-нуклеотиды и все другие нуклеотиды являются 2'-OH и где 3T указывает на инвертированный дезокситимидин. Полученный модифицированный амином аптамер можно конъюгировать с группой PEG, выбранной из группы, состоящей из PEG массой 10 кДа, PEG массой 20 кДа, PEG массой 30 кДа и линейного PEG массой 40 кДа. В некоторых вариантах осуществления, предусмотрена фармацевтическая композиция, содержащая терапевтически эффективное количество аптамера, содержащего нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO:1, SEQ ID NO:2 и с SEQ ID NO:6 по SEQ NO:66, в частности, из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62, и с SEQ ID NO:64 по SEQ ID NO:66, или его соли. Фармацевтическая композиция по этому изобретению может содержать фармацевтически приемлемый носитель или разбавитель. В данном аспекте этого изобретения предусмотрена фармацевтическая композиция для применения в целях лечения, профилактики или смягчения заболевания in vivo, содержащая аптамер, который содержит нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO:2 и с SEQ ID NO:6 по SEQ ID NO:66, в частности, из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62 и с SEQ ID NO:64 по SEQ ID NO:66.
В другом варианте осуществления, предусмотрен способ лечения, профилактики или смягчения заболевания, опосредуемого белком комплемента C5, включающий введение фармацевтической композиции, содержащей аптамер или его соль, где аптамер содержит нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO:2 и с SEQ ID NO:6 по SEQ ID NO:66, в частности, из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62 и с SEQ ID NO:64 по SEQ ID NO:66, позвоночному. В некоторых вариантах осуществления данного аспекта этого изобретения, способ включает введение фармацевтической композиции по этому изобретению млекопитающему, предпочтительно человеку.
В некоторых вариантах осуществления, опосредуемое белком комплемента C5, C5a и/или C5b-9 заболевание, подлежащее лечению, представляет собой острые ишемические заболевания (инфаркт миокарда, инсульт, повреждение при ишемии/реперфузии); острые воспалительные заболевания (инфекционное заболевание, септицемия, шок, острое/сверхострое отторжение трансплантата); хронические воспалительные и/или опосредуемые иммунной системой заболевания, включая диабетическую ретинопатию, дегенерацию желтого пятна, включая экссудативную и неэкссудативную формы AMD, и также включая аллергию, астму, ревматоидный артрит и другие ревматологические заболевания, рассеянный склероз и другие неврологические заболевания, псориаз и другие дерматологические заболевания, миастению, системную красную волчанку (SLE); и подострое/хроническое воспалительное и/или опосредуемое иммунной системой заболевание (включая отторжение трансплантата, гломерулонефрит и другие заболевания почек) и глазные заболевания. В некоторых вариантах осуществления, опосредуемые белком комплемента C5, C5a и/или C5b-9 заболевания, подлежащие лечению, включают активацию комплемента, обусловленную диализом или ситуациями, когда кровь проходит над и/или через синтетическую трубку и/или чужеродный материал. В некоторых вариантах осуществления, опосредуемое белком комплемента C5, C5a и/или C5b-9 заболевание, подлежащее лечению, выбрано из группы, состоящей из повреждения миокарда, связанного с хирургической операцией CABG, повреждения миокарда, связанного с баллонной ангиопластикой, и повреждения миокарда, связанного с рестенозом. В некоторых вариантах осуществления, опосредуемое белком комплемента C5, C5a и/или C5b-9 нарушение, подлежащее лечению, выбрано из группы, состоящей из: повреждения миокарда, связанного с хирургической операцией CABG, повреждения миокарда, связанного с баллонной ангиопластикой, повреждения миокарда, связанного с рестенозом, опосредуемых белком комплемента осложнений, связанных с хирургической операцией CABG, опосредуемых белком комплемента осложнений, связанных с чрескожным вмешательством на коронарных сосудах, пароксизмальной ночной гемоглобинурии, острого отторжения трансплантата, сверхострого отторжения трансплантата, подострого отторжения трансплантата и хронического отторжения трансплантата. В некоторых вариантах осуществления опосредуемое белком комплемента C5, C5a и/или C5b-9 заболевание, подлежащее лечению, представляет собой осложнения, связанные с хирургической операцией CABG. В конкретном варианте осуществления, заболевание, подлежащее лечению, представляет собой повреждение миокарда, связанное с хирургической операцией CABG. В конкретном варианте осуществления способа лечения по этому изобретению, заболевание, симптом которого подлежит уменьшению, стабилизации и/или профилактике, представляет собой глазное нарушение, в частности диабетическую ретинопатию, экссудативную и/или неэкссудативную AMD.
В некоторых вариантах осуществления, способ по этому изобретению включает введение фармацевтической композиции, содержащей аптамер, обладающий нуклеотидной последовательностью, выбранной из группы, состоящей из: SEQ ID NO:2 и с SEQ ID NO:6 по SEQ ID NO:66, в частности, из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62 и с SEQ ID NO:64 по SEQ ID NO:66, пациенту для достижения концентрации аптамера в плазме, которая изменена приблизительно в от 0,5 до 10 раз по сравнению с концентрацией эндогенного белка комплемента C5. В некоторых вариантах осуществления, фармацевтические композиции аптамера вводят для достижения концентрации аптамера в плазме, которая изменена в от приблизительно 0,75 до приблизительно 5 раз, в от 0,75 до приблизительно 3 раз и в от 1,5 до приблизительно 2 раз по сравнению с концентрацией эндогенного белка комплемента C5, в то время как в других вариантах осуществления композицию аптамера вводят для достижения концентрации, эквивалентной концентрации эндогенного белка комплемента. В некоторых вариантах осуществления, фармацевтическую композицию по этому изобретению вводят для достижения концентрации аптамера в плазме приблизительно 5 мкМ, приблизительно 4 мкМ, приблизительно 3 мкМ, приблизительно 2 мкМ, приблизительно 1,5 мкМ, приблизительно 1 мкМ или приблизительно 500 нМ.
Можно применять любое сочетание способа, длительности и частоты введения, которые достаточны для достижения концентраций аптамера в плазме по этому изобретению. В некоторых вариантах осуществления фармацевтическую композицию вводят внутривенно. В некоторых вариантах осуществления, фармацевтическую композицию вводят в виде болюса и/или посредством непрерывной инфузии.
В конкретных вариантах осуществления лечения, профилактики и/или смягчения осложнений, связанных с хирургической операцией CABG, в частности повреждения миокарда, связанного с хирургической операцией CABG, способ по этому изобретению включает введение фармацевтической композиции перед хирургической операцией и продолжение введения в течение по меньшей мере 24 часов, в некоторых вариантах осуществления в течение приблизительно 48 часов или в некоторых вариантах осуществления в течение приблизительно 72 часов. В конкретном варианте осуществления данного аспекта этого изобретения, требуемой концентрации аптамера в плазме, например, в некоторых вариантах осуществления, в два раза превышающей эндогенную концентрацию белка комплемента, достигают посредством введения внутривенного болюса пациенту, подлежащему лечению, до, одновременно или после внутривенной инфузии более низкой дозы аптамера. В других вариантах осуществления данного аспекта этого изобретения, где аптамер/конъюгат обладает достаточно длительным временем полужизни, фармацевтическую композицию аптамера можно вводить один раз или два раза в сутки в качестве внутривенной болюсной дозы.
В другом аспекте этого изобретения, предусмотрены способы диагностики. В одном варианте осуществления, способ диагностики включает контактирование композиции с аптамером, содержащим нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO:2 и с SEQ ID NO:6 по SEQ ID NO:66, в частности, из группы, состоящей из SEQ ID NO:61, SEQ ID NO:62, и с SEQ ID NO:64 по SEQ ID NO:66, и детекцию наличия или отсутствия в композиции белка комплемента C5 или его варианта. В некоторых вариантах осуществления белок комплемента или вариант представляет собой белок или вариант позвоночного, в частности млекопитающего и более конкретно человека. Настоящее изобретение относится к композиции аптамера, имеющей аптамер, содержащий нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO:2 и с SEQ ID NO:6 по SEQ ID NO:66, для применения в качестве диагностического средства in vitro или in vivo. В настоящем изобретении предусмотрен аптамер, содержащий нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO:2 и с SEQ ID NO:6 по SEQ ID NO:66, для применения в целях изготовления фармацевтической композиции.
В другом аспекте этого изобретения, предусмотрен аптамер, содержащий нуклеотидную последовательность, которая на 80% идентична любой из последовательностей, выбранных из группы, состоящей из SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98. В некоторых вариантах осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность, которая на 80% идентична уникальному участку любой из последовательностей, выбранных из группы, состоящей из SEQ ID NO: с 75 по 81 и SEQ ID NO: с 88 по 98. В другом варианте осуществления предусмотрен аптамер, содержащий нуклеотидную последовательность, которая на 90% идентична любой из последовательностей, выбранных из группы, состоящей из SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98. В конкретном варианте осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность, которая на 90% идентична уникальному участку любой из последовательностей, выбранных из группы, состоящей из SEQ ID NO: с 75 по 81 и SEQ ID NO: с 88 по 98. В другом варианте осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность из 40 последовательно расположенных нуклеотидов, идентичную 40 последовательно расположенным нуклеотидам, включенным в любую из последовательностей, выбранных из группы, состоящей из SEQ ID NO: с 75 по 81 и SEQ ID NO: с 88 по 98. В другом варианте осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность из 30 последовательно расположенных нуклеотидов, идентичную 30 последовательно расположенным нуклеотидам, включенным в любую из последовательностей, выбранных из группы, состоящей из SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98. В другом варианте осуществления, предусмотрен аптамер, который специфично связывается с белком C5 комплемента, содержащий нуклеотидную последовательность из 10 последовательно расположенных нуклеотидов, идентичную 10 последовательно расположенным нуклеотидам, включенным в любую из последовательностей, выбранных из группы, состоящей из SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98. В предпочтительном варианте осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность в соответствии с любой из нуклеотидных последовательностей, выбранных из группы, состоящей из: SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98.
В некоторых вариантах осуществления, аптамеры по этому изобретению, описанные выше, далее могут обладать химической модификацией, выбранной из группы, состоящей из: химического замещения в положении сахара; химического замещения в положении фосфата и химического замещения в положении основания последовательности нуклеиновой кислоты. В некоторых вариантах осуществления модификация выбрана из группы, состоящей из: включения модифицированного нуклеотида; 3'-кэппирования, конъюгации с высокомолекулярным неиммуногенным соединением; конъюгации с липофильным соединением и модификации фосфатного остова.
В предпочтительных вариантах осуществления данного аспекта этого изобретения, аптамер модулирует функцию белка C5 комплемента или его варианта. В особенно предпочтительных вариантах осуществления, аптамер ингибирует функцию белка C5 комплемента или его варианта, предпочтительно in vivo, более предпочтительно in vivo у человека. В одном варианте осуществления данного аспекта этого изобретения, функция, модулируемая, предпочтительно ингибируемая, аптамером, представляет собой расщепление белка C5 комплемента.
В некоторых вариантах осуществления другого аспекта, настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество аптамера, который блокирует расщепление белка C5 комплемента in vivo или его соли и фармацевтически приемлемый носитель или разбавитель.
В некоторых вариантах осуществления, предусмотрена фармацевтическая композиция, содержащая терапевтически эффективное количество аптамера, содержащего нуклеотидную последовательность, на 80% идентичную, предпочтительно, на 90% идентичную нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98, или его соли. В некоторых вариантах осуществления, предусмотрена фармацевтическая композиция, содержащая терапевтически эффективное количество аптамера, содержащего нуклеотидную последовательность, на 80% идентичную, предпочтительно, на 90% идентичную уникальному участку нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98, или его соли. В других вариантах осуществления, предусмотрена фармацевтическая композиция, содержащая терапевтически эффективное количество аптамера, обладающего 40, 30 или 10 последовательно расположенными нуклеотидами, идентичными 40, 30 или 10 нуклеотидам, соответственно, нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98. Фармацевтическая композиция по этому изобретению может содержать фармацевтически приемлемый носитель или разбавитель. В данном аспекте этого изобретения, предусмотрена фармацевтическая композиция для применения в целях лечения, профилактики или смягчения заболевания in vivo, где фармацевтическая композиция содержит аптамер, обладающий нуклеотидной последовательностью, выбранный из группы, состоящей из: SEQ ID NO: с 3 по 4, SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98, или его соль. В этом аспекте, предусмотрен аптамер, обладающий нуклеотидной последовательностью, выбранной из группы, состоящей из: SEQ ID NO: с 3 по 4, SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98 для применения в целях изготовления фармацевтической композиции. В этом аспекте, настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество аптамера, который ингибирует расщепление белка C5 комплемента in vivo, или его соли и фармацевтически приемлемый носитель или разбавитель.
В некоторых вариантах осуществления, опосредуемое C5 белком комплемента, C5a и/или C5b-9 заболевание, подлежащее лечению, представляет собой острые ишемические заболевания (инфаркт миокарда, инсульт, повреждение при ишемии/реперфузии); острые воспалительные заболевания (инфекционное заболевание, септицемия, шок, острое/сверхострое отторжение трансплантата); хронические воспалительные и/или опосредуемые иммунной системой заболевания, включая диабетическую ретинопатию, дегенерацию желтого пятна, включая экссудативную и неэкссудативную формы AMD, и также включая аллергию, астму, ревматоидный артрит и другие ревматологические заболевания, рассеянный склероз и другие неврологические заболевания, псориаз и другие дерматологические заболевания, миастению, системную красную волчанку (SLE), подострое/хроническое отторжение трансплантата, гломерулонефрит и другие заболевания почек); и глазные заболевания. В некоторых вариантах осуществления, опосредуемые белком комплемента C5, C5a и/или C5b-9 заболевания, подлежащие лечению, включают активацию комплемента, обусловленную диализом или ситуациями, когда кровь проходит над и/или через синтетическую трубку и/или чужеродный материал. В некоторых вариантах осуществления, опосредуемое белком комплемента C5, C5a и/или C5b-9 заболевание, подлежащее лечению, выбрано из группы, состоящей из повреждения миокарда, связанного с хирургической операцией CABG, повреждения миокарда, связанного с баллонной ангиопластикой, и повреждения миокарда, связанного с рестенозом. В некоторых вариантах осуществления, опосредуемое белком комплемента C5, C5a и/или C5b-9 нарушение, подлежащее лечению, выбрано из группы, состоящей из: повреждения миокарда, связанного с хирургической операцией CABG, повреждения миокарда, связанного с баллонной ангиопластикой, повреждения миокарда, связанного с рестенозом, опосредуемых белком комплемента осложнений, связанных с хирургической операцией CABG, опосредуемых белком комплемента осложнений, связанных с чрескожным вмешательством на коронарных сосудах, пароксизмальной ночной гемоглобинурии, острого отторжения трансплантата, сверхострого отторжения трансплантата, подострого отторжения трансплантата и хронического отторжения трансплантата. В некоторых вариантах осуществления опосредуемое белком комплемента C5, C5a и/или C5b-9 заболевание, подлежащее лечению, представляет собой осложнения, связанные с хирургической операцией CABG. В конкретном варианте осуществления, заболевание, подлежащее лечению, представляет собой повреждение миокарда, связанное с хирургической операцией CABG. В конкретном варианте осуществления способа лечения по этому изобретению, заболевание, симптом которого подлежит уменьшению, стабилизации и/или профилактике, представляет собой глазное нарушение, в частности диабетическую ретинопатию, экссудативную и/или неэкссудативную AMD.
В некоторых вариантах осуществления, способ по этому изобретению включает введение фармацевтической композиции, содержащей аптамер, обладающий нуклеотидной последовательностью, выбранной из группы, состоящей из: SEQ ID NO: с 3 по 4, SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98, пациенту для достижения концентрации аптамера в плазме, которая изменена приблизительно в от 0,5 до 10 раз по сравнению с концентрацией эндогенного белка C5 комплемента. В некоторых вариантах осуществления, фармацевтические композиции аптамера вводят для достижения концентрации аптамера в плазме, которая изменена в от приблизительно 0,75 до приблизительно 5 раз, в от 0,75 до приблизительно 3 раз и в от 1,5 до приблизительно 2 раз по сравнению с концентрацией эндогенного белка C5 комплемента, в то время как в других вариантах осуществления композицию аптамера вводят для достижения концентрации, эквивалентной концентрации эндогенного белка комплемента. В некоторых вариантах осуществления, фармацевтическую композицию по этому изобретению вводят для достижения концентрации аптамера в плазме приблизительно 5 мкМ, приблизительно 4 мкМ, приблизительно 3 мкМ, приблизительно 2 мкМ, приблизительно 1,5 мкМ, приблизительно 1 мкМ или приблизительно 500 нМ.
Можно применять любое сочетание способа, длительности и скорости введения, которые достаточны для достижения концентраций аптамера в плазме по этому изобретению. В некоторых вариантах осуществления фармацевтическую композицию вводят внутривенно. В некоторых вариантах осуществления, фармацевтическую композицию вводят в виде болюса и/или посредством непрерывной инфузии.
В конкретных вариантах осуществления лечения, профилактики и/или смягчения осложнений, связанных с хирургической операцией CABG, в частности повреждения миокарда, связанного с хирургической операцией CABG, способ по этому изобретению включает введение фармацевтической композиции перед хирургической операцией и продолжение введения в течение по меньшей мере 24 часов, в некоторых вариантах осуществления в течение приблизительно 48 часов или в некоторых вариантах осуществления в течение приблизительно 72 часов. В конкретном варианте осуществления данного аспекта этого изобретения, требуемую концентрацию аптамера в плазме, например, в некоторых вариантах осуществления, в два раза превышающей концентрацию эндогенного белка комплемента, достигают посредством введения внутривенного болюса пациенту, подлежащему лечению, до, одновременно или после внутривенной инфузии более низкой дозы аптамера. В других вариантах осуществления данного аспекта этого изобретения, где аптамер/конъюгат обладает достаточно длительным временем полужизни, фармацевтическую композицию аптамера можно вводить один раз или два раза в сутки в качестве внутривенной болюсной дозы.
В другом варианте осуществления предусмотрен диагностический способ, при этом способ включает контактирование композиции, предположительно содержащей белок комплемента C5 или его вариант, с аптамером, содержащим нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98, и детекцию наличия или отсутствия белка C5 комплемента или его варианта. В некоторых вариантах осуществления, белок комплемента или вариант представляют собой белок или вариант позвоночного, в частности млекопитающего, и более конкретно человека. Настоящее изобретение относится к композиции аптамера, имеющей аптамер, содержащий нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO: с 75 по 81, SEQ ID NO:83 и SEQ ID NO: с 88 по 98 для применения в качестве диагностического средства in vitro или in vivo.
В некоторых вариантах осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность, по существу состоящую из нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO:68 и 69. В некоторых вариантах осуществления, предусмотрен аптамер, содержащий нуклеотидную последовательность, состоящую из нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO:68 и 69. В некоторых вариантах осуществления данного аспекта этого изобретения, аптамеры можно применять в диагностическом способе.
В одном варианте осуществления, способ по настоящему изобретению направлен на лечение, стабилизацию и/или профилактику опосредуемого комплементом глазного нарушения, при этом способ включает стадию введения терапевтически эффективного количества аптамера против комплемента субъекту, нуждающемуся в этом. В одном варианте осуществления глазное нарушение, подлежащее лечению, представляет собой дегенерацию желтого пятна. В одном варианте осуществления глазное нарушение, подлежащее лечению, представляет глазное нарушение, связанное с неоваскуляризацией.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Детали одного или нескольких вариантов осуществления этого изобретения указаны в прилагаемом описании, ниже. Несмотря на то, что при применении на практике или тестировании настоящего изобретения можно использовать любые способы и материалы, сходные или эквивалентные способам и материалам, описанным в настоящем документе, предпочтительные способы и материалы описаны в настоящем документе. Другие признаки, цели и преимущества этого изобретения будут очевидны из описания. В описании формы единственного числа также включают множественное число, если контекст явно не подразумевает иное. Если нет иных указаний, все технические и научные термины, используемые в настоящем документе, обладают тем же значением, которое обычно подразумевает специалист в области, к которой относится настоящее изобретение. В случае противоречий следует руководствоваться настоящим описанием.
Средства против C5 и глазные нарушения
Последние данные позволяют предположить, что возрастная дегенерация желтого пятна (AMD) также представляет собой опосредуемое воспалением заболевание, в котором участвует активация комплемента. Связанная со старением дегенерация желтого пятна ("AMD") представляет собой хроническое и прогрессирующее заболевание глаза, и она является основной причиной невосстанавливаемой потери зрения в США, Европе и Японии. AMD характеризуется прогрессирующей деградацией центральной части сетчатки, называемой желтым пятном. Наиболее отчетливым показателем прогрессирования AMD является возникновение друз, желто-белых отложений под сетчаткой, которые представляют собой бляшки материала, источником которого являются метаболические продукты жизнедеятельности клеток сетчатки. Возникновение друз является важным компонентом обеих форм AMD: экссудативной ("влажной") и неэкссудативной ("сухой"). Влажная AMD происходит, когда новые кровеносные сосуды, растущие непосредственно под сетчаткой, проникают в слои сетчатки через мембрану, известную как мембрана Бруха. Этот аномальный рост кровеносных сосудов обычно называют ангиогенезом или (хориоидальной) неоваскуляризацией. Эти новые кровеносные сосуды обладают тенденцией к ломкости, и часто из них происходит кровотечение и проникновение жидкости в желтое пятно, что иногда приводит к внезапному и часто тяжелому нарушению зрения. Несмотря на то, что новые лекарственные средства (например, LucentisTM) могут остановить ангиогенез и вызывать реверсию накопления жидкости, даже восстанавливая зрение у небольшого числа пациентов, неоваскулярное повреждение часто приводит к рубцеванию и/или повреждению клеток сетчатки, вызывая необратимую потерю зрения. Влажной AMD, как правило, предшествует образование друз, которые накапливаются и которые содержат белки комплемента, включая фактор H комплемента (CFH). Наличие множества друз, от промежуточных до крупных, ассоциировано с наибольшим риском прогрессирования в заболевание поздней стадии, характеризующееся географической атрофией и/или неоваскуляризацией. Большинство пациентов с влажной AMD испытывают тяжелое снижение зрения в поврежденном глазу в течение от нескольких месяцев до двух лет после постановки диагноза заболевания, хотя потеря зрения может произойти в течение нескольких часов или суток. Сухая AMD является более постепенной и происходит при медленной атрофии светочувствительных клеток в желтом пятне, постепенно размывая центральное зрение в поврежденном глазу. Потеря зрения усиливается образованием и накоплением друз и иногда повреждением сетчатки, хотя и без аномального роста сосудов и кровотечения.
Существуют компоненты комплемента и другие медиаторы воспаления в друзах и воспалительные клетки, которые обнаруживают в очагах повреждения, вызывающих связанную с AMD потерю зрения (Haines et al. (2005) Science 308: 419-421). AMD сильно ассоциирована с генетическим дефектом в регулирующем комплемент каскаде (Haines et al., 2005). Вариант гена, кодирующего фактор H комплемента (CFH), по-видимому, приводит к повышенному риску AMD в основном или полностью через его влияние на развитие друз, предшествующих неоваскуляризации и потере зрения, ассоциированных с AMD (Edwards et al. (2005) Science 308:421-424). Первичной функцией CFH является снижение активности конвертаз альтернативного каскада посредством связывания и инактивации C3b или посредством стимуляции диссоциации фактора Bb от C3b. (Hageman et al. (2005) PNAS 102(2):7227-7232). Последние генетические данные позволяют предположить, что индивид с друзами по типу влажной AMD обладает 50-70% вероятностью наличия аллеля CFH, результатом чего является высокопредсказательная и сильная корреляция между вмешательством комплемента и лечением AMD (Klein et al. (2005) Science 308: 385-389). Аптамер против C5, который блокирует его активацию, будет блокировать ее, даже если пациент обладает дефектным геном CFH, ассоциированным с AMD. Аналогично, аптамеры, которые ингибируют активность мишеней из альтернативного каскада, таких как фактор B, фактор D и пропердин, компонентов обычного каскада, в частности C3, и компонентов мембраноатакующего комплекса, будут ингибировать активность, даже если пациент обладает дефектным геном CFH, ассоциированным с AMD. Было определено несколько гаплотипов гена фактора комплемента H (CFH), ассоциированных у индивида с риском развития дегенерации желтого пятна (AMD). В частности, замена тирозин-гистидин в 402 аминокислоте фактора комплемента H (CFH) на 1 хромосоме приводит к образованию варианта гена CFH, строго ассоциированного с предрасположенностью к заболеванию. Изменение последовательности находится в области CFH, которая связывает гепарин и C-реактивный белок. Люди, генетический набор которых включает этот вариант гена CFH, обладают большей вероятностью развития AMD. Вариант гена CFH может быть ответственным приблизительно за половину из 15 миллионов случаев дегенерации желтого пятна в США. Шансы развития дегенерации желтого пятна повышаются приблизительно в от 2,5 до 5,5 раз, если индивид обладает вариантом гена CFH.
Ген CFH вовлечен в регуляцию воспалительных каскадов (альтернативный каскад комплемента). Это подразумевает, что воспаление также играет важную роль в развитии дегенерации желтого пятна. Также было выявлено, что уровни в крови воспалительного маркера C-реактивного белка (CRP) повышены при дегенерации желтого пятна (Science. 2005 Apr 15;308(5720):419-21, Science. 2005 Apr 15;308(5720):421-4, Science. 2005 Apr 15;308(5720):385-9). Исходя из расположения замены в связывающем гепарин и CRP участке фактора H, вариант Y402H может нарушать опосредуемое протеогликанами и CRP привлечение фактора H к поверхностям клеток-хозяев, препятствуя способности фактора H снижать активность C3b, находящегося на этих клетках. Беспрепятственное усиление каскада комплемента в ткани хозяина вызывает воспаление в сетчатке и окружающих кровеносных сосудах вследствие неконтролируемого высвобождения C5a и C5b-9. CFH препятствует неконтролируемой активации комплемента и воспалению; таким образом, мутация в CFH приведет к повышению воспаления и его последствий. Посредством снижения чрезмерной активации комплемента (воспалительного каскада), которая происходит при дегенерации желтого пятна, можно замедлить прогрессирование заболевания. Кроме того, детекцию варианта гена когда-нибудь можно будет использовать в сочетании с технологиями визуализации для идентификации индивидов с высоким риском развития развернутой AMD раньше, чем это возможно в настоящее время (JAMA. 2005 Apr 20; 293(15):18410).
Также активация комплемента вовлечена в другие глазные заболевания, такие как диабетическая ретинопатия, и она может усугублять или запускать повреждение сосудов сетчатки (Zhang et al., (2002) Diabetes 51:3499). Пролиферативная диабетическая ретинопатия (PDR) представляет собой осложнение диабета, причиной которого являются изменения кровеносных сосудов сетчатки. Когда кровеносные сосуды сетчатки повреждаются, из них может вытекать кровь и отрастать ломкие, кисточковидные ветви и рубцовая ткань. Они могут нарушать четкость или искажать зрительные изображения, которые сетчатка посылает в головной мозг. Установлено, что 25% диабетиков страдают диабетической ретинопатией и заболеваемость ею возрастает до 60% после 5 лет и до 80% после 10-15 лет диабета I типа. Это заболевание характеризуется гипергликемией, утолщением базальной мембраны, потерей перицитов, микроаневризмами и преретинальной неоваскуляризацией, которые могут привести к слепоте посредством геморрагии и частичного отслоения сетчатки. Непролиферативная диабетическая ретинопатия характеризуется внутриретинальными микроаневризмами, геморрагиями, инфарктами в нервно-фиброзном слое, плотными экссудатами и аномалиями микрососудов. Отек желтого пятна является основным механизмом потери зрения. Он возникает вследствие истечения из микроаневризм в капиллярах желтого пятна (центральной области сетчатки). Истечение может прогрессировать до утолщения желтого пятна, ассоциированного с плотными экссудатами или кистоидными изменениями, и это часто приводит к потере центрального зрения в различной степени. Пролиферативная диабетическая ретинопатия характеризуется неоваскуляризацией сетчатки. Ее классифицируют в соответствии с наличием, расположением, тяжестью и ассоциированной геморрагической активностью неоваскуляризации сетчатки. Она ассоциирована с тяжелой потерей зрения. Патология диабетической ретинопатии может быть свойственна следующим болезненным состояниям. Проблемы с кровообращением приводят к тому, что области сетчатки становятся лишенными кислорода или ишемизированными. Неоваскуляризация приводит к тому, что в стекловидном теле начинают расти новые кровеносные сосуды для поддержания надлежащих уровней кислорода. Кровь, просачивающаяся из вновь образованных капилляров, и образование рубцовой ткани приводят к тракции сетчатки, вызывая небольшие разрывы. После разрывов происходит скопление жидкости под или между слоями сетчатки и отслоение. Пациенты ощущают размытое изображение, плавающие помутнения, вспышки света и внезапную потерю зрения вследствие геморрагии, отека и образования рубцовой ткани.
В норме в недиабетическом глазу происходит конститутивная активация комплемента на низком уровне, на что указывает наличие MAC и регулирующих комплемент белков в глазах недиабетических крыс, показывая, что у диабетических пациентов происходит нарушение регуляции комплемента (Sohn et al., (2000) IOVS 41: 3492). Кроме того, в сосудах сетчатки диабетических доноров-людей выявлено отложение C5b-9, где оно отсутствует у недиабетических доноров-людей (Zhang et al.), в диабетических сетчатках показана сниженная экспрессия CD55 и CD59 (Zhang et al.), и в моче диабетических пациентов, но не у диабетических пациентов, имеется гликозилированный CD59 (Acosta et al., (2002) PNAS 97, 5450-5455). Кроме того, известно, что при диабете I типа активируются система комплемента и сосудистая система. См., например, Hansen, T.K. et al., Diabetes, 53: 1570-1576 (2004). C5a активирует эндотелиальные клетки посредством взаимодействия с иммунной системой и системой комплемента. См., например, Albrecht, E.A. et al., Am J Pathology, 164: 849-859 (2004). Сосудистая система активируется при глазных заболеваниях, включая диабетическую ретинопатию. См., например, Gert, V.B. et al., Invest Opthalmol Vis Sci, 43: 1104-1108 (2002). Также система комплемента активируется при диабетической ретинопатии. См., например, Gert, V.B. et al., Invest Opthalmol Vis Sci, 43: 1104-1108 (2002) и Badouin, C et al., Am J Opthalmol, 105:383-388 (1988).
Увеит относится к воспалению сосудистой оболочки глаза, пигментного сосудистого слоя (или оболочки), расположенного в склере (фиброзной оболочке) и сетчатке (светочувствительном слое, ответственном за зрение) и состоящего из радужной оболочки, ресничного тела и сосудистой оболочки (сосудистого и соединительного слоя, который питает сетчатку). Вследствие комплексной анатомии сосудистой оболочки глаза, существует множество различных типов увеита, и, вследствие анатомической взаимосвязи сосудистой оболочки с другими структурами глаза (например, сетчаткой), увеит часто сопровождается воспалением других тканей (например, ретинитом). Как правило, увеит относится к категории переднего увеита (в основном поражающего радужную оболочку и связанное с ней ресничное тело, и, таким образом, переднюю камеру между радужной оболочкой и роговицей и находящуюся в ней прозрачную жидкую водянистую влагу), промежуточного увеита (поражающего плоскую часть непосредственно за ресничным телом и на переднем "крае" сетчатки), или задний увеит (поражающий заднюю камеру позади радужной оболочки, которая заполнена прозрачным гелеобразным стекловидным телом и находится в прямом контакте с сетчаткой). Увеит может вовлекать любую или все части сосудистой оболочки глаза (например, хориоидит, ирит). Передний увеит является наиболее распространенной формой и часто он ассоциирован с аутоиммунными заболеваниями, такими как ревматоидный артрит. Промежуточный увеит является второй из наиболее распространенных форм. Задний увеит, наименее распространенная форма, может возникать после системной инфекции (например, вирусной, бактериальной, грибковой или паразитической) или он может быть ассоциирован с аутоиммунными заболеваниями. Аутоиммунный увеит также может встречаться без системного вовлечения. Также увеит может возникать в результате травмы (например, ранения, хирургической операции, и т.д.) или вследствие неизвестных причин ("идиопатический"). Независимо от этиологии, любое воспалительное состояние сосудистой оболочки глаза по определению приведет к увеиту.
В норме жидкости и ткани глаза содержат многие компоненты комплемента, такие как фактор B и C2, C4, C3, C5, C6 и C7 в увеальной ткани (Brawman-Mintzer O, Invest Ophthalmol Vis Sci. 1989 Oct;30(10):2240-4), C1, C4, C3 и C5 в водянистой влаге (Mondino BJ, Arch Ophthalmol. 1983 Mar;101(3):465-8) и C1, C4, C2, C3, C5, C6 и C7 в роговице (Mondino BJ, Arch Ophthalmol. 1981 Aug; 99(8): 1430-3). Полагают, что эти компоненты комплемента и ассоциированные с ними регулирующие комплемент белки важны для нормального иммунного надзора в тканях глаза.
Важная роль активации комплемента в патогенезе увеита была показана посредством: повышенной встречаемости аллотипа C4 (C4B2) у пациентов с передним увеитом по сравнению с (контрольными) пациентами с васкулитом сетчатки (Wakefield D, Hum Immunol. 1988 Apr;21(4):233-7.); повышения содержания в плазме C3d и содержащих комплемент иммунных комплексов у пациентов с идиопатическим увеитом (Vergani S, Br J Ophthalmol. 1986 Jan;70(l):60-3); повышения опосредуемой комплементом гемолитической активности и уровней C3 и C4 в слезной жидкости (в слезах) у пациентов с системными воспалительными заболеваниями и сопутствующим увеитом; (Drozdova EA, Vestn Oftalmol. 2004 Jul-Aug;120(4):24-6); и отложения иммунных комплексов и комплемента в увеальном тракте при увеите в качестве инициирующего процесса (O'Connor GR, Trans Ophthalmol Soc U K. 1981 Sep;101 (Pt 3)(3):297-300). Активация комплемента вовлечена во множество воспалительных и/или аутоиммунных заболеваний с глазным компонентом в форме увеита, включая болезнь Бехчета (Cuchacovich M, Clin Exp Rheumatol. 2005 Jul-Aug; 23(4 Suppl 38):S27-34, Bardak Y, Ocul Immunol Inflamm. 2004 Mar;12(l):53-8), гетерохромный иридоциклит Фуха (La Hey E, Am J Ophthalmol. 1992 Jan 15;113(l):75-80), болезнь Фогта-Каянаги-Харада (Sakamoto T, Arch Ophthalmol. 1991 Sep; 109(9): 1270-4), и субретинальный фиброз и хронический увеит (Palestine AG, Ophthalmology. 1985 Jun;92(6):838-44).
Ряд моделей экспериментально индуцированного увеита на животных также показал, что активация комплемента важна в патогенезе увеита (Jha P, Invest Ophthalmol Vis Sci.2006 Mar; 47(3):1030-8, Kasp E, Clin Exp Immunol. 1992 May; 88(2):307-12) и что активация комплемента находится под сильной регуляцией регулирующих комплемент белков (Bardenstein DS, Immunology.2001 Dec; 104(4):423-30, Sohn JH, Invest Ophthalmol Vis Sci. 2000 Oct; 41(ll):3492-502). В этих моделях опосредуемого комплементом увеита устранение комплемента, например, введением фактора яда кобры (CVF) или посредством генетического устранения важных компонентов каскадов активации комплемента (C3), приводит к предотвращению или значительному снижению тяжести индуцированного увеита.
В совокупности, данные указывают на то, что компоненты комплемента и регуляторные белки являются важными нормальными составляющими тканей глаза и сосудистой оболочки глаза, в частности, что активация комплемента ответственна за увеит в экспериментальных моделях аутоиммунного увеита, и что активация комплемента ассоциирована с увеитом у человека. Таким образом, в некоторых вариантах осуществления предусмотрены аптамеры, которые останавливают альтернативный и классический каскады активации комплемента до активации C5 и последующего образования C5a и C5b-9 (MAC), для применения в способах по этому изобретению для снижения встречаемости, длительности и/или тяжести увеита.
Глаукома относится к группе заболеваний, при которых происходит потеря зрения в результате повреждения оптического нерва и сетчатки, обычно ассоциированного с повышением внутриглазного давления ("IOP"). Первичная открытоугольная глаукома представляет собой наиболее распространенную форму, вызываемую постепенным сужением или блокадой дренажных каналов для жидкости (т.е. трабекулярной сети) с течением времени, что приводит к повышению IOP вследствие скопления водянистой влаги. Глаукому классифицируют на две категории: открытоугольная и закрытоугольная глаукома. Первичная открытоугольная глаукома развивается медленно и безболезненно, часто без заметной потери зрения в течение нескольких лет. Вторичную открытоугольную глаукому вызывают другие заболевания (например, увеит или другое воспалительное заболевание, диабет, опухоль, катаракта), повреждение тупым предметом или определенными лекарственными средствами, такими как стероиды. Закрытоугльную глаукому (также называемую узкоугольной глаукомой или острой глаукомой) вызывает смещение положения радужной оболочки, приводящее к внезапной блокаде оттока водянистой влаги и быстрому повышению IOP. Симптомы закрытоугольной глаукомы, помимо потери зрения, включают боль в глазу и тошноту. У некоторых индивидов повреждение нерва происходит без повышения IOP; этот тип глаукомы известен как нормотензивная глаукома (глаукома нормального давления или низкого давления). Причина повреждения нерва при этом типе глаукомы неизвестна.
Роль комплемента в патогенезе глаукомы показана с помощью: 1) повышенной экспрессии в сетчатке мРНК компонентов комплемента (C1q, C1r, C1s, C3) при экспериментальном повышении IOP в модели глаукомы на крысах (Ahmed, F, Brown, KM, Stephan, DA, Morrison, JC, Johnson, EC, Tomarev, SI (2004) IOVS 45, 1247-54); 2) повышенной экспрессии в сетчатке мРНК компонентов комплемента (C4, B, C1q, C3) в модели глаукомы на яванских макаках (Miyahara, T, Kikuchi, T, Akimoto, M, Kurokawa, T, Shibuki, H, Yoshimura, N (2003) IOVS 44, 4347-56); 3) повышенной экспрессии мРНК и белка C1q в глиальных клетках сетчатки в моделях глаукомы на мышах и обезьянах (Stasi, K, Nagel, D, Yang, X, Wang, R, Ren, L, Podos, SM, Mittag, T, Danias, J (2006) IOVS 47, 1024-29); 4) иммуногистохимического окрашивания на C1q в глиальных клетках субъектов-людей (Stasi et al., 2006). Оказалось, что экспрессия компонента комплемента C1q у экспериментальных мышей с глаукомой коррелирует с повышением IOP с течением времени и предшествует повреждению ганглионарных клеток сетчатки, позволяя предположить, что комплемент может участвовать в патогенезе заболевания (Stasi et al., 2006). Лечение аптамером против комплемента, например, аптамером против C5, может обладать защитным эффектом против нейродегенерации при глаукомах высокого и низкого давления.
Таким образом, в некоторых вариантах осуществления настоящее изобретение относится к средствам против C5 для лечения опосредуемых комплементом глазных нарушений. В некоторых вариантах осуществления, средство против C5 по этому изобретению применяют отдельно, в то время как в других вариантах осуществления его применяют в сочетании со средством против VEGF и/или против PDGF.
Другие варианты осуществления настоящего изобретения относятся к аптамерам против комплемента для лечения, стабилизации и/или профилактики опосредуемых комплементом глазных нарушений. Аптамеры против комплемента можно получать способом SELEXTM. В конкретных вариантах осуществления, настоящее изобретение включает введение аптамера против комплемента, например аптамера против C5, субъекту в способе снижения, стабилизации и/или профилактики по меньшей мере одного симптома глазного нарушения, в частности симптома диабетической ретинопатии, экссудативной и/или неэкссудативной AMD.
Специфичные к C5 аптамеры
Специфичные к C5 аптамеры для применения в целях лечения, стабилизации, профилактики и/или снижения симптомов опосредуемых комплементом глазных нарушений можно получать способом SELEXTM. В конкретных вариантах осуществления, настоящее изобретение включает введение средства на основе аптамера против C5 субъекту в способе снижения, стабилизации и/или профилактики по меньшей мере одного симптома глазного нарушения, в частности симптома диабетической ретинопатии, экссудативной и/или неэкссудативной AMD.
Аптамеры представляют собой молекулы нуклеиновых кислот, обладающие аффинностью специфичного связывания с молекулами через взаимодействия, отличные от классического образования пар оснований по принципу Уотсона-Крика.
Аптамеры, подобно пептидам, получаемым с помощью фагового дисплея, или моноклональным антителам ("mAb"), способны специфично связываться с определенными мишенями и модулировать активность мишени, например, аптамеры посредством связывания могут блокировать способность их мишени к осуществлению функций. Созданные посредством процесса селекции in vitro из наборов олигонуклеотидов со случайными последовательностями, аптамеры были получены более чем для 100 белков, включая факторы роста, факторы транскрипции, ферменты, иммуноглобулины и рецепторы. Размер типичного аптамера составляет 10-15 кДа (30-45 нуклеотидов), он связывается со своей мишенью с субнаномолярной аффинностью, и отличает высокосходные мишени (например, аптамеры, как правило, не связывают другие белки из того же семейства генов). В ряде структурных исследований было показано, что аптамеры способны использовать те же типы связывающих взаимодействий (например, образование водородных связей, электростатическую комплементарность, гидрофобные взаимодействия, пространственное вытеснение), какие определяют аффинность и специфичность в комплексах антитело-антиген.
Аптамеры обладают рядом свойств, желательных для применения в качестве средств для диагностики и лечения, включая высокую специфичность и аффинность, биологическую эффективность и превосходные фармакокинетические свойства.
СПОСОБ SELEX TM
Предпочтительным способом получения аптамеров является процесс, называемый "Систематическим перестроением лигандов посредством экспоненциального обогащения" ("SELEXTM"), представленный в общих чертах на Фиг.2. Процесс SELEXTM, способ перестроения молекул нуклеиновых кислот in vitro с высокоспецифичным связыванием с молекулами-мишенями, описан, например, в патентной заявке № 07/536428, поданной 11 июня 1990, в настоящее время аннулированной, в патенте США № 5475096 с названием "Nucleic acid Ligands" и в патенте США № 5270163 (см. также WO 91/19813), с названием "Nucleic acid Ligands". Посредством многократного проведения циклов селекции и амплификации, SELEXTM можно применять для получения аптамеров, также называемых в настоящем документе "нуклеиновыми кислотами-лигандами" с любым требуемым уровнем аффинности связывания мишени.
Процесс SELEXTM основан на уникальном открытии, что нуклеиновые кислоты обладают достаточной способностью к формированию множества двумерных и трехмерных структур и достаточной химической универсальностью, имеющейся у их мономеров, для того, чтобы выступать в качестве лигандов (т.е. формировать специфично связанные пары) практически для любого химического соединения, как мономерного, так и полимерного. В качестве мишеней могут выступать молекулы любого размера или состава.
Процесс SELEXTM основан на способности связывать мишень. Аптамеры, полученные способом SELEXTM, таким образом, будут обладать свойством связывания мишени. Однако сам по себе SELEXTM не осуществляет отбор по другим свойствам аптамеров или мишени, и обоснованно нельзя ожидать, что полученный способом SELEXTM аптамер будет обладать каким-либо свойством, отличным от связывания с требуемой мишенью, хотя можно надеяться, что полученные аптамеры будут обладать другими свойствами. Таким образом, несмотря на то, что можно надеяться, что аптамер будет оказывать конкретный эффект на мишень, помимо связывания с мишенью, данный аптамер может не обладать эффектом или может оказывать несколько эффектов. Например, когда мишень представляет собой белок, который взаимодействует со множеством рецепторов клеточной поверхности аптамер может действовать, либо блокируя, либо усиливая связывание между белком и одним или несколькими такими рецепторами, или он может не обладать каким-либо эффектом ни на одно из взаимодействий. В другом примере мишень может быть каталитической, и аптамер может блокировать или усиливать эффективность каталитической функции или не оказывать эффекта на каталитическую функцию. Однако перед тестированием в соответствующем анализе квалифицированный специалист не способен предсказать, каким свойством, в случае его наличия, в действительности обладает аптамер. Действительно, только связывание мишени не предоставляет информации о функциональном эффекте, в случае его наличия, который может быть произведен в отношении мишени посредством воздействия связывания аптамером.
Изменение свойства молекулы-мишени требует, чтобы аптамер в целях достижения изменения свойства мишени связывался с определенным участком на мишени. В теории SELEXTM может приводить к идентификации большого количества аптамеров, где каждый аптамер связывается в отличающемся участке мишени. На практике связывающие взаимодействия аптамер-мишень часто происходят в одном или относительно небольшом количестве предпочтительных участков связывания на мишени, которые предоставляют стабильные и доступные структурные поверхности для взаимодействия. Более того, когда SELEXTM проводят на физиологической молекуле-мишени, специалист обычно не способен контролировать положение аптамера в отношении мишени. Таким образом, положение участка связывания аптамера на мишени может находиться в области, или вблизи нее, одного из нескольких потенциальных участков связывания, которые могут привести к требуемому эффекту, или оно может не оказывать какого-либо эффекта на молекулу-мишень.
Даже когда выявляют, что аптамер, вследствие его способности связывать мишень, обладает эффектом, отсутствует способ предсказания существования этого эффекта или преждевременного знания о том, каким будет эффект. При проведении эксперимента SELEXTM специалист может только узнать с определенной точностью, что аптамеры, в пределах возможности получения аптамера против мишени, будут обладать свойством связывания мишени. Можно проводить эксперимент SELEXTM в надежде, что некоторые идентифицированные аптамеры также будут оказывать эффект на мишень, помимо связывания с ней, однако это точно неизвестно.
В качестве исходной точки SELEXTM основывается на крупной библиотеке или наборе одноцепочечных олигонуклеотидов, содержащих случайные последовательности. Олигонуклеотиды могут представлять собой модифицированную или немодифицированную ДНК, РНК или гибриды ДНК/РНК. В некоторых примерах набор содержит 100% случайные или частично случайные олигонуклеотиды. В других примерах набор содержит случайные или частично случайные олигонуклеотиды, содержащие по меньшей мере одну постоянную последовательность и/или консервативную последовательность, встроенную в случайную последовательность. В других примерах набор содержит случайные или частично случайные олигонуклеотиды, содержащие по меньшей мере одну постоянную и/или консервативную последовательность на их 5'- и/или 3'-конце, которая может содержать последовательность, общую для всех молекул набора олигонуклеотидов. Постоянные последовательности представляют собой последовательности, общие для олигонуклеотидов в наборе, которые встроены для предварительно выбранной цели, такие как мотивы CpG, дополнительно описанные ниже, участки для гибридизации праймеров ПЦР, промоторные последовательности для РНК-полимераз (например, T3, T4, T7 и SP6), участки рестрикции или гомополимерные последовательности, такие как поли-A- или поли-T-участки, каталитические центры, участки для селективного связывания на аффинных колонках и другие последовательности для упрощения клонирования и/или секвенирования представляющего интерес олигонуклеотида. Консервативные последовательности, в отличие от описанных ранее постоянных последовательностей, являются общими для ряда аптамеров, которые связываются с одной мишенью.
Олигонуклеотиды из набора предпочтительно включают случайные участки последовательности, а также постоянные последовательности, необходимые для эффективной амплификации. Как правило, олигонуклеотиды исходного набора содержат постоянные 5'- и 3'-концевые последовательности, которые фланкируют внутренний участок из 30-50 случайных нуклеотидов. Случайные нуклеотиды можно получать рядом способов, включая химический синтез и отбор по размеру из расщепленных случайным образом нуклеиновых кислот клеток. Изменение последовательностей анализируемых нуклеиновых кислот также можно вносить или увеличивать мутагенезом до или в процессе повторений отбора/амплификации.
Участок случайной последовательности олигонуклеотида может обладать любой длиной и может содержать рибонуклеотиды и/или дезоксирибонуклеотиды, и может включать модифицированные или неприродные нуклеотиды или аналоги нуклеотидов. См., например, патент США № 5958691; патент США № 5660985; патент США № 5958691; патент США № 5698687; патент США № 5817635; патент США № 5672695 и публикацию PCT WO 92/07065. Случайные олигонуклеотиды можно синтезировать из нуклеотидов, связанных фосфодиэфирной связью, с использованием технологий твердофазного синтеза олигонуклеотидов, хорошо известных в данной области. См., например, Froehler et al., Nucl. Acid Res. 14:5399-5467 (1986) и Froehler et al., Tet. Lett. 27:5575-5578 (1986). Случайные олигонуклеотиды также можно синтезировать с использованием жидкофазных способов, таких как способы триэфирного синтеза. См., например, Sood et al., Nucl. Acid Res. 4:2557 (1977) и Hirose et al., Tet. Lett., 28:2449 (1978). Типичный синтез проводят на автоматическом оборудовании для синтеза ДНК с получением 1014-1016 отдельных молекул - количества, достаточного для большинства экспериментов SELEXTM. Достаточно крупные участки случайной последовательности при разработке последовательности повышают вероятность того, что каждая синтезированная молекула, вероятно, будет обладать уникальной последовательностью.
Исходная библиотека олигонуклеотидов может быть получена автоматическим химическим синтезом на устройстве для синтеза ДНК. Для синтеза случайных последовательностей на каждой стадии добавления нуклеотидов в процессе синтеза добавляют смеси всех четырех нуклеотидов, обеспечивая случайное встраивание нуклеотидов. Как указано выше, в одном варианте осуществления случайные олигонуклеотиды содержат полностью случайные последовательности; однако в других вариантах осуществления случайные олигонуклеотиды могут содержать участки неслучайных или частично случайных последовательностей. Частично случайные последовательности можно создавать добавлением четырех нуклеотидов в различных молярных соотношениях на каждой стадии добавления.
Исходная библиотека олигонуклеотидов может представлять собой библиотеку РНК, ДНК, замещенной РНК или ДНК или их комбинаций. В тех случаях, где в качестве исходной библиотеки подлежит применению РНК-библиотека, как правило, ее получают синтезом ДНК-библиотеки, необязательно посредством амплификации ПЦР, с последующей транскрипцией ДНК-библиотеки in vitro, используя РНК-полимеразу T7 или модифицированную РНК-полимеразу T7, и очисткой транскрибированной библиотеки. Затем библиотеку нуклеиновых кислот смешивают с мишенью в условиях, способствующих связыванию, и подвергают поэтапным повторениям связывания, разделения и амплификации с использованием указанной общей схемы отбора, достигая практически любого желательного критерия аффинности связывания и селективности. Более конкретно, начиная со смеси, содержащей исходный набор нуклеиновых кислот, способ SELEXTM включает следующие стадии: (a) контактирование смеси с мишенью в условиях, подходящих для связывания; (b) разделение не связавшихся нуклеиновых кислот и нуклеиновых кислот, которые специфично связались с молекулами-мишенями; (c) диссоциация комплексов нуклеиновая кислота-мишень; (d) амплификация нуклеиновых кислот, диссоциированных из комплексов нуклеиновая кислота-мишень, с получением смеси нуклеиновых кислот, обогащенной лигандом; и (e) повторение стадий связывания, выделения, диссоциации и амплификации на протяжении множества циклов, при желании, с получением высокоспецифичных высокоаффинных нуклеиновых кислот-лигандов для молекулы-мишени. В тех случаях, где отбору подвергаются РНК-аптамеры, способ SELEXTM дополнительно включает следующие стадии: (i) обратная транскрипция нуклеиновых кислот, диссоциированных из комплексов нуклеиновая кислота-мишень, перед амплификацией на стадии (d); и (ii) транскрипция амплифицированных на стадии (d) нуклеиновых кислот перед повторением процесса.
В смеси нуклеиновых кислот, содержащей большое количество возможных последовательностей и структур, для данной мишени существует широкий диапазон аффинности связывания. Те из них, которые обладают наибольшей аффинностью (наиболее низкими константами диссоциации) к мишени, обладают наибольшей вероятностью связаться с мишенью. После разделения, диссоциации и амплификации получают вторую смесь нуклеиновых кислот, обогащенную кандидатами с более высокой аффинностью связывания. Дополнительные этапы отбора постепенно способствуют получению наилучших лигандов до тех пор, пока полученная смесь нуклеиновых кислот не будет преимущественно включать только одну или несколько последовательностей. Затем их можно клонировать, секвенировать и индивидуально тестировать в отношении 1) аффинности связывания в отношении мишени; и 2) способности влиять на функцию мишени.
Циклы селекции и амплификации повторяют до тех пор, пока не достигают требуемой цели. В наиболее общем случае селекцию/амплификацию продолжают до тех пор, пока не достигают отсутствия значительного улучшения силы связывания при повторении цикла. Как правило, способ применяют для анализа приблизительно 1014 различных образцов нуклеиновых кислот, однако его можно применять для анализа вплоть до приблизительно 1018 различных образцов нуклеиновых кислот. Как правило, молекулы нуклеиновой кислоты-аптамера отбирают на от 5 до 20 процедуре цикла. В одном варианте осуществления гетерогенность вносят только на исходных стадиях селекции и она не возникает при повторении процесса.
В одном варианте осуществления способа SELEXTM процесс селекции является настолько эффективным в отношении выделения таких нуклеиновых кислот-лигандов, которые связываются наиболее сильно с определенной мишенью, что требуется только один цикл селекции и амплификации. Такую эффективную селекцию можно проводить, например, в хроматографическом процессе, где способность нуклеиновых кислот связываться с мишенями, связанными на колонке, действует таким образом, чтобы обеспечить достаточную эффективность разделения и выделения нуклеиновых кислот-лигандов с наибольшей аффинностью.
Во многих случаях не обязательно является желательным проведение повторных стадий SELEXTM до тех пор, пока не идентифицируют одну нуклеиновую кислоту-лиганд. Раствор специфичных к мишени нуклеиновых кислот-лигандов может включать семейство структур нуклеиновых кислот или мотивов, которые обладают рядом консервативных последовательностей и рядом последовательностей, которые могут быть замещены или добавлены без значительного влияния на аффинность нуклеиновых кислот-лигандов к мишени. Посредством остановки процесса SELEXTM перед завершением можно определить последовательность ряда членов семейства нуклеиновых кислот-лигандов в растворе.
Известно о существовании множества первичных, вторичных и третичных структур нуклеиновых кислот. Структурами или мотивами, для которых было показано, что они наиболее часто вовлечены во взаимодействия, не соответствующие взаимодействиям по типу Уотсона-Крика, считают петли-шпильки, симметричные и асимметричные выступы, псевдоузлы и большое количество их сочетаний. Практически все известные случаи таких мотивов подтверждают, что они могут быть образованы последовательностью нуклеиновой кислоты не более чем из 30 нуклеотидов. По этой причине часто является предпочтительным, чтобы процессы SELEXTM со смежными случайными сегментами начинались с последовательностей нуклеиновых кислот, содержащих случайный сегмент между приблизительно 20 и приблизительно 50 нуклеотидами и в некоторых вариантах осуществления между приблизительно 30 и приблизительно 40 нуклеотидами. В одном примере последовательность с сегментами 5'-постянный:случайный:3'-постоянный содержит случайную последовательность из от приблизительно 30 до приблизительно 50 нуклеотидов.
Основа способа SELEXTM была модифицирована для достижения ряда конкретных целей. Например, в патенте США № 5707796 описано применение SELEXTM совместно с гель-электрофорезом для селекции молекул нуклеиновых кислот с конкретными структурными элементами, такими как изгиб ДНК. В патенте США № 5763177 описаны способы на основе SELEXTM для селекции нуклеиновых кислот-лигандов, содержащих фотореактивные группы, способные связывать молекулу-мишень и/или образовывать поперечные связи в молекуле-мишени под действием света и/или инактивировать молекулу-мишень под действием света. В патенте США № 5567588 и патенте США № 5861254 описаны способы на основе SELEXTM, с помощью которых достигают высокоэффективного разделения олигонуклеотидов, обладающих высокой и низкой аффинностью к молекуле-мишени. В патенте США № 5496938 описаны способы получения улучшенных нуклеиновых кислот-лигандов после проведения процесса SELEXTM. В патенте США № 5705337 описаны способы ковалентного связывания лиганда с его мишенью.
Также SELEXTM можно применять для получения нуклеиновых кислот-лигандов, которые связываются более чем с одним участком молекулы-мишени, и для получения нуклеиновых кислот-лигандов, которые включают элементы, не являющиеся нуклеиновыми кислотами, которые связываются с конкретными участками мишени. SELEXTM обеспечивает способы выделения и идентификации нуклеиновых кислот-лигандов, которые связываются с любой предполагаемой мишенью, включая биомолекулы больших и небольших размеров, такие как белки, связывающие нуклеиновые кислоты, и белки, для которых не известно о наличии связывания нуклеиновых кислот, как части их биологической функции, а также кофакторы и другие низкомолекулярные соединения. Например, в патенте США № 5580737 описаны последовательности нуклеиновых кислот, идентифицированных с помощью SELEXTM, которые способны с высокой аффинностью связываться с кофеином и со значительно сходным с ним аналогом, теофиллином.
Counter-SELEXTM представляет собой способ повышения специфичности нуклеиновых кислот-лигандов в отношении молекулы-мишени посредством удаления последовательностей нуклеиновых кислот-лигандов с перекрестной реактивностью в отношении одной или нескольких молекул, не являющихся мишенями. Counter-SELEXTM включает следующие стадии: (a) получение предназначенной для анализа смеси нуклеиновых кислот; (b) контактирование предназначенной для анализа смеси с мишенью, где нуклеиновые кислоты с повышенной относительно предназначенной для анализа смеси аффинностью в отношении мишени могут быть отделены от остальной части предназначенной для анализа смеси; (c) отделение нуклеиновых кислот с повышенной аффинностью от остальной части предназначенной для анализа смеси; (d) диссоциация нуклеиновых кислот с повышенной аффинностью и мишени; (e) контактирование нуклеиновых кислот с повышенной аффинностью с одной или несколькими молекулами, не являющимися мишенями, для удаления нуклеиновых кислот-лигандов с определенной аффинностью к молекуле(ам), не являющейся мишенью(ями); и (f) амплификация нуклеиновых кислот с определенной аффинностью только к молекуле-мишени с получением смеси нуклеиновых кислот, обогащенных последовательностями нуклеиновых кислот с относительно более высокой аффинностью и специфичностью в отношении связывания с молекулой-мишенью. Как описано выше для SELEXTM, циклы отбора и амплификации при необходимости повторяют до тех пор, пока не достигнут желаемой цели.
Одной потенциальной проблемой, встречающейся при применении лекарственных средств и вакцин на основе нуклеиновых кислот, является то, что олигонуклеотиды в фосфодиэфирной форме могут быстро деградироваться в жидкостях организма внутриклеточными и внеклеточными ферментами, такими как эндонуклеазы и экзонуклеазы, до проявления требуемого эффекта. Таким образом, способ SELEXTM включает идентификацию нуклеиновых кислот-лигандов с высокой аффинностью, содержащих модифицированные нуклеотиды, придающие улучшенные свойства лиганду, такие как увеличенная стабильность in vivo, или улучшенные свойства, касающиеся доставки. Примеры таких модификаций включают химическое замещение в положениях сахара, и/или фосфата, и/или основания. Идентифицированные посредством SELEXTM нуклеиновые кислоты-лиганды, содержащие модифицированные нуклеотиды, описаны, например, в патенте США № 5660985, в котором описаны олигонуклеотиды, содержащие производные нуклеотидов, химически модифицированные по 2' положению рибозы, 5 положению пиримидинов и 8 положению пуринов, в патенте США № 5756703, в котором описаны олигонуклеотиды, содержащие различные 2'-модифицированные пиримидины, и в патенте США № 5580737, в котором описаны высокоспецифичные нуклеиновые кислоты-лиганды, содержащие один или несколько нуклеотидов, модифицированных с помощью заместителей 2'-амино (2'-NH2), 2'-фтор (2'-F) и/или 2'-O-метил (2'-OMe).
Модификации нуклеиновых кислот-лигандов, предусмотренные для этого изобретения, включают, но не ограничиваются ими, модификации, которые предоставляют другие химические группы, которые обеспечивают дополнительный заряд, поляризуемость, гидрофобность, образование водородных связей, электростатические взаимодействия и гибкость оснований нуклеиновой кислоты-лиганда или нуклеиновой кислоты-лиганда в целом. Модификации с целью получения наборов олигонуклеотидов, которые являются устойчивыми к нуклеазам, также могут включать одну или несколько замещенных межнуклеотидных связей, измененных сахаров, измененных оснований или их сочетание. Такие модификации включают, но не ограничиваются ими, модификации сахара во 2' положении, модификации пиримидина в 5 положении, модификации пурина в 8 положении, модификации экзоциклических аминов, замещение 4-тиоуридина, замещение 5-бром- или 5-йодурацила, модификации остова, модификации фосфоротиоатом или алкилфосфатом, метилирование и необычные сочетания при спаривании оснований, такие как в случае изооснований изоцитидина и изогуанидина. Также модификации могут включать 3'- и 5'-модификации, такие как кэпирование, например добавление 3'-3'-dT кэпа с целью повышения устойчивости к экзонуклеазам (см., например, патенты США №№ 5674685; 5668264; 6207816 и 6229002, все из которых включены в настоящий документ в качестве ссылок в полном объеме).
В одном варианте осуществления предусмотрены олигонуклеотиды, в которых группа P(O)O замещена P(O)S ("тиоатом"), P(S)S ("дитиоатом"), P(O)NR2 ("амидатом"), P(O)R, P(O)OR', CO или CH2 ("формацеталем") или 3'-амином (-NH-CH2-CH2-), где каждый из R или R' независимо представляет собой H или замещенный или незамещенный алкил. Связывающие группы могут быть присоединены к соседним нуклеотидам посредством связи -O-, -N-, или -S-. Необязательно, чтобы все связи в олигонуклеотиде были идентичными.
В дополнительных вариантах осуществления олигонуклеотиды содержат модифицированные группы сахаров, например, одна или несколько гидроксильных групп замещены галогеном, алифатическими группами, или функционализированы в качестве простых эфиров или аминов. В одном варианте осуществления 2' положение остатка фуранозы замещено любым из O-метила, O-алкила, O-аллила, S-алкила, S-аллила или группы гало. Способы синтеза 2'-модифицированных сахаров описаны, например, в Sproat, et al., Nucl. Acid Res. 19:733-738 (1991); Gotten, et al., Nucl. Acid Res. 19:2629-2635 (1991); и Hobbs, et al., Biochemistry 12:5138-5145 (1973). Другие модификации известны среднему специалисту в данной области. Такие модификации могут представлять собой модификации, предшествующие процессу SELEXTM или модификации после процесса SELEXTM (модификации ранее идентифицированных немодифицированных лигандов) или могут быть проведены посредством включения в процесс SELEXTM.
Модификации, предшествующие процессу SELEXTM, или модификации, проведенные посредством включения в процесс SELEXTM, приводят к получению нуклеиновых кислот-лигандов как со специфичностью в отношении их мишени в SELEXTM, так и с повышенной стабильностью, например стабильностью in vivo. Модификации после процесса SELEXTM, проведенные в отношении нуклеиновых кислот-лигандов, могут приводить к повышенной стабильности, например стабильности in vivo, без отрицательного влияния на связывающую способность нуклеиновой кислоты-лиганда.
Способ SELEXTM включает комбинирование отобранных олигонуклеотидов с другими отобранными олигонуклеотидами и неолигонуклеотидными функциональными элементами, как описано в патенте США № 5637459 и в патенте США № 5683867. Кроме того, способ SELEXTM включает комбинирование отобранных нуклеиновых кислот-лигандов с липофильными или неиммуногенными высокомолекулярными соединениями в диагностический или терапевтический комплекс, как описано, например, в патенте США № 6011020, патенте США № 6051698 и публикации PCT WO 98/18480. В этих патентах и заявках описано сочетание широкого ряда форм и других свойств со свойствами эффективной амплификации и воспроизведения олигонуклеотидов и с требуемыми свойствами других молекул.
Также разработан способ идентификации с помощью способа SELEXTM нуклеиновых кислот-лигандов для небольших гибких пептидов. Небольшие пептиды обладают подвижной структурой и, как правило, в растворе находятся в виде множества конформеров в равновесном состоянии, и, таким образом, изначально полагали, что аффинность связывания может быть ограничена потерей конформационной энтропии при связывании с гибким пептидом. Однако применимость идентифицированных нуклеиновых кислот-лигандов в отношении небольших пептидов в растворе была показана в патенте США № 5648214. В этом патенте описана идентификация высокоаффинных РНК-нуклеиновых кислот-лигандов к веществу P, пептиду из 11 аминокислот.
Аптамеры со специфичностью и аффинностью связывания в отношении мишени(ей) по настоящему изобретению, как правило, отбирают с помощью процесса SELEXTM, как описано в настоящем документе. Кроме того, селекцию можно проводить для последовательностей, включающих модифицированные нуклеотиды для стабилизации молекулы аптамера против деградации in vivo.
SELEX TM с 2'-модификацией
Для того чтобы аптамер был пригодным для применения в качестве лекарственного средства или диагностического средства, он должен быть безопасным и стабильным in vivo, и его синтез не должен быть дорогостоящим. РНК- и ДНК-аптамеры дикого типа, как правило, не являются стабильными in vivo, вследствие того, что они подвержены деградации нуклеазами. Если желательно, устойчивость к деградации нуклеазами можно значительно повышать встраиванием модифицирующих групп во 2' положение.
Группы 2'-фтор и 2'-амино успешно встраивали в олигонуклеотидные библиотеки, из которых затем отбирали аптамеры. Однако эти модификации значительно повышают стоимость синтеза конечного аптамера, и они в некоторых случаях могут привести к проблеме безопасности вследствие вероятности того, что модифицированные нуклеотиды могут повторно использоваться в ДНК хозяина при деградации модифицированных олигонуклеотидов и последующем применении нуклеотидов в качестве субстратов для синтеза ДНК.
Аптамеры, которые содержат нуклеотиды с 2'-O-метилом ("2'-OMe"), как представлено в настоящем документе в некоторых вариантах осуществления, устраняют множество из этих недостатков. Олигонуклеотиды, содержащие 2'-OMe-нуклеотиды, являются устойчивыми к воздействию нуклеаз, и их синтез не является дорогостоящим. Несмотря на то, что 2'-OMe-нуклеотиды являются распространенными в биологических системах, в физиологических условиях природные полимеразы не распознают 2'-OMe-NTP в качестве субстратов, таким образом, не существует проблем безопасности в отношении повторного использования 2'-OMe-нуклеотидов в ДНК хозяина. Способы SELEXTM, используемые для получения 2'-модифицированных аптамеров, описаны, например, в условной патентной заявке США с серийным № 60/430761, поданной 3 декабря 2002 года, в условной патентной заявке США с серийным № 60/487474, поданной 15 июля 2003 года, в условной патентной заявке США с серийным № 60/517039, поданной 4 ноября 2003 года, в патентной заявке США № 10/729581, поданной 3 декабря 2003 года, в патентной заявке США № 10/873856, поданной 21 июня 2004 года, с названием "Method for in vitro Selection of 2'-OMe Substituted Nucleic acids", и в условной патентной заявке США с серийным № 60/696295, поданной 30 июня 2005 года, с названием "Improved Materials and Methods for the Generation of Fully 2'-Modified Containing Nucleic Acid Transcripts", все из которых включены в настоящий документ в качестве ссылок в полном объеме.
Настоящее изобретение относится к аптамерам, связывающимся с комплементом и модулирующим его функцию, которые содержат модифицированные нуклеотиды (например, нуклеотиды с модификацией в положении 2') для получения олигонуклеотида, более устойчивого к ферментативной и химической деградации, а также термической и физической деградации, чем немодифицированный олигонуклеотид. В предпочтительном варианте осуществления, указанный компонент комплемента-мишень представляет собой белок C5 комплемента. Несмотря на то, что в литературе существуют различные примеры аптамеров, содержащих 2'-OMe (см., например, Green et al., Current Biology 2, 683-695, 1995), они были получены посредством селекции in vitro из библиотек модифицированных транскриптов, в которых остатки C и U являлись 2'-фтор(2'-F)-замещенными, и остатки A и G представляли собой остатки с 2'-OH. После идентификации функциональных последовательностей каждый остаток A и G анализировали на возможность 2'-OMe замещения, и аптамер повторно синтезировали, со всеми остатками A и G, в которых было допустимо 2'-OMe замещение, в виде остатков с 2'-OMe. В большинстве остатков аптамеров A и G, полученных таким двухстадийным способом, является допустимым замещение остатками с 2'-OMe, хотя, в среднем, приблизительно в 20% остатков замещение не допустимо. Таким образом, аптамеры, полученные с использованием этого способа, предполагают наличие от двух до четырех остатков 2'-OH, и стабильность и стоимость синтеза в результате уравновешиваются. Вследствие включения модифицированных нуклеотидов в реакцию транскрипции, в результате которой образуются стабилизированные олигонуклеотиды, используемые в пулах олигонуклеотидов, из которых аптамеры отбирают и обогащают посредством SELEXTM (и/или любого из его вариантов и усовершенствований, включая варианты и усовершенствования, описанные в настоящем документе), способы по настоящему изобретению устраняют необходимость в стабилизации отобранных олигонуклеотидов аптамеров (например, посредством повторного синтеза олигонуклеотидов аптамеров с модифицированными нуклеотидами).
В одном варианте осуществления, настоящее изобретение относится к аптамерам, содержащим сочетания модификаций 2'-OH, 2'-F, 2'-дезокси и 2'-OMe нуклеотидов ATP, GTP, CTP, TTP и UTP. В другом варианте осуществления настоящее изобретение относится к аптамерам, содержащим сочетания модификаций 2'-OH, 2'-F, 2'-дезокси, 2'-OMe, 2'-NH2 и 2'-метоксиэтилом нуклеотидов ATP, GTP, CTP, TTP и UTP. В другом варианте осуществления настоящее изобретение относится к аптамерам, содержащим 56 сочетаний модификаций 2'-OH, 2'-F, 2'-дезокси, 2'-OMe, 2'-NH2 и 2'-метоксиэтилом нуклеотидов ATP, GTP, CTP, TTP и UTP. В предпочтительном варианте осуществления настоящее изобретение относится к аптамерам, содержащим все или по существу все модифицированные 2'-OMe нуклеотидов ATP, GTP, CTP, TTP и/или UTP.
2'-Модифицированные аптамеры по этому изобретению получают с использованием модифицированных полимераз, например модифицированной полимеразы T7 с частотой встраивания модифицированных нуклеотидов с объемными заместителями в 2' положении фуранозы, более высокой, чем частота полимеразы дикого типа. Например, мутантная T7 полимераза, в которой остаток тирозина в положении 639 заменен фенилаланином (Y639F), легко использует (включает) 2'дезокси-, 2'амино- и 2'фторнуклеотидтрифосфаты (NTP), но не NTP с объемными 2'-заместителями, такими как заместители 2'-OMe или 2'-азидо (2'-N3). Для встраивания объемных 2'-заместителей описана мутантная полимераза T7 с заменой гистидина в 784 положении остатком аланина в дополнение к мутации Y639F (Y639F/H784A), и ее используют в определенных случаях для встраивания модифицированных пиримидиновых NTP. См. Padilla, R. and Sousa, R., Nucleic acids Res., 2002, 30(24): 138. Мутантную РНК-полимеразу T7, в которой остаток тирозина в положении 639 заменен фенилаланином, остаток гистидина в 784 положении заменен аланином и остаток лизина в положении 378 заменен аргинином (Y639F/H784A/K378R), используют в определенных случаях для встраивания модифицированных пуриновых и пиримидиновых NTP, например 2'-OMe NTP, но для транскрипции она требует добавления 2'-OH GTP. См. Burmeister et al., Chemistry and Biology, 2005, 12: 25-33. Без связи с какой-либо теорией, мутация K378R находится не вблизи активного центра полимеразы, и, таким образом, полагают, что она является молчащей мутацией, не оказывающей эффекта на включение модифицированных посредством 2'-OMe NTP. Также описана мутантная полимераза T7 с заменой гистидина в 784 положении остатком аланина (H784A). Padilla et al., Nucleic acids Research, 2002, 30: 138. Как в мутантной полимерезе T7 Y639F/H784A, так и в мутантной полимеразе T7 H784A замена аминокислотным остатком меньшего размера, таким как аланин, делает возможным встраивание более объемных нуклеотидных субстратов, например, 2'-O-Me-замещенных нуклеотидов. См. Chelliserry, K. and Ellington, A.D., (2004) Nature Biotech, 9:1155-60. Были описаны дополнительные РНК-полимеразы T7 с мутациями в активном центре РНК-полимеразы T7, которые более легко встраивают объемные 2'-модифицированные субстраты, например мутантная РНК-полимераза T7 с заменой остатка тирозина в 639 положении лейцином (Y639L).
Главным образом, было выявлено, что в условиях, описанных в настоящем документе, мутантную форму Y693F можно использовать для встраивания всех 2'-OMe-замещенных NTP за исключением GTP, и мутантные РНК-полимеразы T7 Y639F/H784A, Y639F/H784A/K378R, Y639L/H784A, Y639L/H784A/K378R, Y639L, Y639L/K378R, P266L/Y639L/H784A или P266L/Y639L/H784A/K378R в условиях, описанных в настоящем документе, можно использовать для встраивания всех 2'-OMe замещенных NTP, включая 2'-OMe GTP.
2'-Модифицированные олигонуклеотиды можно синтезировать полностью из модифицированных нуклеотидов или частично из модифицированных нуклеотидов. Модификации могут быть одинаковыми или могут отличаться. Модифицированными могут быть все или некоторые нуклеотиды, и те нуклеотиды, которые модифицированы, могут содержать одну модификацию. Модифицированными могут быть все или некоторые нуклеотиды, и те нуклеотиды, которые модифицированы, могут содержать различные модификации, например, все нуклеотиды, содержащие одно основание, могут обладать одним типом модификации, в то время как нуклеотиды, содержащие другие основания, могут обладать другими типами модификаций. Все пуриновые нуклеотиды могут обладать одним типом модификации (или могут быть немодифицированными), в то время все пиримидиновые нуклеотиды могут обладать другим, отличающимся типом модификации (или могут быть немодифицированными). Таким образом, транскрипты или библиотеки транскриптов получают с использованием любого сочетания модификаций, включая, например, рибонуклеотиды (2'-OH), дезоксирибонуклеотиды (2'-дезокси), 2'-аминонуклеотиды (2'-NH2), 2'-фторнуклеотиды (2'-F) и 2'-O-метилнуклетоиды (2'-OMe). Транскрипционную смесь, содержащую 2'-OH A и G и 2'-F C и U, называют смесью "rRfY", и отобранные из нее аптамеры называют аптамерами "rRfY". Транскрипционную смесь, содержащую 2'-OMe C и U и 2'-OH A и G, называют смесью "rRmY", и отобранные из нее аптамеры называют аптамерами "rRmY". Транскрипционную смесь, содержащую дезокси A и G и 2'-OMe U и C, называют смесью "dRmY", и отобранные из нее аптамеры называют аптамерами "dRmY". Транскрипционную смесь, содержащую 2'-OMe A, C и U, и 2'-OH G, называют смесью "rGmH", и отобранные из нее аптамеры называют аптамерами "rGmH". Транскрипционную смесь, альтернативно содержащую 2'-OMe A, C, U и G и 2'-OMe A, U и C и 2'-F G, называют "альтернативной смесью", и отобранные из нее аптамеры называют аптамерами из "альтернативной смеси". Транскрипционную смесь, содержащую 2'-OMe A, U, C и G, где вплоть до 10% G составляют рибонуклеотиды, называют смесью "r/mGmH", и отобранные из нее аптамеры называют аптамерами "r/mGmH". Транскрипционную смесь, содержащую 2'-OMe A, U и C, и 2'-F G, называют смесью "fGmH", и отобранные из нее аптамеры называют аптамерами "fGmH". Транскрипционную смесь, содержащую 2'-OMe A, U и C, и дезокси G, называют смесью "dGmH", и отобранные из нее аптамеры называют аптамерами "dGmH". Транскрипционную смесь, содержащую дезокси A, и 2'-OMe C, G и U, называют смесью "dAmB", и отобранные из нее аптамеры называют аптамерами "dAmB", и транскрипционную смесь, содержащую все нуклеотиды 2'-OH, называют смесью "rN", и отобранные из нее аптамеры называют аптамерами "rN", "rRrY" или "RNA". Транскрипционную смесь, содержащую 2'-OH аденозинтрифосфат и гуанозинтрифосфат и дезоксицитидинтрифосфат и тимидинтрифосфат, называют смесью rRdY, и аптамеры, отобранные из нее, называют аптамерами "rRdY". Аптамер "mRmY", также называемый аптамером "MNA", представляет собой аптамер, содержащий только 2'-OMe нуклеотиды за исключением исходного нуклеотида, который представляет собой 2'-OH гуанозин или гуанозин дикого типа, и его можно получать из олигонуклеотида r/mGmH посредством замены после SELEXTM, когда это возможно, или заменой любых 2'-OH G посредством 2'-OMe-G. Альтернативно, аптамеры mRmY можно идентифицировать посредством SELEXTM mRmY.
Предпочтительный вариант осуществления включает любое сочетание 2'-OH, 2'-дезокси и 2'-OMe нуклеотидов. Более предпочтительный вариант осуществления включает любое сочетание 2'-дезокси и 2'-OMe нуклеотидов. Более предпочтительный вариант осуществления включает сочетание 2'-дезокси и 2'-OMe нуклеотидов, где пиримидины являются 2'-OMe (как в случае dRmY, mRmY или dGmH).
Встраивание модифицированных нуклеотидов в аптамеры по этому изобретению проводят до (перед) процесса селекции (например, модификация перед процессом SELEXTM). Необязательно, аптамеры по этому изобретению, в которых модифицированные нуклеотиды встроены модификацией перед процессом SELEXTM, можно дополнительно модифицировать в процессе модификации после процесса SELEXTM (т.е. модификация после процесса SELEXTM после проведения процесса SELEXTM). Модификации перед процессом SELEXTM приводят к модифицированным нуклеиновым кислотам-лигандам со специфичностью в отношении мишени SELEXTM, а также к улучшенной стабильности in vivo. Модификации после процесса SELEXTM (например, укорочение, делеция, замещение или дополнительные модификации нуклеотидов ранее идентифицированных лигандов, обладающих нуклеотидами, встроенными посредством модификации перед процессом SELEXTM) могут приводить к дополнительному повышению стабильности in vivo без отрицательного влияния на связывающую способность нуклеиновой кислоты-лиганда, имеющей нуклеотиды, встроенные посредством модификации перед процессом SELEXTM.
Для получения пулов 2'-модифицированных (например, 2'-OMe) РНК-транскриптов в условиях, при которых полимераза использует 2'-модифицированные NTP, можно использовать любые из мутантных РНК-полимераз T7 Y693F, Y693F/K378R, Y693F/H784A, Y693F/H784A/K378R, Y693L/H784A, Y693L/H784A/K378R, Y639L, Y639L/K378R, P266L/Y693F/H784A и P266L/Y693F/H784A/K378R. Также в настоящем изобретении можно использовать другие РНК-полимеразы T7, в частности, РНК-полимеразы T7, которые проявляют высокую толерантность в отношении объемных 2'-заместителей. При использовании в управляемой матрицей полимеризации с использованием описанных условий, описанных в настоящем документе, мутантную РНК-полимеразу T7 Y639L/H784A, Y639L/H784A/K378R, P266L/Y693F/H784A или P266L/Y693F/H784A/K378R можно использовать для встраивания всех 2'-OMe NTP, включая 2'-OMe GTP, с более высоким выходом транскрипта, чем достигают с использованием мутантых РНК-полимераз T7 Y639F, Y639F/K378R, Y639F/H784A, Y639F/H784A/K378R, Y639L, Y639L/K378R. Можно использовать мутантные РНК-полимеразы T7 Y639L/H784A, Y639L/H784A/K378R, P266L/Y693F/H784A и P266L/Y693F/H784A/K378R с 2'-OH GTP, для которых не требуется 2'-OH GTP, для достижения высокого выхода 2'-модифицированных, например, содержащих 2'-OMe олигонуклеотидов.
В настоящем изобретении можно использовать другие полимеразы, в частности, полимеразы, которые проявляют высокую толерантность в отношении объемных 2'-заместителей. Такие полимеразы можно подвергать скринингу в отношении этой способности посредством оценки их способности включать модифицированные нуклеотиды в условиях транскрипции, описанных в настоящем документе.
Был выявлен ряд факторов, которые являются важными для условий транскрипции, пригодных в способах, описанных в настоящем документе. Например, лидерная последовательность, включенная в фиксированную последовательность на 5'-конце транскрипционной матрицы ДНК, может быть важна для повышения выхода модифицированных транскриптов, когда для транскрипции используют мутантные РНК-полимеразы T7 Y639F/K378R или Y639F/H784A/K378R, например, в условиях транскрипции dRmY или r/mGmH, описанных ниже. Кроме того, лидерную последовательность можно применять, но не обязательно, для способствования повышению выхода модифицированных транскриптов, например, когда для транскрипции используют мутантную РНК-полимеразу T7 Y639L/H784A или Y639L/H784A/K378R, например, в условиях транскрипции mRmY, описанных ниже. Длина лидерной последовательности, как правило, составляет 6-15 нуклеотидов, и она может состоять только из пуринов или из смеси пуриновых и пиримидиновых нуклеотидов.
Другим важным фактором при получении транскриптов, включающих модифицированные нуклеотиды, является наличие или концентрация 2'-OH-гуанозина (например, GMP, GTP или другого не 2'-OMe не трифосфата). Транскрипцию можно разделить на две фазы: первая фаза представляет собой инициацию, в процессе которой NTP добавляется к 3'-гидроксильному концу GTP (или GMP, или другого не 2'-OMe не трифосфата) с получением динуклеотида, который затем удлиняется приблизительно на 10-12 нуклеотидов; вторая фаза представляет собой элонгацию, в процессе которой после вставки первых приблизительно 10-12 нуклеотидов продолжается транскрипция. Было выявлено, что небольшие количества 2'-OH GTP (или GMP, или другого не 2'-OMe не трифосфата), добавляемые в транскрипционную смесь, содержащую избыток 2'-OMe GTP, являются достаточными для обеспечения инициации транскрипции полимеразой с использованием 2'-OH GTP (или GMP, гуанозина, или другого не 2'-OMe не трифосфата). Таким образом, например, транскрипционная смесь dRmY (содержащая дезоксипурины и 2'OMe-пиримидины) требует добавления небольшого количества GMP для обеспечения инициации транскрипции полимеразой, в то время как в случае транскрипционной смеси r/mGmH (содержащей вплоть до 10% 2'-OH GTP), в транскрипционную смесь можно добавлять небольшое количество GMP, но он не требуется для обеспечения инициации транскрипции полимеразой, поскольку 2'-OH GTP уже находится в транскрипционной смеси. После вхождения транскрипции в фазу элонгации, сниженная дифференциация между 2'-OMe и 2'-OH GTP, и избыток 2'-OMe GTP по сравнению с 2'-OH GTP обеспечивают включение, главным образом, 2'-OMe GTP.
Как описано непосредственно выше, запуск транскрипции 2'-OH-гуанозином (например, GMP, GTP или другим не 2'-OMe не-трифосфатом) является важным. Этот эффект возникает вследствие специфичности полимеразы в отношении инициирующего нуклеотида. В результате, 5'-концевой нуклеотид транскрипта, полученного таким образом, вероятно будет представлять собой 2'-OH G. Предпочтительной концентрацией GMP является 0,5 мМ и более предпочтительно 1 мМ. Было выявлено, что включение PEG, предпочтительно PEG-8000, в реакцию транскрипции является пригодным для максимального повышения включения модифицированных нуклеотидов.
Другим важным фактором при встраивании 2'-OMe замещенных нуклеотидов в транскрипты является использование как двухвалентного магния, так и двухвалентного марганца в транскрипционной смеси. Было выявлено, что различные сочетания концентраций хлорида магния и хлорида марганца влияют на выход 2'-O-метилированных транскриптов, при этом оптимальная концентрация хлорида магния и марганца зависит от концентрации в транскрипционной реакционной смеси NTP, которые образуют комплекс с ионами металлов. Для получения наиболее высокого выхода полностью 2'-O-метилированных транскриптов (т.е. все A, C и U нуклеотиды и приблизительно 90% нуклеотида G содержат 2'-OMe), концентрации приблизительно 5 мМ хлорида магния и 1,5 мМ хлорида марганца являются предпочтительными, если каждый NTP находится в концентрации 0,5 мМ. Если концентрация каждого NTP составляет 1,0 мМ, предпочтительными являются концентрации приблизительно 6,5 мМ магния хлорида и 2,0 мМ хлорида марганца. Если концентрация каждого NTP составляет 2,0 мМ, то предпочтительными являются концентрации приблизительно 9,6 мМ хлорида магния и 2,9 мМ хлорида марганца. В любом случае, отклонение от этих концентраций вплоть до двух раз, тем не менее, обеспечивают получение значительных количеств модифицированных транскриптов.
В некоторых вариантах осуществления является преимущественным запуск транскрипции посредством GMP или гуанозина. Этот эффект возникает вследствие специфичности полимеразы в отношении инициирующего нуклеотида. В результате 5'-концевой нуклеотид любого транскрипта, полученного таким способом, вероятно, окажется 2'-OH G. Предпочтительная концентрация GMP (или гуанозина) составляет 0,5 мМ и более предпочтительно 1 мМ. Также было выявлено, что добавление в реакцию транскрипции PEG, предпочтительно PEG-8000, является полезным для максимального встраивания модифицированных нуклеотидов.
Для максимального встраивания 2'-OMe ATP (100%), UTP (100%), CTP (100%) и GTP (~90%) ("r/mGmH") в транскрипты, предпочтительными являются следующие условия: 200 мМ буфер HEPES, 40 мМ DTT, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 5 мМ MgCl2 (6,5 мМ, где концентрация каждого 2'-OMe NTP составляет 1,0 мМ), 1,5 мМ MnCl2 (2,0 мМ, где концентрация каждого 2'-OMe NTP составляет 1,0 мМ), 500 мкМ 2'-OMe NTP (каждый) (более предпочтительно, 1,0 мМ), 30 мкМ 2'-OH GTP, 500 мкМ 2'-OH GMP, pH 7,5, 200 нМ РНК-полимераза T7 Y639F/H784A, 5 единиц/мл неорганической пирофосфатазы, и полностью состоящая из пуринов лидерная последовательность длиной по меньшей мере 8 нуклеотидов. Как используют в настоящем документе, одну единицу мутантной РНК-полимеразы T7 Y639F/H784A (или любой другой мутантной РНК-полимеразы T7, описанной в настоящем документе) определяют как количество фермента, необходимое для встраивания 1 нмоль 2'-OMe NTP в транскрипт в условиях r/mGmH. Как используют в настоящем документе, одна единица неорганической пирофосфатазы определяется как количество фермента, которое обеспечивает высвобождение 1,0 моль неорганического ортофосфата в минуту при pH 7,2 и 25°C.
Для максимального встраивания (100%) 2'-OH GTP и 2'-OMe ATP, UTP и CTP ("rGmH") в транскрипты предпочтительными являются следующие условия: буфер 200 мМ HEPES, 40 мМ DTT, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 5 мМ MgCl2 (9,6 мМ, где концентрация каждого NTP составляет 2,0 мМ), 1,5 мМ MnCl2 (2,9 мМ, где концентрация каждого NTP составляет 2,0 мМ), 500 мкМ NTP (каждый) (более предпочтительно, 2,0 мМ), 1 мМ 2'-OH GMP, pH 7,5, 200 нМ РНК-полимераза T7 Y639F/K378R, 5 единиц/мл неорганической пирофосфатазы, и полностью состоящая из пуринов лидерная последовательность длиной по меньшей мере 8 нуклеотидов.
Для максимального встраивания в транскрипты 2'-OMe ATP (100%), 2'-OMe UTP (100%), 2'-OMe CTP (100%) и 2'-OMe GTP (100%) ("mRmY" или "MNA") предпочтительными являются следующие условия: 200 мМ буфер HEPES, 40 мМ DTT, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 8 мМ MgCl2, 2,5 мМ MnCl2, 1,5 мМ 2'-OMe NTP (каждый), 1 мМ 2'-OH GMP, pH 7,5, 200 нМ мутантная РНК-полимераза T7 Y639L/H784A/K378R, 5 единиц/мл неорганической пирофосфатазы, и лидерная последовательность, которая повышает выход транскрипции в установленных условиях транскрипции. В одном варианте осуществления, необязательная лидерная последовательность представляет собой лидерную последовательность, полностью состоящую из пуринов. В другом варианте осуществления, необязательная лидерная последовательность представляет собой смесь пуринов и пиримидинов.
Для максимального встраивания (100%) 2'-OH ATP и GTP и 2'-OMe UTP и CTP ("rRmY") в транскрипты предпочтительными являются следующие условия: 200 мМ буфер HEPES, 40 мМ DTT, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 5 мМ MgCl2 (9,6 мМ, где концентрация каждого NTP составляет 2,0 мМ), 1,5 мМ MnCl2 (2,9 мМ, где концентрация каждого NTP составляет 2,0 мМ), 500 мкМ NTP (каждый) (более предпочтительно, 2,0 мМ), 1 мМ 2'-OH GMP, pH 7,5, 200 нМ РНК-полимераза T7 Y639L/H784A/K378R, 5 единиц/мл неорганической пирофосфатазы и лидерная последовательность, полностью состоящая из пуринов, длиной по меньшей мере 8 нуклеотидов.
Для максимального встраивания (100%) в транскрипты 2'-OMe ATP, UTP и CTP ("rGmH"), предпочтительными являются следующие условия: 200 мМ буфер HEPES, 40 мМ DTT, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 5 мМ MgCl2 (9,6 мМ, где концентрация каждого 2'-OMe NTP составляет 2,0 мМ), 1,5 мМ MnCl2 (2,9 мМ, где концентрация каждого 2'-OMe NTP составляет 2,0 мМ), 500 мкМ 2'-OMe NTP (каждый) (более предпочтительно, 2,0 мМ), pH 7,5, 15 единиц/мл РНК-полимеразы T7 Y639F, 5 единиц/мл неорганической пирофосфатазы, и лидерная последовательность, полностью состоящая из пуринов, длиной по меньшей мере 8 нуклеотидов.
Для максимального встраивания (100%) 2'-OMe UTP и CTP ("rRmY") в транскрипты предпочтительными являются следующие условия: 200 мМ буфер HEPES, 40 мМ DTT, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 5 мМ MgCl2 (9,6 мМ, где концентрация каждого 2'-OMe NTP составляет 2,0 мМ), 1,5 мМ MnCl2 (2,9 мМ, где концентрация каждого 2'-OMe NTP составляет 2,0 мМ), 500 мкМ 2'-OMe NTP (каждый) (более предпочтительно, 2,0 мМ), pH 7,5, 15 единиц/мл РНК-полимеразы T7 Y639L/H784A, 5 единиц/мл неорганической пирофосфатазы и лидерная последовательность, полностью состоящая из пуринов, длиной по меньшей мере 8 нуклеотидов.
Для максимального встраивания (100%) дезокси ATP и GTP и 2'-OMe UTP и CTP ("dRmY") в транскрипты предпочтительными являются следующие условия: 200 мМ буфер HEPES, 40 мМ DTT, 2 мМ спермин, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 9,6 мМ MgCl2, 2,9 мМ MnCl2, 2,0 мМ NTP (каждый), 1 мМ 2'-OH GMP, pH 7,5, 200 нМ РНК-полимераза T7 Y639F/K378R, 5 единиц/мл неорганической пирофосфатазы, и лидерная последовательность, полностью состоящая из пуринов, длиной по меньшей мере 8 нуклеотидов.
Для максимального встраивания (100%) 2'-OMe ATP, UTP и CTP и 2'-F GTP ("fGmH") в транскрипты предпочтительными являются следующие условия: 200 мМ буфер HEPES, 40 мМ DTT, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 9,6 мМ MgCl2, 2,9 мМ MnCl2, 2,0 мМ 2'-OMe NTP (каждый), 1 мМ 2'-OH GMP, pH 7,5, 200 нМ РНК-полимераза T7 Y639F/K378R, 5 единиц/мл неорганической пирофосфатазы, и лидерная последовательность, полностью состоящая из пуринов длиной по меньшей мере 8 нуклеотидов.
Для максимального встраивания (100%) дезокси ATP и 2'-OMe UTP, GTP и CTP ("dAmB") в транскрипты предпочтительными являются следующие условия: 200 мМ буфер HEPES, 40 мМ DTT, 2 мМ спермидин, 10% PEG-8000 (мас./об.), 0,01% Triton X-100 (мас./об.), 9,6 мМ MgCl2, 2,9 мМ MnCl2, 2,0 мМ NTP (каждый), 1 мМ 2'-OH GMP, pH 7,5, 200 нМ РНК-полимераза T7 Y639F/K378R, 5 единиц/мл неорганической пирофосфатазы, и лидерная последовательность, полностью состоящая из пуринов, длиной по меньшей мере 8 нуклеотидов.
Для каждого из указанных выше, (a) транскрипцию предпочтительно проводят при температуре от приблизительно 20°C до приблизительно 50°C, предпочтительно от приблизительно 30°C до 45°C и более предпочтительно при приблизительно 37°C в течение по меньшей мере двух часов и (b) используют 50-300 нМ двухцепочечную ДНК-матрицу для транскрипции (200 нМ матрицу используют в цикле 1 для повышения разнообразия (300 нМ матрицу используют в транскрипциях dRmY), и для последующих циклов используют приблизительно 50 нМ, разбавление 1/10 оптимизированной реакционной смеси ПЦР, с использованием условий, описанных в настоящем документе). Предпочтительные ДНК-матрицы для транскрипции описаны ниже (где ARC254 и ARC256 транскрибируются при всех условиях для 2'-OMe и ARC255 транскрибируется при условиях rRmY).

В условиях транскрипции rN по настоящему изобретению, транскрипционная реакционная смесь содержит 2'-OH аденозинтрифосфаты (ATP), 2'-OH гуанозинтрифосфаты (GTP), 2'-OH цитидинтрифосфаты (CTP) и 2'-OH уридинтрифосфаты (UTP). Модифицированные олигонуклеотиды, полученные с использованием транскрипционных смесей rN по настоящему изобретению, содержат по существу только 2'-OH-аденозин, 2'-OH-гуанозин, 2'-OH-цитидин и 2'-OH-уридин. В предпочтительном варианте осуществления транскрипции rN, полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 80% всех нуклеотидов аденозина представляют собой 2'-OH-аденозин, по меньшей мере 80% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, по меньшей мере 80% всех нуклеотидов цитидина представляют собой 2'-OH-цитидин, и по меньшей мере 80% всех нуклеотидов уридина представляют собой 2'-OH-уридин. В более предпочтительном варианте осуществления транскрипции rN, полученные модифицированные олигонуклеотиды по настоящему изобретению содержат последовательность, где по меньшей мере 90% всех нуклеотидов аденозина представляют собой 2'-OH-аденозин, по меньшей мере 90% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, по меньшей мере 90% всех нуклеотидов цитидина представляют собой 2'-OH-цитидин, и по меньшей мере 90% всех нуклеотидов уридина представляют собой 2'-OH-уридин. В наиболее предпочтительном варианте осуществления транскрипции rN модифицированные олигонуклеотиды по настоящему изобретению содержат последовательность, где 100% всех нуклеотидов аденозина представляют собой 2'-OH-аденозин, 100% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, 100% всех нуклеотидов цитидина представляют собой 2'-OH-цитидин, и 100% всех нуклеотидов уридина представляют собой 2'-OH-уридин.
При условиях транскрипции rRmY по настоящему изобретению, транскрипционная реакционная смесь содержит 2'-OH-аденозинтрифосфаты, 2'-OH-гуанозинтрифосфаты, 2'-OMe-цитидинтрифосфаты и 2'-OMe-уридинтрифосфаты. Модифицированные олигонуклеотиды, полученные с использованием транскрипционных смесей rRmY по настоящему изобретению, содержат по существу только 2'-OH-аденозин, 2'-OH-гуанозин, 2'-OMe-цитидин и 2'-OMe-уридин. В предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 80% всех нуклеотидов аденозина представляют собой 2'-OH-аденозин, по меньшей мере 80% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, по меньшей мере 80% всех нуклеотидов цитидина представляют собой 2'-OMe-цитидин и по меньшей мере 80% всех нуклеотидов уридина представляют собой 2'-OMe-уридин. В более предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 90% всех нуклеотидов аденозина представляют собой 2'-OH-аденозин, по меньшей мере 90% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, по меньшей мере 90% всех нуклеотидов цитидина представляют собой 2'-OMe-цитидин и по меньшей мере 90% всех нуклеотидов уридина представляют собой 2'-OMe-уридин. В наиболее предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где 100% всех нуклеотидов аденозина представляют собой 2'-OH-аденозин, 100% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, 100% всех нуклеотидов цитидина представляют собой 2'-OMe-цитидин и 100% всех нуклеотидов уридина представляют собой 2'-OMe-уридин.
При условиях транскрипции dRmY по настоящему изобретению, транскрипционная реакционная смесь содержит 2'-дезоксиаденозинтрифосфаты, 2'-дезоксигуанозинтрифосфаты, 2'-O-метилцитидинтрифосфаты, и 2'-O-метилуридинтрифосфаты. Модифицированные олигонуклеотиды, полученные с использованием условий транскрипции dRmY по настоящему изобретению, содержат по существу только 2'-дезоксиаденозин, 2'-дезоксигуанозин, 2'-O- метилцитидин и 2'-O-метилуридин. В предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды по настоящему изобретению содержат последовательность, где по меньшей мере 80% всех нуклеотидов аденозина представляют собой 2'-дезоксиаденозин, по меньшей мере 80% всех нуклеотидов гуанозина представляют собой 2'-дезоксигуанозин, по меньшей мере 80% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин и по меньшей мере 80% всех нуклеотидов уридина представляют собой 2'-O-метилуридин. В более предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды по настоящему изобретению содержат последовательность, где по меньшей мере 90% всех нуклеотидов аденозина представляют собой 2'-дезоксиаденозин, по меньшей мере 90% всех нуклеотидов гуанозина представляют собой 2'-дезоксигуанозин, по меньшей мере 90% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин и по меньшей мере 90% всех нуклеотидов уридина представляют собой 2'-O-метилуридин. В наиболее предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды по настоящему изобретению содержат последовательность, где 100% всех нуклеотидов аденозина представляют собой 2'-дезоксиаденозин, 100% всех нуклеотидов гуанозина представляют собой 2'-дезоксигуанозин, 100% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, и 100% всех нуклеотидов уридина представляют собой 2'-O-метилуридин.
При условиях транскрипции rGmH по настоящему изобретению, транскрипционная реакционная смесь содержит 2'-OH гуанозинтрифосфаты, 2'-O-метилцитидинтрифосфаты, 2'-O-метилуридинтрифосфаты и 2'-O-метиладенозинтрифосфаты. Модифицированные олигонуклеотиды, полученные с использованием транскрипционных смесей rGmH по настоящему изобретению, содержат по существу только 2'-OH-гуанозин, 2'-O-метилцитидин, 2'-O-метилуридин и 2'-O-метиладенозин. В предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 80% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, по меньшей мере 80% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, по меньшей мере 80% всех нуклеотидов уридина представляют собой 2'-O-метилуридин и по меньшей мере 80% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин. В более предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 90% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, по меньшей мере 90% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, по меньшей мере 90% всех нуклеотидов уридина представляют собой 2'-O-метилуридин и по меньшей мере 90% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин. В наиболее предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где 100% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин, 100% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, 100% всех нуклеотидов уридина представляют собой 2'-O-метилуридин и 100% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин.
При условиях транскрипции r/mGmH по настоящему изобретению транскрипционная реакционная смесь содержит 2'-O-метиладенозинтрифосфат, 2'-O-метилцитидинтрифосфат, 2'-O-метилгуанозинтрифосфат, 2'-O-метилуридинтрифосфат и 2'-OH-гуанозинтрифосфат. Конечные модифицированные олигонуклеотиды, полученные с использованием транскрипционных смесей r/mGmH по настоящему изобретению, содержат по существу только 2'-O-метиладенозин, 2'-O-метилцитидин, 2'-O-метилгуанозин и 2'-O-метилуридин, где совокупность нуклеотидов гуанозина содержит максимум приблизительно 10% 2'-OH-гуанозина. В предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды r/mGmH по настоящему изобретению содержат последовательность, где по меньшей мере 80% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин, по меньшей мере 80% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, по меньшей мере 80% всех нуклеотидов гуанозина представляют собой 2'-O-метилгуанозин, по меньшей мере 80% всех нуклеотидов уридина представляют собой 2'-O-метилуридин, и не более чем приблизительно 10% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин. В более предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 90% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин, по меньшей мере 90% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, по меньшей мере 90% всех нуклеотидов гуанозина представляют собой 2'-O-метилгуанозин, по меньшей мере 90% всех нуклеотидов уридина представляют собой 2'-O-метилуридин, и не более чем приблизительно 10% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин. В наиболее предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где 100% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин, 100% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, 90% всех нуклеотидов гуанозина представляют собой 2'-O-метилгуанозин и 100% всех нуклеотидов уридина представляют собой 2'-O-метилуридин, и не более чем приблизительно 10% всех нуклеотидов гуанозина представляют собой 2'-OH-гуанозин.
При условиях транскрипции mRmY (также предпочтительно MNA) по настоящему изобретению, транскрипционная смесь содержит только 2'-O-метиладенозинтрифосфат, 2'-O-метилцитидинтрифосфат, 2'-O-метилгуанозинтрифосфат, 2'-O-метилуридинтрифосфат. Конечные модифицированные олигонуклеотиды, полученные с использованием транскрипционной смеси mRmY по настоящему изобретению содержат последовательность, где 100% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин, 100% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, 100% всех нуклеотидов гуанозина представляют собой 2'-O-метилгуанозин (исключая первый гуанозин олигонуклеотида) и 100% всех нуклеотидов уридина представляют собой 2'-O-метилуридин.
При условиях транскрипции fGmH по настоящему изобретению транскрипционная реакционная смесь содержит 2'-O-метиладенозинтрифосфаты, 2'-O-метилуридинтрифосфаты, 2'-O-метилцитидинтрифосфаты и 2'-F-гуанозинтрифосфаты. Модифицированные олигонуклеотиды, полученные с использованием условий транскрипции fGmH по настоящему изобретению, содержат по существу только 2'-O-метиладенозин, 2'-O-метилуридин, 2'-O-метилцитидин и 2'-F-гуанозин. В предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 80% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин, по меньшей мере 80% всех нуклеотидов уридина представляют собой 2'-O-метилуридин, по меньшей мере 80% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, и по меньшей мере 80% всех нуклеотидов гуанозина представляют собой 2'-F-гуанозин. В более предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 90% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин, по меньшей мере 90% всех нуклеотидов уридина представляют собой 2'-O-метилуридин, по меньшей мере 90% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, и по меньшей мере 90% всех нуклеотидов гуанозина представляют собой 2'-F-гуанозин. В наиболее предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где 100% всех нуклеотидов аденозина представляют собой 2'-O-метиладенозин, 100% всех нуклеотидов уридина представляют собой 2'-O-метилуридин, 100% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин и 100% всех нуклеотидов гуанозина представляют собой 2'-F-гуанозин.
При условиях транскрипции dAmB по настоящему изобретению транскрипционная реакционная смесь содержит фосфаты, 2'-O-метилцитидинтрифосфаты, 2'-O-метилгуанозинтрифосфаты и 2'-O-метилуридинтрифосфаты. Модифицированные олигонуклеотиды, полученные с использованием транскрипционных смесей dAmB по настоящему изобретению, содержат по существу только 2'-дезоксиаденозин, 2'-O-метилцитидин, 2'-O-метилгуанозин и 2'-O-метилуридин. В предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 80% всех нуклеотидов аденозина представляют собой 2'-дезоксиаденозин, по меньшей мере 80% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, по меньшей мере 80% всех нуклеотидов гуанозина представляют собой 2'-O-метилгуанозин и по меньшей мере 80% всех нуклеотидов уридина представляют собой 2'-O-метилуридин. В более предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды содержат последовательность, где по меньшей мере 90% всех нуклеотидов аденозина представляют собой 2'-дезоксиаденозин, по меньшей мере 90% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, по меньшей мере 90% всех нуклеотидов гуанозина представляют собой 2'-O-метилгуанозин и по меньшей мере 90% всех нуклеотидов уридина представляют собой 2'-O-метилуридин. В наиболее предпочтительном варианте осуществления полученные модифицированные олигонуклеотиды по настоящему изобретению содержат последовательность, где 100% всех нуклеотидов аденозина представляют собой 2'-дезоксиаденозин, 100% всех нуклеотидов цитидина представляют собой 2'-O-метилцитидин, 100% всех нуклеотидов гуанозина представляют собой 2'-O-метилгуанозин и 100% всех нуклеотидов уридина представляют собой 2'-O-метилуридин.
В каждом случае, продукты транскрипции затем можно использовать в процессе SELEXTM в качестве библиотеки для идентификации аптамеров и/или для определения консервативного мотива последовательностей, которые обладают специфичностью в отношении связывания данной мишени. Полученные последовательности уже являются стабилизированными, устраняя эту стадию из процесса для достижения стабилизированной последовательности аптамера, и получая в результате более высоко стабилизированный аптамер. Другим преимуществом процесса 2'-OMe SELEXTM является то, что полученные последовательности, вероятно, обладают меньшим количеством требуемых 2'-OH-нуклеотидов в последовательности, возможно не обладают ими. В случаях, если 2'-OH-нуклеотиды остаются, их можно удалять проведением модификаций после процесса SELEXTM.
Как описано ниже, более низкий, но, тем не менее, пригодный выход транскриптов, полностью включающих 2'-замещенные нуклеотиды, может быть достигнут при условиях, отличных от описанных выше оптимизированных условий. Например, отклонения от описанных выше условий транскрипции включают:
Концентрация буфера HEPES может варьировать от 0 до 1 M. Настоящее изобретение также охватывает другие буферные вещества с pKa между 5 и 10, включая, например, трис(гидроксиметил)аминометан.
Концентрация DTT может варьировать от 0 до 400 мМ. Способы по настоящему изобретению также относятся к применению других восстановителей, включая, например, меркаптоэтанол.
Концентрация спермидина и/или спермина может варьировать от 0 до 20 мМ.
Концентрация PEG-8000 может варьировать от 0 до 50% (мас./об.). Способы по настоящему изобретению также относятся к применению других гидрофильных полимеров, включая, например, PEG с другой молекулярной массой или другие полиалкиленгликоли.
Концентрация Triton X-100 может варьировать от 0 до 0,1% (мас./об.). Способы по настоящему изобретению также относятся к применению других неионных детергентов, включая, например, другие детергенты, включающие другие детергенты Triton-X.
Концентрация MgCl2 может варьировать от 0,5 нМ до 50 мМ. Концентрация MnCl2 может варьировать от 0,15 мМ до 15 мМ. Концентрации как MgCl2, так и MnCl2 должны находиться в описанных диапазонах, и в предпочтительном варианте осуществления они находятся в соотношении MgCl2:MnCl2 от приблизительно 10 до приблизительно 3, предпочтительно соотношение составляет приблизительно 3-5:1, более предпочтительно, соотношение составляет приблизительно 3-4:1.
Концентрация 2'-OMe NTP (каждого NTP) может находиться в диапазоне от 5 мкМ до 5 мМ.
Концентрация 2'-OH GTP может находиться в диапазоне от 0 мкМ до 300 мкМ. В некоторых вариантах осуществления, транскрипция происходит в отсутствие 2'-OH GTP (0 мкМ).
Концентрация 2'-OH GMP, гуанозина или другого 2'-OH G, замещенного в положении, отличном от 2'-положения сахара, может варьировать от 0 до 5 мМ, и где в некоторых вариантах осуществления, 2'-OH GTP не включен в реакцию и требуется 2'-OH GMP и он может находиться в диапазоне от 5 мкМ до 5 мМ.
Концентрация ДНК-матрицы может варьировать от 5 нМ до 5 мкМ.
Концентрация мутантной полимеразы может варьировать от 2 нМ до 20 мкМ.
Неорганическая пирофосфатаза может варьировать от 0 до 100 единиц/мл.
Значение pH может варьировать от pH 6 до pH 9. Способы по настоящему изобретению можно применять на практике со значениями pH, находящимися в диапазоне активности большинства полимераз, которые встраивают модифицированные нуклеотиды.
Реакции транскрипции можно позволять протекать в течение от приблизительно одного часа до недель, предпочтительно от приблизительно 1 до приблизительно 24 часов.
Выбранные аптамеры, обладающие наиболее высокой аффинностью и связыванием, показанными посредством биологических анализов, как описано в примерах ниже, представляют собой пригодные лекарственные средства для лечения состояний, в патогенез которых вовлечен белок C5 комплемента.
МЕДИЦИНСКАЯ ХИМИЯ АПТАМЕРОВ
После идентификации аптамеров, которые связываются с требуемой мишенью, для последующего повышения связывания и/или функциональных характеристик идентифицированных последовательностей аптамеров можно необязательно проводить несколько способов. Аптамеры, которые связываются с требуемой мишенью, идентифицированной посредством процесса SELEXTM (включая 2'-модифицированый SELEXTM), необязательно можно укорачивать с получением минимальной последовательности аптамера (также называемой в настоящем документе "минимизированной конструкцией"), обладающей требуемым связыванием и/или функциональными характеристиками. Одним таким способом является применение программ по сворачиванию и анализа последовательности (например, выравнивание последовательностей клонов, полученных при селекции, для поиска консервативных мотивов и/или ковариации) для обеспечения информации при разработке минимизированных конструкций. Также можно проводить биохимические эксперименты с использованием зондов с целью определения 5'- и 3'-границ последовательности аптамера для обеспечения информации при разработке минимизированных конструкций. Затем минимизированные конструкции можно химически синтезировать и тестировать в отношении связывания и функциональных характеристик по сравнению с неминимизированной последовательностью, из которой они могут быть образованы. Варианты последовательности аптамера, содержащей серии 5', 3' и/или внутренних делеций также можно непосредственно химически синтезировать и тестировать в отношении связывания и/или функциональных характеристик по сравнению с неминимизированной последовательностью аптамера, из которой они были образованы.
Кроме того, для исследования требований к последовательности в последовательности одного активного аптамера (т.е. аптамера, который связывается с требуемой мишенью, идентифицированный посредством процесса SELEXTM, (включая 2'-модифицированный SELEXTM), или одной минимизированной последовательности аптамера, можно использовать активированную повторную селекцию. Активированную повторную селекцию проводят с использованием синтетического, вырожденного пула, который разработан на основе одной представляющей интерес последовательности. Степень вырожденности, как правило, варьирует от 70% до 85% относительно нуклеотидов дикого типа, т.е. отдельной представляющей интерес последовательности. Как правило, с помощью активированной повторной селекции выявляют нейтральные мутации, однако в некоторых случаях изменения последовательности могут привести к повышению аффинности. Затем суммарную информацию о последовательности из клонов, идентифицированных с использованием активированной повторной селекции, можно использовать для идентификации минимального связывающего мотива и способствования попыткам оптимизации.
Последовательности аптамеров, идентифицированные с использованием процесса SELEXTM, (включая 2'-модифицированный SELEX и активированную повторную селекцию) и/или минимизированные последовательности аптамером, также можно необязательно оптимизировать после SELEXTM с использованием медицинской химии аптамеров для проведения случайного или направленного мутагенеза последовательности для повышения аффинности связывания и/или функциональных характеристик, или альтернативно, для определения того, какие положения в последовательности необходимы для активности связывания и/или функциональных характеристик.
Медицинская химия аптамеров представляет собой способ усовершенствования аптамеров, в котором химически синтезируют ряд вариантов аптамеров. Эти группы вариантов, как правило, отличаются от исходного аптамера посредством встраивания отдельного заместителя, и они отличаются друг от друга локализацией этого заместителя. Затем эти варианты сравнивают между собой и с исходным аптамером. Улучшение свойств может быть настолько выраженным, что включение отдельного заместителя может оказаться всем, что необходимо для достижения определенного терапевтического критерия.
Альтернативно информацию, полученную от ряда отдельных вариантов, можно использовать для разработки дополнительных наборов вариантов, в которых одновременно встраивают более одного заместителя. В одной стратегии разработки оценивают все варианты отдельных заместителей, отбирают 4 наилучших и синтезируют и анализируют все возможные двойные (6), тройные (4) и четверные (1) сочетания из этих 4 вариантов отдельных заместителей. Во второй стратегии разработки наилучший отдельный вариант заместителя рассматривают как новый исходный аптамер и синтезируют и анализируют все возможные двойные варианты заместителей, которые включают этот наиболее высоко оцененный отдельный вариант заместителя. Можно использовать другие стратегии, и эти стратегии можно использовать многократно, так чтобы при продолжении идентификации дополнительно улучшенных вариантов количество заместителей постепенно повышалось.
Медицинскую химию аптамеров, в частности, можно использовать в качестве способа исследования локального, скорее чем общего, встраивания заместителей. Вследствие того, что аптамеры выявляют из библиотек, которые получают посредством транскрипции, любые заместители, которые встраивают в процессе SELEXTM, должны быть встроены по всей молекуле. Например, если желательно внести фосфоротиоатные связи между нуклеотидами, то они могут быть встроены в каждый A (или каждый G, C, T, U и т.д.) (замещение по всей молекуле). Аптамеры, где необходимо наличие фосфоротиоатов в некоторых A (или некоторых G, C, T, U и т.д.) (локальное замещение), но не допускается их наличие в других A, нельзя легко сконструировать с помощью этого процесса.
Типы заместителей, которые можно использовать в процессе медицинской химии аптамеров, ограничиваются только возможностью их получения в качестве реагентов для твердофазного синтеза и введения их в схему синтеза олигомеров. Процесс, безусловно, не ограничивается только нуклеотидами. Схемы медицинской химии аптамеров могут включать заместители, которые обеспечивают пространственный объем, гидрофобность, гидрофильность, липофильность, липофобность, положительный заряд, отрицательный заряд, нейтральный заряд, цвиттерионы, поляризуемость, устойчивость к нуклеазам, конформационную жесткость, конформационную подвижность, свойства в отношении связывания белков, массу и т.д. Схемы медицинской химии аптамеров могут включать модификации оснований, модификации сахаров или модификации фосфодиэфирных связей.
При рассмотрении типов заместителей, которые, вероятно, окажутся преимущественными в контексте терапевтического аптамера, может быть желательным введение заместителей, которые находятся в одной или нескольких из следующих категорий:
(1) Заместители уже представленные в организме, например, 2'-дезокси-, 2'-рибо-, 2'-O-метилпурины или пиримидины или 5-метилцитозин.
(2) Заместители, которые уже являются частью одобренных лекарственных средств, например, связанные посредством фосфоротиоата олигонуклеотиды.
(3) Заместители, которые подвергаются гидролизу или деградации до одной из указанных выше двух категорий, например, связанные с метилфосфонатом олигонуклеотиды.
Аптамеры по этому изобретению включают аптамеры, разработанные посредством медицинской химии аптамеров, как описано в настоящем документе.
Аффинность связывания мишени аптамеров по настоящему изобретению можно оценивать посредством серии реакций связывания между аптамером и мишенью (например, белком), в которых меченный меткой 32P аптамер инкубируют с сериями разведений мишени в буферной среде, а затем анализируют с помощью фильтрации через нитроцеллюлозу с использованием вакуумной фильтрационной установки. Называемый в настоящем документе дот-блот анализом связывания, этот способ использует трехслойную фильтрационную среду, состоящую (сверху вниз) из нитроцеллюлозы, нейлонового фильтра и бумаги для блоттинга гелей. РНК, связанная с мишенью, задерживается на нитроцеллюлозном фильтре, в то время как не связанная с мишенью РНК задерживается на нейлоновом фильтре. Бумагу для блоттинга гелей включают в качестве поддерживающей основы для других фильтров. После фильтрации, слои фильтра разделяют, сушат и проявляют на фосфорном экране и количественно определяют с использованием системы для визуализации фосфора. Количественно определенные результаты можно использовать для получения кривых связывания аптамеров, из которых можно вычислять константы диссоциации (KD). В предпочтительном варианте осуществления, буферная среда, используемая для проведения реакций связывания, представляет собой 1X PBS Дульбекко (с Ca++ и Mg++) плюс 0,1 мг/мл BSA.
Как правило, способность аптамера модулировать функциональную активность мишени, т.е. функциональную активность аптамера, можно оценивать с использованием моделей in vitro и in vivo, которые будут варьировать в зависимости от биологической функции мишени. В некоторых вариантах осуществления, аптамеры по настоящему изобретению могут ингибировать известную биологическую функцию мишени, в то время как в других вариантах осуществления аптамеры по этому изобретению могут стимулировать известную биологическую функцию мишени. Функциональную активность аптамеров по настоящему изобретению можно оценивать с использованием моделей in vitro и in vivo, разработанных для определения известной функции мишени компонента комплемента.
Аптамеры по настоящему изобретению можно общепринятыми способами адаптировать для диагностических целей в соответствии с любым количеством способов, используемых специалистами в данной области. Диагностическое применение может включать диагностическое применение как in vivo, так и in vitro. Диагностические средства должны обладать способностью позволить пользователю идентифицировать наличие данной мишени в конкретной области или концентрации. Просто способность образовывать связывающие пары с мишенью могут быть достаточной для обеспечения положительного сигнала для диагностических целей. Специалисты в данной области также способны адаптировать любой аптамер способами, известными в данной области, для включения метки в целях прослеживания наличия такого лиганда. Такую метку можно использовать в ряде диагностических способов.
МОДУЛИРОВАНИЕ ФАРМАКОКИНЕТИКИ И БИОРАСПРЕДЕЛЕНИЯ ЛЕКАРСТВЕННЫХ СРЕДСТВ НА ОСНОВЕ АПТАМЕРОВ
Важно, что фармакокинетические свойства всех лекарственных средств на основе олигонуклеотидов, включая аптамеры, должны соответствовать требуемому фармацевтическому способу применения. Несмотря на то, что для аптамеров, направленных против внеклеточных мишеней, отсутствуют проблемы, связанные с внутриклеточной доставкой (как в случае с лекарственными средствами на основе антисмысловых последовательностей и RNAi), тем не менее, такие аптамеры должны обладать способностью к распределению в органы-мишени и ткани-мишени и поддерживаться в организме (в немодифицированной форме) в течение периода времени, согласующегося с требуемой схемой дозирования.
Таким образом, настоящее изобретение относится к материалам и способам для влияния на фармакокинетику композиций аптамеров, и, в частности, к возможности регулировать фармакокинетику аптамеров. Возможности регулировать (т.е. возможность модулировать) фармакокинетику достигают посредством конъюгации с аптамером модифицирующих групп (например, полимеров PEG) и/или встраивания модифицированных нуклеотидов (например, 2'-фтор или 2'-O-метил) для изменения химического состава нуклеиновой кислоты. Возможность регулировать фармакокинетику аптамеров применяют для улучшения существующих способов терапевтического применения, или, альтернативно, для разработки новых способов терапевтического применения. Например, в некоторых способах терапевтического применения, например, в условиях интенсивной терапии при приступах или противоопухолевой терапии, где может быть желательным быстрое выведение или удаление лекарственного средства, желательным является снижение времени нахождения аптамеров в кровотоке. Альтернативно в других способах терапевтического применения, например, при поддерживающей терапии, где желательной является системная циркуляция лекарственного средства, может быть желательным повышение времени нахождения аптамеров в кровотоке.
Кроме того, возможность регулирования фармакокинетики аптамеров используют для модификации биораспределения лекарственного средства на основе аптамера у субъекта. Например, в некоторых способах терапевтического применения может быть желательным изменение биораспределения лекарственного средства на основе аптамера для направления воздействия на конкретный тип ткани или конкретных орган (или группу органов). В этих способах применения лекарственное средство на основе аптамера предпочтительно накапливается в конкретной ткани или конкретном органе(ах). В других способах терапевтического применения может быть желательным направление воздействия на ткани, в которых выявлен клеточный маркер или симптом, ассоциированный с данным заболеванием, клеточным повреждением или другой патологией, так чтобы лекарственное средство на основе аптамера предпочтительно накапливалось в поврежденной ткани. Например, как описано в находящейся на рассмотрении условной заявке США с серийным № 60/550790, поданной 5 марта 2004 года, с названием "Controlled Modulation of the Pharmacokinetics and Biodistribution of Aptamer Therapeutics", и в безусловной заявке США с серийным № 11/075648, поданной 7 марта 2005 года, и называемой "Controlled Modulation of the Pharmacokinetics and Biodistribution of Aptamer Therapeutics", для направления воздействия на воспаленные ткани используют пегилирование лекарственного средства на основе аптамера (например, пегилирование полимером PEG 20 кДа) таким образом, что пегилированное лекарственное средство на основе аптамера предпочтительно накапливается в воспаленной ткани.
Для определения профилей фармакокинетики и биораспределения лекарственных средств на основе аптамеров (например, конъюгатов аптамеров или аптамеров с измененными химическими группами, такими как модифицированные нуклеотиды) проводят мониторинг множества параметров. Такие параметры включают, например, время полужизни (t1/2), выведение из плазмы (Cl), объем распределения (Vss), площадь под кривой концентрация-время (AUC), максимальную наблюдаемую концентрацию в сыворотке или в плазме (Cmax) и среднее время нахождения (MRT) композиции аптамера. Как используют в настоящем документе, термин "AUC" относится к площади под кривой, отражающей зависимость концентрации лекарственного средства на основе аптамера в плазме от времени после введения аптамера. Значение AUC используют для оценки биодоступности (т.е. процентного количества введенного лекарственного средства на основе аптамера в кровотоке после введения аптамера) и/или общего выведения (Cl) (т.е. скорости, с которой лекарственное средство на основе аптамера удаляется из кровотока) данного лекарственного средства на основе аптамера. Объем распределения связывает концентрацию в плазме лекарственного средства на основе аптамера с количеством аптамера, находящегося в организме. Чем больше Vss, тем больше аптамера обнаруживается вне плазмы (т.е., тем больше экстравазация).
Настоящее изобретение относится к материалам и способам для контролируемого модулирования фармакокинетики и биораспределения устойчивых композиций на основе аптамера in vivo посредством конъюгации аптамера с модулирующей группой, такой как низкомолекулярное соединение, пептид или полимерная концевая группа, или посредством встраивания в аптамер модифицированных нуклеотидов. Как описано в настоящем документе, конъюгация модифицирующей группы и/или измененного нуклеотида(ов) с химической композицией изменяет основные аспекты времени нахождения аптамера в циркуляции и распределения в тканях.
В дополнение к выведению посредством нуклеаз, лекарственные средства на основе олигонуклеотидов подвергаются элиминации посредством почечной фильтрации. По существу, устойчивый к действию нуклеаз олигонуклеотид, введенный внутривенно, как правило, обладает периодом полураспада in vivo <10 мин, если только фильтрация не может быть заблокирована. Блокирование фильтрации можно проводить либо облегчением быстрого распределения из кровотока в ткани, либо повышением кажущейся молекулярной массы олигонуклеотида выше эффективного предела пропускания гломерул по размеру. Конъюгация низкомолекулярных лекарственных средств с полимером PEG (пегилирование), описанная ниже, может значительно удлинить время нахождения аптамеров в кровотоке, таким образом, снижая частоту дозирования и повышая эффективность против мишеней в сосудах.
Аптамеры можно конъюгировать со множеством модифицирующих групп, таких как высокомолекулярные полимеры, например, PEG; пептиды, например, Tat (фрагмент из 13 аминокислот белка HIV Tat (Vives, et al., (1997), J. Biol. Chem. 272(25): 16010-7)), Ant (16-аминокислотная последовательность, полученная из третьей спирали гомеозисного белка Drosophila antennapedia (Pietersz, et al., (2001), Vaccine 19(11-12): 1397-405)) и Arg7 (короткие, положительно заряженные пептиды для проникновения в клетку, состоящие из полиаргинина (Arg7) (Rothbard, et al., (2000), Nat. Med. 6(11): 1253-7; Rothbard, J et al, (2002), J. Med. Chem. 45(17): 3612-8)); и низкомолекулярные соединения, например, липофильные соединения, такие как холестерин. Среди различных описанных в настоящем документе конъюгатов, наиболее значительно свойства аптамеров in vivo изменяются при образовании комплексов с группами PEG. Например, конъюгация лекарственного средства на основе аптамера с полимером PEG массой 20 кДа, препятствует почечной фильтрации и обеспечивает распределение аптамера как в здоровых, так и в воспаленных тканях, как описано в безусловной заявке, приведенной выше (заявка США с серийным № 11/075648, поданная 7 марта 2005 года, и называемая "Controlled Modulation of the Pharmacokinetics and Biodistribution of Aptamer Therapeutics"). Более того, конъюгат полимер PEG массой 20 кДа - аптамер является приблизительно настолько же эффективным в отношении предотвращения почечной фильтрации аптамеров, как и полимер PEG массой 40 кДа. В то время как одним эффектом пегилирования является эффект на выведение аптамера, длительное системное воздействие, обеспечиваемое наличием группы массой 20 кДа, также облегчает распределение аптамера в ткани, в частности, в ткани органов с высокой перфузией и в ткани в области воспаления. Конъюгат аптамер-полимер массой PEG массой 20 кДа направляет распределение аптамера в область воспаления, так что пегилированный аптамер предпочтительно накапливается в воспаленной ткани. В некоторых случаях, пегилированный конъюгат аптамера массой 20 кДа способен проникать внутрь клеток, например, таких как клетки почки.
В целом, эффекты на фармакокинетику и распределение в тканях аптамеров, полученных посредством низкомолекулярных модифицирующих групп, включая холестерин и обеспечивающие проникновение в клетки пептиды, как правило, являются менее выраженными, чем эффекты, полученные в результате пегилирования или модификации нуклеотидов (например, измененный химический состав). Без связи с теорией, полагают, что опосредуемое холестерином связывание с липопротеинами плазмы, предположительно происходящее в случае антисмыслового конъюгата, устранены в конкретном случае собранной структуры конъюгата аптамер-холестерин, и/или относится к аспекту липофильной природы группы холестерина. Подобно холестерину, наличие пептидного маркера Tat обеспечивает выведение аптамера из кровотока, с относительно низкими уровнями конъюгата, появляющегося в почках в течение 48 ч. Другие пептиды (например, Ant, Arg7), для которых было описано в данной области, что они опосредует прохождение через мембраны in vitro, как оказалось, на обеспечивают выведение из кровотока. Однако, подобно Tat, конъюгат Ant в значительной степени накапливается в почках относительно других аптамеров. Без связи с теорией, возможно, что неблагоприятное представление модифицирующих групп в виде пептидов Ant и Arg7 в контексте трехмерно свернутых аптамеров in vivo нарушает способность этих пептидов влиять на транспортные свойства аптамеров.
Также модифицированные нуклеотиды можно использовать для модулирования выведения аптамеров из плазмы. Например, для неконъюгированного аптамера, который включает 2'-F, 2'-OMe и/или фосфоротиоатные стабилизирующие химические группы, которые являются типичными для аптамеров настоящего поколения, поскольку они проявляют высокую степень стабильности в отношении нуклеаз in vitro и in vivo, показано быстрое удаление из плазмы (т.е. быстрое выведение из плазмы) и быстрое распределение в тканях, главным образом, в почках, по сравнению с немодифицированным аптамером.
Производные нуклеиновых кислот, полученные с помощью PAG
Как описано выше, получение производных нуклеиновых кислот с высокомолекулярными неиммуногенными полимерами обеспечивает возможность изменения фармакокинетических и фармакодинамических свойств нуклеиновых кислот, что делает их более эффективными лекарственными средствами. Положительные изменения активности могут включать повышение устойчивости к деградации нуклеазами, снижение фильтрации почками, снижение воздействия со стороны иммунной системы, и изменение распределения лекарственного средства в организме.
Производные композиций на основе аптамеров по этому изобретению можно получать с помощью групп полиалкиленгликоля (PAG). Примеры производных нуклеиновых кислот, полученных с помощью PAG, можно найти в патентной заявке США № 10/718833, поданной 21 ноября 2003 года, которая включена в настоящий документ в качестве ссылки в полном объеме. Типичные полимеры, используемые по этому изобретению, включают полиэтиленгликоль (PEG), также известный как полиэтиленоксид (PEO), и полипропиленгликоль (включая полиизопропиленгликоль). Кроме того, во многих способах применения можно использовать статистические сополимеры или блок-сополимеры различных оксидов алкиленов (например, оксида этилена и оксида пропилена). В своей самой распространенной форме, полиалкиленгликоль, такой как PEG, представляет собой линейный полимер, оканчивающийся с каждой стороны гидроксильными группами: HO-CH2CH2O-(CH2CH2O)n-CH2CH2-OH. Этот полимер, альфа-, омега-дигидроксилполиэтиленгликоль, также может быть изображен в виде HO-PEG-OH, где понятно, что символ -PEG- отображает следующую структурную единицу: -CH2CH2O-(CH2CH2O)n-CH2CH2-, где n, как правило, находится в диапазоне от приблизительно 4 до приблизительно 10000.
Полимеры PAG, пригодные для терапевтического применения, как правило, обладают свойствами растворимости в воде и во многих органических растворителях, они не обладают токсичностью и иммуногенностью. Одним из способов применения PAG является ковалентное связывание полимера с нерастворимыми молекулами для обеспечения растворимости полученного "конъюгата" PAG-молекула. Например, было показано, что нерастворимое в воде лекарственное средство паклитаксел при соединении с PEG становится растворимым в воде. Greenwald, et al. J. Org. Chem., 60:331-336 (1995). Конъюгаты с PAG часто используют не только для повышения растворимости и стабильности, но также для увеличения продолжительности времени полужизни молекул в кровотоке.
Возможность образования производных с PAG, например, конъюгации с PEG, для изменения биораспределения лекарственного средства связана с множеством факторов, включающих кажущийся размер (например, как измеряют в единицах гидродинамического радиуса) конъюгата. Известно, что более крупные конъюгаты (>10 кДа) более эффективно блокируют фильтрацию через почку и последовательно повышают время полужизни в сыворотке небольших макромолекул (например, пептидов, антисмысловых олигонуклеотидов). Было показано, что способность конъюгатов с PEG блокировать фильтрацию возрастает с увеличением размера PEG вплоть до приблизительно 50 кДа (дальнейшее повышение оказывает минимальный положительный эффект, поскольку время полужизни начинает определять опосредуемым макрофагами метаболизмом, а не элиминацией через почки).
Размер полиалкилированных соединений по этому изобретению, как правило, составляет между 5 и 80 кДа, однако можно использовать соединения любого размера, выбор зависит от аптамера и способа применения. Размер других соединений PAG по этому изобретению составляет между 10 и 80 кДа. Размер других соединений PAG по этому изобретению составляет между 10 и 60 кДа. Например, размер полимера PAG может составлять по меньшей мере 10, 20, 30, 40, 50, 60 или 80 кДа. В некоторых вариантах осуществления, PEG представляет собой линейный PEG, в то время как в других вариантах осуществления, PEG представляет собой разветвленный PEG. В других вариантах осуществления PEG представляет собой разветвленный PEG массой 40 кДа, как представлено на Фиг.4. В некоторых вариантах осуществления разветвленный PEG массой 40 кДа присоединен к 5'-концу аптамера, как изображено на Фиг.5.
Настоящее изобретение относится к другому экономичному направлению синтеза конъюгатов высокомолекулярный PEG-нуклеиновая кислота (предпочтительно, аптамер), включающему многократно пегилированные нуклеиновые кислоты. Настоящее изобретение также относится к связанным посредством PEG мультимерным олигонуклеотидам, например, к димеризованным аптамерам. В противоположность лекарственным средствам на основе биологически экспрессируемых белков, лекарственные средства на основе нуклеиновых кислот, как правило, химически синтезируют из активированных мономерных нуклеотидов. Конъюгаты PEG-нуклеиновая кислота можно получать встраиванием PEG с использованием того же повторяющегося синтеза из мономеров. Например, PEG, активированные превращением в форму фосфорамидита, могут быть включены в твердофазный синтез олигонуклеотидов. Альтернативно синтез олигонуклеотидов можно проводить сайт-специфическим встраиванием реакционно-способного участка для присоединения PEG.
АКТИВИРОВАННЫЕ PEG
Получение высокомолекулярных PEG (>10 кДа) может быть сложным, неэффективным и дорогостоящим. В качестве курса в направлении синтеза конъюгатов высокомолекулярный PEG-нуклеиновая кислота, предшествующая работа была сосредоточена на получении активированных PEG c более высокой молекулярной массой. Способ поучения таких молекул вовлекает образование линейного активированного PEG или разветвленного активированного PEG, в случае которого с центром, несущим активированную группу, соединены два или более PEG. Терминальные участки этих молекул PEG с более высокой молекулярной массой, т.е. относительно не реакционно-способные гидроксильные (-OH) группы, можно активировать или превращать в функциональные группы, для присоединения одного или нескольких PEG к другим соединениям в реакционно-способных участках соединения. Разветвленные активированные PEG обладают более чем двумя концами, и в случаях, где активированы два или более концов, такие активированные молекулы PEG с более высокой молекулярной массой называют в настоящем документе мультиактивированными PEG. В некоторых случаях, не все концы в разветвленной молекуле PEG являются активированными. В случаях, где активированными являются любые два конца разветвленной молекулы PEG, такие молекулы PEG называют биактивированными PEG. В некоторых случаях, где активированным является только один конец разветвленной молекулы PEG, такие молекулы PEG называются моноактивированными. В качестве примера этого подхода, описан активированный PEG, полученный присоединением двух монометокси-PEG к лизиновому центру, который затем активируют для реакции, (Harris et al., Nature, vol.2: 214-221, 2003).
Как показано на Фиг.6, линейная молекула PEG является бифункциональной и иногда ее называют "диол PEG". Концевые участки молекулы PEG представляют собой относительно не реакционно-способные гидроксильные группы, группы -OH, которые можно активировать или превращать в функциональные группы, для присоединения PEG к другим соединениям в реакционно-способных участках соединения. Такие активированные диолы PEG в настоящем документе рассматривают как биактивируемые PEG. Например, терминальные группы диола PEG были функционализированы в качестве активного карбонатного сложного эфира для селективной реакции с аминогруппами замещением относительно не реакционно-способных гидроксильных групп -OH активными сложноэфирными группами сукцимидила из N-гидроксисукцинимида. Альтернативно, диолы PEG могут быть активированы с помощью различных групп, включая, без ограничений, α-галоуксусную кислоту, эпигалогидрины, малеаты, тартраты и карбогидраты, которые после соответствующих действий приведут к конъюгации активированного карбонила или эквивалента. Другие методы активации PEG описаны в Roberts et al., (2002) Advanced Drug Deliver Reviews 54:549-476, полностью включенную сюда в качестве ссылки. Дополнительно к активации PEG, используя один из описанных ранее методов, одна или обе терминальные спиртовые функциональные группы молекул PEG могут быть модифицированы для возможности осуществлять различные виды конъюгации с нуклеиновой кислотой. Например, преобразование одной из терминальных спиртовых групп в аминную или тиольную группы позволяет получить конъюгаты мочевины и тиомочевины.
Во многих способах применения является желательным кэпирование молекулы PEG на одном конце по существу не реакционно-способной группой, так чтобы молекула PEG
являлась монофункциональной (или моноактивируемой). В случае белковых лекарственных средств, на которых, как правило, находятся множественные реакционные центры для активированных PEG, бифункциональные активированные PEG приводят к обширному поперечному связыванию, с получением мало функциональных агрегатов. Для получения моноактивированных PEG, одну гидроксильную группу на конце молекулы PEG диола, как правило, замещают не реакционно-способной концевой группой метокси, -OCH3. Другой, не содержащий кэп, конец молекулы PEG, как правило, превращают в реакционно-способную концевую группу, которая может быть активирована для присоединения к реакционно-способному центру на поверхности молекулы, такой как белок.
АПТАМЕРЫ, КОНЪЮГИРОВАННЫЕ С ОДНИМ ИЛИ НЕСКОЛЬКИМИ PEG
Наиболее часто, синтез конъюгатов высокомолекулярный PAG-нуклеиновая кислота проводят добавлением свободного первичного амина к 5'-концу (встраиваемому с использованием модификатора фосфорамидита на последней стадии присоединения в твердофазном синтезе). С использованием этого подхода, реакционно-способный PEG (например, PEG, активированный таким образом, что он будет вступать в реакцию и образовывать связь с амином) объединяют с очищенным олигонуклеотидом и в растворе проводят реакцию присоединения.
Кроме того, высокомолекулярные конъюгаты PAG-нуклеиновая кислота-PAG можно получать реакцией монофункционального активированного PEG с нуклеиновой кислотой, содержащей более одного реакционно-способного участка. В одном варианте осуществления нуклеиновая кислота является биреакционно-способной и содержит два реакционно-способных участка: 5'-аминогруппу и 3'-аминогруппу, встроенные в олигонуклеотид посредством общепринятого синтеза с фосфорамидитом, начиная с твердой положки с 3'-амином: 3'-5'-ди-пегилирование, как представлено на Фиг.6. В альтернативных вариантах осуществления, реакционно-способные участки можно встраивать во внутренние положения, например, с использованием 5 положения пиримидинов, 8 положения пуринов, или 2' положения рибозы в качестве участков для присоединения первичных аминов. В таких вариантах осуществления нуклеиновая кислота может иметь несколько активированных или реакционно-способных участков и ее называют множественно активированной.
Для получения конъюгата нуклеиновая кислота-PEG-нуклеиновая кислота, нуклеиновую кислоту изначально синтезируют таким образом, чтобы она содержала один реакционно-способный участок (например, чтобы она являлась моноактивированной). В предпочтительном варианте осуществления этот реакционно-способный участок представляет собой аминогруппу, встроенную на 5'-конце посредством добавления модификатора фосфорамидита в качестве последней стадии твердофазного синтеза олигонуклеотида. В другом варианте осуществления, синтез проводят с использованием модификатора 3'-амина с внесением амина на 5'-конец, что приводит к 3',5'-диаминолигонуклеотиду. После удаления защитной группы и очистки модифицированного олигонуклеотида, его при высокой концентрации разбавляют в растворе, который сводит к минимуму спонтанный гидролиз активированного PEG. В предпочтительном варианте осуществления концентрация олигонуклеотида составляет 1 мМ и разбавленный раствор содержит 200 мМ буфера на основе NaHCO3, pH 8,3. Синтез конъюгата начинается с медленного поэтапного добавления высоко очищенного активированного PEG. В предпочтительном варианте осуществления, PEG является активированным в качестве п-нитрофенилкарбоната. После реакции конъюгат PEG-нуклеиновая кислота очищают гель-электрофорезом или жидкостной хроматографией для разделения полностью, частично и неконъюгированных молекул.
МНОЖЕСТВО АПТАМЕРОВ, КОНЪЮГИРОВАННЫХ С ОДНИМ PEG
Также настоящее изобретение относится к высокомолекулярным композициям аптамера, в которых две или более группы нуклеиновых кислот ковалентно конъюгированы по меньшей мере с одной группой полиалкиленгликоля. Группы полиалкиленгликоля служат в качестве стабилизирующих групп. Стабилизирующая группа представляет собой молекулу или часть молекулы, которые повышают фармакокинетические и фармакодинамические свойства высокомолекулярных композиций аптамера по этому изобретению. В некоторых случаях, стабилизирующая группа представляет собой молекулу или часть молекулы, которая обеспечивает близкое расположение двух или более аптамеров, или доменов аптамеров, или обеспечивает снижение общей свободы вращения высокомолекулярных композиций аптамера по этому изобретению. Стабилизирующая группа может представлять собой полиалкиленгликоль, такой как полиэтиленгликоль, который может представлять собой линейный или разветвленный гомополимер или гетерополимер. Другие стабилизирующие группы включают полимеры, такие как пептидные нуклеиновые кислоты (PNA). Олигонуклеотиды также могут быть стабилизирующими группами; такие олигонуклеотиды могут включать модифицированыне нуклеотиды, и/или модифицированные связи, такие как фосфоротиоаты.
Стабилизирующая группа может быть неотъемлемой частью композиции аптамера, т.е. она ковалентно связана с аптамером. В композициях, где группа полиалкиленгликоля ковалентно связана с каким-либо концом аптамера, так что полиалкиленгликоль связывает группы нуклеиновых кислот вместе в одну молекулу, полиалкиленгликоль называют линкерной группой. В таких композициях, первичная структура ковалентной молекулы включает линейное расположение нуклеиновая кислота-PAG-нуклеиновая кислота. Одним примером композиции, где стабилизирующая группа PEG служит в качестве линкера, который разделяет различные части аптамера, является композиция, где PEG конъюгирован в единую последовательность аптамера, так чтобы линейное расположение высокомолекулярной композиции на основе аптамера представляло собой, например, расположение нуклеиновая кислота-PEG-нуклеиновая кислота (-PEG-нуклеиновая кислота)n, где n больше или равно 1.
Для получения конъюгата нуклеиновая кислота-PEG-нуклеиновая кислота, нуклеиновую кислоту изначально синтезируют таким образом, чтобы она содержала один реакционно-способный участок (например, чтобы она являлась моноактивированной). В предпочтительном варианте осуществления этот реакционно-способный участок представляет собой аминогруппу, встроенную на 5'-конце посредством добавления модификатора фосфорамидита в качестве последней стадии твердофазного синтеза олигонуклеотида. После удаления защитной группы и очистки модифицированного олигонуклеотида, его при высокой концентрации разбавляют в растворе, который сводит к минимуму спонтанный гидролиз активированного PEG. В предпочтительном варианте осуществления концентрация олигонуклеотида составляет 1 мМ и разбавленный раствор содержит 200 мМ буфера на основе NaHCO3, pH 8,3. Синтез конъюгата начинается с медленного, поэтапного добавления высоко очищенного бифункционального PEG. В предпочтительном варианте осуществления, диол PEG активируют на обоих концах (би-активированный) посредством получения производного в качестве п-нитрофенилкарбоната. После реакции конъюгат PEG-нуклеиновая кислота очищают гель-электрофорезом или жидкостной хроматографией для разделения полностью, частично и неконъюгированных образцов. Для достижения различной длины (или молекулярной массы) можно присоединять несколько связанных молекул PAG (например, в качестве статистических сополимеров или блок-сополимеров) или цепи PAG меньших размеров. Между цепями PAG различной длины можно использовать линкеры, представляющие собой не PAG.
Линкерные домены также могут содержать одну или несколько групп полиалкиленгликоля, присоединенных к ним. Такие PAG могут обладать различной длиной и их можно использовать в соответствующих сочетаниях для достижения требуемой молекулярной массы композиции.
На эффект конкретного линкера может влиять как его химическая структура, так и его длина. Линкер, который является слишком длинным, слишком коротким или вступает в нежелательные пространственные и/или ионные взаимодействия с мишенью, препятствует образованию комплекса между аптамером и мишенью. Линкер, более длинный, чем необходимо для того, чтобы перекрыть расстояние между нуклеиновыми кислотами, может снижать стабильность связывания посредством уменьшения эффективной концентрации лиганда. Таким образом, в целях повышения до максимума аффинности аптамера к мишени часто необходимо оптимизировать состав и длину линкера.
Аптамеры с аффинностью связывания с белком C5 системы комплемента
В некоторых вариантах осуществления, материалы по настоящему изобретению содержат серию нуклеиновых кислот-аптамеры длиной от приблизительно 15 до приблизительно 60 нуклеотидов, которые специфично связываются с белком C5 комплемента и которые функционально модулируют, например блокируют, активность белка C5 комплемента в анализах in vivo и/или в клеточных анализах.
В некоторых вариантах осуществления настоящего изобретения описаны аптамеры, которые способны специфично связывать и модулировать белок C5 комплемента. Эти аптамеры обеспечивают низко токсичную, безопасную и эффективную методику лечения, смягчения и/или профилактики различных связанных с комплементом заболеваний или нарушений, включая, например, связанные с комплементом нарушения сердца (например, повреждение миокарда; опосредуемые компонентом C5 комплемента осложнения, связанные с хирургической операцией по аортокоронарному шунтированию (CABG), такие как постоперационное кровотечение, системная активация нейтрофилов и лейкоцитов, повышенный риск инфаркта миокарда, и повышенная когнитивная дисфункция; рестеноз; и опосредуемые компонентом C5 комплемента осложнения, связанные с чрескожным вмешательством на коронарных сосудах), повреждение при ишемии-реперфузии (например, инфаркт миокарда, инсульт, отморожение), связанные с комплементом воспалительные нарушения (например, астму, артрит, сепсис и отторжение после трансплантации органа), и связанные с комплементом аутоиммунные нарушения (например, миастению, системную красную волчанку (SLE)). Другие показания, при которых является желательным ингибирование C5, включают, например, воспаление легких (Mulligan et al. (1998) J. Clin. Invest. 98:503), экстракорпоральную активацию комплемента (Rinder et al. (1995) J. Clin. Invest. 96:1564), опосредуемую антителом активацию комплемента (Biesecker et al. (1989) J. Immunol. 142:2654), гломерулонефрит и другие заболевания почек, офтальмологические показания, такие как опосредуемое C5 повреждение тканей глаза, например, диабетическая ретинопатия, экссудативная и/или неэкссудативная возрастная дегенерация желтого пятна (AMD), и пароксизмальную ночную гемоглобинурию. Также эти аптамеры можно использовать в диагностике.
В некоторых вариантах осуществления, аптамеры по настоящему изобретению можно применять для низко токсичного, безопасного и эффективного способа лечения, стабилизации и/или профилактики различных связанных с комплементом глазных заболеваний или нарушений в способах по этому изобретению, включая, например, острое или хроническое воспалительное и/или опосредуемое иммунной системой глазное нарушение, воспалительный конъюнктивит, включая аллергический и гигантский папиллярный конъюнктивит, отек желтого пятна, увеит, эндофтальмит, склерит, язвы роговицы, сухой кератит, глаукому, ишемическое заболевание сетчатки, отторжение трансплантата роговицы, осложнения, связанные с внутриглазной хирургической операцией, такой как внутриглазная имплантация искусственной линзы, и воспаление, обусловленное хирургической операцией по поводу катаракты, болезнь Бехчета, иммунокомплексный васкулит, болезнь Фуха, болезнь Фогта-Каянаги-Харада, субретинальный фиброз, кератит, витреоретинальное воспаление, паразитическое заражение/миграцию в глазу, пигментный ретинит, цитомегаловирусный ретинит и хориоидальное воспаление, дегенерацию желтого пятна, возрастную дегенерацию желтого пятна ("AMD"), AMD неэкссудативного ("сухого") типа, или глазное нарушение, связанное с неоваскуляризацией, включая диабетическую ретинопатию или AMD экссудативного ("влажного") типа. Также эти аптамеры можно применять в офтальмологической диагностике.
Эти аптамеры могут включать модификации, как описано в настоящем документе, включающие, например, конъюгацию с липофильными или высокомолекулярными соединениями (например, PEG), включение группы кэпа, включение модифицированных нуклеотидов и модификации фосфатного остова.
В одном варианте осуществления этого изобретения предусмотрен выделенный не встречающийся в природе аптамер, который связывается с белком C5 комплемента. В другом варианте осуществления, предусмотрен выделенный не встречающийся в природе аптамер, который связывается с белком C5 комплемента, для применения в способах по этому изобретению в целях лечения, стабилизации и/или профилактики опосредуемого комплементом глазного нарушения. В некоторых вариантах осуществления, выделенный не встречающийся в природе аптамер имеет константу диссоциации ("Kd") для белка C5 комплемента менее 100 мкМ, менее 1 мкМ, менее 500 нМ, менее 100 нМ, менее 50 нМ, менее 1 нМ, менее 500 пМ, менее 100 пМ, менее 50 пМ. В некоторых вариантах осуществления этого изобретения, константу диссоциации определяют дот-блот титрованием, как описано в примере 1, ниже.
В другом варианте осуществления, аптамеры для применения в способах по этому изобретению модулируют функцию белка C5 комплемента, в частности ингибируют функцию белка C5 комплемента и/или функцию варианта белка C5 комплемента. Как используют в настоящем документе, вариант белка C5 комплемента, охватывает варианты, которые выполняют по существу ту же функцию, что и функция белка C5 комплемента. Вариант белка C5 комплемента предпочтительно обладает по существу той же структурой и в некоторых вариантах осуществления обладает по меньшей мере 80% идентичностью последовательности, более предпочтительно по меньшей мере 90% идентичностью последовательности, и более предпочтительно по меньшей мере 95% идентичностью последовательности с аминокислотной последовательностью белка C5 комплемента, содержащего аминокислотную последовательность, указанную ниже (SEQ ID NO:102) (цитированную в Haviland et al., J Immunol. 1991 Jan 1; 146(1):362-8):


В некоторых вариантах осуществления этого изобретения, идентичность последовательности вариантов мишени определяют с использованием BLAST, как описано ниже. Термины "идентичность последовательностей" в контексте двух или более последовательностей нуклеиновых кислот или белков, относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми или обладают определенным процентным содержанием аминокислотных остатков или нуклеотидов, которые оказываются одинаковыми при сравнении и выравнивании для максимального соответствия, определяемого с использованием одного из представленных ниже алгоритмов сравнения последовательностей или визуальным исследованием. Как правило, при сравнении последовательностей одна последовательность выступает в качестве эталонной последовательности, с которой сравнивают анализируемые последовательности. При использовании алгоритма сравнения последовательностей, анализируемую и эталонную последовательности вводят в компьютер, при необходимости обозначают координаты подпоследовательностей и указывают параметры программы алгоритма для последовательностей. Затем алгоритм сравнения последовательностей вычисляет процентную идентичность последовательностей для анализируемой последовательности(ей) относительно эталонной последовательности, исходя из указанных параметров программы. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма локальной гомологии Smith & Waterman, Adv. Appl. Math. 2: 482 (1981), с помощью алгоритма выравнивания по гомологии Needleman & Wunsch, J Mol. Biol. 48: 443 (1970), с помощью способа поиска на наличие сходства Pearson & Lipman, Proc. Natl. Acad. Sci. USA 85: 2444 (1988), с помощью компьютеризованной реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wis.), или посредством визуального исследования (см., главным образом, Ausubel et al., ниже).
Одним примером алгоритма, который пригоден для определения процентной идентичности последовательностей, является алгоритм, используемый в инструментарии поиска базовых локальных блоков (в дальнейшем в этом документе "BLAST"), см., например, Altschul et al., J. Mol. Biol. 215: 403-410 (1990) и Altschul et al., Nucleic Acids Res., 15: 3389-3402 (1997). Программное обеспечение для проведения анализов BLAST является общедоступным с помощью Национального центра биотехнологической информации (в дальнейшем в этом документе "NCBI"). Параметры по умолчанию, применяемые для определения идентичности последовательностей с использованием программного обеспечения, доступного в NCBI, например, BLASTN (для нуклеотидных последовательностей) и BLASTP (для аминокислотных последовательностей), описаны в McGinnis et al., Nucleic Acids Res., 32: W20-W25 (2004).
В другом варианте осуществления этого изобретения предусмотрен аптамер, который обладает по существу такой же способностью связывать белок C5 комплемента, что и аптамер, содержащий любую из: SEQ ID NO:1-2, 5-67, 75-81, 83 или 88-98. В другом варианте осуществления этого изобретения, аптамер обладает по существу такой же структурой и способностью связывать белок C5 комплемента, что и аптамер, содержащий любую из: SEQ ID NO:1-2, 5-67, 75-81, 83 или 88-98. В другом варианте осуществления, аптамеры по этому изобретению обладают последовательностью, включающей любые химические модификации, в соответствии с любой из SEQ ID NO:2, 5-67, 75-81, 83 или 88-98. В другом варианте осуществления, аптамеры по этому изобретению применяют в качестве активного ингредиента в фармацевтических композициях. В другом варианте осуществления, аптамеры или композиции, содержащие аптамеры по этому изобретению, применяют для лечения различных связанных с комплементом заболеваний или нарушений, включая любое заболевание или нарушение, выбранное из группы, состоящей из: связанных с комплементом нарушений сердца (например, повреждения миокарда; опосредуемых компонентом C5 комплемента осложнений, связанных с аортокоронарным шунтированием (CABG), таких как послеоперационное кровотечение, системная активация нейтрофилов и лейкоцитов, повышенный риск инфаркта миокарда и повышенная когнитивная дисфункция; рестеноза; и опосредуемых компонентом комплемента C5 осложнений, связанных с чрескожным вмешательством на коронарных сосудах), повреждения при ишемии-реперфузии (например, инфаркт миокарда, инсульт, отморожение), связанных с комплементом воспалительных нарушений (например, астмы, артрита, сепсиса и отторжения после трансплантации органа), и связанных с комплементом аутоиммунных нарушений (например, миастении, системной красной волчанки (SLE), воспаления легких, экстракорпоральной активации комплемента, опосредуемой антителом активации комплемента и связанных с комплементом глазных заболеваний, таких как диабетическая ретинопатия, а также связанная со старением дегенерация желтого пятна (AMD).
В одном варианте осуществления, аптамеры против C5 по этому изобретению включают смесь 2'-фтор-модифицированных нуклеотидов, 2'-OMe-модифицированных нуклеотидов ("2'-OMe") и остатков 2'-OH-пуринов. Описательная общая последовательность (SEQ ID NO:1) для модифицированного аптамера против C5 представлена ниже в таблице 1, и его структура представлена на Фиг.3A. Подавляющее большинство пуринов (A и G) модифицированы до 2'-OMe, исключая только два остатка G, которые остаются 2'-OH (остатки, показанные контурным изображением). Заключенные в кружок остатки соответствуют группе пиримидинов, которые можно одновременно модифицировать в 2'-H без существенного изменения активности против C5 аптамера (см. ARC330 в таблице 1, ниже (SEQ ID NO:2, Фиг.3B)). Подчеркнутые остатки, представленные на Фиг.3A, соответствуют пиримидиновым остаткам, которые могут содержать либо 2'-фтор-, либо 2'-H-модификацию (но не 2'-OMe), в то время как заключенные в рамку остатки соответствуют пиримидиновым остаткам, которые могут содержать либо 2'-фтор-, либо 2-OMe-модификацию (но не 2'-H). Остатки, указанные стрелкой (→) должны содержать 2'-фтор-модификацию. Без 2'-фтор-модификации в остатках, указанных стрелкой (→), конечная гемолитическая активность полученного аптамера существенно снижается. В предпочтительном варианте осуществления, аптамер против C5 по этому изобретению содержит нуклеотидную последовательность в соответствии с SEQ ID NO:1.
Примером аптамера против C5 в соответствии с этим изобретением является ARC186 (SEQ ID NO:4), который показан на Фиг.3C и описан в патенте США № 6395888, который включен в настоящий документ в качестве ссылки в полном объеме. Все 21 пиримидиновых остатка ARC186 обладают 2'-фтор-модификациями. Большинство остатков пурина (14 остатков) обладают 2'-OMe-модификациями, за исключением трех 2'-OH пуриновых остатков (показанных контурным изображением на Фиг.3C). Аптамеры против C5 по этому изобретению также могут включать различные сочетания 2'-фтор- и 2'-H-модификаций. Например, другим аптамером против C5 по этому изобретению является ARC330 (SEQ ID NO:2), представленный на Фиг.3B. ARC330 (SEQ ID NO:2) содержит семь 2'-H модификаций (заключенные в кружок остатки на Фиг.3B), 14 пиримидиновых остатков с 2'-фтор-модификациями, 14 пуриновых остатков с 2'-OMe-модификациями, и три 2'-OH пуриновых остатка (показанных контурным изображением на Фиг.3B).
Другие сочетания аптамеров, обладающих сочетанием 2'-фтор-модификаций, 2'-OMe-модификаций, 2'-OH-пуриновых остатков, и конъюгацией с неиммуногенными высокомолекулярными соединениями (например, PEG) различного размера, все из которых образованы из ARC186 (SEQ ID NO:4), описаны в таблице 1, ниже. Настоящее изобретение включает аптамеры, как описано в таблице 1, ниже. Также настоящее изобретение включает аптамеры, описанные ниже, но лишенные указанного 3'-кэпа (например, инвертированного дезокситимидинового кэпа), и/или аптамеры, указанные ниже, но содержащие 3'-кэп (например, инвертированный dT), где 3'-кэп не указан.
Аптамер против C5 для применения в способах, описанных посредством некоторых вариантов осуществления настоящего изобретения, может представлять собой аптамер, содержащий любую из: SEQ ID NO: с 1 по 69, 75, 76, 81, 91, 95 и 96, описанных ниже.
Если нет иных указаний, нуклеотидные последовательности в таблице 1, ниже, приведены в направлении от 5' к 3'. Для каждой из индивидуальных последовательностей в таблице 1, все 2'-OMe-модификации пуринов или пиримидинов указаны посредством "m" перед соответствующим нуклеотидом; все 2'-фтор-модификации пиримидинов указаны посредством "f" перед соответствующим нуклеотидом; все дезокси-модификации пуринов или пиримидинов указан посредством "d" перед соответствующим нуклеотидом; и любой пурин или пиримидин без "m", "f" или "d" перед нуклеотидом указывает на остаток с 2'-OH. Кроме того, "3T" указывает на инвертированный дезокситимидин, "NH" указывает на гексиламиновый линкер, "NH2" указывает на гексиламиновую концевую группу, "PEG" указывает на группу полиэтиленгликоля, обладающую указанной молекулярной массой и "биотин" указывает на аптамер, имеющий биотин, конъюгированный с 5'-концом.
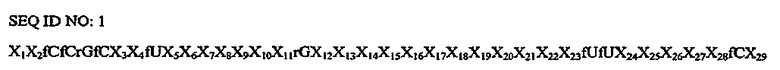





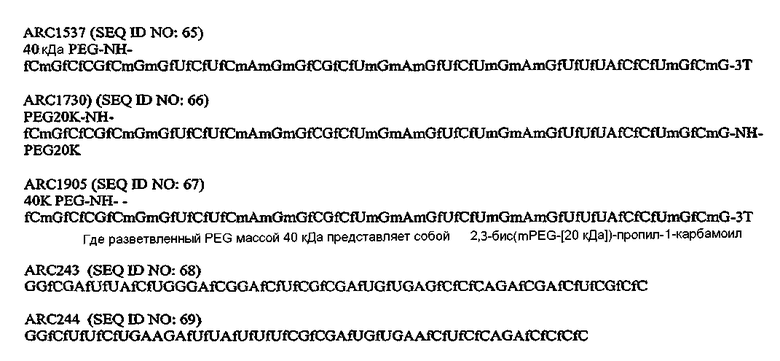
Кроме того, настоящее изобретение относится к аптамерам, представленным в таблице 2, ниже. Аптамеры, представленные в таблице 2, приведены в направлении от 5' к 3', и соответствуют рибонуклеотидной последовательности аптамеров, которые были отобраны в представленных условиях dRmY SELEXTM. В некоторых вариантах осуществления этого изобретения, полученных в результате этой селекции (и отраженных в списке последовательностей) пурины (A и G) являются дезокси и пиримидины (U и C) являются 2'-OMe. В некоторых вариантах осуществления аптамеры содержат кэп (например, 3'-инвертированный dT). В некоторых вариантах осуществления аптамеры содержат PEG.
Аптамеры против C5 dRmY
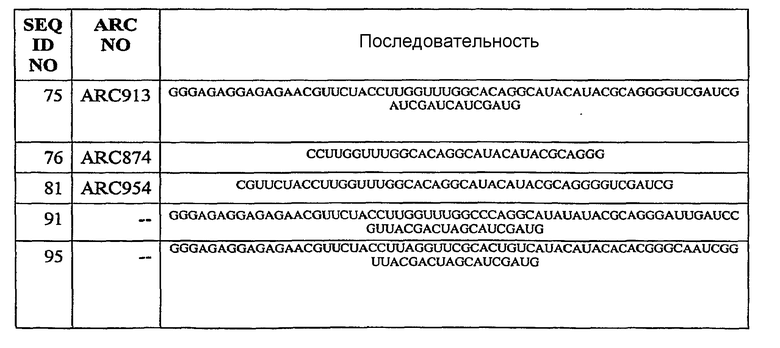
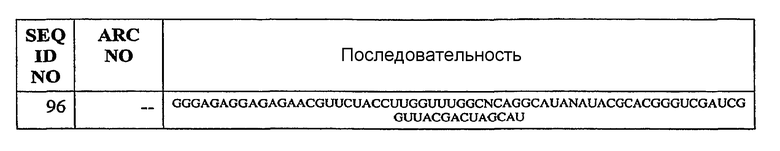
Другие аптамеры по этому изобретению, которые связывают белок C5 комплемента, описаны ниже в примере 3. Специфичные к C5 аптамеры, кроме того, описаны в условных патентных заявках США 60/544542, 60/547747, 60/581685 и 60/608048, все из которых включены в настоящий документ в качестве ссылок в полном объеме.
В некоторых вариантах осуществления лекарственные средства на основе аптамера по настоящему изобретению обладают высокой аффинностью и специфичностью к их мишеням, снижая неблагоприятные побочные эффекты вследствие не встречающихся в природе нуклеотидных замен, если лекарственное средство на основе аптамера разрушается в организме пациентов или субъектов. В некоторых вариантах осуществления, терапевтические композиции, содержащие лекарственные средства на основе аптамера по настоящему изобретению, не содержат или обладают сниженным количеством фторированных нуклеотидов.
Аптамеры по настоящему изобретению можно синтезировать с использованием любых способов синтеза олигонуклеотидов, известных в данной области, включая способы твердофазного синтеза олигонуклеотидов, хорошо известные в данной области (см., например, Froehler et al., Nucl. Acid Res. 14:5399-5467 (1986) и Froehler et al., Tet. Lett. 27:5575-5578 (1986)) и жидкофазные способы, такие как способы триэфирного синтеза (см., например, Sood et al., Nucl. Acid Res. 4:2557 (1977) и Hirose et al., Tet. Lett., 28:2449 (1978)).
Также настоящее изобретение относится к применению средств против C5 по этому изобретению с аптамерами, специфичными к PDGF и/или VEGF и/или распознаваемым ими рецепторов PDGFR и VEGFR, соответственно, в способах по этому изобретению для стабилизации, лечения и/или профилактики глазных нарушений. Таким образом, способы, описанные непосредственно выше, можно применять для создания аптамеров по этому изобретению в целях блокирования связывания лиганда (например, PDGF или VEGF) с его мишенью, такой как распознаваемый им рецептор.
Примеры аптамеров против PDGF для применения в способах по этому изобретению описаны в международной патентной заявке № PCT/US2005/039975, поданной 2 ноября 2005 года, и включенной в настоящий документ в качестве ссылки в полном объеме, в частности, ARC513, ARC594, ARC127 и ARC404, описанные в настоящем документе.
Примеры специфичных к VEGF аптамеров для применения в способах по этому изобретению описаны в патентах США № 5919455, 5932462, 6113906, 6011020, 6051698 и 6147204. Например, особенно пригодный аптамер для применения в способах лечения глазных нарушений в сочетании со средством против C5 по этому изобретению, может представлять собой EYE001 (ранее NX1838) в его пегилированной и непегилированной форме, в частности инъекцию пегаптаниба натрия (MacugenB®, Eyetech Pharmaceuticals, Inc. и Pfizer, Inc. NY, NY).
Средства на основе антител против C5
Средства против C5 по этому изобретению включают антитела-антагонисты, направленные против белка C5 комплемента и их применение в целях лечения опосредуемых C5 глазных нарушений. Антитела-антагонисты C5 по этому изобретению прочно связывают C5 и предотвращают его активацию и расщепление. В конкретных вариантах осуществления, настоящее изобретение относится к введению антитела против C5 субъекту в способе снижения, стабилизации и/или профилактики по меньшей мере одного симптома глазного нарушения, в частности, симптома диабетической ретинопатии, экссудативной и/или неэкссудативной AMD.
Антагонист антитела по этому изобретению включает моноклональные ингибиторные антитела. Моноклональные антитела, или их фрагменты, охватывают все классы иммуноглобулинов, такие как IgM, IgG, IgD, IgE, IgA, или их подклассы, такие как подклассы IgG, или их смеси. Пригодными являются IgG и их подклассы, такие как IgG1, IgG2, IgG2a, IgG2b, IgG3 или IgGM. Пригодные варианты осуществления включают подтипы IgG IgG.sub.1/каппа и IgG.sub.2b/каппа. Фрагменты, которые можно упомянуть, представляют собой все укороченные или модифицированные фрагменты антитела с одним или двумя комплементарными антигену участками связывания, которые показывают высокое связывание и нейтрализующую активность в отношении C5 млекопитающих, такие как части антител, обладающие участком связывания, которые соответствует антителу и образованы легкой и тяжелой цепями, такие как фрагменты Fv, Fab или F(ab')2 или одноцепочечные фрагменты. Особенно пригодными являются укороченные двухцепочечные фрагменты, такие как Fv, Fab или F(ab')2. Эти фрагменты можно получать, например, ферментативными способами посредством устранения Fc-части антитела с помощью ферментов, таких как папаин или пепсин, посредством химического окисления или посредством генетического манипулирования с генами антитела. Также является возможным и преимущественным применение генетически измененных неукороченных фрагментов.
Новые антитела, фрагменты антител, их смеси или производные преимущественно обладают аффинностью связывания в отношении C5 в диапазоне от 1.x.10-7 M до 1.x.10-12 M или от 1.х.10-8 M до 1.x.10-11 M или от 1.x.10-9 M до 5.х.10-10 M.
Гены антитела для генетического манипулирования можно выделять, например, из клеток гибридом, способом, известным специалисту. Для этой цели, антителопродуцирующие клетки культивируют и, когда оптическая плотность клеток становится достаточной, мРНК выделяют из клеток известным способом посредством лизиса клеток тиоцианатом гуанидиния, подкислением ацетатом натрия, экстракцией фенолом, смесью хлороформ/изоамиловый спирт, осаждением изопропанолом и промыванием этанолом. Затем из мРНК синтезируют кДНК с использованием обратной транскриптазы. Синтезированную кДНК можно встраивать, прямо или после генетического манипулирования, например, с посредством сайт-направленного мутагенеза, внесения вставок, инверсий, делеций или заменой оснований, в пригодный вектор животного, грибов, бактериальный или вирусный вектор, и экспрессировать в соответствующем хозяйском организме. Пригодные бактериальные или дрожжевые векторы представляют собой pBR322, pUC18/19, pACYC184, вектор лямбда или дрожжевой вектор mu для клонирования генов и экспрессии в бактериях, таких как E. coli, или в дрожжах, таких как Saccharomyces cerevisiae.
Более того, настоящее изобретение относится к клеткам, которые синтезируют антитела против C5. Они включают клетки животных, грибов, бактерий или клетки дрожжей после трансформации, как упомянуто выше. Они преимущественно представляют собой клетки гибридомы или клетки триомы, как правило, клетки гибридомы. Эти клетки гибридомы можно получать, например, известным способом из животных, иммунизированных с помощью C5, и посредством выделения их антителопродуцирующих B-клеток, селекции этих клеток в отношении связывающих C5 антител и последующего слияния этих клеток, например, с клетками человека или животного, например, клетками миеломы мыши, лимфобластоидными клетками человека или клетками гетерогибридомы (см., например, Koehler et al., (1975) Nature 256:496) или посредством инфицирования этих клеток соответствующими вирусами с получением иммортализованных клеточных линий. Пригодными являются гибридомные клеточные линии, полученные слиянием, и особенно пригодными являются гибридомные клеточные линии мыши. Гибридомные клеточные линии по этому изобретению секретируют пригодные антитела типа IgG. Связывающие антитела mAb по этому изобретению связываются с высокой аффинностью и снижают или нейтрализуют биологическую активность (например, расщепление C5) белка C5 комплемента.
Кроме того, настоящее изобретение относится к производным этих антител против C5, которые сохраняют их активность в отношении ингибирования C5, при изменении одного или нескольких других свойств, связанных с их применением в качестве фармацевтического средства, например, стабильности в сыворотке или эффективности продукции. Примеры таких производных антител против C5 включают пептиды, пептидомиметики, образованные из антигенсвязывающих участков антител, и антитела, фрагменты антител или пептиды, связанные с твердыми или жидкими носителями, такими как полиэтиленгликоль, стекло, синтетические полимеры, такие как полиакриламид, полистирол, полипропилен, полиэтилен или природные полимеры, такие как целлюлоза, сефароза или агароза, или конъюгаты с ферментами, токсинами или радиоактивными или нерадиоактивными маркерами, такими как 3H, 123I, 125I, 131I, 32P, 35S, 14C, 51Cr, 36Cl, 57Co, 55Fe, 59Fe, 90Y, 99mTc, 75Se, или антитела, фрагменты или пептиды, ковалентно связанные с флуоресцентными/хемилюминесцентными метками, такими как родамин, флуоресцеин, изотиоцианат, фикоэритрин, фикоцианин, флуорескамин, хелатирующие металлы агенты, авидин, стрептавидин или биотин.
Новые антитела, фрагменты антител, их смеси и производные можно применять непосредственно, после высушивания, например, лиофилизации, после присоединения к упомянутым выше носителям или после изготовления с другими фармацевтически активными и дополнительными веществами для получения фармацевтических препаратов. Примерами активных и дополнительных веществ, которые можно упомянуть, являются другие антитела, противомикробные активные вещества с микробицидным или микробиостатическим действием, такие как антибиотики в целом, или сульфонамиды, противоопухолевые средства, вода, буферы, физиологические растворы, спирты, жиры, воски, инертные носители или другие вещества, общепринятые для парентеральных продуктов, такие как аминокислоты, загустители или сахара. Эти фармацевтические препараты применяют для лечения заболеваний, и они пригодны для стабилизации, снижения и/или профилактики встречаемости по меньшей мере одного симптома глазных связанных с неоваскуляризацией нарушений и заболеваний, включая AMD (экссудативную и/или неэкссудативную) и диабетическую ретинопатию.
Новые антитела, фрагменты антител, их смеси или производные можно применять для лечения или диагностики непосредственно или после присоединения к твердым или жидким носителям, ферментам, токсинам, радиоактивным или нерадиоактивным меткам или с флуоресцентными/хемилюминесцентными метками, как описано выше.
Моноклональные антитела против C5 человека по настоящему изобретению можно получать любыми способами, известными в данной области. Например, млекопитающее иммунизируют посредством C5 человека. Очищенный C5 человека является коммерчески доступным (например, от Quidel Corporation, San Diego, CA или Advanced Research Technologies, San Diego, CA). Альтернативно C5 человека можно легко очищать из плазмы человека. Млекопитающее, используемое для индукции антитела против C5 человека, не ограничивается, и может представлять собой примата, грызуна (такого как мышь, крыса или кролик), жвачных животных, овцу, козу или собаку.
Далее, антителопродуцирующие клетки, такие как клетки селезенки, извлекают из иммунизированного животного и подвергают слиянию с клетками миеломы. Клетки миеломы хорошо известны в данной области (например, можно использовать клетки p3x63-Ag8-653, NS-0, NS-1 или P3U1). Процесс слияния клеток можно проводить любым общепринятым способом, известным в данной области.
Затем клетки, после процесса слияния, культивируют в селективной среде HAT, чтобы отобрать гибридомы. Затем гибридомы, которые продуцируют античеловеческие моноклональные антитела, подвергают скринингу. Этот скрининг можно проводить, например, посредством "сэндвич"-иммуноферментного твердофазного анализа (ELISA) или сходных с ним, в котором продуцированные моноклональные антитела связываются с лунками, на которых иммобилизованы C5 человека. В этом случае, в качестве вторичного антитела, можно использовать антитело, специфичное к иммуноглобулину иммунизированного животного, которое мечено ферментом, таким как пероксидаза, щелочная фосфатаза, глюкозооксидаза, бета-D-галактозидаза или сходные с ними. Метку можно выявлять посредством реакции фермента для мечения с субстратом и измерения образовавшегося цвета. В качестве субстрата, можно получать 3,3-диаминобензидин, 2,2-диамино-бис-о-дианизидин, 4-хлорнафтол, 4-аминоантипирин, о-фенилендиамин или сходные с ними.
Посредством описанного выше процесса, можно отбирать гибридомы, которые продуцируют антитела против C5 человека. Затем отобранные гибридомы клонируют общепринятым способом лимитирующих разведений или способом с мягким агаром. Если желательно, клонированные гибридомы можно крупномасштабно культивировать с использованием содержащей сыворотку или бессывороточной среды, или их можно инокулировать в брюшную полость мыши и выделять из асцитной жидкости, посредством чего можно получать большое количество клонированных гибридом.
Из отобранных моноклональных антител против C5 человека, антитела, которые обладают способностью предотвращать расщепление C5 (например, в клеточной системе анализа C5), затем отбирают для последующего анализа и манипулирования. Если антитело блокирует расщепление C5, это означает, что тестируемое моноклональное антитело обладает способностью снижать или нейтрализовать активность C5 человека. Следовательно, моноклональное антитело специфично распознает и/или препятствует расщеплению и активации C5.
Моноклональные антитела, описанные в настоящем документе, далее включают гибридные и рекомбинантные антитела, полученные сплайсингом вариабельного (включая гипервариабельный) домена антитела против C5 с константным доменом (например, "гуманизированные" антитела), или легкой цепи с тяжелой цепью, или цепи из одного вида с цепью из другого вида, или слиянием с гетерологичными белками, независимо от вида источника или класса иммуноглобулина или обозначения подкласса, а также фрагменты антител [например, Fab, F(ab)2 и Fv], при условии, что они проявляют требуемую биологическую активность [см., например, патент США № 4816567 и Mage & Lamoyi, в Monoclonal Antibody Production Techniques and Applications, pp.79-97 (Marcel Dekker, Inc.), New York (1987)].
Таким образом, термин "моноклональное" указывает на то свойство полученного антитела, что оно получено по существу из гомогенной совокупности антител, и его не следует истолковывать, как требующий продукции антитела каким-либо конкретным способом. Например, моноклональные антитела, подлежащие применению в соответствии с настоящим изобретением, можно получать способом гибридом, впервые описанным Kohler & Milstein, Nature 256:495 (1975), или его можно получать способами рекомбинантных ДНК (патент США № 4816567). Также "моноклональные антитела" можно выделять из фаговых библиотек, созданных с использованием способов, описанных, например, в McCafferty et al., Nature 348:552-554 (1990).
"Гуманизированные" формы нечеловеческих (например, мышиных) антител представляют собой специфичные химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(ab)2 или другие антигенсвязывающие подпоследовательности антител), которые содержат минимальную последовательность, образованную из нечеловеческого иммуноглобулина. В основном, гуманизированные антитела представляют собой человеческие иммуноглобулины (реципиентное антитело), в которых остатки из определяющих комплементарность участков (CDR) реципиентного антитела заменены остатками из CDR не относящегося не человеку вида (донорского антитела), такого как мышь, крыса или кролик, обладающие требуемой специфичностью, аффинностью и емкостью. В некоторых случаях, остатки Fv каркасной области (FR) иммуноглобулина человека заменены соответствующими нечеловеческими остатками FR. Более того, гуманизированное антитело может содержать остатки, которые отсутствуют как в реципиентном антителе, так и в импортных последовательностях CDR или FR. Эти модификации проводят для дальнейшего усовершенствования и оптимизации параметров антитела. Как правило, гуманизированное антитело будет содержать по существу все из по меньшей мере одного, и, как правило, двух, вариабельных доменов, в которых все или по существу все участки CDR соответствуют участкам CDR нечеловеческого иммуноглобулина и все или по существу все остатки FR представляют собой остатки FR консенсусной последовательности иммуноглобулина человека. Гуманизированное антитело также оптимально будет содержать по меньшей мере часть константного участка иммуноглобулина (Fc), как правило, часть константного участка иммуноглобулина человека.
Способы гуманизации антител, не являющихся человеческими, хорошо известны в данной области. Главным образом, гуманизированное антитело обладает одним или несколькими встроенными в него аминокислотными остатками из источника, который не является человеческим. Эти не являющиеся человеческими аминокислотные остатки часто называют "импортными" остатками, которые, как правило, взяты из "импортной" вариабельной области. Гуманизацию, главным образом, можно проводить в соответствии со способом Winter и коллег (Jones et al., (1986) Nature, 321:522-525; Riechmann et al., (1988) Nature, 332:323-327; Verhoeyen et al., (1988) Science, 239:1534-1536), посредством замены CDR или последовательностей CDR грызуна соответствующими последовательностями антитела человека. Таким образом, такие "гуманизированные" антитела представляют собой химерные антитела, где по существу менее чем нетронутый вариабельный человеческий домен замещен соответствующей последовательностью вида, не относящегося к человеку. На практике, гуманизированные антитела, как правило, представляют собой антитела человека, в которых некоторые остатки CDR и возможно некоторые остатки FR замещены остатками из аналогичных участков антител грызунов.
Выбор вариабельных доменов человека как легких, так и тяжелых цепей, для использования при получении гуманизированных антител является очень важным для снижения антигенности. В соответствии с так называемым способом "наилучшего соответствия", последовательность вариабельного домена антитела грызуна подвергают скринингу против целой библиотеки известных последовательностей вариабельных доменов человека. Затем последовательность человека, которая является наиболее сходной с последовательностью грызуна, принимают в качестве каркасной области (FR) для гуманизированного антитела (Sims et al., 1993) J. Immunol, 151:2296; и Chothia and Lesk, (1987) J. Mol. Biol, 196:901). В другом способе используют конкретную каркасную область, полученную из консенсусной последовательности всех антител человека из конкретной подгруппы легких или тяжелых цепей. Одну и ту же каркасную область можно использовать для нескольких различных гуманизированных антител (Carter et al., (1992) Proc. Natl. Acad. Sci USA, 89:4285; Presta et al., (1993) J. Immunol, 151:2623).
Кроме того, важно, чтобы антитела были гуманизированными с сохранением высокой аффинности к антигену и других положительных биологических свойств. Для достижения этой цели, в соответствии с предпочтительным способом, гуманизированные антитела получают с помощью процесса анализа исходных последовательностей и различных предполагаемых гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются широко доступными и известны специалистам в данной области. Доступными являются компьютерные программы, которые иллюстрируют и выводят на экран возможные трехмерные конформационные структуры выбранных последовательностей иммуноглобулинов-кандидатов. Исследование этих выведенных на экран данных позволяет проводить анализ возможной роли остатков в функционировании предполагаемых последовательностей иммуноглобулинов, т.е. анализ остатков, которые влияют на способность иммуноглобулина-кандидата связывать свой антиген. Таким образом, остатки FR можно отобрать и комбинировать из консенсусной и импортной последовательностей таким образом, чтобы были достигнуты требуемые свойства антител, такие как повышенная аффинность к антигену(ам)-мишени. Как правило, непосредственное и наиболее значительное влияние на связывание антигена оказывают остатки CDR.
Человеческие моноклональные антитела, направленные против C5, также относятся к этому изобретению. Такие антитела можно получать способом гибридом. Клеточные линии миеломы человека и гетеромиеломы человека-мыши для продукции человеческих моноклональных антител описаны, например, Kozbor (1984) J. Immunol., 133, 3001; Brodeur, et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987); и Boemer et al., (1991) J. Immunol., 147:86-95.
В настоящее время является возможным получение трансгенных животных (например, мышей), которые способны, при иммунизации, продуцировать полный набор антител человека в отсутствие эндогенной продукции иммуноглобулинов. Например, описано, что гомозиготная делеция в гене участка присоединения тяжелой цепи антитела (J.sub.H) у химерных мышей и мышей из мутантных зародышевых клеток приводит к полному ингибированию эндогенной продукции антитела. Перенос набора генов иммуноглобулинов эмбрионального типа человека в такую мышь из мутантной зародышевой клетки приведет к продукции антител человека при иммунизации антигеном (см., например, Jakobovits et al., (1993) Proc. Natl. Acad. Sci. (USA), 90: 2551; Jakobovits et al., (1993) Nature, 362:255-258; и Bruggermann et al., (1993) Year in Immuno., 7:33).
Альтернативно, для получения человеческих антител и фрагментов антител in vitro из набора генов вариабельных (V) доменов иммуноглобулинов можно использовать технологию фагового дисплея (McCafferty et al., (1990) Nature, 348: 552-553) (для обзора см., например, Johnson et al., (1993) Current Opinion in Structural Biology, 3:564-571). Для фагового дисплея можно использовать несколько источников сегментов V-генов. Например, Clackson et al. ((1991) Nature, 352: 624-628) выделили набор разнообразных антител против оксазолона из небольшой случайной комбинаторной библиотеки V-генов, образованной из селезенок иммунизированных мышей. Можно конструировать набор V-генов из неиммунизированных доноров-людей, и антитела к набору разнообразных антигенов (включая собственные антигены) можно выделять, по существу в соответствии со способами, описанными Marks et al., ((1991) J. Mol. Biol., 222:581-597, или Griffith et al., (1993) EMBO J., 12:725-734).
В природном иммунном ответе, гены антитела накапливают мутации с высокой частотой (соматическая гипермутация). Некоторые из внесенных изменений обеспечат высокую аффинность, и в процессе последующей иммунизации антигеном предпочтительно реплицируются и дифференцируются B-клетки, представляющие высокоаффинный поверхностный иммуноглобулин. Этот природный процесс можно имитировать с использованием способа, известного как "перетасовка цепей" (см. Marks et al., (1992) Bio. Technol., 10:779-783). В этом способе, аффинность "первичных" человеческих антител, полученных посредством фагового дисплея, можно повышать посредством последовательной замены генов V-участка тяжелой и легкой цепей наборами встречающихся в природе вариантов (наборами) генов V-домена, полученными из неиммунизированных доноров. Этот способ обеспечивает продукцию антител и фрагментов антител с аффинностью в нМ-диапазоне. Стратегия получения очень высоких фаговых наборов антител описана Waterhouse et al., ((1993) Nucl. Acids Res., 21:2265-2266).
Перетасовку генов также можно использовать для получения антител человека из антител грызунов, где антитело человека обладает сходной аффинностью и специфичностью с исходным антителом грызуна. В соответствии с этим способом, который также называют "инпринтингом эпитопа", ген V-домена тяжелой или легкой цепи антител грызуна, полученных способом фагового дисплея, заменяют набором генов V-домена человека, создавая химеры грызуна-человека. Селекция по антигену приводит к выделению вариабельного участка человека, способного восстанавливать функциональный антигенсвязывающий участок, т.е. эпитоп определяет (отпечатывает) выбор партнера. Когда процесс повторяют в целях замены оставшегося V-домена грызуна, получают человеческое антитело (см. PCT WO 93/06213, опубликованную 1 апреля 1993 года). В отличие от традиционной гуманизации антитела грызунов пересадкой CDR, этот способ приводит к полностью человеческим антителам, которые не обладают остатками каркасной области или остатками CDR из грызунов.
Примером моноклонального антитела и фрагмента антитела, которые можно использовать в качестве средства против C5 в способах по этому изобретению, является экулизумаб (также известный как SolirisTM, Alexion, Cheshire, CT) и пекселизумаб (Alexion, Cheshire, CT), соответственно, оба из которых описаны в USSN 6355245, включенной в настоящий документ в качестве ссылки в полном объеме.
Также настоящее изобретение относится к применению средств против C5 по этому изобретению с антителами-антагонистами, направленными против PDGF и/или VEGF и распознаваемых ими рецепторов PDGFR и/или VEGFR, соответственно, в способах по этому изобретению для стабилизации, лечения и/или профилактики глазных нарушений. Таким образом, способы, описанные непосредственно выше, можно применять для создания антител-антагонистов по этому изобретению для блокирования связывания лиганда (например, PDGF или VEGF) с его мишенью, такой как распознаваемый им рецептор. Таким образом, антитело-антагонист PDGF по этому изобретению включает антитела, направленные против PDGF, а также против PDGFR-мишени.
Примеры антител-антагонистов, направленных против VEGF, для применения со средствами против C5 в способах по этому изобретению, представляют собой: бевацизумаб (также известный как Avastin®, Genentech, San Francisco, CA), описанный в патенте США № 6054297, включенный в настоящий документ в качестве ссылки в полном объеме; и ранибизумаб (также известный как Lucentis®, Genentech, San Francisco, CA).
Антисмысловые средства против C5 и рибозимы против C5
Средства против C5 по этому изобретению включают антисмысловые олигонуклеотиды и рибозимы, которые нацелены на C5 и осуществляют ингибирование C5 посредством ингибирования трансляции белка с матричной РНК или посредством нацеливания соответствующей мРНК C5 на деградацию. Также предусмотрено применение антисмысловых средств против C5 и средств на основе рибозимов против C5 в способах лечения глазных нарушений. В конкретных вариантах осуществления, настоящее изобретение относится к введению антисмыслового средства против C5 или средства на основе рибозима против C5 субъекту в способе снижения, стабилизации и/или профилактики по меньшей мере одного симптома глазного нарушения, в частности, симптома диабетической ретинопатии, экссудативной и/или неэкссудативной AMD.
Способы разработки и синтеза антисмысловых олигонуклеотидов и рибозимов известны в данной области. Дополнительное руководство представлено в настоящем документе.
Одной проблемой при конструировании специфичных и эффективных нацеленных на мРНК олигонуклеотидов (антисмысловых ODN) и рибозимов, является идентификация доступных участков для антисмыслового образования пар с мРНК-мишени (которая сама по себе сворачивается во вторичную структуру, частично образующую собственные пары). Сочетание компьютерных алгоритмов предсказания доступности образования пар РНК и молекулярного скрининга обеспечивает возможность создания специфичных и эффективных рибозимов и/или антисмысловых олигонуклеотидов, направленных против большинства мРНК-мишеней. Действительно, было описано несколько подходов для определения доступности молекулы РНК-мишени для антисмысловых ингибиторов или ингибиторов-рибозимов. В одном подходе используется скрининговый анализ in vitro, в котором применяется настолько много антисмысловых одигодезоксинуклеотидов, насколько это возможно (см. Monia et al., (1996) Nature Med., 2:668-675; и Milner et al., (1997) Nature Biotechnol., 15:537-541). В другом подходе используются случайные библиотеки ODN (Ho et al., (1996) Nucleic Acids Res., 24:1901-1907; Birikh et al., (1997) RNA 3:429-437; и Lima et al., (1997) J. Biol. Chem., 272:626-638). Доступные участки можно подвергать мониторингу посредством расщепления РНКазой H (см. Birikh et al., supra; и Ho et al., (1998) Nature Biotechnol., 16:59-63). РНКаза H катализирует гидролитическое расщепление фосфодиэфирного остова РНК-цепи дуплекса ДНК-РНК.
Другой подход, вовлекающий применение набора полуслучайных химерных химически синтезированных ODN, используют для идентификации доступных участков, расщепляемых РНКазой H на синтезированной in vitro РНК-мишени. Затем для идентификации этих участков в молекуле-мишени используют анализ удлинения праймера (см. Lima et al., выше). Другие подходы для выявления антисмысловых мишеней в РНК основаны на компьютерных моделях сворачивания РНК. Было опубликовано несколько сообщений, касающихся применения случайных библиотек рибозимов для скрининга эффективного расщепления (см. Campbell et al., (1995) РНК 1:598-609; Lieber et al., (1995) Mol. Cell Biol., 15: 540-551; и Vaish et al., (1997) Biochem., 36:6459-6501).
Другие подходы in vitro, в которых используются случайные или полуслучайные библиотеки ODN и РНКаза H, могут быть более пригодными, чем компьютерное моделирование (Lima et al., выше). Однако применение синтезированной in vitro РНК не предсказывает доступности антисмысловых ODN in vivo, поскольку последние наблюдения позволяют предположить, что на взаимодействия, связанные с ренатурацией полинуклеотидов, влияют РНК-связывающие белки (см. Tsuchihashi et al., (1993) Science, 267:99-102; Portman et al., (1994) EMBO J., 13:213-221; и Bertrand and Rossi (1994) EMBO J., 13:2904-2912). В патенте США № 6562570, содержание которого включено в настоящий документ в качестве ссылки, представлены композиции и способы для определения доступных участков в мРНК в присутствии клеточного экстракта, который имитирует условия in vivo.
В кратком изложении, этот способ вовлекает инкубацию нативных или синтезированных in vitro РНК с определенными антисмысловыми ODN, рибозимами или ДНКзимами, или с библиотекой случайных или полуслучайных ODN, рибозимов или ДНКзимов, в условиях гибридизации в реакционной смеси, которая включает клеточный экстракт, содержащий эндогенные связывающие РНК белки, или которая имитирует клеточный экстракт вследствие наличия одного или нескольких связывающих РНК белков. Любой антисмысловой ODN, рибозим или ДНКзим, который является комплементарным доступному участку в РНК-мишени, будет гибридизоваться с этим участком. Когда используют определенные ODN или библиотеку ODN, РНКаза H присутствует в процессе гибридизации или ее добавляют после гибридизации для расщепления РНК в области, где произошла гибридизация. РНКаза H может присутствовать, когда используют рибозимы или ДНКзимы, но она не является необходимой, поскольку рибозимы и ДНКзимы расщепляют РНК в области, где произошла гибридизация. В некоторых случаях, используют библиотеку случайных или полуслучайных ODN в клеточных экстрактах, содержащих эндогенную мРНК, связывающие РНК белки и РНКазу H.
Далее, различные способы можно использовать для идентификации тех участков на РНК-мишени, с которыми связались антисмысловые ODN, рибозимы или ДНКзимы и в которых произошло расщепление. Например, для этой цели можно использовать зависимую от терминальной дезоксинуклеотидилтрансферазы полимеразную цепную реакцию (TDPCR) (см. Komura and Riggs (1998) Nucleic Acids Res., 26:1807-11). Для превращения матрицы РНК в ДНК используют стадию обратной транскрипции, с последующей TDPCR. В этом изобретении, 3'-концы, необходимые для способа TDPCR, создают обратной транскрипцией представляющей интерес РНК-мишени с любой пригодной РНК-зависимой ДНК-полимеразой (например, обратной транскриптазой). Это осуществляют гибридизацией первого ODN-праймера (PI) с РНК в области, которая расположена ниже (т.е. в направлении от 5' к 3' на молекуле РНК) исследуемой части молекулы РНК-мишени. Полимераза в присутствии dNTP копирует РНК в ДНК с 3'-конца P1 и завершает копирование в области расщепления, созданной любой антисмысловым ODN/РНКазой H, рибозимом или ДНКзимом. Новая молекула ДНК (называемая первой цепью ДНК) служит в качестве первой матрицы для ПЦР-части способа TDPCR, которую используют для идентификации соответствующей доступной последовательности-мишени, находящейся в РНК.
Затем, например, можно использовать способ TDPCR, т.е. обратно транкрибированную ДНК с гуанозинтрифосфатом (rGTP) подвергают реакции в присутствии терминальной дезоксинуклеотидилтрансферазы (TdT) для добавления (rG)2-4-хвоста на 3'-конце молекул ДНК. Затем лигируют двухцепочечный линкер ODN, имеющий 3'2-4-выступающий конец на одной цепи, который образует пары оснований с (rG)2-4-хвостом. Затем добавляют два ПЦР-праймера. Первый праймер представляет собой линкерный праймер (LP), который является комплементарным цепи линкера TDPCR, который лигирован с (rG)2-4-хвостом (иногда называемый нижней цепью). Другой праймер (P2) может быть таким же, как и P1, однако он может быть вложенным относительно P1, т.е. он комплементарен РНК-мишени в области, которая по меньшей мере частично расположена выше (т.е. в направлении от 3' к 5' на молекуле РНК) области, которая связывается P1, но ниже исследуемой части молекулы РНК-мишени. Следовательно, часть молекулы РНК-мишени, исследуемая в целях определения наличия в ней доступных участков связывания, представляет собой часть, которая расположена выше области, которая комплементарна P2. Затем проводят ПЦР известным образом в присутствии ДНК-полимеразы и dNTP для амплификации сегментов ДНК, определяемых праймерами LP и P2. Затем амплифицированный продукт можно извлекать любым известным способом, а затем секвенировать на автоматическом ДНК-секвенаторе, обеспечивая точную идентификацию участка расщепления. После этой идентификации, антисмысловые ДНК или рибозимы с определенной последовательностью можно синтезировать для применения in vitro или in vivo.
Антисмысловое вмешательство в экспрессию конкретных генов можно поводить с помощью применения синтетических антисмысловых олигонуклеотидных последовательностей (см., например, Lefebvre-d'Hellencourt et al., (1995) Eur. Cyokine Netw., 6:7; Agrawal (1996) TIBTECH, 14: 376; и Lev-Lehman et al., (1997) Antisense Therap. Cohen and Smicek, eds. (Plenum Press, New York)). В кратком изложении, антисмысловые олигонуклеотидные последовательности могут представлять собой короткие последовательности ДНК, как правило, 15-30-мерные, однако они могут быть в пределах 7-мера (см. Wagner et al., (1994) Nature, 372: 333), сконструированные, чтобы они были комплементарными представляющей интерес мРНК-мишени и образовывали дуплекс РНК:AS. Это образование дуплекса может препятствовать процессингу, сплайсингу, транспорту или трансляции соответствующей мРНК. Более того, определенные нуклеотидные последовательности AS могут вызывать клеточную активность РНКазы H при гибридизации с их мРНК-мишенью, что приводит к деградации мРНК (см. Calabretta et al., (1996) Semin. Oncol., 23:78). В этом случае, РНКаза H будет расщеплять РНК-компонент дуплекса и может потенциально высвобождать AS для последующей гибридизации с дополнительными молекулами РНК-мишени. Дополнительный способ действия возникает вследствие взаимодействия AS с геномной ДНК с образованием тройной спирали, которая может быть транскрипционно неактивной.
В качестве неограничивающего примера, добавления или замены для антисмысловой последовательности, рассмотренной в настоящем документе выше, для подавления функции гена можно использовать рибозимы. Это особенно необходимо в случаях, где антисмысловая терапия ограничена с точки зрения стехиометрии. Затем можно использовать рибозимы, которые будут нацелены на ту же последовательность. Рибозимы представляют собой молекулы РНК, которые обладают каталитической способностью в отношении РНК, которые расщепляют специфичный участок в РНК-мишени. Количество молекул РНК, которые расщепляются рибозимом, превышает количество, предсказанное по стехиометрии 1:1 (см. Hampel and Tritz (1989) Biochem., 28: 4929-33; и Uhlenbeck (1987) Nature, 328: 596-600). Таким образом, настоящее изобретение также позволяет применять последовательности рибозимов, нацеленные на доступный домен типов мРНК PDGF или VEGF и содержащие соответствующий каталитический центр. Рибозимы получают и доставляют, как известно в данной области и далее рассмотрено в настоящем документе. Рибозимы можно применять в сочетании с антисмысловыми последовательностями.
Рибозимы катализируют расщепление фосфодиэфирной связи РНК. Идентифицировано несколько структурных семейств рибозимов, включая интроны I группы, РНКазу P, рибозим вируса гепатита дельта, рибозимы в виде "головки молотка" и шпилечный рибозим, первоначально образованный из отрицательной цепи сателлитной РНК вируса кольцевой пятнистости табака (sTRSV) (см. Sullivan (1994) Investig. Dermatolog., (Suppl.) 103: 95S; и патент США № 5225347). Последние два семейства образованы из вироидов и вирусоидов, в которых, как полагают, рибозим отделяет мономеры от олигомеров, полученных в процессе репликации по типу "катящегося кольца" (см. Symons (1989) TEBS, 14: 445-50; Symons (1992) Ann. Rev. Biochem., 61: 641-71). Мотивы рибозимов в виде "головки молотка" и шпилек являются наиболее широко адаптированными для трансращепления мРНК в целях генной терапии. Тип рибозима, используемый в настоящем изобретении, выбирают, как известно в данной области. Шпилечные рибозимы в настоящее время проходят клинические испытания и являются особенно пригодным типом. Как правило, длина рибозима составляет 30-100 нуклеотидов.
Наряду с тем, что рибозимы, которые расщепляют мРНК в области последовательностей специфичного распознавания, можно использовать для разрушения конкретных мРНК, применение рибозимов в виде "головки молотка" является особенно пригодным. Рибозимы в виде "головки молотка" расщепляют мРНК в областях, определяемых фланкирующими участками, которые образуют комплементарные пары оснований с мРНК-мишенью. Единственным требованием является то, чтобы мРНК-мишень обладала следующей последовательностью из двух оснований: 5'-UG-3'. Конструирование и продукция рибозимов в виде "головки молотка" хорошо известны в данной области и описаны более подробно в Haseloff and Gerlach ((1988) Nature, 334: 585).
Рибозимы по настоящему изобретению также включают РНК-эндорибонуклеазы (в дальнейшем в настоящем документе "рибозимы Cech-типа"), такие как РНК-эндорибонуклеазы, которые встречаются в природе в Tetrahymena thermophila (известные как РНК IVS, или L-19 IVS), и которые исчерпывающе описаны Thomas Cech и сотрудниками (см. Zaug et al., (1984) Science, 224:574-578; Zaug и Cech (1986) Science, 231:470-475; Zaug, et al., (1986) Nature, 324:429-433; международная патентная заявка № W088/04300; Been and Cech (1986) Cell, 47:207-216). Рибозимы Cech-типа имеют активный участок из восьми пар, который гибридизуется с последовательностью РНК-мишенью, где после этого происходит расщепление РНК-мишени. Настоящее изобретение относится к тем рибозимам Cech-типа, которые нацелены на последовательности активного участка из восьми пар. Несмотря на то, что настоящее изобретение не ограничивается конкретной теорией и используемым механизмом, применение рибозимов в виде "головки молотка" в этом изобретении может обладать преимуществом над применением направленных против PDGF/VEGF антисмысловых молекул, поскольку последние данные показали, что рибозимы в виде "головки молотка" действуют посредством блокирования трансляции РНК и/или специфичного расщепления мРНК-мишени.
Как и в антисмысловом подходе, рибозимы могут состоять из модифицированных олигонуклеотидов (например, для повышенной стабильности, нацеливания и т.д.) и их доставляют к клеткам, экспрессирующим мРНК-мишень. Пригодный способ доставки вовлекает применение конструкции ДНК, "кодирующей" рибозим под контролем сильного конститутивного промотора pol III или pol II, так что трансфицированные клетки будут продуцировать достаточные количества рибозима для разрушения транскриптов-мишеней и ингибирования трансляции. Поскольку рибозимы, в отличие от антисмысловых молекул, являются каталитическими, для их эффективности требуется более низкая внутриклеточная концентрация.
Как описано выше, устойчивость к нуклеазам, при необходимости, обеспечивают любым способом, известным в данной области, который по существу не препятствует биологической активности антисмысловых олигодезоксинуклеотидов или рибозимов, необходимой для способа применения и доставки (Iyer et al., (1990) J. Org. Chem., 55: 4693-99; Eckstein (1985) Ann. Rev. Biochem., 54: 367-402; Spitzer и Eckstein (1988) Nucleic Acids Res., 18: 11691-704; Woolf et al., (1990) Nucleic Acids Res., 18: 1763-69; и Shaw et al., (1991) Nucleic Acids Res., 18: 11691-704). Как описано выше для аптамеров, неограничивающие репрезентативные модификации, которые можно проводить в антисмысловых олигонуклеотидах или рибозимах в целях повышения устойчивости к нуклеазам, включают модификацию гетероатома фосфора или кислорода в фосфатном остове, короткие алкильные или циклоалкильные связи между сахарами или короткие гетероатомные или гетероциклические связи между сахарами. Они включают, например, получение 2'-фторированных, O-метилированных олигомеров, метилфосфонатов, фосфоротиоатов, фосфородитиоатов и морфолиноолигомеров. Например, антисмысловой олигонуклеотид или рибозим может обладать фосфоротиоатными связями, связывающими от четырех до шести 3'-концевых нуклеотидных оснований. Альтернативно фосфоротиоатные связи могут связывать все нуклеотидные основания. Фосфоротиоатные антисмысловые олигонуклеотиды в норме не показывают значительную токсичность в концентрациях, которые являются эффективными и проявляют достаточное фармакодинамическое время полужизни у животных (см. Agarwal et al., (1996) TEBTECH, 14: 376) и являются устойчивыми к нуклеазам. Альтернативно устойчивость к нуклеазам для AS-ODN можно обеспечить наличием петли из 9 нуклеотидов, образующей последовательность на 3'-конце, обладающую нуклеотидной последовательностью CGCGAAGCG. Применение реакции конъюгации авидин-биотин также можно применять для повышения защиты AS-ODN против деградации нуклеазами сыворотки (см. Boado and Pardridge (1992) Bioconj. Chem., 3: 519-23). В соответствии с этой концепцией средства AS-ODN являются монобиотинилированными на их 3'-конце. При реакции с авидином, они образуют прочные устойчивые к нуклеазам комплексы с улучшенной в 6 раз стабильностью по сравнению с неконъюгированными ODN.
В других исследованиях было показано удлинение антисмысловых олигодезоксинуклеотидов in vivo (Agarwal et al., (1991) Proc. Natl. Acad. Sci. (USA) 88: 7595). Этот процесс, предположительно пригодный в качестве механизма выведения для удаления AS-олигонуклеотидов из кровотока, зависит от существования свободных 3'-концов на присоединенных олигонуклеотидах, в которых происходит удлинение. Таким образом, частичный фосфоротиоат, защита петлей или биотин-авидин в этом важном положении должны быть достаточными для обеспечения стабильности этих AS-олигодезоксинуклеотидов.
В дополнение к применению модифицированных оснований, как описано выше, можно получать аналоги нуклеотидов, где структура нуклеотида принципиально изменена и которые лучше подходят в качестве терапевтических или экспериментальных реагентов. Примером нуклеотидного аналога является пептидная нуклеиновая кислота (PNA), где фосфатный остов дезоксирибозы (или рибозы) в ДНК (или РНК) заменен полиамидным остовом, который является сходным с остовом, находящимся в пептидах. Было показано, что PNA-аналоги являются устойчивыми к деградации ферментами и обладают более длительным существованием in vivo и in vitro. Кроме того, было показано, что PNA связываются более прочно с комплементарной последовательностью ДНК, чем молекула ДНК. Это наблюдение приписывают отсутствию отталкивания зарядов между цепью PNA и цепью ДНК. Другие модификации, которые можно проводить в олигонуклеотидах, включают полимерные остовы, морфолино-полимерные остовы (см., например, патент США № 5034506, содержание которого включено в настоящий документ в качестве ссылки), циклические остовы или ациклические остовы, миметики сахаров или любую другую модификацию, включая модификации, которые могут повысить фармакодинамические свойства олигонуклеотида.
Следующий аспект этого изобретения относится к применению ферментов ДНК для повышения экспрессии мРНК-мишени, поскольку, например, ферменты C5 включают некоторые из механистических признаков технологий как антисмысловых молекул, так и рибозимов. ДНК-ферменты конструируют, чтобы они распознавали конкретную последовательность нуклеиновой кислоты-мишени, подобно антисмысловому олигонуклеотиду, однако подобно рибозиму, они являются каталитическими и специфично расщепляют нуклеиновую кислоту-мишень.
В настоящее время существует два основных типа ДНК-ферментов, и оба из них были идентифицированы Santoro и Joyce (см., например, патент США № 6110462). ДНК-фермент 10-23 содержит структуру петли, которая соединяет два плеча. Два плеча обеспечивают специфичность посредством распознаваний конкретной последовательности нуклеиновой кислоты-мишени, в то время как структура петли обеспечивает каталитическую функцию в физиологических условиях.
В кратком изложении, для конструирования ДНК-фермента, который специфично распознает и расщепляет нуклеиновую кислоту-мишень, специалист в данной области должен сначала идентифицировать уникальную последовательность мишени. Это можно проводить с использованием того же подхода, что описан для антисмысловых олигонуклеотидов. В некоторых случаях, уникальная или по существу уникальная последовательность является G/C-богатой и состоит приблизительно из от 18 до 22 нуклеотидов. Высокое содержание G/C помогает обеспечить прочное взаимодействие между ДНК-ферментом и последовательностью-мишенью.
При синтезе ДНК-фермента, специфичную антисмысловую последовательность для распознавания, которая нацеливает фермент на транскрипт, разделяют таким образом, чтобы она содержала два плеча ДНК-фермента, и между двумя специфичными плечами помещают петлю ДНК-фермента.
Способы получения и введения ДНК-ферментов можно найти, например, в патенте США № 6110462. Аналогично, способы доставки ДНК-рибозимов in vitro или in vivo включают способы доставки РНК-рибозима, как описано в настоящем документе. Кроме того, специалисту в данной области понятно, что, подобно антисмысловым олигонуклеотидам, ДНК-ферменты необязательно можно модифицировать для повышения стабильности и повышения устойчивости к деградации.
Также настоящее изобретение относится к применению средств против C5 по этому изобретению с антисмысловыми средствами, средствами на основе рибозима и/или средствами на основе ДНК-фермента, направленными против экспрессии PDGF и/или VEGF, в способах по этому изобретению для стабилизации, лечения и/или профилактики глазных нарушений. Таким образом, способы, описанные непосредственно выше, можно применять для получения антисмысловых средств, средств на основе рибозима и/или средств на основе ДНК-фермента для блокирования или ингибирования экспрессии PDGF и/или VEGF для применения со средствами против C5 по этому изобретению.
RNAi-средства против C5
В некоторых вариантах осуществления этого изобретения применяют материалы и способы для осуществления подавления C5 посредством интерференции РНК (RNAi). Таким образом, средства против C5 по этому изобретению включают RNAi-средства против C5. Настоящее изобретение относится к применению RNAi-средств против C5 в способах лечения глазных нарушений по этому изобретению. В конкретных вариантах осуществления, настоящее изобретение относится к введению RNAi-средства против C5 в способе снижения, стабилизации и/или профилактики по меньшей мере одного симптома глазного нарушения, в частности, симптома диабетической ретинопатии, экссудативной и/или неэкссудативной AMD.
RNAi представляет собой процесс специфичной в отношении последовательности посттрансляционной репрессии гена, который может происходить в эукариотических клетках. Как правило, этот процесс вовлекает деградацию мРНК с конкретной последовательностью, индуцируемую двухцепочечной РНК (dsRNA), которая гомологична этой последовательности. Например, экспрессия длинной dsRNA, соответствующей последовательности конкретной одноцепочечной мРНК (ss мРНК), будет обеспечивать неустойчивость этого транскрипта, препятствуя посредством этого в экспрессии соответствующего гена. Таким образом, можно подавлять любой выбранный ген посредством введения dsRNA, которая соответствует всей или существенной части мРНК этого гена. Оказалось, что при экспрессии длинной dsRNA, она первоначально подвергается процессингу рибонуклеазой III в более короткие олигонуклеотиды dsRNA длиной не более чем от 21 до 22 пар. Таким образом, RNAi можно достигать посредством введения или экспрессии относительно коротких гомологичных dsRNA. Действительно, применение относительно коротких гомологичных dsRNA может обеспечить определенные преимущества, как описано ниже.
Клетки млекопитающих обладают по меньшей мере двумя каскадами, на которые действует двухцепочечная РНК (dsRNA). В каскаде RNAi (специфичном в отношении последовательности), инициирующая dsRNA первоначально разрушается на короткие интерферирующие (si) РНК, как описано выше. siRNA обладают смысловыми и антисмысловыми цепями приблизительно из 21 нуклеотида, которые образуют si РНК приблизительно из 19 нуклеотидов с выступающими концами из двух нуклеотидов на каждом 3'-конце. Полагают, что короткие интерферирующие РНК обеспечивают информацию о последовательности, которая обеспечивает нацеливание определенной матричной РНК на деградацию. Напротив, неспецифичный каскад запускается dsRNA с любой последовательностью, при условии, что она обладает длиной по меньшей мере приблизительно 30 пар оснований. Неспецифичные эффекты происходят вследствие того, что dsRNA активирует два фермента: PKR (двухцепочечная РНК-активируемая протеинкиназа), которая в ее активной форме фосфорилирует фактор инициации трансляции eIF2, останавливая весь белковый синтез, и 2',5'-олигоаденилатсинтетаза (2',5'-AS), которая синтезирует молекулу, которая активирует РНКазу L, неспецифичный фермент, который нацелен на все мРНК. Неспецифичный каскад может представлять собой ответ хозяина на стресс или вирусную инфекцию, и, как правило, в особенно пригодных способах по настоящему изобретению эффекты неспецифичного каскада минимизируют. По существу, более длинные dsRNA по-видимому требуются для индукции неспецифичного каскада и, таким образом, dsRNA короче приблизительно 30 пар оснований, являются особенно пригодными для достижения репрессии гена посредством RNAi (см., например, Hunter et al., (1975) J. Biol. Chem., 250: 409-17; Manche et al., (1992) Mol. Cell Biol., 12: 5239-48; Minks et al., (1979) J. Biol. Chem., 254: 10180-3; и Elbashir et al., (2001) Nature, 411: 494-8).
Определенные двухцепочечные олигонуклеотиды, используемые для достижения RNAi, обладают длиной менее 30 пар оснований и могут содержать приблизительно 25, 24, 23, 22, 21, 20, 19, 18 или 17 пар оснований рибонуклеиновой кислоты. Необязательно, олигонуклеотиды dsRNA по этому изобретению могут включать 3'-выступающие концы. Неограничивающие иллюстративные 2-нуклеотидные 3'-выступающие концы могут состоять из рибонуклеотидных остатков любого типа и могут даже состоять из остатков 2'-дезокситимидина, которые снижают стоимость синтеза РНК и могут повысить устойчивость к нуклеазам siRNA в клеточной культуральной среде и в трансфицированных клетках (см. Elbashi et al., (2001) Nature, 411: 494-8).
Более длинные dsRNA из 50, 75, 100 или даже 500 пар оснований или более также можно применять в некоторых вариантах осуществления этого изобретения. Иллюстративные концентрации dsRNA для достижения RNAi составляют приблизительно 0,05 нМ, 0,1 нМ, 0,5 нМ, 1,0 нМ, 1,5 нМ, 25 нМ или 100 нМ, хотя можно использовать другие концентрации, в зависимости от природы обрабатываемых клеток, гена-мишени и других факторов, легко определяемых квалифицированным специалистом. Иллюстративные dsRNA можно синтезировать химически или получать in vitro или in vivo с использованием соответствующих экспрессирующих векторов. Иллюстративные синтетические РНК включают РНК из 21 нуклеотида, химически синтезированные с использованием способов, известных в данной области (например, фосформаидиты РНК и тимидинфосфорамидит Expedite (Proligo, Germany)). Из синтетических олигонуклеотидов можно удалять защитные группы и их можно очищать в геле с использованием способов, известных в данной области (см., например, Elbashir et al., (2001) Genes Dev., 15: 188-200). Более длинные РНК можно транскрибировать с промоторов, таких как промоторы РНК-полимеразы T7, известные в данной области. Одна РНК-мишень, помещенная в обеих возможных ориентациях ниже промотора in vitro, будет транскрибировать обе цепи мишени для создания олигонуклеотида dsRNA с требуемой последовательностью мишени.
Конкретная последовательность, используемая для конструирования олигонуклеотидов, может представлять собой любую непрерывную последовательность нуклеотидов, находящуюся в экспрессируемом транскрипте гена мишени (например, C5). Для выбора пригодных последовательностей-мишеней можно использовать программы и алгоритмы, известные в данной области. Кроме того, можно выбирать оптимальные последовательности, как описано дополнительно выше, с использованием программ, созданных для предсказания вторичной структуры указанной одноцепочечной последовательности нуклеиновой кислоты и обеспечения отбора тех последовательностей, которые вероятно встретятся в экспонированных одноцепочечных областях свернутой мРНК. Способы и композиции конструирования пригодных олигонуклеотидов можно найти, например, в патенте США № 6251588, содержание которого включено в настоящий документ в качестве ссылки. Как правило, полагают, что мРНК является линейной молекулой, которая содержит информацию для направления синтеза белка в последовательности рибонуклеотидов. Однако исследования выявили, что в большинстве мРНК существует множество вторичных и третичных структур. Элементы вторичной структуры в РНК образуются, главным образом, посредством взаимодействий по типу Уотсона-Крика между различными участками одной молекулы РНК. Важные вторичные структурные элементы включают внутримолекулярные двухцепочечные участки, шпилечные петли, выпячивание в дуплексной РНК и внутренние петли. Третичные структурные элементы образуются, когда вторичные структурные элементы контактируют друг с другом или с одноцепочечными участками с образованием более комплексной трехмерной структуры. Ряд исследователей измеряли энергии связывания большого количества дуплексных структур РНК и вывели набор правил, которые можно применять для предсказания вторичной структуры РНК (см., например, Jaeger et al., (1989) Proc. Natl. Acad. Sci. (USA) 86:7706 (1989); и Turner et al., (1988) Ann. Rev. Biophys. Biophys. Chem., 17:167). Правила пригодны для идентификации структурных элементов РНК и, в частности, для идентификации одноцепочечных участков РНК, которые могут представлять собой особенно пригодные сегменты мРНК для нацеливания технологий подавления посредством RNAi, рибозима или антсмысловых молекул. Таким образом, можно идентифицировать конкретные сегменты мРНК-мишени для конструирования опосредующих RNAi олигонуклеотидов dsRNA, а также для конструирования соответствующих композиций рибозима и рибозима в виде "головки молотка" по этому изобретению.
Олигонуклеотиды dsRNA можно вводить в клетку посредством трансфекции с гетерологичным геном-мишненью с использованием композиций носителя, таких как липосомы, которые известны в данной области, например, Lipofectamin 2000 (Life Technologies, Rockville Md.), как описано изготовителем для прикрепленных клеточных линий. Трансфекцию олигонуклеотидов dsRNA для нацеливания на эндогенные гены можно проводить с использованием Oligofectamine (Life Technologies). Эффективность трансфекции можно проверять с использованием флуоресцентной микроскопии для клеточных линий млекопитающих после котрансфекции hGFP, кодирующего pAD3 (Kehlenback et al., (1998) J. Cell. Biol., 141: 863-74). Эффективность RNAi после введения dsRNA можно оценивать любым из множества способов анализа. Они включают, но не ограничиваются ими, Вестерн-блот анализ с использованием антител, которые распознают продукт гена-мишени после достаточного времени для обновления эндогенного пула после репрессии синтеза нового белка, и Нозерн-блот анализ для определения уровня существующей мРНК-мишени.
Дополнительные композиции, способы и применение технологии RNAi для использования в этом изобретении представлены в патентах США № 6278039, 5723750 и 5244805, которые включены в настоящий документ в качестве ссылок.
Также настоящее изобретение относится к применению средства против C5 по этому изобретению со средствами RNAi для репрессии PDGF и/или VEGF в способах по этому изобретению для стабилизации, лечения и/или профилактики глазных нарушений. Таким образом, способы, описанные непосредственно выше, можно применять для создания средств RNAi в целях подавления PDGF и/или VEGF для применения со средствам против C5 по этому изобретению.
Белковые и полипептидные средства против C5
В некоторых вариантах осуществления этого изобретения, средство против C5 представляет собой белок или полипептид. Настоящее изобретение относится к применению белкового или полипептидного средства против C5 в способах лечения глазных нарушений. В конкретных вариантах осуществления, настоящее изобретение относится к введению белкового или полипептидного средства против C5 субъекту в способе снижения, стабилизации и/или профилактики по меньшей мере одного симптома глазного нарушения, в частности, симптома диабетической ретинопатии, экссудативной и/или неэкссудативной AMD.
Например, в способах по этому изобретению можно применять TP10 (Avant Immunotherapeutics, Inc. Needham, MA) растворимый укороченный рецептор для комплемента 1 типа и белковые или полипептидные средства против C5, описанные, например, в патентах США № 5212071, 5252216, 5256642, 5456909, 5472939, 5840858, 5856297, 5858969, 5981481, 6057131, 6169068 и 6316604, все из которых включены в настоящий документ в качестве ссылок в полном объеме. В способах по этому изобретению можно применять APT070 (также известный как Mirococept®, Inflazyme Pharmaceuticals, LTD., Richmond, B.C. Canada).
Также настоящее изобретение относится к применению средств против C5 по этому изобретению с белковыми и/или полипептидными средствами против PDGF и/или против VEGF в способах по этому изобретению для стабилизации, лечения и/или профилактики глазных нарушений.
Низкомолекулярные средства против C5
В некоторых вариантах осуществления этого изобретения, средство против C5 представляет собой низкомолекулярное соединение, в частности, низкомолекулярное органическое соединение. Настоящее изобретение относится к применению низкомолекулярных средств против C5 в способах лечения глазных нарушений. В конкретных вариантах осуществления, настоящее изобретение относится к введению низкомолекулярного средства против C5 субъекту в способе снижения, стабилизации и/или профилактики по меньшей мере одного симптома глазного нарушения, в частности, симптома диабетической ретинопатии, экссудативной и/или неэкссудативной AMD.
Также настоящее изобретение относится к применению средств против C5 по этому изобретению с низкомолекулярными средствами против PDGF и/или против VEGF в способах по этому изобретению для стабилизации, лечения и/или профилактики глазных нарушений. Например, средство против C5 по этому изобретению можно применять со средством против PDGF иматиниба мезилатом (GleeveB®, Novartis Pharmaceuticals, Inc. East Hanover, NJ). Средство против C5 по этому изобретению также можно применять со средством против VEGF, таким как сорафениб (Nexavar® Onyx Pharmaceuticals, Inc.Emeryville, CA и Bayer Pharmaceuticals Corportion, West Haven, CT); сунитнаба малат (Sutent®, Pfizer, Inc. NY, NY).
Аптамеры против C3
В некоторых вариантах осуществления, материалы по настоящему изобретению содержат серию нуклеиновых кислот-аптамеров, которые связываются с высокой специфичностью с белком C3 комплемента и которые функционально модулируют, например блокируют, активность белка C3 комплемента в анализах in vivo и/или в клеточных анализах. Эти аптамеры обеспечивают низко токсичный, безопасный и эффективный способ лечения, стабилизации и/или профилактики различных связанных с комплементом глазных заболеваний или нарушений в способах по этому изобретению, включающих, например, острое или хроническое воспалительное и/или опосредуемое иммунной системой глазное нарушение, воспалительный конъюнктивит, включая аллергический и гигантский папиллярный конъюнктивит, отек желтого пятна, увеит, эндофтальмит, склерит, язвы роговицы, сухой кератит, глаукому, ишемическое заболевание сетчатки, отторжение трансплантата роговицы, осложнения, связанные с внутриглазной хирургической операцией, такой как внутриглазная имплантация искусственной линзы, и воспаление, обусловленное хирургической операцией по поводу катаракты, болезнь Бехчета, иммунокомплексный васкулит, болезнь Фуха, болезнь Фогта-Каянаги-Харада, субретинальный фиброз, кератит, витреоретинальное воспаление, паразитическое заражение/миграцию в глазу, пигментный ретинит, цитомегаловирусный ретинит и хориоидальное воспаление, дегенерацию желтого пятна, возрастную дегенерацию желтого пятна ("AMD"), AMD неэкссудативного ("сухого") типа, или глазное нарушение, связанное с неоваскуляризацией, включая диабетическую ретинопатию или AMD экссудативного ("влажного") типа. Эти аптамеры также можно использовать в офтальмологической диагностике.
Эти аптамеры для применения в способах по этому изобретению могут включать модификации, как описано в настоящем документе, включая, например, конъюгацию с липофильными или высокомолекулярными соединениями (например, PEG), встраивание группы кэпа, встраивание модифицированных нуклеотидов и модификации фосфатного остова.
В одном варианте осуществления, предусмотрен выделенный не встречающийся в природе аптамер, который связывается с белком C3 комплемента для применения в способах по этому изобретению для лечения, стабилизации и/или профилактики опосредуемого комплементом глазного нарушения. В некоторых вариантах осуществления, выделенный не встречающийся в природе аптамер для применения в способах по этому изобретению имеет константу диссоциации ("KD") в отношении белка C3 комплемента менее 100 мкМ, менее 1 мкМ, менее 500 нМ, менее 100 нМ, менее 50 нМ, менее 1 нМ, менее 500 пМ, менее 100 пМ, менее 50 пМ. В некоторых вариантах осуществления этого изобретения, константу диссоциации определяют дот-блот титрованием, как описано в примере 2, ниже.
В другом варианте осуществления, аптамеры для применения в способах по этому изобретению модулируют функцию белка C3 комплемента, в частности ингибируют функцию белка C3 комплемента и/или функцию варианта белка C3 комплемента. Как используют в настоящем документе, вариант белка C3 комплемента, включает варианты, которые выполняют по существу ту же функцию, что и функция белка C3 комплемента. Вариант белка C3 комплемента предпочтительно включает по существу ту же структуру и в некоторых вариантах осуществления обладает по меньшей мере 80% идентичностью последовательности, более предпочтительно по меньшей мере 90% идентичностью последовательности, и более предпочтительно по меньшей мере 95% идентичностью последовательности с аминокислотной последовательностью белка C3 комплемента, содержащего аминокислотную последовательность, указанную в De Bruijn, MH and Fey, GH (1985) Human complement component C3: cDNA coding sequence and derived primary structure. Proc Natl Acad Sci USA 82, 708-12.
Другие аптамеры по этому изобретению, которые связывают белок C3 комплемента, далее описаны в патентных заявках США с серийными номерами 6140490, 6395888 и 6566343, все из которых включены в настоящий документ в качестве ссылок в полном объеме.
Аптамеры против C1q
В некоторых вариантах осуществления, материалы по настоящему изобретению содержат серию нуклеиновых кислот-аптамеров, которые связываются с высокой специфичностью с белком C1q комплемента и которые функционально модулируют, например блокируют, активность белка C1q комплемента в анализах in vivo и/или в клеточных анализах. Эти аптамеры обеспечивают низкотоксичный, безопасный и эффективный способ лечения, стабилизации и/или профилактики множества связанных с комплементом глазных заболеваний или нарушений в способах по этому изобретению, включая, например, острое или хроническое воспалительное и/или опосредуемое иммунной системой глазное нарушение, воспалительный конъюнктивит, включая аллергический и гигантский папиллярный конъюнктивит, отек желтого пятна, увеит, эндофтальмит, склерит, язвы роговицы, сухой кератит, глаукому, ишемическое заболевание сетчатки, отторжение трансплантата роговицы, осложнения, связанные с внутриглазной хирургической операцией, такой как внутриглазная имплантация линзы, и воспаление, обусловленное хирургической операцией по поводу катаракты, болезнь Бехчета, иммунокомплексный васкулит, болезнь Фуха, болезнь Фогта-Каянаги-Харада, субретинальный фиброз, кератит, витреоретинальное воспаление, паразитическое заражение/миграцию в глазу, пигментный ретинит, цитомегаловирусный ретинит и хориоидальное воспаление, дегенерацию желтого пятна, возрастную дегенерацию желтого пятна ("AMD"), AMD неэкссудативного ("сухого") типа, или глазное нарушение, связанное с неоваскуляризацией, включая диабетическую ретинопатию или AMD экссудативного ("влажного") типа. Эти аптамеры также можно применять в офтальмологической диагностике.
Эти аптамеры для применения в способах по этому изобретению могут включать модификации, как описано в настоящем документе, включая, например, конъюгацию с липофильными или высокомолекулярными соединениями (например, PEG), встраивание кэпирующей группы, встраивание модифицированных нуклеотидов и модификации фосфатного остова.
В одном варианте осуществления, предусмотрен выделенный не встречающийся в природе аптамер, который связывается с белком C1q комплемента для применения в способах по этому изобретению в целях лечения, стабилизации и/или профилактики опосредуемого комплементом глазного нарушения. В некоторых вариантах осуществления, выделенный не встречающийся в природе аптамер для применения в способах по этому изобретению имеет константу диссоциации ("KD") в отношении белка комплемента C1q менее 100 мкМ, менее 1 мкМ, менее 500 нМ, менее 100 нМ, менее 50 нМ, менее 1 нМ, менее 500 пМ, менее 100 пМ, менее 50 пМ. В некоторых вариантах осуществления этого изобретения, константу диссоциации определяют дот-блот титрованием, как описано в примере 2, ниже.
В другом варианте осуществления, аптамеры для применения в способах по этому изобретению модулируют функцию белка C1q комплемента, в частности ингибируют функцию белка C1q комплемента и/или функцию варианта белка C1q комплемента. Как используют в настоящем документе, вариант белка C1q комплемента включает варианты, которые выполняют по существу такую же функцию, что и функция белка C1q комплемента. Вариант белка C1q комплемента предпочтительно включает по существу ту же структуру и в некоторых вариантах осуществления обладает по меньшей мере 80% идентичностью последовательности, более предпочтительно по меньшей мере 90% идентичностью последовательности, и более предпочтительно по меньшей мере 95% идентичностью последовательности с аминокислотной последовательностью белка C1q комплемента, содержащего аминокислотную последовательность, указанную в Sellar, GC, Blake, DJ и Reid, KB (1991) Characterization and organization of the genes encoding the A-, B- and C-chains of human complement subcomponent C1q. The complete derived amino acid cequence of human C1q. Biochem J. 274, 481-90.
Другие аптамеры по этому изобретению, которые связывают белок C1q комплемента, далее описаны в патентных заявках США с серийными номерами 6140490, 6395888 и 6566343, все из которых включены в настоящий документ в качестве ссылок в полном объеме.
В некоторых вариантах осуществления лекарственные средства на основе аптамера, включая средства против C5, C3 и/или C1q по настоящему изобретению, обладают значительной аффинностью и высокой специфичностью к их мишеням, при снижении вредных побочных эффектов не встречающихся в природе нуклеотидных замен, если лекарственные средства на основе аптамера разрушаются в организме пациентов или субъектов.
Аптамеры против комплемента по настоящему изобретению, включая аптамеры против C5, C3 и/или C1q по этому изобретению, можно синтезировать с использованием любого способа синтеза олигонуклеотидов, известного в данной области, включая способы твердофазного синтеза олигонуклеотидов, хорошо известные в данной области (см., например, Froehler et al., Nucl. Acid Res. 14:5399-5467 (1986) и Froehler et al., Tet. Lett.27:5575-5578 (1986)) и жидкофазные способы, такие как триэфирные способы синтеза (см., например, Sood et al., Nucl. Acid Res. 4:2557 (1977) и Hirose et al., Tet. Lett., 28:2449 (1978)).
Также настоящее изобретение относится к применению аптамеров против комплемента по этому изобретению (включая аптамеры против C5, C3 и/или C1q) с аптамерами к PDGF и/или VEGF и/или распознаваемым ими рецепторами PDGFR и VEGFR, соответственно в способах по этому изобретению для стабилизации, лечения и/или профилактики глазных нарушений.
Примеры аптамеров против PDGF для применения в способах по этому изобретению описаны в международной патентной заявке № PCT/US2005/039975, поданной 2 ноября 2005 года и включенной в настоящий документ в качестве ссылки в полном объеме, в частности ARC513, ARC594, ARC127 и ARC404, описанные в настоящем документе.
Примеры специфичных к VEGF аптамеров для применения в способах по этому изобретению описаны в патентах США №№ 5919455, 5932462, 6113906, 6011020, 6051698 и 6147204. Например, особенно пригодным аптамером для применения при лечении глазных нарушений в сочетании с аптамером против комплемента, в частности с аптамером против C5 по этому изобретению, является EYE001 (ранее NX1838) в его пегилированной и непегилированной форме, в частности инъекция пегаптаниба натрия (Macugen®, Eyetech Pharmaceuticals, Inc. и Pfizer, Inc. NY, NY).
Фармацевтические композиции
Также настоящее изобретение относится к фармацевтическим композициям, содержащим средство против C5, в частности молекулы аптамеров, которые связываются белком C5 комплемента, в частности аптамер, который связывается с белком C5 комплемента и препятствует его расщеплению. В некоторых вариантах осуществления, композиции пригодны для внутреннего применения и включают эффективное количество фармакологически активного соединения по этому изобретению, отдельно или в сочетании, с одним или несколькими фармацевтически приемлемыми носителями. Соединения особенно пригодны, поскольку они обладают очень низкой, если обладают ей, токсичностью.
Композиции по этому изобретению можно применять для лечения или профилактики патологии, такой как заболевание или нарушение, или смягчения симптомов такого заболевания или нарушения у пациента. Например, композиции по настоящему изобретению можно применять для лечения или профилактики патологии, ассоциированной со связанными с комплементом нарушениями сердца (например, повреждением миокарда; опосредуемыми компонентом C5 комплемента осложнениями, связанными с хирургической операцией по аортокоронарному шунтированию (CABG), такими как послеперационное кровотечение, системная активация нейтрофилов и лейкоцитов, повышенный риск инфаркта миокарда и повышенная когнитивная дисфункция; рестенозом; и опосредуемыми C5 осложнениями, связанными с чрескожным вмешательством на коронарных сосудах); повреждением при ишеми-реперфузии (например, инфарктом миокарда, инсультом, отморожением); связанными с комплементом воспалительными нарушениями (например, астмой, артритом, сепсисом и отторжением после трансплантации органа); и связанными с комплементом аутоиммунными нарушениями (например, миастенией, системной красной волчанкой (SLE, или волчанкой); воспалением легких; экстракорпоральной активацией комплемента; опосредуемой антителом активацией комплемента; и опосредуемыми комплементом офтальмологическими показаниями, такими как связанные с неоваскуляризацией глаза нарушения, в частности диабетическая ретинопатия и связанная со старением дегенерация желтого пятна (AMD). В конкретном варианте осуществления, композиции по настоящему изобретению применяют для снижения, стабилизации и/или профилактики симптома опосредуемого C5 глазного нарушения, в частности диабетической ретинопатии, экссудативной и/или неэкссудативной AMD.
В некоторых вариантах осуществления, композиции по этому изобретению можно применять для стабилизации, лечения и/или профилактики патологии, такой как глазное заболевание или нарушение, у пациента. Например, композиции по настоящему изобретению можно применять для стабилизации, лечения и/или профилактики патологии, ассоциированной со связанными с комплементом глазными нарушениями, такими как: острое или хроническое воспаление и/или опосредуемое иммунной системой глазное нарушение, воспалительный конъюнктивит, включая аллергический и гигантский папиллярный конъюнктивит, отек желтого пятна, увеит, эндофтальмит, склерит, язвы роговицы, сухой кератит, глаукома, ишемическое заболевание сетчатки, отторжение трансплантата роговицы, осложнение, связанное с внутриглазной хирургической операцией, такой как внутриглазная имплантация искусственной линзы, и воспаление, обусловленное хирургической операцией по поводу катаракты, болезнь Бехчета, иммунокомплексный васкулит, болезнь Фуха, болезни Фогта-Каянаги-Харада, субретинальный фиброз, кератит, витреоретинальное воспаление, паразитическое заражение/миграция в глазу, пигментный ретинит, цитомегаловирусный ретинит и хориоидальное воспаление, дегенерация желтого пятна, возрастная дегенерация желтого пятна ("AMD"), AMD неэкссудативного ("сухого") типа, или глазное нарушение, связанное с неоваскуляризацией, включая диабетическую ретинопатию или AMD экссудативного ("влажного") типа.
Композиции по этому изобретению пригодны для введения субъекту, страдающему, или предрасположенному к нему, заболеванием или нарушением, которое связано или возникает вследствие белка C5 комплемента, который ингибирует средства против C5 по этому изобретению или с которым средства против C5 по этому изобретению специфично связываются. В некоторых вариантах осуществления, композиции по этому изобретению, в частности, пригодны для ведения субъекту, страдающему, или предрасположенному к нему, заболеванием или нарушением глаз, которое связано или возникает вследствие белка комплемента, который ингибирует аптамеры против комплементы по этому изобретению и/или с которым аптамеры против комплемента по этому изобретению связываются с высокой специфичностью.
В некоторых вариантах осуществления, предусмотрены композиции для лечения субъектов, имеющих опосредуемое комплементом глазное нарушение или предрасположенных к нему. В конкретных вариантах осуществления, предусмотрены композиции для лечения субъектов, страдающих или обладающих риском опосредуемого комплементом глазного нарушения, в частности AMD неэкссудативного типа и/или глазного нарушения, связанного с неоваскуляризацией, в частности, диабетической ретинопатиии и AMD экссудативного типа. В некоторых вариантах осуществления, обладающие риском субъекты представляют собой субъекты, обладающие друзами и/или изменениями пигментации сетчатки, но не обладающие клиническим снижением остроты зрения. Друзы, как правило, выявляют с использованием офтальмоскопа, где они выглядят в виде желтых пятен и частиц против красного фона сетчатки. Клиническое снижение остроты зрения представляет собой демонстрацию снижения зрения на от 1 до 3 строк с использованием карты Early Treatment for Diabetic Retinopathy Study Chart ("карта ETDRS"). Другие изменения зрения, ассоциированные с дегенерацей желтого пятна, включают искажение и/или слепые пятна (скотому), выявляемые с использованием сетки Amsler, изменения адаптации к темноте (диагностический признак состояния здоровья палочковидных клеток) или изменение интерпретации цвета (диагностический признак состояния здоровья колбочковидных клеток). В некоторых вариантах осуществления, субъекты, обладающие риском, представляют собой субъекты, имеющие варианты фактора комплемента H субъекта по сравнению с диким типом. См., например, варианты, описанные Edwards et al., Science vol 308, pp421-422 (2005), Hageman, G. et al., PNAS, vol.102, pp. 7227-7231 (2005), и Haines, J. et al., Science, vol. 308, pp 419-421 (2005). В некоторых вариантах осуществления субъекты, обладающие риском, представляют собой субъектов, имеющих сочетание друз, отсутствия снижения остроты зрения и варианта фактора комплемента H. В некоторых вариантах осуществления, субъекты, обладающие риском, представляют собой субъекты, у которых выявлены друзы. В некоторых вариантах осуществления, субъекты, обладающие риском, представляют собой субъекты, у которых выявлены друзы и имеется клиническое снижение остроты зрения и/или другие изменения зрения.
Композиции по этому изобретению можно применять в способе лечения пациента или субъекта, обладающего патологией, которая, в некоторых предпочтительных вариантах осуществления, представляет собой глазную патологию. Способы по этому изобретению вовлекают введение пациенту или субъекту средства против C5, в частности специфичного к C5 аптамера или композиции, содержащей его, чтобы средство против C5 связывалось с белком C5 комплементом, так чтобы связывание белка C5 комплемента изменяло его биологическую функцию, например, посредством предотвращения его расщепления in vivo, излечивая, посредством этого, опосредуемую C5 патологию. В конкретных вариантах осуществления, связывание средства против C5 по этому изобретению, в частности специфичного к C5 аптамера по этому изобретению, снижает уровень экспрессии VEGF и/или PDGF и/или bFGF и/или других факторов роста, которые стимулируют рост эндотелиальных клеток, в частности, в ткани сетчатки, клеток RPE, сосудов сосудистой оболочки и/или капилляров сетчатки, у пациента, обеспечивая посредством этого излечение опосредуемых VEGF и/или PDGF нарушений, в частности, связанных с неоваскуляризацией глаза нарушений, таких как AMD и/или диабетическая ретинопатия.
В одном варианте осуществления, аптамер против комплемента по этому изобретению, в частности аптамер против C5 по этому изобретению, вводят в глаз или около глаза, субъекту в количестве, достаточном для снижения уровня экспрессии VEGF и/или PDGF в глазу in vivo. В конкретном варианте осуществления способов по этому изобретению, субъект, которому вводят аптамер против комплемента, в частности аптамер против C5 по этому изобретению, идентифицирован как страдающий или обладающий риском связанного с неоваскуляризацией глаза нарушения, при котором сниженная экспрессия VEGF и/или PDGF способствует профилактике, стабилизации и/или снижению по меньшей мере одного симптома глазного нарушения, связанного с неоваскуляризацией.
Пациент или субъект, обладающий глазной патологией, т.е. пациент или субъект, которого лечат способами по этому изобретению, может представлять собой позвоночного, более конкретно млекопитающего, или, более конкретно, человека.
На практике средства против C5 по этому изобретению, в частности специфичные к C5 аптамеры по этому изобретению или их фармацевтически приемлемые соли или пролекарства, вводят в количествах, которые будут достаточными для проявления их требуемой биологической активности, например, ингибирования связывания мишени аптамера с его рецептором, предотвращения расщепления белка-мишени.
Один аспект этого изобретения включает композицию аптамера по этому изобретению в сочетании с другими способами лечения опосредуемых компонентом C5 комплемента нарушений. В одном варианте осуществления настоящее изобретение относится к композиции аптамера по этому изобретению в сочетании с другими лекарственными средствами против опосредуемых комплементом глазных нарушений. Композиция аптамера по этому изобретению может содержать, например, более одного аптамера. В некоторых примерах, композицию аптамера по этому изобретению, содержащую одно или несколько соединений по этому изобретению, вводят в сочетании с другой пригодной композицией, такой как противовоспалительное средство, иммунодепрессант, противовирусное средство или сходные с ними. Более того, соединения по этому изобретению можно вводить в сочетании с цитотоксическим, цитостатическим или химиотерапевтическим средством, таким как алкилирующее средство, антиметаболит, ингибитор митоза или цитотоксический антибиотик, как описано выше. В конкретных вариантах осуществления, пригодным будет средство против C5 по этому изобретению, такое как, главным образом, доступные в настоящее время дозированные формы известных лекарственных средств для применения в таких сочетаниях.
"Комбинированная терапия" (или "котерапия") включает введение средства против C5 по этому изобретению, в частности, композиции специфичного к C5 аптамера по этому изобретению и по меньшей мере второго средства в качестве части определенной схемы лечения, предназначенной для обеспечения благоприятного эффекта вследствие совместного действия этих терапевтических средств. Благоприятный эффект сочетания включает, но не ограничивается ими, фармакокинетическое или фармакодинамическое совместное действие вследствие сочетания терапевтических средств. Введение этих терапевтических средств в сочетании, как правило, проводят в течение определенного периода времени (как правило, в течение некоторого количества минут, часов, суток или недель, в зависимости от выбранного сочетания). В некоторых вариантах осуществления, второе средство может представлять собой средство против VEGF и/или средство против PDGF.
В вариантах осуществления описанных выше способов, где способ дополнительно включает стадию введения субъекту средства против VEGF, средство против VEGF может быть выбрано из группы, состоящей из: молекулы нуклеиновой кислоты, аптамера, антисмысловой молекулы, молекулы RNAi, белка, пептида, циклического пептида, антитела или фрагмента антитела, сахара, полимера и низкомолекулярного соединения.
В вариантах осуществления описанных выше способов, где способ дополнительно включает стадию введения субъекту средства против PDGF, средство против PDGF может быть выбрано из группы, состоящей из: молекулы нуклеиновой кислоты, аптамера, антисмысловой молекулы, молекулы RNAi, белка, пептида, циклического пептида, антитела или фрагмента антитела, сахара, полимера и низкомолекулярного соединения.
В некоторых вариантах осуществления описанных выше способов, где способ дополнительно включает введение субъекту средства против образования сосудов, средство против образования сосудов представляет собой производное порфирина. В некоторых вариантах осуществления производное порфирина представляет собой вертепорфин для инъекции (Visudyne®, Novartis Pharmaceuticals Corporation, East Hanover, NJ). В некоторых вариантах осуществления, способ дополнительно включает стадию активации производного порфирина лазерным излучением.
"Комбинированная терапия" может предполагать, но, как правило, не предполагает, включение введения двух или более из этих терапевтических средств в качестве части отдельных схем монотерапии, которые случайно и произвольно приводят к сочетаниям по настоящему изобретению. Подразумевают, что "комбинированная терапия" охватывает введение этих терапевтических средств последовательно, а именно когда терапевтические средства вводят в различное время, а также введение этих терапевтических средств, или по меньшей мере двух терапевтических средств, по существу одновременно. По существу одновременное введение можно проводить, например, посредством введения субъекту одной капсулы, обладающей фиксированным соотношением каждого терапевтического средства, или в нескольких отдельных капсулах для каждого из терапевтических средств. В другом варианте осуществления, по существу одновременное введение можно проводить, например, посредством введения субъекту одного шприца, обладающего фиксированным соотношением каждого терапевтического средства, или в нескольких отдельных капсулах для каждого из терапевтических средств.
Последовательное или по существу одновременное введение каждого терапевтического средства можно проводит любым пригодным способом, включая, но не ограничиваясь ими, местные способы, пероральные способы, внутривенные способы, внутримышечные способы, глазные способы и прямое всасывание через ткани слизистых мембран. Терапевтические средства можно вводить одним способом или различными способами. Например, первое терапевтическое средство выбранного сочетания можно вводить посредством инъекции, в то время как другие терапевтические средства сочетания можно вводить местно.
Альтернативно, например, все терапевтические средства можно вводить местно или все терапевтические средства можно вводить посредством инъекции. Последовательность, в которой вводят терапевтические средства, не является строго критичной, если нет иных указаний. "Комбинированная терапия" также может охватывать введение терапевтических средств, как описано выше, в дальнейшем сочетании с другими биологически активными ингредиентами. Когда комбинированная терапия далее включает немедикаментозное лечение, немедикаментозное лечение можно проводить в любое пригодное время, при условии достижения благоприятного эффекта вследствие совместного действия сочетания терапевтических средств и немедикаментозного лечения. Например, в соответствующих случаях, благоприятный эффект все еще достигается, когда немедикаментозное лечение отделено по времени от введения терапевтических средств, возможно, на сутки или даже недели.
Терапевтические или фармакологические композиции по настоящему изобретению, главным образом, будут содержать эффективное количество активного компонента(ов) терапевтического средства, растворенного или диспергированного в фармацевтически приемлемой среде. Фармацевтически приемлемые среды или носители включают любые и все растворители, диспергирующие среды, покрытия, антибактериальные и противогрибковые средства, обеспечивающие изотоничность и замедляющие всасывание средства и т.п. Применение таких сред и средств для фармацевтически активных веществ хорошо известно в данной области. Также в терапевтические композиции по настоящему изобретению могут быть включены дополнительные активные ингредиенты.
Изготовление фармацевтических или фармакологических композиций будет известно специалистам в данной области с учетом настоящего описания. Как правило, такие композиции можно получать в качестве инъецируемых препаратов, в качестве либо жидких растворов, либо суспензий; твердых форм, пригодных для растворения, или суспендирования, в жидкости перед инъекцией; в качестве таблеток или других твердых веществ для перорального введения; в качестве капсул с замедленным высвобождением; или в любой другой форме, используемой в настоящее время, включая капли для глаз, кремы, лосьоны, мази, ингалируемые препараты и т.п. Применение стерильных составов, таких как жидкости для промывания на основе физиологического раствора, хирургами, терапевтами или лицами, осуществляющими уход, для обработки конкретной области операционного поля, также может быть особенно пригодным. Также композиции можно доставлять с помощью микроустройства, микрочастицы или губки.
После изготовления терапевтические средства будут вводить способом, совместимым с вводимым составом, и в таком количестве, которое является фармакологически эффективным. Составы легко вводить в различных дозированных формах, таких как тип инъецируемых растворов, описанный выше, однако также можно использовать капсулы для высвобождения лекарственного средства и т.п.
В данном случае количество активного ингредиента и объем композиции, подлежащей введению, зависят от животного-хозяина, подлежащего лечению. Точные количества активного соединения, требуемые для введения, зависят от суждения специалиста и являются индивидуальными для каждого индивида.
Как правило, используют минимальный объем композиции, требуемый для диспергирования активных соединений. Пригодные схемы введения также варьируют, однако их типичность можно обеспечить первоначальным введением соединения и мониторингом результатов, с последующим введением следующих контролируемых доз через следующие промежутки времени.
Например, для перорального введения в форме таблетки или капсулы (например, желатиновой капсулы), активный компонент лекарственного средства можно комбинировать с пероральным нетоксичным фармацевтически приемлемым инертным носителем, таким как этанол, глицерин, вода и т.п. Более того, когда желательно или необходимо, также в смесь можно включать пригодные связующие вещества, смазывающие вещества, дезинтегрирующие вещества и красители. Пригодные связующие вещества включают крахмал, алюмосиликат магния, крахмальную пасту, желатин, метилцеллюлозу, карбоксиметилцеллюлозу натрия и/или поливинилпирролидон, природные сахара, такие как глюкоза или бета-лактоза, кукурузные подсластители, природные и синтетические камеди, такие как гуммиарабик, трагакант или альгинат натрия, полиэтиленгликоль, воски и т.п. Смазывающие вещества, используемые в этих дозированных формах, включают олеат натрия, стеарат натрия, стеарат магния, бензоат натрия, ацетат натрия, хлорид натрия, диоксид кремния, тальк, стеариновую кислоту, ее соль с магнием или кальцием и/или полиэтиленгликоль и т.п. Дезинтегрирующие вещества включают, но не ограничиваются ими, крахмал, метилцеллюлозу, агар, бентонит, крахмалы ксантановых смол, агар, альгиновую кислоту или ее соль с натрием, или шипучие смеси, и т.п. Разбавители включают, например, лактозу, декстрозу, сахарозу, маннит, сорбит, целлюлозу и/или глицин.
Соединения по этому изобретению также можно вводить в таких пероральных дозированных формах, как таблетки или капсулы с постепенным или с замедленным высвобождением, пилюли, порошки, гранулы, эликсиры, настойки, суспензии, сиропы и эмульсии.
Инъецируемые композиции предпочтительно представляют собой водные изотонические растворы или суспензии, и суппозитории преимущественно получают из жировых эмульсий или суспензий. Композиции могут быть стерилизованными и/или они могут содержать адъюванты, такие как консерваторы, стабилизаторы, смачивающие вещества или эмульгаторы, промоторы растворов, соли для регуляции осмотического давления и/или буферы. Кроме того, также они могут содержать другие терапевтически важные вещества. Композиции получают в соответствии с общепринятыми способами смешивания, гранулирования или нанесения покрытия, соответственно, и, как правило, они содержат приблизительно от 0,1 до 75%, предпочтительно приблизительно от 1 до 50%, активного ингредиента.
Жидкие, в частности инъецируемые, композиции, например, можно получать растворением, диспергированием, и т.д. Активное соединение растворяют или смешивают с фармацевтически чистым растворителем, например таким как вода, физиологический раствор, водный раствор декстрозы, глицерин, этанол и т.п., с получением посредством этого инъецируемого раствора или суспензии. Кроме того, можно изготавливать твердые формы, пригодные для растворения в жидкости перед инъекцией.
Соединения по настоящему изобретению можно вводить в виде внутривенной (как болюсной, так и инфузионной), внутрибрюшинной, подкожной или внутримышечной формы, во всех случаях с использованием форм, хорошо известных специалистам в области фармацевтики. Инъецируемые вещества можно получать в виде общепринятых форм, в качестве либо жидких растворов, либо суспензий.
Парентеральное инъецируемое введение, как правило, используют для подкожных, внутримышечных или внутривенных инъекций и инфузий. Кроме того, в одном подходе для парентерального введения используется имплантация систем для замедленного высвобождения или непрерывного высвобождения, которая обеспечивает поддержание постоянного уровня дозирования, в соответствии с патентом США № 3710795, включенным в настоящий документ в качестве ссылки.
Более того, предпочтительные соединения для настоящего изобретения можно вводить в интраназальной форме посредством местного применения пригодных интраназальных носителей, ингалируемых веществ, или трансдермальными способами, с использованием форм трансдермальных кожных пластырей, хорошо известных специалисту в данной области. Для введения в форме трансдермальной системы для доставки, введение дозировки будет, безусловно, постоянным, а не прерывистым, на протяжении схемы дозирования. Другие предпочтительные местные препараты включают кремы, мази, лосьоны, аэрозольные распыляемые вещества и гели, где концентрация активного ингредиента, как правило, может варьировать от 0,01% до 15%, мас./мас. или мас./об.
Для твердых композиций можно использовать эксципиенты, включающие фармацевтические маннит, лактозу, крахмал, стеарат магния, сахарин натрия, тальк, целлюлозу, глюкозу, сахарозу, карбонат магния, и т.п. Активное соединение, определенное выше, также можно изготавливать в качестве суппозиториев с использованием в качестве носителя, например полиалкиленгликолей, например, пропиленгликоля. В некоторых вариантах осуществления, суппозитории преимущественно изготавливают из жировых эмульсий или суспензий.
Соединения по настоящему изобретению также можно вводить в форме липосомальных систем для доставки, таких как небольшие однослойные везикулы, крупные однослойные везикулы и многослойные везикулы. Липосомы могут быть образованы из различных фосфолипидов, включающих холестерин, стеариламин или фосфатидилхолины. В некоторых вариантах осуществления пленка из липидных компонентов гидратирована водным раствором лекарственного средства с образованием липидного слоя, инкапсулирующего лекарственное средство, как описано в патенте США № 5262564. Например, молекулы аптамеров, описанные в настоящем документе, могут быть предоставлены в виде комплекса с липофильным соединением или неиммуногенным высокомолекулярным соединением, изготовленным с использованием способов, известных в данной области. Примеры ассоциированных с нуклеиновой кислотой комплексов представлены в патенте США № 6011020.
Соединения по настоящему изобретению также могут быть соединены с растворимыми полимерами в качестве нацеливаемых носителей лекарственного средства. Такие полимеры могут включать поливинилпирролидон, сополимер пирана, полигидроксипропилметакриламидфенол, полигидроксиэтиласпанамидфенол или полиэтиленоксидполилизин, замещенный остатками пальмитоила. Более того, соединения по настоящему изобретению могут быть соединены с классом биологически деградируемых полимеров, пригодных для достижения контролируемого высвобождения лекарственного средства, например, с полимолочной кислотой, поли-эпсилон-капролактоном, полигидроксимасляной кислотой, полиортоэфирами, полиацеталями, полидигидропиранами, полицианоакрилатами и поперечно-сшитыми или амфипатическими блок-сополимерами гидрогелей.
В предпочтительном варианте осуществления, соединения по настоящему изобретению можно доставлять в отдел глаза посредством инъекции в стекловидное тело, окологлазной, внутрикамерной, субконъюнктивальной или транссклеральной инъекции в полость глаза или непосредственно в глазную или окологлазную ткань(и). Соединения по этому изобретению можно инъецировать в субтеноновое пространство или в ретробульбарное пространство. Соединения по этому изобретению также можно доставлять в отдел глаза или ткань с помощью системной крови и жидкости в глаз и его ткани, и, таким образом, их вводят парентеральной системной инъекцией, внутривенным, внутримышечным или подкожным способами доставки. Субконъюнктивальное, проводимое в стекловидное тело или транссклеральное введение фармацевтических композиций по этому изобретению может быть пригодным в качестве дополнения к системному введению лекарственного средства для лечения глазных заболеваний и/или системных заболеваний с глазными проявлениями. В некоторых вариантах осуществления способов по этому изобретению для стабилизации, лечения и/или профилактики диабетической ретинопатии и/или болезни Бехчета аптамер против комплемента вводят не системно, предпочтительно его вводят в глаз.
Соединения по настоящему изобретению также можно вводить в отдел глаза или ткань в составе образующего депо геля или полимера, или геля, или полимера с замедленным высвобождением посредством хирургической имплантации биологически дуградеруемой полимерной микросистемы, например, микроустройства, микрочастиц или губки, или других транссклеральных устройств с замедленным высвобождением, имплантируемых в процессе лечения глазного заболевания, или глазного устройства для доставки, например, устройства для замедленного высвобождения на основе полимерных контактных линз. Соединения по этому изобретению также можно вводить в отдел или ткань глаза местно, например, в форме глазных капель, в форме контактной линзы, содержащей соединение по этому изобретению, или посредством ионофереза с использованием электрического тока для продвижения лекарственного средства с поверхности в заднюю область глаза.
Если желательно, фармацевтическая композиция, подлежащая введению, также может содержать небольшие количества нетоксичных вспомогательных веществ, таких как смачивающие вещества или эмульгаторы, pH-буферные средства и другие средства, например, такие как ацетат натрия и олеат триэтаноламина.
Схему дозирования для применения аптамеров выбирают в соответствии с различными факторами, включая тип, вид, возраст, вес, пол и медицинские показатели пациента; тяжесть состояния, подлежащего лечению; способ введения; функцию почек и печени пациента и конкретный используемый аптамер или его соль. Терапевт или ветеринар обычной квалификации может легко определить и назначить эффективное количество лекарственного средства, необходимое для профилактики, противодействия или остановки прогрессирования состояния.
Пероральные дозы композиций аптамера по настоящему изобретению, при использовании для достижения указанных эффектов, будут находиться в диапазоне между приблизительно 0,05 и 7500 мг/сутки перорально. Композиции предпочтительно находятся в форме делимых таблеток, содержащих 0,5, 1,0, 2,5, 5,0, 10,0, 15,0, 25,0, 50,0, 100,0, 250,0, 500,0 и 1000,0 мг активного ингредиента. Соединения по настоящему изобретению можно водить в единичной суточной дозе, или общую суточную дозировку можно вводить в разделенных дозах два, три или четыре раза в сутки.
Инфузируемые дозировки, интраназальные дозы и трансдермальные дозы будут находиться в диапазоне между 0,05 и 7500 мг/сутки. Подкожные, внутривенные и внутрибрюшинные дозировки будут находиться в диапазоне между 0,05 и 3800 мг/сутки.
Глазные дозировки композиций аптамера по настоящему изобретению будут находиться в диапазоне между 0,001 и 10 мг/глаз, вводимыми в глаз, например, посредством инъекции в стекловидное тело, от одного раза в неделю до одного раза в три месяца или посредством устройства или состава для замедленного высвобождения.
Эффективные уровни соединений аптамера по настоящему изобретению в плазме находятся в диапазоне от 0,002 мг/мл до 50 мг/мл. Эффективные уровни соединений аптамера по этому изобретению в глазу могут находиться в диапазоне от 20 нМ до 250 мкМ.
Эффективность лечения
Связанные с неоваскуляризацией нарушения
Эффективность лечения связанного с неоваскуляризацией нарушения, например AMD, в частности AMD экссудативного типа или диабетической ретинопатии, оценивают любым принятым способом измерения наличия замедления или уменьшения ангиогенеза. Это включает прямое наблюдение и непрямую оценку, такую как в случае оценки субъективных симптомов или объективных физиологических показателей. Эффективность лечения, например, можно оценивать, исходя из предотвращения, стабилизации и/или реверсии неоваскуляризации, микроангиопатии, истечения из сосудов или сосудистого отека или любого их сочетания. Эффективность лечения для оценки подавления глазного связанного с неоваскуляризацией нарушения также можно определять с точки зрения стабилизации или улучшения остроты зрения.
При определении эффективности средства против C5, отдельно или в сочетании со средством против VEGF и/или средством против PDGF, в отношении стабилизации, снижения симптома и/или профилактики глазного связанного с неоваскуляризацией нарушения, также пациентов может клинически оценивать офтальмолог через несколько дней после инъекции и непосредственно перед последующей инъекцией. Также каждый месяц можно проводить оценку остроты зрения ETDRS, фотографирование на пленку Кодахром и ангиографию с флуоресцеином.
При определении эффективности аптамера против комплемента, отдельно или в сочетании со средством против VEGF и/или средством против PDGF, в отношении стабилизации, снижения симптома и/или профилактики глазного связанного с неоваскуляризацией нарушения, также пациентов может клинически оценивать офтальмолог через несколько суток после инъекции и непосредственно перед последующей инъекцией. Также каждый месяц можно проводить оценку остроты зрения ETDRS, фотографирование дна, оптическую когерентную томографию и ангиографию с флуоресцеином.
Например, в целях оценки эффективности средства против C5, в частности специфичного к C5 аптамера, отдельно или в сочетании со средством против VEGF и/или средством против PDGF, для лечения глазной неоваскуляризации, проводят исследования, вовлекающие введение либо однократной, либо многократных инъекций в стекловидное тело средства против C5, в частности специфичного к C5 аптамера, отдельно или в сочетании со средством против VEGF и/или средством против PDGF, пациентам, страдающим субфовеальной хориоидальной неоваскуляризацией, вторичной по отношению к связанной со старением дегенерацией желтого пятна в соответствии со стандартными способами, хорошо известными в области офтальмологии. Пациентам с субфовеальной хориоидальной неоваскуляризацией (CNV), вторичной по отношению к связанной со старением дегенерации желтого пятна (AMD), можно проводить однократную инъекцию в стекловидное тело средства против C5, в частности специфичного к C5 аптамера и/или специфичного к VEGF аптамера, и/или специфичного к PDGF аптамера. Мониторинг эффективности проводят, например, посредством офтальмологической оценки и/или ангиографии с флуоресцеином. Пациентов, показывающих стабильное или улучшенное зрение через три месяца после лечения, например демонстрирующих улучшение зрение на 3 строки или более по карте ETDRS, считают получающими эффективную дозировку.
Другие глазные нарушения
Лечение воспалительного конъюнктивита, включая аллергический и гигантский папиллярный конъюнктивит, отека желтого пятна, увеита, эндофтальмита, склерита, язв роговицы, сухого кератита, глаукомы, ишемического заболевания сетчатки, диабетической ретинопатии, отторжения трансплантата роговицы, осложнений, связанных с внутриглазной хирургической операцией, такой как врунтриглазная имплантация линзы, и воспаления, обусловленного хирургической операцией по поводу катаракты, болезни Бехчета, болезни Штаргардта, иммунокомплексного васкулита, болезни Фуха, болезни Фогта-Каянаги-Харада, субретинального фиброза, кератита, витреоретинального воспаления, паразитического заражения/миграции глаз, пигментного ретинита, цитомегаловирусного ретинита и хориоидального воспаления, оценивают способами, принятыми в данной области. При определении эффективности аптамера против комплемента, отдельно или в сочетании с другим средством, в отношении стабилизации, снижения симптома и/или профилактики глазного нарушения, также пациентов может клинически оценить офтальмолог. Клиническую оценку можно проводить через несколько суток после инъекции и непосредственно перед последующей инъекцией. Клиническая оценка может включать прямое наблюдение и непрямую оценку, такую как оценка субъективных симптомов или объективных физиологических показателей. Эффективность лечения, например, можно оценивать, исходя из профилактики, стабилизации и/или реверсии истечения из сосудов или сосудистого отека или любого их сочетания. В случае глаукомы, эффективность лечения можно оценивать, исходя из стабилизации состояния здоровья слоя нервных волокон сетчатки или оптического нерва, которое можно подвергать мониторингу с использованием фотографирования дна или оптической когерентной томографии.
Все публикации и патентные документы, цитированные в настоящем документе, включены в настоящий документ в качестве ссылок, как в случае, если бы в отношении каждой публикации или документа было конкретно и отдельно указано, что они включены в данное описание в качестве ссылки. Цитирование публикаций и патентных документов не следует понимать как подтверждение того, что любые из них относятся к предшествующей области, а также оно не обеспечивает никакого подтверждения относительно их содержания или даты. Настоящее изобретение описано в настоящем документе посредством письменного описания, и специалисты в данной области поймут, что настоящее изобретение может быть применено на практике во множестве вариантов осуществления и что вышеизложенное описание и примеры, следующие ниже, предназначены для иллюстрации, а не для ограничения формулы изобретения, которая представлена ниже.
ПРИМЕР 1
Активность аптамера против C5 в классическом и альтернативном каскадах комплемента
Пример 1A. Анализ гемолиза
Тест CH50 измеряет способность системы комплемента в тестируемом образце сыворотки лизировать 50% клеток в стандартизированной суспензии покрытых антителом эритроцитов овцы. 0,2% раствор сыворотки человека смешивали с покрытыми антителом эритроцитами овцы (набор Diamedix EZ Complement CH50 Kit, Diamedix Corp., Miami, FL) в присутствии или отсутствие различных аптамеров против C5. Анализ проводили в соответствии с протоколом набора в забуференном вероналом физиологическом растворе, содержащем кальций, магний и 1% желатин (буфер для комплемента GVB++) и инкубировали в течение 30 минут при 37°С. После инкубации образцы центрифугировали для осаждения интактных эритроцитов. Для количественного определения высвобождения растворимого гемоглобина, которое пропорционально степени гемолиза, проводили измерение оптической плотности супернатанта при 412 нм (OD412) (Green et al., (1995) Chem. Biol. 2:683-95). Для подтверждения того, что аптамеры блокируют активацию C5, некоторые супернатанты после гемолиза анализировали в отношении наличия C5a и C5b-9 посредством ELISA (набор для C5b-9 ELISA, Quidel, San Diego, CA; набор для C5a ELISA, BD Biosciences, San Diego, CA) в соответствии с протоколами набора для ELISA.
Добавление непегилированного аптамера против C5 (ARC186) (SEQ ID NO:4) в реакционную смесь ингибировало гемолиз зависимым от дозы образом, как показано на графике Фиг.7A, с IC50 0,5 ± 0,1 нМ, (см. Фиг.7B), что согласуется с KD, определенной фильтраций через нитроцеллюлозу. При очень высоких концентрациях аптамера (>10 нМ) степень гемолиза была по существу неотличима от фона (без добавления сыворотки), указывая на то, что ARC186 (SEQ ID NO:4) был способен полностью блокировать активность комплемента. Конъюгация аптамера ARC186 (SEQ ID NO:4) с группами PEG массой 20 кДа (ARC657; SEQ ID NO:61), 30 кДа (ARC658; SEQ ID NO:62), разветвленного PEG массой 40 кДа (1,3-бис(mPEG-[20 кДа])-пропил-2-(4'-бутамид) (ARC187; SEQ ID NO:5), разветвленного PEG массой 40 кДа (2,3-бис(mPEG-[20 кДа])-пропил-1-карбамоил) (ARC1905; SEQ ID NO:67), линейного PEG массой 40 кДа (ARC1537; SEQ ID NO:65) и линейного PEG массой 2×20 кДа (ARC1730; SEQ ID NO:66) оказала небольшой эффект на ингибиторную активность аптамера в анализе гемолиза CH50 (Фиг.7A-Фиг.7D).
В дополнительном исследовании ингибиторную активность пегилированного аптамера против C5 ARC1905 (разветвленный PEG массой 40 кДа (2,3-бис(mPEG-[20 кДа])-пропил-1-карбамоил); SEQ ID NO:67) сравнивали с его непегилированным предшественником, ARC672 (SEQ ID NO:63), который содержит концевой 5'-амин, в анализе гемолиза CH50, описанном выше. Раствор сыворотки человека (Innovative Research, Southfield, MI) смешивали с покрытыми антителом эритроцитами овцы (набор Diamedix EZ Complement CH50 Kit, Diamedix Corp., Miami, FL) в присутствии или отсутствие различных концентраций ARC1905 и ARC627, соответственно, так что конечная концентрация сыворотки в каждом анализируемом образце составляла 0,1%, и проводили анализ в соответствии с рекомендованным изготовителем протоколом. Смеси для реакции гемолиза инкубировали в течение 1 часа при 37°C при встряхивании для обеспечения того, чтобы клетки оставались в суспензии. В конце инкубации интактные клетки осаждали центрифугированием (2000 об/мин, 2 мин, комнатная температура), 200 мкл супернатанта переносили в планшет из полистирола с плоским дном (VWR, каталожный #62409-003). Для количественного определения высвобождения растворимого гемоглобина измеряли оптическую плотность при 415 нм (OD415) супернатанта. % Ингибирование при каждой измеренной концентрации аптамера вычисляли с использованием уравнения % инг. = 100 - 100 × (Aобразца-Aбез сыворотки)/(Aбез аптамера-Aбез сыворотки), где Aобразца представляет собой поглощение в образце при различных концентрациях аптамера, Aбез сыворотки представляет собой поглощение вследствие фонового гемолиза в отсутствие сыворотки (контроль со 100% ингибированием) и Aбез аптамера представляет собой поглощение вследствие базальной активности комплемента в отсутствие аптамера (контроль с 0% ингибированием). Значения IC50 определяли из графика % ингибирования против [ингибитор] с использованием уравнения % инг. = (% инг.)максимум × [ингибитор]n/(IC50 n + [ингибитор]n) + фон. Значения IC90 и IC99 вычисляли из значений IC50 с использованием уравнений IC90 = IC50 × [90/(100-90]1/n и IC90 = IC50 × [99/(100-99]1/n. Значения IC50 для ARC1905 и ARC627 в этом параллельном исследовании составляли 0,648±0,0521 и 0,913±0,0679 соответственно (также см. Фиг.58), далее подтверждая, что пегилирование оказывает небольшой эффект, если оказывает его, на функцию аптамера.
Анализ посредством ELISA супернатантов после гемолиза показал, что это функциональное ингибирование коррелировало с блокадой высвобождения C5a. Таким образом, данные гемолиза показывают, что ARC186 (SEQ ID NO:4), и его пегилированные конъюгаты, являются высокоэффективными ингибиторами комплемента, которые функционируют посредством блокирования катализируемой конвертазой активации C5.
Анализы гемолиза с непегилированным материалом показали, что аптамер против C5 не реагирует перекрестно с C5 из ряда не относящихся к приматам видов, включая крысу, морскую свинку, собаку и свинью. Однако значительная ингибиторная активность была выявлена при скрининге сыворотки приматов, включая сыворотку яванской макаки, макаки резус и шимпанзе. Эффективность аптамера против C5 in vitro далее исследовали в сыворотке яванской макаки с использованием ARC658 (SEQ ID NO:62), аналога ARC186 (SEQ ID NO:4) с PEG массой 30 кДа. При непосредственном сравнении (n=3), ARC658 ингибировал активность комплемента человека с IC50 0,21±0,0 нМ и активность комплемента яванской макаки с IC50 1,7 ± 0,4 нМ (Фиг.8). Таким образом ARC658 (SEQ ID NO:62) является в 8±3 раз менее эффективным в сыворотке яванской макаки по сравнению с сывороткой человека при этом определении.
В сходном исследовании эффекты разветвленного пегилированного посредством PEG массой 40 кДа (2,3-бис(mPEG-[20 кДа])-пропил-1-карбамоила) аптамера против C5 ARC1905 (SEQ ID NO:67) на активацию классического каскада комплемента, при анализе посредством гемолиза эритроцитов овцы, исследовали в присутствии сыворотки человека (Innovative Research, Southfield, MT), яванской макаки (Bioreclamation, Hicksville, NY), или крысы (Bioreclamation, Hicksville, NY). Эти анализы проводили в высокоразбавленной сыворотке, 0,1% для человека и яванской макаки, и 0,3% для крысы, в тех же условиях, что и использовали для сравнения ингибиторных эффектов ARC1905 и ARC672 на гемолиз эритроцитов овцы, описанного непосредственно выше. При непосредственном сравнении с помощью ARC1905 достигали полного ингибирования (90-99%) активности комплемента in vitro в сыворотке как человека, так и яванской макаки, в то время как ARC1905 показал небольшую ингибиторную активность, или ее отсутствие, в сыворотке с комплементом крысы (Фиг.59A). Аналогично ARC658, ARC1905 был в ~10 раз менее эффективен против активности комплемента яванской макаки в условиях анализа, что отразилось значениями IC90 и IC99, представленными на Фиг.59B.
Анализы связывания с нитроцеллюлозным фильтром. Отдельные аптамеры метили 32P на 5'-конце посредством инкубации с γ-32P-ATP и полинуклеотидкиназой (New England Biolabs, Beverly, MA). Меченый радиоактивной меткой аптамер очищали от свободного ATP гель-фильтрацией с последующим полиакриламидным гель-электрофорезом. Для измерения аффинности аптамера против C5 меченный радиоактивной меткой аптамер (≤ 10 пМ) инкубировали с возрастающими концентрациями (0,05-100 нМ) очищенного белка C5 (Quidel, San Diego, CA) в фосфатно-солевом буфере, содержащем 1 мМ MgCl2 при комнатной температуре (23°C) и 37°C, в течение промежутков времени 15 мин и 4 ч. Реакции связывания анализировали фильтрацией через нитроцеллюлозу с использованием 96-луночного дот-блот-вакуумного фильтрационного устройства Minifold I (Schleicher & Schuell, Keene, NH). Использовали трехслойный фильтрующий материал, состоящий (сверху вниз) из нитроцеллюлозы Protran (Schleicher & Schuell), нейлона Hybond-P (Amersham Biosciences, Piscataway, NJ) и бумаги для блоттинга гелей GB002 (Schleicher & Schuell). Слой нитроцеллюлозы, который селективно связывает белок относительно нуклеиновой кислоты, предпочтительно задерживал аптамер против C5 в комплексе с белковым лигандом, в то время как не образовавший комплекс аптамер против C5 проходил через нитроцеллюлозу и прикреплялся к нейлону. Бумагу для блоттинга гелей включали просто в качестве поддерживающего материала для других фильтров. После фильтрации фильтрующие слои разделяли, сушили и проявляли на фосфорном экране (Amersham Biosciences) и количественно определяли с использованием системы для визуализации блотов Storm 860 Phosphorimager® (Amersham Biosciences).
Как показано на Фиг.9 и Фиг.10, повышение концентраций C5 повышает долю ARC186, захваченного нитроцеллюлозной мембраной. Зависимость связанного ARC186 от возрастания концентраций C5 хорошо описана посредством модели с одним участком связывания (C5 + ARC186 ↔ C5·ARC186; % связавшегося = Cmax/(1 + KD/[C5]); Cmax представляет собой максимальный % связавшегося аптамера при насыщении [C5]; KD представляет собой константу диссоциации). Кривые связывания ARC186 при двух температурах после инкубации в течение либо 15 мин, либо 4 ч, представлены на Фиг.9 и 10, соответственно. После инкубации в течение 15 мин кривые связывания ARC186 при 23 и 37°C по существу неотличимы с учетом погрешностей, соответствуя значениям KD 0,5-0,6 нМ (Фиг.9). Различия между кривыми связывания при 23 и 37°C становятся более выраженными при увеличении времени. После инкубации в течение 4 ч (Фиг.10) KD, выявленная при 23°C, снижается до 0,08 ± 0,01 нМ, в то время как KD, выявленная при 37°C, остается неизмененной (0,6 ± 0,1 нМ).
Для того чтобы показать основание для потребности в длительной инкубации при комнатной температуре, аффинность при этой температуре далее исследовали с использованием способов кинетики. Скорость обратной реакции, описывающей диссоциацию C5B·ARC186, составляет νrev = k -1[C5B·ARC186], где νrev представляет собой скорость (в единицах M мин-1) и k -1 представляет собой константу скорости диссоциации первого порядка (в единицах мин-1). Скорость прямой реакции, описывающая образование комплекса C5B·ARC186, представляет собой νfor = k 1[C5][ARC186], где νfor представляет собой скорость (в единицах M мин-1) и k 1 представляет собой константу скорости ассоциации второго порядка (в единицах M-1 мин-1). Данные анализировали с использованием допущения псевдо-первого порядка, где концентрация одного реагирующего вещества (C5 в данном случае) находится в значительном избытке относительно другого ([C5] >> [ARC186], и таким образом остается по существу неизменной на протяжении реакции. В этих условиях прямую реакцию описывает уравнение скорости для процесса первого порядка, νfor = k 1'[ARC186], где k 1' = k 1[C5].
Для анализа диссоциации C5·ARC186 меченный радиоактивной меткой ARC186 (≤ 10 пМ) предварительно уравновешивали с 5 нМ белком C5 в фосфатно-солевом буфере, содержащем 1 мМ MgCl2, при комнатной температуре (23°С). Реакцию диссоциации начинали добавлением немеченного ARC186 (1 мкМ), который действует в качестве ловушки для свободного C5, и останавливали разделением фильтрацией через нитроцеллюлозу связавшегося и свободного меченного радиоактивной меткой ARC186. Ход диссоциации ARC186 исследовали варьированием длительности между началом реакции диссоциации и фильтрацией. Ход диссоциации, наблюдаемый в качестве снижения процентного количества меченого радиоактивной меткой ARC186, задержанного на нитроцеллюлозном фильтре (равного процентному количеству связавшегося с C5), хорошо описывается посредством однократного экспоненциального затухания, где % связавшегося ARC186=100 ×  (см. Фиг.11). Значение константы скорости диссоциации k
-1, определенное этим способом, составляет 0,013±0,02 мин-1, что соответствует времени полужизни (t1/2 = ln2/k
-1) 53 ± 8 мин.
(см. Фиг.11). Значение константы скорости диссоциации k
-1, определенное этим способом, составляет 0,013±0,02 мин-1, что соответствует времени полужизни (t1/2 = ln2/k
-1) 53 ± 8 мин.
Для анализа реакции ассоциации измеряли равновесную константу скорости (k eq) для образования C5B·ARC186 в присутствии различных концентраций белка C5 (1-5 нМ). Образование комплекса инициировали смешиванием белка C5 и меченого радиоактивной меткой ARC186 в PBS, содержащем 1 мМ MgCl2 при комнатной температуре (23°C), и останавливали разделением посредством фильтрации через нитроцеллюлозу. Как описано для реакций диссоциации, ход образования комплексов с течением времени исследовали варьированием длительности между инициацией реакции и фильтрацией. Ход равновесия, наблюдаемый в качестве повышения процентного количества меченного радиоактивной меткой ARC186, задержанного на нитроцеллюлозном фильтре, хорошо описывается посредством однократного экспоненциального затухания, где % связавшегося ARC186 = 100 × (1-e-k-1t). Ход равновесия для 1, 2 и 4 нМ C5 представлен на Фиг.12. Как и ожидалось, значение k eq возрастает линейно с [C5] (k eq (1 нМ) = 0,19 ± 0,02 мин-1; k eq (2 нМ) = 0,39 ± 0,03 мин-1; k eq (3 нМ) = 0,59 ± 0,05 мин-1; k eq (4 нМ) = 0,77 ± 0,06 мин-1; k eq (5 нМ) = 0,88 ± 0,06 мин-1). В условиях эксперимента взаимоотношение между k eq, k 1 и k -1 представляет собой k eq=k 1[C5]+k -1. Таким образом, оценку k 1 проводят исходя из наклона графика k eq против [C5] (см. вставку на Фиг.12), в этом случае она составляет 0,18 ± 0,01 нМ-1мин-1.
Эти данные указывают на то, что в условиях низкой концентрации C5 (например, 0,1 нМ), требуется более длительная инкубация для того, чтобы смесь C5 и меченного радиоактивной меткой ARC186 достигла равновесия. В этих условиях, k eq = (0,18±0,01 нМ-1мин-1) (0,1 нМ) + 0,013 мин-1 = 0,03 мин-1, что соответствует времени полужизни 22 мин. Таким образом, для полного уравновешивания (>95%) требуется инкубация в течение практически 2 часов при комнатной температуре (~5 показателей времени полужизни). Кратковременная инкубация (например, 15 мин) значительно занизит истинную аффинность комплекса, как показано выше посредством различия в аффинности, выявленной в случае инкубации в течение 15 мин (KD=0,5 нМ) против 4 часов (KD=0,08 нМ). Альтернативная оценка KD при комнатной температуре может быть вычислена из кинетических данных, исходя из соотношения KD = k -1/k 1 . В этом случае вычисленная KD составляет 0,07±0,01 нМ, что полностью согласуется с KD, определенной термодинамическими способами.
Специфичность ARC186 (SEQ ID NO:4) в отношении C5 также оценивали в анализах фильтрации через нитроцеллюлозу посредством сравнения с компонентами комплемента, расположенными как выше, так и ниже C5 в каскаде комплемента. Очищенные человеческие белки и белковые комплексы приобретали от Complement Technologies (Tyler, TX), включая: C1q (каталожный # A099.18; 2,3 мкМ), C3 (каталожный # A113c.8; 27 мкМ), C5 (каталожный # A120.14; 5,4 мкМ), C5a des Arg (каталожный # A145.6; 60 мкМ), sC5b-9 (каталожный # A127.6; 1 мкМ), фактор B (каталожный # A135.12; 11 мкМ) и фактор H (каталожный № A137.13P; 6,8 мкМ). Реакции связывания осуществляли проведением серийных разведений белка в PBS плюс 1 мМ MgCl2, 0,02 мг/мл BSA и 0,002 мг/мл тРНК, инкубацией в течение 1-4 часов при 25°C или 37°C, с последующим нанесением на устройство для фильтрации через нитроцеллюлозу, как описано выше. Константы диссоциации KD определяли из полулогарифмических графиков % связывания с нитроцеллюлозой против [C5] посредством подстановки данных в уравнение: % связывание с нитроцеллюлозой = амплитуда × [C5]/(KD + [CS]).
Результаты, представленные на Фиг.13, показывают, что аптамер по существу не распознает C5a (KD>>3 мкМ), хотя он проявляет слабую аффинность к растворимому C5b-9 (KD > 0,2 мкМ), вероятно, вследствие взаимодействий с компонентом C5b. Другие компоненты комплемента проявляют от умеренной до слабой аффинности к аптамеру. Неактивированный C3 по существу не связывается с аптамером; однако фактор H (KD ~100 нМ) и, в меньшей степени, C1q (KD>0,3 мкМ) связываются с ним. Взятые вместе, эти данные указывают на то, что ARC186 (SEQ ID NO:4) связывается с высокой аффинностью с C5 человека, главным образом, посредством распознавания домена C5b. Таким образом, ARC186 и его пегилированные производные, например ARC1905, не должны препятствовать образованию C3b, которое является важным для опсонизации бактерий, или свойственному C регуляторным факторам контролю активации.
Конъюгация аптамеров с высокомолекулярными группами PEG обеспечивает возможность пространственного препятствия, ведущего к сниженной аффинности. Модифицированные посредством PEG аптамеры нелегко оценить в отношении прямого связывания посредством анализов фильтрации через нитроцеллюлозу, вследствие тенденции к прикреплению этих аптамеров к нитроцеллюлозе даже в отсутствие белка-мишени. Однако относительную аффинность этих аптамеров можно оценивать из их сравнительной способности конкурировать с меченным радиоактивной меткой непегилированным аптамером (<10 пМ) за связывание с мишенью, при определении посредством фильтрации через нитроцеллюлозу, известной в качестве конкурентного анализа связывания, проводимого при 37°С. По мере повышения концентрации холодного (т.е., не меченного радиоактивной меткой) конкурента процентное количество меченного радиоактивной меткой аптамера, связавшегося с белком-мишенью, повышается. Как показано на Фиг.14, возрастающие концентрации холодного ARC186 (SEQ ID NO:4) или пегилированного аптамера (ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62), и ARC187 (SEQ ID NO:5) (0,05-1000 нМ) явно конкурируют с меченным радиоактивной меткой ARC186 (SEQ ID NO:4) за связывание в присутствии 2 нМ белка C5. Кроме того, кривые титрования для всех четырех аптамеров практически перекрываются, указывая на то, что конъюгация с PEG в случае ARC657, ARC658 и ARC187 оказывает небольшой эффект, или не оказывает его на аффинность аптамера к C5.
В сходном исследовании эффект конъюгации с PEG на связывание с C5 тестировали посредством сравнения ARC672 (ARC186 с 5'-концевым амином; SEQ ID NO:63) с ARC1905 (ARC627, конъюгированный с разветвленным PEG массой 40 кДа (2,3-бис(mPEG-[20 кДа])-пропил-1-карбамоил)) с использованием конкурентного анализа связывания. Получали исходные 10 мкМ растворы каждого аптамера в PBS плюс 1 мМ MgCl2, 0,01 мг/мл BSA, 0,002 мг/мл тРНК и серийно разводили с получением 10X серий образцов, покрывающих >100-кратный диапазон концентрации аптамера. Затем аликвоты по 12 мкл каждого образца добавляли в 96-луночный планшет к 96 мкл меченого радиоактивной меткой 32P ARC186 с получением раствора 1,1X метки и холодного конкурента. Затем 90 мкл раствора метки/конкурента добавляли к 10 мкл 10X белка C5 для инициации реакций. Конечную концентрацию меченного радиоактивной меткой ARC186 в каждой реакционной смеси поддерживали на постоянном уровне. Смеси для реакции связывания достигали равновесия в течение 15-30 мин при 37°C, а затем их фильтровали на устройстве с нитроцеллюлозным фильтром, описанном выше. Для целей анализа данных холодные конкурентные аптамеры рассматривали в качестве конкурентных ингибиторов взаимодействия ARC186/C5; % ингибирования вычисляли посредством нормализации данных относительно контрольных реакций, лишенных конкурента (контроль с 0% ингибированием). Значения IC50 определяли из полулогарифмических графиков % ингибирования против [ARC672] или [ARC1905] подстановкой данных в уравнение: % ингибирование = амплитуда × [конкурент]n/(IC50 n + [конкурент]n).
Как показано на Фиг.60, добавление разветвленного PEG массой 40 кДа (2,3-бис(mPEG-[20 кДа])-пропил-1-карбамоил) оказывало небольшой эффект, или не оказывало его, на аффинность аптамера, при измерении посредством конкурентного связывания. Для ARC672 и ARC1905 были приближенно определены значения KD 0,46 ± 0,149 нМ и 0,71 ± 0,130 нМ соответственно посредством отрезка, отсекаемого на y оси, кривой, соответствующей данным IC50 против C5 на Фиг.60. Оба значения близки к KD, определенной для ARC186 при 37°C.
Температурную зависимость взаимодействия между ARC1905 и C5 также оценивали посредством конкурентного анализа. ARC1905 серийно разбавляли с получением 10X серий образцов, как описано выше. Смесь для реакции связывания уравновешивали в течение 1-4 часов при 25°C или 37°C, а затем фильтровали на устройстве с нитроцеллюлозным фильтром. Процентное ингибирование вычисляли посредством нормализации данных относительно контрольных реакционных смесей, лишенных конкурента (контроль с 0% ингибированием) или лишенных белка C5 (контроль со 100% ингибированием). Значения IC50 определяли из полулогарифмических графиков % ингибирования против [ARC672] или [ARC1905] подстановкой данных в уравнение: % ингибирование = амплитуда × [конкурент]n/(IC50 n + [конкурент]n). Как показано на Фиг.61, ARC1905 связывается с C5 с высокой аффинностью как при 25°C, так и при 37°C. Значения KD 0,15 ± 0,048 нМ и 0,69 ± 0,148 нМ были получены для 25°C и 37°C, соответственно, исходя из отрезка, отсекаемого на оси y, данных IC50 против C5. Оба значения согласовывались со значениями KD, определенными для взаимодействия ARC186/C5, описанного выше.
Пример 1B. Анализ цельной крови
Эффект аптамера против C5 на альтернативный каскад системы комплемента анализировали с использованием следующего анализа цельной крови. Забор крови проводили от здоровых добровольцев-людей в отсутствие антикоагулянта. Аликвоты крови (не содержащей антикоагулянт) инкубировали с возрастающими концентрациями ARC186 (SEQ ID NO:4) в течение 5 часов при комнатной температуре или 37°С. Образцы центрифугировали для выделения сыворотки, и наличие C5b в сыворотке определяли посредством ELISA sC5b-9 (набор C5b-9 ELISA. Quidel, San Diego, CA). Как показано на Фиг.15, активность против комплемента, отражаемая образованием C5b-9, между образцами, инкубированными при различных температурах, отличалась на 3 мкМ. Данные, полученные при комнатной температуре, указывали на то, что концентрация аптамера, требуемая для количественного ингибирования, находится в диапазоне 3-6 мкМ, в то время как описанная концентрация C5 составляет приблизительно 400 нМ. Эти результаты позволяют предположить, что для полного ингибирования активности C5 может быть необходим более чем 10-кратный молярный избыток аптамера против C5 (ARC186; SEQ ID NO:4).
Пример 1C. Активация комплемента зимозаном
Зимозан представляет собой полисахаридный компонент клеточной стенки дрожжей и сильнодействующий активатор альтернативного каскада комплемента. Добавление зимозана к образцам крови, плазмы или сыворотки ex vivo приводит к накоплению продуктов активации комплемента, включая C5a и растворимый вариант C5b-9 (sC5b-9). В образцы неразбавленной сыворотки человека (Center for Blood Research, Boston, MA), цитратной цельной крови человека (Center for Blood Research, Boston, MA) или сыворотки яванской макаки (Charles River Labs, Wilmington, MA) добавляли возрастающие концентрации ARC658 (SEQ ID NO:62), аналога ARC186 с PEG массой 30 кДа (SEQ ID NO:4). Для активации комплемента к образцам добавляли зимозан (Sigma, St. Louis, MO) в 10X суспензии до конечной концентрации 5 мг/мл. После инкубации в течение 15 минут при 37°C частицы зимозана удаляли центрифугированием и степень активации комплемента определяли посредством ELISA для C5a и/или sC5b-9 (набор C5b-9 ELISA, Quidel, San Diego, CA; набор C5a ELISA, BD Biosciences, San Diego, CA).
В отсутствие аптамера обработка зимозаном активирует ~50% C5 сыворотки или цельной крови, по сравнению с активацией ~1% в необработанном образце. Добавление аптамера против C5 в концентрации до 50 нМ (~10% от концентрации C5 в крови) оказало небольшой эффект на образование sC5b-9. Однако последующее титрование C5 возрастающими концентрациями ARC658 (SEQ ID NO:62) ингибировало активацию C5 зависимым от дозы образом, как показано на Фиг.16. В сыворотке или цельной крови человека количественное (~99%) ингибирование было выявлено при 0,8-1 мкМ ARC658 (SEQ ID NO:62), что соответствует ~2 молярным эквивалентам аптамера к C5. Для достижения сравнимого ингибирования в сыворотке яванской макаки требовались более высокие концентрации аптамера. В этом случае 99% ингибирование было достигнуто только в присутствии 10 мкМ аптамера, или ~20 молярных эквивалентов аптамера к C5.
В сходном исследовании тестировали ингибиторные эффекты ARC1905 (пегилированный вариант ARC186 с разветвленным PEG массой 40 кДа (2,3-бис(mPEG-[20 кДа])-пропил-1-карбамоил)) на образцах человека и яванской макаки с использованием зимозана для активации комплемента посредством альтернативного каскада следующим образом. Зимозан A из Saccharomyces cerevisiae был предоставлен Sigma-Aldrich, Inc. (каталожный № Z4250-1G, St. Louis, MO). Зимозан A был предоставлен в качестве порошка и его ресуспендировали в PBS Дульбекко (Gibco, Carlsbad, CA, каталожный № 14190-144) с получением суспензии 50 мг/мл. Замороженная объединенная нормальная сыворотка человека (каталожный № IPLA-SER) была приобретена от Innovative Research (Southfield, MI). Замороженная объединенная сыворотка яванской макаки (каталожный № CYNSRM) была приобретена от Bioreclamation (Hicksville, NY). Флаконы с 5-10 мл сыворотки, предоставленной поставщиком, размораживали при 37°C, разделяли на аликвоты (~1 мл) и хранили при -20°С. Аликвоты размораживали по мере необходимости непосредственно перед применением посредством инкубации при 37°C и хранили на льду в ходе экспериментов. Конечная концентрация сыворотки в каждом анализе составляла ~100%. Изготавливали исходный 20 мкМ раствор ARC1905 в 0,9% физиологическом растворе и серийно разбавляли с получением 10X серий образца, охватывающих ~90-кратный диапазон концентраций аптамера. Также включали образец без аптамера (только физиологический раствор) в качестве отрицательного (0% ингибирование) контроля.
90 мкл сыворотки раскапывали в лунки 96-луночного плашнета для ПЦР (VWR, каталожный № 1442-9596). 10 мкл образца аптамера разбавляли непосредственно в сыворотке при комнатной температуре и перемешивали. 8 мкл зимозана в концентрации 50 мг/мл раскапывали в лунки отдельного 96-луночного планшета для ПЦР. Оба планшета одновременно преинкубировали при 37°C в течение 15 минут. Сразу после преинкубации 80 мкл смеси сыворотка/аптамер добавляли непосредственно к 8 мкл зимозана и перемешивали с получением конечной концентрации зимозана 5 мг/мл. Планшет с реакционной смесью накрывали и инкубировали в течение 15 минут при 37°C. В конце инкубации, реакционную смесь гасили раскапыванием 8 мкл 0,5M ЭДТА в лунки и перемешиванием. Зимозан осаждали центрифугированием (3700 об/мин, 5 мин, комнатная температура) и ~80 мкл супернатанта после гашения переносили в новый 96-луночный планшет для ПЦР и накрывали. Супернатанты быстро замораживали в жидком азоте и хранили при -20°С. Для контроля независимой от зимозана фоновой активации, образцы сыворотки получали и обрабатывали точно как описано выше, за исключением того, что вместо зимозана добавляли 8 мкл физиологического раствора.
Степень активации C5 определяли из относительных уровней C5a, образованного в активированном зимозаном образце, как измеряют посредством ELISA для C5a (ALPCO Diagnostics, Windham, NH, каталожный. № EIA-3327) в соответствии с протоколом набора для C5a ELISA. Набор C5a ELISA включает специфичные для человека реагенты и он предназначен для анализа C5a человека (hC5a) в образцах плазмы или сыворотки. Таким образом, было необходимо охарактеризовать ответ ELISA на C5a яванской макаки с использованием стандартов концентрации для яванской макаки. Для получения набора обычных стандартов аликвоты сыворотки человека или яванской макаки объемом 0,5 мл инкубировали с 5 мг/мл зимозана в течение 15 мин при 37°C, гасили 12,5 мкл 0,5 M ЭДТА и центрифугировали для удаления зимозана. Концентрацию C5a в образце активированной зимозаном сыворотки человека определили как приблизительно 2 мкг/мл hC5a посредством сравнения со стандартными плазмами с hC5a, предоставленными с набором. Концентрацию C5a в образце яванской макаки, выраженную в эквивалентах C5a человека (hC5a экв.), определили как приблизительно 0,6 мкг/мл hC5a экв. Получали серии стандартов, охватывающих диапазон 0,4-400 нг/мл hC5a или 0,12-120 нг/мл hC5a экв. разбавлением в сыворотке крысы (которая не препятствовала ELISA). Стандарты предварительно обрабатывали реагентом для осаждения белка, как указано в протоколе набора для ELISA, и без дальнейшего разбавления наносили на планшет для ELISA. Считывание планшета для ELISA проводили при максимальном поглощении 450 нм (A450) с использованием устройства для измерения поглощения в планшетах VersaMax UV/vis (Molecular Dynamics, Sunnyvale, CA). Значение A450 варьировало в зависимости от концентрации C5a от низкого 0,1-0,2 при низкой концентрации C5a до стабильного ~3,5 при высокой концентрации C5a. Для количественного определения C5a в анализируемых образцах верхняя и нижняя границы определения составляли, соответственно, 25 и 0,78 нг/мл hC5a для человека, и 15 и 0,94 нг/мл hC5a экв. для яванской макаки. A450 против нг/мл hC5a или hC5a экв. наносили, как показано на Фиг.62, и получали стандартную кривую посредством 4-параметрической подгонки данных с использованием уравнения y = ((A - D)/(1 + (x/C)B)) + D.
Непосредственно перед анализом C5a образцы для анализа (включая контроли только с физиологическим раствором и без зимозана) предварительно обрабатывали реагентом для осаждения белка, в соответствии с протоколом набора для ELISA, а затем серийно разбавляли в 0,9% физиологическом растворе. Уровни C5a в неразбавленных образцах для анализа (включая некоторые из контролей без зимозана), как правило, превышали верхнюю границу определения (ULOQ). Таким образом, разведения 1/5, 1/50 и 1/250 тестировали для обеспечения полного диапазона концентраций C5a в анализируемом образце. Уровни C5a количественно определяли посредством сравнения с соответствующей (для человека или яванской макаки) стандартной кривой и корректировали по разбавлению. % Ингибирование для каждой концентрации аптамера вычисляли с использованием уравнения % инг.=100-100×(C5aобразца-C5aбез зимозана)/(C5aтолько с физиологическим раствором-C5aбез зимозана). Значения IC50 определяли из графика % ингибирования против [ARC1905] с использованием уравнения % инг. = (% inh.)максимум × [ARC1905]n/(IC50 n + [ARC1905]n) + фон. Значения IC90 и IC99 вычисляли из значений IC50 с использованием уравнений IC90 = IC50 × [90/(100-90]1/n и IC99 = IC50 × [99/(100-99]1/n.
Степень активации C3 (стадия в обычном каскаде комплемента непосредственно перед C5) определяли из относительных уровней C3a, образованного в каждом активированном зимозаном образце, при измерении посредством ELISA C3a (набор Becton-Dickinson OptiEIA C3a ELISA, каталожный № 550499, Franklin Lakes, NJ) в соответствии с протоколом для набора C3a ELISA.
Непосредственно перед анализом C3a образцы (включая контроли только с физиологическим раствором и без зимозана) серийно разводили в 0,9% физиологическом растворе. ELISA C3a является более чувствительным, чем ELISA для C5a; таким образом, были необходимы разведения 1/500, 1/5000 и 1/25000 для обеспечения полного диапазона концентраций C3a в образце. Стандарты набора, полученные из сыворотки человека, использовали вместо обычных стандартов, изготовленных для анализа C5a. Поскольку уровни C3a не варьировали значительно, специфичные для человека стандарты обеспечили достаточный показатель их относительных уровней.
Данные, полученные из ELISA как для C5a, так и для C3, анализировали с использованием Microsoft Excel, и средние значения % ингибирования наносили на график с использованием Kaleidagraph (v. 3.51, Synergy Software). Значения IC50, IC90 и IC99 определяли с использованием дополнения XLfit 4.1 к Excel. Сравнительные эффекты ARC1905 на активацию комплемента человека и яванской макаки, измеренные посредством этого подхода, обобщены на Фиг.63 и Фиг.64. Как можно видеть из этих фигур, полное ингибирование активации C5 посредством альтернативного каскада является достижимым in vitro посредством ARC1905 в сыворотке как человека, так и яванской макаки. В сыворотке человека концентрация ARC1905, требуемая для 90% ингибирования активации C5 в неразведенном образце, составляла 442 ± 23 нМ, что приблизительно эквивалентно средней молярной концентрации C5. Однако ARC1905 был в 4-6 раз менее эффективен против активности комплемента яванской макаки в условиях анализа, что отражено значениями IC90 и IC99.
Эффекты активации C3 посредством ARC1905, измеренные по уровням C3a, обобщены на Фиг.65. Обоснованием для специфичного нацеливания на конец каскада комплемента является блокирование провоспалительных функций C5a и мембраноатакующего комплекса (MAC) без нарушения связанных с борьбой с патогенами функций предшествующих факторов, достигающих максимального уровня при образовании C3a и C3b. Данные на Фиг.65 демонстрируют, что ARC1905, вплоть до 2 мкМ, не ингибирует образование C3a, и указывают на то, что ARC1905 не оказывает отрицательного влияния на активацию предшествующих компонентов комплемента. С использованием ARC1905 в образцах сыворотки как человека, так и яванской макаки было достигнуто по существу полное блокирование альтернативного каскада активации C5. В условиях этого анализа ARC1905 был приблизительно на порядок менее эффективен в отношении ингибирования активации C5 яванской макаки, чем в отношении активации C5 человека. Без связи с теорией, ингибиторный эффект ARC1905 на активацию комплемента является специфичным к C5, поскольку активация C3 не ингибировалась.
Пример 1D. Модель активации комплемента с петлей трубки
Для тестирования способности аптамера против C5 блокировать активацию комплемента, индуцируемую воздействием чужеродных материалов, встречающимся в контуре искусственного кровообращения, авторы настоящего изобретения использовали модель петли трубки, описанную Nilsson и коллегами (Gong et al., (1996) Journal of Clinical Immunology 16, 222-9; Nilsson et al., (1998) Blood 92, 1661-7). Медицинскую/хирургическую трубку Tygon S-50-HL (с внутренним диаметром 1/4") (United States Plastic Corp. ((Lima, OH), каталожный # 00542) разрезали на фрагменты длиной приблизительно 300 мм (объемом приблизительно 9 мл) и заполняли 5 мл донорской крови человека, содержащей 0,4 единиц/мл гепарина (Celsus) и различные концентрации ARC658 (SEQ ID NO:62) (0-5 мкМ). Каждый отрезок трубки Tygon замыкали в петлю короткими фрагментами (~50 мм) нехирургической силиконовой соединяющей трубки (с внутренним диаметром 3/8") (United States Plastic Corp. (formulation R-3603, каталожный # 00271) как описано в Gong et al. Петли трубок вращали в течение 1 часа со скоростью приблизительно 30 об/мин на водяной бане при 37°C. Затем содержимое петли выливали в конические пробирки из полипропилена, содержащие ЭДТА (конечная концентрация 10 мМ) для гашения активации комплемента. Не содержащую тромбоциты плазму выделяли центрифугированием и анализировали в отношении C5a и C3a посредством ELISA (набор C3a ELISA, Quidel, San Diego, CA; набор C5a ELISA, BD Biosciences, San Diego, CA).
Общая активация комплемента в отсутствие аптамера была небольшой по сравнению с анализом с зимозаном. Как правило, уровни C5a повышались приблизительно на 6 нг/мл после инкубации в течение 1 часа, что соответствует активации <1% доступного C5. Тем не менее, это повышение было воспроизводимым и ингибировалось титрованием посредством ARC658 (SEQ ID NO:62). Как показано на Фиг.17, 300-400 нМ ARC658 (SEQ ID NO:62) было достаточно для достижения 99% ингибирования активации C5, уровня, который является приблизительно эквивалентным или немного меньшим, чем молярная концентрация C5 в крови. Без связи с теорией, несмотря на то, что в этой модели для достижения 99% ингибирования активации C5 требуется меньшее количество аптамера, чем в модели с зимозаном, это наблюдение может отражать значительные отличия активирующих комплемент стимулов, используемых в этих двух анализах. Мониторинг образования C3a также проводили в качестве контроля для подтверждения того, что ARC658 (SEQ ID NO:62) не блокирует активацию стадий, предшествующих C5 в каскаде комплемента. Уровни C3a возросли приблизительно на 4000 нг/мл после инкубации в течение 1 часа, что соответствует приблизительно 10% активации доступного C3. В противоположность образованию C5a небольшое зависимое от дозы ингибирование образования C3a наблюдалось после титрования ARC658 (SEQ ID NO:62), что демонстрирует, что ARC658 (SEQ ID NO:62) специфично блокирует расщепление C5.
Исследование модели с петлей трубки повторяли с аптамером против C5 ARC1905 (SEQ ID NO:67). ARC1905 серийно разбавляли в 0,9% физиологическом растворе с получением 20X серий образцов, охватывающих 100-кратный диапазон концентраций аптамера (конечная концентрация в этом анализе 10-1000 нМ). Образцы, содержащие посторонний аптамер (ARC127), включили для контроля неспецифичных эффектов олигонуклеотида. Образец без аптамера (только с физиологическим раствором) также был включен в качестве отрицательного контроля. Образцы крови одного донора забирали стандартными способами флеботомии от добровольцев из проводящей исследование лаборатории. Цельную кровь забирали от 5 отдельных доноров, непосредственно в шприц объемом 60 мл (Becton-Dickinson, (Franklin Lakes, NJ), каталожный # 309653) и сразу разделяли на аликвоты в бивалирудин (конечная концентрация 20 мкМ) (Prospec-Tany Technogene Ltd., (Israel), серия # 105BIV01) ± аптамер. Антикоагулянт бивалирудин, прямой ингибитор тромбина, использовали вместо гепарина, который препятствует активации комплемента.
Модель петли трубки проводили по существу как описано непосредственно выше. Отрезки трубки ~300 мм (диаметром 1/4", объемом ~9 мл) заполняли 5 мл образцов кровь/аптамер/бивалирудин непосредственно после забора крови от донора. Затем трубки осторожно соединяли в петли с помощью коротких фрагментов (~50 мм) силиконовой соединяющей трубки, с получением объема газа ~4 мл. Петли трубок вращали вертикально при 32 об/мин в процессе инкубации на водяной бане при 37°C в течение 1 часа. После инкубации, все 5 мл образца переносили в 15-мл коническую пробирку (Corning, (Corning, NY), каталожный # 430766), содержащую 100 мкл 0,5 M ЭДТА, с конечной концентрацией ЭДТА 10 мМ. После гашения и центрифугирования собирали 1 мл супернатанта плазмы каждого образца (Eppendorf Centrifuge 5804) при 4°C (3300 об/мин, 20 минут). Супернатанты быстро замораживали в жидком азоте и хранили при -20°С. Для контроля фоновой активации получали образец пре-CPB посредством добавления 5 мл свежей крови непосредственно в 15-мл коническую пробирку на льду, содержащую 100 мкл 0,5 M ЭДТА. Этот образец обрабатывали для получения плазмы и хранили, как описано выше.
Степень активации C5 определяли из относительных уровней C5a, образованного в каждом активированном образце, при измерении посредством ELISA C5a, как описано непосредственно выше. ELISA C5a проводили на неразбавленных образцах плазмы в соответствии с протоколом набора ELISA и уровни C5a в образцах количественно определяли посредством сравнения со стандартами C5a, предоставленными изготовителем. % Ингибирование образования C5a в каждой концентрации аптамера вычисляли с использованием уравнения % инг. = 100 - 100 × (C5aобразца - C5aпре-CPB)/(C5aтолько с физиологическим раствором - C5aпре-CPB). Значения IC50 определяли из графика % ингибирования против [ARC1905] с использованием уравнения % инг. = (% inh.)максимум × [ARC1905]n / (IC50 n + [ARC1905]n) + фон. Значения IC90 и IC99 вычисляли из значений IC50 с использованием уравнений IC90 = IC50 × [90/(100-90]1/n и IC99 = IC50 × [99/(100-99]1/n.
Степень активации C3 определяли из относительных уровней C3a, образованного в каждом активированном образце, определенных посредством ELISA C3a, как описано непосредственно выше. Непосредственно перед анализом C3a образцы (включающие контроли только с физиологическим раствором и пре-CPB) серийно разводили в 0,9% физиологическом растворе. ELISA C3a более чувствителен, чем ELISA для C5a; таким образом, для обеспечения диапазона концентраций C3a в образце было необходимо разведение 1/5000. Уровни C3a в образце количественно определяли сравнением со стандартами набора, и % ингибирование вычисляли, как описано для C5a. Данные анализировали с использованием Microsoft Excel, и средние значения % ингибирования наносили на график с использованием Kaleidagraph (v. 3.51, Synergy Software). Значения IC50, IC90 и IC99 определяли с использованием дополнения XLfit 4.1 к Excel.
Средние эффекты ARC1905 и постороннего аптамера, ARC127, на активацию комплемента у пяти доноров обобщены на Фиг.66. Как показано на Фиг.67, полная блокада активации C5, отражаемая образованием C5a, была достигнута при <500 нМ ARC1905, в то время как посторонний аптамер не оказывал ингибиторного эффекта вплоть до 1 мкМ. Средние значения IC50, IC90 и IC99 в цельной крови составляли 119 ± 28,6 нМ, 268 ± 39,2 нМ и 694 ± 241 нМ, соответственно (Фиг.66). Без связи с теорией, обоснованным является предположение, что ARC1905 исключается из объема клеток крови, который составляет приблизительно 45% от общего объема. Значения IC50, IC90 и IC99, скорректированные для отражения ингибирования C5 в плазме, таким образом, составляли 216 ± 52,0 нМ, 487 ± 71 нМ и 1261 ± 438 нМ. Эти значения согласуются с параметрами, вычисленными для ингибирования посредством ARC1905 индуцируемой зимозаном активации комплемента в сыворотке, что позволяет предположить, что клеточные компоненты крови не препятствуют значительно активности ARC1905 против C5. Образование C3a не ингибировалось посредством ARC1905 или постороннего аптамера вплоть до 1 мкМ. Без связи с теорией, это позволяет предположить, что ARC1905 ни ингибирует реакцию конвертазы C3, ни блокирует другие стадии, которые приводят к активации альтернативного каскада, такие как депонирование C3 и сборка конвертазы.
ПРИМЕР 2
Селекция de novo и последовательности
Селекция C5 с пулом dRmY
Две селекции проводили для идентификации аптамеров dRmY к полноразмерному белку C5 человека. Белок C5 (Quidel Corporation, San Diego, CA или Advanced Research Technologies, San Diego, CA) использовали в полноразмерной ("FL") и частично трипсинизированной ("TP") форме и обе селекции представляли собой прямые селекции против белковых мишеней, иммобилизованных на гидрофобном планшете. Обе селекции привели к пулам значительно обогащенным в отношении связывания полноразмерного C5 против интактного, не подвергнутого селекции пула. Все последовательности, представленные в этом примере, показаны в направлении от 5' к 3'.
Получение пула. Матрицу ДНК с последовательностью TAATACGACTCACTATAGGGAGAGGAGAGAACGTTCTACN(30)GGTCGATCGATCGATCATCGATG (ARC520; SEQ ID NO:70) синтезировали с использованием устройства для синтеза ДНК ABI EXPEDITETM и стандартными способами удаляли защитные группы. Матрицы амплифицировали с 5'-праймером TAATACGACTCACTATAGGGAGAGGAGAGAACGTTCTAC (SEQ ID NO:71) и 3'-праймером CATCGATGATCGATCGATCGACC (SEQ ID NO:72), а затем использовали в качестве матрицы для транскрипции in vitro с РНК-полимеразой T7 с одной мутацией Y639F. Транскрипцию проводили с использованием 200 мМ HEPES, 40 мМ DTT, 2 мМ спермидина, 0,01% TritonX-100, 10% PEG-8000, 9,5 мМ MgCl2, 2,9 мМ MnCl2, 2 мМ NTP, 2 мМ GMP, 2 мМ спермина, 0,01 единиц/мкл неорганической пирофосфатазы и полимеразы T7 с одной мутацией Y639F.
Селекция: в 1 раунде, стадию положительной селекции проводили на связывающих колонках с нитроцеллюлозным фильтром. В кратком изложении, 1 × 1015 молекул (0,5 нмоль) РНК пула инкубировали в 100 мкл буфера для связывания (1X DPBS) с 3 мкМ полноразмерного C5 или 2,6 мкМ частично трипсинизированного C5 в течение 1 часа при комнатной температуре. Комплексы РНК:белок и свободные молекулы РНК разделяли с использованием 0,45-мкм нитроцеллюлозных колонок для центрифугирования от Schleicher & Schuell (Keene, NH). Колонки предварительно промывали 1 мл 1X DPBS, а затем растворы, содержащие РНК:белок, добавляли в колонку и центрифугировали при 1500 g в течение 2 мин. Проводили три промывания 1 мл буфера для удаления неспецифичных связавшихся веществ с фильтров, а затем комплексы РНК:белок, прикрепленные к фильтрам, элюировали два раза промыванием 200 мкл буфера для элюирования (7 M мочевина, 100 мМ ацетат натрия, 3 мМ ЭДТА, предварительно нагретый до 95°С). Элюированную РНК осаждали (2 мкл гликогена, 1 объем изопропанола, 1/2 объема этанола). РНК подвергали обратной транскрипции с помощью системы ThermoScript RT-PCRTM (Invitrogen, Carlsbad, CA) в соответствии с инструкциями изготовителя, с использованием 3'-праймера, описанного выше SEQ ID NO:72, с последующей амплификацией посредством ПЦР (20 мМ Tris pH 8,4, 50 мМ KCl, 2 мМ MgCl2, 0,5 мкМ праймеров SEQ ID NO:71 и SEQ ID NO:72, 0,5 мМ каждого dNTP, 0,05 единиц/мкл Taq-полимеразы (New England Biolabs, Beverly, MA)). Матрицы для ПЦР очищали с использованием колонок Centricep (Princeton Separations, Princeton, NJ) и использовали для транскрипции пула следующего раунда.
В последующих раундах селекции разделение связанной и свободной РНК проводили на гидрофобных планшетах Nunc Maxisorp (Nunc, Rochester, NY). Раунд начинали иммобилизацией 20 пмоль как полноразмерного C5, так и частично трипсинизированного C5 на поверхности планшета в течение 1 часа при комнатной температуре в 100 мкл в 1X DPBS. Затем супернатант удаляли и лунки промывали 4 раза 120 мкл буфера для промывания (1X DPBS). Затем лунки с белком блокировали 1X буфером DPBS, содержащим 0,1 мг/мл тРНК дрожжей и 0,1 мг/мл ДНК спермы лосося в качестве конкурентов. Используемая концентрация пула всегда по меньшей мере в пять раз превышала концентрацию белка. РНК пула также инкубировали в течение 1 часа при комнатной температуре в пустых лунках для удаления любых связывающихся с пластмассой последовательностей, а затем инкубировали в блокированной лунке без белка для удаления любых конкурентных связывающих последовательностей из пула до стадии положительной селекции. Затем РНК пула инкубировали в течение 1 часа при комнатной температуре, и РНК, связанную с иммобилизованным C5, подвергали обратной транскрипции непосредственно в планшете для селекции посредством добавления RT-смеси (3'-праймер, SEQ ID NO:72 и Thermoscript RT, Invitrogen) с последующей инкубацией при 65°С в течение 1 часа. Полученную кДНК использовали в качестве матрицы для ПЦР (Taq-полимераза, New England Biolabs). Амплифицированную матричную ДНК пула обессоливали посредством колонки Centrisep (Princeton Separations) в соответствии с рекомендованными изготовителем условиями и использовали для программы транскрипции РНК пула для следующего раунда селекции. Транскрибированный пул очищали из геля с использованием 10% полиакриламидного геля в каждом раунде.
Мониторинг хода селекции проводили с использованием сэндвич-анализа связывания с фильтром (дот-блот анализа). 5'-меченую посредством 32P РНК пула (следовая концентрация) инкубировали с C5, 1X DPBS плюс 0,1 мг/мл тРНК и 0,1 мг/мл ДНК спермы лосося, в течение 30 минут при комнатной температуре, а затем наносили на нитроцеллюлозу и нейлоновый сэндвич-фильтр в устройстве для дот-блот анализа (Schleicher and Schuell). Процентное количество РНК пула, связанной с нитроцеллюлозой, вычисляли и подвергали мониторингу приблизительно через каждые 3 раунда с однократным скринингом (±300 нМ C5). Определение Kd пула проводили с использованием титрования белка и устройства для дот-блот анализа, как описано выше.
Данные селекции. Обе селекционные смеси были обогащены после 10 раундов относительно интактного пула. См. Фиг.18. На 10 раунде Kd пула составляла приблизительно 115 нМ для селекции с полноразмерным белком и 150 нМ для селекции с трипсинизированным белком, однако степень связывания на уровне плато составляла только 10% в обоих случаях. Пулы R10 клонировали с использованием набора для клонирования TOPO TA (Invitrogen) и секвенировали.
Информация о последовательности: 45 клонов из каждого пула секвенировали. В полноразмерном пуле R10 преобладал один единичный клон ARC913 (SEQ ID NO:75), который составлял 24% пула, остальную часть составляли 2 набора копий и отельные последовательности. Трипсинизированный пул R10 содержал 8 копий одной последовательности ARC913 (SEQ ID NO:75), однако в пуле преобладала другая последовательность (AMX221.A7; 46%). Клон ARC913 (SEQ ID NO:75) имел Kd приблизительно 140 нМ и степень связывания доходила до 20%. См. Фиг.19.
Отдельная последовательность, приведенная в таблице 3, представлена в направлении от 5' к 3' и соответствует рибонуклеотидной последовательности аптамера, которая была отобрана в представленных условиях dRmY SELEXTM. В вариантах осуществления этого изобретения, полученных посредством этой селекции (и как отражено в списке последовательностей), пурины (A и G) являются дезокси и пиримидины (U и C) являются 2'-OMe. Последовательность, приведенная в таблице 3, может содержать или может не содержать кэп (например, 3'-инвертированный dT). Уникальная последовательность аптамера, представленная ниже, начинается с 23 нуклеотида, непосредственно после фиксированной последовательности GGGAGAGGAGAGAACGUUCUAC (SEQ ID NO:73) и продолжается до 3'-фиксированной последовательности нуклеиновой кислоты GGUCGAUCGAUCGAUCAUCGAUG (SEQ ID NO:74).

Анализ гемолиза: Эффект ARC913 (SEQ ID NO:75) на классический каскад системы комплемента анализировали с использованием анализа гемолиза, описанного ранее, по сравнению как с ARC186 (SEQ ID NO:4) (аптамер против C5, положительный контроль), так и с не подвергнутым селекции пулом dRmY (отрицательный контроль). В анализе ингибирования гемолиза, 0,2% раствор цельной сыворотки человека смешивали с покрытыми антителом эритроцитами овцы (Diamedix EZ Complement CH50 Test, Diamedix Corporation, Miami, FL) в присутствии титрованного ARC913 (SEQ ID NO:75). Анализ проводили в забуференном вероналом физиологическом растворе, содержащем кальций, магний и 1% желатин (буфер для комплемента GVB++) и инкубировали в течение 1 ч при 25°С. После инкубации образцы центрифугировали. Измеряли оптическую плотность при 415 нм (OD415) супернатанта. Ингибирование активности в отношении гемолиза выражают в качестве % активности в отношении гемолиза по сравнению с контролем. См. Фиг.20. IC50 аптамера вычислили как приблизительно 30 нМ.
ПРИМЕР 3
Композиция и оптимизация последовательности
Пример 3A
Минимизация ARC913.
Шесть конструкций на основе ARC913 (SEQ ID NO:75) транскрибировали, очищали из геля и тестировали в дот-блотах в отношении связывания с C5. ARC954 был сходным с исходным клоном с Kd 130 нМ и степенью связывания 20%, в то время как ARC874 (SEQ ID NO:76) был единственным другим клоном, который связывался с C5 с Kd 1 мкМ.
Отдельные последовательности, приведенные в таблице 4, представлены в направлении от 5' к 3', они были образованы из аптамеров, отобранных в представленных условиях dRmY SELEX. В вариантах осуществления этого изобретения, образованных при этой селекции (и как отражено в списке последовательностей), пурины (A и G) являются дезокси и пиримидины (U и C) являются 2'-OMe. Каждая из последовательностей, приведенных в таблице 4, может содержать или может не содержать кэп (например, 3'-инвертированный dT).

Пример 3B. Оптимизация ARC913: Активированная повторная селекция
В целях как оптимизации клона ARC913 (SEQ ID NO:75) для аффинности связывания C5, так и определения ключевых связывающих элементов, проводили активированную повторную селекцию. Активированную повторную селекцию используют для исследования требований к последовательности в активном клоне или минимере. Селекцию проводят с синтетическим вырожденным пулом, который был создан, исходя из единичной последовательности. Уровень вырожденности, как правило, варьирует от 70% до 85% нуклеотидов дикого типа. Как правило, наблюдают нейтральные мутации, однако в некоторых случаях изменения последовательности могут приводить к повышению аффинности. Затем комплексную информацию о последовательности можно использовать для идентификации минимального связывающего мотива и способствования попыткам оптимизации.
Получение пула. Матричная последовательность taatacgactcactataGGGAGAGGAGAGAACGTTCTACN(30)GTTACGACTAGCATCGATG (SEQ ID NO:82) была основана на ARC913 (SEQ ID NO:75) и ее синтезировали с каждым остатком, происходящим из случайной области с изменением на уровне 15%, т.е. в каждом случайном положении ("N"), остаток имеет 85% вероятность того, что он будет представлять собой нуклеотид, встречающийся в последовательности дикого типа CTTGGTTTGGCACAGGCATACATACGCAGGGGTCGATCG (SEQ ID NO:83) и вероятность 15% того, что он будет представлять собой один из других трех нуклеотидов.
Матрицу и пул РНК для активированной повторной селекции получали, по существу, как описано выше. Матрицы амплифицировали с праймерами taatacgactcactataGGGAGAGGAGAGAACGTTCTAC (SEQ ID NO:84) и CATCGATGCTAGTCGTAAC (SEQ ID NO:85). Проводили две селекции с полноразмерным C5, одну селекцию проводили с использованием более высокой концентрации соли на стадии промывания. Протокол селекции выполняли, как описано выше, с двумя исключениями: 1) раунд 1 проводили на гидрофобных планшетах (а также все последующие раунды) только с одной положительной стадией; и 2) в процессе селекции совсем не использовали конкурентов. Концентрацию C5 и концентрацию пула РНК поддерживали постоянной на уровне 200 нМ и 1 мкМ, соответственно.
Данные активированной повторной селекции. Селекционные смеси как с нормальной, так и с высокой концентрацией соли были обогащены после 5 раундов относительно интактного пула. На 5 раунде Kd составляла приблизительно 165 нМ для селекции с высокой концентрацией соли и 175 нМ для селекции с нормальной концентрацией соли. На уровне плато степень связывания обеих смесей составляла 20%. Пулы R4 клонировали с использованием набора для клонирования TOPO TA (Invitrogen, Carlsbad, CA) и 48 клонов из каждого пула секвенировали. 12 клонов из каждого пула транскрибировали и анализировали в одноточечном дот-блот анализе при 500 нМ C5. Константы диссоциации (Kd) снова измеряли с использованием дот-блот анализа, описанного ранее. Kd оценивали в отношении 11 наилучших клонов, идентифицированных в одноточечном скрининге, посредством подстановки данных в уравнение: фракция связанной РНК = амплитуда*Kd/(Kd + [C5]). Клоны с тремя наилучшими Kd представляли собой SEQ ID NO:91 (73 нМ), SEQ ID NO:96 (84 нМ) и SEQ ID NO:95 (92 нМ). Последовательности для этих 11 клонов представлены ниже в таблице 5.
Последовательности, приведенные в таблице 5, представлены в направлении от 5' к 3' и соответствуют нуклеотидным последовательностям аптамеров, которые отбирали в представленных условиях dRmY SELEX. В вариантах осуществления этого изобретения, полученных в результате этой селекции (и как отражено в списке последовательностей), соответствующие последовательности, содержащие сочетания остатков dRmY, как указано в тексте, где пурины (A и G) являются дезокси и пиримидины (U и C) являются 2'-OMe. Каждая из последовательностей, приведенных в таблице 5, может содержать или может не содержать кэп (например, 3'-инвертированный dT). Уникальные последовательности каждого аптамера, представленного ниже, начинаются с 23 нуклеотида, непосредственно после 5'-фиксированной последовательности GGGAGAGGAGAGAACGUUCUAC (SEQ ID NO:86) и продолжается до 3'-фиксированной последовательности нуклеиновой кислоты GUUACGACUAGCAUCGAUG (SEQ ID NO:87).

Пример 3C
Модификация ARC186 посредством разветвленного PEG массой 40 кДа
Олигонуклеотид 5' NH2-fCmGfCfCGfCmGmGfUfCfUfCmAmGmGfCGfCfUmGmAmGfUfCfUmGmAmGfUfUfUAfCfCfUmGfCmG-3T-3' (ARC672, SEQ ID NO:63) синтезировали на устройстве для синтеза ДНК Expedite (ABI, Foster City, CA) в соответствии с рекомендованными изготовителем способами с использованием стандартной коммерчески доступной 2'-OMe РНК и 2'-F РНК и TBDMS-защищенных РНК-фосфорамидитов (Glen Research, Sterling, VA) и подложки CPG с инвертированным дезокситимидином. К функциональной группе концевого амина присоединяли 5'-амино-модификатор, 6-(трифторацетиламино)гексил-(2-цианоэтил)-(N,N-диизопропил)фосфорамидит, C6-TFA (Glen Research, Sterling, VA). После удаления защитных групп олигонуклеотиды очищали ионообменной хроматографией на смоле Super Q 5PW (30) (Tosoh Biosciences) и осаждали этанолом.
Модифицированный по амину аптамер после синтеза конъюгировали с различными группами PEG. Аптамер растворяли в растворе вода/DMSO (1:1) до концентрации между 1,5 и 3 мМ. Добавляли натрий-карбонатный буфер, pH 8,5, до конечной концентрации 100 мМ, и олигонуклеотиды подвергали реакции в течение ночи с 1,7-молярным избытком требуемого реагента PEG (например, ARC1905, Sunbright GL2-400NP сложный эфир п-нитрофенилкарбоната массой 40 кДа [NOF Corp, Japan], или ARC187, сложный эфир mPEG2-NHS массой 40 кДа [Nektar, Huntsville AL]), растворенного в равном объеме ацетонитрила. Полученные продукты очищали ионообменной хроматографией на смоле Super Q 5PW (30) (Tosoh Biosciences), и обессоливали с использованием обращенно-фазовой хроматографии на смоле Amberchrom CG300-S (Rohm and Haas) и лиофилизировали. Структура ARC187 (SEQ ID NO:5) представлена на Фиг.21, в то время как структура ARC1905 (SEQ ID NO:67) представлена на Фиг.22.
ПРИМЕР 4
Модель изолированного перфузируемого сердца
Пример 4A
Проверка принципа с помощью ARC186
Средняя концентрация компонента C5 комплемента в плазме человека составляет приблизительно 500 нМ. При воздействии на изолированные сердца мышей, перфузируемые буфером Krebs Heinseleit, 6% плазмы человека, происходит активация каскада комплемента человека, что приводит к расщеплению C5 на C5a и C5b. После этого компонент C5b образует комплекс с компонентами C6, C7, C8 и C9 комплемента, также известными как "мембраноатакующий комплекс" ("MAC" или C5b-9), который повреждает кровеносные сосуды сердца и сердечные миоциты, таким образом вызывая дисфункцию миокарда (повышенное конечно-диастолическое давление, аритмии) и асистолию (Evans et. al., Molecular Immunology, 32, 1183-1195 (1995)). Ранее моноклональные и одноцепочечные антитела, которые блокируют расщепление C5 человека (пекселизумаб или одноцепочечный вариант scFv пекселизумаба) тестировали в этой модели, и было показано, что они ингибируют повреждение и дисфункцию миокарда (Evans et al., 1995).
Эту модель использовали для установления того, что блокирующий C5 аптамер ARC186 (SEQ ID NO: 4), такой как пекселизумаб, ингибирует опосредуемое компонентом C5 комплемента человека повреждение изолированных перфузируемых мышиных сердец. Мышей C57 B1/6 приобретали от Charles River Laboratories, (Wilmington, MA). В кратком изложении, после индукции глубокой анестезии сердце каждой мыши извлекали и устанавливали на затупленную иглу, помещенную в аорту, через которую сердце подвергали постоянной перфузии буфером Krebs Heinseleit. В левый желудочек помещали датчик давления (Mouse Specifics, Boston, MA), который давал возможность непрерывного измерения частоты сердцебиений и внутрижелудочкого давления. После 15-минутного периода уравновешивания, в течение которого были проведены исходные измерения, сердца подвергали последовательной перфузии буфером и 6% плазмой человека +/- аптамер в различных концентрациях (см. Фиг.23). В ходе этих исследований и как описано в Evans et al., авторы настоящего изобретения показали, что в сердцах, которые подвергали перфузии буфером Krebs Heinseleit + 6% плазма человека, остановка происходила при добавлении плазмы к перфузионному раствору в течение 5 минут, в то время как сердца, которые подвергали непрерывной перфузии буфером отдельно, продолжали биться более двух часов. Таким образом, длительность каждого эксперимента была произвольно определена как 15 минут. Схема этого исследования с ARC186 представлена на Фиг.23.
Внутрижелудочное давление подвергали мониторингу и регистрировали непрерывно с получением записи волны давления (Фиг.24 и 25). Наиболее низкая точка смещения соответствует конечно-диастолическому давлению ("EDP") и наиболее высокая точка смещения соответствует систолическому давлению ("SP"). Исходные волны давления представлены слева от вертикальной черной линии, обозначенной "0", показанной на каждой записи. Как опубликовано ранее (Evans et al., 1995), в сердцах, перфузированных 6% плазмой человека, происходило быстрое повышение давления в левом желудочке и диастолического давления, с достижением в завершении асистолии (сердце останавливалось) в течение 5 минут (Фиг.24). Когда в плазму человека добавляли посторонний аптамер в 50-кратном избытке, также наблюдали повышенное EDP и асистолию (Фиг.24).
Когда к системе добавляли ARC186 с молярной эквивалентностью, также происходило резкое повышение EDP, завершавшееся асистолией (Фиг.25). Во всех трех группах сердец, в которых происходило опосредуемое комплементом повреждение, повышение EDP и асистолия, сердца к концу эксперимента были заметно отечными и набухшими. Когда к плазме добавляли ARC186 в 10-кратном или 50-кратном (Фиг.25) молярном избытке, волны давления в желудочке оставались нормальными и асистолию не наблюдали. Кроме того, ранее описанный отек и набухание не были заметны в этих группах.
В процессе каждого эксперимента частоту сердцебиений регистрировали с 5-минутными интервалами, и среднюю частоту сердцебиений для каждой группы в течение каждого промежутка наносили на график. Как показано на Фиг.26, в сердцах, перфузированных без аптамера или с посторонним аптамером, быстро развивалась асистолия, как правило, в течение 5 минут. ARC186, добавленный к системе с молярной эквивалентностью, немного замедлял начало асистолии. Однако в итоге сердца в этой группе в останавливались. ARC186, добавленный к плазме в 10-кратном или 50-кратном молярном избытке, сохранял частоту сердцебиений в продолжении эксперимента.
Процентное повышение массы сердца по сравнению с исходным уровнем вычисляли для репрезентативной выборки остановившихся сердец (без аптамера или 50-кратный молярный избыток постороннего аптамера) и сравнивали с защищенными ARC186 сердцами (10-кратный и 50-кратный молярный избыток ARC186). Как показано на Фиг.27, масса защищенных ARC186 сердец увеличилась значительно меньше, чем масса остановившихся сердец в контрольных группах.
Вследствие того, что ARC186 ингибирует расщепление C5, но не C3, продукты расщепления C3 (C3a), но не продукты расщепления C5 (C5a или C5b) должны быть выявлены в вытекающей жидкости из изолированных сердец, защищенных ARC186. Для того, чтобы прямо показать, что ARC186 ингибировал расщепление плазмы человека C5, относительные уровни белков комплемента человека C5a и C5b (продуктов расщепления C5) и C3a (продукта расщепления C3) измеряли в вытекающем буфере в различных группах с помощью коммерчески доступных наборов ELISA (набор C5b-9 ELISA, Quidel, San Diego, CA; набор C5a и C3a ELISA, BD Biosciences, San Diego, CA). ARC186 ингибировал расщепление C5 в плазме человека и продукцию C5a (Фиг.28) и C5b-9 (Фиг.29) зависимым от дозы образом. В противоположность этому ARC186 не оказывал эффекта на расщепление C3 человека на C3a и C3b (Фиг.30), далее демонстрируя специфичность молекулы к C5.
После получения фрагменты комплемента C3b и C5b депонируются местно в тканях вблизи участка активации комплемента. После завершения экспериментов сердца мышей, замораживали в средах OCT (Sakura Finetek, Torrance, CA), разрезанные, а затем окрашивали с использованием стандартной иммуногистохимии в отношении наличия C3b человека (клон H11, Chemicon, Temecula, CA), C5b-9 человека (клон aE11, DAKO, Carpinteria, CA) или контрольного мышиного IgG (Vector Laboratories, Burlingame, CA). Результаты исследования представлены на Фиг.31.
Как описано в этом исследовании, блокирующий C5 аптамер ARC186 тестировали в модели опосредуемого компонентом C5 комплемента повреждения ткани ex vivo, в которой используются изолированные мышиные сердца, перфузируемые буфером Krebs Heinseleit и 6% гепаринизированная плазма человека, на основе модели, описанной в ранее опубликованном исследовании, в котором тестировали эффекты антитела против C5, пекселизумаба, на систему комплемента (Evans, Molecular Immunol 32:1183, (1995). С использованием этой модели было показано, что блокирующий C5 аптамер (a) ингибировал расщепление C5 плазмы человека (но не C3), (b) ингибировал депонирование C5b человека (но не C3b) на ткани сердца мыши и (c) ингибировал опосредуемую C5b-9 человека дисфункцию миокарда в имеющих отношение к клинике концентрациях (5 мкМ, 10-кратный молярный избыток аптамера против C5). Эти данные показывают, что когда каскад комплемента человека активируется физиологическим образом, блокирующие C5 аптамеры способны ингибировать расщепление C5 плазмы и предотвратить повреждение и дисфункцию миокарда.
Пример 4B
Эффективность пегилированного аптамера
Материалы и способы, используемые в этом исследовании, были точно такими же, как описано в примере 4A, выше. Схема и результаты эксперимента представлены на Фиг.32. В первой половине эксперимента в качестве источника комплемента использовали гепаризинированную плазму человека (Center for Blood Research, Harvard Medical School, Boston, MA), а во второй половине в качестве источника комплемента использовали гепаринизированную плазму яванской макаки (Charles River Laboratories, Wilmington, MA). Пегилированный аптамер (ARC658; SEQ ID NO:62) добавляли в систему в повышенных молярных соотношениях. Несмотря на то, что были получены все необходимые записи давления в желудочках, в таблице представлено наличие или отсутствие повышения конечно-диастолического давления (EDP), независимо от возникновения асистолии и времени до остановки сердца (определяемой как наличие повышенного EDP и асистолии).
В ходе экспериментов с плазмой человека оптимальную дозу AR658 (SEQ ID NO:62) определили как молярно эквивалентную (500 нМ), в то время как в ходе эксперментов с плазмой не относящегося к человеку примата для защиты сердца от опосредуемого C5b повреждения был необходим 50-кратный молярный избыток (25 мкМ) (см. Фиг.32).
Эти данные согласуются с отличием аффинности аптамера против C5 к человеческому C5 относительно C5 не относящегося к человеку примата, показанным с помощью данных in vitro. Без связи с какой-либо теорией, в ходе последующих исследований авторов настоящего изобретения в отношении PK/PD яванской макаки, описанных в примере 5, авторы настоящего изобретения, кроме того, показали, что 30-кратный молярный избыток аптамера был необходим для ингибирования опосредуемого зимозаном расщепления C5 в плазме, далее подтверждая мнение, что аптамер связывает C5 примата с меньшей аффинностью, чем C5 человека.
В совокупности, эти исследования указывают на то, что оба блокирующих C5 аптамера, ARC186 (SEQ ID NO:4) и в большей степени ARC658 (SEQ ID NO:62), эффективны в модели на изолированном перфузируемом сердце мыши. Также эта модель демонстрирует, что для ингибирования опосредуемого C5 плазмы яванской макаки повреждения сердца необходимо было использовать значительно большее количество ARC658 (SEQ ID NO:62) (молярный избыток 30+) по сравнению с опосредуемым C5 человеком повреждением сердца (молярная эквивалентность), далее подтверждая данные in vitro, которые указывают на то, что аптамер обладал более низкой аффинностью к C5 примата. В заключение, эти данные указывают на то, что яванским макакам может быть необходимо дозирование более 30-кратного молярного избытка, для того, чтобы показать блокаду C5 in vivo в процессе исследований PK/PD.
ПРИМЕР 5
МЕТАБОЛИЗМ ЛЕКАРСТВЕННОГО СРЕДСТВА И ФАРМАКОКИНЕТИКА АПТАМЕРОВ ПРОТИВ C5 У ЖИВОТНЫХ
В примерах 5A-5G, все данные, касающиеся концентраций на основе масс, относятся только к молекулярной массе олигонуклеотидной части аптамера, независимо от массы, обеспечиваемой конъюгацией с PEG.
Пример 5A
Метаболическая стабильность ингибитора C5 ARC186 в плазме примата и крысы
Непегилированный олигонуклеотидный предшественник аптамеров (т.е., ARC186; SEQ ID NO:4) тестировали в плазме крысы и яванской макаки (Charles River Labs, Wilmington, MA) в целях оценки его стабильности, показателей кинетики и каскадов деградации. Тестирование проводили с использованием меченого на 5'-конце радиоактивной меткой (32P) аптамера, инкубированного при 37°C в 95% объединенной плазме (цитратной) в течение 50 ч. В выбранные моменты времени, проводили забор аликвот содержащей аптамер плазмы, сразу быстро замораживали в жидком азоте, и хранили при -80°C. Детекцию и анализ аптамера и его метаболитов проводили с использованием экстракции жидкость-жидкость (фенол-хлороформ) с последующим гель-электрофорезом (на 10% денатурирующем полиакриламидном геле для секвенирования) и радиоавтографии высокого разрешения.
На Фиг.33 показан логарифмически-линейный график оставшегося процентного количества полноразмерного аптамера в зависимости от времени инкубации в плазме как крысы, так и яванской макаки. Профиль деградации в обоих видах оказался по существу монофазным, с константой скорости, составляющей приблизительно k ~0,002 ч-1.
Пример 5B
Фармакокинетика ARC657, ARC658 и ARC187 у крысы после внутривенного введения
Для оценки фармакокинетического профиля ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5), и для оценки требуемого уровня и частоты дозирования у приматов и человека проводили фармакокинетическое исследование у катетеризованных крыс Sprague-Dawley (Charles River Labs, Wilmington, MA). Аптамеры изготавливали для инъекции в количестве 10 мг/мл (oligo weight) в стандартном физиологическом растворе и подвергали стерильной фильтрации (0,2 мкм) в предварительно стерилизованный флакон для дозирования в асептических условиях. Используемый способ введения для исследования на крысах представлял собой внутривенный болюс через хвостовую вену в дозе 10 мг/кг. Исследуемые группы состояли из 3 животных на группу, из которых получали серийные образцы крови перед дозированием и в указанные моменты времени в течение 48 часов. Схема исследования представлена на Фиг.34. Образцы крови получали из хирургически имплантированных в яремную вену катетеров, переносили непосредственно в покрытые ЭДТА пробирки, перемешивали переворачиванием и помещали на лед до обработки для получения плазмы.
Плазму собирали центрифугированием пробирок кровь-ЭДТА при 5000 об/мин в течение 5 минут и супернатант (плазму) переносили в свежую предварительно обозначенную пробирку. Образцы плазмы хранили при -80°C до анализа. Анализ образцов плазмы в отношении ARC187 проводили с использованием гомогенного формата анализа с применением прямого добавления аликвот плазмы в анализируемые лунки, содержащие коммерчески доступный флуоресцентный реагент для детекции нуклеиновых кислот OligreenTM (Molecular Probes, Eugene, OR). После инкубации в течение короткого периода времени (5 мин) при комнатной температуре в отсутствие света в планшетах для анализа проводили считывание данных посредством флоресцентного устройства для считывания планшетов (SpectraMax Gemini XS, Molecular Devices, Sunnyvale, CA). Флуоресцентный сигнал от каждой лунки был пропорционален концентрации аптамера в лунке, и концентрации в образце вычисляли посредством интерполяции показателей флуоресценции из стандартной кривой флуоресценция-концентрация (средние значения из кривых для образцов в двух или трех экземплярах). Средние концентрации в плазме получали в каждый момент времени от трех животных в каждой группе. Данные концентрации в плазме против времени подвергали некомпартментному анализу (NCA) с использованием промышленного стандартного программного обеспечения для фармакокинетического моделирования WinNonLinTM v.4.0 (Pharsight Corp., Mountain View, CA). Была проведена оценка следующих первичных фармакокинетических параметров: максимальная концентрация в плазме, Cmax; площадь под кривой концентрация-время, AUC; терминальное время полужизни, t1/2, терминальный клиренс, Cl; и объем распределения в состоянии покоя, Vss.
Данные средней концентрации в плазме против времени представлены на Фиг.35 и нанесены на график на Фиг.36. Данные концентрации против времени подвергали некомпартментному анализу (NCA) с использованием WinNonLinTM v.4.0. Этот анализ привел к значениям, представленным на Фиг.37.
Как и ожидалось, аптамер ARC187 массой 40 кДа (SEQ ID NO:5) обладал наиболее длительным временем полужизни, и аптамер ARC657 массой 20 кДа (SEQ ID NO:61) обладал наиболее коротким временем полужизни. Выявленный Vss относительно известного объема плазмы (~40 мл/кг) подтвердил умеренную степень связывания/секвестрации ARC187 (SEQ ID NO:5) белками и/или матриксом ткани во внесосудистом пространстве. Предполагая необходимость в поддержании 5-кратного молярного избытка аптамера, результаты исследования подтвердили, что ARC187 (SEQ ID NO:5) обладает значительным преимуществом с точки зрения частоты дозирования и общего количества аптамера, необходимого для поддержания требуемых уровней в плазме.
Предшествующие исследования (данные не представлены) у грызунов и приматов с аптамерами со сходным составом показали пропорциональность дозе/линейность в дозах до 30 мг/кг, так что не ожидали, что этот уровень дозирования приведет к нелинейным фармакокинетическим характеристикам.
Пример 5C
Фармакокинетика ARC187 и ARC1905 у мышей после внутривенного введения
Для оценки фармакокинетического профиля олигонуклеотидного остова ARC186 (SEQ ID NO:4), конъюгированного с разветвленным PEG массой 40 кДа, отличающимся от PEG ARC187 (SEQ ID NO:5), проводили фармакокинетическое исследование на самках мышей CD-1 (полученных от Charles River Labs, Wilmington, MA). Аптамеры изготавливали для инъекции в концентрации 2,5 мг/мл (масса олигонуклеотида) в стандартном физиологическом растворе и подвергали стерильной фильтрации (0,2 мкм) в предварительно стерилизованный флакон для дозирования в асептических условиях. Используемый способ введения для исследования на мышах представлял собой внутривенный болюс через хвостовую вену в дозе 10 мг/кг. Исследуемые группы состояли из 3 животных на группу, из которых получали серийные образцы крови перед дозированием (т.е., контрольная группа без дозирования) и в указанные моменты времени в течение 72 часов. Схема исследования представлена на Фиг.38A.
Образцы крови получали посредством терминальной пункции сердца, переносили непосредственно в покрытые ЭДТА пробирки, перемешивали переворачиванием и помещали на лед до обработки для получения плазмы. Плазму собирали центрифугированием пробирок кровь-ЭДТА при 5000 об/мин в течение 5 минут и супернатант (плазму) переносили в свежую предварительно обозначенную пробирку. Образцы плазмы хранили при -80°C до анализа. Анализ образцов плазмы в отношении ARC187 и 1905 проводили с использованием гомогенного формата анализа с применением прямого добавления аликвот плазмы в анализируемые лунки, содержащие коммерчески доступный флуоресцентный реагент для детекции нуклеиновых кислот OligreenTM (Molecular Probes, Eugene, OR). После инкубации в течение короткого периода времени (5 мин) при комнатной температуре в отсутствие света с планшетов для анализа проводили считывание данных посредством флуоресцентного устройства для считывания планшетов (SpectraMax Gemini XS, Molecular Devices, Sunnyvale, CA). Флуоресцентный сигнал от каждой лунки был пропорционален концентрации аптамера в лунке, и концентрации в образце вычисляли посредством интерполяции показателей флуоресценции из стандартной кривой флуоресценция-концентрация (средние значения из кривых для образцов в двух или трех экземплярах). Средние концентрации в плазме получали в каждый момент времени от трех животных в каждой группе. Данные концентрации в плазме против времени подвергали некомпартментному анализу (NCA) с использованием промышленного стандартного программного обеспечения для фармакокинетического моделирования WinNonLinTM v.4.0 (Pharsight Corp., Mountain View, CA). Была проведена оценка следующих первичных фармакокинетических параметров: максимальная концентрация в плазме, Cmax; площадь под кривой концентрация-время, AUC; терминальное время полужизни, t1/2, терминальный клиренс, Cl; и объем распределения в состоянии покоя, Vss. Данные средней концентрации в плазме против времени нанесены на график на Фиг.38B.
Данные концентрации против времени подвергали некомпартментному анализу (NCA) с использованием WinNonLinTM v.4.0. Этот анализ привел к значениям, представленным на Фиг.38С. Как и ожидалось, PEG массой 40 кДа из обоих источников у мышей показал фармакокинетическую эквивалентность.
Те же образцы плазмы для ARC187 и 1905, используемых для анализа oligreen, описанного непосредственно выше, анализировали с использованием валидированного анализа посредством высокоэффективной жидкостной хроматографии (ВЭЖХ) с УФ-детекцией.
Значения средней концентрации в плазме для ARC187 и ARC1905 вычисляли с использованием Microsoft Excel 2003. Когда значения концентрации в плазме были ниже LLOQ биоаналитического анализа перед введением дозы (время 0), приписывали нулевое значение. Значения ниже LLOQ из образцов, взятых после введения дозы, не были включены в вычисления средней концентрации в плазме. Данные средней концентрации в плазме использовали в независимом от модели PK-анализе с использованием WinNonlin, версия 5.1 (Pharsight Corporation, Mountainview, CA). Площадь под кривой концентрация в плазме-время (AUC0-последний) оценивали с использованием линейного правила трапеции. Для вычислений любое значение, которое было ниже LLOQ анализа, за исключением образца перед дозированием, было исключено из вычислений для оценки параметров PK. Кажущееся терминальное время полужизни вычисляли с использованием формулы t1/2 = 0,693/λz, где λz представляет собой константу скорости элиминации, оцененную по регрессии терминального наклона кривой концентрации против времени. Для определения λz использовали по меньшей мере три значения концентрации в плазме после пиковой концентрации на терминальной фазе, и требовалось, чтобы коэффициент определения (r2) составлял ≥ 0,85.
В целом, анализ ВЭЖХ подтверждает анализ oligreen, описанный непосредственно выше, указывая на то, что было выявлено, что ARC1905 и ARC187 являются биологически эквивалентными, исходя из сравнений средних оценок параметров Cmax, AUC0-последний и AUC0-∞. Различия значений AUC0-последний и AUC0-∞ для ARC1905 относительно ARC187 (при измерении посредством ВЭЖХ) находились в пределах критериев приемлемости биоэквивалентности ± 20%.
Пример 5D
Исследование тканевого захвата ингибиторов C5 ARC657, ARC658 и ARC187 у мышей после внутривенного болюсного введения
Самок мышей CD-1 получали от Charles River Labs (Wilmington, MA). Состав ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5) для инъекций был изготовлен в физиологическом растворе в концентрации 5 мг/мл. Составы для дозирования подвергали стерильной фильтрации (0,2 мкм) в предварительно стерилизованные флаконы для дозирования в асептических условиях, и животным вводили внутривенный болюс через хвостовую вену в дозе 25 мг/кг. Исследование состояло из групп по 3 животных для каждого из четырех моментов времени, t= перед введением дозы, через 3, 6, 12 ч. После извлечения крови сосуды каждого животного тщательно промывали (V-30 мл) физиологическим раствором для удаления какой-либо крови, оставшейся в сосудах. Ткани (сердце, печень, почку) собирали, взвешивали, затем гомогенизировали при 50% мас./об. в стандартном физиологическом растворе и хранили при -80°C до анализа.
Анализ ткани в отношении ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5) проводили с использованием анализа по типу ELISA на основе гибридизации. В этом анализе биотинилированный зонд для захвата предварительно иммобилизовывали на лунках 96-луночного микропланшета с концентрацией в растворе для связывания 125 нМ в течение 3 ч. Лунки планшетов промывали 5 раз посредством 1X PBS. Затем планшеты блокировали 150 мкл/лунка 1X SuperBlock в TBS (Pierce Chemical, Rockford, IL). Планшеты снова промывали, накрывали и хранили при 4°C до применения. В отдельных пробирках проводили отжиг образца(образцов) в буфере, содержащем меченый посредством FAM (5'-флуоресцеина) зонд для детекции в образце в концентрации 200 нМ при 90°С в течение 10 мин, а затем гасили на льду. Стандарты концентраций и контрольные образцы плазмы/ткани также подвергали предварительному отжигу с раствором зонда для детекции в образце, а затем раскапывали в анализируемые лунки планшета, содержащие иммобилизованный зонд для захвата с биотином и подвергали отжигу при 45°C в течение 2,5 ч. Затем планшеты снова промывали и заполняли 100 мкл/лунка раствора, содержащего 1X PBS, содержащий 1 мкг/мл моноклонального антитела против флуоресцеина, конъюгированного с пероксидазой хрена (MAb против FITC-HRP, Molecular Probes, Eugene, OR) в 1X PBS, и инкубировали приблизительно в течение 1 ч. Планшеты снова промывали, как указано выше. Затем анализируемые лунки планшета заполняли 100 мкл раствора, содержащего флуорогенный субстрат HRP (QuantaBlu, Pierce Chemical, Rockford, IL), и инкубировали в течение 20-30 мин в отсутствие света. После инкубации в течение 45 минут, добавляли 100 мкл/лунка стоп-раствора для гашения реакции образования флуоресцентного осадка. Считывание с планшетов проводили непосредственно на флуоресцентном устройстве для считывания микропланшетов (SpectraMax Gemini XS, Molecular Devices, Sunnyvale, CA) с возбуждением флуоресценции при 325 нм и испусканием, детектированном при 420 нм. Считывание с каждой лунки проводили 10 раз. Все три аптамера были детектируемыми в ткани сердца в три момента времени (Фиг.39).
Пример 5E
Фармакокинетика и фармакодинамика ингибиторов C5 ARC657, ARC658 и ARC187 у яванской макаки после внутривенного введения - исследование 1
Состав ARC657 (SEQ ID NO:61), ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5) для инъекции был изготовлен в стандартном физиологическом растворе в концентрации 10 мг/мл и составы для дозирования подвергали стерильной фильтрации (0,2 мкм) в предварительно стерилизованные флаконы для дозирования в асептических условиях. Используемый способ введения для исследования у макак представлял собой внутривенный болюс через хирургически имплантированный в бедренную вену катетер в дозе 30 мг/кг (приблизительно 50-кратый молярный избыток). Схема исследования представлена на Фиг.40. Образцы крови получали из катетеров в бедренной вене, переносили непосредственно в пробирки, покрытые цитратом натрия, перемешивали переворачиванием и помещали на лед до центрифугирования для отделения плазмы (3000 об/мин в течение 5 минут). Затем плазму разделяли на аликвоты объемом 250 мкл, которые хранили при -80°C, и одну аликвоту каждого образца оценивали в отношении концентрации аптамера с использованием флуоресцентного анализа OligreenTM, описанного ранее в разделе, касающемся PK у крыс, выше.
Данные первичной концентрации в плазме против времени представлены в форме таблице на Фиг.41. Как и ожидалось, аптамер ARC187 с PEG массой 40 кДа (SEQ ID NO:5) находился в плазме в течение наиболее длительного периода времени, в то время как аптамер ARC657 с PEG массой 20 кДа (SEQ ID NO:61) находился в плазме в течение наиболее короткого периода времени. Исследование данных, представленных на Фиг.41, позволяет предположить, что данные более всего соответствуют двухкомпартментной модели. Таким образом, оценка фармакокинетических параметров, описанных на Фиг.42, была проведена на основе двухкомпартментной модели с использованием промышленного стандартного программного обеспечения для фармакокинетического моделирования WinNonLinTM v.4.0 (Pharsight Corp., Mountain View, CA).
Как показано на Фиг.42, все аптамеры обладали сходным значением Cmax, между 23 и 30 мкМ, указывая на то, что доза аптамера (30 мг/кг) была достаточной для достижения 50-кратного молярного избытка аптамера в плазме относительно концентрации C5 (50-кратный молярный избыток, приблизительно 25 мкМ). Несмотря на отличие молекулярной массы на 10000, ARC657 (PEG массой 20 кДа) (SEQ ID NO:61)
и ARC658 (PEG массой 30 кДа) (SEQ ID NO:62) обладали сходными значениями экспозиции (AUC), t1/2(α) и t1/2 (β). В противоположность этому ARC187 (SEQ ID NO:5)
обладал значительно более высокими значениями экспозиции (AUC), увеличенным t1/2(α) и немного увеличенным t1/2(β), чем другие молекулы.
Затем дополнительные аликвоты образцов плазмы, собранные в ходе фармакокинетического исследования, анализировали in vitro для определения эффективности блокады C5 приматов. Анализ активации зимозаном проводили, как описано выше, для определения количества образовавшегося C5b-9 и C5a приматов, соответственно. Данные наносили на график в нескольких различных форматах, включая концентрацию C5b-9 против времени забора образца (Фиг.43a), концентрацию C5b-9 против концентрации аптамера (Фиг.43b), концентрацию C5a против времени забора образца (Фиг.43c) и концентрацию C5a против концентрации аптамера (Фиг.43d).
Аптамер ARC187 с PEG массой 40 кДа (SEQ ID NO:5) ингибировал расщепление C5 примата (концентрация C5b-9 и C5a) в течение наиболее длительного периода времени (Фиг.43a и 43c). Когда данные C5b-9 и C5a нанесли на график против концентрации аптамера, они показали, что концентрация блокирующего C5 аптамера должна была превышать 30-кратный молярный избыток, независимо от размера молекул PEG, для полного ингибирования расщепления C5 (Фиг.43b и 43d).
В целом, денные исследования PK/PD у яванских макак показывают, что (a) как и ожидалось, по меньшей мере 30-кратный избыток аптамера (концентрация аптамера в плазме приблизительно 15 мкМ) был необходим для ингибирования расщепления C5 in vivo у яванской макаки, независимо от размера группы PEG, (b) блокирующие C5 аптамеры не вызывали очевидной токсичности у этого вида, и (c) когда животным вводили относительно высокие уровни (50-кратный молярный избыток), уровни аптамера в плазме находились в пределах соответствующего диапазона анализа в течение периода забора образцов, позволяя вычислить фармакокинетические параметры.
Пример 5F
Фармакокинетика и фармакодинамика ингибиторов C5 ARC658 и ARC187 у яванских макак после внутривенного введения - исследование 2
Схема исследования 2 была сходной с исследованием 1, описанным выше, со следующими исключениями a) оценивали только два соединения (ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5); b) количество животных было увеличено до четырех в группе; и c) забор образца плазмы через 1 минуту был исключен и заменен забором образца через 144 часа для того, чтобы обеспечить вычисление терминального времени полужизни на основе большего количества результатов. Способы изготовления и дозирования этих двух аптамеров, забора образцов крови и выделения плазмы были идентичны способам, описанным выше в исследовании 1. Схема исследования 2 обобщена на Фиг.44.
После завершения исследования 2, аликвоты плазмы анализировали, как описано в исследовании 1, для определения a) концентрации аптамера в плазме в различные моменты времени после внутривенного введения, и b) эффективности блокады C5.
Концентрацию аптамера в плазме наносили на график как функцию времени (Фиг.45) и первичные данные для ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5) представлены в форме таблицы на Фиг.39 и 40, соответственно. Аптамер ARC187 с PEG
массой 40 кДа (SEQ ID NO:5) находился в плазме в течение наиболее длительного периода времени. Исследование данных, представленных на Фиг.45, указывает на то, что данные более всего соответствуют двухкомпартментной модели. Таким образом, оценка фармакокинетических параметров, описанных на Фиг.46, была проведена на основе двухкомпартментной модели с использованием WinNonLinTM v.4.0 (Pharsight Corp., Mountain View, CA).
При сравнении фармакокинетических параметров, полученных в ходе исследования PK/PD 1 и 2, представленных выше, данные для ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5) были сходны за исключением показателя t1/2(α) для ARC187. Без связи с какой-либо теорией, отличие показателей t1/2(α) для ARC187 в двух исследованиях, вероятно, является следствием небольшого размера выборки в предварительном исследовании.
Как показано на Фиг.46, значения Cmax были сходны для ARC658 (SEQ ID NO:62) и ARC187 (SEQ ID NO:5). Напротив, экспозиция лекарственного средства (AUC) была значительно более высокой у животных, которым вводили ARC187 (SEQ ID NO:5). Также, ARC187 обладал увеличенными показателями t1/2(α) и t1/2(β) по сравнению с ARC658 (SEQ ID NO:62). Эти данные совместно с данными, полученными в ходе исследования PK/PD 1, указывают на то, что блокирующие C5 аптамеры ARC187 могут обеспечить наиболее эффективную блокаду C5 in vivo в данной дозе.
Затем дополнительные аликвоты образцов плазмы, собранных в ходе фармакокинетического исследования, анализировали in vitro для определения эффективности блокады C5 приматов. Как и ранее, анализ активации зимозаном проводили для определения количества образовавшегося C5b-9 и C5a примата, соответственно. Данные наносили на график в качестве концентрации C5b-9 против концентрации аптамера (Фиг.47) и концентрации C5a против концентрации аптамера (Фиг.48). Как ранее показано в ходе исследования PK/PD 1, для полного ингибирования расщепления C5 примата концентрация блокирующего C5 аптамера должна превышать 30-кратный молярный избыток (концентрация аптамера к концентрации C5 в плазме) или составлять приблизительно 15 мкМ, независимо от размера молекулы PEG (Фиг.41 и 42).
При рассмотрении данных, приведенных в таблицах Фиг.39 и 40, очевидно, что после внутривенного болюса 30-мг/кг концентрация ARC658 (SEQ ID NO:62) оставалась выше 15 мкМ приблизительно в течение 4 часов, в то время как концентрация ARC187 оставалась выше 15 мкМ приблизительно в течение 8 часов. Таким образом, с учетом сходной дозы лекарственного средства, аптамер 40K ARC187 обеспечивает клиническую эффективность приблизительно в два раза дольше, чем аптамер 30K ARC658 (SEQ ID NO:62).
В целом, для блокирования превращения C5 in vivo яванским макакам необходимо вводить по меньшей мере 30-кратный молярный избыток аптамера относительно C5 в плазме. Эти данные согласуются с предшествующими исследованиями in vitro (гемолиз) и ex-vivo (изолированное перфузируемое сердце мыши), которые позволяют предположить, что связывающие C5 аптамеры обладают более низкой аффинностью к C5 примата относительно C5 человека. Было показано, что блокирующие C5 аптамеры можно безопасно доставлять в качестве внутривенного болюса в дозе вплоть до 30 мг/кг, которая сопоставима приблизительно с 50-кратным молярным избытком аптамера относительно концентрации C5.
Пример 5G
ARC1905 у яванской макаки после внутривенного болюсного введения
Фармакодинамику ингибиторов C5 ARC1905 оценивали у яванской макаки после внутривенного введения. Состав ARC1905 для инъекции был изготовлен в стандартном физиологическом растворе в концентрации 7,5 мг/мл и составы для дозирования подвергали стерильному фильтрованию (0,2 мкм) в предварительно стерилизованные флаконы в аспетических условиях. Яванским макакам (n=4) проводили введение дозы 0 (контроль в виде физиологического раствора) или 30 мг/кг посредством внутривенного болюсного введения. Образцы крови получали из периферической вены или артериального отверстия и образцы крови (0,5 мл) переносили в пробирки с дикалий(K2)ЭДТА, помещенные на влажный лед, и центрифугировали в течение 30 минут после сбора приблизительно при 4°C.
Образцы плазмы анализировали in vitro для определения эффективности ARC1905 в отношении блокады C5 приматов. Для определения количества образовавшегося C5a примата использовали анализ с зимозаном, описанный ранее в отношении ARC1905 в примере 1C. Снижение значений для C5a после зимозана через 0,5 и 2 часа после дозирования указывает на то, что ARC1905 ингибирует расщепление C5 in vivo у яванской макаки сходным образом с ARC187 при дозировании приблизительно в той же концентрации и с тем же способом введения, при определении in vitro с использованием анализа активации зимозаном.
Пример 5H
Фармакокинетика и фармакодинамика ингибитора C5 ARC187 у яванской макаки после внутривенного болюсного введения и инфузии
Фармакокинетический (PK) и фармакодинамический (PD) профили ARC187 (SEQ ID 20 NO: 5) также оценивали у яванских макак после внутривенного нагрузочного болюса со следующим немедленно началом внутривенной инфузии. Схема этого исследования представлена на Фиг.49.
Нагрузочную болюсную дозу и скорость инфузии, необходимые для достижения надлежащей устойчивой концентрации в плазме, составляющей 1 мкМ, вычисляли с использованием фармакокинетических параметров, полученных из исследования только внутривенного болюса, представленного на Фиг.50.
Всего трем яванским макакам вводили внутривенный болюс ARC187 в концентрации 1 мг/кг с последующим немедленным началом внутривенной инфузии со скоростью 0,0013 мг/кг/мин в течение 48 ч. Образцы цельной крови забирали от 0 до 192 часов после введения, хранили на льду, обрабатывали для получения плазмы, а затем хранили в замороженном виде при -80°С. Концентрацию ARC187 в образцах плазмы определяли с использованием как анализа флуоресцентного окрашивания нуклеиновых кислот (описанного в примере 5B), так и анализа GLP-валидированной жидкостной хроматографии (ВЭЖХ). Способ анализа ВЭЖХ для определения ARC187
в плазме обезьяны был проверен ClinTrials Bio-Research (Montreal, Canada). Валидированное исследование согласовывалось с правилами United States Food and Drug Administration (FDA) Good Laboratory Practice (GLP) (21 CFR §58). Способ анализа ВЭЖХ был проверен в отношении: селективности, линейности, нижней границы количественного определения (LLOQ), уноса, точности и аккуратности анализа, стабильности исходных растворов, стабильности инъецируемых материалов, кратковременной стабильности матрикса, стабильности в отношении размораживания-оттаивания, длительной стабильности матрикса и точности разведения. Пригодный линейный диапазон динамической концентрации анализа был определен как от 0,080 до 50,0 мкМ.
Измеренный профиль PK ARC187 в этих условиях значительно соответствовал вычисленному профилю, полученному с использованием параметров PK только внутривенного болюса (см. Фиг.51). Намеченная концентрация в плазме, составляющая 1 мкМ, устанавливалась в течение <5 мин после дозирования и поддерживалась в течение всего продолжения инфузии. После прекращения инфузии, аптамер показал терминальное время полувыведения, t1/2(β) ~40-60 ч.
Фармакодинамическую активность ARC187 (SEQ ID NO:5) у яванской макаки оценивали ex-vivo с использованием образцов плазмы, полученных в ходе исследования PK в анализе активации зимозаном, описанном ранее, с модификацией, состоящей в том, что образец плазмы яванской макаки разбавляли в 10-раз в 10% плазме человека, а затем обрабатывали 5 мг/мл зимозана. Активацию C5, отражаемую появлением продукта расщепления C5a, измеряли посредством ELISA, специфичного к C5a человека (набор C5a ELISA, BD Biosciences, San Diego, CA). Затем в каждом образце количественно определяли концентрацию активного ARC187 посредством сравнения со стандартной кривой, полученной из анализов с зимозаном с использованием полученных образцов с известными уровнями ARC187 (см. Фиг.52). Это исследование указывает на то, что ARC187 сохраняет его активность против комплемента во время и после инфузии, на уровнях, по существу согласующихся с фармакокинетическим профилем, описанным выше.
Пример 5I
Предсказание требований к дозированию у человека
Требования к дозированию у человека для профилактики, смягчения или лечения осложнений, связанных с хирургической операцией CABG, основаны на следующих доводах: во-первых, пациентам с CABG будут вводить однократную внутривенную болюсную дозу аптамера против C5 перед началом хирургической операции с последующей непрерывной инфузией для установления и поддержания устойчивой концентрации в плазме 1,5 мкМ в течение 24-48 часов после хирургической операции CABG. Болюсная доза и скорость инфузии основаны на вычислениях с использованием фармакокинетических параметров, полученных из ранее описанных исследований только внутривенного болюса и болюса плюс инфузия у яванских макак. Оцененная болюсная доза ARC187 составляет 1 мг/кг, и обусловленная ей скорость инфузии составляет 0,0013 мг/кг/мин. Для этой схемы из болюса плюс 48-ч инфузия ожидаемое требуемое общее количество лекарственного средства составляет 0,4 г для ARC187, где масса относится только к массе олигонуклеотида (см. столбец 7 в таблице Фиг.53). Столбец 2 таблицы, представленной на Фиг.53, относится к массе группы PEG, конъюгированной с олигонуклеотидной частью ARC187, третий столбец относится к молекулярной массе олигонуклеотидной части ARC187 (и она в данном случае будет одинаковой для всех аптамеров, которые содержат ARC186 (SEQ ED NO:4) в качестве их олигонуклеотидной последовательности), столбец 4 относится к молекулярной массе PEG массой 40 кДа конъюгированного с ARC186 (SEQ ID NO:4) через химическую группу амина, как описано в примере 3C, выше, 5 столбец относится к времени полужизни ARC187 α-фазы в двухкомпартментной модели, и шестой столбец относится к времени полужизни β-фазы ARC187 в двухкомпартментной модели.
ПРИМЕР 6
Взаимодействие аптамеров против C5 и гепарина/протамина
Одним предполагаемым применением аптамера против C5 является применение в качестве профилактического средства для предотвращения или смягчения воспалительных побочных эффектов, обусловленных хирургической операцией по аортокоронарному шунтированию (CABG). Как правило, в процессе CABG вводят высокие концентрации антикоагулянта гепарина (3-5 единиц/мл или 1-2 мкМ) для профилактики тромбоза и поддержания проходимости в компонентах обходного насоса; реверсию эффекта гепарина после операции и восстановление нормального гемостаза обеспечивают посредством введения аналогично высоких концентраций протамина (~5 мкМ). С учетом потенциальных опасностей для пациентов какого-либо вмешательства в эффективность какого-либо из этих лекарственных средств было необходимо показать, что аптамеры против C5 (1) не изменяют активность какого-либо лекарственного средства и (2) не оказывают собственных эффектов на гемостаз, которые могут осложнить противосвертывающее лечение у пациента.
Гепарин представляет собой сульфатированный полисахарид с отрицательным полным зарядом и средней молекулярной массой приблизительно 15 кДа, который оказывает ингибиторный эффект на ряд протеаз каскада свертывания крови, запуская взаимодействия с антитромбином. Протамин, высокоположительно заряженный полипептид, способен блокировать активность гепарина посредством мало охарактеризоанного взаимодействия, которое по меньшей мере частично является электростатическим. Функциональная основа ARC187 (SEQ ID NO:5), аналогично гепарину, является высокоанионной. Таким образом, возможно, что ARC187 может неспецифично связываться со связывающими гепарин участками или протамином и препятствовать активности этих молекул. В последующих исследованиях изучали собственные (т.е., гепарин-подобные) противосвертывающие свойства ARC187, эффекты ARC187 на функцию, эффекты ARC187 на нейтрализацию гепарина протамином и эффекты протамина на свойства ARC187 в отношении ингибирования комплемента.
Пример 6A
Эффекты ARC187 на свертывание in vitro
Собственные эффекты ARC187 (SEQ ID NO:5) на способность плазмы к свертыванию исследовали с использованием стандартных клинических тестов для внешних и внутренних звеньев каскада свертывания: протромбинового времени (PT) и активированного частичного тромбопластинового времени (aPTT), соответственно. Как показано на Фиг.54, титрование цитратной плазмы человека концентрациями, значительно превышающими предполагаемые дозы (вплоть до 20 мкМ), не вызвало изменений PT и вызывало только небольшое повышение aPTT.
Для оценки эффектов ARC187 на функции гепарина и протамина in vitro, проводили забор крови от 3 индивидов в гепарин в дозе 4-5 единиц/мл, ассоциированной с уровнями гепарина, используемыми в хирургической операции CABG. Способность к свертыванию в этих образцах оценивали с использованием активированного времени свертывания (ACT), теста коагуляции цельной крови, обычно используемого для мониторинга активности гепарина в ходе хирургической операции. В этих концентрациях гепарина, в отсутствие других добавок, ACT было значительно увеличено по сравнению с исходным уровнем ~150 секунд до ~500 секунд в присутствии 4 Ед/мл гепарина или ~800 секунд в присутствии 5 Ед/мл гепарина. Добавление 10 мкМ ARC187 к этим образцам оказало небольшой эффект на время свертывания, показывая, что ARC187 не препятствует противосвертывающей активности гепарина.
Противосвертывающий эффект гепарина легко нейтрализовывался титрованием протамином вплоть до 6-8 мкМ (4 Е/мл гепарина) или 12 мкМ (5 Е/мл гепарина). Показатели ACT в присутствии гепарина и нейтрализующие концентрации протамина по существу не отличались от исходного уровня. Поскольку основа ARC187, представляющая собой нуклеиновую кислоту (12 кДа), обладает большей молекулярной массой, чем протамин (5 кДа), можно ожидать, что эквимолярные концентрации ARC187, добавленные к протамину, будут достаточными для полной реверсии нейтрализующей активности протамина.
Однако предварительная инкубация протамина приблизительно с эквивалентными концентрациями ARC187 оказала небольшой эффект на ACT. Образцы крови, содержащие нейтрализующие концентрации протамина, показывали сходные значения ACT в присутствии или отсутствие 10 мкМ ARC187, указывая на то, что ARC187 оказывает только слабый эффект на прокоагулянтную активность протамина. Эти результаты обобщены на Фиг.55.
Пример 6B
Эффекты ARC187 на коагуляцию in vivo
Взаимодействия между функцией гепарина и протамина в ходе сопутствующего введения аптамера против C5 ARC187 (SEQ ID NO:5) в клинических дозах гепарина и клинических/субклинических/надклинических дозах протамина исследовали для определения того, препятствует ли наличие субклинических/надклинических концентраций ARC187 в плазме реверсии противосвертывающего действия гепарина посредством протамина. Результаты исследования обобщены на Фиг.56. В кратком изложении исходные значения ACT не изменялись посредством 10 мкМ (т.е., 10-кратного молярного избытка клинической дозы) ARC187 при всех тестируемых дозах гепарина. Аналогично, степень противосвертывающего действия гепарина не изменялась посредством 10 мкМ ARC187. В отсутствие ARC187 минимальная эффективная доза протамина составляла ~30% (клиническая доза=100%). Более того, реверсия противсвертывающего действия гепарина посредством 30% протамина не изменялась посредством 10-кратного молярного избытка клинической дозы (т.е., 10 мкМ) ARC187. Таким образом, на применение ARC187 для ингибирования комплемента в клинических условиях (например, CABG) не должно влиять сопутствующее применение гепарина и протамина в обычных дозах.
Пример 6C
Эффект гепарина и протамина на функцию ARC187, направленную против комплемента
Эффекты гепарина и протамина на активность ARC187 (SEQ ID NO:5), направленную против комплемента, исследовали в образцах цитратной цельной крови, активированных зимозаном, как описано в примере 1. Непосредственно перед активацией зимозана ARC187 титровали в образцы цитратной крови, обработанной в условиях: 1) без обработки (без гепарина или протамина); 2) 4 Ед/мл гепарина; 3) 6 мкМ протамин; 4) 4 Ед/мл гепарина + 6 мкМ протамин. После активации зимозаном активацию C5 количественно определяли посредством измерения с помощью ELISA sC5b-9 в плазме (набор C5b-9 ELISA, Quidel, San Diego, CA). Для каждого условия результаты, выраженные в качестве процентного ингибирования активации C5 против концентрации ARC187, были неотличимы в пределах погрешности (см. Фиг.57). Во всех случаях полного ингибирования достигали посредством 1-2 мкМ ARC187. Таким образом, оказалось, что гепарин и протамин, отдельно или в сочетании, в концентрациях, соответствующих их применению в хирургической операции CABG, влияют на активность ARC187, направленную против комплемента.
ПРИМЕР 7
Хориоидальная неоваскуляризация
Индуцируемую лазером хориоидальную неоваскуляризацию часто используют в качестве модели, связанной со старением дегенерации желтого пятна. Также ее можно использовать в качестве модели диабетической ретинопатии. Эффект введения средств против C5, которые, в предпочтительных вариантах осуществления настоящего изобретения, представляют собой аптамеры против комплемента, описанные в настоящем документе, на предотвращение, а также на стабилизацию и/или регрессию хориоидальной неоваскуляризации, оценивают в этой модели, как описано ниже и Rrzystolik, M.G. et al., Arch OpthalmoL, vol. 120, pp 338-346 (2002).
В случае, когда оценивают профилактику хориоидальной неоваскуляризации, средство против C5, в частности специфичный к C5 аптамер, который связывает и ингибирует функцию белка C5 комплемента яванской макаки, инъецируют в глаз каждой яванской макаки, в то время как в контрольный глаз вводят носитель. Через от нескольких суток до нескольких недель после инъекции аптамера проводят лазерную фотокоагуляцию на обоих глазах каждой яванской макаки. Мониторинг глаз каждого животного проводят посредством офтальмологического исследования, цветной фотосъемки и ангиографии с флуоресцеином. Если встречаемость хориоидальной неоваскуляризации (оцененной ангиографически) значительно ниже в глазу, в который вводили средство против C5, в частности специфичный к C5 аптамер, чем в контрольном глазу, специфичное средство против C5 считают эффективным. Если оценке подлежит профилактика хориоидальной неоваскуляризации в случае введения сочетания средства против C5 и средства против VEGF и/или средства против PDGF, следуют указанному выше процессу, за исключением того, что в один глаз животного вводят средство против C5 и средство против VEGF и/или средство против PDGF от нескольких суток до нескольких недель перед лазерной фотокоагуляцией.
В другом варианте осуществления, когда оценивают профилактику хориоидальной неоваскуляризации, аптамер против комплемента, в частности аптамер, который ингибирует функцию белка комплемента яванской макаки, такого как C5 или C3, инъецируют в стекловидное тело одного глаза каждой яванской макаки, в то время как в контрольный глаз вводят носитель. От нескольких суток до нескольких недель после инъекции аптамера проводят лазерную фотокоагуляцию на обоих глазах яванской макаки. Мониторинг глаз каждого животного проводят посредством офтальмологического исследования, цветной фотосъемки и ангиографии с флуоресцеином. Если встречаемость хориоидальной неоваскуляризации (оцененной ангиографически) значительно ниже в глазу, в который вводили аптамер против комплемента, чем в контрольном глазу, аптамер против комплемента считают эффективным. Если оценке подлежит профилактика хориоидальной неоваскуляризации в случае введения сочетания аптамера против комплемента и средства против VEGF и/или средства против PDGF, следуют указанному выше процессу, за исключением того, что в один глаз животного вводят аптамер против комплемента и средство против VEGF и/или средство против PDGF от нескольких суток до нескольких недель перед лазерной фотокоагуляцией.
Когда оценке подлежит стабилизация и/или регрессия хориоидальной неоваскуляризации, на обоих глазах каждой яванской макаки проводят лазерную фотокоагуляцию. От нескольких суток до нескольких недель после лазерной фотокоагуляции средство против C5 по настоящему изобретению вводят посредством инъекции в стекловидное тело одного глаза каждого животного, в то время как в другой глаз вводят носитель. В одном варианте осуществления это средство против C5 представляет собой аптамер против комплемента. В предпочтительном варианте осуществления указанный аптамер против комплемента представляет собой ингибирующий C5 и/или C3 аптамер. Мониторинг глаз каждого животного проводит посредством офтальмологического исследования, цветной фотосъемки и ангиографии с флуоресцеином. Если встречаемость хориоидальной неоваскуляризации (оцененная ангиографически) является такой же и/или значительно более низкой в глазу, в который вводили средство против C5, в частности аптамер против C5, чем в контрольном глазу, тогда аптамер считают эффективным в отношении стабилизации и/или регрессии, соответственно. Если оценке подлежит стабилизация и/или регрессия хориоидальной неоваскуляризации в отношении лечения сочетанием средства против C5 и средства против VEGF и/или средства против PDGF, следуют указанному выше процессу, за исключением того, что в один глаз животного вводят средство против C5 и средство против VEGF и/или средство против PDGF через от нескольких суток до нескольких недель после лазерной фотокоагуляции.
Аналогично оценке у яванских макак, описанной выше, эффективность средств против C5 и аптамеров против комплемента в отношении профилактики, стабилизации и/или регрессии хориоидальной неоваскуляризации отдельно или в сочетании со средством против VEGF и/или средством против PDGF, можно оценивать у мышей или других видов с использованием средства против C5, которое модулирует белок C5 комплемента мыши, или белок C5 комплемента другого вида. В другом варианте осуществления аналогично той же оценке у яванских макак, описанной выше, эффективность аптамера против комплемента в отношении профилактики, стабилизации и/или регрессии хориоидальной неоваскуляризации отдельно или в сочетании со средством против VEGF и/или средством против PDGF, можно оценивать у мышей или других видов с использованием аптамера против комплемента, который модулирует, в частности ингибирует, представляющий интерес белок комплемента мыши, или представляющий интерес белок комплемента другого вида. См., например, Bora et al., Journal of Immunolgy, 174: 491-497 (2005).
ПРИМЕР 8
Модель неэкссудативной AMD с дегенерацией сетчатки у мышей
Модель на мышах, имеющих мутацию либо в моноцитарном хемоаттрактантном белке 1 (MCP-1 или Ccl-2), либо в распознаваемом им рецепторе C-C-хемокинов 2 (Ccr-2), имитирует симптомы связанной со старением дегенерации желтого пятна человека, включая развитие друз, атрофию фоторецепторов и хориоидальную неоваскуляризацию. См. Ambati et al., Nature Medicine. 2003 Nov 2003; 9(11): 1390-7. Эта модель показывает значительное накопление C5 в пигментном эпителии сетчатки и сосудистой оболочке, указывая на то, что при заболевании экспрессируется комплемент. Кроме того, CD46 (мембраносвязанный регулятор комплемента), витронектин (регулатор MAC) и C3c (продукт деградации C3b) присутствуют в пигментном эпителии сетчатки и/или сосудистой оболочке, подтверждая, что произошла активация комплемента.
Когда оценке подлежит стабилизация и/или регрессия дегенерации сетчатки, аптамер против комплемента по этому изобретению, например аптамер, ингибирующий C5 или C3 мыши, вводят посредством инъекции в один глаз каждого животного, в то время как в другой глаз вводят носитель. Проводят мониторинг глаз в отношении дегенерации сетчатки каждого животного, включая накопление продуктов комплемента в RPE/сосудистой оболочке, развитие аномальной электрофизиологии и/или локализованной атрофии RPE и/или фоторецепторов, и встречаемость хориоидальной неоваскуляризации. Если встречаемость дегенерации сетчатки является такой же и/или значительно более низкой в глазу, в который вводили аптамер против комплемента, в частности аптамер против C5 или против C3, чем в контрольном глазу, аптамер считают эффективным в отношении стабилизации и/или регрессии соответственно.
После описания этого изобретения посредством письменного описания и примеров, специалисты в данной области поймут, что настоящее изобретение можно применять на практике во множестве вариантов осуществления, и что описание и примеры, представленные выше, предназначены только для целей иллюстрации, а не для ограничения следующей ниже формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| АПТАМЕРНЫЕ ТЕРАПЕВТИЧЕСКИЕ СРЕДСТВА, ПРИМЕНИМЫЕ ДЛЯ ЛЕЧЕНИЯ СВЯЗАННЫХ С КОМПЛЕМЕНТОМ РАССТРОЙСТВ | 2006 |
|
RU2406507C2 |
| НОВЫЕ СВЯЗЫВАЮЩИЕ C5a НУКЛЕИНОВЫЕ КИСЛОТЫ | 2013 |
|
RU2645261C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ДЕГО | 2011 |
|
RU2570390C2 |
| АНТИ-C5a-АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2773779C2 |
| АНТИТЕЛА ПРОТИВ C3b И СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СВЯЗАННЫХ С КОМПЛЕМЕНТОМ НАРУШЕНИЙ | 2008 |
|
RU2473563C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| ГУМАНИЗИРОВАННАЯ МОДЕЛЬ НАРУШЕНИЙ СО СТОРОНЫ ПОЧЕК И ПЕЧЕНИ | 2018 |
|
RU2772583C2 |
| SDF-1-СВЯЗЫВАЮЩИЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2007 |
|
RU2590709C2 |
| ГЕНЕТИЧЕСКИЕ ПОЛИМОРФИЗМЫ ПРИ ВОЗРАСТНОЙ ДЕГЕНЕРАЦИИ ЖЕЛТОГО ПЯТНА | 2010 |
|
RU2577726C2 |
| ГЕНЕТИЧЕСКИЕ ПОЛИМОРФИЗМЫ ПРИ ВОЗРАСТНОЙ ДЕГЕНЕРАЦИИ ЖЕЛТОГО ПЯТНА | 2010 |
|
RU2546008C2 |
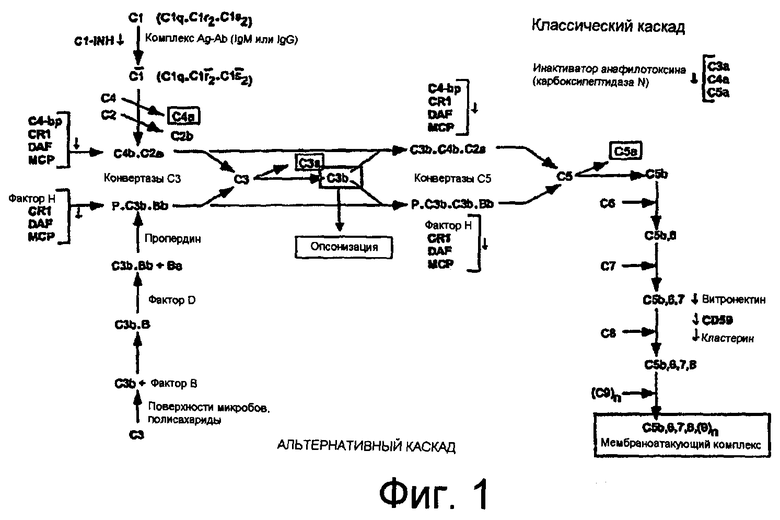
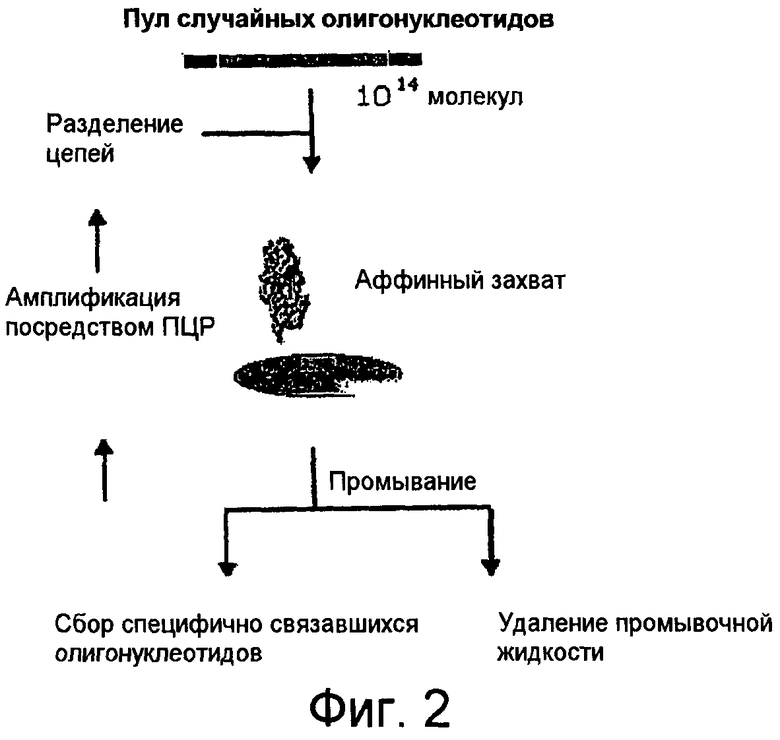
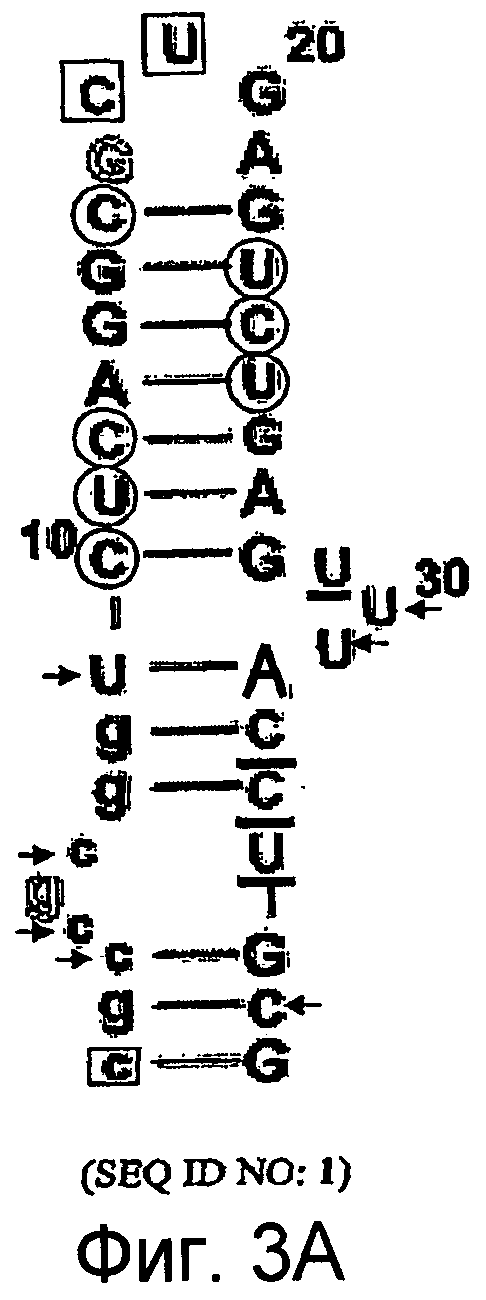
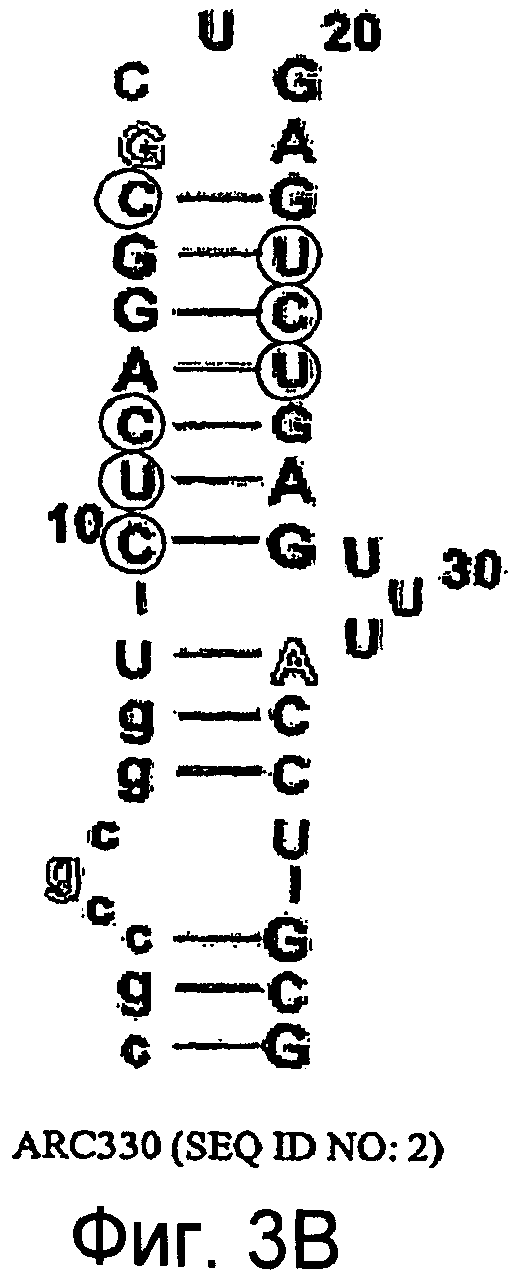
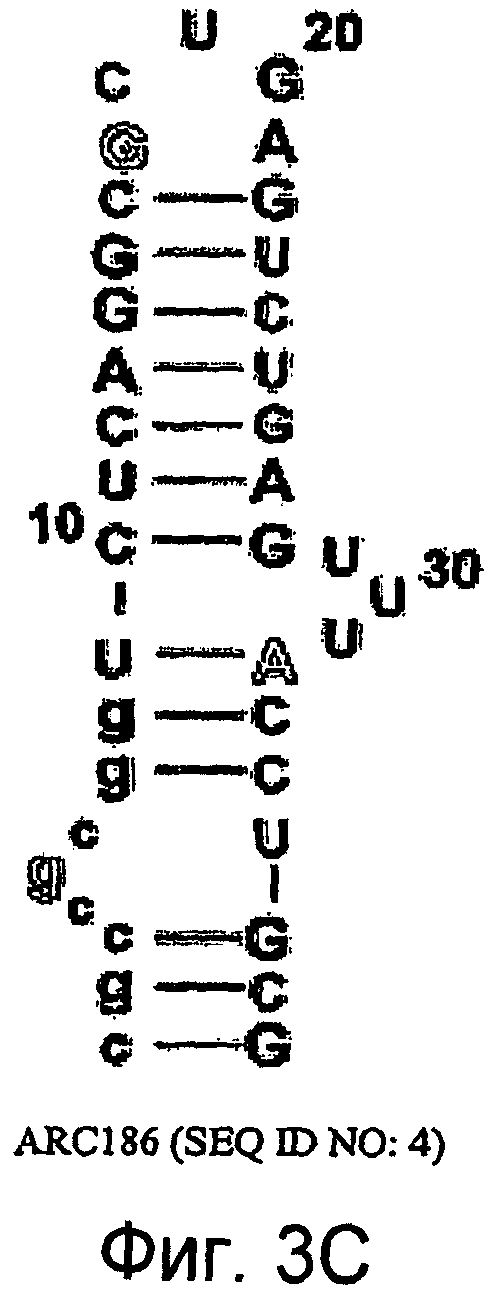
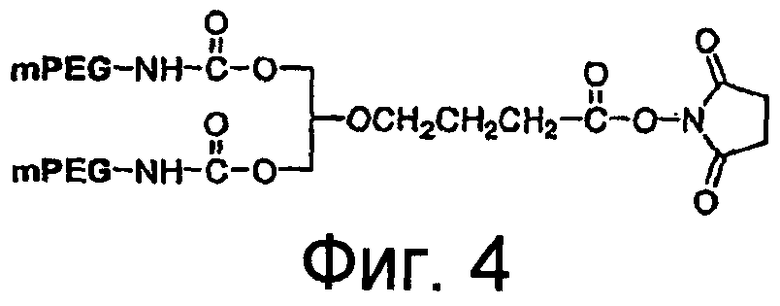
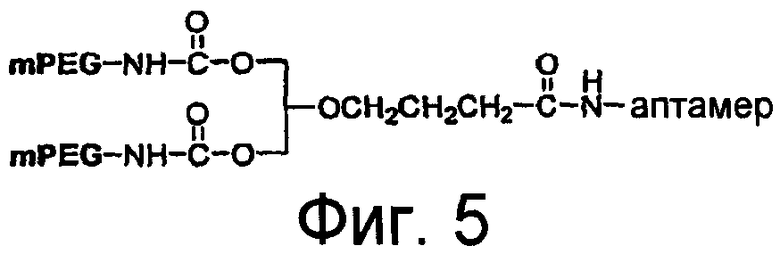
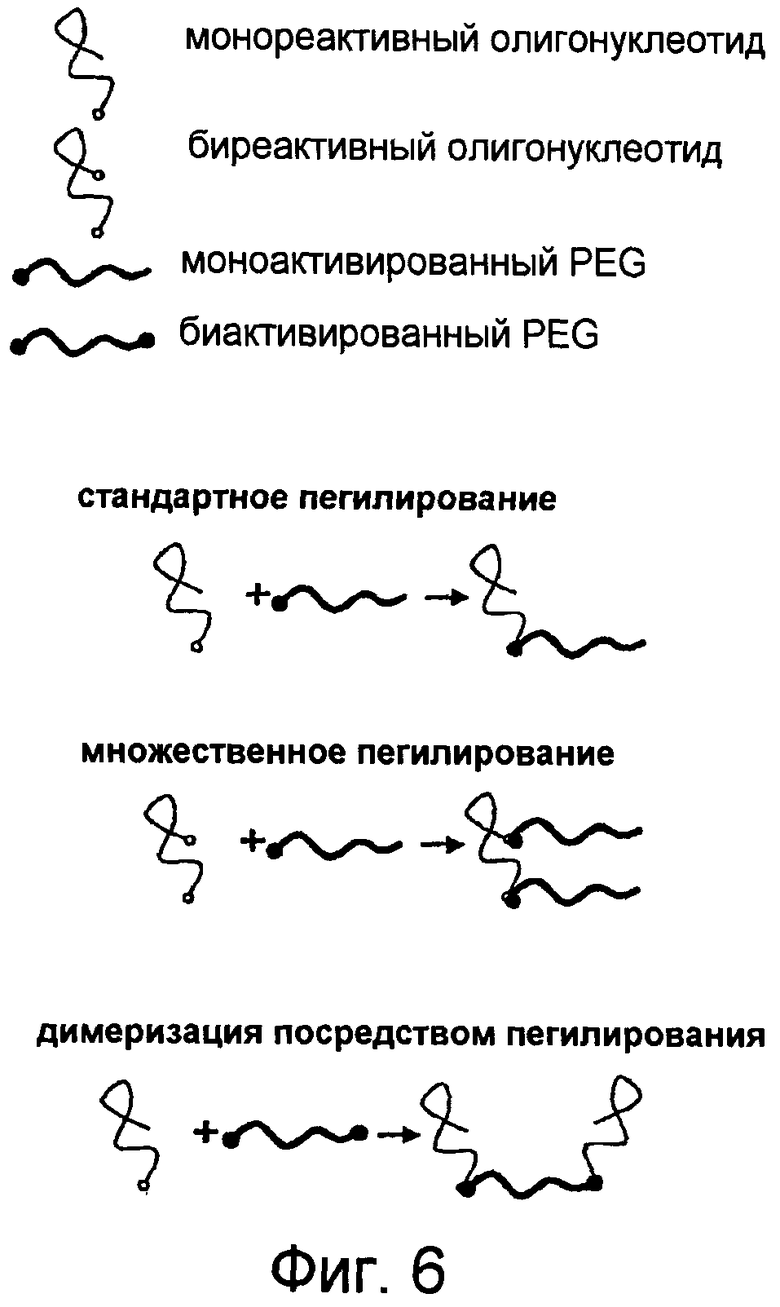
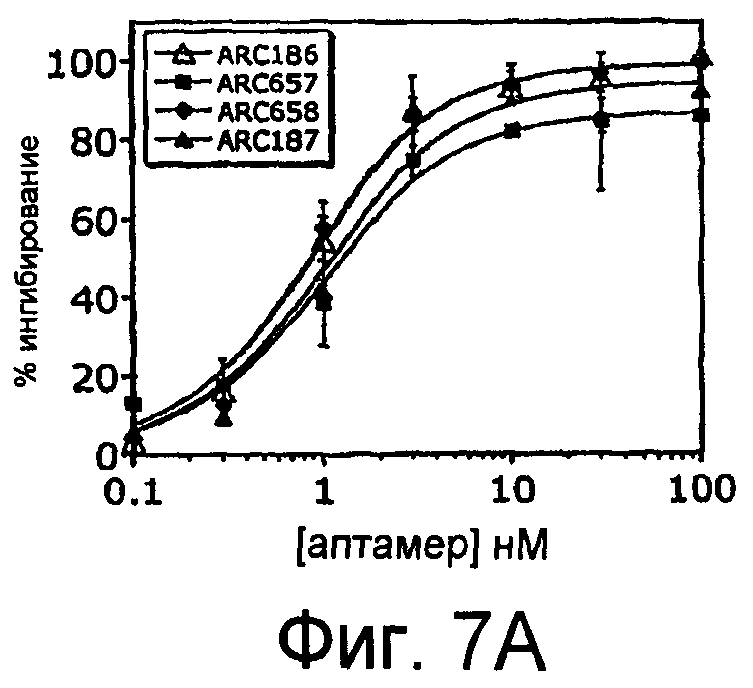

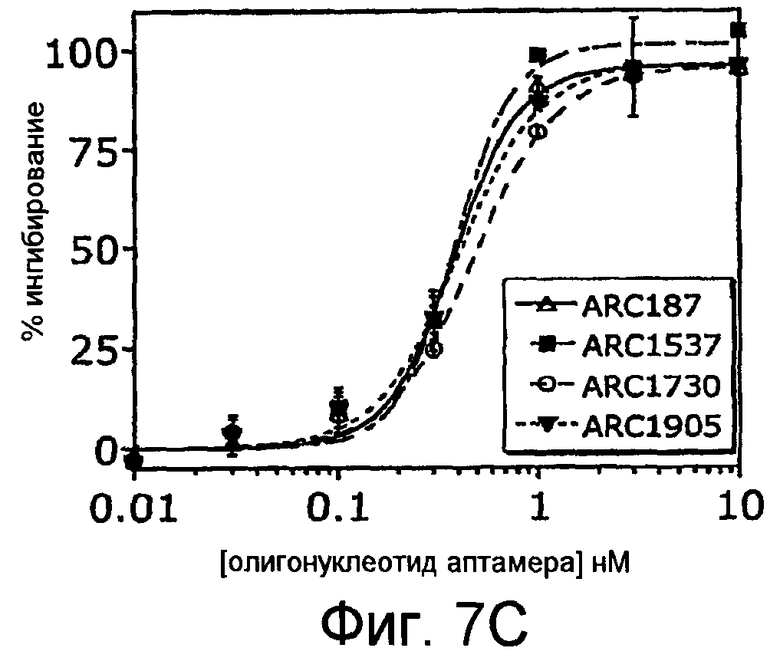

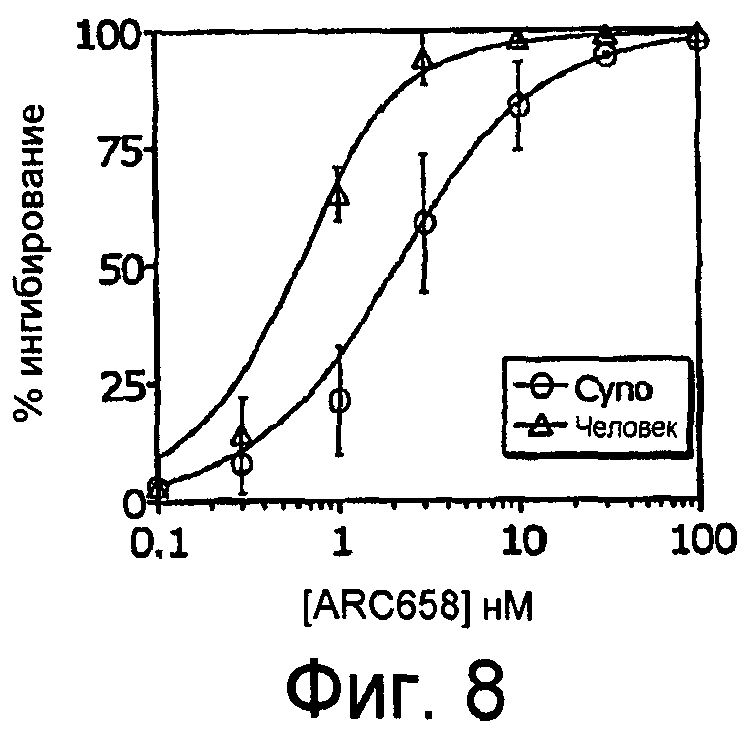
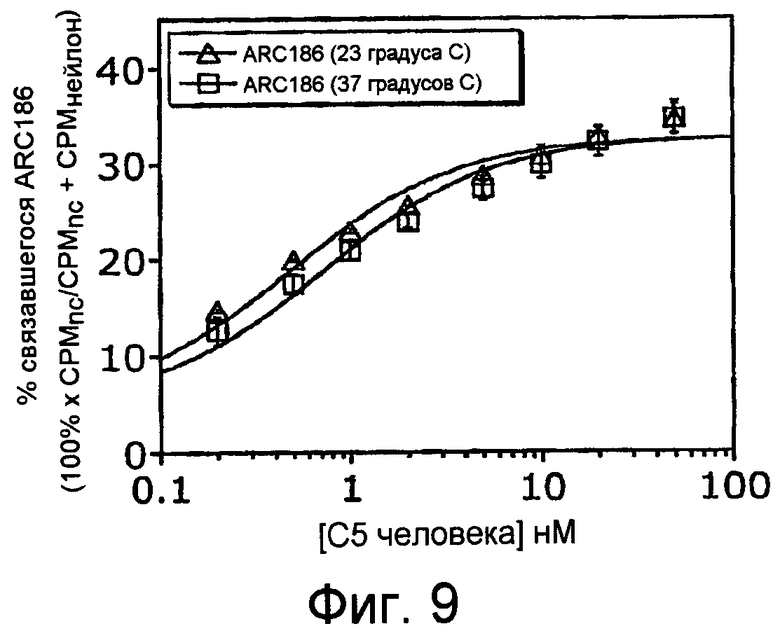
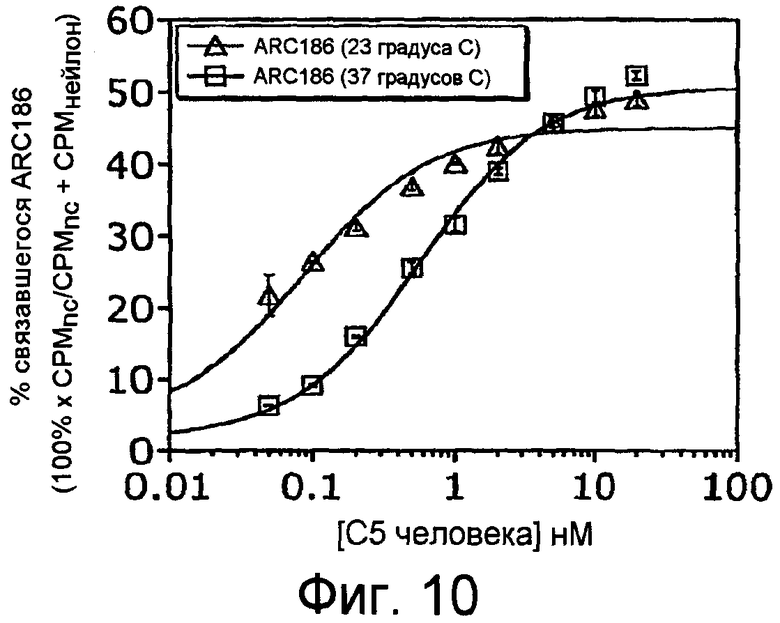
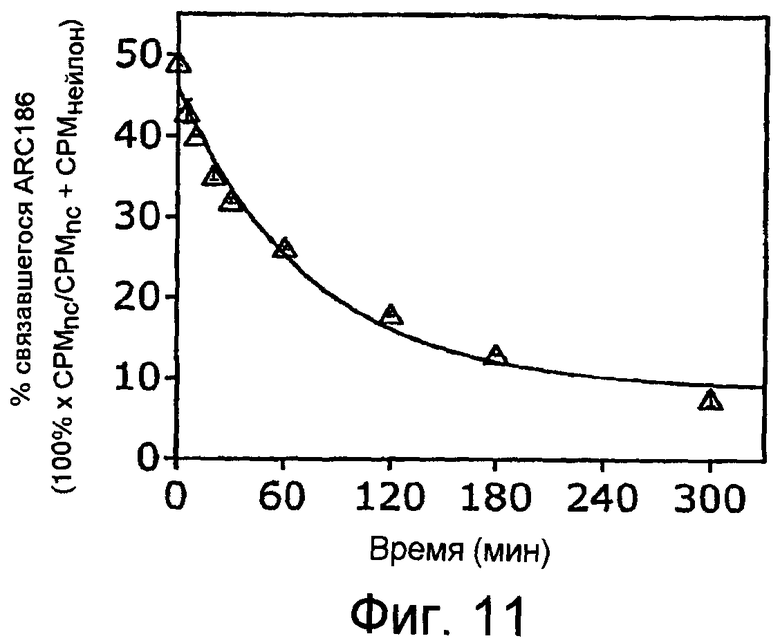
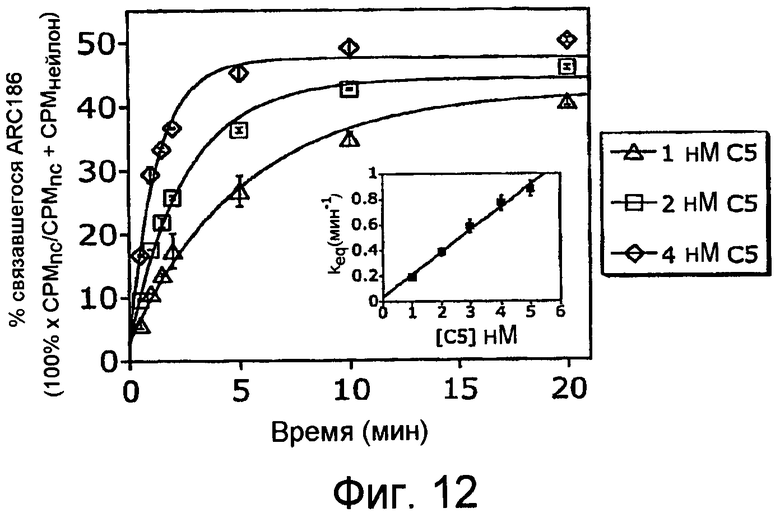
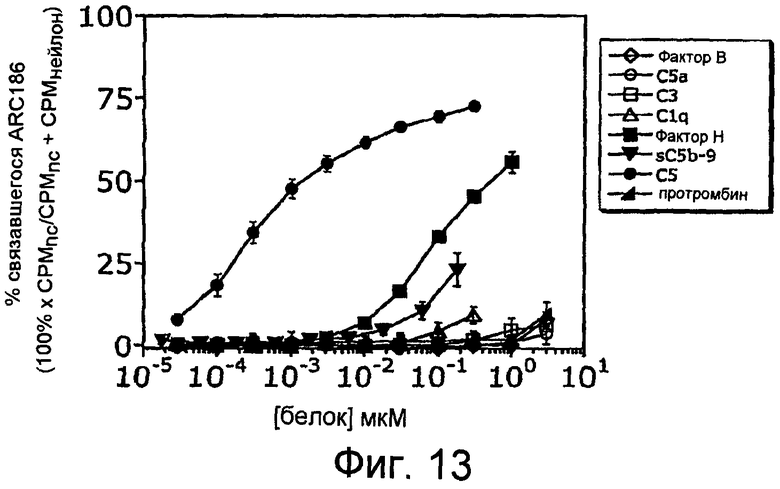
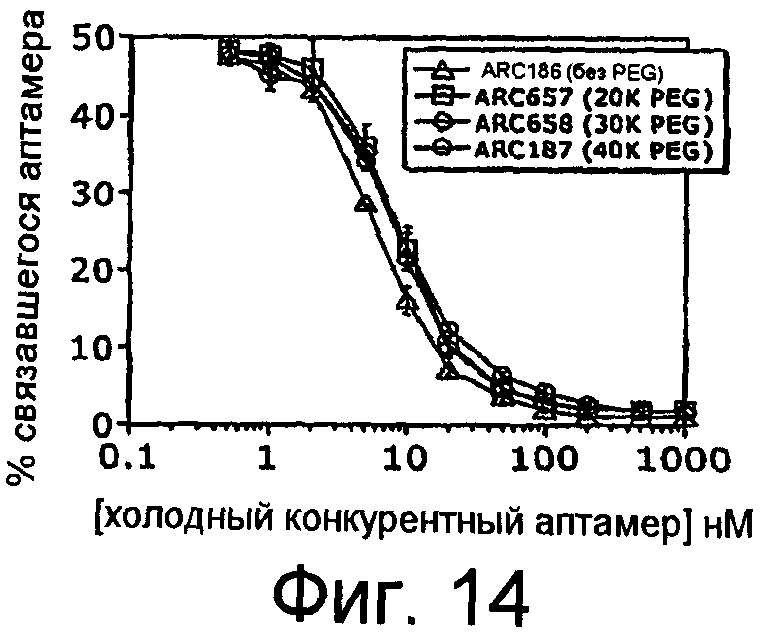
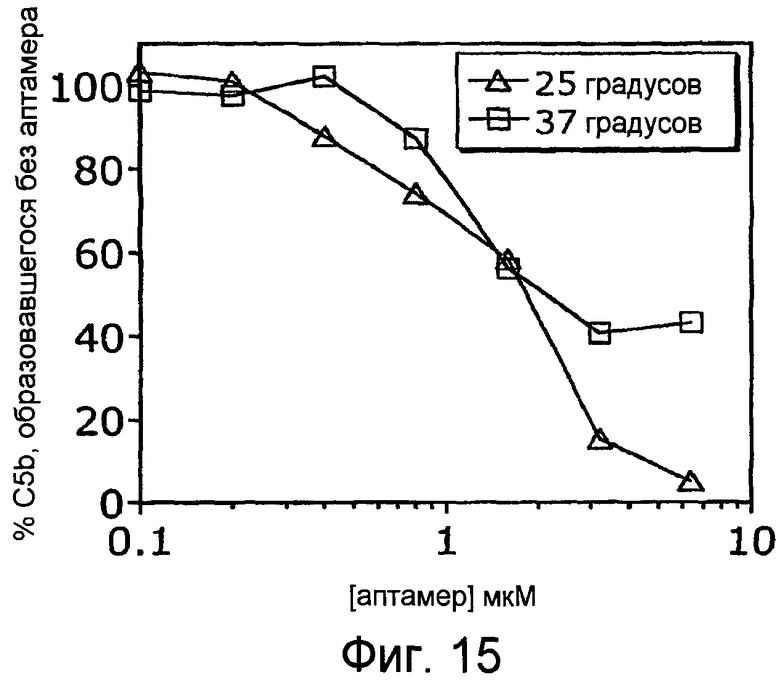
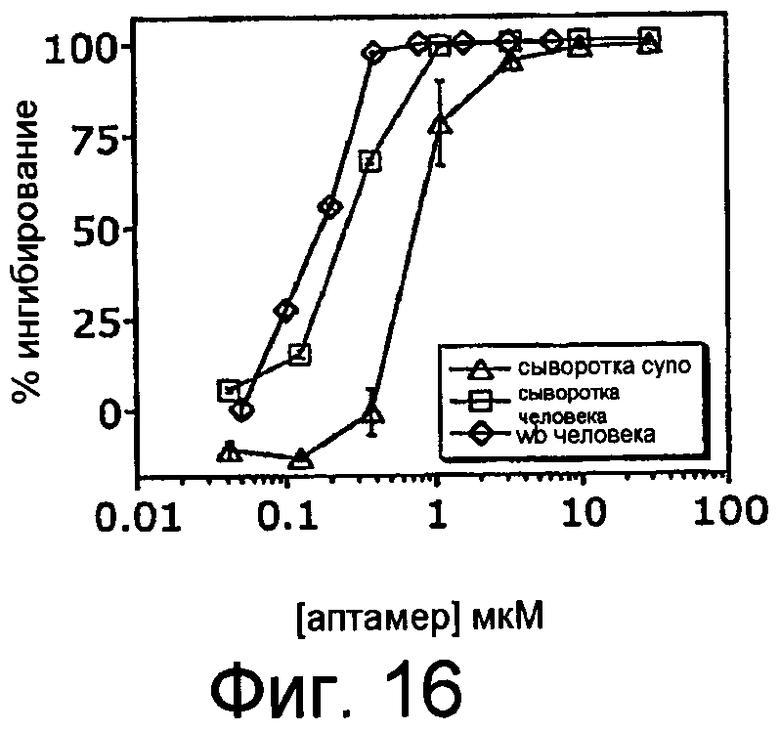
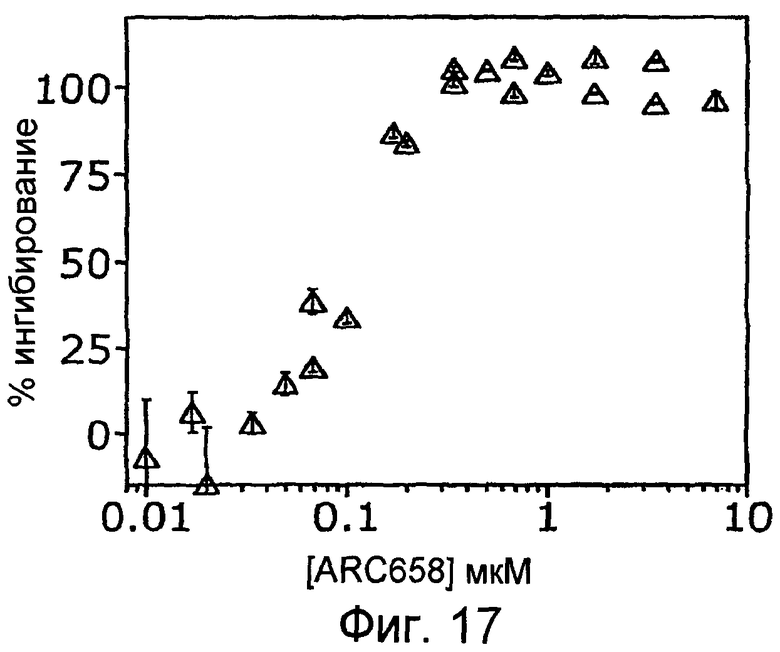
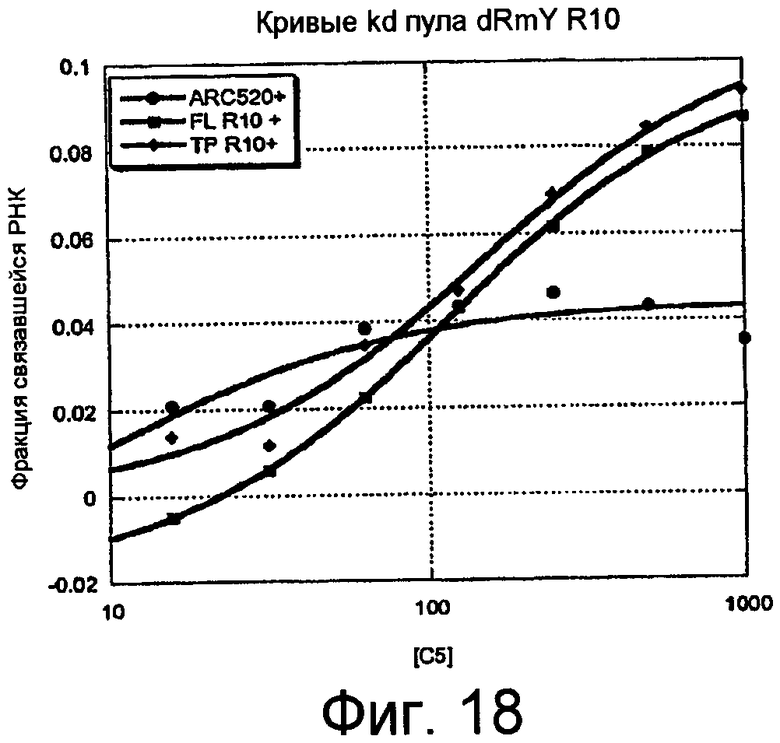
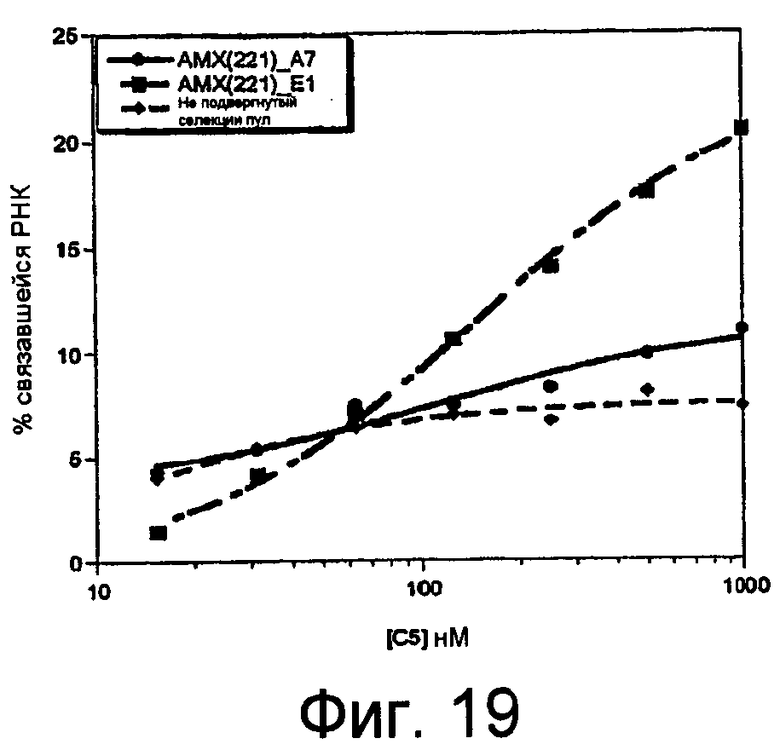
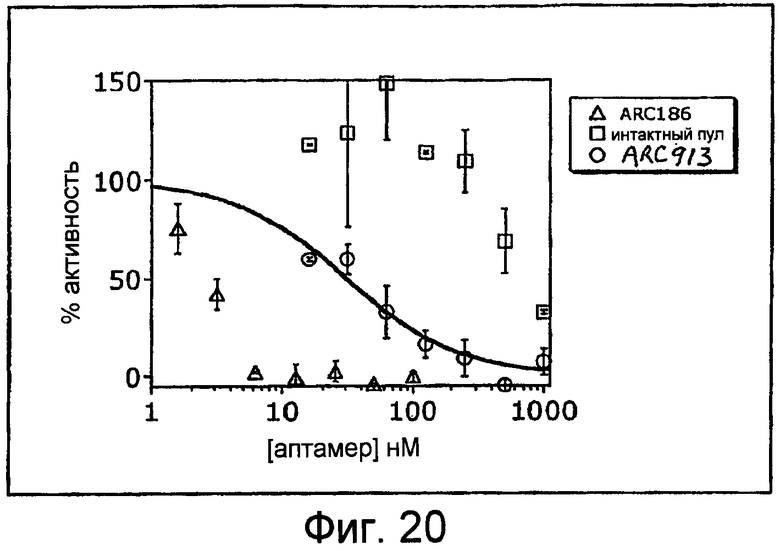
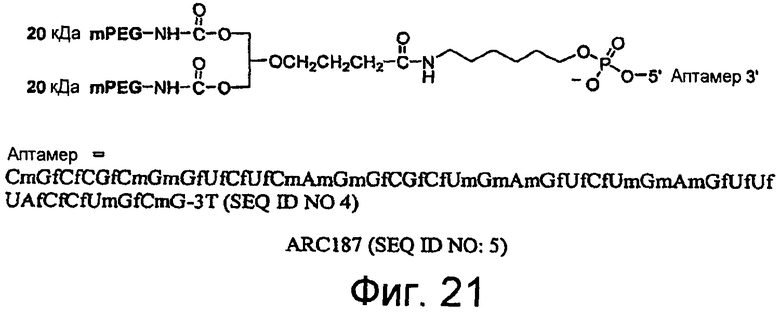
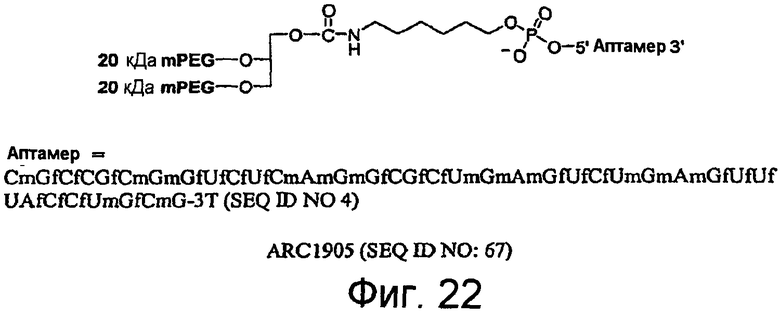

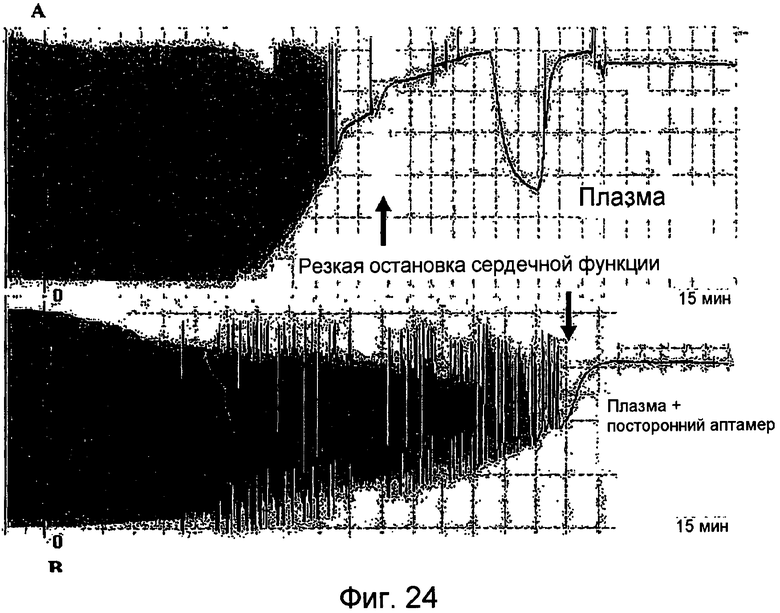
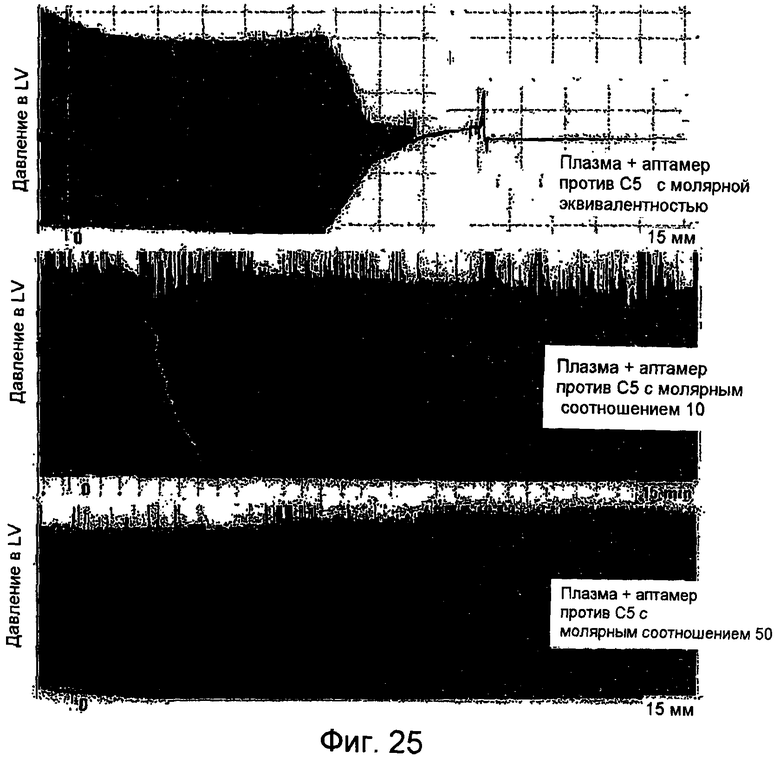
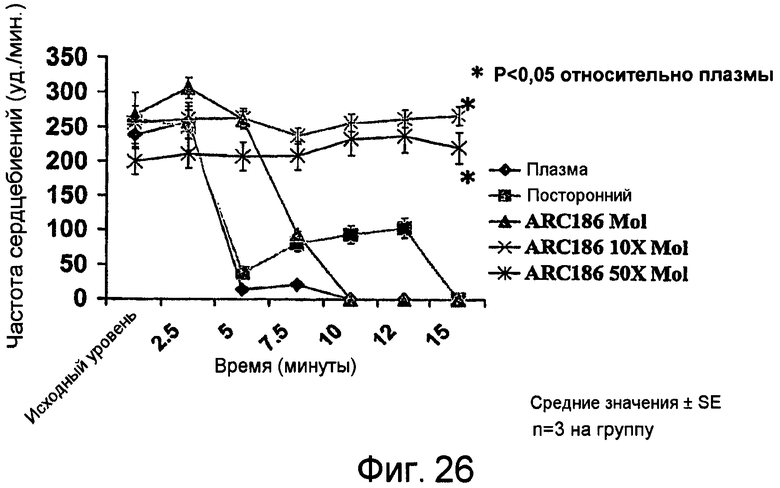
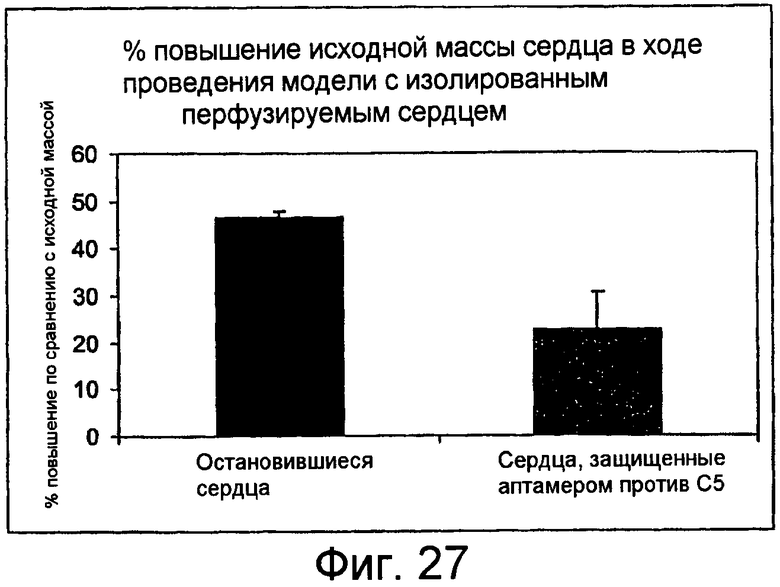
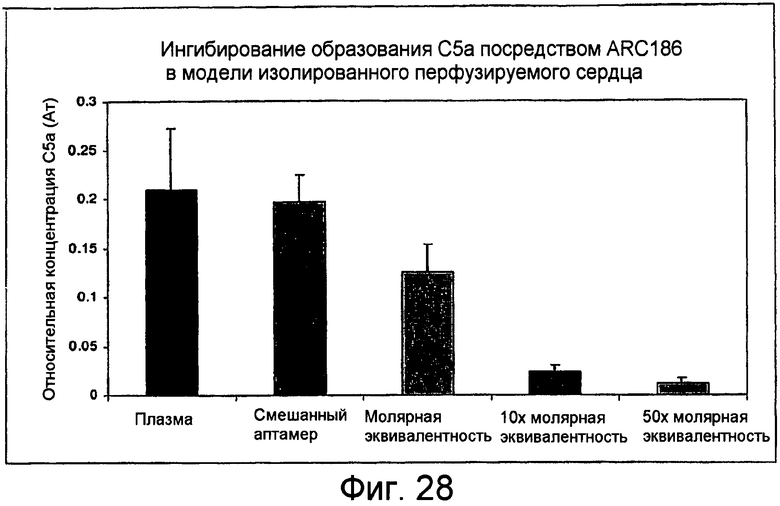
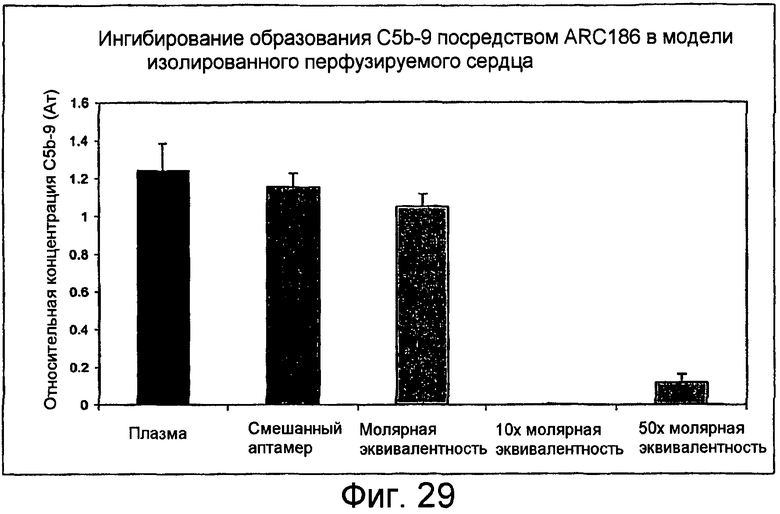
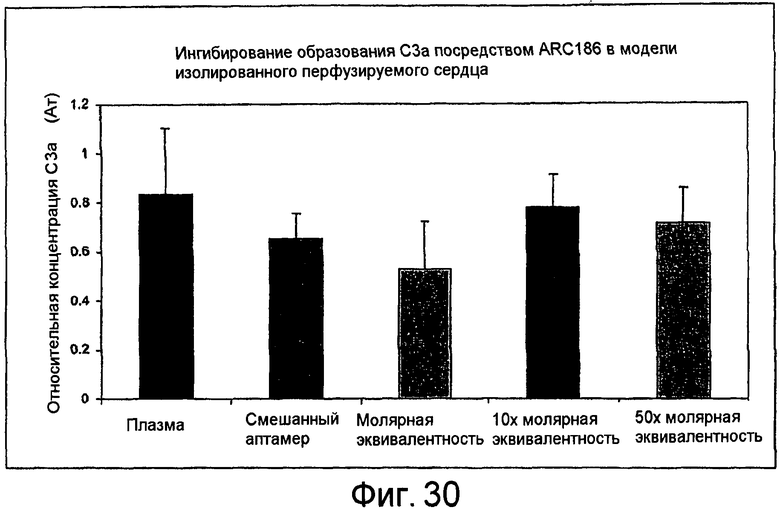

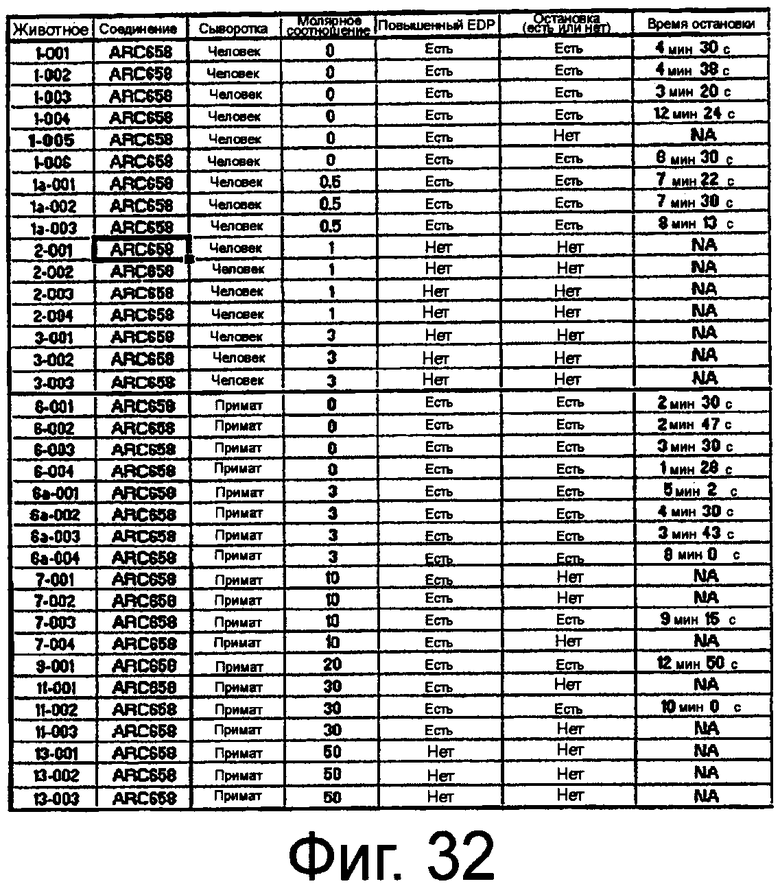
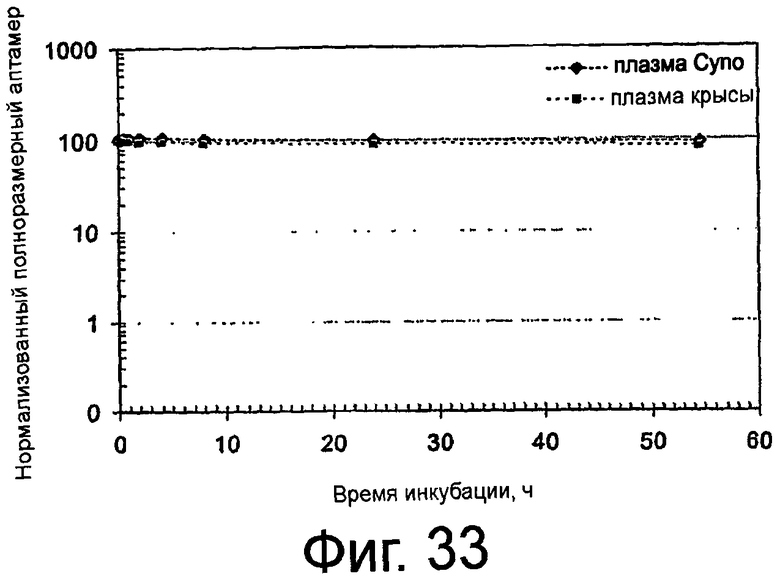

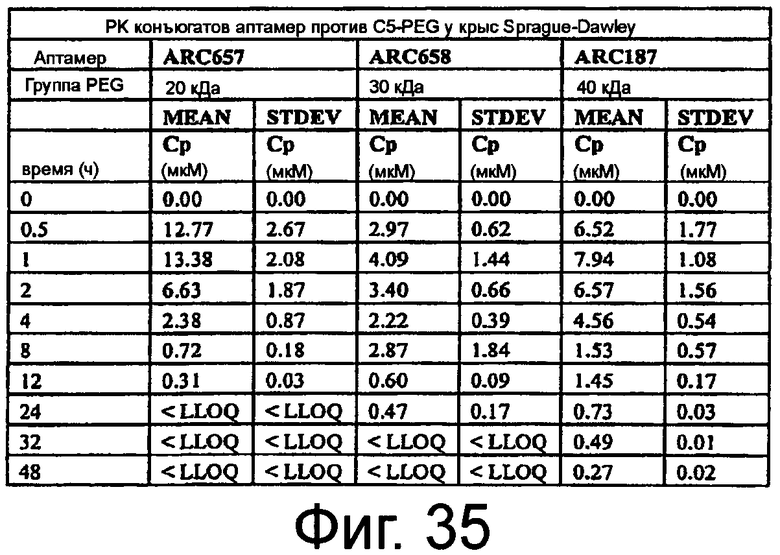
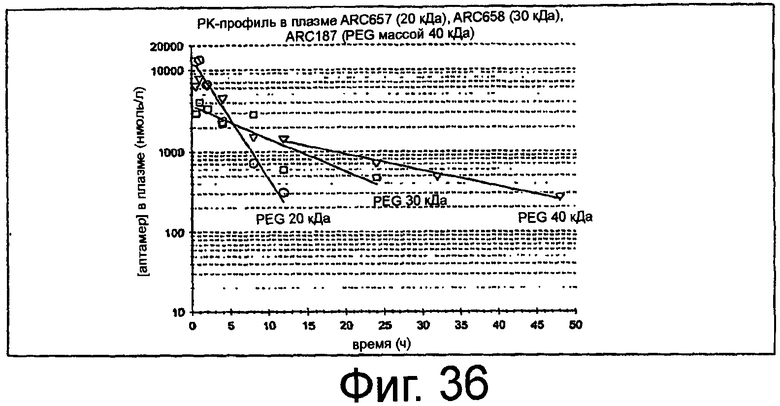
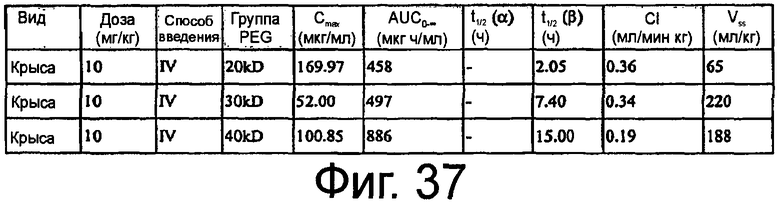
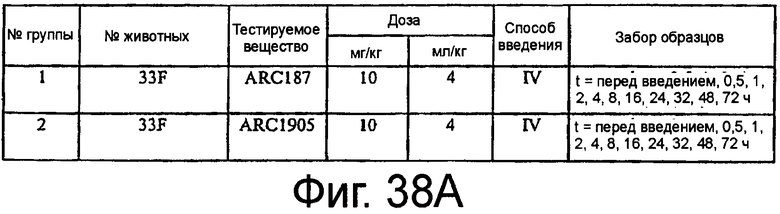
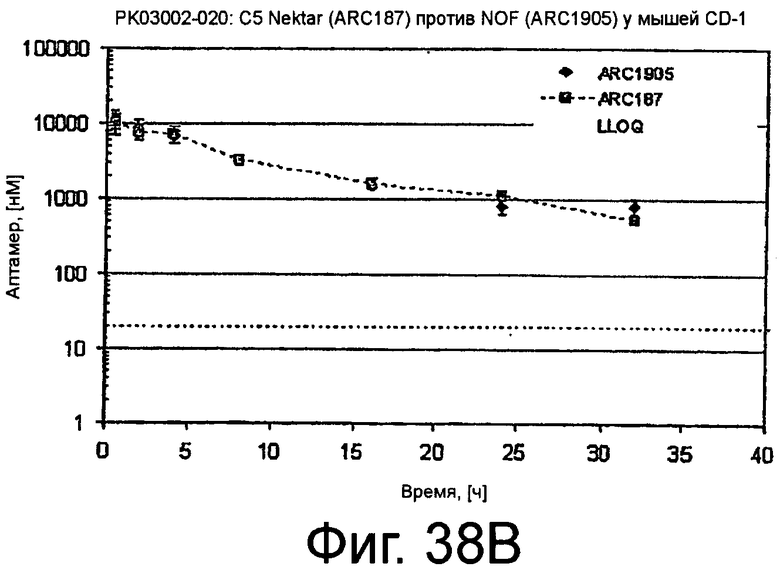
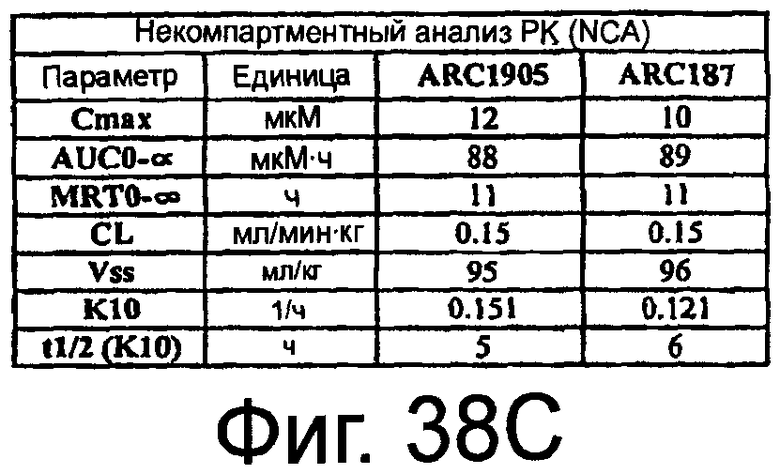
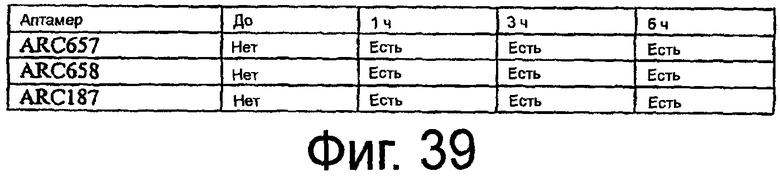
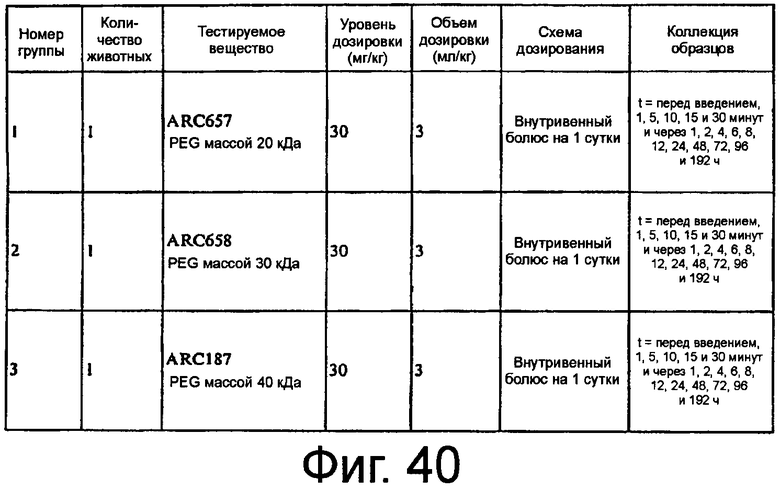
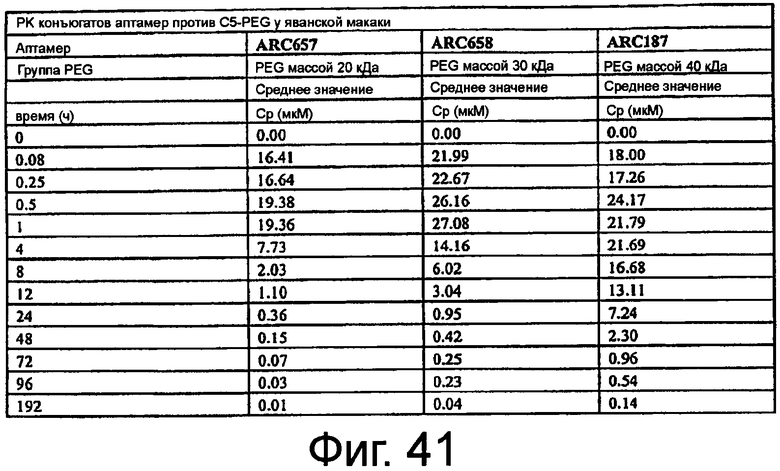
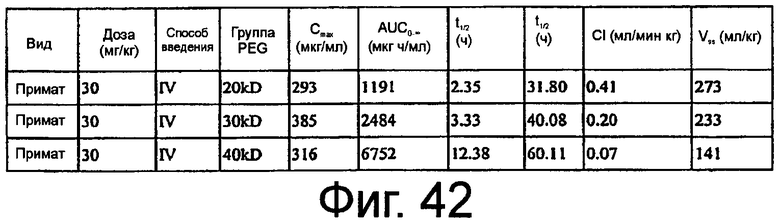
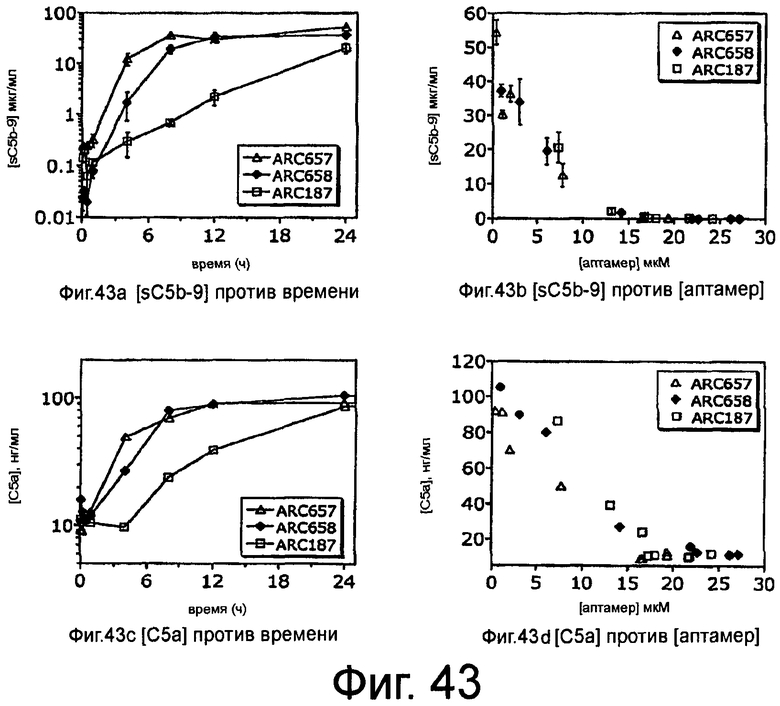
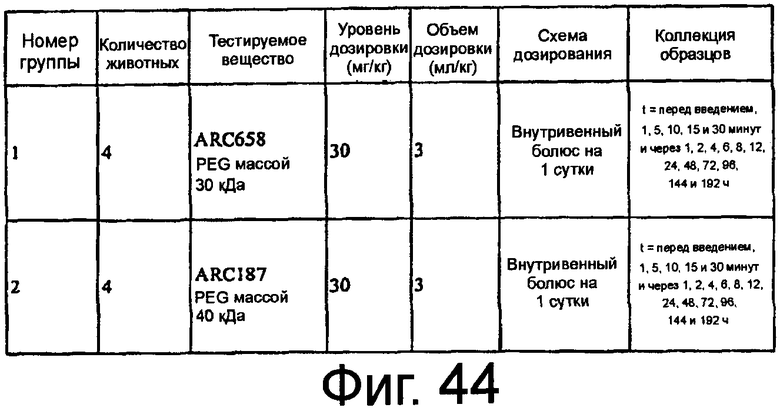
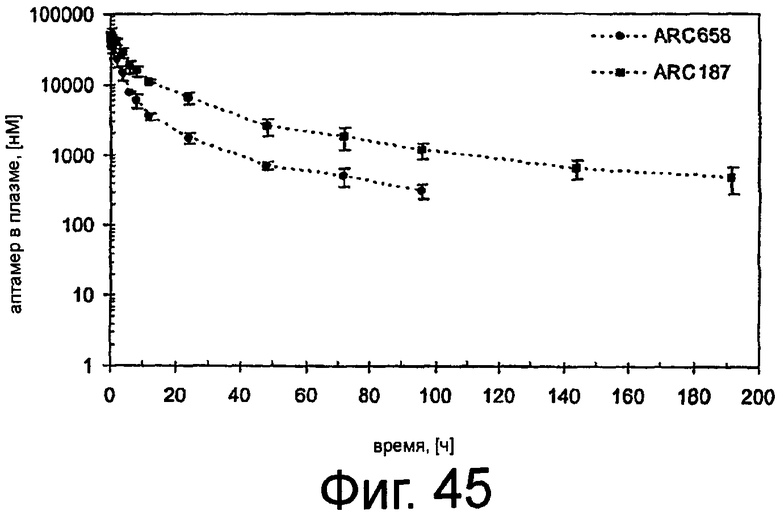
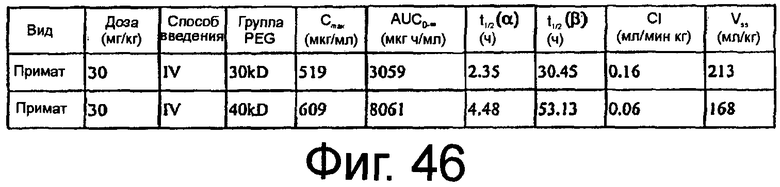
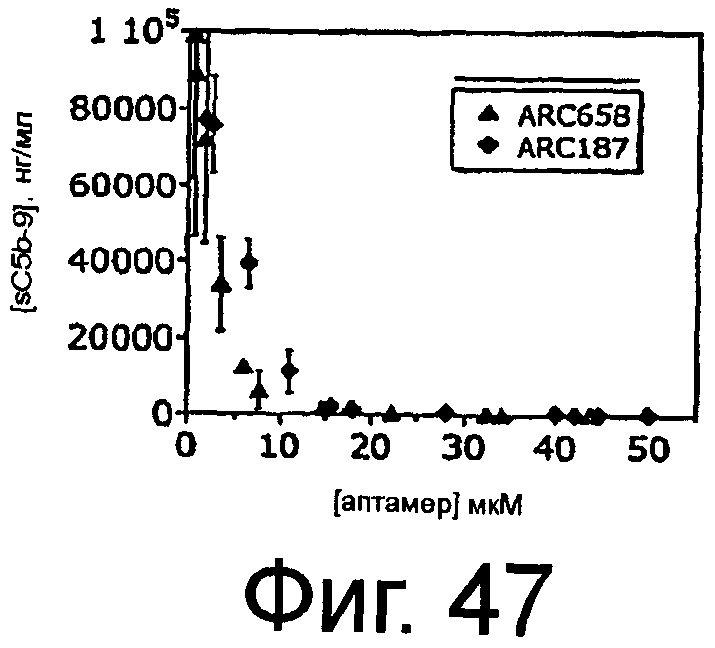
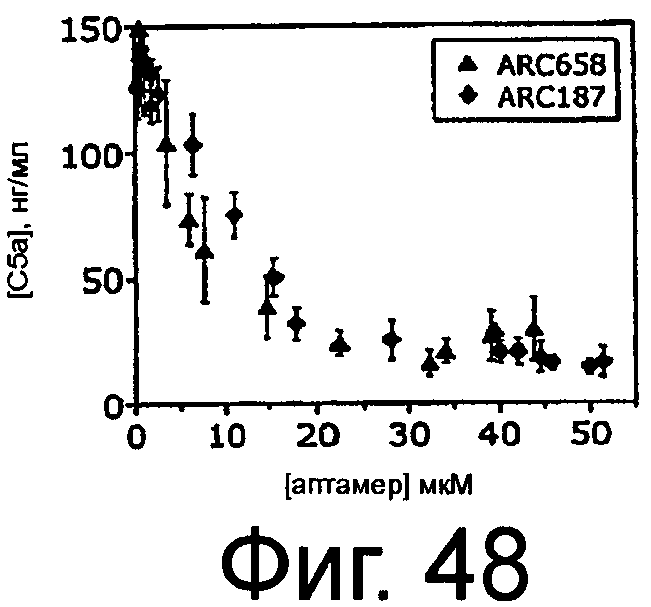
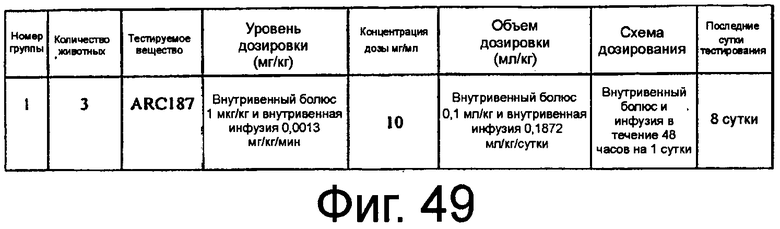
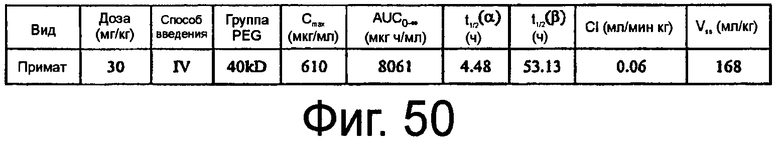
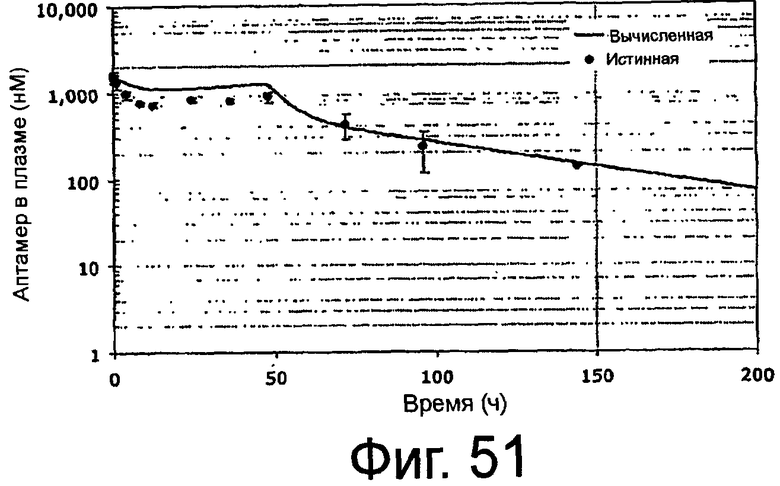
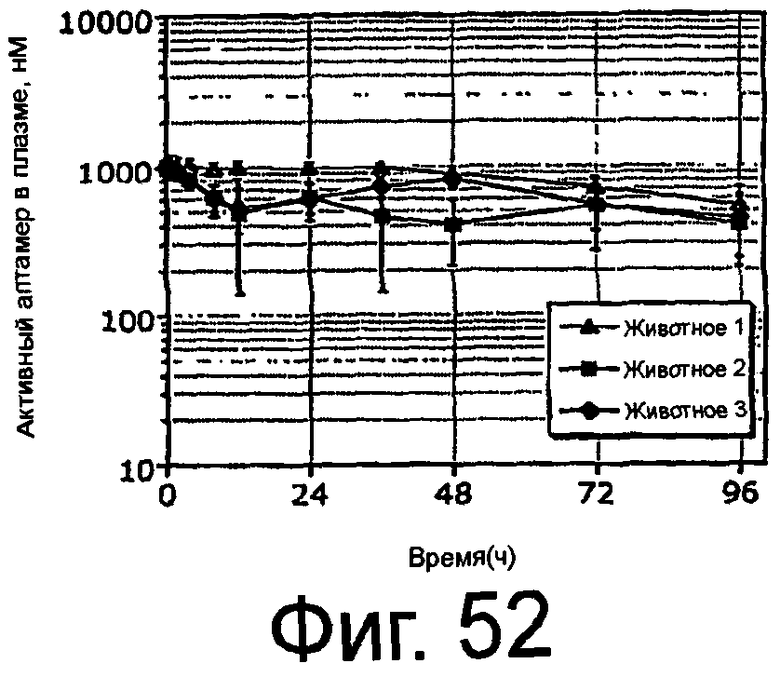
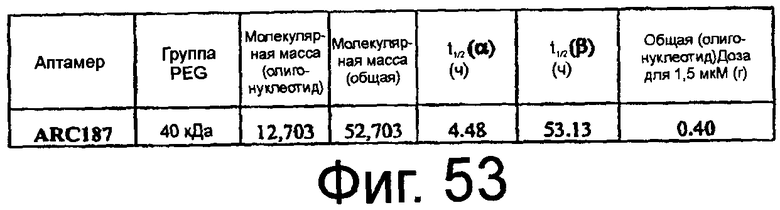
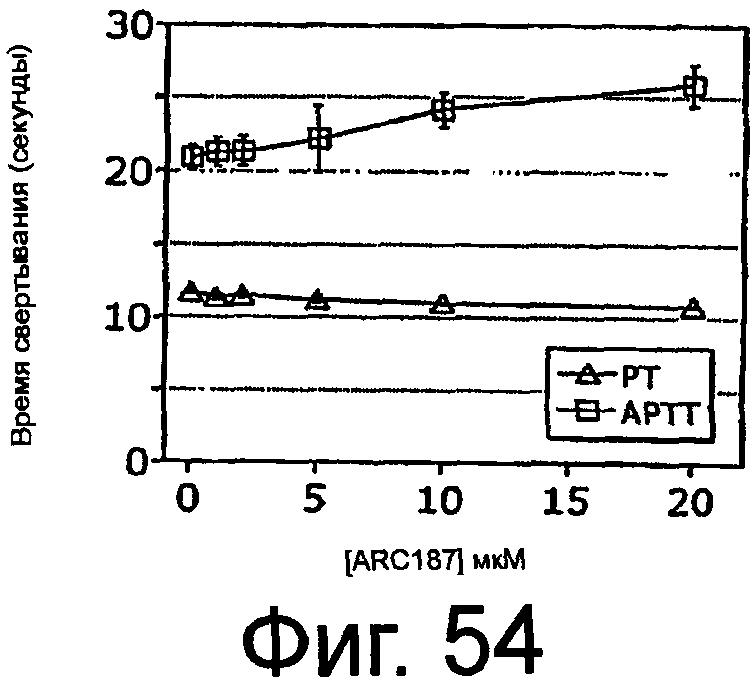

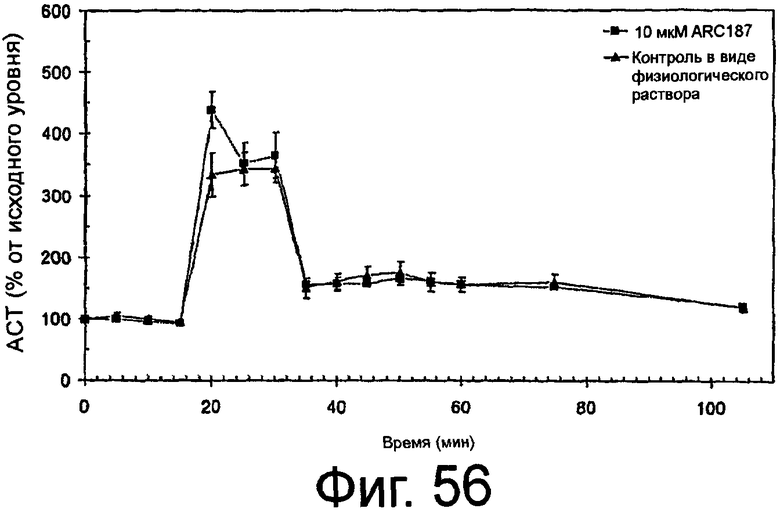
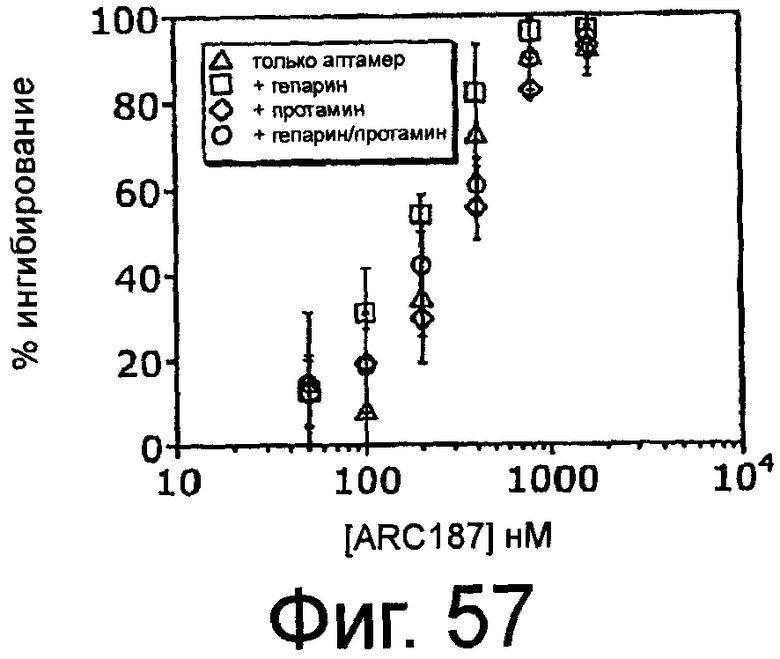
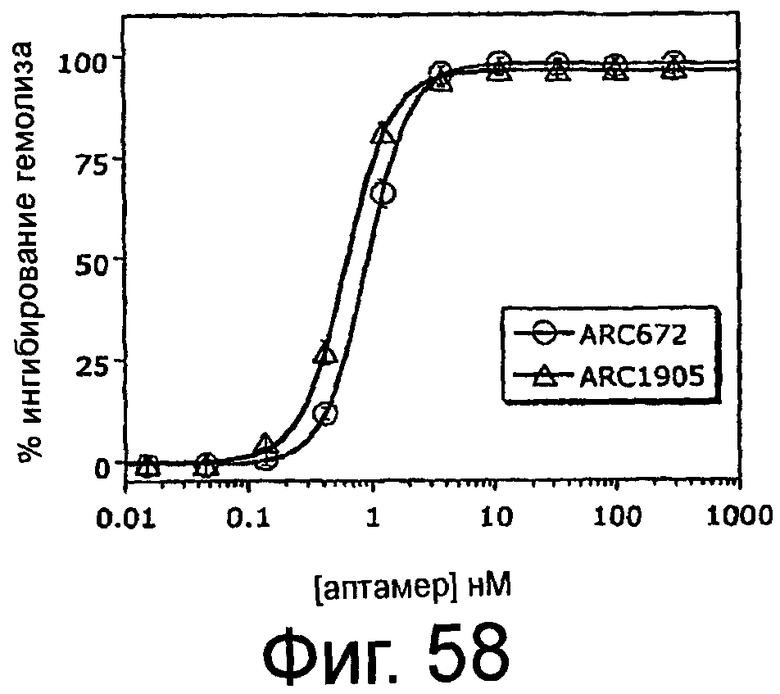
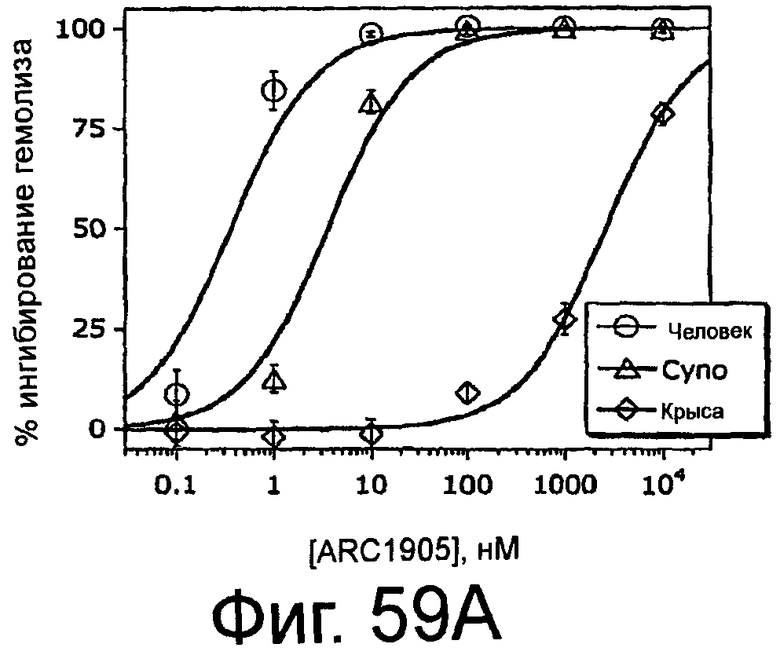
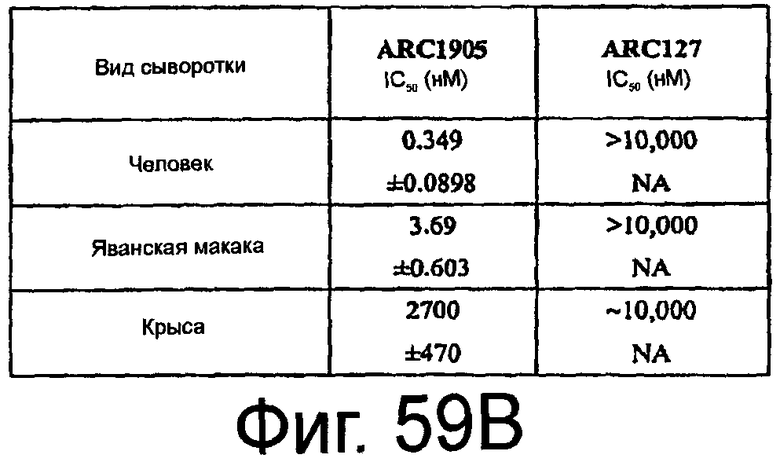
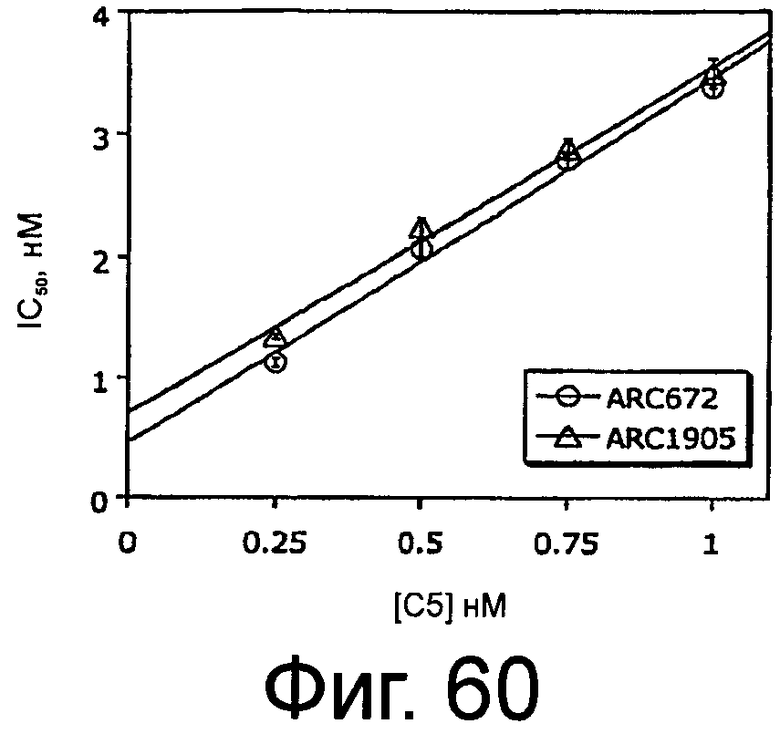
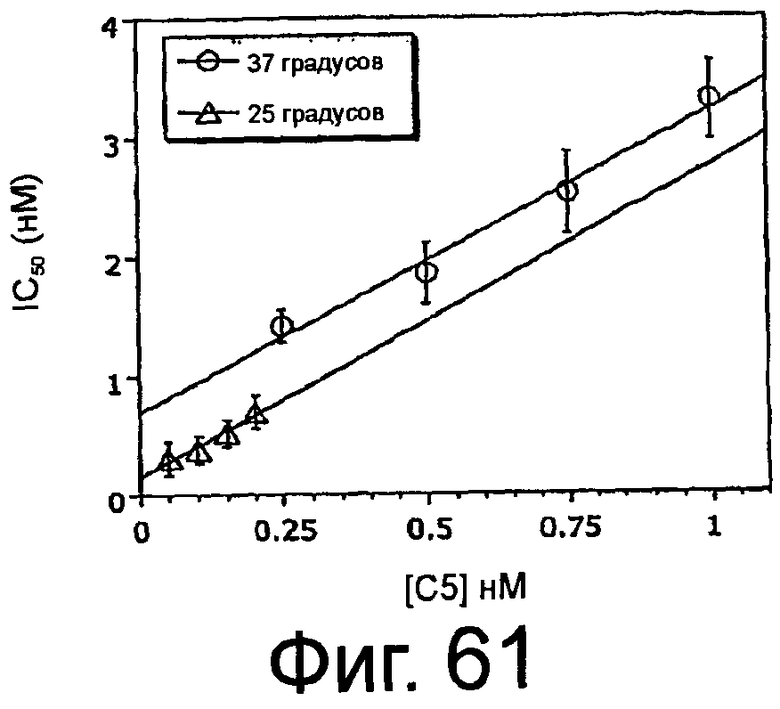
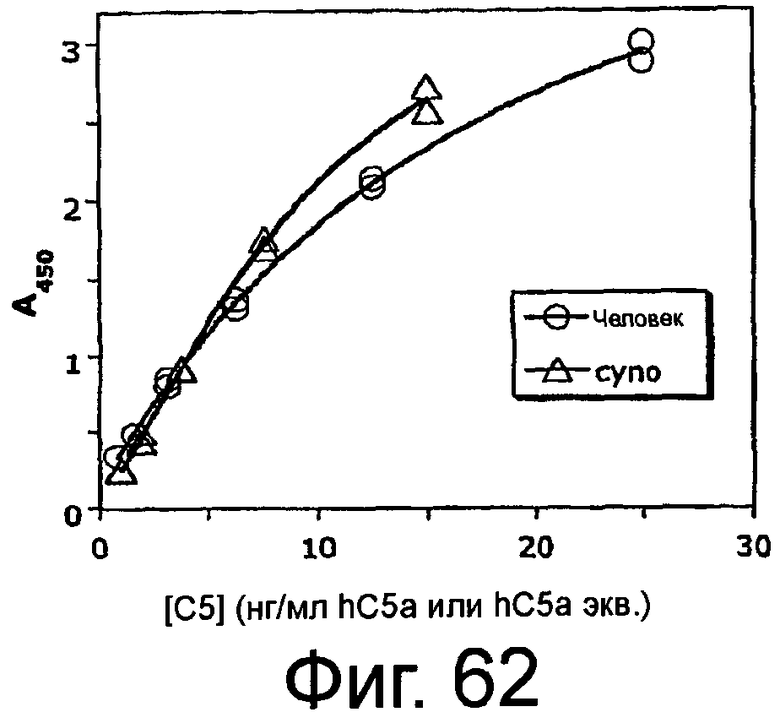

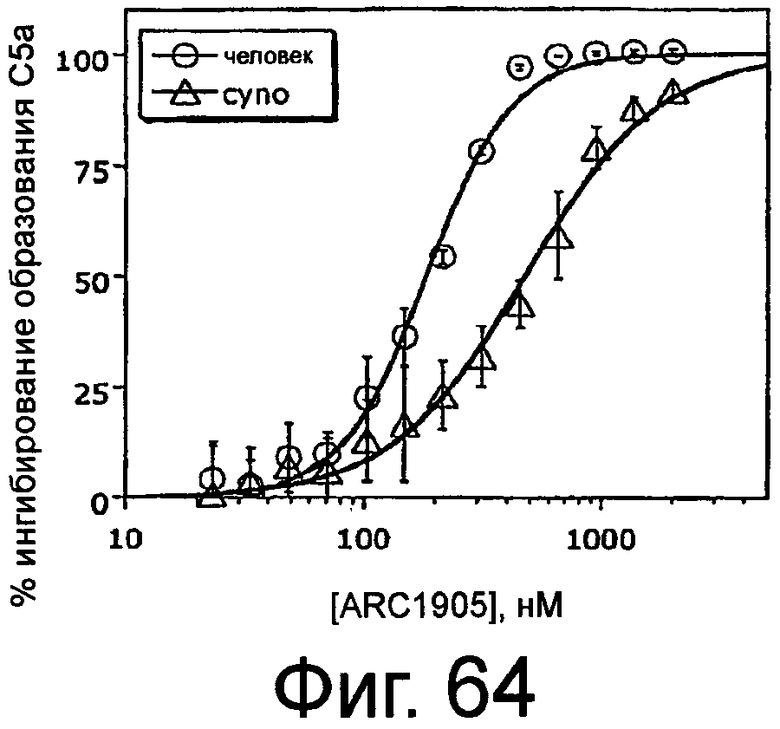
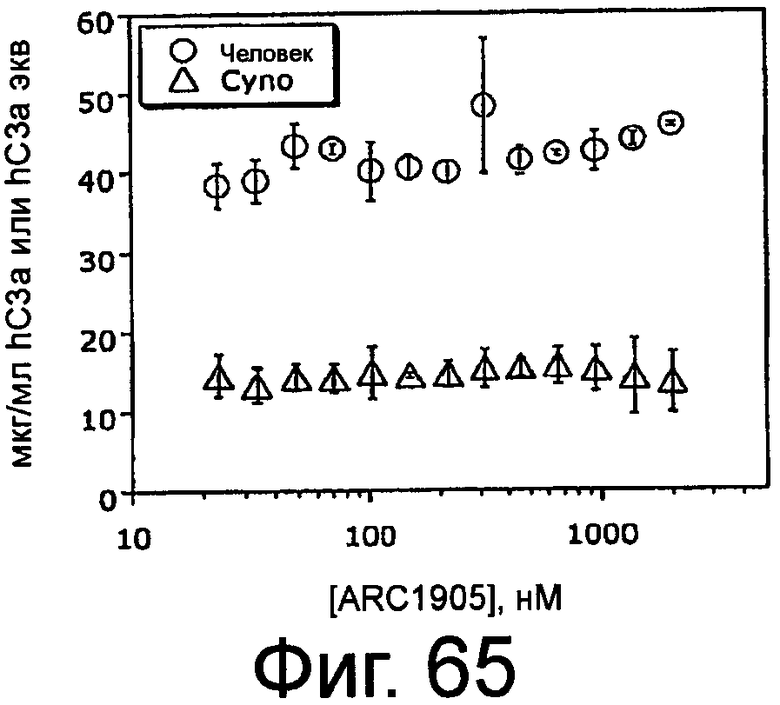
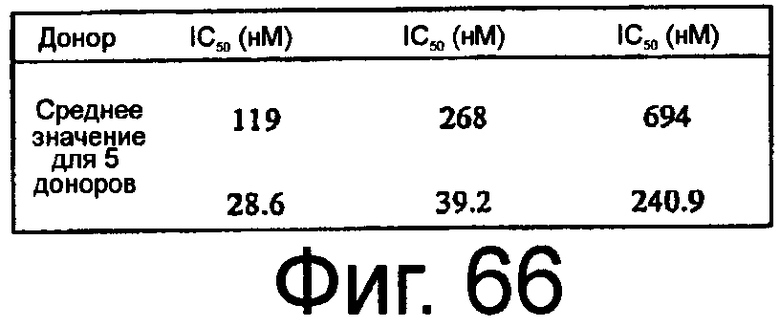
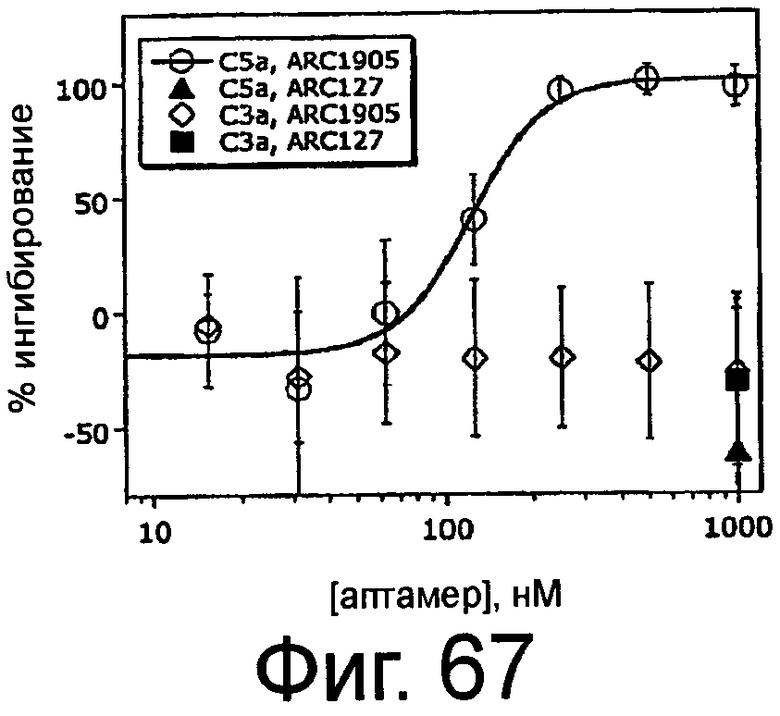
Изобретение относится к медицине, а именно к офтальмологии, и может быть использовано для лечения, стабилизации и/или профилактики дегенерации желтого пятна. Лечение включает введение терапевтически эффективного количества средства против С5 в комбинации со средством против VEGF субъекту, нуждающемуся в этом. При этом средство против С5 связывается с С5 комплементом и имеет последовательность SEQ ID NO:4 или SEQ ID NO:67. Изобретение обеспечивает аддитивный и синергетический эффект указанных средств за счет из комбинирования, что ведет к улучшению остроты зрения, особенно у пациентов с возрастной дегенерацией желтого пятна (AMD) «влажного» типа по сравнению с монотерапией против VEGF. 14 з.п. ф-лы, 67 ил., 8 пр.
1. Способ лечения, стабилизации и/или профилактики дегенерации желтого пятна, при этом способ включает введение терапевтически эффективного количества средства против С5 в комбинации со средством против VEGF субъекту, нуждающемуся в этом, где средство против С5 связывается с С5 комплементом и имеет последовательность SEQ ID NO: 4 или SEQ ID NO: 67.
2. Способ по п.1, где дегенерация желтого пятна, подлежащее лечению, представляет собой возрастную дегенерацию желтого пятна (AMD).
3. Способ по п.2, где AMD представляет собой AMD неэкссудативного типа или AMD экссудативного типа.
4. Способ по п.1, где средство против С5 доставляют внутриглазным введением.
5. Способ по п.1, где средство против С5 доставляют введением в стекловидное тело.
6. Способ по п.1, где средство против С5 доставляют посредством окологлазного введения.
7. Способ по п.1, где средство против С5, подлежащее введению, содержится в составе с замедленным действием.
8. Способ по п.1, где субъектом является человек.
9. Способ по п.1, где средство против VEGF представляет собой антитело антагониста или фрагмент антитела направленного против VEGF.
10. Способ по п.9, где антитело антагониста или фрагмент антитела направленного против VEGF представляет собой бевацизумаб или ранибизумаб.
11. Способ по п.9, где средство против VEGF представляет собой ранибизумаб.
12. Способ по п.9, где средство против VEGF представляет собой бевацизумаб.
13. Способ по п.1, где средство против С5 и средство против VEGF вводятся пациенту, нуждающемуся в этом в количестве, эффективном для стабилизации дегенерации желтого пятна, снижения симптома дегенерации желтого пятна или профилактики симптома дегенерации желтого пятна.
14. Способ по п.1, где средство против С5 и средство против VEGF вводятся последовательно или по существу одновременно.
15. Способ по п.1, где средство против С5 имеет следующую структуру:
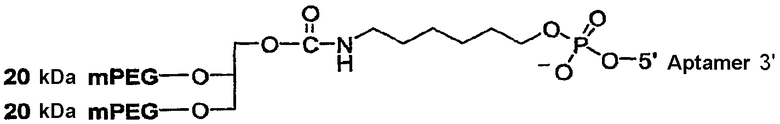
Аптамер=fCmGfCfCGfCmGmGfUfCfUfCmAmGmGfCGfCfUmGmAmGfUf
CfUmGmAmGfUfUfUAfCfCfUmGfCmG-3T (SEQ ID NO: 4),
где fC и fU=2'-фторнуклеотиды, и mG и mA=2'-ОМе-нуклеотиды и все другие нуклеотиды являются 2'-ОН, и ЗТ указывает на инвертированный дезокситимидин.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| СРЕДСТВО, ВОССТАНАВЛИВАЮЩЕЕ ФУНКЦИЮ СЕТЧАТОЙ ОБОЛОЧКИ ГЛАЗА | 1993 |
|
RU2073518C1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Nalini S | |||
| Bora et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

